Tvarová dynamika křemičitých struktur u modelových populací chrysomonád (Synurophyceae)
|
|
|
- Vladislav Jelínek
- před 6 lety
- Počet zobrazení:
Transkript
1 Universita Karlova v Praze Přírodovědecká fakulta Katedra botaniky Bakalářská práce Tvarová dynamika křemičitých struktur u modelových populací chrysomonád (Synurophyceae) Kateřina Janatková 2009 Školitelka: RNDr. Yvonne Němcová, PhD.
2 Abstrakt: Předkládaná bakalářská ráce shrnuje poznatky o taxonomii a geografiie křemitých chrysomonád a jejich distribuci podél gradientů životního prostředí. Křemité chrysomonády jsou především sladkovodní bičíkovci, jejich buňky jsou pokryty křemitými šupinami. Šupiny jsou druhově specifické. Druhy se vyskytují v úzkou škále ekologických podmínek a jsou tak cennými biondikátory nedávného i historického prostředí. Literární rešerše se týká distribuce chrysomonád podél gradientu ph, vodivosti, obsahu živin a teplotních gradientů.je diskutována i skutečnost, že některé druhy mohou způsobovat zápach a zakalení vod. Práce se zmiňuje i o typech rozšíření.praktická část ze zabývá odběry z lokalit na Borkovických blatech a v Novohradských horách. Spolu s kosmopolitními a široce rozšířenými druhy, byla nalezena Mallomonas conspersa. Tento druh dosud byl nalezen pouze v Japonsku a na Nového Zélandu. Nálezy druhů Synura mollispina a M. pumilio var. munda byly rovněž prvními pro Českou republiku. Klíčová slova: chrysomonády s křemičitými šupinami, Synurophyceae, taxonomie, bioindikace,, biogeografie, Mallomonas conspersa 2
3 Abstract: Shape dynamics of silicate structures in experimental populations of chrysophytes (Synurophyceae) Presented Bc. Thesis summarises temporal knowledge on taxonomy and biogeography of silica-scaled chrysophytes and their distribution along environmental gradients. Silicascaled chrysophytes are mainly freshwater flagellates, their cells are covered with imbricated silica scales. The scales are species-specific and thus the taxa with the narrow range of ecological conditions represent a valuable biondicators of recent and historical environments. The literature survey concerns the distribution of chrysophytes along ph, conductivity, nutrient content and temperature gradients. The species causing water tastes and odors are discussed. Types of distribution are underlined. The practical part surveys silica-scaled chrysophytes from Borkovická Blata peat-bog and Novohradské Hory ponds. Along with cosmopolitan and widely distributed species, Mallomonas conspersa was found, This taxon has so far been reported only from Japan and New Zealand. Moreover Synura mollispina and M. pumilio var. munda were recorded for the first time from the territory of the Czech Republic. K ey words : Silica scaled chrysophytes, Synurophyceae, taxonomy, bioindication, geografical distribution, Mallomonas conspersa 3
4 Poděkování Především bych chtěla mnohokrát poděkovat zejména své školitelce RNDr.Yvonne Němcové, PhD. za její nekonečnou trpělivost, kterou projevovala celou dobu při vedení mé práce. Dále pak RNDr. Jiřímu Neustupovi, PhD za to, že mě k této práci přivedl. Děkuji samozřejmě také všem ostatním kamarádům a kolegům, kteří mi během mé práce vycházeli vstříc a neváhali mi s čímkoli pomoci. 4
5 1.Úvod Teoretická část Chrysomonády s křemitými šupinami Taxonomické zařazení Obecná charakteristika Ekologie Distribuce druhů chrysomonád podél gradientu prostředí Výskyt podél gradientu ph Výskyt podél gradientu konduktivity Výskyt podél gradientu trofie Výskyt podél gradientu teploty Bioindikace Chrysomonády jako bioindikátory Využití v paleolimnologii Biogeografie Geograficky podmíněné rozšíření Neutrální teorie ubikvitního rozšíření Praktická část Úvod Odběrové lokality Odběry vzorků a měření Výsledky a diskuze Závěr Použitá literatura:
6 1.Úvod Pojmem chrysomonády s křemitými šupinami označujeme solitérní či koloniální bičíkovce, jejichž zástupci se vyskytují téměř výhradně ve sladkých vodách. První popisy těchto chrysomonád pocházejí z pozorování preparátů ve světelném mikroskopu (LM) již v polovině 19.století (EHRENBERG; PERTY ). V 50. letech 19. století, po vynálezu elektronového mikroskopu, dochází k rozvoji studia křemičitých struktur, k objevům nových druhů chrysomonád a k objasnění celkového vzhledu buněk. První snímky šupin ze skenovacího elektronového mikroskopu (SEM) byly publikovány v roce 1955 (ASMUND 1955, FOTT 1955) a další využívání této metody, spolu s metodou transmisního elektronového mikr oskopu (TEM), přineslo další detailní informace o uspořádání křemitých struktur na povrchu buněk a o povrchových strukturách šupin v závislosti na specifitě pro jednotlivé druhy (KRISTIANSEN 1971). Dnes můžeme za použití EM determinovat jednotlivé druhy chrysomonád podle jejich druhově charakteristických šupin. Vzhledem k tomu, že některé druhy mají úzké rozpětí ideálních podmínek výskytu, jsou tak ideální pro využití jako bioindikátory pro posuzování jak stavu současného, tak i minulého, a to díky dlouhodobému zachovávání šupin i cyst v sedimentu (SIVER 1995, SMOL 1995). Cílem této práce je poskytnout úvod ke studiu ekologie a biogeografie křemičitých chrysomonád. Zaměřuje se na jejich výskyt v rámci gradientů prostředí, na jejich využití jako bioindikátorů a také se zabývá geografickým rozšířením chrysomonád. Cílem praktické části bylo seznámit se s metodikou odběru a určováním druhů. 2. Teoretická část 2.1.Chrysomonády s křemitými šupinami Taxonomické zařazení Jako chrysomonády s křemitými šupinami jsou označovány koloniální či jednotlivě žijící bičíkovci zařazení do dvou tříd v oddělení Heterokontophyta. Jde o třídy Chrysophyceae, a Synurophyceae ANDERSEN (1987). Třída Synurophyceae, kterou se zabývá tato práce, 6
7 obsahuje rody Mallomonas, Synura, Chrysodidymus a Tessellaria (KRISTIANSEN & PREISIG 2001). V dnešní době se tradiční taxonomická rozdělení dostávají do rozporu s novými molekulárními daty, a je tomu tak i u této skupiny organismů. Ta byla, jako jiné skupiny, zprvu klasifikována podle morfologických znaků různých vegetačních stadií. Jedním z prvních systémů navrhl roku 1957 BOURELLY (ANDERSEN 2007). V tomto systému byly všechny rody šupinatých chrysomonád řazeny do čeledi Synuraceae. Během dalších výzkumů bylo tento systém několikrát rozčleněn a pozměněn. V souvislosti s molekulárními a biochemickými daty se ukazuje, že není možné využívat v taxonomii pouze morfologické znaky vegetativních stadií, neboť tato stadia mají značnou variabilitu v závislosti na prostředí a samotné znaky jsou mnohdy obtížně identifikovatelné (např.druhý bičík je v některých případech minimálně vyvinutý a nepostřehnutelný). Morfologická a molekulární data spolu nesouhlasí při kvalifikaci vyšších skupin (podtřídy, řády), ale v případě nižších taxonomických skupin (čeleď, rod) spolu korelují (ANDERSEN 2007). Třídy Chrysophyceae a Synurophyceae jsou od sebe tradičně oddělovány na základě biochemických a morfologických rozdílů, a nejnovější molekulární studie toto rozdělení podporují, i když zatím jen slabě. Naproti tomu stojí molekulární data o rodu Tesselaria, který byl dříve řazen do třídy Synurophyceae, ale molekulární data ukazují, že spadá přesně mezi dvě třídy Synurophyceae a Chrysophyceae. Na úrovni rodů Mallomonas a Synura skupiny Synurophyceae nebylo přesné rozdělení potvrzeno, ale oproti tomu jsou diskutovány nálezy šupin těchto rodů 47 mil. let staré, které jsou morfologií srovnatelné s tvarem šupin jak je známe dnes. Je tedy otázka, zda toto zachování přesné morfologie nedoprovází i zachování genu na úrovni druhů (ANDERSEN 2007). Na úrovni druhů dochází k novým objevům, kdy se ukazuje, že v některých případech může jít o druhové komplexy (např. Synura petersenii) s více či méně odlišenou morfologií šupin (KYNČLOVÁ, et al. 2008, subm.) Obecná charakteristika Třídy Chrysophyceae a Synurophyceae (podle tradičního členění) (ANDERSEN 1987) mají, jako ostatní zástupci oddělení Heterokontophyta, některé společné znaky jako jsou dva bičíky, jeden delší, se dvěma řadami mastigonem, a druhý krátký, pokrytý pouze jemným vlášením. Dále jsou to chloroplasty se 4 obalnými membránami, thylakoidy tvořící skupiny po třech a zásobní látkou je chrysolaminaran. Přítomny jsou pigmenty chlorofyl a, c, β-karoten, 7


 a poté jsou exocytozou dopraveny na povrch buňky. Tvar a velikost šupin se podle umístění na buňce liší.")

8 xantofyly a zlatavou barvu typickou pro tyto dvě třídy dodává dominantní fukoxanthin (KALINA & VÁŇA 2005). Přesto, že se v řadě dalších znaků liší, je pro obě tyto třídy typické vytváření křemičitých cyst s pórem uzavřeným zátkou. Některé morfologické a biochemické znaky jsou pro třídy Chrysophycae a Synurophyceae odlišné a jsou základem pro jejich taxonomické oddělení (ANDERSEN 2007). Pro zástupce třídy Synurophyceae, kterými se tato práce zabývá, jsou typické buňky kryté pancířem šupin, tvořených z polymerovaného oxidu křemičitého. Buňky kromě šupin vytvářejí ze stejného materiálu i další struktury, jako ostny či cysty. Všechny tyto struktury mají endogenní původ. Šupiny vznikají v SDV (silica deposition vesicle) a poté jsou exocytozou dopraveny na povrch buňky. Tvar a velikost šupin se podle umístění na buňce liší. Všechny šupiny mají bazální desku s případnou sekundární vrstvou a všechny nesou druhově specifickou ornamentaci. Rod Synura tvoří kolonie několika buněk spojených posteriorními konci. Šupiny jsou tvarem a dorsální ornamentací druhově specifické a tvarově odlišné podle umístění na buňce. Vzájemně na sebe nasedají a zcela tak obepínají buňku. Na distálním konci se pak může vytvářet ostének, nebo kýl, s původem ve stejném SDV jako samotná šupina. (KRISTIANS 2005). EN Synura echinulata Synura petersenii d 1 Obr.1. Morfologie šupin rodu Synura: a.osten (scale), kýl (S. (S. petersenii), b. žebra (ribs), c. přehnutý proximální konec, d. póry na bazální desce, d1:kýl Buňky rodu Mallomonas žijí jednotlivě a k některým jejich šupinám je připevněn osten. Jejich uspořádání na buňce je stejné, jako je tomu u rodu Synura. Šupiny jsou většinou bipolární, na spodní straně hladké. Na vrchní straně je pak sekundární vrstva tvořící žebra, papily či jiné struktury. Přes obě vrstvy mohou procházet drobné póry. Nejčastější šupiny bývají tzv. tripartitní, rozdělené na tři části. Povrch šupiny je dělen žebrem ve tvaru V (V-rib) na štít a postraní okraje. Na těchto plochách se pak vytvářejí druhově specifické ornamentace. 8
.")
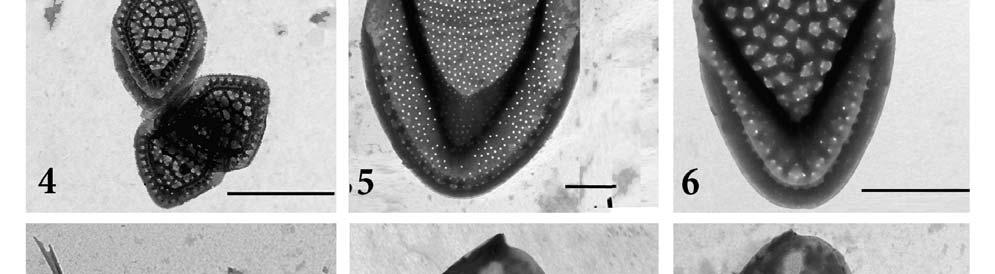
9 Na distálním konci se nachází dóm (dome), na který se přikládá osten. Osten má původ v samostatném SDV, kde vznikne stočením původně plochého útvaru, a k šupině se připojí až dodatečně na povrchu buňky (KRISTIANSEN 1986). Ostny jsou stejně jako šupiny důležité pro taxonomii. Obr.2.1. celá buňka: 1.šupiny s dómem, 2. šupiny bez dómu, 3. posteriorní šupiny, 4. ostny; 2.2. tripartitní šupina rodu Mallomonas: 1. posteriorní přehnutý kraj, 2. V-rib, 3. štít, 4. dóm, 5. postraní žebra. Převzato ze SIVER 2003 Obr. 3. Zobrazení nejčastějších typů ostnů. Převzato ze SIVER (1991) 9

10 Cysty (Obr.4) jsou stejně jako šupiny a ostny tvořeny endogenně, splýváním SDV a váčků Golgiho aparátu. Poté co se obal cysty uvnitř buněk dostatečně zpevní, přesune se volným otvorem dovnitř celý obsah protoplastu bičíkovce. Cysta se nakonec uzavře v místě póru zátkou, která je pevná, ale není křemičitá. Povrch cyst je opět druhově charakteristický hladký, různě strukturovaný, či s osténky (SIVER 1991). Fakt, že morfologie cyst je druhově charakteristická a že se dobře dochované cysty nacházejí i v sedimentech, je předurčuje k roli vhodných bioindikátorů v paleolimnologických výzkumech, při studiích historických jezerních sedimentů. (SMOL 1995). Obr. 4. Cysty šupinatých chrysomonád, s viditelnými póry a různou povrchovou ornamentací (papily nebo ostny). Ze SIVER (2003) 2.2. Ekologie Chrysomonády se jako striktní autotrofové vyskytují v planktonu sladkých vod, až na některé výjimky v rodu Paraphysomonas, kde najdeme i druhy planktonu salinních vod. Vzhledem ke značnému globálnímu rozšíření zástupců těchto skupin chrysomonád je obtížné přesně vymezit jejich ekologické nároky (SIVER 1995). Chrysomonády byly dříve označovány jako organismy chladných, oligotrofních vod s maximem rozvoje výskytu na jaře (KRISTIANSEN 1975). Toto je však zobecnění, které nelze aplikovat na všechny zástupce, pouze na skupinu jako celek, protože podmínky výskytu se druh od druhu liší, a některé druhy jako takové mají úzkou ekologickou valenci (SIVER 1995). Velké množství druhů chrysomonád můžeme tedy najít i ve fytoplanktonu eutrofních vod (KRISTIANSEN 1985). Nejvyšší počet druhů se pak nachází v lokalitách s mírným a nižším ph, nízkou alkalinitou, 10
11 konduktivitou, mírným obsahem živin a rozpuštěných huminových kyselin (SIVER 1995). Zástupce nacházíme v jezerech, tůních, rašeliništích, slepých ramenech řek i přímo v tekoucích vodách (říčky či vysychavé potůčky) (např.veselá & PICHRTOVÁ, in press) Výskyt chrysomonád na lokalitách je nejvíce ovlivněn ph, alkalinitou a konduktivitou vody. Tyto parametry ovlivňují druhové složení na lokalitě, neboť výskyt jednotlivých druhů je specifický podle tolerance rozpětí gradientu prostředí (SIVER 1991). Teplota vody pak ovlivňuje sezónní přítomnost a abudanci populací chrysomonád ve fytoplanktonu (SIV ER & HAMER 1992). Někteří zástupci rodů Synura a Mallomonas mohou ve vodních nádržích vytvářet vodní květy a způsobovat nepříjemný zápach vody připomínající rybinu (WHIPPLE et al. 1984), nebo negativně ovlivnit další organismy, ať už konkurencí o světlo a živiny nebo svými metabolity. Nejčastěji tyto zákaly způsobují M. acaroides, M. crassisquama, M. caudata (KRISTIANSEN 1971) a druhy rodu Synura (NICHOLLS & GERATH 1985). Tendence tvořit zákaly mají populace žijící v severnějších oligotrofních jezerech (SANDGREN 1988) a to zejména z důvodu, že při nižších teplotách a hodnotách osvětlení rychleji jejich populace rostou rychleji než populace jiných druhů ve přítomných ve fytoplanktonu. Problémy se zákaly je potřeba řešit zejména při výskytu ve vodohospodářských nádržích s pitnou vodou, kdy mohou kromě vytváření zápachu narušovat svou přítomností filtrační a sedimentační procesy při konečných úpravách pitné vody (NICHOLLS ET GERATH 1985). 3. Distribuce druhů chrysomonád podél gradientu prostředí Data o druhovém složení a proměnných prostředí lze využít k diskriminaci lokalit a zhodnocení relativního významu jednotlivých proměnných pro distribuci druhů ve zkoumaných lokalitách. Mezi nejpoužívanější statistické metody zpracování dat patří přímé a nepřímé ordinační metody. Mnohorozměrné statistické metody se používají také k odvození inferenčních modelů. V souboru zkoumaných jezer se naměří požadované hodnoty prostředí a určí se zastoupení jednotlivých druhů chrysomonád z vrchní vrstvy sedimentu. Tyto dva soubory dat (druhové složení a hodnoty prostředí) se pak použijí ke stanovení optimálních hodnot pro výskyt jednotlivých druhů a šíři tolerance podél gradientu zkoumané hodnoty prostředí. Použitím 11
12 inferenčních modelů na soubory dat z hlubších sedimentů lze pak vysledovat historické změny na lokalitě (SIVER 2003) Výskyt podél gradientu ph Hodnota ph je nejvýznamnějším parametrem pro výskyt jednotlivých druhů, což bylo opakovaně prokázáno v mnoha studiích (SIVER 1987, SIVER 1988, SIVER & HAMER 1989, SIVER & SMOL 1993). Druhy s úzkým rozpětím tolerance ph rozdělil Husted (1939) do několika skupin podle výskytu podél gradientu ph (SIVER 1991): 1. acidobiontní omezený výskyt pouze na kyselých vodách s ph < acidofilní nejčastější výskyt při ph < indiferentní převážně v hodnotách ph kolem alkalifilní maximum výskytu při ph > alkalibiontní výskyt omezen na vody s ph > 7.0 Tab1. Zobrazuje průměr naměřených hodnot ph, konduktivity (množství ropzuštěných solí) a teploty popisující prostředí, kde byly druh nalezeny. Spolu s následným zařazením taxonů do skupin, které definoval Husted (1939); ACB- acidobiontní, ACF acidofilní, IND- indiferentní, ALKF- alkalifilní, ALKB- alkalibiontní; Chrysosphaerella C.; Synura S.;Spiniferomonas Sp.; Mallomonas M. Převzato ze SIVER & HAMER (1989). 12

.")
13 Podle zařazení nalezených druhů do těchto skupin lze říci jaká hodnota ph se na stanovišti vyskytovala či vyskytuje. Acidobiontní skupinu nejčastěji reprezentují Mallomonas acaroides var. muskokana, M. canina, M. hindonii, M.pugio, M. paludosa, M. hamata, Synura echinulata, a S. sphagnicola. Acidofilní skupinu tvoří například M. punctifera, M. akromonas, M. galeiformis, M. crassisquama, M. caudata, S. spinosa, S. uvella,. Na druhé straně gradientu kolem hodnot ph 7.0 se vyskytují M. corymbosa, M. tonsurata, M. alpina. Druhy M. acaroides var. acaroides a M. pseudocoronata se zřídkakdy vyskytují v prostředí s ph klesajícím pod 7.0 (SIVER 1988). Hodnoty ph na rozšíření chrysomonád nepůsobí přímo, ale pravděpodobně limitují chrysomonády množstvím dostupného anorganického uhlíku. Ten se ve vodě vyskytuje jako - rozpuštěný CO ále disociuje na HCO 3 a CO 2-2, který reaguje s vodou za vzniku H 2 CO 3 a d 3. Zastoupení těchto forem uhlíku udává výslednou hodnotu ph. Chrysomonády nejsou pravděpodobně schopné využívat pro fotosyntézu ion HCO3 -, který se vyskytuje při vysokých hodnotách ph. Potřebují zejména CO 2, který je dostupný naopak při nižším ph (BHATTI & COLMAN 2008). To je jednak přímo limituje a nepřímo pak nejsou při nižších hladinách CO2 schopny konkurovat jiným planktonním druhům. Může to být jeden z důvodů jejich jarního maxima výskytu a naopak případného letního minima (SAXBY-ROUEN 1998). Klesající hodnoty fotosyntetické aktivity u dvou druhů rodu Synura lze vidět na Obr.5. Obr.5. Hodnoty O 2 (mev. l - 1)spotřebovávaného při fotosyntetické aktivitě v závislosti na okolním ph vody. Převzato z práce BHATTI & COLMAN (2008). 13


 Nejčastěji se chrysomonády vyskytují při nižších hladinách konduktivity a jejich množství se stoupajícími")

14 3.2. Výskyt podél gradientu konduktivity Výskyt chrysomonád je dále ovlivňován konduktivitou. Konduktivita udává míru vodivosti vody dané množstvím rozpuštěných solí. Na změnách hodnot konduktivity lze zpětně sledovat změny v okolí vod jako např. osidlování lokalit, stavby silnic, přítomnost továren atd. a jejich vliv na zvyšování rozpuštěných živin ve vodě. Hodnoty konduktivity jsou v takových případech průkaznější, neboť takovéto změny se na hodnotách ph nemusejí vůbec projevit (SIVER 1993). Základními kationy ovlivňujícími vodivost jsou Na +, Ca ++, Mg ++ a K +. Poměry koncentrací těchto iontů se v závislosti na hodnotě konduktivity liší, ale obecně platí, že při hodnotách >90 µs. cm -1 dominuje rozpuštěný Na + (viz Tab. 2). Vliv rozpuštěného H + na hodnoty konduktivity je minimální a zanedbatelný (SIVER 1995) Nejčastěji se chrysomonády vyskytují při nižších hladinách konduktivity a jejich množství se stoupajícími hodnotami klesá. V hodnotách nad 200 µs. cm -1 se pak téměř nevyskytují (viz Tab.2.) (SUSHIL et al 1999). Tab.2. Hodnoty konc. rozpuštěných kationů v závislosti na hodnotě specifické konduktivity. Ze SIVER Podobně jako u hodnot ph lze jednotlivé druhy seřadit do skupin podle hodnot konduktivity, kterou tyto druhy preferují. Skupina druhů zahrnující např. Mallomonas acaroides var. muskokana, M. punctifera, M. galeiformis a Synura sphagnicola reprezentuje spodní hladinu gradientu a málokdy je najdeme ve vodách s hodnotami nad 75 µs. cm -1. Uprostřed gradientu, přibližně v rozmezí hodnot 75 µs. cm -1 až 125 µs. cm -1, pak najdeme 14
15 například druhy M. hamata, M. crassisquama, M. caudata, M. akromonas, Synura echinulata, S. spinosa. V poslední skupině se nachází druhy které se vyskytují ve vyšších gradientech a prakticky se nevyskytují v prostředí s konduktivitou klesající pod 50 µs. cm -1. Jsou to třeba M. corymbosa, M. tonsurata, M. alpina. M. acaroides var. Acaroides. Ve skupinách jsou jako příklady uváděny druhy, které jsou ve studiích nejčastěji nacházeny, a které mají úzkou ekologickou valenci Výskyt podél gradientu trofie Dalším signifikantním parametrem výskytu je pro chrysomonády hladina trofie. Aby bylo možno postihnout gradient trofie, byly vytvořeny indexy trofie (trophic score) (SIVER & MA RSCIANO 1996). Podle množství celkového fosforu, dusíku a chlorofylu-a a hloubku viditelnosti Secchiho desky (prostupnost světla vodním sloupcem) se lokality řadí do 6 skupin (FRINK & NOVELL 1984): 1.Oligotrofní největší množství výskytu chrysomonád 2. Mírně mesotrofní 3. Mesotrofní 4. Silně mesotrofní 5. Eutrofní 6. Silně eutrofní pouze vzácné nálezy chrysomonád Tab.4. Tabulka jednotlivých trofických skupin a jejich charakteristikpodle 4 proměnných. Ze SIVER & MARSCIANO


16 Druhy lze pak podle jejich tolerance rozmístit podél gradientu trofie. Mallomonas paludosa, M. retrosa, M. acaroides var. muskonana a Synura lapponica jsou typičtí zástupci oligotrofních vod. Naproti tomu M. heterospina, M. acaroides var. acaroides a Synura curtispina se vyskytují při vysokých hladinách trofie. M. tonsurata a M. corymbosa se vyskytují ve vodách s různou mírou eutrofizace, ale nikdy je nenajdeme ve vodách oligotrofních. Naopak M. elongata nebo S. uvella jsou schopny tolerovat jak lehce eutrofní vody, tak je nacházíme i na oligotrofních lokalitách. Rozložení druhů podél gradientu spolu s jejich optimy výskytu znázorňuje Obr.6. Obr.6. Výskyt druhů podle jejich tolerance k hodnotám trofie spolu s jejich optimálními hodnotami pro výskyt. Převzato ze SIVER & MARSCIANO (1996). V silně eutrofizovaných vodách se chrysomonády téměř vůbec nevyskytují. Existuje několik teorií pokoušejících se tento fakt vysvětlit. Může to být z důvodu zvýšené konkurence s ostatními druhy fytoplanktonu, např. se sinicemi, které dokážou efektivněji využít větší množství dostupných živin. (SANDGREN 1988). Dále může jít o souhru více faktorů, které 16
17 v eutrofních nádržích panují. Jedná se spíše o nádrže mělké, tudíž s vyššími teplotami vody, což je v rozporu s ideálními podmínkami výskytu pro chrysomonády, jak bylo uvedeno dříve (metalimnium oligotrofních chladnějších jezer). V mělkých nádržích nemohou chrysomonády tak snadno uniknout predátorům do větších metalimnických hloubek, mělké nádrže se také snadněji promíchávají a ohřívají a to pro chrysomonády s optimem růstu spíše v nižších teplotách není ideální (WATSON & MCCAULY 2005). Rovněž bývá pravidlem, že vysoce eutrofní lokality mají i vyšší ph a konduktivitu, a optima výskytu chrysomonád na těchto dvou parametrech opět leží spíše ve středních a nižších hodnotách (SUSHIL et al.) Výskyt podél gradientu teploty Teplota hraje v rozšíření chrysomonád také svou roli. Má vliv zejména na sezónní vývoj početnosti populací na dané lokalitě (SIVER 1995, SIVER & HAMER 1992). Druhy se nacházejí ve velkém rozmezí teplot, ale jejich optima se opět druhově liší (ROIJACKERS & KESSELS 1986): 1. Eurytermní druhy se širokým spektrem tolerance teplot 2. Stenotermní druhy s úzkým rozpětím tolerance 3. Oligotermní optima v nižších teplotách 4. Polytermní optima ve vyšších teplotách Teplotní optima jsou pro druhy důležitá z hlediska rychlosti jejich růstu. Bylo prokázáno, že největší rychlosti dosahují spíše při nižších teplotách. Pokud jsou jejich podmínky růstu ideální, mohou tak přerůst a konkurenčně vytlačit jiné druhy fytoplanktonu a stát se dominantami nejen na jaře (KRISTINASEN 1975), ale i v průběhu celého roku (SIVER & HAMER 1992). Záleží také na tom, jaké druhy se spolu na lokalitě nacházejí (Obr.7). Pokud máme vedle sebe dva druhy se stejnou valencí teplot, pak jejich společná optima mohou znásobit rychlost vývoje chrysomonád v celkovém fytoplanktonu (typ II), a naopak při snížení či zvýšení teploty za hranici jejich tolerance mohou zcela z fytoplanktonu vymizet (typ I a III). Pokud má jeden z druhů jiné optimum teploty než druhý, budou se chrysomonády s velkou pravděpodobností nacházet ve fytoplanktonu celoročně (typ IV) (pokud by jejich výskyt záv isel na teplotě) a to v podobě chladno- či teplomilnějšího druhu v závislosti na ročním období (NICHOLLS & GERATH 1985). 17

18 Obr.7. Různé typy sezónní distribuce chrysomonád ve fytoplanktonu. Typy distribucí jsou vysvětleny v textu. Převzato ze NICHOLLS & GERATH Bioindikace 4.1. Chrysomonády jako bioindikátory Bioindikátory jsou organismy, pomocí nichž je možné monitorovat vývoj stavů životního prostředí či ekosystému. Faktorem určujícím změny prostředí jsou pak změny abundance a druhového složení těchto organismů (KRISTIANSEN 2005). Chrysomonády, stejně jako jiné bioindikační organismy musejí splňovat několik podmínek. Bioindikátory musí být taxonomicky přesně definované, snadno identifikovatelné a jejich výskyt se musí vyznačovat úzkou ekologickou valencí podél gradientu sledovaného faktoru prostředí (KRISTIANSEN 1986). Mnoho druhů křemičitých chrysomonád tyto podmínky splňuje, a navíc se jejich cysty a šupiny, podle kterých je lze zpětně určit, dobře zachovávají v sedimentech Využití v paleolimnologii Dříve se v paleolimnologii používaly pouze křemičité schránky rozsivek, v posledních 15 letech se však začaly ve studiích používat i šupiny a cysty chrysomonád (SUSHIL et al. 1999). 18
 a k posouzení klimatických změn (SMOL 2005). Takovéto studie byly prováděny například na jezerech v Connecticutu (SIVER et al.")
19 Cysty jsou nejčastěji využívány ve studiích, kde se porovnávají dnešní stavy vod s podmínkami z preindustriální doby a v Evropě z doby před jejím osídlením (SIVER et al. 1999) a k posouzení klimatických změn (SMOL 2005). Takovéto studie byly prováděny například na jezerech v Connecticutu (SIVER et al. 1999; MARSCIANO & SIVER 1993), na jezerech v pohoří Adirondack v USA (SMOL et al. 1983; SIVER 1987) nebo v jezerech ve Vermontu a v New Hempshire (SIVER & LOTT 2000). Vzhledem k tomu, že chybí historická data o faktorech prostředí z minulých dob, využívají se křemité struktury, získané z vrtů do starších vrstev sedimentu, pro určení jednak typu změny, a jednak jakým faktorem byla tato změna způsobena zda přírodním vývinem ekosystému, nebo antropogenními vlivy (SIVER et al 1999). Například v Minnesotě byly v sedimentech z doby před 8,500 lety nalezeny struktury podobné cystám chrysomonád a to na místě, kde se věřilo že bývala travnatá prérie (SIVER 2003). Unikátním nálezem jsou šupiny z vrtu Girrafe Pipe z jezerního sedimentu v Kanadě. Ve vrtu hlubokém 163 m bylo nalezeno několika chrysofytních šupin. Nálezy byly z hloubek 95.6m a 127,6m. Oba nálezy jsou datované z doby středního Eocénu, tedy z doby asi před 47 mil. lety. Na základě znalostí ekologických nároků druhů chrysomonád se zdá, že zatímco druhy z hloubky 127,6 m pocházejí z jezera, v hloubce 95,6m se již nacházejí druhy z mělkého rašeliniště (SIVER & WOLF 2005). Obr.8. A, B: nálezy šupin z Eocénu srovnané s nálezy s dnešní doby (C, D) Ze SIVER & WOLF Struktury podobné cystám chrysomonád se našly i v mořských sedimentech z období druhohor (145 mil. 65 mi.l let). Vzhledem k tomu, že nálezy ze starších vrstev se 19

 či pro sledování klimatických změn (SIVER 1993).")
20 neobjevily, mohlo by to znamenat, že v tomto období započala silifikace u této skupiny řas (SIVER & WOLF 2005). Mnoho studií se rovněž zabývá studiem změn v posledních desetiletích v kvalitě vody, zejména v temperátních jezerech Severní Ameriky a Evropy.Chrysomonády se využívají jako indikátory změn v kyselých depozicích (CHARLES AND SMOL 1988, SIVER AN HAMER 1999, CUMMING et al 1991), při stanovení míry acidifikace způsobené např. nárůstem CO 2 z emisí (MARSCIANO & SIVER 1993, SIVER &LOTT 2000) či pro sledování klimatických změn (SIVER 1993). Například ze studie prováděné na jezerech v Connecticutu vyplývá, že v posledních 100 letech se hodnota ph zvedla v rozpětí 0,4-1,1 a hladina konduktivity se zvýšila přes 100µS. cm -1. Jako jedním z hlavních důvodů bylo vykácení značného množství lesního porostu v okolí těchto jezer(obr.9) a jejich následné obhospodařování (SIVER et al. 1999). Obr.9. Změny trofie a ph v závislosti na kácení okolních lesů. Přítomnost lesních ploch uvedena v %. Ze SIVER et al Biogeografie 5.1. Geograficky podmíněné rozšíření Podle KRISTIANSENA (2001) je rozšíření chrysomonád závislé na konkrétních podmínkách prostředí, na tom, jak se v minulosti měly možnost rozšiřovat ze svých stanovišť a na možnostech, které takovéto šíření zprostředkují. Podle této teorie jsou tedy druhy geograficky rozmístěny, ať už v klimatických zónách nebo v rámci kontinentů. Podle tohoto rozšíření se řadí do několika kategorií: 20
21 1. kosmopolitní druhy nalezené na všech kontinentech; např. M. tonsurata 2. široce rozšířené chybí na 1-2 kontinentech (často je tímto kontinentem Austrálie) 3. bipolární s výjimkou tropického pásu výskyt na obou polokoulích, např. M. alata 4. tropické- pouze v tropických oblastech 5. arktické a temperátní rozšíření pouze v temperátních či arktických zónách severní polokoule, např. M. hamata 6. rozptýlené pouze v rámci dvou nebo tří kontinentů, např. Synura mollispina 7. endemické výskyt omezen na jediný kontinent O možnostech rozšiřování mezi vzdálenějšími lokalitami se vedou spory. Bičíkatá stádia mají jen malou pravděpodobnost, že by se mohla bez vyschnutí dostat do větších vzdáleností. Přenášeny jsou tedy pravděpodobně křemičité stomatocysty. Jednou z možností je přenos cyst na nebo uvnitř těla ptáků, což by ale nevysvětlovalo celkové geografické rozšíření, nebo ť na mnoho míst, kde se chrysomonády vyskytují, nevedou žádné migrační trasy ptáků (WEE et. al. 1993). Na tato místa by se pak mohly stomatocysty dostat spolu se vzdušnými mikročásticemi unášenými větrem (KRISTIANSEN 1996). Nezanedbatelným faktorem pro šíření chrysomonád je pak člověk - převozem ryb nebo vodních rostlin z místa na místo, mezi kontinenty a jinak izolovanými ostrovy pak v zásobách sladké vody na lodích (KRISTIANSEN 2001). Posledním faktorem minulých desetiletí může být transport druhů na nedostatečně vymytých planktonních sítích či jiných pomůckách vědců cestujícími mezi více lokalitami (KRISTIANSEN 2007) Neutrální teorie ubikvitního rozšíření Naproti této teorii staví FINLEY & CLARKE tzv. neutrální teorii ubikvitního rozšíření druhů. Týká se všech mikroorganismů menších než 1mm a toho, že skupiny mikroorganismů jsou velice staré a tudíž již měli ve své historii dost času a možností na to, aby se globálně rozšířily. Tato teorie také předpokládá velkou lokální diversitu mikroorganismů, která se se zvětšující plochou mění jen málo (FENCHEL & FINLAY 2004). Pokud nebyly na některých lokalitách nalezeny, je to zapříčiněno nevhodnými podmínkami prostředí, které zde nedovolují druhu přežít. Rozšíření chrysomonád tak, jak je publikoval KRISTIANSEN (2001) není v rozporu s touto teorií kromě faktu, že druhy nacházející se v severní temperátní zóně se nevyskytují na jižní polokouli. Model ubikvitního rozšíření na vybraných zástupcích rodu Mallomonas a pravděpodobnost jejich výskytu na jižní polokouli testovali ve své práci ŘEZÁČOVÁ & 21
22 NEUSTUPA (2007). Srovnávali nálezy v jedné aluviální tůni, soustavě několika aluviálních tůní, nálezy v ČR a nálezy publikovanými ve světě. U druhů nalezených ve zkoumaných tůních pak byla v závislosti na počtu jejich předchozích nálezů vypočtena pravděpodobnost, s jakou by mohly být nalezeny i na polokouli jižní. U velké většiny druhů byla tato pravděpodobnost značně vysoká. Pokud tomu tak u některých druhů není, může to být zapříčiněno buď tím, že druh je z evolučního hlediska příliš mladý, než aby se stačil globálně rozšířit. Další možností je menší schopnost některých druhů se šířit, což může být způsobeno např. menší odolností jejich cyst. Také připadá v úvahu možnost, že šíření cyst je vázáno na určitou skupinu zvířat, která se sice v areálu dosavadního výskytu druhu pohybuje, ale nemůže tento areál opustit. Druhy chrysomonád tak nebudou mít možnost se mimo tento areál rozšířit (ŘEZÁČOVÁ & NEUSTUPA 2007). 22

23 6. Praktická část 6.1. Úvod Chrysomonády s křemitými šupinami jsou často významnou součástí fytoplanktonu sladkovodních jezer a nádrží. Cílem práce bylo zvládnutí metodiky odběru a následného zpracování vzorků ze vhodných přírodních lokalit, na nichž je možné předpokládat výskyt křemičitých chrysomonád, a následné vyhodnocení výskytu nalezených druhů v kontextu přírodních podmínek, ve kterých byly nalezeny zde a v dosud publikovaných studiích. Rašeliniště Borkovická Blata a rybníky v lesích Novohradských hor se zdály jako ideální lokality, na niž lze předpokládat hojnější výskyt chrysomonád Odběrové lokality Lokality Borkovická Blata a Novohradské hory jsou unikátními biotopy. Rašeliniště Borkovická Blata se nachází na jihu Čech poblíž Veselí nad Lužnicí. Rašeliniště staré více jak deset tisíc let bylo od poloviny dvacátých let využíváno pro těžbu rašeliny, která probíhala do nedávné doby. Vzorky byly sbírány z mělkých rašelinných jezírek vzniklých předchozí těžbou rašeliny. Nacházejí se na části rašeliniště o výměře asi 50 ha, které je od roku 1980 chráněné. Novohradské hory představují cennou nedotčenou přírodní oblast u česko rakouských hranic (Obr. 10.) Tato oblast byla v letech uzavřena pro veřejnost a v roce 2000 zde byla založena Přírodní rezervace Novohradské Hory. Rybníky, nebo lesní nádrže, z nichž byly vzorky odebírány, byly vystavěny v 18. století k plavení dřeva Obr. 10. Odběrová místa: LokalityBorkovická blata-č.1-3, Lokality Novohradské Hory č.4-5. Hlavní chemicko-fyzikální parametry na zkoumaných lokalitách je zobrazen v Tab.5., a na Obr. 11 jsou fotografie některých odběrových míst. 23







24 Obr. 11. Fotografie odběrových lokalit 1-Uhlíšťský rybník, 2-Huťský rybník 3-5 Borkovická Blata 5 24
25 Lokalita Datum odběru ph kond. Teplota NH3 P N (µs.c m-1) ( C) (µg/l) (µg/l) (µg/l) 1 Borkovická blata,tůň , ,7 2,16 0,31 2,4 2 Borkovická blata, tůň , ,61 0,23 6,9 3 Borkovická blata, tůň ,9 85 6,8 1,8 0,35 2,8 4 Novohradské hory, Uhlíšťský rybník , ,2 2,8 0,18 1,8 5 Novohradské hory, Huťský rybník , ,7 0,2 1,7 Tab.5. Hlavní chemicko-fyzikální parametry na zkoumaných lokalitách 6.3. Odběry vzorků a měření Vzorky byly odebírány planktonní sítí o velikosti ok 20 µm a zároveň s planktonem byla injekční stříkačkou odebírána i svrchní část sedimentu. Na rašeliništích a na březích rybníků s vodní vegetací byl z rostlin ždímán metafyton. Zvlášť pak byl odebrán vzorek nezčeřené vody pro pozdější spektofotometrická měření hodnot celkového fosforu, křemíku a dusíku. Všechny vzorky byly po příjezdu do laboratoře prohlédnuty pod světelným mikroskopem a ty, které obsahovaly šupinaté chrysomonády byly v příslušném množství nakapány na měděné síťky pokryté formvarovou vrstvou. Tyto síťky pak byly prohlíženy pod transmisním elektronový mikroskopem JEOL 1011 pro determinaci druhové složení těchto lokalit. Zbytek vzorku byl zafixován Lugolovým roztokem pro případné opětovné využití. Měření ph, konduktivity a teploty vody probíhalo přímo v lokalitě a to pomocí terénního měřícího přístroje Combo ph EC (Hanna Instruments). V laboratoři byly pak ze vzorků čisté nezčeřené vody měřeny další parametry vodní ho prostředí. Zjišťovalo se množství celkového dusíku, celkového fosforu za využití spektrofotometru DR/890 Colorimetr (Hach). Naměřené hodnoty jsou uvedeny v Tab Výsledky a diskuze Zkoumané lokality mají podobné hodnoty ph, ale hodnoty vodivosti se liší. Tyto hodnoty jsou vyšší v lokalitách z Borkovických Blat, kde se pohybují kolem 100 μs. cm -1. V rybnících Novohradských hor jsou hodnoty pouze okolo 50 μs. cm -1. Nicméně druhové složení chrysomonád se mezi těmito dvěma lokalitami příliš neliší. Druhy nalezené v jednotlivých 25
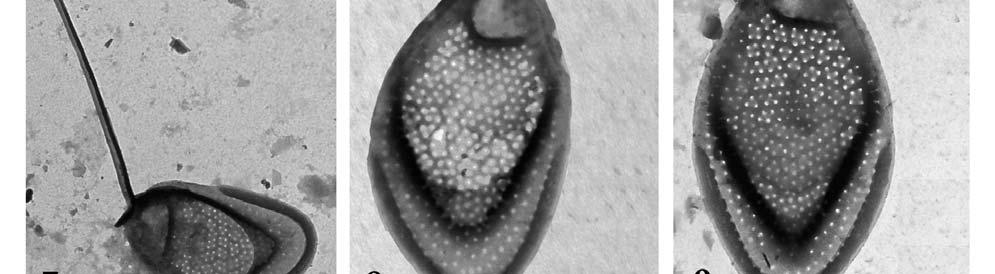
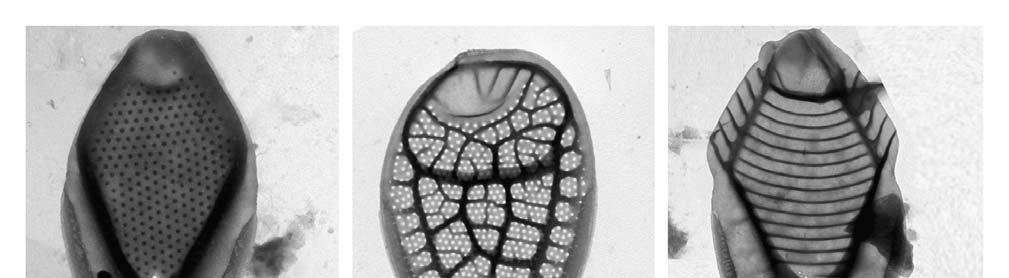
26 lokalitách jsou shrnutu v Tab.6. Bylo zaznamenáno celkem 22 druhů chrysomoná d z třídy Synurophyceae. Počet druhů na lezených na Borkovických Blatech ( Lokality 1-3) se pohyboval v rozmezí 2-12, v Novohradských Horách (lokality 4-5) byl nalezen shodný počet 11 druhů. Mezi nejčastější druhy patřila Synura petersenii, která byla nalezena ve vše ch 5 lokalitách. Dalšími hojnými druhy nalezenými na 4 lokalitách byly S. spinosa a Mallomonas papillosa. Většina druhů z těchto lokalit je ph indiferentní s širokým rozšířením. Byly nalezeny i některé vzácné druhy (S. mollispina, M. retifera, M. kalinae). Druh Mallomonas conspersa byl České republice nalezena poprvé. Fotografie šupin podobných šupinám M. conspersa byly dříve publikovány v Japonsku, jako neidentifikovaný druh (TAKAHASHI 1959) a později byl druh popsán z Nového Zélandu (DÜRRSCHMIDT 1986). Před tím byl také publikován záznam o jejím nálezu v Japonsku, ale nebyl doložen fotografiemi šupin (ITO1991). Šupiny toho druhu jsou velice podobné šupinám druhu M. retifera. Druhy by se měly lišit ve velikosti bu něk a šupin a (M. conspersa: velikost buňky 8-15 x 5-7 μm, velikost šupin: x 1,5-2 μm; M. retifera : velikost buňky x 5-9 μm, velikost šupin 3,2-3,7 x 2-2,4 μm), zde ale byly nalezeny šupiny obou druhů o shodné velikosti. Oba druhy byly nalezeny na téže lokalitě (viz Tab. 9). SIVER (1991) se na základě podobné morfologie domnívá, že tyto dva vzácné druhy mohou představovat jeden a týž druh. Nicméně je zde několik znaků jasně odlišujících šupiny těchto dvou druhů. Například šupiny M. retifera mají pod dómem umístěno příčné žebro, což je na první pohled odlišuje od šupin M. conspersa. Ty mají naopak z boku nad dóm vybíhající osten (zub) který šupiny M. retifera postrádají (viz. Tab.8., Obr 7-9). Zatímco druh M. retifera byl hlášen z Evropy (např. HARTMANN & STEINBERG 1989, PICHRTOVÁ et al. 2007), Jižní Ameriky (DÜRRSCHMIDT 1980, 1982) a východní Asie (ITO 1990), Mallomonas conspersa byl považován za mimořádně vzácný druh. Tento druh může být široce či kosmopolitně rozšířen, ovšem ve srovnání s ostatními druhy se vyskytuje jen vzácně. Mallmonas conspersa byl popsán jako druh z malé vodní tůně v mírně zalesněném rašeliništi, s ph 5,6 a vodivostí 90 μs / cm (DÜRRSCHMIDT 1986). Tyto přírodní podmínky odpovídají hodnotám a prostředí, kde byl M. conspersa nalezen v této studii (Tab.5 a 6.) Druh Mallomonas kalinae je široce rozšířen, ale vyskytuje se vzácně, nalézt ho lze především při vyšších teplotách.dürrschmidt & CROOME 1985, ho určili jako Mallomonas cf. rasilis, popsán byl až roku 2006 (ŘEZÁČOVÁ 2006). M. pumilio je druh s prokázaný výskytem převážně při teplotách nižších než 13 C, a s bipolárním rozšířením (SIVER 1991). M. schwemmelei je možno popsat jako zimní druh temperátních oblastí. Četné populace M. schwemmlei byly objeveny v Huťském rybníce (Lok. 5) pod ledem. 26
27 Druhy Lokality M. tonsurata TEILING X S ynura echinulata KORSHIKOV Fig. 3 X 5 X S. mollispina (J.B.PETERSEN & J.B.HANSEN) L.S.PÉTERFI & MOMEU Fig. 2 X S. petersenii KORSHIKOV Figs X X X X X S. sphagnicola (KORSHIKOV) KORSHIKOV X X S. spinosa KORSHIKOV X X X X S. uvella EHRENBERG Fig. 1 Mallomonas. acaroides PERTY emend. Fig. 15 X X X X IWANOFF M. calceolus D.E.BRADLEY X M. conspersa DÜRRSCHMIDT Figs 7-9 X M. costata DÜRRSCHMIDT Fig. 6 X M. crassisquama (ASMUND) FOTT X M. heterospina J.W.G.LUND Fig.11 X X M. kalinae ŘEZÁČOVÁ Fig. 10 X M. multis etigera DÜRRSCHMIDT X M. ouradion K.HARRIS & D.E.BRADLEY X M. papillosa K.Harris & D.E.BRADLEY e mend. K.HARRIS Figs X X X X M. pumilio var. pumilio K.HARRIS & D.E.BRADLEY emend. ASM., CRONB. & D ÜRRS. X X Mallomonas pumilio var. munda ASMUND, C RONBERG & DÜRRSCHMIDT Fig. 4 X X M retifera DÜRRSCHMIDT Fig. 7 X M. schwemmlei GLENK emend. GLENK & F OTT Figs X M. striata K.HARRIS & D.E.BRADLEY Fig. 12 X X X Tab. 6. Seznam druhů a jejich výskyt ve zkoumaných lokalitách 27



28 28



29 29
30 7. Závěr V dnešní době se studiem šupinatých chrysomonád zabývá stále více studií. Chrysomonády se ve stále větší míře úspěšně využívají jako bioindikátory přírodních podmínek, což je umožněno úzkou ekologickou valencí jednotlivých druhů a jejich snadnou identifikací. Mnoho studií to potvrzuje a dále se snaží určit co nejpřesnější rozpětí těchto podmínek a poznat ekologické nároky jednotlivých druhů co nejpřesněji. Zároveň je nezbytné posoudit přesnost morfologického určování druhů podle křemitých struktur, zda toto tradiční členění skutečně odpovídá fylogenetickému členění druhů. Proto se některé studie snaží potvrdit či vyvrátit toto tradičního morfologického členění srovnáváním morfologických dat spolu s daty molekulárními. Co se geografického rozšíření týče, mnoho druhů bylo nalezeno po celém světě a jejich rozšíření je tedy kosmopolitní, ale některé druhy se stále na mnoha místech nalézt nepodařilo, a tak zůstává otázkou, zda je jejich rozšíření ubikvitní, či geograficky podmíněné. V praktické části se podařilo odebrat a zpracovat vzorky ze dvou vybraných lokalit a následně určit jejich druhové složení. Hodnoty podmínek prostředí, ve kterých byly druhy nalezeny, odpovídaly předchozím publikovaným nálezům. Výsledkem praktické části je floristický článek (JANATKOVÁ & NĚMCOVÁ 2009). 30
31 8. Použitá literatura: AND ERSEN. R. A. (1987): Synurophyceae species nov., a new clase of algae. Amer. J. Bot. 74: ANDERSEN. R. A. (2007):Molecular systematics of the chrysophyceae and Synurophyceae - Unravelling the algae: the past, present, and future of algal systematics, edit. Juliet Brodie, Jane Lewis, CRC Press, 15: ASMUND, B. (1955): Electron microscope observations on Mallomonas caudata and some remarks on its occurence in four Danish ponds. Bot. Tidsskr. 52: ASMUND, B. & KRISTIANSEN, J. (1986) The genus Mallomonas (Chrysophyceae). Opera Botanica 85: BHATTI S., COLMAN B. (2008): Inorganic carbon acquisition in some synurophyte algae. Physiol Plant. 133: CUMMING B.F., SMAOL J.P. & BIRKS H.J.B. (1990): The relationship between sedimentary chrysophyte scales (Chrysophycaea and Synurophyceae) anod limnological characteristics in 25 Norwegiam lakes Nordic Journal of Botany 11: CUMMING, B.F., SMOL, J.P., & BIRKS, H.J.B. (1992). Scaled chrysophytes. (Chrysophyceae and Synurophyceae) from Adirondack drainage lakes and their relationship to enviromental variables DIXIT, S.S., DIXIT, A.S. & SMOL, J.P. (1999): Lake sediment chrysophyte scales from the northeastern U.S.A. and their relationship to environmental variables. J. Phycology 35: DÜRRSCHMIDT, M. (1980): Studies on the Chrysophyceae from Rio Cruces, Prov. Valdivia, South Chile by Scanning and transmission microscopy. Nova Hedwigia 33: DÜRRSCHMIDT, M. (1982): Mallomonas parvula sp. nov. and Mallomonas retifera sp. nov. (Chrysophyceae, Synuraceae) from South Chile. Can. J. Bot. 60: DÜRRSCHMIDT, M. (1986): New species of the genus Mallomonas (Mallomonadaceae, Chrysophyceae) from New Zealand. In: Kristiansen, J. & Andersen, R. A. (eds): Chrysophytes. Aspects and Problems, pp., Cambridge University Press DÜRRSCHMIDT, M. & CROOME, R. (1985): Mallomonadaceae (Chrysophyceae) from Malaysia and Australia. - Nord. J. Bot. 5: FOTT, B. (1955): Scales of Mallomonas observed in the elektron microscope. Preslia 27:
32 FENCHEL T. & INLAY B. J. (2004): Species Diversity in Space and Time - Bioscience 54, 777, Cambridge Univ. Press, Cambridge, FINLAY B.J., CLARKE K.J. (1999): Ubiquitous dispersal of microbial species. Nature 400: 828 ITO, H. KALIN KRISTI KRISTIAN KRISTI KRISTI KRISTIAN (1990): Chrysophytes in the southern part of Hyogo prefecture, Japan (I). v současné biologii, 606 pp. Universita Karlova v Praze, Nakladatelství Karolinum, Praha ANSEN, J. (1971): A Mallomonas bloom in a Bulgarian mountain lake. Nova SEN, J. (1986): The ultrastructural base sof chrysophyte systematics and phylogeny ANSEN J. (2001): Biogeography of silica-scaled chrysophytes - Nova Hedwigia Beih ANSEN J. (2002): The genus Mallomonas (Synurophyceae) a taxonomic survey Biodivers Conserv 17: KRISTIANSEN, J. & PREISIG H. R. (EDS.) (2001): Encyclopedia of Chrysophyte genera. Bibl.Phycol. 110 KRISTI ANSEN, J. (1975): On the occurence of the species of Synura (Chrysophyceae) Verh. Internat. Verein. Limnol. 1: MARSI NICHO PICHRT ROIJACK ŘEZÁČ Chrysophyte flora in three ponds and reservoir. - Jap. J. Phycol. 38: A, T. & VÁŇA, J. (2005): Sinice, řasy, houby, mechorosty a podobné organismy Hedwigia 21: CRC Crit. Rev. Plant Sci. 4: : based on the ultrastructure of silica scales and bristles. Opera Bot 139: KRISTIANSEN, J. (2005): Golden Algae, 166 pp. Gantner Verlag Kommanditgesellschaft SEN, J. (2007): Dispersal and biogeography of silica-scaled chrysophytes CANO, L.J. AND P.A. SIVER. (1993): A paleolimnological assessment of Lake Acidification in five Connecticut lakes. J. Paleolimnology 9: LLS, K. H. & GERRATH, J. F (1985): The taxonomy of Synura (Chrysophyceae) in Ontario with special reference to taste and odour in water supplies. - Canad. J. Bot. 63: OVÁ, M. & VESELÁ, J. (in press): Silica-scaled chrysophytes in the Elbe Sandstone Region, Czech Republic.- Fottea.SUSHIL et al 1999 ERS, R. M. & KESSELS, H. (1986): Ecological characteristics of scale-bearing Chrysophyceae from the Netherlands. - Nord. J. Bot. 6: OVÁ, M. (2006): Mallomonas kalinae (Synurophyceae), a new species of alga from 32
33 ŘEZÁČ northern Bohemia, Czech Republic. - Preslia 78: OVÁ, M. & NEUSTUPA J. (2007): Distribution of the Genus Mallomonas Synurophyceae) Ubiquitous Dispersal in Microorganisms Evaluated. Protist 157( 4 SANDG REN C. D. (1988): The ecology of chrysophyte flagellates: their growth and perennation strategies as freshwater phytoplankton. In: Sandgren, C. D. (ed): Growth and reproductive strategies of freshwater phytoplankton: 9-104, Cambridge University Press, Cambridge. SAXBY SIVER, SIVER, SIVER, SIVER -ROUEN K. J. & LEADBEATER B.S., REYNOLDS C. S. (1998): The relationship between the growth of Synura petersenii (Synurophyceae) and components of the dissolved inorganic carbon system. Phycologia 37: P. A. (1987a): The distribution and variation of Synura species (Chrysophyceae) in P.A. (1987): Distribution of scaled chrysophytes in 17 Adirondack (New York) lakes P. A. (1988): The distribution of scale chrysophytes along a ph gradient, Canadian P. A. (1989): The distribution of scaled chrysophytes along a ph gradient. Can J Bot 67: SIVER, P. A. (1991): The biology of Mallomonas: Morphology, Taxonomy and Ekology. Kluwer Academic Publishers SIVER, P.A. (1993): Inferring lakewater specific conductivity with scaled chrysophytes. Limnol. Oceanogr. 38: SIVER, P. A. (1995): The distribution of chrysophytes along enviromental gradients: thein use as biological idicators. In SANDGREN, C.D., SMOL, J.P & KRISTIANSEN, J. (eds): Chrysophyte algae, pp. Cambridge University Press, Cambridge. SIVER, P.A. (1999): Morphological observations of synurophycean algae from some acidic habitats, including the description of a sub-specific taxon. Nordic Journal of Botany 19: SIVER, Connecticut, USA. - Nord. J. Bot. 7: with special reference to ph. Can. J. Bot. 66: Journal of botany 59 P.A. (2002): Paleolimnology: Use of siliceous structures of chrysophytes as biological indicators in freshwater systems. In: The Encyclopedia of Environmental Microbiology. G. Bitton, Section Editor. John Wiley. Pages SIVER, P.A. & J.S. HAMER. (1989): Use of extant populations of scaled chrysophytes for the inference of lakewater ph. Can. J. Fish. Aquat. Sci. 47: SIVER, P.A., LOTT, A.M., CASH, E., MOSS, J. & L.J. MARSICANO. (1999): Century 33
34 SIVER, SIVER changes in Connecticut, U.S.A., lakes as inferred from siliceous algal remains and their relationship to land-use changes. Limnology and Oceanography 44: of scaled chrysophytes in Vermont and New Hampshire (USA) lakes and their utility to infer lake water chemistry. Nordic Journal of Botany 20: Nova Hedwigia Beih 114: SIVER, P.A., RICARD, R., GOODWIN, R. & A.E. GIBLIN.(2003): Estimating historical in-lake alkalinity generation and its relationship to lake chemistry as inferred from algal microfossils. J. Paleolimnology 29: SIVER, P. A. & SMOL, J. P. (1993): The use of scaled chrysophytes in long term monitoring programs for the detection of changes lakewater acidity Water, Air and Soil polution 71: SIVER, P. A. & LOTT A. M.(2000): Preliminary investigations on the distribution P. A., MARSICANO L. J. (1996): Inferring lake tropic status using scaled chrysophytes. P.A. & A.P. WOLFE (2005): Eocene scaled chrysophytes with pronounced modern affinities. Int. J. Plant Sci. 166(3): SMOL, J. P. (1995): Apli cation of chrysophytes to problems in paleoecology. In: SANDGREN, C.D., SMOL, J.P & KRISTIANSEN, J. (eds): Chrysophyte algae, , Cambridge University Press, Cambridge. SMOL, J. P., CHARLES D. F., & D. R. WHITEHEAD. (1984): Mallomonadacean (Chrysophyceae) assemblages and their relationships with limnological characteristics in 38 Adirondack (New York) Lakes. Can. J. Bot 62: TAKAH WEE, J ASHI, E. (1959): Studies on genera Mallomonas, Synura, and other plankton in fresh- water by electron microscope (1). Bull. Yamagata Univ. Agr. Sci. 3: L., BOOTH, D. J.. &. BOSSIER, M. A. (1993): Synurophyceae from Southern. Atlantic Coastal Plain of North America: A preliminary survey in Louisiana, USA. Nord. J. Bot. 11:
Seznam příloh Příloha 1: Příloha 2: Příloha 3: Příloha 4: Příloha 5: Příloha 6: Příloha 7:
 Seznam příloh Příloha 1: Mapa sledovaných lokalit Příloha 2: Fotografie lokalit Příloha 3: Nalezené druhy křemičitých chrysomonád Příloha 4: Seznam druhů a jejich výskyt na sledovaných lokalitách Příloha
Seznam příloh Příloha 1: Mapa sledovaných lokalit Příloha 2: Fotografie lokalit Příloha 3: Nalezené druhy křemičitých chrysomonád Příloha 4: Seznam druhů a jejich výskyt na sledovaných lokalitách Příloha
Chrysomonády. Yvonne. katedra botaniky Př.F. UK Praha
 Chrysomonády - bičíkovci se skleněnou nou schránkou Yvonne Němcová katedra botaniky Př.F. UK Praha Třída zahrnuje jednotlivě žijící nebo koloniální sladkovodní bičíkovce. Jejich výživa je fotoautotrofní.
Chrysomonády - bičíkovci se skleněnou nou schránkou Yvonne Němcová katedra botaniky Př.F. UK Praha Třída zahrnuje jednotlivě žijící nebo koloniální sladkovodní bičíkovce. Jejich výživa je fotoautotrofní.
Vliv faktorů prostředí na morfologickou variabilitu křemičitých struktur chrysomonád (Synurophyceae)
") Univerzita Karlova v Praze Přírodovědecká fakulta Katedra botaniky Bakalářská práce Vliv faktorů prostředí na morfologickou variabilitu křemičitých struktur chrysomonád (Synurophyceae) Martina Pichrtová
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra botaniky Bakalářská práce Vliv faktorů prostředí na morfologickou variabilitu křemičitých struktur chrysomonád (Synurophyceae) Martina Pichrtová
CHRYSOMONÁDY A SLUNIVKY S KŘEMITÝMI ŠUPINAMI MONTÁNNÍCH A SUBALPINSKÝCH RAŠELINIŠŤ KRKONOŠ
 OPERA CORCONTICA 37: 334 340, 2000 CHRYSOMONÁDY A SLUNIVKY S KŘEMITÝMI ŠUPINAMI MONTÁNNÍCH A SUBALPINSKÝCH RAŠELINIŠŤ KRKONOŠ Silica scaled chrysophytes and heliozoans of the Krkonoše Mountains alpine
OPERA CORCONTICA 37: 334 340, 2000 CHRYSOMONÁDY A SLUNIVKY S KŘEMITÝMI ŠUPINAMI MONTÁNNÍCH A SUBALPINSKÝCH RAŠELINIŠŤ KRKONOŠ Silica scaled chrysophytes and heliozoans of the Krkonoše Mountains alpine
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka
 primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
Univerzita Karlova v Praze Přírodovědecká fakulta. Studijní program: Biologie Katedra botaniky
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Katedra botaniky Pavla Slámová Skrytá diverzita autotrofních bičíkovců na příkladu zlativek (Synurophyceae) Hidden diversity
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Katedra botaniky Pavla Slámová Skrytá diverzita autotrofních bičíkovců na příkladu zlativek (Synurophyceae) Hidden diversity
1. Cíle práce 1. Cíle práce
 Summary Presented diploma thesis concerns the chrysophyte flora and its seasonal periodicity of two floodplain pools of river Lužnice in the Southern Bohemia and one pond in the Eastern Bohemia. The samples
Summary Presented diploma thesis concerns the chrysophyte flora and its seasonal periodicity of two floodplain pools of river Lužnice in the Southern Bohemia and one pond in the Eastern Bohemia. The samples
S postupným nárůstem frekvence lokalit se zjevnou nadprodukcí (tzv. hypertrofie) přechází definice v devadesátých letech do podoby
 přechází definice v devadesátých letech do podoby") Eutrofizace je definována jako proces zvyšování produkce organické hmoty ve vodě, ke které dochází především na základě zvýšeného přísunu živin (OECD 1982) S postupným nárůstem frekvence lokalit se zjevnou
Eutrofizace je definována jako proces zvyšování produkce organické hmoty ve vodě, ke které dochází především na základě zvýšeného přísunu živin (OECD 1982) S postupným nárůstem frekvence lokalit se zjevnou
4 ROKY HYDROBIOLOGA NA MOSTECKÉM JEZEŘE
 4 ROKY HYDROBIOLOGA NA MOSTECKÉM JEZEŘE JANA ŘÍHOVÁ AMBROŽOVÁ, BARBORA KOFROŇOVÁ VŠCHT ÚTVP TECHNICKÁ 5, PRAHA 6 UJEP FŽP KPV KRÁLOVA VÝŠINA 7, ÚSTÍ NAD LABEM V rámci řešeného projektu TA ČR č. TA 01020592,
4 ROKY HYDROBIOLOGA NA MOSTECKÉM JEZEŘE JANA ŘÍHOVÁ AMBROŽOVÁ, BARBORA KOFROŇOVÁ VŠCHT ÚTVP TECHNICKÁ 5, PRAHA 6 UJEP FŽP KPV KRÁLOVA VÝŠINA 7, ÚSTÍ NAD LABEM V rámci řešeného projektu TA ČR č. TA 01020592,
Pedagogická činnost pro jmenovací řízení
 Pedagogická činnost pro jmenovací řízení Jiří Neustupa Pedagogická činnost Bakalářský studijní program Biologie (studijní obory Biologie, Ekologická a evoluční biologie, Molekulární biologie a biochemie
Pedagogická činnost pro jmenovací řízení Jiří Neustupa Pedagogická činnost Bakalářský studijní program Biologie (studijní obory Biologie, Ekologická a evoluční biologie, Molekulární biologie a biochemie
Výběr substrátu při odběru fytobentosu
 Výběr substrátu při odběru fytobentosu lze ovlivnit výsledky monitoringu? Mgr. Lenka Šejnohová BU AVČR, Brno Označení společenstev dle typu substrátů epifyton taxony uzpůsobené k osidlování mechorostů,
Výběr substrátu při odběru fytobentosu lze ovlivnit výsledky monitoringu? Mgr. Lenka Šejnohová BU AVČR, Brno Označení společenstev dle typu substrátů epifyton taxony uzpůsobené k osidlování mechorostů,
Třída: XANTHOPHYCEAE
 HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc.
 KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc. Úvod do problematiky Fytoplankton=hlavní producent biomasy, na kterém
KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc. Úvod do problematiky Fytoplankton=hlavní producent biomasy, na kterém
Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů
 Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta TŘÍDA: Dinophyceae ŘÁD: Gonyaulacales Ceratium (TP) ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta
Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta TŘÍDA: Dinophyceae ŘÁD: Gonyaulacales Ceratium (TP) ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta
Jan POTUŽÁK a Kateřina KOLÁŘOVÁ. Povodí Vltavy, státní podnik, VHL České Budějovice
 Jan POTUŽÁK a Kateřina KOLÁŘOVÁ Povodí Vltavy, státní podnik, VHL České Budějovice Mapy a umístění rybník Zhejral VN Karhov Rybník Zhejral (49 º 13'12.975''N; 15º18 48.557''E) Zatopená plocha: 14,46 ha
Jan POTUŽÁK a Kateřina KOLÁŘOVÁ Povodí Vltavy, státní podnik, VHL České Budějovice Mapy a umístění rybník Zhejral VN Karhov Rybník Zhejral (49 º 13'12.975''N; 15º18 48.557''E) Zatopená plocha: 14,46 ha
Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus TŘÍDA: Synurophyceae Synura
Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus TŘÍDA: Synurophyceae Synura
Monitoring stavu vody ve vodní nádrži v parku Pod Plachtami
 Sdružení Flos Aquae Monitoring stavu vody ve vodní nádrži v parku Pod Plachtami Autorský kolektiv: Ing. Eliška Maršálková, Ph.D. Ing. Marcela Lagová Prof. Ing. Blahoslav Maršálek, CSc. Brno, květen 2013
Sdružení Flos Aquae Monitoring stavu vody ve vodní nádrži v parku Pod Plachtami Autorský kolektiv: Ing. Eliška Maršálková, Ph.D. Ing. Marcela Lagová Prof. Ing. Blahoslav Maršálek, CSc. Brno, květen 2013
Vliv abiotických a biotických stresorů na vlastnosti rostlin 2015, ČZU Praha
 Vliv abiotických a biotických stresorů na vlastnosti rostlin 2015, ČZU Praha Sándor T. Forczek #, Josef Holík #, Luděk Rederer &, Václav Koza & # Ústav experimantální botaniky AV ČR, v.v.i. & Povodí Labe
Vliv abiotických a biotických stresorů na vlastnosti rostlin 2015, ČZU Praha Sándor T. Forczek #, Josef Holík #, Luděk Rederer &, Václav Koza & # Ústav experimantální botaniky AV ČR, v.v.i. & Povodí Labe
Změny trofického potenciálu a koncentrace chlorofylu a v řece Jihlavě a v nádržích Dalešice a Mohelno od jejich napuštění
 Czech Phycology, Olomouc, 2: 115-124, 22 115 Změny trofického potenciálu a koncentrace chlorofylu a v řece Jihlavě a v nádržích Dalešice a Mohelno od jejich napuštění Changes of the trophic potential and
Czech Phycology, Olomouc, 2: 115-124, 22 115 Změny trofického potenciálu a koncentrace chlorofylu a v řece Jihlavě a v nádržích Dalešice a Mohelno od jejich napuštění Changes of the trophic potential and
PROBLEMATIKA VZORKOVÁNÍ PŘÍRODNÍCH KOUPACÍCH VOD
 PROBLEMATIKA VZORKOVÁNÍ PŘÍRODNÍCH KOUPACÍCH VOD Petr Pumann, Tereza Pouzarová Státní Vodárenská biologie 2012 Praha, 1.-2.2.2012 Státní Zdroje dat IS PiVo data od roku 2004 Programy zkoušení způsobilosti
PROBLEMATIKA VZORKOVÁNÍ PŘÍRODNÍCH KOUPACÍCH VOD Petr Pumann, Tereza Pouzarová Státní Vodárenská biologie 2012 Praha, 1.-2.2.2012 Státní Zdroje dat IS PiVo data od roku 2004 Programy zkoušení způsobilosti
NÁDRŽ KLÍČAVA VZTAH KVALITY VODY A INTENZITY VODÁRENSKÉHO VYUŽÍVÁNÍ
 Citace Duras J.: Nádrž Klíčava vztah kvality a intenzity vodárenského využití. Sborník konference Pitná voda 2010, s. 271-276. W&ET Team, Č. Budějovice 2010. ISBN 978-80-254-6854-8 NÁDRŽ KLÍČAVA VZTAH
Citace Duras J.: Nádrž Klíčava vztah kvality a intenzity vodárenského využití. Sborník konference Pitná voda 2010, s. 271-276. W&ET Team, Č. Budějovice 2010. ISBN 978-80-254-6854-8 NÁDRŽ KLÍČAVA VZTAH
Ekosystém. tok energie toky prvků biogeochemické cykly
 Ekosystém tok energie toky prvků biogeochemické cykly Ekosystém se sestává z abiotického prostředí a biotické složky (společenstva) a jejich vzájemných interakcí. Ekosystém si geograficky můžeme definovat
Ekosystém tok energie toky prvků biogeochemické cykly Ekosystém se sestává z abiotického prostředí a biotické složky (společenstva) a jejich vzájemných interakcí. Ekosystém si geograficky můžeme definovat
Využití zásoby živin a primární produkce v eutrofních rybnících
 Využití zásoby živin a primární produkce v eutrofních rybnících Libor Pechar a kolektiv Jihočeská Univerzita v Českých Budějovicích Zemědělská fakulta, Laboratoř aplikované ekologie a ENKI o.p.s., Třeboň
Využití zásoby živin a primární produkce v eutrofních rybnících Libor Pechar a kolektiv Jihočeská Univerzita v Českých Budějovicích Zemědělská fakulta, Laboratoř aplikované ekologie a ENKI o.p.s., Třeboň
Malý test znalostí odběrových skupin - fotografie
 Malý test znalostí odběrových skupin - fotografie správné odpovědi, vy a komentáře PT#V/8/2016 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 15. 6. 2016 Účastníci programu
Malý test znalostí odběrových skupin - fotografie správné odpovědi, vy a komentáře PT#V/8/2016 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 15. 6. 2016 Účastníci programu
Užití země v České republice v letech 1994 až 2012 Karel Matějka IDS, Na Komořsku 2175/2a, 143 00 Praha 4, Česká republika matejka@infodatasys.
 Užití země v České republice v letech 1994 až 2012 Karel Matějka IDS, Na Komořsku 2175/2a, 143 00 Praha 4, Česká republika matejka@infodatasys.cz Po roce 19 došlo k výrazné změně hospodářských poměrů v
Užití země v České republice v letech 1994 až 2012 Karel Matějka IDS, Na Komořsku 2175/2a, 143 00 Praha 4, Česká republika matejka@infodatasys.cz Po roce 19 došlo k výrazné změně hospodářských poměrů v
Botanika bezcévných rostlin pro učitele 2. praktické cvičení
 Botanika bezcévných rostlin pro učitele 2. praktické cvičení ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus (TP) TŘÍDA: Synurophyceae Synura petersenii
Botanika bezcévných rostlin pro učitele 2. praktické cvičení ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus (TP) TŘÍDA: Synurophyceae Synura petersenii
Univerzita Karlova v Praze Přírodovědecká fakulta. Studijní program: Biologie Studijní obor: Botanika. Bc. Jana Kreidlová
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Botanika Bc. Jana Kreidlová Synura echinulata (Synurales) tvarová variabilita křemičitých šupin přírodních populací
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Botanika Bc. Jana Kreidlová Synura echinulata (Synurales) tvarová variabilita křemičitých šupin přírodních populací
Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA)
") HYDROBOTANIKA Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA) Celková charakteristika Relativně malá skupinka co do počtu druhů (500 recentních druhů v 50 rodech), ale zejména mořští zástupci mají značný ekologický
HYDROBOTANIKA Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA) Celková charakteristika Relativně malá skupinka co do počtu druhů (500 recentních druhů v 50 rodech), ale zejména mořští zástupci mají značný ekologický
Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA)
") HYDROBOTANIKA Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA) Celková charakteristika Relativně malá skupinka co do počtu druhů (500 recentních druhů v 50 rodech), ale zejména mořští zástupci mají značný ekologický
HYDROBOTANIKA Oddělení: HAPTOPHYTA (PRYMNESIOPHYTA) Celková charakteristika Relativně malá skupinka co do počtu druhů (500 recentních druhů v 50 rodech), ale zejména mořští zástupci mají značný ekologický
STANOVENÍ TVARU A DISTRIBUCE VELIKOSTI ČÁSTIC MODELOVÝCH TYPŮ NANOMATERIÁLŮ. Edita BRETŠNAJDROVÁ a, Ladislav SVOBODA a Jiří ZELENKA b
 STANOVENÍ TVARU A DISTRIBUCE VELIKOSTI ČÁSTIC MODELOVÝCH TYPŮ NANOMATERIÁLŮ Edita BRETŠNAJDROVÁ a, Ladislav SVOBODA a Jiří ZELENKA b a UNIVERZITA PARDUBICE, Fakulta chemicko-technologická, Katedra anorganické
STANOVENÍ TVARU A DISTRIBUCE VELIKOSTI ČÁSTIC MODELOVÝCH TYPŮ NANOMATERIÁLŮ Edita BRETŠNAJDROVÁ a, Ladislav SVOBODA a Jiří ZELENKA b a UNIVERZITA PARDUBICE, Fakulta chemicko-technologická, Katedra anorganické
Univerzita Hradec Králové Přírodovědecká fakulta katedra biologie
 Univerzita Hradec Králové Přírodovědecká fakulta katedra biologie Populační ekologie, stanovištní nároky a generativní reprodukce kriticky ohrožených druhů cévnatých rostlin ČR (Adenophora liliifolia,
Univerzita Hradec Králové Přírodovědecká fakulta katedra biologie Populační ekologie, stanovištní nároky a generativní reprodukce kriticky ohrožených druhů cévnatých rostlin ČR (Adenophora liliifolia,
Třída: RAPHIDOPHYCEAE
 HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
NORMY PRO BIOLOGICKÉ METODY
 NORMY PRO BIOLOGICKÉ METODY Ing. Lenka Fremrová Sweco Hydroprojekt a.s. 1 ČSN EN 16698 Návod pro kvantitativní a kvalitativní odběr vzorků fytoplanktonu z vnitrozemských vod Norma popisuje postupy odběru
NORMY PRO BIOLOGICKÉ METODY Ing. Lenka Fremrová Sweco Hydroprojekt a.s. 1 ČSN EN 16698 Návod pro kvantitativní a kvalitativní odběr vzorků fytoplanktonu z vnitrozemských vod Norma popisuje postupy odběru
6. Tzv. holocenní klimatické optimum s maximálním rozvojem lesa bylo typické pro a) preboreál b) atlantik c) subrecent
 preboreál b) atlantik c) subrecent") 1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
135 str., 70 barevných fotografií, 3 pérovky, 52 druhů nejohroženějších mexických kaktusů, fotografie z naleziště, množení, pevná vazba, anglicky,
 135 str., 70 barevných fotografií, 3 pérovky, 52 druhů nejohroženějších mexických kaktusů, fotografie z naleziště, množení, pevná vazba, anglicky, 890,- Kč. 315 str., druhé vydání dobré příručky sloužící
135 str., 70 barevných fotografií, 3 pérovky, 52 druhů nejohroženějších mexických kaktusů, fotografie z naleziště, množení, pevná vazba, anglicky, 890,- Kč. 315 str., druhé vydání dobré příručky sloužící
Konference Vodárenská biologie 2019, února 2019, Interhotel Olympik, Praha
 Konference Vodárenská biologie 2019, 6. 7. února 2019, Interhotel Olympik, Praha (neboli top-down effect ) je založena na ovlivnění potravního řetězce vodního ekosystému: dravé ryby plaktonožravé ryby
Konference Vodárenská biologie 2019, 6. 7. února 2019, Interhotel Olympik, Praha (neboli top-down effect ) je založena na ovlivnění potravního řetězce vodního ekosystému: dravé ryby plaktonožravé ryby
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162
 ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
Sdružení Flos Aquae SLEDOVÁNÍ ZMĚN V MNOŽSTVÍ A SLOŽENÍ FYTOPLANKTONNÍCH SPOLEČENSTEV V BRNĚNSKÉ ÚDOLNÍ NÁDRŽI V OBDOBÍ KVĚTEN ŘÍJEN 2010
 Sdružení Flos Aquae SLEDOVÁNÍ ZMĚN V MNOŽSTVÍ A SLOŽENÍ FYTOPLANKTONNÍCH SPOLEČENSTEV V BRNĚNSKÉ ÚDOLNÍ NÁDRŽI V OBDOBÍ KVĚTEN ŘÍJEN 21 Autorský kolektiv: Ing. Eliška Maršálková, Ph.D. Doc. Ing. Radovan
Sdružení Flos Aquae SLEDOVÁNÍ ZMĚN V MNOŽSTVÍ A SLOŽENÍ FYTOPLANKTONNÍCH SPOLEČENSTEV V BRNĚNSKÉ ÚDOLNÍ NÁDRŽI V OBDOBÍ KVĚTEN ŘÍJEN 21 Autorský kolektiv: Ing. Eliška Maršálková, Ph.D. Doc. Ing. Radovan
Rybníky a malé vodní nádrže jako součást kulturního dědictví z pohledu kvality vodního prostředí
 MINISTERSTVO KULTURY ČESKÉ REPUBLIKY Rybníky a malé vodní nádrže jako součást kulturního dědictví z pohledu kvality vodního prostředí Miloš ROZKOŠNÝ, Miriam DZURÁKOVÁ, Hana HUDCOVÁ, Pavel SEDLÁČEK Výzkumný
MINISTERSTVO KULTURY ČESKÉ REPUBLIKY Rybníky a malé vodní nádrže jako součást kulturního dědictví z pohledu kvality vodního prostředí Miloš ROZKOŠNÝ, Miriam DZURÁKOVÁ, Hana HUDCOVÁ, Pavel SEDLÁČEK Výzkumný
Tlumení rozvoje sinic a řas pomocí mikrobiálněenzymatického
 Tlumení rozvoje sinic a řas pomocí mikrobiálněenzymatického preparátu SEKOL Lakus aqua Pokusná aplikace na vodní nádrži Pod Santonem vegetační sezóna 2007 Zemědělská vodohospodářská zpráva Brno 2007 Zpracoval:
Tlumení rozvoje sinic a řas pomocí mikrobiálněenzymatického preparátu SEKOL Lakus aqua Pokusná aplikace na vodní nádrži Pod Santonem vegetační sezóna 2007 Zemědělská vodohospodářská zpráva Brno 2007 Zpracoval:
PRACOVNÍ LIST EVVO - VODA
 Projekt Integrovaný vzdělávací systém města Jáchymov Mosty indikátor 06.43.19 PRACOVNÍ LIST EVVO - VODA Úkol: Fyzikální a chemická analýza vody Princip: Vlastním pozorováním získat poznatky o vlastnostech
Projekt Integrovaný vzdělávací systém města Jáchymov Mosty indikátor 06.43.19 PRACOVNÍ LIST EVVO - VODA Úkol: Fyzikální a chemická analýza vody Princip: Vlastním pozorováním získat poznatky o vlastnostech
fytopatogenů a modelování
 Mapování výskytu fytopatogenů a modelování škod na dřevinách v lesích ČR Dušan Romportl, Eva Chumanová & Karel Černý VÚKOZ, v.v.i. Mapování výskytu vybraných fytopatogenů Introdukce nepůvodních patogenů
Mapování výskytu fytopatogenů a modelování škod na dřevinách v lesích ČR Dušan Romportl, Eva Chumanová & Karel Černý VÚKOZ, v.v.i. Mapování výskytu vybraných fytopatogenů Introdukce nepůvodních patogenů
Univerzita Karlova v Praze Přírodovědecká fakulta. Katedra botaniky
 Univerzita Karlova v Praze Přírodovědecká fakulta Katedra botaniky Kateřina Bartošová Stomatocysty a šupiny chrysofyt a jejich využití v paleolimnologii Chrysophycean cysts and scales in paleolimnological
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra botaniky Kateřina Bartošová Stomatocysty a šupiny chrysofyt a jejich využití v paleolimnologii Chrysophycean cysts and scales in paleolimnological
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková
 Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Malý test znalostí odběrových skupin - fotografie
 Malý test znalostí odběrových skupin - fotografie správné odpovědi, vyhodnocení a komentáře PT#V/8/2018 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 3. 7. 2018 Účastníci
Malý test znalostí odběrových skupin - fotografie správné odpovědi, vyhodnocení a komentáře PT#V/8/2018 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 3. 7. 2018 Účastníci
ZHODNOCENÍ DLOUHODOBÉHO VÝVOJE KVALITY VODY VE ZBYTKOVÝCH JEZERECH SHP
 ZHODNOCENÍ DLOUHODOBÉHO VÝVOJE KVALITY VODY VE ZBYTKOVÝCH JEZERECH SHP I. PŘIKRYL ENKI O.P.S. TŘEBOŇ PROJEKT VITA-MIN 18.06.2019, Most UMÍSTĚNÍ JEZER 2 BARBORA A MALÉ LOMY V SEVEROČESKÉ PÁNVI JSOU DESÍTKY
ZHODNOCENÍ DLOUHODOBÉHO VÝVOJE KVALITY VODY VE ZBYTKOVÝCH JEZERECH SHP I. PŘIKRYL ENKI O.P.S. TŘEBOŇ PROJEKT VITA-MIN 18.06.2019, Most UMÍSTĚNÍ JEZER 2 BARBORA A MALÉ LOMY V SEVEROČESKÉ PÁNVI JSOU DESÍTKY
ZHORŠENÍ JAKOSTI VODY V NÁDRŽI NOVÁ ŘÍŠE VODÁRENSKÁ BIOLOGIE 2017 RODAN GERIŠ, DUŠAN KOSOUR POVODÍ MORAVY, S.P.
 ZHORŠENÍ JAKOSTI VODY V NÁDRŽI NOVÁ ŘÍŠE VODÁRENSKÁ BIOLOGIE 2017 RODAN GERIŠ, DUŠAN KOSOUR POVODÍ MORAVY, S.P. Jeden z autorů Vás vítá na prezentaci přímo z nádrže... Nová Říše pohled na povodí Základní
ZHORŠENÍ JAKOSTI VODY V NÁDRŽI NOVÁ ŘÍŠE VODÁRENSKÁ BIOLOGIE 2017 RODAN GERIŠ, DUŠAN KOSOUR POVODÍ MORAVY, S.P. Jeden z autorů Vás vítá na prezentaci přímo z nádrže... Nová Říše pohled na povodí Základní
ROSTLINNÍ PREDÁTOŘI. Vliv eutrofizace na vodní svět. Co se vám bude hodit vědět
 ROSTLINNÍ PREDÁTOŘI Vliv eutrofizace na vodní svět Vzpomeňte si, jak jste šli v létě na výletě krajinou, kde bylo několik vodních zdrojů jako řeky, rybníky, potůčky, popř. jezera. Možná si vzpomenete,
ROSTLINNÍ PREDÁTOŘI Vliv eutrofizace na vodní svět Vzpomeňte si, jak jste šli v létě na výletě krajinou, kde bylo několik vodních zdrojů jako řeky, rybníky, potůčky, popř. jezera. Možná si vzpomenete,
Úvod do biochemie. Vypracoval: RNDr. Milan Zimpl, Ph.D.
 Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Vizuální hodnocení vodního květu sinic
 Vizuální hodnocení vodního květu sinic Petr Pumann Seminář Vyhodnocení programů zkoušení způsobilosti v oblasti orientační senzorické analýzy a odběrů pitné a koupací vody 20.6.2007, Praha, Statní Co to
Vizuální hodnocení vodního květu sinic Petr Pumann Seminář Vyhodnocení programů zkoušení způsobilosti v oblasti orientační senzorické analýzy a odběrů pitné a koupací vody 20.6.2007, Praha, Statní Co to
Vysoká eutrofizační účinnost fosforu původem z odpadních vod v nádrži Lipno
 Vysoká eutrofizační účinnost fosforu původem z odpadních vod v nádrži Lipno Josef Hejzlar Petr Znachor Zuzana Sobolíková Vladimír Rohlík Biologické centrum AV ČR, v. v. i. Hydrobiologický ústav České Budějovice
Vysoká eutrofizační účinnost fosforu původem z odpadních vod v nádrži Lipno Josef Hejzlar Petr Znachor Zuzana Sobolíková Vladimír Rohlík Biologické centrum AV ČR, v. v. i. Hydrobiologický ústav České Budějovice
MAKROZOOBENTOS NA HNĚDOUHELNÝCH VÝSYPKÁCH (BENTHIC INVERTEBRATES OF POST-MINING SPOIL HEAPS)
") Hydrochemický monitoring vod MAKROZOOBENTOS NA HNĚDOUHELNÝCH VÝSYPKÁCH (BENTHIC INVERTEBRATES OF POST-MINING SPOIL HEAPS) Michal Straka, Denisa Němejcová, Marek Polášek Výzkumný ústav vodohospodářský TGM,
Hydrochemický monitoring vod MAKROZOOBENTOS NA HNĚDOUHELNÝCH VÝSYPKÁCH (BENTHIC INVERTEBRATES OF POST-MINING SPOIL HEAPS) Michal Straka, Denisa Němejcová, Marek Polášek Výzkumný ústav vodohospodářský TGM,
Katedra botaniky PřF UK, Benátská 2, Praha 2, CZ
 OPERA CORCONTICA 37: 341 346, 2000 ŘASY KRKONOŠSKÝCH RAŠELINIŠŤ Algae of the peat bogs in the Krkonoše Mountains NOVÁKOVÁ SYLVIE Katedra botaniky PřF UK, Benátská 2, 128 01 Praha 2, CZ Během dvouletého
OPERA CORCONTICA 37: 341 346, 2000 ŘASY KRKONOŠSKÝCH RAŠELINIŠŤ Algae of the peat bogs in the Krkonoše Mountains NOVÁKOVÁ SYLVIE Katedra botaniky PřF UK, Benátská 2, 128 01 Praha 2, CZ Během dvouletého
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu
 TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie
 synekologie b) autekologie c) demekologie") 1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
Třída: RAPHIDOPHYCEAE
 HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
Možné dopady klimatické změny na dostupnost vodních zdrojů Jaroslav Rožnovský
 Český hydrometeorologický ústav, pobočka Brno Kroftova 43, 616 67 Brno e-mail:roznovsky@chmi.cz http://www.chmi.cz telefon: 541 421 020, 724 185 617 Možné dopady klimatické změny na dostupnost vodních
Český hydrometeorologický ústav, pobočka Brno Kroftova 43, 616 67 Brno e-mail:roznovsky@chmi.cz http://www.chmi.cz telefon: 541 421 020, 724 185 617 Možné dopady klimatické změny na dostupnost vodních
+ Fytoplankton (producenti) Zooplankton, zoobentos (konzumenti 1.řádu) Ryby (konzumenti 2.řádu)
 Zooplankton, zoobentos (konzumenti 1.řádu) Ryby (konzumenti 2.řádu)") Trend budování nových rybníků, tůněk a nádrží Sukcese společenstva jako předmět zájmu z pohledu rybářství i ochrany přírody Požadovány komplexní studie ekosystému Fyzikálně-chemické parametry + Fytoplankton
Trend budování nových rybníků, tůněk a nádrží Sukcese společenstva jako předmět zájmu z pohledu rybářství i ochrany přírody Požadovány komplexní studie ekosystému Fyzikálně-chemické parametry + Fytoplankton
Ústřední kontrolní a zkušební ústav zemědělský Oddělení půdy a lesnictví
 Ústřední a zkušební ústav zemědělský Oddělení půdy a lesnictví Analýza a vyhodnocení účinnosti leteckého vápnění, provedeného v roce 2008 v Krušných horách v okolí Horního Jiřetína, po pěti letech od data
Ústřední a zkušební ústav zemědělský Oddělení půdy a lesnictví Analýza a vyhodnocení účinnosti leteckého vápnění, provedeného v roce 2008 v Krušných horách v okolí Horního Jiřetína, po pěti letech od data
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu
 TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
3. HYDROLOGICKÉ POMĚRY
 Tunel Umiray Macua, Filipíny hydrogeologický monitoring Jitka Novotná1, Pavel Blaha2, Roman Duras3 1 GEOtest, a.s., Brno, Šmahova 112 novotna@geotest.cz 2 GEOtest, a.s., Brno, Šmahova 112 blaha@geotest.cz
Tunel Umiray Macua, Filipíny hydrogeologický monitoring Jitka Novotná1, Pavel Blaha2, Roman Duras3 1 GEOtest, a.s., Brno, Šmahova 112 novotna@geotest.cz 2 GEOtest, a.s., Brno, Šmahova 112 blaha@geotest.cz
2. Kteří z odborníků mohli vyslovit následující tvrzení?
 1. Vysvětli pojmy: - EKOLOGIE, OCHRANA PŘÍRODY, ZDROJ, PODMÍNKA Ekologie věda, která se zabývá zkoumáním vztahů mezi organismy a prostředím a mezi organismy vzájemně. 1 bod Ochrana přírody lidská péče
1. Vysvětli pojmy: - EKOLOGIE, OCHRANA PŘÍRODY, ZDROJ, PODMÍNKA Ekologie věda, která se zabývá zkoumáním vztahů mezi organismy a prostředím a mezi organismy vzájemně. 1 bod Ochrana přírody lidská péče
Pseudodendromonas vlkii (VLK) BOURRELLY poutník po systému
 BOURRELLY poutník po systému") Czech Phycology, Olomouc, 2: 33-37, 2002 33 Pseudodendromonas vlkii (VLK) BOURRELLY poutník po systému Pseudodendromonas vlkii (VLK) BOURRELLY a taxonomic wanderer Sylvie Nováková Katedra botaniky PřF
Czech Phycology, Olomouc, 2: 33-37, 2002 33 Pseudodendromonas vlkii (VLK) BOURRELLY poutník po systému Pseudodendromonas vlkii (VLK) BOURRELLY a taxonomic wanderer Sylvie Nováková Katedra botaniky PřF
I. Morfologie toku s ohledem na bilanci transportu plavenin a splavenin
 I. Morfologie toku s ohledem na bilanci transportu plavenin a splavenin I.1. Tvar koryta a jeho vývoj Klima, tvar krajiny, vegetace a geologie povodí určují morfologii vodního toku (neovlivněného antropologickou
I. Morfologie toku s ohledem na bilanci transportu plavenin a splavenin I.1. Tvar koryta a jeho vývoj Klima, tvar krajiny, vegetace a geologie povodí určují morfologii vodního toku (neovlivněného antropologickou
TWINNING PROJEKT CZ01/IB-EN-01
 Projekt / Komponent TWINNING CZ/01 IB-EN-01 Environmentální monitoring Autoři Konecny, Weber, Hunt Ze dne / Verze 29. října 02/2.0 Směrný dokument Info Fáze 2 Reference / Strana EM strategický směrný dokument
Projekt / Komponent TWINNING CZ/01 IB-EN-01 Environmentální monitoring Autoři Konecny, Weber, Hunt Ze dne / Verze 29. října 02/2.0 Směrný dokument Info Fáze 2 Reference / Strana EM strategický směrný dokument
Lesní hospodářství a ochrana biodiversity v ČR základní východiska v kontextu právních předpisů
 Lesní hospodářství a ochrana biodiversity v ČR základní východiska v kontextu právních předpisů Petr Kjučukov ČZU v Praze, Fakulta lesnická a dřevařská Katedra ekologie lesa Kašperské Hory 19.5.2016 Biodiversita
Lesní hospodářství a ochrana biodiversity v ČR základní východiska v kontextu právních předpisů Petr Kjučukov ČZU v Praze, Fakulta lesnická a dřevařská Katedra ekologie lesa Kašperské Hory 19.5.2016 Biodiversita
Voda jako životní prostředí - světlo
 Hydrobiologie pro terrestrické biology Téma 6: Voda jako životní prostředí - světlo Sluneční světlo ve vodě Sluneční záření dopadající na hladinu vody je 1) cestou hlavního přísunu tepla do vody 2) zdrojem
Hydrobiologie pro terrestrické biology Téma 6: Voda jako životní prostředí - světlo Sluneční světlo ve vodě Sluneční záření dopadající na hladinu vody je 1) cestou hlavního přísunu tepla do vody 2) zdrojem
POSSIBLE USING OF FLOTATION FOR REMOVAL OF PHYTO PLANKTON WITHIN PROCESSING OF DRINKING WATER
 OVĚŘENÍ POUŽITÍ FLOTACE PŘI ODSTRAŇOVÁNÍ FYTOPLANKTONU V PROCESU ÚPRAVY PITNÉ VODY EVA KYNCLOVÁ POSSIBLE USING OF FLOTATION FOR REMOVAL OF PHYTO PLANKTON WITHIN PROCESSING OF DRINKING WATER ABSTRAKT Biologické
OVĚŘENÍ POUŽITÍ FLOTACE PŘI ODSTRAŇOVÁNÍ FYTOPLANKTONU V PROCESU ÚPRAVY PITNÉ VODY EVA KYNCLOVÁ POSSIBLE USING OF FLOTATION FOR REMOVAL OF PHYTO PLANKTON WITHIN PROCESSING OF DRINKING WATER ABSTRAKT Biologické
Projekt SGS Využití nástrojů GIS k vyhodnocení agrárních valů v Českém středohoří
 Projekt SGS Využití nástrojů GIS k vyhodnocení agrárních valů v Českém středohoří RNDr. Iva Machová, Ph.D., Mgr. Ing. Petr Novák, Bc. Markéta Kučerová Druhová skladba dřevin v keřovém patře agrárních valů
Projekt SGS Využití nástrojů GIS k vyhodnocení agrárních valů v Českém středohoří RNDr. Iva Machová, Ph.D., Mgr. Ing. Petr Novák, Bc. Markéta Kučerová Druhová skladba dřevin v keřovém patře agrárních valů
Vliv přístupnosti živina na vegetaci rašelinných okrajů rybníků Třeboňské pánve
 Vliv přístupnosti živina na vegetaci rašelinných okrajů rybníků Třeboňské pánve Jana Navrátilová spoluřešitel: : Michal Hájek Cíle: Zjištění vlivu přístupnosti živin na jednotlivé typy vegetace rašelinných
Vliv přístupnosti živina na vegetaci rašelinných okrajů rybníků Třeboňské pánve Jana Navrátilová spoluřešitel: : Michal Hájek Cíle: Zjištění vlivu přístupnosti živin na jednotlivé typy vegetace rašelinných
Malý test na znalosti odběrových skupin ODPOVĚDI. PT#V/6/2010 Odběry vzorků koupaliště ve volné přírodě
 Malý test na znalosti odběrových skupin ODPOVĚDI PT#V/6/2010 Odběry vzorků koupaliště ve volné přírodě 1) Víte, co je to (velikost několik cm) a jak byste to popsali do odběrového protokolu? Jedná se o
Malý test na znalosti odběrových skupin ODPOVĚDI PT#V/6/2010 Odběry vzorků koupaliště ve volné přírodě 1) Víte, co je to (velikost několik cm) a jak byste to popsali do odběrového protokolu? Jedná se o
Nárostová společenstva řas a sinic ve vodních ekosystémech I. Jana Veselá Přírodovědecká fakulta UK Centrum pro cyanobakterie a jejich toxiny
 Nárostová společenstva řas a sinic ve vodních ekosystémech I. Jana Veselá Přírodovědecká fakulta UK Centrum pro cyanobakterie a jejich toxiny Rozsivky Bacillariophyta, Diatoms, Kieselalgen Kam patří v
Nárostová společenstva řas a sinic ve vodních ekosystémech I. Jana Veselá Přírodovědecká fakulta UK Centrum pro cyanobakterie a jejich toxiny Rozsivky Bacillariophyta, Diatoms, Kieselalgen Kam patří v
Pedagogická činnost Jiří Neustupa
 Pedagogická činnost Jiří Neustupa Výuka v bakalářském a magisterském studijním programu Biologie, v bakalářském studijním programu Molekulární biologie a biochemie organismů a v doktorském studijním programu
Pedagogická činnost Jiří Neustupa Výuka v bakalářském a magisterském studijním programu Biologie, v bakalářském studijním programu Molekulární biologie a biochemie organismů a v doktorském studijním programu
Zákon 114/1992 Sb., o ochraně přírody a krajiny
 Zákon 114/1992 Sb., o ochraně přírody a krajiny Účelem zákona je přispět k udržení a obnově přírodní rovnováhy v krajině, k ochraně rozmanitostí forem života, přírodních hodnot a krás a k šetrnému hospodaření
Zákon 114/1992 Sb., o ochraně přírody a krajiny Účelem zákona je přispět k udržení a obnově přírodní rovnováhy v krajině, k ochraně rozmanitostí forem života, přírodních hodnot a krás a k šetrnému hospodaření
trubicovitá pletivná vláknitá, větvená vláknitá
 ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
Fylogeneze a morfologie bezcévných rostlin 4. praktické cvičení. Přehled pozorovaných objektů
 Fylogeneze a morfologie bezcévných rostlin 4. praktické cvičení ŘÍŠE: Chromista Přehled pozorovaných objektů ODDĚLENÍ: Heterocontophyta (syn. Chromophyta) TŘÍDA: Bacillariophyceae podtřída: Coscinodiscophycidae
Fylogeneze a morfologie bezcévných rostlin 4. praktické cvičení ŘÍŠE: Chromista Přehled pozorovaných objektů ODDĚLENÍ: Heterocontophyta (syn. Chromophyta) TŘÍDA: Bacillariophyceae podtřída: Coscinodiscophycidae
Monitoring odpadních vod z čistírny odpadních vod - vyhodnocení kvality
 ZÁVĚRY MONITORINGŮ ZA ROK 2015 Na základě kapitoly 11.2. Integrovaného povolení Čj: MSK 20001/2008 pro výrobní závod Hyundai Motor Manufacturing Czech s.r.o. (HMMC) tímto zveřejňujeme závěry z monitoringů
ZÁVĚRY MONITORINGŮ ZA ROK 2015 Na základě kapitoly 11.2. Integrovaného povolení Čj: MSK 20001/2008 pro výrobní závod Hyundai Motor Manufacturing Czech s.r.o. (HMMC) tímto zveřejňujeme závěry z monitoringů
Malý test znalostí odběrových skupin - fotografie a živé vzorky
 Malý test znalostí odběrových skupin - fotografie a živé vzorky správné odpovědi, vyhodnocení a komentáře PT#V/8/2014 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 7.6.2014
Malý test znalostí odběrových skupin - fotografie a živé vzorky správné odpovědi, vyhodnocení a komentáře PT#V/8/2014 Odběry vzorků přírodní koupaliště Připravil: Petr Pumann, Státní zdravotní ústav, 7.6.2014
Výsledky ichtyologického průzkumu nádrže Nová Říše v roce 2013
 Výsledky ichtyologického průzkumu nádrže Nová Říše v roce 213 HYDROBIOLOGICKÝ ÚSTAV Biologické centrum AV ČR, v.v.i. Na Sádkách 7 České Budějovice 375 tel.: +42 385 31 262 fax: +42 385 31 248 email: hbu@hbu.cas.cz
Výsledky ichtyologického průzkumu nádrže Nová Říše v roce 213 HYDROBIOLOGICKÝ ÚSTAV Biologické centrum AV ČR, v.v.i. Na Sádkách 7 České Budějovice 375 tel.: +42 385 31 262 fax: +42 385 31 248 email: hbu@hbu.cas.cz
Mnohobuněčné kvasinky
 Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Metodika hodnocení EP silně ovlivněných a umělých vodních útvarů kategorie jezero. RNDr. Jakub Borovec, Ph.D. a kolektiv
 Metodika hodnocení EP silně ovlivněných a umělých vodních útvarů kategorie jezero RNDr. Jakub Borovec, Ph.D. a kolektiv Praha, 5.2.2014 Cíl výzkumného úkolu: metodický postup, který bude: - splňovat požadavky
Metodika hodnocení EP silně ovlivněných a umělých vodních útvarů kategorie jezero RNDr. Jakub Borovec, Ph.D. a kolektiv Praha, 5.2.2014 Cíl výzkumného úkolu: metodický postup, který bude: - splňovat požadavky
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
REVITALIZACE RYBNÍKŮ ŽELEZNÁ STUDNIČKA BIOLOGICKOU CESTOU 2016
 REVITALIZACE RYBNÍKŮ ŽELEZNÁ STUDNIČKA BIOLOGICKOU CESTOU 2016 Zpráva o aplikaci přípravku PTP PLUS na rybníkách v katastru Železné studničky (Mestské lesy v Bratislave). Úvod Začátkem letošního roku jsme
REVITALIZACE RYBNÍKŮ ŽELEZNÁ STUDNIČKA BIOLOGICKOU CESTOU 2016 Zpráva o aplikaci přípravku PTP PLUS na rybníkách v katastru Železné studničky (Mestské lesy v Bratislave). Úvod Začátkem letošního roku jsme
Podnebí a počasí všichni tyto pojmy známe
 Podnebí a počasí všichni tyto pojmy známe Obsah: Podnebí Podnebné pásy Podnebí v České republice Počasí Předpověď počasí Co meteorologové sledují a používají Meteorologické přístroje Meteorologická stanice
Podnebí a počasí všichni tyto pojmy známe Obsah: Podnebí Podnebné pásy Podnebí v České republice Počasí Předpověď počasí Co meteorologové sledují a používají Meteorologické přístroje Meteorologická stanice
Průběh průměrných ročních teplot vzduchu (ºC) v období na stanici Praha- Klementinum
 v období na stanici Praha- Klementinum") Změna klimatu v ČR Trend změn na území ČR probíhá v kontextu se změnami klimatu v Evropě. Dvě hlavní klimatologické charakteristiky, které probíhajícím změnám klimatického systému Země nejvýrazněji podléhají
Změna klimatu v ČR Trend změn na území ČR probíhá v kontextu se změnami klimatu v Evropě. Dvě hlavní klimatologické charakteristiky, které probíhajícím změnám klimatického systému Země nejvýrazněji podléhají
Potravní síť Společenstvo
 Potravní síť Společenstvo Potravní řetězec Predátor 2 Predátor 1 Predátor 3 Herbivor 2 Herbivor 3 Herbivor 4 Herbivor 5 Herbivor 1 Producent 1 Producent 2 Potravní síť potravní síť Topografická potravní
Potravní síť Společenstvo Potravní řetězec Predátor 2 Predátor 1 Predátor 3 Herbivor 2 Herbivor 3 Herbivor 4 Herbivor 5 Herbivor 1 Producent 1 Producent 2 Potravní síť potravní síť Topografická potravní
ROSTLINNÁ BUŇKA A JEJÍ ČÁSTI
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Název lokality Stehelčeves 53,91 41,01 40,92 48,98 89,84 55,06 43,67 Veltrusy 13,82 14,41
 Název lokality 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 Stehelčeves 53,91 41,01 40,92 48,98 89,84 55,06 43,67 Veltrusy 13,82 14,41 Kromě meteorologických podmínek má na koncentrace suspendovaných
Název lokality 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 Stehelčeves 53,91 41,01 40,92 48,98 89,84 55,06 43,67 Veltrusy 13,82 14,41 Kromě meteorologických podmínek má na koncentrace suspendovaných
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
Sinice v koupacích vodách ČR v letech
 Sinice v koupacích vodách ČR v letech 26 216 Petr Pumann, Filip Kothan, Tereza Pouzarová Vodárenská biologie 217 1. 2. 2. 217, Praha Problémy spojené s kvalitou přírodní koupacích vod Zdravotně významné
Sinice v koupacích vodách ČR v letech 26 216 Petr Pumann, Filip Kothan, Tereza Pouzarová Vodárenská biologie 217 1. 2. 2. 217, Praha Problémy spojené s kvalitou přírodní koupacích vod Zdravotně významné
BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA
 MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA GEOGRAFICKÝ ÚSTAV BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA Diplomová práce Jan Kučera Vedoucí práce: Mgr. Daniel Seidenglanz, Ph.D. Brno 2013 Bibliografický záznam Autor:
MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA GEOGRAFICKÝ ÚSTAV BRNO KOMPLEXNÍ DOPRAVNÍ ANALÝZA Diplomová práce Jan Kučera Vedoucí práce: Mgr. Daniel Seidenglanz, Ph.D. Brno 2013 Bibliografický záznam Autor:
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
ON-LINE KVANTIFIKACE SINIC V SUROVÉ VODĚ
 ON-LINE KVANTIFIKACE SINIC V SUROVÉ VODĚ Mgr. ZLATICA NOVOTNÁ Doc. Ing. BLAHOSLAV MARŠÁLEK, CSc. Ing. MARTIN TRTÍLEK Ing. TOMÁŠ RATAJ CENTRUM PRO CYANOBAKTERIE A JEJICH TOXINY, BÚ AVČR Photon System Instrument,
ON-LINE KVANTIFIKACE SINIC V SUROVÉ VODĚ Mgr. ZLATICA NOVOTNÁ Doc. Ing. BLAHOSLAV MARŠÁLEK, CSc. Ing. MARTIN TRTÍLEK Ing. TOMÁŠ RATAJ CENTRUM PRO CYANOBAKTERIE A JEJICH TOXINY, BÚ AVČR Photon System Instrument,
Jaro 2010 Kateřina Slavíčková
 Jaro 2010 Kateřina Slavíčková Obsah: 1. Biologické vědy. 2. Chemie a fyzika v biologii koloběh látek a tok energie. 3. Buňka, tkáně, pletiva, orgány, orgánové soustavy, organismus. 4. Metabolismus. 5.
Jaro 2010 Kateřina Slavíčková Obsah: 1. Biologické vědy. 2. Chemie a fyzika v biologii koloběh látek a tok energie. 3. Buňka, tkáně, pletiva, orgány, orgánové soustavy, organismus. 4. Metabolismus. 5.
Prezentace je určena k seznámení se se základními ekologickými pojmy.
 Registrační číslo projektu: Šablona/číslo materiálu: Jméno autora: Třída/ročník CZ.1.07/1.5.00/34.0996 III/2 VY_32_INOVACE_BI601 Mgr. Lucie Křepelová 3. ročník Datum vytvoření: 10.2.2014 Vzdělávací oblast:
Registrační číslo projektu: Šablona/číslo materiálu: Jméno autora: Třída/ročník CZ.1.07/1.5.00/34.0996 III/2 VY_32_INOVACE_BI601 Mgr. Lucie Křepelová 3. ročník Datum vytvoření: 10.2.2014 Vzdělávací oblast:
ANALYTICKÝ PRŮZKUM / 1 CHEMICKÉ ANALÝZY DROBNÝCH KOVOVÝCH OZDOB Z HROBU KULTURY SE ZVONCOVÝMI POHÁRY Z HODONIC METODOU SEM-EDX
 / 1 ZPRACOVAL Mgr. Martin Hložek TMB MCK, 2011 ZADAVATEL David Humpola Ústav archeologické památkové péče v Brně Pobočka Znojmo Vídeňská 23 669 02 Znojmo OBSAH Úvod Skanovací elektronová mikroskopie (SEM)
/ 1 ZPRACOVAL Mgr. Martin Hložek TMB MCK, 2011 ZADAVATEL David Humpola Ústav archeologické památkové péče v Brně Pobočka Znojmo Vídeňská 23 669 02 Znojmo OBSAH Úvod Skanovací elektronová mikroskopie (SEM)
Ekologický a chemický stav útvarů povrchových vod v MOPO
 Petr Tušil Ekologický a chemický stav útvarů povrchových vod v MOPO Výzkumný ústav vodohospodářský T. G. Masaryka, v.v.i. Podbabská 30/ 2582, 160 00 Praha 6 +420 220 197 111 info@vuv.cz, www.vuv.cz, Pobočka
Petr Tušil Ekologický a chemický stav útvarů povrchových vod v MOPO Výzkumný ústav vodohospodářský T. G. Masaryka, v.v.i. Podbabská 30/ 2582, 160 00 Praha 6 +420 220 197 111 info@vuv.cz, www.vuv.cz, Pobočka
World of Plants Sources for Botanical Courses
 Prostorové šíření organismů Bariéry šíření a prostory pro šíření Bariéry šíření veškeré jevy znesnadňující prostorové šíření nemusí nutně znemožňovat šíření působí jako filtr závisí na prostředí závisí
Prostorové šíření organismů Bariéry šíření a prostory pro šíření Bariéry šíření veškeré jevy znesnadňující prostorové šíření nemusí nutně znemožňovat šíření působí jako filtr závisí na prostředí závisí