MODERNÍ BIOFYZIKÁLNÍ METODY:
|
|
|
- Alois Dvořák
- před 9 lety
- Počet zobrazení:
Transkript
1 MODERNÍ BIOFYZIKÁLNÍ METODY: POKROČILÉ PRAKTICKÉ VZDĚLÁVÁNÍ V EXPERIMENTÁLNÍ BIOLOGII Operační program Vzdělávání pro konkurenceschopnost Číslo projektu: CZ.1.07/2.3.00/ Praktický kurz pokročilých metod experimentální biologie konaný na Katedře biologie a ekologie Přírodovědecké fakulty Ostravské univerzity v Ostravě v termínu

2 Program praktického kurzu Pondělí Úterý Středa Čtvrtek Pátek 9-10 přednáška přednáška přednáška přednáška přednáška dopoledne PCR kolonií, specifická detekce plasmidu v buňce. Agarosová gelová elfo.. Detekce jednořetězcových úseků v plasmidové DNA. Interakce s látkami reagujícími s cukrfosfátovou páteří, s bázemi. Využití reparačních enzymů k detekci poškození DNA. Fluorescenční mikroskopie, konfokální mikroskopie Olympus. Elektronová mikroskopie, fluorescenční mikroskopie, AFM, konfokální mikroskopie. odpoledne PCR kolonií, specifická detekce plasmidu v buňce. Agarosová gelová elfo pokračování. Detekce jednořetězcových úseků v plasmidové DNA - pokračování. Interakce s látkami reagujícími s cukrfosfátovou páteří, s bázemi. Využití reparačních enzymů k detekci poškození DNA pokačování. Gelová elektroforéza. Fluorescenční mikroskopie, konfokální mikroskopie Olympus. Vyhodnocení výsledků, hodnocení kurzu účastníky.

3 PCR kolonií Princip: Polymerázová řetězová reakce je metoda, která umožňuje namnožit požadovanou a specifickou sekvenci genomové DNA bez jejího předchozího klonování ve vektorech. Princip metody je založen na replikaci nukleových kyselin. Podstatou je cyklicky se opakující enzymová syntéza nových řetězců vybraných úseků DNA prostřednictvím termostabilní DNA-polymerázy. Požadovaný úsek DNA je vymezen dvěma oligodeoxyribonukleotidy (primery) o délce cca nt. Tyto primery jsou navrženy tak, aby se po denaturaci dsdna vázaly na protilehlé řetězce a vytvořily startovací místa pro syntézu DNA. Po přidání DNA-polymerázy a dntp probíhá syntéza nových vláken na obou templátových řetězcích protisměrně. K syntéze DNA se používají termostabilní DNA-polymerázy izolované z termostabilních mikroorganismů. Tyto enzymy zůstávají v nativním stavu i za teplot, při nichž DNA denaturuje. To umožňuje, aby se syntéza DNA cyklicky opakovala. Průběh PCR lze rozdělit na tři cyklicky se opakující děje s odlišnými nároky na teplotu: a) Denaturace dvouřetězcových molekul DNA (94 C) b) Připojení primerů k odděleným řetězcům DNA (30-65 C) c) Syntéza nových řetězců DNA prostřednictvím DNA-polymerázy (65-75 C) Polymerázová řetězová reakce probíhá v zařízení nazývaném termocykler. V něm se automaticky podle předem zvoleného programu mění teplota v daných časových intervalech. Postupným opakováním jednotlivých cyklů dochází k amplifikaci zvoleného úseku DNA. Počet molekul roste vzhledem k počtu cyklů exponenciálně (2 n ; n = počet cyklů), výsledkem může být až miliarda kopií vybraného úseku DNA. Použití polymerázové řetězové reakce není omezeno na izolovanou DNA, byly vyvinuty techniky, které umožňují amplifikovat zvolený úsek bez předchozí separace nukleových kyselin. Toho se využívá například při tvorbě rekombinantních plasmidů při detekci jednotlivých klonů. Další využití této techniky spočívá v přímé identifikaci bakteriálních kmenů pomocí specifických primerů. Při PCR kolonií z agarové plotny sterilní špičkou odebereme dobře narostlou a oddělenou kolonie, rozsuspendujeme ji v destilované vodě a přeneseme do sterilní PCR zkumavky do směsi pro PCR včetně termostabilní DNA-polymerázy. První denaturační krok při PCR vyvolá lýzi bakterií a uvolněná DNA může být amplifikována. Výtěžek je nižší než při použití izolované DNA, ale dostatečný pro detekci jednotlivých klonů. Materiál a chemikálie: - Plasmidová DNA (pbs, ppgm1) - 10 mm datp, dctp, dgtp, dttp - Taq DNA polymeráza + pufr - Primery - Termocykler - Pipety, špičky - Ledová lázeň - Termostat - Rukavice - Agarosový gel, pufr - Elektroforetická aparatura - Zdroj napětí

4 - Dokumentační systém Pracovní postup: 1 Z agarové plotny setřeme sterilní špičkou dobře narostlou kolonii a suspendujeme v 10 µl destilované vody. Ze suspenze odebereme 1 µl, který přidáme ke směsi PCR. Složení směsi a množství jednotlivých komponent uvádí tabulka. 2 Zkumavky vložíme do termocykleru. 3 Termocykler naprogramujeme na následující hodnoty: a. Krok 1: i. denaturace 94 C 120 sec ii. počet cyklů 1 b. Krok 2: i. Denaturace DNA 94 C 30 sec ii. Hybridizační primerů s templářovou DNA 65 C 30 sec iii. Syntéza nových řetězců 72 C 60 sec iv. Počet cyklů 30 Spustíme start. 4 Po skončení amplifikace zkontrolujeme výtěžek namnoženého úseku pomocí gelové elektroforézy. Na agarosový gel naneseme po 5 µl reakční směsi vzorků. Délku získaných fragmentů určíme porovnáním polohy signálů s polohou fragmentů markerové DNA o známé velikosti. Výtěžek odhadneme z intenzity fluorescence srovnáním s fluorescencí známého množství DNA. 5 Z délky získaných fragmentů Pozn.: 1) V případě odlišných koncentrací výchozích komponent je třeba příslušné objemy přepočítat. Pokud produkty PCR použijeme pouze jako kontrolní vzorky pro gelovou elektroforézu a naším cílem není dosáhnout vysokého výtěžku, můžeme snížit objemy všech látek 5-10 x, tj. PCR provádíme v µl. Snížení spotřeby primerů, templátové DNA i enzymu vede k finančním úsporám. Abychom mohli pracovat v tak malých objemech, je nezbytné mít modernější přístroj s vyhřívaným víkem. U cyklerů první generace, u kterých vyhřívané víko není běžnou součástí, hrozí při příliš malých objemech vypaření vzorků a jejich kondenzace na víčku mikrozkumavky. Nedoporučuje se provádět PCR v objemech větších než 100 µl, protože u velkých objemů je potřeba delších inkubačních časů, aby bylo dosaženo stejné teploty ve všech částech vzorku. Protože i termostabilní DNA-polymeráza v průběhu opakovaných cyklů PCR postupně ztrácí svou aktivitu, při prodloužení inkubačních časů bychom museli do reakční směsi přidat více enzymu. 2) PCR je velmi citlivá metoda, pomocí které dokážeme mnohonásobně zmnožit vybraný úsek DNA, nacházející se ve vzorku ve velmi malém množství. Odvrácenou tváří obrovské citlivosti této metody je možnost vzniku falešně-pozitivních výsledků nebo nespecifických produktů v případě sebemenší kontaminace ať už přímo vzorku nebo prostřednictvím pipet, špiček, přidávaných roztoků apod. Z tohoto důvodu musíme při PCR dbát zvýšené opatrnosti, veškeré roztoky pipetujeme sterilními špičkami, pracovní plochu předem důkladně vyčistíme a při manipulaci se vzorky nikdy nepracujeme bez rukavic.

5 Tabulka PCR: Vzorek č.: Plasmid pbs (100 µg/ml) (Kolonie A) Plasmid ppgm1 (100µg/ml) (Kolonie B) Plasmid c-myc (100µg/ml) (Kolonie C) 10 x cc pufr pro Taq DNA-pol Primer B500for (10 µm) Primer B900 (10 µm) Primer PU27 (10 µm) Primer PY27 (10 µm) Primer p53-con (10 µm) Primer p53-con-com (10 µm) Směs dntp (10 mm) 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 voda 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8 13,8
 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 Primer B900 (10 µm) 1 1 1 - - - - - - - - - - - - Primer PU27 (10 µm) - - - 1 - - - 1 - - - 1 - - - Primer PY27")
6 Detekce jednořetězcových úseků v plasmidové DNA pomocí nukleázy S1 Princip: Některé sekvence DNA mohou za určitých podmínek vytvářet struktury, které jsou odlišné od běžné Watsonovy-Crickovy pravotočivé dvoušroubovice. Mezi tyto tzv. lokální otevřené struktury patří levotočivá DNA, křížová forma DNA, triplex, tetraplex. Předpokládá se, že tyto struktury hrají důležitou roli při regulaci buněčných procesů a expresi genů. Aby lokální otevřené struktury DNA vznikly, musí být v molekule DNA vhodná sekvence [např. levotočivá DNA vzniká v sekvencích v nichž se střídá purinová báze s bází pyrimidinovou (da-dc) n.(dg-dt) n, triplex vyžaduje polypurin.polypyrimidinové úseky (dg n.dc n nebo da n.dt n ), křížová struktura se tvoří v palindromatických sekvencích (sekvence, které se čtou stejně na jednom i druhém řetězci), apod.] a dostatečně vysoké superhelikální napětí. Také okolní prostředí musí splňovat podmínky, které jsou vyžadovány pro vznik jednotlivých struktur: křížová forma často vyžaduje přítomnost hořečnatých iontů, triplex snadněji vzniká v mírně kyselém (ph 5) prostředí nebo v přítomnosti hořečnatých nebo zinečnatých iontů, tetraplex potřebuje draselné ionty Všechny lokální otevřené struktury, jakkoliv se od sebe navzájem liší, mají společné to, že aspoň malá jejich část, nebo úsek k lokální otevřené struktuře přilehlý, obsahuje jednořetězcovou formu. Této vlastnosti se využívá při detekci otevřených struktur. Jedna z metod využívá strukturně selektivní nukleázu S1, která štěpí jednořetězcové úseky, zatímco dvoušroubovici nechává netknutou. Štěpením DNA nukleázou S1 dostaneme z původní superhelikální molekuly molekulu lineární nebo otevřenou. Vhodným výběrem restrikční endonukleázy, kterou necháme štěpit vzniklou lineární nebo otevřenou DNA, a následným elektroforetickým rozdělením získaných fragmentů můžeme určit vzdálenost, ve které se lokální struktura nacházela vůči použité restrikční endonukleáze A B Obr.: (A) Štěpení superhelikálního plasmidu obsahujícího křížovou strukturu nukleázou S1. V dráhách 1, 5 a 6 jsou produkty štěpení nukleázou S1 a restrikční endonukleázou ScaI u plasmidů
 n.(dg-dt) n, triplex vyžaduje polypurin.polypyrimidinové úseky (dg n.dc n nebo da n.")
7 ppgm2 (dráha 1), pbluescript SK- (dráha 5) a ppgm1 (dráha 6). Plasmid pbluescript SK- (pbsk-) neobsahuje sekvenci schopnou vytvořit lokální otevřenou strukturu a slouží jako kontrola. Plasmid ppgm1 vytváří křížovou strukturu méně ochotně v porovnání s plasmidem ppgm2, což se na gelu projeví menším množstvím štěpené DNA. Specifické produkty štěpení jsou na obrázku označeny šipkou. Proužky v dráze 3 vznikly společným štěpením plasmidu pbluescript SK- restrikčními endonukleázami ScaI a HindIII a slouží jako kontrola detekce specifických fragmentů. Dráha 2 linearizovaný pbsk-, dráha 4 superhelikální pbsk-. (B) Křížová struktura plasmidu ppgm2, černě jsou zvýrazněny nukleotidy, kterými se plasmidy ppgm1 a ppgm2 od sebe liší. Materiál: - nukleáza S1 - restrikční endonukleáza - pufry pro S1 nukleázu a restrikční endonukleázu - plasmidy ppgm1, ppgm2, ppgm3 - srážecí směs (EtOH + 3 M CH 3 COONa, ph 5.0) - 80% vychlazený etanol (-20 C) - 10 mm TE; ph voda pro molekulární biologii - elektroforetický pufr (TAE, TBE) - agarosa - mikrozkumavky 1.5 ml - pipety - špičky - rukavice - mrazicí box 80 C - termostat na 37 C - stolní centrifuga - vortex - horizontální elektroforetická aparatura - zdroj napětí Postup: Tab. 1: Vzorek č.: Pozn Plasmid pbs pbs pbs ppgm1 ppgm2 pbs Štěpící enzym - ScaI S1/ScaI S1/ScaI S1/ScaI HindIII/ScaI 1. Do 1.5 ml zkumavek napipetujeme 2 µl plasmidové DNA (1 µg), podle tabulky 2 přidáme (vzorky 3-5) 4 µl 5 x koncentrovaného pufru pro nukleázu S1, 1 µl nukleázy S1 (2U/ µl) a doplníme vodou do 20 µl. Necháme inkubovat 30 minut při 37 C.
 Křížová struktura plasmidu ppgm2, černě jsou zvýrazněny nukleotidy, kterými se plasmidy ppgm1 a ppgm2 od sebe liší.")
8 Tab. 2: Vzorek č.: pbs ppgm ppgm Pufr pro S1 (5x konc.) Pufr pro ScaI (10x konc.) Nukleáza S1 (2U/µl) ScaI (2U/µl) HindIII (2U/µl) Destilovaná voda Vzorky 3-5 přesrážíme: přidáme 2 µl 3 M octanu sodného, ph 5,0 a 65 µl 100% EtOH (-20 C). Vložíme vzorky na 10 minut do hlubokomrazícího boxu (-80 C), stočíme je 15 minut při x g a odstraníme supernatant. Vysráženou DNA promyjeme 200 µl 80% EtOH (-20 C) a opět centrifugujeme při x g (10 minut). Odstraníme supernatant a vzorky vysušíme za sníženého tlaku. 3. DNA ve vzorcích 3-5 rozpustíme v 17 µl vody, přidáme 2 µl 10 x koncentrovaného pufru pro restrikční endonukleázu a 1 µl enzymu (2U/ µl). Necháme štěpit 60 minut při 37 C. Všechny vzorky mají objem 20 µl. 4. Ke vzorkům přidáme 5 µl 6 x koncentrovaného nanášecího pufru, naneseme je na 1% agarosový gel a provádíme elektroforézu 1 hodinu při 120 V. 5. Po skončení elektroforézy gel vložíme do roztoku ethidiumbromidu (1 µg/ml), po 15 minutách jej vyjmeme, promyjeme a zdokumentujeme pomocí gelového dokumentačního systému. Poznámka: - Vzorek 1: kontrola, na gelu indikuje elektroforetickou pohyblivost superhelikální formy plasmidu ppgm1. - Vzorek 2: kontrola, na gelu ukazuje elektroforetickou pohyblivost lineární formy daného plasmidu. - Vzorky 3-5: vzorky, u nichž byla provedena detekce pomocí nukleázy S1. Jednotlivé plasmidy pbs, ppgm1, a ppgm2 se liší pouze 20 pb dlouhou sekvencí v restrikčním místě HindIII (u pbs sekvence chybí, u ppgm1 a ppgm2 se vložená sekvence liší ve dvou nukleotidech). Tato sekvence u jednotlivých plasmidů jeví rozdílnou ochotu vytvářet křížovou strukturu s volnými tyminy, které mohou být modifikovány oxidem osmičelým. - Vzorek 6: kontrola, štěpení endonukleázami HindIII a ScaI vytváří fragmenty, které se svou velikostí velmi blíží očekávaným fragmentům u modifikovaných vzorků. Porovnáním polohy fragmentů u modifikovaných vzorků a kontroly snadno zjistíme, který plasmid byl štěpen nukleázou S1 a tedy pravděpodobně tvořil křížovou strukturu.
, stočíme je 15 minut při 14 000 x g a odstraníme supernatant.")
9 Detekce poškození DNA - detekce jednořetězcových zlomů v plasmidové DNA, detekce modifikace bazí DNA, jako nositelka genetické informace, je v buňce vystavena častým interakcím s látkami, které ji mohou negativně ovlivnit. Tyto látky, které mohou pocházet z buněčného metabolismu nebo z vnějšího prostředí, můžeme zjednodušeně rozdělit do dvou základních skupin: 1) Látky štěpící cukr-fosfátovou kostru 2) Látky modifikující báze Do první skupiny látek, řadíme především radikály a také oxidační činidla. Reakce těchto látek s DNA vede většinou k jedno nebo dvou-řetězcovým zlomům. Radikály jsou vedlejším produktem metabolismu a jsou v buňce velmi časté. Jsou to velmi reaktivní sloučeniny a s DNA v buňce reagují poměrně často. Proto je v poslední době často zmiňována důležitost antioxidantů, které s radikály a jim podobnými látkami v buňce reagují a chrání tak nejen DNA před často ireverzibilním poškozením. Zlomy v DNA jsou snadno detekovatelné pomocí gelové elektroforézy. Přerušení řetězce DNA vede ke změně její elektroforetické mobility. Plasmidová DNA je totiž ve své nativní formě v tzv. superhelikální nadšroubovicové konformaci (scdna). Tato forma se vyznačuje velkou kompaktností (molekula DNA zaujímá při stejné hmotnosti menší objem oproti tzv. otevřené kružnicové ocdna), proto migruje agarozovým gelem rychleji. Při přerušení jednoho z řetězců dvoušroubovice dojde k přechodu z kompaktní scdna do rozměrnější ocdna. Změnu si můžeme představit jako rozmotání klubka s bavlnkou a snahu protáhnout kompaktní klubko, nebo nekompaktní shluk stejně velkým otvorem malých rozměrů. Do látek poškozujících (modifikujících) báze spadá poměrně velká a různorodá skupina chemikálií. Nejčastější modifikace vyvolávají látky s oxidačním, případně alkylačním mechanismem působení. Zvláště v případě methylací mohou být fyziologické důsledky velmi vážne, protože nativní methylace DNA hraje významnou úlohu v epigenetických regulačních procesech. Buňky mají pro opravu poškozené DNA vyvinut poměrně rozsáhlý aparát enzymů. Proces opravy probíhá zpravidla podle schématu rozpoznání modifikace > vystřižení modifikované báze (nukleotidu) -> doplnění chybějící báze (nukleotidu) podle komplementárního řetězce -> ligace přerušeného řetězce (pokud k přerušení došlo). Chceme-li využít reparačních enzymů pro detekci poškození DNA, vynecháváme poslední dva kroky. Vystřižení modifikovaného nukleotidu vede zpravidla k přerušení cukrfosfátové páteře DNA a tedy k přechodu do ocdna. Pro vytvoření volných radikálů v laboratorních podmínkách nejčastěji využíváme tzv. Fentonovy reakce: Me n+ + H 2 O 2 -> Fe n+1 + OH - + OH Ve cvičení bude ke štěpení DNA použit roztok manganistanu draselného, jako druhé štěpící agens použijeme hydroxylové radikály, generované měďnatými ionty (analogie klasické Fentonovy reakce). Pro detekci modifikace bází DNA využijeme dimethylsulfát (methylace guaninu) a reparační enzym Endonukleáza III.

10 Materiál: Roztok měďnatých iontů (50mM) Zásobní roztok KMnO 4 (0,1M) Fosfátový pufr ph 7,4 (0,2M) Plasmidová DNA (pbluesktript (SK - )) TAE (50x) Peroxid vodíku (H 2 O 2 ) Dimethyl sulfát 1% (DMS) Endonukleáza III + pufr 3M octan sodný 96%, 70% ethanol Agaróza Nanášecí pufr Minicentrifuga Termostat Aparatura pro gelovou elektroforézu Chemické sklo Mikrozkumavky Automatické pipety, špičky Destilovaná voda Pracovní postup: A) Vliv oxidačních činidel 1) Připravíme si koncentrační řadu roztoku manganistanu draselného v koncentracích: 100mM, 50mM a 10mM. 2) Do mikrozkumavek 1-4 napipetujeme dle tabulky 3) Krátce zcentrifugujeme (5 s, pomocí tlačítka shortspin ) 4) Vše necháme inkubovat na 37 C po dobu 30 min. 5) Do všech mikrozkumavek napipetujeme 5 µl nanášecího pufru. 6) Celý objem reakční směsi naneseme na připravený agarozový gel. Tab. 1 Rozpis pipetování pro KMnO 4 Zkumavka 1 Zkumavka 2 Zkumavka 3 Zkumavka 4 (kontrola) DNA (200ng/µl) 1 µl 1 µl 1 µl 1 µl Fosfátový pufr 5 µl 5 µl 5 µl 5 µl (200 mm) KMnO 4 2 µl (100 mm) 2 µl (50 mm) 2 µl (10 mm) - H 2 O 12 µl 12 µl 12 µl 14 µl
 Do mikrozkumavek 1-4 napipetujeme dle tabulky 3) Krátce zcentrifugujeme (5 s, pomocí tlačítka shortspin ) 4) Vše necháme inkubovat na 37 C po dobu 30 min.")
11 B) Vliv volných radikálů 1) Připravený roztok měďnatých iontů zředíme na 10 mm. 2) Roztok peroxidu zředíme na 0.3% 3) V mikrozkumavce smícháme: Zkumavka 5 Zkumavka 6 Zkumavka 7 DNA (200ng/µl) 1 µl 1 µl 1 µl Cu 2+ 2 µl 2 µl - H 2 O 2 2 µl - 2 µl H 2 O 15 µl 17µl 17µl 4) Krátce zcentrifugujeme (5 s, pomocí tlačítka shortspin ) 5) Inkubujeme po dobu 30 minut při 37 C. 6) Do všech mikrozkumavek napipetujeme 5µl nanášecího pufru 7) Naneseme na připravený agarozový gel. 8) Do osmého startu naneseme 3 µl standardu (generuler) Obr.1: Poškození plasmidové DNA působením manganistanu draselného. (1) Kontrola DNA, (2) DNA + 12 mm KMnO4, (3) DNA + 6 mm KMnO4, (4) DNA + 3 mm KMnO4, (5) DNA + 2 mm KMnO4, (6) DNA + 1 mm KMnO4, (7) DNA + 0,1 mm KMnO4, (8) DNA + 0,01 mm KMnO 4. C) Detekce modifikace bází 1) V mikrozkumavce smícháme 5µl DNA 2µl fosfátového pufru 2µl 1% DMS 11µl H 2 O 2) Pro negativní kontrolu připravíme stejnou směs, roztok DMS naradíme dest. vodou 3) Směs necháme inkubovat 30min při 37 C 4) Do obou zkumavek přidáme 5µl 3M octanu sodného a 80 µl 96% ethanolu 5) Zkumavky necháme inkubovat 20min na -80 C 6) Centrifugujeme 30min při x g 7) Odpipetujeme obsah zkumavky a necháme minutu schnout dnem vzhůru na filtračním papíru (ubrousku) 8) Přidáme 80 µl 70% ethanolu a centrifugujeme 15min při x g 9) Odpipetujeme obsah zkumavky, necháme minutu schnout dnem vzhůru a vložíme na 5min do exikátoru
 Do osmého startu naneseme 3 µl standardu (generuler) 1 2 3 4 5 6 7 8 Obr.1: Poškození plasmidové DNA působením manganistanu draselného.")
12 10) Do obou zkumavek přidáme 30 µl dest. vody, chvíli protřepeme na vortexu Body 4-10 slouží k tzv. srážení DNA, pomocí tohoto postupu se DNA v organickém prostředí usadí jako sraženina na dně zkumavky a zbytek reakční směsi je odstraněn. V tomto případě provádíme srážení, abychom si byli jisti, že z reakční směsi odstraníme nezreagovaný DMS, který by mohl inhibovat činnost reparačních enzymů, které použijeme v dalším kroku. 11) Z obou zkumavek odebereme 15 µl směsi a přeneseme do dvou čistých eppendorfek 12) K oběma novým zkumavkám přidáme 2 µl pufru pro Endo III a 3 µl roztoku enzymu (naředěného na koncentraci 1u/µl) 13) Směs necháme inkubovat 30min při 37 C 14) Připravíme si 1% agarozový gel 15) Ke směsím přidáme vždy 1/6 objemu nanášecího pufru 16) Naneseme tak, aby dvojice vzorků (s enzymem a bez enzymu) byly na gelu vedle sebe Obr.2: Detekce modifikace bazí za využití Endo III dráhy 1) Kontrolní DNA 2) DNA + 0.1%DMS 3) DNA + 0.1%DMS + 1u EndoIII 4) DNA + 0.1%DMS+3u EndoIII 5) DNA + 0.1%DMS + 5u Endo III 6) DNA + 5u Endo III
 Z obou zkumavek odebereme 15 µl směsi a přeneseme do dvou čistých eppendorfek 12) K oběma novým zkumavkám přidáme 2 µl pufru pro Endo III a 3 µl roztoku enzymu (naředěného na koncentraci 1u/µl)")
13 Základy světelné mikroskopie

14 Účel mikroskopu 1)Zvětšení 2)Rozlišení 3)Kontrast
Rozlišení")
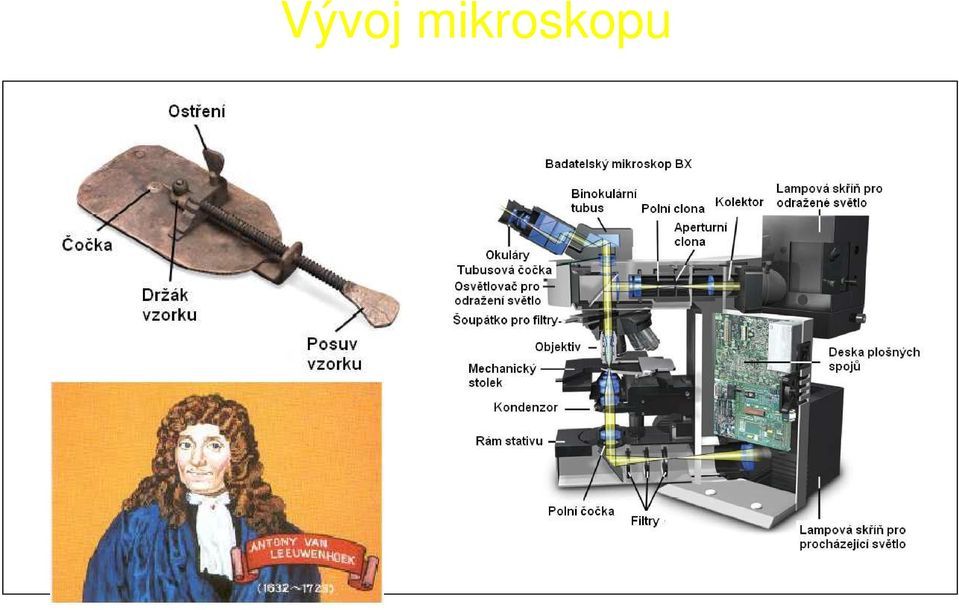
15 Vývoj mikroskopu
16 Princip mikroskopu
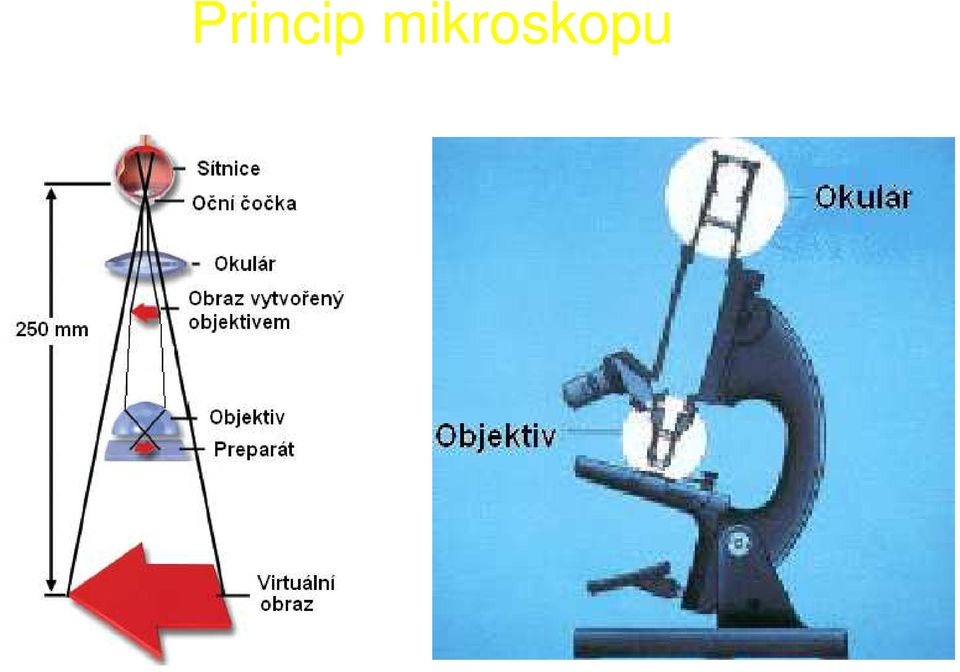
17 Zvětšení mikroskopu Celkové zvětšení mikroskopu = zvětšení objektivu zvětšení okuláru Příklad: Objektiv 20, okulár 10 celkové zvětšení 200

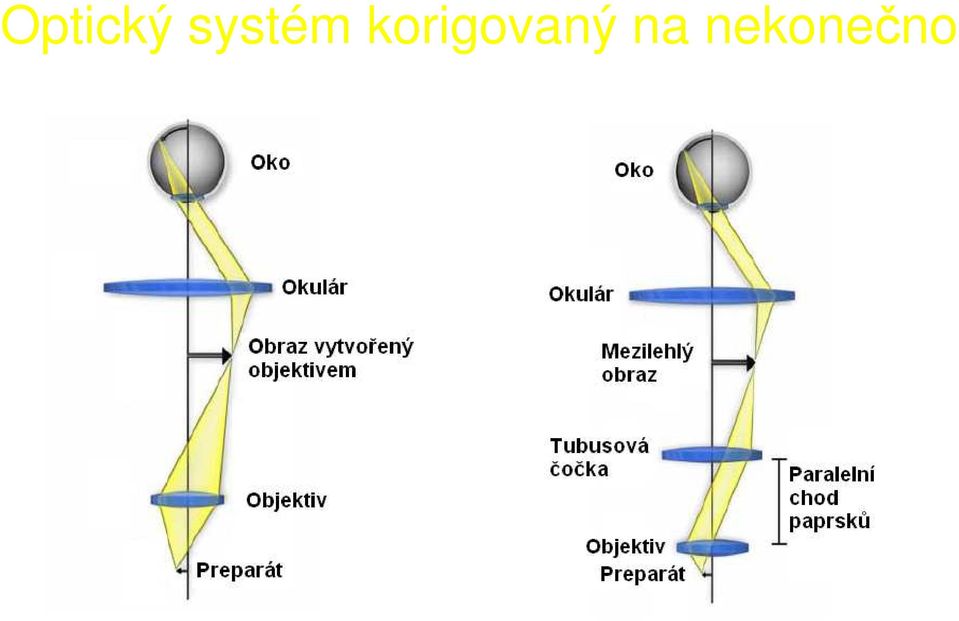
18 Optický systém korigovaný na nekonečno
19 Sestava 1

20 Sestava 2

21 Anatomie mikroskopu
22 1) Objektiv
23 Objektivy Nejdůležitější optická součást mikroskopu. Vytváří reálný převrácený obraz preparátu. Značení objektivů
24 Zvětšení objektivů Typy imerze 4, 5 (proužek blíže k preparátu) Olej 40, 50 Voda , 250
25 Typy objektivů Objektiv by měl poskytovat co nejkvalitnější zobrazení. Kvalita objektivu je určena stupněm korekce optických aberací a rovinností obrazového pole. Plan Ach Fl Apo
26 Základní optické aberace Chromatická vada Sférická vada
27 Korekce objektivů Achromáty - chromaticky pro modré a červené - sférická vada pro zelené světlo - nejlepší obraz pro monochromatické zobrazení přes zelený filtr Fluority (semi-apochromáty) - chromaticky pro modré, zelené a červené světlo - sférická vada pro modré a zelené světlo - vhodné pro barevné zobrazení Apochromáty - chromaticky pro krátkovlnné modré, modré, zelené a červené světlo - sférická vada pro krátkovlnné modré, modré a zelené světlo - nejvyšší kvalita barevného zobrazení
28 Rovinnost obrazového pole Důležitá zejména pro záznam obrazu. Rozdíl mezi Ach a Plan Ach
29 Numerická apertura objektivu Numerická apertura (N.A.) = n sinα N.A. 1 nelze dosáhnout bez imerze. Se zvětšením roste numerická apertura objektivu.
30 Hloubka pole Je maximální axiální interval, uvnitř kterého jsou jednotlivé hladiny preparátu zároveň zaostřené. Hloubka pole klesá s rostoucí numerickou aperturou objektivu.
31 2) Okuláry
32 Číslo pole (F.N.) Okuláry dále zvětšují obraz vytvořený objektivem. Pevná clona je v místě mezilehlého obrazu. Ramsdenův typ okuláru umožňuje snadnou montáž měřítka pozor na prach! Průměr clony v mm udáváčíslo pole. Výpočet průměru zorného pole v rovině preparátu: D = F.N. / zvětšení objektivu
33 Prázdné zvětšení Celkové zvětšení 1000 N.A. Příklad 1: Objektiv 40, N.A. = 0,65 Okuláry = O.K. Příklad 2: Objektiv 100, N.A. = 1,3 Okuláry = 1500 > 1300 Okuláry 12, ,5 = O.K. Pozor na další zvětšení při přenosu digitálního obrazu na monitor!
34 3) Kondenzor
35 Funkce kondenzoru Soustřeďuje světlo ze světelného zdroje do kužele, který: 1) rovnoměrně osvětluje preparát 2) odpovídá numerické apertuře objektivu Při změně objektivu je vhodné upravit N.A. kondenzoru aperturní clonou. N.A. kondenzoru = 80% N.A. objektivu
36 Typy kondenzorů Abbeův - nejjednodušší, N.A. = 1,25 - není korigován chromaticky ani sféricky - vhodný pro jednoduché objektivy Aplanaticko-achromatický korigován pro obě vady Speciální kondenzory pro různé kontrastní metody (DF, Ph, DIC, RC, POL). Výklopnáčočka pro velmi malá zvětšení (do 4 ).
37 Typy mikroskopů Podle povahy pozorovaných preparátů se postupně vyvinuly různé typy mikroskopů: Klasický (Up-right) Invertovaný Stereo Speciální systémy (např. konfokální)
38 Mikroskopy up-right Pro pozorování v procházejícím světle Pro pozorování v odraženém světle Sklíčkové preparáty Mikroskopy třídy CX2, BX2, AX
39 Invertované mikroskopy Pro pozorování v procházejícím světle Pro pozorování v odraženém světle Preparáty v kultivačních miskách, mikrotitrační destičky Mikroskopy třídy CKX2, IX2
40 Stereomikroskopy Pro pozorování v odraženém světle Pro pozorování v procházejícím světle 3-D preparáty Mikroskopy třídy SZ2, SZX
41 Speciální systémy Například: Konfokální mikroskop FV1000, FV10i Laserový mikrodisekční systém MicroBeam Laserové cytometry LSC, icyte, icys Systém Cell-R pro metodu TIRFM
42 Kontrastní metody Slouží k zvýšení kontrastu obrazu tak, aby byl dobře pozorovatelný. Nejpoužívanější metody: Temné pole Fázový kontrast Polarizované světlo Reliéfní kontrast Diferenciální interferenční kontrast Fluorescence
43 Fluorescence
44 Druhy výbojek
45 Princip konfokální mikroskopie LASER Pin Hole Detector Objectiv Above focus plane Focus plane Below focus plane
46 Fluorescence/konfokál Fluorescenční mikroskopie Konfokální mikroskopie
47 Mikroskop = stavebnice
48 Vybrané informační zdroje M. Abramowitz, Microscope: Basics and Beyond. Olympus America, Inc., Melville, New York, Revised M. Abramowitz, Fluorescence Microscopy: The Essentials. Olympus America, Inc., Melville, New York, M. Abramowitz, Reflected Light Microscopy: An Overview. Olympus America, Inc., Melville, New York, Olympus Microscopy Resource Center:
49 OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 1
50 Vlastnosti Jedinečný kompaktní design Plně automatizovaný konfokální mikroskop Vysoká kvalita obrazu Jednoduchá obsluha Interaktivní nápověda rychlé zaškolení OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 2
51 Vlastnosti Objektivy: UPLSAPO 10 Ph NA 0.40 UPLSAPO 60 Water Ph NA 1.20 UPLSAPO 60 Oil Ph NA 1.35 Optický zoom: pro objektiv 10 1 až 6, krok 0,1 Optický zoom: pro objektiv 60 1 až 10, krok 0,1 Rozsah zvětšení: 10 až 600 Motorizovaný x-y stolek i ostření OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 3
52 Vlastnosti Lasery: 405 nm (18mW), 473 nm (12,5mW), 559 nm (15mW), 635 nm (10mW) Dvoukanálový spektrální detektor TD pro fázový kontrast Autofokus & automatická expozice Mapování OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 4
53 FV10i OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 5
54 Schéma FV10i Temná místnost Skenovací jednotka Mikroskop Antivibrační deska Lasery OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 6
55 Produktová řada pouze dva modely FV10i-O: objektiv UPLSAPO 60 O-Ph snímání obrazů s vysokým rozlišením FV10i-W: objektiv UPLSAPO 60 W-Ph, inkubátor (37 C, CO 2 ) snímání živých buněk a časosběrné snímání OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 7
56 Pracovní prostředí OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 8
57 Intuitivní ovládání Vložit vzorek OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 9
58 Intuitivní ovládání Vložit vzorek Zvolit oblast OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 10
59 Intuitivní ovládání Vložit vzorek Zvolit oblast Nasnímat obraz OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 11
60 Režimy snímání Obrazy jsou průběžně snímány v předdefinovaných intervalech => časosběrné snímání Obrazy jsou opakovaně snímány v různých rovinách ostrosti lze vytvořit 3D obraz Kombinace proostřování a časosběrného snímání Automatickéčasosběrné snímání ve více předdefinovaných oblastech Kombinace všech tří předchozích režimů OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 12
61 Summary Shrnutí FV10i All-in-one integrovaný systém Zabudovaná temná místnost Úspora místa Mobilita Intuitivní ovládací software Interaktivní nápověda Jednoduchá obsluha rychlé zaškolení OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 13
62 Děkuji za pozornost OLYMPUS CZECH GROUP Nový konfokální mikroskop OLYMPUS FluoView FV10i 14
63 Fluorescenční mikroskopie v genotoxikologii Comet assay; Micronucleus test; Metoda FISH
64 SCGE Single Cell Gel Electrophoresis / Comet assay Charakteristika: Molekulárně genetická metoda umožňující detekovat poškození DNA typu: - jednoretězcových zlomů DNA (SSB ) (zlomy přímé, ale i zlomy přechodné, které vznikají v důsledku neúplné excizní opravy), - alkali-labilní polohy (ALS), - křížové DNA vazby(cross-linking)- DNA/DNA nebo DNA/protein, - oxidační a alkylační poškození DNA - poškození DNA související s apoptósou Analýzu lze provést v buňkách všech organismů - bakteriálních, u kvasinek, v rostlinných buňkách, živočišných i lidských. A ve všech typech buněk - v proliferujících i neproliferujících, ve stabilizovaných buněčných líniích, primokulturách, lymfocytech, v buňkách z biopsií. K analýze postačuje relativně malé množství buněk (< ).
65 Princip SCGE Molekula DNA má záporný náboj, proto v elektrickém poli migruje směrem k anodě - kladný pól. Rychlost migrace DNA v elektrickém poli závisí na velikosti molekuly a tedy na počtu zlomů. V Comet assay se využívá při identifikaci malých fragmentů DNA vzniklých působením mutagenů v jednotlivých buňkách.
66 Průběh SCGE testu Mikroskopování Buňky Fixace na agarozový gel Barvení Lyze buněk V gelu zůstává superhelikální DNA + zbytky jaderné membrány a matrix Neutralizace Aklalické rozvolnění při ph >13. Zlomy navodí vznik relaxované DNA ELEKTROFORÉZA VZNIK KOMETY
67 VZNIK KOMETY DENATURACE ELEKTROFORÉZA IZOLOVANÉ IZOLOVANÉ JÁDRO JÁDRO JÁDRO JÁDRO S S RELAXOVANOU RALAXOVANOU DNA DNA KOMETA KOMETA
68 Mikroskopické hodnocení Velikost a tvar komety závisí na míře poškození DNA
69 Hodnocení SCGE Head DNA (%) Tail DNA (%) Tail Length (µm) Tail Moment (µm)
70 Mikroskopická analýza Hodnocení se provádí fluorescenčním mikroskopem při zvětšení 25x nebo 40x. K barvení se používají fluorescenční barvy s afinitou k DNA. Každý fluorochrom vyžaduje vlastní excitační a emisní filtry.
71 a) Vizuální hodnocení preparátů Analyzuje se 100 komet na jeden gel (celkově 300 hodnot). Komety se zařazují do 5 kategorií podle tvaru komety.. Stanovuje se průměrná hodnota poškození (P) P = Σ (0 x a) + (1 x b) + (2 x c) + (3 x d) + (4 x e) kde: 0 až 4 označení kategorie poškození a = počet buněk kategorie 0 b = počet buněk kategorie 1 c = počet buněk kategorie 2 d = počet buněk kategorie 3 e = počet buněk kategorie 4
72 Pět kategorií hodnocení komet Extrémní hodnoty poškození při vizuálním hodnocení jsou : 0 nebylo zaznamenáno poškození všechny komety patří do třídy 4.
73 b) Hodnocení obrazovou analýzou Hodnotí se obvykle menší počet komet minimálně na sklíčko. Průměrnou hodnotu stanoví program. Program obvykle stanoví %DNA v kometě, délku chvostu komety a tzv. tail moment
74 počítačové zpracování
75 In vitro SCGE test Lze detekovat somatické i gametické mutace Teoreticky lze využít kterýchkoli buněk Nejčastěji se využívá buněčných kultur - bakterie, kvasinky - nebo buněčných linií exp. zvířat myší lymfocyty, buňky ovariií, plicní buňky čínského křečka, hepatocyty. Lze využít i rostlinných buněk např. Nicotiana tabacum var. xanthi Lze aplikovat in vitro metabolickou aktivaci postmitochondriální frakcí S9 Buňky jsou vystavovány koncentrické řadě sledovaného mutagenu lze zjistit závislost dávka efekt Positivní výsledek indikuje, že v testovaných podmínkách substance indukuje poškození DNA. Výhodou testu je, že není zapotřebí znát karyotyp.
76 In vivo SCGE test Výhoda hodnotí se malý počet buněk z příslušné tkáně lze stanovit lokální genotoxickou citlivost. Lze hodnotit buňky laboratorních zvířat, ale i zvířat z volného prostředíryby apod., včetně lidských leukocytů a lymfocytů.
77 Aplikační možnosti SCGE (Dusinska et Colins 2008)
78 Využití SCGE v optimalizaci léčby některých CA onemocnění Výsledky hodnocení individualní citlivosti tumoru k radiaci a chemoterapeutickým látkám s cílem ušít efektivní léčbu na míru ( radiosensitivita (Fisher et al., 2007, chemo-sensitivity (Smith et al., 2007) tab. podle McKenna 2008)
79 Perspektivy využití SCGE v ekotoxikologii Většina testů je založena na hodnocení genotoxicity v laboratorních podmínkách na experimentálních organismech problém s interpretací výsledků a zevšeobecněním jejich platnosti. In vivo Comet assay dovoluje hodnotit změny DNA v buňkách různých organismů ovlivněných mutageny in situ v reálných podmínkách. Lze hodnotit nejen somatické, ale i gametické změny DNA ve spermatických buňkách tím se rozšiřuje využití na hodnocení vlivu genotoxikantů a na reprodukční schopnosti a fekunditu. Využití v ekotoxikologii bude však vyžadovat optimalizaci metody pro přirozenou biotu, validaci a inter-laboratorní harmonizaci testu, výběr vhodných druhů žijících v různých nikách s různými potravními a reprodukčními strategiemi.
80 Využití SCGE v prevenci ochraně zdraví Hodnocení protektivních effektů různých potravních faktorů v chemo-preventivních studiích (Bichler et al., 2007). V kombinaci s určitými bakteriálními enzymy (např.formamidopyrimidin glykosylasou, endonukleasou III, uracil-dna glykosylasou), které rozpoznávají oxidační poškození purinových a pyrimidinových bazí, se využívá SCGE pro identifikaci oxidativních DNA změn, které mohou být příčinou různých zdravotních problémů (Collins et al., 2001, Kruman et al., 2002).
81 Mikronukleus test (MN) in vitro krátkodobý test Charakteristika MN test je založen na tvorbě mikrojader ve stimulovaných živočišných nebo lidských periferních lymfocytech. Vyžaduje, aby se buňky dělily. Hodnocení se provádí v metafázi/anafázi mitózy. Mikrojádra vznikají: - v důsledku chromozómových zlomů a jsou pak tvořena acentrickým fragmentem, - poruchou funkce dělícího vřeténka (mikrojádro je tvořeno celým chromozómem nezačleněným do nově vytvořeného jádra). Mikrojádra se analyzují výhradně v těch lymfocytech, které prošly pouze jedním buněčným dělením. Analyzuje se frekvence mikrojader ve dvoujaderných buňkách.
82 Možnosti využití MN-testu Mikrojádra jsou považována za biomarker efektu a uplatňující se v risk assessmentu (rakoviny) Umožňuje identifikaci chromozomových + genomových mutací Rozlišení jednojaderných a dvoujaderných buněk se provádí tzv. cyto-b assay po aplikaci cytochalasinu-b V jednojaderných buňkách jsou změny vzniklé in vivo před kultivací buněk. Ve dvoujaderných buňkách jde o změny před kultivací buněk + změny vytvořené v průběhu kultivace, tj. v průběhu první in vitro mitozy.
83 Princip MN
84 Fluorescenční in situ hybridizace (FISH) Charakteristika Metoda umožňuje detekci získaných strukturálních chromozómových přestaveb - aberací v savčích a lidských buňkách. Stabilní chromozomální přestavby nezpůsobují ztrátu chromozomálního materiálu, mohou se při dělení přenášet z mateřské na dceřiné buňky, kumulují se v organismu a jejich počet je odrazem dlouhodobé expozice genotoxickými látkami. Je nutno znát karyotyp Metoda FISH je nejčastěji využívána v diagnostice, prognostice; v hodnocení průběhu a efektu léčby některých chorob, rakoviny a studiu genetických syndromů.
85 Princip FISH Lymfocyty jsou pozorovány v metafázi. Preparáty jsou hybridizovány s fluorescenčně značenými celochromozomovými sondami (tzv. painting).
86
87 Mikroskopická analýza Chromozómové aberace se hodnotí ve fluorescenčním mikroskopu při zvětšení 1 000x za použití imerzního oleje. Vyhodnocuje se 1000 metafázních buněk u každého vzorku (daného jedince). Analyzují se jen na první pohled úplné metafáze s dobrým hybridizačním signálem. Je velmi vhodné snímat aberantní buňky pomocí CCD kamery do počítačového programu umožňujícího analýzu obrazu
88 Přeskupeni na chromozomech 9 a 22 Hodnocení translokace BCR / ABL pro chronickou myeloitidu
89 Značení buněk 4 různými sondami
90 Kombinace testů MN a FISH
91 Baterie testů Comet assay - MN - FISH Kombinací lze docílit upřesnění charakteru poškození DNA. Vzniká-li kometa budou také vznikat mikrojádra? Kombinace Comet - MN dovoluje upřesnit charakter poškození DNA (tj. zda jde o chromozomovou aberaci nebo vznik acentrického chromozomu). V kombinaci s fluorescenční in situ hybridizací (Comet-FISH), můžeme determinovat sekvence nebo specifické genetické změny a reparace (McKenna et al., 2003, Kumaravel and Bristow, 2005)