MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMETÁLNÍ BIOLOGIE. Bakalářská práce
|
|
|
- Denis Bezucha
- před 6 lety
- Počet zobrazení:
Transkript
1 MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMETÁLNÍ BIOLOGIE Bakalářská práce Brno 2016 Adelheid Skočovská

2 MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMETÁLNÍ BIOLOGIE MNOŽENÍ ORCHIDEJÍ V PODMÍNKÁCH IN VITRO Bakalářská práce Adelheid Skočovská Vedoucí práce: RNDr. Jaroslava Dubová, CSc. Brno
3 Bibliografický záznam Autor: Název práce: Studijní program: Studijní obor: Vedoucí práce: Adelheid Skočovská Přírodovědecká fakulta, Masarykova univerzita Ústav experimentální biologie Množení orchidejí v podmínkách in vitro Experimentální biologie Speciální biologie RNDr. Jaroslava Dubová, CSc. Akademický rok: 2015/2016 Počet stran: 47 Klíčová slova: In vitro, množení orchidejí, asymbiotické klíčení 3
4 Bibliographic Entry Author: Title of Thesis: Degree programme: Field of Study: Supervisor: Adelheid Skočovská Faculty of Sciences, Masaryk University Department of Experimental Biology In vitro orchid multiplication Experimental biology Special biology RNDr. Jaroslava Dubová, CSc. Academic Year: 2015/2016 Number of Pages: 47 Keywords: In vitro, orchid multiplication, asymbiotic germination 4
5 Abstrakt Tato bakalářská práce pojednává o množení terestrických, temperátních orchidejí metodou asymbiotického výsevu semen in vitro. Je to moderní metoda pro kultivaci a množení těchto často ohrožených rostlin v laboratorních podmínkách. V práci jsem se zabývala především shromážděním a nastudováním odborné literatury týkající se daného tématu, abychom takto získané poznatky mohla otestovat v experimentech, jež zahrnuje praktická část. Díky těmto prvotním pokusům se mi podařilo získat představu o vhodnosti či nevhodnosti použitých metod a vybrat nejlepší postupy pro další práci. Abstract This thesis is focused on reproducing of terrestrial orchids from temperate zone using asymbiotic seed germination in vitro. This is a modern laboratory method to cultivate and reproduce these endangered plants. In thesis I focused especially on collecting and studying specialized literature and using the knowledge during experiments at the practical part of this thesis. Thanks to those experiments I was able to get a vision of suitability of used methods and after this I was able to choose the best method for my further work. 5
 zadání")
6 Místo tohoto listu vložte kopii oficiálního (podepsaného) zadání práce. 6
7 Poděkování Na tomto místě bych chtěla velmi poděkovat vedoucí mé práce RNDr. Jaroslavě Dubové, CSc. za cenné rady, věnovaný čas, ochotu, trpělivost a v neposlední řadě pomoc s experimenty a za možnost jejich praktického provedení. Ráda bych také poděkovala RNDr. Milanu Balážovi, Ph.D. za věnovaná semena orchidejí pro mé experimenty. Prohlášení Prohlašuji, že jsem svoji bakalářskou práci vypracovala samostatně s využitím informačních zdrojů, které jsou v práci citovány. Brno 3. května 2016 Adelheid Skočovská 7
8 Obsah Seznam použitých zkratek LITERÁRNÍ REŠERŠE Čeleď Orchidaceae Stručná historie množení orchidejí in vitro Vlastnosti semen SEMENA Desinfekce, ošetření, klíčení Narušení povrchu semen, desinfekce Máčení, bobtnání semen Vývojové fáze klíčení semen Faktory ovlivňující klíčení Asymbiotické a symbiotické výsevy Živná média Složky médií Ztužování médií Hodnota ph Příprava médií Propagace a následná kultivace nových rostlin Charakteristika propagace jednotlivých druhů in vitro Testování životaschopnosti semen Převod rostlin do nesterilního,,ex vitro prostředí Cíl MATERIÁL A METODY Vybrané druhy orchidejí botanická charakteristika druhů Orchis morio L. syn. Anacamptis morio (L.) R. M. Bateman (vstavač kukačka) Dactylorhiza fuchsii (Druce) Soó subsp. fuchsii (prstnatec Fuchsův pravý) Orchis mascula L. subsp. signifera (Vest) Soó (vstavač mužský znamenaný) Platanthera bifolia (L.) Rich. (vemeník dvoulistý) Ošetření semen a jejich výsevy Použitá média příprava, složení Podmínky kultivace Sledování vývoje kultury Výsledky První pokus Druhý pokus Třetí pokus Diskuse Shrnutí Literatura
9 ÚVOD Čeleď Orchidaceae (vstavačovité) zahrnuje odhadem kolem druhů rostlin, proto je považována v rámci rostlinné říše za druhově nejbohatší čeleď. Současně se však jedná o rostliny nejvíce ohrožené. Většina se vyskytuje v tropických oblastech, zejména v amerických a asijských tropických pralesech (Průša, 2005). V těchto oblastech nalezneme především epifytní druhy, oproti tomu temperátní orchideje (a tedy i orchideje naší flóry) patří mezi zemní orchideje, tedy terestrity (Ježek, 2010). Všechny orchideje, bez ohledu na místo původu, jsou alespoň ve fázi klíčení existenčně závislé na úzce specializovaných druzích mykorhizních hub jde o tzv. orchideoidní mykorhizu (Gryndler et al. 2004, Ježek, 2010). Orchideje byly snad od pradávna předmětem fascinace mnoha lidí, především díky svým květům. Snad právě proto se je lidé začali pokoušet s větším či menším úspěchem množit (Ježek, 2010). Lidé postupem času získali mnoho poznatků o množení orchidejí v umělých, tedy in vitro podmínkách. Existují dva základní typy kultivace in vitro a to symbiotická (s využitím mykorhizní houby) a asymbiotická (McKendrick, 2000). V mojí práci jsem se zabývala asymbiotickou kultivací terestrických orchidejí naší flóry. Vůbec první pokusy s asymbiotickými kultivacemi se zabývaly množením tropických druhů orchidejí, které se vyznačují menší závislostí na mykorhize (zpravidla pouze ve fázi klíčení), avšak v posledních dvaceti letech se objevuje stále více studií o asymbiotických kultivacích terestrických orchidejí (Kauth et al. 2008). Orchideje mírného pásu, a tedy i naši zástupci, nedovedou odolávat konkurenčním tlakům expanzivních druhů trav a dřevin (proto je nutné pravidelné kosení lokalit jejich výskytu a odstraňování náletů dřevin). Škodlivý vliv na ně má i eutrofizace krajiny a jedovaté imise, jež poškozují mykorhizu ( kvetenacr.cz). Z těchto důvodů jsou téměř všechny druhy z čeledi vstavačovité v naší přírodě velmi ohroženými druhy a tak jsou zákonem chráněny (CITES). Je tedy přísně zakázáno jakékoli poškozování těchto rostlin (trhání, vyrývání, ap.) (Procházka, 2010). 9
10 Seznam použitých zkratek ABA = kyselina abscisová BA = benzylaminopurin C1 = kriticky ohrožený taxon C2 = silně ohrožený taxon C2t = silně ohrožený, ustupující taxon C3 = ohrožený taxon C4 = taxon vyžadující pozornost NAA = kyselina naftyloctová SDV = sterilní destilovaná voda TTC = trifenyl tetrazolium chlorid MS médium = Murashige Skoog médium 10
11 1 LITERÁRNÍ REŠERŠE 1.1 Čeleď Orchidaceae Rostlinná čeleď Orchidaceae, česky vstavačovité, zahrnuje odhadem kolem druhů a většina jejích zástupců patří mezi nejkrásnější rostliny planety. Většina vstavačovitých roste v tropických a subtropických oblastech Ameriky, Asie, Afriky a Austrálie. V České republice je dosud zaznamenáno 70 druhů a poddruhů orchidejí (Ježek, 2010). Orchideje lidi fascinovaly od nepaměti. Byly známy už ve starověku, kdy se využívalo kořenových hlíz některých druhů k výrobě salepu (sušené a rozemleté hlízy některých orchidejí), jemuž se připisovaly afrodiziakální účinky. Pojmenování rostliny získaly právě díky těmto pověrám, vychází totiž z latinského slova,,orchis, tedy česky varle, což poukazuje na podobnost podzemních hlíz s mužskými pohlavními žlázami (Ježek, 2010). Jsou to vytrvalé byliny s listnatou či jen šupinovitou lodyhou a obrovstou variabilitou v morfologii. Tato mnohotvárnost je dána rozdílnými životními formami (terestrické a epifytické druhy), odlišnými formami výživy (autotrofní, mixotrofní nebo obligátně heterotrofní), obýváním nejrůznějších podnebních pásem (tropy, subtropy, mírné pásmo, tundra) a biotopů (mokřady, suché louky, křoviny, světlé i stinné lesy) (Průša, 2005). Květy vyrůstají v úžlabí listů a jsou většinou oboupohlavné, zygomorfní (tzn. souměrné podle jedné osy, zrcadlově symetrické), trojčetné. Prostřední okvětní plátek je často zveličelý a vytváří,,pysk. Pylová zrna jsou obvykle slepena v tzv. brylky s lepkavou destičkou, díky níž se při opylování přichytí na tělo hmyzu, který tak snadno přenese velké množství pylu do květu dalšího jedince a zajistí tím efektivní opylování a následný vývoj semen (Novák a Skalický, 2009). Většina orchidejí totiž patří k cizosprašným (alogamickým) rostlinám. Plodem je nejčastěji tříchlopňová až šestichlopňová suchá tobolka vyvíjející se ze spodního semeníku, která obsahuje ohromné množství mikroskopických semen bez endospermu, s nedokonale vyvinutým embryem a tenkým síťnatým osemenním vyplněným vzduchem. Nepatrná velikost a velké množství semen zajišťuje jejich snadné šíření. Klíčení semen v přírodních podmínkách je možné pouze za přítomnosti symbiotické houby (Novák a Skalický, 2009; Průša, 2005). Všechny orchideje jsou existenčně závislé na úzce specializovaných druzích hub. Jedná se o tzv. orchidejovou mykorhizu, což je jeden z typů endomykorhizy (hyfy houby pronikají do buněk kořenů rostlin) (Gryndler et al., 2004). Semena orchidejí jsou schopna pouze 11
12 nabobtnat a vytvořit nepravé vlasové kořínky (rhizoidy), potom končí možnost jejich samostatného vývoje. Houbové vlákno pronikne do spodní části klíčícího semene a začne jej vyživovat, postupně vzniká kulovitý útvar protokorm, který následně zezelená a na jeho horní části se objeví vrcholový pupen, u báze se vytvářejí pravé kořeny, které jsou osídleny mykobiontem, jež zde zůstává po celý život rostliny (Ježek, 2010). Orchideje můžeme rozdělit například podle způsobu jejich života a to na terestrické, nebo-li zemní, a epifytní orchideje (Ježek, 2010). Terestrické orchideje rostou v zemi, odkud získávají vodu a živiny. Mezi terestrickými orchidejemi můžeme najít jak tropické zástupce (např. rod Paphiopedilum), tak zástupce mírného pásma. Tyto orchideje se nevyvíjejí celoročně, ale pouze sezónně a nepříznivou část roku (zimu) přežívají zataženy v podzemních orgánech, kterými jsou kořenové hlízy či oddenky. Prýty mají vždy vzpřímené, zakončené klasnatým či hroznovitým květenstvím, skládajícím se z různého počtu drobných květů. Rostliny jsou v naprosté většině celý život vázané na mykorhizu, proto je jejich kultivace v umělých podmínkách náročná. Tato závislost je u dospělých rostlin zpravidla mnohem silnější, než u tropických zástupců. Mezi terestrické orchideje patří i zástupci naší flóry (Ježek, 2010). Většina zástupců čeledi ale patří mezi epifytní druhy, tedy druhy žijící na jiných rostlinách (většinou na stromech). Tyto orchideje mohou přežívat pouze v mrazuprostých oblastech. Navzdory dřívějšímu přesvědčení epifytní orchideje na svých hostitelích neparazitují, jsou ve smyslu výživy naprosto samostatné a stromy využívají pouze jako podložku pro svůj růst a do pletiv hostitele nijak nezasahují. Hostitelům mohou škodit snad jen druhotně, a to že jim stíní, případně také svou hmotností. Asi nejdůležitějším orgánem epifytních druhů jsou tzv. vzdušné kořeny, plnící hned několik funkcí. Kořeny slouží nejen k ukotvování rostlin na stanovišti, ale i jako absorpční a zásobní orgány, plní asimilační funkci (u některých druhů totiž kořeny obsahují chlorofyl) a vytvářejí prostředí pro rozvoj hyf symbiotických hub. Vodu a minerální živiny přijímají pouze mladé kořenové špičky, starší části kořenů jsou pokryty tzv. velamenem, což je pletivo tvořené vrstvou tlustostěnných buněk naplněných vzduchem. Velamen chrání kořeny před ztrátou vody a propůjčuje jim charakteristickou bílou barvu. Po ovlhčení však velamen velmi rychle nasává vodu, díky čemuž propustí více světla ke spodnějším vrstvám buněk obsahujících chlorofyl (Ježek, 2010). 12
13 1.2 Stručná historie množení orchidejí in vitro Zájem o množení orchidejí začal již v roce 1800, kdy britští pěstitelé orchidejí umístili nodální segmenty stonku Phalaenopsis na organické substráty, jako je rašelina či listovka. Tyto prvotní pokusy unikly pozornosti botaniků a na dlouhou dobu upadly v zapomnění (Arditi, 2008). Bernard a Burgeff (1899) poznali roli mykorhizních hub pro klíčení semen orchidejí. Jejich experimenty se symbiotickou germinací spočívaly ve výsevu semen do květináče k mateřské rostlině (Kauth et al., 2008). Na základě experimentů Bernarda a Burgeffa Lewis Knudson dále zkoumal klíčení semen orchidejí. Pro kultivaci použil médium s přídavkem 1 % sacharózy a podařilo se mu vyklíčit semena většiny epifytických druhů orchidejí (Knudson, 1922 in Kauth et al. 2008). Tímto experimentem Knudson demonstroval, že orchidejová semena mohou klíčit in vitro bez přítomnosti mykorhizní houby, tzv. asymbiotická germinace (Kauth et al. 2008). Pro kultivaci semen, jež se mu nepodařilo v prvotních pokusech vyklíčit, vyvinul Knudson nové médium, které je dodnes široce používáno, jako tzv. Knudson C médium (Knudson, 1946 in Kauth et al., 2008). Moderní historie mikropropagace orchidejí se začala psát v roce 1949, kdy Gavino Rotor vyvinul novou, jednoduchou metodu pro vegetativní množení Phalaenopsis. Pro kultivaci použil Knudsonovo C médium (Knudson, 1946), tímto položil základy množení orchidejí v in vitro prostředí. Svou práci však publikoval v populárně naučném časopise American Orchid Society Bulletin, což je pravděpodobně důvodem, že jeho objevy unikly vedecké veřejnosti (Arditi, 2008). Asymbiotické klíčení semen in vitro je ideální pro studium růstu a vývoje semen i semenáčků orchidejí. Ačkoliv se první pokusy s asymbiotickou kultivací zaměřovaly na tropické, epifytní orchideje, v uplynulých 20 letech se prováděly četné experimenty i s terestrickými druhy (Kauth et al., 2008). Zatímco asymbiotická kultivace je oblíbenou metodou pro klíčení a následné množení orchidejí i v zahradnickém průmyslu, symbiotická kultivace získává svou popularitu v konzervačních a rekultivačních projektech (Kauth et al., 2008). 1.3 Vlastnosti semen Semena orchidejí jsou velmi malá, měří u terestrických orchidejí včetně testy pouze mm na délku a mm na šířku, u epifytních orchidejí jsou rozměry i menší, nazývají se proto,,prachová semena. Navíc semena obsahují jen velmi malé nebo vůbec 13
.")
14 žádné zásoby živin. Semena se vyvíjejí v plodech orchidejí, což je nejčastěji suchá tobolka, ve velkém množství ( na tobolku) (Rasmussen, 1995; Pierik, 1987). Semena obsahují embrya obalená suchým, síťovitým osemením (testou). Testa je mrtvé pletivo, obklopující embryo, prostor mezi ní a embryem je vyplněn vzduchem, obvykle má podlouhlý tvar, sbíhající se na koncích do špičky. Buňky osemení obsahují látky typu suberinu a díky tomu vytváří repelentní, lipofilní obal embrya se zvrásněným povrchem (viz. obr. 1), díky čemuž brání absorbci vody embryem. Tato rezistence k absorbci vody je výraznější u terestrických orchidejí, repelentní vrstva udržuje semena v půdě v suchém stavu po dlouhou dobu. Hicks A. J. (2009) Obr. 1: SEM povrchu semen orchidejí Embrya orchidejí mají oválný až kulovitý tvar a skládají se z cca 100 buněk, které mají izodiametrický tvar a jednoduchou strukturu. U většiny orchidejí nejsou embrya téměř vůbec diferencovaná: nemají vytvořené dělohy, vegetační vrchol, hypokotyl, základ kořene a ani endosperm (nebo je jen velmi málo vyvinutý) (Pierik, 1987; Sekerka et al., 2006). Díky redukovanému endospermu tak nemají semena orchidejí dostatečnou zásobu živin pro klíčení a další vývoj, to je odsuzuje k závislosti na mykorhizní houbě. Semena jsou schopna pouze nabobtnat a po nabobtnání musí v přírodě dojít k infekci semene mykorhizní houbou, která po navázání mykorhizy dodává embryu látky potřebné pro růst a vývoj (Rasmussen, 1995). Avšak dostupná data (Rasmussen, 1995) ukazují na možnost, že závislost na mykorhizní houbě je dána nedostatečně rychlým metabolismem embryí, která tedy nejsou limitována nedostatkem živin, ale jejich pomalým rozkladem. U některých druhů bylo totiž experimentálně prokázáno, že první fáze klíčení mohou úspěšně probíhat v prostředí čisté vody (např. někteří zástupci rodů Dactylorhiza, Orchis), tedy bez vnějších zdrojů živin. Tyto pokusy dokazují, že ve zralých semenech je pravděpodobně uloženo malé, koncentrované množství živin. Buňky embryí obsahují lipidová a proteinová tělíska, vzácně škrobová zrna. Toto je však druhově specifické (Rasmussen, 1995). 14
15 1.4 SEMENA Desinfekce, ošetření, klíčení Narušení povrchu semen, desinfekce Semena orchidejí mají na svém povrchu osemení, které je nepropustné pro vodu, a tak zajišťuje fyzickou dormanci semen. Tato dormance může být zrušena různými stimuly, např. tmou, teplotním režimem, v přírodě klimatickými podmínkami (kolísání teploty, způsobující zamrzání a rozmrzání), vnějším chemickým signálem. Po dozrání se u semen objevuje i morfologická dormance (Rasmussen, 1995). Před výsevem v laboratorních podmínkách je nutné semena vydesinfikovat a narušit jejich povrch, aby mohla přijímat vodu. Oba tyto požadavky (desinfekce a prolomení bariéry hydrofobity) lze spojit v jediný krok a to ošetřením roztokem chemické látky (Malmgren, 1996). Pro tyto účely se používají roztoky chlornanu vápenatého (Ca(OCl) 2 ) či chlornanu sodného (NaOCl). Jako alternativa se někdy používá peroxid vodíku (H 2 O 2 ) nebo ethanol (Rasmussen, 1995). Roztoky chlornanů se používají nejčastěji v koncentraci %. Jde o nestálé, alkalické roztoky, proto se doporučuje roztoky připravovat bezprostředně před použitím. Koncentrace roztoku a optimální doba působení se liší druh od druhu. Čas ošetření musí být dostatečně dlouhý, aby proběhla dekontaminace semen a narušení jejich povrchu, ne však příliš dlouhý, protože by pak mohlo dojít k poškození embryí. Účinek chemického ošetření zlepšuje přidání detergentu do vodného roztoku sterilizačního činidla. Pro tyto účely se obecně používá detergent Tween 80 v koncentraci do 0.5 % (Rasmussen, 1995). Chlornany mají na klíčení semen orchidejí pozitivní vliv. Chlornan sodný je silnějším oxidačním činidlem, než chlornan vápenatý (Rasmussen, 1995). Chlornan vápenatý značně stimuluje vývoj protokormů, naopak chlornan sodný rychlost klíčení inhibuje, pravděpodobně způsobuje poškození embryí. Důvod vyšší četnosti klíčení semen ošetřených Ca(OCl) 2 není zcela uspokojivě vysvětlen. Stimulační účinek ošetření povrchu semen na klíčení může mít několik vysvětlení: možný fyziologický efekt spočívá ve vymytí exogenního inhibitoru kyseliny abscisové (ABA) ze semen, tím přerušení dormance a iniciaci klíčení, dalším účinkem je narušení suberinových látek z osemení, díky čemuž se stává propustným pro vodu a umožní embryím lépe vodu nasávat (Vejsadová, 2006). Pro narušování osemení se při stratifikaci semen někdy používá i kyselina sírová. Tato aplikace byla testována i u orchidejí a při ošetření semen % roztokem H 2 SO 4 15
16 před aplikací chlornanového roztoku byl pozorován výraznější účinek narušení povrchu semen ve srovnání s použitím samotného chlornanu (Malmgren, 1992 in Rasmussen, 1995). Kromě chemické cesty narušení osemení byly popsány další procedury vedoucí k narušení povrchu semen. Mezi tyto metody patří např.: degradace buněčné stěny buněk osemení prostřednictvím enzymů (Lindén, 1992 in Rasmussen, 1995), mechanické narušení testy sterilními nástroji (Butcher a Marlow, 1989 in Rasmussen, 1995), ultrazvuk (Miyoshi a Mii, 1987 in Rasmussen, 1995), obroušení semen v písku. Všechna tato ošetření (ať už mechanická či chemická) vedou k vybělení testy do barvy slonovinové kosti (Rasmussen, 1995). Je prokázáno, že pouze vybělená semena mohou bobtnat a klíčit, zatímco nahnědlá až hnědá semena nejeví známky klíčení (Vejsadová, 2006). U symbiotických kultivací dojde k narušení osemení v oblasti mikropyle hyfou mykorhizní houby (Rasmussen, 1995) Máčení, bobtnání semen Po chemickém či mechanickém ošetření se osemení stává propustným pro vodu, která může k embryu pronikat přes mikropyle a suspenzor (Rasmussen, 1995). Jelikož nejsou embrya diferencována, jsou schopna pouze nabobtnat, což je ale důležité pro jejich další vývoj (Pierik, 1987). Dlouhá doba namáčení semen ve vodě má podobný účinek jako odstranění či částečné rozložení osemení. U některých druhů bylo v minulosti s úspěchem využito několikaměsíční namáčení semen, ale není zcela jasné, zda tato cesta není vhodná pouze pro testované druhy (Burgeff, 1936, 1954 in Rasmussen, 1995). Samotné dlouhé namáčení však zřejmě nemůže vyřešit všechny problémy spojené s porušením dormantního stavu semen (Rasmussen, 1995). Někteří výzkumníci shledávali výhodu v použití tekutého média pro ošetření před výsevem nebo přímo jako substrát pro výsev (Liddell, 1944; Kano, 1968; Miyoshi a Mii, 1987 in Rasmussen, 1995). Pozdější pokusy však tuto výhodu nepotvrdily, naopak klíčivost byla horší a při míchání média v průběhu kultivace neklíčila prakticky žádná semena (Rasmussen, 1995). Kromě zvýšení propustnosti testy je také pravděpodobné, že namáčením se současně uvolňují ze semen inhibiční látky. V semenech druhů, které neklíčí snadno, byla zjištěna asi pětkrát vyšší hladina ABA než u snadno klíčících druhů (van der Kinderen, 1987 in Rasmussen, 1995). Například po sterilizaci semen Dactylorhiza maculata trvající dvě hodiny nebyla v semenech detekována prakticky žádná ABA. Naopak aplikace ABA 16
17 inhibovala klíčení těchto semen, proto bylo potvrzeno, že endogenní hladina ABA je spojena s dormancí semen i u orchidejí (Van Waes, 1984 in Rasmussen, 1995) Vývojové fáze klíčení semen Drobná semena orchidejí mají velmi redukované embryo skládající se jen z malého počtu buněk. U embryí není patrná diferenciace, tedy nejsou zřetelné základy vegetačního vrcholu, hypokotylu ani kořene a nejsou vyvinuté ani dělohy. Rovněž endosperm, který by obsahoval zásobní látky (glycidy nebo tuky) jako zásobu energie pro embryo, není vyvinut, takže semena neobsahují téměř žádné zásobní látky jsou schopná pouze nabobtnat a následně musí v přírodních podmínkách dojít k infekci embrya příslušnou mykorhizní houbou (Pierik, 1987). Semena obsahují pouze malé množství lipidů, které se změní při klíčení v nepatrné množství cukrů (Průša, 2005). U epifytních orchidejí se vzniklý protokorm (což je shluk buněk, na němž již lze rozeznat prýtový meristém) zvětšuje v zelenou, tedy autotrofní, kuličku, z níž po dosažení určité velikosti vyrůstá první list a posléze i kořen. U terestrických druhů bývá vývoj složitější. Terestrické orchideje obvykle klíčí mnohem pomaleji, než tropické epifytní druhy (Pierik, 1987). Začne-li semeno klíčit, protokorm se prodlužuje a vyvíjí se z něj protáhlé tělísko, které protrhne osemení. Na spodní části tohoto tělíska se následně vytvoří kořenové vlásky (rhizoidy). Později na horní části útvaru vzniká hrbol, jež se změní v duté ouško (dělohu), v němž se tvoří první list. Nezelené tělísko se později mění v kořenovou hlízku. U většiny našich druhů orchidejí probíhá další vývoj tak, že se na pupenu hlízky tvoří další listy a silnější kořen, osídlený mykorhizní houbou, který semenáček zatahuje do půdy (Průša, 2005). U klíčení semen orchidejí bylo popsáno 6 vývojových stádií embryí (Yamazaki a Miyoshi, 2006 in Bektas et al., 2013), která jsou dokumentována na obr. 2. Jako fáze 0, tedy,,neklíčící stádium byl popsán stav, kdy nedochází k žádnému růstu embrya. Po nabobtnání se embryo zvětšuje a vyplňuje prostor testy. Tento stav byl označen jako fáze 1 = předklíčící stádium. Jako klíčící stádium, tedy fáze 2 se udává stav, kdy se embryo uvolňuje z osemení. Ve fázi 3 označované jako protokormové stádium je embryo zcela uvolněné z osemení. V následující fázi 4 neboli rhizoidovém stádiu dochází na povrchu protokormu k vývoji rhizoidů. V posledním stadiu, fázi 5 neboli: prýtovém stádiu, se na protokormu viditelně diferencuje prýt. 17

18 Obr. 2. Vývojová stádia klíčení embrya orchidejí: 1 - fáze 1 předklíčící stádium = zvětšené embryo vyplňuje prostor testy, 2 - fáze 2 klíčící stádium = embryo se uvolňuje z osemení, 3 - fáze 3 protokormové stádium = embryo zcela uvolněné z osemení, 4 - fáze 4 rhizoidové stádium = rhizoidy na povrchu protokormu, 5 - fáze 5,,prýtové stádium = viditelně diferencovaný prýt na protokormu. Upraveno podle: Faktory ovlivňující klíčení Klíčení a růst orchidejí ovlivňuje velké množství faktorů, jako jsou vlhkost, teplota, světlo, složení atmosféry apod. Vliv těchto faktorů je navíc silně druhově závislý. U terestrických orchidejí je klíčení a růst obtížnější, než u jejich epifytních (tropických) kolegů (Pierik, 1987). Důležitým faktorem je teplota a také průběh její periody. I rostliny pěstované zcela uměle ve sterilních podmínkách in vitro mají stejné požadavky na teplotu, jako rostliny v přírodních podmínkách (Malmgren, 1996). Teplotu kultivace je třeba přizpůsobit konkrétnímu druhu, ale současně je nutné mít na paměti, že teplota vhodná pro kultivaci mateřské rostliny nemusí vyhovovat novému kříženci, ap. (Arditi, 2008). Na teplotě závisí rychlost i procento klíčení (Rasmussen, 1995). Teplota vhodná pro klíčení většiny druhů se pohybuje v rozmezí C (Pierik, 1987). Při teplotách nad nebo pod optimem některá semena setrvávají v dormantním stavu a neklíčí, ačkoli není snížena jejich životaschopnost (Rasmussen, 1995). V přírodě dochází k sezónnímu kolísání teplot, které je nutné při kultivaci zohlednit (Rasmussen, 1995). Proto je u některých druhů k prolomení dormance nutné chladové ošetření (Pierik, 1987). Chladové ošetření se provádí obvykle udržováním semen při teplotě 5 C po dobu 4 5 týdnů. Toto ošetření patrně aktivuje metabolické dráhy, které jsou zodpovědné za organogenezi semenáčků a formování stabilní symbiotické asociace s mykorhizní houbou. Fyziologický účinek chladového ošetření pravděpodobně spočívá buď ve snížení endogenní hladiny ABA nebo ve zvýšení endogenní koncentrace cytokininů. Obě události mohou stimulovat klíčení. V přírodě je důvodem nutnosti chladové periody 18
19 (zimy) pro iniciaci klíčení nejspíš zabránění klíčení bezprostředně po rozptýlení semen. Cykly zahrnující chlazení, mrznutí a rozmrzání semen během zimy s sebou přinášejí změkčení testy, vyluhování inhibitorů klíčení a iniciaci fyziologických změn embrya (Rasmussen, 1995). V mikropropagaci orchidejí má ohromný význam světlo a to jak fotoperioda či úplná tma, tak i kvalita světla (barva, spektrální složení) a jeho zdroj (přírodní, zářivky, žárovky, atd.) (Arditi, 2008). Na druhou stranu ale není vliv světla na klíčení semen orchidejí zcela vysvětlen. Jeho vliv je rozdílný mezi druhy, velkou roli hrají světelné podmínky panující v prostředí přirozeného výskytu dané rostliny (Rasmussen, 1995). Mnoho výzkumníků preferuje inkubaci vysetých semen terestrických orchidejí ve tmě. Jejich závěry vycházejí ze zjištění, že i nízká intenzita světla klíčení semen inhibuje (Rasmussen, 1995). Mladé rostliny rostoucí ve tmě neprodukují chlorofyl a zezelenají až po přenosu na světlo. Při převodu je však nutné postupovat opatrně a rostliny zvykat na vyšší intenzitu záření postupně (Arditi, 2008). Fotoperioda je obvykle v rozmezí hodin (Pierik, 1987). Vhodnou fotoperiodu pro kultivaci daného druhu je třeba vyzkoušet experimentálně, je-li však již vyvinuta metodika pro daný druh, je vhodné použít fotoperiodu doporučovanou autorem (Arditi, 2008). Doporučuje se nízká hodnota ozářenosti, a to µmol m -2 s -1 (Pierik, 1987). Pro růst a zdárný vývoj rostlin orchidejí je také důležité spektrální složení světla, které je (jako u všech ostatních rostlin) v rozmezí vlnových délek nm, je to tzv. fotosynteticky aktivní radiace. Tyto vlnové délky jsou absorbovány asimilačními pigmenty (Taiz a Zeiger, 2010). Reakce semen orchidejí na světlo se zdá být komplikovaná a druhově specifická. U druhů, které jsou obtížně klíčitelné, může být jedním z problémů velká citlivost na světlo. Dodnes nejsou bohužel známy vlnové délky, které aktivují fytochromy v semenech orchidejí. Bylo by totiž praktické stanovit tzv. bezpečné vlnové délky, tedy takové světlo v němž lze s citlivými semeny pracovat bez rizika inhibice jejich klíčení (Rasmussen, 1995). Podle předběžných pozorování byla u orchidejí zjištěna citlivost zejména v červené oblasti spektra, která je podstatná pro následný vývoj semenáčků a mladých rostlin (Eiberg, 1970; Rasmussen a Rasmussen, 1991 in Rasmussen, 1995). Složení atmosféry v kultivačních nádobách také velmi ovlivňuje klíčení semen a následný vývoj rostlin. Bylo pozorováno vyšší procento klíčení v případech, kdy kultivační nádoby byly hermeticky uzavřené (Nakamura, 1962; Kano, 1968; McIntyre et al.,
20 in Rasmussen, 1995). Je to zajímavé, protože klíčení je proces aerobní, navíc při symbiotické kultivaci kyslík (O 2 ) vyžadují nejen semena, ale i mykorhizní houba. Současně jsou oba tyto organismy producenty oxidu uhličitého(co 2 ). Proto v atmosféře kultury obsah O 2 klesá, obsah CO 2 naopak roste, ať už se jedná o symbiotickou či asymbiotickou kultivaci. Podle Rasmussenové (Rasmussen, 1995) se jedná o analogii procesů, které se v přírodě odehrávají ve vzduchových kapsách v půdě. Obsah O 2 v půdě v přírodních podmínkách je obvykle nižší, než v atmosféře (méně než 16 %). Podle dostupných informací není klíčení orchidejí nijak zvlášť negativně ovlivňováno sníženou koncentrací O 2, s vyjímkou úplné anoxie (Rasmussen, 1995). Obsah CO 2 v půdě může činit 3 4 % oproti 0.03 % ve vzduchu nad povrchem půdy. Navíc se jeho koncentrace může lokálně zvyšovat v bezprostřední blízkosti půdních mikroorganismů, mezi které patří i mykorhizní houby. Z tohoto důvodu lze očekávat, že vyšší koncentrace CO 2 může působit na klíčení semen orchidejí stimulačně. Tento možný účinek však není dostatečně prozkoumán (Rasmussen, 1995) Asymbiotické a symbiotické výsevy Existují dva základní typy kultivace klíčení semen in vitro: asymbiotická a symbiotická (McKendrick, 2000). Asymbiotické výsevy se obvykle používají pro množení tropických orchidejí, které jsou takto snáze množitelné ve srovnání s orchidejemi terestrickými. Média, používaná pro asymbiotické kultivace (bez asistence houby) musí obsahovat veškeré anorganické i organické živiny ve formě okamžitě přijatelné pro orchideje (McKedrick, 2000). Do těchto médií se často přidávají i různé růstové regulátory typu auxinů a cytokininů (jako NAA, BA) a různá organická aditiva (např. kokosová voda, ananasová šťáva) (Bustam, 2014). Ovšem i všechny terestrické druhy v rámci rodů Dactylorhiza, Gymnadenia, Platanthera, Nigritella, Coeloglossum, Ophrys, Serapias, Himantoglossum a Anacamptis mohou být s úspěchem pěstovány asymbioticky (Malmgren, 1996). Asymbiotické výsevy jsou velmi populární metodou, široce využívanou zejména v zahradnickém průmyslu. Avšak rozdílné druhy mají různé požadavky na média a vykazují specifické reakce, proto je nutné médium vždy optimalizovat pro daný druh (Bustam, 2014). Nevýhodou těchto kultivací je vyšší výskyt kontaminací oproti výsevům symbiotickým (Chávez, 2015), což je dáno bohatějším médiem, které ve srovnání s jednoduchými médii pro symbiotické výsevy podporuje lépe i růst mikroorganismů.. 20
21 U symbiotických výsevů se médium inokuluje vhodnou mykorhizní houbou (například rod Rhizoctonia) současně s výsevem semen. Houba médium prorůstá, následně kolonizuje klíčící semena a vyživuje protokormy do doby, než se na nich vytvoří první listy a stanou se (více či méně) autotrofní. Tato metoda se často využívá pro množení terestrických orchidejí (McKendrick, 2000). Velkou výhodou této metody je, že používaná média jsou ve srovnání s médii určenými pro asymbiotické výsevy velmi jednoduchá (jedno z nejpopulárnějších médií se skládá pouze z ovesné mouky s přídavkem kvasničného extraktu). Navíc rostliny získané touto cestou jsou často silnější a více rezistentní k jiným houbovým infekcím než jejich asymbioticky namnožené protějšky (McKendrick, 2000). Kultivace za přítomnosti mykorhizní houby je preferována, protože mykobiont nejen podporuje klíčení, ale současně pomáhá předcházet kontaminacím patogenních mikroorganismů (Chávez, 2015). Symbiotické klíčení vyžaduje izolaci, purifikaci a následnou kultivaci symbiotické houby. Tyto mykorhizní houby se izolují z divoce rostoucích orchidejí, z pelotonů v jejich kořenech (Bustam, 2014). Nevýhodou symbiotických kultivací je specificita mykorhizy pro daný druh orchideje je potřeba vhodná mykorhizní houba, jinak se nemůže vyvinout symbiotický vztah, ba dokonce může vzniknout i vztah parazitický, kdy houba semenáčky zahubí (McKendrick, 2000). Proto, není-li k dispozici vhodný mykobiont, je asymbiotické klíčení lepším řešením (Chávez, 2015). Navíc bylo zjištěno, že při správně zvolených podmínkách lze asymbiotickou cestou dosáhnout stejné úspěšnosti, jako cestou symbiotickou (Bustam, 2014). 1.5 Živná média Kultury explantátů v in vitro podmínkách jsou heterotrofní a jejich výživu proto musí zajišťovat komplexní médium, obsahující jednak složku anorganických solí zahrnující jak makroelementy, tak mikroelementy, ale dále i celou řadu látek organických, které reprezentují především sacharidy jako zdroj organického uhlíku, proteinové nebo kvasniční hydrolyzáty, či aminokyseliny jako zdroj organického dusíku. Další nezbytnou složkou médií jsou také vitamíny a případně další organické látky (jako např. kokosová voda). Vývoj kultur je regulován přídavkem různých růstových regulátorů nebo fytohormonů (Šebánek a Sladký, 1988). V průběhu let byla mnohými autory publikována celá řada médií rozmanitého složení, která byla navíc četnými pracovníky modifikována pro účely různých experimentů i taxonů (Kováč, 1992). Živná média se používají buď tekutá nebo jsou 21
22 ztužována přídavkem agaru nebo jiných gelujících látek. Pro kultivaci orchidejí in vitro se dříve využívala ztužená média. Ale s rozvojem třepaček, rotátorů a podobných zařízení začali výzkumníci upřednostňovat spíše tekutá média, která mají pozitivní vliv především na proliferaci a vývoj protokormů (Rao, 1977) Složky médií Anorganické látky Makroelementy jsou prvky, které rostlina potřebuje v relativně velkém množství a patří mezi ně: vápník, hořčík, dusík, fosfor, draslík a síra. Tyto živiny se přidávají ve formě solí, jejichž poměr v každém médiu poskytuje přesně vybalancované koncentrace a kombinace vhodných živin (Arditi, 2008). Makroelementy se přidávají v milimolárních koncentracích (Šebánek a Sladký, 1988). Mikroelemnty jsou živiny, které rostliny vyžadují pouze v malém množství (Arditi, 2008). Patří mezi ně: železo, mangan, zinek, bróm, měď a molybden (Kováč, 1992). Do médií se přidávají v mikromolárních koncentracích (Šebánek a Sladký, 1988). Různá kultivační média se velmi liší v použití a obsahu mikroelementů. Jejich koncentrace a přítomnost či nepřítomnost však zřejmě není kritickým faktorem pro vývoj kultur. Nicméně je důležité mít na paměti, že mnoho z těchto mikroživin je pro rostliny ve vyšších koncentracích toxické. Soli makroelementů i mikroelementů jsou termostabilní a mohou být sterilizovány autoklávováním (Arditi, 2008). Velmi důležitým prvkem pro asymbiotické klíčení semen orchidejí je dusík a to jak jeho koncentrace, tak i jeho formy. Výzkumy ukazují, že větší čerstvé hmotnosti kultur orchidejí bylo dosaženo při vyšším poměru amonných a nitrátových iontů. Ačkoli jsou amonné ionty pro některé tropické druhy stimulující, pro terestrické druhy mohou naopak působit inhibičně. Ke zlepšení vývoje semenáčků může přispět organický dusík (Kauth et al., 2008). Jako zdroj organického dusíku lze použít jednotlivé aminokyseliny, ale jelikož mohou být metabolizovány až po určitém čase, semena je nemohou využít jakožto prvotní zdroj dusíku (Kauth et al., 2008). Avšak v kontrastu s tímto poznatkem několik výzkumníků naopak navrhuje, že by snad mohl být dusík ve formě aminokyselin lépe dostupný pro klíčící semena a vyvíjející se protokormy, na rozdíl od anorganického dusíku (van Waes & Deberberg, 1986b; Malmgren, 1993; Anderson, 1996; Malmgren, 1996; Stewart a Kane, 2006a in Kauth et al., 2008). 22
23 Organické látky Semena orchidejí mají jen minimální zdroje zásob sacharidů a z toho důvodu je jejich exogenní zdroj v in vitro kulturách nezbytným předpokladem pro klíčení (Kauth et al., 2008). Jako nejčastější zdroj organického uhlíku a energie se v médiích využívá sacharózy (Kováč, 1992). Sacharóza se používá obvykle v koncentraci 1-3 % (w/v), někdy se nahrazuje směsí glukózy a fruktózy (oba cukry v koncentraci 0,5-1,5 %), tato složka je extrémně důležitá, jako zdroj energie, zvláště pro semena klíčící ve tmě (Pierik, 1987). V přírodě mají semena k dispozici pro klíčení a první fáze vývoje embrya dva zdroje sacharidů: malé množství obsažené v embryu a druhým zdrojem jsou sacharidy získané od mykorhizní houby. Semena některých orchidejí obsahují glukoproteiny, jejichž hydrolýzou lze uvolnit glukózu, což by mohlo vysvětlovat, proč některé druhy dovedou klíčit jen v čisté vodě (Rasmussen, 1995). Ve studii, provedené Ernsem a Arditim (Erns a Arditi, 1990), v níž byl zkoumán vliv různých zdrojů sacharidů na klíčení a vývoj orchidejí bylo zjištěno, že nejsou vhodné sacharidy s dlouhými řetězci. Pokusy prováděli s glukózou (monosacharid) a maltoheptaózou (oligosacharid). Výsledky ukázaly, že mnohem větší procento semen vyklíčilo na médiu obsahující pouze glukózu. Navíc byla čerstvá hmotnost kultivovaných rostlin na médiu s maltoheptaózou mnohem nižší a přežívání semenáčků horší. Tento jev je způsoben pravděpodobně absencí enzymů v semenech orchidejí, které by dovedly štěpit vazby složitějších cukrů (Kauth et al., 2008). Vitamíny, které si rostliny samy syntetizují, jsou nezbytné pro jejich správný růst a vývoj slouží jako katalyzátory v řadě metabolických procesů. V heterotrofních kulturách in vitro mohou být některé vitamíny limitujícím faktorem růstu (Kováč, 1992), proto jsou tyto látky často diskutovanou komponentou médií pro kultury orchidejí (Pierik, 1987). Do médií pro kultivaci orchidejí se nejčastěji přidávají následující vitamíny: kyselina nikotinová, pyridoxin (vitamín B 6 ) a thiamin (vitamín B 1 ), někdy také biotin, kyselina listová a kyselina panthotenová. Není zcela jasné, zda jsou všechny, či alespoň některé z těchto vitamínů orchidejemi vyžadovány, nicméně složení médií nelze měnit bez předběžného vyzkoušení (Arditi, 2008). Vitamíny nejsou termorezistentní, a proto je nelze autoklávovat (Ten Ham, 1971 in Arditi, 2008), ale velmi často se média s obsahem vitamínů sterilizují teplem. Existuje několik názorů na autoklávování vitamínů: 1) vitamíny mohou zcela či zčásti toto ošetření 23
24 přežít, 2) explantáty orchidejí nevyžadují vitamíny, 3) cokoli zůstane z vitamínů po autoklávování aktivní může uspokojit požadavky explantátů (Arditi, 2008) Další organická aditiva Růst explántátových kultur často stimulují různé organické přísady, přidávané do média (Kováč, 1992). Při kultivaci orchidejí se používají např.: banánový homogenát, kokosová voda, kvasniční extrakt, salep (mouka získaná z hlíz orchidejí rodu Orchis včetně druhů Orchis mascula a Orchis militaris, tyto hlízy obsahují škrobnatý polysacharid glukomanan), ananasový džus, rajčatový džus, bramborový homogenát (Pierik, 1987). Tyto látky označujeme jako nedefinovatelné organické složky, protože mají velmi variabilní složení, a proto nelze přesně definovat, které podpůrné látky dané médium s jejich přídavkem, obsahuje. Organická aditiva mohou mít výrazný účinek na růst a vývoj orchidejí in vitro, který je výraznější u asymbiotických kultur (Rasmussen, 1995). Organická aditiva pravděpodobně obsahují rostlinné hormony, aminokyseliny, proteiny, sacharidy i neznámé růstové faktory. Rozdílné rostlinné druhy vyžadují odlišná aditiva a pro dosažení co nejlepších výsledků je třeba jejich kombinace zvolit experimentálně k vypracování nejvhodnějšího složení média (Malmgren, 1996). Nejčastěji zmiňovaným přírodním aditivem při kultivaci orchidejí in vitro v souvislosti se zlepšením germinační a proliferační fáze semen je tzv. kokosová voda (Bustam, 2014). Kokosová voda je tekutý endosperm semen kokosovníku ořechoplodého (Cocos nucifera). V médiu pro pěstování orchidejí byla poprvé použita v roce 1951 (Mariat, 1951 in Arditi, 2008). Působí pozitivně na proliferaci kultur (Arditi, 2008), byl pozorován rychlejší vývoj a menší mortalita mladých rostlin (Van Waes, 1984 in Rasmussen, 1995), udává se také stimulační vliv na vývoj hlíz a kořenů (McIntyre et al., 1974 in Rasmussen). Tyto pozitivní vlivy pravděpodobně zajišťují obsažené fytohormony, fytohormonům podobné látky, hexitoly, aminokyseliny a mnoho dalších látek, které na rostliny působí buď samy o sobě nebo je důležitá jejich kombinace (Arditi, 2008). Kromě fytohormonů kokosová voda obsahuje aminokyseliny, 2,6 4,4 % bílkovin, sacharidy, z nichž většina jsou cukry (především sacharóza) a malé množství škrobu. Z minerálních látek jsou hlavními prvky fosfor, vápník a draslík (Thaiphanit a Anprung, 2015). Obvykle se využívá kokosová voda ze zeleného, tedy nezralého plodu, ačkoli může být použit i tekutý endosperm plodů zralých. Po zmrazení lze toto aditivum uchovávat dlouhou dobu a může být autoklávováno (Arditi, 2008). 24
25 Fytohormony Růstové regulátory používané v in vitro kulturách lze rozdělit do čtyř základních skupin: auxiny, cytokininy, gibereliny a kyselina abscisová. O působení fytohormonů na růst kultur nerozhoduje pouze koncentrace těchto látek, ale často i jejich vzájemný poměr. Poměry vedoucí k žádoucím účinkům závisí na druhu rostliny, kultivaru, ale i typu explantátu (Kováč, 1992). Princip vlivu rostlinných růstových regulátorů na vývoj asymbiotických kultur orchidejí nebyl doposud zcela objasněn. Klíčení často podporují cytokininy (jmenovitě např.: benzylaminopurin (BA), zeatin a kinetin) a auxiny (kokrétně např.: kyselina naftyoctová (NAA), prekurzory ethylénu). Ačkoli není dobře vysvětlena role cytokininů v klíčení semen orchidejí, jejich nízké koncentrace podporují klíčení u asymbiotických výsevů mnoha terestrických druhů orchidejí. V přírodních podmínkách cytokininy vyvíjející se rostlině poskytuje pravděpodobně mykorhizní houba. Ovšem množství cytokininů, které je detekovatelné produkují pouze dva druhy mykorhizních hub, u ostatních je produkce možná, avšak v tak nízké koncentraci že ji nelze zaznamenat. To bude nejspíš důvodem, proč klíčení semen podporují pouze nízké koncentrace těchto fytohormonů např. 1 µm zeatin a kinetin (Stewart a Kane, 2006a in Kauth et al., 2008; Kauth et al., 2008). V jiných pracech bylo popsáno, že růstové regulátory nejsou pro klíčení semen orchidejí zapotřebí, naopak jejich přídavek často vede k nežádoucím efektům, jako je tvorba kalusu nebo adventivních prýtů (Pierik, 1987) Aktivní uhlí Aktivní uhlí má na kultury příznivý vliv, bez ohledu na složení média (Vij et al., 1998 in Murdad, 2006). Je charakteristické extrémně velkým specifickým povrchem (2 000 m 2 g -1 ), proto se používá pro jeho dobrou schopnost adsorbce látek. Díky této vlastnosti v kultuře adsorbuje toxické, morfogeneticky aktivní či škodlivé látky, jako jsou fenolické a karboxylové sloučeniny, ethylén, atd. produkované pletivy kultury. Na druhou stranu může adsorbovat i vitamíny a fytohormony a snižovat tak jejich aktivitu (Arditi, 2008). Podle jiných autorů přídavek aktivního uhlí v médiu zvyšuje ph a udržuje jeho hodnotu stabilní v průběhu kultivace, zvyšuje příjem dusíku, zlepšuje růst, redukuje inhibiční vliv exogenních cytokininů na růst kořenů (Eymar at al. in Murdad, 2006), zlepšuje zdravotní stav kultur (Pan a Staden, 1998 in Murdad, 2006), provzdušňuje médium a tmavá barva simuluje podobnost s půdním prostředím (Arditi, 2008). Přežívání semenáčků je podpořeno aktivním 25
26 uhlím zejména u druhů, které mají tendenci produkovat fenoly, např.: Anacamptis pyramidalis, Dactylorhiza spp., Epipactis spp., Gymnadenia spp. atd. (Werckmeister, 1971 in Rasmussen, 1995) Ztužování médií Nejčastěji využívanou látkou pro zpevnění médií pro klíčení a mikropropagaci orchidejí in vitro je po desetiletí agar. V poslední době se začalo více využívat i gumu gellan (E 418) dodávanou pod různými obchodními názvy (např.: Gelrit (Sigma) nebo Phytagel (Merck)) (Arditi, 2008). Většina typů agaru je vhodná pro komerční použití, ale pro experimentální účely je zapořebí použít agar vyšší kvality (Arditi, 2008). Oproti jiným gelujícím látkám má agar řadu výhod. Po smíchání s vodou se gel vytvoří při C a tuhne při 45 C, je tedy stabilní při teplotách obvyklých pro kultivaci. Navíc nereaguje s ostatními složkami médií a není rozkládán rostlinnými enzymy. Obvykle používaná koncentrace agaru je 8 10 g.l -1 (Kováč, 1992). Guma gellan je náhrada agaru, jedná se o přírodní anionický polysacharid, produkovaný bakteriální fermentací (bakterií Pseudomonas elodea). Je to polymer tetrasacharidu (glukóza glukuronová kyselina glukóza rhamnosa). Tato látka vyžaduje pro vytvoření gelu přítomnost bivalentních iontů Mg 2+, Ca 2+, vytváří pak bezbarvý, čistý gel (Arditi, 2008). Čirost gelu umožňuje rychlou detekci případných mikrobiálních kontaminací (Kováč, 1992). Používá se v poloviční koncentraci ve srovnání s agarem a není vhodné její opakované autoklávování. Obvykle používaná koncentrace pro kultury rostlin je 1,2 2 g.l -1 (Arditi, 2008) Hodnota ph Různé druhy orchidejí mohou mít rozdílné požadavky na ph substrátu nebo média, podle místa jejich přirozeného výskytu (Rasmussen, 1995). Hodnoty ph pro in vitro kultivaci orchidejí by se měly pohybovat v rozmezí (Pierik, 1987). Jako dolní mez se udává ph 3 nebo 4, klesne-li pod tuto hodnotu je růst značně retardovaný. Za horní mez bývá označována hodnota 6.5 (Fast, 1980 in Rasmussen, 1995). Pro kalcifilní, evropské druhy terestrických orchidejí je vhodné hodnotu ph upravit na 7.0 (Rasmussen, 1995). 26
27 Hodnotu ph je důležité při přípravě média zkontrolovat (Kováč, 1992). Použijeme-li jako organické aditivum ananasový džus, je nutné ph upravit pomocí např. hydroxidem sodným (NaOH) (Malmgren, 1996) Příprava médií Kultivační média se připravují většinou ze zásobních roztoků jeho jednotlivých složek (Kováč, 1992). Zásobní roztoky jsou zpravidla koncentrovanější roztoky jednotlivých látek nebo jejich směsí a při přípravě média se pak používají alikvotní podíly zásobního roztoku obsahující požadovanou koncentraci dané složky (Šebánek a Sladký, 1988). Při přípravě média se postupuje následovně: v destilované vodě rozpustíme (podle receptu na dané medium), správné množství makroelementů, přidáme mikroelementy, inositol a jiná organická aditiva (obsahuje-li medium tyto složky), přidáme správné množství cukru. Objem doplníme do 1 l a zkontrolujeme ph. Pro dokončení přípravy média přidáme zpevňující látku (např. agar), chceme-li ztužené medium, a provedeme sterilizaci v autoklávu. Po sterilizaci přidáváme do mírně vychladlého média termolabilní fytohormony, aminokyseliny, vitamíny a další termolabilní látky, které musí být sterilizovány filtrací. Ještě horké, vysterilizované medium rozlijeme do sterilních kultivačních nádob (Arditi, 2008). Jinou možností je příprava média z práškových směsí, ve kterých již dodavatelské firmy garantují přesnou kombinaci jednotlivých složek. 1.6 Propagace a následná kultivace nových rostlin Charakteristika propagace jednotlivých druhů in vitro Orchis Semena rodu Orchis si zachovávají životaschopnost více než rok a obvykle nejsou náročná na narušení povrchu semen (Rasmussen, 1995). Přesně podle předpokladu nereagují semena pozitivně na chladové ošetření, jsou to totiž druhy v přírodě klíčící v létě a na podzim (Riether, 1990 in Rasmussen, 1995). Nahrazením anorganického dusíku v médiu za organický zdroj lze dosáhnout vyšší klíčivosti jejich semen (Rasmussen, 1995). Semenáčky obvykle nevyžadují chladové ošetření k prolomení dormance pupenů. Pasážování je vhodné provádět každých 6 týdnů, pro udržení vysoké růstové rychlosti. Přežívání rostlin po převodu do nesterilního prostředí obvykle zlepšuje chladové ošetření in vitro kultur, před tímto převodem (Rasmussen, 1995). Rostliny lze přivést ke květu 3. jaro od vyklíčení semen (Vermeulen, 1947 in Rasmussen, 1995). 27
28 Rozdílné výsledky experimentů se semeny tohoto rodu jsou přisuzovány nevhodným podmínkám skladování semen, díky čemuž dochází k rychlému snižování jejich životaschopnosti (Rasmussen, 1995) Dactylorhiza Semena tohoto rodu jsou obvykle snadno skladovatelná a životaschopnost si zachovávají až několik let. Pro klíčení je podstatná především dostatečně vysoká vzdušná vlhkost, mnohem méně důležité je potom složení média. Ale pokud je v médiu použit organický dusík namísto anorganického, dosahuje se vyššího procenta klíčení. V prvních dnech po výsevu je klíčení inhibováno světlem (Rasmussen, 1995). Protokormy jsou po dlouhou dobu svého vývoje intolerantní ke světlu a i mladé semenáčky, jsou-li přeneseny na světlo příliš brzy, mohou v důsledku poškození zahynout (Rasmussen, 1995). U starších rostlin je fotoperioda důležitá pro zdárný vývoj, u některých druhů může být dokonce vyžadována vernalizace pro indukci rašení bohatě olistěného prýtu. Vždy je ale třeba uvažovat druhovou specifičnost (Rasmussen, 1995). Špatné klíčení semen některých druhů tohoto rodu je přisuzováno obvykle špatné kvalitě semen, případně jsou-li semena vzniklá po samooplození (Rasmussen, 1995) Platanthera Semena druhů rodu Platanthera mohou být skladována v suchu v CaCl 2 při 4 C až 31 měsíců bez znatelné ztráty životaschopnosti. Klíčení by mělo zlepšit namáčení 2 3 měsíce ve vodě (Burgeff, 1954 in Rasmussen, 1995), případně dlouhá desinfekce (až 10 hodin) roztokem Ca(OCl) 2 (Van Waes, 1984 in Rasmussen, 1995). Klíčení podporuje také přítomnost organického dusíku v živném médiu (Van Waes a Debergh, 1986b in Rasmussen, 1995). Klíčení některých druhů tohoto rodu je velmi pomalé, může trvat 3 12 měsíců, navíc je inhibováno světlem. V prvních 4 6 měsících klíčící semena vykazují jen velmi nepatrné zvětšení embrya uvnitř testy, teprve až po vytvoření rhizoidů embryo zezelená a růstová rychlost se rapidně zvýší (Stoutamire, 1964 in Rasmussen, 1995). Ke klíčení všech životaschopných semen nedochází dříve, než po 1 roce. Právě neschopnost zaznamenat tuto extrémně pomalou rychlost klíčení bývá důvodem negativních výsledků testů klíčení semen 28
29 druhů rodu Platanthera. Velké problémy s klíčením plynou pravděpodobně z nedostatečně popsaných mechanismů dormance semen (Rasmussen, 1995). Rozvoj listnatého stonku výrazně stimuluje chladové ošetření (2 5 C) (Borris, 1970; Hadley, 1970a in Rasmussen, 1995), ačkoliv se prýt u některých druhů vyvíjí i bez tohoto ošetření. Všeobecně ale platí, že přežívání rostlin po převodu do nesterilního prostředí je vyšší, jsou-li rostliny bezprostředně před převodem ošetřeny chladem (Rasmussen, 1995). Fáze kvetení je u skleníkových rostlin dosaženo obvykle po 3 4 letech od vyklíčení semen (Stoutamire, 1974; Fast, 1985 in Rasmussen, 1995) Testování životaschopnosti semen Schopnost semen vyklíčit závisí na jejich životaschopnosti. Nejčastěji se životaschopnost semen stanovuje pomocí chemické látky trifenyl tetrazolium chlorid tzv. TTC test (Kauth et al., 2008). Tato rychlá chemická metoda se využívá čím dál častěji než klasická metoda stanovení klíčivosti semen (SGC = seed germination count) (Vujanovic et al., 2000). Trifenyl tetrazolium chlorid (TTC) způsobuje zčervenání embryí životaschopných semen (Vujanovic et al., 2000). Test je založen na stanovení aktivity dehydrogenáz, které jsou přítomny v živých, respirujících buňkách. Tyto enzymy redukují TTC na formazan, což je rudá, ve vodě nerozpustná sloučenina ( 2020seedlabs.ca). Přítomnost formazanu v buňkách indikuje mitochondiální respiraci, a tak umožňuje rozlišení životaschopných a neživotaschopných semen (Ribeiro de Souza et al., 2010). Mezi výhody této metody patří zejména rychlost vyhodnocení výsledky jsou k dispozici během 24 až 48 hodin ( 2020seedlabs.ca) a nízká cena metody. Dále tuto metodu neovlivňuje faktor přítomnosti mykorhizní houby, který může zkreslovat výsledky standartních klíčících testů. Navíc se test zaměřuje na fyziologický stav embryí semen (Ribeiro de Souza et al., 2010). Z důvodu nepropustnosti testy semen terestrických orchidejí pro vodu je nutné před provedením testu semena ošetřit např. 10 % roztokem NaOCl (Vujanovic et al., 2000). U epifytních orchidejí bývají v testě často praskliny, které usnadňují prostup roztoků k embryím (Kauth et al., 2008). Jako alternativa hodnocení viability semen může být použit test s houbou Fusarium sp., která produkuje červený pigment. Ke zbarvení embryí semen terestrických orchidejí dochází po infekci touto houbou. Výsledky ukazují, že metoda je srovnatelná s TTC testem, ale před 29
30 širším využitím by bylo třeba zdokonalit metodiku. Pozornost by měla být věnována zejména druhové specifičnosti houby (pro určitý druh terestrické orchideje) a dále izolačním a testovacím postupům (Vujanovic et al., 2000). 1.7 Převod rostlin do nesterilního,,ex vitro prostředí Převod rostlin do nesterilního prostředí je asi nejnáročnějším krokem v kultivaci terestrických orchidejí. Semenáčky jsou zvyklé na velmi zhýčkané podmínky růstu. V kultivační místnosti je, oproti vnějším podmínkám, nižší intenzita světla, navíc jsou světelné i teplotní podmínky poměrně konstantní, relativní vzdušná vlhkost v kultivačních nádobách je vysoká ( %), takže semenáčky žijí v prostředí chráněném proti houbovým a bakteriálním infekcím a herbivorům (McKendrick, 2000; Rasmussen, 1995 ). Rostliny in vitro by měly být před vyjmutím z chráněného prostředí silné a životaschopné, jak jen je to možné (McKendrick, 2000). Nejvhodnější je převádět rostliny s co nejvyšším poměrem biomasy kořenů k nadzemní části (= root-schoot ratio ), proto můžeme podmínky in vitro přizpůsobit tak, aby vedly k maximalizaci tohoto poměru. Růst kořenů můžeme podpořit například infekcí symbioickou houbou, oddálením počátku fotosyntézy (udržováním rostlin co nejdelší dobu kultivace ve tmě, a to až do vzniku prvního listu), nízkou teplotou (10 C), u některých druhů i snížením koncentrace solí v médiu (Rasmussen, 1995). Asymbioticky pěstované semenáčky je možné určitou dobu před vylahvováním kultivovat v médiu inokulovaném mykorhizní houbou, pro zlepšení přežívání mladých rostlin (McKendrick, 2000). Z důvodu tak velké odlišnosti prostředí in vitro a ex vitro je nutné mladé rostliny před zasazením do zeminy postupně zvykat na nové prostředí aklimatizovat (Rasmussen, 1995). Po převodu rostlin do ex vitro prostředí je nutné semenáčky chránit zejména před vysokou intenzitou slunečního záření, které může popálit listy a vést až k úhynu rostlin. Mladé semenáčky uvyknuté na vysokou vzdušnou vlhkost nemají dostatečně vyvinutou kutikulu na listech, a tak je třeba je také pomalu zvykat na sušší prostředí. Z toho důvodu musíme rostliny chránit před vysycháním a to udržováním vyšší vzdušné vlhkosti v jejich okolí nepřímým rosením, umístěním do,,stanu z plastové fólie, ap. (McKendrick, 2000). Rostlinám hrozí, kromě poškození slunečním zářením a vysycháním, i nebezpečí v podobě nežádoucí infekce parazitickými mikroorganismy (Rasmussen, 1995). Tomu lze částečně předejít namáčením kořenů rostlin do fungicidu před jejich zasazením do substrátu (McKendrick, 2000). 30
31 Jak již bylo zmíněno výše, rostliny je třeba před převodem do nesterilního prostředí postupně aklimatizovat na jiné světelné, teplotní a především vlhkostní podmínky. U každého druhu je třeba podmínky aklimatizace přizpůsobit konkrétním potřebám. Obecně však lze postupovat tak, že rostliny přeneseme z kultivační místnosti do venkovního prostředí v uzavřených kultivačních nádobách. Postupně (v řádech dnů a týdnů) uvolňujeme víčka a postupně vpouštíme vzduch z venkovního prostředí. Pravidelně kultury kontrolujeme z důvodu případné infekce. Po aklimatizaci v kultivačních nádobách rostliny vyjmeme, opatrně odstraníme veškerý agar a semenáčky zasadíme do vhodného substrátu (McKendrick, 2000). Po přesazení pokračujeme v uvykání rostlin na nové prostředí a to udržováním vyšší vzdušné vlhkosti v jejich okolí (viz. výše), kterou postupně snižujeme (McKendrick, 2000). Pro aklimatizaci je vhodné upřednostnit umístění mladých rostlin do skleníku a ne přímo do venkovního prostředí. Ve skleníku se rostliny mohou snáze adaptovat na sezónní rytmy. Okamžitý přenos rostlin do venkovního prostředí vede často k jejich úhynu (Rasmussen, 1995). 31
32 2 Cíl Cílem teoretické části bakalářské práce byla jednak literární rešerše prací zabývajících se množením orchidejí v podmínkách in vitro. V praktické části bylo hlavním záměrem otestování základních laboratorních postupů přípravy živných médií a jejich sterilizace, desinfekce rostlinného materiálu (konkrétně semen terestrických orchidejí) a vlastních postupů manipulace ve sterilním prostředí laminárního boxu pro úspěšné asymbiotické kultivace terestrických orchidejí in vitro. 32
33 3 MATERIÁL A METODY 3.1 Vybrané druhy orchidejí botanická charakteristika druhů Výběr experimentálních druhů orchidejí byl ovlivněn jednak tím, která semena byla k dispozici a také obdobím (jeho vhodností) pro jejich výsev. První výsevy byly prováděny na podzim, a tak byly zvoleny druhy přirozeně klíčící v tomto období. To proto, že podle literatury je vhodné respektovat přirozené biorytmy rostlin i v umělých podmínkách (Malmgren, 1996; http.:// lidaforsgarden. com). Semena pro experimenty laskavě věnoval ze své sbírky RNDr. Milan Baláž, Ph.D. Zralé tobolky nasbírané v přirozených podmínkách, byly skladovány v papírových sáčcích za pokojové teploty. Semena uvolněná ze zralých, prasknutých tobolek byla použita k in vitro výsevům. Orchideje patří mezi taxony, které jsou u nás ohrožené. Nová verze červeného seznamu naší flóry navazuje na seznam cévnatých rostlin české květeny (Danihelka et al. 2012) a zohledňuje nové poznatky o výskytu cévnatých rostlin a jeho dynamice. V klasifikaci ohrožení používá již dříve zavedené kategorie škály ohrožených taxonů: C1 kriticky ohrožený taxon, C2 - silně ohrožený taxon, C3 - ohrožený taxon, C4 taxon vyžadující pozornost Orchis morio L. syn. Anacamptis morio (L.) R. M. Bateman (vstavač kukačka) Orchis morio je vzhledem a morfologií velmi variabilní druh, vytváří mnoho poddruhů a forem. Jedná se o vytrvalou bylinu s 8 20 (někdy až 40) cm dlouhou lodyhou s malými, kulovitými hlízami. Květenství se skládá ze 7 25 květů nejčastěji nachové, často růžové nebo ojediněle i bílé barvy (Procházka a Velíšek, 1983). Podle nevýrazně hnědě pruhované květní přilby připomínající šaškovskou čepici bylo odvozeno jeho druhové jméno morio (= šašek). Kvete od března do června. Dolní listy jsou kopinaté, až 14 cm dlouhé, horní kratší, pochvovité (Ježek, 2010). Orchis morio roste od nížin do podhůří, typicky na loukách, pastvinách, křovinatých stráních a lesních okrajích. Vyhovují mu bazické i kyselé, mírně vlhké půdy (Průša, 2005). U nás dříve patřil k nejhojnějším vstavačům, dnes je častější jen v Pošumaví, horním Podyjí a předhůří Karpat ( V současnosti je v Červeném seznamu hodnocen jako kriticky ohrožený druh naší květeny (C1) ( Procházka (2010) ještě uvádí 33
34 stupeň jeho ohrožení jako C2. Jeho areál je ostrůvkovitý v Evropě (včetně České republiky), v severní Africe i Malé Asii ( Dactylorhiza fuchsii (Druce) Soó subsp. fuchsii (prstnatec Fuchsův pravý) Dactylorhiza fuchsii je vytrvalá bylina, cm vysoká s přímou lodyhou. (Burnie, 2007). Květy jsou středně velké, světle fialové, růžové či bílé, vždy s tmavší kresbou na pysku, skládající se v kuželovitý klas až 14 cm dlouhý (Procházka a Velíšek, 1983). Kvete v červnu a červenci. Listy na konci zaoblené až tupě špičaté s tmavými skvrnami (Burnie, 2007). Roste od podhůří do hor, výjimečně i v pahorkatině. Vyhovují mu vlhčí, vápnité půdy. Typicky se vyskytuje na loukách, pastvinách, rašeliništích, ve světlých lesích a na lesních okrajích (Průša, 2005). Variabilita tohoto druhu (D. fuchsii) je ohromná (tvar pysku, počet květů, délka spodních listen, ap.) (Průša, 2005). Na území České Republiky je nejhojněji rozšířený právě poddruh D. f. subsp. fuchsii, celkově jde o nejrozšířejnější druh z prstnatců u nás. Nicméně i přes to se jedná z hlediska ohrožení o vzácnější druh, vyžadující další pozornost (C4a) ( botany.cz) Orchis mascula L. subsp. signifera (Vest) Soó (vstavač mužský znamenaný) Orchis mascula je vytrvalá bylina, vysoká cm s kulovitými až vejčitými hlízami. Listy jsou podlouhle kopinaté, přízemní mají u báze tmavě červené tečky, které se vyskytují i na spodní části lodyhy (Procházka a Velíšek, 1983). Květy uspořádané v klasu jsou světle nachové nebo růžové, pysk je na bázi bělavý až nazelenalý, tmavě tečkovaný. Kvete od května do června ( Listy jsou přízemní, kopinaté, se skvrnami či bez nich (Ježek, 2010). Roste od pahorkatin do hor na chudých loukách, ve světlých lesích a lesních okrajích. Vyhovují mu až mírně suché zásadité půdy (Průša, 2005). V Čechách roste jen vzácně, na Moravě roztroušeně (Procházka, 2010). V Červeném seznamu je hodnocený jako silně ohrožený, ustupující druh (C2t) ( Platanthera bifolia (L.) Rich. (vemeník dvoulistý) Platanthera bifolia je ekologicky přizpůsobivá, silně vonná orchidej. V dospělosti je málo závislá na výživě prostřednictvím hub (mykorhize). Nepříznivé období přečkává díky dvojici podlouhlých, vejčitých hlíz (Ježek, 2010). Jedná se o vytrvalou bylinu, cm vysokou s holou květní lodyhou, na nichž jsou při bázi dva hnědé šupinovité objímavé lístky 34
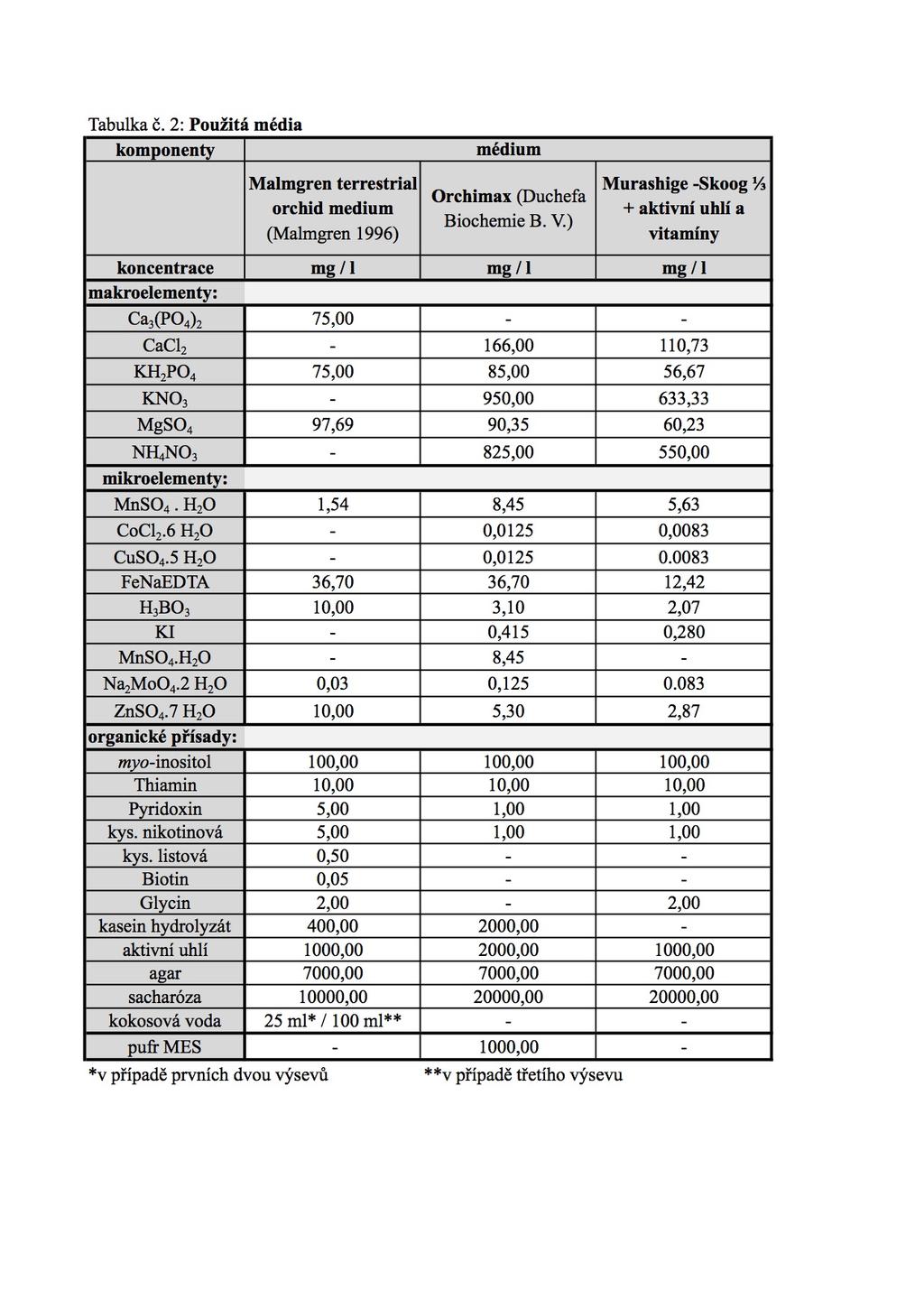
35 a nad nimi pak dva vstřícné, široce oválné, světle zelené listy (Procházka a Velíšek, 1983). Má řídké a bohaté květenství dlouhé do 25 cm sestávající se z zelenobílých květů. Kvete od května do srpna (Ježek, 2010). Roste na loukách, křovinatých stráních, vřesovištích a světlých lesích od nížin do hor. Vyhledává vlhčí, humózní půdy (Průša, 2005). Vyskytuje se v Evropě, severní Africe, Malé Asii, na Kavkaze a v Persii. V České republice je hodnocený jako ohrožený druh naší flóry (C3) ( 3.2 Ošetření semen a jejich výsevy Práce byla prováděna sterilně v laminárním boxu Gelaire. Bobtnání a desinfekce semen bylo prováděno pomocí plastových injekčních stříkaček o objemu 20 ml, jejichž ústí bylo uzavřeno pomocí síťky Uhelon. K semenům, nasypaným do injekční stříkačky, byla nejprve nasáta sterilní destilovaná voda (SDV), která byla posléze nahrazena 7 % desinfekčním roztokem chlornanu vápenatého. Doba bobtnání semen a působení roztoku chlornanu vápenatého pro desinfekci a narušení hydrofóbního povrchu je uvedena v tab.1. U třetího pokusu byl přibližně v polovině průběhu desinfekce vyměněn 7 % roztok Ca(Cl) 2 za nový. Po ukončení sterilizace byla semena opakovaně promyta v SDV a společně s malým množstvím poslední oplachovací vody byla vyseta na povrch připraveného média. Tabulka č. 1. Doba desinfekce semen v jednotlivých experimentech druh doba desinfekce [min] 1. výsev 2. výsev 3. výsev Anacamptis morio Dactylorhiza fuchsii Orchis mascula Platanthera bifolia Použitá média příprava, složení Pro kultivaci výsevů semen orchidejí bylo testováno několik variant agarem ztužených živných médií: Malmgren terrestrial orchid medium (PT068) s přídavkem kokosové vody (Malmgren, 1996), modifikované médium podle autorů Murashige a Skoog (Murashige a Skoog, 1962) s třetinovou koncentrací živin, přídavkem aktivního uhlí 35
36 a vitamínů dle Gamborga a komerčně dodávaná směs určená pro pěstování orchidejí - Orchimax od firmy Duchefa Biochemie B.V. Konkrétní složení použitých médií je uvedeno v Tabulce č. 2. Kultivační média byla připravena navážením hotové směsi nebo jednotlivých složek, ty byly následně rozmíchány v destilované vodě, pro ztužení byl přidán rozehřátý agar. Výsledné ph bylo upraveno pomocí KOH. Takto připravená média byla rozlita do kultivačních nádob a následně autoklávována nebo bylo médium nejprve autoklávováno a následně sterilně rozléváno ve flow-boxu do sterilních plastových Petriho misek. Všechny složky, včetně vitamínů a kokosové vody, byly přidávány před autoklávováním. 3.4 Podmínky kultivace V prvních dvou pokusech byly jako kultivační nádoby použili sklenice od dětské výživy o objemu 210 ml. Při třetím pokusu byly použity plastové Petriho misky o průměru 9 cm. Po výsevu byla semena uložena ve tmě při 21 C. Poté, co protokormy dosáhly 5. vývojové fáze, tedy když došlo ke zřetelné diferenciaci prýtu (nebo alespoň vrcholového pupenu) na jejich vrcholové části, byly pasážovány na shodné médium, na kterém byly doposud kultivovány. Následně byly kultury přesunuty ke kultivaci na světle v kultivačním regálu osvětleném zářivkami při fotoperiodě 8/12 hodin (světlo/tma) při ozářenosti 43 µmol.m -2.s -1 a teplotě 21 C. 3.5 Sledování vývoje kultury Všechny kultury byly po výsevu pravidelně kontrolovány kvůli výskytu infekcí. Případně infikované kultury byly buď zcela vyřazeny, nebo jsem se pokusila (v závislosti na rozsahu) infekci odstranit. Dále byl sledován postupný vývoj semen a protokormů. V průběhu kultivace byla prováděna fotodokumentace semen pomocí preparačního mikroskopu Leica S8 APO se zabudovaným LED osvětlením (Leica Microsystems, Německo) a digitálního fotoaparátu Olympus, model C5050Z (OLYMPUS OPTICAL CO., LTD, Japonsko). Fotodokumentace starších kultur, tedy protokormů v pokročilejších vývojových stádiích, byla prováděna v režimu makro, fotoaparátem Panasonic DMC-FZ50. Pokusy nebyly hodnoceny žádnou statistickou metodou. Jednalo se totiž o prvotní, ne příliš rozsáhlé testovací experimenty, jejiž cílem bylo vyzkoušení metod a stanovení vhodných 36
37 laboratorních postupů přípravy živných médií, desinfekcí a výsevů semen i pasážování kultur pro množení terestrických orchidejí in vitro. Získané poznatky by měly být využity při další práci, kdy bude již možné sledovat konkrétní cíle v experimentech většího rozsahu (např. porovnání různých médií, způsobů kultivace...), při kterých již bude možné využívat statistické metody pro jejich vyhodnocení. 37
38 38




39 Obr. 3. Obr. 4. Obr. 4.: semeno Anacamptis morio, 3 týdny kultivace; zv. 8x 2.,,klíčící fáze vývoje embrya měřítko představuje 0,5 mm Obr. 3.: semeno Dactylorhiza fuchsii, 3 týdny kultivace; zv. 8x 1.,,předklíčící fáze vývoje embrya měřítko představuje 0,5 mm Obr. 5. Obr. 6. Obr. 5.: Anacamptis morio, 21 týdnů kultivace ve tmě na modifikovaném MS médiu; 5.,,prýtová fáze vývoje embrya měřítko představuje 0,5 cm Obr. 6.: Anacamptis morio, 21 týdnů kultivace na modifikovaném MS médiu, stav po dvou dnech kultivace na světle patrné zezelenání prýtu; 5.,,prýtová fáze vývoje embrya měřítko představuje 0,5 cm
40 4 Výsledky 4.1 První pokus První pokus byl založen Po desinfekci byla semena vysévána na tři typy médií (viz. kapitola 3.3) připravených ve sklenicích od dětské výživy o objemu 210 ml. Již po 5 dnech kultivace bylo při mikroskopickém pozorování možné pozorovat nabobtnalá embrya v 1.,,předklíčící fázi vývoje embryí, u všech druhů (obr. 3). Po 3 týdnech kultivace, kdy byla provedena fotodokumentace, bylo možné mikroskopickým pozorováním u některých druhů najít 2.,,klíčící fázi vývoje embryí (obr. 4). Během kultivace docházelo však k rychlému vysychání kultur, což se projevilo zpomalením, či úplným zastavením vývoje semen. Tato situacie byla řešena aseptickým přidáváním SDV do kultur (1 2 ml) a překrytím uzávěrů kultivačních nádob hliníkovou fólií pro snížení výparu vody přes molitanový uzávěr. V prvním experimentu byl zaznamenán největší výskyt infekcí, což bylo způsobeno především nedostatečnou desinfekcí semen. První protokormy, embrya v 3.,,protokormovém a 4.,,rhizoidovém stádiu vývoje embryí byly patrné po 14 týdnech kultivace u druhu Anacamptis morio, Dactylorhiza fuchsii a Orchis mascula. Druh Anacamptis morio vytvářel největší protokormy, u Dactylorhiza fuchsii byl výskyt těchto útvarů nejhojnější. Nejdrobnější protokormy tvořil druh Orchis mascula, což je logické vzhledem k tomu, že tento druh měl i nejmenší rozměry semen z vybraných druhů. Nejhorších výsledků bylo dosaženo u druhu Platanthera bifolia, jehož semena velmi rychle vysychala a nepokračovala ve vývoji. Po 21 týdnech kultivace v některých kulturách pouze ojedinělá embrya dospěla do 5.,,prýtové vývojové fáze (obr.5), proto bylo provedeno jejich pasážování na čerstvé médium. K pasáži byly použity pouze protokormy z kultury Anacamptis morio a Orchis mascula, které byly pěstované na modifikovaném médiu Murashige Skoog. Tyto kultury byly přeneseny v kultivační místnosti na světlo, kde již po dvou dnech došlo k zezelenání prýtové části protokormů (obr. 6). Pozorování a posuzování vývoje kultur bylo velmi ztíženo kultivací v silnostěnných lahvích od dětské výživy nebylo tak možné jejich hodnocení pod preparačním mikroskopem ani postupná fotodokumentace. Mikroskopicky byl tedy sledován pouze vývoj u těch semen, 39
41 jež byla v infikovaných kulturách určených k likvidaci. Proto nebylo možné v tomto pokusu pozorovat postupně všechny vývojové fáze embryí. 40


42 Obr. 7. Obr. 8. Obr. 7.: Dactylorhiza fuchsii, 17 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; 5.,,prýtová fáze vývoje embrya; měřítko představuje 0,5 cm Obr. 8.: Dactylorhiza fuchsii, 17 týdnů kultivace na Malmgren Terrestrial Orchids medium, stav po dvou dnech kultivace na světle patrné zezelenání prýtu; 5.,,prýtová fáze vývoje embrya; měřítko představuje 0,5 cm
43 4.2 Druhý pokus Druhý pokus byl založen Doba desinfekce byla mírně prodloužena u semen Anacamptis morio a Orchis mascula. Po desinfekci byla semena stejným způsobem vysévána na tři různá média ve stejném typu kultivačních nádob, jako tomu bylo v prvním pokusu. Po několika dnech kultivace byla pozorována nabobtnalá embrya u všech testovaných druhů nacházela se v 1.,,předklíčící fázi vývoje embryí. Pokud jde o výskyt infekcí, byly zaznamenány v počátcích experimetu, ale jejich výskyt nebyl tak masivní, jako u prvního pokusu. Předpokládám, že toto zlepšení nastalo díky prodloužené době desinfekce. Problémy s rychlým vysycháním kultur byly však stejné jako u 1. pokusu, a proto byla i u tohoto 2. experimentu asepticky dle potřeby přidávána do kultur SDV a víčka kultivačních nádob byla překryta hliníkovou fólií. Po 9 týdnech kultivace bylo pozorovány první protokormy, tj. 3.,,protokormové stádium, především v kulturách druhu Anacamptis morio. Několik drobných protokormů vytvořila i semena druhu Dactylorhiza fuchsii. U semen druhu Orchis mascula byla embrya nabobtnalá. Avšak kultury semen Platanthera bifolia nevykazovaly opět žádné známky klíčení. V 11 týdnu kultivace semena druhů Anacamptis morio a Dactylorhiza fuchsii vytvořila protokormy 4.,,rhizoidové vývojové fáze. Vývoj zbývajících dvou druhů se bohužel zastavil, patrně došlo k přílišnému vyschnutí média a přeschnutí semen. Pokud hodnotíme vývoj kultur z hlediska použitých médií, bylo nejlepších výsledků dosaženo po výsevu semen na médium pro terestrické orchideje podle Malmgrena (Malmgren Terrestrial Orchid Medium) (Malmgren 1996). V 17. týdnu kultivace v jedné kultuře Anacamptis morio na Malmgren Terrestrial Orchid Medium dospěl protokorm do 5.,,prýtové vývojové fáze s prorostlým vrcholovým pupenem (obr.7). Byl tedy přepasážován na čerstvé médium a umístěn na fotoperiodu. Zde po dvou dnech došlo k zezelenání prýtové části (obr.8). Pozorování bylo opět, jako v prvním experimentu ztíženo umístěním kultur v silnostěnných kultivačních nádobách. 41






44 Obr. 9. Obr. 9.: semeno Platanthera bifolia, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 8x 1.,,předklíčící fáze vývoje embrya měřítko představuje 0,5 mm Obr. 11. Obr. 11.: semena Anacamptis morio, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 6,3x nalevo 2.,,klíčící,napravo 3.,,protokormová fáze vývoje embrya měřítko představuje 0,5 mm Obr. 13. Obr. 13.: protokorm Anacamptis morio, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 4x 4.,,rhizoidová fáze vývoje embrya měřítko představuje 0,5 mm Obr. 10. Obr. 10.: semena Dactylorhiza fuchsii, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 4x napravo 1.,,předklíčící, nalevo 2.,,klíčící fáze vývoje embrya měřítko představuje 0,5 mm Obr. 12. Obr. 12.: semeno Platanthera bifolia, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 8x praskání testy - přechod mezi 1. a 2. fází vývoje embrya měřítko představuje 0,5 mm Obr. 14. Obr. 14.: semena Dactylorhiza fuchsii, 6 týdnů kultivace ve tmě na Malmgren Terrestrial Orchids medium; zv. 4x 5.,,rhizoidová fáze vývoje embrya měřítko představuje 0,5 mm
Tkáňové kultury rostlin. Mikropropagace
 Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Oceněné rostlinné hnojivo!
 Oceněné rostlinné hnojivo! Powder Feeding Organická a minerální hnojiva byla v zemědělství používána po tisíce let, ovšem co se týká zemědělské techniky a zdrojů, tak ty se příliš nezměnily. Co ho dělá
Oceněné rostlinné hnojivo! Powder Feeding Organická a minerální hnojiva byla v zemědělství používána po tisíce let, ovšem co se týká zemědělské techniky a zdrojů, tak ty se příliš nezměnily. Co ho dělá
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty.
 o značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty. Podobné složení živých organismů Rostlina má celkově více cukrů Mezidruhové rozdíly u rostlin Živočichové
o značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty. Podobné složení živých organismů Rostlina má celkově více cukrů Mezidruhové rozdíly u rostlin Živočichové
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
Odborná škola výroby a služeb, Plzeň, Vejprnická 56, Plzeň. Číslo materiálu 19. Bc. Lenka Radová. Vytvořeno dne
 Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Krása TC. mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze.
 Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze. Kalusové kultury, protokormy. I když kalusové kultury nejsou metodou běžnou při rozmnožování
Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze. Kalusové kultury, protokormy. I když kalusové kultury nejsou metodou běžnou při rozmnožování
Vladimír Vinter
 Embryo (zárodek) Vývoj embrya (embryogeneze) trvá různě dlouhou dobu (např. u pšenice 20-25 dnů). U některých rostlin jsou embrya zcela nediferencovaná, např. u orchidejí. Zygota je výrazně polární buňka
Embryo (zárodek) Vývoj embrya (embryogeneze) trvá různě dlouhou dobu (např. u pšenice 20-25 dnů). U některých rostlin jsou embrya zcela nediferencovaná, např. u orchidejí. Zygota je výrazně polární buňka
OBSAH 1 ÚVOD... 7. 1.1 Výrobek a materiál... 7 1.2 Přehled a klasifikace materiálů pro výrobu... 8 2 ZDROJE DŘEVA... 13
 OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
Půdní úrodnost, výživa a hnojení
 Půdní úrodnost, výživa a hnojení Faktory ovlivňující růst a vývoj rostlin Přírodní faktory ovlivňující růst a vývoj rostlin významně ovlivňují úspěch či neúspěch budoucí rostlinné produkce. Ovlivňují se
Půdní úrodnost, výživa a hnojení Faktory ovlivňující růst a vývoj rostlin Přírodní faktory ovlivňující růst a vývoj rostlin významně ovlivňují úspěch či neúspěch budoucí rostlinné produkce. Ovlivňují se
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Obecná charakteristika hub
 Fyziologie hub Prvá část: Charakteristiku hub na základě výživy Ekologická charakteristika výživy hub Chemické zdroje výživy hub Druhá část Fyziologie růstu a rozmnožování Způsoby stanovení růstu, způsoby
Fyziologie hub Prvá část: Charakteristiku hub na základě výživy Ekologická charakteristika výživy hub Chemické zdroje výživy hub Druhá část Fyziologie růstu a rozmnožování Způsoby stanovení růstu, způsoby
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
SACHARIDY FOTOSYNTÉZA: SAHARIDY JSOU ORGANICKÉ SLOUČENINY SLOŽENÉ Z VÁZANÝCH ATOMŮ UHLÍKU, VODÍKU A KYSLÍKU.
 SACHARIDY SAHARIDY JSOU ORGANICKÉ SLOUČENINY SLOŽENÉ Z VÁZANÝCH ATOMŮ UHLÍKU, VODÍKU A KYSLÍKU. JSOU TO HYDROXYSLOUČENINY, PROTOŽE VŠECHNY OBSAHUJÍ NĚKOLIK HYDROXYLOVÝCH SKUPIN -OH. Sacharidy dělíme na
SACHARIDY SAHARIDY JSOU ORGANICKÉ SLOUČENINY SLOŽENÉ Z VÁZANÝCH ATOMŮ UHLÍKU, VODÍKU A KYSLÍKU. JSOU TO HYDROXYSLOUČENINY, PROTOŽE VŠECHNY OBSAHUJÍ NĚKOLIK HYDROXYLOVÝCH SKUPIN -OH. Sacharidy dělíme na
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
MTI Cvičení č. 2 Pasážování buněk / Jana Horáková
 MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
Ukázka knihy z internetového knihkupectví www.kosmas.cz
 Ukázka knihy z internetového knihkupectví www.kosmas.cz U k á z k a k n i h y z i n t e r n e t o v é h o k n i h k u p e c t v í w w w. k o s m a s. c z, U I D : K O S 1 8 0 0 1 1 U k á z k a k n i h
Ukázka knihy z internetového knihkupectví www.kosmas.cz U k á z k a k n i h y z i n t e r n e t o v é h o k n i h k u p e c t v í w w w. k o s m a s. c z, U I D : K O S 1 8 0 0 1 1 U k á z k a k n i h
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje. 26.2.2010 Mgr.
 Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje 26.2.2010 Mgr. Petra Siřínková ABIOTICKÉ PODMÍNKY ŽIVOTA SLUNEČNÍ ZÁŘENÍ TEPLO VZDUCH VODA PŮDA SLUNEČNÍ
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje 26.2.2010 Mgr. Petra Siřínková ABIOTICKÉ PODMÍNKY ŽIVOTA SLUNEČNÍ ZÁŘENÍ TEPLO VZDUCH VODA PŮDA SLUNEČNÍ
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ
 PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ LIGNOSULFONÁTY Lignin představuje heterogenní amorfní polymer potřebný pro pevnost a tuhost dřevnatých buněčných stěn rostlin. Po celulóze je to druhá nejrozšířenější látka
PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ LIGNOSULFONÁTY Lignin představuje heterogenní amorfní polymer potřebný pro pevnost a tuhost dřevnatých buněčných stěn rostlin. Po celulóze je to druhá nejrozšířenější látka
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU
 Opatření děkana Přírodovědecké fakulty Masarykovy univerzity č. 12 / 2018 Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU (ve znění účinném od 15.12.2018)
Opatření děkana Přírodovědecké fakulty Masarykovy univerzity č. 12 / 2018 Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU (ve znění účinném od 15.12.2018)
Opatření děkana č. 1/2012 Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU
 Opatření děkana č. 1/2012 Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU Bakalářské, diplomové a rigorózní práce odevzdávané k obhajobě na Přírodovědecké
Opatření děkana č. 1/2012 Pokyny pro vypracování bakalářských, diplomových a rigorózních prací na Přírodovědecké fakultě MU Bakalářské, diplomové a rigorózní práce odevzdávané k obhajobě na Přírodovědecké
Možnosti reintrodukce domácích orchidejí
 Možnosti reintrodukce domácích orchidejí Botanická zahrada Hl. m. Prahy, Trojská 800/196, 171 00 Praha 7 Troja Katedra experimentální biologie rostlin, PřF UK, Viničná 5, 128 44 Praha 2 jan.ponert@botanicka.cz
Možnosti reintrodukce domácích orchidejí Botanická zahrada Hl. m. Prahy, Trojská 800/196, 171 00 Praha 7 Troja Katedra experimentální biologie rostlin, PřF UK, Viničná 5, 128 44 Praha 2 jan.ponert@botanicka.cz
INDUKCE TVORBY MIKROHLÍZEK BRAMBORU IN VITRO
 Úloha č. 14 Indukce tvorby hlíz in vitro na jednonodálních segmentech rostlin bramboru -1 - INDUKCE TVORBY MIKROHLÍZEK BRAMBORU IN VITRO TUBERIZACE BRAMBOR (Solanum tuberosum) Tuberizace je morfogenetický
Úloha č. 14 Indukce tvorby hlíz in vitro na jednonodálních segmentech rostlin bramboru -1 - INDUKCE TVORBY MIKROHLÍZEK BRAMBORU IN VITRO TUBERIZACE BRAMBOR (Solanum tuberosum) Tuberizace je morfogenetický
Fotosyntéza (2/34) = fotosyntetická asimilace
 = fotosyntetická asimilace") Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
EU peníze středním školám
 EU peníze středním školám Název projektu Registrační číslo projektu Název aktivity Název vzdělávacího materiálu Číslo vzdělávacího materiálu Jméno autora Název školy Moderní škola CZ.1.07/1.5.00/34.0526
EU peníze středním školám Název projektu Registrační číslo projektu Název aktivity Název vzdělávacího materiálu Číslo vzdělávacího materiálu Jméno autora Název školy Moderní škola CZ.1.07/1.5.00/34.0526
Obsah 5. Obsah. Úvod... 9
 Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
Biochemie, Makroživiny. Chemie, 1.KŠPA
 Biochemie, Makroživiny Chemie, 1.KŠPA Biochemie Obor zabývající se procesy uvnitř organismů a procesy související s organismy O co se biochemici snaží Pochopit, jak funguje život Pochopit, jak fungují
Biochemie, Makroživiny Chemie, 1.KŠPA Biochemie Obor zabývající se procesy uvnitř organismů a procesy související s organismy O co se biochemici snaží Pochopit, jak funguje život Pochopit, jak fungují
Složky potravy a vitamíny
 Složky potravy a vitamíny Potrava musí být pestrá a vyvážená. Měla by obsahovat: základní živiny cukry (60%), tuky (25%) a bílkoviny (15%) vodu, minerální látky, vitaminy. Metabolismus: souhrn chemických
Složky potravy a vitamíny Potrava musí být pestrá a vyvážená. Měla by obsahovat: základní živiny cukry (60%), tuky (25%) a bílkoviny (15%) vodu, minerální látky, vitaminy. Metabolismus: souhrn chemických
Růst a vývoj rostlin
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
2) Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.
 Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.") Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Vliv teploty. Mezofilní mik. Termoofilní mik. Psychrofilní mik. 0 C 10 C 20 C 30 C 40 C 50 C 60 C 70 C teplota
 Vliv teploty Jeden z hlavních faktorů ovlivňující téměř všechny životní pochody mik. Každý mik. žije v určitém teplotním rozmezí je dáno: Minimální teplotou nejnižší teplota, při které mik. roste a množí
Vliv teploty Jeden z hlavních faktorů ovlivňující téměř všechny životní pochody mik. Každý mik. žije v určitém teplotním rozmezí je dáno: Minimální teplotou nejnižší teplota, při které mik. roste a množí
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka
 primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
RŮST A VÝVOJ ROSTLIN. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1
 RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
Střední škola gastronomie, hotelnictví a lesnictví Bzenec, náměstí Svobody 318. Profilová část maturitní zkoušky
 Střední škola gastronomie, hotelnictví a lesnictví Bzenec, náměstí Svobody 318 Obor: 29 42 M / 01 Analýza potravin Třída: AN4A Období: jaro 2013 Profilová část maturitní zkoušky 1. Povinná volitelná zkouška
Střední škola gastronomie, hotelnictví a lesnictví Bzenec, náměstí Svobody 318 Obor: 29 42 M / 01 Analýza potravin Třída: AN4A Období: jaro 2013 Profilová část maturitní zkoušky 1. Povinná volitelná zkouška
Zdroje informací.
 Zahradní orchideje Zpracováno podle: Pavel Sekerka; Jan Ponert; Jiří Obdržálek (2006): Orchideje na zahradě, dá se sehnat jako e-kniha Plejonky a další chladnomilné orchideje; Pavel Sekerka a kol.; Grada;
Zahradní orchideje Zpracováno podle: Pavel Sekerka; Jan Ponert; Jiří Obdržálek (2006): Orchideje na zahradě, dá se sehnat jako e-kniha Plejonky a další chladnomilné orchideje; Pavel Sekerka a kol.; Grada;
Úvod do biochemie. Vypracoval: RNDr. Milan Zimpl, Ph.D.
 Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
ZEOTECH MINI prémiové trávníkové hnojivo
 ZEOTECH MINI prémiové trávníkové hnojivo Postupný a vyrovnaný růst Excelentní barva pro Vaše greeny Vyvinuto profesionály pro profesionály Není to jednoduché vždy udržet greeny pěkné, zdravé a hratelné,
ZEOTECH MINI prémiové trávníkové hnojivo Postupný a vyrovnaný růst Excelentní barva pro Vaše greeny Vyvinuto profesionály pro profesionály Není to jednoduché vždy udržet greeny pěkné, zdravé a hratelné,
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
DUM VY_52_INOVACE_12CH33
 Základní škola Kaplice, Školní 226 DUM VY_52_INOVACE_12CH33 autor: Kristýna Anna Rolníková období vytvoření: říjen 2011 duben 2012 ročník, pro který je vytvořen: 9. vzdělávací oblast: vzdělávací obor:
Základní škola Kaplice, Školní 226 DUM VY_52_INOVACE_12CH33 autor: Kristýna Anna Rolníková období vytvoření: říjen 2011 duben 2012 ročník, pro který je vytvořen: 9. vzdělávací oblast: vzdělávací obor:
Oligobiogenní prvky bývají běžnou součástí organismů, ale v těle jich již podstatně méně (do 1%) než prvků makrobiogenních.
 než prvků makrobiogenních.") 1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2
 Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2 Obsah tématu: 1) Vzdušný obal země 2) Složení vzduchu 3) Tlak vzduchu 4) Vítr 5) Voda 1) VZDUŠNÝ OBAL ZEMĚ Vzdušný obal Země.. je směs
Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2 Obsah tématu: 1) Vzdušný obal země 2) Složení vzduchu 3) Tlak vzduchu 4) Vítr 5) Voda 1) VZDUŠNÝ OBAL ZEMĚ Vzdušný obal Země.. je směs
Obchodní akademie a Jazyková škola s právem státní jazykové zkoušky Písek
 Obchodní akademie a Jazyková škola s právem státní jazykové zkoušky Písek Pracovní list DUMu v rámci projektu Evropské peníze pro Obchodní akademii Písek", reg. č. CZ.1.07/1.5.00/34.0301, Číslo a název
Obchodní akademie a Jazyková škola s právem státní jazykové zkoušky Písek Pracovní list DUMu v rámci projektu Evropské peníze pro Obchodní akademii Písek", reg. č. CZ.1.07/1.5.00/34.0301, Číslo a název
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Střední škola obchodu, řemesel a služeb Žamberk. Výukový materiál. zpracovaný v rámci projektu. EU Peníze SŠ
 Střední škola obchodu, řemesel a služeb Žamberk Výukový materiál zpracovaný v rámci projektu EU Peníze SŠ Registrační číslo projektu: CZ.1.07/1.5.00/34.0130 Šablona: III/2 Ověřeno ve výuce dne: 19. 10.
Střední škola obchodu, řemesel a služeb Žamberk Výukový materiál zpracovaný v rámci projektu EU Peníze SŠ Registrační číslo projektu: CZ.1.07/1.5.00/34.0130 Šablona: III/2 Ověřeno ve výuce dne: 19. 10.
sladká symfonie cukrů
 CO JE STROM? poznej své bratrance Stromy jsou rostliny, které se naučily dosahovat vysokého vzrůstu. Slouží jim k tomu samonosný, dřevěný kmen, rok za rokem sílící. Stejně jako vše živé na Zemi pocházejí
CO JE STROM? poznej své bratrance Stromy jsou rostliny, které se naučily dosahovat vysokého vzrůstu. Slouží jim k tomu samonosný, dřevěný kmen, rok za rokem sílící. Stejně jako vše živé na Zemi pocházejí
Protimrazová ochrana rostlin
 Protimrazová ochrana rostlin Denní variabilita teploty Každý den představuje sám o sobě jedinečnou vegetační sezónu Denní teplota Sluneční záření Vyzářená energiedlouhovlnná radiace Východ slunce Západ
Protimrazová ochrana rostlin Denní variabilita teploty Každý den představuje sám o sobě jedinečnou vegetační sezónu Denní teplota Sluneční záření Vyzářená energiedlouhovlnná radiace Východ slunce Západ
Nutriční aspekty konzumace mléčných výrobků
 Nutriční aspekty konzumace mléčných výrobků Prof. MVDr. Lenka VORLOVÁ, Ph.D. a kolektiv FVHE VFU Brno Zlín, 2012 Mléčné výrobky mají excelentní postavení mezi výrobky živočišného původu - vyšší biologická
Nutriční aspekty konzumace mléčných výrobků Prof. MVDr. Lenka VORLOVÁ, Ph.D. a kolektiv FVHE VFU Brno Zlín, 2012 Mléčné výrobky mají excelentní postavení mezi výrobky živočišného původu - vyšší biologická
Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.)
") Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.) Botanická charakteristika: ČESNEK KUCHYŇSKÝ (ALLIUM SATIVUM L.) Pravlastí je Džungarsko (severní Čína) v Střední Asii,
Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.) Botanická charakteristika: ČESNEK KUCHYŇSKÝ (ALLIUM SATIVUM L.) Pravlastí je Džungarsko (severní Čína) v Střední Asii,
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Živé organismy. Člověk Ostatní živočichové Rostliny Houby
 Živá příroda Člověk Živé organismy Člověk Ostatní živočichové Rostliny Houby Znaky života rostlin Rostliny: Dýchají Přijímají vodu a živiny Vylučují odpadní látky Rozmnožují se Rostou Vyvíjejí se Rostliny
Živá příroda Člověk Živé organismy Člověk Ostatní živočichové Rostliny Houby Znaky života rostlin Rostliny: Dýchají Přijímají vodu a živiny Vylučují odpadní látky Rozmnožují se Rostou Vyvíjejí se Rostliny
MIKROORGANISMY EDÍ. Ústav inženýrstv. enýrství ochrany ŽP FT UTB ve Zlíně
 MIKROORGANISMY A OCHRANA ŽIVOTNÍHO PROSTŘED EDÍ Ústav inženýrstv enýrství ochrany ŽP FT UTB ve Zlíně Důvody využívání mikroorganismů v procesech ochrany životního prostřed edí jsou prakticky všudypřítomné
MIKROORGANISMY A OCHRANA ŽIVOTNÍHO PROSTŘED EDÍ Ústav inženýrstv enýrství ochrany ŽP FT UTB ve Zlíně Důvody využívání mikroorganismů v procesech ochrany životního prostřed edí jsou prakticky všudypřítomné
Krása TC. mé zkušenosti s technikami mikropropagace v obrazech.
 Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Kultivace stonkových segmentů. Meristémové kultury. Kultivace vzrostných vrcholků. Kultivace rostlin z axilárních pupenů. Hegemonie vrcholu.
Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Kultivace stonkových segmentů. Meristémové kultury. Kultivace vzrostných vrcholků. Kultivace rostlin z axilárních pupenů. Hegemonie vrcholu.
RNDr. David Novotný Ph.D Biologické přípravky na bázi bakterií
 RNDr. David Novotný Ph.D. +420 777 664 756 Biologické přípravky na bázi bakterií Zabýváme se přípravky na bázi symbiotických půdních bakterií PGPR (Plant grow-promoting rhizobacteria) Fungujeme na českém
RNDr. David Novotný Ph.D. +420 777 664 756 Biologické přípravky na bázi bakterií Zabýváme se přípravky na bázi symbiotických půdních bakterií PGPR (Plant grow-promoting rhizobacteria) Fungujeme na českém
Orchideoidní mykorhiza
 Metody studia kořenových systémů Orchideoidní mykorhiza není žádný idylický vztah.. 14. 3. 2012 Tamara Těšitelová Orchidaceae mladá čeleď ~ 100mil let druhově extrémně bohatá: přes 20000 druhů velká bohatost
Metody studia kořenových systémů Orchideoidní mykorhiza není žádný idylický vztah.. 14. 3. 2012 Tamara Těšitelová Orchidaceae mladá čeleď ~ 100mil let druhově extrémně bohatá: přes 20000 druhů velká bohatost
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
žák zvládne základní informace o glukóze, sacharóze a škrobu, pochopí základní schéma fotosyntézy Spec. vzdělávací potřeby Stupeň a typ vzdělávání
 Subjekt Speciální ZŠ a MŠ Adresa U Červeného kostela 110, 415 01 TEPLICE Číslo op. programu CZ. 1. 07 Název op. programu OP Vzdělávání pro konkurenceschopnost Číslo výzvy 21 Název výzvy Žádost o fin. podporu
Subjekt Speciální ZŠ a MŠ Adresa U Červeného kostela 110, 415 01 TEPLICE Číslo op. programu CZ. 1. 07 Název op. programu OP Vzdělávání pro konkurenceschopnost Číslo výzvy 21 Název výzvy Žádost o fin. podporu
Moderní odborníci na výživu věří, že plody jujuby jsou bohaté na vitamíny a mají vysokou nutriční a medicínskou hodnotu.
 Datlový sirup TIENS Datlový sirup Čínští lékaři věří, že Jujuba Udržuje lidi fit Doplňuje energii Posiluje játra, slezinu a žaludek Vyživuje krev Zklidňuje nervy Moderní odborníci na výživu věří, že plody
Datlový sirup TIENS Datlový sirup Čínští lékaři věří, že Jujuba Udržuje lidi fit Doplňuje energii Posiluje játra, slezinu a žaludek Vyživuje krev Zklidňuje nervy Moderní odborníci na výživu věří, že plody
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP. Šárka Poláková
 STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
Pokuste se vlastními slovy o definici pojmu Sacharidy: ? Které sacharidy označujeme jako cukry?
 Pokuste se vlastními slovy o definici pojmu Sacharidy: Sacharidy jsou polyhydroxyderiváty karbonylových sloučenin (aldehydů nebo ketonů).? Které sacharidy označujeme jako cukry? Jako tzv. cukry označujeme
Pokuste se vlastními slovy o definici pojmu Sacharidy: Sacharidy jsou polyhydroxyderiváty karbonylových sloučenin (aldehydů nebo ketonů).? Které sacharidy označujeme jako cukry? Jako tzv. cukry označujeme
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
VESMÍR. za počátek vesmíru považujeme velký třesk před 13,7 miliardami let. dochází k obrovskému uvolnění energie, která se rozpíná
 VESMÍR za počátek vesmíru považujeme velký třesk před 13,7 miliardami let dochází k obrovskému uvolnění energie, která se rozpíná vznikají první atomy, jako první se tvoří atomy vodíku HVĚZDY první hvězdy
VESMÍR za počátek vesmíru považujeme velký třesk před 13,7 miliardami let dochází k obrovskému uvolnění energie, která se rozpíná vznikají první atomy, jako první se tvoří atomy vodíku HVĚZDY první hvězdy
NÁVRH METODIKY PRO TESTOVÁNÍ ODOLNOSTI STAVEBNÍCH HMOT PROTI NAPADENÍ PLÍSNĚMI
 NÁVRH METODIKY PRO TESTOVÁNÍ ODOLNOSTI STAVEBNÍCH HMOT PROTI NAPADENÍ PLÍSNĚMI PROPOSAL OF METHODOLOGY FOR TESTING RESISTANCE OF BUILDING MATERIALS AGAINST MOLD INFESTATION Ilona Kukletová, Ivana Chromková
NÁVRH METODIKY PRO TESTOVÁNÍ ODOLNOSTI STAVEBNÍCH HMOT PROTI NAPADENÍ PLÍSNĚMI PROPOSAL OF METHODOLOGY FOR TESTING RESISTANCE OF BUILDING MATERIALS AGAINST MOLD INFESTATION Ilona Kukletová, Ivana Chromková
prokaryotní Znaky prokaryoty
 prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
Název školy: Střední odborná škola stavební Karlovy Vary Sabinovo náměstí 16, 360 09 Karlovy Vary Autor: Hana Turoňová Název materiálu:
 Název školy: Střední odborná škola stavební Karlovy Vary Sabinovo náměstí 16, 360 09 Karlovy Vary Autor: Hana Turoňová Název materiálu: VY_32_INOVACE_04_BUŇKA 1_P1-2 Číslo projektu: CZ 1.07/1.5.00/34.1077
Název školy: Střední odborná škola stavební Karlovy Vary Sabinovo náměstí 16, 360 09 Karlovy Vary Autor: Hana Turoňová Název materiálu: VY_32_INOVACE_04_BUŇKA 1_P1-2 Číslo projektu: CZ 1.07/1.5.00/34.1077
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie
 synekologie b) autekologie c) demekologie") 1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
a) pevná fáze půdy jíl, humusové částice vážou na svém povrchu živiny v podobě iontů
 pevná fáze půdy jíl, humusové částice vážou na svém povrchu živiny v podobě iontů") Otázka: Minerální výživa rostlin Předmět: Biologie Přidal(a): teriiiiis MINERÁLNÍ VÝŽIVA ROSTLIN - zahrnuje procesy příjmu, vedení a využití minerálních živin - nezbytná pro život rostlin Jednobuněčné
Otázka: Minerální výživa rostlin Předmět: Biologie Přidal(a): teriiiiis MINERÁLNÍ VÝŽIVA ROSTLIN - zahrnuje procesy příjmu, vedení a využití minerálních živin - nezbytná pro život rostlin Jednobuněčné
Rozmanitost podmínek života na Zemi Podnebné pásy
 Podnebné pásy Tropický mezi obratníky - Vhlké vnitřní tropy: - bez střídání ročních období - silné srážky, -průměrná roční teplota nad 20 C -Vnější tropy: -přechod k subtropům - období dešťů a období sucha
Podnebné pásy Tropický mezi obratníky - Vhlké vnitřní tropy: - bez střídání ročních období - silné srážky, -průměrná roční teplota nad 20 C -Vnější tropy: -přechod k subtropům - období dešťů a období sucha
Nádherné akvarijní rostliny díky perfektně nastavené péči. Systémová péče o rostliny
 Nádherné akvarijní rostliny díky perfektně nastavené péči Systémová péče o rostliny sera péče o rostliny Vyvážený a kompletní systém pro individuální a účinnou péči o rostliny sera florena pro vodní rostliny,
Nádherné akvarijní rostliny díky perfektně nastavené péči Systémová péče o rostliny sera péče o rostliny Vyvážený a kompletní systém pro individuální a účinnou péči o rostliny sera florena pro vodní rostliny,
BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA
 MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA GEOGRAFICKÝ ÚSTAV BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA Diplomová práce Jan Kučera Vedoucí práce: Mgr. Daniel Seidenglanz, Ph.D. Brno 2013 Bibliografický záznam Autor:
MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA GEOGRAFICKÝ ÚSTAV BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA Diplomová práce Jan Kučera Vedoucí práce: Mgr. Daniel Seidenglanz, Ph.D. Brno 2013 Bibliografický záznam Autor:
6. Tzv. holocenní klimatické optimum s maximálním rozvojem lesa bylo typické pro a) preboreál b) atlantik c) subrecent
 preboreál b) atlantik c) subrecent") 1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
Tabulace učebního plánu. Obecná chemie. Vzdělávací obsah pro vyučovací předmět : Ročník: 1.ročník a kvinta
 Tabulace učebního plánu Vzdělávací obsah pro vyučovací předmět : CHEMIE Ročník: 1.ročník a kvinta Obecná Bezpečnost práce Názvosloví anorganických sloučenin Zná pravidla bezpečnosti práce a dodržuje je.
Tabulace učebního plánu Vzdělávací obsah pro vyučovací předmět : CHEMIE Ročník: 1.ročník a kvinta Obecná Bezpečnost práce Názvosloví anorganických sloučenin Zná pravidla bezpečnosti práce a dodržuje je.
VY_52_Inovace_242 Vzdělávací oblast: Člověk a příroda Vzdělávací obor: Chemie Ročník: 8, 9 Projekt EU peníze školám Operačního programu Vzdělávání
 Sacharidy VY_52_Inovace_242 Vzdělávací oblast: Člověk a příroda Vzdělávací obor: Chemie Ročník: 8, 9 Projekt EU peníze školám Operačního programu Vzdělávání pro konkurenceschopnost Sacharidy název z řeckého
Sacharidy VY_52_Inovace_242 Vzdělávací oblast: Člověk a příroda Vzdělávací obor: Chemie Ročník: 8, 9 Projekt EU peníze školám Operačního programu Vzdělávání pro konkurenceschopnost Sacharidy název z řeckého
ROSTLINNÁ BUŇKA A JEJÍ ČÁSTI
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Stimulátor biologické aktivity rhizosféry Dostaňte z kořenů to nejlepší. explorer 20
 Stimulátor biologické aktivity rhizosféry Dostaňte z kořenů to nejlepší 20 Jednejte již při výsevu Úspěšná sklizeň kukuřice se připravuje již od prvních stádií růstu. Cílem je co nejdříve umožnit efektivní
Stimulátor biologické aktivity rhizosféry Dostaňte z kořenů to nejlepší 20 Jednejte již při výsevu Úspěšná sklizeň kukuřice se připravuje již od prvních stádií růstu. Cílem je co nejdříve umožnit efektivní
Vyjádření fotosyntézy základními rovnicemi
 FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
5. Bioreaktory. Schematicky jsou jednotlivé typy bioreaktorů znázorněny na obr. 5.1. Nejpoužívanějšími bioreaktory jsou míchací tanky.
 5. Bioreaktory Bioreaktor (fermentor) je nejdůležitější částí výrobní linky biotechnologického procesu. Jde o nádobu různého objemu, ve které probíhá biologický proces. Dochází zde k růstu buněk a tvorbě
5. Bioreaktory Bioreaktor (fermentor) je nejdůležitější částí výrobní linky biotechnologického procesu. Jde o nádobu různého objemu, ve které probíhá biologický proces. Dochází zde k růstu buněk a tvorbě
Dlouhodobě uvolňovaný dusík v různém zastoupení. Najdete zde hnojiva se zastoupením 30 50 % dlouhodobě
 Produktová řada hnojiv FENIX přináší vašemu trávníku přesně takové živiny, které potřebuje. Při vývoji hnojiv FENIX jsme se zaměřili na požadavky a poznatky, které přicházejí od profesionálních trávníkářů.
Produktová řada hnojiv FENIX přináší vašemu trávníku přesně takové živiny, které potřebuje. Při vývoji hnojiv FENIX jsme se zaměřili na požadavky a poznatky, které přicházejí od profesionálních trávníkářů.
Číslo a název projektu Číslo a název šablony
 Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Bc. Hana Potyšová květen 2012
 Bc. Hana Potyšová květen 2012 Zkouška životnosti životnost osiva je souhrn vlastností určujících aktivitu osiva v různých podmínkách prostředí vyrovnanost klíčení a růstu klíčků schopnost vzcházet za nepříznivých
Bc. Hana Potyšová květen 2012 Zkouška životnosti životnost osiva je souhrn vlastností určujících aktivitu osiva v různých podmínkách prostředí vyrovnanost klíčení a růstu klíčků schopnost vzcházet za nepříznivých
UŽITNÝ VZOR CZ U1 A01G 24/60 ( ) C09K 17/00 ( ) A01N 65/03 ( ) A01N 63/02 ( ) (11) Číslo dokumentu:
 C09K 17/00 ( ) A01N 65/03 ( ) A01N 63/02 ( ) (11) Číslo dokumentu:") UŽITNÝ VZOR (19) ČESKÁ REPUBLIKA (21) Číslo přihlášky: 2018-34978 (22) Přihlášeno: 21.05.2018 (47) Zapsáno: 17.07.2018 (11) Číslo dokumentu: 31 921 (13) Druh dokumentu: U1 (51) Int. Cl.: A01G 24/60 (2018.01)
UŽITNÝ VZOR (19) ČESKÁ REPUBLIKA (21) Číslo přihlášky: 2018-34978 (22) Přihlášeno: 21.05.2018 (47) Zapsáno: 17.07.2018 (11) Číslo dokumentu: 31 921 (13) Druh dokumentu: U1 (51) Int. Cl.: A01G 24/60 (2018.01)
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
BRASSAVOLA BRASAVOLA BRASSAVOLA BRASAVOLA
 BRASSAVOLA BRASAVOLA BRASSAVOLA BRASAVOLA DRUHY VHODNÉ PRO PĚSTOVÁNÍ V DOMÁCÍCH PODMÍNKÁCH 50 ORCHIDEJE VHODNÉ DO DOMÁCÍCH PODMÍNEK 51 DRUHY VHODNÉ PRO PĚSTOVÁNÍ V DOMÁCÍCH PODMÍNKÁCH Druhy a kultivary
BRASSAVOLA BRASAVOLA BRASSAVOLA BRASAVOLA DRUHY VHODNÉ PRO PĚSTOVÁNÍ V DOMÁCÍCH PODMÍNKÁCH 50 ORCHIDEJE VHODNÉ DO DOMÁCÍCH PODMÍNEK 51 DRUHY VHODNÉ PRO PĚSTOVÁNÍ V DOMÁCÍCH PODMÍNKÁCH Druhy a kultivary
Buňka cytologie. Buňka. Autor: Katka www.nasprtej.cz Téma: buňka stavba Ročník: 1.
 Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Ústřední kontrolní a zkušební ústav zemědělský Oddělení půdy a lesnictví
 Ústřední a zkušební ústav zemědělský Oddělení půdy a lesnictví Analýza a vyhodnocení účinnosti leteckého vápnění, provedeného v roce 2008 v Krušných horách v okolí Horního Jiřetína, po pěti letech od data
Ústřední a zkušební ústav zemědělský Oddělení půdy a lesnictví Analýza a vyhodnocení účinnosti leteckého vápnění, provedeného v roce 2008 v Krušných horách v okolí Horního Jiřetína, po pěti letech od data
Využití hydroabsorbentů při školkařské produkci i výsadbě dřevin
 Využití hydroabsorbentů při školkařské produkci i výsadbě dřevin Petr Salaš Zahradnická fakulta Mendelovy univerzity v Brně se sídlem v Lednici Školkařské dny 2018; Lísek, 11.1.2018 Co je hydroabsorbent?
Využití hydroabsorbentů při školkařské produkci i výsadbě dřevin Petr Salaš Zahradnická fakulta Mendelovy univerzity v Brně se sídlem v Lednici Školkařské dny 2018; Lísek, 11.1.2018 Co je hydroabsorbent?
2.01 Aerobní/anaerobní reakce aneb kvasinky v akci. Projekt Trojlístek
 2. Vlastnosti látek a chemické reakce 2.01 Aerobní/anaerobní reakce aneb kvasinky v akci. Projekt úroveň 1 2 3 1. Předmět výuky Metodika je určena pro vzdělávací obsah vzdělávacího předmětu Chemie. Chemie
2. Vlastnosti látek a chemické reakce 2.01 Aerobní/anaerobní reakce aneb kvasinky v akci. Projekt úroveň 1 2 3 1. Předmět výuky Metodika je určena pro vzdělávací obsah vzdělávacího předmětu Chemie. Chemie
OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.)
") Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
J. Kubíček FSI Brno 2018
 J. Kubíček FSI Brno 2018 Fosfátování je povrchová úprava, kdy se na povrch povlakovaného kovu vylučují nerozpustné fosforečnany. Povlak vzniká reakcí iontů z pracovní lázně s ionty rozpuštěnými z povrchu
J. Kubíček FSI Brno 2018 Fosfátování je povrchová úprava, kdy se na povrch povlakovaného kovu vylučují nerozpustné fosforečnany. Povlak vzniká reakcí iontů z pracovní lázně s ionty rozpuštěnými z povrchu