d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění
|
|
|
- Stanislav Novák
- před 5 lety
- Počet zobrazení:
Transkript

1 MBRO ) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR


2 d) Vznik gamet Životní cyklus rostliny 2 Mikrosporogeneze Megasporogeneze

 - kóduje protein")

 Meióza (2x) Tetráda haploidních buněk 4 volné")

3 Vývoj samčího gametofytu - mikrosporogeneze 3 Mikrosporogeneze = tvorba samčího gametofytu = pylových zrn Sporofytická buňka Update 2015 Komunikace mezi vegetativní buňkou a generativní buňkou: ABA-hypersensitive germination3 (AHG3) - kóduje protein fosfatázu přepisována ve veget. buňce ale překládána v generativní buňce. Tapetální iniciační buňka Tapetum Sporogenní iniciační buňka (pylová mateřská) Meióza (2x) Tetráda haploidních buněk 4 volné mikrospory Mitóza Kalóza = 1,3-ß-glukan Vegetativní buňka Generativní buňka Kaláza = 1,3-ß-D-glukanáza Tapetální buňky produkují kalázu, polymery a pigmenty 2 spermatické buňky Mitóza



 Plant Cell, Tissue")
4 4 Samčí pohlavní buňka - pylové zrno Exina = sporopolenin = polymer fenolů; extrémně rezistentní k chemickým látkám; geny, které utváření exiny kódují, jsou málo známy Fosilní nálezy pylových zrn Vývojová biologie Pylové protoplasty Fellner M (1995) Plant Cell, Tissue and Organ Culture 42:
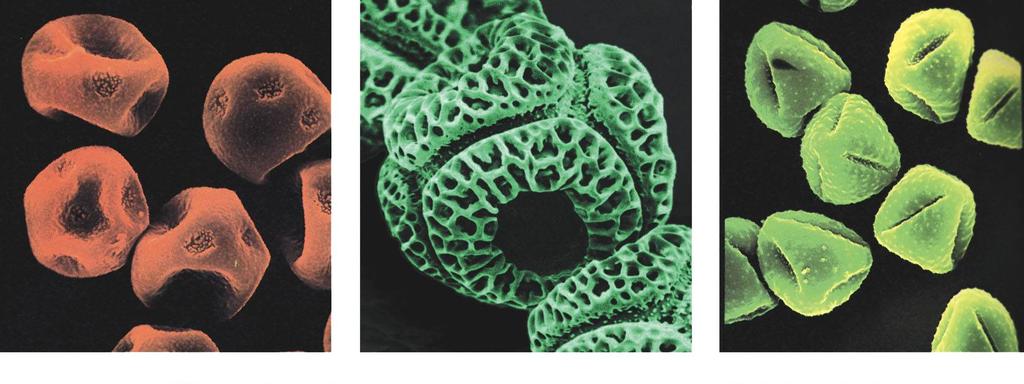
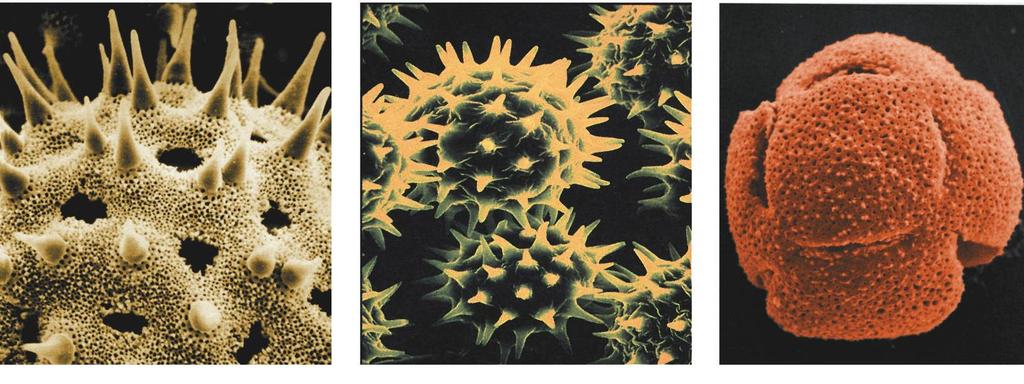
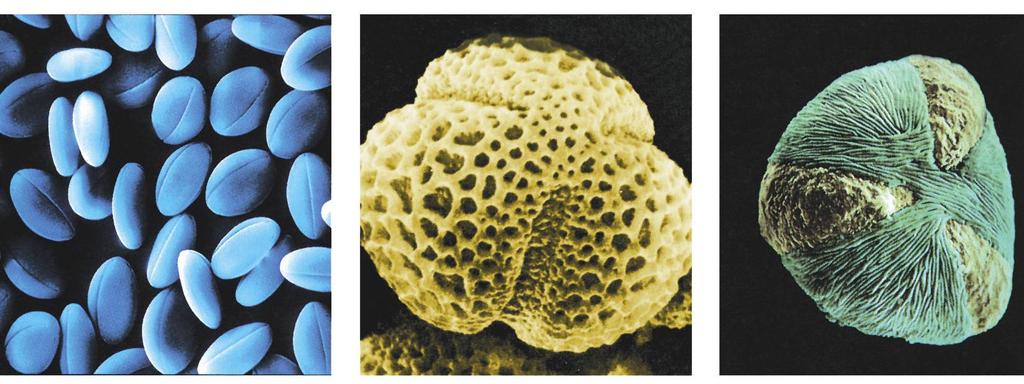

5 5 Pestré uspořádání exiny pylových zrn Praktické využití (např. kriminalistika)
")

 1 centrální")

6 Vývoj samičího gametofytu - megasporogeneze 6 Meióza (2x) Mitóza (3x) Nezralé vajíčko Megaspora Embryonální vak: 7 buněk 3 antipodální (AC) 2 synergické (SC) 1 centrální (CC) 1 vaječná (EC) PN 2 polární jádra SEN jádro sek. endospermu


7 Je problematické najít geny specificky exprimované v samičím gametofytu: problematické izolovat vajíčko od sporofytického pletiva => obtížné izolovat mrna a vytvořit cdna knihovny 7 Orchidej - experimentální rostlina pro studium genů specificky exprimovaných ve vajíčku - synchronizovaný vývoj vajíček - vajíčko se vyvíjí dlouhou dobu (11 týdnů) => možnost izolovat vajíčka v různých stádiích vývoje => mrna v různých etapách vývoje => možnost determinovat expresi během vývoje Nadeau JA et al. (1996) Plant Cell 8:
8 e) Mutace ve vývoji gametofytu 8 Analýza sterilních mutantů Identifikace genů Pylově sterilní mutanti s defektem ve vývoji pylu - většinou recesivní, homozygotní - defekt v meióze, netvoří pyl - defekt ve vývoji tapetálních buněk - deformace prašníků pyl se neuvolňuje, nebo pozdě - defekt v pylovém otvoru pyl nemůže klíčit - defekt ve vývoji sporofytu pyl nemůže klíčit Sterilní mutanti s defektem ve vývoji vajíčka - defekt ve vývoji sporofytu - defekt ve vývoji megaspory - ovlivněn vývoj vajíčka a ovlivněno oplodnění - defekt ve vývoji embryonálního vaku a vajíčka ovlivněno oplodnění
9 9 Mutace ve vývoji samčího gametofytu 60 90% genů exprimovaných v gametofytu je exprimováno i ve sporofytu Sekvenování Arabidopsis genomu (2000) 26 tisích genů 350 genů exprimovaných specificky v prašnících Specifické pro pyl Nespecifické pro pyl
 (např.")
10 TDF1 (DEFECTIVE IN TAPETAL DEVELOPMENT AND FUNCTION 1) transkripční faktor MYB; klíčový v regulaci vývoje tapeta 10 DYT1 (DYSFUNCTIONAL TAPETUM 1) protein DYT1 se váže přímo na promotorovou oblast genu TDF1 WT tdf1 Trangen TDF1 DYT1 Promotor TDF1 Exprese genů tvorby pylové stěny (exiny) (např. AMS, MS1, TEK, MS188) TDF1 - DYT1 je lokalizován v cytoplazmě i jádře - C-terminální doména BIF je nezbytná pro dimerizaci, nukleární lokalizaci a schopnost aktivovat transkripci - Prostorově časová subcelulární lokalizace DYT1 je důležitá pro samotnou funkci DYT1
 dráhy zapojený v biosyntéze sterolů MVA důležitý pro vývoj organel tapetálních buněk => pylová zrna fkp1")
; exprimován v prašníkových pletivech před koncem")
11 fkp1 pylová zrna bez povrchové vrstvy 11 FKP1 (FLAKY POLLEN 1) kóduje 3-hydroxy-3-methylglutaryl-coenzym A syntázu = enzym mevalonátové (MVA) dráhy zapojený v biosyntéze sterolů MVA důležitý pro vývoj organel tapetálních buněk => pylová zrna fkp1 nemají povrchovou vrstvu, která vzniká z rozpadlých tapetálních buněk quartet (qrt) tetrády se nerozdělují a uvolňují se celé z prašníku QRT kóduje enzym s pektin metylesterázovou aktivitou (PME); exprimován v prašníkových pletivech před koncem meiózy


12 12 T-DNA mutant cals5 narušená fertilita, degenerované mikrospory CALS5 kóduje kalóza-syntázu syntéza kalózy exina WT cals5-3 cals5-4 cals5-5 Knockout mutant CYP703A2 (cytochrom P450) CYP703A2 katalyzuje hydroxylaci kys. laurové Sporopolenin bloky hydroxylované kyseliny laurové WT CYP703A2

 transkripční faktor regulující tvorbu exiny, pyl.")
13 MGP1 (MALE GAMETOPHYTE DEFECTIVE 1) kóduje F a d podjednotku mitochondriální F 1 F 0 -ATP syntázy u Arabidopsis. 13 mgp1 mutant destrukce mitochondrií v pylových zrnech a k zániku pylových zrn MS1 (MALE STERILITY1) transkripční faktor regulující tvorbu exiny, pyl. cytosolu a tapeta MS2 (MALE STERILE2) kóduje reduktázu mastných kyselin nutnou pro tvorbu exiny
 membránový protein nezbytný pro tvorbu exiny Guan Z-F et al.")
14 MGT4, MGT5, MGT9 (MAGNESIUM TRANSPORTER 4, 5, 9) nezbytné pro vývoj pylu Arabidopsis; mgt4-1 nevyvinutá pylová zrna, rostliny sterilní 14 MGT4 lokalizován v ER, exprimován v pylových zrnech ve dvoubuněčném stádiu až do zralého pylového zrna RPG1 (RUPTURE POLLEN GRAIN1) membránový protein nezbytný pro tvorbu exiny Guan Z-F et al. (2008) Plant Physiol 147: WT rpg1 TIR1, AFB1 AFB3 auxinové receptory; mutanti vytváří krátké tyčinky prašníku a předčasné zrání pylu

 + (funkce známa) 9")
15 15 Mutace ve vývoji samičího gametofytu Mutace zapojené v meiotickém a mitotickém buněčném cyklu samičí gametofyt 23 genů 14 nespecifických ( ) + (funkce známa) 9 specifických ( )


16 Specifické geny 16 Arabidopsis mutant fem2 zastaven vývoj vajíčka před megagametogenezí Arabidopsis mutant gfa nedochází k fúzi jader centrální buňky fem2 gfa Arabidopsis mutant gcd1 narušené zrání vaječné buňky Mitochondriální protein GCD1 je nezbytný pro konečné zrání samičích gamet.
 Science 324: 1684-1689 Distribuce auxinů v embryonálním vaku je")

17 17 Pagnussat GC et al. (2009) Science 324: Distribuce auxinů v embryonálním vaku je polarizovaná - tvoří se gradient koncentrace auxinů. Na základě tohoto gradientu auxin určuje identitu buněk: Vysoká koncentrace auxinu Nízká koncentrace auxinu Synergické buňky a vaječná buňka Antipodální buňky
18 18 Cytoplazmatická samčí sterilita (CMS) = pylová sterilita přenášená pouze samičími orgány Odpovědné geny jsou většinou součástí chloroplastového či mitochondriálního genomu. Ve všech známých případech je CMS je způsobena expresí abnormálních proteinů v mitochondriích prašníků. Mechanismus, jakým abnormální proteiny ovlivňují mitochondrie, není znám. Mitochondrie v prašníku mají vliv na vývoj pylu. Pokud je exprese abnormálního proteinu redukována, fertilita je obnovena. Ve všech CMS systémech existují jaderné geny, které potlačují expresi (tvorbu) abnormálních proteinů v prašníku.


19 CMS-T systém u kukuřice abnormální mitochondriální protein URF13 19 Obnovení fertility vyžaduje 2 jaderné geny: Rf1 a Rf2 Aldehyd dehydrogenáza Aldehyd?
20 20 Klíčení pylu Pro klíčení vyžaduje vysušené pylové zrno vlhkost 1) Rostliny s vlhkou bliznou pylové zrno bere vlhkost z blizny 2) Rostliny se suchou bliznou vlhkost je zajištěna lipidy na povrchu pylového zrna. Lipidy hrají významnou roli v klíčení pylu. Arabidopsis mutant cer má defektní lipidovou vrstvu klíčí pouze za extrémní vlhkosti Arabidopsis mutant fiddlehead má odlišné lipidy (vysokomolekulární) v epidermálních buňkách listů. Pylová zrna WT na těchto listech klíčí!!! Flavonoidy na povrchu pylového zrna hrají roli v klíčení pylu. Rostliny kukuřice s mutací v genu, kódujícím enzym biosyntézy flavonoidů, jsou self-sterilní.



21 21 Mechanismus klíčení pylu není dosud přesně znám Ca 2+ gradient pozorován při růstu pylové láčky

22 22 Není známo, jak je Ca 2+ signál přeložen do finální reakce prodlužování pylové láčky. cngc18 nulový mutant s pylovou sterilitou CNGC18 kóduje kationtový kanál regulovaný cyklickými nukleotidy GFP:CNGC18 analýza cngc18: komplementovaný GFP:CNGC18 Frietsch S et al. (2007) PNAS 104:
 Tonoplast Vakuolární hlavičky Exprese TIP1;3-GFP v klíčícím")
23 TIP1;3, TIP5;1 jsou nejrozšířenější vysoce exprimované proteiny ve zralých pylových zrnech 23 TIP1;3 exprimován ve vezikulech a vakuolách vegetativní buňky TIP5;1 exprimován ve vakuolách spermatických buněk Tetráda Mikrospora 2-buněčný pyl 3-buněčný pyl Exprese TIP1;3-GFP ve zrajícím pylovém zrně (jádra obarvena modře) Tonoplast Vakuolární hlavičky Exprese TIP1;3-GFP v klíčícím pylovém zrně; exprese v tonoplastu vegetativní buňky a vakuolárních hlavičkách pylové láčky Mutanti tip1;3 a tip5;1 ukazují krátké pylové láčky pouze v podmínkách při nedostatku dusíku Update 2016 Di Giorgio JAP et al. (2016) Plant Cell 28: TIP1;3 a TIP5;1 jsou zapojeny v metabolické dráze N během klíčení pylu NIP4;1, NIP4;2 nezbytné pro vývoj pylu a opylení; mutanti - nízký počet semen, redukované klíčení pylu, redukovaný růst pylové láčky; NIP4;1 velice nízká exprese ve zralém pylu, NIP4;2 nejvyšší exprese během růstu pylové láčky C-terminální domény NIP4;1, NIP4;2 fosforylovány pylově specifickými CPK (Ca 2+ - dependentní protein kinázy) => modifikace permeability pro H 2 O NIP4;1 transportuje rovněž amoniak, močovinu, H 3 BO 3, H 2 O 2



24 Klíčení se dá indukovat in vitro na médiu obsahujícím cukr, kys. boritou, Ca Klíčení in vivo je vždy rychlejší Další faktory (?) pocházející z blizny, hrající roli v klíčení pylu
 WT Mutant bri1 BRI1")
25 Další faktory ovlivňující klíčení pylové láčky rostlinné hormony 25 Gibereliny stimulují prodlužovací růst GA deficientní mutanti Mutanti s defektem v GA signaling trpasličí vzrůst, defekt ve vývoji prašníků a pylu Overexprese enzymu deaktivující GA Inhibice růstu pylové láčky Brasinosteroidy stimulují prodlužovací růst Mutant cpd CPD kóduje cytochrom P450 (biosyntéza BRs) WT Mutant bri1 BRI1 kóduje receptor BRs Zastaveno prodlužování pylové láčky BRs a BRs signaling nutné pro růst pylové láčky bri1 Clouse et al. (1996) Plant Physiol 111:
26 f) Opylení, oplodnění 26 Obě spermatické buňky vnikají do jedné ze synergických buněk (SC). Dochází k dvojímu oplodnění: 1. Oplodnění: 1. spermatická buňka oplodní haploidní vaječnou buňku (EC) => diploidní zygota PN CC 2. Oplodnění: 2. spermatická buňka oplodní diploidní centrální buňku (CC) = spojí se s jádry (PN) => triploidní endosperm EC SC
 Current Opinion in Plant Biology 16: 1-8 Update 2016 Peptidy sekretované synergickou buňkou: Pereira AM et al.")
27 1) Co nutí a směřuje pylovou láčku k embryonálnímu vaku? 27 PN CC EC SC Beale KM, Johnson MA (2013) Current Opinion in Plant Biology 16: 1-8 Update 2016 Peptidy sekretované synergickou buňkou: Pereira AM et al. (2016) Molecular Plant 9: Kukuřice: EGG APPARATUS1 (ZmEA1) Torénie: LURE1/LURE2 Arabidopsis: AtLURE1 Mutant jagger synergidy nedegenerují, neustálá produkce LURE => polytubey JAGGER kóduje arabinogalactan protein AGP4 (typ glykoproteinu); pravděpodobně vychytává Ca 2+ ; oscilace hladiny Ca 2+ je důležitá právě pro proniknutí pylové láčky, uvolnění spermatických buněk a oplodnění.
28 28 Liu J et al. (2013) Current Biology 23: 1-6 LIP1 a LIP2 (Lost In Pollen tube guidance 1 a 2) kódují cytoplazmatickou receptor-like kinázu; ukotvená k neznámému transmembránovému proteinu receptoru. Neznámý receptor přijímá peptidy LURE a EA1. Vypnutí LIP1 a LIP2 vede ke ztrátě orientace růstu pylové láčky.
 Dimer proteinů RLK MIK1 a MDIS1; vazba LURE1 vede ke vzniku dimeru a k autofosforylaci MIK1 a přenosu P z MIK1 na MDIS1 2) RLK PRK6 interaguje s druhým PRK6 nebo s PRK3,")
29 Identifikován receptor atraktantů LURE1 a 2 29 Update 2016 Takeuchi M, Higashiayma T (2016) Nature 531: Wang T et al. (2016) Nature 531: Wang T et al. Takeuchi M, Higashiayma T 1) Dimer proteinů RLK MIK1 a MDIS1; vazba LURE1 vede ke vzniku dimeru a k autofosforylaci MIK1 a přenosu P z MIK1 na MDIS1 2) RLK PRK6 interaguje s druhým PRK6 nebo s PRK3, s quanin-exchange faktorem (GEFs), který aktivuje RhoGTPase protein from plants (ROPs). Vazba LURE na tyto transmembránové receptory => nasměrování růstu a vniknutí pylové láčky do synergidy

 Neznámý ligand + receptor FERONIA Zastavení růstu pylové láčky, uvolnění")
30 2) Co způsobuje přijetí pylové láčky, její prasknutí a uvolnění spermatických buněk? 30 Mutant feronia pylová láčka vniká do synergické buňky, ale nepraská a neuvolňuje spermatické buňky Synergické buňky exprimují protein FERONIA (FER, receptor-like kináza) Neznámý ligand + receptor FERONIA Zastavení růstu pylové láčky, uvolnění spermatických buněk
31 Struktura a funkce FERONIA 31 SS signální peptid; ECD extracelulární doména; TM transmembranová doména MALA, MALB tandemové malectin-like domény; exjm extracelulární juxtamembranová oblast FERONIA kontroluje růst a samičí fertilitu, zprostředkuje patogenem indukované reakce, nutný pro normální vývoj buněčné stěny. RALF = Rapid ALkalinization Factor = malý peptidový růstový regulátor, inhibuje růst hypokotylu a kořene; první známý ligand receptoru FERONIA THESEUS1 příbuzný k FERONIA funguje jako odhadce stavu buněčné stěny ANXUR1 a 2 homology FERONIA zajišťují integritu pylové láčky a samčí sterilitu Interakce pylové láčky a synergických buněk vede k programové smrti (PCD) pylové láčky a jedné ze synergid. Synergidy kontrolují tento proces - koordinují hladiny Ca 2+ synergid v reakci na dynamiku Ca 2+ v pylové láčce a růstové chování láčky. FERONIA je vyžadován k iniciaci a modulaci reakcí k Ca 2+ a jejich spojení s PCD.
. Tam se ale otočí a směřuje zpátky k mikropyle.")
32 MBR 32 Mutant lorelei - defekt v uvolnění spermatických buněk Pylová láčka mutanta po dosažení embryonálního vaku nepraská, ale pokračuje v růstu uvnitř embryonálního vaku směrem k centrální buňce (CC). Tam se ale otočí a směřuje zpátky k mikropyle. cc central cell my micropylární konec ch chalaziální konec LORELEI (LRE) - exprimován v synergických buňkách LORELEI je glucosylphosphatidylinositol (GPI)-anchored protein umožňuje samičímu gametofytu rozpoznat vniknutí kompatibilní pylové láčky a umožňuje uvolnění spermatických buněk.

33 Update 2016 Liu X et al. (2016) Plant Cell 28: Protein LORELEI (LRE) je exprimován v tzv. filiform aparátu membránová struktura, která je prvním místem styku pylové láčky se samičím gametofytem. Ve filiform aparátu je rovněž exprimován receptor FERONIA; proteiny LORELEI a FERONIA spolupracují; LORELEI indukuje expresi FERONIA.

34 34 Beale KM, Johnson MA (2013) Current Opinion in Plant Biology 16: 1-8 Přiblížení pylové láčky k vaječné buňce Ca 2+, exprese proteinů FER, LRE a NTE Prasknutí pylové láčky, uvolnění spermatických buněk

35 MBR 35 anx1anx2 pylová láčka praská ještě předtím, než dosáhne samičího gametofytu Geny ANXUR1 a ANXUR2 homology genu FERONIA Funkce ANX1 a ANX2 : konstitutivní inhibice prasknutí pylové láčky; specificky exprimované v pylové láčce Pylová láčka dosahuje samičího gametofytu Aktivace dráhy FERONIA Deaktivace ANX1/ANX2 Prasknutí pylové láčky, uvolnění spermatických buněk. Kessler SA, Grossniklaus (2011) Current Opinion in Plant Biology 14:
36 3) Co způsobuje fúzi gamet a zablokování pronikání další pylové láčky? 36 Beale KM, Johnson MA (2013) Current Opinion in Plant Biology 16: 1-8 Mechanizmus fúze gamet není znám. Signál z EC1 vede k relokalizaci HAP2 k membráně zapojení neznámých proteinů ve fúzi gamet. Dva navržené mechanizmy zablokování dalšího oplodnění: 1) Přerušení tvorby atraktantů sekretovaných synergickou buňkou 2) Uvolnění odpuzovačů sekretovaných vaječnou buňkou
RVR ) Vývoj květu a kontrola kvetení. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění
 Vývoj květu a kontrola kvetení. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění") 2015 6) Vývoj květu a kontrola kvetení 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění 2 d) Vznik gamet Životní cyklus rostliny Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
2015 6) Vývoj květu a kontrola kvetení 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění 2 d) Vznik gamet Životní cyklus rostliny Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
MBR ) Reprodukce rostlin. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění
 Reprodukce rostlin. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění") MBR1 2015 4) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění d) Vznik gamet Životní cyklus rostliny 2 Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
MBR1 2015 4) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění d) Vznik gamet Životní cyklus rostliny 2 Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2) Reprodukce rostlin 2015 2 d) Vznik gamet d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění Životní cyklus rostliny Mikrosporogeneze Megasporogeneze 3 Vývoj samčího gametofytu - mikrosporogeneze
1 2) Reprodukce rostlin 2015 2 d) Vznik gamet d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění Životní cyklus rostliny Mikrosporogeneze Megasporogeneze 3 Vývoj samčího gametofytu - mikrosporogeneze
MBR ) Reprodukce rostlin. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění
 Reprodukce rostlin. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění") MBR1 2016 4) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění d) Vznik gamet Životní cyklus rostliny 2 Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
MBR1 2016 4) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění d) Vznik gamet Životní cyklus rostliny 2 Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
2) Reprodukce rostlin
 Reprodukce rostlin") 2010 2) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction
2010 2) Reprodukce rostlin 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Pohlavní rozmnožování. Gametogeneze u rostlin a živočichů.
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Pohlavní rozmnožování Gametogeneze u rostlin a živočichů. 2/65 Pohlavní rozmnožování obecně zajišťuje variabilitu druhu
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Pohlavní rozmnožování Gametogeneze u rostlin a živočichů. 2/65 Pohlavní rozmnožování obecně zajišťuje variabilitu druhu
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Růst a vývoj rostlin - praktikum
 Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Organely v pylu a pylových láčkách 2. Jak atraktivní jsou vajíčka? 3. Aktivita promotorů v gametofytu 4. Vývojové mutace pylu Teoretický úvod Samčí gametofyt
Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Organely v pylu a pylových láčkách 2. Jak atraktivní jsou vajíčka? 3. Aktivita promotorů v gametofytu 4. Vývojové mutace pylu Teoretický úvod Samčí gametofyt
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Růst a vývoj rostlin - praktikum
 Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Organely v pylu a pylových láčkách 2. Jak atraktivní jsou vajíčka? 3. Aktivita promotorů v gametofytu 4. Vývojové mutace pylu Teoretický úvod Samčí gametofyt
Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Organely v pylu a pylových láčkách 2. Jak atraktivní jsou vajíčka? 3. Aktivita promotorů v gametofytu 4. Vývojové mutace pylu Teoretický úvod Samčí gametofyt
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Přijímací zkoušky BGI Mgr. 2016/2017. Počet otázek: 30 Hodnocení každé otázky: 1 bod Čas řešení: 60 minut. Varianta B
 Přijímací zkoušky BGI Mgr. 2016/2017 Počet otázek: 30 Hodnocení každé otázky: 1 bod Čas řešení: 60 minut Varianta B A1. Čepička na 5' konci eukaryotické mrna je tvořena a. 7-methylguanosin trifosfátem
Přijímací zkoušky BGI Mgr. 2016/2017 Počet otázek: 30 Hodnocení každé otázky: 1 bod Čas řešení: 60 minut Varianta B A1. Čepička na 5' konci eukaryotické mrna je tvořena a. 7-methylguanosin trifosfátem
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Reprodukční orgány II
 Reprodukční orgány II Plant cell (1993) 5:1139-1146 Reprodukční orgány II Střídání generací u semenných rostlin Vývoj samčího gametofytu Mikrosporogeneze Mikrogametogeneze Interakce sporofytu s gametofytem
Reprodukční orgány II Plant cell (1993) 5:1139-1146 Reprodukční orgány II Střídání generací u semenných rostlin Vývoj samčího gametofytu Mikrosporogeneze Mikrogametogeneze Interakce sporofytu s gametofytem
Samčí gametofyt. mikrosporogeneze mikrogametogeneze
 Samčí gametofyt mikrosporogeneze mikrogametogeneze Clarkia xantania (Onagraceae) Am. J. Bot. blizna s čnělkou http://www.botany.org/ plantimages větší prašníky menší prašníky Photo: C. J. Runions Cornell
Samčí gametofyt mikrosporogeneze mikrogametogeneze Clarkia xantania (Onagraceae) Am. J. Bot. blizna s čnělkou http://www.botany.org/ plantimages větší prašníky menší prašníky Photo: C. J. Runions Cornell
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
Anatomie, histologie a embryologie
 Anatomie, histologie a embryologie Témata: - Opylení a oplození - Dvojí oplození u krytosemenných rostlin - Zygota, vývin embrya a semene Vývoj samčích a samičích gamét v květných orgánech Po dokončení
Anatomie, histologie a embryologie Témata: - Opylení a oplození - Dvojí oplození u krytosemenných rostlin - Zygota, vývin embrya a semene Vývoj samčích a samičích gamét v květných orgánech Po dokončení
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 03 Reprodukční vývoj apomixie doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Pohlavní amfimixis megasporogeneze megagametogeneze mikrosporogeneze mikrogametogeneze
Bi8240 GENETIKA ROSTLIN Prezentace 03 Reprodukční vývoj apomixie doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Pohlavní amfimixis megasporogeneze megagametogeneze mikrosporogeneze mikrogametogeneze
Otázka 22 Rozmnožování rostlin
 Otázka 22 Rozmnožování rostlin a) Nepohlavně (vegetativně): 1. Způsoby rozmnožování u rostlin: typ množení, kdy nový jedinec vzniká z jediné buňky, tkáně, nebo části orgánu o některé rostliny vytvářejí
Otázka 22 Rozmnožování rostlin a) Nepohlavně (vegetativně): 1. Způsoby rozmnožování u rostlin: typ množení, kdy nový jedinec vzniká z jediné buňky, tkáně, nebo části orgánu o některé rostliny vytvářejí
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
REGULACE TRANSLACE DEGRADACE BÍLKOVIN. 4. Degradace bílkovin. 4. Degradace bílkovin. 4. Degradace bílkovin
 4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
5) Klíčení semen a mobilizace rezerv
 Klíčení semen a mobilizace rezerv") MBR2 2016 5) Klíčení semen a mobilizace rezerv a) Klíčení semen b) Mobilizace rezerv 1 Podmínky prostředí, které mohou zrušit dormanci 2 Dormantní semena mohou za působení určitých vnějších podmínek klíčit:
MBR2 2016 5) Klíčení semen a mobilizace rezerv a) Klíčení semen b) Mobilizace rezerv 1 Podmínky prostředí, které mohou zrušit dormanci 2 Dormantní semena mohou za působení určitých vnějších podmínek klíčit:
3) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
- význam: ochranná funkce, dodává buňce tvar. jádro = karyon, je vyplněné karyoplazmou ( polotekutá tekutina )
") Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů
 Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů") MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 04 Inkompatibilita doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Inkompatibilní systémy vyšších rostlin Neschopnost rostlin tvořit semena Funkční gamety zachovány
Bi8240 GENETIKA ROSTLIN Prezentace 04 Inkompatibilita doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Inkompatibilní systémy vyšších rostlin Neschopnost rostlin tvořit semena Funkční gamety zachovány
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Prokaryota x Eukaryota. Vibrio cholerae
 Živočišná buňka Prokaryota x Eukaryota Vibrio cholerae Dělení živočišných buněk: buňky jednobuněčných organismů (volně žijící samostatné jednotky) buňky mnohobuněčných větší morfologické i funkční celky
Živočišná buňka Prokaryota x Eukaryota Vibrio cholerae Dělení živočišných buněk: buňky jednobuněčných organismů (volně žijící samostatné jednotky) buňky mnohobuněčných větší morfologické i funkční celky
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 2 1 2017 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gunawardena,
2 1 2017 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gunawardena,
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
3) Růst a vývoj. a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů KFZR 1
 Růst a vývoj. a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů KFZR 1") 1 2010 3) Růst a vývoj a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů Raghavan V (2006) Double Fertilization. Embryo and Endosperm Development In Flowering Plants. Springer.
1 2010 3) Růst a vývoj a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů Raghavan V (2006) Double Fertilization. Embryo and Endosperm Development In Flowering Plants. Springer.
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin Nejnovější review: Latrasse D et al. (2016) Journal of Exp. Botany 67: 5887-5900 Mira M et
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin Nejnovější review: Latrasse D et al. (2016) Journal of Exp. Botany 67: 5887-5900 Mira M et
The cell biology of rabies virus: using stealth to reach the brain
 The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
Vladimír Vinter
 Přehled vývojových cyklů cévnatých rostlin U nejstarších psilofytních rostlin se gametofyt pravděpodobně morfologicky neodlišoval od sporofytu. Rozdíl byl pouze v počtu chromozomů a také v tom, že na gametofytu
Přehled vývojových cyklů cévnatých rostlin U nejstarších psilofytních rostlin se gametofyt pravděpodobně morfologicky neodlišoval od sporofytu. Rozdíl byl pouze v počtu chromozomů a také v tom, že na gametofytu
ANATOMIE REPRODUKČNÍCH ÚTVARŮ
 ANATOMIE REPRODUKČNÍCH ÚTVARŮ životní cyklus - Bryofyta - Lycopodiofyta - Polypodiofyta - Equisetofyta - Spermatofyta vývoj samčího i samičího gametofytu z hlediska fylogeneze i ontogeneze Bryophyta životní
ANATOMIE REPRODUKČNÍCH ÚTVARŮ životní cyklus - Bryofyta - Lycopodiofyta - Polypodiofyta - Equisetofyta - Spermatofyta vývoj samčího i samičího gametofytu z hlediska fylogeneze i ontogeneze Bryophyta životní
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Inovace studia molekulární. a buněčné biologie
 Inovace studia molekulární I n v e s t i c e d o r o z v o j e v z d ě l á v á n í a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární I n v e s t i c e d o r o z v o j e v z d ě l á v á n í a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Endocytóza o regulovaný transport látek v buňce
 . Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
. Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
13. ONTOGENEZE III.: REPRODUKCE
 13. ONTOGENEZE III.: REPRODUKCE 13.1. VÝVOJ KVĚTU Při vývoji květních orgánů nastávají ve vrcholech podstatné změny organogeneze a růstu orgánů. Listový nebo pupenový původ květních orgánů je sice patrný
13. ONTOGENEZE III.: REPRODUKCE 13.1. VÝVOJ KVĚTU Při vývoji květních orgánů nastávají ve vrcholech podstatné změny organogeneze a růstu orgánů. Listový nebo pupenový původ květních orgánů je sice patrný
Buňky, tkáně, orgány, soustavy
 Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Samičí gametofyt. Gyneceum Typy placentace, stavba vajíčka Megasporogeneze (vývoj megaspor) Megagametogeneze (vývoj zárodečného vaku)
 Megagametogeneze (vývoj zárodečného vaku)") Samičí gametofyt Gyneceum Typy placentace, stavba vajíčka Megasporogeneze (vývoj megaspor) Megagametogeneze (vývoj zárodečného vaku) Gyneceum Pestík (pistillum) je samičí pohlavní orgán vznikající srůstem
Samičí gametofyt Gyneceum Typy placentace, stavba vajíčka Megasporogeneze (vývoj megaspor) Megagametogeneze (vývoj zárodečného vaku) Gyneceum Pestík (pistillum) je samičí pohlavní orgán vznikající srůstem
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Vodní režim rostlin. Obsah vody, RWC, vodní potenciál a jeho komponenty: Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy,
 Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
MEMBRÁNOVÉ STRUKTURY EUKARYONTNÍCH BUNĚK
 MEMBRÁNOVÉ STRUKTURY EUKARYONTNÍCH BUNĚK PLASMATICKÁ MEMBRÁNA EUKARYOTICKÝCH BUNĚK Všechny buňky (prokaryotické a eukaryotické) jsou ohraničeny membránami zajišťujícími integritu a funkci buněk Ochrana
MEMBRÁNOVÉ STRUKTURY EUKARYONTNÍCH BUNĚK PLASMATICKÁ MEMBRÁNA EUKARYOTICKÝCH BUNĚK Všechny buňky (prokaryotické a eukaryotické) jsou ohraničeny membránami zajišťujícími integritu a funkci buněk Ochrana
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
Univerzita Karlova v Praze - 1. lékařská fakulta. Buňka. Ústav pro histologii a embryologii
 Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Vakuola. Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich
 Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Rostlinná cytologie. Přednášející: RNDr. Jindřiška Fišerová, Ph.D. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") 2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Růst a vývoj rostlin - praktikum
 Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Lokalizace aktivity promotorů gametofytických genů 2. Gametofytické mutace 3. Umlčování mrna v pylových láčkách Teoretický úvod Samčí gametofyt krytosemenných
Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Lokalizace aktivity promotorů gametofytických genů 2. Gametofytické mutace 3. Umlčování mrna v pylových láčkách Teoretický úvod Samčí gametofyt krytosemenných
Růst a vývoj rostlin - praktikum
 Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Lokalizace aktivity promotorů gametofytických genů 2. Gametofytické mutace 3. Umlčování mrna v pylových láčkách Teoretický úvod Samčí gametofyt krytosemenných
Růst a vývoj rostlin - praktikum Blok II Úlohy 1. Lokalizace aktivity promotorů gametofytických genů 2. Gametofytické mutace 3. Umlčování mrna v pylových láčkách Teoretický úvod Samčí gametofyt krytosemenných
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
PROČ ROSTLINA KVETE Při opylení
 - Při opylení je pylové zrno přeneseno u nahosemenných rostlin na nahé vajíčko nebo u krytosemenných rostlin na bliznu pestíku. - Květy semenných rostlin jsou přizpůsobeny různému způsobu opylení. - U
- Při opylení je pylové zrno přeneseno u nahosemenných rostlin na nahé vajíčko nebo u krytosemenných rostlin na bliznu pestíku. - Květy semenných rostlin jsou přizpůsobeny různému způsobu opylení. - U