TRANSGENOZE ROSTLIN. vnášení cizorodých genů či jiné genetické informace do rostlin a manipulace s nimi (GMO)
|
|
|
- Jan Konečný
- před 6 lety
- Počet zobrazení:
Transkript

1 TRANSGENOZE ROSTLIN vnášení cizorodých genů či jiné genetické informace do rostlin a manipulace s nimi (GMO)
2 transformace rostlinné buňky TRANSGENOZE trvalá - změna genetické informace je přenášená na potomstvo (infekce Agrobacteriem tumefaciens) TRANSGENOZE dočasná - genetická informace je změněna pouze u vegetativní části rostliny a nepřenáší se na potomstvo (biolistické metody, virální infiltrace)



3 Agrobacterium tumefaciens patogenní bakterie přirozeně se vyskytující v kořenovém systému bobovitých rostlin přítomnost baktérie se projevuje tumory tzv. crown gall tissue Agrobacterium je schopné napadat jakékoliv pletivo většiny dvouděložných rostlin a za určitých podmínek i rostlin jednoděložných
4 Ti plazmid (tumor inducing) Agrobacteria tumefaciens (200 kb) ipt,aux,nop syntéza cytokininů auxinů opinů nopalin další geny pro biosyntézu cytokininů, permeasy, přenašeče přes membránu, enzymy metabolizující opiny a ostatní enzymy umožňující infekci

5 rostlina při zranění produkuje fenolické látky (acetosyringon), ty se váží na specifické receptory na membráně Agrobacteria VirA a způsobují transkripci genu s vir regionu Ti plazmidu Agrobacteria

6 aktivace vir regulonu VirD2 protein se váže na 5 T-DNA (transferred) denaturace ss T-DNA z Ti plazmidu VirE2 proteiny stabilizují ss T-DNA VirD2 a VirE2 proteiny obsahují sekvenci potřebnou pro cílení do jádra rostlinné buňky VirE2 se váže na proteiny VIP1 a VIP2 které zacílí T-DNA řetězec do chromatinu rostlinného chromozomu

7 začlenění T-DNA do chromozomu začlenění T-DNA do hostitelského chromozomu je víceméně náhodné nejčastěji se začlení jedna T-DNA do jedné buňky, muže však nastat i několikanásobné začlenění kolem centromer je začlenění méně pravděpodobné čím blíže telomerám tím větší pravděpodobnost Arabidopsis thaliana
: nese větší část Ti plazmidu, všechny geny potřebné pro symbiózu s")

8 binární vektory odvozené od Ti plazmidu pro genetickou modifikaci rostlin Ti plazmid je rozdělen na dva: pomocný plazmid (velký): nese větší část Ti plazmidu, všechny geny potřebné pro symbiózu s hostitelem a přenos T-DNA klonovací plazmid (malý): nese pouze T-DNA, do jejíž části se naklonuje požadovaná gen.informace, replikační počátky pro E.coli a Agrobacterium a potřebné selekční markery
9 klonovací plazmid pbin-hyg-tx polylinker resistence k hygromycinu B (selekce transgenních rostlin) 35S promotor tetracyklinový supresor T-DNA replikační počátky ori požadovaný gen se překlonuje do polylinkeru a konstrukt se namnoží v E.coli. Pro přenos je nutná jen 25bp repetice z konce původní T-DNA. (kanamycinová selekce) transformace do Agrobacteria nesoucí pomocný vektor (selekce Kan, Rimf, Gent) transformace rostliny a selekce transgenního potomstva (selekce hygromycinová) resistence ke kanamycinu pomocný plazmid ma resistenci k rimfapicinu a gentamycinu

10 transgenoze Agrobacteriem standardní postupy pro řadu dvouděložných užitkových rostlin (rajče, hořčice, fazole). nevýhoda: zdlouhavá regenerace přes tkáňové kultury ne u všech se dá použít germ-line transformace jako u Arabidopsis transformace T-DNA se dá využít i jako inzertní mutageneze. T-DNA přeruší důležitý gen (mutace) a tuto mutaci lze pak pomocí markerů na T-DNA snadno lokalizovat. Existují databanky mutantních (transgenních) rostlin Arabidopsis, kdy máte mutaci (inzerci) přímo v genu který studujete.
.")
11 transgenoze namáčením květů (floral dipping, germ-line) A) rostliny Arabidopsis se vypěstují do fertilního stádia (3-5 v květináči) B) lodyhy s květy se vymáchají v kultuře Agrobacteria nesoucí binární systém vektorů s naklonovaným transgenem (5-10 min., přidává se smáčedlo). C) rostliny se nechají pořádně okapat a vrátí do skleníku, kde dozrají (vytvoří semena) D) transgenní semena se selektují vyklíčením na živném médiu obsahující selekční marker (hygromycin) např. Arabidopsis thaliana, řepka, vojtěška
.")
12 transgenoze namáčením květů (floral dipping) Agrobacterium se dostává do vyvíjejících se květů a T-DNA je včleněna do samičího gametofytu a přenesená přes vajíčko do další generace (nikdy ne do samčích pohl. buněk, transgenní semena jsou proto vždy heterozygotní na vložený transgen). zlepšení : vakuová infiltrace přidávání smáčedla silikon Silwet L-77 účinnost: až 20% transgenního potomstva GUS (uida gen) X-Gluc 5-bromo-4-chloro-3-inolyl- -D-glucuronic acid -glucuronidasa modrý precipitát

13 transgenoze pomocí listových disků např. tabák metoda používaná u většiny dvouděložných rostlin



14 regenerace transgenních tabákových rostlin z kalusu vzniklých na listových discích



15 tabák Arabidopsis

16 TRANSFORMACE TOPOLU






17 TOP 10 ZEMĚDĚLSKÝCH PLODIN které se nejvíce transformují a budou transformovat 2006 budoucnost 1. Kukuřice setá 2. Lilek brambor 3. Řepka olejka 4. Sója luštinatá 5. Bavlník 6. Lilek rajče 7. Tabák viržinský 8. Rýže setá 9. Pšenice setá 10. Topol osika 1. Rýže setá 2. Sója luštinatá 3. Pšenice setá 4. Lilek brambor 5. Kukuřice setá 6. Tabák viržinský 7. Banánovník 8. Réva vinná 9. Čajovník a Kávovník 10. Mák polní
 izolace nevyvinutých embryí z obilniny mechanické narušení odstranění embryonální osy")

")
18 transgenoze jednoděložných např. ječmen (Tingay et al.,1997) izolace nevyvinutých embryí z obilniny mechanické narušení odstranění embryonální osy kokultivace s Agrobacteriem kultivace na kalusotvorném médiu detekce reportérového genu v T0 i T1 generaci efektivita do 4% (např. z 1282 embryí vyselektováno 54 transformovaných lini poprvé chřest (1988) problém: jednoděložné obsahují daleko méně fenolických látek zlepšení: ošetření acetosyringonem (aktivace vir regionu) supervirulentní kmeny Agrobacteria: mají zmnožený gen pro virg, který je navíc pod konstitutivním promotorem

19 Metoda transformace ječmene


20 Selekce homozygotní linie T0 generace všichni transformanti jsou heterozygotní selekce semen na nutričním mediu obsahující selekční antibiotikum T1 generace T-DNA inzerce na jednom lokusu ¼ 00 homozygotů ½ M0+0M heterozygotů ¼ MM homozygotů 3:1 75% T-DNA inzerce na dvou lokusech 1/14 oo 00 homozygotů 3/14 oo Mx homo-heterozygotů 3/14 mx 00 homo-heterozygotů 9/14 mx MX hetero-homozygotů 9:3:3:1 93% T-DNA inzerce na třech lokusech 1/64 oo homozygotů 27/64 homo-heterozygotů 36/64 MM hetero-homozygotů 27:9:9:3:9:3:3:1 98% T2 generace selekce semen na nutričním mediu obsahující selekční antibiotikum 100% 75% HOMOZYGOTNÍ LINE
 smíchají se s roztokem")
21 přímá transgenoze TRANSGENOZE PROTOPLASTŮ odstranění buněčné stěny a inkubace protoplastů s roztokem DNA, DNA vniká do protoplastu endocytózou polyethylenglykol elektroporace makroinjekce protoplasty se dají udělat ze všech rostlin lze z nich však jen velmi obtížně regenerovat celé rostliny BIOLISTICKÉ METODY TRANSGENOZE používají se zlaté nebo wolframové kuličky (2 m) smíchají se s roztokem plazmidové DNA s transgeny, odpaří se vstřelují se do pletiva pod vakuem transgen společně s reportérovým genem (GUS) snadná detekce transformace
 5%")
22 10% pronikne do pletiva 50% zůstane v epidermis (netvoří kalus) 1% přežívá a dochází k integraci do genomu 95% cytoplazma nedochází k transkripci) 5% jádro


23 genetická puška pokud se nepodaří z buněk zregenerovat kalus, transgenoze je pouze dočasná
24 SELEKČNÍ MARKERY PRO ROSTLINNOU TRANSGENOZI 1.ANTIBIOTIKA KANAMYCIN gen nptii pro neomycinfosfotransferasu. Rostliny se buď stříkají, nebo se dává přímo do média. HYGROMYCIN B gen aphiv pro hygromycinfosfotransferasu silně toxický i pro člověka inhibitor proteosyntesy 2. HERBICIDY nejpoužívanější GLYFOZÁT inhibitor syntézy aromatických AK (je esenciální u autotrofních organismů) sp. 5-enolpyruvylšikimát-3-fosfátsyntasa (EPSP) tři strategie: 1) overexprese EPSP 2) exprese mutantní ne tak citlivé formy EPSP 3) exprese glyfozátoxidoreduktasy (degradace glyfozátu) transgenní potomstvo se sprejuje glyfozátem, přežívají pouze rostliny nesoucí současně s transgenem jeden ze tří způsobů rezistence
25 SELEKČNÍ MARKERY PRO ROSTLINNOU TRANSGENOZI 3. ZDROJ SACHARIDŮ v raných stádiích vývoje rostlin v in vitro podmínkách je závislost na zdroji uhlíku v médiu (sacharóza). Selekční marker: v médiu je místo sacharózy manóza. Hexokinasou vzniká manóza-6-fosfát, který není schopen podléhat následné glykolýze a je pro rostlinu toxický, kumuluje fosfor. Selekční transgen: fosfomanozaizomerasa přeměňuje manóza-6-fosfát na fruktóza-6-fosfát (použito u tabáku, cukrovky a kukuřice) 4. SELEKCE ROSTLINNÝMI HORMONY transgen isopentenyltransferasa produkuje nadbytek cytokininů (nemusí se přidávat do selekčního média) které podporují tvorbu prýtů z transformovaného pletiva, u tabáku a salátu bylo zjištěno že tento nadbytek nemá vliv na celkovou morfologii transgenní rostliny.
26 ODSTRANĚNÍ SELEKČNÍCH TRANSGENů Z ROSTLIN někdy je třeba tento transgen odstranit aby nenarušoval vývoj rostliny (ipt,nptii) využití bakteriofágového systému Crelox, přirozený exscisní systém, gen cre kóduje rekombinasu (transpozasu) a lox symbolizují krátké cílové sekvence excise.
27 SLOŽKY TRANSGENŮ PROMOTOR konstitutivní 35 S - nejpoužívanější, virového původu, nebezpečí KOSUPRESE ubikvitin - rostlinného původu, exprese kolísá během ontogeneze actin - rostlinného původu, vysoká ale kolísavá exprese pletivově specifické 2A11 - z rajčete, plodově specifický Lhcb3 - listově specifický, světlem regulovaný z Arabidopsis RAmy3D - amylasa z rýže, aktivovaný sacharosou B-HORD a Zea - zásobní proteiny z obilek, hordein z ječmene a zein z kukuřice inducibilní In2-2 - je indukovaný benzensulfonamidovým herbicidem Safener tet - indukce tetracyklinem, nejpoužívanější pop - indukovaný glukokortikoidem DEXAMETHASONE, živočišný systém přenesený do rostliny ER - promotor lidského estradiolového receptoru
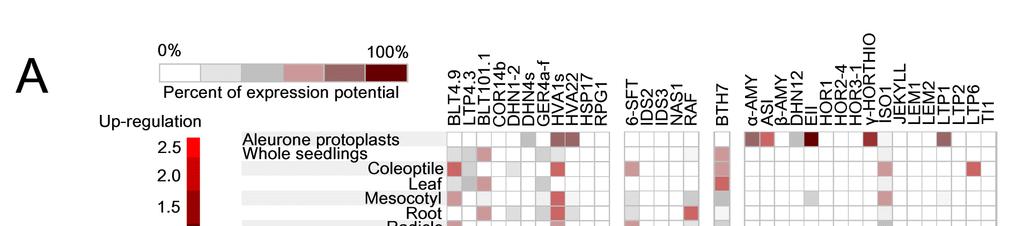



28 Pletivově specifické promotory u jednoděložných
29 SLOŽKY TRANSGENŮ POLYADENYLAČNÍ SIGNÁL a TERMINÁTOR v klonovací kazetě je kromě promotorové sekvence i terminátor transkripce s polyadenylačním signálem nejčastěji používaný terminátor z agrobakteriálního genu pro nopalin syntasu používá se jak u jedno tak dvouděložných rostlin další používané terminátory: 3 nekódující sekvence rostlinného genu pro pšeničný histon H3 může se použít i vlastní 3 nekódující sekvence transgenu, ale ta nemusí vždy fungovat, je jednodušší mít univerzální již v plasmidu
30 SLOŽKY TRANSGENŮ INTRONY inkorporace intronu do transgenu může zvyšovat sílu exprese transgenu v rostlině např. intron I z genu pro rýžový aktin nebo intron I z genu pro kukuřičný ubikvitin 5 NEKODUJÍCÍ REGION Kozákové sekvence a podobné sekvence zvyšují postranskripční stabilitu mrna a tím zvyšují míru exprese NUKLEOTIDOVÉ SLOŽENÍ pokud vkládáme do rostliny gen z baktérie, hrozí nebezpečí použití odlišného genetického kódu, vyštěpení neexistujícího intronu atd. geny ze vzdálených taxonů se proto musí často syntetický upravovat (mutacemi) aby bylo dosaženo dostatečné exprese v rostlinném hostiteli (např. u Bt toxinu z baktérie), nebo se současně řeší umělou syntézou genů
:")

31 odchylky v genetickém kódu snižují výtěžek heterologní exprese jiná preference: kodón pro arginin (6 různých): CGU CGA CGG CGC AGA AGG E. coli Arabidopsis th. H. sapiens AGA 2.2% AGA 18.9% AGA 11.9% AGG 1.6% AGG 11.0% AGG 12.1%

32 PŘÍMÁ TRANSFORMACE CHLOROPLASTŮ začlenění transgenu do chloroplastové DNA VÝHODY: dědí se pouze matrilineárně, tzn. transgen se nepřenáší pylem. k začleňování transgenu dochází pouze na specifických místech DNA homologní rekombinací vysoká exprese, až několik procent celkových proteinů lepší pro expresi bakteriálních genů v rostlinách (stejný genetický kód) NEVÝHODY: provedení: používá se biolistické metody velice silné genetické dělo hnané héliem, musí prorazit dvojitou chloroplastovou membránu, velice malá účinnost specifické vektory: obsahují část genetické informace chloroplastu, aby mohla proběhnout homologní rekombinace selekční marker antibiotikum spektinomycin Limitace: lze provést pouze u několika rostlinných druhů
 B) TOLERANCE VŮČI STRESU C) MOLEKULÁRNÍ FARMÁŘENÍ (rostlinná buňka jako bioreaktor) 1. plantibodies a jedlé vakcíny 2.")
33 VYUŽITÍ TRANSGENOZE ROSTLIN A) ZMĚNA VLASTNOSTÍ ZEMĚDĚLSKÝCH PLODIN 1. rezistence k herbicidům 2. rezistence k hmyzím škůdcům 3. rezistence k virům 4. změna obsahu olejů 5. oddálení dozrávaní plodů 6. kontrola opylování 7. zvyšování nutriční hodnoty (zlatá rýže) B) TOLERANCE VŮČI STRESU C) MOLEKULÁRNÍ FARMÁŘENÍ (rostlinná buňka jako bioreaktor) 1. plantibodies a jedlé vakcíny 2. produkce terapeutik rostlinami 3. produkce biopolymerů rostlinami
34 rezistence ke hmyzím škůdcům Bt - rostliny 1902 byla v Japonsku objevena baktérie Bacillus thuringiensis, která způsobovala úhyn bource morušového 1987 byl z této baktérie izolován krystalický protein -endotoxin, který po projití trávicím traktem hmyzu, způsobuje jeho rozklad se začala tato baktérie využívat jako insekticid, postřiky byly drahé a dlouho neúčinkovaly 1985 připraven transgenní tabák overexprimující Bt-toxin (nízká exprese % všech listovým proteinů), důvod: nestabilita bakteriální mrna v rostlině synteticky upravené geny pro -endotoxin - byl snížen obsah AT bází - pozměněn genetický kód tak aby vyhovoval rostlinám - vloženy introny exprese se zvýšila až 1000x, většinou se využívá konstitutivní 35S promotor existuje řada genů pro Bt-toxin (CryIA- CryIIIB) s různou specifitou vůči různým hmyzím druhům


35 Bt-rostliny a herbicid-rezistentní rostliny 40% pěstované kukuřice v USA jsou Bt-transgenní odrůdy 80% světové produkce sóji je ROUND-UP odrůda běžná rostlina bramboru napadená mandelinkou Bt-brambora
 1999 NEJVĚTŠÍ SKANDÁL GMO případ motýlka Danaus plexipus tento motýl patří v USA mezi ohrožené druhy publikovaná")

36 rizika Bt-rostliny i samotný endotoxin byly testovány na lidech, psech, morčatech, čolcích, krysách, rybách a ptácích bez žádných příznaků byly prováděny alergologické testy nebyl pozorován ani žádny vliv na hmyzích predátorech (1996 včela, slunéčko) 1999 NEJVĚTŠÍ SKANDÁL GMO případ motýlka Danaus plexipus tento motýl patří v USA mezi ohrožené druhy publikovaná práce kdy byla prokázána zvýšená mortalita larev po poprášení rostlin klejichy (potrava) pylem z Bt kukuřice v roce 1999 byl však zaznamenán nárůst populace tohoto motýlka nedochází k časové koincidenci kvetení kukuřice a vývoje larev GREENPEACE vydává seznam evropských motýlů ohrožených endotoxinem v Bt-rostlinách snaha o zavádění Bt-rostlin s matrilineární dědičnosti
... běžná kanola Roundup Ready kanola")
37 GMO rezistentní k herbicidům Roundup Ready bavlna, kukuřice, sója a jarní řepka (kanola)... běžná kanola Roundup Ready kanola


38 zlatá rýže další využívané transgeny největší evropský projekt GMO ovlivněná celá metabolická dráha geranylgeranyl pyrofosfát fytoensyntasa fytoen fytoendesaturasa fytoendesaturasa normální zrna rýže -karoten -karoten desaturasa lykopen lykopen- β-cyklasa b-karoten endogenní lidské geny zrná rýže s β-karotenem Vitamin A
 nejsou obratlovci schopni syntetizovat, ale")
39 projekt - zlatá rýže rýže je schopná syntetizovat β-karoten jen v zelených částech v obilkách je posledním meziproduktem geranylgeranylpyrofosfát do genomu rýže vneseny pod specifickým glutinovým promotorem s cílením do endospermu tři geny: fytoendesaturasa z baktérie Erwinia fytoensyntasa z narcisu lykopen- β-cyklasa β-karoten (provitamín A) nejsou obratlovci schopni syntetizovat, ale potřebují ho pro syntézu retinalu (součást pigmentu rhodopsinu) a k.retinové (reguluje morfogenezi) avitaminóza způsobuje dětskou slepotu, snižování imunity a je příčinou asi půl milionové dětské úmrtnosti především v rozvojových zemích jihovýchodní Asie silné námitky antibiotechnologických aktivistů není třeba, jsou tabletky - může dojít k předávkování atd.

40 USA 2. Argentina 3. Brazílie 4. Kanada 5. Indie 6. Čína 7. Paraguay 8. JAR 9. Uruguay 10. Filipíny 11. Austrálie 12. Španělsko 13. Mexiko 14. Kolumbie 15. Chile 16. Francie 17. Honduras 18. Česko 19. Portugalsko 20. Německo 21. Slovensko 22. Rumunsko USA 2. Brazílie 3. Argentina 4. Kanada 5. Indie 6. Paraguay 7. Pákistán 8. Čína 9. JAR 10. Uruguay 11. Bolivie 12. Austrálie 13. Filipíny 14. Barma 15. Španělsko 16. Súdán 17. Mexiko 18. Kolumbie 19. Vietnam 20. Honduras 21. Chile 22. Portugalsko 23. Bangladéš 24. Kostarika 25. Slovensko 26. Česko 27. Rumunsko 28. Burkina Faso

41
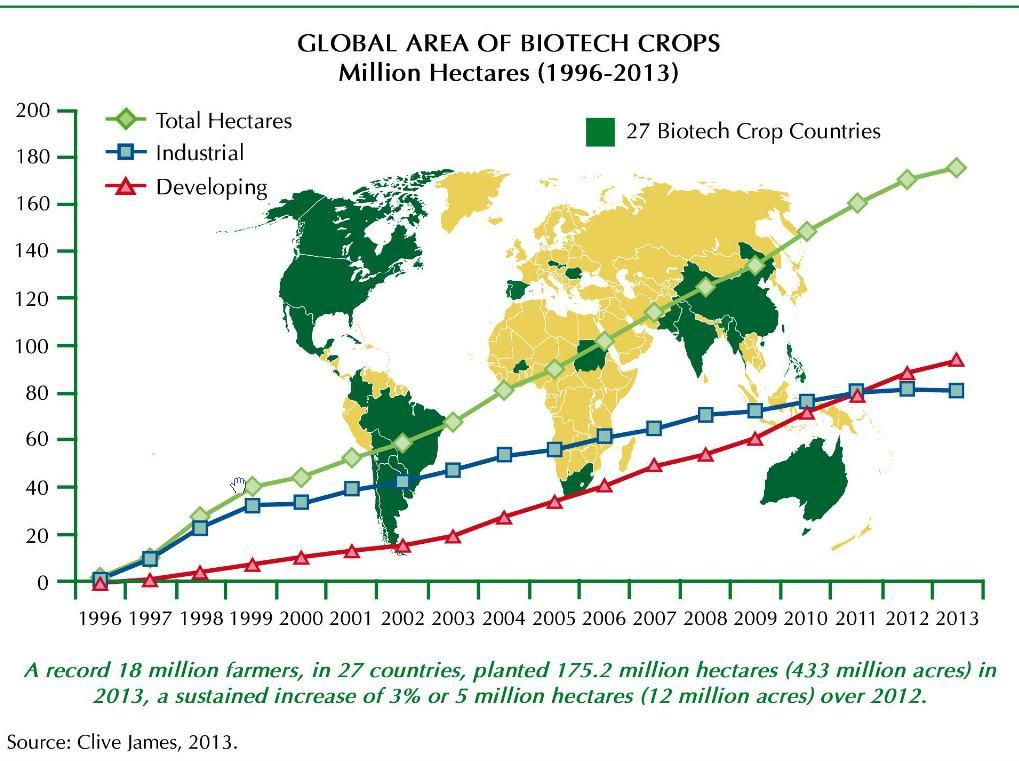
42
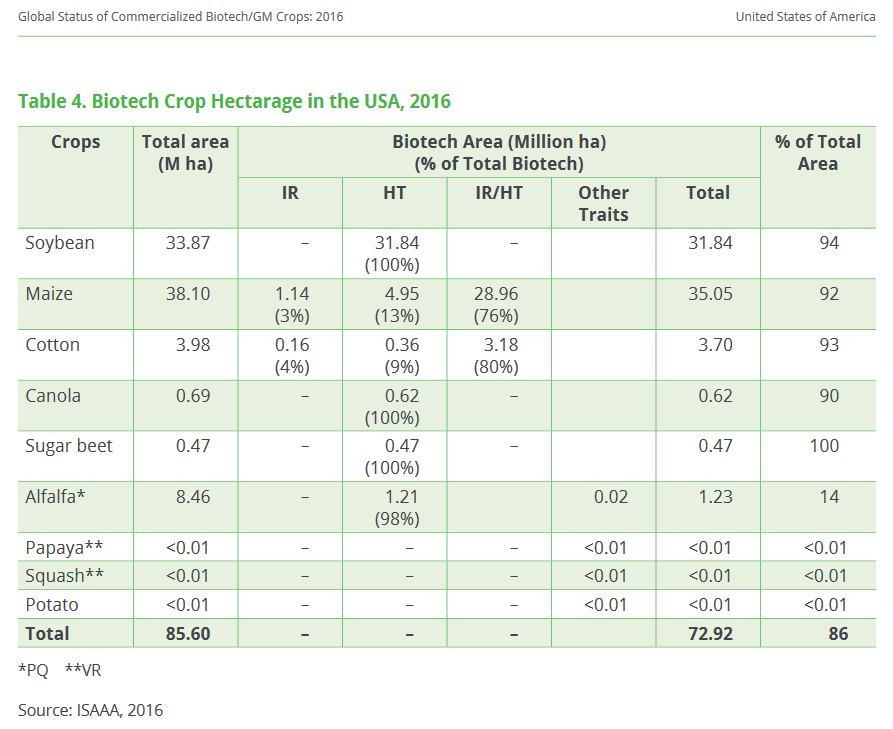
43










44 Polní experiment s GM ječmenem v Olomouci
45 situace v roce 2013 (GM plodiny komerčně pěstovány od ) GM plodiny na více než hektarech (cca rozloha Libye) 27 zemí (v roce přibyla Kuba, Barma, Burkina Faso, Súdán) z toho 19 rozvojových a 8 vyspělých odrůda brambor Amflora po 13 letech nová schválená odrůda v EU 2012 kukuřice odolná vůči suchu 2019 zlatá rýže Round up a další herbicid tolerantní Sója Kukuřice Bavlna Vojtěška Řepa Psineček (bentgrass) Len Rýže Pšenice Bt a tolerantní vůči hmyzu Kukuřice bavlna Rajče Dýně Topol Lilek Rýže Řepka (high-laurate, male-sterile) Rajče (FlavrSavr, ethylen) Karafiáty (umělá barva) Brambor (high amylopektin) Čekanka (male-sterile) Meloun (oddálení senescence) Růže (změna barvy květů) Cukrová třtina (tolerantní vůči suchu) Tabák (nicotin free) Tolerantní vůči virům Papaja Dýně Sweet Pepper Fazol Slivoň
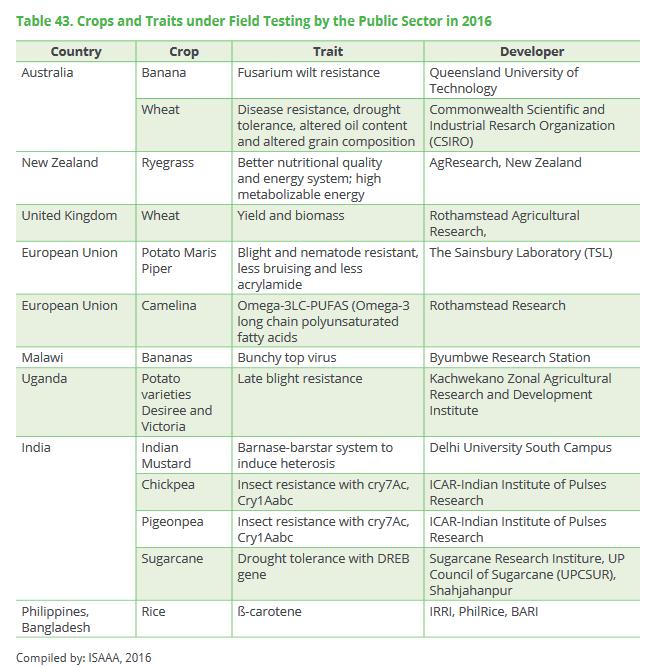
46
 nebo mohou být extrahovány a použity v diagnostice či medicíně výrazně se sníží náklady a čas na tvorbu protilátek (hybridomové buňky)")
47 molekulární farmáření v rostlinách připravené živočišné protilátky nebo části protilátek v rostlinách lze produkovat i monoklonální protilátky protilátky jsou vylučovány do mezibuněčného prostoru mohou sloužit přímo v rostlině (jako ochrana proti patogenům) nebo mohou být extrahovány a použity v diagnostice či medicíně výrazně se sníží náklady a čas na tvorbu protilátek (hybridomové buňky) plantibodies

48 plantibodies hlavní problém rozdíly v N-glykosylaci proteinů (protilátek) u živočichů a rostlin plantibodies mají jinak glykosylovanou strukturu a po aplikaci do zvířete či člověka vyvolávají nechtěnou imunitní odpověď řešení: společně s geny pro Ab je do rostliny vnesen gen pro lidskou β-1,4- galaktosyltransferasu rostlina pak produkovala polidštěné protilátky jako transgen stačí vložit pouze malou variabilní oblast tzv. Fv domény z lehkého a těžkého řetězce spojenou krátkým peptidem scfv single chain variable fragment
49 GLYKOSYLACE Rostliny: β(1,2)xylosa (1,3)fukosa Člověk: (1,6)fukosa β(1,2)galaktosa k.sialová řešení: stabilní genetická úprava glykosylačních enzymů v rostlinné buňce Ko et al. Plant-produced microbial vaccines. Springer, 2009.

50 Některé lidské proteiny produkované suspenzními rostlinnými kulturami terapeutický rostlina výtěžek Typ produkce promoter rok protein mg/l alfa-antitrypsin rýže 247 sekrece RAmy3D 2005 lidský růstový hormon hgh tabák 35 sekrece CaMV35S 2010 interferon α2b tabák 28 sekrece CaMV35S 2007 interleukin-12 rýže 31 sekrece RAmy3D 2010 glukocerebrosidasa mrkev 30 vakuolarní CaMV35S 2007 lysozym rýže 3% intra-buněčně RAmy3D 2002 lactoferrin ženšen tabák 4.3% intra-buněčně SWPA2 2006
 lektin sněženky GNA")
51 rezistence ke hmyzím škůdcům transgeny rostlinného původu inhibitory trávicích enzymů (amylas a proteas) hmyzu, lektiny (proteiny vážící sacharidy), chitinasy, syntéza alkaloidů INHIBITORY PROTEAS sójový inhibitor serinových proteas inhibitor trypsin proteas z vigny inhibitor proteas z rajčete INHIBITORY AMYLAS amylasový inhibitor z fazole LEKTINY (účinné proti mšicím) lektin sněženky GNA (Galantus nivalis aglutinin) lektin hrachu chitinasa z fazole

52 rezistence k virům hlavním způsobem rezistence je exprese genu pro plášťový protein daného viru 1986 první transgenní rostliny tabáku rezistentní vůči TMV (existuje faktor, který rozbalí virus a aktivuje ho, tento faktor se při infekci vychytá plášťovým proteinem produkovaným rostlinou. Korelace: čím větší exprese tím vyšší rezistence byly nalezeny i jiné původní geny způsobující rezistenci, ty byly mapovány a použity jako transgeny proti RNA virům se využívá protismyslná RNA strategie



53 papája na Havaji papaya ringspot virus (PRV) ovlivňoval úrodu na Havaji od roku 92 do roku 96 klesla produkce díky viru o 40% 1996 první transgenní papája overexprimující plášťový protein pod 35S promotorem připravená biolistickou metodou od roku 1998 nárůst produkce
54 transgenní rostliny tolerantní vůči stresu VODNÍ STRES při nedostatku vody (i zasolení) se zvyšuje osmotický tlak buněk, dochází k poklesu fotosyntézy a fotoredukci kyslíku za vzniku aktivních kyslíkových radikálů poškození a) nadprodukce osmoprotektivních látek glycinbetain transgen cholindehydrogenasa z E.coli (tabák, brambor) prolin transgen semialdehyddehydrogenasa k. glutamové z vigny manitol transgen manitol-1-fosfátdehydrogenasa fruktany chimerický transgen fruktozyltransferasa z Bacillus subtilis a vakuolární signální sekvence z karboxypeptidasy ze Saccharomyces b) ovlivnění signální dráhy pro dehydratační stres u Arabidopsis thaliana byla objevena signální dráha společná pro stres dehydratací a chladem současně bylo zjištěno, že geny reagující na tyto stresy mají ve svých promoterech konzervovanou sekvenci TACCGACAT tzv. DRE (dehydratation responsive element). Byl nalezen transkripční faktor aktivující tuto sekvenci DREB1A, a jeho gen použit jako transgen. + transgenní rostliny daleko odolnější k suchu, mrazu zasolení - zpomalení růstu (může se použít specifický promotor indukovatelný stresem)
55 GENOVÉ UMLČOVÁNÍ (gene silencing; knock-down)
 založena na specifické degradaci mrna Science, prosinec")
56 genové umlčování TGS (transcriptional gene silencing) nedochází k transkripci transgenu může jí předcházet transkripce promotorové sekvence a následná inhibice promoteru komplementárním duplexem RNA-DNA nebo častěji metylace promotorové sekvence PTGS (post-transcriptional gene silencing) založena na specifické degradaci mrna Science, prosinec 2002

57 TGS a PTGS read-through transcription DNA-DNA pairing methylation chromatin remodelling methyltransferases chromatin remodelling factors RNA-DNA pairing RNA degradation RNA-RNA pairing dsrna
 pod silným virálním promotorem. Transgen interferoval s vlastním genem pro tento enzym a docházelo k jeho vzájemnému umlčování.")
58 poprvé pozorováno 1990 u transgenní petúnie vědci se pokoušeli ztmavit květy petúnie overexpresí genu pro chalkonát syntasu (enzym produkující černý antokyanin) pod silným virálním promotorem. Transgen interferoval s vlastním genem pro tento enzym a docházelo k jeho vzájemnému umlčování. prpl prpl
59 PRINCIP RNAi hairpin RNA Virus dsrna Transgene RNA dsrna sirna (short interfering RNAs, nt long) HO p HO p p OH p HO OH p Cleavage of dsrna by dicer-like enzyme p p HO OH p OH (DICER contains dsrna cleavage activity, helicase and dsrna binding activity) RNase III like protein systém funguje v cytoplasmě a zřejmě existuje jako rezistence vyvinutá proti RNA virům sirna/protein complex HO p p OH sirna is unwound, single-stranded RNA formed mrna target RISC complex (RNA-induced silencing complex) sequence-specific target recognition p p OH OH RISC se automaticky generuje v rostlině v přítomnosti sirna imunitní odpověď na úrovni NK mrna degradation continued on next slide
60 RNA dependentní RNA polymerasa Single-stranded sirnas can also be used as primers to make more dsrna by host RNA polymerase New dsrna formed from target mrna p OH Dicer (as in previous slide) HO p p p OH p HO OH OH p p OH HO p HO p význam RNAi STUDIUM FUNKCE GENů Etc. Model was modified from Roth et al., (2004) Virus Research 102: TERAPEUTICKÉ VYUŽITÍ knock-out nebo inhibice funkce genu přežije organismus? jaké pozorujeme fenotypové změny? léčba rakoviny (viz. genová terapie)
61 dvě strategie: a) KLASICKÁ PCR s dvěma typy primerů a postupné klonování (vektor phannibal) b) REKOMBINANTNÍ jedno PCR a klonování s následnou rekombinací (BP klonasa, vektor
 je způsobováno hlavně degradaci pektinu (polysacharid, polymer kys.")


62 ODDÁLENÍ DOZRÁVÁNÍ PLODů rajčata první transgenní komerční odrůda FLAVR SAVR zrání plodů rajčat (i ostatních) je způsobováno hlavně degradaci pektinu (polysacharid, polymer kys. D-galakturonové) v buněčné stěně plodů při zrání se aktivuje polygalakturonasa která rozkládá pektin 1994 transgenní odrůda lilku rajčete FlavrSavr s knock-outovanou polygalakturonasou (na 10%) transgen produkuje protismyslnou RNA zvýšena trvanlivost rajčat po sklizni o 2 týdny větši rezistence vůči houbovým a bakteriálním chorobám transgenic control + ethylene transgenic

63 další využívané transgeny bezkofeinová káva 7-metylxantin N-metyltransferasa a theobromin N-metyltransferasa JAPONSKO 2003 exprese genů inhibována metodou RNAi druh Cofea canephora transformován Agrobaktériem transformované rostliny stejné jako wild type Cofea canephora se pěstuje na Madagaskaru a má 2x větší obsah kofeinu než nejrozšířenější Cofea arabica v mladých listech: theobromin 30-80% kofein 50-70%

.")
64 další využívané transgeny MODRÁ RůŽE 2004 vědci z firmy FLORIGENE (Calgene Pacific) pracovali na vývoji transgenní modré růže po několik let. Nakonec použili jako transgen gen pro flavonoid 3', 5'- hydroxylasu z petúnie pod specifickým promotorem s cílením do květů a transformovali čínskou čajovou růži (Rosa x hybrida). Transgen způsobil syntézu modrého flavonoidu delfinidinu.
65 transgen
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Biotechnologie rostlin, kultivace rostlinných buněk
 Katedra biochemie, Přírodovědecká fakulta Univerzita Palackého v Olomouci Ivo FRÉBORT Biotechnologie rostlin, kultivace rostlinných buněk I. část Šlechtění kulturních plodin Zemědělské plodiny jsou výsledkem
Katedra biochemie, Přírodovědecká fakulta Univerzita Palackého v Olomouci Ivo FRÉBORT Biotechnologie rostlin, kultivace rostlinných buněk I. část Šlechtění kulturních plodin Zemědělské plodiny jsou výsledkem
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
GENETICKY MODIFIKOVANÉ
 GENETICKY MODIFIKOVANÉ ROSTLINY (GMR) Lukáš Fischer Katedra experimentální biologie rostlin PřF UK Geny základ vlastností organismů Změny genetické informace rostlin a definice genetické modifikace dle
GENETICKY MODIFIKOVANÉ ROSTLINY (GMR) Lukáš Fischer Katedra experimentální biologie rostlin PřF UK Geny základ vlastností organismů Změny genetické informace rostlin a definice genetické modifikace dle
Modifikace dědičné informace rostlin I. modifikace
 Modifikace dědičné informace rostlin I Klasická genetická modifikace Lukáš Fischer, KEBR Legislativa: Genetická modifikace (GM) = vnesení genetické informace (úseku DNA) či změna > 20 nt způsobem, který
Modifikace dědičné informace rostlin I Klasická genetická modifikace Lukáš Fischer, KEBR Legislativa: Genetická modifikace (GM) = vnesení genetické informace (úseku DNA) či změna > 20 nt způsobem, který
Organizace a kontrola pěstování GM plodin v ČR. Ing. Jana Trnková MZe, odbor rostlinných komodit
 Organizace a kontrola pěstování GM plodin v ČR Ing. Jana Trnková MZe, odbor rostlinných komodit Geneticky modifikované plodiny GM plodiny, transgenní rostliny změněn dědičný materiál (DNA) pomocí genových
Organizace a kontrola pěstování GM plodin v ČR Ing. Jana Trnková MZe, odbor rostlinných komodit Geneticky modifikované plodiny GM plodiny, transgenní rostliny změněn dědičný materiál (DNA) pomocí genových
Geneticky modifikované organismy
 Geneticky modifikované organismy Ivo Frébort KBC/BAM Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
Geneticky modifikované organismy Ivo Frébort KBC/BAM Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
GENETICKY MODIFIKOVANÉ ORGANISMY. Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN
 GENETICKY MODIFIKOVANÉ ORGANISMY Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN VERTIKÁLNÍ PŘENOS VLASTNOSTÍ DĚDIČNOST považoval člověk za samozřejmou zákonitost Evoluce
GENETICKY MODIFIKOVANÉ ORGANISMY Prof. Jaroslav DROBNÍK Přírodovědecká fakulta Karlovy Univerzity Sdružení BIOTRIN VERTIKÁLNÍ PŘENOS VLASTNOSTÍ DĚDIČNOST považoval člověk za samozřejmou zákonitost Evoluce
Obsah přednášky. 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin
 Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin") Obsah přednášky 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin 2 Zákon č. 78/2004 Sb. Směrnice Evropského parlamentu a Rady 2001/18/ES ze dne 12.3.
Obsah přednášky 1) Zákon č. 78/2004 2) GMO ve světě 3) GMO v EU 4) Situace s nakládáním v ČR 5) Reakce zájmových skupin 2 Zákon č. 78/2004 Sb. Směrnice Evropského parlamentu a Rady 2001/18/ES ze dne 12.3.
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 09 Genetické modifikace pro zlepšení výţivy člověka doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Nový trend zlepšení výţivy lidí Výţiva a zdraví člověka Prevence
Bi8240 GENETIKA ROSTLIN Prezentace 09 Genetické modifikace pro zlepšení výţivy člověka doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Nový trend zlepšení výţivy lidí Výţiva a zdraví člověka Prevence
vnášení cizorodých genů či jiné genetické informace do rostlin a manipulace s nimi
 TRANSGENOZE ROSTLIN vnášení cizorodých genů či jiné genetické informace do rostlin a manipulace s nimi (GMO) TRANSGENOZE trvalá - změna genetické informace je přenášená na potomstvo (infekce Agrobakteriem)
TRANSGENOZE ROSTLIN vnášení cizorodých genů či jiné genetické informace do rostlin a manipulace s nimi (GMO) TRANSGENOZE trvalá - změna genetické informace je přenášená na potomstvo (infekce Agrobakteriem)
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin Geneticky modifikované rostliny v zemědělské praxi - přínosy a rizika Bakalářská práce Vedoucí práce: Ing. Tomáš
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav biologie rostlin Geneticky modifikované rostliny v zemědělské praxi - přínosy a rizika Bakalářská práce Vedoucí práce: Ing. Tomáš
Geneticky modifikované potraviny a krmiva
 Geneticky modifikované potraviny a krmiva Co je to geneticky modifikovaný organismus (GMO)? Za GMO je považován organismus, s výjimkou člověka, jehož dědičná informace uložená v DNA byla změněna pomocí
Geneticky modifikované potraviny a krmiva Co je to geneticky modifikovaný organismus (GMO)? Za GMO je považován organismus, s výjimkou člověka, jehož dědičná informace uložená v DNA byla změněna pomocí
Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme
 Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme Doc. RNDr. Jindřich Bříza, CSc. ÚMBR BC AV ČR,v.v.i. & katedra genetiky PřF JU Branišovská 31, 370 05 České Budějovice GM crops
Geneticky modifikované rostliny - proč je potřebujeme a jak je získáváme Doc. RNDr. Jindřich Bříza, CSc. ÚMBR BC AV ČR,v.v.i. & katedra genetiky PřF JU Branišovská 31, 370 05 České Budějovice GM crops
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém
 Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Nové směry v rostlinných biotechnologiích
 Nové směry v rostlinných biotechnologiích Tomáš Moravec Ústav Experimentální Botaniky AV ČR Praha 2015-05-07 Praha Prvních 30. let transgenních rostlin * V roce 2014 byly GM plodiny pěstovány na ploše
Nové směry v rostlinných biotechnologiích Tomáš Moravec Ústav Experimentální Botaniky AV ČR Praha 2015-05-07 Praha Prvních 30. let transgenních rostlin * V roce 2014 byly GM plodiny pěstovány na ploše
Investujeme do naší společné budoucnosti
 Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Modifikace dědičné informace rostlin
 Modifikace dědičné informace rostlin Lukáš Fischer, KFR PřF UK Jak zlepšit vlastnosti rostlin Principy a klasické způsoby přípravy geneticky modifikovaných rostlinných buněk a celých rostlin Genový přenos
Modifikace dědičné informace rostlin Lukáš Fischer, KFR PřF UK Jak zlepšit vlastnosti rostlin Principy a klasické způsoby přípravy geneticky modifikovaných rostlinných buněk a celých rostlin Genový přenos
STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010. M. Mendlová, V. Ostrý, J. Ruprich
 STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010 M. Mendlová, V. Ostrý, J. Ruprich Státní zdravotní ústav v Praze Centrum zdraví, výživy a potravin Oddělení analýzy
STUDIE GENOMON VÝSKYT GENETICKY MODIFIKOVANÝCH POTRAVIN V TRŽNÍ SÍTI V ČR V ROCE 2010 M. Mendlová, V. Ostrý, J. Ruprich Státní zdravotní ústav v Praze Centrum zdraví, výživy a potravin Oddělení analýzy
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Molekulární biotechnologie. Nový obor, který vznikl koncem 70. let 20. století (č.1)
") Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO)
") Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO) Stanovisko Věc: Stanovisko ČK GMO k žádosti o dovoz řezaných květů karafiátů linie FLO-40685-2 do EU Stav
Česká komise pro nakládání s geneticky modifikovanými organismy a genetickými produkty (ČK GMO) Stanovisko Věc: Stanovisko ČK GMO k žádosti o dovoz řezaných květů karafiátů linie FLO-40685-2 do EU Stav
Molekulární biotechnologie č.9. Cílená mutageneze a proteinové inženýrství
 Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Transgenoze a reverzní genetika. Metody transformace rostlinných buněk Rekombinace
 Transgenoze a reverzní genetika Metody transformace rostlinných buněk Rekombinace Využití transgenoze 2 Mutageneze (ztráta funkce) Využití transgenoze 3 Charakterizace promotoru na základě exprese reportérového
Transgenoze a reverzní genetika Metody transformace rostlinných buněk Rekombinace Využití transgenoze 2 Mutageneze (ztráta funkce) Využití transgenoze 3 Charakterizace promotoru na základě exprese reportérového
Transgeneze u ptáků: očekávání vs. realita
 Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Biologické základy rostlinné produkce
 Zemědělství Biologické základy rostlinné produkce C3 C4 CAM Typy fotosyntézy C3-C4 Účinnost fotosyntézy ze 100% slunečního světla je 47% mimo použitelné vlnové délky ze zbylých 53% (400--700nm) -30%-fotonů
Zemědělství Biologické základy rostlinné produkce C3 C4 CAM Typy fotosyntézy C3-C4 Účinnost fotosyntézy ze 100% slunečního světla je 47% mimo použitelné vlnové délky ze zbylých 53% (400--700nm) -30%-fotonů
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rostlinné biotechnologie
 Fyziologie rostlin Letní semestr 2013 Rostlinné biotechnologie Lukáš Fischer klasické šlechtění příprava geneticky modifikovaných rostlin příklady praktického využití GM rostlin využití GM rostlin v exp.
Fyziologie rostlin Letní semestr 2013 Rostlinné biotechnologie Lukáš Fischer klasické šlechtění příprava geneticky modifikovaných rostlin příklady praktického využití GM rostlin využití GM rostlin v exp.
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE)
") MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
ZÁKLADY BAKTERIÁLNÍ GENETIKY
 Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Nové genové techniky, potraviny a monitoring
 21. Konference Monitoringu 2016, SZÚ, Milovy, 6.10. 2016 Nové genové techniky, potraviny a monitoring Veronika Kýrová Vladimír Ostrý Pavla Surmanová Ivana Procházková - Jiří Ruprich Podpořeno MZ ČR RVO
21. Konference Monitoringu 2016, SZÚ, Milovy, 6.10. 2016 Nové genové techniky, potraviny a monitoring Veronika Kýrová Vladimír Ostrý Pavla Surmanová Ivana Procházková - Jiří Ruprich Podpořeno MZ ČR RVO
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Transformace rostlin. úvod metody
 Transformace rostlin úvod metody Biotechnologie metody využívající různé organismy ve prospěch člověka a současně pozměňující jejich vlastnosti tak, aby byly co nejlépe využitelné výrobní postupy: získávání
Transformace rostlin úvod metody Biotechnologie metody využívající různé organismy ve prospěch člověka a současně pozměňující jejich vlastnosti tak, aby byly co nejlépe využitelné výrobní postupy: získávání
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Modifikace dědičné informace rostlin II
 Modifikace dědičné informace rostlin II Lukáš Fischer, KFR PřF UK Obsah přednášky Jak zlepšit vlastnosti rostlin Principy přípravy GMR Příprava genových konstruktů Genový přenos do jaderného a plastidového
Modifikace dědičné informace rostlin II Lukáš Fischer, KFR PřF UK Obsah přednášky Jak zlepšit vlastnosti rostlin Principy přípravy GMR Příprava genových konstruktů Genový přenos do jaderného a plastidového
Klonování a genové inženýrství
 Klonování a genové inženýrství KBC/BAM Ivo Frébort Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
Klonování a genové inženýrství KBC/BAM Ivo Frébort Klonování a genetické modifikace Sci-fi Skutečnost Dolly the Sheep Genetické modifikace a baktérií a kvasinek - Běžná praxe Nadexprese proteinů Velkoobjemové
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Použití transgenoze při šlechtění rostlinje třeba se obávat?
 Poslanecká sněmovna parlamentu ČR 3. května 2017 Použití transgenoze při šlechtění rostlinje třeba se obávat? Mgr. Tomáš Moravec, PhD., Ústav Experimentální Botaniky AV ČR Laboratoř virologie Praha Modifikování
Poslanecká sněmovna parlamentu ČR 3. května 2017 Použití transgenoze při šlechtění rostlinje třeba se obávat? Mgr. Tomáš Moravec, PhD., Ústav Experimentální Botaniky AV ČR Laboratoř virologie Praha Modifikování
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Biotechnologie a genové inženýrství rostlin (BAGIR)
") Biotechnologie a genové inženýrství rostlin (BAGIR) zodpovídá: spolupřednášející: rozsah: počet kreditů: semestr: stupeň: Prof.RNDr. Zdeněk Opatrný CSc. Doc.RNDr. Jindřich Bříza, CSc. Ing. Miluše Dvoržáková-Kusendová
Biotechnologie a genové inženýrství rostlin (BAGIR) zodpovídá: spolupřednášející: rozsah: počet kreditů: semestr: stupeň: Prof.RNDr. Zdeněk Opatrný CSc. Doc.RNDr. Jindřich Bříza, CSc. Ing. Miluše Dvoržáková-Kusendová
Molekulární biotechnologie č.10c. Využití poznatků molekulární biotechnologie. Využití škrobu, cukrů a celulózy.
 Molekulární biotechnologie č.10c Využití poznatků molekulární biotechnologie. Využití škrobu, cukrů a celulózy. Využití škrobu, cukrů a celulózy Zejména v potravinářském průmyslu Škrob je hydrolyzován
Molekulární biotechnologie č.10c Využití poznatků molekulární biotechnologie. Využití škrobu, cukrů a celulózy. Využití škrobu, cukrů a celulózy Zejména v potravinářském průmyslu Škrob je hydrolyzován
GENETIKA dědičností heredita proměnlivostí variabilitu Dědičnost - heredita podobnými znaky genetickou informací Proměnlivost - variabilita
 GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316
 Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Využití houbových organismů v genovém inženýrství MIKROORGANISMY - bakterie, kvasinky a houby využíval
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Využití houbových organismů v genovém inženýrství MIKROORGANISMY - bakterie, kvasinky a houby využíval
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
BAKTERIÁLNÍ REZISTENCE
 BAKTERIÁLNÍ REZISTENCE Petr Zouhar, Fyziologický ústav AV ČR, v. v. i.; UK v Praze, PřF, Katedra fyziologie V této úloze se v hrubých rysech seznámíte s některými metodami používanými v běžné molekulárně
BAKTERIÁLNÍ REZISTENCE Petr Zouhar, Fyziologický ústav AV ČR, v. v. i.; UK v Praze, PřF, Katedra fyziologie V této úloze se v hrubých rysech seznámíte s některými metodami používanými v běžné molekulárně
Autor: Clive James, zakladatel a předseda představenstva ISAAA. Věnováno památce zesnulého laureáta Nobelovy ceny za mír, Normanovi Borlaugovi
 (Globální přehled o komerčně pěstovaných biotechnologických/gm plodinách: 2009) Autor: Clive James, zakladatel a předseda představenstva ISAAA Věnováno památce zesnulého laureáta Nobelovy ceny za mír,
(Globální přehled o komerčně pěstovaných biotechnologických/gm plodinách: 2009) Autor: Clive James, zakladatel a předseda představenstva ISAAA Věnováno památce zesnulého laureáta Nobelovy ceny za mír,
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Zdrojem je mrna. mrna. zpětná transkriptáza. jednořetězcová DNA. DNA polymeráza. cdna
 Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Protinádorová imunita. Jiří Jelínek
 Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Rostliny a lidstvo. Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin
 Rostliny a lidstvo Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin Čím se živíme Čím se živíme Čím se živíme Čím se živíme Čím se živíme
Rostliny a lidstvo Reklama na: MB130P77 Rostliny a rozkvět a pád lidské civilizace MB130P19I Biotechnologie a genové inženýrství rostlin Čím se živíme Čím se živíme Čím se živíme Čím se živíme Čím se živíme
R o z h o d n u t í. rozhodlo
 Ministerstvo životního prostředí odbor environmentálních rizik Praze dne 30. dubna 2007 Čj.: 4675/ENV/07 R o z h o d n u t í Ministerstvo životního prostředí jako správní úřad příslušný podle 5 zákona
Ministerstvo životního prostředí odbor environmentálních rizik Praze dne 30. dubna 2007 Čj.: 4675/ENV/07 R o z h o d n u t í Ministerstvo životního prostředí jako správní úřad příslušný podle 5 zákona
Speciace neboli vznik druhů. KBI/GENE Mgr. Zbyněk Houdek
 Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
GENETICKÁ TRANSFORMACE OBILOVIN
 GENETICKÁ TRANSFORMACE OBILOVIN Pokročilé biochemické a biotechnologické metody 2.11.2015 K. Holubová ÚVOD Kukuřice pšenice rýže - ječmen Obiloviny: potrava, krmivo, průmyslní výroba Klimatické změny:
GENETICKÁ TRANSFORMACE OBILOVIN Pokročilé biochemické a biotechnologické metody 2.11.2015 K. Holubová ÚVOD Kukuřice pšenice rýže - ječmen Obiloviny: potrava, krmivo, průmyslní výroba Klimatické změny:
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/ B.Mieslerová (KB PřF UP v Olomouci)
") Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 2011 B.Mieslerová (KB PřF UP v Olomouci) VYUŽITÍ HOUBOVÝCH ORGANISMŮ V GENOVÉM INŽENÝRSTVÍ MIKROORGANISMY
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 2011 B.Mieslerová (KB PřF UP v Olomouci) VYUŽITÍ HOUBOVÝCH ORGANISMŮ V GENOVÉM INŽENÝRSTVÍ MIKROORGANISMY
Oligobiogenní prvky bývají běžnou součástí organismů, ale v těle jich již podstatně méně (do 1%) než prvků makrobiogenních.
 než prvků makrobiogenních.") 1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
doc. RNDr. Milan Bartoš, Ph.D.
 doc. RNDr. Milan Bartoš, Ph.D. Konference Klonování a geneticky modifikované organismy Parlament České republiky, Poslanecká sněmovna 7. května 2015, Praha Výroba léků rekombinantních léčiv Výroba diagnostických
doc. RNDr. Milan Bartoš, Ph.D. Konference Klonování a geneticky modifikované organismy Parlament České republiky, Poslanecká sněmovna 7. května 2015, Praha Výroba léků rekombinantních léčiv Výroba diagnostických
Proč nemáme vakcínu proti HIV-1?
 UAB THE UNIVERSITY OF ALABAMA AT BIRMINGHAM UAB Proč nemáme vakcínu proti HIV-1? Jiří Městecký Department of Microbiology University of Alabama at Birmingham USA Pandemie HIV/AIDS 1983 2004 2010 2020 Popsání
UAB THE UNIVERSITY OF ALABAMA AT BIRMINGHAM UAB Proč nemáme vakcínu proti HIV-1? Jiří Městecký Department of Microbiology University of Alabama at Birmingham USA Pandemie HIV/AIDS 1983 2004 2010 2020 Popsání
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Klonování. Co to vlastně je?
 Kristýna Boháčová Klonování Co to vlastně je? Klonování je podle běžné definice vytváření nového jedince geneticky identického (shodného) s předlohou Tyto dva jedinci se poté označují jako klony Tento
Kristýna Boháčová Klonování Co to vlastně je? Klonování je podle běžné definice vytváření nového jedince geneticky identického (shodného) s předlohou Tyto dva jedinci se poté označují jako klony Tento
RNA interference (RNAi)
") Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE
 ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
Transformace ptdna tabáku genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace
 Transformace ptdna tabáku fúzním genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace Jiřich ich BřízaB 1,, Josef Vlasák 1, Štěpán n Ryba, Viera Ludvíkov ková 3, Hana Niedermeierová
Transformace ptdna tabáku fúzním genem E7/GUS a eliminace selekčního genu za využití homologní rekombinace Jiřich ich BřízaB 1,, Josef Vlasák 1, Štěpán n Ryba, Viera Ludvíkov ková 3, Hana Niedermeierová
GM kukuřice. 0,0004% DNA kukuřice
 Kolik je DNA v krmivech a potravinách? 0,005 až 0,02 % sušiny GM kukuřice cizí gen 4000 písmen 0,0004% DNA kukuřice Hypotetický příklad: brojler Krmná dávka - jen kukuřice Veškerá kukuřice jen GMO Brojler
Kolik je DNA v krmivech a potravinách? 0,005 až 0,02 % sušiny GM kukuřice cizí gen 4000 písmen 0,0004% DNA kukuřice Hypotetický příklad: brojler Krmná dávka - jen kukuřice Veškerá kukuřice jen GMO Brojler
Hodnocení rizik geneticky modifikovaných rostlin pro životní prostředí
 Klasifikace: Draft Pro vnitřní potřebu VVF Oponovaný draft Pro vnitřní potřebu VVF Finální dokument Pro oficiální použití Deklasifikovaný dokument Pro veřejné použití Název dokumentu: Hodnocení rizik geneticky
Klasifikace: Draft Pro vnitřní potřebu VVF Oponovaný draft Pro vnitřní potřebu VVF Finální dokument Pro oficiální použití Deklasifikovaný dokument Pro veřejné použití Název dokumentu: Hodnocení rizik geneticky
Základní škola Karviná Nové Město tř. Družby 1383
 Základní škola Karviná Nové Město tř. Družby 1383 Projekt OP VK oblast podpory 1.4 Zlepšení podmínek pro vzdělávání na středních školách Registrační číslo projektu: CZ.1.07/1.4.00/21.3526 Název projektu:
Základní škola Karviná Nové Město tř. Družby 1383 Projekt OP VK oblast podpory 1.4 Zlepšení podmínek pro vzdělávání na středních školách Registrační číslo projektu: CZ.1.07/1.4.00/21.3526 Název projektu: