OXIDATIVNÍ FOSFORYLACE
|
|
|
- Zdeňka Krausová
- před 8 lety
- Počet zobrazení:
Transkript
1 OXIDATIVNÍ FOSFORYLACE

2 OBSAH Mitochondrie Elektronový transport Oxidativní fosforylace Kontrolní systém oxidativního metabolismu.

3 Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů Dýchací řetězec - oxidativní fosforylace Oxidativní fosforylace je proces tvorby ATP při přenosu elektronů z NADH a FADH 2 na kyslík přes řadu přenašečů elektronů. NADH a FADH 2 jsou vytvořené při glykolýze, oxidaci mastných kyselin a cyklu trikarboxylových kyselin Oxidativní fosforylace je hlavním zdrojem energie aerobních organismů.

4 Významné reakce katalyzované NAD(P)H dehydrogenasami
H")
5 Mitochondrie Mitochondrie jsou místem eukaryotního oxidačního metabolismu. Vnější membrána Vnitřní membrána Kristy Matrix Mezimembránový prostor Drsné endoplazmatické retikulum Mitochondrie obsahují: pyruvátdehydrogenasu enzymy citrátového cyklu enzymy katabolismu mastných kyselin enzymy spolu s proteiny elektronového transportního řetězce a oxidativní fosforylace.

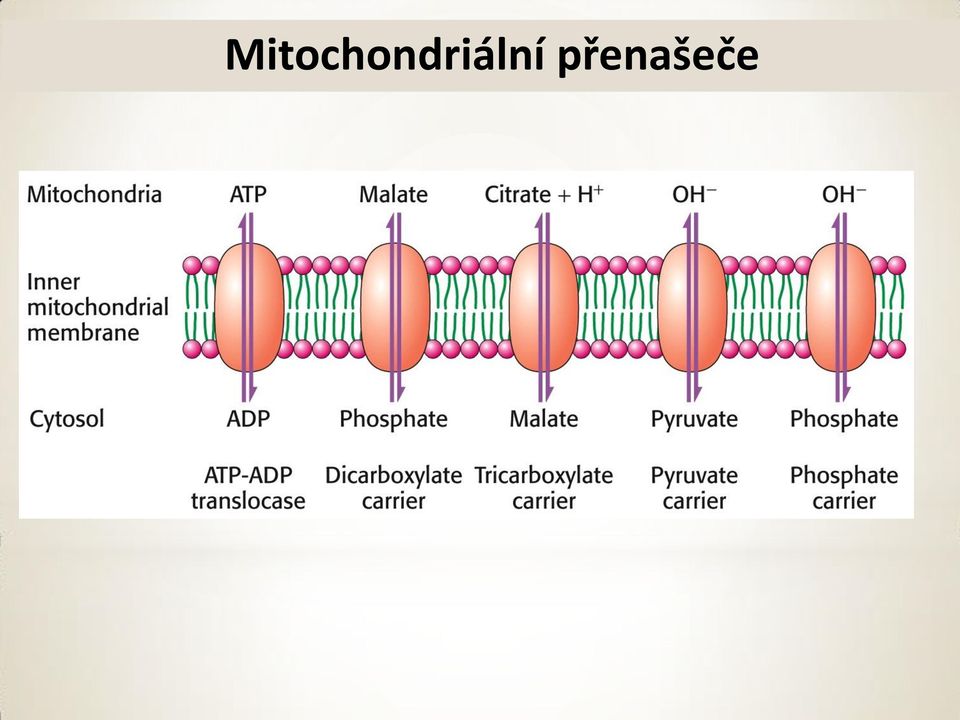
6 Mitochondriální transportní systém Vnější mitochondriální membrána, stejně jako bakteriální, obsahuje poriny, proteiny, které dovolují volnou difúzi molekul do 10 kd. Vnitřní membrána, která je hmotnostně složena ze 75 % z proteinů, volně propouští O 2, CO 2 a H 2 O. Jinak obsahuje řadu transportních proteinů, které kontrolují průchod metabolitů jako jsou ATP, ADP, pyruvát, Ca 2+ a fosfát. Redukované ekvivalenty (NADH) se transportují z cytosolu do mitochondrie, buď malát-aspartátovým člunkem nebo glycerol-3- fosfátovým člunkem. NADH transportované glycerol-3-fosfátovým člunkem poskytuje po oxidaci jen 2 ATP.
 se transportují z cytosolu do mitochondrie, buď malát-aspartátovým člunkem nebo glycerol-3- fosfátovým člunkem.")
7 Mitochondriální přenašeče
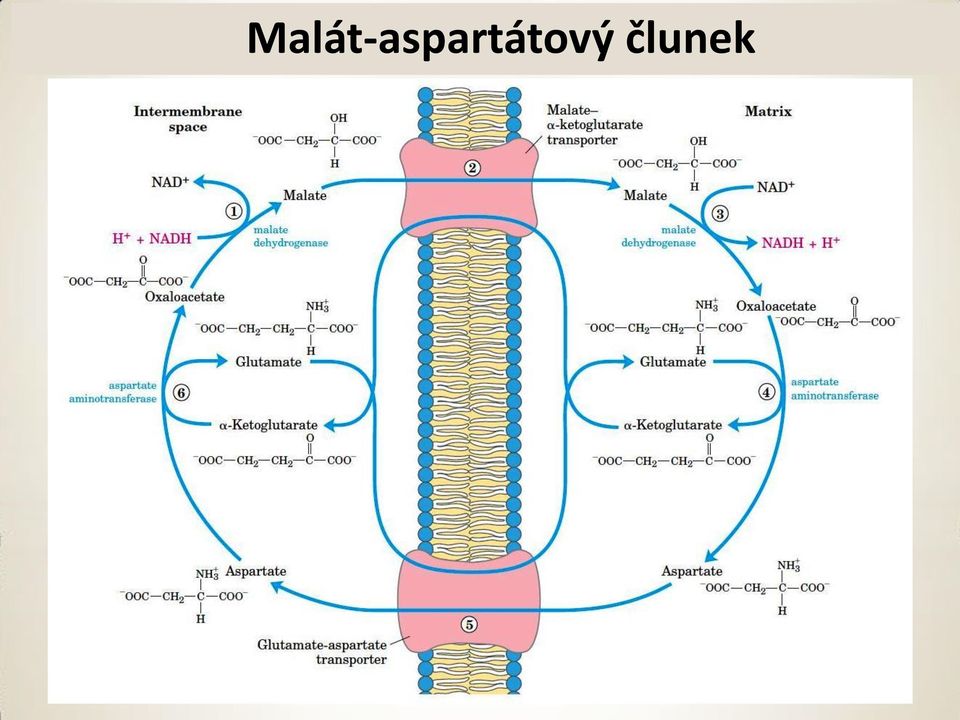
8 Malát aspartátový člunek Malát aspartátový člunek (srdce a játra). Reversibilní závisí na poměru NADH/NAD + v cytoplasmě a matrix

9 Malát-aspartátový člunek
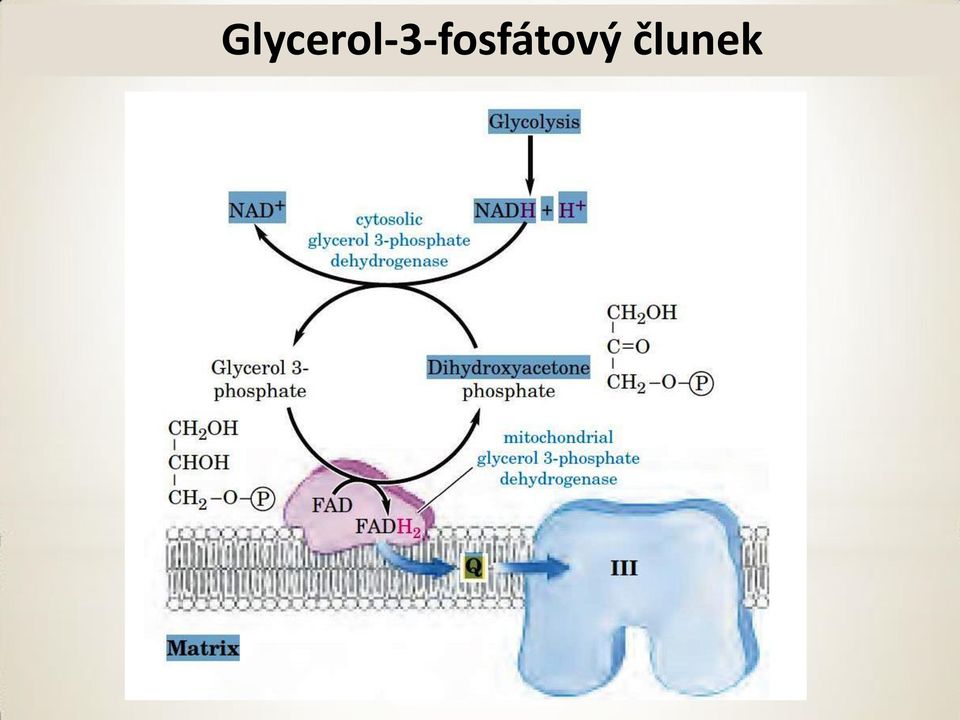
10 Glycerol-3-fosfátový člunek Glycerol-3-fosfátový člunek ve svalech(shuttle). Redukce dihydroxyacetonfosfátu na glycerol-3-fosfát a poté mitochondriální glycerol-3-fosfátdehydrogenasa. Tvoří se pouze 2 ATP!!!

11 Glycerol-3-fosfátový člunek
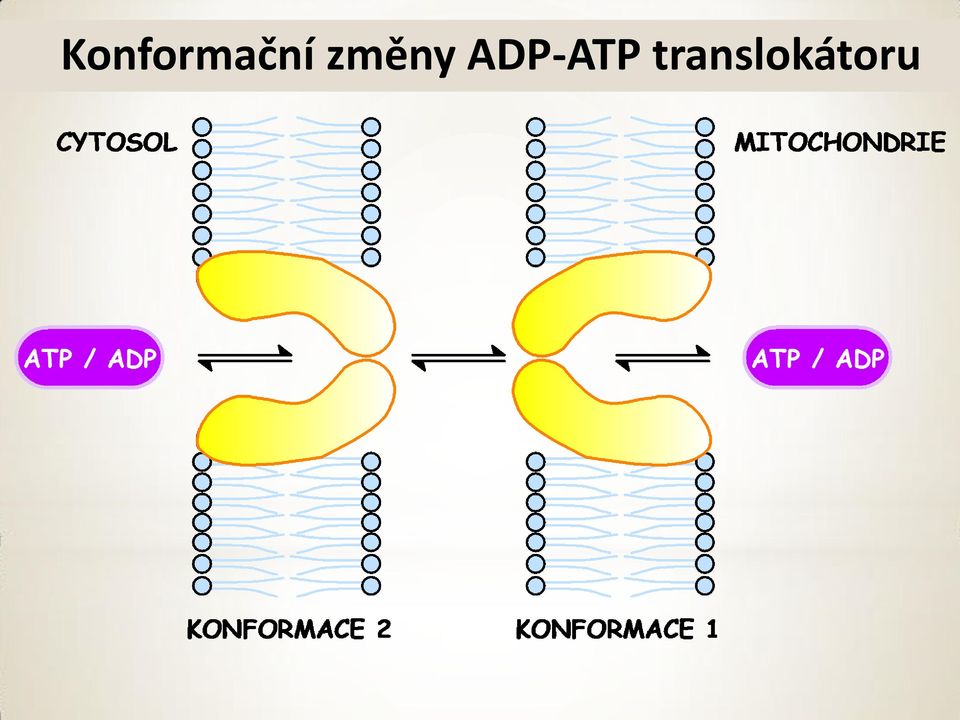
12 Translokátor ADP ATP Většina v mitochondrii vytvořeného ATP se využívá v cytoplasmě. Vnitřní mitochondriální membrána obsahuje ADP-ATP translokator transportující ATP do cytosolu a ADP z cytosolu do mitochondrie. Translokator je dimer identických 30 kd podjednotek s jedním vazebným místem pro ADP i ATP, které vzájemně kompetují. Translokátor mění konformaci při vazbě buď ADP nebo ATP. Export ATP (4 záporné náboje) proti importu ADP (3 záporné náboje) je elektrogenní antiport poháněný rozdílem membránového potenciálu.

13 Mechanismus mitochondriální ATP-ADPtranslokasy Mechanismus mitochondriální ATP-ADPtranslokasy (14 % proteinů mitochondriální membrány) ANTIPORTER. ATP je transportován 30 x rychleji ven, než ADP dovnitř. Čtvrtina energie získaná ox. fosforylací jde na konto exportu ATP. Inhibice atraktylosid (rostlinný glykosid) a bongkreková kyselina (antibiotikum z plísně).


14 Konformační změny ADP-ATP translokátoru
15 Transport fosfátů K syntéze ATP z ADP a P i je nutné transportovat fosfát z cytosolu do mitochondrie. Fosfátový nosič lze charakterizovat jako elektrochemický P i -H + symport poháněný rozdílem (D ph) (transmembránový protonový gradient). Vytvořený transmembránový protonový gradient nevede pouze k syntéze ATP, ale také k transportu ADP a P i.
 (transmembránový protonový gradient).")
16 1. Vstup do matrix je poháněn membránovým potenciálem (uvniř negativní) 2. Antiport s H + Transport Ca 2+ Mitochondrie působí jako pufr pro cytosolový Ca 2+

17 Elektronový transport / Termodynamika Elektronové nosiče přenášející elektrony z NADH a FADH 2 na kyslík jsou lokalizovány ve vnitřní mitochondriální membráně. Oxidace NADH je silně exergonická. Měřítkem afinity substrátu k elektronům je standardní redukční potenciál ( o ). Čím vyšší hodnota, tím větší afinita k elektronům. Poloreakce oxidace NADH kyslíkem jsou: NAD + + H e - NADH ½ O H e - H 2 O o = - 0, 315 V o = 0, 815 V NADH je donor elektronů, O 2 akceptor. Celková reakce: ½ O 2 + NADH + H + H 2 O + NAD + D o = 0, 815 V (-0, 315 V) = 1, 130 V!!!

18 Elektronový transport / Termodynamika Redukční potenciál páru H + : H 2 je definován jako 0 voltů. Např. NADH jako silné redukční činidlo předává elektrony a má tak negativní redukční potenciál. Silné oxidační činidlo jako je O 2 přijímá elektrony a má tak pozitivní redukční potenciál. V biochemických textech je redukční potenciál uváděn při koncentraci H + = 10-7 M (ph 7), zatímco v chemických 1 M (ph 0) Redukční potenciál v biochemii je tak označen jako E o (obdobně jako D G o ) Vztah mezi změnou standardní volné (Gibbsovy) energie a změnou redukčního potenciálu: D G o = - nfde o n = počet přenášených elektronů, F je Faradayův náboj (96, 48 kj.mol -1.V -1 ) a DE o je ve voltech.
, zatímco v chemických 1 M (ph 0) Redukční potenciál v biochemii je tak označen jako")
19 Elektronový transport / Termodynamika Oxidace NADH poskytuje D G o = kj.mol -1 volné energie. K syntéze jednoho molu ATP z ADP a P i je třeba 30, 5 kj.mol -1. Energetický rozdíl D G o = kj.mol -1 nelze přímo převést na několik ATP. Ve vnitřní mitochondriální membráně je soustava tří proteinových komplexů přes které putují elektrony z NADH ke kyslíku. Celková volná energie je tak rozdělena na tři menší části z nichž každá se podílí na tvorbě ATP oxidativní fosforylací. Oxidace NADH poskytne přibližně 3 ATP. Termodynamická efektivita za standardních biochemických podmínek je 42 %. (3 x 30,5 kj.mol -1 x 100 / 218 kj.mol -1 ).

20 Komplexy sekvence elektronového transportu
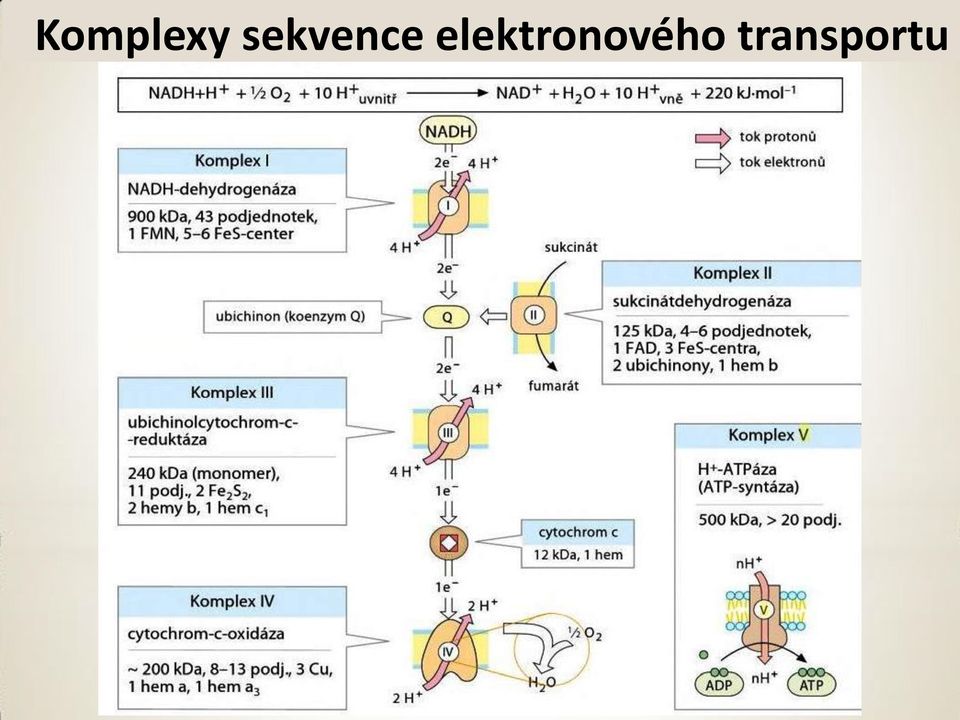
21 Komplexy sekvence elektronového transportu Komplexy jsou řazeny podle vzrůstajícího redukčního potenciálu. Komplex I: Katalyzuje oxidaci NADH koenzymem Q (CoQ). NADH + CoQ (oxidovaný) NAD + + CoQ (redukovaný) D o = 0, 360 V; D G o = - 69, 5 kj.mol -1 Komplex II: Katalyzuje oxidaci FADH 2 koenzymem Q. FADH 2 + CoQ (oxidovaný) FAD + CoQ (redukovaný) D o = 0, 085 V; D G o = - 16, 4 kj.mol-1 Redoxní reakce neposkytuje dostatečné množství energie pro tvorbu ATP. Funkcí je, pouze předávat elektrony z FADH 2 do elektronového transportního řetězce.
22 Komplexy sekvence elektronového transportu Komplex III: Katalyzuje oxidaci CoQ (red.) cytochromem c. CoQ (redukovaný) + 2 cytochrom c (oxidovaný) CoQ (oxidovaný) + 2 cytochrom c (redukovaný) D o = 0, 190 V; D G o = - 36, 7 kj.mol -1 Komplex IV: Katalyzuje oxidaci redukovaného cytochromu c kyslíkem terminálním akceptorem elektronů. 2 cytochrom c (red.) + ½ O 2 2 cytochrom c (oxid.) + H 2 O D o =0, 580 V; D G o = kj.mol -1.
23 Komplexy sekvence elektronového transportu
24

25 Mitochondriální elektronový transport
26 Mitochondriální elektronový transport
27 Komplex I: NADH koenzym Q oxidoreduktasa Komplex I je největší protein v mitochondriální membráně 43 podjednotek o celkové hmotnosti 900 kd. Komplex I obsahuje jednu molekulu FMN a 6 až 7 komplexů (klastrů) železo-síra. Klastry jsou prostetickými skupinami proteinů železo-síra nebo jinak nehemové proteiny. FMN je pevně vázaný na proteiny; zatímco CoQ je, díky svému hydrofobnímu isoprenoidnímu řetězci, volně pohyblivý v lipidové dvojvrstvě membrány. U savců obsahuje řetězec CoQ 10 C 5 isoprenoidních jednotek (Q 10 ). U některých organismů je řetězec CoQ kratší 6 nebo 8 jednotek.
28 Struktura NADH-Q oxidoreduktasy (Komplex I) Matrix Skládá se z části lokalizované v membráně a ramene v matrix. NADH se oxiduje v rameni a elektrony jsou přeneseny do membrány, kde redukují CoQ.
29 Klastry železo-síra. Přechod elektronů mezi Fe 2+ a Fe 3+
30 Oxidační stavy FMN a koenzymu Q (CoQ) 2- H 2 C OPO 3 H 3 C H 3 C H 3 C C C C CH 3 CH 3 CH 3 H CH 2 O H 3 C N N O H 3 CO CH 3 C H 3 H N O N H CH 3 H 3 CO CH 2 CH C CH 2 H n O Isoprenoidní jednotky Flavinmnonukleotid (FMN) (oxidovaná nebo chinonová forma) H Koenzym Q (CoQ) nebo ubichinon (oxidovaná nebo chinonová forma) H H R O H 3 C N N O H 3 CO CH 3 C H 3 N N H H 3 CO R H H O FAMNH (radikálová nebo semichinonová forma) H OH Koenzym QH nebo ubisemichinon (radikálová nebo semichinonová forma) H H R H OH H 3 C N N O H 3 CO CH 3 C H 3 N N H H 3 CO R H H O OH FMNH 2 (redukovaná nebo hydrochinonová forma) Koenzym QH 2 nebo ubichinol (redukovaná nebo hydrochinonová forma)
31 Reakce katalyzovaná NADH-Q oxidoreduktasy NADH + Q + 5 H + matrix NAD + + QH H + cytoplasma První krok: vazba NADH a přenos dvou elektronů na FMN (FMNH 2 ). Druhý krok: elektrony jsou přeneseny na soustavu sloučenin železo síra. Třetí krok: elektrony jsou přeneseny na vázaný CoQ. Tok dvou elektronů z NADH na vázaný CoQ přes komplex I pumpuje čtyři elektrony z matrix. Redukce CoQ na CoQH 2 vyžaduje další dva protony z matrix. Čtvrtý krok: pár elektronů z vázaného CoQH 2 se přenáší na 4Fe 4S a protony se uvolňují do cytoplasmy. Pátý krok: elektrony z 4Fe 4S se přenáší na mobilní CoQ v hydrofobní části membrány. To vede k odebrání dalších dvou protonů z matrix.
32 Přenos elektronů a translokace protonů v komplexu I. Tok elektronů z NADH na CoQ probíhá stupňovitě mezi redox centry. NADH se účastní vždy jen dvouelektronové výměny. FMN a CoQ mohou přenášet jak dva, tak jeden elektron a proto tvoří elektronovou spojku mezi dvouelektronovým NADH a jednoelektronovými akceptory cytochromy. Při toku elektronů mezi redox centry komplexu I jsou transportovány čtyři protony z matrix do mezimembránového prostoru.
33 Spojení přenosu elektron proton. Redukce chinonu (CoQ) na CoQH 2 vede k odčerpání dvou protonů z matrix
34 Komplex II: sukcinát-koenzym Q Komplex II obsahuje enzym citrátového cyklu sukcinátdehydrogenasu. Přenáší elektrony z FADH 2 na CoQ. Komlex II obsahuje sukcinátdehydrogenasu s kovalentně vázaným FAD, několik klastrů Fe-S a jeden cytochrom b 560. Komplex I a II nejsou v sérii. Komplex II přenáší elektrony ze sukcinátu na CoQ. CoQ je mobilní sběrač elektronů. oxidoreduktasa Cytochrom b 560 slouží pravděpodobně k odstraňování kyslíkatých radikálů vznikajících vedlejšími reakcemi.
35 Struktura sukcinát-koenzym Q oxidoreduktasy
36 Cytochromy-hemové transportní proteiny Během transportu elektronů alternují mezi Fe 2+ a Fe 3+. PROTEIN CH 3 Cys HO H 2 C CH CH 2 CH C CH 2 H 3 CH 3 CH 2 CH CH 3 H 3 C S CH CH 3 Cys 2 3 S H 3 C 1 N + N 4 CH CH 2 H 3 C N + N CH CH 2 H 3 C N + N CH CH 3 Fe 2+ Fe 2+ Fe 2+ O CH 8 N N + 5 CH 3 H 3 C N N + CH 3 H 3 C N N + CH CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH COO COO Hem a - - COO COO Hem b (Fe-protoporfyrin IX) - - COO COO Hem c
37 ABSORBANCE Absorpční spektra hemových skupin redukovaných Fe 2+ cytochromů Soretovy pásy Cytochrom a Cytochrom b Cytochrom c Cytochrom c Cytochrom c Vlnová délka (nm)
38 Komplex III: koenzym Q cytochrom c oxidoreduktasa Komplex III (také jako cytochrom bc 1 ) transportuje elektrony z redukovaného CoQH 2 na cytochrom c. Obsahuje: Dva cytochromy typu b, jeden c 1 a jeden klastr [2Fe- 2S], znám také jako Rieskeho centrum. Transport elektronů a pumpování protonů probíhá tzv. Q cyklem. CoQH 2 podstupuje dva cykly reoxidace za tvorby stabilních meziproduktů semichinonů CoQ. Existují dvě nezávislá centra pro koenzym Q: Q o vážící CoQH 2 (blíže vnější straně membrány out) a Q i (blíže vnitřní straně membrány inner) vážící semichinon CoQ..
39 Struktura koenzym Q cytochrom c oxidoreduktasy
40 Q-cytochrom c oxidoreduktasa Druhá protonová pumpa. Funkcí komplexu III je katalyzovat přenos elektronů z QH 2 na oxidovaný cytochrom c za současného pumpování protonů z matrix. QH Cyt c ox + 2 H + matrix Q + 2 Cyt c red + 4 H + cytoplasma
41 První cyklus: CoQH 2 + cytochrom c 1 (Fe 3+ ) CoQ. + cytochrom c 1 (Fe 2+ ) + 2 H + (mezimembránový prostor) Druhý cyklus - nejdříve další CoQH 2 opakuje první cyklus. CoQH 2 + CoQ. + cytochrom c 1 (Fe 3+ ) + 2 H + (z matrix) CoQ + CoQH 2 + cytochrom c 1 (Fe 2+ ) + 2 H + (do mezimembrány) Z každých dvou CoQH 2 vstupující do Q cyklu, je jeden regenerován. Celková reakce: Reakce Q cyklu CoQH cytochrom c 1 (Fe 3+ ) + 2H + (z matrix) CoQ + 2 cytochrom c 1 (Fe 2+ ) + 4 H + (do mezimembránového prostoru). Elektrony jsou následně přenášeny na cytochrom c, který je transportuje do komplexu IV.
42 Q cyklus první část M E Z I M E M B R Á N O V Ý Z KOMPLEXU I QH 2 QH 2 2H + Q - e - e - FeS-protein b Q - L e - P R e - b e - H O Q Q S c 1 Q Q T o i O R PRVNÍ CYKLUS M A T R I X
43 Q cyklus druhá část M E Z I M E M B R Á N O V Ý Z KOMPLEXU I QH 2 QH 2 2H + Q - QH 2 e - e - e - FeS-protein b L e - Q - P R O S T O R e - b H Q Q c 1 Q o DRUHÝ CYKLUS Q i 2H + M A T R I X
44 Q cyklus
45 Komplex IV: cytochrom c oxidasa Katalyzuje jednoelektronovou oxidaci čtyřech redukovaných cytochromů c a současnou čtyřelektronovou redukci jedné molekuly O 2. 4 Cytochrom c (Fe 2+ ) + 4 H + + O 2 4 cytochrom c (Fe 3+ ) + 2 H 2 O Obsahuje čtyři redoxní centra: cytochrom a, cytochrom a 3, atom mědi značený jako Cu B a dvojici atomů mědi označovaných jako Cu A centrum. Redukce O 2 cytochrom c oxidasou je postupný složitý proces.
46 Komplex IV: cytochrom c oxidasa Cytochrom c oxidasa má dva kanály translokace protonů. Čtyři tzv. chemické nebo skalární protony jsou odňaty z matrix během redukce kyslíku za tvorby dvou molekul vody. Proces je spojen s translokací čtyř tzv. pumpovaných nebo vektorových protonů z matrix do mezimembránového prostoru. Při reakci komplexu IV je celkově transportováno přes vnitřní mitochondriální membránu osm protonů. 8 H + (matrix) + O cytochrom c (Fe 2+ ) 4 cytochrom c (Fe 3+ ) + 2 H 2 O + 4 H + (do mezimembránového prostoru)
47 Struktura cytochrom c oxidasy
48 Transport protonů z matrix cytochrom c oxidasou Čtyři protony do cytoplasmy a čtyři na tvorbu vody
49 Reaktivní kyslíkaté radikály (ROS) Čtyřelektronová redukce kyslíku cytochrom c oxidasou je rychlá a precizní. Přesto vznikají kyslíkaté radikály, které reagují s různými buněčnými součástmi. Nejznámější je superoxidový radikál: O 2 + e - O 2 -. Superoxidový radikál je prekurzorem silnějších oxidačních radikálů jako jsou protonovaný (hydroxoniový) O 2 -. HO 2. a hydroxylový radikál. OH
50 Reaktivní kyslíkaté radikály (ROS) Antioxidační mechanismus: superoxiddismutasa (SOD) přítomná téměř ve všech buňkách. Katalyzuje přechod O 2 -. na peroxid vodíku. Vytvořený peroxid vodíku je degradován katalasou na vodu a kyslík: 2 H 2 O 2 2 H 2 O + O 2 nebo glutathionperoxidasou: 2 GSH + H 2 O 2 GSSG + 2 H 2 O Dalšími potenciálními antioxidanty jsou rostlinné sloučeniny jako askorbát (vitamin C) a -tokoferol (vitamin E). Pravděpodobně chrání rostliny před oxidačním stresem během fotosyntézy, kdy je H 2 O fotolýzou rozkládána na O 2, protony a elektrony.
51 Redoxní systémy dýchacího řetězce

52 Komplex V: ATP synthasa Oxidativní fosforylace. Energetické spojení. Volná energie transportu protonů se uplatní při tvorbě ATP.
53 Komplex V: ATP synthasa
54 Chemiosmotická hypotéza oxidativní fosforylace Mitchell, P., Coupling of phosphorylation to electron and hydrogen transfer by a chemiosmotic type of mechanism. Nature 191, (1961). Peter Mitchell ( ), Nobelova cena za Fyziologii a medicínu Volná energie elektronového transportu je realizována pumpováním H + z mitochondriální matrix do mezimembránového prostoru za tvorby elektrochemického H + gradientu přes membránu. Elektrochemický gradient je posléze uplatněn při syntéze ATP. Tato volná energie se nazývá protonmotivní síla.
55 Chemiosmotická hypotéza oxidativní fosforylace Naměřený membránový potenciál přes membránu jaterní mitochondrie je 0, 168 V (v matrix negativní). Naměřené ph v matrix je o 0,75 jednotky vyšší než v mezimembránovém prostoru. DG pro transport protonů z matrix je 21, 5 kj.mol -1 endergonní proces. Rušení gradientu, syntéza ATP, je proces exergonní!!
56 Procesy oxidace NADH a tvorby ATP NADH + ½ O 2 + H + H 2 O + NAD + D G o = - 220, 1 kj/mol ADP + P i + H + ATP + H 2 O D G o = + 30, 5 kj/mol
57 ATP synthasa protony pumpující ATP synthasa, F1Fo-ATPasa Multipodjednotkový transmembránový protein o celkové molekulové hmotnosti 450 kd. Složena ze dvou funkčních jednotek (komponent), F o a F 1. (F o vyslovuj ef ó - o znamená na oligomycin citlivá komponenta). protonvodivá (Fo) v membráně katalytická (F1) v matrix
58 ATP synthasa protony pumpující ATP synthasa, F1Fo-ATPasa Oligomycin B Antibiotikum produkované streptomycety Inhibuje ATP synthasu vazbou na F o Brání transportu H + kanálkem F o
59 ATP synthasa protony pumpující ATP synthasa, F1Fo-ATPasa. F o je ve vodě nerozpustný transmembránový protein obsahující více jak osm různých typů podjednotek. F 1 je vodě rozpustný periferní membránový protein složený z pěti typů podjednotek, které lze jednoduše oddělit od F o působením močoviny. Rozpuštěná F 1 komponenta (oddělená od F o ) hydrolyzuje, ale nesyntetizuje ATP. F 1 komponenta ATPsynthasy má podjednotkové složení 3 3 e. Další dva polipeptidy b2 a d tvoří stator, kterým jsou podjednotky a přichyceny k F o F o komponenta je složena z 12 peptidů c procházejících membránou a jedné podjednotky a.
60 Model F1Fo-ATPasy
61 Model F1Fo-ATPasy
62 Mechanismus syntézy ATP Rotace podjednotky o 120 o proti směru hodinových ručiček mění strukturu tří podjednotek. Podjednotka s nově syntetizovaným ATP je ve formě T (tight) nemůže uvolnit ATP. Pohybem podjednotky se převede na O formu, uvolní ATP a váže nové ADP a P i Tok protonů přes ATPsynthasu vede k uvolnění pevně vázaného ATP Role protonového gradientu nespočívá v syntéze ATP, ale v jeho uvolnění ze synthasy!!! ATP se tvoří i bez protonmotivní síly, ale neuvolňuje se (hydrolyzuje)

63 Mechanismus rotace F1Fo-ATPasy Protony z mezimembránového prostoru vstupují do podjednotky c, reagují s podjednotkou a vystupují ven až se c kruh otočí o jednu otočku, kdy se podjednotka c znovu setká s podjednotkou a.

64 Pohyb protonů přes membránu pohání rotaci c kruhu
65 Mechanismus vazebné změny tvorba ATP z ADP a Pi F 1 komponenta má tři reaktivní katalytické protomery ( jednotky), každý ve jiném konformačním stavu. L - váže substrát a produkt slabě (L = loosely) T - váže pevně (T = tightly) O - neváže vůbec, je otevřený (O = open) Uvolněná energie translokací protonů se realizuje přechodem mezi těmito stavy. Fosfoanhydridová vazba ATP je syntetizována jen ve stavu T a ATP se uvolňuje ve stavu O. Tři stupně: ADP a P i se váží do stavu L. Průchod protonů mění konformaci L na T. ATP je syntetizované ve stavu T, druhé ATP se dostává pohybem rotoru do stavu O a oddisociuje.
66 Mechanismus vazebné změny tvorby ATP 1. Vazba ADP a Pi k L-místu 2. Energeticky závislá konformační změna přemění vazebná místa L na T, T na O a O na L 3. Syntéza ATP v místě T a uvolnění ATP z místa L
67 Poměr P/O Poměr P/O reprezentuje relaci mezi množstvím syntetizovaného ATP (P) a množství redukovaného kyslíku (O). Tok dvou elektronů přes komplexy I, III a IV vede ke translokaci 10 protonů. Zpětný tok těchto protonů přes ATPasu poskytuje 3 ATP. Elektrony z FADH 2, vynechávají komplex I, vedou ke translokaci 6 protonů, což poskytuje jen 2 ATP. U aktivně respirujících mitochondrií nebývá poměr P/O celé číslo. Peter Hikle prověřoval P/O poměry a prokázal, že aktuální hodnoty jsou blíže číslů 2,5 a 1,5.
68 Kontrolní mechanismy oxidativní fosforylace Dospělý člověk spotřebuje denně kj metabolické energie. To odpovídá volné energii hydrolýzy 200 molů ATP na ADP a P i. Celkové množství v těle přítomného ATP je méně než 0,1 molu!!! ATP musí být nutně recyklován a jeho produkce regulována, protože se neprodukuje nikdy více ATP než je potřeba. V lidském těle se denně vytvoří 65 kg ATP Celkový obsah AMP + ADP + ATP je jen 3-4 g Každá molekula ADP se za den několikatisíckrát fosforyluje a pak defosforyluje
69 Regulace oxidativní fosforylace poměrem ATP/ADP regulace akceptorem Oxidativní fosforylace (od NADH k cytochromu c) je v rovnováze: ½ NADH + cytochrom c (Fe 3+ ) + ADP + P i ½ NAD + + cytochrom c (Fe 2+ ) + ATP D G 0 K eq = ([NAD + ] / [NADH]) ½.[c 2+ ] / [c 3+ ]. [ATP] /[ADP].[P i ] V cytcytochrom c oxidasové reakci je konečný krok řetězce elektronů ireversibilní. Cytochromoxidasa je řízena dostupností substrátů [NADH] / [NAD + ] a [ATP] / [ADP].[P i ]. Čím vyšší je poměr [NADH] / [NAD + ] a nižší [ATP] / [ADP].[P i ], tím vyšší je koncentrace redukovaného cytochromu c [c 2+ ] a vyšší cytochrom c oxidasová aktivita.
70 Regulace oxidativní fosforylace glykolýza, citrátový cyklus Vliv poměru NADH/NAD + Inhibice fosfofruktokinasy citrátem
71 Regulace oxidativní fosforylace poměrem ATP/ADP regulace akceptorem
72 Regulace oxidativní fosforylace poměrem ATP/ADP regulace akceptorem
73 Respirační kontrola Elektrony jsou trasportovány na O 2 tehdy, když je současně ADP fosforylováno na ATP. Kontrola hladinou ADP.
74 Inhibitory blokující elektronový transportní řetězec -0.4 NADH NAD + 2e - ( V) -0.2 KOMPLEX I D = V (DG = kj.mol -1 ) ADP + P i Rotenon, amytal ATP 0 ( V) Sukcinát 2e - FADH 2 KOMPLEX II CoQ ( V) D (V) +0.2 Fumarát KOMPLEX III D = V (DG = kj.mol -1 ) ADP + P i ATP Antimycin A Cytochrom c ( V) KOMPLEX IV D = V (DG = -112 kj.mol -1 ) 2e - ADP + P i ATP CN H + + 1/2 O 2 H 2 O ( V)
75 Inhibitory blokující elektronový transportní řetězec
76 Inhibitory blokující elektronový transportní řetězec Rotenon rostlinný toxin používaný indiány na Amazonce k lovu ryb, také insekticid) Amytal barbiturát. Antimycin antibiotikum. Proč lze inhibici rotenonem zrušit přídavkem sukcinátu??
77 Inhibitory blokující elektronový transportní řetězec OCH 3 CH 3 H 3 CO CH H O O C H 2 CH 3 HN CH 2 CH 2 CH 3 O H O O CH 2 O N H O Rotenon H CH 3 Amytal O O N C - NH O CH 3 Kyanid H 3 C OH O O CH 3 HN CHO (CH 2 ) 5 O CH 2 CH 3 Antimycin A H 3 C
78 Rozpojovače oxidativní fosforylace. Probíhá respirace netvoří se ATP
79 Rozpojovače oxidativní fosforylace. Probíhá respirace netvoří se ATP MATRIX Vysoká [H + ] CYTOSOL Nízká [H + ] O - OH OH O - H + + NO 2 NO 2 Difuze NO 2 NO 2 + H + NO 2 NO 2 NO 2 NO 2 2,4-Dinitrofenol (DNP) 2,4-Dinitrofenol (DNP)
80 Funkce odpojovacího proteinu UCP-1 Thermogenin UCP-1 protein) hnědá adiposní tkáň bohatá na mitochondrie (zazimující živočichové, novorozenci, zvířata adaptovaná na zimu) dimer (2x32 kda) u zvířat adaptovaných na chlad 15 % proteinů mitochondriální membrány hnědé adiposní tkáně
81 Funkce odpojovacího proteinu UCP-1
82 Regulace na úrovni proteinkinasy závislé na AMP Proteinkinasa závislá na AMP v játrech svalech a CNS Při vysoké spotřebě ATP je ADP přeměňován adenylátkinasou na ATP a AMP AMP aktivuje proteinkinasu závislou na AMP fosforylacíe řady enzymů - aktivace katabolických drah - inaktivace anabolických drah 2 adenylátkinasa 3 proteinkinasa závislá na AMP
83 Produkce ATP
Elektronový transport a oxidativní fosforylace
 Elektronový transport a oxidativní fosforylace Mitochondrie Elektronový transport xidativní fosforylace Kontrolní systém oxidativního metabolismu. Moto: dpočívající lidské tělo spotřebuje asi 420 kj energie
Elektronový transport a oxidativní fosforylace Mitochondrie Elektronový transport xidativní fosforylace Kontrolní systém oxidativního metabolismu. Moto: dpočívající lidské tělo spotřebuje asi 420 kj energie
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Aerobní odbourávání cukrů+elektronový transportní řetězec
 Aerobní odbourávání cukrů+elektronový transportní řetězec Dochází k němu v procesu jménem aerobní respirace. Skládá se z kroků: K1) Glykolýza K2) oxidativní dekarboxylace pyruvátu K3) Krebsův cyklus K4)
Aerobní odbourávání cukrů+elektronový transportní řetězec Dochází k němu v procesu jménem aerobní respirace. Skládá se z kroků: K1) Glykolýza K2) oxidativní dekarboxylace pyruvátu K3) Krebsův cyklus K4)
Citrátový cyklus a Dýchací řetězec. Milada Roštejnská Helena Klímová
 Citrátový cyklus a Dýchací řetězec Milada oštejnská elena Klímová 1 bsah 1 Citrátový cyklus Citrátový cyklus (reakce) Citrátový cyklus (schéma) espirace (dýchání) Vnější a vnitřní respirace Dýchací řetězec
Citrátový cyklus a Dýchací řetězec Milada oštejnská elena Klímová 1 bsah 1 Citrátový cyklus Citrátový cyklus (reakce) Citrátový cyklus (schéma) espirace (dýchání) Vnější a vnitřní respirace Dýchací řetězec
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Oxidativní fosforylace (9). Prof.RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Oxidativní fosforylace (9). Prof.RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Metabolické dráhy. František Škanta. Glykolýza. Repetitorium chemie X. 2011/2012. Glykolýza. Jaký je osud pyruátu bez přítomnosti kyslíku?
 Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy
 Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
Dýchací řetězec (DŘ)
") Dýchací řetězec (DŘ) Vladimíra Kvasnicová animace na internetu: http://vcell.ndsu.nodak.edu/animations/etc/index.htm http://vcell.ndsu.nodak.edu/animations/atpgradient/index.htm http://www.wiley.com/college/pratt/0471393878/student/animations/oxidative_phosphorylation/index.html
Dýchací řetězec (DŘ) Vladimíra Kvasnicová animace na internetu: http://vcell.ndsu.nodak.edu/animations/etc/index.htm http://vcell.ndsu.nodak.edu/animations/atpgradient/index.htm http://www.wiley.com/college/pratt/0471393878/student/animations/oxidative_phosphorylation/index.html
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Transport elektronů a oxidativní fosforylace
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
MitoSeminář II: Trochu výpočtů v bioenergetice. Souhrn. MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK
 MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
Enzymologie. Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů.
 ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
DYNAMICKÁ BIOCHEMIE. Daniel Nechvátal :: www.gymzn.cz/nechvatal
 DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
Sacharidy a polysacharidy (struktura a metabolismus)
") Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
UNIVERZITA PALACKÉHO V OLOMOUCI. Přírodovědecká fakulta Katedra biochemie
 UNIVERZITA PALACKÉHO V OLOMOUCI Přírodovědecká fakulta Katedra biochemie Dehydrosilybin jako protonofor: vztah struktury k biologické aktivitě DIPLOMOVÁ PRÁCE Autor: Hana Popelková Studijní program: N1406
UNIVERZITA PALACKÉHO V OLOMOUCI Přírodovědecká fakulta Katedra biochemie Dehydrosilybin jako protonofor: vztah struktury k biologické aktivitě DIPLOMOVÁ PRÁCE Autor: Hana Popelková Studijní program: N1406
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
9. Dýchací řetězec a oxidativní fosforylace. mitochondriální syntéza ATP a fotosyntéza
 9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV
 CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV a) Chemické složení a. biogenní prvky makrobiogenní nad 0,OO5% (C, O, N, H, S, P, Ca.) - mikrobiogenní pod 0,005%(Fe,Zn, Cu, Si ) b. voda 60 90% každého organismu - 90% příjem
CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV a) Chemické složení a. biogenní prvky makrobiogenní nad 0,OO5% (C, O, N, H, S, P, Ca.) - mikrobiogenní pod 0,005%(Fe,Zn, Cu, Si ) b. voda 60 90% každého organismu - 90% příjem
VAKUOLA. membránou ohraničený váček membrána se nazývá tonoplast. běžná u rostlin, zvířata specializované funkce či její nepřítomnost
 VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
>>> E A1 + E A2. . aktivační energie potřebná k reakci bez přítomnosti katalyzátoru E A E A1. energie potřebná ke vzniku enzym-substrátového komplexu
 Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
CYKLUS TRIKARBOXYLOVÝCH KYSELIN A GLYOXYLÁTOVÝ CYKLUS
 YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty
 sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
Bp1252 Biochemie. #8 Metabolismus živin
 Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů.
 Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů. Šárka Gregorová, 2013 Poznámka: protože se tyhle dvě státnicové otázky z velké
Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů. Šárka Gregorová, 2013 Poznámka: protože se tyhle dvě státnicové otázky z velké
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Energetický metabolismus rostlin
 Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
PRODUKCE VOLNÝCH RADIKÁLŮ V MITOCHONDRIÍCH
 UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ KATEDRA BIOLOGICKÝCH A BIOCHEMICKÝCH VĚD PRODUKCE VOLNÝCH RADIKÁLŮ V MITOCHONDRIÍCH BAKALÁŘSKÁ PRÁCE AUTOR PRÁCE: Kateřina Zvoníčková VEDOUCÍ PRÁCE:
UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ KATEDRA BIOLOGICKÝCH A BIOCHEMICKÝCH VĚD PRODUKCE VOLNÝCH RADIKÁLŮ V MITOCHONDRIÍCH BAKALÁŘSKÁ PRÁCE AUTOR PRÁCE: Kateřina Zvoníčková VEDOUCÍ PRÁCE:
Přednáška 6: Respirace u rostlin
 Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
Procvičování aminokyseliny, mastné kyseliny
 Procvičování aminokyseliny, mastné kyseliny Co je hlavním mechanismem pro odstranění aminoskupiny před odbouráváním většiny aminokyselin: a. oxidativní deaminace b. transaminace c. dehydratace d. působení
Procvičování aminokyseliny, mastné kyseliny Co je hlavním mechanismem pro odstranění aminoskupiny před odbouráváním většiny aminokyselin: a. oxidativní deaminace b. transaminace c. dehydratace d. působení
Ukázky z pracovních listů z biochemie pro SŠ A ÚVOD
 Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Energetický metabolismus rostlin
 Energetický metabolismus rostlin Sylabus - témata (Fischer, Šantrůček) 1. Základy energetiky v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta,
Energetický metabolismus rostlin Sylabus - témata (Fischer, Šantrůček) 1. Základy energetiky v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta,
Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY
 Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
12-Fotosyntéza FRVŠ 1647/2012
 C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
B4, 2007/2008, I. Literák
 B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
TERMODYNAMICKÁ ROVNOVÁHA, PASIVNÍ A AKTIVNÍ TRANSPORT
 TERMODYNAMICKÁ ROVNOVÁHA, PASIVNÍ A AKTIVNÍ TRANSPORT Termodynamická rovnováha systému je charakterizována absencí spontánních procesů. Poněvadž práce může být konána pouze systémem, který směřuje ke spontánní
TERMODYNAMICKÁ ROVNOVÁHA, PASIVNÍ A AKTIVNÍ TRANSPORT Termodynamická rovnováha systému je charakterizována absencí spontánních procesů. Poněvadž práce může být konána pouze systémem, který směřuje ke spontánní
Úvod do buněčného metabolismu Citrátový cyklus. Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK
 Úvod do buněčného metabolismu Citrátový cyklus Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK METABOLISMUS = přeměna látek v organismu - má stránku chemickou (látkovou) - reakce anabolické
Úvod do buněčného metabolismu Citrátový cyklus Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK METABOLISMUS = přeměna látek v organismu - má stránku chemickou (látkovou) - reakce anabolické
Biotransformace Vylučování
 Biotransformace Vylučování Toxikologie Ing. Lucie Kochánková, Ph.D. Biotransformace proces chemické přeměny látek v organismu zpravidla enzymaticky katalyzované reakce vedoucí k látkám tělu vlastním nebo
Biotransformace Vylučování Toxikologie Ing. Lucie Kochánková, Ph.D. Biotransformace proces chemické přeměny látek v organismu zpravidla enzymaticky katalyzované reakce vedoucí k látkám tělu vlastním nebo
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Respirace - buněčné dýchání (katabolismus)
") Respirace - buněčné dýchání (katabolismus) Schéma základního energetického metabolismu rostlinné buňky Fotosyntéza Fotochemie Calvinův cyklus Respirace Glykolýza (+ β-oxidace, ) Krebsův cyklus Dýchací
Respirace - buněčné dýchání (katabolismus) Schéma základního energetického metabolismu rostlinné buňky Fotosyntéza Fotochemie Calvinův cyklus Respirace Glykolýza (+ β-oxidace, ) Krebsův cyklus Dýchací
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Univerzita Karlova v Praze - 1. lékařská fakulta. Buňka. Ústav pro histologii a embryologii
 Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
E ŘEŠENÍ KONTROLNÍHO TESTU ŠKOLNÍHO KOLA
 Ústřední komise Chemické olympiády 48. ročník 2011/2012 ŠKOLNÍ KOLO kategorie A a E ŘEŠENÍ KONTROLNÍ TESTU ŠKOLNÍ KOLA KONTROLNÍ TEST ŠKOLNÍ KOLA (60 BODŮ) ANORGANICKÁ CEMIE 16 BODŮ Úloha 1 8 bodů Napište
Ústřední komise Chemické olympiády 48. ročník 2011/2012 ŠKOLNÍ KOLO kategorie A a E ŘEŠENÍ KONTROLNÍ TESTU ŠKOLNÍ KOLA KONTROLNÍ TEST ŠKOLNÍ KOLA (60 BODŮ) ANORGANICKÁ CEMIE 16 BODŮ Úloha 1 8 bodů Napište
Lékařská chemie a biochemie modelový vstupní test ke zkoušce
 Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
CZ.1.07/2.2.00/ Obecný metabolismus. Energetický metabolismus (obecně) (1).
 (1).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74. Katedra experimentální biologie rostlin, Z. Lhotáková
 FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Metabolismus sacharidů. VY_32_INOVACE_Ch0216.
 Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Předmět: KBB/BB1P; KBB/BUBIO
 Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno:
 Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
POZNÁMKY K METABOLISMU SACHARIDŮ
 POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
Redoxní děj v neživých a živých soustavách
 Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Vzdělávací materiál. vytvořený v projektu OP VK. Anotace. Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20. Číslo projektu:
 Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
BUŇKA A ENERGIE. kajman brýlový Caiman crocodilus Kostarika, 2004. Biologie 6, 2015/2016, Ivan Literák
 BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
NaLékařskou.cz Přijímačky nanečisto
 alékařskou.cz Chemie 2016 1) Vyberte vzorec dichromanu sodného: a) a(cr 2 7) 2 b) a 2Cr 2 7 c) a(cr 2 9) 2 d) a 2Cr 2 9 2) Vypočítejte hmotnostní zlomek dusíku v indolu. a) 0,109 b) 0,112 c) 0,237 d) 0,120
alékařskou.cz Chemie 2016 1) Vyberte vzorec dichromanu sodného: a) a(cr 2 7) 2 b) a 2Cr 2 7 c) a(cr 2 9) 2 d) a 2Cr 2 9 2) Vypočítejte hmotnostní zlomek dusíku v indolu. a) 0,109 b) 0,112 c) 0,237 d) 0,120
Buněčný metabolismus. J. Vondráček
 Buněčný metabolismus J. Vondráček Téma přednášky BUNĚČNÝ METABOLISMUS základní dráhy energetického metabolismu buňky a dynamická podstata jejich regulací glykolýza, citrátový cyklus a oxidativní fosforylace,
Buněčný metabolismus J. Vondráček Téma přednášky BUNĚČNÝ METABOLISMUS základní dráhy energetického metabolismu buňky a dynamická podstata jejich regulací glykolýza, citrátový cyklus a oxidativní fosforylace,
BIOKATALYZÁTORY I. ENZYMY
 BIOKATALYZÁTORY I. Obecné pojmy - opakování: Katalyzátory látky, které ovlivňují průběh katalyzované reakce a samy se přitom nemění. Dělíme je na: pozitivní (aktivátory) urychlující reakce negativní (inhibitory)
BIOKATALYZÁTORY I. Obecné pojmy - opakování: Katalyzátory látky, které ovlivňují průběh katalyzované reakce a samy se přitom nemění. Dělíme je na: pozitivní (aktivátory) urychlující reakce negativní (inhibitory)
METABOLISMUS SACHARIDŮ
 METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
ANABOLISMUS SACHARIDŮ
 zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Kofaktory enzymů. T. Kučera. (upraveno z J. Novotné)
") Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
- metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy a jejich životním prostředím
 Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Seznam použitých zkratek (v abecedním pořadí)
") Seznam použitých zkratek (v abecedním pořadí) A Adenin AD Alkoholdehydrogenasa ADP Adenosindifosfát AMP Adenosinmonofosfát ATP Adenosintrifosfát BM Bazální metabolismus ytosin AM rassulacean Acid Metabolism
Seznam použitých zkratek (v abecedním pořadí) A Adenin AD Alkoholdehydrogenasa ADP Adenosindifosfát AMP Adenosinmonofosfát ATP Adenosintrifosfát BM Bazální metabolismus ytosin AM rassulacean Acid Metabolism
Metabolismus aminokyselin. Vladimíra Kvasnicová
 Metabolismus aminokyselin Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze dalších
Metabolismus aminokyselin Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze dalších
Antioxidační ochrana. Stárnutí. Antioxidanty v potravě
 Antioxidační ochrana. Stárnutí. Antioxidanty v potravě MUDr. Jan Pláteník, PhD Ústav lékařské biochemie a laboratorní diagnostiky 1.LF UK Ionizační záření: Hydroxylový radikál vzniká ionizací vody: H 2
Antioxidační ochrana. Stárnutí. Antioxidanty v potravě MUDr. Jan Pláteník, PhD Ústav lékařské biochemie a laboratorní diagnostiky 1.LF UK Ionizační záření: Hydroxylový radikál vzniká ionizací vody: H 2
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Citrátový cyklus. VY_32_INOVACE_Ch0218.
 Vzdělávací materiál vytvořený v projektu P VK ázev školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: ázev projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK ázev školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: ázev projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Konsultační hodina. základy biochemie pro 1. ročník. Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa
 Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
ENZYMY. Charakteristika enzymaticky katalyzovaných reakcí:
 ENZYMY Definice: Enzymy (biokatalyzátory) jsou jednoduché či složené makromolekulární bílkoviny s katalytickou aktivitou. Urychlují reakce v organismech tím, že snižují aktivační energii (Ea) potřebnou
ENZYMY Definice: Enzymy (biokatalyzátory) jsou jednoduché či složené makromolekulární bílkoviny s katalytickou aktivitou. Urychlují reakce v organismech tím, že snižují aktivační energii (Ea) potřebnou
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
Úvod do biologie rostlin Transport látek TRANSPORT. Krátké, střední, dlouhé vzdálenosti
 Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Ukládání energie v buňkách
 Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
33.Krebsův cyklus. AZ Smart Marie Poštová
 33.Krebsův cyklus AZ Smart Marie Poštová m.postova@gmail.com Metabolismus Metabolismus je souhrn chemických reakcí v organismu. Základní metabolické děje jsou: a) katabolické odbourávací (složité látky
33.Krebsův cyklus AZ Smart Marie Poštová m.postova@gmail.com Metabolismus Metabolismus je souhrn chemických reakcí v organismu. Základní metabolické děje jsou: a) katabolické odbourávací (složité látky
Otázka: Metabolismus. Předmět: Biologie. Přidal(a): Furrow. - přeměna látek a energie
: Furrow. - přeměna látek a energie") Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Repetitorium chemie 2016/2017. Metabolické dráhy František Škanta
 Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
Cukry (Sacharidy) Sacharidy a jejich metabolismus. Co to je?
 Sacharidy a jejich metabolismus. Co to je?") Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
AMPK AMP) Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze
 Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze") AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Metabolismus. Source:
 Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Propojení metabolických drah. Alice Skoumalová
 Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Centrální metabolické děje
 Základy biochemie KB / B entrální metabolické děje Inovace studia biochemie prostřednictvím elearningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KB / B entrální metabolické děje Inovace studia biochemie prostřednictvím elearningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
ení s chemickými látkami. l rní optiky
 OPTICKÉ SENSORY Základem je interakce světeln telného zářenz ení s chemickými látkami. l Při i konstrukci katalytických biosensorů se používaj vají: optické techniky: absorbance fluorescence luminiscence
OPTICKÉ SENSORY Základem je interakce světeln telného zářenz ení s chemickými látkami. l Při i konstrukci katalytických biosensorů se používaj vají: optické techniky: absorbance fluorescence luminiscence
MASTNÉ KYSELINY, LIPIDY (BIOSYNTÉZA)
") MASTNÉ KYSELINY, LIPIDY (BISYNTÉZA) BSAH Syntéza mastných kyselin cholesterolu žlučové kyseliny isoprenoidy steroly Rozdíly mezi odbouráváním a syntézou mastných kyselin 1. Syntéza mastných kyselin probíhá
MASTNÉ KYSELINY, LIPIDY (BISYNTÉZA) BSAH Syntéza mastných kyselin cholesterolu žlučové kyseliny isoprenoidy steroly Rozdíly mezi odbouráváním a syntézou mastných kyselin 1. Syntéza mastných kyselin probíhá
Mitochondriální genom, úloha mitochondrií v buněčném metabolismu, signalizaci a apoptóze
 Mitochondriální genom, úloha mitochondrií v buněčném metabolismu, signalizaci a apoptóze MUDr. Jan Pláteník, PhD březen 2007 Mitochondrie:... původně fagocytované/parazitující bakterie čtyři kompartmenty:
Mitochondriální genom, úloha mitochondrií v buněčném metabolismu, signalizaci a apoptóze MUDr. Jan Pláteník, PhD březen 2007 Mitochondrie:... původně fagocytované/parazitující bakterie čtyři kompartmenty:
Fotosyntéza Světelné reakce. Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni
 Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
Biochemie jater. Vladimíra Kvasnicová
 Biochemie jater Vladimíra Kvasnicová Obrázek převzat z http://faculty.washington.edu/kepeter/119/images/liver_lobule_figure.jpg (duben 2007) Obrázek převzat z http://connection.lww.com/products/porth7e/documents/ch40/jpg/40_003.jpg
Biochemie jater Vladimíra Kvasnicová Obrázek převzat z http://faculty.washington.edu/kepeter/119/images/liver_lobule_figure.jpg (duben 2007) Obrázek převzat z http://connection.lww.com/products/porth7e/documents/ch40/jpg/40_003.jpg
ENZYMY A NUKLEOVÉ KYSELINY
 ENZYMY A NUKLEOVÉ KYSELINY Autor: Mgr. Stanislava Bubíková Datum (období) tvorby: 28. 3. 2013 Ročník: devátý Vzdělávací oblast: Člověk a příroda / Chemie / Organické sloučeniny 1 Anotace: Žáci se seznámí
ENZYMY A NUKLEOVÉ KYSELINY Autor: Mgr. Stanislava Bubíková Datum (období) tvorby: 28. 3. 2013 Ročník: devátý Vzdělávací oblast: Člověk a příroda / Chemie / Organické sloučeniny 1 Anotace: Žáci se seznámí