Oscilační aktivita humánního mozku
|
|
|
- Hana Beránková
- před 7 lety
- Počet zobrazení:
Transkript
1 Oscilační aktivita humánního mozku Daniela Sochůrková doktorandská práce LF MU Brno
2 Obsah Předmluva 1. Úvod 1.1. Oscilace 1.2. Problém zvaný generátor 1.3. Role thalamu hlavní subkortikální generátor? 1.4. Lokální vlastnosti neuronální sítě 1.5. Rezonantní chování mozku 2. Metodologie 2.1. Možnosti zpracování EEG signálu 2.2. Frekvenční specificity oscilací 2.3. Kvantifikace ERD/ERS v čase a v prostoru Časový průběh Zobrazení průběhu ERD/ERS v prostoru Výběr individuálního frekvenčního pásma 2.4. Interpretace výkonových změn Event-related desynchronizace (ERD) Event-related synchronizace (ERS) Simultánní výskyt ERD a ERS 3. Bazální a aplikovaný výzkum 3.1. Elektrofyziologie pohybu a kortex 3.2. Elektrofyziologie pohybu a subkortikální struktury 3.3. Oscilace v epileptologii 4. Závěr 2
3 5. Seznam použitých zkratek 6. Poděkování 7. Literatura 8. Přílohy 8.1. Publikace 8.2. Spoluautorství 8.3. Obrázek č. 9a-f 3
4 Název: Oscilační aktivita humánního mozku Autor: Daniela Sochůrková Předmluva K obsahu předmluvy k doktorandské práci mě přivedlo jedno letní nedělní odpoledne. V Setkávání, v pravidelném programu rozhlasové stanice Vltava, mluvil Jan Sokol. Filosof Jan Sokol. Vysvětloval, že vše kolem nás a my sami je založeno na rytmicitě, na střídání fází: systola-diastola, vdech-výdech, všední-sváteční, zima-léto, spánek-bdění synchronizacedesynchronizace. Nositelem významu, v nejširším slova smyslu, je dle něj pak ROZDÍL. Proslov ještě doplnil citátem z Jana Patočky: Člověk žije v AMPLITUDĚ Nevím, do jaké míry byl a je Jan Sokol obeznámen s neustálou oscilační aktivitou našich mozků, s faktem, že oscilace je právě i v tomto případě nahlížena coby nositelka významu, nositelka informace. Ale ať již tomu bylo jakkoliv, byl to jen, řekněme, metafyzický pohled na problematiku, kterou se elektrofyziologové snaží uchopit a vysvětlit od dob Bergerových, od 30. let minulého století, od jím prvně popsané desynchronizace nad týlními oblastmi mozku při otevření očí. Trvalo však dalších 50 let, než pánové Pfurtscheller a Aranibar rozložili nepravidelnou křivku naší mozkové aktivity na jednotlivá frekvenční pásma a začali pozorovat, počítat, co se v nich odehrává při té či oné činnosti, které oblasti našeho mozku se nejspíše zapojují do dané aktivity a jak. Pojmy synchronizacedesynchronizace tak dostaly nový rozměr. Rozměr, jenž svým obsahem začal od té doby velmi výrazně přispívat do studnice vědomostí týkající se výzkumu fungování neuronálních sítí. Já jsem měla možnost studovat tuto problematiku téměř z největší blízkosti díky analýze elektrické aktivity mozku přímo ve zdroji za pomocí vnořených makroelektrod. Výsledky dlouholetého výzkumu jsou shrnuty v následujícím textu. 4
5 1. Úvod Mozková aktivita může být zaznamenávána a analyzovaná pomocí širokého spektra přístupů. V mé práci se budu zabývat hlavně jednou z metod elektrofyziologie na podnětu závislé desynchronizaci a synchronizaci (Event-related synchronization and desynchronization; ERD/ERS), která je používána ke studiu mozkových oscilací. Krom vysvětlení pojmu, metody a obeznámení se světovým písemnictvím uvedu výsledky vlastních výzkumů. Výzkum byl zaměřen na studium volní motoriky a na studium některých kognitivních funkcí. Volená paradigmata umožňovala současnou analýzu tzv. evokovaných i indukovaných změn (blíže viz dále v textu). Hlavním cílem práce bylo stanovení shodnosti či rozdílnosti jejich distribuce a potažmo interpretace významu existence obou elektrofyziologických fenoménů Oscilace Ze záznamu elektrické aktivity mozku je patrné, že registrovaný signál je průnikem nejrůznějších frekvenčních komponent, oscilací, jejichž intenzita v čase je nestálá, odvislá, obecně, od vnitřních a vnějších podmínek. Jaký je fyziologický význam oscilací, kde jsou oscilace vytvářeny, generovány, jak souvisejí s patologickými stavy mozkové činnosti na to vše a na další otázky se pokouší přinést odpověď jejich analýza pomocí metody ERD/ERS. V klidovém, relaxovaném stavu bez mentálního úsilí, při zavřených očích je nad zadními mozkovými kvadranty zaznamenávána EEG aktivita u většiny lidí ve formě vysoce koherentní, téměř sinusoidální 10 Hz oscilace. Berger v roce 1930 jako první přišel 5
6 s pozorováním, že tato oscilace se zcela změní, sníží amplitudu, tedy tzv. se desynchronizuje, suprimuje, při otevření očí. Považoval to již tenkrát za odraz aktivace vizuálního kortexu. Stejná odezva byla dále popisována u mentálního úsilí, v situacích zvýšené pozornosti, při vizuálním podnětu apod. Jasper a Andrew pak o několik let později, v roce 1938, popisují obdobnou reaktivitu ale centrálního mu rytmu po taktilním podnětu. V roce 1949 opět Jasper, s Penfieldem, demonstrovali supresi centrální beta aktivity po volním pohybu a to na datech získaných elektrokortikograficky. Gastaut pak roku 1952 publikuje údaje o supresi mu rytmu při kontralaterálním volním či pasivním pohybu či i somatosensorické stimulaci. Teprve ale roku 1977, po zavedení výpočtu kvantifikace amplitudových respektive výkonových změn ve zvolených frekvenčních pásmech Pfurtschellerem a Aranibarem se rozkrývání změn v oscilacích dostává do popředí zájmu u výzkumu zaměřeného na fungování neuronálních sítí. Postulují se hypotézy předpokládající existenci zdrojů oscilací, existenci neuronálních sítí, jejichž funkce a funkčnost jsou určovány změnami v činnosti jejich zdrojů, generátorů. Pro uchopení představy o oscilační aktivitě mozku je nezbytné si uvědomit, že komunikace mezi neurony probíhá v nejrůznějších časových a prostorových rovinách, jež se navzájem prolínají. Dynamika těchto neokortikálních procesů je měřitelná metodami s krátkou či dlouhou časovou základnou. Ty první z nich informují lépe přímo o dynamice a patří mezi ně elektroencefalografie (EEG) a magnetoencefalografie (MEG). Naopak, metody s dlouhou časovou základnou (PET, SPECT, fmri, MRS) jsou vhodné více pro studium metabolických pochodů, jejich výraznou výhodou je možnost lepší reálné prostorové zobrazitelnosti. 6
7 1.2. Problém zvaný generátor Co vlastně měří EEG? Lapidárně lze konstatovat, že měří sumaci inhibičních a excitačních postsynaptických potenciálů. Kortikální pyramidové buňky, uspořádané do vrstev a propojené mnoha spoji, jsou zdrojem lokálních elektrických proudů, potažmo elektromagnetických polí; summa těchto změn pak vytváří tzv. neokortikální dipólovou vrstvu. Aktivita dipólu, změna jeho prostorové orientace, lokalizace dipólu se odráží ve výsledném získávaném elektroencefalogramu (EEG). Okamžitý moment dipólu je určován aktivitou neuronálních buněk v jednotce objemu o rozměru několika milimetrů. Tato jednotka je považována za zdroj respektive za generátor. Sumace aktivity jednotlivých generátorů, která může být koherentní či nekoherentní (oscilace dipólu na stejné či jiné frekvenci), se matematicky sčítá resp. odečítá. Koherence je potom jev makroskopicky sledovatelný při posuzování změny aktivity mozku mezi různými oblastmi. I přesto, že výše uvedené vysvětlení pojmu generátor oscilací se jeví poměrně snadno pochopitelné, opak je pravdou; problém totiž vyplývá z faktu, že změny v oscilování nejsou, na rozdíl od evokovaných potenciálů (ERP, SSEP, BAEP, VEP, MEP), fázově koherentní děje a že fázový zvrat (phase reversal), jenž je považován za průkaz generátoru evokovaného potenciálu, nebyl nikdy podán jako jednoznačný důkaz přítomnosti generátoru oscilací. Oscilační změny jsou zejména dány radiálními dipóly, které rychle mění amplitudu, orientaci i lokalizaci v rozsahu několika čtverečních milimetrů či centimetrů v závislosti na oscilační frekvenci. Lze si tedy zdroje kortikálních oscilací představit jako ohraničenou skupinu neuronů, které vytvářejí díky synaptickým spojům a pravděpodobně i thalamické aferentaci synchronizovanou rytmickou aktivitu. Projevem činnosti takového generátoru může být pokles nebo nárůst amplititudy respektive výkonu aktivity; pro přítomnost generátoru svědčí 7
8 nejen vysoký absolutní výkon v daném frekvenčním pásmu, ale také právě jeho reaktivita (Andrej Stančák, ústní sdělení) Role thalamu hlavní subkortikální generátor? Myšlenka, že rytmická EEG aktivita je primárně určena činností pacemakerů, pochází z šedesátých let (Andersen and Andersson, 1968). Jahnsen a Llinás přinášejí v roce 1984 důkaz o spontánní oscilatorní aktivitě některých typů thalamických neuronů i v případě, že je zablokovaná synaptická transmise. Thalamus je rytmogeneticky vysoce variabilní. Je schopný produkovat oscilace nízké frekcence (1-4 Hz), střední (12-17 Hz) a vysoké (20-60 Hz). Hlavní roli při generování má pak nucleus reticularis thalami projikující exklusivně do dalších jader thalamu s následným účinkem na tzv. relay a inhibiční interneurony. Chování retikulárních neuronů je odvislé od klidového membránového potenciálu; hypepolarizace vede k rytmickým výbojům v delta pásmu, klidové napětí kolem -60 uv je asociováno s oscilační aktivitou spindle-like 7-14 Hz, a naopak, depolarizace resp. klidový membránový potenciál vyšší než -60 uv koresponduje s vysokofrekvenční aktivitou kolem 40 Hz. Je legitimní považovat takové skupiny neuronů za autonomní, z tohoto hlediska i srovnatelné se specializovanými pacemakerovými buňkami srdečního svalu. V reálném stavu ale neexistují tyto skupiny neuronů (a samozřejmě žádné jiné) izolovaně, nýbrž jsou zapojeny do různých okruhů, jsou součástí neuronálních sítí. Tedy vždy, když se hovoří o mechanismech odpovědných za rytmické, oscilatorní chování mozku, je třeba vzít v potaz dynamiku neuronální sítě, její zpětněvazebný charakter a nepřeceňovat intrinsické vlastnosti různých neuronálních skupin (Lopes da Silva, 1991; Buzsáki, 1991). 8
9 Záhy ale bylo rozpoznáno, že rytmická aktivita ovšem není generována jen v subkortikálních oblastech, nýbrž i v oblastech kortikálních. Experimentální modely, většinou animální, z osmdesátých a devadesátých let, prokazují existenci oscilací neokortikálních interneuronů v rozpětí Hz, vyšší frekvence je pak dosahováno v případě jistého stupně depolarizace navozené nejspíše vlivem aktivity thalamu na neokortex. V žádném z modelů se však nepotvrdila existence předpokládané uniformní neokortikální oscilatorní aktivity kolem 40 Hz ani nebyl nalezen hlavní pacemaker kortikální aktivity, tak jak se dá snad označit thalamus za hlavní generátor oscilací mezi strukturami subkortikálními. I přesto lze ale neokortex vzhledem k velké denzitě krátkých intrakortikálních (do 1mm) a kortikokortikálních (do 20 cm) vláken považovat za separovaný, samostatně fungující systém, do něhož vstup ze středního mozku lze mít za externí kontrolu (Braitenberg, 1978; Katznelson, 1981). Nejoprávněnější je ale nejspíše nahlížet neokortex a thalamus coby komplementární struktury při generování mozkové rytmické aktivity Lokální vlastnosti neuronální sítě Lokální vlastnosti neuronální sítě jsou určovány intrinsickými a extrinsickými vlastnostmi. Za ty první jsou považovány lokální okruhy, excitační či inhibiční. V thalamu např. mají neurony retikulární formace a lokální interneurony inhibiční (GABA-ergní) vliv na tzv. thalamokortikální okruh. Zevní, extrinsická kontrola, je pak hlavně cholinergní, méně již serotoninergní a noradrenergní. Výsledkem cholinergního působení je utlumení inhibice thalamokortikálního okruhu, snížení výskytu pomalých oscilací a naopak objevení se spánkových vřetének. Dále např. při terapii barbituráty a tedy posílení GABA-ergní vazby se objevují nízkofrekvenční oscilace. Oba výše citované modely jsou nezbytně 9
10 zjednoduššujícími komplexnost reálné situace s mnoha faktory, jejichž výsledkem je potom oscilatorní aktivita thalamokortikální neuronální sítě. Otázkou stále zůstává, co je vlastně anatomickým a fyziologickým substrátem synchronní aktivity simplifikovaně vyjádřeno je to skupina neuronů navzájem propojených excitačními a inhibičními synapsemi. Kvantitativní anatomie (Braitenberg a Schuz, 1991) prokazuje existenci cca 8000, z 85% excitačních synapsí, na oblast tzv. 1 mm cross-section tvořených krátkými intrakortikálními spoji. Vzniklý akční potenciál jednoho neuronu vyvolává excitaci jiného, trvající 5-15 msec, s časovým zpožděním 1 msec. Akční potenciál jednoho neuronu může ale v jiných případech být zodpovědný za inhibici jiných neuronů, inhibice bývá pak časově delší, msec. Variace a permutace nahlíženo z úhlu modelování neuronálních sítí jsou pak samozřejmě nesčetné, nutná je regulace. Regulace oscilatorních aktivit je velmi komplexní, využívající velké množství vložených relé, ať už v paralelním či sériovém uspořádání. Důvodem tak vysoké složitosti systému je požadavek zajištění nesmírně jemných nuancí potřebných ve všech sférách mozkové činnosti (kognitivní funkce, motorika se svým pyramidovým a extrapyramidovým systémem, regulace spánku-bdění, atd ). Těmto nárokům vychází vstříc velké množství specializovaných podskupin neuronů, jež mají či mohou mít různé intrinsické membránové vlastnosti a také různé vlastnosti resonantní. Externí kontrola funkcí oscilátorů je pak umožněna právě díky komplexnosti neuronálních sítí pouze relativně omezeným možstvím neuromediátorů a neurotransmiterů. Neuromodulátory jsou rozdělitené do dvou skupin: a/ desynchronizujicí (ACh, NA, DA) a b/ synchronizující (5-HT). Obě skupiny neuromodulátorů a jejich ovlivnění oscilací mají klinickou aplikaci, byť mnohdy ještě hodně spekulativní: idiopatická Parkinsonova choroba s nedostatkem dopaminu a tudíž s pozdní desynchronizací (Defebvre a spol, 1994), klinicky pozorovatelnou jako poruchu rychlé iniciace motoriky a kognice, lze nahlížet jako hypersynchronní stav. Naopak, hyposynchronní stav s předpokládanými četnými lokálními generátory bez vzájemné souhry je hypotetickým 10
11 elektrofyziologickým podkladem např. tikové chorby Gilles de la Tourette (zvýšená hladina monoaminů?) či i např. schizofrenie Resonantní chování mozku K pojmu oscilace se pojí, fyzikálně, i pojem resonance. Názornou pomůckou pro objasnění této složité problematiky budiž nám strunné nástroje (Obrázek č. 1). Struna vibruje sice na jedné, základní, dominantní frekvenci, ale fyzikálně, ale event. i hudebně při velmi disponovaném sluchu, lze rozpoznat tzv. alikvóty, tedy harmonické frekvence, jejichž míra, intenzita, počet atd. je dána místem úderu do struny. Struna samotná vydává zvuk velmi malé intenzity. Amplifikace je umožněna transferem oscilací přes kobylku nástroje na jeho tělo. Vibrace dřeva a vzduchu v dutině nástroje poté přispívají k výslednému zvuku nástroje, aniž by nutně, ba naopak tomu tak většinou nebývá, tělo nástroje vibrovalo na stejné základní frekvenci jako struna. Také samozřejmě dochází k procesu opačnému, k přenosu vibrací zpět z těla na strunu nástroje. Výsledná souhra pak určuje barvu (timbre) vyluzovaného zvuku, jeho intenzitu atd. Takto lze přibližně popsat pojem resonance, coby de facto přenos mechanické energie z jedné struktury na druhou, jejich vzájemné ovlivňování se, vlastní produkci harmonických či subharmonických frekvencí, a v neposlední řadě lineárnost (přímost) či nelineárnost (nepřímost) interakcí mezi zúčastněnými strukturami. Důležitým, a při studiu mozkových oscilací relevantní poznatkem, je ještě fakt, že budou-li všechny charakteristiky např. dvou nástrojů stejné s výjimkou délky strun, bude nástroj s delšími strunami schopný produkovat zvuk i v nižším frekvenčním pásmu. Dalším modelem, směřujícím k usnadnění pochopení oscilací v gamma pásmu, je představa velmi krátkých strun, jež jsou připevněny na struny dlouhé a jež mohou být zdrojem harmonických oscilací 11
12 velmi vysokých frekvencí. A hlavně; ať už se jedná o tu či onu délku struny, potažmo tedy vlnovou frekvenci, o ten či onen model, účel je jediný - zajistit včasný přenos informace, v případě hudebního nástroje tedy zvuk, v případě mozku informaci nejrůznějšího charakteru více či méně relevantní pro danou konkrétní situaci. Obrázek č. 1 (použito z Nuneze) 12
13 Vše výše popsané se tedy dá volně aplikovat na mozkové oscilace. Vzájemný poměr excitatorních a inhibičních synapsí a externí kontrola jsou potom faktory určující přítomnost či nepřítomnost oscilací v určitých frekvenčních pásmech. Otázkou opětnou je lokalizace oscilačních generátorů, které jsou pak schopné resonovat s jiným neuronálním systémem. Lokalizace generátorů, jak již bylo zmíněno, není dodnes vyjasněna. Poměrně jasně mluví elektrofyziologické nálezy pro existenci thalamických generátorů ovlivňujících neokortex, existence samostatných kortikálních generátorů je však hůře prokazatelná. Dalším zajímavým a důležitým aspektem oscilací je jejich mezidruhová charakteristika. Obratlovci vykazují tzv. low pass behavior, většina spektrálního výkonu mozkové aktivity leží pod 100 Hz. Bezobratlí, naopak, mají maximum spektrálního výkonu nad 100 Hz. Předpokládá se tedy, že výsledné EEG, vysokofrekvenční, odráží aktivitu individuálních malých skupin neuronů s malou mírou interkonektivity či komplexnosti systému, která se naopak jeví být pravděpodobná u pomalých oscilací nebo u ostrých vrcholů (spectral peak) různých frekvenčních pásem. Tyto dva elektrofyziologické fenomény, pomalé oscilace a ostré vrcholy, jsou odrazem vysoce koherentního chování. Je hypoteticky možné, že právě přechod z převážně vysokofrekvenčního, ostrůvkovitého a na malou vzdálenost přenos informace zajišťujícího systému do systému převážně nízkofrekvenčního, jenž je schopen zajistit přenos informace na větší vzdálenost, propojit mezi sebou více oblastí, byl fylogeneticky významným krokem ve vývoji a zdokonalování lidského mozku. To ale nikterak neznamená, že by lidský mozek nebyl schopen a také neosciloval na vysokých frekvencích a že vysokofrekvenční oscilace nemají pro člověka důležitost (blíže viz kapitola 3.3.). Výzkumy posledních let při stálém zlepšování technických parametrů EEG rekordérů a analyzátorů naopak prokazují existence oscilací o frekvencích řádově až do 500 Hz ( fast ripples ). Jedná se ale díky fyzikálním vlastnostem oscilací jen o tzv. lokální resonance. 13
14 Rytmická aktivita nad 30 Hz je výrazem činnosti vzato prostorově neuronální skupiny s milimetrovým rozměrem. Takové oblasti mohou makroskopicky představovat např. thalamický generátor či kortikální kolumnu. Opakovaně bylo prokázáno (Llinas, 1992), že rytmická aktivita kolem 40 Hz je s vysokou pravděpodobností součástí či samotným substrátem negativní zpětné kličky, že je aktivitou nasedající jako malá lokální struna na velkou strunu zpětnovazebného okruhu a to v případě, že gain zpětnovazebné kličky je dostatečně velký. V jaké míře je při generování této rytmické aktivity nezbytný thalamus, je neobjasněno, předpokládá se ale, že jeho význam je zásadní. Lokální resonance, na rozdíl od regionálních resp. globálních oscilací, nemusí být koherentní na vzdálenost větší než několik milimetrů. Ale právě tyto lokální vysokofrekvenční koherence představující vysoký stupeň časově-prostorové specifické aktivity v neokortexu jsou nezbytné při kognitivních procesech, spoluurčují např. adekvátní sociální chování. 14
15 2. Metodologie 2.1. Možnosti zpracování EEG signálu Neuronální populace reagují na zevní či vnitřní podnět časově vázanou odpovědí. Analýza této reakce je možna více elektrofyziologickými postupy, jež se dají ve své podstatě rozdělit do dvou skupin. Prvním, a v praxi denně požívaným přístupem, je metoda zprůměrnění úseků EEG podle daného podnětu. Výsledkem pak je, díky zvýšení poměru signál/šum, křivka s vykreslením na podnět nejen časově ale i fázově vázané odpovědi. Široce a všeobecně jsou tyto odpovědi nazývány evokovanými potenciály (EP) a jsou nahlíženy jako výsledek reorganizace fází mozkové aktivity v určitých neuronálních populacích (Sayers et al., 1974). Výzkumy však přinesly informace o tom, že za určitých okolností, při použití určitých paradigmat je nalézána fázově vázaná, pomalá, pozvolná odpověď (tzv. DC shift) i již před očekávaným podnětem (contingent negative variation, CNV, Obrázek č.2) či před volním pohybem (Bereitschaftspotential, BP, Obrázek č.3). Tento první přístup k elektrofyziologické analýze EEG však nikterak není schopen, a také si neklade za cíl, zhodnotit, co se děje právě s oním šumem pozadí. V tomto případě již nestačí jednoduché lineární metody, nýbrž je potřeba použít frekvenční analýzy, jež umožní zjistit, co se odehrává v jednotlivých vybraných frekvenčních pásmech v době před, během a po podnětu. Pokles či zvýšení amplitudy respektive výkonu ve vybraném frekvenčním pásmu pak představuje na podnětu závislou, ale fázově nevázanou odpověď, jež je nahlížena jako obraz snížení či zvýšení synchronie určité neuronální populace. Právě Pfurtscheller a Aranibar (1977) zvolili pro pokles synchronie pojem Event-related desynchronization ( na podnětu závislá synchronizace, ERD) a Pfurtscheller pak v roce 1992 přidává pojem Event-related 15
16 synchronization (ERS) pro stav zvýšené synchronie. Důležité je poznamenat, že oba fenomény jsou zjišťovatelné nejen analýzou EEG křivky, ale i pomocí magnetické encefalografie (MEG). Obrázek č. 2 Obrázek č. 3 16
17 K pochopení obou elektrofyziologických dějů je potřeba vycházet z faktu, že neuronální sítě se mohou nacházet v různých stavech synchronie, že mohou oscilovat na různých frekvencích. V případě podnětu, ať už vnitřního či vnějšího, dochází ke specifickým změnám mozkové oscilační aktivity, určitá neuronální populace se stává terénem pro zpracování informace. Změnám, k nimž dochází na pozadí mozkové aktivity díky změnám parametrů kontrolujících oscilace, se pak říká změny indukované. Naopak, změny, na nichž se více podílí ascendentní senzorické aferentace, jsou nazývány změny evokované (Obrázek č.4). Předpokládané parametry, jež určují oscilační, a tedy indukovanou, aktivitu jsou: a/ intrinsické vlastnosti buněčných membrán neuronů a dynamika synaptických procesů b/ intenzita a rozsah interkonekcí mezi elementy neuronální sítě, často vytvářenými zpětnovazebnými okruhy; zpětnovazebné okruhy jsou principiálně dvojí: thalamokortikální a kortiko-kortikální c/ modulace neuronální sítě lokálními a systémovými neurotransmitery Obrázek č. 4 (použito z Pfurtschellera a da Silvy) 17
18 2.2. Frekvenční specificity oscilací Amplituda oscilací je proporcionální množství resp. počtu synchronně aktivních neuronálních elementů. Platí, že frekvence oscilací negativně koreluje s jejich amplitudou. Důležitým aspektem tohoto elektrofyziologického dogmatu je potom možnost šíření a tedy i registrovatelnost změn ve vybraných frekvenčních pásmech. Změny stavu neokortexu na makroskopické úrovni (jako např. densita synaptických akčních potenciálů) se mohou projevit jako šířící se vlny (traveling waves) odrážející postupnou synaptickou aktivaci. Čím nižší bude frekvenční pásmo, tím bude propagace oscilací možná na větší vzdálenost a také bude větší jejich schopnost překonávat překážky ve formě různých tkání (CSF, mozkové pleny, skelet ) s jejich výrazně odlišnými vodivostními charakteristikami. Tyto fyzikální parametry oscilací jsou směrodatné pro možnost jejich registrace a potažmo analýzy. Kupříkladu při registraci pomocí povrchových skalpových elektrod je z důvodu tkáňových charakteristik velmi nesnadné analyzovat změny ve frekvenčním pásmu gamma, neboť skalpová propagace gamma aktivity je výrazně omezena. Intrakraniální registrace obchází výše zmíněné tkáňové bariéry, ale naráží zase na na jiné problémy; elektrokortikografická analýza (ECoG) kromě nadále přítomné bariéry ve formě měkkých plen a cerebrospinální tekutiny snímá elektrickou aktivitu z poměrně velké plochy několika čtverečních centimetrů, což stěžuje lokalizaci oscilací, intracerebrální registrace je pak sice nejpřesnější, aktivita neuronálních buněk je snímána naopak ale z tak malého objemu tkáně, že např. nepřítomnost gamma oscilací na kontaktu hluboké elektrody nevylučuje přítomnost generátoru o několik milimetrů vedle. 18
19 2.3. Kvantifikace ERD/ERS v čase a v prostoru Metoda ERD/ERS je principiálně založena na kvantifikaci amplitudových respektive výkonových změn v čase. Absolutní hodnota daných parametrů má jen omezený význam (předpokládá se důležitost klidového výkonu u oscilátorů); naopak relativní hodnota s jejími průběžnými změnami je relevatní informací při posuzování oscilačních změn. Blíže je kvantifikace popsána v následujících podbodech Časový průběh K vyjádření ERD/ERS se používá procentuální, tedy relativní vyjádření změny amplitudy resp. výkonu ve zvolené frekvenci (Obrázek č. 5). Změna se pak vyjadřuje pro vybraný časový interval a vztahuje se k hodnotě amplitudy resp. výkonu bazálnímu, jenž se vypočítává z úseku EEG křivky nejméně několik sekund před podnětem. Stejně jako při získávání evokovaného potenciálu je i zde nutné analyzovat dostatečné množství dat, respektive úseků EEG získaných opakování uloženého úkolu ve zvoleném paradigmatu. 19
20 Obrázek č. 5 Faktický způsob zpracování hrubých EEG dat je pak možný mnohý (Obrázek č. 6): 1/ výkonová analýza frekvenčního pásma 2/ intertrial variance 3/ Hilbertova transformace 4/ komplexní demodulace 5/ wavelet transformace atd. Klasická, tedy výkonová analýza (Pfurtscheller and Aranibar, 1977) je prováděna v následujících krocích: a/ filtrace hrubých EEG b/ umocnění hodnot amplitud druhou mocninou pro získání výkonových hodnot c/ zprůměrnění všech úseků resp. jejich hodnot výkonu d/ vyjádření procentuální změny výkonu referenční versus vybraný časový interval 20
21 ERD/ERS (%) = (A-R)/R x 100 V referenčním úseku je hodnotě výkonu přisouzena nulová hodnota. Obrázek č. 6 (použito z Pfurtschellera) Tato klasická metoda nedokáže eliminovat příměs fázově vázaných odpovědí, které mohou výrazně ovlivňovat analýzu indukovaných odpovědí v oscilačních sítích. Odstranění resp. odečtení této odpovědi je dosaženo pomocí metody 2/, kdy před výkonovou analýzou je provedeno běžné způměrnění přeběhů a výsledný evokovaný potenciál je matematicky odečten od každého úseku zvlášť. Teprve poté se provádí výkonová analýza. Opakovaně v textu zmiňuji, že ERD/ERS hodnotí změny amplitudy RESPEKTIVE výkonu v čase ve vybraném frekvenčním pásmu. Obě varianty, jak amplitudová tak i výkonová analýza, jsou 21
22 možné a správné. Rozdílné na nich je, jak ukázala i naše práce P3 and ERD/ERS in a visual oddball paradigm: a depth EEG study from the mesial temporal structures (akceptováno pro tisk J Psychophysiology), že výkonová analýza akcentuje, díky umocňování na kvadrát, rozdíly v hodnocených časových intervalech či hodnoty v jednotlivých bodech vzorkovací frekvence registrovaných dat (např. rozdíl mezi hodnotou 0,5 a 1,5 bude jiný, než v případě porovnávání druhých mocnin obou hodnot). Vezme-li se tento fakt v úvahu, je amplitudová analýza korektnější a přesnější. Použití výkonové analýzy je ovšem legitimní, neboť je-li výkon v referenčním intervalu označen relativně za nulový, výchozí, ERD ve vyšetřovaném intervalu by dosahovala záporných hodnot, což z úhlu pohledu fyziologie je logický non-sens. Další omezení klasické výkonové analýzy spočívá v tom, že hodnoty jsou získávány coby průměr hodnot ve zvoleném časovém okně; toto posuvné okno je nasazené na začátek analyzovaného úseku, má svojí určitou délku (nejčastěji 1 sec) a pohybuje se vždy o určitý úsek (např. o 125 ms). Aktuální hodnota výkonu se pak vypočítává jako průměr 8 hodnot (vždy pro každou 125. ms celého posuvného okna) a hodnota se umísťuje doprostřed posuvného okna. Oproti tomu metoda komplexní demodulace je postavená na vytvoření obálky nad každým bodem vzorkovací frekvence, informuje tedy zcela přesně o průběhu amplitudových či výkonových změn v čase. Stanovení začátku nástupu změny výkonu se stanovuje dle následujícího principu (Obrázek č. 6a): 22
23 Zobrazení průběhu ERD/ERS v prostoru Záměrem ERD/ERS metody, stejně ale tak jako standardního EEG či EP je určit, kde se nachází zdroj, generátor studovaných elektrofyziologických jevů. V případě analýzy dat získaných z vnořených elektrod tento problém částečně odpadá, z charakteru křivky evokovaného potenciálu či DC shifts (CNV, BP) se dá usuzovat, zda se kontakt elektrody nachází blízko, vzdáleně či popřípadě i přímo ve zdroji generování evokované odpovědi; hovořit však o generátorech oscilačních změn je velmi obtížné, dodnes de facto nebylo stanoveno, jak by elektrofyziologicky měly generátory vypadat. Při analýze skalpových dat je pak nutno obdržet tzv. reference-free data, konvertování dat se provádí více možnými způsoby: a/ pomocí tzv. common average reference b/ pomocí tzv. Laplacian reference c/ pomocí tzv. local average reference Takto upravená data lze pak použít pro anatomické či reálné mapování elektroencefalografických změn, získané hodnoty zjišťovaných parametrů lze mnohem přesněji lokalizovat (Obrázek č. 7). 23
24 Obrázek č. 7 (použito z Pfurtschellera) Výběr individuálního frekvenčního pásma Frekvenční analýza a potažmo analýza amplitudivých či výkonových změn je nejcitlivější, zvolí-li se co nejužší frekvenční pásmo (omezení je fyzikálně dané). Stejně jako různé podněty vyvolávají změny v různých frekvenčních pásmech, existuje i interindividuální variabilita při reaktivnosti na stejný podnět. Je tedy zcela zásadně důležité stanovit pro daný vyšetřovaný subjekt individuální frekvenční pásmo a vyjadřovat se ke změnám vztahujícím se právě k tomuto pásmu. K tomuto účelu lze použít následující metody: a/ detekce nejreaktivnějšího pásma pomocí srovnání dvou krátkých úseků výkonových spekter (bazální interval, inkriminovaný interval kolem podnětu) v několika přebězích 24

25 b/ kontinuální tzv. wavelet transformace c/ definováním frekvenčního pásma relativně k výkonovému maximu, vrcholu (peak) celé frekvenční spektrální analýzy (Obrázek č. 8) Obrázek č Interpretace výkonových změn Proces od popisu fenoménu ke klinickému významu oscilačních změn je velmi obtížný. Jak synchronizace, tak i desynchronizace mohou odrážet (či být přímým terénem ) fyziologických nebo patologických dějů. Navíc, jak potvrzují výzkumy, se oba děje vyskytují 25
26 většinou současně. Zcela rozhodující pro interpretaci pak bude zjištění, v kterém frekvenčním pásmu je daný jev pozorován. Samozřejmě, nelze hodnotit jen výskyt či nevýskyt daného fenoménu, dobu jeho vzniku, jeho lokalizaci, lateralizaci, nýbrž je nutné hodnotit míru, procento dané změny. Většina studií přináší výsledky, že mezi kvalitou zvládnutí požadovaného úkolu a mírou desynchronizace či synchronizace existuje vztah přímé úměrnosti. Proti těmto výsledkům je postavena hypotéza tzv. neural efficiency - je-li subjekt schopen efektivněji využít neuronální sítě, již musí aktivovavat během zadaného úkolu, pak oscilační změny budou fokalizovanější a méně výrazné. Faktorem, jenž zcela nutně ovlivňuje pohled na synchronizaci a desynchronizaci je údaj Braitenberga a Schuze (1991). Dle těchto autorů, jak již bylo výše uvedeno, je cca 85% kortikálních neuronů excitatorních, zbylé jsou inhibiční. Inhibice je však nesmírně důležitá nejméně ze dvou důvodů; optimalizace energetických nároků a kontrola excitatorních procesů. Hypoteticky dalším důvodem je nezbytná kontrola míry informačních vstupů do neuronální sítě např. již zpracovávající informaci. Proces synchronizace a tedy deaktivace, aktivní inhibice je například pozorován během NREM spánku. Všechny spánkové, fyziologické grafoelementy odrážejí změnu oscilačních charakteristik ve smyslu koherentně se chovajících neuronálních skupin; vertexové ostré vlny, spánková vřeténka, K komplexy, vysokovoltážní delta vlny v hlubokých stádiích NREM spánku. Otázka místa generování této aktivity je vyřešena je parciálně; Steriade a Llinas (1988) považují např. spánková vřeténka za výraz bloku synaptické transmise přes thalamus v časných spánkových stádiích. 26
27 Event-related desynchronizace (ERD) Přechod ze stavu synchronizace do stavu se synchronizací nižší je všeobecně nazírán jako výraz aktivace určité konkrétní skupiny neuronů. Tento závěr je podepřen výsledky mnohých studií, v nejrůznějších paradigmatech, ať už z dat získaných skalpovým snímáním, nebo z elektrokortikografií či vnořených elektrod. Nejrozsáhlejším fenoménem je desynchronizace v pásmu alfa (8-12 Hz). Odráží, vysoce pravděpodobně, všeobecnou aktivaci kortikálních oblastí implikovaných při zpracování daného úkolu. Desynchronizace ve vyšších frekvenčních pásmech jsou, jak již vyplývá z fyzikálních vlastností oscilací, jevy mnohem více prostorově ohraničené, více somatotopičtější. Beta desynchronizace (12-30 Hz) je fenomén standardně patřící k motorickým paradigmatům. Z analýz skalpových dat se dlouho předpokládalo, že centrální mozková rýha je dělící strukturou mezi alfa a beta oscilacemi (první byvše postcentrální a druhá precentrálním fenoménem), data získaná intracerebrálními registrací ale vyvracejí tento předpoklad, beta aktivita je nacházena i nad zadními mozkovými kvadranty a vice versa. Desynchronizace v delta, theta a gamma pásmu nebývá až na výjimky popisována. Delta pásmo není v oblasti oscilací příliš předmětem zájmu, theta pásmo vykazuje nestandardně synchronizaci či desynchronizaci, nepanuje všeobecný konsensus, jenž by komentoval změnu v tomto frekvenčním pásmu v motorických či kognitivních paradigmatech. Gamma pásmo vykazuje synchronizaci, desynchronizace je popsána v literatuře pouze u Szurhaje a spol. (2003, 2005). K poslednímu lze dodat, že výsledky našeho výzkumu (nepublikovaná data, Obrázek č.9a-f) z elektrokortikografických dat nad rolandickou oblastí potvrzují existenci gamma desynchronizace v precentrální oblasti v motorickém paradigmatu (self-paced pohyb, Bereitschaftpotential paradigma). 27
28 ERD tedy může být interpretována jako elektrofyziologický korelát kortikálních oblastí aktivovaných ve zpracovávání sensorických či kognitivních procesů a/nebo motorického úkolu (Pfurtscheller a spol., 1992). Předpokládá se, že výraznější (výkonově) či rozsáhlejší (prostorově) ERD odrážejí zavzetí širší neuronální sítě nebo většího množství neuronů jedné sítě při zpracování informací Event-related synchronizace (ERS) ERS odráží synchronizované chování skupiny neuronů. Platí (Nuneze, 1995), že čím nižší je frekvenční pásmo, tím větší musí být počet koherentně se chovajících neuronů, aby změna byla registrovatelná. Např. v pásmu alfa je nezbytná synchronizace neuronů na ploše minimálně několika čtverečních centimetrů (Cooper a spol., 1965; da Silva, 1991), jak udávají výsledky analýz skalpových dat. Fenomén synchronizace, detekovatelný jako navýšení amplitudy resp. výkonu, je interpretačně mnohem složitější než fenomén synchronizace. Příčinou je fakt, že stav synchronizace může odrážet situaci funkční inhibice (vide supra spánkové grafoelementy) či naopak vysoce aktivní stav např. před provedením nebo během provádění určitého úkolu; také epileptický záchvat se svým EEG obrazem je výrazem intenzivní synchronizace. Nejranější studie přinášející popis synchronizace byly uveřejněny v 60. letech minulého století; bylo pozorováno navýšení amplitudy resp. výkonu v centrálních oblastech při vizuálním podnětu a naopak synchronizace v okcipitálních oblastech v okamžiku pohybu. Synchronizace tak je v těchto situacích chápána jako výraz klidového stavu (idling state). Stejná interpretace je pak aplikována na synchronizaci v alfa, ale hlavně v beta pásmu po ukončení volního pohybu (Pfurtscheller, 1992). Beta ERS nad centrálními či v centrálních oblastech je interpretována 28
29 jako shift motorických neuronů z aktivovaného do deaktivovaného stavu (Pfurtscheller a da Silva, 1999). Dle Cassima a spol (2000) je beta ERS také ovlivněná či přímo daná aferentním somestetickým impulsem. Důkazem podporujícím tuto teorii je magnetická encefalografie, kdy při stimulaci motorického kortexu v intervalu ERS po dokončení pohybu není možné získat žádnou motorickou odpověď, neboť kortex je ve stádiu aktivní funkční inhibice. Stejná pozorování jsou pak popisována v kognitivních parametrech - ERS v alfa pásmu je nahlížena jako obraz kognitivní inaktivity. Ovšem interpretace tohoto jevu je recentně nabízena i zcela opačně; Knyazev a spol. (2005) prokazují, že vyšší individuální výkon v alfa pásmu během klidového stavu měřeno v referenčním intervalu by mohl znamenat větší připravenost alfa systému ke zpracování informací. Na souboru pacientů vystavených různé míře anxiety prokazují, že připravenost systému vyjádřena coby vyšší ERS v referenčním intervalu a vyšší ERD v intervalu během úkolu koreluje s lepším výkonem v požadovaném paradigmatu. Výsledky analýz výkonových změn v pásmu theta jsou také kontroverzní, je popisována jak desynchronizace tak i synchronizace, nejčastěji je pak toto frekvenční pásmo analyzováno v kognitivních paradigmatech s důrazem na mnestické funkce. V temporálních oblastech je popisována synchronizace, jež je nahlížena jako substrát, na němž se odehrávají fyziologické pochody spojené s činností paměťových center. Oscilace v gamma pásmu, de facto vždy jen ve smyslu synchronizace, je interpretačně nejsložitějším, ale současně i nejzajímavějším jevem. První popis gamma oscilací se datuje ze 40. let (Adrian, 1942). Gamma pásmo je mnohými autory považováno za tzv. binding frekvenci, tedy oscilaci, jejíž funkcí je komunikace. Pro správné vykonání určitého úkolu, ať už z hlediska jeho včasnosti a kvality, je zapotřebí rychlá např. sensorimotorická integrace. Singer roku 1993 přichází s teorií, že dolní frekvenční pásma (alfa, beta) jsou příliš pomalá pro vyšší úroveň zpracování signálu ( higher level of processing ) a že právě ve frekvenci gamma, a to kolem 40 Hz, dochází k patřičnému, rychlému synchronizování mezi prostorově 29
30 vzdálenými skupinami neuronů. Naopak, Kopell přichází v roce 2000 s postojem, že gamma oscilace nejsou schopny (fyzikálně) se šířit na velké vzdálenosti a že předpokládaná funkce spjatá s přenosem informací bude na větší vzdálenosti zajišťována spíše beta oscilacemi. Gamma synchronizace - a v tom panuje konsensus - reflektuje stav aktivního zpracování informace, což je úplně v protikladu s chápáním alfa či beta synchronizace, které ale, uvažuje se, jsou nezbytným předpokladem pro možnost vzniku ERS v gamma pásmu. Úloha a lokalizace generátorů těchto rytmů zůstává i nadále značně nejasná. Dle Basar-Eroglua (1996) však budou sehrávat zásadní roli právě při kognitivních úlohách a při zpracování sensorické informace. Byly vysloveny další mnohé hypotézy týkající se funkčního korelátu gamma oscilací: předpokládá se, že gamma synchronizace odráží zaměřenou pozornost (Bouyer a spol., 1987), plánování pohybu (Donoghue a spol., 1998), eferentaci ke svalům (Salenius a spol, 1996) či sensorimotorickou integraci (Aoki a spol, 1999). Zdá se, že se jedná o elementární signály s mnohočetnými funkčními koreláty (Schurmann a spol, 1997). Pfurtscheller navrhnul před několika lety (1996), že beta synchronizace po pohybu může odrážet tzv. resetující mechanismus resp stav resetování motorického kortexu, stejný mechanismus byl jím navrhnut i pro gamma oscilace. Oba fenomény však byly pozorovány i v somatosensorickém kortexu a tudíž představa resetování v této oblasti, zvláště pak v gamma pásmu a zvláště v čase somatosensorické aferentace je málo pravděpodobné, že by primární somatosensorický kortex byl ve stavu funkčního klidu. Stejně tak ale gamma synchronizace v motorickém kortexu byla pozorována před i během pohybu, je tedy vysoce pravděpodobné, že nárůst výkonu v tomto frekvenčním spektru odráží jiné fyziologické pochody než stav tzv. funkční inhibice. Szurhaj a spol. ve své práci z roku 2005 spekulují, že tyto dva časové patterny ERS během a po pohybu odrážejí dva různé typy aferentace; proprioceptivní a povrchový kožní. Titíž autoři popisují gamma oscilace v okamžiku začátku pohybu v motorickém kortexu a předpokládají, že je v tomto případě odrazem efferent 30
31 support to the muscles. Navíc ale, primární motorický kortex vysoce pravděpodobně také dostává přímé aferentní impulsy (Geyer a spol., 1996; Balzamo a spol., 2004), Szurhaj a spol. z této skutečnosti poté vyvozují, že gamma ERS v M1 po ukončení pohybu může odrážet aferentní sensorickou zpětnou vazbu. Singerovu teorii autoři této práce opětně zakomponovávají do úvahy o tzv. binding oscilaci ve frekvenčním pásmu kolem 40 Hz, neboť kortikální representace např. ruky není jednou, koncentrovanou oblastí, nýbrž je spíše skupinkami od sebe i relatině vzdálených neuronů, jež mohou komunikovat různými způsoby a mezi nimi i pomocí gamma oscilací. Byť tato práce zmiňuje fakt, že gamma ERS je pozorována současně v M1 i v S1 kortexu, časová souvislost resp. posloupnost či synchronnost mezi M1 a S1 a potažmo i koherence, není v této práci zkoumána. Jak vidno, je problematika gamma oscilací nesmírně složitá a zatím velmi spekulativní a hypotetická Simultánní výskyt ERD a ERS Již klasická elektroencefalografie, ještě v dobách, kdy nebyla zavedena ERD/ERS kvantifikace, přinášela pozorování, že při otevření očí dochází ke snížení amplitudy nad okcipitálním kortexem zatímco amplituda v centrálních oblatsech zůstává stejná či se navyšuje (Berger 1930), a dále, že při volním či pasivním pohybu dochází ke snížení amplitudy centrálního mu rytmu zatímco v okcipitálních oblastech je amplitudový enhacement (Jasper a Penfield, 1938). Bylo tedy patrno, že oba fenomény se mohou vyskytovat simultánně a že budou odrážet různý funkční stav jednotlivých kortikálních oblastí. Stejný fenomén jako výše popsané antagonistické chování, současný výskyt nárůstu a poklesu výkonu odrážející klidový a aktivní stav různých korových oblastí specializovaných pro jiné modality (rolandický versus okcipitální kortex), lze pozorovat i uvnitř jedné oblasti. 31
32 Při pohybu rukou dochází k alfa a beta desynchronizaci nad motorickou oblastí pro ruku a naopak k synchronizaci nad oblastí pro nohu a vice versa (Pfurtscheller a Neuper, 1994). Funkčním významem indikovaných oscilací v alfa pásmu se komplexně zabývá Pfurtscheller ve své práci z roku Resumuje, že fenomén 10 Hz ERD může být v jiné kortikální lokalizaci doprovázen 10 Hz ERS, nazývá tento obraz pak focal ERD/surround ERS. Tyto výsledky analýz skalpových jsou ale jen obtížně doložitelné výsledky snímání oscilačních změn direktně v inkriminovaných oblastech. I při provedení in situ MRI s vnořenými elektrodami víme poměrně málo o funkčním uspořádáním např. motorické oblasti, nejsou-li provedena současně také funkční vyšetření (fmri, PET, SPECT). Topizace např. motorické oblasti např. pro ruku a pro nohu je elektrofyziologicky de facto umožněna pouze pomocí evokovaných potenciálů či pomocí přímé bipolární stimulace kontaktů vnořených elektrod resp. subdurálních gridů či stripů. Indikace k těmto vyšetřením je vysoce specifická a zřídkavá. Jinými slovy do dnešního dne neexistuje studie, jež by přinášela výsledky analýzy oscilačních změn primárního sensomotorického kortexu získaných přímou, invazivní metodou za podmínky dostatečně přesné topizace oblasti pro ruku a pro nohu. Naše výsledky z ojedinělé kazuistiky (nepublikovaná data, Obrázek č.9) u pacienta s gridem nad sensorimotorickou oblastí, jenž prováděl volní, self-paced pohyb jak rukou, tak i nohou, ipsi- i kontralaterálně ke straně implantace subdurálního gridu a u nějž lokalizace elektrod gridu byla identifikována stimulací, jsou následující: alfa i beta ERD jsou somatotopické dle Penfieldova homunkula, oba fenomény lze pozorovat pre- i postcentrálně, jsou přítomny při ipsi- i kontralaterálním pohybu, ale kontralaterální pohyb vyvolává resp. je odrážen procentuálně i lokalizačně rozsáhlejší a časnější oscilatorními změnami. V premotorické oblasti pak byla pozorována beta ERS nastupující již v době před započetím pohybu a to v místě, kde byl zaznamenán i pomalý DC shift přípravný motorický potenciál (motor readiness potential, Bereitschaftspotential). 32
33 V neposlední řadě lze současný výskyt různých fenoménů pozorovat v oblasti jednoho kontaktu elektrody (skalpové, ECoG, SEEG elektrody). Kontakt umístěný např. nad primární motorickou oblastí vykazuje v období těsně před, během a těsně po skončení pohybu (rozmezí +/- 2 sekundy) ERD/ERS ve třech frekvenčních pásmech; alfa ERD, beta ERD/ERS a gamma ERS. K sychronizaci v gamma pásmu může docházet krátce před pohybem. Míra amplitudové změny, měřeno absolutně, je pak různá; největší v alfa pásmu a nejmenší v pásmu gamma. Důležité je však pochopit, že snížení míry synchronie v jednom frekvenčním pásmu umožňuje vystoupení do popředí a i event. enhancement, jiné frekvence. Efekt změn oscilatorních aktivit je potom odvislý na frekvečních pásmech, ve kterých se odehrává, na lokalizaci oscilátoru, na časovém vztahu např. k podnětu. Proto může synchronizace být jednou například odrazem stavu funkční inhibice (beta ERS po ukončení pohybu), jindy naopak substrátem probíhajících mnestických procesů (theta ERS v hipokampu) či substrátem epileptického záchvatu (synchronizace v beta pásmu a výše). Z výše řečeného též vyplývá, že určující kvalitou je normalita neuronální sítě. V případě patologických stavů, jdoucích od molekulární po globální anatomickou úroveň, může být fungování neuronální sítě zcela změněno. Příkladem jsou např. degenerativní procesy se ztrátou specializovaných skupin neuronů, nebo epilepsie s abnormální synchronní aktivitou neuronů sítě. 3. Bazální a aplikovaný výzkum 3.1. Elektrofyziologie pohybu a kortex Motorika, volní či pasivní, spontánní či řízená s dalšími a dalšími možnými obměnami zaujímala v elektrofyziologickém výzkumu vždy dominantní postavení. V čase desetiletích 33
34 probíhalo bádání, jak následující v krátkosti popsané mezníky nastiňují, spíše po dvou liniích - sledovaly se evokované změny získávané zpětným zprůměrněním EEG dat podle trigeru a analyzovaly se změny týkající se pozadí, EEG šumu, oscilací. Jen ojediněle a spíše recentně byla stejná EEG data podrobena oběma analýzám. Již v roce 1938 Jasper a Andrew popisují reaktivitu centrálního mu rytmu po taktilním podnětu. Jasper s Penfieldem v roce 1949 demonstrují supresi centrální beta aktivity po volním pohybu a to na datech získaných elektrokortikograficky. Gastaut roku 1952 a Chatrian roku 1959 publikují údaje o supresi mu rytmu při kontralaterálním volním či i pasivním pohybu či i somatosensorické stimulaci (Obrázek č. 8). Obrázek č. 8 (použito z Pfurtschellera) Kornhuber a Deecke popisují roku 1965 na zprůměrněných datech opakovaného volního pohybu tzv. přípravný motorický potenciál (RP readinnes potential, BP - 34
35 Bereitschaftspotential) coby pozvolný DC posun před pohybem, nález potvrzují později Shibasaki (1980), Neshige (1988) a další. Přípravný motorický potenciál (RP, BP) je pozorován bilaterálně, symetricky nad centrálními oblastmi se začátkem cca 1,5-3 s před pohybem, jenž v čase velmi krátce před pohybem (0,5 s) nabývá kontralaterálního zvýraznění ve formě tzv. NS (negative slope, viz Obrázek č. 2). Roku 1994 Toro a spolupracovníci popisují další složku potenciálu, jenž doprovází již vlastní pohyb frontal peak of the motor potential fpmp, jenž byl také pojmenován movement accompanying potential roku 1998 Rektorem. Jiným paradigmatem při zkoumání motoriky je paradigma pozor, připrav se a jdi nebo nejdi (CNV - contingent negative variation či jeho obměna go/no-go paradigma) zohledňující fenomén anticipace (Obrázek č. 1). Problematika oscilací je kupodivu až do 70. let tedy upozaděna. Aby bylo tedy možné lépe popisovat, co se děje s oscilačním šumem na pozadí, bylo nutné ho v prvém kroku kvantifikovat. Prvenství v kvantifikaci oscilačních změn souvisejících s motorikou pak náleží od roku 1977 Pfurtschellerovi a Aranibarovi. Teprve od tohoto roku, po zavedení výpočtu kvantifikace amplitudových respektive výkonových změn ve zvolených frekvenčních pásmech, šlo pozorovat, že mu rytmus (čili alfa pásmo) a beta rytmus reagují jinak; jednak časově a jednak lokalizačně. Alfa desynchronizace nastává cca 2 s před pohybem, zatímco beta teprve cca 1,5 s, obojí nad kontralaterální centrální oblastí, a teprve v okamžiku započetí pohybu je desynchronizace patrná i nad ipsilaterální centrální oblastí. Průběh i během pohybu a těsně po pohybu je jiný; alfa rytmus po své maximální desynchronizaci během pohybu se pozvolna vrací k k úrovni amplitudy resp. výkonu v klidovém stavu, kdežto beta rytmus ihned po ukončení pohybu vykazuje často i vysokou míru synchronizace. Ze skalpových záznamů, pomocí matematického modelování, se předpokládalo, že za generování změn v beta pásmu je odpovědna precentrální oblast a za změny v alfa pásmu oblast poscentrální. Beta desynchronizace byla opakovaně popisována taktéž nad středním vertexem, v oblasti, která 35
36 dle systému uložení skalpových elektrod odpovídá supplementární motorické oblasti. Ze skalpového EEG také jako první a prozatím jediný popisuje Pfurtscheller a Neuper (1992) gamma oscilace kolem 40Hz nad k pohybu kontralaterální centrální oblastí v čase kolem jeho začátku. Jednoznačným důvodem zřídkavosti popisu tohoto fenoménu z EEG dat získaných snímaním ze skalpu jsou fyzikální vlastnosti jak gamma oscilací (relativně malá amplitudová změna a extrémně vysoká fokusace), tak i filtračních funkcí struktur, jež dělí snímací elektrodu od mozkového kortexu. Souhrnně, dle Pfurtschellera (2003) lze na skalpu, v oblasti primárního sensorimotorického kortexu, standardně rozlišovat následující oscilační změny: a/ kontralaterálně dominantní alfa a beta ERD před pohybem b/ oboustranně symetrická alfa a beta ERD během pohybu c/ kontralaterálně dominantní beta ERS v první sekundě po ukončení pohybu. Již z těchto údajů o indukovaných (ERD/ERS) a evokovaných (BP) změnách lze uvažovat o různém elektrofyziologickém významu obou fenoménů. Tyto údaje z analýz skalpových dat o časové a lokalizační rozdílnosti ale nejsou vždy zcela kongruentní s výsledky intrakraniálních registrací. Jak tedy vypadají výsledky dat získaných intrakraniálně, ať už subdurální či intracerebrální registrací? Světové písemnictví nabízí pouze nečetné publikace, jež se zabývají danou problematiku ve více či méně modifikovaných paradigmatech a jež zkoumají buď jen indukované změny (ERD/ERS) či i změny evokované (BP, CNV). Analýzu elektrokortikografických dat přináší Arroyo (1993), Toro (1994), Crone (1998), Aoki (1999), Ohara (2000), a Pfurtscheller (2003). Intracerebrální registrace je ještě vzácnější; kromě kolegů z francouzského Lille (Szurhaj a spol a 2005) se jedná pouze o brněnské pracoviště pod vedením profesora Rektora. Prvně, intracerebrální data nepřinášejí závěry tak striktní jako skalpové studie. Skutečností je, že skalpová data jsou získávána registrováním zdravých dobrovolníků, kdežto data intrakraniální jsou obdržena během preoperační invazivní monitorace pacientů 36
37 s farmakorezistentní epilepsií (velmi sporadicky se jedná o jiný důvod implantace intrakraniálních kortikálních elektrod), u nichž může být fungování neuronálních sítí modifikováno mnoho let trvající patologickou epileptickou aktivitou. Všechna tato fakta jsou ale brána při interpretací výsledků v potaz a lze i tak postulovat některé závěry a hypotézy. Intrakraniální registrace naopak využívá různé přednosti, jichž není možné docílit u skalpového EEG (blíže vide supra v této kapitole). Souhrnně pak lze konstatovat, že intrakraniální výzkum, včetně našeho, se zaměřil několika směry: a/ je tzv. suplementární motorická oblast suplementární či supramotorickou? Jaká je časová posloupnost mezi aktivací SMA, M1 a S1, případně dalších oblastí? b/ jsou při volním (BP paradigma) či řízeném pohybu (CNV paradigma) zavzaty v činnost jen oblasti motorické či i extramotorické? Je rozdíl v distribuci evokovaných a indukovaných změn? c/ jaká je distribuce gamma oscilací, jejich lokalizace a časový výskyt v souvislosti s pohybem, jejich funkční význam? d/ je centrální rýha skutečně dělící strukturou mezi alfa a beta oscilacemi? Otázek je samozřejmě více, odpovědi na výše uvedené jsou rozebírány v následujícím textu. Předně - nezdá se, že by centrální rýha byla dělící čárou mezí oběma rytmy, ale výsledky studií se liší. Pfurtscheller popisuje beta ERD nejen precentrálně, ale i postcentrálně, což je právě v kontradikci se zažitým schématem a s výsledky studie MEG (Salmelin a Hari, 1994), v níž byla za generátor mu rythmu označena primární somatosensorická area a beta rytmu primární motorická area. Alfa ERD je autorem pozorována nad posteriorním parietálním kortexem, nad inferiorním parietálním lalokem. SMA vykazovala stejné změny jako nálezy Ohary z r ve smyslu beta ERD. Beta ERS u kolegů z Rakouska byla zaznamenána výraznější a navíc pozorovali gamma ERS. Skutečně zajímavým výsledkem, 37
38 jenž koreluje s nálezem Croneho (1998) a Arroya (1993), je gamma ERS v roszáhlém pásmu Hz nad M1/S1. Obdobný je pak nález Croneho gamma ERS Hz nad M1/S1 s lokalizačně velmi striktním, somatotopickým nálezem. Existenci takto širokého pásma, jež současně reaguje ve smyslu gamma ERS pak Pfurtscheller vysvětluje následovně: předpokládá, že přeci jen subdurálně uložený kontakt elektrody představuje sumaci aktivity poměrně rozsáhlého množství kortikálních modulů resp. skupin neuronů, což může znamenat, že každý modul může oscilovat na jiné frekvenci, souhrnná aktivita se pak promítá do širšího frekvenčního rozpětí. Kolegové z Francie (Szurhaj a spol.) se v práci z roku 2003 soustředili na výzkum v sensorimotorických oblastech (gyrus precentralis, gyrus postcentralis, frontální laterální a frontální mesiální kortex) u 3 pacientů. Kontrolní analýza skalpových dat u všech pacientů korelovala přibližně se všeobecným postulátem o průběhu desynchronizace; výsledky však byly nehomogenní. Změny byly pozorovány v alfa a beta pásmu, gamma oscilace Hz byly beze změny. Intracerebrálně alfa a beta pásmo desynchronizovalo nejprve v precentrální oblasti, následně pak v oblasti postcentrálního gyru; synchronizace v beta pásmu byla pozorována jak pre- tak i postcentrálně; v gamma pásmu byla pozorována desychronizace (jev v literatuře prvně popisovaný), jak v precentrální, tak u jiného subjektu i v postcentrální oblasti, pokles byl poměrně malý, okolo 20 % a nastal těsně před pohybem či v okamžiku pohybu. V každém případě je ale nutno poukázat na fakt, že interindividuální rozdíly byly velké; latence ERD varírovaly mezi v romezí cca 1,5 s (stejné nálezy byly pak popisovány Pfurtschellerem na skalpu ( 2250 do 750 ms), Oharou (-2700 do 400 ms)). V roce 2005 popisují Szurhaj a spol. gamma ERS pozorována nejen v postcentrální ale i precentrální oblasti (studované frekvenční pásmo Hz). ERS mělo v M1/S1 následující charakter: začátek byl buď spolu v okamžiku začátku pohybu a ERS přetrvávalo i po jeho ukončení, nebo ERS začala po skončení pohybu. 38
39 Japonští kolegové u 4 pacientů zkoumali reaktivitu oscilací a výskyt Bereitschaftspotential. Data byla snímána elektrokortikograficky a volní pohyb byl prováděn kontralaterálně i ipsilaterálně k uložení elektrody. Výsledky ukázaly byť je nezbytné uvážit malý počet subjektů opět poměrně velkou variabilitu nálezů; BP začínal buď dříve v SMA než v M1/S1, nebo nebylo rozdílu. Stejně nad SMA varírovala i lateralizace; začátek BP byl dříve buď s kontralaterálním pohybem, nebo nebylo stranového rozdílu, nebo začínal dříve i ipsilaterálně. BP nad M1/S1 pak začínal, pokud byl hodnocen k oběma stranám pohybu, kontralaterálně dříve. ERD v beta pásmu bylo pozorováno vždy ponejprve nad SMA, teprve poté desynchronizovala primární motorická a primární somatosensitivní area. ERS v beta a gamma pásmu byla nad M1/S1 pozorována striktně jen u kontralaterálního pohybu, nad SMA pak byla nekonstatní a pokud, tak se vyskytovala u ipsi- i kontralaterálního pohybu. Výsledek tedy vysoce nehomogenní, nicméně prokazující mnoho věcí; jednak to, že ipsilaterálně k pohybu dochází k desynchronizaci již před pohybem (což nekoreluje se zažitým dogmatem z EEG skalpového, že začátek ipsilaterální desynchronizace je až se začátkem pohybu), dále z této práce pro autory vyplývá, že suplementární motorická area hraje úlohu spíše supramotorickou vzhledem k tomu, že jak přípravný motorický potenciál (BP), tak i oscilační změny jsou většinou nejprve pozorovány nad SMA a teprve později nad M1/S1. Takto jednoznačný závěr přeci jen je zatím v literatuře osamělý, Szurhaj a spol. a ani naše práce neprokazují (SEEG data) tak jasnou časovou posloupnost aktivace. V naší práci ( Intracerebral recording of cortical activity related to self paced voluntary movements: A Bereitschaftspotential and Event-related de-/synchronization. A SEEG study. akceptováno k publikaci) jsme jednak, jak bylo výše zmíněno, neprokázali primární aktivaci SMA před M1/S1 (nízký počet subjektů, nehomogenní data, podrobněji viz Příloha, Publikace), ale naopak jsme zkonstatovali, že indukované změny (ERD/ERS) jsou mnohem rozsáhlejší, lokalizačně, než změny evokované (EP), které jejichž výskyt je omezen na oblast M1/S1, 39
40 SMA, méně již cingulární, premotorickou a dorzolaterální prefrontální oblast, zatímco oscilační změny (alfa a beta pásmo) jsou v BP paradigmatu pozorovány i v laterálním temporálním neokortexu, v meziálních temporálních strukturách, v orbitofrontálním kortexu a v oblasti lobulus parietalis inferior. Tyto oscilační změny jsou pak přítomny jak při kontralaterálním, tak i při ipsilaterálním pohybu. Vyjadřují, dle naší interpretace, kognitivní procesy spjaté s volním pohybem v BP paradigmatu jako jsou pozornost, paměť a odhad časového intervalu. Lze tedy říct, že dosud provedené intrakraniální studie nebyly schopny přinést kongruentní výsledek týkající se posloupnosti aktivace SMA a M1/S1. Dále je možné zkonstatovat, že somatotopika a oscilace korelují lépe ve vyšších frekvenčních pásmech (beta a gamma) než v nižším alfa pásmu, jež bude pravděpodobně jen odrážet kortikální aktivaci (Steriade a Llinas, 1988). Autoři z Lille (Szurhaj a spol., 2003, 2005) i na základě studie Cassima z roku 2004 dále předpokládají, že ERS nemusí vždy nutně znamenat funkčně klidový stav; tedy i naopak, že funkční areaktivita nemusí být reflektována vždy formou ERS. Své tvrzení dokládají pozorováním, že beta ERS lze pozorovat i během pohybu a stejně jako gamma ERS může plnit úlohu při zpracovávání somestetických aferentací. V závěru práce konstatují, že reaktivita mu a beta rytmů je rozdílná, ale jejich lokalizace se překrývá; lze tedy předpokládat, že odrážejí různé mechanismy aktivace. Autoři také navrhují, že SEEG analýza kortikálních rytmů by mohla doplnit funkční mapování sensorimotorických oblastí u pacientů před plánovaným neurochirurgickým výkonem. Relativně, ale skutečně jen relativně na okraji zájmu stojí dvě níže uvedené práce. Obě se totiž snaží odpovědět na jednu z nejzákladnějších otázek elektrofyziologie: proč při unilaterálních pohybu dochází k aktivaci i ipsilaterální centrální oblasti, aniž by byl pozorován zrcadlový pohyb? Co tedy odráží a jaký je funkční význam ipsilaterální aktivace? Stančák a spolupracovníci se zabývali hledáním souvislosti mezi velikostí corpus callosum a 40
41 rozsahem ERD/ERS v oblasti sensomotorických oblastí. Analýza dat získaných ze skalpu, ale s hustou densitou rozmístěných elektrod (82 elektrod), v paradigmatu repetitivních kutánních stimulací v nepravidelných intervalech pak přinesla tyto výsledky: alfa a beta ERD byla pozorována oboustranně nad S1/M1, post-stimulus beta ERD byla opět pozorována oboustranně, frontálně. Dále byla pozorována habituace na aplikované podněty vyjádřena poklesem desynchronizace. A taktéž - hodnota ipsilaterální synchronizace v beta pásmu korelovala pozitivně s velikostí střední části corpus callosum. Tento poslední poznatek, přinášející další elektrofyziologický důkaz interhemisferální komunikace, je velmi zajímavý a může mít praktický dopad i pro epileptologii. Studii, obdobnou, řekněme oscilačních změn a vzájemné hemisferální souhry, u pacientů s nebo bez epilepsie a s agenezí corpus callosum, však dle mé obeznámenosti s literaturou nikdo zatím neučinil (Stancak a spol., 2003). Rau roku 2003 si klade otázku, jak reaguje ipsilaterální primární motorický kortex během volní fázické extense prstů HKK. Paradigma bylo ale významně obohaceno o aplikaci magnetické stimulace kortexu (TMS) v intervalu ms od počátku pohybu. Výsledek studie pak je následující: ipsilaterální ERD během pohybu je spojena se zvýšenou excitabilitou a se zvýšenou intrakortikální facilitací v intervalu ms. Během ERS po dokončení pohybu nebyly pozorovány žádné změny v kortikální excitabilitě. Autoři z těchto nálezů usuzují na aktivní podíl ipsilaterálního motorického kortexu na kontrole pohybu Elektrofyziologie pohybu a subkortikální struktury Podíl subkortikálních struktur, zvláště pak bazálních ganglií na přípravě pohybu byl, na rozdíl od kortexu, studován elektrofyziologicky již méně. Data jsou získávána teprve recentně, díky terapeutické metodě DBS (deep brain stimulation) u extrapyramidových 41
42 onemocnění, speciálně pak u Parkinsonovy nemoci. Většina současného elektrofyziologického výzkumu je zaměřená na určování patologických a fyziologických typů oscilací v jednotlivých subkortikálních strukturách, zkoumání jejich klinického korelování ať už s parkinsonským či dyskinetickým syndromem a samozřejmě směřování k terapeutickému výstupu implantování stimulátorů pracujích na kýžené frekvenci, jak již se dnes v několika světových centrech děje. V níže uvedeném přehledu jsou zmíněny hlavně práce, jež poukazují právě na přítomnost různých typů oscilací. Prací, zabývajících se současným výskytem oscilačních změn a evokovaných změn (EP v širokém slova smyslu, CNV, BP) je již výrazně méně, brněnské pracoviště je v tomto ohledu téměř ojedinělé a přispívá tak výraznou měrou do studnice vědomostí zabývající se fyziologií subkortikálních struktur a vzájemnou interakcí kortiko-subkortikální. Paradiso a spol. (2004) hodnotili jak oscilační změny, tak i pomalý DC posun odpovídající Bereitschaftspotentialu v thalamu (6 pacientů s tremorem, 1 pacient myoklonusdystonie). Registrovali BP u 5 ze 7 subjektů v thalamu, s latencí kolem 2 sec, jež odpovídala latenci na skalpu, pouze ale u aktivního pohybu, pohyb pasivní nebyl doprovázen žádným BP. Fázový zvrat či maximum amplitudové bylo pak pozorováno ve ventrálním laterálním jádru. Frekvenční analýza získaných dat (5 subjektů) ukázala výkonový pokles před pohybem v alfa i beta pásmu (beta ERD v thalamu i na skalpu kolem 2,5 s před pohybem, alfa ERD v thalamu až kolem 1 s před pohybem v datech tzv grand average). Dále, beta pásmo vykazovalo thalamo-kortikální koherenci až do okamžiku cca 0,5 s před začátkem pohybu, naopak v alfa pásmu nebyla koherentní aktivita pozorována. Vzhledem k uložení skalpové elektrody (FCz) pak autoři vyvozují, že suplementární motorická area a motorický thalamus interagují během přípravy pohybu hlavně v pásmu beta. O gamma pásmu se práce nezmiňuje. Kuhn roku 2004 publikoval práci na téma oscilačních změn v nucleus subthalamicus v go/no-go paradigmatu u 8 pacientů s Parkinsonovou chorobou. V paradigmatu go 42
43 referovali o poklesu výkonu v beta pásmu kolem 20 Hz před pohybem, přičemž začátek desychronizace silně pozitivně koreloval s reakčním časem. Po skočení pohybu pak byla pozorována synchronizace v daném frekvenčním pásmu. Změny spjaté s no-go situací vykazovaly předčasné ukončení beta desynchronizace po v tomto případě no-go stimulu a časnou beta synchronizaci. Subthalamické jádro a stupeň jeho synchronního chování v beta pásmu tak byl v této práci považován za významnou determinantu při rozhodování, zda pohyb bude nadále připravován a event. i započat. Neurofyziologové z Queen Square (Doyle a spol., 2005) studovali vliv podání levodopy na oscilační aktivitu nucleus subthalamicus. Vycházeli z hypotézy, že zvýšená míra synchronizace v tomto jádru, jež je významnou součástí okruhu v bazálních gangliích, může přispívat ke zpomalení motorické inicace u idiopatické Parkinsonovy choroby. U 14 pacientů pak v paradigmatu volního pohybu (BP paradigma) pozorovali zvýšení ERD a prodloužení jeho trvání po aplikaci levodopy oproti situaci bez administrace léku. Brown v roce 2002 uveřejňuje studii s výsledky sledování oscilačních změn u pacientů s Parkinsonovou chorobou a s implantovanými elektodami v subthalamickém jádru a/nebo v globus pallidum internum (6 pacientů). V klidovém stavu, bez pohybu, dominovala v situaci off aktivita pod 5 Hz a dále spektrálním maximem na 20 Hz. V klidovém stavu, ale s dopaminergní medikací, bylo naopak spektrální maximum na 70 Hz. Další výzkum se týkal paradigmatu CNV, ve dvou situacích: ve stavu off a při dopaminergní stimulaci. Ve stavu off docházelo jak po prvním, přípravném, varujícím stimulu, tak i po imperativním k supresi aktivity na 20 Hz. Naopak, pakliže při medikaci byla klidová oscilace s maximem na 70 Hz, při pohybu, po imperativním stimulu, docházelo k nárůstu výkonu, k synchronizaci. Zajímavá a důležitá také byla otázka tzv. coupling s GPi a s kortexem studovaná pomocí frekvenčních koherencí. Výsledkem pak byl nález stejných změn, jaké se odehrávaly v subthalamickém jádru. Brown ve studii zmiňuje Croneho, který roku 1998 publikoval nález 43
44 gamma oscilací stejné frekvence z oblasti primárního sensorimotorického kortexu. Recentně pak byly publikovány další práce zmiňující obdobné nálezy ať již z dat získaných intracerbrální (Szurhaj a spol., 2003; Szurhaj a spol., 2005) či subdurální registrací (Pfurtscheller a spol., 2003). Chybění těchto oscilací a naopak maximum na 20 Hz pak Brown považuje za elektrofyziologický aspekt patologie tohoto neurodegenerativního onemocnění. Levy ve stejném roce jako Brown, tedy 2002, uveřejňuje studii ze subthalamického jádra u 15 pacientů s idiopatickou Parkinsonovou chorobou, frekvenční analýzu provádí do 55 Hz a výsledkem je závěr o přítomnosti patologické oscilace v off stavu ve frekvenčním pásmu Hz. Nesmírně zajímavá a stále nedořešená je otázka, jakým způsobem probíhá spolupráce mezi bazálními ganglii a kortexem. Elektrofyziologie přináší na tomto poli nenahraditelnou, k funkčně zobrazovacím metodám (PET, SPECT, fmri) komplementární informaci. Nezastupitelnost elektrofyziologie stojí na lepším zachycení časového průběhu změn, nevýhodou naopak je horší prostorová rezoluce. Většina studií je založena na analýze skalpových dat, matematické modelování je pak vždy jen nutně aproximací reálné lokalizace hledaných intracerebrálních generátorů evokovaných či indukovaných odpovědí. Data získaná přímo intracerebrální registrací jsou samozřejmě lokalizačně velmi přesná, nicméně dávají informaci o elektrofyziologických pochodech jen ve velmi restriktovaném vzorku mozkové tkáně a tudíž také neumožňují žádanou prostorovou představu o pochodech v neuronálních sítích. Brněnské pracoviště v čele s profesorem Rektorem se pokouší opakovaně dát alespoň parciální odpovědi na výše uvedené otázky a to jak na poli elektrofyziologickém, tak i v posledních letech na poli zobrazovacím. Tak v rozsáhlé práci Cognitive potentials in the basal ganglia-frontocortical circuits. An intracerebral study. (Rektor a spol., 2004) je analyzován výskyt kognitivních potenciálů (P300) resp. výskyt s kognitivními pochody 44
45 souvisejících potenciálů (BP, CNV) současně v bazálních gangliích a v motorickém i nemotorickém kortexu. Přítomnost generátorů (registrace přímo v generátoru odpovídá fázovému zvratu mezi dvěma kontakty vnořené elektrody, registrace v přímé blízkosti blízkosti generátoru odpovídá amplitudivému gradientu na sousedních kontaktech vnořené elektrody) všech potenciálů pak byla prokázána v bazálních ganglich, v kortexu byl kognitivní P300 potenciál registrován v nejrůznějších lokalizacích, výskyt CNV a BP byl výrazně omezen na centrální kortikální arey. Časově pak výskyt P300 potenciálu u všech volených paradigmat (auditivní a vizuální oddball paradigma a auditivní a vizuální CNV paradigma) kromě vizuální P300 vykazoval statisticky významně kratší latence u generátorů v bazálních gangliích než u generátorů kortikálních. Dnes již v jistém slova smyslu historický Alexanderův model (1986) motorické kortiko-subkortikální kličky při provádění motorického úkonu je výsledkem této studie podporován, výskyt s pohybem souvisejících kognitivních aktivit monitorovaných podle výskytu pomalých potenciálů (BP, CNV) je výrazně vymezen na oblast pre- a motorického kortexu a bazálních ganglií. Takováto segregace funkce se naopak zdá málo pravděpodobná při zpracovávání sensorických informací (P300 paradigma, vlna P300 v rámci CNV paradigmatu). Generátory vlny P300 jsou v kortikálních strukturách distribuovány velmi hustě a široce, také v bazálních gangliích (ve studii explorován hlavně putamen) není distribuce interindividuálně a i intraindividuálně jednotná. Představa jednoduché kličky, jednoduché neuronální sítě se tak zdá nepravděpodobná. Parent a Cicchetti v roce 1998 uvažují spíše o vnitřní, na konkrétní dané úloze odvisející organizaci systému bazální ganglia kortex. Zdá se tedy, že bazální ganglia, hlavně striatum, mohou plnit integrující úlohu při zpracovávání kognitivní informace a to jak v motorické i nemotorické úloze. Tento závěr podporuje i naše další práce, uveřejněná roku 2003 (Sochůrková and Rektor, 2003). Při předoperačním explorování pacientů s farmakorezistentní epilepsií mohou být ojediněle, díky trajektorii implantovaných hlubokých elektrod, některé 45
46 kontakty umístěny v bazálních gangliích. Studovali jsme takto, kazuisticky, současný výskyt oscilačních změn a přípravného motorického potenciálu v BP paradigmatu. Fenomén výrazné alfa ERD a beta ERD/ERS byl pozorován v putamen (trajektorie hluboké elektrody přes bazální ganglia) jak při volním kontralaterálním, tak ale i ipsilaterálním pohybu akrem horní končetiny, synchronizace po ukončení pohybu byla výraznější ipsilaterálně. Naopak, BP potenciál nebyl v tomto místě putamen vůbec registrován. I toto pozorování oscilačních změn je tedy, zdá se, dalším potvrzením teorie paralelní kortiko-subkortiko-kortikální aktivace (Alexander 1994). Dalším z příkladů klinické aplikace, respektive výzkumu změn resonantního chování mozku u jasné klinické diagnozy přinášejí Defebvre a jeho spolupracovníci již roku Parkinsonova choroba je onemocněním primárně subkortikálním, avšak s nezbytným dopadem na kortiko-subkortikální okruhy, jejichž alterace se projevuje známou klinickou sémiologií. Cílem studie bylo zjištění, jak toto neurodegenerativní onemocnění pozměňuje elektrofyziologické parametry u volního pohybu. Přípravný motorický potenciál (BP) byl poprvé popsán v roce 1964 Kornhuberem a Deeckem. Objevuje se, na skalpových datech, jako pomalý negativní potenciál začínající oboustranně a symetricky nad centrálními oblastmi (Shibasaki a spol., 1980) 1-1,5 s před pohybem, aby asi 0,5 s před započetím pohybu vykazoval kontralaterální preponderanci. Na skalpových datech je pak popisována nejvyšší amplituda nad vertexem a předpoládá se, že odráží aktivitu suplementární motorické oblasti, jež je zapojena do programování pohybu přes thalamo-kortikální okruh. Studie zabývající se alterací BP u Parkinsonovy choroby jsou víceré, závěry nejsou však jednotné. Deecke (1977) a Shibasaki (1978) pozorovali snížení amplitudy BP nad k akinetické straně kontralaterálním centrální kortexem, Barrett (1986) naopak žádný rozdíl mezi skupinou starších zdravých osob a pacientů s IPD neviděl. Defebvre (1994) přichází se stejným výsledkem jako posledně jmenovaný. Oběma posledně jmenovaným studiím by se však dalo vytknout, že studovali 46
47 výskyt BP u pacientů léčených L-dopa. Ovšem zcela odlišně pak vypadala analýza ERD/ERS; desynchronizace nastávala u skupiny subjektů s onemocněním IPD signifikantně později (1250 ms oproti 1750 ms u zdravé skupiny) a byla patrna ipsilaterálně již 0,5 s před pohybem zatímco normálně se objevuje v této oblasti až od začátku pohybu. Defebre uzavírá, že s může jednat o kompenzatorní aktivitu jak ipsilaterálního motorického kortexu, tak vzhledem k relativně vyššímu potenciálu BP frontocentrálně i SMA. Jelikož desynchronizace je interpretována jako indikátor kortikální aktivace (Pfurtscheller a Berghold, 1989), jeví se alespoň v této elektrofyziologické studii sledování změn indukovaných (ERD/ERS) přínosnější než sledování změn evokovaných (BP) Oscilace v epileptologii Difúzní oploštění EEG křivky a nástup nízkovoltážní vysokofrekvenční aktivity je již standardně uznávaným elektroencefalografikcým počátkem epileptického záchvatu. Pojem desynchronizace je implicitně běžně používán. Desynchronizací se v tomto případě rozumí tak, jak odpovídá pojmu viditelné snížení amplitudy EEG při běžném rozpětí frekvenčních filtrů. Teprve ale při bližší analýze je patrné, že daná oblast sice neosciluje na klidové frekvenci, ale že přešla na modus oscilování ve vyšších frekvenčních pásmech. Tyto oscilace se pohybují, jak ukazují recentní výzkumy využívající možnosti téměř neomezeného horního frekvenčního filtru, nejen minimálně na hranici beta-gamma pásma, ale mnohem výše. V terminologii se pak objevují další pojmenování frekvenčních pásem: dolní gamma (30-50 Hz), horní gamma ( Hz), ripples ( Hz, Buszaki a spol., 1992), fast ripples ( Hz, Bragin a spol. 1999). Světové písemnictví nabízí výsledky několika studií, jež si kladou otázku, jaký je význam těchto vysokofrekvenčních oscilací v epileptologii. Fisher a 47
48 spol. v roce 1992 studovali frekvenční komponenty do 150 Hz u pěti pacientů s implantovanými subdurálními gridy v rámci předoperační přípravy léčby epilepsie. Elektrodekrementální vzorec na začátku záchvatu vykázal dvojnásobný nárůst výkonu ve spektru Hz a ve spektru Hz dokonce pětinásobek výkonu. Autoři předpokládali, že aktivita nad 40 Hz představuje sumaci akčních potenciálů, harmonické frekvence synaptických potenciálů nebo transientní ostré komponenty synaptických potenciálů. Dále, nález této aktivity nebyl pozorován jinde než v oblastech kontaktů odpovídající zoně počátku záchvatů. Nabízí se tedy využití zkoumání těchto vysokofrekvenčních oscilací v klinické epileptologii. Skupina neurofyziologů z Los Angeles (Bragin a spol., 1999) prohlubuje roku 1999 výše uvedené závěry. Ripples jsou nacházeny v CA1 a v entorhinálním kortexu u normálních laboratorních krys, ale i u krysích modelů mesiotemporální epilepsie (kindling, kainát) a u pacientů s MTLE, kteří byli vyšetřování pomocí hlubokých elektrod. Tato aktivita pravděpodobně reprezentuje aktivitu inhibičních interneuronů (IPSP). Intermitentní oscilace v ještě vyšším frekvenčním pásmu ( Hz, fast ripples ) byly ale pozorovány jen u laboratorních zvířat po aplikaci kainátu a u pacientů s epilepsií. Navíc, další analýza hovořila pro fakt, že fast ripples jsou generovány hlavními pyramidovými buňkami a ne interneurony a že tato aktivita může přispívat k synchronním výbojům u pacientů s epilepsií. Etiopatogenetické podklady mohou být následující: sprouting mechových vláken se synaptickou reorganizací, tvorba gap juntions mezi lokálními clustery neuronů s volumeconducted elektrickými proudy. Všechny tyto jevy se hypoteticky mohou podílet na hypersynchronii u epilepsie a mohou být histopatogenetickým podkladem pro vznik patologických velmi rychlých oscilací, fast ripples. Jak vidno, opět jsme konfrontováni s fyziologickou a patologickou synhronizací. 48
49 Dále například Adjamian (2004) studoval ERD/ERS ze skalpových dat nad primárním vizuálním kortexem u zdravých jedinců v případě expozice strukturovanému podnětu, jenž je známý svojí tendencí indukovat dyskomfort, zrakové iluze, bolest hlavy a až i vyvolat patternsensitivní epileptický reflexní záchvat. Zjistili pak, že s pocity dyskomfortu a iluzí časově koincidují s ERS v gamma pásmu (30-60 Hz) nad primárním zrakovým kortexem. Vyvozuje, že právě gamma oscilace mohou hrát významnou roli při iniciaci epileptického paroxysmu. Výzkum oscilací ve vysokých frekvenčních pásmech se z elektrofyziologického hlediska jeví být nejdůležitějším tématem současného bádání v epileptologii; má-li se někdy v budoucnosti podařit vyrobit analyzátor zavčas informující o blížícím se záchvatu s terapeutickou konsekvencí ať už ve formě aplikace antiepileptika či uvedení v činnost např. implantovaného stimulátoru je výzkum vysokofrekvenčních oscilací primárním předmětem zájmu Oscilace a kognitivní funkce Pojem kognice je nesmírně široký a zahrnuje velké množství mozkových funkcí. Studovaná paradigmata se zaměřují na pozornost, různé druhy paměti, inteligenci, sexuální rozdíly, anticipaci. Alfa pásmo je v kognici výrazným předmětem zájmu, uveřejněných studií je velké množství. Standardně je s větší či menší mírou u všech volených výzkumných paradigmat nacházena desynchronizace v pásmu alfa (8-12 Hz) nad oblastmi, které jsou implikovány, předpokládaně, ve zpracování daného úkolu. Nicméně je důležité zkonstatovat, že i v takto úzkém frekvenčním pásmu lze rozlišovat odlišnou reaktivitu v 8-10 Hz a Hz pásmu; desynchronizace v tzv. dolní alfě je přítomna v případě téměř jakéhokoliv paradigmatu a 49
50 pravděpodobně odráží všeobecnou připravenost a pozornost. Oproti tomu, desynchronizace v tzv. horní alfě je pozorována v paradigmatech, ve kterých je vyžadována cílená, fokusovaná pozornost a její rozsah je oproti dolní alfě omezený, restriktovaný, více lokalizovaný. Pozornost v paradigmatu kožně aplikovaného bolestivého stimulu studovali Ohara a spol (2004) u 4 pacientů se subdurálními elektrodami. V případě, že pacienti věnovali pozornost bolestivému stimulu, byla nad primární somatosensorickou oblastí, ale i parasylvicky pozorována významnější desynchronizace než za situace, kdy byly použity distraktory a pozornost k bolestivému stimulu byla snížena. Autoři pak předpokládají, že thalamokortikání okruhy hrají úlohu při modulaci bolesti a že právě tyto okruhy jsou zodpovědné za generování na podnětu závislém poklesu výkonu v určitém frekvenčním pásu (ERD). Kromě studování oscilačních změn v paradigmatech s pozorností, je zájem zaměřen také na paměťové funkce ( různá míra paměťové zátěže, pracovní paměť, dlouhodobá paměť, různá onemocnění ovlivňující paměť..) a na inteligenci. Již v začátku devadesátých letech minulého století se výzkumu oscilací a paměťovým schopnostem věnuje skupina kolem Klimesche v Salzburku; opakovaně uveřejňují údaje o nárůstu výkonu v theta pásmu ve fronto-temporálních oblastech u subjektů s lepšími mnestickými schopnostmi. My jsme potvrdili a blíže topizovali jejich výsledky analýzou dat z intracerebrální registrace (viz práce P3 and ERD/ERS in a visual oddball paradigm: a depth EEG study from the mesial temporal structures, v tisku pro Int J Psychophysiol). Zdá se, že hipokampální theta rytmus je základním substrátem při vytváření paměťových stop. Do jaké míry je pak např. i principiální oscilací v anatomickém Papezově okruhu, jenž je historickým, ale doposud akceptovaným modelem fungování paměti, se za současného stavu znalostí nedá určit. Recentně na poli kognitivního zkoumání dominuje pojem tzv. neurální účinnosti (neural efficiency). Je totiž poměrně zásadní otázkou, zda neurony, neuroglie, resp. neuronální sítě jsou schopny pracovat energeticky úsporně, tedy aktivovat jen úzce vymezenou oblast, 50
51 nebo naopak zda rozsáhlejší aktivace je výhodnější z hlediska výstupní kvality zpracování zevního či vnitřního podnětu. Jinými slovy: je inteligentnější jedinec s úzce vymezenou aktivací kortexu nebo naopak s aktivací rozsáhlou? Studovat inteligenci lze samozřejmě velmi obtížně, paradigmata se v zásadě zaměřují na snáze uchopitelné a definovatelné parametry jako je např. právě pracovní paměť. Grabner (2004) studoval souvislost mezi inteligencí a mírou kortikální aktivace v kontextu pracovní paměti v alfa pásmu. Shledal negativní korelaci mezi inteligencí a rozsahem kortikální aktivace u mužů. Neubauer dále rozvíjí studium inteligenčního kvocientu, pohlaví a hypotézu neurální efektivnosti. Roku 2005 uveřejňuje výsledek studie, ve které prokazuje korelaci mezi výkonem žen ve verbálním paradigmatu a výkonem mužů s paradigmatu s prostorovou manipulací. Obě skupiny vykazovaly negativní vztah mezi mírou kortikální aktivace a dosaženými výsledky. Výsledky kognitivních studií jsou ale protichůdné, alespoň pokud se týče horního alfa pásma; rakouská skupina nachází u inteligentnějších či lépe řečeno u subjektů s lepšími výsledky v kognitivních paradigmatech rozsáhlejší ERD ve zmíněném frekvenčním pásmu, zatímco jiné výzkumné skupiny referují o výsledcích, jež podporují hypotézu neurální efektivnosti (neural efficiency), kdy naopak ohraničenější kortikální aktivace mluví pro schopnost lépe, inteligentněji využívat energetický potenciál. V práci, ve které se skupina kolem Klimesche pokouší odpovědět na tuto otázku, jsou obsaženy následující výsledky: skupina s vyšším IQ všeobecně vykazovala výraznější aktivaci (ERD) v theta pásmu. V alfa pásmu, horním, pak pozorovali dva stavy; v případě lehkého kognitivního úkolu měla skupina s vyšším IQ menší aktivaci, ale při těžším úkolu nastával stav opačný, alfa desynchronizace Hz byla mohutnější. Zcela jiný pohled na funkční význam alfa ERS mají kolegové z Ruska (Knyazev a spol., 2005). Na rozdíl od zaběhnuté představy, že synchronizace představuje stav kognitivní inaktivity či aktivní inhibice zpracování sensorické informace navrhují, že zvýšená míra synchronie během referenčního intervalu může znamenat vyšší připravenost alfa systému na následující 51
52 informační vstup a nutnost jeho zpracování. Studovali svou hypotézu na dvou skupinách subjektů; s vyšší a nižší mírou anxiety, jimž byl prezentován akustický podět (tón 1000 Hz) nebo neutrální slovo. Subjekty s vyšší mírou anxienty pak vykazovali vyšší absolutní výkon v alfa pásmu v referenčním intervalu a současně také vyšší alfa desynchronizaci při zpracování informace. Samozřejmě, i v oblasti experimentálního výzkumu si badatelé kladou otázku praktického využití jejich metody, jejich poznatků. Elektrofyziologie, její mnohé metody, tvoří rutinní součást neurologické praxe. Přesto ale tomu tak u metody ERD/ERS není či dosud není. Snad pro příliš velkou časovou náročnost při zpracovávání dat nebo pro prozatím nedostatečně homogenní data či nestandardizovaný protokol vyšetřování u jednotlivých afekcí či podezření na ně. Ale takový osud se netýká jen hledání oscilačních změn EEG. Kognitivní P300, motorický Bereitschaftspotential, contingent negative variation jsou dalšími příklady elektrofyziologických parametrů, u nichž prozatím dominuje výzkum nad běžnou denní praxí. Babiloni a jeho kolegové (2005) v Římě jsou jedním z příkladů pokusu o klinickou aplikaci metody ERD/ERS. Srovnávali průběh oscilačních změn středočárové 6-12 Hz aktivity (magnetoencefalografická studie) u pacientů mírnou formou Alzheimerovy choroby (mini mental state 17-24) a u pacientů s vaskulární demencí oproti zdravým jedincům. Maximum alfa desynchronizace pak bylo u pacientů s demencí pozorováno později, současně bylo ale výraznější než u jedinců z kontrolní skupiny. Zvolené paradigma bylo modifikované CNV s interstimulus intervalem delším než je obvyklé (zde 3,5-5,5 s). Taková modifikace pak poměrně dobře imituje situace z běžného života, i když lze namítnout, že spíše než mnestické funkce jako takové v sobě zvolené paradigma zahrnuje pozornost a pracovní paměť. Autoři nenašli rozdíl v ERD/ERS mezi mírnou formou Alzheimerovy choroby či vaskulární demencí. Uzavírají pak, že i relativně počátečních stádiích obou onemocnění jsou již patrny změny v oscilační mozkové aktivitě. V jiné práci Karrasch srovnával v auditivně-verbálním 52
53 paradigmatu pacienty s tzv. mild cognitive deficit (MCD) se skupinou s diagnozou Alzheimerovy choroby. Jak v situaci ukládání, tak i později při vybavování informace měli pacienti s pokročilejším stádiem degenerativního onemocnění méně výraznou desynchronizaci v alfa a beta pásmu v levostranných anterotemporálních oblastech. Dalším zkoumaným fenoménem je anticipace, jev v běžném životě nesmírně využívaný a potřebný. Anticipovat, čili řekněme předvídat, očekávat např. bolest je obzvláště důležité. Výzkumem oscilačních změn v souvislosti s aplikací bolestivých podnětů v různých paradigmatech se věnuje skupina kolem profesora Stančáka v Praze, skupina kolem Claudia Babiloniho v Římě (2005). Ti posledně jmenovaní zkoumali reaktivitu EEG v paradigmatu go/no-go, kdy určujícím podnětem pro rozhodnutí subjektu byl právě bolestivý podnět. Vyšší procento alfa desynchronizace nad kontralaterálním primárním sensorimotorickým kortexem pak bylo pozorováno právě v souvislosti s aplikací bolestivého podnětu a reflektovalo anticipatorní mozkovou aktivitu. Neuper si klade otázku (2005), zda pozorované změny ve spektrální analýze, potažmo ERD/ERS změny v kognitivních paradigmatech, jsou stabilní, individuálně, v čase. Během dvou let opakovaně vyšetřovali tytéž subjekty, aby závěrem jejich bádání byl poznatek, že změna v alfa pásmu 8-10 Hz v parieto-okcipitálních oblastech je změnou nejkonzistnější, že procento ERD/ERS změn zůstává také stabilní, ale že intraindividuálně varírují změny v ostatních frekvenčních složkách a variabilita se týká také lokalizace změn do jednotlivých mozkových regionů. Toto pozorování samozřejmě může snižovat možnost uplatnitelnosti metody ERD/ERS v praxi. 53
54 4. Závěr Výzkum fungování lidského mozku patří mezi jednu u nejvíce fascinujících oblastí lidského poznávání. Máme k dispozici stále více a více sofistikovanějších berliček ve formě zobrazovacích a elektrofyziologických metod, a přesto se jen ale vlastně jak jinak krůček po krůčku posunujeme vpřed při rozkrývání fungování jednotlivých oblastí, jejich vzájemných a mnohdy dle anatomických modelů neočekávaných propojení, rozkrýváme složitost neuronálních sítí. Nejen že nám mnohdy chybějí informace, KTERÉ oblasti mozku jsou vzájemně funkčně popojeny, nýbrž jsme teprve na počátku chápání JAK k této komunikaci mnohdy i relativně velmi vzdálených oblastí mozku dochází. Zdá se velmi pravděpodobné, že substrátem komunikace budou oscilace už i vzhledem k tomu, že neexistuje fyziologický stav mozku, jenž by nebyl charakterizovatelný, popsatelný rezonantním chováním v určitém frekvenčním pásmu. Neexistuje fyziologický stav elektrického ticha. Jak si tedy vlastně představit komunikaci neuronů? Asi jako permanentní oscilování větších či menších buněčných skupin na určité frekvenci s putováním vln (traveling waves). Frekvence se může měnit dle požadavků daných zpracováváním určitého úkolu v rámci některé z neuronálních sítí. Zdá se, že základní proces aktivace mozkových oblastí je fenomén prostorově mnohem rozsáhlejší, než by se na první pohled zdálo; nutnost být připraven, tak jak to odráží hlavně alfa desynchronizace, je upřednostěna nad prostorově restriktovanou a energeticky snad méně náročnou aktivací. I tak jednoduchý úkol, jako je volní pohyb akrem končetiny neznamená, jak by se mohlo nabízet, jen aktivaci primárního kontralaterálního sensorimotorického kortexu. Právě chápání oscilací, rezonantního chování mozku nám snad umožní více proniknout k meritu věci: k rozpoznání, jak funguje lidský mozek za fyziologických i patologických stavů. Náš výzkum, přímo ve zdroji díky intracerebrálně vnořeným elektrodách, přinesl další informace do mozaiky 54
55 vědomostí. Největším přáním elektrofyziologa by pak byla praktická aplikace těchto poznatků tak, jak jsem se pokoušela nastínit v předešlých kapitolách. 5. Seznam použitých zkratek ERD event-related desynchronization ERS event-related synchronization EEG elektroencefalografie, elektroencefalogram MEG - magnetoencefalografie PET - pozitronová emisní tomografie SPECT single foton emisní tomografie fmri funkční magnetická rezonance MRS magnetická rezonanční spektrofotometrie ERP event-related potential SSEP somatosensorický evokovaný potenciál BAEP sluchový evokovaný potenciál VEP zrakový evokovaný potenciál MEP motorický evokovaný potenciál ACh - acetylcholin NA - noradrenalin DA - dopamin 5-HT - serotonin DC shift - CNV contingent negative variation BP - Bereitschaftpotential 55
56 RP Readiness potential DBS deep brain stimulation P3 kognitivní P3 (P300) potenciál MTLE mesial temporal lobe epilepsy IPSP inhibitory post-synaptic potential 6. Poděkování Vyjadřuji poděkování všem, kdo mi pomaháli a kdo se podíleli ať už na výzkumných publikacích, tak i na sepsání doktorandské práce. Největší dík pak patří panu Chlapíčkovi. 7. Literatura Adjamian P, Holliday IE, Barnes GR, Hillebrand A, Hadjipapas A, Singh KD. Induced visual illusions and gamma oscillations in human primary visual cortex. Eur J Neurosci. 2004;20: Adrian ED. Olfactory reactions in the brain of the hedgehog. J Physiol. 1942;100: Alexander GE. Basal ganglia-thalamocortical circuits: their role in control of movements. J. Clinical Neurophysiol 1994;11: Andersen P, Andersson SA, Lomo T. Thalamo-cortical relations during spontaneous barbiturate spindles. Electroencephalogr Clin Neurophysiol. 1968;24:90. Aoki F, Fetz EE, Shupe L, Lettich E, Ojemann GA. Increased gamma-range activity in human sensorimotor cortex during performance of visuomotor tasks. Clin Neurophysiol 1999;110:
57 Arroyo S, Lesser RP, Gordon B, Uematsu S, Jackson D, Webber R. Functional significance of the mu rhythm of human cortex: an electrophysiologic study with subdural electrodes. Electroencephalogr Clin Neurophysiol. 1993;87: Babiloni C, Brancucci A, Capotosto P, Arendt-Nielsen L, Chen AC, Rossini PM. Expectancy of pain is influenced by motor preparation: a high-resolution EEG study of cortical alpha rhythms. Behav Neurosci. 2005;119: Babiloni C, Cassetta E, Chiovenda P, Del Percio C, Ercolani M, Moretti DV, Moffa F, Pasqualetti P, Pizzella V, Romani GL, Tecchio F, Zappasodi F, Rossini PM. Alpha rhythms in mild dements during visual delayed choice reaction time tasks: a MEG study. Brain Res Bull. 2005;65: Balzamo E, Marquis P, Chauvel P, Regis J. Short-latency components of evoked potentials to median nerve stimulation recorded by intracerebral electrodes in the human pre- and postcentral areas. Clin Neurophysiol ;115: Barrett G, Shibasaki H, NeshigeR. Cortical potential shifts preceding voluntary movement are normal in parkinsonism. Electroencephalogr Clin Neurophysiol. 1986;63: Basar-Eroglu C, Struber D, Schurmann M, Stadler M, Basar E. Gamma-band responses in the brain: a short review of psychophysiological correlates and functional significance. Int J Psychophysiol. 1996;24: Berger H. Ueber das Elektrenkephalogramm des Menschen II. J. Psychol. Neurol., 1930, 40: Bouyer JJ, Montaron MF, Vahnee JM, Albert MP, Rougeul A. Anatomical localization of cortical beta rhythms in cat. Neuroscience. 1987;22:
58 Bragin A, Engel J Jr, Wilson CL, Fried I, Mathern GW. Hippocampal and entorhinal cortex high-frequency oscillations ( Hz) in human epileptic brain and in kainic acid--treated rats with chronic seizures. Epilepsia. 1999;40: Braitenberg V. Cortical architectonics: General and areal. In: Architectonics of the Cerebral Cortex. (M.A.B. Brazier and Petsche, H., Eds.) New York: Raven Press. Braitenberg V and Schuz A. Anatomy of the Cortex. Statistics and Geometry. New York: Springer-Verlag,1991. Brown P, Kupsch A, Magill PJ, Sharott A, Harnack D, Meissner W. Oscillatory local field potentials recorded from the subthalamic nucleus of the alert rat. Exp Neurol. 2002;177: Buzsaki G. The thalamic clock: emergent network properties. Neuroscience. 1991;41: Buzsaki G, Horvath Z, Urioste R, Hetke J, Wise K.High-frequency network oscillation in the hippocampus.science.1992;256: Cassim F, Szurhaj W, Sediri H, Devos D, Bourriez J, Poirot I, Derambure P, Defebvre L, Guieu J. Brief and sustained movements: differences in event-related (de)synchronization (ERD/ERS) patterns. Clin Neurophysiol. 2000;111: Chatrian G, Petersen MC, Lazarte JA. The blocking of the rolandic wicket rhythm and some central chnages realated to movement. Elctroenceph. Clin. Neurophysiol., 1959, 11: Cooper R, Winter AL, Crow HJ, Walter WG. Comparison of subcortical, cortical and scalp activityusing chronically indwelling electrodes in man. Electroencephalogr. Clin. Neurophysiol. 1965;18:
59 Crone N, Miglioretti D, Gordon B, Lesser R. Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. II. Event-related synchronization in the gamma band. Brain 1998;121: Crone N, Miglioretti D, Gordon B, Sieracki J, Wilson M, Uematsu S, Lesser R. Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. I. Alpha and beta event-related desynchronization. Brain 1998;121: Cooper R, Winter AL, Crow HJ, Walter WG. Comparison of subcortical, cortical and scalp activityusing chronically indwelling electrodes in man. Electroencephalogr. Clin. Neurophysiol. 1965;18: Defebvre L, Bourriez JL, Dujardin K, Derambure P, Destee A, Guieu JD. Spatiotemporal study of Bereitschaftspotential and event-related desynchronization during voluntary movement in Parkinson's disease. Brain Topogr. 1994;3: Donoghue JP, Sanes JN, Hatsopoulos NG, Gaal G. Neural discharge and local field potential oscillations in primate motor cortex during voluntary movements. J Neurophysiol. 1998;79: Doyle LM, Kuhn AA, Hariz M, Kupsch A, Schneider GH, Brown P. Levodopa-induced modulation of subthalamic beta oscillations during self-paced movements in patients with Parkinson's disease. Eur J Neurosci. 2005;5: Fisher RS, Webber WR, Lesser RP, Arroyo S, Uematsu S. High-frequency EEG activity at the start of seizures. J Clin Neurophysiol. 1992;3: Gastaut H. Etude electrocortocographique de la reactivite des rhythmes rolandiques. Rev: Neurol., 1952;87:
60 Geyer S, Ledberg A, Schleicher A, Kinomura S, Schormann T, Burgel U, Klingberg T, Larsson J, Zilles K, Roland PE. Two different areas within the primary motor cortex of man. Nature. 1996;382: Grabner RH, Fink A, Stipacek A, Neuper C, Neubauer AC. Intelligence and working memory systems: evidence of neural efficiency in alpha band ERD. Brain Res Cogn Brain Res. 2004;20: Jahnsen H, Llinas R.Electrophysiological properties of guinea-pig thalamic neurones: an in vitro study. J Physiol. 1984;349: Jasper HH and Andrew HL. Electro-encephalography III. Nornal differentiation of occipital and precentral regions in man. Arch. Neurol. Psychiatry, 1938;39: Jasper HH and Penfield W. Electrocorticograms in man: effect of the voluntary movement upon the electrical activity of the precentral gyrus. Arch. Psychiatry Z. Neurol., 1949;183: Karrasch M, Laine M, Rinne JO, Rapinoja P, Sinerva E, Krause CM. Brain oscillatory responses to an auditory-verbal working memory task in mild cognitive impairment and Alzheimer's disease. Int J Psychophysiol Jun 17 Katznelson RD. Normal modes of the brain: Neuroanatomical basis and a physiological theoretical model. In: Electric Fields of the Brain: The Neurophysics of EEG. (P.L. Nunez, Author) New York: Oxford University Press, Klimesch W. Memory processes, brain oscillations and EEG synchronization. Int J Psychophysiol 1996;24: Knyazev GG, Savostyanov AN, Levin EA.Alpha synchronization and anxiety: Implications for inhibition vs. alertness hypotheses. Int J Psychophysiol. 2005;3:
61 Kopell N, Ermentrout GB, Whittington MA, Traut RD. Gamma rhythms and beta rhythms have different synchronization properties. Proc Natl Acad Sci USA. 2000;97: Kornhuber HH, Deecke L. Changes in the brain potential in voluntary movements and passive movements in ma: readiness potential and reafferent potentials. Pflugers Arch Gesamte Physiol Menschen Tiere. 1965;10:1-17. Kuhn AA, Williams D, Kupsch A, Limousin P, Hariz M, Schneider GH, Yarrow K, Brown P. Event-related beta desynchronization in human subthalamic nucleus correlates with motor performance. Brain. 2004;127: Levy R, Ashby P, Hutchison WD, Lang AE, Lozano AM, Dostrovsky JO. Dependence of subthalamic nucleus oscillations on movement and dopamine in Parkinson's disease. Brain. 2002;125: Llinas RR. Oscillations in CNS neurons: A possible role for cortical interneurons in generationof 40-Hz oscillations. In: Induced Rhythms in the Brain (E. Basar, Bullock TH, Eds.) Boston: Birkhauser, Lopez da Silva F. Neural mechanisms underlying brain waves: from neural membranes to networks. Electroenceph. Clin. Neurophysiol. 1991;79: Neshige R, Luders H, Friedman L, Shibasaki H. Recording of movement-related potentials from the human cortex. Ann Neurol. 1988;3: Neubauer AC, Grabner RH, Fink A, Neuper C. Intelligence and neural efficiency: Further evidence of the influence of task content and sex on the brain-iq relationship. Brain Res Cogn Brain Res ; Neuper C, Grabner RH, Fink A, Neubauer AC. Long-term stability and consistency of EEG event-related (de-)synchronization across different cognitive tasks. Clin Neurophysiol. 2005;7:
62 Nunez PL. Neocortical Dynamics and Human EEG Rhythms. New York Oxford. Oxford University Press Ohara S, Ikeda A, Kunida T, Yazawa S, Baba K, Nogamine T, Taki W, Hashimoto N, Mihara T, Shibasaki. Movement-related change of electrocorticographic activity in human supplementary motor area proper. Brain 2000;123: Ohara S, Crone NE, Weiss N, Lenz FA. Attention to a painful cutaneous laser stimulus modulates electrocorticographic event-related desynchronization in humans. Clin Neurophysiol 2004;7: Paradiso G, Cunic D, Saint-Cyr JA, Hoque T, Lozano AM, Lang AE, Chen R. Involvement of human thalamus in the preparation of self-paced movement. Brain. 2004;127: Parent A and Cicchetti F. The current model of basal ganglia organization under scrutiny. Mov Disord 1998;13: Pfurtscheller G. Event-related synchronization (ERS): an electrophysiological correlate of cortical areas at rest. Electroenceph clin Neurophysiol 1992;83: Pfurtscheller G. Induced oscillations in the alpha band: functional meaning. Epilepsia. 2003;44Suppl12:2-8. Pfurtscheller G, Aranibar A. Event-related cortical desynchronization detected by power measurements of the scalp EEG. Electroenceph clin Neurophysiol 1977;42: Pfurtscheller G, Berghold A. Patterns of cortical activation during planning of voluntary movement. Electroencephalogr Clin Neurophysiol. 1989;3: Pfurtscheller G, Lopes da Silva FH. Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol 1999;110:
63 Pfurtscheller G, Neuper C. Simultaneous EEG 10 Hz desynchronization and 40 Hz synchronization during finger movements. Neuroreport. 1992;3: Pfurtscheller G, Stancak A Jr, Neuper C. Event-related synchronization (ERS) in the alpha band--an electrophysiological correlate of cortical idling: a review. Int J Psychophysiol. 1996;24: Rau C, Plewnia C, Hummel F, Gerloff C. Event-related desynchronization and excitability of the ipsilateral motor cortex during simple self-paced finger movements. Clin Neurophysiol. 2003;10: Rektor I, Louvel J, Lamarche M. Intracerebral recording of potentials accompanying simple limb movements: a SEEG study in epileptic patients. Electroencephalogr Clin Neurophysiol. 1998;4: Rektor I, Bares M, Kanovsky P, Brazdil M, Klajblova I, Streitova H, Rektorova I, Sochurkova D, Kubova D, Kuba R, Daniel P. Cognitive potentials in the basal gangliafrontocortical circuits. An intracerebral recording study. Exp Brain Res. 2004;158: Salenius S, Salmelin R, Neuper C, Pfurtscheller G, Hari R. Human cortical 40 Hz rhythm is closely related to EMG rhythmicity. Neurosci Lett. 1996;213: Sayers BM, Beagley HA. Objective evaluation of auditory evoked EEG responses. Nature. 1974;251: Shibasaki H, Barret G, Halliday E, Halliday AM. Components of the movement-related cortical potential and their scalp topography. Electroenceph clin Neurophysiol 1980;48:
64 Schurmann M, Basar-Eroglu C, Basar E. Gamma responses in the EEG: elementary signals with multiple functional correlates. Neuroreport. 1997;8: Singer W. Synchronization of cortical activity and its putative role in information processing and learning. Annu Rev Physiol. 1993;55: Sochurkova D, Rektor I. Event-related desynchronization/synchronization in the putamen. An SEEG study. Exp Brain Res. 2003;3: Stancak A, Svoboda J, Rachmanova R, Vrana J, Kralik J, Tintera J. Desynchronization of cortical rhythms following cutaneous stimulation: effects of stimulus repetition and intensity, and of the size of corpus callosum. Clin Neurophysiol. 2003;114: Steriade M, Llinas RR. The functional states of the thalamus and the associated neuronal interplay. Physiol Rev. 1988;68: Steriade M, Llinas RR. The functional states of the thalamus and the associated neuronal interplay. Physiol Rev ;68: Szurhaj W, Derambure P, Labyt E, Cassim F, Bourriez JL, Isnard J, Guieu JD, Mauguiere F. Basic mechanisms of central rhythms reactivity to preparation and execution of a voluntary movement: a stereoelectroencephalographic study. Clin Neurophysiol 2003;1: Szurhaj W, Bourriez JL, Kahane P, Chauvel P, Mauguiere F, Derambure P. Intracerebral study of gamma rhythm reactivity in the sensorimotor cortex. Eur J Neurosci 2005;5: Toro C, Deuschl G, Thatcher R, Sato S, Kufta C, Hallett M.Event-related desynchronization and movement-related cortical potentials on the ECoG and EEG. Electroencephalogr Clin Neurophysiol. 1994;93:
65 8. Přílohy 8.1. Publikace A/ Sochurkova D, Rektor I. Event-related desynchronization/synchronization in the putamen. An SEEG study. Exp Brain Res. 2003;3: B/ Daniela Sochurková, Milan Brázdil, Pavel Jurák, Ivan Rektor. P3 and ERD/ERS in a visual oddball paradigm: a depth EEG study from the mesial temporal structures (akceptováno pro tisk v Int J Psychophysiol I/2006) C/ Daniela Sochůrková, Ivan Rektor, Pavel Jurák, Andrej Stančák. Intracerebral recording of cortical activity related to self paced voluntary movements: A Bereitschaftspotential and Event-related de-/synchronization. SEEG study. (akceptováno pro tisk v Exp Brain Res) 65
66 A/ Research Notes Event-related desynchronization/synchronization in the putamen. A SEEG case study. D. Sochurkova, I. Rektor 1st Department of Neurology, Masaryk University, St. Anne s Hospital, Brno, Czech Republic Author for correspondence: Daniela Sochurkova 1 st Department of Neurology Masaryk University Pekarska 53, Brno, Czech Republic Tel.: Fax: daniela.sochurkova@fnusa.cz 66
67 Abstract Event-related desynchronization (ERD) and synchronization (ERS) were studied during the invasive exploration of an epileptic surgery candidate. An electrode that was targeted in the amygdalo-hippocampal complex passed through the putamen with several contacts. During a simple self-paced motor task, we observed in the putamen a power decline (ERD) in both the alpha and beta frequency bands, and a rebound phenomena (ERS) in the beta frequency band, concurrent with the movement of each hand. This is the first report of ERD/ERS in the basal ganglia. Keywords: Event-related desynchronization/synchronization, basal ganglia, putamen, selfpaced movement Introduction Event related desynchronization/synchronization (ERD/ERS, Pfurtscheller and Aranibar, 1977) are well-studied electrophysiological phenomena related to cortical activation and activity suppression. ERD/ERS have been studied in relation to movement preparation and performance, both with scalp and intracranial recordings. Using subdural grids or depth electrodes, ERD/ERS have been found in the primary sensorimotor cortex, in the supplementary motor cortex, in the lateral temporal cortex, and in the cingulate gyrus (Crone et al. 1998; Ohara et al. 2000; Toro et al. 1994). This is the first report of ERD/ERS recorded in the basal ganglia. Our report is based on a unique observation of EEG data, which was acquired from some electrode contacts located in the putamen during the presurgical invasive monitoring (SEEG) of a drug-resistant epileptic patient. The basal ganglia are not a target for SEEG exploration in epileptic patients; nevertheless, some contacts of a diagonal depth electrode targeted into the amygdalo-hippocampal complex may occasionally pass through 67
68 the basal ganglia. Given the fact that there are no other reports of ERD/ERS in the basal ganglia, and that the probability of recording from these structures in human subjects in the future is low, we decided to publish this report of a recording in a single subject. Case study An invasive video-eeg study was performed on a 28 year old right-handed man, who had been suffering from drug-resistant mesiotemporal lobe epilepsy for 13 years, with perinatal anoxia, and with a history of febrile seizures. The MRI revealed mesiotemporal sclerosis and a stripy gliosis between the roof of the lateral ventricle and the precentral region on the right side. The patient remained unresponsive to conventional forms of antiepileptic drug therapy. Prior to the epilepsy surgery, he was recommended by a special commission for invasive video-eeg monitoring. He gave his informed consent for the electrophysiological testing. The study received the approval of the local ethics committee. Intracerebral depth electrodes were implanted to localise the seizure onset zone prior to the surgical treatment. Five multicontact platinum electrodes were implanted (4 orthogonal; 1 diagonal) using Talairach s stereotactic co-ordinate system (Talairach et al. 1967; Talairach and Tournoux 1988). MicroDeep (DIXI, Briançon, France) semiflexible electrodes, each with a diameter of 0.8 mm, a contact length 2 mm, and an intercontact interval of 1.5 mm were used for invasive EEG monitoring. Contacts at the electrode were always numbered from the medial to the lateral side, and from the inner to the outer side, respectively. The exact position of the electrodes and their contacts in the brain were verified with the electrodes in situ using postplacement MRI. A diagonal electrode (X) passed from the frontal cortex through the basal ganglia into the amygdalo-hippocampal complex. The implantation of diagonal electrodes is a safe and well-established method that has used in epileptology for decades. The electrode is never targeted into the basal ganglia, but in some cases the electrode passes through the basal 68
69 ganglia with one or several contacts. This technique is used only exceptionally in our department; the placement of orthogonal electrodes, with the electrodes controlled by MRI while in situ, is prefered. Four orthogonal electrodes (C, O, L, and P) were inserted into the middle part of the hippocampus, fusiform gyrus, middle temporal gyrus, anterior cingulate gyrus, orbital gyruses, inferior frontal gyrus, middle frontal gyrus, posterior cingulate gyrus, and the supramarginal and angular gyrus. All of the explored sites were on the right side. No adverse effect and no sequels of depth electrode implantation were seen. A Cpz scalp electrode (10-20 International System) was placed on the vertex. Linked earlobes were used as a reference. The surface EMG of the movements of both hands were recorded bipolarly using sintered electrodes placed over the m. flexor digitorum superficialis in the ulnar portion. Electrode impedance was no more than 5 kω for all of the electrodes. Electrical potentials (DC amplifiers; bandpass Hz; sampling rate 256 Hz) were continuously recorded using an 88-channel EEG system (BrainScope, M&I, Prague, Czech Republic). During the preoperative video-eeg monitoring, the patient performed the modified Bereitschaftspotential paradigm with a brisk self-paced flexion of the fingers (Kornhuber and Deecke 1965, Rektor et al. 1994). The subject voluntarily performed movements at irregular intervals ranging from 12 to 15 seconds. He was instructed to fix his gaze on a point about 2 meters in front of him, and to avoid any movements other than those requested. Two separate sessions, each approximately 15 minutes, were performed for each hand movement. About 40 artefact-free trials, each lasting 10 seconds, were recorded with the movements of each hand. The trials for both hands were analysed (ERD/ERS computation) in five different frequency bands for each hand (7-10, 16-24, 24-28, 28-32, Hz), using software designed by Stančák (Stančák 2000). Data were smoothed by the Blackman window (1 s) prior to the fast Fourier transform computation. Adjacent 1 s epochs overlapped by 125 ms (32 samples). 69
70 Seventy-two spectral estimates, representing 4.5 s of the pre-movement period and 4.5 s of the post-movement period, were obtained for each 10 s trial. Spectral arrays from all of the trials were then averaged separately for each electrode. The frequency components displaying a significant (P< 0.05) power decrease or increase were identified. The sampling rate was 256 Hz. The spectral analysis did not show any significant change in the higher frequency bands. A simple averaging procedure for each session was performed to measure readiness potential (BP, Bereitschaftspotential). RP was defined as a clear rise or decrease from the baseline, with a slope greater than ± 10 µv/s. The baseline for both ERD/ERS and RP computation was defined during the first 1000 ms of each of the 10 s trials. Results EMG bursts corresponding to the brisk fingers flexions lasted at about 650 ms. The frequency bands which displayed the highest oscillatory changes were chosen as bands of interest. In both scalp and intracerebral recordings, ERD/ERS was widely displayed in alpha and beta frequency bands. In the alpha band (8-12 Hz), the most important power decline was for 8 Hz; therefore a band 7-10 Hz was further analysed. In the beta band, ERD/ERS phenomena was of the highest percentage in Hz band. In scalp Cpz, an alpha ERD (55%; onset approximately 1 s before the movement onset) and a beta ERD (40%; onset 500 ms before the movement onset) were recorded. A mild beta ERS (24-28 Hz; 25%, data not shown) was observed. The ERD/ERS findings on the scalp were similar to those reported in previous literature (Pfurtscheller and Berghold, 1989). In intracranial recordings through orthogonal electrodes, ERD/ERS were displayed in several cortical areas and structures (in the amygdalo-hippocampal formation, in the posterior cingulate gyrus, and in the supramarginal gyrus; these results will be reported elsewhere, in conjuction with recordings of other subjects; this report is focused on the unique observation of ERD/ERS in the putamen). 70
71 The highest percentage of power changes (see Fig.1) from all the explored structures was seen in the putamen, with movements of each hand; the power changes concerned all of the investigated frequency bands except the Hz band. In the alpha frequency band (7-10 Hz), a power decline started 2 s before the movement onset, the highest level of change (50%) was 2 s after the movement onset, and reached the baseline approximately 3-4 s after the movement offset. In the beta bands, the ERD/ERS phenomena was more significant than the alpha power changes; the onset of power decline (ERD) was also approximately 2 s before the movement onset, the highest level of change (75%) was 500 ms after the movement onset, a high percentage ( %) beta rebound (ERS) was observed 1500 ms after the movement offset; the baseline power value was reached in the subsequent 2 s. The beta rebound (ERS) was approximately 50 % higher in the ipsilateral hand movement than in the contralateral hand movement (130%). Bereitschaftspotential was not displayed in the explored putaminal contacts in this patient. Discussion This is the first presentation of a recording of ERD/ERS in the basal ganglia. In this case, no BP was recorded in the putamen. However, we know from recordings in other patients that BP can be generated in the basal ganglia (Rektor et al. 2001). In this case, the contacts were probably in a part of the putamen where BP is not displayed at variance with ERD/ERS. There was no reason to believe that any pathological change of the basal ganglia activity was present, and no epileptic activity was recorded in this structure, either. Nevertheless, a bias due to plasticity changes in epilepsy patients cannot be fully excluded. BP and ERD/ERS represent different physiological processes related to movement preparation and execution, at least in the putamen. The finding of power changes in the putamen with bilateral hand movements indicate that ERD/ERS is not exclusively a cortical phenomena. It seems that the 71
72 ERD/ERS is displayed in the cortical as well as in the subcortical parts of the corticosubcortico-cortical circuitry (Alexander 1994). This fits with the suggestion that there is a slow, long-lasting parallel activation of cortical and subcortical structures before and during motor tasks (Rektor 2000). References Alexander GE (1994) Basal ganglia-thalamocortical circuits: their role in control of movements. J. Clinical Neurophysiol 11: Crone NE, Miglioretti DL, Gordon B, Sieracki JM, Wilson MT, Uematsu S, Lesser RP (1998) Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. I. Alpha and beta event-related desynchronization. Brain 121: Kornhuber HH, Deecke L. (1964) Hirnpotentialaenderung beim Menschen vor und nach Willkuerbewegungen, dargestellt mit Magnetbandspeicherungund Rueckwaertsanalyse. Pfluegers Archiv; 281: 52. Ohara S, Ikeda A, Kunieda T, Yazawa S, Baba K, Nagamine T, Taki W, Hashimoto N, Mihara T, Shibasaki H (2000) Movement-related change of electrocorticographic activity in human supplementary motor area proper. Brain 123: Pfurtscheller G, Aranibar A (1977) Event-related cortical desynchronization detected by power measurements of scalp EEG. Electroenceph clin Neurophysiol 42: Pfurtscheller G, Berghold A (1989) Patterns of cortical activation during planning of voluntary movement. Electroenceph clin Neurophysiol 72: Rektor I, Fève A, Buser P, Bathien N, Lamarche M (1994) Intracerebral recording of movement related readiness potentials: an exploration in epileptic patients. Electroenceph clin Neurophysiol 90: Rektor I (2000) Parallel information processing in motor systems: intracerebral recordings of readiness potential and CNV in human subjects. Neural Plasticity 7:
73 Rektor I, Bareš M, Kubová D (2001) Movement related potentials in the basal ganglia. A SEEG readinnes potential study. Clin. Neurophysiol 112 : Stančák A Jr. (2000) The electroencephalographic β synchronization following extension and flexion finger movements in humans. Neuroscience Letters 284:41-44 Talairach J, Szikla G, Tournoux P, Prosalentis A, Bordas-Ferrer M, Covello J, et al. (1967) Atlas d anatomie stéréotaxique du télencephale. Masson, Paris. Toro C, Deuschl G, Thatcher R, Sato S, Kufta C, Hallett M (1994) Event-related desynchronization and movement-related cortical potentials on the ECoG and EEG. Electroenceph clin Neurophysiol 93: Acknowledgements We wish to thank to Dr. Stančák providing us with appropriate software, and Mr. Daniel for his technical assistance. We would also like to extend our thanks to the Epilepsy Center Staff, including: Dr. Novák and Dr. Chrastina (neurosurgery), Dr. Pažourková and Dr. Krupa (neuroradiology), Dr. Brázdil and Dr. Kuba (neurology). This research was supported by MS CR Research Program Legend Figure 1. ERD/ERS recorded from the diagonal electrode passing from the fusiform gyrus (X1, 2, 3), through the hippocampus (X4), the lateral part of amygdala (X5, 6), the white matter (X7, 8, X15), and the putamen (X9,10,11,12,13,14). ERD/ERS with the respect to the movement onset, with left and right hand movements, are displayed. The position of the electrode is shown on the right. The time scale is 10 s, the mark indicates the movement onset at 5 s. Příloha obrázek 73
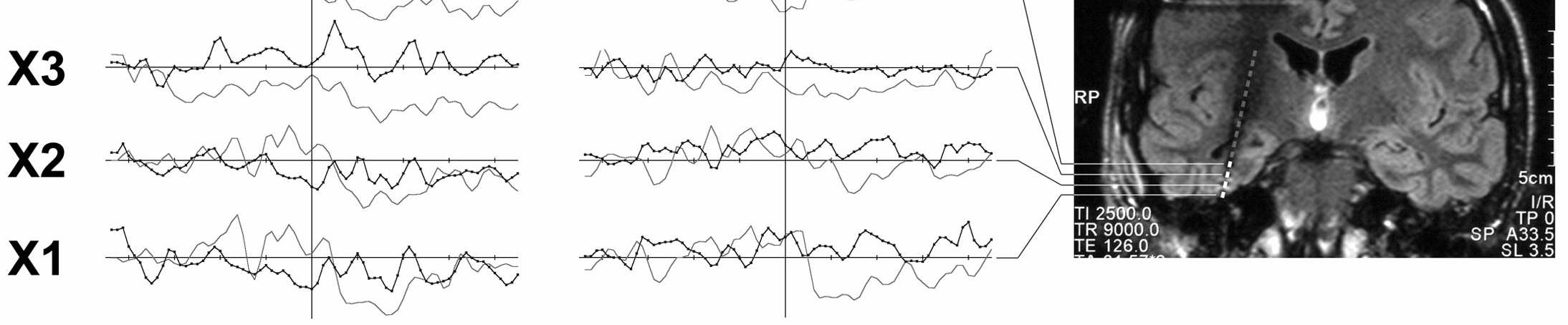
74 74
75 B/ P3 and ERD/ERS in a visual oddball paradigm: a depth EEG study from the mesial temporal structures Daniela Sochurková a, *, Milan Brázdil a, Pavel Jurák b, Ivan Rektor a a First Department of Neurology, Saint-Anne s University Hospital, Brno, Czech Republic b Institute of Scientific Instruments, Academy of Sciences, Brno, Czech Republic * Corresponding author Tel.: Fax: dasoch@med.muni.cz Acknowledgements We wish to thank Prof. Pfurtscheller for his kind advice, Mr. Daniel for technical assistance, and Dr. Tyrlík for statistical analysis. Our thanks also go to Dr. Novák and Dr. Chrastina (neurosurgery), and to Dr. Pazourková, Dr. Feitová and Dr. Krupa (neuroradiology). This research was supported by the Yamanouchi European Foundation, by MS CR Research Program and by GACR grant No. 102/05/
76 P3 and ERD/ERS in a visual oddball paradigm: a depth EEG study from the mesial temporal structures Objective: The present study intracerebrally investigates the concurrent occurrence of eventrelated potentials (ERPs) and event-related desynchronization/synchronization phenomena (ERD/ERS) in subjects performing a visual oddball task. Methods: Evoked (P3) and induced (ERD/ERS) changes were studied in six subjects with drug resistant epilepsy. Depth EEG activity from mesiotemporal limbic structures (amygdala, hippocampus and parahippocampal gyrus) was analysed. We used an averaging of raw data to obtain ERPs, and an averaging of the amplitude/power envelope (complex demodulation) in five frequency bands - theta, alpha 1, alpha 2, beta, and gamma - to obtain ERD/ERS. The P3 component of ERP and ERD/ERS to target and non-target stimuli were evaluated. Results: Even though both P3 and ERD/ERS phenomena were observed in all investigated mesiotemporal regions, the most pronounced findings were revealed in the hippocampus. A P3 component of ERP was repeatedly observed in the hippocampus after target stimuli. Significant changes of the oscillatory hippocampal activity were found: ERD in the alpha 1 and alpha 2 frequency bands. Evident, near-significant, changes were observed throughout the hippocampus in the theta band. No apparent ERD/ERS changes in the beta or in the gamma frequency band were seen in our intracerebral data. The distribution of the P3 and ERD/ERS maxima differed slightly in adjacent contacts, but the time of their occurrence was the same. Conclusions: The cognitive processing of visual information is simultaneously accompanied by several electrophysiological phenomena. In addition to ERPs, apparent changes in oscillatory activity were detected in the intracerebral recordings from mesiotemporal limbic 76
77 structures. Timing of both neuronal processes seems to be simultaneous, but spatial distribution which differ could reflect different network being involved in the task.used. Keywords: oddball paradigm; visual P3; intracerebral recordings; event-related de/synchronization. 1. Introduction There are many ways to analyse the electrophysiological data. Most EEG research focuses on event-related potentials (ERPs) which reflect evoked EEG activity or on changes in the EEG power spectrum (event-related desynchronization/synchronization; ERD/ERS; Pfurtscheller and Aranibar, 1977) which reflect induced EEG activity. In cognitive studies, both phenomenas are often investigated but they are rarely studied simultaneously. The most studied waveform of ERPs in cognitive domaine is a P300 or P3 component. It is viewed as an indicator of up-dating working memory (Donchin and Coles, 1988), or as an indicator of the closure of the recognition demand, or as a reflection of a decision (Smith et al., 1990; Paller et al., 1987; Brázdil et al., 1999). It is a long latency large positive potential with a centro-parietal distribution in scalp recordings, which is consistently elicited by rare (target) stimuli in an oddball task. An oddball task is based on a random alternating presentation of target (rare) and non-target (frequent) stimuli, in which the subject is asked to pay attention only to the rare stimuli, and to indicate the presence of this stimuli as soon as possible according to a given instruction (simple motor response, mental counting, etc.). The precise localisation of P3 generators have been found in studies with depth electrodes in a number of sites (mesial temporal lobe, lateral temporal lobe, dorsolateral prefrontal cortex.. etc); however, there are many areas where the presence of this evoked potential has not yet been ascertained (McCarthy et al., 1989; Halgren et al., 1995, 1998; Brázdil et al., 1999). 77
78 Power changes of the background oscillatory activity in cognitive tasks were investigated in different paradigmas. Theta power increase (ERS, event-related synchronization) is considered to reflect short-term(episodic) memory (Klimesch et al., 1996, 1999; Krause et al. 2001). The alpha power decline (ERD, event-related desynchronization) was observed in conditions when alertness and attention (Dujardin et al., 1993) or long-term (semantic) memory (Klimesch et al., 1996; 1999) were demanded. Recently, many authors report the importance of gamma oscillations in cognitive tasks either in scalp or in intracranial EEG recordes analyses (Howard et al, 2003; Sederberg et al, 2003; Babiloni et al, 2004; Fitzgibbon et al., 2004; Goffaux et al, 2004). The simultaneous analysis of both P3 and ERD/ERS phenomena has been conducted in some scalp recordings, in visual, visuo-verbal and auditory oddball tasks. Sergeant et al. (1987) focused their interest on the alpha frequency band and P3 waveform changes influenced by different task variables in a visual oddball paradigm. With a higher cognitive load, they observed the growing power decline, its longer duration and a longer P3 latency. Significantly larger P3 and delta and theta increase in the same time interval was observed in visuo-verbal oddball task (Klimesch et al., 2000). In P3 paradigm studies of Basar-Eroglu et al. (1992, 2001) the delta and theta power increase was found and this power change was independent of the modality of the stimulation. Gurtubay et al. (2001) examined the characteristics of induced gamma band oscillatory responses during P3 development in an auditory oddball task. They found a late oscillatory activity peaking at 37 Hz with a latency around 360 ms appearing only after target stimuli, in scalp EEGs. The latency of this activity had a high correlation to the latency of the simultaneously analysed P3 waveform. It has been shown in motor tasks that slow potentials (motor readiness potential, contingent negative variation) and oscillatory changes do not reflect the same neuronal processes (Defebvre et al., 1994; Stancak et al., 2000; Filipovic et al., 2001). The aim of our 78
79 study was to investigate the concurrent occurrence of both P3 and ERD/ERS in EEGs recorded intracerebrally from mesial temporal structures in a visual oddball task in epilepsy patients during the preoperative invasive monitoring and to analyze the time and spatial relationship between them. Hippocampal formation is known to play a crucial role in cognitive processing, therefore this structure has been chosen for our analysis. A postplacement MRI permitted a rigorous localisation of the implanted electrodes. Even the intracerebral recordings give information about changes in small limited area we suppose that this type of analysis is of importance for the research concerning identification of anatomical but also functional substrate in cognitive assessment. The direct recording within the candidate structures is the most precise method for localising P3 and oscillatory changes generated structures. 2. Subjects and methods 2.1. Subjects Six patients (five males and one female) with an average age of 29.0 years (ranging from 23 to 36 years), all suffering from drug-resistant epilepsy, participated in the study (Table 1). All subjects had normal or corrected-to-normal vision. Informed consent was obtained from each subject prior to the experiment, and the study received the approval of the local ethics committee Procedure The standard visual oddball paradigm was used. Subjects were seated comfortably in a moderately lighted room with a monitor screen positioned approximately 100 cm in front of their eyes. During the examination, they were requested to maintain visual focus on the small fixation point in the centre of the screen, and to minimise blinking. Two types of emotionally 79
80 neutral stimuli - target and frequent - were presented in the centre of the screen in random order. The experimental stimuli consisted of yellow uppercase letters X (target) and O (frequent) on a white background. The ratio of target to frequent stimuli was 1:5. The duration of stimulus exposure was always 200 ms. The interstimulus interval varied between 2 and 5 seconds. Two task blocks were performed, each of duration approximatively 10 minutes, the pause between trials was 15 minutes. Subjects were instructed to respond to the target stimulus as quickly and as accurately as possible by pressing a microswitch button in the dominant hand, and they were requested to simultaneously count the targets in a silent manner EEG recording The EEG signal was simultaneously recorded from various intracerebral structures, using the 64-channel Brain Quick EEG system (Micromed). All recordings were monopolar, using a reference electrode on the right processus mastoideus. A single control CPz scalp electrode (situated between Cz and Pz) was used. EEGs were amplified with a bandwidth of Hz for further P3 analysis. The sampling rate was 128 Hz. Depth electrodes were implanted to localise the seizure origin prior to surgical treatment. Each patient received 4-9 orthogonal platinum electrodes using the Talairach methodology (Talairach et al., 1967). In some patients, an additional diagonal electrode was inserted stereotactically into the amygdalo-hippocampal complex opposite to the presumed epileptogenic zone. These diagonal electrodes were always targeted into the mesiotemporal structures via a frontal approach, passing through the basal ganglia. Standard MicroDeep semiflexible electrodes (DIXI) with diameters of 0.8 mm, contact lengths of 2 mm, and intercontact intervals of 1.5 mm were used for invasive EEG monitoring. Contacts at the electrode (5-15) were always numbered from the medial to the lateral side. Their positions were indicated in relation to the axes defined by the Talairach system using the "x,y,z" 80
81 format, where "x" = lateral, mm to midline, positive right hemisphere; "y" = anteroposterior, mm to the AC line, positive anterior; and "z" = vertical, mm to the AC-PC line, positive up. The exact positions of the electrodes and their contacts in the brain were verified using postplacement MRI with electrodes in situ Data analysis The EEG data from all fifty electrode contacts inserted into mesiotemporal structures were analysed. Twenty-three contacts were placed in the hippocampus (11 on the right side; 12 on the left), seventeen in the parahippocampal gyrus (11 on the right side; 6 on the left) and ten in the amygdala (4 on the right side; 6 on the left). The number of contacts across the structures varied from one to five. ScopeWin software was used for both EEG signal analyses. Further processing was performed off-line with artefact-free EEG periods. The rejecting of artefact trials was done after conspicious visual analysis by experienced electrophysiologist. The recordings from lesional anatomical structures and epileptogenic zones were not included in the analysis. A minimum of 20 trials were obtained for each contact for each task block. Evoked changes (P3) time domain analysis: Two-second EEG periods were averaged offline using the stimulus onset as the trigger (-200 ms and ms from stimulus onset). Trials were independently averaged for both the target and the frequent stimuli. The main ERP components in the latency range of ms were identified by visual inspection, and quantified by latency and amplitude measures. In intracerebral findings, the most interesting results were the findings of polarity reversal or steep voltage gradients, which uniquely proved the focal origin of the waveform (for details, see Brázdil et al., 1999).. Induced changes (ERD/ERS joint time-frequency analysis: The length of the analysed EEG period was again two seconds (-500 ms and ms from stimulus onset). An interval preceding the stimulus onset by 500 ms was chosen as the reference baseline. The methodology of complex demodulation (CD) was used for ERD/ERS quantification (Nogawa 81
82 et al., 1976). The result of CD is an instantaneous amplitude of selected frequency range oscillations. First, the intertrial variance method was applied to avoid the admixture of ERPs in the spectral analysis (Kalcher and Pfurtscheller, 1995). Subsequently, each EEG period underwent a complex demodulation. Both amplitude and power analysis was used, partly to lower the risk of result distortion as a result of non-homogeneous data, and partly to compare both possibilities of ERD/ERS expression. An average of the demodulated trials in the chosen frequency bands (theta 4-7 Hz, alpha Hz, alpha Hz, beta Hz, gamma Hz) was submitted to further analysis. In the reference interval, the mean value was counted. In the post-stimulus interval, the maximal/minimal values and its latency was detected and statistically compared with the mean value. SPSS software was used for statistical analysis. A model consisting of relations among frequent/target events, baseline value/amplitude or power values of ERD/ERS, 4-7 Hz /8-10 Hz /10-12 Hz frequency bands, and hippocampus/parahippocampal gyrus/amygdala structure was assessed. Multivariante analyses (MANOVA statistics with repeated measures) and Wilcoxon and Kruskal Wallis non-parametric test were demanded for such a model. 3. Results All of the patients satisfactorily co-participated in the study. The mean reaction time by pressing a microswitch button in the dominant hand was ± ms (Table 1) Evoked changes (P3) In scalp recordings, after the frequent stimuli only mild non-significant changes were found. After the target stimuli, the individual and grand average scalp ERPs across all six subjects revealed a typical P3 response in the centro-parietal region (CPz) (Fig. 1). The mean 82
83 peak latency of the P3 response was ± ms, and the mean N2/P3 peak-to-peak amplitude was 11.5 ± 4.00 µv. Intracerebrally, P3 waveforms were observed in all investigated mesiotemporal structures. The mean latencies of these P3 potentials were ± ms in the hippocampus, ± ms in the amygdala, and ± ms in the parahippocampal gyrus. The amplitudes of individual as well as averaged intracerebral P3 waveforms after target stimuli were generally several times greater than in scalp recordings (up to 300 µv). However, these amplitudes did not seem to be very consistent in the investigated structures. In the hippocampus, fifty percent of the P3 potentials were seen in the form of a phase reversal. In the amygdala and in the parahippocampal gyrus, generators were less frequently observed (twenty and twelve percent, respectively). The latencies among explored structures did not significantly differ Induced changes (ERD/ERS) In the scalp CPz electrode, apparent changes in oscillatory responses after target stimuli were seen in some subjects: ERS in the theta frequency band with a mean peak latency of ± ms (mean power increase of 48.3 ± 27.10%); ERD in alpha 1 (mean peak latency of ± 5.66 ms; mean power decrease of 48.6 ± 13.00%); ERD in alpha 2 (mean peak latency of ± ms, mean power decrease of 31.3 ± 16.69%); and ERD in the beta frequency band (mean peak latency of ± ms; mean power decrease 60.9 ± 8.62%). However, these findings did not reach a statistical significance across the group data. No consistent ERD or ERS phenomena were proven extracranially from our data in the gamma frequency band. Intracerebrally, clear-cut oscillatory responses were observed in both amplitude and power spectral analysis in the theta, alpha 1 and alpha 2 frequency bands after the target stimuli. These ERD/ERS phenomena were revealed in all the investigated mesiotemporal 83
84 structures, with the most pronounced findings in the hippocampal formations (Fig.2). At the same time, no apparent changes were seen in the beta or gamma bands. The observed amplitude and power changes strongly depended on the investigated frequency band: in the theta frequency band, mostly ERS, and in both of the alpha frequency bands, ERDs were displayed (Table 2). In amplitude analysis, we found a near-significant difference in amplitude changes following frequent/target events among the three frequency bands (F(d.f.=2,13) =3.687, p= 0.054). We also found that the amplitude change following frequent/target events varies nearsignificantly among the three frequency bands in dependence upon the differences among the hippocampus, parahippocampal gyrus and amygdala structures. (F(d.f.=4,26) =2.512, p = 0.066). Since the number of explored subjects was low, we used non-parametric tests in order to support these results. We did not find any significant difference among the three structures when the individual amplitudes of the baseline and of the ERD/ERS values were taken into account. Instead, we found significant or near-significant differences between: the target baseline and target amplitude values ERD in 8-10 Hz frequency band when the hippocampus was addressed (Wilcoxon SRT z=-2.2; p= 0.031); and the non-target baseline and non-target amplitude values ERD in 8-10 Hz frequency band when the parahippocampal gyrus was addressed (Wilcoxon SRT z=-1.96; p= 0.055). We did not find any significant differences among the amygdala. In order to find differences between the ERD/S value baseline value in relation to target and non-target events, we subtracted the individual amplitude value from the baseline value. Results in delta for target events were compared to results in delta for nontarget events. We found near-significant differences in the 4-7 Hz band (Wilcoxon SRT z=- 1.99; p=0.063) and the 8-10 Hz band (Wilcoxon SRT z=-1.99; p=0.063) among hippocampus measurements, with amplitude increase in theta frequency band 4-7 Hz and amplitude decline 84
85 in low alpha frequency band 8-10 Hz. We did not find any analogous difference when the parahippocampal gyrus and amygdala were taken into account. The same statistical model was used in the power analysis, in which we found nearsignificant differences in the power change following frequent/target stimuli among the three frequency bands (F(d.f.=2,13) =3.487, p=0.062). In contrast to the amplitude analysis, our study did not reveal that the power change following frequent/target events varies among the three frequency bands in dependence upon the differences among the hippocampus, parahipocampal gyrus and amygdala structures (F(d.f.=4,26) =1.022, p= 0.414). We have used the non-parametric tests in order to support these results also in power analysis. We did not find any significant difference among three structures when individual baseline and power ERD/S values were taken into account. Instead, we found significant or near-significant differences in a form of power decline between: target baseline and target power values when the hippocampus was addressed in the 8-10 Hz frequency band (Wilcoxon SRT z=-2.2; p= 0.031), and in the Hz frequency band (Wilcoxon SRT z=-2.2; p= 0.031); between the non-target baseline and non-target power values in 8-10 Hz frequency band when parahippocampal gyrus was addressed (Wilcoxon SRT z=-2.24; p= 0.023). We did not find any significant differences in the amygdala. In order to investigate differences between the power ERD/ERS value baseline value relation among target and non-target events, we subtracted the individual power value from the baseline value. The results in delta for target events were compared to results in delta for non-target events. We found near-significant differences in the 4-7 Hz band (Wilcoxon SRT z=-1.99; p=0.063) and significant differences in the 8-10 Hz frequency band (Wilcoxon SRT z=-1.99; p=0.031) among hippocampus measurements. We did not find any analogous difference when the parahippocampal gyrus and amygdala were taken into account. 85
86 Even though the results from amplitude and power statistical analysis differ slightly, the main characteristic of results are the same. This finding is even more important from the point of view that both amplitude and power values of ERD/ERS were measured separately by independent observers. Concerning the time domain, the courses of intracerebral ERD/ERS waveforms were smooth, with simple peaks. The onset of amplitude/power changes was 200 ms ± 100 ms after a target stimulus onset for all investigated frequency bands and structures. The latencies of maximal oscillatory changes apparently varied across the structures and analysed frequency bands. No significant differences in peak latencies were revealed by statistics. The return to the baseline was observed within the analysed 1.5 second period, usually at about 1200 ms (Fig.3). We did not find statistical differences among the latencies of P3-like potentials and maxima of ERD/ERS phenomena (Table 2). Concerning spatial distribution, a rigorous assessment of P3 and of ERD/ERS in each contact of one electrode showed the different distribution of the observed phenomenon in one structure (Fig.4). 4. Discussion In our study, mesiotemporal structures displayed both of concerned electrophysiological phenoma (ERPs, ERD/S). P3 potentials were observed in all investigated structures; in the hippocampus, the highest number of ERPs generators was found. Oscillatory changes were observed in theta (ERS), low (ERD) and high alpha (ERD) frequency bands. The most pronounced findings were elicited in the hippocampus. The study was designed in such a manner to give a possibility to compare both phenomena in sense of spatial distribution and time evolution. In order to achieve the 86
87 comparability of both evoked and induced activities, we met three conditions. First, we used same data for computation of P3 and ERD/ERS; second, we avoided as much as possible the false interpretation of spectral analysis caused by the admixture of phase-locked (evoked) activity when background oscillatory changes was evaluated. We applied the method of intertrial variance (Kalcher and Pfurtscheller, 1995), in which the phase-locked evoked activity (P3) resulting from the average of all trials is subtracted from the raw data in each trial before the spectral analysis is performed. The third condition concerned the time scale. We were comparing the maxima of amplitude/power in both phenomena, so the most accurate assessment of evolution in time was necessary. For this purpose, we used the methodology of complex demodulation in the chosen frequency bands, where an amplitude/power value in each sampling rate is acquired (Nogawa et al., 1976). In studies concerning ERD/ERS, a power analysis is usually carried out. The EEG signal has a low signal-to-noise ratio (SNR). The averaging of instantaneous power gives better results than the averaging of instantaneus amplitude. This is why power analyse is routinely used. Bearing in mind that power analysis can emphasises the differences, we applied complex demodulation in the same chosen frequency bands in both the amplitude and power analysis. Both computation produced comparable results, however we observed statistical significance being more important in power analysis. Control scalp electrodes placed between Cz and Pz (according to the international system) gave us the opportunity to compare our results with previous scalp studies. P3 response to the target stimuli was clearly revealed in this region. Oscillatory changes were observed for the theta band (ERS), alpha band (ERD), and beta band (ERD). Taking into account the placement of scalp electrodes, the beta band decline, which was not elicited in the deep mesial structures of temporal lobe, is probably a reflection of the motor activity required in response to the labelled target stimuli. On the other hand, slow oscillatory activity in the 87
88 theta band rising from these structures is more probably accessible in scalp recording because of electrophysiological properties of slow oscillations. Gamma oscillatory activity was not observed in scalp recordings. All deep structures explored in this study displayed P3 and oscillatory changes. The highest number of P3 generator was found in hippocampus with the latency comparable with previously reported results (Halgren et al., 1995; Brázdil et al., 1999). It is suggested that hippocampus is implied in at least two of neurocognitive network (directed attention and memory) which are necessary in such a simple visual discrimination task as an oddball paradigm. Simultaneously to these evoked responses the oscillatory changes were also observed, in particular in hippocampus. The most reliable was the desynchronization in alpha 8-10 Hz frequency band which is thought to reflect the general attentional demand in elaborated task. The more focused attention which is thought to be reflected by Hz desynchronization and the episodic memory demand depictable by 4-7 Hz synchronization were less consistent when the values of ERD/ERS were statistically analysed even though the number of such a observations was high (Table 2). It is known that shift in oscillatory activity is strongly frequency and functionally dependent and that at the same time interval desynchronization in one and synchronization in other frequency band can be observed (Pfurtscheller and da Silva, 1999). In view of the chosen paradigm, with a motor task and mental counting, we cannot be certain whether the observed ERD/ERS were influenced only by memory and attentive demands to recognise a target stimuli, or whether they were also influenced by the influenced by the required responses, motor with pressing a switch and mental with counting of targets number. Furthermore, we did not find any oscillatory changes in the gamma band in any of the investigated structures. The supposed binding frequency was reported mainly around 40 Hz, though higher gamma oscillations fulfilling this likely function were also noticed. In our study, with a sampling frequency of 128 Hz, we were 88
89 unable to carry out these high gamma bands. Even in the chosen Hz band, no amplitude/power in the deep structures of the temporal lobes were seen which is in the contrast with recently reported studies (Howard et al, 2003; Sedeberg et al., 2003; Babiloni et al, 2004). Finally, we did not found the difference in the time of appearance and the time evolution of both phenomena. But the spatial distribution of P3 and of ERD/ERS in each contact of one electrode differed in one structure. This finding is the only result of our study which can support the hypothesis that evoked and induced responses did not reflect the same neuronal process and that they may be given by activation of different neuronal groups. It has to be also considered that our patients were treated by antiepileptic drugs for many years, that even they were during depth EEG monitoring without treatment we could not exclude the possibility of drugs effect on neuronal circuitry functions. From the same point of view, the long lasting epileptic syndrome could be responsible for a reorganisation of neuronal network. Our results from intracerebral EEG data could shed a bit more light on the theories dealing with the oscillatory changes in structures involved in a cognitive task. The most interesting results of our study were: first, the greater precision in the distinction in the rhythmical responses to an event within the mesiotemporal structures, when a distinction between hippocampus, parahippocampal gyrus and amygdala could be made; and second, the differences between the theta and the lower and upper alpha bands were obvious. Our results do not contradict previous cognitive studies with scalp recordings from electrodes situated over the temporal lobe, the modelling of underlying neuronal networks functions from scalp recordings is supported by real findings from intracerebral data in our study. 89
90 References Babiloni C, Babiloni F, Carducci F, Cappa S, Cincotti F, Del Percio C, Miniussi C, Moretti DV, Pasqualetti P, Rossi S, Sosta K, Rossini PM. Human cortical EEG rhythms during long-term episodic memory task. A high-resolution EEG study of the HERA model. Neuroimage. 2004;4: Babiloni C, Bares M, Vecchio F, Brazdil M, Jurak P, Moretti DV, Ubaldi A, Rossini PM, Rektor I. Synchronization of gamma oscillation increases functional connectivity of human hippocampus and inferior-middle temporal cortex during repetitive visuomotor events. Eur J Neurosci 2004;11: Basar-Eroglu C, Basar E, Demiralp T, Schurmann M. P300-response: possible psychophysiological correlates in delta and theta frequency channels. A review. Int J Psychophysiol. 1992;2: Basar E, Basar-Eroglu C, Karakas S, Schurmann M. Gamma, alpha, delta, and theta oscillations govern cognitive processes. Int J Psychophysiol. 2001;39: Basar-Eroglu C, Demiralp T, Schurmann M, Basar E. Topological distribution of oddball 'P300' responses. Int J Psychophysiol. 2001;2-3: Brázdil M, Rektor I, Dufek M, Daniel P, Jurák P, Kuba R. The role of frontal and temporal lobes in visual discrimination task - depth ERP studies. Clin Neurophysiol 1999;29: Defebvre L, Bourriez JL, Dujardin K, Derambure P, Destee A, Guieu JD. Spatiotemporal study of Bereitschaftspotential and event-related desynchronization during voluntary movement in Parkinson's disease. Brain Topogr. 1994;3:
91 Dujardin K, Derambure P, Defebvre L, Bourriez JL, Jacquesson JM, Guieu JD. Evaluation of event-related desynchronization (ERD) during a recognition task: effect of attention. Electroenceph clin Neurophysiol 1993;86: Donchin E, Coles MGH. Is the P300 component a manifestation of context updating? Behav. Brain Sci 1988;11: Filipovic SR, Jahanshahi M, Rothwell JC. Uncoupling of contingent negative variation and alpha band event-related desynchronization in go/no-go task. Clin Neurophysiol 2001;112: Fitzgibbon SP, Pope KJ, Mackenzie L, Clark CR, Willoughby JO. Cognitive tasks augment gamma EEG power. Clin Neurophysiol. 2004;8: Goffaux V, Mouraux A, Desmet S, Rossion B. Human non-phase-locked gamma oscillations in experience-based perception of visual scenes. Neurosci Lett. 2004;1:14-7. Gurtubay IG, Alegre M, Labarga A, Malanda A, Iriarte J, Artieda J. Gamma band activity in an auditory oddball paradigm studied with the wavelet transform. Clin Neurophysiol 2001;112: Halgren E, Baudena P, Clarke JM, Heit G, Marinkovic K, Deveaux B, et al. Intracerebral potentials to rare target and distractor auditory and visual stimuli. II. Medial, lateral and posterior temporal lobe. Electroencephal clin Neurophysiol 1995;94: Halgren E, Marinkovic K, Chauvel P. Generators of the late cognitive potentials in auditory and visual oddball tasks. Electroenceph clin Neurophysiol 1998;106: Howard MW, Rizzuto DS, Caplan JB, Madsen JR, Lisman J, Aschenbrenner-Scheibe R, Schulze-Bonhage A, Kahana MJ. Gamma oscillations correlate with working memory load in humans. Cereb Cortex. 2003;12:
92 Kalcher J, Pfurtscheller G. Discrimination between phase-locked and non-phase locked eventrelated EEG activity. Electroenceph clin Neurophysiol 1995;94: Klimesch W. Memory processes, brain oscillations and EEG synchronization. Int J Psychophysiol 1996;24: Krause CM, Salminen PA, Sillanmaki L, Holopainen IE. Event-related desynchronization and synchronization during a memory task in children. Clin Neurophysiol. 2001;12: McCarthy G, Wood CC, Williamson PD, Spencer DD. Task-dependent field potentials in human hippocampal formation. J Neurosci 1989;9: Niedermayer E, Lopes da Silva F. Electroencephylography. Basic Principles, Clinical Applications and Related Fields. 1 st ed. Baltimore: Urban & Schwarzenberg, Nogawa T, Katayama K, Tabata Y, Ohshio T, Kawahara T. Changes in amplitude of the EEG induced by a photic stimulus. Electroenceph clin Neurophysiol 1976;40: Paller KA, Kutas M, Mayes AR. Neural correlates of encoding in an incidental learning paradigm. Electroenceph Clin Neurophysiol 1987;67: Pfurtscheller G, Aranibar A. Event-related cortical desynchronization detected by power measurement of scalp EEG. Electroenceph clin Neurophysiol 1977;42: Pfurtscheller G, Klimesch W. Functional topography during a visuoverbal judgement task studied with event-related desynchronization mapping. J Clin Neurophysiol 1992;9(1): Pfurtscheller G. Event-related synchronization (ERS): an electrophysiological correlate of cortical areas at rest. Electroenceph clin Neurophysiol 1992;83:
93 Pfurtscheller G. Quantification of ERD and ERS in the time domain. In: Pfurtscheller G, da Silva FHL, editors. Handbook of Electroenceph clin Neurophysiol 1999; vol.6: Pfurtscheller G, Lopes da Silva FH. Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol 1999;110: Sederberg PB, Kahana MJ, Howard MW, Donner EJ, Madsen JR. Theta and gamma oscillations during encoding predict subsequent recall. J Neurosci Nov 26;23(34): Sergeant J, Geuze R, Winsum W. Event-Related Desynchronization and P300. Psychophysiology 1987;3: Stancak A Jr, Feige B, Lucking CH, Kristeva-Feige R. Oscillatory cortical activity and movement-related potentials in proximal and distal movements.clin Neurophysiol. 2000;4: Smith ME, Halgen E, Sokolik M, Baudena P, Mussolino A, Liégois-Chauvel C, et al. The intracranial topography of teh P3 event-related potential elicited during auditory oddball. Electroenceph clin Neurophysiol 1990;76: Talairach J, Szikla G, Tournoux P, Prosalentis A, Bordas-Ferrer M, Covello J, et al. Atlas d anatomie stéréotaxique du télencephale. Paris: Masson; Legends Fig.1 Patient No. 5. CPz scalp electrode; thin line averaged non-target response; thick line averaged target response; S - stimulus 93
94 Fig.2 Patient No. 2. Individual ERD/ERS phenomena in analysed frequency bands in the hippocampus; amplitude analysis; the percentage of ERD/ERS is expressed; approximate coordinates in the Talairach axes of x = 30 mm, y= 18 mm, z = 13 mm; S stimulus; arrows indicated maxima of P3 or ERD/ERS Fig.3 The hippocampal ERD/ERS in the theta frequency band from six subjects; power analysis, the percentage of ERD/ERS is expressed; S - stimulus Fig.4 Patient No. 6. Different distribution of P3 and ERD/ERS phenomena across the posterior part of the right-side hippocampus; Talairach s co-ordinates, x = 24 to 33 mm, y = 31 mm, z = 5 mm; S stimulus; arrows indicated maxima of P3 or ERD/ERS Table 1 Patient characteristics. Table 2 Mean peak latencies of ERPs and ERD/ERS phenomena recorded at different anatomical sites across the mesial aspect of the temporal lobe Příloha obrázky a tabulky Fig 1 94
95 Fig 2 95

96 Fig 3 Fig 4 96
97 Table 1 Table 2 97
98 C/ Intracerebral recording of cortical activity related to selfpaced voluntary movements: A Bereitschaftspotential and Event-related de-/synchronization. SEEG study. Running title: SEEG analysis of movement-related potentials and rhythms Daniela Sochůrková 1, Ivan Rektor 1, Pavel Jurák 2, Andrej Stančák 3 1 1st Department of Neurology, Masaryk University, St. Anne s Hospital, Brno, Czech Republic 2 Laboratory of NMR Electronics, Institute of Scientific Instruments, Academy of Science, Brno, Czech Republic 3 Department of Normal, Pathological and Clinical Physiology, Third Faculty of Medicine, Charles University, Prague, Czech Republic Address correspondence to: Daniela Sochůrková, M.D. Department of Neurology, St. Anne s Hospital Pekařská 53 Brno phone: fax: daniela.sochurkova@fnusa.cz 98
99 Aknowledgement We wish to thank Mr. Daniel for technical assistance and Dr. Tyrlík for statistical analysis. Our thanks also go to Dr. Novák and Dr. Chrastina (neurosurgery), and to Dr. Pazourková, Dr. Feitová and Dr. Krupa (neuroradiology). 99
100 Abstract To analyze the distribution of the cortical electrical activity related to self paced voluntary movements, i.e. the movement-related readiness potentials (Bereitschaftspotential, BP) and the event-related desynchronization (ERD) and synchronization (ERS) of cortical rhythms using intracerebral recordings.eeg was recorded in 14 epilepsy surgery candidates during pre-operative video-stereo-eeg monitoring. Subjects performed self-paced hand movements, with their right and left fingers in succession. EEG signals were obtained from a total of 501 contacts using depth electrodes located in primary and non-primary cortical regions. In accordance with previous studies, BP was found consistently in the primary motor (M1) and somatosensory (S1) cortex, the supplementary motor area (SMA), and in a few recordings also in the cingulate cortex and in the dorsolateral prefrontal and premotor cortex. ERD and ERS of alpha and beta rhythms were also observed in these cortical regions. The distribution of contacts showing ERD or ERS was larger than the distribution of those showing BP. In contrast to BP, ERD and ERS frequently occurred in the lateral and mesial temporal cortex and the inferior parietal lobule. The number of contacts and cortical regions showing ERD and ERS and not BP suggests that the two electrophysiological phenomena are differently involved in the preparation and execution of simple voluntary movements. Substantial differences between BP and ERD in spatial distribution and the widespread topography of ERD/ERS in temporal and higher-order motor regions suggest that oscillatory cortical changes are coupled with cognitive processes supporting movement tasks, such as memory, time interval estimation, and attention. Key words: SEEG; Bereitschaftspotential; Event-related desynchronization; Event-related synchronization 100
101 Introduction Voluntary movements are associated with patterned cortical responses manifesting in haemodynamic and electrophysiological measures. Movement-related haemodynamic changes revealed by functional magnetic resonance imaging were found in a number of cortical regions involving the primary sensorimotor cortex, the supplementary motor area, the cingulate cortex, the inferior parietal cortex, the ventral and dorsolateral premotor cortex, and the basal ganglia (Cunnington et al. 2003; Deiber et al. 1999; Sammer et al. 2005; Stancak et al. 2003; Stephan et al. 1999; Toma et al. 2002). Electrical potentials and magnetic fields were observed in similar cerebral structures (Rektor 2000, 2003; Ball et al. 1999; Cui et al. 2000; Pedersen et al. 1998; Rektor et al. 1994, 1998, 2001b; Toma et al. 2002). Electrocortical changes during the movement preparation and execution are of two basic types (Pfurtscheller and Lopes da Silva 1999): i) Phase-locked responses corresponding to the averaged electroencephalographic potentials known as movement-related potentials. The major pre-movement potential is the Readiness Potential, also called the Bereitschaftspotential (Deecke et al. 1969; Kornhuber and Deecke 1965; Neshige et al. 1988; Shibasaki et al. 1980; ). The averaged electrical activity recorded during movement execution is called movement accompanying potential (Rektor et al. 1998) or frontal peak of the motor potential (fpmp) (Toro et al. 1994). ii) Non-phase-locked changes, manifesting in an amplitude decrease of the alpha (Chatrian et al. 1959; Gastaut et al. 1952; Pfurtscheller and Aranibar 1979) and beta rhythms (Pfurtscheller 1981; Jasper and Penfield 1949; Pfurtscheller et al. 1996; Salmelin and Hari 1994; Stancak and Pfurtscheller 1995), followed by a post-movement amplitude increase of beta rhythms 101
102 (Pfurtscheller 1981; Pfurtscheller et al. 1996; Salmelin and Hari 1994; Stancak and Pfurtscheller 1995). In the gamma frequency band, an amplitude increase at the movement onset or offset is reported (Ohara et al. 2001; Pfurtscheller et al. 2003; Szurhaj et al. 2005) embedded in the alpha and/or beta power decrease. The amplitude decrease and increase of cortical rhythms related to an external or internal event is known as event-related desynchronization (ERD) and synchronization (ERS), respectively (Pfurtscheller 1992; Pfurtscheller and Aranibar 1977, 1979). The topography of BP and ERD/ERS revealed both similarities and differences (Defebvre at al. 1994; Derambure et al. 1999; Ohara et al. 2001; Stancak et al. 2000; Toro et al. 1994). In scalp recordings, BP appears more than 1 s prior to the movement over the vertex and over the primary sensorimotor areas, where oscillatory changes are reported to start over the contralateral primary sensorimotor area, and over the ipsilateral sensorimotor area shortly before movement onset. However, comparisons of BP and ERD from surface EEG recordings are limited by the filtering properties of the skull, skin and fluids, which attenuate portions of the signals during their passage from the cortex to the scalp (Nunez et al. 1997; Pfurtscheller and Cooper 1975) and may smear the differences in cortical distribution of BP and ERD. Intracranial recordings reliably showed oscilatory changes in the primary sensorimotor cortex (Crone et al. 1998a, b; Ohara et al. 2000; Toro et al. 1994) and the SMA (Ohara et al. 2000), in which premovement negative potentials also occur (Ohara et al. 2000; Toro et al. 1994), as well as in the basal ganglia (Sochurkova and Rektor 2003). Movement-related cortical potentials and ERD/ERS were also identified in the inferior parietal cortex, the cingulate cortex, and the premotor cortex (Szurhaj et al. 2003). While the evoked (averaged) activity during the movement preparation and execution has been extensively studied in widespread cortical and subcortical structures (for review, see Rektor 2003), the cortical 102
103 distribution of the induced ERD/ERS has remained unexplored in large parts of the cortex. Moreover the mutual relationship of the two phenomena (BP and ERD/ERS) has been studied only in a few locations and remains largely unknown. To shed more light on the distribution of movement-related potentials and the desynchronization and synchronization of cortical rhythms, we analyzed the electrocortical signals obtained using stereo-eeg in a group of patients with intractable epilepsy. We studied the location of BP and ERD/S in the primary sensorimotor cortex, the supplementary motor area (SMA), the higher-order motor areas (dorsal and ventrolateral premotor cortex, cingulate cortex, inferior parietal cortex), the lateral and mesial temporal cortices, and the orbitofrontal cortex. Methods Subjects Fourteen patients (eight males and six females; 11 right-handed, 3 left-handed) ranging in age from 16 to 43 years (mean: 29.1 years), all suffering from intractable epilepsy, participated in the study (Table 1). Using intracerebrally inserted electrodes, recordings were obtained from the patients during their examinations in the diagnostic phase of preparation for epilepsy surgery. These candidates for epilepsy surgery were patients who had remained unresponsive to conventional forms of therapy, and who were recommended by a special commission for stereotactic exploration. A neuropsychological examination excluded cognitive disturbances and dementia in each patient. All the patients had normal motor performance, normal hearing, and normal or corrected-to-normal vision. The experimental procedure was explained to the patients. Informed written consent was obtained from each patient. The experiment was approved by the local ethics committee. 103
104 Experimental procedure Patients rested in a supine position on a bed during the recordings. They were instructed to perform voluntary brisk self-paced fingers flexions with their right or left fingers at irregular intervals ranging from 12 to 15 s. The time estimation of the interval was to be done without counting or otherwise estimating the time. If the movement frequency was high, the patient was asked to increase the interval between the movements. Each subject was given the opportunity to practice movements prior to the recording to obtain a consistent EMG pattern. They were instructed to fix their gaze at a point placed about 2 m in front of them, and to avoid all movement except the fingers movements. Each subject performed at least two blocks of movements with either hand. The duration of one block of movement was 15 minutes. Ten minutes of rest separated successive blocks of movements. In one subject, only right-hand movements were recorded. Recordings Intracerebral depth electrodes were implanted to localize the seizure onset zone prior to the surgical treatment. Between four and seven multicontact MicroDeep (DIXI, Briançon, France) semiflexible platinum electrodes (each with a diameter of 0.8 mm, a contact length of 2 mm, and an intercontact interval of 1.5 mm) were inserted orthogonally in each patient using Talairach s stereotactic co-ordinate system (Talairach et al., 1967;Talairach and Bancaud, 1973; Talairach and Tournoux, 1988). The exact positions of the electrodes and their contacts in the brain were verified with the electrodes in situ using postplacement MRI in each patient. Electrical potentials (bandpass Hz, sampling rate 256 Hz) were recorded in continuous mode from 88 leads (Brain Scope, M&I Prague). In two patients, diagonal electrodes were also used: in accordance with clinical symptomatology and MRI findings, 104
105 these electrodes passed through the occipital regions to the hippocampal formation of both sides or from the frontal region to the hippocampal formation. A total of 501 sites were recorded by means of multicontact depth electrodes: 8 in the primary sensorimotor cortex (BA 1, 4), 8 in the supplementary motor area (BA 8), 51 in the premotor cortex (BA 6), 83 in the dorsolateral prefrontal cortex (BA 9, 45, 46), 64 in the cingulate gyrus (BA 32, 24, 31), 141 in the lateral temporal cortex, 80 in the mesiotemporal structures, 14 in the lobulus parietalis inferior (BA 40), and 52 in the orbitofrontal cortex (BA 11, 47). Surface EEG from the Cpz electrode (10-20 International system) was recorded in nine subjects. Linked ear platinum electrodes were used as a reference. Surface EMG was recorded bipolarly using sintered electrodes placed over the m. flexor digitorum superficialis of both forearms to check the presence of mirror EMG activity. Electrode impedance was 5 kω or less for all electrodes. Data processing EEG data were evaluated off-line. At the beginning of the movement, defined by an EMG onset burst, a marker was inserted during the visual review of each of the individual trials. Only trials containing brisk EMG onset and termination bursts were accepted for further analysis. Trials containing EEG artefacts (spikes, spike-waves, rhythmic activity, and slow waves) were excluded from further analysis. Data were reduced into 10 s epochs: 5 s preceding and 5 s following the onset of EMG activity. The baseline for both BP and ERD/ERS was set between 5 and 3.5 s before the onset of movement. In every session, the trails numbered between 40 and 60. The presence of BP in averaged EEG potentials was visually determined by two independent observers. The BP records were considered for their various features, polarity, duration, and slope values. For the sake of reproducibility, BP had to occur in each of two 105
106 blocks of movements. The BP in scalp recordings is a slow increasing negative potential with the onset appearing between 3.0 and 1.5 s before the movement onset. In intracerebral recordings, due to variances in the positions of the electrode contact and of the dipole generator, the potentials can occur with positive or negative polarities. When the BP exhibits two distinct slopes, the intersection point of these slopes is considered to be the starting point of the second (NS, negative slope; Shibasaki et al. 1980) component. We tried to determine the presence of the two classically described slopes. Further, absolute amplitudes were measured from the baseline. The distance from the electrode to the generator heavily influences the amplitude of intracerebrally recorded potentials, and thus the differences of amplitude can only be compared intraindividually. A clearly larger amplitude in one structure than in its neighbouring structures, particularly an amplitude involving a steep voltage gradient and polarity inversion over a short distance, implied a local BP generator. A steep voltage gradient probably indicated that the electrode was in the immediate vicinity of the generator. A polarity inversion probably indicated that the depth electrode was recording from directly inside the generating tissue. The potentials that had amplitudes that remained unchanged over several consecutive contacts were considered as far-field potentials; these findings were not taken into consideration for further statistical analysis. For each electrode and trial, power spectral densities were computed in sliding 1 s data epochs (256 samples) using the Fast Fourier Transform (FFT). Data were smoothed by the Blackman window prior to the FFT computation. The adjacent 1 s epochs overlapped by 125 ms (32 samples). Eighty-nine spectral estimates representing 4.5 s of the pre-movement period and 4.5 s of the period after the movement onset were obtained for each 10 s trial. Spectral arrays from all trials were then averaged separately for each electrode. The frequency components displaying a significant (P< 0.05) power decrease or increase (Jenkins and Watts, 1968) were identified. A significant power decrease/increase 106
107 was declared if the 95% confidence intervals surrounding both the reference spectrum and the movement spectrum did not overlap. The time courses of absolute power series were computed from absolute power using the reference power as 0% (Pfurtscheller and Aranibar 1977). The frequency bands of interest were 8-12 Hz, Hz, Hz, Hz and Hz. These frequency bands were subject-specific and were determined according to the frequency bands of the subjects. Statistical analysis Our research concerned basically categorical data (the presence or absence of BP or ERD/ERS) obtained from an inequal number of sites in different subjects and cortical regions. Relative frequencies (counts) of positively identified BP, ERD, and ERS in each subject, in each contact, and in each cortical region were submitted for statistical analysis. The relative counts were obtained as the percent number of contacts showing a phenomenon in a particular region. The hypotheses were set in followed way: i. observed movement-related phenomena are lateralised with respect to the side of movement ii. the distribution of observed phenomena differs between explored brain areas The Kruskal-Wallis test was used to test the differences in occurrence of various electrocortical phenomena in contralateral and ipsilateral cortical regions. The Wilcoxonsigned rank test was applied to test the differences in relative counts of BP, ERD, and ERS in different cortical regions. A 95% confidence level was always used. Results 107
108 The EMG burst durations were ± ms (mean ± SD) and ± ms for the dominant and non-dominant hand, respectively. None of the subjects showed any mirror EMG activity, regardless of the type of movement. Figure 1 shows the waveforms of BP and ERD/ERS in different cortical areas in one patient; the contacts are overlaid on the real MR image. The summary of the contacts showing BP and ERD/ERS across the subjects is given in Figure 2. Figure 3 shows the waveforms of BP and ERD/ERS in different cortical areas; the contacts are overlaid on the anatomical MR image. The number of contacts presenting all three types of observed phenomena (BP, alpha ERD/ERS, and beta ERD/ERS) did not differ, whether the movement was performed with contralateral or ipsilateral hand. However, we found significant differences among the nine explored areas for BP in the side contralateral (Kruskal-Wallis test x 2 (d.f.=8)=29.5; p=0.001) and ipsilateral to the movement (Kruskal-Wallis test x 2 (d.f.=8)=26.9; p=0.001). Significant differences did not appear for alpha or beta ERD/ERS. In each explored brain area, the results were found as follows: 3.1. M1/S1 The primary sensorimotor cortex was explored in 2 subjects in a total of 8 contacts (4 contacts in M1, 4 contacts in S1). BP was observed in 4 contacts (1 in M1, 3 in S1, relative count 50%). In contralateral movements, BP was present in the form of a long BP (mean 1.5 s before the movement onset), and in ipsilateral movements in the form of a brief NS (negative slope, 0.5 s before the movement onset). The 8-12 Hz ERD was displayed in all contacts (relative count 100%); the onset was in the interval -1.5 to -1.0 s before the contralateral movement and -0.5 to 0.0 s before the ipsilateral movement. The Hz ERD started at the movement onset and was present in all contacts (relative 108
109 count 100%). The Hz rebound was observed in S1 (relative count 75%). We did not observe beta rebound in M1. However, due to the small number of contacts and electrodes placed in the primary sensorimotor cortex, the presence of ERS could not be reliably evaluated.. Also, due to the small number of patients with electrodes in S1 and M1, a statistical analysis was not performed for these areas. Partial data are shown in Figure SMA The SMA was explored in 3 subjects, in a total of 8 contacts (in one subject, two electrodes with two contacts on each was inserted into SMA). BP was observed in 4 contacts in 2 subjects (relative count 50%) in a form of bilaterally similar negative slope (NS ) or asymmetrical BP (3.0 s before the movement onset in the cortex contralateral to the side of the movement and 1.5 s before the movement onset in the cortex ipsilateral to the side of movement). The 8-12 Hz ERD was observed in 6 contacts in 2 subjects (relative count 75%). It appeared in one subject for each side of the movements; in the other subjects it appeared only during the contralateral movements, starting 1.0 to 0.5 s before the movement onset. The 8-12 Hz ERS was observed in both contralateral and ipsilateral movement in the same subject in whom the 8-12 Hz ERD was observed. The Hz ERD was observed in one subject in whom the 8-12 Hz ERD was also found. The Hz ERS was observed in 4 contacts, in two subjects. The subject with two electrodes placed in the SMA (4 contacts) displayed the Hz ERS only on the site placed in the rostral part of the SMA, while the contact in the caudal part of the SMA displayed only a brief beta decline. Due to the small number of patients and the enormously high intra- and interindividual variability, a statistical analysis was not performed. Partial data are shown in Figure
110 3.3. Premotor cortex The lateral premotor cortex (Brodmann area 6) was explored in 5 subjects, in a total of 51 sites. BP was found in 2 subjects, in 4 contacts (relative count 7.8%) in both contralateral and ipsilateral movements, starting 1.5 to 1.0 s before the movement onset and showing a slight tendency to appear earlier in contralateral movements. The 8-12 Hz ERD was found in 3 subjects. In one subject, the 8-12 Hz ERD was present only in contralateral movements, and in two subjects it was present in both contralateral and ipsilateral movements. The power decline started either equally 1.0 to 0.5 s before the movement onset, or 0.5 s before the movement onset on the side contralateral to the movement, and at movement onset on the side ipsilateral to the movement. The 8-12 Hz ERD and Hz ERD were both observed in 18 contacts in contralateral movements (relative count 35.3%), and in 7 contacts in ipsilateral movements (relative count 13.7%). One of the subjects also showed postmovement Hz ERS, which was equal in contralateral and ipsilateral finger movements. The Hz ERS was observed in 5 contacts in movements of either hand (relative count 9.8%). Statistical analysis did not reveal any significant differences. Partial data are shown in Figure Dorsolateral prefrontal cortex The dorsolateral prefrontal cortex (Brodmann areas 9, 45 and 46) was investigated in 5 subjects (in 3 subjects, the DLPFC in both hemispheres was explored), in a total of 83 sites. BP was found in 5 subjects; in all 5 of them during contralateral movements and in 4 of them during ipsilateral movements. There was a non-significant difference between the number of sites of BP registration in contralateral (20) versus ipsilateral movements (18) (relative count 24.1% and 21.7% respectively); the amplitude and latencies (1.5 to 1.0 s before movement onset) did not differ. ERD/ERS was observed in 2 subjects. The 8-12 Hz ERD was displayed in 35 contacts (relative count 42.2%) in contralateral movements and in 18 contacts (relative 110
111 count 21.7%) in ipsilateral movements; the onset was -0.5 to 0.0 s before the movement onset. The Hz ERD band changes were observed in 13 contacts (relative count 15.7%) in movements of either hand; the power decline started at the movement onset. Statistical analysis did not reveal any significant differences. Partial data are shown in Figure Cingulate gyrus The cingulate gyrus was explored, in its anterior and posterior part, in 12 subjects (in 3 subjects, the cingulate gyrus in both hemispheres was investigated), in a total of 64 sites. BP was present in 4 subjects during contralateral movements (in 7 contacts, relative count 10.9%), and in 3 subjects in ipsilateral movements (in 6 contacts, relative count 9.4%). There were no amplitude or duration differences in the subjects displaying BP in both cingulate gyri. The BP onset varied interindividually, between -2.0 to -0.5 s before the movement onset. ERD/ERS was seen in 5 subjects. The 8-12 Hz ERD was observed in 20 contacts in contralateral movements (relative count 31.3%) and in 18 contacts in ipsilateral movements (relative count 28.1%). The power decline started at the movement onset for both sides of movement Hz ERD was observed in 1 subject in 2 contacts for each side of the movement (relative count 3.1%); ERS in the same frequency band was displayed in 11 contacts for each side of the movement (relative count 17.4%). The onset of beta ERS was in the period s before movement onset on the side ipsilateral to the movement, and during the movement on the side contralateral side to the movement. Statistical analysis did not reveal any significant differences Lateral temporal cortex The lateral temporal cortex (superior, medial and inferior temporal gyrus) was explored in 9 subjects, in a total of 141 sites. BP was not observed. ERD/ERS was observed 111
112 in 7 subjects. The 8-12 Hz ERD was found in 53 contacts in contralateral movements (relative count 37.6%) and in 32 contacts in ipsilateral movements (relative count 22.7%). The power decline started in the interval s before the movement onset for each side of the movement. The Hz ERD was observed in 25 contacts in contralateral movements (relative count 17.7%) and in 20 contacts in ipsilateral movements (relative count 14.2%). The beta ERD started in the same period as alpha ERD. In 2 contacts (relative count 1.4%), a post-movement Hz ERS was also present for movements of either fingers. The statistical analysis showed significant differences in the relative occurrence of the 8-12 Hz ERD and BP, with a greater preponderance toward the 8-12 Hz ERD over the BP in contralateral movements (Wilcoxon sign ranks test z = -2.2, p < 0.031). The contrast in the occurrence of BP and the Hz ERD/ERS in contralateral movements was not significant (z = - 2.0, p < 0.063). In ipsilateral finger movements, the difference in the relative occurrence of BP and the 8-12 Hz ERD/ERS was statistically significant (Wilcoxon sign ranks test z = - 2.2, p < 0.031). The analysis for all three phenomena in this area compared with other areas was also statistically significant in contralateral movement (Friedman test x 2 (d.f.=2)=10.3; p=0.002) Mesiotemporal cortex The mesiotemporal cortex (amygdala, hippocampal formation, and parahippocampal gyrus) was explored in 11 subjects, in a total of 80 contacts. No BP was observed. ERD/ERS was seen in 7 subjects. The 8-12 Hz ERD was found in 26 contacts in contralateral movements (relative count 32.5%) and in 20 contacts in ipsilateral movements (relative count 25.0%). The alpha power decline started in the interval s before the movement onset. The Hz ERD, starting in the same time interval as the alpha power decline for each side 112
113 of movement, was observed in 19 contacts in contralateral movements (relative count 23.8%) and in 14 contacts in ipsilateral movements (relative count 17.5%). The post-movement Hz power increase was observed in 6 contacts after contralateral movements and ipsilateral movements (relative count 7.5%). The statistical analysis revealed significant differences when all three phenomena in the contralateral hand movement were compared with other areas (Friedman test x 2 (d.f.=2)=7.6; p=0.025) Lobulus parietalis inferior The lobulus parietalis inferior (Brodmann area 40) was explored in 3 subjects, in 14 contacts total. BP was not present. ERD/ERS was observed in 2 subjects. The 8-12 Hz and the Hz ERD was found in contralateral and ipsilateral movements in 12 contacts (relative count 85.7%). The onset of decline in both spectra was around 1.0 s before the movement onset in contralateral movements and 0.5 s before the movement in ipsilateral movement. Due to the small number of patients with electrodes in this area, an individual statistical analysis was not performed Orbitofrontal cortex The orbitofrontal cortex (Brodmann area 11, 47) was explored in 5 subjects, in 46 contacts total. BP was not observed. ERD/ERS was seen in 2 subjects. The 8-12 Hz ERD was observed in 7 contacts in contralateral movements (relative count 15.2%) and in 3 contacts in ipsilateral movements (relative count 6.5%). The power decline started at the movement onset in each side of movement. The other frequency bands did not display any oscillatory changes. Due to the small number of patients with electrodes in this area, an individual statistical analysis was not performed. 113
114 3.10. Scalp Cpz The control Cpz electrode (International 20/21 system), or Cz and Cp electrodes, were placed in 9 subjects. BP was seen in 6 subjects, the onset was observed in the interval s before the movement. In all nine subjects, oscillatory changes were observed. The 8-12 Hz ERD was present in 8 of them in movements on each side (relative count 88.9%), the power decline started in the interval s prior to the movement. The Hz ERD was observed in 6 subjects (relative count 66.7%); the power decline onset was observed at the movement onset. In one subject, the Hz ERS was observed starting at the movement onset of each side (relative count 11.1%). Individual statistical analysis was not performed. Discussion Previous studies showed that self-paced finger movements are accompanied by slow negative potentials and amplitude changes of cortical rhythms in various motor cortical regions. This study demonstrates the occurrence of movement-related oscillatory changes in the lateral and mesial temporal cortex and in the orbitofrontal and inferior parietal cortices in addition to the motor cortices. In the non-motor regions the slow potentials were not displayed ERD/ERS and BP in primary sensorimotor cortex and higher-order motor regions As in previous intracerebral studies (Ohara et al. 2000; Szurhaj et al. 2003; Szurhaj et al. 2005), BP and ERD/ERS were observed in the primary sensorimotor cortices and in the SMA. BP and ERD/ERS occurred during contralateral or ipsilateral movements with a preponderance of amplitude and latencies for the contralateral hand movements. The contralateral preponderance of alpha and beta ERD found in our intracerebral recordings is in full accordance with intracranial and scalp EEG studies (Babiloni et al. 1999; 114
115 Pfurtscheller and Aranibar 1979; Pfurtscheller et al. 1997; Salenius et al. 1995; Stancak and Pfurtscheller 1996a, b; Stancak et al. 2000). Gamma band changes in our study we regarded only the Hz frequency band were not found, neither in S1/M1, nor in SMA, nor in extra-motor regions. The number of explored patients with depth electrodes in S1/M1 or in SMA was, however, too small (2 subjects in the former group, 3 subjects in the latter) to reach any conclusions ERD/ERS and BP in higher-order motor regions In addition to the studies dealing primarily with S1/M1 and SMA, we also explored the premotor, dorsolateral prefrontal, orbitofrontal, inferior parietal, and cingulate cortices. BP was seldom observed in the premotor, dorsolateral prefrontal, and cingulate cortices. It was not present in the orbitofrontal and inferior parietal cortices. On the other hand, oscillatory changes, alpha and beta ERD, were observed in all of these cortical structures. Given the known role of higher-order motor regions in cognitive processes, the involvement of these areas might reflect several of these functions. One of them is the time estimation by an internal clock It has been hypothesized (Alexander et al. 1986) that cortical systems with reciprocal pathways to the basal ganglia (supplementary motor area, frontal eye fields, and dorsolateral prefrontal cortex) are candidates supporting time perception, given the role of the of the striatum in interval timing. SMA may directly mediate interval timing (Rao et al. 1997). Further, the cerebelar dentate nucleus, which is also implicated in timing, projects to the DLPFC and premotor cortex (Middleton and Strick 1994). The dorsolateral premotor cortex can support timing because of its putative role in working memory and attention (Posner and Dehaene 1994). The inferior parietal cortex might also mediate time perception (Poizner et al. 1995). 115
116 It seems possible that the activation of higher-order motor regions reflected as oscillatory changes could be at least partly explained by the requirements of attention, working memory, and timing in performed task ERD/ERS and BP in temporal cortices BP was absent in the lateral and mesial temporal cortex. In contrast, the occurrence of alpha and beta ERD were statistically significant in these regions. Any oscillatory changes were found in the explored gamma band Hz. The ERD and ERS occurred bilaterally in the temporal cortex during unimanual movements, and the relative occurrence of ERD/ERS in the ipsilateral and contralateral cortices was similar in spite of a mild contralateral preponderance. The term self-paced obviously does not mean spontaneous. The process is self-paced because no external cue is necessary to trigger it. The experimental findings led to conclusions that voluntary acts can be initiated by unconscious cerebral processes before conscious intention appears. According to Libet, the conscious volitional control may operate not to initiate the volitional process but to select and control it, either by permitting or trigerring the final motor outcome of the unconsciously initiated process or by vetoing the progression to actual motor activation (Libet 1985). The long unconscious period of BP is associated with preparation to act in the near future, while the process with an onset 0.5 sec before an act is associated with voluntary choice and the endogenous urge to act. It has been shown (Libet et al. 1982) that the processes assotiated with attention, cognition and expectancy could not solely account for BP. The BP probably reflects a goal directed process; i.e. it is related directly to the execution of movement. Nevertheless, several processes occur in the premovement period that are not necessarily goal directed. Self-paced movement is preceded by the measurement of the inter-movement period by an internal clock, 116
117 by the decisional process to move, and also by the preparation and the initiation for the movement itself (Rektor et al. 2001a). Mesiotemporal structures, with their memory functions, including the internal estimation of time (Drane et al. 1999; Kesner et al. 2001; Perbal et al. 2001; Vodolazhskaia et al. 2001) appear to be involved in these processes. The absence of BP in temporal structures indicates that ERD/ERS reflects other functions, which are related probably not to the movement itself but to other aspects of the task. In contrast to the BP, the ERD and ERS in mesiotemporal structures could reflect the memory functions involved in repetitive movement; oscillatory changes were found over the temporal regions in scalp studies in memory tasks with an attentional load (Klimesch et al. 1996). The bilateral distribution of power changes in the temporal lobes suggests a large contribution of extramotor regions even in a simple motor task. Anatomical studies reveal that the temporal cortex might act in co-operation with the motor regions. The functional human tractography showed connections between the primary motor cortex and the posterior superior and middle temporal gyri in the left hemisphere in right-handed subjects (Guye et al. 2003). Even with the known limitations of fmri, the results of the above-mentioned study could support our data concerning the functional cooperation between temporal structures and primary motor cortex General discussion Our results could be biased by two phenomena typical for intracerebral studies. The first is the low spatial resolution of intracerebral recordings. This is compensated in our studies by the high number of recording sites in the regions of interests. The intracranial explorations in human subjects are targeted according to clinical data and some regions remained insufficiently explored in our study, namely the motor areas. However, these regions have been already explored by other authors (Ohara et al. 2000; Pfurtscheller et al. 2003; Szurhaj et al. 2003, 2005). On the other hand, our study could present the results from 117
118 regions with no reported recordings in previous studies. The second possible bias is the fact that our recordings were performed in epileptic patients with epileptic foci. Though the recordings from epileptic tissue were excluded from evaluations, there still remains the question of a possible influence of cortical excitability in subjects with epilepsy. We could imagine that the presence of ERD and ERS in the temporal lobe during movement might be influenced by the increased excitability in the temporal cortex in some patients. Although this possibility cannot be fully excluded, a previous ERD study showed normal ERD alpha-erd over the peri-rolandic in patients with temporal lobe epilepsy (Derambure et al. 1997). The influence of epilepsy on the presence of ERD and ERS in the temporal lobe during voluntary movements is also improbable because our recordings were obtained from the temporal lobe ipsilateral as well as contralateral to the epileptogenic zone, and no difference was observed. To conclude, phase-locked and non-phase-locked movement-related electrophysiological responses may share common neurophysiological mechanisms (Derambure et al. 1999). However, according to our data, if this is so, it is only in some cortical regions. While the BP is tightly related to the motor control areas, ERD/ERS represent broader physiological aspects of motor execution and control. The evoked electrical activity probably reflects the regional activation, while the more widespread ERD/ERS may reflect the spread of task relevant information across relevant areas. The substantial differences between BP and ERD in spatial distribution, and especially the widespread topography of ERD/ERS in temporal and higher-order motor regions, suggest that oscillatory cortical changes could be related to cognitive processes supporting movement tasks, such as memory, time interval estimation, and attention (Dujardin et al. 1993). The timing of the studied phenomena indicates that both of them cover the preparation as well as the execution of the movement. The ERD/ERS as well as the BP could be recorded during the unconscious as well as conscious parts of movement preparation. The cortico-subocortical circuitries of 118
119 two studied electrical phenomena are probably different. While the oscillatory changes - at least in the alpha frequency band - depend mainly on thalamo-cortical feedback loop (Lopes da Silva 1991; Steriade and Llinas 1988), the BP is generated in a number of subcortical structures cerebellum, basal ganglia, and thalamus (Rektor et al. 2001b; Shibasaki et al. 1986; Tarkka et al. 1993). Our results are in concordance with the view that both primary sensorimotor and supplementary motor areas are involved in the preparation, initiation, and execution of simple unilateral volitional movement (Ikeda et al. 1992; Urbano et al. 1996; Urbano et al. 1998). References Alexander GE, DeLong MR, Strick PL (1986) Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 9: Babiloni C, Carducci F, Cincotti F, Rossini PM, Neuper C, Pfurtscheller G, Babiloni F (1999) Human movement-related potentials vs desynchronization of EEG alpha rhythm: a high-resolution EEG study. Neuroimage 6: Chatrian GE, Petersen MC, Lazarte JA (1959) The blocking of the rolandic wicket rhythm and some central changes related to movement. Electroencephalogr Clin Neurophysiol Suppl.3: Cunnington R, Windischberger C, Deecke L, Moser E (2003) The preparation and readiness for voluntary movement: a high-field event-related fmri study of the Bereitschafts- BOLD response. Neuroimage 20:
120 Crone N, Miglioretti D, Gordon B, Sieracki J, Wilson M, Uematsu S, Lesser R (1998a) Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. I. Alpha and beta event-related desynchronization. Brain 121: Crone N, Miglioretti D, Gordon B, Lesser R (1998b) Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. II. Event-related synchronization in the gamma band. Brain 121: Cui RQ, Huter D, Egkher A, Lang W, Lindinger G, Deecke L (2000) High resolution DC- EEG mapping of the Bereitschaftspotential preceding simple or complex bimanual sequential finger movement. Exp Brain Res 134:49-57 Deecke L, Scheid P, Kornhuber HH (1969) Distribution of readiness potential, pre-motion positivity, and motor potential of the human cerebral cortex preceding voluntary finger movements. Exp Brain Res 2: Defebvre L, Bourriez JL, Dujardin K, Derambure P, Destee A, Guieu JD (1994) Spatiotemporal study of Bereitschaftspotential and event-related desynchronization during voluntary movement in Parkinson's disease. Brain Topogr. 3: Deiber M-P, Honda M, Ibanez V, Sadato N, Hallett M (1999) Mesial motor area in selfinitiated versus externally triggered movements examined with fmri: effect of movement type and rate. J Neurophysiol 81: Derambure P, Bourriez JL, Defebvre L, Cassim F, Josien E, Duhamel A, Destée A, Guieu JD (1997) Abnormal cortical activation during planning of voluntary movement in patients with epilepsy with focal motor seizures: Event-related desynchronization study of electroencephalographic mu rhythm. Epilepsia 38: Derambure P, Defebvre L, Bourriez JL, Cassim F, Guieu JD (1999) Event-related desynchronization and synchronization. Reactivity of electrocortical rhythms in 120
121 relation to the planning and execution of voluntary movement. Neurophysiol Clin 29:53-70 Drane DL, Lee GP, Loring DW, Meador KJ (1999) Time perception following unilateral amobarbital injection in patients with temporal lobe epilepsy. J Clin Exp Neuropsychol 21: Dujardin K, Derambure P, Defebvre L, Bourriez JL, Jacquesson JM, Guieu JD (1993) Evaluation of event-related desynchronization (ERD) during a recognition task: effect of attention. Electroenceph Clin Neurophysiol 86: Gastaut H (1952) Electrocorticographic study of the reactivity of rolandic rhythm. Rev Neurol 87: Guye M, Parker GJ, Symms M, Boulby P, Wheeler-Kingshott CA, Salek-Haddadi A, Barker GJ, Duncan JS (2003) Combined functional MRI and tractography to demonstrate the connectivity of the human primary motor cortex in vivo. Neuroimage 19: Ikeda A, Luders HO, Burgess RC, Shibasaki H (1992) Movement-related potentials recorded from supplementary motor area and primary motor area. Role of supplementary motor area in voluntary movements. Brain 115: Jasper HH, Penfield W (1949) Electrocorticograms in man: effect of voluntary movement upon the electrical activity of the precentral gyrus. Arch Psychiatr. Z. Ges. Neurol 183: Jenkins GM and Watts DG (1968) Spetral Analysis and its Applications. Holden-Day, San Francisco Kesner RP, Hopkins RO (2001) Short-term memory for duration and distance in humans: role of the hippocampus. Neuropsychology 15:58-68 Klimesch W, Doppelmayr M, Russegger H, Pachinger T (1996) Theta band power in the human scalp EEG and the encoding of new information. Neuroreport 7: Kornhuber HH, Deecke L (1965) Hirnopotentialänderung bei 121
122 Willkürbewegungen und passiven Bewegungen des Menschen: Bereitschaftspotential und reafferente Potentiale. Plügers Arch 284:1-17 Libet B, Wright EW Jr, Gleason CA (1982) Readiness-potentials preceding unrestricted "spontaneous" vs. pre-planed voluntary acts. Electroencephalogr Clin Neurophysiol 54: Libet B (1985) Unconscious cerebral initiative and the role of conscious will in voluntary action. Behav Brain Sci 8: Lopes da Silva F (1991) Neural mechanisms underlying brain waves: from neural membranes to networks. Electroencep Clin Neurophysiol 79: Middleton FA, Strick PL (1994) Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 21: Neshige R, Lüders H, Shibasaki H (1988) Recording of movement-related potential from scalp and cortex in man. Brain 111: Nunez P, Srinivasan R, Westdorp A, Wijesinghe R, Tucker D, Silberstein R, Cadusch P (1997) EEG coherency. I: Statistics, reference electrode, volume conduction, Laplacians, cortical imaging, and interpretation at multiple scales. Electroencephalogr Clin Neurophysiol 103: Ohara S, Ikeda A, Kunida T, Yazawa S, Baba K, Nogamine T, Taki W, Hashimoto N, Mihara T, Shibasaki (2000) Movement-related change of electrocorticographic activity in human supplementary motor area proper. Brain 123: Ohara S, Mima T, Baba K, Ikeda A, Kunieda T, Matsumoto R, Yamamoto J, Matsuhashi M, Nagamine T, Hirasawa K, Hori T, Mihara T, Hashimoto N, Salenius S, Shibasaki H (2001) Increased synchronization of cortical oscillatory activities between human supplementary motor and primary sensorimotor areas during voluntary movements. J Neurosci 23:
123 Pedersen JR, Johannsen P, Bak CK, Kofoed B, Saermark K, Gjedde A (1998) Origin of human motor readiness field linked to left middle frontal gyrus by MEG and PET. Neuroimage 8: Perbal S, Ehrle N, Samson S, Baulac M, Pouthas V (2001) Time estimation in patients with right or left medial-temporal lobe resection. Neuroreport 5: Pfurtscheller G (1981) Central beta rhythm during sensorimotor activities in man. Electroencephalogr Clin Neurophysiol. 3: Pfurtscheller G (1992) Event-related synchronization (ERS): an electrophysiological correlate of cortical areas at rest. Electroenceph clin Neurophysiol 83:62-69Pfurtscheller G, Cooper R (1975) Frequency dependence of the transmission of the EEG from cortex to scalp. Electroencephalogr Clin Neurophysiol 38:93-96 Pfurtscheller G, Aranibar A (1977) Event-related cortical desynchronization detected by power measurements of the scalp EEG. Electroenceph clin Neurophysiol 42: Pfurtscheller G, Aranibar A (1979) Evaluation of event-related desynchronization (ERD) preceding and following voluntary self-paced movement. Electroencephalogr Clin Neurophysiol 2: Pfurtscheller G, Stancak A Jr, Neuper C (1996) Post-movement beta synchronization. A correlate of an idling motor area? Electroencephalogr Clin Neurophysiol 4: Pfurtscheller G, Neuper C, Andrew C, Edlinger G (1997) Foot and hand area mu rhythms. Int J Psychophysiol 26: Pfurtscheller G, Lopes da Silva FH (1999) Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol 11: Pfurtscheller G, Graimann B, Huggins JE, Levine SP, Schuh LA (2003) Spatiotemporal patterns of beta desynchronization in corticographic data during self-paced movement. Clin Neurophysiol 7:
124 Poizner H, Clark MA, Merians AS, Macauley B, Gonzalez Rothi LJ, Heilman KM (1995) Joint coordination deficits in limb apraxia. Brain 118: Posner MI, Dehaene S (1994) Attentional networks. Trends Neurosci 17:75-79 Rao SM, Bobholz JA, Hammeke TA, Rosen AC, Woodley SJ, Cunningham JM, Cox RW, Stein EA, Binder JR (1997) Functional MRI evidence for subcortical participation in conceptual reasoning skills. Neuroreport 8: Rektor I (2000) Parallel information processing in motor systems: intracerebral recordings of readiness potential and CNV in human subjects. Neural Plasticity 7:65-72 Rektor I (2003) Intracerebral recordings of the Bereitschaftspotential and related potentials in cortical and subcortical structures in human subjects. In: Eds. M. Jahanshahi and M. Hallett. The Bereitschaftspotential. Kluwer Academic/Plenum Publishers, Rektor I, Feve A, Buser P, Bathien N, Lamarche M (1994) Intracerebral recording of movement realted readiness potentials: an exploration in epileptic patients. Electroenceph Clin Neurophysiol 90: Rektor I, Louvel J, Lamarche M (1998) Intracerebral recording of potentials accompanying simple limb movements: a SEEG study in epileptic patients. Electroenceph Clin Neurophysiol 107: Rektor I, Bareš M, Kanovský P, Kukleta M (2001a) Intracerebral recording of readiness potential induced by a complex motor task. Movement Disorders 16: Rektor I, Kanovský P, Bareš M, Louvel J, Lamarche M (2001b) Evoked potentials, ERP, CNV, readiness potential, and movement accompanying potential recorded from the posterior thalamus in human subjects. A SEEG study. Neurophysiologie Clinique/Clinical Neurophysiology 31:
125 Salenius S, Kajola M, Thompson WL, Kosslyn S, Hari R (1995) Reactivity of magnetic parieto-occipital alpha rhythm during visual imagery. Electroencephalogr Clin Neurophysiol 6: Salmelin R Hari R (1994) Spatiotemporal characteristics of sensorimotor neuromagnetic rhythms related to thumb movement. Neuroscience 2: Sammer G, Blecker C, Gebhardt H, Kirsch P, Stark R, Vaitl D (2005) Acquisition of typical EEG waveforms during fmri: SSVEP, LRP, and frontal theta. Neuroimage 24: Shibasaki H, Barret G, Halliday E, Halliday AM (1980) Components of the movement-related cortical potential and their scalp topography. Electroenceph Clin Neurophysiol 48: Shibasaki H, Barrett G, Neshige R, Hirata I, Tomoda H (1986) Volitional movement is not preceded by cortical slow negativity in cerebellar dentate lesion in man. Brain Res 19: Sochurkova D, Rektor I (2003) Event-related desynchronization/synchronization in the putamen. An SEEG study. Exp Brain Res 3: Stancak A (2000) The electroencephalographic beta synchronization following extension and flexion finger movements in humans. Neuroscience Letters Stancak A Jr, Pfurtscheller G (1995) Desynchronization and recovery of beta rhythms during brisk and slow self-paced finger movements in man. Neurosci Lett 1-2:21-24 Stancak A Jr, Pfurtscheller G (1996a) Mu-rhythm changes in brisk and slow self-paced finger movements. Neuroreport 6: Stancak A Jr, Pfurtscheller G (1996b) Event-related desynchronisation of central betarhythms during brisk and slow self-paced finger movements of dominant and nondominant hand. Brain Res Cogn Brain Res 3:
126 Stancak A, Cohen ER, Seidler RD, Duong TQ, Kim SG (2003) The size of corpus callosum correlates with functional activation of medial motor cortical areas in bimanual and unimanual movements. Cereb Cortex 13: Stephan KM, Binkofski F, Halsband U, Dohle C, Wunderlich G, Schnitzler A, Tass P, Posse S, Herzog H, Sturm V, Zilles K, Seitz RJ, Freund HJ (1999) The role of ventral medial wall motor areas in bimanual co-ordination. Brain 122: Steriade M, Llinas RR (1988) The functional states of the thalamus and associated neuronal interplay. Physiol Rev 68: Szurhaj W, Derambure P, Labyt E, Cassim F, Bourriez JL, Isnard J, Guieu JD, Mauguiere F (2003) Basic mechanisms of central rhythms reactivity to preparation and execution of a voluntary movement: a stereoelectroencephalographic study. Clin Neurophysiol 1: Szurhaj W, Bourriez JL, Kahane P, Chauvel P, Mauguiere F, Derambure P (2005) Intracerebral study of gamma rhythm reactivity in the sensorimotor cortex. Eur J Neurosci 5: Talairach J, Szikla G, Tournoux P, Prosalentis A, Bordas-Ferrer M, Covello J (1967) Atlas d anatomie stéréotaxique du télencephale. Paris: Masson Talairach J, Bancaud J (1973) Stereotaxic approach to epilepsy. Methodology of anatomofunctional stereotaxic investigation. Prog Neurol Surg 5: Talairach J, Tournoux P (1988) Co-Planar Stereotaxic Atlas of the Human Brain. Georg Thieme Verlag, Stuttgart, p. 122 Tarkka IM, Massaquoi S, Hallett M (1993) Movement-related cortical potentials in patients with cerebellar degeneration. Acta Neurol Scand 88: Toma K, Matsuoka T, Immisch I, Mima T, Waldvogel D, Koshy B, Hanakawa T, Shill H, Hallett M (2002) 126
127 Generators of movement-related cortical potentials: fmri-constrained EEG dipole source analysis. Neuroimage 17: Toro C, Deuschle G, Thatcher R, Sato S, Kufta C, Hallett M (1994) Event-related desynchronization and movement-related cortical potentials on the ECoG and EEG. Electroenceph Clin Neurophysiol 93: Urbano A, Babiloni C, Onorati P, Babiloni F (1996) Human cortical activity related to unilateral movements. A high resolution EEG study. NeuroReport 8: Urbano A, Babiloni C, Onorati P, Carducci F, Ambrosini A, Fattorini L, Babiloni F (1998) Responses of human primary sensorimotor and supplementary motor areas to internally-triggered unilateral and simultaneous bilateral one-digit movements. A high resolution EEG study. European Journal of Neuroscience 40: Vodolazhskaia MG, Beier EV (2001) Impairment of internal estimation time and emotional behaviour in rats with lesioned striatum and hippocampus Ross Fiziol Zh Im I M Sechenova87:
128 Legends Figure 1 BP and ERD waveforms in selected cortical regions during self-paced right- and left finger movements in patient No. 13. All MRI scans with electrodes placed in the right hemisphere. ACG anterior cingulate gyrus. BA 6 Brodmann area 6. MTC mesial temporal cortex. LTC lateral temporal cortex. Timescale of waveforms 10 seconds (-5/+5 sec), baseline zero at the movement onset. ERD/ERS is in the upper part of each picture: on the left side with ipsilateral movements and on the right side with contralateral movements towards the electrode placement. Thick line ERD/ERS in 8-12 Hz frequency band. Dotted thin line ERD/ERS in Hz frequency band. ERD/ERS expressed in relative value (in percentage of power changes with respect to the baseline). BP potentials are in the lower part of each picture: on the left side with ipsilateral movements and on the right side with contralateral movements towards the electrode placement. Absolute value of BP (in uv; with respect to the baseline). Figure 2 Distribution of BP (red circles), alpha-erd (green circles), and beta ERD and/or ERS (blue circles) in subjects. Full circles mean the presence of observed phenomena; and empty circles indicate the absence. The circles are a schematic representation in Talairach s brain. Figure 3 Distribution of BP and ERD in different cortical regions in contralateral and ipsilateral finger movements. Three-dimensional MRI reconstruction scans. M1/S1 primary sensorimotor cortex. DLPFC dorsolateral prefrontal cortex. 128
129 IPL inferior parietal lobulus. ACG anterior cingulate gyrus. BA 9, 11 Brodmann areas 9 and 11. SMA supplementary motor area. Timescale of waveforms 10 seconds (-5/+5 sec), zero at the movement onset. The ERD/ERS is in the upper part of each picture: on the left side with contralateral movements and on the right side with ipsilateral movements towards the side of the SEEG registration. Thick line ERD/ERS in 8-12 Hz frequency band. Dotted thin line ERD/ERS in Hz frequency band. ERD/ERS expressed in relative value (in percentage of power changes with respect to the baseline). BP potentials are in the lower part of each picture: on the left side with contralateral movements and on the right side with ipsilateral movements towards the side of the SEEG registration. Absolute value of BP (in uv; with respect to the baseline). Table 1 T CPS temporal complex partial seizure; F CPS frontal complex partial seizure; ATMR anteromedial temporal resection; LE - lesionectomy; MTS mesiotemporal sclerosis; FCD focal cortical dysplazia; M male; F female; R right; L left Table 2 M1/S1 primary sensorimotor cortex. SMA supplementary motor area. BA 6 Brodmann area 6 (premotor cortex). DLPFC dorsolateral prefrontal cortex. CG cingulate gyrus. LTC lateral temporal cortex. MTC mesial temporal cortex. IPL inferior parietal lobulus. OFC orbitofrontal cortex. Cpz centroparietal scalp electrode (international 10/20 system; No s/c number of subjects/contacts; α contra alpha ERD contralateral to the movement; α ipsi - alpha ERD ipsilateral to the movement; β contra - beta ERD contralateral to the 129

130 movement; β ipsi - beta ERD ipsilateral to the movement; n.p. not performed; n.s. not statistically significant Fig 1 130
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti EEG - elektroencefalogram Skalpové EEG Intrakraniální EEG > 1 cm < 1 cm Lokální potenciály Extracelulární akční potenciály ~ 1 mm ~ 1 um EEG - elektroencefalogram
Elektrofyziologické metody a studium chování a paměti EEG - elektroencefalogram Skalpové EEG Intrakraniální EEG > 1 cm < 1 cm Lokální potenciály Extracelulární akční potenciály ~ 1 mm ~ 1 um EEG - elektroencefalogram
Elektroencefalografie
 Elektroencefalografie EEG vzniká součinností neuronů thalamu a kortexu thalamus - funkce generátoru rytmů hlavní zdroj EEG - elektrická aktivita synaptodendritických membrán v povrchních vrstvách kortexu
Elektroencefalografie EEG vzniká součinností neuronů thalamu a kortexu thalamus - funkce generátoru rytmů hlavní zdroj EEG - elektrická aktivita synaptodendritických membrán v povrchních vrstvách kortexu
VYŠETŘENÍ NERVOVÉHO SYSTÉMU. seminář z patologické fyziologie
 VYŠETŘENÍ NERVOVÉHO SYSTÉMU seminář z patologické fyziologie Osnova Morfologické vyšetřovací metody (zobrazovací diagnostika) 1 Počítačová (výpočetní) tomografie 2 Pozitronová emisní tomografie (PET) 3
VYŠETŘENÍ NERVOVÉHO SYSTÉMU seminář z patologické fyziologie Osnova Morfologické vyšetřovací metody (zobrazovací diagnostika) 1 Počítačová (výpočetní) tomografie 2 Pozitronová emisní tomografie (PET) 3
10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální
 10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální iktální periodické Evokované potenciály sluchové (AEP) zrakové
10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální iktální periodické Evokované potenciály sluchové (AEP) zrakové
SIMULTÁNNÍ EEG-fMRI. EEG-fMRI. Radek Mareček MULTIMODÁLNÍ FUNKČNÍ ZOBRAZOVÁNÍ. EEG-fMRI. pozorování jevu z různých úhlú lepší pochopení
 SIMULTÁNNÍ Radek Mareček MULTIMODÁLNÍ FUNKČNÍ ZOBRAZOVÁNÍ pozorování jevu z různých úhlú lepší pochopení některé jevy jsou lépe pozorovány pomocí jedné modality, pozorovatele však zajímá informace obsažená
SIMULTÁNNÍ Radek Mareček MULTIMODÁLNÍ FUNKČNÍ ZOBRAZOVÁNÍ pozorování jevu z různých úhlú lepší pochopení některé jevy jsou lépe pozorovány pomocí jedné modality, pozorovatele však zajímá informace obsažená
Etiologie epilepsie. Epilepsie nevychází z centra jizvy nebo postmalatické pseudocysty, ale spíše z jejího okraje, kde přežívají poškozené neurony.
 Epilepsie Epilepsie Chronické mozkové onemocnění charakterizované opakujícím se výskytem (nevyprovokovaných) epileptických záchvatů. Ojedinělý epileptický záchvat může být vyprovokován i u člověka bez
Epilepsie Epilepsie Chronické mozkové onemocnění charakterizované opakujícím se výskytem (nevyprovokovaných) epileptických záchvatů. Ojedinělý epileptický záchvat může být vyprovokován i u člověka bez
Vlastnosti neuronových sítí. Zdeněk Šteffek 2. ročník 2. LF UK v Praze
 Vlastnosti neuronových sítí Zdeněk Šteffek 2. ročník 2. LF UK v Praze 7. 3. 2011 Obsah Neuronální pooly Divergence Konvergence Prolongace signálu, kontinuální a rytmický signál Nestabilita a stabilita
Vlastnosti neuronových sítí Zdeněk Šteffek 2. ročník 2. LF UK v Praze 7. 3. 2011 Obsah Neuronální pooly Divergence Konvergence Prolongace signálu, kontinuální a rytmický signál Nestabilita a stabilita
Spánek. Neurobiologie chování a paměti. Eduard Kelemen. Národní ústav duševního zdraví, Klecany
 Spánek Neurobiologie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Spánek Spánková stadia a architektura spánku Role spánku při konsolidaci paměti behaviorální studie Role spánku
Spánek Neurobiologie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Spánek Spánková stadia a architektura spánku Role spánku při konsolidaci paměti behaviorální studie Role spánku
Bioelektromagnetismus. Zdeněk Tošner
 Bioelektromagnetismus Zdeněk Tošner Bioelektromagnetismus Elektrické, elektromagnetické a magnetické jevy odehrávající se v biologických tkáních elektromagnetické vlastnosti tkání chování vzrušivých tkání
Bioelektromagnetismus Zdeněk Tošner Bioelektromagnetismus Elektrické, elektromagnetické a magnetické jevy odehrávající se v biologických tkáních elektromagnetické vlastnosti tkání chování vzrušivých tkání
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti RNDr. Eduard Kelemen, PhD Národní ústav duševního zdraví, Klecany Elektrofyziologické metody studia chování a paměti EEG a LFP théta, gama, pomalé
Elektrofyziologické metody a studium chování a paměti RNDr. Eduard Kelemen, PhD Národní ústav duševního zdraví, Klecany Elektrofyziologické metody studia chování a paměti EEG a LFP théta, gama, pomalé
Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie
 Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie Záznam elektrické aktivity mozku Buněčné mechanizmy vzniku Principy registrace Základní vlastnosti Experimentální využití
Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie Záznam elektrické aktivity mozku Buněčné mechanizmy vzniku Principy registrace Základní vlastnosti Experimentální využití
Elektroencefalografie. X31LET Lékařská technika Jan Havlík Katedra teorie obvodů
 Elektroencefalografie X31LET Lékařská technika Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Elektroencefalografie diagnostická metoda, umožňující snímání a záznam elektrické aktivity mozku invazivní
Elektroencefalografie X31LET Lékařská technika Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Elektroencefalografie diagnostická metoda, umožňující snímání a záznam elektrické aktivity mozku invazivní
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti (EEG) oscilace v mozku Théta rytmus Gamma rytmus Sharp waves ripples Pomalé oscilace Spánková vřetena Hans Berger První nahrávání EEG u lidí (1924)
Elektrofyziologické metody a studium chování a paměti (EEG) oscilace v mozku Théta rytmus Gamma rytmus Sharp waves ripples Pomalé oscilace Spánková vřetena Hans Berger První nahrávání EEG u lidí (1924)
ZPRACOVÁNÍ A ANALÝZA BIOSIGNÁLŮ V. Institut biostatistiky a analýz
 ZPRACOVÁNÍ A ANALÝZA BIOSIGNÁLŮ V. ELEKTROENCEFALOGRAM ELEKTROENCEFALOGRAM ELEKTROENCEFALOGRAM (EEG) je (grafická) reprezentace časové závislosti rozdílu elektrických potenciálů, snímaných z elektrod umístěných
ZPRACOVÁNÍ A ANALÝZA BIOSIGNÁLŮ V. ELEKTROENCEFALOGRAM ELEKTROENCEFALOGRAM ELEKTROENCEFALOGRAM (EEG) je (grafická) reprezentace časové závislosti rozdílu elektrických potenciálů, snímaných z elektrod umístěných
Elektroencefalografie. X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů
 Elektroencefalografie X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Elektroencefalografie diagnostická metoda, umožňující snímání a záznam elektrické aktivity
Elektroencefalografie X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Elektroencefalografie diagnostická metoda, umožňující snímání a záznam elektrické aktivity
Zpracování informace v NS Senzorická fyziologie
 Zpracování informace v NS Senzorická fyziologie doc. MUDr. Markéta Bébarová, Ph.D. Fyziologický ústav, Lékařská fakulta, Masarykova univerzita Tato prezentace obsahuje pouze stručný výtah nejdůležitějších
Zpracování informace v NS Senzorická fyziologie doc. MUDr. Markéta Bébarová, Ph.D. Fyziologický ústav, Lékařská fakulta, Masarykova univerzita Tato prezentace obsahuje pouze stručný výtah nejdůležitějších
8. PŘEDNÁŠKA 20. dubna 2017
 8. PŘEDNÁŠKA 20. dubna 2017 EEG systém rozložení elektrod 10/20 základní typy zapojení požadavky na EEG přístroj analýza EEG a způsoby zobrazení ontogeneze normální EEG úvod ke cvičení montáž, filtrace,
8. PŘEDNÁŠKA 20. dubna 2017 EEG systém rozložení elektrod 10/20 základní typy zapojení požadavky na EEG přístroj analýza EEG a způsoby zobrazení ontogeneze normální EEG úvod ke cvičení montáž, filtrace,
Senzorická fyziologie
 Senzorická fyziologie Čití - proces přenosu informace o aktuálním stavu vnitřního prostředí a zevního okolí do formy signálů v CNS Vnímání (percepce) - subjektivní vědomá interpretace těchto signálů na
Senzorická fyziologie Čití - proces přenosu informace o aktuálním stavu vnitřního prostředí a zevního okolí do formy signálů v CNS Vnímání (percepce) - subjektivní vědomá interpretace těchto signálů na
Elektroencefalografie v intenzivní péči. Petr Aulický
 Elektroencefalografie v intenzivní péči Petr Aulický Elektroencefalografie (EEG) Metoda pro vyšetření mozkové elektrické aktivity Hlavní význam v epileptologii Pomocná metoda nutná korelace s klinickým
Elektroencefalografie v intenzivní péči Petr Aulický Elektroencefalografie (EEG) Metoda pro vyšetření mozkové elektrické aktivity Hlavní význam v epileptologii Pomocná metoda nutná korelace s klinickým
Laboratorní úloha č. 8: Elektroencefalogram
 Laboratorní úloha č. 8: Elektroencefalogram Cíle úlohy: Rozložení elektrod při snímání EEG signálu Filtrace EEG v časové oblasti o Potlačení nf a vf rušení o Alfa aktivita o Artefakty Spektrální a korelační
Laboratorní úloha č. 8: Elektroencefalogram Cíle úlohy: Rozložení elektrod při snímání EEG signálu Filtrace EEG v časové oblasti o Potlačení nf a vf rušení o Alfa aktivita o Artefakty Spektrální a korelační
SIMULTÁNNÍ EEG-fMRI. Radek Mareček
 SIMULTÁNNÍ EEG-fMRI Radek Mareček MULTIMODÁLNÍ FUNKČNÍ ZOBRAZOVÁNÍ pozorování jevu z různých úhlú lepší pochopení některé jevy jsou lépe pozorovány pomocí jedné modality, pozorovatele však zajímá informace
SIMULTÁNNÍ EEG-fMRI Radek Mareček MULTIMODÁLNÍ FUNKČNÍ ZOBRAZOVÁNÍ pozorování jevu z různých úhlú lepší pochopení některé jevy jsou lépe pozorovány pomocí jedné modality, pozorovatele však zajímá informace
Analýzy intrakraniálního EEG signálu
 BSG 2018 Analýzy intrakraniálního EEG signálu Ing. Radek Janča, Ph.D. jancarad@fel.cvut.cz Fakulta elektrotechnická České vysoké učení technické v Praze Česká republika Analýzy ieeg signálu 2/38 Epilepsie
BSG 2018 Analýzy intrakraniálního EEG signálu Ing. Radek Janča, Ph.D. jancarad@fel.cvut.cz Fakulta elektrotechnická České vysoké učení technické v Praze Česká republika Analýzy ieeg signálu 2/38 Epilepsie
CW01 - Teorie měření a regulace
 Ústav technologie, mechanizace a řízení staveb CW01 - Teorie měření a regulace ZS 2010/2011 SPEC. 2.p 2010 - Ing. Václav Rada, CSc. Ústav technologie, mechanizace a řízení staveb Teorie měření a regulace
Ústav technologie, mechanizace a řízení staveb CW01 - Teorie měření a regulace ZS 2010/2011 SPEC. 2.p 2010 - Ing. Václav Rada, CSc. Ústav technologie, mechanizace a řízení staveb Teorie měření a regulace
Biologické signály. X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů
 Biologické signály X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Biologické signály mají původ v živém organismu jsou vyvolány buď samotnými životními projevy
Biologické signály X31ZLE Základy lékařské elektroniky Jan Havlík Katedra teorie obvodů xhavlikj@fel.cvut.cz Biologické signály mají původ v živém organismu jsou vyvolány buď samotnými životními projevy
VY_32_INOVACE_E 15 03
 Název a adresa školy: Střední škola průmyslová a umělecká, Opava, příspěvková organizace, Praskova 399/8, Opava, 746 01 Název operačního programu: OP Vzdělávání pro konkurenceschopnost, oblast podpory
Název a adresa školy: Střední škola průmyslová a umělecká, Opava, příspěvková organizace, Praskova 399/8, Opava, 746 01 Název operačního programu: OP Vzdělávání pro konkurenceschopnost, oblast podpory
Řízení svalového tonu Martina Hoskovcová
 Řízení svalového tonu Martina Hoskovcová Neurologická klinika a Centrum klinických neurověd Universita Karlova v Praze, 1. lékařská fakulta a Všeobecná fakultní nemocnice v Praze Svalový tonus Reflexně
Řízení svalového tonu Martina Hoskovcová Neurologická klinika a Centrum klinických neurověd Universita Karlova v Praze, 1. lékařská fakulta a Všeobecná fakultní nemocnice v Praze Svalový tonus Reflexně
Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem. MUDr.
 Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem MUDr. Tomáš Veverka Neurologická klinika Lékařské fakulty Univerzity Palackého a Fakultní
Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem MUDr. Tomáš Veverka Neurologická klinika Lékařské fakulty Univerzity Palackého a Fakultní
Katedra biomedicínské techniky
 ČESKÉ VYSOKÉ UČENÍ TECHNICKÉ V PRAZE FAKULTA BIOMEDICÍNSKÉHO INŽENÝRSTVÍ Katedra biomedicínské techniky BAKALÁŘSKÁ PRÁCE 2008 Jakub Schlenker Obsah Úvod 1 1 Teoretický úvod 2 1.1 Elektrokardiografie............................
ČESKÉ VYSOKÉ UČENÍ TECHNICKÉ V PRAZE FAKULTA BIOMEDICÍNSKÉHO INŽENÝRSTVÍ Katedra biomedicínské techniky BAKALÁŘSKÁ PRÁCE 2008 Jakub Schlenker Obsah Úvod 1 1 Teoretický úvod 2 1.1 Elektrokardiografie............................
Evokované potenciály. Principy, možnosti a meze, indikace. Doc. MUDr. Pavel Urban, CSc.
 Evokované potenciály Principy, možnosti a meze, indikace Doc. MUDr. Pavel Urban, CSc. Rozdělení evokovaných potenciálů Podle typu podnětu Podle doby vzniku Podle lokalizace generátorů Near-field x far-field
Evokované potenciály Principy, možnosti a meze, indikace Doc. MUDr. Pavel Urban, CSc. Rozdělení evokovaných potenciálů Podle typu podnětu Podle doby vzniku Podle lokalizace generátorů Near-field x far-field
Techniky detekce a určení velikosti souvislých trhlin
 Techniky detekce a určení velikosti souvislých trhlin Přehled Byl-li podle obecných norem nebo regulačních směrnic detekovány souvislé trhliny na vnitřním povrchu, musí být následně přesně stanoven rozměr.
Techniky detekce a určení velikosti souvislých trhlin Přehled Byl-li podle obecných norem nebo regulačních směrnic detekovány souvislé trhliny na vnitřním povrchu, musí být následně přesně stanoven rozměr.
Fyziologická akustika. fyziologická akustika: jak to funguje psychologická akustika: jak to na nás působí
 Fyziologická akustika anatomie: jak to vypadá fyziologická akustika: jak to funguje psychologická akustika: jak to na nás působí hudební akustika: jak dosáhnout libých počitků Anatomie lidského ucha Vnější
Fyziologická akustika anatomie: jak to vypadá fyziologická akustika: jak to funguje psychologická akustika: jak to na nás působí hudební akustika: jak dosáhnout libých počitků Anatomie lidského ucha Vnější
Spánek. kurz Neurobiologie chování a paměti. RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR
 Spánek kurz Neurobiologie chování a paměti RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR Spánek Spánková stadia a spánková architektura Role spánku při konsolidaci
Spánek kurz Neurobiologie chování a paměti RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR Spánek Spánková stadia a spánková architektura Role spánku při konsolidaci
Ultrazvuková defektoskopie. Vypracoval Jan Janský
 Ultrazvuková defektoskopie Vypracoval Jan Janský Základní principy použití vysokých akustických frekvencí pro zjištění vlastností máteriálu a vad typické zařízení: generátor/přijímač pulsů snímač zobrazovací
Ultrazvuková defektoskopie Vypracoval Jan Janský Základní principy použití vysokých akustických frekvencí pro zjištění vlastností máteriálu a vad typické zařízení: generátor/přijímač pulsů snímač zobrazovací
ZPRACOVÁNÍ A ANALÝZA EEG. Jan Prokš. Katedra teorie obvodů, Fakulta elektrotechnická, České vysoké učení technické v Praze.
 ZPRACOVÁNÍ A ANALÝZA EEG Jan Prokš Katedra teorie obvodů, Fakulta elektrotechnická, České vysoké učení technické v Praze. Anotace Číslicové zpracování signálů je relativně mladým technickým oborem, který
ZPRACOVÁNÍ A ANALÝZA EEG Jan Prokš Katedra teorie obvodů, Fakulta elektrotechnická, České vysoké učení technické v Praze. Anotace Číslicové zpracování signálů je relativně mladým technickým oborem, který
Transkraniální elektrostimulace jako způsob rehabilitace částo nemocných dětí
 Transkraniální elektrostimulace jako způsob rehabilitace částo nemocných dětí Abstrakt V tomto článku se poprvé zkoumala účinnost transkraniální elektrické stimulace (TES) v programu rehabilitace dětí
Transkraniální elektrostimulace jako způsob rehabilitace částo nemocných dětí Abstrakt V tomto článku se poprvé zkoumala účinnost transkraniální elektrické stimulace (TES) v programu rehabilitace dětí
SIGNÁLY A LINEÁRNÍ SYSTÉMY
 SIGNÁLY A LINEÁRNÍ SYSTÉMY prof. Ing. Jiří Holčík, CSc. holcik@iba.muni.cz @iba.muni.cz,, Kamenice 3, 4. patro, dv.č.44.44 INVESTICE Institut DO biostatistiky ROZVOJE VZDĚLÁVÁNÍ a analýz XI. STABILITA
SIGNÁLY A LINEÁRNÍ SYSTÉMY prof. Ing. Jiří Holčík, CSc. holcik@iba.muni.cz @iba.muni.cz,, Kamenice 3, 4. patro, dv.č.44.44 INVESTICE Institut DO biostatistiky ROZVOJE VZDĚLÁVÁNÍ a analýz XI. STABILITA
Příloha 2. Informovaný souhlas účastníka výzkumu
 Přílohy: Příloha 1 vyjádření etické komise Příloha 2 informovaný souhlas účastníka výzkumu Příloha 3 srovnání dynamiky opory u běhu Příloha 4 komponenty reakční síly podložky při akcelerovaném běhu 1.
Přílohy: Příloha 1 vyjádření etické komise Příloha 2 informovaný souhlas účastníka výzkumu Příloha 3 srovnání dynamiky opory u běhu Příloha 4 komponenty reakční síly podložky při akcelerovaném běhu 1.
1. Základy teorie přenosu informací
 1. Základy teorie přenosu informací Úvodem citát o pojmu informace Informace je název pro obsah toho, co se vymění s vnějším světem, když se mu přizpůsobujeme a působíme na něj svým přizpůsobováním. N.
1. Základy teorie přenosu informací Úvodem citát o pojmu informace Informace je název pro obsah toho, co se vymění s vnějším světem, když se mu přizpůsobujeme a působíme na něj svým přizpůsobováním. N.
Elektromagnetický oscilátor
 Elektromagnetický oscilátor Již jsme poznali kmitání mechanického oscilátoru (závaží na pružině) - potenciální energie pružnosti se přeměňuje na kinetickou energii a naopak. T =2 m k Nejjednodušší elektromagnetický
Elektromagnetický oscilátor Již jsme poznali kmitání mechanického oscilátoru (závaží na pružině) - potenciální energie pružnosti se přeměňuje na kinetickou energii a naopak. T =2 m k Nejjednodušší elektromagnetický
Aktivační metody v průběhu EEG - výhody a rizika. Petr Zlonický
 Aktivační metody v průběhu EEG - výhody a rizika Petr Zlonický Aktivační metody - výhody Používají se k vyprovokování, zvýraznění nebo lepšímu definování abnormální aktivity při normálním nebo neprůkazném
Aktivační metody v průběhu EEG - výhody a rizika Petr Zlonický Aktivační metody - výhody Používají se k vyprovokování, zvýraznění nebo lepšímu definování abnormální aktivity při normálním nebo neprůkazném
Návrh frekvenčního filtru
 Návrh frekvenčního filtru Vypracoval: Martin Dlouhý, Petr Salajka 25. 9 2010 1 1 Zadání 1. Navrhněte co nejjednodušší přenosovou funkci frekvenčního pásmového filtru Dolní propusti typu Bessel, která bude
Návrh frekvenčního filtru Vypracoval: Martin Dlouhý, Petr Salajka 25. 9 2010 1 1 Zadání 1. Navrhněte co nejjednodušší přenosovou funkci frekvenčního pásmového filtru Dolní propusti typu Bessel, která bude
VÍŘIVÉ PROUDY DZM 2013 1
 VÍŘIVÉ PROUDY DZM 2013 1 2 VÍŘIVÉ PROUDY ÚVOD Vířivé proudy tvoří druhou skupinu v metodách, které využívají ke zjišťování vad materiálu a výrobků působení elektromagnetického pole. Na rozdíl od metody
VÍŘIVÉ PROUDY DZM 2013 1 2 VÍŘIVÉ PROUDY ÚVOD Vířivé proudy tvoří druhou skupinu v metodách, které využívají ke zjišťování vad materiálu a výrobků působení elektromagnetického pole. Na rozdíl od metody
Neurologie pro fyzioterapeuty: vstupní přednáška. Jan Roth
 Neurologie pro fyzioterapeuty: vstupní přednáška Jan Roth Obecný úvod Neurologie je lékařský obor zabývající se diagnostikou, terapií a prevencí nemocí a poruch centrální nervové soustavy (mozek, mícha),
Neurologie pro fyzioterapeuty: vstupní přednáška Jan Roth Obecný úvod Neurologie je lékařský obor zabývající se diagnostikou, terapií a prevencí nemocí a poruch centrální nervové soustavy (mozek, mícha),
TERMINOLOGIE ... NAMĚŘENÁ DATA. Radek Mareček PŘEDZPRACOVÁNÍ DAT. funkční skeny
 PŘEDZPRACOVÁNÍ DAT Radek Mareček TERMINOLOGIE Session soubor skenů nasnímaných během jednoho běhu stimulačního paradigmatu (řádově desítky až stovky skenů) Sken jeden nasnímaný objem... Voxel elementární
PŘEDZPRACOVÁNÍ DAT Radek Mareček TERMINOLOGIE Session soubor skenů nasnímaných během jednoho běhu stimulačního paradigmatu (řádově desítky až stovky skenů) Sken jeden nasnímaný objem... Voxel elementární
Obsah. Předmluva...13
 Obsah Předmluva...13 1 Pohyb jako základní projev života...17 1.1 Pohyb obecně...17 1.2 Pohybové chování...17 1.3 Vliv pohybu na životní pochody...18 1.4 Vztah pohybu k funkci CNS...19 1.5 Psychomotorické
Obsah Předmluva...13 1 Pohyb jako základní projev života...17 1.1 Pohyb obecně...17 1.2 Pohybové chování...17 1.3 Vliv pohybu na životní pochody...18 1.4 Vztah pohybu k funkci CNS...19 1.5 Psychomotorické
OPVK CZ.1.07/2.2.00/
 18.2.2013 OPVK CZ.1.07/2.2.00/28.0184 Cvičení z NMR OCH/NMR Mgr. Tomáš Pospíšil, Ph.D. LS 2012/2013 18.2.2013 NMR základní principy NMR Nukleární Magnetická Resonance N - nukleární (studujeme vlastnosti
18.2.2013 OPVK CZ.1.07/2.2.00/28.0184 Cvičení z NMR OCH/NMR Mgr. Tomáš Pospíšil, Ph.D. LS 2012/2013 18.2.2013 NMR základní principy NMR Nukleární Magnetická Resonance N - nukleární (studujeme vlastnosti
Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy)
 mozek mícha Periferní nervový systém (nervy)") Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Jak se měří rychlost toku krve v cévách?
 Jak se měří rychlost toku krve v cévách? Princip této vyšetřovací metody je založen na Dopplerově jevu, který spočívá ve změně frekvence ultrazvukového vlnění při vzájemném pohybu zdroje a detektoru vlnění.
Jak se měří rychlost toku krve v cévách? Princip této vyšetřovací metody je založen na Dopplerově jevu, který spočívá ve změně frekvence ultrazvukového vlnění při vzájemném pohybu zdroje a detektoru vlnění.
(XXIX.) Vyšetření vzpřímeného postoje
 Vyšetření vzpřímeného postoje") (XXIX.) Vyšetření vzpřímeného postoje Fyziologie II - cvičení Fyziologický ústav LF MU, 2016 Michal Pásek Řízení vzpřímeného postoje centrálním nervovým systémem spočívá v neustálé korekci výchylek našeho
(XXIX.) Vyšetření vzpřímeného postoje Fyziologie II - cvičení Fyziologický ústav LF MU, 2016 Michal Pásek Řízení vzpřímeného postoje centrálním nervovým systémem spočívá v neustálé korekci výchylek našeho
Akustika. Teorie - slyšení. 5. Přednáška
 Akustika Teorie - slyšení 5. Přednáška Sluchové ústrojí Vnitřní a vnější slyšení Zpěv, vlastní hlas Dechové nástroje Vibrace a chvění Ucho Ucho je složeno z ucha vnějšího, středního a vnitřního. K vnějšímu
Akustika Teorie - slyšení 5. Přednáška Sluchové ústrojí Vnitřní a vnější slyšení Zpěv, vlastní hlas Dechové nástroje Vibrace a chvění Ucho Ucho je složeno z ucha vnějšího, středního a vnitřního. K vnějšímu
Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy)
 mozek mícha Periferní nervový systém (nervy)") Buňka Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Buňka Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Emise vyvolaná působením fotonů nebo částic
 Emise vyvolaná působením fotonů nebo částic PES (fotoelektronová spektroskopie) XPS (rentgenová fotoelektronová spektroskopie), ESCA (elektronová spektroskopie pro chemickou analýzu) UPS (ultrafialová
Emise vyvolaná působením fotonů nebo částic PES (fotoelektronová spektroskopie) XPS (rentgenová fotoelektronová spektroskopie), ESCA (elektronová spektroskopie pro chemickou analýzu) UPS (ultrafialová
Projekt: Inovace oboru Mechatronik pro Zlínský kraj Registrační číslo: CZ.1.07/1.1.08/ Vlnění
 Projekt: Inovace oboru Mechatronik pro Zlínský kraj Registrační číslo: CZ.1.07/1.1.08/03.0009 Vlnění Vhodíme-li na klidnou vodní hladinu kámen, hladina se jeho dopadem rozkmitá a z místa rozruchu se začnou
Projekt: Inovace oboru Mechatronik pro Zlínský kraj Registrační číslo: CZ.1.07/1.1.08/03.0009 Vlnění Vhodíme-li na klidnou vodní hladinu kámen, hladina se jeho dopadem rozkmitá a z místa rozruchu se začnou
Rychlost pulzové vlny (XII)
") Rychlost pulzové vlny (XII) Definice pulzové vlny Pulzová vlna vzniká během srdeční revoluce, kdy dochází za systoly k vypuzení krve z levé komory do velkého oběhu. Arteriální systém se s tímto rychle
Rychlost pulzové vlny (XII) Definice pulzové vlny Pulzová vlna vzniká během srdeční revoluce, kdy dochází za systoly k vypuzení krve z levé komory do velkého oběhu. Arteriální systém se s tímto rychle
Základy spektroskopie a její využití v astronomii
 Ing. Libor Lenža, Hvězdárna Valašské Meziříčí, p. o. Základy spektroskopie a její využití v astronomii Hvězdárna Valašské Meziříčí, p. o. Krajská hvezdáreň v Žiline Světlo x záření Jak vypadá spektrum?
Ing. Libor Lenža, Hvězdárna Valašské Meziříčí, p. o. Základy spektroskopie a její využití v astronomii Hvězdárna Valašské Meziříčí, p. o. Krajská hvezdáreň v Žiline Světlo x záření Jak vypadá spektrum?
ELEKTRICKÉ POLE V BUŇKÁCH A V ORGANISMU. Helena Uhrová
 ELEKTRICKÉ POLE V BUŇKÁCH A V ORGANISMU Helena Uhrová Hierarichické uspořádání struktury z fyzikálního hlediska organismus člověk elektrodynamika Maxwellovy rovnice buňka akční potenciál fenomenologická
ELEKTRICKÉ POLE V BUŇKÁCH A V ORGANISMU Helena Uhrová Hierarichické uspořádání struktury z fyzikálního hlediska organismus člověk elektrodynamika Maxwellovy rovnice buňka akční potenciál fenomenologická
Elektroencefalografie. doc. MUDr. Jan Pokorný, DrSc.
 Elektroencefalografie doc. MUDr. Jan Pokorný, DrSc. Elektroencefalografie (EEG) EEG = základní elektrofyziologická metoda pro vyšetření mozkové aktivity. Podstata EEG: registruje časoprostorové změny mozkových
Elektroencefalografie doc. MUDr. Jan Pokorný, DrSc. Elektroencefalografie (EEG) EEG = základní elektrofyziologická metoda pro vyšetření mozkové aktivity. Podstata EEG: registruje časoprostorové změny mozkových
Spektrální charakteristiky
 Spektrální charakteristiky Cíl cvičení: Měření spektrálních charakteristik filtrů a zdrojů osvětlení 1 Teoretický úvod Interakcí elektromagnetického vlnění s libovolnou látkou vzniká optický jev, který
Spektrální charakteristiky Cíl cvičení: Měření spektrálních charakteristik filtrů a zdrojů osvětlení 1 Teoretický úvod Interakcí elektromagnetického vlnění s libovolnou látkou vzniká optický jev, který
Osnova. Idea ASK/FSK/PSK ASK Amplitudové... Strana 1 z 16. Celá obrazovka. Konec Základy radiotechniky
 Pulsní kódová modulace, amplitudové, frekvenční a fázové kĺıčování Josef Dobeš 24. října 2006 Strana 1 z 16 Základy radiotechniky 1. Pulsní modulace Strana 2 z 16 Pulsní šířková modulace (PWM) PAM, PPM,
Pulsní kódová modulace, amplitudové, frekvenční a fázové kĺıčování Josef Dobeš 24. října 2006 Strana 1 z 16 Základy radiotechniky 1. Pulsní modulace Strana 2 z 16 Pulsní šířková modulace (PWM) PAM, PPM,
Akustika. Teorie - slyšení. 5. Přednáška
 Akustika Teorie - slyšení 5. Přednáška http://data.audified.com/downlpublic/edu/zha_pdf.zip http://data.audified.com/downlpublic/edu/akustikaotazky03.pdf http://data.audified.com/downlpublic/edu/jamusimulatorspro103mac.dmg.zip
Akustika Teorie - slyšení 5. Přednáška http://data.audified.com/downlpublic/edu/zha_pdf.zip http://data.audified.com/downlpublic/edu/akustikaotazky03.pdf http://data.audified.com/downlpublic/edu/jamusimulatorspro103mac.dmg.zip
TRÉNINK DOVEDNOSTÍ LUDĚK BUKAČ TRENÉR ČSLH
 TRÉNINK DOVEDNOSTÍ LUDĚK BUKAČ TRENÉR ČSLH DOVEDNOSTI Spektrum herní činnosti tvoří hokejové dovednosti. Bruslení, střelba, přihrávání, kličkování. Opakováním se dovednost stává návykem. Dovednostní návyky
TRÉNINK DOVEDNOSTÍ LUDĚK BUKAČ TRENÉR ČSLH DOVEDNOSTI Spektrum herní činnosti tvoří hokejové dovednosti. Bruslení, střelba, přihrávání, kličkování. Opakováním se dovednost stává návykem. Dovednostní návyky
Stručný úvod do spektroskopie
 Vzdělávací soustředění studentů projekt KOSOAP Slunce, projevy sluneční aktivity a využití spektroskopie v astrofyzikálním výzkumu Stručný úvod do spektroskopie Ing. Libor Lenža, Hvězdárna Valašské Meziříčí,
Vzdělávací soustředění studentů projekt KOSOAP Slunce, projevy sluneční aktivity a využití spektroskopie v astrofyzikálním výzkumu Stručný úvod do spektroskopie Ing. Libor Lenža, Hvězdárna Valašské Meziříčí,
Zvuk a jeho vlastnosti
 PEF MZLU v Brně 9. října 2008 Zvuk obecně podélné (nebo příčné) mechanické vlnění v látkovém prostředí, které je schopno vyvolat v lidském uchu sluchový vjem. frekvence leží v rozsahu přibližně 20 Hz až
PEF MZLU v Brně 9. října 2008 Zvuk obecně podélné (nebo příčné) mechanické vlnění v látkovém prostředí, které je schopno vyvolat v lidském uchu sluchový vjem. frekvence leží v rozsahu přibližně 20 Hz až
Pokročilé operace s obrazem
 Získávání a analýza obrazové informace Pokročilé operace s obrazem Biofyzikální ústav Lékařské fakulty Masarykovy univerzity Brno prezentace je součástí projektu FRVŠ č.2487/2011 (BFÚ LF MU) Získávání
Získávání a analýza obrazové informace Pokročilé operace s obrazem Biofyzikální ústav Lékařské fakulty Masarykovy univerzity Brno prezentace je součástí projektu FRVŠ č.2487/2011 (BFÚ LF MU) Získávání
Příloha č. 5 k nařízení vlády č. 361/2007 Sb. (Zapracovaná změna provedená NV č. 68/2010 Sb. a změna č. 93/2012 Sb.)
") Příloha č. 5 k nařízení vlády č. 361/2007 Sb. (Zapracovaná změna provedená NV č. 68/2010 Sb. a změna č. 93/2012 Sb.) Fyzická zátěž, její hygienické limity a postup jejich stanovení ČÁST A Přípustné a průměrné
Příloha č. 5 k nařízení vlády č. 361/2007 Sb. (Zapracovaná změna provedená NV č. 68/2010 Sb. a změna č. 93/2012 Sb.) Fyzická zátěž, její hygienické limity a postup jejich stanovení ČÁST A Přípustné a průměrné
VYUŽITÍ MATLABU PRO PODPORU VÝUKY A PŘI ŘEŠENÍ VÝZKUMNÝCH ÚKOLŮ NA KATEDŘE KOMUNIKAČNÍCH A INFORMAČNÍCH SYSTÉMŮ
 VYUŽITÍ MATLABU PRO PODPORU VÝUKY A PŘI ŘEŠENÍ VÝZKUMNÝCH ÚKOLŮ NA KATEDŘE KOMUNIKAČNÍCH A INFORMAČNÍCH SYSTÉMŮ Markéta Mazálková Katedra komunikačních a informačních systémů Fakulta vojenských technologií,
VYUŽITÍ MATLABU PRO PODPORU VÝUKY A PŘI ŘEŠENÍ VÝZKUMNÝCH ÚKOLŮ NA KATEDŘE KOMUNIKAČNÍCH A INFORMAČNÍCH SYSTÉMŮ Markéta Mazálková Katedra komunikačních a informačních systémů Fakulta vojenských technologií,
4.2.3 ŠÍŘE FREKVENČNÍHO PÁSMA CHOROVÉHO ELEMENTU A DISTRIBUČNÍ FUNKCE VLNOVÝCH NORMÁL
 4.2.3 ŠÍŘE FREKVENČNÍHO PÁSMA CHOROVÉHO ELEMENTU A DISTRIBUČNÍ FUNKCE VLNOVÝCH NORMÁL V předchozích dvou podkapitolách jsme ukázali, že chorové emise se mohou v řadě případů šířit nevedeným způsobem. Připomeňme
4.2.3 ŠÍŘE FREKVENČNÍHO PÁSMA CHOROVÉHO ELEMENTU A DISTRIBUČNÍ FUNKCE VLNOVÝCH NORMÁL V předchozích dvou podkapitolách jsme ukázali, že chorové emise se mohou v řadě případů šířit nevedeným způsobem. Připomeňme
Epilepsie. Silvia Čillíková FEL ČVUT. 9th May 2006
 Epilepsie Silvia Čillíková FEL ČVUT 9th May 2006 Úvod Epilepsie (zkr. epi) je skupina poruch mozku projevujících se opakovanými záchvaty (paroxysmy) různého charakteru Je to relativně běžné onemocnění,
Epilepsie Silvia Čillíková FEL ČVUT 9th May 2006 Úvod Epilepsie (zkr. epi) je skupina poruch mozku projevujících se opakovanými záchvaty (paroxysmy) různého charakteru Je to relativně běžné onemocnění,
Modelování a aproximace v biomechanice
 Modelování a aproximace v biomechanice Během většiny lidské aktivity působí v jednom okamžiku víc než jedna skupina svalů. Je-li úkolem analyzovat síly působící v kloubech a svalech během určité lidské
Modelování a aproximace v biomechanice Během většiny lidské aktivity působí v jednom okamžiku víc než jedna skupina svalů. Je-li úkolem analyzovat síly působící v kloubech a svalech během určité lidské
Středofrekvenční terapie (SF)
") Středofrekvenční terapie (S) pojem zavedl Gildemeister dráždění (střídavým) proudem o frekvenci 1-100 khz neodpovídá dělení rádiových frekvencí (v USA ani ve T!) harmonický střídavý proud; bipolární impulzy
Středofrekvenční terapie (S) pojem zavedl Gildemeister dráždění (střídavým) proudem o frekvenci 1-100 khz neodpovídá dělení rádiových frekvencí (v USA ani ve T!) harmonický střídavý proud; bipolární impulzy
(VIII.) Časová a prostorová sumace u kosterního svalu. Fyziologický ústav LF MU, 2016 Jana Svačinová
 Časová a prostorová sumace u kosterního svalu. Fyziologický ústav LF MU, 2016 Jana Svačinová") (VIII.) Časová a prostorová sumace u kosterního svalu Fyziologický ústav LF MU, 2016 Jana Svačinová Kontrakce příčně pruhovaného kosterního svalu Myografie metoda umožňující registraci kontrakce svalů
(VIII.) Časová a prostorová sumace u kosterního svalu Fyziologický ústav LF MU, 2016 Jana Svačinová Kontrakce příčně pruhovaného kosterního svalu Myografie metoda umožňující registraci kontrakce svalů
3.2.5 Odraz, lom a ohyb vlnění
 3..5 Odraz, lom a ohyb vlnění Předpoklady: 304 Odraz a lom vlnění na rozhranní dvou prostředí s různou rychlostí šíření http://www.phy.ntnu.edu.tw/ntnujava/index.php?topic=16.0 Rovinná vlna dopadá šikmo
3..5 Odraz, lom a ohyb vlnění Předpoklady: 304 Odraz a lom vlnění na rozhranní dvou prostředí s různou rychlostí šíření http://www.phy.ntnu.edu.tw/ntnujava/index.php?topic=16.0 Rovinná vlna dopadá šikmo
Zkoušení velkých výkovků a digitální ultrazvukové přístroje
 - 1 - Zkoušení velkých výkovků a digitální ultrazvukové přístroje Ultrazvuková kontrola Ing. Jaroslav Smejkal, Testima, spol. s r.o. zpracováno dle materiálů GE IT Krautkramer Zkoušení výkovků není jednoduchou
- 1 - Zkoušení velkých výkovků a digitální ultrazvukové přístroje Ultrazvuková kontrola Ing. Jaroslav Smejkal, Testima, spol. s r.o. zpracováno dle materiálů GE IT Krautkramer Zkoušení výkovků není jednoduchou
Neuromuskulární jitter v diagnostice okulární myastenie gravis
 Neuromuskulární jitter v diagnostice okulární myastenie gravis S. Voháňka J. Bednařík Z. Kadaňka Neurologická klinika FN Brno Základní technické aspekty a požadavky SF EMG Elektrody Filtry (25 µm) (500
Neuromuskulární jitter v diagnostice okulární myastenie gravis S. Voháňka J. Bednařík Z. Kadaňka Neurologická klinika FN Brno Základní technické aspekty a požadavky SF EMG Elektrody Filtry (25 µm) (500
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.4 Prvky elektronických obvodů Kapitola
Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.4 Prvky elektronických obvodů Kapitola
Jiří Brus. (Verze 1.0.1-2005) (neupravená a neúplná)
 (neupravená a neúplná)") Jiří Brus (Verze 1.0.1-2005) (neupravená a neúplná) Ústav makromolekulární chemie AV ČR, Heyrovského nám. 2, Praha 6 - Petřiny 162 06 e-mail: brus@imc.cas.cz Transverzální magnetizace, která vykonává precesi
Jiří Brus (Verze 1.0.1-2005) (neupravená a neúplná) Ústav makromolekulární chemie AV ČR, Heyrovského nám. 2, Praha 6 - Petřiny 162 06 e-mail: brus@imc.cas.cz Transverzální magnetizace, která vykonává precesi
Zvuk. 1. základní kmitání. 2. šíření zvuku
 Zvuk 1. základní kmitání - vzduchem se šíří tlakové vzruchy (vzruchová vlna), zvuk je systémem zhuštěnin a zředěnin - podstatou zvuku je kmitání zdroje zvuku a tím způsobené podélné vlnění elastického
Zvuk 1. základní kmitání - vzduchem se šíří tlakové vzruchy (vzruchová vlna), zvuk je systémem zhuštěnin a zředěnin - podstatou zvuku je kmitání zdroje zvuku a tím způsobené podélné vlnění elastického
Elektronický systém a programové vybavení pro detekci a optimalizaci pulzů kardiostimulátoru
 Elektronický systém a programové vybavení pro detekci a optimalizaci pulzů kardiostimulátoru Milan Štork Katedra aplikované elektroniky a telekomunikací & Regionálním inovační centrum pro elektrotechniku
Elektronický systém a programové vybavení pro detekci a optimalizaci pulzů kardiostimulátoru Milan Štork Katedra aplikované elektroniky a telekomunikací & Regionálním inovační centrum pro elektrotechniku
Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách
 Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách Difuze Vyrovnávání koncentrací látek na základě náhodného pohybu Osmóza (difuze rozpouštědla) Dva roztoky o rúzné koncentraci oddělené
Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách Difuze Vyrovnávání koncentrací látek na základě náhodného pohybu Osmóza (difuze rozpouštědla) Dva roztoky o rúzné koncentraci oddělené
fluktuace jak dob trvání po sobě jdoucích srdečních cyklů, tak hodnot Heart Rate Variability) je jev, který
 je jev, který") BIOLOGICKÉ A LÉKAŘSKÉ SIGNÁLY VI. VARIABILITA SRDEČNÍHO RYTMU VARIABILITA SRDEČNÍHO RYTMU VARIABILITA SRDEČNÍHO RYTMU, tj. fluktuace jak dob trvání po sobě jdoucích srdečních cyklů, tak hodnot okamžité
BIOLOGICKÉ A LÉKAŘSKÉ SIGNÁLY VI. VARIABILITA SRDEČNÍHO RYTMU VARIABILITA SRDEČNÍHO RYTMU VARIABILITA SRDEČNÍHO RYTMU, tj. fluktuace jak dob trvání po sobě jdoucích srdečních cyklů, tak hodnot okamžité
AMBULANTNÍ MONITOROVÁNÍ TK V LÉČBĚ HYPERTENZE DIABETIKŮ
 AMBULANTNÍ MONITOROVÁNÍ TK V LÉČBĚ HYPERTENZE DIABETIKŮ Ve svém sdělení bych vás chtěla seznámit s principem ambulantního monitorování TK a s jeho využitím u hypertenzních diabetiků. Nesprávně bývá toto
AMBULANTNÍ MONITOROVÁNÍ TK V LÉČBĚ HYPERTENZE DIABETIKŮ Ve svém sdělení bych vás chtěla seznámit s principem ambulantního monitorování TK a s jeho využitím u hypertenzních diabetiků. Nesprávně bývá toto
Prognóza počtu a věkové struktury obyvatel MČ Praha-Satalice do roku 2025
 Prognóza počtu a věkové struktury obyvatel MČ Praha-Satalice do roku 2025 Březen 2016 Zpracoval: RNDr. Tomáš Brabec, Ph.D. Institut plánování a rozvoje hl. m. Prahy Sekce strategií a politik, Kancelář
Prognóza počtu a věkové struktury obyvatel MČ Praha-Satalice do roku 2025 Březen 2016 Zpracoval: RNDr. Tomáš Brabec, Ph.D. Institut plánování a rozvoje hl. m. Prahy Sekce strategií a politik, Kancelář
Vztah výpočetní techniky a biomedicíny
 Vztah výpočetní techniky a biomedicíny počítač - nástroj pro vývoj nových přístrojů počítač -součást přístrojových systémů počítač - nástroj pro zpracování informací přispívá k metody, techniky a teorie
Vztah výpočetní techniky a biomedicíny počítač - nástroj pro vývoj nových přístrojů počítač -součást přístrojových systémů počítač - nástroj pro zpracování informací přispívá k metody, techniky a teorie
Jaký význam má kritický kmitočet vedení? - nejnižší kmitočet vlny, při kterém se vlna začíná šířit vedením.
 Jaký význam má kritický kmitočet vedení? - nejnižší kmitočet vlny, při kterém se vlna začíná šířit vedením. Na čem závisí účinnost vedení? účinnost vedení závisí na činiteli útlumu β a na činiteli odrazu
Jaký význam má kritický kmitočet vedení? - nejnižší kmitočet vlny, při kterém se vlna začíná šířit vedením. Na čem závisí účinnost vedení? účinnost vedení závisí na činiteli útlumu β a na činiteli odrazu
9. Léčiva CNS - úvod (1)
") 9. Léčiva CNS - úvod (1) se se souhlasem souhlasem autora autora ál školy koly -techlogic techlogické Jeho Jeho žit bez bez souhlasu souhlasu autora autora je je ázá Nervová soustava: Centrální nervový
9. Léčiva CNS - úvod (1) se se souhlasem souhlasem autora autora ál školy koly -techlogic techlogické Jeho Jeho žit bez bez souhlasu souhlasu autora autora je je ázá Nervová soustava: Centrální nervový
Speciální spektrometrické metody. Zpracování signálu ve spektroskopii
 Speciální spektrometrické metody Zpracování signálu ve spektroskopii detekce slabých signálů synchronní detekce (Lock-in) čítaní fotonů měření časového průběhu signálů metoda fázového posuvu časově korelované
Speciální spektrometrické metody Zpracování signálu ve spektroskopii detekce slabých signálů synchronní detekce (Lock-in) čítaní fotonů měření časového průběhu signálů metoda fázového posuvu časově korelované
Mechanické kmitání a vlnění
 Mechanické kmitání a vlnění Pohyb tělesa, který se v určitém časovém intervalu pravidelně opakuje periodický pohyb S kmitavým pohybem se setkáváme např.: Zařízení, které volně kmitá, nazýváme mechanický
Mechanické kmitání a vlnění Pohyb tělesa, který se v určitém časovém intervalu pravidelně opakuje periodický pohyb S kmitavým pohybem se setkáváme např.: Zařízení, které volně kmitá, nazýváme mechanický
ELT1 - Přednáška č. 6
 ELT1 - Přednáška č. 6 Elektrotechnická terminologie a odborné výrazy, měřicí jednotky a činitelé, které je ovlivňují. Rozdíl potenciálů, elektromotorická síla, napětí, el. napětí, proud, odpor, vodivost,
ELT1 - Přednáška č. 6 Elektrotechnická terminologie a odborné výrazy, měřicí jednotky a činitelé, které je ovlivňují. Rozdíl potenciálů, elektromotorická síla, napětí, el. napětí, proud, odpor, vodivost,
FVZ K13138-TACR-V004-G-TRIGGER_BOX
 TriggerBox Souhrn hlavních funkcí Synchronizace přes Ethernetový protokol IEEE 1588 v2 PTP Automatické určení možnosti, zda SyncCore zastává roli PTP master nebo PTP slave dle mechanizmů standardu PTP
TriggerBox Souhrn hlavních funkcí Synchronizace přes Ethernetový protokol IEEE 1588 v2 PTP Automatické určení možnosti, zda SyncCore zastává roli PTP master nebo PTP slave dle mechanizmů standardu PTP
2. Vlnění. π T. t T. x λ. Machův vlnostroj
 2. Vlnění 2.1 Vlnění zvláštní případ pohybu prostředí Vlnění je pohyb v soustavě velkého počtu částic navzájem vázaných, kdy částice kmitají kolem svých rovnovážných poloh. Druhy vlnění: vlnění příčné
2. Vlnění 2.1 Vlnění zvláštní případ pohybu prostředí Vlnění je pohyb v soustavě velkého počtu částic navzájem vázaných, kdy částice kmitají kolem svých rovnovážných poloh. Druhy vlnění: vlnění příčné
Ing. Jiří Fejfar, Ph.D. Dálkový průzkum Země
 Ing. Jiří Fejfar, Ph.D. Dálkový průzkum Země strana 2 Co je DPZ Dálkový průzkum je umění rozdělit svět na množství malých barevných čtverečků, se kterými si lze hrát na počítači a odhalovat jejich neuvěřitelný
Ing. Jiří Fejfar, Ph.D. Dálkový průzkum Země strana 2 Co je DPZ Dálkový průzkum je umění rozdělit svět na množství malých barevných čtverečků, se kterými si lze hrát na počítači a odhalovat jejich neuvěřitelný
MUDr. Jozef Jakabčin, Ph.D.
 MUDr. Jozef Jakabčin, Ph.D. RAO LAO AKCE RYTMUS FRQ OSA QRS P QRS QT ST T Patologické změny ARYTMIE Šíření aktivace v pravé a následně levé síni P vlna je zápis splynutí dvou vln Aktivace pravé
MUDr. Jozef Jakabčin, Ph.D. RAO LAO AKCE RYTMUS FRQ OSA QRS P QRS QT ST T Patologické změny ARYTMIE Šíření aktivace v pravé a následně levé síni P vlna je zápis splynutí dvou vln Aktivace pravé
Didaktické metody Metodou
 Didaktické metody Metodou rozumíme záměrné, plánovité uspořádání úkolů a činností sportovce tak, aby vzhledem k spolupůsobícím podmínkám byl co nejefektivněji dosažen tréninkový cíl. Z aspektu trenéra
Didaktické metody Metodou rozumíme záměrné, plánovité uspořádání úkolů a činností sportovce tak, aby vzhledem k spolupůsobícím podmínkám byl co nejefektivněji dosažen tréninkový cíl. Z aspektu trenéra
Relativistické jevy při synchronizaci nové generace atomových hodin. Jan Geršl Český metrologický institut
 Relativistické jevy při synchronizaci nové generace atomových hodin Jan Geršl Český metrologický institut Objasnění některých pojmů Prostoročas Vlastní čas fyzikálního objektu Souřadnicový čas bodů v prostoročase
Relativistické jevy při synchronizaci nové generace atomových hodin Jan Geršl Český metrologický institut Objasnění některých pojmů Prostoročas Vlastní čas fyzikálního objektu Souřadnicový čas bodů v prostoročase
Elektrická impedanční tomografie
 Biofyzikální ústav LF MU Projekt FRVŠ 911/2013 Je neinvazivní lékařská technika využívající nízkofrekvenční elektrické proudy pro zobrazení elektrických vlastností tkaní a vnitřních struktur těla. Různé
Biofyzikální ústav LF MU Projekt FRVŠ 911/2013 Je neinvazivní lékařská technika využívající nízkofrekvenční elektrické proudy pro zobrazení elektrických vlastností tkaní a vnitřních struktur těla. Různé
Kognitivní poruchy u RS. Eva Havrdová 1.LF UK a VFN
 Kognitivní poruchy u RS Eva Havrdová 1.LF UK a VFN Celkový přehled Výskyt kognitivních poruch: 43 70 % pacientů s RS ve všech stadiích choroby Lépe koreluje s progresí nemoci, atrofií CNS Důležitá determinanta
Kognitivní poruchy u RS Eva Havrdová 1.LF UK a VFN Celkový přehled Výskyt kognitivních poruch: 43 70 % pacientů s RS ve všech stadiích choroby Lépe koreluje s progresí nemoci, atrofií CNS Důležitá determinanta
Základy a aplikace digitálních. Katedra radioelektroniky (13137), blok B2, místnost 722
, blok B2, místnost 722") Základy a aplikace digitálních modulací Josef Dobeš Katedra radioelektroniky (13137), blok B2, místnost 722 dobes@fel.cvut.cz 6. října 2014 České vysoké učení technické v Praze, Fakulta elektrotechnická
Základy a aplikace digitálních modulací Josef Dobeš Katedra radioelektroniky (13137), blok B2, místnost 722 dobes@fel.cvut.cz 6. října 2014 České vysoké učení technické v Praze, Fakulta elektrotechnická
Příloha č. 1. amplitudová charakteristika filtru fázová charakteristika filtru / frekvence / Hz. 1. Určení proudové hustoty
 Příloha č. 1 Při hodnocení expozice nízkofrekvenčnímu elektromagnetickému poli (0 Hz 10 MHz) je určující veličinou modifikovaná proudová hustota J mod indukovaná v tělesné tkáni. Jak je uvedeno v nařízení
Příloha č. 1 Při hodnocení expozice nízkofrekvenčnímu elektromagnetickému poli (0 Hz 10 MHz) je určující veličinou modifikovaná proudová hustota J mod indukovaná v tělesné tkáni. Jak je uvedeno v nařízení
doc. Dr. Ing. Elias TOMEH Elias Tomeh / Snímek 1
 doc. Dr. Ing. Elias TOMEH e-mail: elias.tomeh@tul.cz Elias Tomeh / Snímek 1 Frekvenční spektrum Dělení frekvenčního pásma (počet čar) Průměrování Časovou váhovou funkci Elias Tomeh / Snímek 2 Vzorkovací
doc. Dr. Ing. Elias TOMEH e-mail: elias.tomeh@tul.cz Elias Tomeh / Snímek 1 Frekvenční spektrum Dělení frekvenčního pásma (počet čar) Průměrování Časovou váhovou funkci Elias Tomeh / Snímek 2 Vzorkovací
ELCOM, a.s. Energie Pro Budoucnost Ampér 2017
 ELCOM, a.s. Energie Pro Budoucnost Ampér 2017 Doc. Ing. Daniel Kaminský, CSc. Ing. Jiří Hula, Ph.D. Úrovně monitoringu elektrické energie Globální měřicí systémy pro monitoring fázorů (WAMS) na nadnárodní
ELCOM, a.s. Energie Pro Budoucnost Ampér 2017 Doc. Ing. Daniel Kaminský, CSc. Ing. Jiří Hula, Ph.D. Úrovně monitoringu elektrické energie Globální měřicí systémy pro monitoring fázorů (WAMS) na nadnárodní