Mendelova univerzita v Brně
|
|
|
- Libor Tábor
- před 5 lety
- Počet zobrazení:
Transkript

1 Mendelova univerzita v Brně Lesnická a dřevařská fakulta Ústav nauky o dřevě Xylogeneze buku lesního (Fagus sylvatica L.) na Drahanské vrchovině Bakalářská práce Brno, 2011 Václav Osička
2 Prohlašuji, že jsem bakalářskou práci na téma: Xylogeneze buku lesního (Fagus sylvatica L.) zpracoval sám a uvedl jsem všechny použité prameny. Souhlasím, aby moje bakalářská práce byla zveřejněna v souladu s 47b Zákona č. 111/1998 Sb., o vysokých školách a uložena v knihovně Mendelovy univerzity v Brně, zpřístupněna ke studijním účelům ve shodě s Vyhláškou rektora MZLU a archivaci elektronické podoby závěrečných prací. Autor kvalifikační práce se dále zavazuje, že před sepsáním licenční smlouvy o využití autorských práv díla s jinou osobou (subjektem) si vyžádá písemné stanovisko univerzity o tom, že předmětná licenční smlouva není v rozporu s oprávněnými zájmy univerzity a zavazuje se uhradit případný příspěvek na úhradu nákladů spojených se vznikem díla dle řádné kalkulace V Brně, dne: podpis studenta: 2
3 Děkuji panu Ing. Vladimíru Grycovi, Ph.D. za ochotu, vstřícnost, praktické rady a odborné vedení mé práce a dále panu Ing. Hanuši Vavrčíkovi, Ph.D. za cenné informace, které mi poskytoval při psaní mé studie. V neposlední řadě mé poděkování patří panu Ing. Ladislavu Menšíkovi, Ph.D. z Ústavu ekologie lesa za poskytnutí odborných údajů týkajících se výzkumné plochy Rájec Domanka. 3
4 Abstrakt Václav Osička, Xylogeneze buku lesního (Fagus sylvatica L.) na Drahanské vrchovině Studie se zabývá xylogenezí buku lesního (Fagus sylvatica L.) na Drahanské vrchovině, výzkumná plocha Rájec Domanka. Předmětem práce je zjištění dynamiky radiálního přírůstu xylému na buněčné úrovni a kambiální aktivity v průběhu vegetačního období V pravidelných týdenních intervalech v mezi proběhl odběr mikrovývrtů trephorem po obvodu kmene z vybraných vzorníkových stromů. Trvalé preparáty byly zkoumány pomocí světelného mikroskopu. Dělivá činnost kambia začala mezi při průměrné teplotě vzduchu 9,1 C předchozího týdne. Kambium dosáhlo v průběhu maximální šířky (12 buněk kambiální zóny). Kambiální činnost byla ukončena v rozmezí při průměrné teplotě vzduchu 17,72 C za předcházející týden. Průměrná šířka plně vytvořeného letokruhu byla dle Gompertzovy funkce 1887 μm. Průměrný vypočtený denní přírůst xylému činil 33,40 μm a průměrná doba nezbytná pro vytvoření většiny buněk xylému dle Gompertzovy funkce byla 60 dní. Klíčová slova: xylogeneze, buk lesní (Fagus sylvatica L.), mikrovývrt, trephor, světelný mikroskop, kambium, letokruh, xylém Abstract Václav Osička, Xylogenesis of common beech (Fagus sylvatica L.) on the Drahan highland The studies is about xylogenesis of common beech (Fagus sylvatica L.) on the Drahan highland, Rájec Domanka site. The object of the work is findings of the dynamic of radial xylem growth on cell level and of the cambial activity at intervals of vegetation period in At permanent week periods between there was taking of microdrilling by trephor around perimeter from tree samples. The preparations were investigatid by light microscope. The camibal cell activity began 4
5 between at the temperature 9,1 C in the last week. The cambial got maximal width 12 cell. The cambial activity was ended between at the temperature 17,72 C in the last week. The average width of the fully created annual ring was by Gompertz function 1887 μm. The average day growth was 33,40 μm and the time necessary for creation of most xylem cells by Gompertz function was 60 days. Keywords: xylogenesis, common beech (Fagus sylvatica L.), microdrill, trephor, light miscroskope, cambium, annual ring, xylem 5
6 Obsah 1 Úvod Cíl práce Literární přehled Taxonomie druhu Fagus sylvatica L Dendrologická charakteristika buku lesního (Fagus sylvatica L.) Popis Ekologie a rozšíření Význam Anatomická stavba dřeva listnatých dřevin Cévy Tracheidy Libriformní vlákna Parenchymatické buňky Dřeňové paprsky Axiální dřevní parenchym Makroskopická a mikroskopická stavba dřeva buku lesního (Fagus sylvatica L.) Makroskopická stavba Mikroskopická stavba Kambium Diferenciace anatomických elementů dřeva Rostlinná buňka Buněčná stěna Chemické složení buněčné stěny Struktura buněčné stěny Xylogeneze výsledky předcházejících výzkumů Materiál a metodika Charakteristika výzkumné plochy Rájec Domanka Odběr vzorků Úprava vzorků před krájením Krájení vzorků na mikrotomu Podvojné barvení Vyhodnocení preparátů pod mikroskopem Výsledky Vyhodnocení výsledků vzorníku B Vyhodnocení výsledků vzorníku B Vyhodnocení výsledků vzorníku B Srovnání výsledků všech vzorníků Zachycení tvorby sekundární buněčné stěny pomocí polarizačního světla Aplikace Gompertzovy funkce
7 6 Diskuse Závěr Summary Literatura Přílohy
8 1 Úvod Dřevo je jedním z nejstarších přírodních materiálů, kterému v přírodě náleží velice specifické postavení. Již od nepaměti ho začal člověk využívat ke svému životu, proto se na něj nemůžeme dívat jako na pouhou spotřební surovinu. Každý druh dřeviny se vyznačuje charakteristickou stavbou dřeva a s ní souvisejícími vlastnostmi, které zpravidla určují, dle aktuální poptávky, k jakým účelům se bude využívat. Proto, abychom mohli vhodně a hospodárně dřevo využít, musíme důkladně znát jeho fyzikální a mechanické vlastnosti, chemické složení a v neposlední řadě růstové schopnosti dřevin, které jsou závislé na klimatických a půdních podmínkách. V poslední době se stále více diskutuje o globálních klimatických změnách a jejich dopadech na světový ekosystém. Pod pojmem klimatické změny si nelze představit pouze oteplování, ale především intenzivní srážky, abnormální výkyvy počasí, velká sucha, nárazový vítr atd. Všechny uvedené faktory mají vliv jak na vitalitu, tak na růst a celkovou produkci lesních dřevin. Na lesní dřeviny, jakož to dlouhověké rostliny, mají klimatické změny podstatně větší vliv, nežli na rychle se obnovující generace rostlin žijících jedno či několik vegetačních období. Se zvyšujícím se zájmem o užívání přírodních materiálů a díky přirozeným, nenahraditelným vlastnostem dřeva, kterými se nevyznačuje téměř žádný jiný materiál, roste v globálním měřítku zájem o tuhle výjimečnou surovinu a proto je účelné se zabývat růstovými schopnosti dřevin na daných lokalitách. 8
9 2 Cíl práce Cílem práce bylo analyzovat v pravidelných týdenních intervalech radiální tvorbu xylému a charakteristiku kambiální činnosti buku lesního (Fagus sylvatica L.) během vegetačního období 2010 s ohledem na věk daného jedince a klimatické podmínky na Drahanské vrchovině, výzkumná plocha Rájec Domanka. Dále bylo cílem analyzovat časový interval vzniku cév, proces jejich diferenciace a čas jejich plné lignifikace. 9
10 3 Literární přehled 3.1 Taxonomie druhu Fagus sylvatica L. Jako první buk lesní (Fagus sylvatica L.) popsal ve svých spisech Karl Linné (1753). Buk tehdy zařadil do XXI. třídy Rostliny o jediné tyčince, nebo tyčinka stojí na stopečce, pod níž je listen. Uvedl, že:,, Buk, Fagus sylvatica L., o kůře šedé, hladké. Listy vejčité, lysé, lesklé, jen po okraji pýřité. Prašné květy v jehnědě kulovité, na dlouhé stopce převislé. Plodem ostnitá tobolka, čtyř chlopňová; chová tří hrané bukvice ʼʼ. Říše: Plantea (Rostliny) Podříše: Tracheobionta (Cévnaté rostliny) Oddělení: Magnoliophyta (Krytosemenné rostliny) Třída: Rosopsida (Vyšší dvouděložné rostliny) Řád: Fagales (Bukotvaré) Čeleď: Fagaceae (Bukovité) Rod: Fagus (Buk) Druh: Fagus sylvatica L. (Buk lesní) 3.2 Dendrologická charakteristika buku lesního (Fagus sylvatica L.) Popis Jedná se o strom velkých rozměrů, s rovným válcovitým kmenem, s tenkou, šedou borkou. Koruna je vysoko nasazená, u volně rostoucích jedinců je zpravidla kulovitá, v porostu metlovitá. Vlivem okusu zvěří vytváří i zakrslé keřovité porosty. Buk dosahuje výšek kolem m a průměru kmene až 1,5 m. Dožívá se maximálně věku let. Kmen bývá vysoko do koruny průběžný a větve odstávají v ostrém úhlu. Na volném prostranství je obvykle kmen široce větvený. Letorosty jsou zprohýbané, bělavě pýřité, později úplně lysé. Střídavé listy jsou eliptické, 5 10 cm dlouhé, 10

11 celokrajné až mělce zubaté, zašpičatělé, na bázi zaokrouhlené až široce klínovitě zúžené. Listy jsou ve stínu ploše rozložené, s tenkou čepelí. Na podzim se buk nápadně barví; nejdříve žlutě, pak červeně a posléze tmavohnědě. Samčí květy se nachází v paždí listů v dlouze stopkatých svazečcích, samičí květy po dvou v červené číšce zevně porostlé dlouze chlupatými, později dřevnatějšími výrůstky. Buk je dřevina jednodomá, plodem jsou trojboké nažky (bukvice) uložené po dvou v dřevnaté číšce otvírající se třemi chlopněmi. Na volném prostranství začíná buk plodit mezi 20. a 40. rokem. Plodná období se vyskytují nepravidelně ve více intervalech (5 až 10 let). Výmladnost je hojná jen v mládí a přetrvává cca do let. Kořenový systém označujeme jako srdčitý. Díky danému (srdčitému) kořenovému systému buk netrpí vývraty, bývá v půdě velmi dobře zakotven. Výjimku tvoří živná stanoviště, na kterých buk vytváří mělký kořenový systém. Na kořenech se nachází ektotrofní mikorrhiza, která zajišťuje lepší příjem vody a minerálních látek (Úradníček et al. 2001). Obr. 1. Dendrologické vyobrazení buku lesního (Kőhler, 1914) Ekologie a rozšíření Buk patří mezi C stratégy, dřeviny s vysokou konkurenční schopností. Snáší i silný zástin, proto na příznivých stanovištích vytlačuje buk většinu ostatních dřevin, což vede ke vzniku čistých bučin. Náhlé vystavení kmenů ze zástinu plnému slunci má za následek korní spálu. Buk má střední nároky na vláhu v půdě. Buk nesnáší stanoviště s vysokou hladinou spodní vody. Vyžaduje dostatek srážek a zvlášť v letním období musí mít dostatečnou relativní vlhkost vzduchu. Proto na chladném severu stačí buku asi 500 mm srážek, zatímco na jihu areálu jeho výskytu musí srážky obnášet nejméně mm ročně (Úradníček a Chmelař 1995). Je indiferentní k geologickému podkladu roste skoro na všech druzích hornin. Nevyskytuje se na suchých písčitých 11
12 půdách, na silně oglejených a uléhavých půdách, dále pak mu nevyhovují půdy rašelinné a jílovité. Nejlepší bučiny jsou na dobrých humózních půdách. Buk často dává přednost vápencům, pokud je dostatek srážek. Buk má také značné nároky na provzdušněnost půdy. Je středně citlivý na znečištění ovzduší a nesnáší jakékoliv znečistění půdy (Nekolová, 2002). Buk je dřevina, které vyhovuje mírně oceanické klima. Na východě končí rozšíření buku na hranici výslovně kontinentálního klimatu a i ve střední části areálu se vyhýbá oblastem s příliš suchými a horkými léty. Nedaří se mu v mrazových kotlinách a v místech s nebezpečím pozdních mrazů (Úradníček a Chmelař 1995). Celé naše území leží uvnitř areálu buku a proto tuto dřevinu nalezneme ve všech středohořích a horských oblastech hercynské i karpatské části států. Buk vytváří v našich podmínkách v nadmořských výškách cca od m často nesmíšené porosty, na spodní hranici rozšíření se mísí s dubem a na horní se smrkem a jedlí (Úradníček et al. 2001) Význam Buk s celkovým zastoupením 7,21 % ( ha) z plošné rozlohy listnatých dřevin ČR patří spolu s dubem k našim nejdůležitějším hospodářským dřevinám (Zpráva o stavu lesa a lesního hospodářství České republiky, 2009). Cenné sortimenty získáváme především z oddenkové části kmene, ostatní části se zpracovávají na palivo či celulózu. Bukové dřevo má všestranné použití (Úradníček et al. 2001). Vyrábí se z něho dýhy a překližky, pražce, parkety, sudy, nábytek, hračky atd. Je využíván v celulózo papírenském průmyslu. Speciální využití se týká u nás tradiční výroby ohýbaného nábytku. Nevýhodou je silná sesýchavost a rychlé zvětrávání dřeva. U buku se často vyskytuje nepravé jádro, které dřevo znehodnocuje (Úradníček a Chmelař 1995). Bukové dřevo nepravého jádra (bez příznaků hniloby) se svými fyzikálními a mechanickými vlastnostmi neliší od zdravého dřeva, avšak některá nepravá jádra mají vyšší obsah vlhkosti. Je-li zdravé, lze ho použít bez omezení na výrobu různých dřevařských výrobků (Nekolová, 2002). 12
13 3.3 Anatomická stavba dřeva listnatých dřevin Dřevo (xylém) je v obecném pojetí centrální část kmenů (větví, kořenů) dřevin, která se nachází mezi kambiem a dření. Je produktem dělivého pletiva kambia. Dřevem se dále rozumí systém rostlinných pletiv s převážně zdřevnatělými buněčnými stěnami (Gričar, 2007). Ve srovnání s dřevem jehličnatým je dřevo listnatých dřevin vývojově mladší a má složitější anatomickou stavbu. Dřevo listnáčů je tvořeno větším počtem typů buněk, které jsou velikostí a tvarem úzce specializovány a přizpůsobeny své funkci. Dřevo listnatých dřevin se skládá z cév (tracheje), tracheid (cévic), libriformních vláken a parenchymatických buněk (Šlezingerová a Gandelová 2002) Cévy Počátek tvorby letokruhu je u buku charakterizován vznikem cév (Oladi a Pourtahmasi 2010). Cévy (tracheje) jsou typické vodivé elementy vyskytující se jen v listnatých dřevinách, kde mají v bělové části dřeva specifickou vodivou funkci (vedou vodu spolu s rozpuštěnými minerálními látky od kořenů do koruny stromů). V jádrovém dřevě plní funkci zásoby vody, můžou být vyplněny jádrovými látkami, thylami, popřípadě mohou být vyplněny pouze vzduchem (Chovanec, 1977). Thyly jsou vakovité útvary vyrůstající z okolních parenchymatických buněk vyklenutím a dorůstáním dvojtečky. Cévy se skládají z tzv. cévních článků. Jedná se o systém vertikálně uložených mrtvých buněk, mezi kterými se rozrušily membrány dvojteček. U buku nám cévy tvoří tzv. letokruhovou síť. Nepřerušená, tenkostěnná céva složená z cévních článků většinou nepřesahuje 10 mm, ale u některých dřevin dosahuje větší délky (u buku 0,8 až 2 m). Z celkového objemu bukového dřeva zaujímají cévy % (Požgaj et al. 1997). Zbytky příčných přehrádek mezi jednotlivými cévními články nazýváme perforace. Pokud se v přehrádce vytvořil pouze jeden velký otvor a zbytek původní buněčné stěny je ve tvaru prstence na vnitřním obvodě cévy, nazývá se taková perforace jednoduchá. Jestliže po částečném rozrušení buněčné stěny v ní zůstane řada přehrádek, mezi kterými jsou otvory, nazýváme takovou perforaci složenou (popřípadě žebříčkovitou či síťovitou). U buku může být perforace jak jednoduchá (v jarním dřevě) tak žebříčkovitá i síťová (v letním dřevě). Stěny cév mohou mít jednoduché či spirálovité 13
14 ztluštěniny. Ztlustlá místa obvykle dřevnatí a dodávají dřevu pevnost (Matovič, 1981) Tracheidy Tracheidy (cévice) vytváří ve dřevě listnáčů přechodné typy anatomických elementů s funkcí vodivou, mechanickou, popřípadě zásobní. Dle tvaru a funkčního zaměření rozlišujeme tracheidy cévovité, vláknité a vazicentrické. Jejich zastoupení ve dřevě listnáčů je velmi rozdílné, v případě buku je výskyt všech tří typů tracheid velmi výrazný (Gandelová et al. 2009). Cévovité tracheidy tvoří přechod mezi cévními články cév listnáčů a jarními tracheidami dřeva jehličnanů. Doprovázejí cévy a slouží k vedení vody. Jsou celistvé, tlustostěnné, bez koncových otvorů a s malými cévními články. Jejich délka zpravidla nepřesahuje 5 mm (Wagenführ, 1989). Vazicentrické tracheidy jsou krátké buňky nepravidelného tvaru, které se vyskytují v bezprostřední blízkosti cév. Nemůžou samostatně vytvářet žádné axiální vodivé dráhy (Požgaj et al. 1997). Vláknité tracheidy představují přechod mezi letními tracheidami jehličnanů a libriformními vláknami listnáčů. Jedná se o uzavřené, zašpičatělé, tlustostěnné buňky, které se tvarově velmi podobají libriformním vláknům. Mají především mechanickou funkci, ale můžou plnit i funkci jak vodivou tak zásobní. Ve dřevě buku tvoří podstatnou část základního pletiva (Šlezingerová a Gandelová 2002) Libriformní vlákna Libriformní vlákna jsou axiálně uložené buňky, s proměnlivou tloušťkou buněčné stěny, které jsou specializované na mechanicky zpevňovací funkci. Diferencovaná libriformní vlákna tvoří mrtvé buňky vyplněné vzduchem či vodou. Libriformní vlákna jsou často na koncích pevně propojeny. U buku jsou konce vláken zoubkovaná, čímž dosahují pevnějšího spojení. V podélném směru můžou být vzájemně stlačeny tak, že se navzájem překrývají. Na příčném řezu vytváří hranatý, ojediněle kulatý tvar (Wagenführ, 1989). Podíl libriformních vláken ve dřevě je velmi rozdílný, odvíjí se od druhu dřeviny. U buku dosahuje podíl libriformních vláken 37 až 44 %. Po diferenciaci z kambia 14
15 libriformní vlákna ihned odumírají, výjimku tvoří javor, u kterého se v okolí cév vyskytují živá vlákna s méně zdřevnatělými buněčnými stěnami. Zpravidla mají vřetenovitý a rourkovitý tvar, který je variabilní jak mezi dřevinami tak i v rámci jednoho druhu dřeviny (Požgaj et al. 1997). Délka libriformních vláken a tloušťka jejich buněčných stěn se zvyšuje ve směru od jarního k letnímu dřevu (ke konci vegetačního období nepatrně klesají). Délka libriformních vláken se udává 0,2 2,2 mm, šířka μm. U dřevin s roztroušeně pórovitou stavbou dřeva je průměrný délkový přírůst v rámci letokruhu %. Tloušťka buněčných stěn a délka se také zvyšují ve směru od dřeně k obvodu kmene (s věkem), zatímco od báze kmene ke koruně se tyto dva rozměry snižují (Šlezingerová a Gandelová 2002) Parenchymatické buňky Parenchymatické buňky u listnatých dřevin zaujímají 8 35 % z celkového objemu dřeva. Jsou více zastoupeny ve dřevě listnáčů a taky jsou tvarově různorodější jak u jehličnanů. Plní ve dřevě funkci zásobní (ukládá škroby, tuky, bílkoviny) a vodivou. Hojně se zde vyskytují krystaly anorganických látek. Dle orientace rozeznáváme dva základní typy dřevního parenchymu: axiální nebo-li podélný dřevní parenchym (buňky orientované podél osy kmene) a radiální (buňky orientované kolmo na osu kmene a směrovány ke dřeni), který nám tvoří dřeňové paprsky. V důsledku poranění kambia se vytváří tzv. traumatický parenchym, který obklopuje poraněná místa a ve dřevě se vytvářejí dřeňové skvrny (Matovič, 1981) Dřeňové paprsky Dřeňové paprsky jsou u listnatých dřevin hojněji zastoupeny a jsou v podstatě mohutnější než u jehličnanů. Jsou tvořeny výhradně živými parenchymatickými buňkami seskupeny kolmo na letokruhy. Na stěnách buněk dřeňových paprsků jsou četné jednoduché tečky, zejména v místech styku s cévami. Slouží k vedení a ukládání zásobních látek parenchymatických ve směru buněk kolmém na rozlišujeme podélnou dva osu základní kmene. typy Dle tvaru dřeňových paprsků: homogenní a heterogenní. Homogenní jsou složené z buněk morfologicky (tvarově) stejného typu buď z tzv. stojatých (buňky jsou orientované delší osou svisle) 15
16 nebo ležatých (obdelníkovité buňky orientované delší stranou horizontálně). Ve dřevě buku se vyskytují homogenní dřeňové paprsky jako tomu je u většiny našich listnáčů (Gandelová et al. 2009). Heterogenní dřeňové paprsky jsou tvořeny z morfologicky odlišných typů buněk. Jedná se o kombinaci ležatých, stojatých a čtvercových parenchymatických buněk. Axiálně protáhlé (stojaté) buňky se většinou vyskytují na obvodu dřeňových paprsků. V rámci stromu se šířka a výška dřeňového paprsku ve směru od obvodu kmene ke dřeni snižuje a naopak od báze ke koruně zvyšuje (Matovič, 1981) Axiální dřevní parenchym Zastoupení axiálního (podélného) dřevního parenchymu je ve dřevě listnáčů velmi variabilní. V průměru se zastoupení pohybuje v rozmezí 10 % objemu dřeva. Je tvořen parenchymatickými buňkami a to buď hranolovitého, krychlovitého, nebo vřetenovitého tvaru, jejichž podélná osa či charakteristické seskupení je orientováno rovnoběžně s podélnou osou kmene. Nepostradatelnou zásobní funkci vykonávají v bělovém dřevě nebo v obvodových částech kmene. Ukládají zejména škrob, v jádrovém dřevě můžou obsahovat jádrové látky. Buňky podélného parenchymu jsou s buňkami dřeňových paprsků propojeny tečkami, přes které je do nich ukládán škrob a jiné zásobní látky (Wagenführ, 1989). Parenchymatické buňky podélného dřevního parenchymu jsou ve dřevě různě uspořádány. Obecně se používají dvě základní členění. V prvním případě se rozlišuje dle kontaktu parenchymatických buněk s ostatními anatomickými elementy dřeva axiální parenchym apotracheální, paratracheální a svazkový. V druhém případě členění vychází opět z kontaktu parenchymatických buněk s vodivými elementy a rovněž se zohledňuje kontakt s ostatními elementy podílející se na tvorbě základního pletiva. Dle těchto faktorů rozlišujeme axiální dřevní parenchym na paratracheidní, paratracheální kontaktní a interfibrilární (Šlezingerová a Gandelová 2002). 16
17 3.4 Makroskopická a mikroskopická stavba dřeva buku lesního (Fagus sylvatica L.) Makroskopická stavba Letokruh se skládá z vrstvy jarního a letního dřeva. Jarní dřevo vzniká na začátku vegetačního období, je světlejší, řidší a pórovitější. Později tvořící se letní dřevo je tmavší a užší. Optický kontrast mezi vrstvou tmavého letního dřeva a světlého jarního dřeva následujícího vegetačního období umožňuje letokruh ohraničit (Lesnický naučný slovník, 1994). Buk řadíme mezi roztroušeně pórovité dřeviny s vyzrálým dřevem a bělí. Tyhle dřeviny mají z dřevin mírného pásu nejméně výrazné letokruhy. Letokruhy jsou nejčastěji zvýrazněné jen velmi úzkou tmavší (nebo světlejší) vrstvou pozdního letního dřeva (Požgaj et al. 1997). Cévy jsou nezávisle na velikosti rovnoměrně rozptýleny po celé šířce letokruhu. Menší cévy nejsou na jednotlivých řezech makroskopicky zřetelné. Dřeňové paprsky jsou široké, velmi dobře zřetelné na transverzálním řezu jako různě široké světlé proužky, paprsčitě se rozbíhající ze dřeně kolmo na letokruhy. Na radiálním řezu jsou dřeňové paprsky nápadné jako hustě nad sebou rozložená zrcátka. Na tangenciálním řezu rozlišujeme dřeňové paprsky jako svislé, načervenalé čárky, až 2 mm vysoké (Grosser, 1977). Bukové dřevo patří mezi středně těžká dřeva (ρ = 685 kg.m-3) a dřeva středně tvrdá (61 MPa). Barva dřeva je narůžovělá až červenohnědá. Dřevo je méně trvanlivé a málo odolné proti abiotickým činitelům. Snadno se impregnuje, paří a moří, obtížněji se suší (Šlezingerová, 1999) Mikroskopická stavba Cévy jsou velmi četné a hustě seřazené, ojediněle se vyskytují radiálních skupinkách po čtyrech. Širší cévy v jarním dřevě mají jednoduchou perforaci, užší cévy v pozdním dřevě mají žebříčkovitou perforaci. V cévách se vyskytují jak thyly tak jádrové látky. Dřeňové paprsky jsou heterogenní, značně široké, vysoké a jsou ve dvou různých velikostech: jednovrstevné a menší počet je mnohovrstevných (až 25 vrstev). Na hranici letokruhu jsou vždy rozšířené. Buňky dřeňových paprsků mají na vztyčených stěnách s cévami velké jednoduché tečky. Dřevo obsahuje velmi četné, vláknité, cévovité a vazicentrické tracheidy, které tvoří přechody k libriformním vláknům. Dřevní parenchym doprovází tracheje i tracheidy a je i bohatě rozptýlen mezi dřevními vlákny. 17
18 Typ axiálního parenchymu apotrachealního je hraniční, difúzní (rozptýleně nakupený) a axiálního parenchymu paratrachealního je skupinový (Balabán, 1955). 3.5 Kambium Termín kambium představuje laterální (boční) dělivé pletivo, tvořící souvislou vrstvu mezi dřevní a lýkovou částí vodivého systému produkující centripetálně (směrem ke dřeni) nové buňky dřeva a centrifugálně (směrem k obvodu kmene) nové buňky lýka (Gryc et al. 2010). Kambium obecně zahrnuje jak kambiální iniciály (tj. buňky, které dodávají kambium trvalý meristematický charakter) tak deriváty dřeva a lýka (buňky z nich oddělené) (Matovič 1981). Počet vrstev nediferencovaných buněk kambia (celé kambiální zóny) se odvíjí od druhu dřeviny a dle daného vegetačního období. V období vegetačního klidu se vrstva kambiální zóny skládá z 3 až 6 řad buněk, v průběhu vegetačního období může mít v některých dřevinách 15 až 20 řad buněk (Požgaj et al. 1997). Kambium obstarává tvorbu buněk dřeva, lýka, dřeňových paprsků, tloušťkový přírůst kmene (přírůst v radiálním směru) a současně zabezpečuje tvorbu kambiálních iniciál s ohledem na zvyšující se obvod kmene (Šlezingerová a Gandelová, 2002). Rozeznáváme dva základní typy kambiálních iniciál. Jedná se o vřetenovité kambiální iniciály a iniciály dřeňových paprsků. Vřetenovité kambiální iniciály tvoří protáhlé, axiálně orientované buňky, na průřezu zploštělého obdelníkovitého nebo mnohoúhelníkovitého tvaru, na obou koncích šikmo nebo střechovitě ukončené. Dle druhu dřeviny vykazují značnou tvarovou i rozměrovou různorodost. U listnatých dřevin se délka vřetenovitých kambiálních iniciál pohybuje v rozmezí 0,14 1,62 mm (Chovanec et al. 1977). S věkem se jejich délka zvyšuje. Takřka stejnorozměrné (izodiametrické) iniciály dřeňových paprsků vznikající příčným dělením nebo postranním oddělením (menší radiální překážkou) z vřetenovitých iniciál. Oba typy kambiálních iniciál vykazují stejnou buněčnou strukturu, která se podobá struktuře parenchymatických buněk (Gandelová et al. 2009). Kambium zahajuje svoji činnost dříve než začne vlastní dělení buněk. Začátek činnosti kambia se odvíjí od druhu dřeviny a je v úzké korelaci s činností pupenů. U listnatých dřevin s roztroušeně pórovitou stavbou dřeva je kambium činné několik týdnů při bázi pupenů. Po vyrašení listů začíná činnost ve výhoncích, postupuje do větví 18
19 a potom na bázi kmene. Je tedy zřejmé, že rychlost aktivace kambia bezipetálně do pupenů je různá a závislá jak na druhu dřeviny, tak na vnějších podmínkách stanoviště (Matovič, 1981). V závislosti na klimatických podmínkách pracuje kambium zejména jen ve vegetačním období. V našich klimatických podmínkách se jedná cca o 6 měsíců v roce. V období zimního vegetačního klidu jsou jeho dílčí schopnosti omezeny. Vzhledem k tomu se rozlišuje klidová a aktivní fáze kambia (Wagenführ, 1989). Doba činnosti kambia je závislá na druhu dřeviny, stanovišti a konkrétních klimatických podmínkách příslušného vegetačního období (limitující faktory jsou zejména délka fotoperiody, teplota a srážky). Na začátku vegetačního období je tvorba nových buněk pozvolná, je-li kambium již plně aktivováno probíhá dělení rychleji a koncem vegetačního období postupně ustává. K prvnímu dělení v kambiu dochází u přezimujících mateřských buněk dřeva a lýka vytvořených v předcházejícím vegetačním období. U dřevin s roztroušeně pórovitou stavbou dřeva se dělí asi s měsíčním předstihem mateřské buňky lýka a až potom dřeva (Gandelová et al. 2009). 3.6 Diferenciace anatomických elementů dřeva Proces diferenciace v určitý anatomický element dřeva nastává v okamžiku, kdy mateřské buňky dřeva, oddělené kambiálními iniciálami nebo deriváty mateřských buněk, začnou zvětšovat svoje rozměry a ztratí meristematický charakter (dělivou schopnost). Na základě daného procesu přestávají být součástí kambiální zóny. Anatomické elementy dřeva orientované vertikálně (rovnoběžně s podélnou osou kmene) vznikají tehdy, jakmile dojde k diferenciaci buněk oddělených vřetenovitými kambiálními iniciálami, případně mateřskými buňkami dřeva. Diferenciací buněk oddělovaných iniciálami dřeňových paprsků se tvoří anatomické elementy dřeva orientované horizontálně kolmo na podélnou osu kmene (Šlezingerová a Gandelová 2002). Z kambiální vrstvy dceřiných buněk dřeva se postupně diferencují různé typy buněk. Proces diferenciace má zhruba tři fáze. a) Po dělení probíhá povrchový, nebo plošný růst buněk, dokud nenabudou finální tvar a rozměr. Z vřetenovité kambiální iniciály se může u listnatých dřevin vytvořit libriformní vlákna, široká céva nebo parenchymatické buňky axiálního parenchymu. Diferenciací buněk oddělených iniciálami dřeňových 19
20 paprsků se u listnatých dřevin vytváří parenchymatické buňky dřeňových paprsků či parenchymatické buňky horizontálních kanálků. b) V druhé fázi probíhá tloustnutí buněčné stěny (tvorba sekundární buněčné stěny S1, S2, S3) ukládáním krystalické celulózy a buňka přechází již do fáze dozrávání. c) Po ukončení přírůstu buněčné stěny nastává její lignifikace. Délka procesu diferenciace se převážně pohybuje v období 3 až 4 týdnů. V procesu diferenciace jsou veškeré buňky živé. Ve většině případů buňky, které se vytvořily z vřetenovitých iniciálů ( libriformní vlákna, tracheje, tracheidy) ztrácejí po fázi lignifikace životnost. Naproti tomu většina parenchymatických buněk si zachovává buněčný obsah a životnost na několik let, tedy na různě dlouhou dobu, která závisí na dřevině a na růstových podmínkách (Požgaj et al. 1997). Obr. 2. Schéma diferenciace tracheid jarního a letního dřeva jehličnanů, kde je znázorněna kambiální zóna (A), dělící se floém (B), dělící se xylém (C), zóna radiálního rozšiřování (D), zóna dozrávajících buněk (E), zóna zralého dřeva (F), letní dřevo předcházejícího letokruhu (G) (Wodzicky, 1971). 20
21 Z výše uvedeného obrázku (Obr.2) je patrné, že nově tvořící se letokruh můžeme podle charakteru diferencujících se buněk na základě znaků patrných na příčném řezu dřeva rozdělit na zóny: a) zónu dělících se mateřských buněk dřeva eventuálně jejich derivátů v kambiální zóně b) zónu diferenciační, zahrnující fázi radiálního rozšiřování buněk a dozrávajících buněk c) zónu zralého dřeva Jednotlivé fáze procesu diferenciace anatomických elementů dřeva je možná časově rozlišit tj. stanovit dobu radiálního rozšiřování buněk a dobu dozrávání buněk. Časové intervaly jsou však platné jen pro konkrétní stromy a stanoviště, na kterém rostly v dané vegetační době (Gandelová et al. 2009). 3.7 Rostlinná buňka Rostlinná buňka je základní struktura a fyziologická jednotka rostlinného organismu, která je tvořena buněčnou stěnou a protoplastem. Protoplast obsahuje cytoplazmu, mající charakter bezbarvé tekuté látky, v níž je jádro (nucleus) a ostatní buněčné organely (Chovanec et al. 1977). Přítomnost buněčné stěny je základní charakteristikou rozlišující rostlinné a živočišné buňky. U živých rostlinných buněk je cytoplazmatický obsah v pohybu jedná se o cytoplazmatické proudění, neboli cyklózu. Cyklóza zabezpečuje výměnu látek uvnitř buňky i mezi buňkou a prostředím (Procházka, 2006) Buněčná stěna Produktem základní cytoplazmy je buněčná stěna a všechny její stavební složky jsou výsledkem metabolické aktivity této základní cytoplazmy buněk. Vznik buněčné stěny je spjat s dělením buněk (cytokinezí), která navazuje na jaderné dělení (káryokinézu) (Gandelová et al. 2009). Dle přítomnosti buněčné stěny rozlišujeme rostlinné a živočišné buňky. Buněčná stěna omezuje velikost protoplastu a má specifické funkce, které jsou nezbytné jak pro existenci buňky, tak pro celistvost rostlin (Procházka, 2006). 21
.")
22 Chemické složení buněčné stěny Dřevo je různorodá látka s buněčnou strukturou, která úzce souvisí s jeho celým chemickým složením (Chovanec et al. 1977). Buněčná stěna dřevin se skládá ze tří základních složek: celulózy, hemicelulózy a ligninu. Orientační podíl všech tří složek je patrný z následujícího obrázku (Obr. 3). Základní charakteristickou složkou buněčných stěn je celulóza, která určuje jejich architekturu (Procházka, 2006). Obr. 3. Přibližný podíl celulózy, hemicelulózy a ligninu v jednotlivých vrstvách buněčné stěny listnatých dřevin (Blažej, 1975). Celulóza je hlavní stavební složkou rostlinných buněčných stěn a je v přírodě nejrozšířenějším sacharidem. Zpravidla je doprovázena ligninem, hemicelulózami, pektiny, rostlinnými gumami, které zpevňují celulosová vlákna (Jankovský, 1999). Celulóza tvoří fibrilární kostru buněčných stěn. Vláknité makromolekuly celulózy se pomocí vodíkových vazeb seskupují ve vyšší strukturální jednotky (fibrily), které jsou odlišně uloženy v jednotlivých vrstvách buněčné stěny (Gandelová et al. 2009). Dlouhé a tenké řetězce molekul glukózy vytváří mikrofibrily, široké nm. Celulóza má krystalické vlastnosti, což určuje uspořádání molekul celulózy v tzv. micelách tvořící mikrofibrily (Procházka, 2006). Hemicelulóza je jedna z hlavních sacharidických složek dřeva a zároveň i složka nejméně stálá. Je to dáno její amorfní nadmolekulovu stavbou, která oproti krystalické celulóze je méně rezistentní k hydrolýze. Proti celulóze má menší molekulovou hmotnost. Ve dřevě buku lesního (Fagus sylvatica L.) je hemicelulózy obsaženo cca 22,5 % (Jankovský, 1999). Hemicelulóza zahrnuje dvě hlavní skupiny pentózany a hexózany, které rozdělují polysacharidy buněčné stěny dle vlastních cukrových složek, přičemž pentózany náleží listnatým a hexózany jehličnatým dřevinám (Melcerová, 22
23 1970). Mezi celulózou a hemicelulózami existují chemické vazby. Hemicelulózy působí jako tmelící (penetrační) faktor celulózové kostry buněčných stěn a částečně se také podílí na vlastní struktuře (Gandelová et al. 2009). Další význačnou složkou buněčné stěny je lignin, který je kromě celulózy hojně vyskytujícím se polymerem v rostlinách (Procházka, 2006). Pod pojmem lignin rozumíme aromatický podíl dřeva, který není možno hydrolyzovat kyselinami a který vyvolává zdřevnatění (Melcerová, 1970). Je uložen v submikroskopických dutinách celulózové kostry, kterou díky své trojrozměrné struktuře dobře vyplňuje. Po stránce fyzikální slouží lignin ke zvýšení pevnosti buněčné stěny (Gandelová et al. 2009). Důležitou složkou buněčné stěny je i glykoprotein, který je bohatý na aminokyselinu hydroxyprolin. Mimo jiné se nachází v buněčných stěnách i kutin, suberin a vosky, tvořící vnější ochranné pletiva rostlinného těla. Buněčné stěny mohou být inkrustovány anorganickými látkami (SiO2, CaCO3) (Procházka, 2006) Struktura buněčné stěny Rostlinné buněčné stěny se liší tloušťkou v závislosti na úloze buněk v rostlině a v závislosti na jejich stáří. Ve všech rostlinných buňkách rozlišujeme dvě vrstvy: střední lamelu a primární stěnu (Procházka, 2006). První viditelnou strukturou při buněčném dělení, která odděluje dceřiné cytoplasty je plazmatická destička. V důsledku následné diferenciace buněčné stěny se mění plazmatická destička na střední lamelu (SL), která vzniká jako první přehrádka mezi dceřinými buňkami. Střední lamelu tvoří hlavně pektiny a hemicelulóza (později silně lignifikuje) (Gandelová et al. 2009). Na střední lamelu je základní cytoplazmou vyloučena další souvislá vrstva primární stěna (Gandelová et al. 2009). Primární stěna (P) kromě celulózy, hemicelulózy a pektinů obsahuje glykoprotein. U primární stěny může docházet k lignifikaci. Po skončení růstu buněk ve fázi jejich dozrávání se vytváří sekundární buněčná stěna (S). Vytváří se tzv. apozicí (přikládáním nových vrstev (lamel), na již vytvořené struktury), čímž dochází k tloustnutí buněčné stěny směrem dovnitř buněk. Tato stěna na rozdíl od primární stěny neobsahuje glykoprotein a pektinové látky, díky kterým se stává neohebná (neelastická). Důležitou částí sekundární stěny je lignin, který 23
. Obr. 4. Struktura buněčné stěny (Požgaj et al. 1997). Tab.")
24 u dřevin tvoří % sušiny (Procházka, 2006). Sekundární stěna se skládá ze tří vrstev (S1, S2, S3), které se liší jak tloušťkou, chemickým složením, tak orientací celulózních fibril. Ze strany lumenu buněk je vrstva S3 pokryta bradavičnatou vrstvičkou označovanou jako vrstva S4, nebo tak W vrstva (Gandelová et al. 2009). Obr. 4. Struktura buněčné stěny (Požgaj et al. 1997). Tab. 1. Tloušťka (μm) a podíl (%) vrstev buněčné stěny jednotlivých anatomických elementů dřeva buku lesního (Wagenfűhr, 1989). libriformní vlákna vláknité tracheidy axiální dřevní parenchym parenchym dřeň. paprsků (%) (μm) (%) (μm) (%) 0,07 1 0,06 4 0,5 27 0, , ,5 50 4, , , , ,1 2 0, , ,09 5 0,07 3 vrstvy buněčné stěny (μm) (%) (μm) (%) (μm) P 0,07 5 0,25 25 S1 0,24 16 S2 0,99 67 S3 0,17 W cévy 24
25 3.8 Xylogeneze výsledky předcházejících výzkumů Vzhledem k jednoduché anatomické stavbě byla ve většině případů z časového hlediska činnost kambia a diferenciace xylémové části v závislosti na vnějších faktorech zkoumána především u jehličnatých dřevin. Analýza problémů týkajících se činnosti kambia u listnáčů s roztroušeně či kruhovitě pórovitou stavbou dřeva je v závislosti k jejích složitější anatomické stavbě značně obtížná. Danou problematikou u jehličnatých dřevin, konkrétně na smrku ztepilém (Picea abies (L.) Karst.), se ve stejném geomorfologickém celku na výzkumných plochách Rájec Jestřebí zabýval Matovič (Matovič, 1985), Horáček (Horáček et al. 1996), Gryc (Gryc et al. 2010). Matovič (1985) uvedl, že aktivace kambia začala až při průměrné teplotě vzduchu 8 ºC po dobu několik dnů, latentní vrstva kambia se skládala z 2 5 buněk a v průběhu vegetačního období dosáhlo kambium 3 6 buněk. Naproti tomu Gryc et al. (2010) stanovili zahájení činnosti kambia u jednotlivých sledovaných stromů od 16. do , kdy v týdnu před neklesla teplota pod 6,2 ºC a počet buněk v kambiální zóně v průběhu vegetačního období dosahoval stejných hodnot jako u Matoviče (1985). Matovič (1975) zkoumal charakteristikou činnosti kambia a jeho xylémových derivátů u 1 6 letých semenáčků jasanu úzkolistého (Fraxinus angustifolia Vahl.) a jasanu ztepilého (Fraxinus excelsior L.). Zjistil, že počátek činnosti kambia bylo možno pozorovat pří déle trvacích teplotách 9 11 ºC před zahájením této činnosti a že kambiální produkce stoupá postupně s věkem. Naopak počet buněk kambia v rámci schopností jedince a jeho sociálního postavení v porostu postupně klesá (Matovič, 1975). Ve Slovinské republice, v lesích Panška Reka se zabývali kambiální aktivitou a diferenciací xylémové částí buku lesního (Fagus sylvatica L.) Čufar a Prislan (2008). Zjistili, že kambium (počet buněk v kambiální zóně) započalo svou činnost v průběhu druhé poloviny dubna ( ), maximálního rozměru dosáhlo koncem května a od do ukončilo svoji dělivou činnost. Autoři ale neuvádějí průměrné denní teploty související s činností kambia. Průměrná šířka letokruhu byla u sledovaných vzorníků dle Gompertzovy funkce 2552 μm a čas nezbytný pro jeho formaci byl 100 dní (Čufar a Prislan 2008). Stejný výzkum v lesích Iránu na buku východním (Fagus orientalis Lipsky) v průběhu vegetačního období 2008 provedli Oladi et al. (2010). Začátek činnosti kambia se pohyboval u všech vzorníků od 16. do , maximálního 25
26 počtu kambiálních buněk dosáhlo okolo , ale konec činnosti kambia byl datován až od konce srpna do poloviny listopadu (Oladi et al. 2010). Analýzu činnosti kambia a xylémové části provedli v městském prostředí Ljubljany u javoru mléče (Acer platanoides L.) v závislosti na vnějších faktorech Marion et al. (2008). Daný druh dřeviny byl rozdělen do dvou skupin a to na stromy poškozené (či vnějším faktorem negativně ovlivněné) a na stromy vitální, zdravé. Kambiální aktivita u poškozených stromů byla zahájena již začátkem dubna (před 19. 4) a ukončena v polovině července ( ), přičemž u zdravých stromů javoru byla reaktivace kambia stanovena v druhé polovině dubna ( ) a konec činnosti kambia začátkem září ( ). Celková doba trvání činnosti kambia byla u zdravých stromů o 7 týdnů delší (Marion et al. 2008). Horáček et al. (2003) v lužních lesích v blízkosti Lednice na Moravě stanovili na dubu letním (Quercus robur L.) začátek kambiální aktivity již před při průměrné teplotě vzduchu vyšší jak 8 ºC, přičemž ukončení činnosti kambia váže na datum, kdy nebyl zaznamenán žádný radiální přírůst xylému (konec diferenciace dřevních vláken), tedy od konce září do začátku října. Naproti tomu Gričar (2007) zaznamenala u dubu (Quercus sessiliflora Salisb.) reaktivaci kambia koncem března (30. 3.) a ukončení její činnosti stanovil na druhou polovinu srpna (23. 8.). Čufar et al. (2011) sledovali v průběhu vegetačního období 2008 kambiální aktivitu a radiální přírůst xylému u kaštanovníku (Castanea sativa Mill.). Vzorníky pocházely z nadmořské výšky 850 m n. m., z lesní oblasti Itálie Monti Cimini. Aktivace kambia začala již začátkem dubna, kdy latentní kambium obsahovalo 7 6 buněk. Počet buněk kambiální zóně se v průběhu vegetačního období měnil, přičemž kambium dosáhlo svého maxima (10 12 buněk) na přelomu dubna a května. Konec kambiální činnosti, kdy kambium obsahovalo 5 6 buněk, zaznamenali na začátku října. Nejintenzivnější radiální přírůst byl zachycen od druhé poloviny května do první poloviny června. Průměrná šířka plně vytvořeného letokruhu byla 2027 μm (Čufar et al. 2011). 26
27 4 Materiál a metodika 4.1 Charakteristika výzkumné plochy Rájec Domanka Jedná se o smíšený buko smrko jedlový porost ve věku okolo 130 let, který náleží k geomorfologickému celku Drahanská vrchovina, spadající do oblasti Brněnská vrchovina. Porost se nachází se v nadmořské výšce kolem 630 m n. m., (PLO) 30, v jedlo bukové LVS na stanovišti původních smíšených lesů. Zeměpisná poloha N: ,371 a E: ,492. Daný porost spadá do lesů Mensdorff Pouilly (MP), Lesy Benešov u Boskovic, LHC MP lesy, polesí Benešov, lesní úsek 07 Boskovice. Půdním typem je kambizem modální oligotrofní (Kamd ) s formou povrchového humusu moder. Fyziologická hloubka půdního profilu je cm (Menšík, 2010). Z typologického hlediska se porost nachází na lesním typu 5S1 svěží jedlová bučina šťavelová (Plíva, 1987). V porostu výzkumné plochy se vyskytuje buk lesní (Fagus sylvatica L.) (50%), smrk obecný (Picea abies (L.) Karst.) (43%), jedle bělokorá (Abies alba Mill.) (7%) s vtroušenou borovicí lesní (Pinus sylvestris L.) a modřínem opadavým (Laryx decidua Mill.) s zakmeněním porostu 7 (Menšík, 2010). Následující graf (Obr. 5) ukazuje vztah průměrné teploty vzduchu k průměrné půdní vlhkosti v průběhu roku Obr. 5. Klimadiagram znázorňující vztah průměrné teploty vzduchu k průměrné půdní vlhkosti v roce
. Jednalo se o reprezentativní vzorníky z mýtního porostu, které byly rovnoměrně rozmístěny po ploše.")
28 4.2 Odběr vzorků Odběr vzorků byl uskutečněn ze tří vybraných vzorníkových stromů (značení: B1, B2, B3) buku lesního (Fagus sylvatica L.). Jednalo se o reprezentativní vzorníky z mýtního porostu, které byly rovnoměrně rozmístěny po ploše. Jedinci nebyli nijak výrazně poranění a nebyly na nich zpozorovány žádné fyziologické změny. Odběr probíhal v pravidelných týdenních intervalech v průběhu vegetačního období od do Z každého vzorníku byly týdně odebrány dva vzorky. Vzorky byly odebrány ve výčetní výšce speciálním nástrojem trephorem (Obr. 6), spirálovitě kolem obvodu kmene. Jedná se o nástroj sloužící k odběru vzorků (mikrovývrtů) o průměru 2 mm a délce 10 mm (Rossi et al. 2006). Před samotným odběrem bylo potřeba u dřevin s hrubší borkou dlátem odstranit kůru, přičemž nesmělo dojít k poranění kambia. Řezací část trephoru byla zaražena do dřeva pomocí gumového kladiva. Trephor musel být orientován kolmo na podélnou osu kmene, aby nedošlo ke zkreslení daného přírůstu. Následně byl nástroj kroutivým pohybem vyjmut z kmene společně s mikrovývrtem. Za pomocí speciálně přizpůsobené jehly byl mikrovývrt přesunut do střední části trephoru, odkud byl přemístěn do plastových kazetek uzpůsobených pro manipulaci se vzorkem. Plastové kazetky byly popsány datem odběru a daným označením vzorníku. Obr. 6. Trephor nástroj pro odběr mikrovývrtů z kmene. 28
29 Mikrovývrty v kazetkách byly ihned po odběru vloženy do fixačního roztoku FAA (100 ml roztoku FAA obsahuje: 90 ml 50 70% etanolu, 5 ml formaldehydu, 5 ml ledové kyseliny octové), aby došlo k usmrcení živých buněk (Dušková, 1995). Díky tomu byla zachycena tvorba nového dřeva (letokruhu) přesně k datu odběru. V laboratoři po týdenní fixaci byly kazetky se vzorky vyjmuty z roztoku, opláchnuty a vloženy do 30% etanolu, ve kterém zůstaly do následného zpracování. 4.3 Úprava vzorků před krájením Odebrané mikrovývrty z terénu bylo potřeba před krájením na mikrotomu zkrátit pomocí žiletky na potřebnou velikost. Před manipulací se vzorky byly popsány dle jednotlivých dat odběru plastové, uzavíratelné zkumavky, do kterých byl vložen jeden ze dvou vzorků, jako náhrada při případném poškození vzorového mikrovývrtu. Plastové kazetky s mikrovývrty byly vyjmuty z etanolu. Makroskopicky kvalitnější vzorek byl pinzetou přemístěn na sací papír, který byl rozprostřen na dřevěném prkýnku. Druhý vzorek byl přesunut do již připravené plastové zkumavky, kde byl za pomocí pipety zalit 30% etanolem. Vzorový mikrovývrt musel být chycen pinzetou za dřevní část (xylém), aby nedošlo k poškození kambiální zóny. Šetrnou manipulací byla odlišena tmavší transverzální část, která byla označena tužkou. Označení transverzální části mikrovývrtu je důležité pro rozlišení správné orientace vzorku při zalévání do parafinu, aby při následném krájení na mikrotomu mohl být uskutečněn příčný řez. Takto označený mikrovývrt byl na straně xylému zkrácen mezi 2 3 letokruhem od kambia. Strana floému byla zkrácena jen o nepatrnou část (cca 2 mm). Zredukovaný mikrovývrt byl vrácen do plastové kazetky a přemístěn do nádoby s 30% etanolem. Aby došlo při zalévání k maximálnímu nasycení vzorků parafinem a k jejich spojení s okolní vrstvou vosku, bylo nutné provést dehydrataci vzorků pomocí etanolové řady. Jedná se o řadu složenou celkem ze 7 nádob se vzestupnou koncentrací etanolu. První dvě nádoby obsahují 70% etanol, další dvě 90% etanol, dále pak následuje nádoba s 95% etanolem a nádoba obsahující 100% etanol. V nádobě se 70% etanolem byly vzorky vloženy 120 min., v ostatních nádobách 90 min.. Po dehydrataci byly vzorky umístěny do Petriho misek s parafinem a následně přesunuty do sušárny, která byla nastavena na teplotu 65 C. V sušárně, kde se ponechaly přibližně 4 hodiny, došlo k maximálnímu nasycení vzorků parafinem. Tím pádem bylo docíleno vytlačení 29
30 přebytečné vody a zpevnění pro krájení na rotačním mikrotomu. Po uplynutí doby potřebné k nasycení mikrovývrtů parafinem následovalo vlastní zalití vzorků. K tomuto účelu bylo použito zařízení Leica EG Jedná se o přístroj pro tavení parafinu s vyhřívanou pracovní plochou, na kterém lze libovolně nastavit teplotu pro tavení (v našem případě se jednalo o 65 C). Jako příslušenství pro tavící přístroj byla použita elektricky vyhřívaná pinzeta Leica EG F. K správnému zalití byla potřeba Petriho miska s chladnou vodou a ocelové formičky sloužící k umístnění vzorků. Vlastní zalití vzorků se skládalo z několika částí. Nejdříve bylo nutné na vyhřívané pracovní ploše nahřát ocelové formičky a zbavit je přebytečného zaschlého parafinu. Po nahřátí vaniček byla ze sušárny vyjmuta Petriho miska s mikrovývrtem v parafinu. Ta se položila na zahřátou pracovní plochu. Do ocelové formičky se na dno napustil parafin (vrstva cca 2 mm). Vyhřívanou pinzetou se šetrnou manipulací rozlišila již označená transverzální část mikrovývrtu, vzorek se pinzetou uchopil za xylémovou část a byl přemístěn do připravené ocelové formičky. Musel být umístěn úhlopříčně do středu formičky tak, aby označená část transverzálního řezu směrovala nahoru. Úhlopříčné uložení bylo nutné proto, aby při krájení na mikrotonu vznikal postupný odpor mikrovývrtu na čepelku. Následně byla ocelová formička s mikrovývrtem za neustálého přidržování vzorku vyhřívanou pinzetou posazena do Petriho misky se studenou vodou. V okamžiku, kdy parafin nepatrně ztuhnul, se zafixoval vzorek do požadované polohy. V tom mohlo dojít k odložení vyhřívané pinzety a k dolití formičky cca do 3/4 parafinem. Dále pak byla do formičky umístěna část plastové kazetky a formička byla dolita parafinem až po okraj. Takto zalitý preparát byl přemístěn na 15 min. do lednice k úplnému ztuhnutí. Po ztuhnutí parafinu byla šetrně odstraněna ocelová formička. Dále před krájením na mikrotomu bylo potřeba zalitý vzorek upravit na požadovanou velikost. Nožem s odlamovací čepelí značky Stanley byl parafin odstraněn po obvodu plastové kazetky a potom byl parafin odřezán kolem mikrovývrtu na plošku cca 1 1 cm. Následně byl takto upravený vzorek (Obr. 7) zatrimován na mikrotomu a vložen do kádinky s čistou vodou z důvodu změkčení vzorku a pro usnadnění následného krájení. 30


31 Obr. 7. Kazetka se zalitým mikrovývrtem v parafinu. 4.4 Krájení vzorků na mikrotomu Jelikož bylo cílem vytvořit kvalitní trvalé preparáty o tloušťce řezu 12 μm, byl pro tento účel použit rotační mikroton Leica RM 2235 (Vavrčík a Gryc 2004). Obr. 8. Zleva: vodní lázeň Leica HI 1210, rotační mikrotom Leica RM Plastová kazetka se vzorkem zalitým v parafinu byla upnuta do adaptéru rotačního mikrotomu tak, aby čepelka, která krájí výsledný pás preparátu, zařezávala nejdříve do dřeva. Rychlým otáčením klikou mikrotomu byl získán souvislý pás řezů o dané 31
32 tloušťce. Čepelka se musela po každém vzorku posunout, aby nedošlo vlivem ztupení k případnému znehodnocení preparátu. Za pomocí pinzety a preparační jehly byl pás řezů přemístěn do vyhřívané vodní lázně Leica HI 1210 (Obr. 8), ve které již byla zahřátá voda na 40 C. Díky teplé vodě došlo k rozpínání parafinu k narovnání jednotlivých řezů. Reprezentativní řezy (4) byly následně přemístěny za pomocí štětce na podložní sklíčko. Na podložní sklíčko byl již nanesen Albumine Mallory (směs vajíčkového bílku s glycerínem v poměru 1:1). Ten zajistil přilnutí daných řezů k podložnímu sklíčku. Tímto postupem vytvořené preparáty byly na několik minut (cca 10 min.) umístěny do sušárny, kde byly při teplotě 70 C zbaveny zbylé vody. 4.5 Podvojné barvení Po zbavení preparátů přebytečné vody následovalo podvojné barvení a tvorba trvalých preparátů. Podložní sklíčka byla umístěna po dvanácti do kontejnerů, pomocí kterých byly přemísťovány v přesných časových intervalech (Tab. 2) do jednotlivých nádob s chemikáliemi. V prvních dvou nádobách se nacházel Bio clear, pomocí kterého došlo k odstranění pozůstalého parafinu. Následovaly dvě nádoby se 100% etanolem. Potom byly kontejnery přemístěny do nádoby, která obsahovala safranin. Roztok safraninu barvil lignifikované (mrtvé) buňky do červena. Po zbarvení byly kontejnery ze safraninu přemístěny do 100% etanolu, ve kterém byly pouze propláchnuty. Po etanolu následovala nádoba s Astra blue. Roztok Astra blue barvil buňky, které byly v době odběru živé do modra. Dále pak byly kontejnery přemístěny do dvou nádob se 100% etanolem a následně do nádoby se xylenem, ve kterých byly opět pouze propláchnuty. Po vyjmutí preparátů z kontejnerů bylo opticky zkontrolováno zbarvení vzorků. Vzorky byly po šetrném osušení zakápnuty uzavíracím médiem. Jako uzavírací médium byl použit kanadský balzám jedná se o přírodní pryskyřici, která se získává z jedle (Abies balsamea (L.) Mill.) (Ives, 2001). Dále byly vzorky za pomocí preparační jehly zakryty krycím sklíčkem. Takto vyrobené preparáty byly ve finální fázi přemístěny na ocelový plát. Na krycí sklíčko se vložil magnet, který z okolí vzorku vytěsnil přebytečný vzduch a vytlačil přebytečný kanadský balzám. Tímto způsobem vytvořené trvalé preparáty (Obr. 9) byly připraveny k vyhodnocování pod mikroskopem. 32
 1) Bio-clear 30 6) Etanol 2) Bio-clear 30 7) Astra blue 3) Etanol 15 8) Etanol proplach 4) Etanol 15 9) Etanol proplach 5) Safranin 5 10) Xylen proplach 4.")
33 Obr. 9. Trvalý preparát. Tab. 2. Použité časové intervaly pro podvojné barvení. Roztok Časový interval (min.) Roztok Časový interval (min.) 1) Bio-clear 30 6) Etanol 2) Bio-clear 30 7) Astra blue 3) Etanol 15 8) Etanol proplach 4) Etanol 15 9) Etanol proplach 5) Safranin 5 10) Xylen proplach 4.6 proplach 3 Vyhodnocení preparátů pod mikroskopem Vyhodnocení trvalých preparátů bylo provedeno pod světelným mikroskopem Leica DMLS. Díky digitální kameře Leica DFC 280 byl uskutečněn přenos obrazu přímo do počítače, kde se probíhalo snímání vzorků. Snímání a následné vyhodnocování snímků v PC bylo uskutečněno v programu ImageJ. Pomocí daného programu byly snímky z digitální kamery ukládány a bylo možno na nich provádět jednotlivá měření. Byl zjišťován průměrný počet buněk v kambiální zóně a celkový radiální přírůst xylému v průběhu vegetačního období. Dále se rozlišovaly první cévy ve fázi radiálního rozšiřování a první plně vytvořené cévy z hlediska jejich rozměru a tvaru. Na základě zbarvení buněk byl rozlišen začátek a konce lignifikace. 33
34 Naměřené hodnoty byly zpracovány pomocí programu OpenOffice.org 3.3, ve kterém byly zpracovány i výsledné grafy. Pomocí Gompetrzovy funkce (Obr. 10) zpracované v programu SigmaPlot ; Table Curve 2D v5.01 byly naměřené hodnoty proloženy vyrovnávací křivkou, která nám vytvořila optimální model hodnot pro daný vzorník. Obr. 10. Gompertzova funkce. Kde: y = rostoucí suma buněk po týdnech t = čas ve dnech A = horní asymptota maximálního počtu buněk β = x osová osa umístění parametru k = míra změny parametru 34
35 5 Výsledky 5.1 Vyhodnocení výsledků vzorníku B1 Počet a tvar buněk kambiální zóny se v průběhu celého roku značně liší (Obr. 13). V latentním kambiu se nacházelo 5 6 buněk s buněčnou stěnou obdelníkovitého tvaru a s malým radiálním rozměrem. Počátek kambiální aktivity, kdy docházelo ke zvětšování radiálního rozměru kambiálních buněk a k zvyšování jejich počtu byl stanoven k při průměrné teplotě vzduchu předcházejícího týdne 7,19 C. Tehdy vzrostl počet buněk kambiální zóny v průměru na 7. V průběhu vegetačního období se počet kambiálních buněk postupně zvyšoval, přičemž nejvyšší kambiální aktivita byla sledována na přelomu května a června ( ), kdy průměrný počet buněk v kambiální zóně dosáhl hodnoty 11. Deaktivace kambia, kdy docházelo k postupnému ustálení počtu kambiálních buněk byl sledován od 12. do při průměrné teplotě vzduchu 18,8 C za předcházející týden. Kambium tehdy obsahovala v průměru 7 buněk. Počet buněk kambia vzrostl od doby jeho aktivace, do doby jeho maximálního rozměru o 4 buňky. V období od do se postupně snížil počet kambiálních buněk a kambium obsahovalo až po datum posledního odběru vzorků průměrně 4 5 buněk, jejichž morfologie byla podobná buňkám latentního kambia. Dne , kdy dosahovalo kambium svého maximálního rozměru, byl naměřen první radiální přírůst xylému. Stejného dne byly rozpoznány první cévy ve fázi radiálního rozšiřování. K datu byly rozlišeny první plně vytvořené cévy z hlediska jejich finálního rozměru a tvaru. Začátek lignifikace byl stanoven k , kdy docházelo k první lignifikaci buněčných stěn libriformních vláken. Konec lignifikace byl dle červeného zbarvení veškerých anatomických elementů dřeva rozpoznán koncem srpna (26. 8.). Šířka plně vytvořeného letokruhu byla 1386 μm a čas nezbytný pro jeho formaci byl 98 dní. 5.2 Vyhodnocení výsledků vzorníku B2 Na začátku vegetačního období byla pozorována změna tvaru kambiálních buněk oproti buňkám latentního kambia. Kambium začalo zvětšovat svůj radiální rozměr a docházelo k tzv. bobtnání kambia. Latentní kambium se skládalo průměrně 35
36 z 5 kambiálních buněk se štíhlou buněčnou stěnou a malým radiálním rozměrem. Reaktivace kambia s 6 buňkami v kambiální zóně byla stanovena ke konci dubna (29. 4.) při průměrné teplotě vzduchu předchozího týdne 11,01 C. Kambium dosáhlo svého maxima v období od do s průměrně 12 buňkami v kambiální zóně. Deaktivace kambia, kdy docházelo k zeštíhlování buněčných stěn a k snižování počtu buněk kambiální zóny byla stanovena v období od do s průměrnými 7 buňkami v kambiální zóně. Průměrná teplota předchozího týdne byla tehdy 16,65 C. Počet buněk kambia se zvýšil od doby jeho aktivace po dobu jeho maximálního rozměru o 6 buněk. V období od do se počet buněk v kambiu ustálil a kambium až po datum posledního odběru obsahovalo v průměru 5 6 buněk s malým radiálním rozměrem a poměrně tenkou buněčnou stěnou. První zaznamenaný radiální přírůst xylému se váže k , což bylo tři týdny před datem, kdy kambium začalo dosahovat svých maximálních rozměrů. Ke stejnému datu byly pozorovány první cévy ve fázi radiálního rozšiřování. První plně vytvořené cévy z hlediska jejich rozměru a tvaru byly rozlišeny začátkem června (3. 6.). Lignifikace buněčných stěn začal , kdy docházelo (dle červeného zbarvení safraninem) k prvnímu dřevnatění libriformních vláken. Dne 2. 9., kdy došlo k celistvému červenému zbarvení nově vytvořeného letokruhu byl stanoven konec lignifikace anatomických elementů dřeva. Šířka plně vytvořeného letokruhu činila 1976 μm a čas potřebný pro jeho vytvoření byl 105 dní. 5.3 Vyhodnocení výsledků vzorníku B3 Latentní kambium s typickou rektangulární buněčnou stěnou obsahovalo 5 6 buněk. Dříve než došlo k dělení buněk kambiální zóny, bylo možné pozorovat, jak kambiální buňky začínají zvětšovat svůj radiální rozměr a dochází k tloustnutí jejich buněčné stěny (tzv. k bobtnání kambia). Začátek kambiální aktivity byl stanoven k při průměrné teplotě vzduchu předcházejícího týdne 7,19 C, kdy vzrostl počet buněk kambia na 7. Během vegetačního období počet buněk v kambiální zóně postupně narůstal. Nejvyšší aktivita kambia byla pozorována ve dnech od do s průměrně 11 buňkami v kambiální zóně. Rozdíl mezi aktivací kambia a dobou jejího maxima jsou 4 kambiální buňky. Činnost kambia byla ukončena ve dnech od do , kdy začal postupně klesat počet buněk v kambiální zóně (kambium obsahovalo 6 7 buněk). Teplota 36
37 vzduchu před ukončením činnosti kambia se v průběhu předcházajícího týdne pohybovala okolo 18,8 C. Od začátku září (9. 9.) do data posledního odběru se počet buněk kambia ustálil s průměrnými 5 buňkami v kambiální zóně, jejichž tvar a velikost se podobal buňkám klidového kambia. První měřitelný radiální přírůst xylému byl zaznamenán dne , což odpovídá době, ve které dosahovalo kambium svého maximálního rozměru. Ve stejný den se objevily první cévy ve fázi radiálního rozšiřování. První dvě vytvořené cévy (z hlediska rozměru a tvaru) byly rozlišeny začátkem června (3. 6.). Počátek lignifikace dle prvního červeného zbarvení buněčných stěn libriformních vláken byl stanoven k , zcela lignifikované buněčné stěny veškerých anatomických elementů byly determinovány začátkem září (2. 9.). Radiální přírůst plně vytvořeného letokruhu činil 2184 μm a čas potřebný pro jeho vylišení se rovnal 112 dní. 5.4 Srovnání výsledků všech vzorníků Kambiální aktivita (zvyšování počtu kambiálních buněk) u vzorníků B1 a B3 začala dne při průměrné teplotě vzduchu předcházejícího týdne 7,19 C, u vzorníku B2 kambium zahájilo svou činnost o týden později, tedy při teplotě 11,01 C. Průměrná týdenní teplota při které kambium zahájilo svou dělivou činnost je 9,1 C. Buňky při reaktivaci kambia byly charakteristické svým větším radiálním rozměrem a svým tzv. nabobtnalým tvarem oproti buňkám latentního kambia. Počet buněk kambia se v době aktivace zvýšil na 6 7. Latentní kambium v průměru obsahovalo s typickou rektangulární buněčnou stěnou a malý radiálním rozměrem 5 6 buněk. Nejvyšší kambiální aktivita byla pozorována v období od do s průměrně 12 kambiálními buňkami. Počet buněk vzrostl od doby aktivace kambia po dobu jeho maximálního rozměru v průměru o 5 buněk. Konec činnosti kambia, kdy docházelo k snižování počtu buněk v kambiální zóně byl stanoven v rozmezí od 12. do s průměrně 6 buňkami kambia. Teplota vzduchu se za předcházející týden pohybovala od 16,65 do 18,8 C, přičemž průměrná teplota dosáhla hodnoty 17,72 C. Průběh tvorby buněk kambiální zóny v během vegetačního období 2010 je patrný z následného grafu (Obr. 11). 37
38 16 14 Počet buněk B1 B2 B3 Ø Den v roce Obr. 11. Počet buněk kambiální zóny v průběhu vegetačního období 2010 (den 80 = , den 320 = ). První měřitelný radiální přírůst xylému byl u všech vzorníků zaznamenán v květnu od 13. do , přičemž první cévy ve fázi radiálního rozšiřování byly popsány ve stejném časovém intervalu. Plně vytvořené cévy z hlediska jejich finálního rozměru a tvaru byly determinovány u všech vzorníků začátkem června (3. 6.). Začátek lignifikace byl stanoven v červnu od 17. do , konec lignifikace buněčných stěn byl pozorován od do 2. 9., tedy na přelomu srpna a září. Šířka plně vytvořeného letokruhu se pohybovala od 1386 do 2184 μm s průměrnou hodnotou 1848 μm a průměrný čas nezbytný pro jeho vytvoření byl 105 dní. Následující graf (Obr. 12) vystihuje radiální přírůst xylému v průběhu vegetačního období
![Radiální přírust dřeva[μm] 3000 2500 2000 B1 B2 B3 Ø 1500 1000](/docs-images/91/106004062/images/39-0.jpg "500 0 80 100 120 140 160 180 200 220 240 260 280 Den v roce")

39 Radiální přírust dřeva[μm] B1 B2 B3 Ø Den v roce Obr. 12. Analýza radiálního přírůstu xylému v průběhu vegetačního období 2010 (den 80 = , den 280 = ). 39
40 40
 13. 5. = CC s 9 buňkami, zóna rozšiřujících se buněk (PC), céva ve fázi radiálního rozšiřování (V); c) 10. 6. = kambium s 7 8")
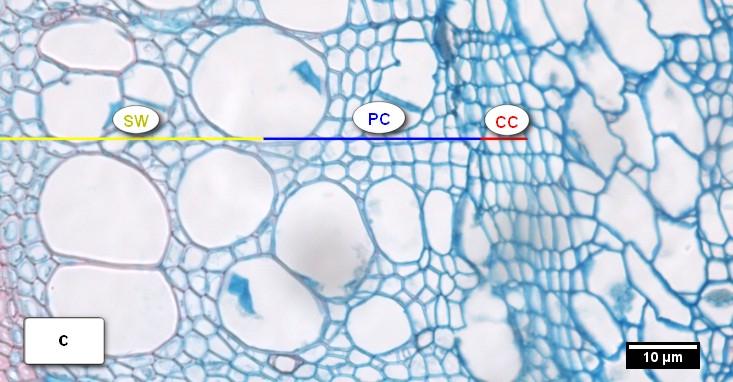
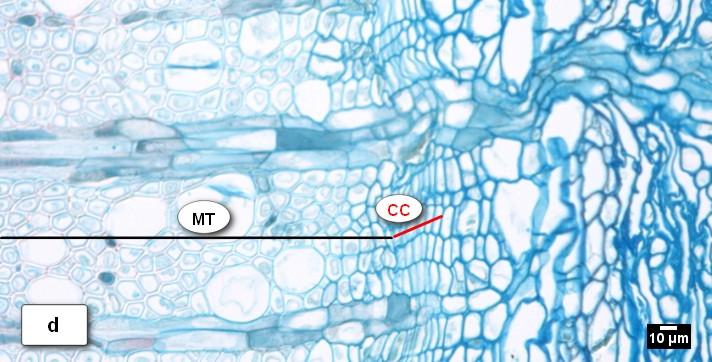
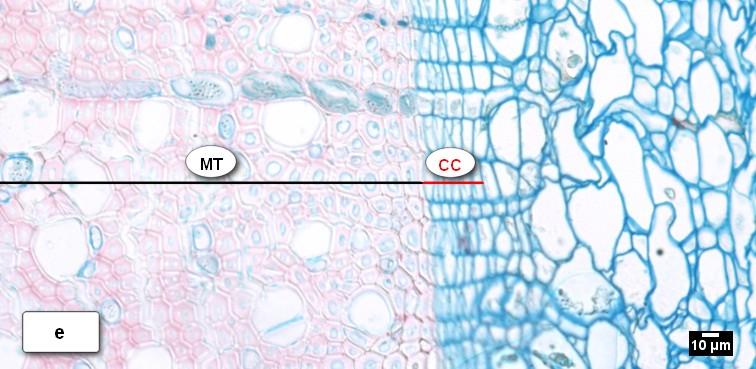
41 Obr. 13. Vzhled kambiální zóny (CC) a průběh radiálního přírůstu xylému u buku lesního (Fagus sylvatica L.) během vegetačního období 2010 k: a) = latentní kambium s 4 5 buňkami; b) = CC s 9 buňkami, zóna rozšiřujících se buněk (PC), céva ve fázi radiálního rozšiřování (V); c) = kambium s 7 8 buňkami, zóna dozrávání buněk (SW); d) = kambiální zóna složena z 8 9 buněk, zóna zralých buněk (MT); e) = kambium přecházející do latentní fáze s 5 6 buňkami; f) = latentní kambium s 4 buňkami. 41
")
 před aplikací polarizačního světla, kde je znázorněna kambiální zóna (CC), zóna rozšiřujících se buněk (PC), zóna dozrávání")
42 5.4.1 Zachycení tvorby sekundární buněčné stěny pomocí polarizačního světla Pomocí polarizace na světelném mikroskopu byla zachycena tvorba sekundární buněčné stěny S1, 2, 3, která díky inkrustaci krystalickou celulózou pod polarizačním světlem jasně reflektovala (Obr. 14). Obr. 14. Rozlišení tvorby sekundární buněčné stěny k pomocí polarizace; a) před aplikací polarizačního světla, kde je znázorněna kambiální zóna (CC), zóna rozšiřujících se buněk (PC), zóna dozrávání buněk (SW), buňka s primární buněčnou stěnou (P) a sekundární buněčnou stěnou (S); b) po aplikaci polarizačního světla. 42
43 5.5 Aplikace Gompertzovy funkce Pomocí Gompertzovy funkce byly naměřené hodnoty radiálního přírůstu xylému proloženy vyrovnávací křivkou, která nám vytvořila optimální model radiálního přírůstu xylému v průběhu vegetačního období 2010 na výzkumné ploše Rájec Domanka. Dle modelu Gompertzovy funkce bylo potřeba v průměru 60 dnů pro vytvoření většiny buněk xylému, přičemž hodnoty u všech vzorníků se pohybovaly v rozpětí od 46 do 75 dnů. Maximální vypočtený přírůst xylému dosahoval v průměru 1887 μm (od 1226 do 2467 μm). Modelový denní přírůst xylému v období aktivního růstu vzorníků se rovnal v průměru 41,3 μm, jednotlivé denní přírůsty vzorníků (Obr.18) se pohybovaly v rozmezí od 40,3 do 43,83 μm. Kambium zahájilo svou činnost dle Gompertzovy funkce koncem dubna (od 22. do ), aktivita kambia byla ukončena v průběhu září (od do ). Kambium bylo dle Gompertze aktivní v průměru 147 dní. Výsledné parametry jsou shrnuty v následující tabulce (Tab.3) Denní přírůst [μm] Radiální přírůst [μm] 1400 T D G Počet dní Obr. 15. Aplikace Gompertzovy funkce u vzorníku B1, kde: T znamená radiální přírůst, D denní přírůst a G Gompertzovu vyrovnávací křivku (den 80 = , den 320 = ). 43
44 Radiální přírůst [μm] Denní přírůst [μm] T D G Počet dní Denní přírůst [μm] Radiální přírůst [μm] Obr. 16. Aplikace Gompertzovy funkce u vzorníku B2, kde: T znamená radiální přírůst, D denní přírůst a G Gompertzovu vyrovnávací křivku (den 80 = , den 320 = ). T D G Počet dní Obr. 17. Aplikace Gompertzovy funkce u vzorníku B3, kde: T znamená radiální přírůst, D denní přírůst a G Gompertzovu vyrovnávací křivku (den 80 = , den 320 = ). 44
45 70 Denní přírůst [μm] B1 B2 B Den v roce Obr. 18. Srovnání denního přírůstu xylému u všech vzorníků (den 80 = , den 320 = ). Tab. 3. Výsledné parametry xylému vypočteny dle Gompertzovy funkce v průběhu vegetačního období Vzorník Maximální radiální přírůst xylému (μm) Průměrný denní přírůstu xylému (μm) Délka tvorby většiny buněk xylému (den) B ,31 75 B ,83 46 B ,03 60 Ø ,
46 6 Diskuse Tato studie se zabývá analýzou radiálního přírůstu xylému a charakteristikou činností kambia u buku lesního (Fagus sylvatica L.) v průběhu vegetačního období 2010 na Drahanské vrchovině, výzkumná plocha Rájec Domanka (Obr. 19). Dále bylo předmětem práce analyzovat začátek a konec lignifikace anatomických elementů dřeva a určit spojitost jednotlivých fází xylogeneze s klimatickými podmínkami dané lokality. Morfologie a počet buněk v kambiální zóně se během vegetačního období značně liší (Obr. 13). Buňky latentního kambia s typickou rektangulární buněčnou stěnou bylo možné vzhledem k jejich zploštělému tvaru jasně rozlišit již od nabobtnalých buněk aktivního kambia. V našem případě se latentní kambium skládalo z 5 6 buněk, v čem se shodujme s výsledky Oladi et al. (2010), jež provedli výzkum na buku východním (Fagus orientalis Lispsky) ve smíšeném porostu v nadmořských výškách od 650 do 1600 m n. m.. Gričar (2007), která provedla studii kambiální aktivity na jedli a smrku, zaznamenal v průměru 7 9 buněk latentního kambia. Daný rozdíl si lze vysvětlit odlišnou (jednoduchou) anatomickou stavbou dřeva jehličnatých dřevin. Určit přesný začátek kambiální aktivity, tedy proces, kdy dochází k zeštíhlování buněčných stěn kambia, k radiálnímu rozšiřování kambiálních iniciál a k zvyšování jejich počtu, je vzhledem k týdenním intervalům odběru vzorků značně obtížné. V období koncem dubna ( ) při průměrné předcházející týdenní teplotě vzduchu 9,1 C bylo zaznamenáno zvyšování počtu kambiálních buněk. Počet buněk se při reaktivaci kambia zvedl v průměru na 7. Lze konstatovat, že reaktivace kambia začala současně v rozmezí jednoho týdne koncem dubna s příznivými klimatickými podmínkami, především s optimální teplotu vzduchu (průměrná teplota vzduchu se rovná v dubnu 7,62 C a v květnu 10,23 C). Čufar a Prislan (2008) zaznamenali počátek kambiální aktivity v lesích Panška Reka (400 m n. m.) již v druhém dubnovém týdnu a počet buněk v kambiu po jeho reaktivaci stanovili na 10. Jelikož neuvedli průměrné denní teploty a stanovištní poměry vztahující se k zjištěným údajům, lze předpokládat, že tento rozdíl byl v našem případě způsoben odlišnými geografickými podmínkami daných lokalit. Horáček et al. (2003) na dubu letním (Quercus robur L.) stanovili počátek kambiální 46
47 aktivity na období před při průměrné teplotě vyšší jak 8 C. Vzhledem k tomu, že se jedná o dřevinu s kruhovitě pórovitou stavbou dřeva z lužních lesů v nadmořské výšce 165 m n. m., lze se domnívat, že primárně teplota vzduchu a nadmořská výška jsou hlavními faktory ovlivňující reaktivaci kambia na začátku vegetačního období (Begum et al. 2007). Kambium v průběhu vegetačního období svůj počet buněk téměř zdvojnásobilo, tj. v průměru dosahovalo kolem 8 10 buněk. Ke stejnému závěru dospěli i Oladi et al. (2010) u vzorníků buku východního (Fagus orientalis Lipsky). Eschrich (1995) sledoval nejvyšší kambiální aktivitu (počet buněk v kambiální zóně) buku lesního (Fagus sylvatica L.) v období na konci května a první polovině června. To se shoduje s našimi výsledky u vzorníků B1 a B3, přičemž u vzorníku B2 kambium dosahovalo vyšších hodnot až do první poloviny července. Konec kambiální aktivity se v našem případě vztahuje na datum, kdy kambium přestalo zvyšovat svůj počet buněk a jejich morfologii se podobala latentním buňkám kambia. Deaktivace kambia byla stanovena v rozmezí při průměrné teplotě vzduchu 17,72 C za předcházejí týden. Vzhledem k nadprůměrným teplotám vzduchu na naší výzkumné ploše došlo v rozmezí tří měsíců (červen srpen) k rapidnímu úbytku půdní vlhkosti, jež mohl mít za následek deaktivaci činnosti kambia. Horáček et al. (2003) u dubu vázali konec činnosti kambia k datu, kdy nezaznamenali žádný radiální přírůst xylému, tedy na přelomu září a října. Naproti tomu Čufar a Prislan (2008) uvedli konec kambiální činnosti o dva týdny dříve, než tomu bylo v našem případě. Lze usoudit, že vzhledem k nižší nadmořské výšce rostou průměrné denní teploty a tím pádem začíná dříve vegetační období. Délka trvání činnosti kambia byla v průměru u našich vzorníků 16 týdnů. Čufar a Prislan (2008) uvedli délku trvání činnosti kambia o 3 týdny delší. Naproti tomu Marion et al. (2007) zjistili u vzorníků z městské oblasti Ljubljana, že doba činnosti kambia javoru mléče (Acer platanoides L.) trvala u zdravých stromů 20 týdnů a u poškozených 13 týdnů. Zjištěné údaje nevztahují ke klimatickým podmínkám, uvádí pouze nadmořskou výšku (323 m n. m.). Na základě daných údajů lze usoudit, že rozdíl výsledných hodnot je způsobený vyšší průměrnou teplotou vzduchu městské oblasti v průběhu vegetačního období. Vzhledem k předpokládané vyšší průměrné teplotě vzduchu v průběhu vegetačního období v městské oblasti Ljubljana mohl mít především rozhodující vliv na ukončení činnosti kambia úbytek půdní vlhkosti. Kambium u námi zkoumaných vzorníků po deaktivaci 47
48 obsahovalo 5 6 buněk. V tom se shodujeme s výsledky Čufar et al. (2011), jež provedli stejný výzkum na kaštanovníku setém. Při porovnání kambiální aktivity s předcházejícím výzkumy na dané téma jsme došli k závěru, že začátek a konec kambiální aktivity je plně závislý na geografických a tím pádem na klimatických podmínkách (především na teplotě vzduchu a půdní vlhkosti). Jak již bylo výše uvedeno, v období od začátku června do konce srpna byl na výzkumné lokalitě Rájec Domanka zaznamenán značný úbytek půdní vlhkosti. V tomto období můžeme pozorovat pokles počtu kambiálních buněk u vzorníku B1. Konec kambiální aktivity byl stanoven cca v polovině srpna, jež lze opět spojit s daným přísuškem. Pro upřesnění závislosti úbytku půdní vlhkosti a průběhu činnosti kambia by bylo zapotřebí kratší intervaly v odběru vzorků z většího počtu vzorníků, abychom byli schopni určit přesnou korelaci mezi oběma hodnotami. První radiální přírůst xylému a první cévy ve fázi radiálního rozšiřování byly zaznamenány v průběhu května ( ). V tom se shodujeme s výsledky Čufar a Prislan (2008). Naproti tomu Marion et al. (2007) uvedli u zdravých javorů první přírůst o dva týdny dříve. První plně vytvořené cévy (mající konečný rozměr a vytvořenou sekundární buněčnou stěnu) byly pozorovány již začátkem června, tj. 5 týdnů po jejich objevení se ve fázi radiálního rozšiřování. Zjištěné výsledky se shodují s dříve publikovanými závěry Čufar a Prislan (2008) pro buk lesní z lokality Panška Reka. Lignifikace buněčných stěn začala již v průměru 10 dní po úplné diferenciaci prvních cév, což odpovídá výsledkům Marion et al. (2007) u zdravých javorů a Oladi et al. (2010) u buku východního. Lignifikace anatomických elementů pokračovala i po ukončení aktivní činnosti kambia (Prislan et al. 2009, Oladi et al. 2010), v průměru dnů, přičemž konec lignifikace (dle celkového zbarvení xylémové části do červena) byl stanoven na přelomu srpna a září. V tom se mírně lišíme od výsledků Oladi et al. (2010) jež uvedli, že lignifikace byla ukončena ve stejné nadmořské výšce u buku východního na přelomu září a října. V den úplné lignifikace buněk nově vytvořeného letokruhu byl stanoven celkový radiální přírůst xylému, který dle mikroskopického měření dosahoval hodnot od μm, v průměru 1848,7 μm. Radiální přírůst xylému je charakterizován typickou S křivkou. Modifikace růstových křivek je ovlivňována klimatickými činiteli, zejména vzájemnými poměry mezi výší teplot a srážek (Matovič, 1975). 48
49 Na základě aplikace Gompertzovy funkce byl vytvořen optimální model hodnot radiálního přírůstu xylému v průběhu vegetačního období Oscilace naměřených hodnot, které nám křivka vyrovnává, je způsobena spirálovitým odběrem vzorků po obvodu kmene. V případě buku se také velmi často vyskytuje reakční dřevo tahové. Tahové dřevo je charakteristické vyšší tvorbou buněk. U některých mikroskopických řezů bylo identifikováno tahové dřevo pomocí přítomnosti tzv. tahových (želatinových) vláken, která obsahovala tzv. G vrstvu. Maximální radiální přírůst dle Gompertzovy funkce se pohyboval v rozmezí μm, s průměrnou hodnotou 1887 μm. Výsledná průměrná hodnota maximálního radiálního přírůstu 1877 μm je větší o 38,3 μm od hodnoty zjištěné mikroskopickým měřením, což činní odchylku cca 2 %. Lze konstatovat, že výsledná křivka Gompertzovy funkce dobře popisuje radiální přírůst xylému v průběhu vegetačního období a vytváří nám optimální model hodnot pro daný druh dřeviny. Doba nezbytná pro vytvoření většiny buněk xylému dle Gompertze byla v průměru 60 dnů, přičemž hodnoty se pohybovaly od dnů. Čufar a Prislan (2008) uvedli dle Gompertzovy funkce průměrný maximální přírůst xylému 2552 μm a průměrnou dobu nezbytnou pro vytvoření většiny buněk xylému 100 dní. Je patrné, že naše bukové vzorníky prokazovaly menší maximální radiální přírůst a tomu odpovídající kratší dobu nezbytnou pro vytvoření většiny buněk xylému. Tento rozdíl mohl být způsoben odlišnými geografickými, klimatickými či stanovištními podmínkami. Sass a Eckstein (1995) vyslovili hypotézu, která říká, že tvrdá dřeva vytvoří vodivý systém již na začátku vegetačního období z důvodů pravděpodobných nepříznivých podmínek během zbylé doby aktivního růstu. To je patrné z vyrovnávací křivky Gompertzovy funkce u našich vzorníků. V průměru 70 % letokruhu je vytvořeno již v první polovině července, což může být např. k přísušku, který se vyskytoval na naší výzkumné ploše značně prospěšné pro celkovou formaci xylému. U vzorníku B1 byl stanoven značně menší denní a tím pádem i celkový radiální přírůst xylému. I když se jednalo o vzorníky přibližně stejných dimenzí, nelze z toho vyvodit závěr, že všechny stromy stejných dendrometrických hodnot přirůstají v průběhu vegetačního období přibližně stejným tempem. 49
50 7 Závěr Na Drahanské vrchovině, výzkumné ploše Rájec Domanka, byl uskutečněn výzkum týkající se kambiální aktivity a diferenciace xylémové části buku lesního (Fagus sylvatica L.) v průběhu vegetačního období Jedná se o jednu z prvních prací v České republice, zabývající se u buku tvorbou xylému na buněčné úrovni. Kambiální aktivita u všech vzorníků buku začala v rozmezí od 22. do při průměrné teplotě vzduchu předcházejícího týden 9,1 C, přičemž kambiální zóna se při reaktivaci skládala z 6 7 buněk. Počet buněk kambia vzrostl od doby jeho reaktivace po dobu jeho maximálního rozměru v průměru o 5 buněk. Činnost kambia u všech vzorníků byla ukončena v období od 12. do při průměrné teplotě předcházajícího týdne 17,72 C. Kambium se skládalo při své deaktivaci v průměru z 6 buněk, což odpovídalo počtu buněk latentního kambia. První cévy ve fázi radiálního rozšiřovaní se objevily od 13. do a shodně 3.6. dosáhly svého plného rozměru. Lignifikace buněčných stěn pokračovala ještě v průměru dnů po ukončení kambiální činnosti. Průměrný vypočtený denní přírůst činil 33,40 μm. Dle Gompertzovy funkce se maximální radiální přirůst rovnal 1887 μm, přičemž se lišil od hodnoty zjištěné mikroskopickým měřením pouze o 2 %. Daný výzkum se týká pouze jednoho vegetačního období. Pro upřesnění výsledných hodnot tvorby dřeva na buněčné úrovni by bylo zapotřebí pokračovat s výzkumem i v následujících letech. Na základě průměrných hodnot zjištěných v průběhu několika vegetačních období by bylo vhodné vytvořit modely, které popíší spojitost mezi radiálním přírůstem dřeva na buněčné úrovni ve vztahu ke klimatickým (teplota vzduchu, půdní vlhkost) a geografickým podmínkám. 50
51 8 Summary The research of common beech during vegetation period 2010 took place on the Drahanhighland, Rájec Domanka. site. The goal of the work was to analyse radial growing of xylem, characteristic cambial activity, beginning and end of lignification of each anatomical wood element, knowing of the first cells in the phase radial spreading and to establich first fully created blood vessels in light of thier size. In permanent week intervals from to there was taking from three beech samples gyrate from the perimeter of stem. Taking took place by special tool trephor. Thanks to immediate putting of the samples to fixative solution, the living cells died and there was creating of new annual ring. After one week the put samples in fixative solution there was their shortening and their transversal sides were marked. Before cutting the samples one the microton there was dehydration thanks to their ethanol row and each sample was cast into plastic cassette by paraphin. Cutting was done by rottate microton. On each slide there were put four cuts. Slides wire into plastic containers, which were then put into wessels with chemicals. The solution of saphrinin coloured dead cells to red. The solution of astra blue coloured living cells to blue. These colored preparations were stained by Canadian balzam and covered by slide. The evaluation of preparations took place under light microscope. The cambial activity of all samples began at intervals at temperature of the last week 9,1 C degres. The cambial yone was made of 6 7 cells during the reactivisation. The cambial activity was finished between at the average temperature 17,72 C degres in the last week. The first cells in the phase of the first spreading appeared from , and the fully size was one The lignification of cell walls continued days after the end of cambial activity. The maximal growth was at intervals μm, the average 1887 μm by Gompertz function. The result of radial growth 1887 μm is 38,3 μm bigger than numbers found out by microscopic measuring, which is 2% departure. 51
52 9 Literatura [1] BALABÁN, K. (1955): Nauka o dřevě. Č. 1, Anatomie dřeva. Praha, 216 s. [2] BEGUM, S., NAKABA, S., ORIBE, Y., KUBO, T., RUNADA, R. (2007): Induction of cambial reactivation by localized heating in a deciduous hardwood hybrid poplar (Populus sieboldii P. Grandidentata). Ann Bot 100: s. [3] BLAŽEJ, A. (1975): Chémia dreva. Bratislava, 221 s. [4] ČUFAR, K., CHERUBINI, M., GRIČAR, J., PRISLAN, P., SPINA, S., ROMAGNOLI, M. (2011): Xylem end phloem formation in chestnut (Castanea sativa Mill.) during the 2008 growing season. Dendrochronologia, Articke in Press, s. [5] ČUFAR, K., PRISLAN, P. (2008): Cambial activity and wood formation in beech (Fagus sylvatica) during the 2006 growth season. Wood research 53 (4): 12 s. [6] DUŠKOVÁ, J. (1995): Vybrané kapitoly z anatomie rostlin pro farmaceuty. Univerzita Karlova Praha, 34 s. [7] ESCHRICH, W. (1995): Funktionalle Planzenanatomie. Springer Varlag. Berlin, Heidelberg, 393 s. [8] GANDELOVÁ, L., HORÁČEK, P., ŠLEZINGEROVÁ, J. (2008): Nauka o dřevě, 2. vydání. MZLU Brno, 184 s. [9] GRIČAR, J. (2007): Xylo-end Phloemogenesis in Silver Fir (Abies alba Mill.) and Norway Spurce (Picea abies (L.) Karst.). Studia foresalia Sloveniica, Professional and Scientific Works, Ljubljana, 106 s. [10] GRIČAR, J. (2007): Xylem end phloem formation im sessile oak from Slovenia im Wood research, 53 (2): 11 s. [11] GROSSER, D. (1977): Die Hőlzer Mitteleuropas: Ein mikrophotographischer Lehratlas. 1. Auflage. Berlin: Springer Verlag Berlin Heidelberg: 208 s. [12] GRYC, V., VAVRČÍK, H., VICHROVÁ, G. (2010): Monitoring tvorby dřeva na buněčné úrovni u smrku ztepilého. Lesnická práce, 11: s.. [13] HORÁČEK, P., ŠLEZINGEROVÁ, J., GANDELOVÁ, L. (2003): Analysis of cambial 52
53 activity and formation of wood in Quercus robur. Journal of Forestr Science, 49 (9): s. [14] HORÁČEK, P., ŠLEZINGEROVÁ, J., GANDELOVÁ, L. (1999): Effects of environment on the xylogenesis of Norway spruce (Picea abies (L.) Karst) In: Wimmer, R., Vetter, 45 R. E. (eds.): Tree ring analysis: biological methodological, end enviromental aspects. CABI International, Universyty Press, Cambridge., s. [15] CHOVANEC, D., KRUTEL, F., HROMADA, E., FUCHSOVÁ, O., ČUNDERLÍK, I. (1977): Náuka o dreve 1. Vysoká škola lesnícka a drevárska. Zvolen, 255 s. [16] IVES, E. (2001): A Guide to Wood Microtomy; Making quality microslides of wood sections. Suffolk Offset Martlesham Suffolk, 114 s. [17] JANKOVSKÝ, M. (1999): Chemie dřeva. Praha, Česká zemědělská univerzita, 211 s. [18] KŐHLER, F. E. (1914): Atlas zur Pharmacopoea. Garaunterhaums, Berlin,120 s. [19] Lesnický naučný slovník: 1. díl A O. Předseda redakční rady Zdeněk Poleno. 1. Praha. Ministerstvo zemědělství, s., ISBN [20] LINNE, C., BERNARD, A. J. (1989): Atlas botanický: (dle soustavy Linnéovy). Praha, 68 s. [21] MARION, L., GRIČAR, J., OVEN P. (2007): Wood formation in urban Norway maple trees studied by the micro coring method. Dendrochronologia, s. [22] MATOVIČ, A. (1988): Nauka o dřevě. Vysoká škola zemědělská, 159 s. [23] MATOVIČ, A. (1975): Činnost kambia a charakteristika jeho xylémových derivátů u Fraxinus angustifolia Vahl. A Fraxinus excelsior L. Závěrečná zpráva výzkumného úkolu. Vysoká škola zemědělská Brno, 85 s. [24] MATOVIČ, A. (1985): Tvorba xylému v různých gradientech prostředí nižších vegetačních stupňů. Závěrečná zpráva výzkumného úkolu. Vysoká škola zemědělská Brno, 63 s. [25] MELCEROVÁ, A. (1970): Chémia dreva. Zvolen, 123 s. 53
54 [26] MENŠÍK, L. (2010): Frakcionace humusových látek lesních půd. disertační práce, MZLU Brno, 210 s. [27] NEKLOVÁ, R. (2002): Listnaté dřeviny od A do Z. Díl 1., Acer-Kolkwitzia. Praha, 367 s. [28] OLADI, R., POURTAHMASI, K., ECKSTEIN, D., BRÄUNING, A. (2010): Seasonal dynamics of wood formation in Oriental beech(fagus orientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest, Iran. Springer Verlag, 9 s. [29] PLÍVA, K. (1987): Typologický klasifikační systém ÚHUL. ÚHUL Brandýs nad Labem, 52 s. [30] POŽGAJ, A., CHOVANEC, D., KURJATKO, S., BABÍK, M. (1997): Štruktúra a vlastnosti dreva, 1. vydanie, Príroda, Bratislava, 486 s. [31] PRISLAN, P., Koch, G., ČUFAR, K., GRIČAR, J., SCHMITT, U. (2009): Topochemical investigations of cell walls in developing xylem of beech (Fagus sylvatica L.). Holzforschung, 63: s. [32] PROCHÁZKA, S. (2006): Botanika, Morfologie a fyziologie rostlin. MZLU Brno, 241 s. ISBN [33] ROSSI, S., ANFODILLO, T., MENARDI, R., (2006): Trephor: A new tool for sampling microcores from tree stems. IAWA Jurnal, 27 (1): s. [34] SASS, U., ECKSTEIN, D. (1995): The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. Trees, 9: s. [35] ŠLEZINGEROVÁ, J. (1999): Stavba dřeva cvičení. MZLU Brno, 129 s. ISBN [36] ŠLEZINGEROVÁ, J., GANDELOVÁ, L. (2002): Stavba dřeva. MZLU Brno, 187s. ISBN [37] ÚRADNÍČEK, L. (2009): Dřeviny České republiky. Lesnická práce, 367 s. ISBN [38] ÚRADNÍČEK, L., CHMELAŘ, J. (1995): Dendrologie lesnická. Část 2, Listnáče (Angiospermae). 1. MZLU Brno, 119 s. ISBN
55 [39] VAVRČÍK, H., GRYC, V. (2004): Metodika výroby mikroskopických preparátů ze vzorků dřeva. Sborník Mendelovy zemědělské a lesnické univerzity v Brně. LII, č. 4, s. [40] WAGENFŰHR, R. (1966): Anatomie des Holzes. Leipzig, VEB Fachbuchverlag, 377 s. [41] WODZICKY, T. J. (1971): Mechanism of xylem Differentiation in Pinus sylvestris L., Journal of Exp. Botany, 22: s. [42] Zpráva o stavu lesa a lesního hospodářství ČR [online] citováno 26. května Dostupné na World Wide Web: < z2009.pdf>. 55


56 10 Přílohy Obr. 19. Výzkumná plocha Rájec Domanka Obr. 20. Etanolová řada. 56


57 Obr. 21. Světelný mikroskop Leica DMLS. Obr. 22. Digestoř s chemikáliemi pro podvojné barvení. 57
MIKROSKOPICKÁ STAVBA DŘEVA
 MIKROSKOPICKÁ STAVBA DŘEVA JEHLIČNANY starší jednoduchá stavba pravidelnost JEHLIČNANY LISTNÁČE letní tracheida libriformní vlákno kambiální iniciála jarní tracheida tracheida parenchym céva parenchym
MIKROSKOPICKÁ STAVBA DŘEVA JEHLIČNANY starší jednoduchá stavba pravidelnost JEHLIČNANY LISTNÁČE letní tracheida libriformní vlákno kambiální iniciála jarní tracheida tracheida parenchym céva parenchym
MIKROSKOPICKÁ STAVBA DŘEVA LISTNÁČE
 MIKROSKOPICKÁ STAVBA DŘEVA LISTNÁČE JEHLIČNANY LISTNÁČE letní tracheida libriformní vlákno kambiální iniciála jarní tracheida tracheida parenchym céva parenchym LISTNATÉ DŘEVINY vývojově mladší složitější
MIKROSKOPICKÁ STAVBA DŘEVA LISTNÁČE JEHLIČNANY LISTNÁČE letní tracheida libriformní vlákno kambiální iniciála jarní tracheida tracheida parenchym céva parenchym LISTNATÉ DŘEVINY vývojově mladší složitější
Vznik dřeva přednáška
 Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
(cv04) Metody výroby mikroskopických preparátů z rostlinných pletiv (2. část) Preparáty pro dokumentaci tvorby dřeva a lýka
 Metody výroby mikroskopických preparátů z rostlinných pletiv (2. část) Preparáty pro dokumentaci tvorby dřeva a lýka") Mikroskopické techniky rostlinných pletiv (cv04) Metody výroby mikroskopických preparátů z rostlinných pletiv (2. část) Preparáty pro dokumentaci tvorby dřeva a lýka Osnova této prezentace Tvorba xylému
Mikroskopické techniky rostlinných pletiv (cv04) Metody výroby mikroskopických preparátů z rostlinných pletiv (2. část) Preparáty pro dokumentaci tvorby dřeva a lýka Osnova této prezentace Tvorba xylému
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 8 Mikroskopická stavba
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 8 Mikroskopická stavba
OBSAH 1 ÚVOD... 7. 1.1 Výrobek a materiál... 7 1.2 Přehled a klasifikace materiálů pro výrobu... 8 2 ZDROJE DŘEVA... 13
 OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
Mikroskopická stavba dřeva jehličnatých dřevin cvičení
 Mikroskopická stavba dřeva jehličnatých dřevin cvičení 2 Mikroskopická stavba dřeva Rostlinný organismus - základní stavební jednotkou jsou buňky (= anatomické elementy) různého typu (např. parenchymatická
Mikroskopická stavba dřeva jehličnatých dřevin cvičení 2 Mikroskopická stavba dřeva Rostlinný organismus - základní stavební jednotkou jsou buňky (= anatomické elementy) různého typu (např. parenchymatická
Biologické základy péče o stromy II.
 Biologické základy péče o stromy II. Ing. Jaroslav Kolařík, Ph.D. Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 PLETIVA VODIVÁ - lýko
Biologické základy péče o stromy II. Ing. Jaroslav Kolařík, Ph.D. Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 PLETIVA VODIVÁ - lýko
(cv03) Metody výroby mikroskopických preparátů z rostlinných pletiv
 Metody výroby mikroskopických preparátů z rostlinných pletiv") Mikroskopické techniky rostlinných pletiv (cv03) Metody výroby mikroskopických preparátů z rostlinných pletiv Osnova této prezentace příprava vzorků měkčení vzorků mikrotomy výroba preparátů barvení řezů
Mikroskopické techniky rostlinných pletiv (cv03) Metody výroby mikroskopických preparátů z rostlinných pletiv Osnova této prezentace příprava vzorků měkčení vzorků mikrotomy výroba preparátů barvení řezů
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 7 Mikroskopická stavba
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 7 Mikroskopická stavba
Makroskopická stavba dřeva
 Makroskopická stavba dřeva přednáška 2 Definice juvenilního dřeva nachází se u jehličnatých i listnatých dřevin výsledek normálních fyziologických pochodů centrální část kmene odlišná stavba a vlastnosti
Makroskopická stavba dřeva přednáška 2 Definice juvenilního dřeva nachází se u jehličnatých i listnatých dřevin výsledek normálních fyziologických pochodů centrální část kmene odlišná stavba a vlastnosti
5. Anatomická a morfologická stavba dřeva
 5. Anatomická a morfologická stavba dřeva Stonek Stonek je vegetativní orgán vyšších rostlin, jehož základními funkcemi je růstem prodlužovat rostlinu ve směru pozitivního heliotropismu, nést listy a generativní
5. Anatomická a morfologická stavba dřeva Stonek Stonek je vegetativní orgán vyšších rostlin, jehož základními funkcemi je růstem prodlužovat rostlinu ve směru pozitivního heliotropismu, nést listy a generativní
Identifikace neznámých vzorků dřev REJVÍZ, MALÉ MECHOVÉ JEZÍRKO
 MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ FAKULTA LESNICKÁ A DŘEVAŘSKÁ, ÚSTAV NAUKY O DŘEVĚ ZEMĚDĚLSKÁ 3, 613 00 BRNO,, TEL: + 420 545 134 547 Identifikace neznámých vzorků dřev REJVÍZ, MALÉ MECHOVÉ
MENDELOVA ZEMĚDĚLSKÁ A LESNICKÁ UNIVERZITA V BRNĚ FAKULTA LESNICKÁ A DŘEVAŘSKÁ, ÚSTAV NAUKY O DŘEVĚ ZEMĚDĚLSKÁ 3, 613 00 BRNO,, TEL: + 420 545 134 547 Identifikace neznámých vzorků dřev REJVÍZ, MALÉ MECHOVÉ
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 6 Makroskopická stavba
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 6 Makroskopická stavba
Faktory ovlivňující strukturu dřeva
 Faktory ovlivňující strukturu dřeva přednáška 1 strana 2 2 Připomenutí základních poznatků strana 3 3 Dřevo definice Dřevo (xylém) definice soubor rostlinných pletiv, která se u dřevin nachází mezi kambiem
Faktory ovlivňující strukturu dřeva přednáška 1 strana 2 2 Připomenutí základních poznatků strana 3 3 Dřevo definice Dřevo (xylém) definice soubor rostlinných pletiv, která se u dřevin nachází mezi kambiem
OCELOVÉ A DŘEVĚNÉ PRVKY A KONSTRUKCE Část: Dřevěné konstrukce
 OCELOVÉ A DŘEVĚNÉ PRVKY A KONSTRUKCE Část: Dřevěné konstrukce Přednáška č. 1 Doc. Ing. Antonín Lokaj, Ph.D. VŠB Technická univerzita Ostrava, Fakulta stavební, Katedra konstrukcí, Ludvíka Podéště 1875,
OCELOVÉ A DŘEVĚNÉ PRVKY A KONSTRUKCE Část: Dřevěné konstrukce Přednáška č. 1 Doc. Ing. Antonín Lokaj, Ph.D. VŠB Technická univerzita Ostrava, Fakulta stavební, Katedra konstrukcí, Ludvíka Podéště 1875,
Praktické cvičení č. 10.
 Praktické cvičení č. 10. Cvičení 10. - Stonek II. b 3 příklady stavby druhotně tloustnoucích stonků u nahosemenných a krytosemenných rostlin - Picea abies (L.) Karsten - smrk ztepilý - Tilia L. sp. - lípa
Praktické cvičení č. 10. Cvičení 10. - Stonek II. b 3 příklady stavby druhotně tloustnoucích stonků u nahosemenných a krytosemenných rostlin - Picea abies (L.) Karsten - smrk ztepilý - Tilia L. sp. - lípa
Archeologie starého dřeva a spálenišť
 MINIATLAS obsahuje dvě části. MINIATLAS mikroskopie dřeva a uhlíků pro učitele a studenty Příloha k úloze Archeologie starého dřeva a spálenišť První obsahuje výběr z anatomických obrázků různých pozorovacích
MINIATLAS obsahuje dvě části. MINIATLAS mikroskopie dřeva a uhlíků pro učitele a studenty Příloha k úloze Archeologie starého dřeva a spálenišť První obsahuje výběr z anatomických obrázků různých pozorovacích
DŘEVO pracovní list II.
 DŘEVO pracovní list II. Autor : Marie Provázková Stručný popis : Pracovní list seznamující žáky s druhy dřeva, jeho stavbou a využitím. Obsahuje různé typy úkolů - doplňovačky, přivazovačku,výpočtovou
DŘEVO pracovní list II. Autor : Marie Provázková Stručný popis : Pracovní list seznamující žáky s druhy dřeva, jeho stavbou a využitím. Obsahuje různé typy úkolů - doplňovačky, přivazovačku,výpočtovou
Výřez kmenem listnáče. parenchymatická medula
 Xylotomie (nauka o struktuře a vlastnostech dřeva) Dřevo (z technického hlediska) = lignifikované vodivé pletivo kmenů stromů (deuteroxylém) vznikající dostředivým dělením buněk kambia. Kmeny manoxylické:
Xylotomie (nauka o struktuře a vlastnostech dřeva) Dřevo (z technického hlediska) = lignifikované vodivé pletivo kmenů stromů (deuteroxylém) vznikající dostředivým dělením buněk kambia. Kmeny manoxylické:
Určování dřev podle makroskopických znaků
 Dřevo a jeho ochrana Určování dřev podle makroskopických znaků cvičení Dřevo a jeho ochrana 2 Zadání Úkoly: 1) Identifikujte základní řezy dřevem na vzorcích 2) Na vzorcích vyhledejte základní a doplňkové
Dřevo a jeho ochrana Určování dřev podle makroskopických znaků cvičení Dřevo a jeho ochrana 2 Zadání Úkoly: 1) Identifikujte základní řezy dřevem na vzorcích 2) Na vzorcích vyhledejte základní a doplňkové
VY_52_INOVACE _76 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Dub zimní
: Praktika z přírodopisu Ročník: 6., 7. Dub zimní") VY_52_INOVACE _76 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Dub zimní Dub zimní Quercus petraea ( Natt. ) Liebl. Strom s poněkud prohýbaným kmenem,
VY_52_INOVACE _76 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Dub zimní Dub zimní Quercus petraea ( Natt. ) Liebl. Strom s poněkud prohýbaným kmenem,
ZÁKLADY ARBORISTIKY. Barbora Vojáčková, a kol. Mendelova univerzita v Brně Lesnická a dřevařská fakulta. Skriptum 2013
 ZÁKLADY ARBORISTIKY Barbora Vojáčková, a kol. Skriptum 2013 Mendelova univerzita v Brně Lesnická a dřevařská fakulta 1 2 Mendelova univerzita v Brně Lesnická a dřevařská fakulta 2013 Učební text pro předmět
ZÁKLADY ARBORISTIKY Barbora Vojáčková, a kol. Skriptum 2013 Mendelova univerzita v Brně Lesnická a dřevařská fakulta 1 2 Mendelova univerzita v Brně Lesnická a dřevařská fakulta 2013 Učební text pro předmět
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 11 Rozpoznávání dřevin
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 11 Rozpoznávání dřevin
Úvod k lesním ekosystémům
 Úvod k lesním ekosystémům Lesní ekosystémy jsou nejdůležitějšími klimaxovými ekosystémy pro oblast střední Evropy, která leží v zóně temperátního širokolistého lesa. Této zóně se vymykají malé plochy jehličnatého
Úvod k lesním ekosystémům Lesní ekosystémy jsou nejdůležitějšími klimaxovými ekosystémy pro oblast střední Evropy, která leží v zóně temperátního širokolistého lesa. Této zóně se vymykají malé plochy jehličnatého
DUB ZIMNÍ (Quercus petraea)
") DUB ZIMNÍ (Quercus petraea) Velmi statný strom dosahující 30 35 m výšky, s nepravidelnou korunou se silnými větvemi, kmen má válcovitý s hrubou, šedohnědou, podélně a hluboko trhlinatou borkou. Listy má
DUB ZIMNÍ (Quercus petraea) Velmi statný strom dosahující 30 35 m výšky, s nepravidelnou korunou se silnými větvemi, kmen má válcovitý s hrubou, šedohnědou, podélně a hluboko trhlinatou borkou. Listy má
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 9 Submikroskopická stavba
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 9 Submikroskopická stavba
Vitální barvení, rostlinná buňka, buněčné organely
 Vitální barvení, rostlinná buňka, buněčné organely Vitální barvení používá se u nativních preparátů a rozumíme tím zvýšení kontrastu určitých buněčných složek v živých buňkách, nebo tkáních pomocí barvení
Vitální barvení, rostlinná buňka, buněčné organely Vitální barvení používá se u nativních preparátů a rozumíme tím zvýšení kontrastu určitých buněčných složek v živých buňkách, nebo tkáních pomocí barvení
STAVBA ROSTLINNÉHO TĚLA
 STAVBA DŘEVA STAVBA ROSTLINNÉHO TĚLA JEDNODĚLOŽNÉ ROSTLINY X DVOJDĚLOŽNÉ ROSTLINY JEDNODĚLOŽNÉ ROSTLINY palmy, bambus Nemohou druhotně tloustnout (přirůstat)!! DVOUDĚLOŽNÉ ROSTLINY mají sekundární dělivé
STAVBA DŘEVA STAVBA ROSTLINNÉHO TĚLA JEDNODĚLOŽNÉ ROSTLINY X DVOJDĚLOŽNÉ ROSTLINY JEDNODĚLOŽNÉ ROSTLINY palmy, bambus Nemohou druhotně tloustnout (přirůstat)!! DVOUDĚLOŽNÉ ROSTLINY mají sekundární dělivé
Ing. Lubomír Kacálek III/2 Inovace a zkvalitnění výuky prostřednictvím ICT VY_32_INOVACE_TDŘ0513Vady dřeva I. vady struktury dřeva
 Název školy Číslo projektu Autor Název šablony Název DUMu Stupeň a typ vzdělání Vzdělávací obor Tematický okruh Druh učebního materiálu Cílová skupina Anotace Klíčová slova Střední odborná škola Luhačovice
Název školy Číslo projektu Autor Název šablony Název DUMu Stupeň a typ vzdělání Vzdělávací obor Tematický okruh Druh učebního materiálu Cílová skupina Anotace Klíčová slova Střední odborná škola Luhačovice
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 15 Modřín Ing. Hana
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 15 Modřín Ing. Hana
MENDELOVA UNIVERZITA V BRNĚ LESNICKÁ A DŘEVAŘSKÁ FAKULTA ÚSTAV NAUKY O DŘEVĚ
 MENDELOVA UNIVERZITA V BRNĚ LESNICKÁ A DŘEVAŘSKÁ FAKULTA ÚSTAV NAUKY O DŘEVĚ MONITORING TVORBY DŘEVA BUKU LESNÍHO (Fagus sylvatica L.) NA BUNĚČNÉ ÚROVNI NA VÝZKUMNÉ PLOŠE RÁJEC HOLÍKOV BAKALÁŘSKÁ PRÁCE
MENDELOVA UNIVERZITA V BRNĚ LESNICKÁ A DŘEVAŘSKÁ FAKULTA ÚSTAV NAUKY O DŘEVĚ MONITORING TVORBY DŘEVA BUKU LESNÍHO (Fagus sylvatica L.) NA BUNĚČNÉ ÚROVNI NA VÝZKUMNÉ PLOŠE RÁJEC HOLÍKOV BAKALÁŘSKÁ PRÁCE
Klíč k makroskopickému určování vybraných dřev jehličnatých a listnatých dřevin
 Mendelova univerzita v Brně Lesnická a dřevařská fakulta Ústav nauky o dřevě Stavba dřeva Klíč k makroskopickému určování vybraných dřev jehličnatých a listnatých dřevin praktická pomůcka do cvičení Tento
Mendelova univerzita v Brně Lesnická a dřevařská fakulta Ústav nauky o dřevě Stavba dřeva Klíč k makroskopickému určování vybraných dřev jehličnatých a listnatých dřevin praktická pomůcka do cvičení Tento
Praktické cvičení č. 1.
 Praktické cvičení č. 1. Cvičení 1. 1. Všeobecné pokyny ke cvičení, zápočtu a zkoušce Bezpečnost práce 2. Mikroskopie - mikroskop a mikroskopická technika - převzetí pracovních pomůcek - pozorování trvalého
Praktické cvičení č. 1. Cvičení 1. 1. Všeobecné pokyny ke cvičení, zápočtu a zkoušce Bezpečnost práce 2. Mikroskopie - mikroskop a mikroskopická technika - převzetí pracovních pomůcek - pozorování trvalého
Rostlinná pletiva. Milan Dundr
 Rostlinná pletiva Milan Dundr Pletiva soubory buněk vykonávají stejné funkce přibližně stejný tvar a velikost Rozdělení pletiv - podle tvaru buněk a tloustnutí bun. stěny PARENCHYM tenké buněčné stěny
Rostlinná pletiva Milan Dundr Pletiva soubory buněk vykonávají stejné funkce přibližně stejný tvar a velikost Rozdělení pletiv - podle tvaru buněk a tloustnutí bun. stěny PARENCHYM tenké buněčné stěny
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 5 Části kmene Příčný
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 5 Části kmene Příčný
VY_52_INOVACE_78 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7.
: Praktika z přírodopisu Ročník: 6., 7.") VY_52_INOVACE_78 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Jasan ztepilý Jasan ztepilý Fraxinus excelsior L. Strom vysoký s přímým kmenem, s řídkou,
VY_52_INOVACE_78 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Jasan ztepilý Jasan ztepilý Fraxinus excelsior L. Strom vysoký s přímým kmenem, s řídkou,
Stromy. Řešení. Pracovní list
 Stromy Anotace: seznamuje žáky s vnější a vnitřní stavbou stromu a se základními rozdíly mezi jehličnatými a listnatými, opadavými a Zároveň vysvětluje rozdíl mezi krytosemennými a nahosemennými rostlinami.
Stromy Anotace: seznamuje žáky s vnější a vnitřní stavbou stromu a se základními rozdíly mezi jehličnatými a listnatými, opadavými a Zároveň vysvětluje rozdíl mezi krytosemennými a nahosemennými rostlinami.
Stonek. Stonek příčný řez nahosemenná rostlina borovice (Pinus)
") Stonek Stonek příčný řez nahosemenná rostlina borovice (Pinus) Legenda: 1 dřeň, 2 dřevo (xylém), 3 dřeňový paprsek, 4 pryskyřičný kanálek v xylému, 5 lýko (floém), 6 primární kůra, 7 pryskyřičný kanálek
Stonek Stonek příčný řez nahosemenná rostlina borovice (Pinus) Legenda: 1 dřeň, 2 dřevo (xylém), 3 dřeňový paprsek, 4 pryskyřičný kanálek v xylému, 5 lýko (floém), 6 primární kůra, 7 pryskyřičný kanálek
1) Pokud dlouhé svisle zavěšené těleso (např. lano) neunese svou vlastní tíhu, jakým opatřením nedosáhneme zlepšení?
 Pokud dlouhé svisle zavěšené těleso (např. lano) neunese svou vlastní tíhu, jakým opatřením nedosáhneme zlepšení?") 1) Pokud dlouhé svisle zavěšené těleso (např. lano) neunese svou vlastní tíhu, jakým opatřením nedosáhneme zlepšení? a) Zvětšením průřezu tělesa b) Zkrácením tělesa c) Použitím pevnějšího materiálu d)
1) Pokud dlouhé svisle zavěšené těleso (např. lano) neunese svou vlastní tíhu, jakým opatřením nedosáhneme zlepšení? a) Zvětšením průřezu tělesa b) Zkrácením tělesa c) Použitím pevnějšího materiálu d)
Modřín opadavý. VY_52_INOVACE_81 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7.
: Praktika z přírodopisu Ročník: 6., 7.") Modřín opadavý VY_52_INOVACE_81 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Modřín opadavý Larix decidua Miller Strom velkých rozměrů, opadavý jehličnan,
Modřín opadavý VY_52_INOVACE_81 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Modřín opadavý Larix decidua Miller Strom velkých rozměrů, opadavý jehličnan,
Buňka cytologie. Buňka. Autor: Katka www.nasprtej.cz Téma: buňka stavba Ročník: 1.
 Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
ROSTLINNÁ PLETIVA I. Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list.
 je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list.") ROSTLINNÁ PLETIVA I Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list. Orgány jsou složeny lž z buněk, které tvoří uvnitř orgánů ů odlišná uskupení pletiva.
ROSTLINNÁ PLETIVA I Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list. Orgány jsou složeny lž z buněk, které tvoří uvnitř orgánů ů odlišná uskupení pletiva.
Jak psát závěrečnou práci na LDF
 17. 3. 2014, Brno Připravil: Hanuš Vavrčík Náležitosti a členění na kapitoly strana 2 Čím se řídit? Směrnice děkana č. 2/2007 O úpravě písemných prací a o citaci dokumentů užívaných v kvalifikačních pracích
17. 3. 2014, Brno Připravil: Hanuš Vavrčík Náležitosti a členění na kapitoly strana 2 Čím se řídit? Směrnice děkana č. 2/2007 O úpravě písemných prací a o citaci dokumentů užívaných v kvalifikačních pracích
Makroskopická stavba dřeva
 Makroskopická stavba dřeva přednáška 2 Ústav nauky o dřevě budova B, 6. NP zaměření ústavu: anatomická stavba dřeva dendrochronologie arboristika fyzikální a mechanické vlastnosti dřeva sušení a modifikace
Makroskopická stavba dřeva přednáška 2 Ústav nauky o dřevě budova B, 6. NP zaměření ústavu: anatomická stavba dřeva dendrochronologie arboristika fyzikální a mechanické vlastnosti dřeva sušení a modifikace
Užitková tropická dřeva. Makroskopická stavba dřeva
 Makroskopická stavba dřeva Znaky makroskopické stavby dřeva - základní letokruhy a přírůstové zóny dřeňové paprsky cévy pryskyřičné kanálky dřeňové skvrny suky - doplňkové barva (jádro, běl, vyzrálé dřevo)
Makroskopická stavba dřeva Znaky makroskopické stavby dřeva - základní letokruhy a přírůstové zóny dřeňové paprsky cévy pryskyřičné kanálky dřeňové skvrny suky - doplňkové barva (jádro, běl, vyzrálé dřevo)
PROJEKT: OP VK 1.4 61345741. Název materiálu: NAHOSEMENNÉ ROSTLINY - JEHLIČNANY VY_52_INOVACE_K1_P36_3
 Název materiálu: NAHOSEMENNÉ ROSTLINY - JEHLIČNANY VY_52_INOVACE_K1_P36_3 Anotace Autor Jazyk Očekávaný výstup Speciální vzdělávací potřeby Klíčová slova Druh učebního materiálu Druh interaktivity Cílová
Název materiálu: NAHOSEMENNÉ ROSTLINY - JEHLIČNANY VY_52_INOVACE_K1_P36_3 Anotace Autor Jazyk Očekávaný výstup Speciální vzdělávací potřeby Klíčová slova Druh učebního materiálu Druh interaktivity Cílová
TÉMA: Stromy. (listnaté) Vytvořil: Mgr. Aleš Sucharda Dne: 30. 10. 2011 VY_32_inovace/7_402
 Vytvořil: Mgr. Aleš Sucharda Dne: 30. 10. 2011 VY_32_inovace/7_402") TÉMA: Stromy (listnaté) Vytvořil: Mgr. Aleš Sucharda Dne: 30. 10. 2011 VY_32_inovace/7_402 1 Anotace: Výukový materiál prezentace seznamuje žáky s listnatými stromy Jazyk: Český jazyk Očekávaný výstup:
TÉMA: Stromy (listnaté) Vytvořil: Mgr. Aleš Sucharda Dne: 30. 10. 2011 VY_32_inovace/7_402 1 Anotace: Výukový materiál prezentace seznamuje žáky s listnatými stromy Jazyk: Český jazyk Očekávaný výstup:
MENDELOVA UNIVERZITA V BRNĚ. Lesnická a dřevařská fakulta. Ústav nauky o dřevě
 MENDELOVA UNIVERZITA V BRNĚ Lesnická a dřevařská fakulta Ústav nauky o dřevě Monitoring tvorby dřeva buku lesního (Fagus sylvatica L.) na buněčné úrovni DIPLOMOVÁ PRÁCE 212/213 Bc. Tomáš Ondrouch Prohlášení
MENDELOVA UNIVERZITA V BRNĚ Lesnická a dřevařská fakulta Ústav nauky o dřevě Monitoring tvorby dřeva buku lesního (Fagus sylvatica L.) na buněčné úrovni DIPLOMOVÁ PRÁCE 212/213 Bc. Tomáš Ondrouch Prohlášení
Identifikace dřeva. Mikroskopické techniky rostlinných pletiv
 Mikroskopické techniky rostlinných pletiv Identifikace dřeva Osnova této prezentace identifikace dřeva makroskopická identifikace recentního dřeva mikroskopická identifikace recentního dřeva mikroskopická
Mikroskopické techniky rostlinných pletiv Identifikace dřeva Osnova této prezentace identifikace dřeva makroskopická identifikace recentního dřeva mikroskopická identifikace recentního dřeva mikroskopická
Mikroskopická stavba dřeva listnatých dřevin cvičení
 Dřevo a jeho ochrana Mikroskopická stavba dřeva listnatých dřevin cvičení Dřevo a jeho ochrana 2 Mikroskopická stavba dřeva Listnaté dřeviny - vývojově mladší -> anatomické elementy již specializovány
Dřevo a jeho ochrana Mikroskopická stavba dřeva listnatých dřevin cvičení Dřevo a jeho ochrana 2 Mikroskopická stavba dřeva Listnaté dřeviny - vývojově mladší -> anatomické elementy již specializovány
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 17 Dub Ing. Hana Márová
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 17 Dub Ing. Hana Márová
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 13 Jedle Ing. Hana Márová
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 13 Jedle Ing. Hana Márová
Stavba stonku. Stavba stonku
 Stavba stonku Stonek je nadzemní část rostliny, která nese listy, pupeny a generativní orgány (květ, plod a semeno). Její další funkcí je ukládání zásob, zajištění transportu živin a případně má i funkci
Stavba stonku Stonek je nadzemní část rostliny, která nese listy, pupeny a generativní orgány (květ, plod a semeno). Její další funkcí je ukládání zásob, zajištění transportu živin a případně má i funkci
VY_52_INOVACE_73 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7.
: Praktika z přírodopisu Ročník: 6., 7.") VY_52_INOVACE_73 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Borovice lesní Borovice lesní, sosna Pinus silvestris L. Strom středních rozměrů, vždyzelený,
VY_52_INOVACE_73 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Borovice lesní Borovice lesní, sosna Pinus silvestris L. Strom středních rozměrů, vždyzelený,
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková
 Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
I kov, či keramika mají svoji strukturu, ale ve vlastnostech jsou v porovnání se dřevem velmi homogenní.
 Obsah: Cílem této části předmětu je přiblížit Vám přírodní dřevo a dřevní kompozity z hlediska jejich vlastností, abyste byli schopni při vaší pedagogické činnosti, ale i v praktickém životě použít dřevo
Obsah: Cílem této části předmětu je přiblížit Vám přírodní dřevo a dřevní kompozity z hlediska jejich vlastností, abyste byli schopni při vaší pedagogické činnosti, ale i v praktickém životě použít dřevo
VEGETATIVNÍ ORGÁNY ROSTLIN
 VEGETATIVNÍ ORGÁNY ROSTLIN 13 Soubory určitých pletiv vytvářejí u rostlin rostlinné orgány, a to buď vegetativního nebo generativního charakteru. Vegetativní orgány slouží rostlinám k zajištění růstu,
VEGETATIVNÍ ORGÁNY ROSTLIN 13 Soubory určitých pletiv vytvářejí u rostlin rostlinné orgány, a to buď vegetativního nebo generativního charakteru. Vegetativní orgány slouží rostlinám k zajištění růstu,
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO. Modelová rostlina suchozemská semenná neukončený růst specializované části
 Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Přehled fyzikálních vlastností dřeva
 Dřevo a jeho ochrana Přehled fyzikálních vlastností dřeva cvičení Dřevo a jeho ochrana 2 Charakteristiky dřeva jako materiálu Anizotropie = na směru závislé vlastnosti Pórovitost = porézní materiál Hygroskopicita
Dřevo a jeho ochrana Přehled fyzikálních vlastností dřeva cvičení Dřevo a jeho ochrana 2 Charakteristiky dřeva jako materiálu Anizotropie = na směru závislé vlastnosti Pórovitost = porézní materiál Hygroskopicita
Mikroskopické preparační techniky pro analýzu rostlinných stonků
 Mikroskopické preparační techniky pro analýzu rostlinných stonků Cílem cvičení je demonstrace rychlé a efektivní mikroskopické preparační metody, která byla vyvinuta na základě letitých zkušeností v laboratořích
Mikroskopické preparační techniky pro analýzu rostlinných stonků Cílem cvičení je demonstrace rychlé a efektivní mikroskopické preparační metody, která byla vyvinuta na základě letitých zkušeností v laboratořích
Název: VNITŘNÍ STAVBA STONKU
 Název: VNITŘNÍ STAVBA STONKU Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2. a 3. (1. ročník vyššího
Název: VNITŘNÍ STAVBA STONKU Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2. a 3. (1. ročník vyššího
Stavba dřeva. Reakční dřevo. přednáška
 Reakční dřevo přednáška 2 Definice 3 Reakční dřevo používáme pro označení tlakového a tahového dřeva. tlakové dřevo se tvoří u jehličnatých dřevin tahové dřevo se tvoří u listnatých dřevin Místo výskytu
Reakční dřevo přednáška 2 Definice 3 Reakční dřevo používáme pro označení tlakového a tahového dřeva. tlakové dřevo se tvoří u jehličnatých dřevin tahové dřevo se tvoří u listnatých dřevin Místo výskytu
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce
 BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
Škody zvěří na lesních porostech
 Škody zvěří na lesních porostech Odhady zastoupení jedinců poškozených zvěří byly získány na základě dat pozemního šetření druhého cyklu Národní inventarizace lesů. Šetření bylo provedeno na počtu 7 772
Škody zvěří na lesních porostech Odhady zastoupení jedinců poškozených zvěří byly získány na základě dat pozemního šetření druhého cyklu Národní inventarizace lesů. Šetření bylo provedeno na počtu 7 772
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.16 Vady dřeva Kapitola 2 Čelní trhliny
Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.16 Vady dřeva Kapitola 2 Čelní trhliny
VY_32_INOVACE_097. VÝUKOVÝ MATERIÁL zpracovaný v rámci projektu EU peníze školám
 VY_32_INOVACE_097 VÝUKOVÝ MATERIÁL zpracovaný v rámci projektu EU peníze školám Registrační číslo projektu: CZ. 1.07. /1. 5. 00 / 34. 0696 Šablona: III/2 Název: Topol osika - prezentace Vyučovací předmět:
VY_32_INOVACE_097 VÝUKOVÝ MATERIÁL zpracovaný v rámci projektu EU peníze školám Registrační číslo projektu: CZ. 1.07. /1. 5. 00 / 34. 0696 Šablona: III/2 Název: Topol osika - prezentace Vyučovací předmět:
Rostlinná pletiva podle tvaru buněk a síly buněčné stěny Úvod - Doplňte chybějící místa v textu:
 Praktické cvičení č. 5 Téma: Pletiva (protokol byl sestaven z pracovních listů, které vytvořila Mgr. Pavla Trčková a jsou součástí DUM) Materiál a pomůcky: Bezová duše, sítina, hruška, stonek hluchavky,
Praktické cvičení č. 5 Téma: Pletiva (protokol byl sestaven z pracovních listů, které vytvořila Mgr. Pavla Trčková a jsou součástí DUM) Materiál a pomůcky: Bezová duše, sítina, hruška, stonek hluchavky,
TŘÍLAMELOVÉ PARKETY. Může obsahovat dobře srostlé suky o průměru až 3 mm, ovšem jen
 TŘÍLAMELOVÉ PARKETY Dub Select Dřevo s poměrně homogenní barvou. Mezi jednotlivými lamelami mohou být patrné malé barevné odchylky. Materiál může obsahovat dřeňové paprsky. Může obsahovat dobře srostlé
TŘÍLAMELOVÉ PARKETY Dub Select Dřevo s poměrně homogenní barvou. Mezi jednotlivými lamelami mohou být patrné malé barevné odchylky. Materiál může obsahovat dřeňové paprsky. Může obsahovat dobře srostlé
vznik: během růstu stromu během těžby a dopravy během uskladnění postihují kvalitu, zejména fyzikální a mechanické vlastnosti
 VADY SUROVÉHO DŘÍVÍ VADA = změna vnějšího vzhledu dřeva, porušení jeho pravidelné struktury, odchylky od normální stavby dřeva, které nepříznivě ovlivňují jeho účelové využití. postihují kvalitu, zejména
VADY SUROVÉHO DŘÍVÍ VADA = změna vnějšího vzhledu dřeva, porušení jeho pravidelné struktury, odchylky od normální stavby dřeva, které nepříznivě ovlivňují jeho účelové využití. postihují kvalitu, zejména
Vypracoval Mgr. David Mikoláš, 22. 9. 2008 DŘEVO
 Vypracoval Mgr. David Mikoláš, 22. 9. 2008 DŘEVO CO JE TO DŘEVO Dřevo je pevné pletivo stonků vyšších rostlin, které označujeme jako dřeviny. Vzniká v rostlinách z meristémových buněk. CHEMICKÉ SLOŽENÍ
Vypracoval Mgr. David Mikoláš, 22. 9. 2008 DŘEVO CO JE TO DŘEVO Dřevo je pevné pletivo stonků vyšších rostlin, které označujeme jako dřeviny. Vzniká v rostlinách z meristémových buněk. CHEMICKÉ SLOŽENÍ
VY_52_INOVACE_84 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Topol osika
: Praktika z přírodopisu Ročník: 6., 7. Topol osika") VY_52_INOVACE_84 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Topol osika Topol osika (osika obecná) Populus tremula L. Strom středních rozměrů se
VY_52_INOVACE_84 Vzdělávací oblast: člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Topol osika Topol osika (osika obecná) Populus tremula L. Strom středních rozměrů se
VY_52_INOVACE_106 Vzdělávací oblast: Člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7.
: Praktika z přírodopisu Ročník: 6., 7.") VY_52_INOVACE_106 Vzdělávací oblast: Člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Třešeň ptačí Třešeň ptačí Prunus avium (L.) Moench. Strom středně velký, opadavý,
VY_52_INOVACE_106 Vzdělávací oblast: Člověk a příroda Vzdělávací obor (předmět): Praktika z přírodopisu Ročník: 6., 7. Třešeň ptačí Třešeň ptačí Prunus avium (L.) Moench. Strom středně velký, opadavý,
Dřevo hlavní druhy dřeva, vlastnosti, anizotropie
 Dřevo hlavní druhy dřeva, vlastnosti, anizotropie Dřevo Dřevo je vnitřní zdřevnatělá část kmenu, větví a kořenů bez kůry a lýka. Strom obsahuje 70 až 90 objemových % dřeva. Tvorba dřevní hmoty probíhá
Dřevo hlavní druhy dřeva, vlastnosti, anizotropie Dřevo Dřevo je vnitřní zdřevnatělá část kmenu, větví a kořenů bez kůry a lýka. Strom obsahuje 70 až 90 objemových % dřeva. Tvorba dřevní hmoty probíhá
Název: VNITŘNÍ STAVBA KOŘENE
 Název: VNITŘNÍ STAVBA KOŘENE Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2. a 3. (1. ročník vyššího
Název: VNITŘNÍ STAVBA KOŘENE Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2. a 3. (1. ročník vyššího
Průmyslové plantáže tvrdých listnáčů a jehličnatých dřevin
 Průmyslové plantáže tvrdých listnáčů a jehličnatých dřevin Ing. Václav Hurt, Ph.D. Prof. Ing. Petr Kantor, CSc. Přednáška byla uskutečněna v rámci předmětu Pěstování účelových lesů a projektu INOBIO Tento
Průmyslové plantáže tvrdých listnáčů a jehličnatých dřevin Ing. Václav Hurt, Ph.D. Prof. Ing. Petr Kantor, CSc. Přednáška byla uskutečněna v rámci předmětu Pěstování účelových lesů a projektu INOBIO Tento
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 25 Topol Ing. Hana Márová
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 25 Topol Ing. Hana Márová
DŘEVINY DŘEVOZPRACUJÍCÍHO PRŮMYSLU
 DŘEVINY DŘEVOZPRACUJÍCÍHO PRŮMYSLU Rozeznávací znaky: - Na stojato odlišnosti jednotlivých znaků (kůra, listy, jehličí) - V řezu textura, barva, vůně - K přesnému určení slouží dendrologické klíče (vylučovací
DŘEVINY DŘEVOZPRACUJÍCÍHO PRŮMYSLU Rozeznávací znaky: - Na stojato odlišnosti jednotlivých znaků (kůra, listy, jehličí) - V řezu textura, barva, vůně - K přesnému určení slouží dendrologické klíče (vylučovací
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení kvality sadebního materiálu cvičení Zadání: Objednavatel si ve školce ŠLP MENDELU
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení kvality sadebního materiálu cvičení Zadání: Objednavatel si ve školce ŠLP MENDELU
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.15 Konstrukční materiály Kapitola 4 Dřeviny
Zvyšování kvality výuky technických oborů Klíčová aktivita V. 2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V. 2.15 Konstrukční materiály Kapitola 4 Dřeviny
Sešit pro laboratorní práci z biologie
 Sešit pro laboratorní práci z biologie téma: Les autor: Mgr. Alena Hyánková vytvořeno při realizaci projektu: Inovace školního vzdělávacího programu biologie a chemie registrační číslo projektu: CZ.1.07/1.1.38/01.0002
Sešit pro laboratorní práci z biologie téma: Les autor: Mgr. Alena Hyánková vytvořeno při realizaci projektu: Inovace školního vzdělávacího programu biologie a chemie registrační číslo projektu: CZ.1.07/1.1.38/01.0002
Zvyšování kvality výuky technických oborů
 Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 21 Buk Ing. Hana Márová
Zvyšování kvality výuky technických oborů Klíčová aktivita V.2 Inovace a zkvalitnění výuky směřující k rozvoji odborných kompetencí žáků středních škol Téma V.2.18 Dřeviny Kapitola 21 Buk Ing. Hana Márová
Praktické cvičení č. 8.
 Praktické cvičení č. 8. Cvičení 8. - Kořen 1. Homorhizie (kapraďorosty, jednoděložné rostliny) 2. Allorhizie (většina nahosemenných a dvouděložných rostlin) 3. Mykorhiza (ektotrofní, endotrofní) 4. Vzrostný
Praktické cvičení č. 8. Cvičení 8. - Kořen 1. Homorhizie (kapraďorosty, jednoděložné rostliny) 2. Allorhizie (většina nahosemenných a dvouděložných rostlin) 3. Mykorhiza (ektotrofní, endotrofní) 4. Vzrostný
Závislost hustoty dřeva na šířce letokruhu a procentu letního dřeva
 prosinec 2009, Brno Závislost hustoty dřeva na šířce letokruhu a procentu letního dřeva Lesnická xylologie cvičení strana 2 Lesnická xylologie 2 Osnova cvičení 1) Teorie 2) Cíl cvičení 3) Materiál a metodika
prosinec 2009, Brno Závislost hustoty dřeva na šířce letokruhu a procentu letního dřeva Lesnická xylologie cvičení strana 2 Lesnická xylologie 2 Osnova cvičení 1) Teorie 2) Cíl cvičení 3) Materiál a metodika
HABR OBECNÝ (Carpinus betulus)
") STROMY A KEŘE HABR OBECNÝ (Carpinus betulus) roste v lesích nižších poloh má šedou hladkou kůru listy jsou podobné buku, ale s pilovitým okrajem má jehnědovité květenství plodem jsou oříšky s křídlem
STROMY A KEŘE HABR OBECNÝ (Carpinus betulus) roste v lesích nižších poloh má šedou hladkou kůru listy jsou podobné buku, ale s pilovitým okrajem má jehnědovité květenství plodem jsou oříšky s křídlem
Nauka o materiálu. Přednáška č.2 Poruchy krystalické mřížky
 Nauka o materiálu Přednáška č.2 Poruchy krystalické mřížky Opakování z minula Materiál Degradační procesy Vnitřní stavba atomy, vazby Krystalické, amorfní, semikrystalické Vlastnosti materiálů chemické,
Nauka o materiálu Přednáška č.2 Poruchy krystalické mřížky Opakování z minula Materiál Degradační procesy Vnitřní stavba atomy, vazby Krystalické, amorfní, semikrystalické Vlastnosti materiálů chemické,
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
LABORATORNÍ PRÁCE Č.
 Úkol A: Pozorování parenchymu suknice cibule kuchyňské Pomůcky: cibule kuchyňská, pomůcky k mikroskopování a) Rozřízněte cibuli, vyjměte jeden vnitřní zdužnatělý list. b) Z vnitřní strany listu sejměte
Úkol A: Pozorování parenchymu suknice cibule kuchyňské Pomůcky: cibule kuchyňská, pomůcky k mikroskopování a) Rozřízněte cibuli, vyjměte jeden vnitřní zdužnatělý list. b) Z vnitřní strany listu sejměte
POLEDNÍK MONITORING LOKALITY PONECHANÉ SAMOVOLNÉMU VÝVOJI
 POLEDNÍK MONITORING LOKALITY PONECHANÉ SAMOVOLNÉMU VÝVOJI David Janik *, Dušan Adam, Pavel Unar, Tomáš Vrška, Libor Hort, Pavel Šamonil, Kamil Král Výzkumný ústav Silva Taroucy pro krajinu a okrasné zahradnictví,
POLEDNÍK MONITORING LOKALITY PONECHANÉ SAMOVOLNÉMU VÝVOJI David Janik *, Dušan Adam, Pavel Unar, Tomáš Vrška, Libor Hort, Pavel Šamonil, Kamil Král Výzkumný ústav Silva Taroucy pro krajinu a okrasné zahradnictví,
Jak psát závěrečnou práci na LDF
 28. 2. 2019, Brno Připravil: Hanuš Vavrčík Jak psát závěrečnou práci na LDF Závazné dokumenty Zásady psaní závěrečných prací Další doporučení Jak psát závěrečnou práci na LDF Závazné dokumenty Jak psát
28. 2. 2019, Brno Připravil: Hanuš Vavrčík Jak psát závěrečnou práci na LDF Závazné dokumenty Zásady psaní závěrečných prací Další doporučení Jak psát závěrečnou práci na LDF Závazné dokumenty Jak psát
Přírodní společenstva v České republice: LES. Martin Chlumský Dis. 3. ročník BI-TV PdF UK Praha Ing. Helena Jedličková Ph.D.
 Přírodní společenstva v České republice: LES Martin Chlumský Dis. 3. ročník BI-TV PdF UK Praha Ing. Helena Jedličková Ph.D. PdF MU Brno Martin Chlumský 2 Cíle hodiny Společenstvo lesa Student / ŢÁK ZŠ:
Přírodní společenstva v České republice: LES Martin Chlumský Dis. 3. ročník BI-TV PdF UK Praha Ing. Helena Jedličková Ph.D. PdF MU Brno Martin Chlumský 2 Cíle hodiny Společenstvo lesa Student / ŢÁK ZŠ:
Poznáváme vybrané druhy původních listnatých dřevin během celého roku Pomůcka k určování deseti druhů dřevin
 Poznáváme vybrané druhy původních listnatých dřevin během celého roku Pomůcka k určování deseti druhů dřevin Libuše Vodová Pedagogická fakulta MU Předkládaný materiál je koncipován jako podpůrný prostředek
Poznáváme vybrané druhy původních listnatých dřevin během celého roku Pomůcka k určování deseti druhů dřevin Libuše Vodová Pedagogická fakulta MU Předkládaný materiál je koncipován jako podpůrný prostředek
Diagnostika poškození lesních dřevin. Habituální diagnostika, defoliace, ukázky symptomů základních typů poškození
 Diagnostika poškození lesních dřevin Habituální diagnostika, defoliace, ukázky symptomů základních typů poškození ICP - Forest Lesprojekt 15 Picea abies defoliace smrk 10 % 30 % 15 % 40 % defoliace smrk
Diagnostika poškození lesních dřevin Habituální diagnostika, defoliace, ukázky symptomů základních typů poškození ICP - Forest Lesprojekt 15 Picea abies defoliace smrk 10 % 30 % 15 % 40 % defoliace smrk
Vakuola. Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich
 Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 39 Lesy v ČR Pro potřeby projektu
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 39 Lesy v ČR Pro potřeby projektu
Dřevo přírodní polymer
 Dřevo přírodní polymer De Havilland DH 98 Mosquito Morane-Saulnier M.S.406 Plymax duralové pláty pokryté překližkou živá tkáň velmi protáhlá, takže vypadají jako vlákna vlákna u jehličnatých vláken až
Dřevo přírodní polymer De Havilland DH 98 Mosquito Morane-Saulnier M.S.406 Plymax duralové pláty pokryté překližkou živá tkáň velmi protáhlá, takže vypadají jako vlákna vlákna u jehličnatých vláken až
Nadzemní biomasa a zásoba uhlíku
 Nadzemní biomasa a zásoba uhlíku V poslední době vzniká velice aktuální potřeba kvantifikace množství biomasy a uhlíku především ze dvou důvodů. Prvním je požadavek stanovení množství uhlíku vázaného v
Nadzemní biomasa a zásoba uhlíku V poslední době vzniká velice aktuální potřeba kvantifikace množství biomasy a uhlíku především ze dvou důvodů. Prvním je požadavek stanovení množství uhlíku vázaného v
NAVRHOVÁNÍ DŘEVĚNÝCH KONSTRUKCÍ DŘEVO, VLASTNOSTI DŘEVA část 1.
 Téma: NAVRHOVÁNÍ DŘEVĚNÝCH KONSTRUKCÍ DŘEVO, VLASTNOSTI DŘEVA část 1. Vypracoval: Ing. Roman Rázl TE NTO PR OJ E KT J E S POLUFINANC OVÁN EVR OPS KÝ M S OC IÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY.
Téma: NAVRHOVÁNÍ DŘEVĚNÝCH KONSTRUKCÍ DŘEVO, VLASTNOSTI DŘEVA část 1. Vypracoval: Ing. Roman Rázl TE NTO PR OJ E KT J E S POLUFINANC OVÁN EVR OPS KÝ M S OC IÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY.
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných