MENDELOVA UNIVERZITA V BRNĚ Lesnická a dřevařská fakulta. Ústav lesnické botaniky, dendrologie a geobiocenologie BAKALÁŘSKÁ PRÁCE
|
|
|
- Andrea Šimková
- před 6 lety
- Počet zobrazení:
Transkript
1 MENDELOVA UNIVERZITA V BRNĚ Lesnická a dřevařská fakulta Ústav lesnické botaniky, dendrologie a geobiocenologie BAKALÁŘSKÁ PRÁCE Brno 2013 Vršanová Marie
2 MENDELOVA UNIVERZITA V BRNĚ Lesnická a dřevařská fakulta Ústav lesnické botaniky, dendrologie a geobiocenologie Vodní stres jako příčina změn histologické struktury jehlic smrku ztepilého BAKALÁŘSKÁ PRÁCE Brno 2013 Vršanová Marie
3
4 PROHLÁŠENÍ Prohlašuji, že jsem bakalářskou práci na téma: Vodní stres jako příčina změn histologické struktury jehlic smrku ztepilého zpracovala sama a uvedla jsem všechny použité prameny. Souhlasím, aby moje bakalářská práce byla zveřejněna v souladu s 47b Zákona č. 111/1998 Sb., o vysokých školách a uložena v knihovně Mendelovy univerzity v Brně, zpřístupněna ke studijním účelům ve shodě s Vyhláškou rektora MENDELU o archivaci elektronické podoby závěrečných prací. Autor kvalifikační práce se dále zavazuje, že před sepsáním licenční smlouvy o využití autorských práv díla s jinou osobou (subjektem) si vyžádá písemné stanovisko univerzity o tom, že předmětná licenční smlouva není v rozporu s oprávněnými zájmy univerzity a zavazuje se uhradit případný příspěvek na úhradu nákladů spojených se vznikem díla dle řádné kalkulace. V Brně dne:.... Vršanová Marie
5 PODĚKOVÁNÍ Na prvním místě bych ráda poděkovala především vedoucímu bakalářské práce panu Ing. Romanu Gebauerovi Ph.D. za umožnění vypracování bakalářské práce pod jeho vedením. Za čas strávený při odborných konzultacích, za rady, ochotu a trpělivost. Také bych chtěla poděkovat panu Ing. Radku Šmudlovi Ph.D za pomoc se statistickým zpracováním dat a RNDr. Mileně Martinkové CSc. za cenné informace, které mi poskytla. Poděkování také patří mé rodině a přátelům, kteří mě ve studiu podporovali. Děkuji.
6 ABSTRAKT Jméno posluchače: Marie Vršanová Název bakalářské práce: Vodní stres jako příčina změn histologické struktury jehlic smrku ztepilého Stres suchem ovlivňuje jak lesní porosty, tak i stromy ve městech. Cílem této bakalářské práce bylo zjistit reakci histologických strukturu jehlic smrku ztepilého na uměle vyvolaný stres suchem a na základě literárních zdrojů zdokumentovat a vybrat vhodná opatření pro zabránění stresu suchem ve městě. Vzorky pro tuto práci byly odebrány ze tří stromů z plochy stresované suchem a tří stromů kontrolních. Odebírali se jehlice z letošního přírůstu větví z báze i z vrchu koruny stromu. U provedeného příčného řezu každé jehlice se zjišťovala výška, šířka, plocha centrálního válce a plocha jehlice. Statistické analýzy prokázaly u všech znaků jehlic z vrchu koruny rozdíl mezi stresovanou a kontrolní plochou. U jehlic z báze koruny se potvrdil rozdíl jen u znaku plocha jehlice. Dále se potvrdil rozdíl mezi jehlicemi z vrchu a z báze koruny. Vodní stres tedy u smrku ztepilého vyvolává reakce, které vedou ke snížení růstu jehlic, čímž se rostlina snaží na nedostatek vody adaptovat. Pro stromy ve městech je stres suchem jedním z limitujících faktorů růstu. Nejdůležitějším opatřením je výběr vhodného taxonu pro dané stanoviště, úprava půdního prostředí, nebo závlaha. Klíčová slova: vodní stres, histologie, městské prostředí, závlaha
7 ABSTRACT Water stress as the cause of histological structures changes of Norway spruce needles Water stress influences forests and also urban trees. The aim of this thesis was to determine the reaction of Norway spruce histological structures to artificially induced water stress and to find out and evaluate suitable measures to water stress avoidance in urban areas based on literary sources. The samples were acquired from three trees from drought stressed plot and three trees from control plot. The needles were acquired from shoots from basal and top of the crown. From the needles cross-section was found out (height, width, and needle area and needle cylinder). Statistical analysis confirmed the difference between control and drought-stressed area for needles from the top of the crown. For the needles from the base of the crown was the difference confirmed only for needle area. Also the difference between needles from base and from top was confirmed. Water stress causes reaction in Norway spruce needles, which lead to needle growth reduction as an adaptation to water deficit. For trees in the urban environment drought stress is one of the factors limiting their growth. The most important measure is the selection of a suitable taxon for concrete site, or the soil improvement or irrigation. Key words: water stress, histological structures, urban areas, irrigation
8 Obsah 1. Úvod Cíl práce Literární přehled Význam vody v rostlině Příjem a transport vody Vodní potenciál Transport vody v kontinuu půda-rostlina-atmosféra Kohezní teorie Stres Vodní stres Vliv vodního stresu na anatomii a fyziologii Působení vodního stresu během vegetace Specifika městského prostředí a rizika vodního stresu Materiál a metody Výsledky Diskuse Závěr Summary Seznam použité literatury... 37
9 1. Úvod V důsledku současných klimatických změn je v dnešní době nedostatek vody velmi diskutovaným tématem a lze předpokládat, že jeho vliv na rostliny bude čím významnější i v oblastech, kde bylo doposud vody dostatek. Stres suchem ovlivňuje nejen lesní ekosystémy, ale i porosty a solitérní stromy v urbanizovaném prostředí. Tato bakalářská práce se bude zabývat možnými vlivy vodního stresu na růst stromu a na reakce, které uvnitř strom vyvolává. V praktické části půjde o měření rozdílů ve velikosti histologických struktur jehlic smrku ztepilého mezi suchem stresovanými a kontrolními stromy. V teoretické části se z literárních zdrojů získají obecné informace o vodě v rostlině, vodním stresu a návrzích vhodných opatření, kterými lze zamezit stresu suchem v městském prostředí. 10
10 2. Cíl práce Stres suchem je v dnešní době velkým problémem nejen v oblasti zemědělství či lesního hospodářství, ale je i jedním z nejvýznamnějších limitujících faktorů existence stromů v městském prostředí. Cílem této bakalářské práce je zjistit reakci histologických strukturu jehlic smrku ztepilého na uměle vyvolaný stres suchem a na základě literárních zdrojů zdokumentovat a vybrat vhodná opatření pro zabránění stresu suchem v městském prostředí. Půjde o zjištění, které struktury jsou zapojeny do adaptací pro přežití rostliny v podmínkách dlouhotrvajícího stresu suchem. Hypotéza je, že se stres suchem projeví snížením růstu jehlic. To je spojeno se sníženou plochou pro fotosyntézu (pokles asimilátů) a tím i s poklesem odolnosti vůči patogenům. 11
11 3. Literární přehled 3.1. Význam vody v rostlině Voda je základní a nezbytnou složkou všech organismů na Zemi, jelikož všechny buněčné funkce jsou adaptovány na fyzikální a chemické vlastnosti vody. Voda se na stavbě rostlinného těla podílí nejvíce ze všech látek, například listy čerstvé zeleniny obsahují % vody a dužnaté plody až %. U rostlin i krátkodobý pokles množství vody v listech pod cca 60 % vede k nevratnému poškození pletiv a k smrti orgánu (Procházka a kol., 2003). Stromy jako ostatní vyšší rostliny jsou rostliny homoihydrické a musí oproti poikilohydrickým (mechy, lišejníky) rostlinám udržet ve svém těle vysoký obsah vody, aby mohly přežít. Z těchto důvodů obsahují homoihydrické rostliny vakuoly, které dokážou zadržet velké množství vody a do určité míry omezují vliv vnějšího prostředí na protoplazmu. Buňky těchto rostlin, ale nejsou schopny snášet dlouhodobé vysušení. Proti náhlým změnám vlhkostních poměrů vnějšího prostředí, které by mohl zapříčinit vysušení, slouží homoihydrickým rostlinám jednak ochranná vrstva kutikuly, která snižuje výpar a také průduchy, které regulující transpiraci (Larcher a kol., 1988). Voda v rostlině funguje jako rozpouštědlo, umožňuje příjem, transport a metabolismus látek. Díky termoregulační schopnosti a schopnosti výparu při různé teplotě stabilizuje teplotu rostliny a chrání rostlinu proti přehřátí. Většina denní spotřeby vody rostlin je použita na transpiraci a k vlastním biochemickým procesům slouží pouze malá část. Z 1000 g přijaté vody 990 g prochází rostlinou jako tranzitní voda, která dosycuje rostlinná pletiva a nahrazuje vodu vydanou při transpiraci. Dalších 8-9 g tvoří konstituční vodu v koloidech a pouze 1-2 g se využije na tvorbu sušiny (Procházka a kol., 2009). 12
12 3.2. Příjem a transport vody V průběhu své evoluce museli rostliny vytvořit specializovaná pletiva, kterými by mohly rychle transportovat vodu na dlouhé vzdálenosti. Proto u těchto rostlin vznikly vodivé cévní svazky složené z lýkové (floémové) a dřevní (xylémové) části. U nahosemenných rostlin jsou buňkami odpovědnými za transport vody z kořenů na místa spotřeby cévice a u krytosemenných rostlin cévice a především cévy (Procházka a kol., 2009). Suchozemské rostliny jsou vystaveny neustálému výparu vody z listů a je pro ně nutné zajistit její pravidelný příjem. Největší podíl na příjmu vody pro rostlinu má kořenový systém. Nejaktivněji přijímají vodu mladé kořeny, v části kde ještě nedošlo k vytvoření sekundární kůry. Důležité je, že v této částí kořene jsou již plně vyvinuty vodivé elementy xylému. Přesto významný příjem vody je i díky starším kořenům, u kterých sice sorpční aktivita klesá, ale oproti jemným kořenům mají rozsáhlý povrch (Procházka a kol., 2009) Vodní potenciál O pohybu vody v rostlině rozhoduje vodní potenciál (Ѱ). Vodní potenciál se rovná rozdílu volné energie jednotky objemu matričné vázané (nebo tlak vyrovnávající nebo osmoticky poutané) vody a jednotky objemu čisté vody. Vodní potenciál vyjadřuje výsledek vzájemného působení osmotického, matričního, tlakového a gravitačního potenciálu a udává se v jednotkách tlaku, tj. v pascalech (Pa). Na výsledné hodnotě vodního potenciálu se nejvíce podílí tlakový a osmotický potenciál. Tlakový potenciál může být kladný, nulový a záporný. Ve vodivých elementech je při plné transpiraci záporný a transport vody pak probíhá v podtlaku. Osmotický potenciál je vždy negativní, neboť rozpuštěné látky ve vodě potenciál vody snižují. Vodní potenciál se zpravidla měří Scholanderovou tlakovou komorou. U odebraného vzorku (letorost, list, dlouhá jehlice) se postupně zvyšuje tlak až do doby než začne být vytlačována voda ven z xylému. Tlak v komoře nutný k vytlačení vody z xylému se rovná záporné hodnotě vodního potenciálu ve vzorku (Tyree a Zimmermann, 2002). Pro měření vodního potenciálu jsou nejdůležitější dvě denní doby: ráno před východem slunce, kdy je vodní potenciál nejvyšší a zpravidla je roven vodnímu potenciálu v půdě. Druhé měření se provádí uprostřed dne, v období největších evaporačních požadavků, kdy je vodní potenciál nejnižší. Tato hodnota 13
13 udává maximální míru stresu rostliny v průběhu dne (Gebauer a kol., 2010). Hodnota vodního potenciálu nás tak informuje o tom, zda a v jaké míře rostlina trpí suchem (Procházka a kol., 2009). Pro rostlinu s dostatečnou zásobou vody jsou charakteristické hodnoty vodního potenciálu listů před východem slunce přibližně do - 0,5 MPa. Poměrně silný vodní stres je kolem hodnoty - 2MPa. Procházka a kol. (2003) uvádějí, že o silný stres se jedná už při hodnotách pod - 1,5MPa, kdy klesá turgorový tlak v buňkách listů na nulu, a listy začínají vadnout. Při dlouhotrvajícím vodním potenciálu nižším než - 2,5 MPa dochází k odumírání rostlinu. Tato čísla jsou však pouze orientační, protože záleží na konkrétním rostlinném druhu, ekotypu, výšce odběru vzorku, řádu odebrané větve apod. (Gebauer a kol., 2010) Transport vody v kontinuu půda-rostlina-atmosféra Pohyb vody v rostlině funguje na principu rozdílných potenciálů. Voda proudí z místa o vyšším potenciálu do místa s nižším potenciálem (Larcher a kol., 1988). Jestliže vložíme buňku do vody s vyšším vodním potenciálem, pak v důsledku rozdílných vodních potenciálů začíná buňka pasivně přijímat vodu do vakuoly, její objem se zvyšuje a na buněčnou stěnu začíná působit tlakový potenciál. Jakmile se vodní potenciály vyrovnají, ustává pohyb vody do buňky a vytváří se tzv. dynamická rovnováha, kdy se množství vody v buňce nemění, přestože molekuly vody stále procházejí membránou v obou směrech (Procházka a kol., 2009). V rámci rostliny se jedná se o souvislou cestu vody s půdy do kořenů, listů a atmosféry. V půdě je totiž vodní potenciál nejvyšší, neboť koncentrace solí v půdním roztoku je velmi malá a voda putuje do místa s nejnižším vodním potenciálem, což je atmosféra (Procházka a kol., 2009). Výdej vodní páry z povrchu orgánů rostliny (převážně listů) do atmosféry se nazývá transpirace. Listy jsou jen zřídka plně nasyceny vodou a mají proto nízký vodní potenciál. Transpirace se projevuje jako sání, které uvede vodu do pohybu směrem vzhůru do těla rostliny. Působí také na nasávání vody kořeny a má mnohem větší význam než kořenový vztlak. Transpirace je regulovaná průduchy, které dokážou na základě několika signálů měnit velikost dýchací skuliny (Procházka a kol., 2009). 14
14 Kohezní teorie Kohezní teorie uvádí, že hybnou silou transportu vody je podtlak ve vodivých elementech xylému, který je způsobený vypařováním vody z listů. Mechanismus, který umožňuje pohyb vody proti gravitaci vzhůru, souvisí se stavbou buněčných stěn listového mezofylu. Póry mezi mikrofibrilami celulózy jsou úzké a voda v nich má ostrý úhel smáčení a díky vysokému povrchovému napětí vody se vytváří negativní tlak. Velikost tohoto negativního tlaku je závislá na průměru vodivých elementů. Čím jsou kapiláry tenčí, tím větší podtlak v nich vzniká a voda může být transportována do velkých výšek, současně je v nich větší soudržnost vodního sloupce než v širších trubicích. Na druhou stranu v úzkých kapilárách vzrůstá také odpor kladený toku vody. Průřez vodivých elementů rostlin tedy musí být kompromisem, vzniklým v průběhu evoluce, při kterém je zaručena dostatečná vodivost současně s provozní bezpečností vodivých drah. Přesto může dojít k přerušení kontinuálních vodních sloupců vlivem kavitací (dutin) vlivem extrémní transpirace při zvyšování teploty nebo při mrznutí a následném tání. Určitou ochranou proti vzniku kavitací jsou perforované přepážky vodivých elementů, které zabraňují šíření bublinek vzduchu na větší vzdálenosti. Rostlina se také snaží eliminovat možné negativní důsledky vzniku kavitací tím, že jednotlivé cévy a cévice nejsou uloženy paralelně vedle sebe, ale různě se ve kmeni proplétají a voda vcházející z jedné části kořene je rozváděna do různých částí koruny. Kapacita vodních cest je však předimenzována a vyřazení i více vodivých elementů z provozu nemusí výrazně ohrozit život rostliny (Procházka a kol., 2009) Stres Obecně řečeno každý faktor zhoršující funkce organismu lze označit jako stres. Každý rostlinný druh toleruje určité rozpětí jednotlivých faktorů a interval mezi minimální a maximální tolerovanou hodnotou se nazývá ekologická valence faktoru. Optimum je taková hodnota faktoru, při které se organizmus nejlépe vyvíjí. Jakékoli přírodní podmínky, které se vymykají optimu daného druhu, do jisté míry zhoršují jeho životní podmínky. Přesněji se termín stres používá, pokud se jedná o skutečné poškození či dokonce odumření rostliny nebo pokud vyvolal v rostlině specifickou odpověď (Bláha a kol., 2003). Rostliny jsou ve vnějším prostředí vystaveny velkému počtu stresujících faktorů sucho, mráz, zasolení apod., a jelikož jsou rostliny trvale umístěny na jednom místě 15
15 bez možnosti pohybu, nemůžou těmto stresujícím vlivům uniknout. Rostliny tedy museli ve svém vývoji zaujmout jinou strategii, jak se stresem vyrovnat a to systémem různých reakcí. U rostlin se může vyvinout resistence vůči stresu (např. schopnost přežít nízký vodní potenciál), kterou lze dosáhnout buď, vyhnutím se stresu nebo tolerancí ke stresu a případně jejich kombinací. (Öpik a Rolfe, 2005). Vyhýbání se stresu je schopnost předcházení (zabránění) stresu vytvářením ekvivalentních vnitřních podmínek v rostlině. Například některé rostliny dokážou v suchých podmínkách otevřením průduchů pouze v průběhu noci snížit míru ztráty vody na velmi nízkou úroveň. Tolerance ke stresu je schopnost přežít vnitřní stresy, např. nízký obsah vody v buňce. U rostlin mírného pásma se ukazuje, že tolerance k vodnímu stresu se v průběhu roku pravidelně mění a je největší v zimních měsících. V zimě jsou rostliny vystaveny vodnímu deficitu ve zmrzlé půdě, která sice má dostatek vody, ale ve formě nepřístupné pro rostliny (Öpik a Rolfe, 2005). Schopnost vyrovnat se s environmentálními stresy se stala limitujícím faktorem pro růst, přežití a geografické rozšíření rostlin. Některé rostliny jsou pozoruhodně odolné vůči jednotlivým druhům stresu jako např. arktická či pouštní vegetace. Na druhou stranu některé druhy odumírají již při teplotě 10 C (Öpik a Rolfe, 2005) Vodní stres Vodní stres je definován jako vystavení rostliny nízkému vnějšímu vodnímu potenciálu. Tento problém nepřímo ovlivňuje i člověka, protože sucho způsobuje největší škody v zemědělství a vysoce převyšuje všechny další vlivy (Öpik a Rolfe, 2005). Důvodem, proč vodní stres tak významně negativně ovlivňuje růst a produktivitu rostlin, je to, že voda je v přírodě v neustálém koloběhu a na rozdíl od minerálních látek zůstává v půdě nebo v rostlinách krátkou dobu (Procházka a kol., 2003). Další příčinou nedostatku vody dostupné pro rostliny jsou nejčastěji klimatické poměry a průběh počasí. Vlastní příjem vody rostlinou je závislý také na obsahu živin a solí v půdě, ale i na půdní reakci (Bláha a kol., 2003). Většina vyšších rostlin prodělá ve svém vývoji období velmi nízkého obsahu vody v buňkách a to ve fázi semena (5 20 %). V průběhu klíčení se odolnost vůči suchu postupně ztrácí a po zbytek rostlinného života se nikdy nedostane na podobnou míru resistence. Skoro všechny rostliny jsou v průběhu svého života vystaveny nějakému stupni vodního stresu v důsledku změn oslunění v průběhu dne. Získávání 16
16 CO 2 pro fotosyntézu skrz stomata je nevyhnutelně spojeno se ztrátou vody do atmosféry a určitou ztrátou turgoru (Fitter a Hay, 2002). Vypořádání se s tímto stresem je běžnou součástí života rostlin a to i druhů, které žijí mimo aridní oblasti. Míra vodnímu stresu, kterému je rostlina vystavena se nejspolehlivěji určuje z charakteristik vycházejících ze stavu vody v rostlině: vodní potenciál buněk či vodní sytostní deficit. Tyto údaje jsou spolehlivější než údaje o vodě v prostředí jako je vlhkost půdy a vzduchu nebo vodní potenciál půdy (Procházka a kol., 2003). Také Fitter a Hay (2002) uvádějí, že neexistuje údaj charakterizující prostředí rostliny, který by mohl být použit ke kvantifikaci vodního stresu, kterému je rostlina vystavena. Vodní stres se tedy vyjadřuje jako stav, kdy obsah vody v buňce klesne pod hodnotu turgoru (pod hodnotu optima pro růst rostlin) Vliv vodního stresu na anatomii a fyziologii Vodní stres vyvolává v rostlině soustavu reakcí a jeho vliv na funkčnost rostliny závisí na míře nedostatku vody. Jelikož je voda součástí buněčné stavby a procesů, je těžké rozlišovat mezi primárním poškozením přímo vyplývajícím z vodního deficitu a sekundárními vlivy, které jsou důsledkem primárního poškození (Öpik a Rolfe, 2005). Vliv vodního deficitu je závislý na čase - poškození se s časem zvětšuje. Nejprve rostlina reaguje na vodní stres snížením růstu a později dochází ke změnám v metabolismu (Öpik a Rolfe, 2005). Reakce rostliny na vodní stres se děje na úrovni molekulární, buněk, pletiv, jednotlivých orgánů až celé rostliny (Fitter a Hay, 2002). Uzavírání průduchů Nedostatek vody v listech vede k poklesu turgoru ve svěracích buňkách průduchů, který má za následek jejich uzavření a tím se rostlina snaží zabránit dalšímu výparu. K uzavírání průduchů dochází při vodním potenciálu - 0,5 až - 1,0 MPa a vede ke změnám dalších fyziologických procesů jako je fotosyntéza a respirace (Procházka a kol., 2009). To znamená, že fotosyntéza je redukována, ještě než vodní stres výrazně ovlivní další buňky a orgány v rostlině (Fitter a Hay, 2002). 17
17 Vadnutí rostlin ztráta turgoru Turgor u rostlin má hlavní úlohu při růstu a prodlužování buněk, otevírání průduchů, pohybu listů a květních obalů. Pokles turgoru vede ke ztrátě pevnosti a pružnosti, nejdříve ztrácejí turgor listy. Vadnutí může být dočasné, to je při velmi vysoké teplotě a nízké relativní vlhkosti vzduchu, kdy ztráty vody transpirací jsou nahrazeny příjmem vody v noci. K úplnému vadnutí dochází, pokud půda neobsahuje přístupnou vodu a vodní deficit vzniklý přes den se v průběhu noci nenahradí. Postupně začne docházet k snižování obsahu vody ve všech orgánech. Začnou odumírat kořenové vlásky a tím se poruší kontakt mezi půdou a kořenem a rostlina ztratí schopnost přijímat vodu, i při opětovné závlaze (Procházka a kol., 2009). Zastavení růstu Extrémně náchylný na vodní stres je růst, který může být omezen v momentě, jakmile obsah vody v buňkách klesne pod hranici plného nasycení, tedy dříve než by se dal očekávat dopad na metabolické aktivity. Toto je pravděpodobně přímý efekt poklesu turgoru, protože expanze buněk závisí na uvolňování buněčné stěny pod tlakem turgoru (Öpik a Rolfe, 2005). Růst začíná zvětšováním objemu buňky absorpcí vody do vakuol a zvětšováním plochy povrchu buněčné stěny. Nedostatkem vody je dlouživý růst inhibován ve fázi, kdy na primární buněčné stěně probíhá proces vkládání nových stavebních látek mezi staré, tzv. plošný růst buněčných stěn. K měření zpomalení růstu dochází již, když turgor klesne na 0,1 až 0,2 MPa. K zastavení růstu dojde dříve než ke zjevnému vadnutí. Proto se v rostlině můžou dočasně hromadit nevyužité asimiláty (Bláha a kol., 2003). Metabolické změny, změny aktivity enzymů Při dalším poklesu vodního potenciálu buněk zhruba na hodnotu - 0,2 až - 0,8 MPa dochází k rychlým změnám aktivity enzymů, snižuje se aktivita enzymů nitrátreduktázy, naopak stoupá činnost alfa amylázy, ribonukleázy, hydrolázy. Snižuje se syntéza proteinů a cytokyninů a zpomaluje se buněčné dělení. Při větším poklesu vodního potenciálu k hodnotám okolo - 1,0 MPa dochází u mnoha druhů k tvorbě aminokyselin, prolinů, cukrů, alkoholů a další sloučenin (Bláha a kol., 2003). Hlavní význam těchto syntéz je zřejmě zvýšení osmotického tlaku v buňkách (Öpik a Rolfe, 2005). 18
18 Rostliny si vytvořily systém, který jim umožňuje na vodní stres reagovat dříve, než dojde k větším ztrátám vody. Jde o reakci zprostředkovanou kyselinou abscisovou (ABA), která se uvolňuje z mezofylových buněk již při velmi mírném vodním stresu a je vedena ke svěracím buňkám průduchů (Procházka a kol., 2009). Při nedostatku vody patří ABA také k významným mediátorům exprese genu pro stresové proteiny. Regulací genové aktivity se zvyšuje nebo snižuje tvorba enzymů i strukturálních proteinů, nepřímo pak i ostatních metabolitů (Bláha a kol., 2003). Fotosyntéza Při pokračujícím nedostatku vody se začínají projevovat další metabolické změny, a to především u fotosyntézy a transportních pochodů v buňce. Fotosyntéza může být limitována dvěma způsoby, a to stomatální a nestomatální inhibicí souběžně. O ovlivňování činnosti fotosyntetických enzymů vodním stresem existují v literatuře rozpory. Je řadu údajů o relativní odolnosti fotosyntetických enzymů ke škodlivým účinkům vodního stresu, ale i literární prameny, které naznačují přímý vliv sucha na fotosyntetické enzymy. Při postupném vysychání se snižuje hydratace protoplazmy a tím i fotosyntetická kapacita. Příjem CO2 dosahuje normálních rychlostí jen v úzkém rozsahu dostatečného zásobování vodou. Mimo tento rozsah příjem CO2 začíná klesat a nakonec se zcela zastaví. Křivka závislosti výměny plynů na ztrátě vody ukazuje dva důležité kritické body: body přechodu z plného výkonu do oblasti omezení a bod, ve kterém je výměna plynů nulová. První kritický bod odpovídá takovému nedostatku vody, při kterém se průduchy začínají zavírat, difúzní odpor průduchů tak začne převyšovat reziduální odpor. Pokud po dosažení prvního kritického bodu rostlina opět získá vodu, nastává rychlá obnova výměny CO2. Druhý kritický bod odpovídá stavu, kdy jsou průduchy úplně uzavřeny, a projevuje se i přímý účinek nedostatku vody na protoplazmu. Příjem CO2 z vnějšku už neprobíhá, ale CO2 uvolněný při dýchání může být znovu vázán. Po dosažení tohoto stavu už obnovení přísunu vody nevede k okamžité obnově fotosyntézy. Obnova se opožďuje a za určitých podmínek se už nemůže po silné vyschnutí původní fotosyntetická kapacita obnovit (Bláha a kol., 2003). Navíc dlouhodobé snížení fotosyntézy vede k nedostatku asimilátů a tím k dalšímu omezení růstu. Přebytku absorbované sluneční energie vede k vzniku reaktivních forem kyslíku, k poškození membrán a tím k dalšímu zhoršení potenciálu pro fotosyntézu (Öpik a Rolfe, 2005). 19
19 Respirace Respirace je vodním stresem ovlivňována méně než fotosyntéza, avšak byl zaznamenán vliv na omezení tvorby ATP. Zatímco proces fotosyntézy je utlumen, respirace v rostlině neustává a to urychluje prohlubování nedostatku živin (Öpik a Rolfe, 2005). Transpirace Průběh transpirace během stresu suchem má charakteristické chování. Na počátku stresu se transpirace snižuje v poledních hodinách a později se opět obnovuje. Při pokračujícím nedostatku vody se přestane objevovat opětovné odpolední zvýšení transpirace a ještě později se průduchy otevírají jen ráno, při relativní ochlazení. Nakonec, ale ještě v době, kdy mají rostliny dostatečný obsah vody, zcela ustává stomatální transpirace a rostliny transpirují pouze skrz kutikulou (Bláha a kol., 2003) Působení vodního stresu během vegetace Rozhodující pro rostlinu mírného pásma je, zda sucho nastalo v průběhu vegetace nebo zda rostlina roste v relativním suchu od počátku vegetace. Sucho, které začne působit v průběhu vegetace, má totiž podstatnější vliv na metabolismus rostliny. Pokud rostlina od počátku roste v suchu, má možnost se na stres adaptovat například hlouběji pronikajícím kořenovým systémem, silnější kutikulou, menším počtem průduchů anebo menší listovou plochu. V případě dlouhodobého nedostatku vody od začátku vegetace dochází k inhibici tvorby kořenového systému. Zpočátku dochází k jeho prodlužování do větších hloubek, ale na úkor tvorby postranních kořenů a kořenového vlášení. Jestliže vodní stres i nadále pokračuje, dochází k redukci kořenového systému, přestává se vytvářet kořenové vlášení a nakonec se zcela zastaví růst kořenů a kořen uhyne. Co se týče nedostatku vody u nadzemních orgánů, pokud k němu dojde na začátku růstového cyklu, bude listová plocha redukována, stejně tak i uhlíkový přírůstek po celou dobu růstového období. Pokud se snižuje turgor během vývoje květenství, redukuje se vývoj květů a tím je ohrožena celá reprodukce. Pokud dojde k nedostatku vody až během dozrávání plodů, dosavadní rostlinný vývoj nebude ovlivněn, pouze se sníží hmotnost semen a může se zvýšit opad plodů. U dřevin často 20
20 mají velký význam vlhkostní poměry i v předchozím roce a rostliny často reagují se zpožděním (Bláha a kol., 2003) Specifika městského prostředí a rizika vodního stresu Stromy jako všechny organismy jsou součástí prostředí, ve kterém se vyskytují. Své prostředí ovlivňují a jsou jím také ovlivňovány. Jsou adaptovány na růst v porostu i ve volné krajině a na vlivy, které je tam mohou potkat, byly v průběhu evoluce připravovány. Avšak z hlediska délky existence rostlin stromového růstu na této planetě, se před poměrně krátkou dobou staly součástí prostředí pro ně velmi nehostinného a nepřirozeného a to prostředí městského. Pro člověka je městská zeleň velmi důležitá a z velké části ji vnímáme podvědomě. Vegetační prvky ve městě mají blahodárný vliv nejen na fyzické, ale i duševní zdraví obyvatel. Stromy díky své velikosti a dlouhověkosti jsou jednou z nejdůležitějších přírodních složek, které můžeme v městském prostředí použít, a svou velikosti ovlivňují větší část prostředí. Ve městě je však nedostatek prostoru a možnosti výsadby stromu jsou omezené. Městské prostředí je silně extrémní stanoviště s několika významnými stresovými vlivy (Kolařík a kol., 2003). Mezi hlavní stresové faktory městského prostředí patří: Dostupnost vody v půdním prostoru Provzdušněnost půdy Skladba půd a jejich ph Kontaminace půdy Klimatické poměry Znečištění vzduchu Intenzita některých faktorů významně kolísá v čase. V takovém případě má zásadní význam i časový průběh vlivu v kombinaci s fenologickými fázemi vývoje dřeviny. Extrémní podmínky ve městě je proto třeba zohlednit u pěstebních opatření. Například nedostatek vody patří k nejdůležitějším faktorům, které se podílejí na špatném stavu dřevin ve městech, zejména v prvních týdnech a měsících po výsadbě. (Kolařík a kol., 2003) 21
21 4. Materiál a metody Lokalita a odběr vzorků V Norsku byly vybrány dvě plochy vysázené 20tiletým smrkem ztepilým (Picea abies L. Karsten.). Jedna plocha byla kontrolních a druhá byla stresovaná suchem. Sucho bylo navozeno stříškou, která byla umístěna pod korunou stromů. Od začátku experimentu do termínu odběru uplynuly dva roky. Na každé ploše byly vybrány tři stromy. V srpnu 2010 byly z těchto stromů Ing. Josefem Urbanem odebrány vzorky větví z vrcholu a báze koruny. Z každého stromu a pozice bylo odebráno pět větví. Celkem bylo tedy pro každou plochu odebráno 15 větví z vrcholu a 15 z báze koruny. Ty byly fixovány v roztoku FAA (90 ml 70% etanolu, 5 ml kyseliny octové a 5 ml 40% formaldehydu). Vzorky byly poté převezeny na Mendelovu univerzitu v Brně k dalšímu zpracování. Vyhotovení řezů a mikrosnímků Z fixovaných vzorků bylo z letošního přírůstu odebráno sedm jehlic. Na bázi každé jehlice byly provedeny ruční příčné řezy. Tyto řezy byly obarveny na přítomnost ligninu pomocí HCl a fluoroglucinolu (Němec et al. 1962). Na zhotovené řezy byl přikápnut glycerol, aby nedošlo k vyschnutí vzorků. Poté byly mikroskopické preparáty překryty krycím sklíčkem a sledovány pod mikroskopem Olympus BX51 (zvětšení 200x a 400x) propojeným s digitálním fotoaparátem Olympus E-330 v laboratoři na Ústavu lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU v Brně. Vyhodnocení řezů Nasnímané preparáty byly analyzovány v počítačovém programu ImageTool3.00. U každého vzorku se zjišťovala výška a šířka jehlice, plocha centrálního válce a plocha jehlice na příčném řezu (Obr. 1). Plocha centrálního válce byla měřena i s endodermisem. 22






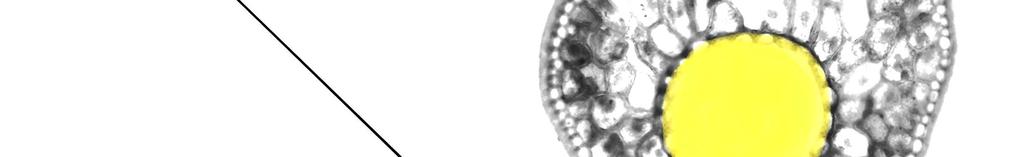


22 Obr. 1 Schematické znázornění měřených anatomických veličin výšky a šířky s vyznačeným umístěním xylému a floému. Celková plocha jehlice je značena zelenou barvou a plocha centrálního válce má žlutou barvu. 23
23 Statistická analýza Statistická analýza dat byla provedena v programu STATISTIKA a QCExpert, kde byly spočítány základní charakteristiky souborů pro jednotlivé znaky. Pro zjištění shody mezi jednotlivými soubory dat bylo použito testování statistických hypotéz. V prvním kroku byl sledován rozdíl mezi kontrolní a stresovanou plochou. Protože hodnoty splňovaly požadavek normálního rozdělení, mohl být použit klasický F-test pro ověření nulové hypotézy na shodu rozptylu a poté studentův t-test na shodu průměrů. Stejné testy byly použity pro posouzení rozdílů u jehlic mezi vrcholem a bázi koruny. 24
24 5. Výsledky Výška jehlice Výsledky analýzy rozdílů výšky jehlic z vrchu a z báze koruny mezi kontrolní a suchem stresovanou plochou jsou uvedeny v tabulce č. 1. Medián výšek jehlic z vrchu koruny z kontrolní plochy je 1168 µm. Interkvartilové rozpětí, v němž se nachází střední polovina všech dat, je µm. Medián výšek jehlic z vrchu koruny ze suchem stresované plochy je 969 µm. Interkvartilové rozpětí je µm. Rozložení hodnot výšky jehlice z vrcholové části koruny je uvedeno v obr. 2. Rozdíl středních hodnot výšky jehlic z vrchu koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně liší (P < 0,001). Medián výšek jehlic z báze koruny z kontrolní plochy je 873 µm. Interkvartilové rozpětí je µm. Medián výšek jehlic z báze koruny ze suchem stresované plochy je 872 µm. Interkvartilové rozpětí je µm. Rozložení hodnot výšky jehlice z báze koruny je uvedeno v obr. 2. Rozdíl středních hodnot výšky jehlic z báze koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně neliší (P = 0, 959). Obr. 2 Rozložení hodnot pro parametr výška jehlice. Levý obrázek zachycuje výšku jehlic z vrchu koruny a pravý výšku jehlic na bázi koruny. 25
25 Šířka jehlice Výsledky analýzy rozdílů šířky jehlic z vrchu a z báze koruny mezi kontrolní a suchem stresovanou plochou jsou uvedeny v tabulce č. 1. Medián šířek jehlic z vrchu koruny z kontrolní plochy je 974 µm. Interkvartilové rozpětí, v němž se nachází střední polovina všech dat, je µm. Medián šířek jehlic z vrchu koruny ze suchem stresované plochy je 1044 µm. Interkvartilové rozpětí je µm. Rozložení hodnot šířky jehlice z vrcholové části koruny je uvedeno v obr. 3. Rozdíl středních hodnot šířky jehlic z vrchu koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně liší (P < 0,001). Medián šířek jehlic z báze koruny z kontrolní plochy je 536 µm. Interkvartilové rozpětí je µm. Medián šířek jehlic z báze koruny ze suchem stresované plochy je 526 µm. Interkvartilové rozpětí je µm. Rozložení hodnot šířky jehlice z báze koruny je uvedeno v obr. 3. Rozdíl středních hodnot šířky jehlic z báze koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně neliší (P = 0,411). Obr. 2 Rozložení hodnot u parametru šířka jehlice. Levý obrázek zachycuje jehlice z vrchu koruny a pravý jehlice z báze koruny. 26
26 Plocha jehlice Výsledky analýzy rozdílů plochy jehlic z vrchu a z báze koruny mezi kontrolní a suchem stresovanou plochou jsou uvedeny v tabulce č. 1. Medián plochy jehlic z vrchu koruny z kontrolní plochy je 0,756 mm 2. Interkvartilové rozpětí, v němž se nachází střední polovina všech dat 0,728-0,783 mm 2. Medián plochy jehlic z vrchu koruny ze suchem stresované plochy je 0,630 mm 2. Interkvartilové rozpětí je 0,605-0,6546 mm 2. Rozložení hodnot plochy jehlice z vrcholové části koruny je uvedeno v obr. 3. Rozdíl středních hodnot plochy jehlic z vrchu koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně liší (P < 0,001). Medián plochy jehlic z báze koruny z kontrolní plochy je 0,342 mm 2. Interkvartilové rozpětí je0,327-0,356 mm 2. Medián plochy jehlic z báze koruny ze suchem stresované plochy je 0,315 mm2. Interkvartilové rozpětí je 0,305-0,325 mm 2. Rozložení hodnot plochy jehlice z báze koruny je uvedeno v obr. 4. Rozdíl středních hodnot plochy jehlic z báze koruny mezi kontrolní a suchem stresovanou plochou se v tomto případě také statisticky významně liší (P = 0,002). Obr. 4 Rozložení hodnot u parametru plocha jehlice. Levý obrázek zachycuje jehlice z vrchu koruny a pravý z báze koruny. 27

27 Plocha centrálního válce Výsledky analýzy rozdílů plochy centrálního válce jehlic z vrchu a z báze koruny mezi kontrolní a suchem stresovanou plochou jsou uvedeny v tabulce č. 1. Medián plochy centrálního válce z vrchu koruny z kontrolní plochy je µm2. Interkvartilové rozpětí, v němž se nachází střední polovina všech dat µm2. Medián plochy jehlic z vrchu koruny ze suchem stresované plochy je µm2. Interkvartilové rozpětí je µm2. Rozložení hodnot plochy centrální válce jehlice z vrcholové části koruny je uvedeno v obr. 5. Rozdíl středních hodnot plochy centrálního válce jehlic z vrchu koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně liší (P < 0,001). Medián plochy centrálního válce jehlic z báze koruny z kontrolní plochy je µm2. Interkvartilové rozpětí je µm2. Medián plochy centrálního válce z báze koruny ze suchem stresované plochy je µm2. Interkvartilové rozpětí je µm2. Rozložení hodnot plochy centrální válce jehlice z báze koruny je uvedeno v obr. 5. Rozdíl středních hodnot plochy centrálního válce jehlic z báze koruny mezi kontrolní a suchem stresovanou plochou se statisticky významně neliší (P = 0,377). Obr. 5 Rozložení hodnot parametru plocha centrálního válce jehlice. Levý obrázek zachycuje jehlice z vrchu koruny a pravý z báze koruny. 28
28 Tabulka 1. Výsledky testů porovnávající rozdíly všech znaků jehlic z vrchu a z báze koruny mezi kontrolní a suchem stresovanou plochou Porovnávané znaky pro vrch Průměr Pravděpodobnost výška (μm) stresovaná 969 kontrolní 1168 < 0,001 šířka (μm) stresovaná 1045 kontrolní 974 < 0,001 p. jehlice (mm 2 ) stresovaná 0,630 kontrolní 0,756 < 0,001 p. cent. válce (μm 2 ) stresovaná kontrolní < 0,001 Porovnávané znaky pro bázi Průměr Pravděpodobnost výška (μm) stresovaná 872 kontrolní 873 0,959 šířka (μm) stresovaná 526 kontrolní 536 0,411 p. jehlice (mm 2 ) stresovaná 0,315 kontrolní 0,342 0,002 p. cent. válce (μm 2 ) stresovaná kontrolní ,377 Rozdíl mezi vrchem a bází koruny U všech porovnávaných znaků výška, šířka, plocha jehlice a plocha centrálního válce byl prokázán statisticky významný rozdíl mezi vrcholem a bázi koruny na obou plochách (Tab. 2 a obr. 6). Tabulka 2. Výsledky testů porovnávající rozdíly u jehlic mezi vrcholem a bází koruny pro jednotlivé znaky. Kontrolní plocha Porovnávané Pravděpodobnost sloupce Stresovaná plocha Porovnávané Pravděpodobnost sloupce výška (μm) < 0,001 výška (μm) < 0,001 šířka (μm) < 0,001 šířka (μm) < 0,001 p. jehlice (mm 2 ) < 0,001 p. jehlice (mm 2 ) < 0,001 p. cent. válce (μm 2 ) < 0,001 p. cent. válce (μm 2 ) < 0,001 29
29 Obr. 6 Krabicové grafy porovnání výšky jehlice z vrchu a báze koruny 30
30 6. Diskuse Výsledkem analýzy porovnání čtyř znaků u jehlic smrku, který byl pěstován na ploše stresované suchem a na kontrolní ploše, bylo nepřijetí nulové hypotézy o rovnosti průměrů (obr.2,3,4,5 a tab. 1). To znamená, že vodní stres měl staticky významný vliv na snížení růstu jehlic. K podobným závěrům došli Grill a kol. (2004) u borovice kanárské a Gebauer a kol. (2010) u smrku ztepilého. Důvodem je, že vodní stres má mimo jiné vliv na pokles turgoru, což vede k omezení růstu rostlinných buněk. Při nedostatku vody se stěny buněk rozpínají méně a velikost buňky je v těchto podmínkách menší. Ke zpomalení růstu dochází již, když turgor klesne na 0,1 až 0,2 MPa. K zastavení růstu tedy dochází dříve, než je patrný vodní stres na vadnutí listu či dříve než jsou ovlivněny metabolické procesy. (Bláha a kol. 2003). U stresovaných jehlic z báze se sledované znaky mimo plochu jehlice nelišily od hodnot z kontrolní plochy (obr.2,3,4,5 a tab. 1). Důvodem této rozdílné reakce báze a vrchu koruny je pravděpodobně rozdílná strategie v distribuci vody v rámci koruny stromu. Strom podporuje vedení vody do horní části koruny, blíže ke slunci, v místě nejaktivnější fotosyntézy. Vodivé elementy na vrchu koruny jsou proto větší, mají menší hydraulický odpor a vedou vodu rychleji (Tyree a Zimmermann 2002). Směrem k bázi zastíněné koruny se vodivé elementy zmenšují a při nedostatku vody jsou to první větve, které odumírají (Gebauer a kol. 2010). Větve na bázi koruny proto rostou na hranici přežití jak u kontrolní tak u stresované plochy. To může být důvodem nepatrných rozdílů mezi plochami. Pokud dojde k zlepšení světelných podmínek prýtů rostoucích na bázi koruny, trvá i několik let než dojde k přeměně anatomické struktury a tím k zvýšení transportu vody k nově exponovaným jehlicím (Kneeshaw a kol. 2002). Velmi významným důsledkem nedostatku vody u rostlin je také ztráta aktivity obranných mechanismů a zvýšení náchylnosti k napadení patogeny (Martinková a Čermák, 1994). Příkladem může být houbová choroba Sphaeropsis sapinea, která způsobuje prosychání výhonů všech druhů borovic a může vést k odumření stromu. Její výskyt se dává do souvislosti právě s vodním deficitem v půdě v předjarním a jarním období (Atlas poškození dřevin, 2013, [on-line]). 31
31 Návrh péče o dřeviny ve městě Negativní vliv stresu suchem na růst, zjištěný na příkladu smrku ztepilého, můžeme obecně aplikovat i na stromy rostoucích ve městech, protože se jedná o prostředí většinou výrazně ovlivněné nedostatkem vody. Město je prostředí pro strom nepřirozené a značně nepříznivé, ve kterém dochází k působení více stresových faktorů naráz. K suchu se například může přidat nedostatek živin, vysoké ph a zasolení. Jestliže vysazujeme strom ve městě, je nesmírně důležité nejprve provést důkladnou analýzu stanoviště, kde chceme stromy sázet. Bereme v úvahu klimatické poměry lokality (nadmořská výška, úhrn srážek, výpar, směr proudění), ale i velikost a vlastnosti kořenového prostoru (půdní typ, ph, obsah živin apod.). Tyto informace nám poslouží k analýze šancí stromu na ujmutí se na daném stanovišti a kolik péče bude potřeba na jeho udržení. (Martinková, ústní sdělení) Základním prostředkem, jak zamezit, aby byl strom stresován suchem, je výběr vhodného druhu. Protože obecně v městském prostředí panují jiné klimatické podmínky např. vlivem jevu tepelného ostrova, je mnohdy vhodnější použít druhy cizokrajné a z pohledu vodního deficitu nejlépe teplomilné a odolné suchu. Příkladem nepůvodních druhů dřevin, nenáročných, sucho snášejících a běžně se vyskytujících jsou např. dřezovec trojtrnný, svitel latnatý nebo zástupci rodu trnovník. Z domácích dřevin vhodných do městského prostředí lze zmínit velmi oblíbený kultivar třešně křovité ' Globosa ' používaný do stromořadí nebo některé kultivary hrušně obecné. Mezi jehličnany odolné suchu patří smrk omorika, smrk pichlavý, borovice Heldreichova nebo borovice lesní (Málek, Horáček, Kiesenbauer, 2012). Závlahy, ochranné sítě, kdy provádět závlahu Příčinou nedostatečné zásoby vody a špatné provzdušenosti půd v městském prostředí je velké množství zpevněných ploch, které v podstatě vodu nepropustí. Doporučuje se proto použít v předpokládaném kořenovém prostoru buď závlahové nebo provzdušňovací prvky a na zvláště nepříznivých stavištích oba systémy zkombinovat (Smýkal a kol., 2008). Závlahové systémy se mají obvykle zřizovat u stromů o obvodu kmene přes 20 cm, u jehličnanů a keřových tvarů stromů výšky přes 2 m. Avšak na špatných stanovištích je používáme bez ohledu na obvod a výšku. Závlahové systémy 32
32 napomáhají lepšímu ujmutí dřeviny na stanovišti a k rychlejšímu překonání povýsadbového šoku. (Smýkal a kol., 2008). Dle Kolaříka a kol.(2003) není třeba do půdy instalovat speciální zavlažovací zařízení, je-li dostatečně velká výsadbová jáma a jsou-li zajištěny dobré vlastnosti půdy, stačí pouze vhodně tvarovat vyvýšené okraje kořenové mísy, které zachytí zálivkovou vodu dodanou na povrch kořenové mísy. Instalace zavlažovacích zařízení má své opodstatnění v extrémních podmínkách městských center či v místech, kde nejsou dostatečně velké kořenové mísy a kde se předpokládá pravidelná zálivka jako součást povýsadbové péče. Na druhou stranu bychom dávky zálivkové vody měli omezovat pouze na nezbytně nutné minimum, protože jinak se snižuje či dokonce ustává činnost obranných mechanismů dřevin vůči suchu. Navíc vysoký obsah vody ve výsadbové jámě může v prvních letech po výsadbě způsobit hnilobu kořenů a následně i úhyn jedince. Intenzita zálivky Potřeba zálivky se po dobu odeznívání povýsadbového šoku stanoví podle vývoje koruny, turgoru listu a vlhkosti půdy pod mulčem nebo pod povrchem půdy. Pokud jsou již listy povadlé, je to symptomem kritického nedostatku vody a zálivka je neodkladná. Závlahová dávka se stanovuje v takovém množství, aby se provlhčila prokořenělá vrstva půdy. Menší dávky vody podporují rozvoj kořenového systému pouze ve vrchní vrstvě, což se negativně projeví při přísušku. Vysoké dávky zase na druhé straně vyplavují živiny. Soustavné přelévání vytěsní z půdy vzduch a dojde k poškození kořenů nedostatkem kyslíku nebo až k jejich uhynutí. Příznaky přelití jsou stejné jako při nedostatku vody, listy vadnou a postupně usychají, protože kořeny nejsou schopny přijímat vodu. Zálivka po odeznění výsadbového šoku by se měla omezit pouze na kritické období sucha (Smýkal a kol., 2008). Zálivka je zahrnuta i ve standardu Výsadby stromů, jako součást povýsadbové péče o stromy. Podle standartu by měla být závlahová mísa udržovaná minimálně 2 roky po výsadbě a po celou dobu, kdy probíhá zálivka stromu. Kvalita vody se řídí ČSN a její kvalita se musí pravidelně kontrolovat. Zálivka by se měla provádět tak dlouho, dokud neodezní povýsadbový šok. Délka povýsadbového šoku je stanovena na dobu 1 roku na každých 80 mm obvodu kmene. Na extrémních stanovištích se však řídíme konkrétními podmínkami a závlaha probíhá až do řádného zakořenění. V případech, kdy není možné propojení kořenového prostoru na rostlý terén je nutné 33
33 strom zavlažovat po celou dobu jeho existence. Ve většině případů je vhodný cyklus 6-8 zálivek během prvního vegetačního období po výsadbě. Ve druhém se roce se četnost zálivek snižuje na 3-6. Zálivka nesmí probíhat vodou pod tlakem, aby nedocházelo k vymývání půdy. Přesto však nejde přesně určit kolik, a jak často by se měly stromy zalévat, vše se odvíjí od požadavků daného taxonu, vlastnostech stanoviště a aktuálního počasí (Martinková, ústní sdělení). Zavlažovací systémy Zálivku lze provádět ručně či automatizovaně, nebo s pomocí speciálních zavlažovacích zařízení. Smýkal a kol. (2008) rozděluje zavlažovací systémy na horizontální, vertikální, kombinované a automatické závlahy. U horizontální závlahy se perforovaná trubka se umísťuje horizontálně po obvodu kořenového systému a u vertikální závlahy se perforované trubky osazují kolmo tak, aby vyústění bylo v úrovni s terénem. Tento systém až pětinásobně zrychluje zasakování zálivkové vody oproti zálivce do závlahové mísy. Kombinované závlahy se zřizují zejména ve zpevněných plochách a obvykle se skládají z horizontálních vrchních trubek uložených v soustředných kružnicích vzájemně propojených. Kolařík a kol. (2003) popisuje způsob závlahy pomocí kořenové (závlahové) sondy. Díky pravidelné zálivce je bezprostřední okolí této sondy hustě a jemně prokořeněno, má dostatek vody i vzduchu. Je-li však pravidelná zálivka přerušena, jsou tyto kořeny snadno poškozovány suchem. Toto nebezpečí lze snížit uzavřením otvoru na povrchu neprodyšným uzávěrem, čímž se ale naopak může výrazně snížit větrání, není-li zajištěno jinak. Když dojde k ucpání sondy, je třeba její nadzemní část odstranit nebo ji zasypat substrátem, který bude propustný pro vzduch, aby mohlo docházet k větrání, ale na druhou stranu nedocházelo k vysoušení kořenů v blízkosti sondy. Na exponovaných místech, např. v centrech měst s množstvím přehřátých ploch a v sušších oblastech se doporučuje zřízení automatické závlahy. K jednotlivým stromům s malými kořenovými mísami se osvědčila kapková závlaha. (Smýkal a kol, 2008) 34
34 7. Závěr Stres suchem se nejvíce projevil u jehlic z vrcholové části koruny snížením hodnot u všech sledovaných parametrů (výška, šířka, plocha jehlice a plocha centrálního válce). Z výsledků analýzy dále vyplývá, že rozdíly mezi kontrolní a suchem stresovanou plochou u jehlic z báze koruny jsou staticky nevýznamné s výjimkou parametru plocha jehlice. To může být způsobeno růstem bazálních větví v podmínkách na hranici přežití jak u kontrolní tak u stresované plochy. Z hlediska stromů v městském prostředí je stres suchem jedním z nejvýznamnějších limitujících faktorů jejich růstu. Nejdůležitějším opatřením je výběr vhodného taxonu pro podmínky daného stanoviště a také úprava půdního prostředí před výsadbou. Automatickou součástí péče o vysazený strom je závlaha, která pomůže překonat povýsadbový šok a zvýší šance ujmutí stromu na stanovišti. 35
35 8. Summary The impact of drought stress was more pronounced in needles from crown top. Histological structures as height, width, needle area and central cylinder were lower in the stress needles and thus needle growth was restricted during drought. The results also show that the difference between control and drought-stressed plot were statistically not significant (except of needle area) in the case of needles from crown base. This could be consequence that shoots in the crown base lives close to critical point for survival on both plots. In terms of trees in the urban environment drought stress is one of the most important factors limiting their growth. The most important measure is the selection of a suitable taxon for concrete site condition and the soil improvement before planting. Irrigation needs to be included in the after-planting care to overcome the post-planting shock and increase the chances for tree survive on site. 36
36 9. Seznam použité literatury AUTORSKÝ KOLEKTIV., Standardy péče o přírodu a krajinu. Arboristické standardy. Řada A. Výsadba stromů. Brno. Mendelova univerzita v Brně. 47 s. BLÁHA, L. a kol., Rostlina stres. Praha. Výzkumný ústav rostlinné výroby.180 s FITTER, A., HAY, R., Environmental physiology of plants 3rd edition. London. Academic Press. 367 s. GEBAUER, R. a kol., Metodika sledování vitality lesních ekosystémů vystavených suchu k podpoře trvale udržitelného lesnictví. Brno. Mendelova univerzita v Brně. 56 s. GRILL a kol., Effects of drought on needle anatomy of Pinus canariensis. Flora 199: KNEESHAW a kol., Patterns of above- and below-ground response of understory conifer release 6 years after partial cutting. Canadian Journal of Forest Research 32 KOLAŘÍK, J. a kol., Péče o dřeviny rostoucí mimo les I. Vlašim. ČSOP. 261 s. KOLAŘÍK, J. a kol., Péče o dřeviny rostoucí mimo les II. Vlašim. ČSOP. 696 s. LARCHER, W., HOUBA, M., BAUER, V., Fyziologická ekologie rostlin. 1. vyd. Praha. Academia. 361 s. MÁLEK, Z., HORÁČEK, P., KIESENBAUER, Z., Stromy pro sídla a krajinu. Olomouc: Petr Baštan ve spolupráci s firmou Arboeko.357 s. MARTINKOVÁ, M., ČERMÁK, J., Fyziologická predispozice lesních dřevin k napadení fytofágy a možnosti obranných reakcí. In: Kůrovcová kalamita: příčiny, rozsah, ochrana. Brno, VŠZ v Brně. ÖPIK, H., ROLFE, S. A., The physiology of flowering plants. 4. vyd. New York. Cambridge University Press. 392 s. PROCHÁZKA, S. a kol., Fyziologie rostlin. Praha. Academia. 484 s. PROCHÁZKA, S. a kol., Botanika: Morfologie a fyziologie rostlin. Brno. Mendelova zemědělská a lesnická univerzita v Brně. 242 s. 4 SMÝKAL, F. A kol., Arboristika II. Mělník. VOŠ Za a SZaŠ Mělník. 260 s. TYREE MT, ZIMMERMANN MH., Xylem structure and the ascent of sap. Springer, Berlin 37
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
 Život rostlin (i ostatních organismů) je neoddělitelně spjat s vodou stálou a nenahraditelnou složkou rostlinného těla. první rostliny vznikly ve vodním prostředí, kde velmi dlouho probíhala jejich evoluce;
Život rostlin (i ostatních organismů) je neoddělitelně spjat s vodou stálou a nenahraditelnou složkou rostlinného těla. první rostliny vznikly ve vodním prostředí, kde velmi dlouho probíhala jejich evoluce;
VODNÍ REŽIM ROSTLIN. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1
 VODNÍ REŽIM ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1 Význam vody pro rostlinu: Rozpouštědlo, transport látek. Účastní se fotosyntézy a dýchání. Termoregulační
VODNÍ REŽIM ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1 Význam vody pro rostlinu: Rozpouštědlo, transport látek. Účastní se fotosyntézy a dýchání. Termoregulační
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Fyziologické a anatomické přizpůsobení sazenic na stres suchem - metody studia stresu
 Fyziologické a anatomické přizpůsobení sazenic na stres suchem - metody studia stresu Doc. Roman Gebauer; Ing. Roman Plichta, Ph.D.; Doc. Josef Urban Fakulta lesnická a dřevařská, Ústav lesnické botaniky,
Fyziologické a anatomické přizpůsobení sazenic na stres suchem - metody studia stresu Doc. Roman Gebauer; Ing. Roman Plichta, Ph.D.; Doc. Josef Urban Fakulta lesnická a dřevařská, Ústav lesnické botaniky,
Transport živin do rostliny. Radiální a xylémový transport. Mimokořenová výživa rostlin.
 Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Vodní režim rostlin. Obsah vody, RWC, vodní potenciál a jeho komponenty: Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy,
 Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Obsah vody v rostlinách
 Transpirace 1/39 Obsah vody v rostlinách Obsah vody v protoplazmě (její hydratace) je nezbytný pro normální průběh životních funkcí buňky. Snížení obsahu vody má za následek i omezení životních dějů (pozorovatelné
Transpirace 1/39 Obsah vody v rostlinách Obsah vody v protoplazmě (její hydratace) je nezbytný pro normální průběh životních funkcí buňky. Snížení obsahu vody má za následek i omezení životních dějů (pozorovatelné
Měření odporu transportních cest, stupně jejich integrace a embolizace
 Měření odporu transportních cest, stupně jejich integrace a embolizace Vít Gloser Cvičení z fyziologie rostlin pro pokročilé Základní principy xylémového transportu vody (1) Tok vody v xylému je možný
Měření odporu transportních cest, stupně jejich integrace a embolizace Vít Gloser Cvičení z fyziologie rostlin pro pokročilé Základní principy xylémového transportu vody (1) Tok vody v xylému je možný
Vodní režim rostlin. Transport vody v xylemu. Kohezní teorie. Transport půda-rostlina-atmosféra. Metody měření. Kavitace
 Vodní režim rostlin Transport vody v xylemu Transport půda-rostlina-atmosféra Kohezní teorie Kavitace Metody měření Longitudinální transport v systému půda-rostlina-atmosféra Hnací síla gradient vodního
Vodní režim rostlin Transport vody v xylemu Transport půda-rostlina-atmosféra Kohezní teorie Kavitace Metody měření Longitudinální transport v systému půda-rostlina-atmosféra Hnací síla gradient vodního
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
Biologie - Kvinta, 1. ročník
 - Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
- Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
Jak psát závěrečnou práci na LDF
 17. 3. 2014, Brno Připravil: Hanuš Vavrčík Náležitosti a členění na kapitoly strana 2 Čím se řídit? Směrnice děkana č. 2/2007 O úpravě písemných prací a o citaci dokumentů užívaných v kvalifikačních pracích
17. 3. 2014, Brno Připravil: Hanuš Vavrčík Náležitosti a členění na kapitoly strana 2 Čím se řídit? Směrnice děkana č. 2/2007 O úpravě písemných prací a o citaci dokumentů užívaných v kvalifikačních pracích
Funkce vody v rostlinném těle. Růstová (hydratační) Metabolická Termoregulační Zásobní Transportní (tranzitní) Volná a vázaná voda
 Metabolická Termoregulační Zásobní Transportní (tranzitní) Volná a vázaná voda") VODNÍ REŽIM ROSTLIN Funkce vody v rostlinném těle Vyjádření stavu vody v rostlině Vodní stav rostlinné buňky Příjem a vedení vody rostlinou Výdej vody rostlinou Hospodaření rostliny s vodou Funkce vody
VODNÍ REŽIM ROSTLIN Funkce vody v rostlinném těle Vyjádření stavu vody v rostlině Vodní stav rostlinné buňky Příjem a vedení vody rostlinou Výdej vody rostlinou Hospodaření rostliny s vodou Funkce vody
Kořenový systém plodin jako adaptační opatření na sucho
 Sucho a degradace půd v České republice - 2014 Brno 7. 10. 2014 Kořenový systém plodin jako adaptační opatření na sucho Vodní provoz polních plodin Ing. Jana Klimešová Ing. Tomáš Středa, Ph.D. Mendelova
Sucho a degradace půd v České republice - 2014 Brno 7. 10. 2014 Kořenový systém plodin jako adaptační opatření na sucho Vodní provoz polních plodin Ing. Jana Klimešová Ing. Tomáš Středa, Ph.D. Mendelova
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Stav lesních půd drama s otevřeným koncem
 Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
SOUČASNÉ PROBLÉMY OBNOVY LESŮ A STAV KOŘENOVÉHO SYSTÉMU LESNÍCH DŘEVIN V ZÁVISLOSTI NA MĚNÍCÍM SE PODNEBÍ
 SOUČASNÉ PROBLÉMY OBNOVY LESŮ A STAV KOŘENOVÉHO SYSTÉMU LESNÍCH DŘEVIN V ZÁVISLOSTI NA MĚNÍCÍM SE PODNEBÍ VÝCHODISKA Propracované a odzkoušené postupy jsou neúspěšné, ztráty po obnově až 6 %, snižuje se
SOUČASNÉ PROBLÉMY OBNOVY LESŮ A STAV KOŘENOVÉHO SYSTÉMU LESNÍCH DŘEVIN V ZÁVISLOSTI NA MĚNÍCÍM SE PODNEBÍ VÝCHODISKA Propracované a odzkoušené postupy jsou neúspěšné, ztráty po obnově až 6 %, snižuje se
Vlastnosti vody. Voda má jednoduché chemické složení (H 2 O) Kyslík s vodíkem je spojen kovalentní vazbou polárního charakter.
 Kyslík s vodíkem je spojen kovalentní vazbou polárního charakter.") Vlastnosti vody Voda má jednoduché chemické složení (H 2 O) δ - δ - H O H δ + δ + Kyslík s vodíkem je spojen kovalentní vazbou polárního charakter vodíkové vazby vodíkové můstky δ - δ + δ - δ + δ - δ+
Vlastnosti vody Voda má jednoduché chemické složení (H 2 O) δ - δ - H O H δ + δ + Kyslík s vodíkem je spojen kovalentní vazbou polárního charakter vodíkové vazby vodíkové můstky δ - δ + δ - δ + δ - δ+
2) Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.
 Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.") Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Meteorologické faktory transpirace
 Člověk ve svém pozemském a kosmickém prostředí Zlíč 17. - 19. květen 2016 Meteorologické faktory transpirace Ing. Jana Klimešová Ing. Tomáš Středa, Ph.D. Mendelova univerzita v Brně Vodní provoz polních
Člověk ve svém pozemském a kosmickém prostředí Zlíč 17. - 19. květen 2016 Meteorologické faktory transpirace Ing. Jana Klimešová Ing. Tomáš Středa, Ph.D. Mendelova univerzita v Brně Vodní provoz polních
Otázky k předmětu Globální změna a lesní ekosystémy
 Otázky k předmětu Globální změna a lesní ekosystémy 1. Jaké jsou formy šíření energie v klimatickém systému Země? (minimálně 4 formy) 2. Na čem závisí množství vyzářené energie tělesem? (minimálně 3 faktory)
Otázky k předmětu Globální změna a lesní ekosystémy 1. Jaké jsou formy šíření energie v klimatickém systému Země? (minimálně 4 formy) 2. Na čem závisí množství vyzářené energie tělesem? (minimálně 3 faktory)
Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim
 Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník
Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník
Bohumil Koníček vedoucí odboru správy majetku Městský úřad Neratovice Kojetická Neratovice. Martinov
 Bohumil Koníček vedoucí odboru správy majetku Městský úřad Neratovice Kojetická 1028 277 11 Neratovice Dendrologické posouzení dřeviny, rostoucí na pozemku č. 185/17 k.ú. Neratovice Quercus robur dub letní
Bohumil Koníček vedoucí odboru správy majetku Městský úřad Neratovice Kojetická 1028 277 11 Neratovice Dendrologické posouzení dřeviny, rostoucí na pozemku č. 185/17 k.ú. Neratovice Quercus robur dub letní
Systémový přístup v pohledu na stromy
 Systémový přístup v pohledu na stromy Petr Horáček Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Obsah Strana 3 1 Východiska 2 Současnost
Systémový přístup v pohledu na stromy Petr Horáček Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Obsah Strana 3 1 Východiska 2 Současnost
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie
 synekologie b) autekologie c) demekologie") 1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
Učební texty Univerzity Karlovy v Praze. Jana SlavíKová JitKa Švíglerová. Fyziologie DÝCHÁNÍ. Karolinum
 Učební texty Univerzity Karlovy v Praze Jana SlavíKová JitKa Švíglerová Fyziologie DÝCHÁNÍ Karolinum Fyziologie dýchání doc. MUDr. Jana Slavíková, CSc. MUDr. Jitka Švíglerová, Ph.D. Recenzovali: prof.
Učební texty Univerzity Karlovy v Praze Jana SlavíKová JitKa Švíglerová Fyziologie DÝCHÁNÍ Karolinum Fyziologie dýchání doc. MUDr. Jana Slavíková, CSc. MUDr. Jitka Švíglerová, Ph.D. Recenzovali: prof.
ROSTLINNÁ FYZIOLOGIE OSMOTICKÉ JEVY
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Vznik dřeva přednáška
 Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
Biologie. Pracovní list č. 6 žákovská verze Téma: Transpirace u rostlin. Lektor: Mgr. Naděžda Kurowská. Student a konkurenceschopnost
 www.projektsako.cz Biologie Pracovní list č. 6 žákovská verze Téma: Transpirace u rostlin Lektor: Mgr. Naděžda Kurowská Projekt: Reg. číslo: Student a konkurenceschopnost CZ.1.07/1.1.07/03.0075 Teorie:
www.projektsako.cz Biologie Pracovní list č. 6 žákovská verze Téma: Transpirace u rostlin Lektor: Mgr. Naděžda Kurowská Projekt: Reg. číslo: Student a konkurenceschopnost CZ.1.07/1.1.07/03.0075 Teorie:
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Základní komponenty modelu lesa. Jan Kadavý
 Základní komponenty modelu lesa Jan Kadavý Základní členění modelů Zdroj: Fabrika, Pretzsch 20011: Analýza a modelovanie lesných ekosystémov. Klasifikace modelů Předmět prezentace Zdroj: Fabrika, Pretzsch
Základní komponenty modelu lesa Jan Kadavý Základní členění modelů Zdroj: Fabrika, Pretzsch 20011: Analýza a modelovanie lesných ekosystémov. Klasifikace modelů Předmět prezentace Zdroj: Fabrika, Pretzsch
Zpráva o testu dřevin na pozemku ve Stachách na Šumavě
 Ústřední kontrolní a zkušební ústav zemědělský Oddělení půdy a lesnictví Zpráva o testu dřevin na pozemku ve Stachách na Šumavě Průběžná zpráva Zpracoval: Ing. Dušan Reininger, Ph.D Dr.Ing. Přemysl Fiala
Ústřední kontrolní a zkušební ústav zemědělský Oddělení půdy a lesnictví Zpráva o testu dřevin na pozemku ve Stachách na Šumavě Průběžná zpráva Zpracoval: Ing. Dušan Reininger, Ph.D Dr.Ing. Přemysl Fiala
Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2
 Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2 Obsah tématu: 1) Vzdušný obal země 2) Složení vzduchu 3) Tlak vzduchu 4) Vítr 5) Voda 1) VZDUŠNÝ OBAL ZEMĚ Vzdušný obal Země.. je směs
Pracovní list č. 3 téma: Povětrnostní a klimatičtí činitelé část 2 Obsah tématu: 1) Vzdušný obal země 2) Složení vzduchu 3) Tlak vzduchu 4) Vítr 5) Voda 1) VZDUŠNÝ OBAL ZEMĚ Vzdušný obal Země.. je směs
Ing. Matěj Orság Vodní bilance rychle rostoucích dřevin
 Ing. Matěj Orság Vodní bilance rychle rostoucích dřevin 16. května 2013, od 9.00 hod, zasedací místnost děkanátu AF (budova C) Akce je realizována vrámci klíčové aktivity 02 Interdisciplinární vzdělávání
Ing. Matěj Orság Vodní bilance rychle rostoucích dřevin 16. května 2013, od 9.00 hod, zasedací místnost děkanátu AF (budova C) Akce je realizována vrámci klíčové aktivity 02 Interdisciplinární vzdělávání
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení kvality sadebního materiálu cvičení Zadání: Objednavatel si ve školce ŠLP MENDELU
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení kvality sadebního materiálu cvičení Zadání: Objednavatel si ve školce ŠLP MENDELU
HODNOCENÍ ZDRAVOTNÍHO STAVU A PROVOZNÍ BEZPEČNOSTI STROMŮ
 HODNOCENÍ ZDRAVOTNÍHO STAVU A PROVOZNÍ BEZPEČNOSTI STROMŮ Luděk Praus Ústav nauky o dřevě, LDF MENDELU v Brně Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018
HODNOCENÍ ZDRAVOTNÍHO STAVU A PROVOZNÍ BEZPEČNOSTI STROMŮ Luděk Praus Ústav nauky o dřevě, LDF MENDELU v Brně Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018
Průduchy regulace příjmu CO 2
 Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Dynamický fluidní model membrány 2008/11
 Dynamický fluidní model membrány 2008/11") RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány Membránový
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány Membránový
Biologické základy péče o stromy II.
 Biologické základy péče o stromy II. Ing. Jaroslav Kolařík, Ph.D. Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 PLETIVA VODIVÁ - lýko
Biologické základy péče o stromy II. Ing. Jaroslav Kolařík, Ph.D. Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 PLETIVA VODIVÁ - lýko
Jak psát závěrečnou práci na LDF
 28. 2. 2019, Brno Připravil: Hanuš Vavrčík Jak psát závěrečnou práci na LDF Závazné dokumenty Zásady psaní závěrečných prací Další doporučení Jak psát závěrečnou práci na LDF Závazné dokumenty Jak psát
28. 2. 2019, Brno Připravil: Hanuš Vavrčík Jak psát závěrečnou práci na LDF Závazné dokumenty Zásady psaní závěrečných prací Další doporučení Jak psát závěrečnou práci na LDF Závazné dokumenty Jak psát
Protimrazová ochrana rostlin
 Protimrazová ochrana rostlin Denní variabilita teploty Každý den představuje sám o sobě jedinečnou vegetační sezónu Denní teplota Sluneční záření Vyzářená energiedlouhovlnná radiace Východ slunce Západ
Protimrazová ochrana rostlin Denní variabilita teploty Každý den představuje sám o sobě jedinečnou vegetační sezónu Denní teplota Sluneční záření Vyzářená energiedlouhovlnná radiace Východ slunce Západ
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková
 Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
 Závlahové režimy Řízení závlahového režimu = stanovení optimální velikosti závlahové dávky a termínu jejího dodání Kvalifikované řízení závlahových režimů plodin - jeden ze základních předpokladů rentability
Závlahové režimy Řízení závlahového režimu = stanovení optimální velikosti závlahové dávky a termínu jejího dodání Kvalifikované řízení závlahových režimů plodin - jeden ze základních předpokladů rentability
Metody řízení závlahy ve sklenících a kontejnerovnách. Tomáš Litschmann
 Metody řízení závlahy ve sklenících a kontejnerovnách Tomáš Litschmann Zásadní informace, nutné pro efektivní řízení závlahy Kolik vody rostliny spotřebují Kolik vody je v půdě (substrátu) Kolik vody dodává
Metody řízení závlahy ve sklenících a kontejnerovnách Tomáš Litschmann Zásadní informace, nutné pro efektivní řízení závlahy Kolik vody rostliny spotřebují Kolik vody je v půdě (substrátu) Kolik vody dodává
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
VAKUOLY - voda v rostlinné buňce
 VAKUOLY - voda v rostlinné buňce Úvod: O vakuole: Vakuola je membránová struktura, která je součástí většiny rostlinných buněk. Může zaujímat 30-90% objemu buňky. Vakuola plní v rostlinné buňce mnoho důležitých
VAKUOLY - voda v rostlinné buňce Úvod: O vakuole: Vakuola je membránová struktura, která je součástí většiny rostlinných buněk. Může zaujímat 30-90% objemu buňky. Vakuola plní v rostlinné buňce mnoho důležitých
Posouzení stavu stromu a návrh pěstebních opatření Lípa Nová Ves
 Posouzení stavu stromu a návrh pěstebních opatření Lípa Nová Ves Zhotovitel: Bc. Robert Kohout Čajkovského 549/12 67401 Třebíč IČ: 68662416 DIČ: CZ7501154551 Tel: +420 777 142 814 Email: robert@arbokohout.cz
Posouzení stavu stromu a návrh pěstebních opatření Lípa Nová Ves Zhotovitel: Bc. Robert Kohout Čajkovského 549/12 67401 Třebíč IČ: 68662416 DIČ: CZ7501154551 Tel: +420 777 142 814 Email: robert@arbokohout.cz
SLEDOVÁNÍ JARNÍCH FENOLOGICKÝCH FÁZÍ U BUKU LESNÍHO VE SMÍŠENÉM POROSTU KAMEROVÝM SYSTÉMEM
 SLEDOVÁNÍ JARNÍCH FENOLOGICKÝCH FÁZÍ U BUKU LESNÍHO VE SMÍŠENÉM POROSTU KAMEROVÝM SYSTÉMEM Bednářová, E. 1, Kučera, J. 2, Merklová, L. 3 1,3 Ústav ekologie lesa Lesnická a dřevařská fakulta, Mendelova
SLEDOVÁNÍ JARNÍCH FENOLOGICKÝCH FÁZÍ U BUKU LESNÍHO VE SMÍŠENÉM POROSTU KAMEROVÝM SYSTÉMEM Bednářová, E. 1, Kučera, J. 2, Merklová, L. 3 1,3 Ústav ekologie lesa Lesnická a dřevařská fakulta, Mendelova
Úbytek stratosférického ozónu a pozorované abiotické poškození rostlin u nás
 Úbytek stratosférického ozónu a pozorované abiotické poškození rostlin u nás Libuše Májková, Státní rostlinolékařská správa Opava Tomáš Litschmann, soudní znalec v oboru meteorologie a klimatologie, Moravský
Úbytek stratosférického ozónu a pozorované abiotické poškození rostlin u nás Libuše Májková, Státní rostlinolékařská správa Opava Tomáš Litschmann, soudní znalec v oboru meteorologie a klimatologie, Moravský
Volitelný předmět Habituální diagnostika
 Tomáš Žid tomas.zid@mendelu.cz LDF MENDELU Volitelný předmět Habituální diagnostika Představení a náplň předmětu Harmonogram předmětu Ukončení předmětu zápočtem Studijní literatura Představení a náplň
Tomáš Žid tomas.zid@mendelu.cz LDF MENDELU Volitelný předmět Habituální diagnostika Představení a náplň předmětu Harmonogram předmětu Ukončení předmětu zápočtem Studijní literatura Představení a náplň
6. Tzv. holocenní klimatické optimum s maximálním rozvojem lesa bylo typické pro a) preboreál b) atlantik c) subrecent
 preboreál b) atlantik c) subrecent") 1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
1. Ekologie zabývající se studiem populací se nazývá a) synekologie b) autekologie c) demekologie 2. Plocha lesa na planetě dle statistiky ročně: a) stoupá cca o 11 mil. ha b) klesá cca o 16 mil. ha c)
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Odborná informace stav hlohů vysazených v linii mezi chodníkem a komunikací v Revoluční ulici v Nýřanech.
 Odborná informace stav hlohů vysazených v linii mezi chodníkem a komunikací v Revoluční ulici v Nýřanech. Druh: Crataegus laevigata hloh obecný Parc.č.: 1316/1 Katastrální území: Nýřany Tato odborná informace
Odborná informace stav hlohů vysazených v linii mezi chodníkem a komunikací v Revoluční ulici v Nýřanech. Druh: Crataegus laevigata hloh obecný Parc.č.: 1316/1 Katastrální území: Nýřany Tato odborná informace
Zpráva o testu klonů topolů a vrb na pozemku ve Stachách na Šumavě
 Ústřední kontrolní a zkušební ústav zemědělský Oddělení bezpečnosti půdy a lesnictví Zpráva o testu klonů topolů a vrb na pozemku ve Stachách na Šumavě Průběžná zpráva Zpracoval : Ing. Dušan Reininger
Ústřední kontrolní a zkušební ústav zemědělský Oddělení bezpečnosti půdy a lesnictví Zpráva o testu klonů topolů a vrb na pozemku ve Stachách na Šumavě Průběžná zpráva Zpracoval : Ing. Dušan Reininger
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
7/12. Vlhkost vzduchu Výpar
 7/12 Vlhkost vzduchu Výpar VLHKOST VZDUCHU Obsah vodní páry v ovzduší Obsah vodní páry závisí na teplotě vzduchu Vzduch obsahuje vždy proměnlivé množství vodních par Vodní pára vzniká ustavičným vypařováním
7/12 Vlhkost vzduchu Výpar VLHKOST VZDUCHU Obsah vodní páry v ovzduší Obsah vodní páry závisí na teplotě vzduchu Vzduch obsahuje vždy proměnlivé množství vodních par Vodní pára vzniká ustavičným vypařováním
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
Faktory ovlivňující strukturu dřeva
 Faktory ovlivňující strukturu dřeva přednáška 1 strana 2 2 Připomenutí základních poznatků strana 3 3 Dřevo definice Dřevo (xylém) definice soubor rostlinných pletiv, která se u dřevin nachází mezi kambiem
Faktory ovlivňující strukturu dřeva přednáška 1 strana 2 2 Připomenutí základních poznatků strana 3 3 Dřevo definice Dřevo (xylém) definice soubor rostlinných pletiv, která se u dřevin nachází mezi kambiem
Vliv aplikace kompostu na povrchový odtok vody při dešťových srážkách
 ..16 Vliv aplikace kompostu na povrchový odtok vody při dešťových srážkách Výzkumný ústav zemědělské techniky, v.v.i. Ing. Pavel Kovaříček, CSc. Metody měření povrchového odtoku Měření při simulovaných
..16 Vliv aplikace kompostu na povrchový odtok vody při dešťových srážkách Výzkumný ústav zemědělské techniky, v.v.i. Ing. Pavel Kovaříček, CSc. Metody měření povrchového odtoku Měření při simulovaných
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Rostlinná buňka jako osmotický systém
 Rostlinná buňka jako osmotický systém Voda se do rostlinné buňky i z ní pohybuje pouze pasivně, difusí. Hnací silou difuse vody jsou rozdíly tzv. vodního potenciálu ( ). Vodní potenciál je chemický potenciál
Rostlinná buňka jako osmotický systém Voda se do rostlinné buňky i z ní pohybuje pouze pasivně, difusí. Hnací silou difuse vody jsou rozdíly tzv. vodního potenciálu ( ). Vodní potenciál je chemický potenciál
2. Otázky k zamyšlení
 Úloha č. 3: Měření vodního a osmotického potenciálu psychrometricky 1. Co je to vodní potenciál (Ψ w ) systému půda(voda) rostlina atmosféra? Vodní potenciál Ψ w je definován jako aktivita vody v systému.
Úloha č. 3: Měření vodního a osmotického potenciálu psychrometricky 1. Co je to vodní potenciál (Ψ w ) systému půda(voda) rostlina atmosféra? Vodní potenciál Ψ w je definován jako aktivita vody v systému.
Na květen je sucho extrémní
 14. května 2018, v Praze Na květen je sucho extrémní Slabá zima v nížinách, podprůměrné srážky a teplý a suchý duben jsou příčinou současných projevů sucha, které by odpovídaly letním měsícům, ale na květen
14. května 2018, v Praze Na květen je sucho extrémní Slabá zima v nížinách, podprůměrné srážky a teplý a suchý duben jsou příčinou současných projevů sucha, které by odpovídaly letním měsícům, ale na květen
ZÁKLADY ARBORISTIKY. Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.
 ZÁKLADY ARBORISTIKY Proč ochrana při stavební činnosti Legislativní souvislosti Co je součástí Projektová příprava Ochranná opatření Kontrola Skutečnost x teorie Park x stavba Standard o přírodu a krajinu
ZÁKLADY ARBORISTIKY Proč ochrana při stavební činnosti Legislativní souvislosti Co je součástí Projektová příprava Ochranná opatření Kontrola Skutečnost x teorie Park x stavba Standard o přírodu a krajinu
ANALÝZY HISTORICKÝCH DEŠŤOVÝCH ŘAD Z HLEDISKA OCHRANY PŮDY PŘED EROZÍ
 Rožnovský, J., Litschmann, T. (ed): Seminář Extrémy počasí a podnebí, Brno, 11. března 24, ISBN 8-8669-12-1 ANALÝZY HISTORICKÝCH DEŠŤOVÝCH ŘAD Z HLEDISKA OCHRANY PŮDY PŘED EROZÍ František Toman, Hana Pokladníková
Rožnovský, J., Litschmann, T. (ed): Seminář Extrémy počasí a podnebí, Brno, 11. března 24, ISBN 8-8669-12-1 ANALÝZY HISTORICKÝCH DEŠŤOVÝCH ŘAD Z HLEDISKA OCHRANY PŮDY PŘED EROZÍ František Toman, Hana Pokladníková
SOUHRNNÝ PŘEHLED nově vytvořených / inovovaných materiálů v sadě
 SOUHRNNÝ PŘEHLED nově vytvořených / inovovaných materiálů v sadě Název projektu Zlepšení podmínek vzdělávání SZŠ Číslo projektu CZ.1.07/1.5.00/34.0358 Název školy Střední zdravotnická škola, Turnov, 28.
SOUHRNNÝ PŘEHLED nově vytvořených / inovovaných materiálů v sadě Název projektu Zlepšení podmínek vzdělávání SZŠ Číslo projektu CZ.1.07/1.5.00/34.0358 Název školy Střední zdravotnická škola, Turnov, 28.
NALEZENÍ A OVĚŘENÍ PROVOZNĚ VYUŽITELNÉ
 Lesy České republiky, s. p., Hradec Králové V Ý ZK U M N É P R O J E K T Y GRANTOVÉ SLUŽBY LČR Projekt NALEZENÍ A OVĚŘENÍ PROVOZNĚ VYUŽITELNÉ METODY PRO HODNOCENÍ AKTUÁLNÍHO FYZIOLOGICKÉHO STAVU SADEBNÍHO
Lesy České republiky, s. p., Hradec Králové V Ý ZK U M N É P R O J E K T Y GRANTOVÉ SLUŽBY LČR Projekt NALEZENÍ A OVĚŘENÍ PROVOZNĚ VYUŽITELNÉ METODY PRO HODNOCENÍ AKTUÁLNÍHO FYZIOLOGICKÉHO STAVU SADEBNÍHO
PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ
 PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ LIGNOSULFONÁTY Lignin představuje heterogenní amorfní polymer potřebný pro pevnost a tuhost dřevnatých buněčných stěn rostlin. Po celulóze je to druhá nejrozšířenější látka
PŘÍPRAVKY NA BÁZI LIGNOSULFONÁTŮ LIGNOSULFONÁTY Lignin představuje heterogenní amorfní polymer potřebný pro pevnost a tuhost dřevnatých buněčných stěn rostlin. Po celulóze je to druhá nejrozšířenější látka
FYZIOLOGIE ROSTLIN Laboratorní práce č. 3
 Téma: Vodní režim rostlin FYZIOLOGIE ROSTLIN Laboratorní práce č. 3 Pozn: Úkoly 1-3 vyhodnoťte po 24 hodinách až týdnu. Prodiskutujte výsledky nejprve teoreticky, poté srovnejte s výsledkem skutečným.
Téma: Vodní režim rostlin FYZIOLOGIE ROSTLIN Laboratorní práce č. 3 Pozn: Úkoly 1-3 vyhodnoťte po 24 hodinách až týdnu. Prodiskutujte výsledky nejprve teoreticky, poté srovnejte s výsledkem skutečným.
Podmínky působící na organismy: abiotické - vlivy neživé části prostředí na organismus biotické - vlivy ostatních živých organismů na život jedince, m
 Přednáška č. 4 Pěstitelství, základy ekologie, pedologie a fenologie Země Podmínky působící na organismy: abiotické - vlivy neživé části prostředí na organismus biotické - vlivy ostatních živých organismů
Přednáška č. 4 Pěstitelství, základy ekologie, pedologie a fenologie Země Podmínky působící na organismy: abiotické - vlivy neživé části prostředí na organismus biotické - vlivy ostatních živých organismů
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Klima jako jeden z půdotvorných faktorů, dopady sucha
 Sucho a degradace půd v České republice - 2014 Brno 7. 10. 2014 Klima jako jeden z půdotvorných faktorů, dopady sucha Vítězslav Vlček, Jan Hladký, Eduard Pokorný, Martin Brtnický Mendelova univerzita v
Sucho a degradace půd v České republice - 2014 Brno 7. 10. 2014 Klima jako jeden z půdotvorných faktorů, dopady sucha Vítězslav Vlček, Jan Hladký, Eduard Pokorný, Martin Brtnický Mendelova univerzita v
Předmět: Hospodářská úprava lesů II
 Předmět: Hospodářská úprava lesů II Komplexní analýza přírodního prostředí 1.1 Přírodní prostředí a jeho složky Prostředí organizmu - vše co působí z okolního prostoru na organizmus faktory - abiotické
Předmět: Hospodářská úprava lesů II Komplexní analýza přírodního prostředí 1.1 Přírodní prostředí a jeho složky Prostředí organizmu - vše co působí z okolního prostoru na organizmus faktory - abiotické
Měření transpirace prostřednictvím transpiračního proudu a operačních struktur dřevin významných z hlediska vodního provozu
 Měření transpirace prostřednictvím transpiračního proudu a operačních struktur dřevin významných z hlediska vodního provozu J.Čermák, N.Naděždina Mendelova zemědělská a lesnická univerzita v Brně Transpirace
Měření transpirace prostřednictvím transpiračního proudu a operačních struktur dřevin významných z hlediska vodního provozu J.Čermák, N.Naděždina Mendelova zemědělská a lesnická univerzita v Brně Transpirace
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Posouzení zdravotního stavu a návrh péče o jilmy horské v obci Lazinov, k. ú. Lazinov.
 " Znalecký posudek č. 142/2009 Posouzení zdravotního stavu a návrh péče o jilmy horské v obci Lazinov, k. ú. Lazinov. I Zpracoval: Doc. Ing. Luboš Úradníček, CSc. Křížkovského 31 60300 Brno Znalecký posudek
" Znalecký posudek č. 142/2009 Posouzení zdravotního stavu a návrh péče o jilmy horské v obci Lazinov, k. ú. Lazinov. I Zpracoval: Doc. Ing. Luboš Úradníček, CSc. Křížkovského 31 60300 Brno Znalecký posudek
Dendrologický posudek
 Dendrologický posudek Datum: 10.9.2018 Objednatel posudku: starostka obce Hoříněves, pí. Kuthanová Posuzované dřeviny Posuzovanými dřevinami jsou lípy velkolisté (Tilia platyphyllos), rostoucí v centru
Dendrologický posudek Datum: 10.9.2018 Objednatel posudku: starostka obce Hoříněves, pí. Kuthanová Posuzované dřeviny Posuzovanými dřevinami jsou lípy velkolisté (Tilia platyphyllos), rostoucí v centru
Anotace: Materiál je určen k výuce přírodopisu v 7. ročníku ZŠ. Seznamuje žáky se základy obecné botaniky. Materiál je plně funkční pouze s použitím
 Anotace: Materiál je určen k výuce přírodopisu v 7. ročníku ZŠ. Seznamuje žáky se základy obecné botaniky. Materiál je plně funkční pouze s použitím internetu. kormus rinyofyty pletivo tkáň kořen stonek
Anotace: Materiál je určen k výuce přírodopisu v 7. ročníku ZŠ. Seznamuje žáky se základy obecné botaniky. Materiál je plně funkční pouze s použitím internetu. kormus rinyofyty pletivo tkáň kořen stonek
GLOBÁLNÍ OTEPLOVÁNÍ A JEHO DOPADY
 GLOBÁLNÍ OTEPLOVÁNÍ A JEHO DOPADY 2010 Ing. Andrea Sikorová, Ph.D. 1 Globální oteplování a jeho dopady V této kapitole se dozvíte: Co je to globální oteplování. Jak ovlivňují skleníkové plyny globální
GLOBÁLNÍ OTEPLOVÁNÍ A JEHO DOPADY 2010 Ing. Andrea Sikorová, Ph.D. 1 Globální oteplování a jeho dopady V této kapitole se dozvíte: Co je to globální oteplování. Jak ovlivňují skleníkové plyny globální
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce
 BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
BUNĚČNÁ STĚNA - struktura a role v rostlinné buňce Buněčná stěna O buněčné stěně: Buněčná stěna je nedílnou součástí každé rostlinné buňky a je jednou z charakteristických struktur odlišujících buňku rostlinnou
Řez stromů. David Hora, DiS. Předcertifikační školení certifikace. European Tree Worker. Evropský arborista
 Řez stromů David Hora, DiS. Předcertifikační školení certifikace European Tree Worker Evropský arborista Vztah: lidé stromy - lidé stromy dokáží žít bez nás my bez nich ne, doprovází nás celou historií
Řez stromů David Hora, DiS. Předcertifikační školení certifikace European Tree Worker Evropský arborista Vztah: lidé stromy - lidé stromy dokáží žít bez nás my bez nich ne, doprovází nás celou historií
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
OBSAH 1 ÚVOD... 7. 1.1 Výrobek a materiál... 7 1.2 Přehled a klasifikace materiálů pro výrobu... 8 2 ZDROJE DŘEVA... 13
 OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
Stomatální vodivost a transpirace
 Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Podmínky a zdroje. Michal Hejcman
 Podmínky a zdroje Michal Hejcman Úplná energetická bilance porostu Q N =I k +I d -I e -λ*e-h-p-f+r Q N je čistý příjem energie do porostu I k - iradiace(ozářenost) ve viditelném a UV spektru, v noci je
Podmínky a zdroje Michal Hejcman Úplná energetická bilance porostu Q N =I k +I d -I e -λ*e-h-p-f+r Q N je čistý příjem energie do porostu I k - iradiace(ozářenost) ve viditelném a UV spektru, v noci je
Úvod do biologie rostlin Úvod PŘEHLED UČIVA
 Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO. Modelová rostlina suchozemská semenná neukončený růst specializované části
 Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Možné dopady klimatické změny na dostupnost vodních zdrojů Jaroslav Rožnovský
 Český hydrometeorologický ústav, pobočka Brno Kroftova 43, 616 67 Brno e-mail:roznovsky@chmi.cz http://www.chmi.cz telefon: 541 421 020, 724 185 617 Možné dopady klimatické změny na dostupnost vodních
Český hydrometeorologický ústav, pobočka Brno Kroftova 43, 616 67 Brno e-mail:roznovsky@chmi.cz http://www.chmi.cz telefon: 541 421 020, 724 185 617 Možné dopady klimatické změny na dostupnost vodních
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11. *Ivana FELLNEROVÁ, PřF UP Olomouc*
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11 Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11 Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.)
") CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
Primární produkce. Vazba sluneční energie v porostech Fotosyntéza Respirace
 Primární produkce Vazba sluneční energie v porostech Fotosyntéza Respirace Nadzemní orgány procesy fotosyntetické Podzemní orgány funkce akumulátoru (z energetického hlediska) Nadzemní orgány mechanická
Primární produkce Vazba sluneční energie v porostech Fotosyntéza Respirace Nadzemní orgány procesy fotosyntetické Podzemní orgány funkce akumulátoru (z energetického hlediska) Nadzemní orgány mechanická
NÁVRH ZELENĚ. Obnova alejí na Ústředním hřbitově v Brně. Ing. Petr FÖRCHTGOTT
 NÁVRH ZELENĚ Obnova alejí na Ústředním hřbitově v Brně Ing. Petr FÖRCHTGOTT PROSINEC 2014 OBSAH Úvod Současný stav... str. 3 Rozsah a cíl návrhu... str. 3 Návrh Odstranění... str. 4 Výsadba... str. 5 Výkaz
NÁVRH ZELENĚ Obnova alejí na Ústředním hřbitově v Brně Ing. Petr FÖRCHTGOTT PROSINEC 2014 OBSAH Úvod Současný stav... str. 3 Rozsah a cíl návrhu... str. 3 Návrh Odstranění... str. 4 Výsadba... str. 5 Výkaz
Transport v rostlinách. Kateřina Schwarzerová Olga Votrubová
 Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
2. Použitá data, metoda nedostatkových objemů
 Největší hydrologická sucha 20. století The largest hydrological droughts in 20th century Příspěvek vymezuje a porovnává největší hydrologická sucha 20. století. Pro jejich vymezení byla použita metoda
Největší hydrologická sucha 20. století The largest hydrological droughts in 20th century Příspěvek vymezuje a porovnává největší hydrologická sucha 20. století. Pro jejich vymezení byla použita metoda
Vybrané technologie povrchových úprav. Základy vakuové techniky Doc. Ing. Karel Daďourek 2006
 Vybrané technologie povrchových úprav Základy vakuové techniky Doc. Ing. Karel Daďourek 2006 Střední rychlost plynů Rychlost molekuly v p = (2 k N A ) * (T/M 0 ), N A = 6. 10 23 molekul na mol (Avogadrova
Vybrané technologie povrchových úprav Základy vakuové techniky Doc. Ing. Karel Daďourek 2006 Střední rychlost plynů Rychlost molekuly v p = (2 k N A ) * (T/M 0 ), N A = 6. 10 23 molekul na mol (Avogadrova
Vliv zhoršeného zdravotního stavu smrkového porostu v důsledku globálních klimatických změn na reálný efekt celospolečenských funkcí lesa
 Vliv zhoršeného zdravotního stavu smrkového porostu v důsledku globálních klimatických změn na reálný efekt celospolečenských funkcí lesa Jiří Schneider Alice Melicharová Petr Kupec Jitka Fialová Ilja
Vliv zhoršeného zdravotního stavu smrkového porostu v důsledku globálních klimatických změn na reálný efekt celospolečenských funkcí lesa Jiří Schneider Alice Melicharová Petr Kupec Jitka Fialová Ilja
Disponibilní vodní zdroje a jejich zabezpečenost
 Adam Vizina (VÚV, ČZU), Martin Hanel (ČZU, VÚV), Radek Vlnas (ČHMÚ, VÚV) a kol. Disponibilní vodní zdroje a jejich zabezpečenost Výzkumný ústav vodohospodářský T. G. Masaryka veřejná výzkumná instituce,
Adam Vizina (VÚV, ČZU), Martin Hanel (ČZU, VÚV), Radek Vlnas (ČHMÚ, VÚV) a kol. Disponibilní vodní zdroje a jejich zabezpečenost Výzkumný ústav vodohospodářský T. G. Masaryka veřejná výzkumná instituce,