rostlin a její využit ití pro produkci nových odrůd
|
|
|
- Bohuslav Müller
- před 9 lety
- Počet zobrazení:
Transkript
1 Základní principy transgenoze rostlin a její využit ití pro produkci nových odrůd Doc. RNDr. Jindřich ich Bříza, B CSc. BC AV ČR, v.v.i. a PřF JU České Budějovice

2 Šlechtění rostlin v neolitu umělý výběr na výsev používána semena z nejlepší ších rostlin na konci 17. st. (po poznání pohlavnosti rostlin) přibyla p druhá metoda křížení ve 20. st. pak mutační šlechtění,, využit ití tkáňových kultur a nakonec genové inženýrstv enýrství nástroje gen. inženýrstv enýrství - molekulárn rní mapování (umožň žňuje přesnp esně sledovat přenos mol. znaku v zastoupení nepřesn esně sledovatelného znaku např.. odolnosti vůčv ůči houb.. onemocnění) - transgenoze (vnáš ášení klonovaných genů do rostlinného genomu)

3 Šlechtění rostlin na základz kladě charakteru použit ité DNA lze rozčlenit genetické modifikace na několik n typů: i) xenogenní modifikace do rostl. genomu včleněna na syntetická DNA ii) transgenní modifikace do genomu včleněna na DNA pocházej zející z různých r často fylogeneticky velmi vzdálených druhů iii) intragenní modifikace do genomu vložen gen pocházej zející ze stejného rostlinného druhu, ale ostatní sekvence (promotor, terminátor) tor) pocházej zejí z druhů jiných, ale navzájem křižitelnýchitelných iv) cisgenní modifikace do genomu integrován n gen (kóduj dující sekvence včetnv etně jejich nativních regul. sekvencí) ) ze stejného druhu nebo druhu křižitelného
 cisgenní modifikace do genomu integrován n gen (kóduj dující sekvence včetnv etně jejich nativních regul.")
4 Agrobacterium tumefaciens nepřímá metoda prostřednictv ednictvím m bakterií rodu Agrobacterium přímá metoda prostřednictv ednictvím m DNA 1) Bakterie Agrobacterium tumefaciens - několik druhů půdních bakterií indukuje morfogenetické změny rostlin (rody Agrobacterium, Rhizobium, Bradyrhizobium a Azorhizobium) - bakterie rodu Rhizobium, Bradyrhizobium a Azorhizobium přeměněné v bakteroidy se stávaj vají trvalou součást stí pletiv kořenových hlízek leguminózn zních rostlin - u rodu Agrobacterium však bylo prokázáno, že e bakterie vnáš ášejí své specifické geny aža do rostlinného genomu

5 Agrobacterium tumefaciens - vnáš ášené geny jsou lokalizovány na bakt. plazmidu nazývaném m Ti (tumor inducing), přenp enášená část plazmidu se označuje jako T-DNA T (transferred( DNA)

6 Agrobacterium tumefaciens - T-DNA plazmidu Ti přinp ináší do rostl. buněk: i) geny pro nové cesty biosyntézy auxinů a cytokininů (dochází tím m k dediferenciaci rostl. buněk, takže transformovaná pletiva rostou jako nediferencované nádory - crown galls)

7 Agrobacterium tumefaciens ii) geny pro syntézu nádorovn dorově specifických látek l opinů případně agrocinopinů (ty slouží pro příslup slušný typ bakterií jako zdroj C, N případně P a energie; jsou vylučov ovány do okolních netransformovaných pletin a do půdy) p - zákl. typy Ti plazmidů jsou oktopinový a nopalinový,, další šími pak leucinamopinový, sukcinamopinový a kukumopinový

8 Agrobacterium tumefaciens rod Agrobacterium má tyto druhy: i) A. tumefaciens (infikuje několik n stovek převp evážně dvoudělo ložných druhů rostlin) ii) A. rhizogenes iii) A. rubi (způsobuje cane galls na maliníku) iv) A. vitis (infikuje vinnou révu) r v) A. radiobacter (nevirulentní,, někdy n za druh nepovažov ován) Prvním m krokem zachycení buněk A. tumefaciens na rostl. buňkách je navázání bakterií prostřednictv ednictvím m specifických bakteriáln lních receptorů typu vitronektinů.. Ty jsou kódovk dovány geny chva, chvb, psca, exoc a att umíst stěnými na bakt. chromosomu.
 Prvním m krokem zachycení buněk A. tumefaciens na rostl.")
9 Agrobacterium tumefaciens Ti plazmid má dva úseky nezbytné pro indukci nádorn dorů: i) T-DNA (vstupuje do rostl. buněk, geny pro vlastní integraci do hostitelské DNA nemá) ii) úsek virulence (vir oblast; obsahuje geny vedoucí k přenosu T-DNA T do rostl. buněk k a její integraci do genomu rostliny) Vir oblast mám délku 35 kb a je tvořena minimáln lně 8 operony vira, virb, virc, virg, vird, vire, virf a virh,, jež kódují zhruba 35 polypeptidů Přenos T-DNA T do rostl. buněk k indukují fenolické látky typu acetosyringonu (mj. acetovanilon, kys. sinapová, kys. lysergová, katechol, kys. ferulová). Tyto látky l jsou produkovány poraněnými nými buňkami většiny v dvoudělo ložných rostlin, ne však v buňkami většiny v rostlin jednodělo ložných.
.")
10 Agrobacterium tumefaciens

11 Agrobacterium tumefaciens Přenos T-DNA T začíná od pravé hraniční 25 bp sekvence, kde mezi 3. a 4. bázíb vznikne zlom v dolním řetězci T-DNA, T a to činností VirD1-VirD2 VirD2 endonukleázov zového komplexu. Do rostl. buňky přechp echází T-DNA v jednořet etězcové formě (T- strand) ) s navázanou molekulou VirD2 na 5` konci a asociovanými molekulami VirE2 po celé své délce (T- komplex).
 ) s navázanou molekulou VirD2 na")
12 Agrobacterium tumefaciens
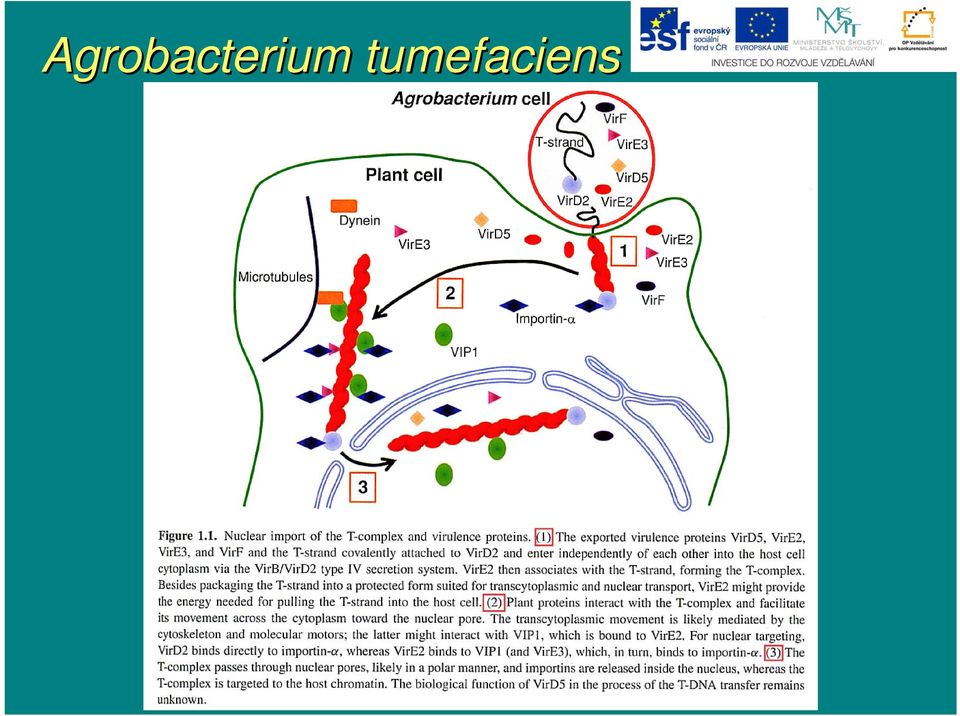
13 Agrobacterium tumefaciens
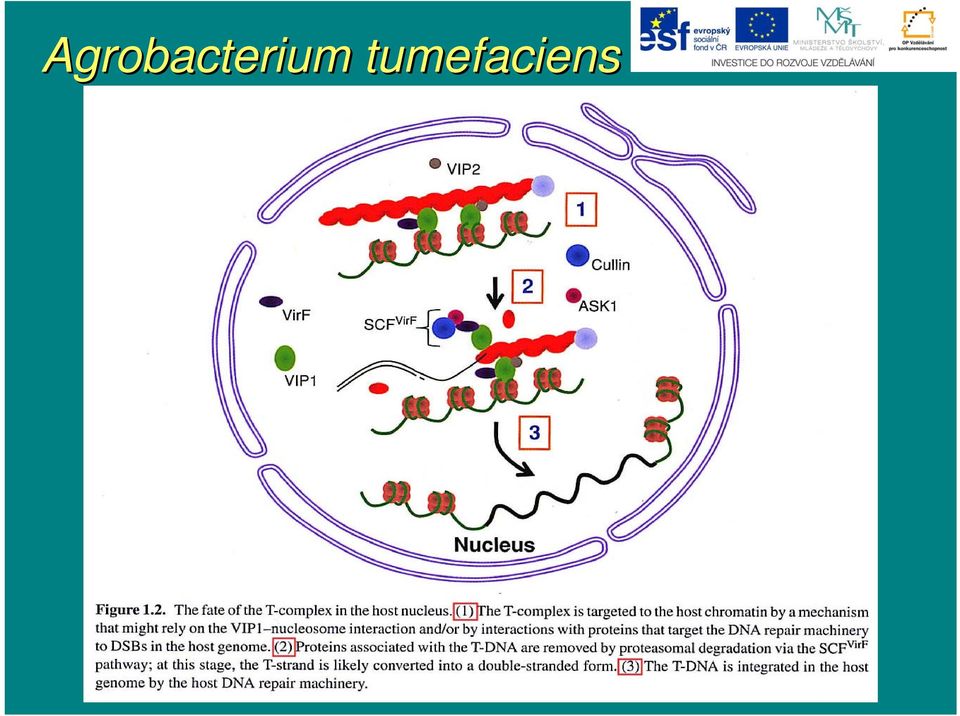
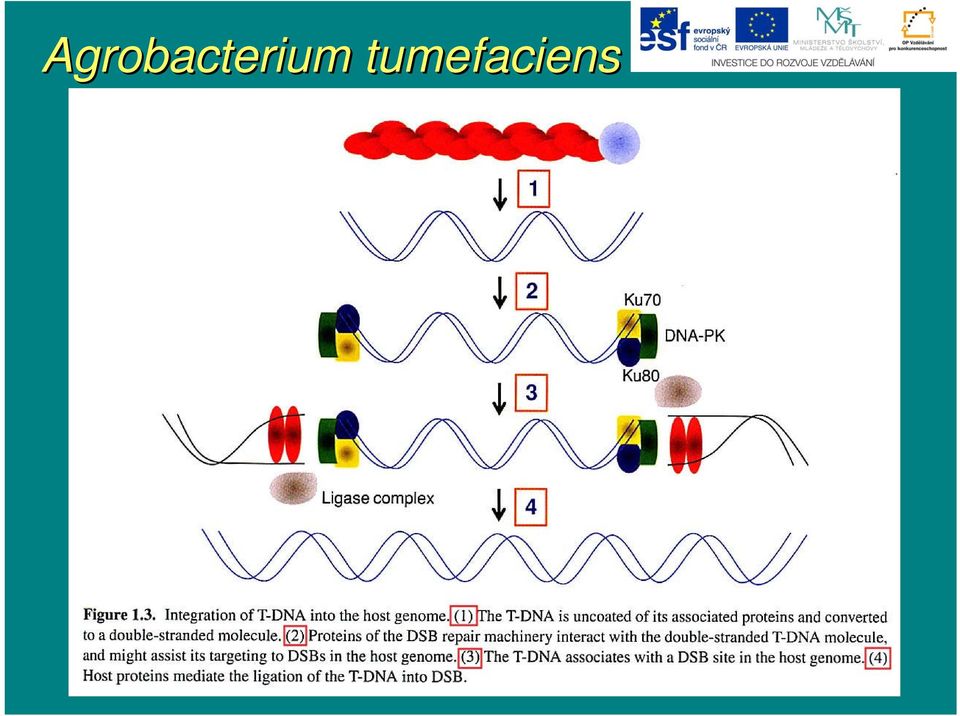
14 Agrobacterium tumefaciens Současn asné znalosti o integraci T-DNA T jsou spojeny do tohoto modelu: i) T-DNA T je zbavena asociovaných proteinů a je konvertována na do dvouřet etězcové molekuly replikačními nástroji hostitelské buňky ii) ) proteiny reparace DSB (Ku70, Ku80 aj.) interagují s dvouřet etězcovou DNA a asistují při i jejím m směrov rování k dvouřet etězcovým zlomům m hostitelského genomu iii) ) v tomto stádiu můžm ůže e zřejmz ejmě také docházet ke spojování několika molekul T-DNA T vedoucí nakonec k mnohačetn etné inserci T-DNAT iv) ) T-DNA T nacházej zející se v místm stě DSB hostitelské buňky je spojena s rostl. genomem ligázovým reparačním komplexem
 ) T-DNA T nacházej zející se v místm stě DSB hostitelské")
15 Agrobacterium tumefaciens
16 Agrobacterium tumefaciens
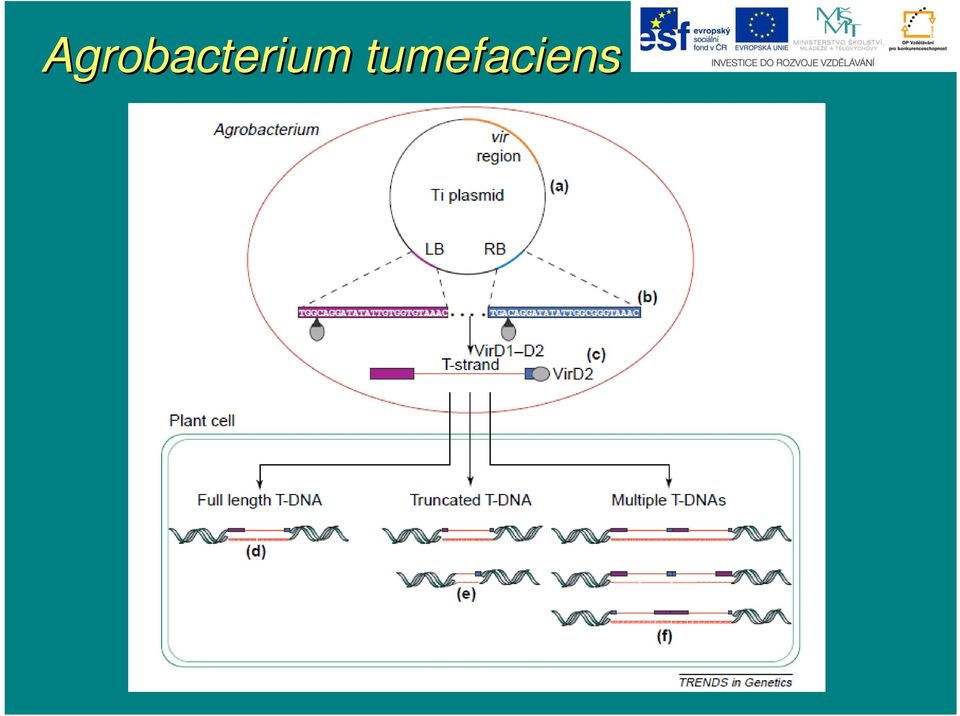
17 Agrobacterium tumefaciens T-DNA plasmidu Ti - nopalinová z jediného úseku - oktopinová z levé a pravé části s meziúsekem sekem,, jenž neobsahuje žádné geny a vstupuje do rostl. genomu fakultativně

18 Agrobacterium tumefaciens - společné geny T-DNA T nopalinových a oktopinových plazmidů Ti tvoří tzv. jádro j (core( core) ) T-DNAT - u oktopinových plazmidů je jádro j tvořeno levým úsekem T-T DNA (T L -DNA) - geny očíslovo slovány podle velikosti jimi kódovank dované mrna Gen 1 (iaam)) kódujek tryptofanmonooxygenázu pro první stupeň nové biosyntetické dráhy auxinů Gen 2 (iaah)) kóduje k indolylacetamidhydrolázu pro druhý stupeň nové syntézy auxinů Gen 3 (ocs)) kóduje k oktopinsyntázu pro syntézu oktopinu z argininu a pyruvátu Gen 4 (ipt)) kóduje k izopentenyltransferázu zu pro první stupeň syntézy cytokininů Gen 5 kóduje produkt účastnící se nové syntézy auxinů
) kóduje k oktopinsyntázu pro syntézu oktopinu z argininu a pyruvátu Gen 4 (ipt)) kóduje k izopentenyltransferázu zu pro první stupeň")
19 Agrobacterium tumefaciens Gen 6a kóduje transportní protein pro sekreci opinů Gen 6b kóduje protein účastnící se nové syntézy cytokininů Gen 7 jeho funkce zatím m neznámá Nopalinová T-DNA mám vlevo od core genů 6 genů (a, b, c, d, e, acs), kde jenom o posledním m je známo, že e kóduje k syntézu agrocinopinů,, funkce ostatních je neznámá. Pravá část oktopinových plasmidů (T R -DNA) obsahuje 5 genů (0` aža 4`). Geny 1` a 2` se podílej lejí na syntéze manopinu,, 0` na syntéze agropinu,, funkce ostatních je zatím m neznámá. T-DNA je vymezena vysoce konzervativními přímými p repeticemi o délce d bp,, jež se nazývají pravá (B R ) a levá hranice (B L ).

20 Agrobacterium rhizogenes 2) Bakterie Agribacterium rhizogenes - vnáš ášené geny jsou lokalizovány na bakt. plazmidu nazývaném Ri (root inducing) - transformovaná pletiva rostou jako kořeny (hairy( roots) - základními typy Ri plazmidů jsou manopinový a agropinový
 - základními typy Ri plazmidů")
21 Agrobacterium rhizogenes - hairy roots jsou schopny proliferace in vitro na médiu m bez růstových látekl - z obou typů transformovaných pletiv (crown( galls, hairy roots) mohou diferencovat transformované nebo i netransformované výhony
22 Agrobacterium rhizogenes - k diferenciaci dochází u hairy roots nikoliv z kořenových špiček, ale ze starší části kořene - téměř všechny kmeny A. rhizogenes mají 3 komponenty Ri plazmidu: i) malý (Mr( Mr~90 MD) s geny pro utilizaci opinů ii) ) velký (Mr( Mr~150 MD) s další šími geny pro využit ití opinů a pro využit ití agrocinopinů iii) kointegrovaný vzniklý spojením m předchp edcházejícíchch - T-DNA manopinových plazmidů je tvořena jedním úsekem, agropinových dvěma (T L a T R ) úseky oddělených segmentem, který do rostlinných buněk k nevstupuje - geny na T L -DNA: celkem asi 18, z nichž 4 (rola( rola, rolb, rolc, rold) ) podrobně prostudovány působí zvýšen ení senzitivity k auxinům
23 Agrobacterium rhizogenes - geny na T R -DNA: nejdůle ležitější jsou geny pro syntézu auxinů, homologní s geny 1 (iaam( iaam) ) a 2 (iaah( iaah) ) v T-DNA T A. tumefaciens, a geny pro syntézu agropinu a nedávno objeven analog genu rolb z T L -DNA
24 Vektory Pro využit ití bakterií Agrobacterium jako vektorů transgenů je třeba odstranit původnp vodní geny a místo m nich (tj. mezi hraniční sekvence T-DNA) T vložit geny, které chceme do rostl. genomu integrovat. A) Intermediárn rní (kointegrativní)) vektory - intermediárn rní vektor je malý plazmid s krátkými úseky s levou a pravou hraniční sekvencí,, mezi nimiž jen vložen zájmový gen a gen selekční - je schopen replikace v E. coli,, nikoliv však v v agrobakteriu - po vnesení do agrobakteria (transformací nebo konjugací) ) se trvale udrží jen v buňkách, ve kterých došlo k rekombinaci mezi homologními mi sekvencemi Ti plazmidu a intermediárn rního vektoru
25 Vektory
26 Vektory
27 Vektory B) Binárn rní vektory - jejich konstrukce založena na skutečnosti, že e schopnost indukovat nádor n mám i Ti plazmid,, který je rozdělen na dva menší ší: i) jeden obsahuje úsek virulence, ale nikoliv T-DNA, T a počátek replikace pro agrobakterium (Ti plazmid zbavený T-DNA) T ii) ) druhý obsahuje T-DNA T a počátek replikace pro agrobakterium (vektorový plazmid); mám vhodná restrikční místa pro inzerci cizorodé DNA - příkladem kmene A. tumefaciens nesoucí Ti plazmid bez T-T DNA je kmen LBA4404, jenž obsahuje plazmid zvaný pal4404, odvozený od Ti plazmidu ptiach5 delecí T- DNA úseku
28 Vektory
29 Vektory - jedním m z prvních binárn rních vektorů byl v r pbin19, který byl v r vylepšen en doplněním m o reportérový rový gen za vzniku pbi121 - tyto dva vektory a jejich deriváty jsou stále nejpopulárn rnější (v letech byly použity v pracech zabývajících ch se transformací vyšší šších rostlin ve 40 % případp padů)
30 Vektory
31 Vektory - za posledních 20 let byla vyvinuta řada systémů binárn rních vektorů,, ovšem díky d omezení velikosti vkládan dané DNA nesou vedle selekčního genu jen geny zájmovz jmové - toto omezení je dáno d monocistronickou expresí eukaryotních ORF, což vede ke značné velikosti vkládan dané DNA a komplikuje klonovací postupy - v r byl publikován n modulárn rní vektorový systém psat, který umožň žňuje vložen ení až 7 ORF do jedné T-DNA a jež je založen na tzv. homing (naváděcích) ch) endonukleázách ch; ; ty rozpoznávaj vají velmi raritní sekvence (např.. I-SceI SceI 18 bázovou b sekvenci, která se teor.. vyskytuje každých 7 x bp) - binárn rní vektor obsahuje 6 různých r raritních sekvencí a přes p různé expresní kazety psat jsou do nich bez komplikací vkládány jednotlivé zájmové geny
32 Vektory
33 Vektory C) Superbinárn rní vektory - vektory, které vedle T-DNA T obsahují ještě některé z genů vir oblasti - využívá se poznatku, že e některn které z produktů vir genů vykazují dávkový efekt, tj. čím m více v je těchto t produktů v buňce přítomno, tím t m většív je účinnost transformace - tyto vektory hrály v pol let stěž ěžejní úlohu při p i rozšiřov ování spektra druhů transformovatelných agrobakteriem o obiloviny
34 Nepřímá transformace používá se agrobakterium s vhodným binárn rním (superbinárním)) vektorem z akceptorové rostliny se používaj vají různé orgány resp. pletiva: listové disky, dělod ložní listy, stonky explantát se kultivuje s bakteriemi agrobakteria v řádech minut aža dní a pak se přenese p na médium m obsahující selekční agens, antibiotikum pro eliminaci agrobakteria a růstové látky (RL) v poměru vedoucím m k organogenezi (vzniku prýtů) na selekčním m médiu m bez RL pak prýty zakořen ení a obvykle ponesou stabilně integrovaný transgen (ověř ěření PCR a Southernovou hybridizací)
35 Nepřímá transformace
36 Nepřímá transformace při i transientní (dočasné) in planta expresi dojde k případnp padné expresi v rostl. buňce během b několika n hodin aža dní a záhy odezní ověř ěřování funkčnosti nosti konstruktů,, rychlá jednorázov zová produkce velkého množstv ství produktu jedna z metod pro indukci transientní exprese využívá agrobakterium agroinfiltrace - tekutá kultura agrobakteria s binárn rním m vektorem se vakuově nebo pomocí inj.. stříka kačky ky infiltruje do listu
37 Nepřímá transformace další metody pro transientní expresi: - mikroprojektily zlata nebo wolframu o velikosti zhruba 1 μm m s navázanou plazmidovou DNA; vnáš ášejí se buď ručním m přístrojem p Helios Gene Gun přímo na živé rostlině nebo do oddělených explantátů ve vakuové komoře e (PDS-1000 He); limitující je předevp edevším m vysoká vstupní investice
38 Nepřímá transformace další metody pro transientní expresi: - virové vektory,, v nichž je zájmový z gen vložen do virového genomu; ; obzvláš áště účinný je sytém magnifection,, ve kterém m virové replikony nesoucí zájmový gen jsou naklonovány ny do T-DNA T a do rostliny jsou vnáš ášeny již zmíněnou nou agroinfiltrací; ; během b množen ení viru je zájmový z gen exprimován spolu s virovými geny, což vede k masivní produkci požadovan adovaného produktu
39 Přímá transformace různé metody začaly aly být vyvíjeny jako alternativa k agrobakteriové transgenozi pro druhy, u nížn Agrobacterium nefungovalo jako první vznikla metoda transformace protoplastů (PP) - omezeno na druhy se zvládnutou regenerací PP - DNA přijp ijímána endocytózou zou,, k její stimulaci lze použít t buď PEG nebo elektroporaci - DNA lze do jader PP vložit mikroinjekcemi biolistika (biologická balistika listika) - jde o bombardování buněk k nebo pletiv mikropartikulemi (~1 μm) zlata nebo wolframu, na nichž je navázan zaná plazmidová DNA
40 Přímá transformace - jen % mikroprojektilů pronikne alespoň do epidermis, ale je třeba t proniknout aža do buněk k mezofylu; pokud se tak stane, jen u 5 % z těchto t buněk k se projektil dostane až do jádra, j ale většina v ( %) těchto t buněk k po zásahu stejně odumírá - pokud se mikroprojektil s DNA dostane aža do jádra j a buňka zásah přežije, p je frekvence integrece vnesené DNA do jaderné DNA (alespoň) ) u tabáku značně vysoká (u tabáku integrací na jeden výstřel - 0,5 mg Au s 0,8 μg g DNA) vsakování roztoku DNA dehydratovanými rostl. pletivy - metoda stále studována a optimalizována (není pro rutinní transgenozi)
41 Přímá transformace přenos zprostředkovaný liposomy (lipofekce) - uvnitř liposomů roztok DNA, aplikováno na zralá pylová zrna, lze použít t na PP - úspěšných prací poskrovnu přenos zprostředkovaný alginátovými mikrokapičkami kami - umožň žňuje přenos p velmi velkých fragmentů DNA do PP, a to navíc c s řádově větší transformační účinností než PEG
42 Přímá transformace technologie transformace pylu i) biolistika jednobuněč ěčného pylu - jednobuněč ěčné mikrospory jsou bombardovány v G1 fázi, f po dozrání mikrospor je pyl použit k opylení in vivo a semena testována na selekčním m médium
43 Přímá transformace ii) biolistika mikrospor nebo nezralých pylových zrn reprogramovaných stresem k tvorbě sporofytu - embryogenní mikrospory či i nezralá pylová zrna jsou transformována na a vzniklé embryogenní buňky dávajd vají vznik haploidnímu embryu a rostlinám m (diploidizace( spontánn nně či chem.. indukována) na)
44 Cisgenoze Cisgenoze je definována na jako přenos p genů s jejich nativními regulačními sekvencemi metodami genového inženýrstv enýrství z druhu přirozenp irozeně křižitelného nebo z druhu téhot hož. Jde téžt o vytvořen ení nových transformačních protokolů bez použit ití (bakteriáln lních) selekčních markerů u generativně množených druhů kotransformací,, u vegetativně množených druhů odstranitelnými selekčními geny nebo použit itím m vektorů bez markerových genů (marker-gene-free vectors). Díky tomu by na takto geneticky upravené rostliny nemuselo být nahlíženo jako na GMO, ale jako na klasickými metodami vyšlecht lechtěné odrůdy dy se všemi v legislativními dopady a s příznivým míněním m m veřejnosti. ejnosti.
45 Cisgenoze V praxi bude cisgenoze vhodná zvláš áště pro rezistentní šlechtění vegetativně množených vysoce heterozygotních plodin (brambory, jabloně), kde zdrojem genů rezistence jsou příbuzné plané druhy.
46 Využit ití GM plodin GM crops are not the problem, but part of the solution to sustainably feeding 9 billion people. (Professor Giles Oldroyd,, Senior Plant Scientist,, John Innes Centre, UK) Do roku 2030 je třeba t celosvětov tově zvýšit výrobu potravin nejméně o 50 %, což se nedá zvládnout pouze klasickými šlechtitelskými postupy. Genetické modifikace budou tedy muset být jednou z technologií,, jak se s tímto t požadavkem úspěšně vyrovnat.
47 Využit ití GM plodin Přes legislativní problémy v určitých částech světa plocha GM plodin stále roste, protože e přinp inášejí vyšší zisk pěstitelp stitelům m a vyšší užitnou hodnotu spotřebitel ebitelům.
48 Využit ití GM plodin Drtivá většina v současnosti v praxi využívaných vaných GM plodin nese geny tolerance k herbicidům m (HT) nebo geny pro tvorbu δ-endotoxinu z B. thuringiensis (BT).
49 Využit ití GM plodin Z hlediska plodin vede sója s následovann sledovaná kukuřic icí a bavlníkem.
50 Využit ití GM plodin Podíl l nejrozší šířenějších GM plodin na jejich celkové světov tové produkci:
51 Využit ití GM plodin Přehled zemí s povoleným polním m pěstovp stováním m viz obr.:
52 Využit ití GM plodin Přicházející GM plodiny: i) kukuřice s genem pro fytázu (Čína) podstatná část fosforu ve zralých obilovinách a olejninách je obsažena ve formě fytátu tu; ; vzhledem k tomu, že monogastrická zvířata mají málo střevn evní fytázy zy,, je jejich využit ití fosforu z obilovin a olejnin omezené a fytát,, který není zvířaty využit, je vylučov ován n v hnoji a způsobuje problémy se znečištěním m prostřed edí; ; kyselina fytová dále můžm ůže e snížit biologickou dostupnost další ších prvků Ca a Zn ii) kukuřice se sníženým obsahem fytátu tu v semeni (USA, Pioneer Hi-Bred International) iii) Bt rýže e se zvýšenou rezistencí vůči i zavíje ječům m (Čína)( iv) SmartStax kukuřice (Monsanto( Monsanto,, 2010) nese 8 různých r transgenů (2 pro rezistenci k glyfosátu resp. glufosinátu tu) ) a 6 genů BT směrovaných proti různým r živočišným škůdcům
53 Využit ití GM plodin v) kukuřice se zvýšenou odolností vůči i suchu (Monsanto( Monsanto); očekávané tržní zavedení 2012 vi) geny zlepšuj ující nutriční vlastnosti plodin sója se zvýšeným obsahem 3-ω3 mastné kyseliny - kyseliny stearidonové (Monsanto), která je stabilnější než rybí tuk a v lidském či i zvířec ecím m těle t je konvertována na na polynenasycené 3-ω mastné kyseliny s příznivým p účinkem na vaskulárn rní systém vii) zlatá rýže e 2 2 s velmi vysokým obsahem β-karoténu bude zavedena 2012
54 Využit ití GM plodin Příklad vývoje GM plodin firmou Monsanto
55 Využit ití GM plodin
56 Využit ití GM plodin
57 Využit ití GM plodin
58 Využit ití GM plodin
59 Využit ití GM plodin Perspektivní GM plodiny: i) rajčata ata s genem Bs2 z papriky pro rezistenci vůčv ůči Xanthomonas sp.. v polních podmínk nkách ii) rajčata ata rezistentní vůči Phytophtora infestans s geny rezistence z planých druhů rajčat at iii) rajčata ata se zvýšenou odolností k bakteriáln lním m chorobám m (vč. Ralstonia solanacearum) ) s genem efr z arabidopsis iv) banánovn novník k je nyní devastován n chorobou sigatoga způsobovanou houbou Mycosphaerella fijiensis téměř vešker kerá světov tová produkce banánů je tvořena extrémn mně citlivou odrůdou dou Cavendish; žádný GM rezistentní banánovn novník ještě není hotov, ale několik n nadějných postupů je testováno
60 Využit ití GM plodin Perspektivní GM plodiny: v) rajčata ata s výrazně vyšší šším m (3x) obsahem antioxidantů (antokyanů a flavonolů) ) díky d expresi dvou genů pro transkripční faktory z hledíku (purple tomato)
61 Využit ití GM plodin Perspektivní GM plodiny: vi) zvýšen ení účinnosti příjmu p N rostlinami k omezení hnojení a znečišťov ování povrchových i spodních vod alaninaminotransferasa je slibným kandidátem; v delší ším horizontu je žádoucí vytvořit GM plodin se schopností vytvořit symbiotický vztah s dusík k fixujícími bakteriemi leguminóz vii) zavedení rezistence vůčv ůči i rzi trávn vní (Puccinia graminis) ) do pšenice; geny rezistence jsou dostupné v planých druzích, ze kterých budou klonovány ny a vneseny do kultivarů pšenice
62 Důvody využit ití GM plodin umožní nutné zvýšen ení světov tové produkce potravin přes p pokles rozlohy obdělávateln vatelné půdy umožní produkci potravin na půdách p dosud pro zemědělskou produkci nevhodných (sucho, zasolení) redukují environmentáln lní dopady rostlinné zemědělsk lské výroby snižov ováním m počtu pesticidních ch ošeto etření a tím t snižov ováním m potřeby fosilních paliv na jednotku produkce redukují spotřebu paliv díky d aplikaci úspornějších metod obdělávání půdy zlepšuj ují nutriční hodnotu potravin a krmiv zlepšuj ují vlastnosti surovin pro průmyslov myslové využit ití umožní zvýšit produkci rostl. hmoty jakožto obnovitelného zdroje energie
Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme
 Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme Doc. RNDr. Jindřich Bříza, CSc. ÚMBR BC AV ČR,v.v.i. & katedra genetiky PřF JU Branišovská 31, 370 05 České Budějovice GM crops
Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme Doc. RNDr. Jindřich Bříza, CSc. ÚMBR BC AV ČR,v.v.i. & katedra genetiky PřF JU Branišovská 31, 370 05 České Budějovice GM crops
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
GENETICKY MODIFIKOVANÉ ORGANISMY. Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN
 GENETICKY MODIFIKOVANÉ ORGANISMY Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN VERTIKÁLNÍ PŘENOS VLASTNOSTÍ DĚDIČNOST považoval člověk za samozřejmou zákonitost Evoluce
GENETICKY MODIFIKOVANÉ ORGANISMY Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN VERTIKÁLNÍ PŘENOS VLASTNOSTÍ DĚDIČNOST považoval člověk za samozřejmou zákonitost Evoluce
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin Geneticky modifikované rostliny v zemědělské praxi - přínosy a rizika Bakalářská práce Vedoucí práce: Ing. Tomáš
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin Geneticky modifikované rostliny v zemědělské praxi - přínosy a rizika Bakalářská práce Vedoucí práce: Ing. Tomáš
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Geneticky modifikované organismy
 Geneticky modifikované organismy Ivo Frébort KBC/BAM Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
Geneticky modifikované organismy Ivo Frébort KBC/BAM Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
Modifikace dědičné informace rostlin
 Modifikace dědičné informace rostlin Lukáš Fischer, KFR PřF UK Jak zlepšit vlastnosti rostlin Principy a klasické způsoby přípravy geneticky modifikovaných rostlinných buněk a celých rostlin Genový přenos
Modifikace dědičné informace rostlin Lukáš Fischer, KFR PřF UK Jak zlepšit vlastnosti rostlin Principy a klasické způsoby přípravy geneticky modifikovaných rostlinných buněk a celých rostlin Genový přenos
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316
 Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Tradice šlechtění šlechtění zlepšování pěstitelsky, technologicky a spotřebitelsky významných vlastností
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Tradice šlechtění šlechtění zlepšování pěstitelsky, technologicky a spotřebitelsky významných vlastností
Transformace ptdna tabáku genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace
 Transformace ptdna tabáku fúzním genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace Jiřich ich BřízaB 1,, Josef Vlasák 1, Štěpán n Ryba, Viera Ludvíkov ková 3, Hana Niedermeierová
Transformace ptdna tabáku fúzním genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace Jiřich ich BřízaB 1,, Josef Vlasák 1, Štěpán n Ryba, Viera Ludvíkov ková 3, Hana Niedermeierová
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Transgenoze a reverzní genetika. Metody transformace rostlinných buněk Rekombinace
 Transgenoze a reverzní genetika Metody transformace rostlinných buněk Rekombinace Využití transgenoze 2 Mutageneze (ztráta funkce) Využití transgenoze 3 Charakterizace promotoru na základě exprese reportérového
Transgenoze a reverzní genetika Metody transformace rostlinných buněk Rekombinace Využití transgenoze 2 Mutageneze (ztráta funkce) Využití transgenoze 3 Charakterizace promotoru na základě exprese reportérového
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém
 Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Modifikace dědičné informace rostlin I. modifikace
 Modifikace dědičné informace rostlin I Klasická genetická modifikace Lukáš Fischer, KEBR Legislativa: Genetická modifikace (GM) = vnesení genetické informace (úseku DNA) či změna > 20 nt způsobem, který
Modifikace dědičné informace rostlin I Klasická genetická modifikace Lukáš Fischer, KEBR Legislativa: Genetická modifikace (GM) = vnesení genetické informace (úseku DNA) či změna > 20 nt způsobem, který
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Metodický list č. 1. TÉMA: Ekologicky šetrné zemědělství PĚSTOVÁNÍ ROSTLIN. Ochrana krajiny
 32 TÉMA: Cíl: uvědomit si vazby mezi zemědělstvím, přírodou a životním prostředím, seznámit žáky s prioritami současné zemědělské výroby v souladu s ochranou životního prostředí Základní pojmy: meliorace,
32 TÉMA: Cíl: uvědomit si vazby mezi zemědělstvím, přírodou a životním prostředím, seznámit žáky s prioritami současné zemědělské výroby v souladu s ochranou životního prostředí Základní pojmy: meliorace,
Sněť kukuřičná - nejrozšířenější choroba kukuřice. Ustilago maydis (DC.) Corda 1842
 Corda 1842") Sněť kukuřičná - nejrozšířenější choroba kukuřice Ustilago maydis (DC.) Corda 1842 Úvod Houbový patogen Ustilago maydis (DC.) Corda je původcem sněti kukuřičné, která je dnes celosvětově nejrozšířenější
Sněť kukuřičná - nejrozšířenější choroba kukuřice Ustilago maydis (DC.) Corda 1842 Úvod Houbový patogen Ustilago maydis (DC.) Corda je původcem sněti kukuřičné, která je dnes celosvětově nejrozšířenější
Zaměření bakalářské práce (témata BP)
") Zaměření bakalářské práce (témata BP) Obor: Buněčná a molekulární diagnostika - zadává katedra - studenti si témata losují Obor: molekulární biologie a genetika - témata BP vychází z vybraného tématu DP
Zaměření bakalářské práce (témata BP) Obor: Buněčná a molekulární diagnostika - zadává katedra - studenti si témata losují Obor: molekulární biologie a genetika - témata BP vychází z vybraného tématu DP
Nové směry v rostlinných biotechnologiích
 Nové směry v rostlinných biotechnologiích Tomáš Moravec Ústav Experimentální Botaniky AV ČR Praha 2015-05-07 Praha Prvních 30. let transgenních rostlin * V roce 2014 byly GM plodiny pěstovány na ploše
Nové směry v rostlinných biotechnologiích Tomáš Moravec Ústav Experimentální Botaniky AV ČR Praha 2015-05-07 Praha Prvních 30. let transgenních rostlin * V roce 2014 byly GM plodiny pěstovány na ploše
Asociace půdních MO s cévnatými rostlinami
 Asociace půdních MO s cévnatými rostlinami Fylosféra Sphingomonas, Xantomonas Rizosféra+rizoplan -růst stimulují Pseudomonas, Achromobacter (auxiny, CK, gibereliny, glykolipidy) -dusík fixují volně žijící
Asociace půdních MO s cévnatými rostlinami Fylosféra Sphingomonas, Xantomonas Rizosféra+rizoplan -růst stimulují Pseudomonas, Achromobacter (auxiny, CK, gibereliny, glykolipidy) -dusík fixují volně žijící
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
GENETICKY MODIFIKOVANÉ
 GENETICKY MODIFIKOVANÉ ROSTLINY (GMR) Lukáš Fischer Katedra experimentální biologie rostlin PřF UK Geny základ vlastností organismů Změny genetické informace rostlin a definice genetické modifikace dle
GENETICKY MODIFIKOVANÉ ROSTLINY (GMR) Lukáš Fischer Katedra experimentální biologie rostlin PřF UK Geny základ vlastností organismů Změny genetické informace rostlin a definice genetické modifikace dle
Havarijní plán PřF UP
 Havarijní plán PřF UP v němž se nakládá s geneticky modifikovanými organismy (GMO), zpracovaný podle 20, odst. 4 zákona č. 78/2004 Sb. pro pracoviště kateder Buněčné biologie a genetiky a Oddělení molekulární
Havarijní plán PřF UP v němž se nakládá s geneticky modifikovanými organismy (GMO), zpracovaný podle 20, odst. 4 zákona č. 78/2004 Sb. pro pracoviště kateder Buněčné biologie a genetiky a Oddělení molekulární
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Krása TC. mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze.
 Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze. Kalusové kultury, protokormy. I když kalusové kultury nejsou metodou běžnou při rozmnožování
Krása TC mé zkušenosti s technikami mikropropagace v obrazech. Nepřímá organogeneze, nepřímá somatická embryogeneze. Kalusové kultury, protokormy. I když kalusové kultury nejsou metodou běžnou při rozmnožování
Kyselina hyaluronová. Kyselina hyaluronová. Streptococcus equi subsp. produkovaná kyselina hyaluronová a. Autor prezentace: Mgr.
 Kyselina hyaluronová Streptococcus equi subsp. zooepidemicus a jím produkovaná kyselina hyaluronová a glukuronidáza Marcela Tlustá Biotechnologická laborato Meyer a Palmer, 1934 Extracelulární matrix,
Kyselina hyaluronová Streptococcus equi subsp. zooepidemicus a jím produkovaná kyselina hyaluronová a glukuronidáza Marcela Tlustá Biotechnologická laborato Meyer a Palmer, 1934 Extracelulární matrix,
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Geneticky modifikované potraviny a krmiva
 Geneticky modifikované potraviny a krmiva Co je to geneticky modifikovaný organismus (GMO)? Za GMO je považován organismus, s výjimkou člověka, jehož dědičná informace uložená v DNA byla změněna pomocí
Geneticky modifikované potraviny a krmiva Co je to geneticky modifikovaný organismus (GMO)? Za GMO je považován organismus, s výjimkou člověka, jehož dědičná informace uložená v DNA byla změněna pomocí
Transformace rostlin. úvod metody
 Transformace rostlin úvod metody Biotechnologie metody využívající různé organismy ve prospěch člověka a současně pozměňující jejich vlastnosti tak, aby byly co nejlépe využitelné výrobní postupy: získávání
Transformace rostlin úvod metody Biotechnologie metody využívající různé organismy ve prospěch člověka a současně pozměňující jejich vlastnosti tak, aby byly co nejlépe využitelné výrobní postupy: získávání
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Organizace a kontrola pěstování GM plodin v ČR. Ing. Jana Trnková MZe, odbor rostlinných komodit
 Organizace a kontrola pěstování GM plodin v ČR Ing. Jana Trnková MZe, odbor rostlinných komodit Geneticky modifikované plodiny GM plodiny, transgenní rostliny změněn dědičný materiál (DNA) pomocí genových
Organizace a kontrola pěstování GM plodin v ČR Ing. Jana Trnková MZe, odbor rostlinných komodit Geneticky modifikované plodiny GM plodiny, transgenní rostliny změněn dědičný materiál (DNA) pomocí genových
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
HYCOL. Lis tová hno jiva. HYCOL-Zn kulturní rostliny. HYCOL-Cu kulturní rostliny. HYCOL-E OLEJNINA řepka, slunečnice, mák
 Lis tová hno jiva n e j ž e n e... víc HYCOL do e kol o g ic ké p ro d u kce BIHOP-K+ HYCOL-BMgS HYCOL-NPK chmel, kukuřice, mák HYCOL-E OBILNINA řepka, slunečnice, mák zelenina, slunečnice pšenice, ječmen,
Lis tová hno jiva n e j ž e n e... víc HYCOL do e kol o g ic ké p ro d u kce BIHOP-K+ HYCOL-BMgS HYCOL-NPK chmel, kukuřice, mák HYCOL-E OBILNINA řepka, slunečnice, mák zelenina, slunečnice pšenice, ječmen,
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Rostliny a lidstvo. Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin
 Rostliny a lidstvo Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin Čím se živíme Čím se živíme Čím se živíme Čím se živíme Čím se živíme
Rostliny a lidstvo Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin Čím se živíme Čím se živíme Čím se živíme Čím se živíme Čím se živíme
R o z h o d n u t í. rozhodlo
 Ministerstvo životního prostředí odbor environmentálních rizik Praze dne 30. dubna 2007 Čj.: 4675/ENV/07 R o z h o d n u t í Ministerstvo životního prostředí jako správní úřad příslušný podle 5 zákona
Ministerstvo životního prostředí odbor environmentálních rizik Praze dne 30. dubna 2007 Čj.: 4675/ENV/07 R o z h o d n u t í Ministerstvo životního prostředí jako správní úřad příslušný podle 5 zákona
Použití transgenoze při šlechtění rostlinje třeba se obávat?
 Poslanecká sněmovna parlamentu ČR 3. května 2017 Použití transgenoze při šlechtění rostlinje třeba se obávat? Mgr. Tomáš Moravec, PhD., Ústav Experimentální Botaniky AV ČR Laboratoř virologie Praha Modifikování
Poslanecká sněmovna parlamentu ČR 3. května 2017 Použití transgenoze při šlechtění rostlinje třeba se obávat? Mgr. Tomáš Moravec, PhD., Ústav Experimentální Botaniky AV ČR Laboratoř virologie Praha Modifikování
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Rostlinné orgány. Kořen (radix)
") - jsou tvořeny soubory pletiv - vyznačují se určitou funkcí a stavbou Rostlinné orgány Rostlinné orgány vegetativní (vyživovací) kořen, stonek, list - funkce : zajištění výživy, růstu a výměny látek s
- jsou tvořeny soubory pletiv - vyznačují se určitou funkcí a stavbou Rostlinné orgány Rostlinné orgány vegetativní (vyživovací) kořen, stonek, list - funkce : zajištění výživy, růstu a výměny látek s
KOMISE EVROPSKÝCH SPOLEČENSTVÍ ZPRÁVA KOMISE EVROPSKÉMU PARLAMENTU A RADĚ. o odvětví lnu a konopí {SEK(2008) 1905}
 1905}") CS CS CS KOMISE EVROPSKÝCH SPOLEČENSTVÍ V Bruselu dne 20.5.2008 KOM(2008) 307 v konečném znění ZPRÁVA KOMISE EVROPSKÉMU PARLAMENTU A RADĚ o odvětví lnu a konopí {SEK(2008) 1905} CS CS ZPRÁVA KOMISE EVROPSKÉMU
CS CS CS KOMISE EVROPSKÝCH SPOLEČENSTVÍ V Bruselu dne 20.5.2008 KOM(2008) 307 v konečném znění ZPRÁVA KOMISE EVROPSKÉMU PARLAMENTU A RADĚ o odvětví lnu a konopí {SEK(2008) 1905} CS CS ZPRÁVA KOMISE EVROPSKÉMU
Obsah 5. Obsah. Úvod... 9
 Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce
 BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 09 Genetické modifikace pro zlepšení výţivy člověka doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Nový trend zlepšení výţivy lidí Výţiva a zdraví člověka Prevence
Bi8240 GENETIKA ROSTLIN Prezentace 09 Genetické modifikace pro zlepšení výţivy člověka doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Nový trend zlepšení výţivy lidí Výţiva a zdraví člověka Prevence
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010. M. Mendlová, V. Ostrý, J. Ruprich
 STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010 M. Mendlová, V. Ostrý, J. Ruprich Státní zdravotní ústav v Praze Centrum zdraví, výživy a potravin Oddělení analýzy
STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010 M. Mendlová, V. Ostrý, J. Ruprich Státní zdravotní ústav v Praze Centrum zdraví, výživy a potravin Oddělení analýzy
Monitorování hladiny metalothioneinu a thiolových sloučenin u biologických organismů vystavených působení kovových prvků a sloučenin
 Laboratoř Metalomiky a Nanotechnologií Monitorování hladiny metalothioneinu a thiolových sloučenin u biologických organismů vystavených působení kovových prvků a sloučenin Ing. Kateřina Tmejová, Ph. D.,
Laboratoř Metalomiky a Nanotechnologií Monitorování hladiny metalothioneinu a thiolových sloučenin u biologických organismů vystavených působení kovových prvků a sloučenin Ing. Kateřina Tmejová, Ph. D.,
Hodnocení rizik geneticky modifikovaných rostlin pro životní prostředí
 Klasifikace: Draft Pro vnitřní potřebu VVF Oponovaný draft Pro vnitřní potřebu VVF Finální dokument Pro oficiální použití Deklasifikovaný dokument Pro veřejné použití Název dokumentu: Hodnocení rizik geneticky
Klasifikace: Draft Pro vnitřní potřebu VVF Oponovaný draft Pro vnitřní potřebu VVF Finální dokument Pro oficiální použití Deklasifikovaný dokument Pro veřejné použití Název dokumentu: Hodnocení rizik geneticky
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
POŽADAVKY NA KVALITU SUROVIN PRO WELLNESS GASTRONOMII
 POŽADAVKY NA KVALITU SUROVIN PRO WELLNESS GASTRONOMII Měli bychom dbát nejen na nutriční hodnotu stravy, ale i na mikrobiologickou a chemickou čistotu! V současnosti se celkem dbá na základní hygienické
POŽADAVKY NA KVALITU SUROVIN PRO WELLNESS GASTRONOMII Měli bychom dbát nejen na nutriční hodnotu stravy, ale i na mikrobiologickou a chemickou čistotu! V současnosti se celkem dbá na základní hygienické
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Univerzita Karlova v Praze Přírodovědecká fakulta. Katedra experimentální biologie rostlin
 Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Studijní program: Chemie Studijní obor: Chemie a biologie se zaměřením na vzdělávání Julie Andrová TRANSGENNÍ PLODINY
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Studijní program: Chemie Studijní obor: Chemie a biologie se zaměřením na vzdělávání Julie Andrová TRANSGENNÍ PLODINY
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
GM kukuřice. 0,0004% DNA kukuřice
 Kolik je DNA v krmivech a potravinách? 0,005 až 0,02 % sušiny GM kukuřice cizí gen 4000 písmen 0,0004% DNA kukuřice Hypotetický příklad: brojler Krmná dávka - jen kukuřice Veškerá kukuřice jen GMO Brojler
Kolik je DNA v krmivech a potravinách? 0,005 až 0,02 % sušiny GM kukuřice cizí gen 4000 písmen 0,0004% DNA kukuřice Hypotetický příklad: brojler Krmná dávka - jen kukuřice Veškerá kukuřice jen GMO Brojler
Metody snižov. Prostřed emisních aspektů (smog, prach, CO 2. ším. často neustále nabývá na významu. znivě,, a to i lní.
 Metody snižov Prostřed edí ve kterém žijeme, je kromě běžně diskutovaných emisních aspektů (smog, prach, CO 2, )) zatíženo další ším, často opomíjeným, který však v neustále nabývá na významu. Je prokázáno
Metody snižov Prostřed edí ve kterém žijeme, je kromě běžně diskutovaných emisních aspektů (smog, prach, CO 2, )) zatíženo další ším, často opomíjeným, který však v neustále nabývá na významu. Je prokázáno
Příloha 2. Přehled řešených projektů v roce 2008
 Příloha 2. Přehled řešených projektů v roce 2008 Výzkumné záměry Kód Rok řešení MZE0002700601 MZE0002700602 MZE0002700603 Principy vytváření, kalibrace a validace trvale udržitelných a produktivních systémů
Příloha 2. Přehled řešených projektů v roce 2008 Výzkumné záměry Kód Rok řešení MZE0002700601 MZE0002700602 MZE0002700603 Principy vytváření, kalibrace a validace trvale udržitelných a produktivních systémů
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
R o z h o d n u t í. Univerzitě Palackého v Olomouci, Křížkovského 8, Olomouc. se p r o d l u ž u j e
 Rozhodnutí nabylo právní moci dne 3. ledna 2017. ODESÍLATEL: Ing. Karel Bláha, CSc. ředitel odboru environmentálních rizik a ekologických škod Ministerstvo životního prostředí Vršovická 65 100 10 Praha
Rozhodnutí nabylo právní moci dne 3. ledna 2017. ODESÍLATEL: Ing. Karel Bláha, CSc. ředitel odboru environmentálních rizik a ekologických škod Ministerstvo životního prostředí Vršovická 65 100 10 Praha
Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO)
") Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO) Stanovisko Věc: Stanovisko ČK GMO k žádosti o dovoz řezaných květů karafiátů linie FLO-40685-2 do EU Stav
Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO) Stanovisko Věc: Stanovisko ČK GMO k žádosti o dovoz řezaných květů karafiátů linie FLO-40685-2 do EU Stav
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE)
") MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
GENETICKÁ TRANSFORMACE OBILOVIN
 GENETICKÁ TRANSFORMACE OBILOVIN Pokročilé biochemické a biotechnologické metody 2.11.2015 K. Holubová ÚVOD Kukuřice pšenice rýže - ječmen Obiloviny: potrava, krmivo, průmyslní výroba Klimatické změny:
GENETICKÁ TRANSFORMACE OBILOVIN Pokročilé biochemické a biotechnologické metody 2.11.2015 K. Holubová ÚVOD Kukuřice pšenice rýže - ječmen Obiloviny: potrava, krmivo, průmyslní výroba Klimatické změny:
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Obsah přednášky. 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin
 Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin") Obsah přednášky 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin 2 Zákon č. 78/2004 Sb. Směrnice Evropského parlamentu a Rady 2001/18/ES ze dne 12.3.
Obsah přednášky 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin 2 Zákon č. 78/2004 Sb. Směrnice Evropského parlamentu a Rady 2001/18/ES ze dne 12.3.
Spermatogeneze saranče stěhovavé (Locusta migratoria)
") Spermatogeneze saranče stěhovavé (Locusta migratoria) Vývoj pohlavních buněk u živočichů zahrnuje několik dějů, které zajistí, že dojde k redukci a promíchání genetického materiálu a vzniklé buňky jsou
Spermatogeneze saranče stěhovavé (Locusta migratoria) Vývoj pohlavních buněk u živočichů zahrnuje několik dějů, které zajistí, že dojde k redukci a promíchání genetického materiálu a vzniklé buňky jsou
Molekulární biotechnologie. Nový obor, který vznikl koncem 70. let 20. století (č.1)
") Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Klonování gen a genové inženýrství
 Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Nové genové techniky, potraviny a monitoring
 21. Konference Monitoringu 2016, SZÚ, Milovy, 6.10. 2016 Nové genové techniky, potraviny a monitoring Veronika Kýrová Vladimír Ostrý Pavla Surmanová Ivana Procházková - Jiří Ruprich Podpořeno MZ ČR RVO
21. Konference Monitoringu 2016, SZÚ, Milovy, 6.10. 2016 Nové genové techniky, potraviny a monitoring Veronika Kýrová Vladimír Ostrý Pavla Surmanová Ivana Procházková - Jiří Ruprich Podpořeno MZ ČR RVO
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
WWW.HOLUB-CONSULTING.DE
 WWW.HOLUB-CONSULTING.DE Kukuřice jako monokultura způsobující ekologické problémy Jako například: půdní erozi díky velkým rozestupům mezi jednotlivými řadami a pozdnímu pokrytí půdy, boj proti plevelu
WWW.HOLUB-CONSULTING.DE Kukuřice jako monokultura způsobující ekologické problémy Jako například: půdní erozi díky velkým rozestupům mezi jednotlivými řadami a pozdnímu pokrytí půdy, boj proti plevelu
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Biosyntéza nukleových kyselin. VY_32_INOVACE_Ch0219.
 Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Klasifikace plazmid. Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: kb
 Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: 1-1000 kb Základní typyplazmid : kryptické - funkce neznámá epizomální - reverzibilní intergace do chromozomu hostitele konjugativní - schopné
Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: 1-1000 kb Základní typyplazmid : kryptické - funkce neznámá epizomální - reverzibilní intergace do chromozomu hostitele konjugativní - schopné
Více než 1,5 miliardy lidí na zemi trpí chronickými bolestmi. Existuje východisko z tohoto pekla?
 Více než 1,5 miliardy lidí na zemi trpí chronickými bolestmi. Existuje východisko z tohoto pekla? 100% Přírodní produkt pro podporu zdraví a úlevu od jakékoliv bolesti. Patentovaná technologie bylin na
Více než 1,5 miliardy lidí na zemi trpí chronickými bolestmi. Existuje východisko z tohoto pekla? 100% Přírodní produkt pro podporu zdraví a úlevu od jakékoliv bolesti. Patentovaná technologie bylin na
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Speciace neboli vznik druhů. KBI/GENE Mgr. Zbyněk Houdek
 Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
Kontrola genové exprese
 Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE
 MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE BRNO 2011 HELENA BŘUSKOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Tvorba vektorových konstruktů pro indukci
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE BRNO 2011 HELENA BŘUSKOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Tvorba vektorových konstruktů pro indukci
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/28.0032
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Seminář Rostlinné biotechnologie kolem nás Božena Navrátilová LS 2014/2015 Seminář
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Seminář Rostlinné biotechnologie kolem nás Božena Navrátilová LS 2014/2015 Seminář
Dekompozice, cykly látek, toky energií
 Dekompozice, cykly látek, toky energií Vše souvisí se vším Živou hmotu tvoří 3 hlavní organické složky: - Bílkoviny, cukry, tuky Syntézu zajišťuje cca 20 biogenních prvků - Nejdůležitější C, O, N, H, P
Dekompozice, cykly látek, toky energií Vše souvisí se vším Živou hmotu tvoří 3 hlavní organické složky: - Bílkoviny, cukry, tuky Syntézu zajišťuje cca 20 biogenních prvků - Nejdůležitější C, O, N, H, P
ANOTACE vytvořených/inovovaných materiálů
 ANOTACE vytvořených/inovovaných materiálů Číslo projektu Číslo a název šablony klíčové aktivity Tematická oblast Formát Druh učebního materiálu Druh interaktivity CZ.1.07/1.5.00/34.0722 III/2 Inovace a
ANOTACE vytvořených/inovovaných materiálů Číslo projektu Číslo a název šablony klíčové aktivity Tematická oblast Formát Druh učebního materiálu Druh interaktivity CZ.1.07/1.5.00/34.0722 III/2 Inovace a
OCHRANA MOSTNÍCH OBJEKTŮ PROTI JEJICH DESTRUKCI VLIVEM POVODŇOVÝCH OVÝCH UDÁLOST LOSTÍ
 projekt ISPROFONDU 5006210046 OCHRANA MOSTNÍCH OBJEKTŮ PROTI JEJICH DESTRUKCI VLIVEM POVODŇOVÝCH OVÝCH UDÁLOST LOSTÍ Ing. Petr Bouška, Ph.D. Ing.. Pavel Balvín Prof. Ing. Pavel Gabriel, DrSc. Doc.. Ing.
projekt ISPROFONDU 5006210046 OCHRANA MOSTNÍCH OBJEKTŮ PROTI JEJICH DESTRUKCI VLIVEM POVODŇOVÝCH OVÝCH UDÁLOST LOSTÍ Ing. Petr Bouška, Ph.D. Ing.. Pavel Balvín Prof. Ing. Pavel Gabriel, DrSc. Doc.. Ing.
NÁRODNÍ PROGRAM KONZERVACE A VYUŽÍVÁNÍ GENETICKÝCH ZDROJŮ ROSTLIN, ZVÍŘAT A MIKROORGANISMŮ VÝZNAMNÝCH PRO VÝŽIVU, ZEMĚDĚLSTVÍ A LESNÍ HOSPODÁŘSTVÍ
 Ministerstvo zemědělství, Praha 1, Těšnov 17 Č.j.: 33 083/03-3000 NÁRODNÍ PROGRAM KONZERVACE A VYUŽÍVÁNÍ GENETICKÝCH ZDROJŮ ROSTLIN, ZVÍŘAT A MIKROORGANISMŮ VÝZNAMNÝCH PRO VÝŽIVU, ZEMĚDĚLSTVÍ A LESNÍ HOSPODÁŘSTVÍ
Ministerstvo zemědělství, Praha 1, Těšnov 17 Č.j.: 33 083/03-3000 NÁRODNÍ PROGRAM KONZERVACE A VYUŽÍVÁNÍ GENETICKÝCH ZDROJŮ ROSTLIN, ZVÍŘAT A MIKROORGANISMŮ VÝZNAMNÝCH PRO VÝŽIVU, ZEMĚDĚLSTVÍ A LESNÍ HOSPODÁŘSTVÍ
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
ITÍ OBNOVITELNÝCH ZDROJŮ ENERGIE Z POHLEDU LEGISLATIVY. Pavel Noskievič
 VYUŽIT ITÍ OBNOVITELNÝCH ZDROJŮ ENERGIE Z POHLEDU LEGISLATIVY Pavel Noskievič Zelená kniha Evropská strategie pro udržitelnou, konkurenceschopnou a bezpečnou energii COM (2006) 105, 8.března 2006 Tři i
VYUŽIT ITÍ OBNOVITELNÝCH ZDROJŮ ENERGIE Z POHLEDU LEGISLATIVY Pavel Noskievič Zelená kniha Evropská strategie pro udržitelnou, konkurenceschopnou a bezpečnou energii COM (2006) 105, 8.března 2006 Tři i
Oligobiogenní prvky bývají běžnou součástí organismů, ale v těle jich již podstatně méně (do 1%) než prvků makrobiogenních.
 než prvků makrobiogenních.") 1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1. Chloroplasty jsou: a. v buňkách rostlin b. v buňkách živočichů c. v buňkách bakterií
 1. Chloroplasty jsou: a. v buňkách rostlin b. v buňkách živočichů c. v buňkách bakterií 2. Rostlinná buňka je: a. autotrofní b. heterotrofní c. schopna fagocytózy 3. Splynutím pohlavních buněk vzniká:
1. Chloroplasty jsou: a. v buňkách rostlin b. v buňkách živočichů c. v buňkách bakterií 2. Rostlinná buňka je: a. autotrofní b. heterotrofní c. schopna fagocytózy 3. Splynutím pohlavních buněk vzniká:
Amoxicillin trihydrate. Amoxicillin trihydrate. Trihydrát amoxicilinu. Amoxicillin trihydrate. Amoxicillin trihydrate. Amoxicillin trihydrate
 Příloha I Seznam názvů, lékových forem, sil veterinárních léčivých přípravků, živočišných druhů, způsobů podání, indikací/ochranných lhůt a žadatelů/držitelů rozhodnutí o registraci v členských státech
Příloha I Seznam názvů, lékových forem, sil veterinárních léčivých přípravků, živočišných druhů, způsobů podání, indikací/ochranných lhůt a žadatelů/držitelů rozhodnutí o registraci v členských státech