Už od počátků molekulární biologie a centrálního dogmatu bylo jasné, že produktem některých genů jerna a ne protein. Týkalo se to ribosomálních genů,
|
|
|
- Ján Kovář
- před 9 lety
- Počet zobrazení:
Transkript
1
2

3 Už od počátků molekulární biologie a centrálního dogmatu bylo jasné, že produktem některých genů jerna a ne protein. Týkalo se to ribosomálních genů, genů pro trna, splicesomálnía snorna, které se účastnily syntézy proteinů a dalších buněčných funkcí. Záhy po osekvenováníprvních genomů a jejich transkriptomůse zjistilo, že velkou část polya+ RNA nelze přiřadit kžádnému protein kódujícímu genu (u myši a člověka cca 50%). RNA polymeráza je přítomna vpodstatě na všech místech vgenomu, a i oblasti, které byly považovány za transkripčně neaktivní (pouště) jsou transkribovány (pervasivní transkripce). Spektrum těchto nekódujících RNAs(non-codingRNA) je veliké, liší se velikostí (long > 200 bp, a short bp) a stabilitou. Jedním zpřekvapení, která vyplynula z analýz genomů a transkriptomů, byl fakt, že nekódující RNA (non-protein-codingrna, ncrna) je vdrtivé většině exprimována vývojově specificky. Savčí transkriptomyse skládají zobrovsky variabilního spektra RNA, zahrnujícího překrývající se transkriptyzintronů i mezigenovýchoblastí, sensei antisense, srůznými pozicemi startu a konce, splicingema expresním patternemlišícím se mezi buňkami, typicky exprimovanými vmalém množství. Tyto transkriptybyly často považovány za pozadí při sekvenování, zřejmě díky nízké hladině transkriptu, což může být dané i omezeným množstvím buněk, kde ktranskripci konkrétního úseku dochází. Nové sensitivní technika sekvenovánícdnaodhalily, že domnělé genové pouště jsou transkribované a produkují velké množství sestřihovaných produktů. Technikami, které nejsou založeny na izolaci RNA pomocí oligo(dt), se ukázalo, že téměř ½ RNA vlidských buňkách nemá polyakonec, takže byla pominuta ztechnických důvodů. Tato RNA je zřejmě syntetizována RNA polymerázou III a strukturou se značně liší od polya+ RNA, syntetizované RNA polymerázou II.
 jsou transkribovány (pervasivní transkripce).")
4
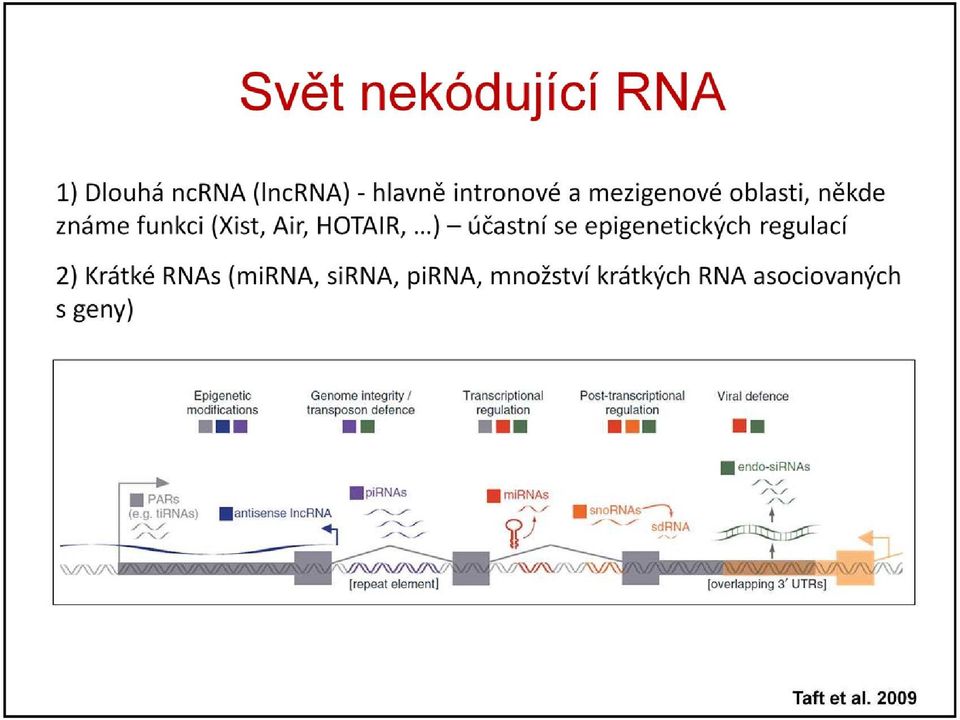
5 Velké množství RNAs bylo nalezeno na hranicích různých částí genů: promoter associated srnas(pasrs), promoterassociatedlrnas(palrs), terminator-associatedrnas(tasrs), transcription-initiation RNAs(tiRNAs), transcription start site-associated RNAs(tssa- RNAs), promoter upstream transcripts(prompts). Není jisté jakou mají tyto RNAs asociované spromotorem funkci. Možné je, že mohou udržovat chromatin ve stavu přístupném transkripci a RNA polymerázu aktivní, což by obojí urychlilo transkripci genu vpřípadě potřeby. -tirnaje nová skupina malých RNAs, odvozených zmíst iniciace transkripce (downstreamod těchto míst), které jsou jen u zvířat. Jejich možnou funkcí je možná označit/regulovat pozici prvního nukleosomu. - splice site RNAs(spliRNA) jsou malé RNAs lokalizovaných na místech sestřihu (splice sites), opět byly nalezeny jen u zvířat. splirnai tirnajsou exprimovány velmi slabě, čili snadno uniknou sekvenování. Takových typů lokusspecifických RNAsmůže být mnoho, je potřeba je objevit. Byly nalezeny u člověka, myši, drozofily, háďátka a houbovce, ale ne u rostlin.
, které jsou jen u zvířat. Jejich možnou funkcí je možná označit/regulovat pozici prvního nukleosomu.")

6 První mirna(lin-4) byla objevena u C. elegans, má funkci vnačasování vývoje (developmentaltiming) a nekóduje protein, jen RNA (60 bp prekurzor a 22 bpmirna). Princip regulace pomocí lin-4 je následující: lin-4 mirna se páruje s 3 UTR mrnalin-14, což způsobuje silencing. Toto bylo původně považováno za ojedinělý jev, ale pak byly objeveny další případy.

7 Princip RNAi Malé regulační RNAsjsou nekódující RNAs, které umlčují geny nebo ničí cílovou RNA na základě komplementarity sekvencí. Jsou 3 hlavní skupiny: sirna(shortinterferingrna), mirna(microrna) a pirna(piwi interacting RNA). sirnasnižuje množství cílové RNA prostřednictvím jejího štěpení. Jsou odvozeny zdlouhých dsrnas, které vznikají při replikaci viru, šíření mobilního elementu nebo experimentální RNAi. Tato RNA je naštěpenarnázouiii jménem Dicer(RNázyIII štěpí dsrna), která funguje jako pravítko a odměřuje délku výsledných produktů, takže vznikají fragmenty dlouhé bp. Jeden řetězec (guidestrand) ze vznikéhofragmentu je následně vložen do komplexu RISC (RNA-inducedsilencingcomplex) na protein Argonaute. Toto vložení se odehrává na RISC loadingcomplex, který se skládá zargonaute, Dicerua dsrnabindingprotein. Druhý řetězec sirna(passangerstrand) je naštěpenargonautem a ostraněn. Argonaut poté využije guidestrandkvazbě cílové RNA, která musí být perfektně komplementární a naštěpí ji. RNA je poté uvolněna. Kromě této funkce existuje ještě endogenní sirna, která je kódována genomem. Přestože exogenní sirnajsou známé relativně dlouho, endogenní sirnas(endosirnas) byly objeveny relativně nedávno u savců a octomilky (mimo zvířata i u rostlin a kvasinek).tato endo-sirnase účastní obrany proti virům, umlčení transposonů, chromatin remodelingua post-transkripční regulace genů. mirna jsou malé RNA kódované vlastním genomem. U rostlin je princip umlčení mrna stejný jako u sirna, ale u zvířat je RNA jsou dva způsoby jak s RNA naložit. mirnaje
, která funguje jako pravítko a odměřuje délku výsledných produktů, takže vznikají fragmenty dlouhé 21-25 bp.")
8 transkribována zmirnagenů ve formě primárního transkriptu(pri-mirna) se strukturami stem-loop dlouhými cca bp. Vlásenkové struktury jsou v jádře naštěpené komplexem Drosha-DGCR8, čímž vzniká pre-mirna(droshaje RNázaIII). Vcytoplazmě je tato struktura dále štěpena Dicerem, takže vzniká mirna-mirna* duplex (mirna= guide, mirna* = passanger). Guide strand je naložen na Argonauta a hledá komplementání RNA. Podle stupně komplementarity je buď cílová mrnanaštěpena(téměř perfektní komplementarita), nebo je potlačena translace mrna(částečná kompementarita). pirnajsou krátké bprna, které umlčují transposonyvživočišných zárodečných buňkách. pirnajsou jednořetězcové, takže nepotřebují Dicer. pirnaindukují štěpení transkriptu mobilního elementu (ME), což je provedeno proteiny ze skupiny PIWI(= p- element inducedwimpytestis) patřící dorodiny Argonaute (PIWI, Aubergine, Argonaute 3). PIWInebo AUBštěpí sensetranskript, vzniká sensepirna, ta asociuje sago3, ten štěpí antisensetranskripttransposonu, vznikne antisensepirna, asociuje spiwia AUB, což vede kštěpení sensetranskriptutransposonu. 7
, nebo je potlačena translace mrna(částečná kompementarita).")
9 pirnasjsou malé RNA, které jsou exprimovány vzárodečné linii mnoha živočichů. Jsou asociované spiwiproteiny, které patří do rodiny AGO proteinů, specificky exprimovaných vzárodečné linii. Stávají se součásti pirisckomplexu, který brání expanzi MEs, což je nezbytné pro správný vývoj gonád a tato funkce je u mnohobuněčných živočichů konzervovaná. Proteiny z PIWI rodiny D. melanogaster(piwi, Aubergine (AUB)) jsou potřeba pro samčí i samičí fertilitu, AGO3 pro samičí fertilitu a částečně pro samčí. U myši knock-outgenů PIWIrodiny (MILI, MIWI2) vede kaktivaci LINEsa LTR retrotransposonůvsamčí zárodečné linii, zastavení gametogeneze a kompletní sterilitě samce. Na samici nemá vliv. Sterilita je zřejmě způsobena aktivitou MEs, která vede kdvouřetězcovýmzlomům, které mají za následek zastavení vkontrolním bodě buněčného cyklu, což vede k ukončení dělení buněk. Navíc byl pozorován pokles metylace CpG, která je potřeba pro umlčení MEs, čili myši bez těchto genů nejsou schopny ustavit metylaci MEsde novo během spermatogeneze, což je zřejmě další role pirisc(navést metylační mašinérii k MEs). pirnasekvence jsou velice početné (stovky tisíc až miliony), ale zabírají relativně malé oblasti genomu (několik kbp až více než 200 kbp), vyskytují se v klastrech.
 vede kaktivaci LINEsa LTR retrotransposonůvsamčí zárodečné linii, zastavení gametogeneze a kompletní sterilitě samce. Na samici nemá vliv.")
10 pirna může ovlivňovat expresi i protein kódujících genů. Příkladem je pirna lokus Su(ste) u drozofily, který je lokalizovaný na Y chromosomu, kde se zřejmě vyvinul zkopie genu Stellate, lokalizovaného na X. Su(ste) reguluje expresi Stellate genů(repetitivní protein kódující geny), které jsouexprimovány vtestesa při overesxpresise zjejich produktů tvoří krystaly, které poškozují buňky a brání spermatogenezi. Dalším genem regulovaným pirnaje fasciclin3(kódující immunoglobulin-likecell adhesionmolecule) exprimovaný vovariích drozofily. OverexpreseFas3brání promíchání somatických a zárodečných buněk vováriícha vede kselhání oogeneze. Knock-outPIWIproteinů vede koverexpresifas3, což napovídá, že pirnaslouží kjeho regulaci.
 exprimovaný vovariích drozofily.")
11 Hybridní dysgeneze je jev, při kterém je potomstvo křížení určitých linií neplodné, zatímco potomstvo reciprokého křížení je v pořádku. Slavným případem je křížení drozofilíchlinií P (nesoucí transposonzvaný P-element) a M (P-element nemají). Při křížení samice P se samcem M je potomstvo plodné, při reciprokém křížení (samice M a samec P) je všechno potomstvo neplodné (dystrofie gonád, aktivace P elementů, chromosomální přestavby, vyšší mutageneze). Dlouho se předpokládalo, že na umlčení P-elementu je zodpovědný nějaký maternálnífaktor, který samice umísťují do vajíček. Dnes víme, že tímto faktorem je pirna, která iniciuje vlastní amplifikaci a tím vede kochraně před MEsod samce.
. Dlouho se předpokládalo, že na umlčení P-elementu je zodpovědný nějaký maternálnífaktor, který samice umísťují do vajíček.")
12

13 Distribuce mirna v genomu není náhodná: 1) mirna geny jsou často organizovány v klastrech (např. u člověka cca 40% mirna genů). Důvodem je zřejmě to, že takto organizované geny lze koexprimovatjako jednu pri-mirna, i když ne vždy to tak je. 2) Více než polovina savčích mirnaje vintronech genů, které regulují, a mnoho znich je s tímto genem koexprimováno. 3) Mnoho mirnaje lokalizovánove fragilních místech a oblastech, které jsou spojené srakovinou, což naznačuje, že se účastní procesů spojených sbuněčným růstem, proliferací a diferenciací. -Expresní patternmirnaje různý některé jsou exprimovány vurčitou dobu během vývoje, zatímco jiné jsou exprimovány kontinuálně. Množství produktů mirnagenů je až 500xvíc než mrna. -Protože se jedná o jednoduché a malé geny, vznikají častěji než klasické protein kódující geny a představují tak pružnější mechanismus regulace genomu. Je zajímavé, že přestože mirnanajdeme u živočichů i u rostlin, není známa žádná mirna, která by se vyskytovala u obou těchto skupin.
 Mnoho mirnaje lokalizovánove fragilních místech a oblastech, které jsou spojené srakovinou, což naznačuje, že se účastní procesů spojených sbuněčným růstem, proliferací a diferenciací.")
14 Pro interakci mirnas mrnaje klíčová oblast umístěná v pozicích 2-7 od 5' konce mirna, tzv. seed. Na základě komplementarity mezi seedoblastí a cílovou oblastí v (obvykle) 3' UTRmRNAdochází k interakci těchto RNA, což vede k represi translace zúčastněné mrna. Pokud je seednebo cílová oblast zasažena mutací, může to vést k nerozpoznání mrnaa tím k selhání regulace genu. Nejčastějším cílem mirnaje 3 UTR, ale jsou známy i cíle v 5 UTRa kódující oblasti, ty jsou ale mnohem méně významné. Jedna mrnamůže mít více cílových oblastí pro několik mirnas, jejich umlčovací účinek je pak silnější.

15 U živočichů snižuje mirnaexpresi cílových mrnadvěma způsoby v závislosti na stupni komplementarity mezi seeda cílovou oblastí. Perfektní nebo téměř perfektní komplementarita vede k rozštěpení a degradaci mrna, zatímco částečná komplementarita vede k inhibici translace. Na rozdíl od rotlin, kde je mrnatakto degradována, u živočichů převládainhibice translace.

16 Nepřítomnost mirnau jednobuněčných organismů a další data indikují, že mirnasjsou klíčové pro diferenciaci buněk i tkání a pro udržení buněk v diferencovaném stavu. -Nediferencované nebo málo diferencované buňky nepotřebují mirnakpřežití (myší ESCbez mirnapřežijí, ale nediferencují se, zárodečné buňky zebřičky nepotřebují mirna k přežití a udržení linie. -Většina mirnanení exprimována během raného vývoje zebřičky, kdy jsou buňky nediferencované, ale vykazují vysokou tkáňově specifickou expresi později, kdy se určují buněčné typy. -Mnoho nádorových buněk redukuje expresi mirnave srovnání spůvodní diferencovanou buňkou Př. let-7 mirnaje downregulovanáu různých rakovin plic a zpokusů na tkáňových kulturách víme, že je schopná potlačovat růst nádorových buněk. let-7 může sloužit i pro predikci vývoje rakoviny, protože pacienti snejnižší expresí let-7 mají i nejmenší šanci na přežití. -Transfekce HeLabuněk mir-1 (svalově specifická) a mir-124 (mozkově specifická) změní expresní profil buněk na svalové/mozkové.

17 let-7 byla jednou zprvních objevených mirna, je u metazoí vysoce konzervovaná a funguje jako hlavní regulátor vývoje a diferenciace vrannémembryu i vkomplexních tkáních dospělce, jako je mozek (u Danio, Ceanorhabditis, myši a drozofily. Cílem let-7 jsou významné regulátory buněčného cyklu, jako Ras a Cdk6. Každý krok biogeneze let-7 je pečlivě regulovaný. Diferenciační faktory jako je Notch, indukují transkripci, pluripotenčnífaktory jako c-myc, transkripci reprimují. Pluripotenčnífaktor LIN28se váže na smyčku (loop) pri-let-7, což inhibuje Droshu. Naopak let-7 inhibuje LIN28, c-myc a c-mycaktivatinggene IMP-1 vazbou na jejich 3 UTR.

18

19 Genová duplikace mirnas, které jsou homologické a mají téměř identické seedsekvence, jsou řazeny do rodin. Vlidském genomu je vrodinách o 2 a více členech 1/3 mirnas, u myši 38%, u C. elegans10%, drozofily 13%. Tyto mirnarodiny jsou paralognísekvence, které vznikly duplikací původní mirnagenu. Pokud se jednalo o lokální duplikaci, jsou geny řazeny těsně za sebou v tandemu a často jsou součástí stejného transkruptu. Pokud to nebyla lokální duplikace, jsou geny klokalizoványna vydálenýchčástech genomu, třeba i na jiných chromosomech. Většina ne-lokálních duplikací u obratlovců je důsledkem duplikace celého genomu, které se vevoluci obratlovců odehrály. Duplikace genu následovaná subfunkcionalizacínebo neofunkcionalizacíjsou největším zdrojem nových mirna.

20 Introny Mnoho mirnasse vyskytuje vintronech protein-kódujících genů (1/2 lidských, 20% u C. elegansvintronech ve stejné orientaci jako hostitelský gen). Introny jsou výhodným místem pro vznik nových mirnas, protože není potřeba nový promotor (jsou koexprimoványs genem, i když některé mirnasvintronech mají vlastní promotor). Bylo pozorováno, že nové mirnasspecifické pro určitou skupinu jsou častěji lokalizované v intronech (intronic exaptation).
.")
21 AntisensetranskriptymiRNAsmají pochopitelně schopnost skládat se do vlásenek, takže, pokud jsou transkribovány, mohou získat vlastní novou funkci. Produkce antisense mirnasbyla skutečně pozorována a je zdrojem nových mirnagenů.
22 Pseudogeny, snornas a trnas vliteratuře informace o mirnasztěchto zdrojů x problematické, protože snornasa trnasse taky mohou účastnit mirnamašinérie, takže je obtížné rozlišit co je skutečná mirnaa o by-productnormální biosyntézy snornaa trna. TEs U člověka identifikováno 278 mirnas, které jsou určitě/možná ztes. Je jich více mezi mladšími, méně konzervovanými mirnasa mirnasspecifickými pro určitou skupinu živočichů (linagespecific). Problematická je jejich lokalizace vgenomu kvůli repetitivnímu okolí a taky možná některé piwirnas chybně považovány za mirnas.
23 Vlidském genomu byly předpovězeny stovky tisíc mirnas+ velká část genomu se transkribuje (pervasivetranscription, všudypřítomná transkripce). Čili vgenomu je mnoho substrátu pro vznik nových mirnas. Vlásenka, kterou by zpracovala Droshaa Dicermusí mít přesnou sekundární strukturu, což je splněno jen u malé části sekvencí, a u těch se prokázalo, že jsou skutečné mirnas. Stovky sekvencí vgenomu tvoří suboptimálnívlásenky ty jsou na půl cesty. Čili pro vznik nové mirnaje potřeba více kroků. Nové mirnasmají náhodné cíle (seedje krátká sekvence), což může být fatální. Model vzniku mirnasje tento: nová mirnaje nejprve transkribována slabě a jen vurčitých tkáních, aby se omezil negativní efekt náhodné interakce scílovými geny. Postupně jsou škodlivé transkriptyodstraněny selekcí a je zesílena transkripce a doba působení nové mirna. Tento model je podpořen faktem, že konzervované sekvence jsou obecně transkribovány silněji než méně konzervované mirnas. Ztoho také vyplývá, že mirnas, které jsou transkribovány slabě, jsou teprve ve fázi testování a nemají žádnou regulační roli.
24
25 mirnapassangerstrandsbyly považovány za biologicky inertní, ale vbuňkách se mohou vyskytovat vrelevantním množství (i když méně, než guidestrands), být asociované sargonaoutema regulovat expresi genů. Představují tedy materiál pro diverzifikaci funkce mirna. U duplikovaných mirnagenů může dojít kposunu transkripce ve prospěch druhého ramene a tím kzisku nové funkce. Armswitchingje častý jev, pozorovaná u mnoha rodin mirnadrozofily a háďátka, u 11% ortologůu hmyzu, a je považován za jeden ze základních mechanismů diverzifikace mirna. Mechanismus, jak ktomu dochází, není jasný.
26 Zatímco výměna ramen nemění sekvenci genu, jen frekvenci přepisování jednotlivých ramen, posun vlásenky mění gen výměnou jednoho ramene za jinou sekvenci. To se děje především výměnou mezi členy mirnarodiny, které jsou lokalizovány blízko sebe. Tento jev byl identifikován u 8 z15 mirnarodin nematoda je to obecný mechanismus evoluce mirna.
27 Zdá ze, že RNA je prostředníkem interakcí mezi prostředím a epigenomem. RNA je předmětem editinguzávislém na kontextu. Editingspočívá vdeaminaci bází a je prováděn dvěmitřídami enzymů: ADAR (adenosin deaminázami) -změna A ani, který se chová jako G, a APOBEC(ApoBeditingcomplex) měnící C anu u RNA i DNA, je specifický pro obratlovce. U živočichů existují 3 ortologyadar: ADAR1a ADAR2, které jsou u bezobratlých i obratlovců, jejich knockoutu myši je letální, a ADAR3, specifický pro mozky obratlovců. O regulaci editinguse moc neví, ale je prokázáno, že je napojen na signální dráhy a tím na signály vnějšího prostředí. A>I editingbyl poprvé pozorován a generationago vdůležitých receptorech neuronů, jako receptorech glutamatu, GABA a serotoninu, kde mění sekvenci aminokyselin za účelem lepšího vyladění elektrofyziologických vlastností synapsí. Původně to bylo považováno za okrajovou věc, ale vroce 2004 bylo publikováno několik prací, které prokázaly, že se A>I editingtýká tisíců transkriptů. Většina editovaných míst je vnekódujících oblastech, což znamená, že editingovlivňuje regulační dráhy založené na RNA, čili potenciálně epigenetické procesy, které jsou základem fungování mozku a učení. Tyto studie navíc ukázaly, že u člověka došlo kmasivnímu nárůstu RNA editinguvporovnání smyší, navíc víc než 90% editace je v Alu. Alu jsou sekvence odvozené z funkční RNA sekvence (7SL RNA signal recognition particle), jsou specifické pro primáty a invadovalyjejich genomy vněkolika vlnách. Dnes tvoří 10,5% lidského genomu. Spolu snimi se vevoluci primátů zvyšovala i frekvence RNA editingua recentní (po rozdělení šimpanze a člověka) de novo inserce Alu jsou soustředěny hlavně do genů spojených sfunkcí neuronů a neurologických onemocnění. Alu byly půpodněpovažovány za odpadní (junk) DNA, ale tato nová zjištění přinášejí mnohem zajímavější interpretaci. Alu byly použity jako substrát pro RNA editinga umožnily zvýšení transkriptomickéa epigenomicképlasticity nutné kinterakci mezi prostředím a epigenomem. APOBECbyla poprvé objevena kvůli modifikaci mrnaapolipoproteinub, kde změnou C na U dělá předčasný stop kodon, a tím zkrácený protein ve střevech, zatímovjátrech je jeho nezkrácená verze. Z5 známých rodin APOBECjsou 2 specifické pro savce: 1) AID, která generuje hypermutacevimunoglobulinech AIS a je potřebná
28 k reprogramování buněk k pluripotenci. 2) APOBEC2 je potřebná k diferenciaci svalů. Zajímavé je, že mezi nervovým systémem a AIS je řada paralel. Např. přítomnost imunoglobulinových domén vmnoha povrchových receptorech neuronů, což indikuje, že se AIS savců vyvinul znervového systému a používají stejné mechanismy k vyladění svých receptorů. Zajímavá je rodina APOBEC3, která vznikla po oddělení vačnatců a placentálůa velmi expandovala vlinii primátů. Jednou její možnou funkcí je obrana proti mobilním elementům, což ovšem nevysvětluje její expanzi právě u primátů. Alternativní možností je, že se účastní domestikace transposonůvneuronech (APOBEC3G), což je podpořeno pozorováním, že de novo transpozice L1se odehrávají vkmenových buňkách neuronů a tím přispívají kindividuálnímu somatickému mozaicismuvmozku a komplexitě neuronové sítě. 26
29
30 Vlivem mutací vznikají v genech neustále nová cílová místa pro mirna. Pokud je jejich existence nevýhodná, jsou odstraněny selekcí. Pokud ale vznikají opakovaně příliš často, bude selekčně výhodné pozměnit cílové místo např. tím, že dojde k zkrácení 3 UTR, čímž ubude prostor pro vznik nových cílových míst. Skutečně bylo pozorováno, že housekeepergeny (např. ribosomalprotein-codinggenes) mají kratší 3 UTR, zatímco geny, jejichž exprese je časově a tkáňově specifická (např. transkripční faktory) mají 3 UTRmnohem delší s mnohem větším množstvím predikovaných mirnacílových míst.
31 Polymorfismus cílové oblasti může ovlivnit regulaci genu vytvořením nebo likvidací cílového místa pro mirna. U člověka byla nalezena řada SNPsv potenciálních cílových oblastech mirna, které by mohly roly při vzniku chorob, a přibývá důkazů o roli SNPsve vzniku rakoviny. Příkladem dopadu vzniku nového místa pro mirnaje změna v regulaci genu pro myostatin. Protein myostatinfunguje jako růstový diferenciační faktor inhibující vznik svalů a porucha v jeho regulaci, nulové mutace v kódující sekvenci genu nebo jeho receptoru vedou k nadměrnému růstu svaloviny. Přirozené nulové mutace v kódující sekvenci byly pozorovány u skotu, chrtů a člověka, u kterých způsobují nárůst svaloviny (a síly) až na dvojnásobek. U ovčího plemene Texelsheepse silně vyvinutou svalovinou došlo ke změně regulace genu pro myostatinvznikem nového cílového místa v3 UTRpro mir-1 a mir-126, což má podobné účinky na nárůst svaloviny jako vyřazení myostatinového genu.
32 Cichlidy představují druhově nejbohatší čeleď obratlovců. Více než 3000 druhů se nachází voblasti Střední a Jižní Ameriky, Africe, Madagaskaru a jižní Indii. Jsou výborným modelem studia speciace, opakovaně předvedly rychlou radiaci a sympatrickou speciaci. Nejzkoumanější jsou cichlidy zvelkých jezer ve východní Africe, kde se nachází 2000 druhů. Stovky druhů se vyvinuly zjedné linie během 1 milionu let. Jejich genomy jsou velice podobné -diverzita nukleotidů je 0,26%, (což je srovnatelné s diverzitou mezi člověkem a šimpanzem, zatímco mezi liniemi laboratorních zebřiček je diverzita 0,48%), ale diverzita morfologie a chování (zbarvení, potravní chování a rozmnožování, organizace mozku) je na úrovni obratlovčího řádu. Ukazuje se, že řada rozdílů mezi genomy linií malawských cichlid je lokalizována v cílových oblastech mirnagenů (86% cílových míst vykazuje variabilitu), čili za diverzitu cichlid by mohly stát regulace genů pomocí mirna. Konkrétní příklady změn v regulacích genů prostřednictvím mirna: 1) mir-181a mir23aregulují hoxa10, který je u ryb exprimovaný vpárových ploutvích a přilehlých svalech u myší (exprese ve svalech nohou) je při vytrvalostním tréninku mir-181 je upregulovaný a mir23a downregulovaný. Dravé cichlidy mají vcílovém místě mutaci, která je odlišuje od ostatních cichlid. 2) Změny ve smyslových orgánech -rozdíly vcílových místech pro mir-34 -váže se na gen crb1, který se podílí na morfogenezi a sensitivitě fotoreceptorů vevoluci cichlid zrak velmi důležitý pro ekologii a výběr partnera. -mir-200arozdíly ve vazbě na triobp(trio and F-actinbindingprotein), jehož mutace způsobuje ztrátu sluchu u východoafrických cichlid hraje sluch roli ve výběru partnera. 3) Imunita 4) Vývoj kostí a zubů

33 Evoluce mirna u živočichů Z osekvenovaných genomů máme informace o mirna různých skupin, z nichž vyplývá že: 1) mirnajako regulátory genů se objevily už na začátku evoluce živočichů. Např. mir-100 je u všech eumetazoí. 2) početná sada mirna(34) u prvo+ druhoústýchje zřejmě svědkem četných inovací na bázi bilaterií(další expanze mirna byla na bázi obratlovců a na linii placentálů 3) mirnagen, který se objeví u společného předka, je jen málokdy ztracen u dceřiných linií. Např. u druhoústýchbylo identifikováno pouze 12 ztrát mirnas, u skupiny Drosophilaobscuraztráta 9 mirnas. To v kombinaci se snadností, sjakou se generují nové mirnasvede kneustálému nárustumirnaběhem evoluce. Studium genomů zástupců druhoústých ukázala, že každá linie má cca 1/3 vlastních mirnas. Existuje přímá korelace mezi počtem mirnaa morfologickou komplexitou! To napovídá, že mirnabyla klíčová pro vznik komplexních organismů.
34 Na základě znalosti mirnasspolečných všem druhoústýma prvoústýmživočichům lze zrekonstruovat mirnasjejich společného předka. Na obrázku jsou seřazené do skupin podle místa exprese u nereidky.
35
, Porifera(houbovci) a Bilateria.")
36 Pro vznik nových mirnaobecně platí, že relativně snadno vznikají, ale vzácně se ztrácejí, což znich dělá výborný markerfylogeneze, který může pomoci vyřešit nejasnosti ve fylogenetických vztazích některých skupin. Příkladem je vztah mezi skupinami Cnidaria (žahavci), Porifera(houbovci) a Bilateria. Podle ESTsvypadají jejich vztahy takto: ((Porifera+ Cnidaria) + Bilateria), zatímco podle morfologie a molekulární biologie takto ((Cnidaria+ Bilateria) + Porifera). Podle mirnapatří Cnidariaa Bilateriak sobě, protože sdílejí mir-100, který chybí u houbovců, dále Cnidarianemají houbovčímirnasa houbovci žahavčí.
, protože buď mirnaa/nebo geny důležité")
37 Žebernatky jsou skupinou mnohobuněčných živočichů, jejíž fylogenetická pozice stále není jasná. Jednou z možností je umístění na bázi mnohobuněčných živočichů. Argumentem, který by pro toto možnost mohl svědčit, je absence mirnaa genů pro Droshua Pashu(protein důležitý pro fungování Droshy, u obratlovců zvaný DGCR8), protože buď mirnaa/nebo geny důležité pro její zpracování jsou přítomny u všech ostatních skupin mnohobuněčných živočichů, zatímco Choanoflagellata, sesterská skupina mnohobuněčných živočichů, je nemá. mirnaa její zpracování by tak vznikla až po odštěpení žebernatek od zbytku mnohobuněčných živočichů. Alternativním vysvětlením je druhotná ztráta mirnau žebernatek a genů pro její zpracování. Salpingoeca rosetta + Monosiga brevicollis(choanoflagellata) Capsaspora owczarzaki(filasterea) Sphaeroforma arctica(ichthyosporea)
38 mirnapatrně hrála roli i vevoluci lidského mozku. Vpráci Somelet al je popsán experiment, při kterém byla zkoumána RNA v přední části čelního laloku a mozečku člověka, šimpanze a makaka rhezusevrůzných postnatálních obdobích (materiál pro izolaci RNA byl odebraný znáhle zemřelých lidí a zvířat). Zjistilo se, že většina rozdílů mezi druhy je vexpresi konstitutivních genů (tj. těch, jejichž exprese není závislá na fázi vývoje), cca 80%, což není až tak zajímavé, protože rychlost jejich evoluce je u člověka i šimpanze srovnatelná. Mnohem zajímavější ale je, že vývojově specifické geny se vyvíjejí u člověka 3-5xrychleji než u šimpanze a děje se tak přes změny vregulačních faktorech, hlavně v mirnas. Evoluce je 2xrychlejší u předního laloku než u mozečku, tedy vmístě, které je zodpovědné za sociální inteligenci, plánování, abstraktní myšlení a operační paměť (workingmemory) (vs. mozeček -motorika, paměť a řeč). Byly nalezeny tři mirnas, které se podílejí na regulaci genů vneuronech, ztoho dvě vca 2+ signálních drahách, které se účastní plasticity neuronů, čili mirnamůže být zodpovědná za rozvoj kognitivních schopností člověka, jako je paměť a učení.
39 Protože mirnareguluje množství genů a účastní se proliferace a diferenciace buněk, je jasné, že změny v její expresi mohou způsobit rakovinu. Přibývají informace o změnách exprese mirnau konkrétních rakovin, což z nich zároveň dělá zajímavý potenciální markera cíl protinádorové terapie. mirnamůže způsobit vznik rakoviny dvěma způsoby: 1) ztrátou funkce u tumor supresorových mirnasvede voverexpresionkogenů, 2) ziskem funkce, kdy posiluje funkci onkogenů. SNPsmohou mít vliv na obojí a to přímo či nepřímo. Přímý efekt zahrnuje mutace, které ovlivňují processing mirna, a nepřímý efekt, kdy SNPs vregulační oblasti mirnaovlivní expresi mirna, nebo vcílové oblasti regulovaného genu, kde buď zanikne cílové místo, nebo vznikne nové. SNPsasociované sfungováním mirnatedy mohou posloužit jako biomarkery tendence rozvoje některých rakovin, předpovědět účinky léčby nebo učinit prognózu postupu nemoci. Podle mirbase(2013) bylo dosud identifikováno prekurzorovýcha zralých (matured) mirnav193 eukaryotních druzích a virech, ztoho u člověka bylo izolováno 1600 prekurzorovýcha 2042 zralých mirna. 50% všech mirnagenů bylo nalezeno vnestabilních oblastech genomu, které jsou často vrakovinných tkáních deletované, amplifikované nebo chybně exprimované.
40 Cca 1/3 mirnagenů člověka je organizována vklastrech (390 genů v113 klastrech), které jsou ko-exprimovány, takže chyba vregulaci jednoho člena ovlivňuje i ostatní geny vklastru. Změny vexpresi mirnamezi normální a rakovinnou tkání jsou často způsobeny tím, že jsou příslušné geny lokalizovány voblastech schromosomální nestabilitou (amplifikace, delece, translokace, blízká oblast častých zlomů). Některé mirnajsou přímo regulovány tumor supresory nebo onkogeny a účastní se jejich signálních drah. Např. TP53 reguluje mir-34a, RAS reguluje mir-21 a MYC reguluje mir klastr. Změny vregulaci mirnasjsou spojeny i sepigenetickými změnami, konkrétně metylace a tím změna regulace mirnagenů. Př: mir-127 při rakovině močového měchýře a mir-9-1 při rakovině prsu. Chromosomální přestavby mohou způsobit rakovinu i tím, že se onkogen dostane pod vliv promotoru mirna genu. Příkladem je mir-142, marker hematopoetických buněk, který se nachází na 17. chromosomu blízko zlomové bodu, který je často příčinou translokace t(8;17). Ta způsobuje agresivní leukemii B buněk kvůli upregulaci genu MYC, který se traslokací dostane pod vliv promotory mir-142.
41 Nejpřímočařejší změnou vedoucí ke změně transkripce mirna genu je mutace v promotorové oblasti, na kterou si sedá transkripční faktor.
42 SNPs v mirna prekurzorech mirna biogeneze zahrnuje štěpení RNázami III Drosha a Dicer. Pro toto štěpení je klíčové správné poskládání mirna do vlásenky, takže mutace ovlivňující párování a tím složení (folding) vlásenky, mohou inhibovat následné zpracování pri-a/nebo pre-mirna. Konkrétní případ tohoto jevu byl pozorován u dvou pacientů schronickou lymfatickou leukémií (CLL). CLL je často způsobena homozygotní delecí voblasti 13q13.4. Tato oblast kóduje polycystronický transkript, ze kterého vznikají mir-15a a mir-16-1, mirnas, které jsou dynamicky exprimované během buněčného cyklu a regulují geny stimulující posun vbuněčném cyklu a apoptózu, čili tyto mirna fungují jako tumor supresory. Bodová mutace vgenu mir-16-1 patrně způsobila chybu ve zpracování mir-16-1 a tím i vznik leukémie.
43
44 Jednou z příčin rakoviny mohou být mutace v genech účastnících se biogeneze mirna včetně RNA editinga terminální modifikace, které změní cíl mirna, zpracování a stabilitu, ale zatím se o tom moc neví. Inhibice biogeneze mirnavede kvážným defektům vývoje a je u mnoha organizmů letální. Porovnáním rakovinných tkání a jejich zdravých protějšků se zjistilo, že mirnasje vtumoru méně než ve zdravé tkáni. To ukazuje, že mirnasfungují spíše jako tumor supresory než jako onkogeny. 27% různých tumorů je hemyzygotníchpro deleci Dicerua globální knockdownmirnasvyřazením Diceruzvyšuje onkogenetický potenciál již transformovaných buněk a rychlost rozvoje tumoru. U myší bylo prokázáno, že k rozvoji tumoru přispělo i snížení množství Diceru vbuňce.
45 Změna ve fungování mirnamůže být založena i na změně cílové sekvence. Ktomu může dojít buď bodovou mutací, translokací nebo zkrácením 3 UTR. Translokace může oddělit mirnavazebná místa od regulovaných onkogenů. Bodové mutace vytvoří/zruší stávající rozpoznávací místa, zkrácením 3 UTRmohou zaniknout rozpoznávací místa mirna. Komplementarita sekvencí je klíčová pro vyhledání cílového místa na genu regulovaným mirna. Takže mutace vytvářející nové nebo ničící stávající vazebné místo významně ovlivňuje regulaci příslušného genu. Let-7 je tumor supresorová mirna, která reguluje onkogeny KRAS, MYCa HMGA2. Exprese let-7je u řady typů rakovin snížena a u rakoviny plic je navíc spojena se špatnou prognózou. Bodová mutace ve vazebném místě let-7v 3 UTRgenu KRAS vede koverexpresikras, což znamená vyšší riziko melanomu a některých rakovin plic, vaječníků a prsu.
46 44
47
Malé RNA a regulace genomu
 Malé RNA a regulace genomu Změna paradigmatu It appears that the genetic programming of humans and other complex organisms has been misunderstood for the past 50 years, due to the assumption that most
Malé RNA a regulace genomu Změna paradigmatu It appears that the genetic programming of humans and other complex organisms has been misunderstood for the past 50 years, due to the assumption that most
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNA molekuly. Analýza genové exprese pomocí cytometrických (a jiných) metod. Analýza exprese a funkce microrna. Úrovně regulace genové exprese
 metod. Analýza exprese a funkce microrna. Úrovně regulace genové exprese") Analýza genové exprese pomocí cytometrických (a jiných) metod Studium exprese a funkce microrna Eva Slabáková, Ph.D. Bi9393 Analytická cytometrie 12.11.2013 Oddělení cytokinetiky Biofyzikální ústav AVČR,
Analýza genové exprese pomocí cytometrických (a jiných) metod Studium exprese a funkce microrna Eva Slabáková, Ph.D. Bi9393 Analytická cytometrie 12.11.2013 Oddělení cytokinetiky Biofyzikální ústav AVČR,
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
RNA interference (RNAi)
") Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
ONKOGENETIKA. Spojuje: - lékařskou genetiku. - buněčnou biologii. - molekulární biologii. - cytogenetiku. - virologii
 ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Biomarkery - diagnostika a prognóza nádorových onemocnění
 Biomarkery - diagnostika a prognóza nádorových onemocnění O. Topolčan,M.Pesta, J.Kinkorova, R. Fuchsová Fakultní nemocnice a Lékařská fakulta Plzeň CZ.1.07/2.3.00/20.0040 a IVMZČR Témata přednášky Přepdpoklady
Biomarkery - diagnostika a prognóza nádorových onemocnění O. Topolčan,M.Pesta, J.Kinkorova, R. Fuchsová Fakultní nemocnice a Lékařská fakulta Plzeň CZ.1.07/2.3.00/20.0040 a IVMZČR Témata přednášky Přepdpoklady
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
The cell biology of rabies virus: using stealth to reach the brain
 The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
DMPK (ZNF9) V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M
 V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M") FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
Na rozdíl od genomiky se funkční genomika zaměřuje na dynamické procesy, jako je transkripce, translace, interakce protein - protein.
 FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1
 Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin.
 Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin. Nukleové base - purinové a pyrimidinové Ribonukleosidy - base + ribosa Deoxyribonukleosidy base + 2 - deoxyribosa Nukleotidy,
Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin. Nukleové base - purinové a pyrimidinové Ribonukleosidy - base + ribosa Deoxyribonukleosidy base + 2 - deoxyribosa Nukleotidy,
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Genetika zvířat - MENDELU
 Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
Syntéza a postranskripční úpravy RNA
 Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
EPIGENETICKÁ DĚDIČNOST
 Epigenetika EPIGENETICKÁ DĚDIČNOST Pojmenovaná britským biologem Conradem Waddingtonem r. 1940, Něco navíc, mimo v porovnání s mendelistickou genetikou, Umožňuje buňkám s identickým genotypem vznik odlišných
Epigenetika EPIGENETICKÁ DĚDIČNOST Pojmenovaná britským biologem Conradem Waddingtonem r. 1940, Něco navíc, mimo v porovnání s mendelistickou genetikou, Umožňuje buňkám s identickým genotypem vznik odlišných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
GENETIKA. Dědičnost a pohlaví
 GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Kontrola genové exprese
 Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
NGS analýza dat. kroužek, Alena Musilová
 NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Cytogenetika. chromosom jádro. telomera. centomera. telomera. buňka. histony. páry bazí. dvoušroubovice DNA
 Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Vztah genotyp fenotyp
 Evoluce fenotypu II Vztah genotyp fenotyp plán? počítačový program? knihovna? genotypová astrologie (Jablonka a Lamb) Modely RNA - různé vážení: A-U, G-C, G-U interakcí, penalizace za neodpovídající si
Evoluce fenotypu II Vztah genotyp fenotyp plán? počítačový program? knihovna? genotypová astrologie (Jablonka a Lamb) Modely RNA - různé vážení: A-U, G-C, G-U interakcí, penalizace za neodpovídající si
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
9. Evo-devo. Thomas Huxley ( )
") 9. Evo-devo Můžeme žasnout nad procesem, kterým se z vajíčka vyvine dospělý jedinec, ale bez problémů přijímáme tento proces jako každodenní fakt. Je to pouze nedostatek fantazie, který nám brání pochopit,
9. Evo-devo Můžeme žasnout nad procesem, kterým se z vajíčka vyvine dospělý jedinec, ale bez problémů přijímáme tento proces jako každodenní fakt. Je to pouze nedostatek fantazie, který nám brání pochopit,
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
Atestace z lékařské genetiky inovované otázky pro rok A) Molekulární genetika
 Molekulární genetika") Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
1.12.2009. Buněčné kultury. Kontinuální kultury
 Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace ztrácejí diferenciační
Primární kultury - odvozené přímo z excise tkáně buněčné linie z různých organizmů, tkání explantované kultury jednobuněčné suspense lze je udržovat jen po omezenou dobu během kultivace ztrácejí diferenciační
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Kosterní svalstvo tlustých a tenkých filament
 Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
Laboratoř molekulární patologie
 Laboratoř molekulární patologie Ústav patologie FN Brno Prof. RNDr. Jana Šmardová, CSc. 19.11.2014 Složení laboratoře stálí členové Prof. RNDr. Jana Šmardová, CSc. Mgr. Květa Lišková Mgr. Lenka Pitrová
Laboratoř molekulární patologie Ústav patologie FN Brno Prof. RNDr. Jana Šmardová, CSc. 19.11.2014 Složení laboratoře stálí členové Prof. RNDr. Jana Šmardová, CSc. Mgr. Květa Lišková Mgr. Lenka Pitrová
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Genová etiologie nemocí
 Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
M A T U R I T N Í T É M A T A
 M A T U R I T N Í T É M A T A BIOLOGIE ŠKOLNÍ ROK 2017 2018 1. BUŇKA Buňka základní strukturální a funkční jednotka. Chemické složení buňky. Srovnání prokaryotické a eukaryotické buňky. Funkční struktury
M A T U R I T N Í T É M A T A BIOLOGIE ŠKOLNÍ ROK 2017 2018 1. BUŇKA Buňka základní strukturální a funkční jednotka. Chemické složení buňky. Srovnání prokaryotické a eukaryotické buňky. Funkční struktury
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Investujeme do naší společné budoucnosti
 Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Indukovaná pluripotence. Petr Vodička Liběchov 16/11/2016
 Indukovaná pluripotence Petr Vodička Liběchov 16/11/2016 Totipotentní Pluripotentní Lidské ES Myší ES LIF + FBS Feeder = vrstva podpůrných buněk Myší embryonální fibroblasty, SNL, STO bfgf + SR Feeder
Indukovaná pluripotence Petr Vodička Liběchov 16/11/2016 Totipotentní Pluripotentní Lidské ES Myší ES LIF + FBS Feeder = vrstva podpůrných buněk Myší embryonální fibroblasty, SNL, STO bfgf + SR Feeder
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika. Genetika. Nauka o dědid. dičnosti a proměnlivosti. molekulárn. rní buněk organismů populací
 Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
VYUŽITÍ CYTOLOGICKÝCH A MOLEKULÁRNĚ BIOLOGICKÝCH METOD PŘI DETEKCI NÁDORŮ Definice problematiky Profil přístupů Nádorová heterogenita
 VYUŽITÍ CYTOLOGICKÝCH A MOLEKULÁRNĚ BIOLOGICKÝCH METOD PŘI DETEKCI NÁDORŮ Definice problematiky Profil přístupů Nádorová heterogenita Všechny nádory mají genetický podtext, ale některé mají tento podtext
VYUŽITÍ CYTOLOGICKÝCH A MOLEKULÁRNĚ BIOLOGICKÝCH METOD PŘI DETEKCI NÁDORŮ Definice problematiky Profil přístupů Nádorová heterogenita Všechny nádory mají genetický podtext, ale některé mají tento podtext
DUM č. 10 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 10 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 26.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Procesy následující bezprostředně po transkripci.
projekt GML Brno Docens DUM č. 10 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 26.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Procesy následující bezprostředně po transkripci.
Evoluční genetika 2/1 Zk/Z
 Evoluční genetika 2/1 Zk/Z Radka Reifová, Pavel Munclinger, Zuzana Musilová Prezentace a materiály k přednášce http://web.natur.cuni.cz/zoologie/biodiversity/ Evoluční genetika Obor vzniklý propojením
Evoluční genetika 2/1 Zk/Z Radka Reifová, Pavel Munclinger, Zuzana Musilová Prezentace a materiály k přednášce http://web.natur.cuni.cz/zoologie/biodiversity/ Evoluční genetika Obor vzniklý propojením
Biologie - Oktáva, 4. ročník (přírodovědná větev)
") - Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
- Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
b) Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?
 Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?") 1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
Systém HLA a prezentace antigenu. Ústav imunologie UK 2.LF a FN Motol
 Systém HLA a prezentace antigenu Ústav imunologie UK 2.LF a FN Motol Struktura a funkce HLA historie struktura HLA genů a molekul funkce HLA molekul nomenklatura HLA systému HLA asociace s nemocemi prezentace
Systém HLA a prezentace antigenu Ústav imunologie UK 2.LF a FN Motol Struktura a funkce HLA historie struktura HLA genů a molekul funkce HLA molekul nomenklatura HLA systému HLA asociace s nemocemi prezentace
GENETIKA. zkoumá dědičnost a proměnlivost organismů
 GENETIKA zkoumá dědičnost a proměnlivost organismů Dědičnost: schopnost organismů uchovávat informace o své struktuře a funkčních schopnostech a předávat je svým potomkům Proměnlivost (variabilita) je
GENETIKA zkoumá dědičnost a proměnlivost organismů Dědičnost: schopnost organismů uchovávat informace o své struktuře a funkčních schopnostech a předávat je svým potomkům Proměnlivost (variabilita) je
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Vakcíny z nádorových buněk
 Protinádorové terapeutické vakcíny Vakcíny z nádorových buněk V. Vonka, ÚHKT, Praha Výhody vakcín z nádorových buněk 1.Nabízejí imunitnímu systému pacienta celé spektrum nádorových antigenů. 2. Jejich
Protinádorové terapeutické vakcíny Vakcíny z nádorových buněk V. Vonka, ÚHKT, Praha Výhody vakcín z nádorových buněk 1.Nabízejí imunitnímu systému pacienta celé spektrum nádorových antigenů. 2. Jejich
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební