FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN
|
|
|
- Dana Irena Konečná
- před 6 lety
- Počet zobrazení:
Transkript

1 Univerzita J. E. Purkyně v Ústí nad Labem Přírodovědecká fakulta FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN KBI/P301, KBI/K301, KBI/P221 Milan Gryndler Lenka Němcová Ústí nad Labem 2013 (aktualizace 2017) Obor: biologie
2 Klíčová slova: Prokaryota, eukaryota, endosymbióza, systematika, morfologie, ekologie, fylogeneze, algologie, mykologie, lichenologie a bryologie. Anotace: Cílem kurzu je poskytnout základní informace o morfologii (stavba buňky a stélky), systematice, ekologii, historickém vývoji a významu organismů, které tradičně studují vědecké obory algologie, mykologie, lichenologie a bryologie. Důraz je kladen především na vývojové vztahy jednotlivých skupin organismů, na jejich roli v ekosystémech a na propojení s praktickou aplikací (biotechnologie, medicína, zemědělství, bioindikace apod.). Projekt Mezioborové vazby a podpora praxe v přírodovědných a technických studijních programech UJEP Registrační číslo projektu: CZ.1.07/2.2.00/ Tento projekt byl podpořen z Evropského sociálního fondu a státního rozpočtu České republiky. UJEP v Ústí nad Labem, 2013 (aktualizace 2017) Autor: Doc. RNDr. Milan Gryndler, CSc. RNDr. Lenka Němcová, CSc.
3 ÚVOD Karl von Linné (v latinském přepisu Carolus Linnaeus, , Species Plantarum 1753) dělil rostliny na tajnosnubné (nekvetoucí, Cryptogamia), nazývané též stélkaté (Thallobionta, Thallophyta, thallus je česky stélka), později též bezcévné, nebo (rozuměj vývojově) nižší, a jevnosnubné (kvetoucí, Phanerogamia), s tělem rozčleněným na kořen, stonek a listy (rozčleněné tělo se nazývá cormus, proto též Cormobionta, Cormophyta), též označované jako cévnaté nebo (rozuměj vývojově) vyšší (nauka o nich se nazývá též fanerogamologie). Bezcévné rostliny je společné označení pro sinice, řasy, houby, houbám podobné organismy, lišejníky a mechorosty (studuje je kryptogamologie). Nejedná se tedy o systematickou nebo vývojovou skupinu, ale pouze o souhrnné a nepřesné (nezahrnuje pouze rostliny - Plantae, ale zástupce několika dalších říší) označení několika skupin stélkatých organismů, jejichž vývoj je spojen s nejstarší historií naší planety. Uvedené skupiny byly ještě nedávno považovány za různorodou směsici s neurčitým postavením na začátku rostlinného či živočišného systému. Nové metody a obory (elektronová mikroskopie, molekulární systematika) prokázaly, jak složité jsou vývojové vztahy mezi těmito skupinami. Bezcévné rostliny studují vědecké obory algologie, mykologie, lichenologie a bryologie. Algologie (fykologie) se zabývá systematikou, ekologií a historickým vývojem jak autotrofních prokaryotických sinic (říše Bacteria, cyanologie), tak autotrofních eukaryotických zástupců říší Excavata, Rhizaria, Chromalveolata a Plantae. Řasy v původním pojetí jsou heterogenní polyfyletickou skupinou fotoautotrofních stélkatých organismů se schopností oxygenní fotosyntézy díky fotosyntetickým pigmentům. Mykologie studuje systematiku, ekologii a historický vývoj vlastních hub (Fungi z říše Opisthokonta) a houbám podobných heterotrofních zástupců říší Excavata, Rhizaria, Amoebozoa a Chromalveolata. Houby v širokém smyslu slova (houby s. l.) tvoří heterogenní polyfyletickou skupinu s heterotrofní (holozoickou nebo osmotrofní) výživou. Produktem metabolismu vlastních hub je polysacharid glykogen, což spolu s dalšími znaky na úrovni biochemické (syntéza lyzinu) a ultrastrukturální (lysozomy) houby (Fungi) přibližuje spíše k živočichům než k rostlinám, se kterými byly v minulosti tradičně spojovány. Stélka (thallus) je obecný název vegetativního těla jednobuněčných i mnohobuněčných organismů, které postrádá diferencovaná pletiva a zpravidla také chybí spojení mezi buňkami pomocí plazmodesmat. Stélka tedy není členěna na kořen, stonek a listy, ale funkční obdobu těchto orgánů tvoří rhizoidy, kauloid a fyloidy. U pokročilejších typů (např. chaluhy, mechy) dochází k diferenciaci plektenchymatických nebo pseudoparenchymatických pletiv a primitivních cévních svazků (např. u mechů hydroidy jsou analogické trachejím cévních svazků, leptoidy jsou analogické sítkovicím). Název stélka má historickou hodnotu a byl používán jako protiklad k rostlinnému tělu sporofytů suchozemských rostlin, nazývanému kormus. Co je úkolem systematiky? 1. Zjistit a správně interpretovat všechny dostupné informace o studovaných organismech. 2. Rozlišit jednotlivé skupiny organismů, utřídit je podle určitých hledisek a srozumitelně je označit (taxonomie a nomenklatorika). 3. A to nejdůležitější: na základě uvedeného se snažit o pochopení vývoje, pochopení fylogenetických vztahů, které by výsledné uspořádání (systém) mělo vyjadřovat. Co je vlastně systém? Systém je obecně výsledek lidské intelektuální činnosti, která směřuje k uspořádání příbuzných objektů, jevů či pojmů podle určitých zvolených principů. Biologický systém není seznam, ale vyjádření názoru na danou skupinu organismů na základě aktuálních 1
4 znalostí, a směřuje k poznání historického vývoje (fylogeneze) i současného stavu dotyčné skupiny (ke studiu: Systematická biologie pracuje se 2 hlavními typy systémů: se systémy hierarchickými a fylogenetickými (kladistickými). Hierarchické systémy jsou historicky starší. Základní jednotkou těchto systémů je druh. Snaha o třídění organismů (klasifikaci) je velmi stará (Platón, Aristoteles). Nutnost třídění byla vyvolána praktickými důvody - využití pro zajištění existence člověka (např. rostliny jedlé, jedovaté). Hromadění poznatků vedlo k potřebě podrobnějšího třídění organismů, nejdříve na základě morfologických znaků. Linné popsal všechny dosud známé živočichy a rostliny, popsal četné nové druhy. Jeho velkou zásluhou je zavedení binomické nomenklatury. Spis Species plantarum (1753) je závazným seznamem platných jmen některých řas, hub, mechů a všech cévnatých rostlin. Jeho systém byl umělý, založený pouze na morfologických znacích stavba květu, stavba stélky (např. vláknité druhy řas i hub zařadil do rodů Conferva a Byssus). Fylogenetické systémy jejich teoretickým základem je teorie o původu druhů, formulovaná Ch. Darwinem. Termín fylogeneze poprvé použil E. Haeckel a rozuměl tím historický vývoj druhů a celých skupin organismů. Své fylogenetické představy zakládal na srovnání ontogenetického vývoje, zejména srovnávací embryologie, morfologie a anatomie, značný význam přikládal paleontologickým nálezům. Vztahy mezi organismy vyjadřoval pomocí fylogenetických stromů, v nichž výchozí, vývojově jednodušší organismy, jsou předky vývojově dokonalejších organismů. Z hlediska fylogeneze lze rozlišit tři kategorie taxonů: 1. Taxony monofyletické zahrnují výlučného společného předka tohoto taxonu a všechny jeho potomky. 2. Taxony parafyletické zahrnují svého společného předka, ale ne všechny jeho potomky. Společný předek tedy není výlučný pro tento taxon, ale byl předkem i taxonu jiného. 3. Taxony polyfyletické nezahrnují svého společného předka, jsou jen seskupením podobných organismů bez vývojových souvislostí. Značný pokrok přinesla numerická taxonomie, která byla předpokladem pro vznik kladistiky. Základní taxon je větev (klad). Získané výsledky jsou graficky vyjádřeny pomocí rozvětvených diagramů (kladogramů, dendrogramů). Klad (clade) je skupina organismů, která se skládá ze společného předka a všech jeho přímých potomků, a představuje jednu "větev" na kladogramu, tzv. "stromě života". V současné době se fylogenetické studium stalo doménou molekulární taxonomie. 2
5 Jaký je rozdíl mezi moderními hierarchickými systémy a kladistikou? Pro obojí je společná snaha vytvořit systém organismů vyjadřující jejich historický vývoj a příbuzenské vztahy. Základní rozdíl spočívá v teorii a metodách. A co je to nomenklatorika? Pomocná disciplína systematické biologie, která se zabývá volbou, použitím a změnami vědeckých jmen taxonů všech kategorií. Vědecká jména jsou latinská (nebo latinizovaná) a mají mezinárodní platnost. Pravidla určující práci se jmény taxonů jsou přesně normována v mezinárodních nomenklatorických kódech. Existuje několik nomenklatorických kódů: Mezinárodní kód botanické nomenklatury - International Code of Botanical Nomenclature (dále MKBN), resp. International Code of Nomenclature for algae, fungi, and plants Mezinárodní kód zoologické nomenklatury (International Code of Zoological Nomenclature) Mezinárodní kód nomenklatury prokaryot (International Code of Nomenclature of Procaryota) Fylogenetický kód biologické nomenklatury (PhyloCode: A Phylogenetic Code of Biological Nomenclature) Zatímco vědecké pojmenování (nomenklatura) je latinské, platí jednotně na celém světě a řídí se mezinárodními pravidly, tzv. kódy, různé etnické skupiny mají vlastní národní názvosloví. V případě druhu je vědecké pojmenování binominální: rodové jméno + druhové epitetum a součástí jména je platná zkratka autora jména. Správné jméno spolu se správným taxonomickým hodnocením představuje ve formě vrcholné zkratky určitou základní charakteristiku taxonu. Např.: Amanita muscaria (L.) Pers. - muchomůrka červená (muchotrávka červená, Roter Fliegenpilz) Některé základní zásady MKBN: 1. Botanická nomenklatura nezávisí na nomenklatuře zoologické. 2. Použití jmen taxonomických skupin v hodnotě druhu až do hodnoty čeledi se řídí pomocí nomenklatorických typů. Nomenklatorický typ je prvek, s nímž je trvale spojeno jméno taxonu bez ohledu na to, zda se jedná o správné jméno nebo synonymum. Nomenklatorický typ jména druhu je jediný, trvale konzervovaný vzorek (herbářová položka, mikroskopický preparát, popř. publikovaná ilustrace). Nemůže to být pouze živá kultura, jako je tomu u bakterií. 3. Nomenklatura taxonů je založena na prioritě, tj. přednost při volbě platného jména mají vždy nejstarší platně publikovaná jména (pozn.: protože se stále objevují historické publikace, byly zavedeny tzv. výchozí publikace, např. autorů Linné, Persoon, Fries). 4. Každý taxon s přesným vymezením, zařazením a hodnotou může nést jediné správné jméno, a to nejstarší jméno, které je v souladu s pravidly Kódu. Ostatní platně publikovaná jména, která nemají prioritu nebo jsou z hlediska dnešního taxonomického pojetí již zastaralá, se nazývají synonyma. V dosahu působnosti daného kódu nesmí existovat stejná jména pro různé taxony (homonyma). Vědecká jména jsou latinská a důležitou součástí jména jsou autorské zkratky za jménem, které jméno jednoznačně určují. 5. Článek 59 - stanoví volbu jména hub s pleomorfickým životním cyklem. Tím se rozumí životní cyklus vřeckatých a stopkovýtrusých hub, ve kterém je zařazena anamorfa, nepohlavní forma, která se rozmnožuje nepohlavními výtrusy a teleomorfa, jejíž rozmnožování je provázeno pohlavním procesem. Správné jméno pro holomorfu (tj. jméno zahrnující anamorfu i teleomorfu) je nejstarší oprávněné jméno platné pro teleomorfu. Př. Venturia inaequalis - holomorfa a teleomorfa, Spilocaea pomi - pouze anamorfa. 3

6 Přehled základních systematických kategorií: taxonomická kategorie koncovka příklad DRUH (species) nemá Chlorella vulgaris, Boletus edulis ROD (genus) nemá Amanita, Volvox ČELEĎ (familia) -aceae Amanitaceae, Volvocaceae ŘÁD (ordo) -ales Agaricales, Volvocales TŘÍDA (classis) -phyceae Chlorophyceae (řasy) -mycetes Agaricomycetes (houby) -opsida Sphagnopsida (mechorosty a cévnaté r.) ODDĚLENÍ (divisio, phyllum) -phyta Chlorophyta (řasy, mechorosty, cévnaté r.) -mycota Basidiomycota (skupiny, které studuje mykologie) ŘÍŠE (regnum) nemá Chromalveolata, Fungi, Plantae NADŘÍŠE, (doména, imperium) nemá Archea, Prokarya, Eukarya Kolik říší živých organismů rozlišujeme? Odpověď souvisí s aktuální úrovní znalostí o fylogenezi živých organismů. Zatímco klasické systémy byly založeny na morfologii, později způsobu výživy a organelové výbavy buněk, současné systémy vycházejí z výsledků ultrastrukturálních studií a zejména molekulárně genetických analýz (ke studiu: Linné publikoval v díle Systema naturae (1735) systém 2 říší živých organismů (Regnum vegetabile a Regnum animalia). Jeho systém byl umělý, založený pouze na morfologických znacích, ale byl velmi stabilní a v obecném povědomí se udržel až do 50. let 20. stol. Již ve 2. pol. 19. stol. se však objevily první pokusy o úpravu Linného systému. Problematickou skupinou byly mikroskopické jednobuněčné organismy (bakterie a protista) 4
 následován Hoggem (1860), jenž ji pojmenoval Protoctista. O šest let později definoval Haeckel (1866) říši Protista.")
7 podobné buď rostlinám, nebo živočichům, ale lišící se od nich přinejmenším velikostí. Pokrok v poznání těchto organismů umožnil rozvoj mikroskopických technik. Jako první vyčlenil pro tyto organismy samostatnou říši Protozoa Owen (1858) následován Hoggem (1860), jenž ji pojmenoval Protoctista. O šest let později definoval Haeckel (1866) říši Protista. Pojmenování a vymezení skupiny se v průběhu let sice měnily, nicméně je zřejmé, že kromě dvou Linnéových říší existuje minimálně jedna další. 1. pol. 20. stol. přinesla 2 významné změny v systematice, jednak rozdělení organismů na prokaryota a eukaryota, jednak vyčlenění hub z říše rostlin (Whittaker 1969) (Fungi poprvé navrhl Fries 1821, dlouho odmítáno). Whittekerův systém je velmi srozumitelný, rozeznává jednu prokaryotickou (Monera) a čtyři eukaryotické říše (Protista, Plantae, Animalia, Fungi), jež definuje počtem buněk či jader v nich a převažujícím způsobem výživy (fotosyntéza u autotrofů a absorpce živin z prostředí příp. příjem potravy a její zpracování pomocí trávicí soustavy u heterotrofů). Poslední třetina 20. stol. přináší nové metodické přístupy (elektronová mikroskopie 70. léta a molekulární fylogenetika 80. a 90. léta), velký význam má formulace endosymbiotické teorie rozpracovaná Margulisovou (Margulis 1975). Nové poznatky uvedené do kontextu endosymbiotické teorie změnily pohled biologů především na skupinu jednobuněčných eukaryot, jejich diverzitu a postavení v rámci eukaryotického stromu života. 5
. http://www.microbe.")

8 Tzv. Univerzální systém organismů je založen na srovnání sekvencí nukleotidů v molekule DNA a ribozomální RNA prokaryotních a eukaryotních buněk, což vedlo k závěru, že organismy lze rozdělit do 3 velkých skupin: domén, čili impérií (Woese et al. 1990). Thomas Cavalier-Smith (*1942) je anglický evoluční biolog, který publikoval více než 160 prací týkajících se především klasifikace jednobuněčných eukaryot, vyšší systematiky eukaryot a vzniku eukaryotické buňky. Nejčastěji používaným je jeho systém z r se dvěma superříšemi Prokaryota a Eukaryota (neuznává Archea) a 5 eukaryotickými říšemi. Za zásadní je považováno vytyčení nové říše Chromista, již definoval dvěma znaky: jednak přítomností plastidu s chlorofylem c obaleného čtyřmi membránami (z nich vnější je kontinuální s ER), jednak přítomností mastigonemat (jemných vlásků ) na bičíku. 6

9 Zařazení probíraných skupin do systému Cavalier-Smith 1998 (viz Kalina, Váňa 2010): Imperium Prokarya, říše: Bacteria, oddělení Cyanobacteria. Imperium Eukarya, říše: Protozoa, oddělení Euglenophyta, Dinophyta, Chlorarachniophyta, Acrasiomycota, Myxomycota, Plasmodiophoromycota, říše: Chromista, oddělení Cryptophyta, Heterokontophyta, Haptophyta, Peronosporomycota, říše: Fungi, oddělení Chytridiomycota, Microsporidiomycota, Zygomycota, Ascomycota, Basidiomycota, říše: Plantae, oddělení Glaucophyta, Rhodophyta, Chlorophyta, Charophyta, Bryophyta, Anthocerotophyta, Marchantiophyta. Cavalier-Smithův systém nepracoval s monofyletickými taxony, problematická je hlavně říše Protozoa jako sběrná skupina pro jednobuněčná eukaryota, která se nedala zařadit do jiné říše. Molekulárně genetické studie a respektování požadavku na monofylii taxonů přineslo řadu koncepcí, např. systém Simpson a Roger (2004). Rozlišují 6 eukaryotických říší (superskupin), chybí říše Protozoa, zástupci zařazeni do jiných říší. Trojice tradičních říší Animalia, Fungi a Chromista se stala součástí říší nově ustavených. Říše Plantae je beze změny. 7

10 V roce 2012 byla zveřejněna revidovaná podoba konceptu superskupin (Adl et al. 2012). Stabilní pozici si udržely Opisthokonta a Amoebozoa a přibylo důkazů o monofylii superskupiny Archaeplastida. Rozpadla se superskupina Chromalveolata, čímž vzala za své Cavalier-Smithova chromalveolátní hypotéza. Ta předpokládala společný původ všech organismů nesoucích sekundární plastid vzniklý pohlcením ruduchy. Taxony Stramenopila a Alveolata vytvořily spolu s dřívější superskupinou Rhizaria novou superskupinu SAR a skupiny Haptophyta a Cryptophyta získaly status skupin s nejasnou systematickou pozicí (incertae sedis). Studie Derreleho et al. (2015) navíc zpochybnila monofylii superskupiny Excavata, uvnitř níž pravděpodobně leží kořen eukaryotického stromu (tj. počátek eukaryotické evoluce neboli napojení eukaryot na prokaryotické domény). V důsledku toho se Excavata nejeví jako přirozená skupina, ale jako základní evoluční stupeň, ze kterého ostatní eukaryotické skupiny vyšly. Je zřejmé, že další změny na úrovni superskupin lze očekávat. 8
11 ROZMNOŽOVÁNÍ ŘAS A HUB - ZÁKLADNÍ POJMY Rozmnožování - jedna ze základních vlastností organismů. Význam: udržování druhu, zvětšování počtu jedinců a šíření na nová území. Je důležitým evolučním činitelem, s průběhem rozmnožování úzce souvisí variabilita, mutace, přenos genetického materiálu. Nepohlavní rozmnožování - vznikají rozmnožovací buňky (nepohlavní výtrusy, spory), které se vyvíjejí ve výtrusnicích (sporangiích) nebo na koncích vegetativních vláken, kde dochází k přeměně vegetativních buněk ve spory. Spory mohou být pohyblivé: zoospory, planospory, či nepohyblivé: u řas autospory, aplanospory, u hub např. konidie. Vznikají při mitotickém dělení jádra mitospory: haplo- či diplo- (příklad: haplomitosporangiospory). Zoospory podle postavení bičíků rozlišujeme na pleurokontní (bičíky umístěné na boku buňky), akrokontní (bičíky v přední části) a opisthokontní (bičíky na zadním konci). Vegetativní rozmnožování je rovněž spojeno s mitózou, tvoří se jednobuněčné či vícebuněčné částice (části vláken, úlomky mycelia), které dorůstají do tvaru původní rostliny. V některých případech se na stélkách tvoří specializované rozmnožovací částice (gemy: parožnatky, mechorosty; soredie, isidie: lišejníky). Při pohlavním rozmnožování probíhá pohlavní proces (výměna genetické informace mezi jedinci), který začíná plazmogamií (splynutím cytoplazmy) dvou odlišně laděných individuí (gamet, specializovaných pohlavních orgánů nebo buněk vegetativní stélky) a pokračuje karyogamií (splynutím jader) za vzniku diploidního jádra, ve kterém ihned nebo po několika mitózách dochází k meiotickému dělení spojenému s rekombinací genetické informace (= podstata pohlavního rozmnožování). Výsledkem splynutí dvou odlišně laděných gamet může být zygota, ale proces může mít i jinou vnější formu, v níž splývají např. cytoplazmy pohlavních orgánů gametangií (viz dále). Zygota často utvoří tlustou stěnu - zygospora (hypnospora) a přetrvá tak i dlouhé klidové období. Pohlavní buňky (gamety) vznikají: buď přeměnou vegetativních buněk, nebo ve specializovaných buňkách (gametangiích). Jsou pohyblivé či nepohyblivé (planogamety a aplanogamety). Lze rozlišit druhy oboupohlavné (tj. obojí gametangia na 1 stélce, monoecické) a jednopohlavné (diecické). Dále druhy homothalické - na stélce vyrostlé z jednoho výtrusu vznikají gametangia obojího pohlaví, jejich jádra, ač geneticky stejná, se účastní plazmo- a karyogamie, heterothalické - stélky se rozpadají do dvou skupin kmenů, + a -. Mezi příslušníky stejného kmenu nemůže dojít k pohlavnímu procesu, i když jsou tam obojí gametangia, samčí a samičí. Pohlavní proces je bržděn homologickým párem genů inkompatibility. Podle toho, jaké struktury při průběhu pohlavního procesu splývají, rozlišujeme následující typy: gametogamie buď izogamie (splývají tvarově stejné pohlavní buňky, gamety), nebo anizogamie (splývají tvarově odlišné gamety), nebo oogamie (splývá velká nepohyblivá vaječná buňka oosféra s podstatně menším androgametem, což může být pohyblivý spermatozoid nebo nepohyblivá spermacie) gametangiogamie - splývání specializovaných pohlavních orgánů - gametangií - připojených ke stélce, rozlišujeme izogametangiogamii (gametangia jsou zhruba stejná) a anizogametangiogamii (gametangia se podstatně tvarově a velikostí liší) gameto-gametangiogamie (spermatizace) - splývá gametangium s volnou gametou somato-gametangiogamie - splývá gametangium s nediferencovanou somatickou buňkou stélky 9
 ŽIVOTNÍ CYKLY (ŽC) ŘAS A HUB ŽC dělíme podle toho, zda v nich je nebo není zařazen pohlavní proces, zda je vegetativní buňka haploidní nebo diploidní a zda je po splynutí cytoplazmy")
12 somatogamie - splynutí protoplastů vegetativních buněk, hyfogamie, hologamie (např. konjugace) ŽIVOTNÍ CYKLY (ŽC) ŘAS A HUB ŽC dělíme podle toho, zda v nich je nebo není zařazen pohlavní proces, zda je vegetativní buňka haploidní nebo diploidní a zda je po splynutí cytoplazmy (plazmogamii) v pohlavním procesu časově oddáleno splynutí jader (karyogamie). Chybí-li v ŽC pohlavní proces, má 4 fáze: růst, replikace DNA, RNA, mitotické dělení jádra, dělení protoplastu. Pohlavní proces vnáší do ŽC nový prvek, střídání jaderných fází. ŽC pak začíná zygotou a končí tvorbou gamet. Podle podílu haploidní a diploidní generace v životním cyklu rozlišujeme: 1. řasy a houby s převahou haploidní generace - vegetativní stélka je haploidní, zygota je jedinou diploidní buňkou, při jejím klíčení proběhne meioza a vyroste nová haploidní stélka (př. spájivky, parožnatky, mukorovité houby) Haplontní ŽC heterothalického druhu Rhizopus stolonifer (pohlavním procesem je gametangiogamie): 2. ŽC s převahou diploidní generace - ve vegetativním stavu jsou stélky diploidní, haploidní fáze omezena pouze na gamety (př. rozsivky) 10
: gamety (n) zygota (2n) diploidní sporofyt (S, 2n) meioza haploidní meiospory (n) gametofyt (G, n).")
13 Diplontní ŽC centrické rozsivky (pohlavním procesem je oogamie): 3. v ŽC navazuje generace haploidní na diploidní, tj. rodozměna (metageneze): gamety (n) zygota (2n) diploidní sporofyt (S, 2n) meioza haploidní meiospory (n) gametofyt (G, n). G a S jsou často samostatní jedinci, kteří se rozmnožují nepohlavními sporami, mohou být morfologicky podobní - izomorfická rodozměna (př. Ulva lactuca) nebo se G a S tvarově liší - heteromorfická rodozměna (př. mechorosty) Izomorfická rodozměna zelené řasy Ulva lactuca (pohlavním procesem je anizogamie): 11

, dikaryotická fáze je omezena na určitá vývojová stádia či celá vegetativní stélka je dikaryotická (př.")
14 Heteromorfická rodozměna chaluhy Laminaria digitata (pohlavním procesem je oogamie): 4. u některých hub je mezi haploidní a diploidní fázi vložena fáze dikaryotická (v buňce je dvojice pohlavně rozlišených jader + a -: dikaryon, mitóza probíhá u obou jader současně), dikaryotická fáze je omezena na určitá vývojová stádia či celá vegetativní stélka je dikaryotická (př. vřeckovýtrusé houby, stopkovýtrusé houby). Dikaryotický ŽC stopkovýtrusé houby (pohlavním procesem je somatogamie, resp. hyfogamie): 12
15 PŘEHLED HLAVNÍCH PROBÍRANÝCH JEDNOTEK (podle Simpson, Roger 2004 a Hibbett et al. 2007): 1. doména (nadříše): Archea (Archaebacteria) 2. doména (nadříše): Prokarya (Prokaryota) Eubacteria (bakterie) oddělení Cyanobacteria (Cyanophyta, Cyanoprokaryota, sinice) 3. doména (nadříše): Eukarya (Eukaryota) Excavata oddělení Acrasiomycota (akrasie, buněčné hlenky) oddělení Euglenophyta (krásnoočka, eugleny, Euglenozoa) Rhizaria oddělení Plasmodiophoromycota (nádorovky, plazmodiofory) oddělení Chlorarachniophyta (Chlorarachnea, Reticulophylosa) Amoebozoa oddělení Myxomycota (hlenky, Mycetozoa, Eumycetozoa, Gymnomycota) Chromalveolata oddělení Cryptophyta oddělení Dinophyta (obrněnky, Dinozoa, Dinoflagellata) oddělení Heterokontophyta oddělení Haptophyta oddělení Peronosporomycota Opisthokonta (incl. Fungi) oddělení Chytridiomycota oddělení Blastocladiomycota oddělení Neocallimastigomycota oddělení Microsporidiomycota oddělení Glomeromycota pododdělení Mucoromycotina pododdělení Entomophthoromycotina oddělení Ascomycota oddělení Basidiomycota Plantae oddělení Glaucophyta oddělení Rhodophyta oddělení Chlorophyta oddělení Charophyta oddělení Anthocerotophyta oddělení Marchantiophyta oddělení Bryophyta 13
16 DOMÉNA (NADŘÍŠE): ARCHEA Nepatří mezi nižší rostliny, ale jsou důležité v kontextu systematiky nejvyšších taxonomických jednotek Eukaryot a Bakterií. Jsou to extremofilní organismy, z nichž řada obývá prostředí s extrémní kyselostí, vysokou teplotou, vysokým tlakem a absencí kyslíku. Hypertermofilní archea žijí při teplotě vyšší než 100 C kolem vulkanických vývěrů na dně moří v hloubce asi 10 km pod hladinou. Termofilní druhy obývají termální prameny ve vulkanických oblastech, teplota kolem 90 C. Metanogenní a halofilní archea žijí v anaerobních bažinách, kde rozkladem organické hmoty vzniká metan, popř. v trávicím traktu skotu a termitů. Na základě vlastností, které považujeme za podstatné, nelze sestrojit spolehlivý fylogenetický strom, který by vymezoval vznik Archeí vývojem směrem od Bakterií nebo Eukaryot, nebo který by ukazoval na to, že jsou předky některé z těchto skupin. Někteří autoři odmítají oprávněnost impéria Archea a soubor znaků, který je charakterizuje, považují za sekundární adaptaci k vysokým teplotám a aciditě prostředí. Stáří Archeí není známo a odhady se u jednotlivých autorů velmi liší. Podle jedněch jsou Archea nejstaršími organismy, podle jiných jsou ale fosilní eubakterie staré asi 3,5 mld let, zatímco Archea a Eukaryota pouze 850 mil let. Je třeba vzít v úvahu, že Archea žijí v ekologických nikách, jejichž podmínky jsou podobné těm, které byly na Zemi před více než 3 mld let: anaerobní s vysokou teplotou, aciditou, koncentrací solí, metanu ( Vlastnosti podobné s Eukaryoty: DNA je organizovaná v nukleozómech, proteosyntetický aparát a transkripční aparát je podobný. Vlastnosti podobné s Bakteriemi: absence jádra, cytoskeletu a vnitřních membrán, přítomny rotující bičíky. Vlastnosti unikátní: glycerol je s mastnými kyselinami v membránových fosfolipidech spojen éterovou vazbou, nikoli esterovou jakou bakterií a eukaryot. Fosfolipidové mastné kyseliny jsou výrazně větvené. Je možné, že příčina neschopnosti jednoznačně odvodit postup vývoje 3 domén v podobě fylogenetického stromu spočívá v tom, že vývoj ve skutečnosti takto neprobíhal. Alternativním názorem je Woese-Kandlerova hypotéza, která předpokládá, že vývoj všech 3 domén začal nezávislou kombinací genetického materiálu získaného z prvotní nestabilní formy života. 14
17 ŘASY JAKO BIOLOGICKÁ SKUPINA Řasy (latinsky Algae, řecky Phykos) jsou včetně sinic zařazeny do 4, resp. 5 říší, přesto mají řadu podobných vlastností jako biotická skupina. Přehled řasových oddělení a jejich současné zařazení: Cavalier- Smith 1998 Simpson a Roger 2004 Impérium Říše Oddělení Říše Prokarya Bacteria Cyanobacteria Bacteria Eukarya Protozoa Euglenophyta Excavata Protozoa Dinophyta Chromalveolata Protozoa Chlorarachniophyta Rhizaria Chromista Cryptophyta Chromalveolata Chromista Heterokontophyta Chromalveolata Chromista Haptophyta Chromalveolata Plantae Glaucophyta Plantae Plantae Rhodophyta Plantae Plantae Chlorophyta Plantae Plantae Charophyta Plantae Řasy představují etapu ve fylogenezi autotrofních organismů, vázaných na vodní prostředí. Jsou to převážně fotoautotrofní organismy, jejichž oxygenní fotosyntéza (předchůdcem je bakteriální anoxygenní fotosyntéza) je založena na podobné funkci a struktuře fotosyntetického aparátu, přítomnosti a složení fotosyntetických pigmentů. Chlorofyly formy: a, b, c, d, e, f. Chlorofyl a Chlorofyl b Chlorofyl c1 Chlorofyl c2 Chlorofyl d Chlorofyl e Chlorofyl f všechny rostliny včetně mechů a řas a všechny sinice vyšší rostliny, zelené řasy, sinice a krásnoočka hnědé řasy hnědé řasy ruduchy, sinice některé různobrvky některé sinice Chlorofyl a je hlavním fotosyntetickým pigmentem. U mnoha sinic je jediným chlorofylem obsaženým v tylakoidech. U ostatních oddělení jsou chlorofyly 2, vzácně 3. Chlorofyly a+b jsou u euglen, chlorarachniofyt a všech zelených rostlin (Viridiplantae). Chlorofyly a+c jsou u fotoheterotrofních zástupců říše Chromalveolata. Chlorofyly a+d jsou u některých ruduch. Karotenoidy (karoteny a xantofyly) jsou součástí světlosběrné antény absorbující světelné záření v širší oblasti než chlorofyly, a tím zvyšují přísun fotonů pro fotosyntézu, chrání buňky před UV zářením, eliminují volné radikály. β-karoten je přítomen u všech řas, α a γ jen u některých. U řas se vyskytují některé specifické xantofyly (peridinin u obrněnek, fukoxantin u zlativek, rozsivek a hnědých řas). Fikobiliny vyskytují se u sinic, ruduch, glaukofyt, skrytěnek. 15
18 OBECNÉ ZÁKONITOSTI VE STAVBĚ ŘASOVÉ STÉLKY Eukaryotické řasy vykazují podobný vývoj buněk a stélek. Typy řasových stélek jsou považovány za organizační stupně ve vývoji řas. Organizačním stupňům bývá přisuzován význam v systému, zpravidla na úrovni řádu. Druhy stélek Monadoidní - jednobuněčná jednojaderná stélka bičíkovců, na apikálním konci vyrůstá 1 nebo více bičíků, na povrchu plazmatická membrána, periplast, pelikula nebo BS, v plazmě pulsující vakuoly (regulují osmotický tlak), v plastidu zpravidla stigma. Podobnou stavbu mají i nepohlavní zoospory a planogamety (izogamety a spermatozoidy). Rhizopodová - jednobuněčná, jedno- či mnohojaderná (plazmodium) měňavkovitá stélka, tvoří panožky (pseudopodia), které slouží k pohybu a k zachycování potravy (ve třídách Chrysophyceae, Xanthophyceae, oddělení Chlorarachniophyta jako samostatné druhy, jindy jako stádia životních cyklů). Kapsální (gloeomorfní) - jednojaderná, odvozená od monadoidní, někdy má pseudocilie (bičíky, které druhotně ztratily pohyblivost), na povrchu BS a vrstva slizu. Kapsální buňky v životním cyklu bičíkovců se označují jako palmelová stádia. Kokální - jednobuněčná, jednojaderná nebo mnohojaderná (cenoblastická) s pevnou BS. U rozsivek křemitá frustula, chlamys u některých zelenivek. Kokální jsou také zygoty a zygospory. Trichální mnohobuněčná, vláknitá, tvořená jednojadernými buňkami. Buňky v pravých vláknech spojeny plasmodesmaty, která prostupují jemnými otvory v příčných přehrádkách, vlákna nevětvená či větvená se stejnocennými větvemi. U zástupců řádů Microsporales a Oedogoniales se vyskytují nepravá vlákna = pseudofilamenta, jejich buněčné stěny jsou dvoudílné, plasmodesmata chybějí. Heterotrichální - odvozená od trichální, větvená, s funkčně i morfologicky rozlišenými větvemi (hlavní a postranní - Draparnaldia nebo plazivá a vzpřímená - Trentepohlia). Pseudoparenchymatická (pletivná) - odvozena od vláknité, někdy morfologická a funkční diferenciace na rhizoidy, kauloid a fyloidy, popř. funkční diferenciace pletiv Laminaria. Sifonální - vakovitá nebo vláknitá, jedna mnohojaderná makroskopická buňka (Botrydium, Vaucheria) nebo stélka tvořena hustě propletenými a funkčně rozlišenými vlákny sifonoblasty (Caulerpa). Sifonokladální - vláknitá či vakovitá, jednoduchá či větvená, sestává z četných mnohojaderných buněk (Cladophora). Stélka parožnatek mnohobuněčná vzpřímená stélka s přeslenitým uspořádáním větví. Vrcholová buňka se dělí a odčleňuje buňky nodální (uzlinové, z nich vyrůstají větve, gametangia a jejich obaly, korová vrstva) a internodální (článkové, které rostou do délky). V jednotlivých odděleních a třídách jsou organizační stupně zastoupeny v různé míře (Heterokontophyta, Chlorophyta x Cryptophyta, Bacillariophyceae). VÝSKYT SINIC A ŘAS Sinice a řasy zaujímají důležité místo v koloběhu látek a energie probíhajícím v přírodě. Jsou hlavními primárními producenty organické hmoty ve vodním prostředí, malá část druhů se adaptovala na život v terestrických podmínkách. Základní životní prostředí, v němž řasy prodělávaly svůj vývoj, bylo moře a kontinentální vody. Vodní prostředí, vyznačující se stabilitou řady vnějších faktorů, mělo vliv na utváření stélek i způsob rozmnožování řas. V biotopech tvoří 16
19 sinice a řasy druhově charakteristická společenstva, vykazující sezónní dynamiku (výskyt vázán na určité roční období). Vnitrozemské (kontinentální vody) a) Přehrady, rybníky, tůně, jezera, hlavní společenstva jsou: Fytoplankton (plankton - organismy volně se vznášející ve vodě aktivním pohybem nebo pomocí vodních proudů, s hustotou málo odlišnou od 1 a menší než 1 cm) - sinice a řasy osidlují osvětlené vodní vrstvy a sestupují až do hlubin s minimální intenzitou FAR, odpovídající 0,05-0,001 % povrchové intenzity. Rozlišovány jsou různé velikostní frakce buněk fytoplanktonu (velikostní rozmezí se u různých autorů liší) - mikroplankton μm v mořích, μm v kontinentálních vodách, - nanoplankton 2 20 μm v mořích, 2 50 μm v kontinentálních vodách, - pikoplankton (ultraplankton) 2 μm. Rozvoj nastává záhy zjara, jarní maximum je v květnu až červnu, přítomni jsou bičíkovci (skrytěnky, chrysomonády, zelení bičíkovci) a některé rozsivky, letní maximum je v červenci a srpnu, často se v této době objevuje vodní květ (společenstvo sinic, které se vznášejí u hladiny - Microcystis, Gomphosphaeria, Anabaena, Chroococcus, Aphanizomenon), který provází málo jiných řas. V letním období se ale vyskytují i kokální zelené řasy, které působí zelené vegetační zbarvení vody (Scenedesmus, Coelastrum, Oocystis, Pediastrum), též obrněnky (Ceratium, Peridinium) a zlativky (Dinobryon, Synura, Mallomonas). Perifyton - porůstá ponořené rostliny (epifytické řasy), vodní živočichy (epizoické řasy), kameny (epilitické řasy), bahno (epipelon - velmi bohatá společenstva hlavně rozsivek, ale též kulovité kolonie sinice Gloeotrichia či vláknité spájivky, Cladophora). Metafyton - pobřežní společenstvo rybníků a tůní, kde rostou planktonní řasy se shluky vláknitých řas a vyššími rostlinami. Bentos - na povrchu dna, rozsivky, sinice, parožnatky a část ročního období spájivky. b) Rašelinné tůně (ph 4,5-6,5) - převažuje bentos a perifyton, charakteristickou skupinou jsou krásivky (Closterium, Micrasterias...), rozsivky a krásnoočka (Euglena mutabilis). c) Kaluže, bazény, jímky - na hladině nádrží chráněných před větrem se tvoří neustonická blanka z pomalu se pohybujících krásnooček (Euglena viridis), i v silně znečištěných vodách. Též některé sinice a zelení bičíkovci působící vegetační zabarvení, na stěnách akinety bičíkovce Haematococcus. d) Proudící voda - organismy reagují různě na proudění vody (rheoxenní pohyb vody nesnášejí, rheofilní klidně tekoucí vody,15-70 cm/s, rheobiontní rychle tekoucí vody, cm/s, torentikolní bystřiny a peřeje), v rychle proudící vodě chybí fytoplankton, omezen prouděním je i perifyton. Charakteristický je výskyt Hydrurus foetidus, Hildebrandia, Lemanea. Střední a dolní toky potoků a řek: malé proudění vody, většinou značné znečištění - Cladophora, rozsivky. e) Vodní biotopy s extrémními podmínkami - termální prameny ( C) - mnoho rozpustných minerálních solí, nejasný je mechanismus, jak zde žijící organismy zabraňují koagulaci bílkovin, hlavně sinice, některé rozsivky a jednobuněčné ruduchy. Mořské biotopy Obsah soli (převážně NaCl) - salinita - se stanoví jako hmotnostní obsah NaCl v 1 l vody, normální je promile, tj. 3-3,3 g/l. Hypertonické prostředí vyrovnávají řasy vysokou osmotickou hodnotou cytosolu. 17

20 Společenstva makrofytických řas (chaluh, ruduch a zelených řas) osidlují hlavně skalnatá pobřeží a vykazují výraznou vertikální zonaci, určovanou světelnými podmínkami /Figure-1-Vertical-distribution-of-the-major-seaweeds-along-the-eulittoral-and.png Litorál - je pásmo, jehož klimatické podmínky jsou silně ovlivněny blízkým oceánem či mořem, a kde v důsledku toho často žijí specifické litorální organismy: epilitorál, není omýván vodou, supralitorál, oblast příboje dostřiková zóna, eulitorál trvale omývaný vodou - přílivová zóna, vystaven mechanickému působení vln a rychle se měnícím teplotním podmínkám, na pobřeží s výrazným odlivem (Bílé moře) též periodickému obnažování, infralitorál (sublitorál) je i při velkých odlivech pod vodou, spodní hranice sublitorálu se určuje podle spodní hranice růstu řas, tj 0,05 % intenzity světla, dopadajícího na hladinu. Mořský fytoplankton je tvořen hlavně rozsivkami, obrněnkami, sinicemi a vápenatými bičíkovci. Druhové zastoupení je charakteristické pro jednotlivé oceány, moře, dokonce i proudy a je dáno především teplotou vody (eury- a stenotermní řasy). Nejproduktivnějšími oblastmi jsou pobřeží (red tides, white water). Terestrické biotopy Společným znakem je závislost růstové rychlosti, průběhu buněčných a životních cyklů na vlhkosti prostředí, která je hlavním limitujícím faktorem. a) Půdní sinice a řasy - osidlují povrchovou vrstvu půdy do cm, jsou důležitou složkou edafonu: zelené řasy (Chlorella, Chlorococcum, Trebouxia), různobrvky (Botrydium granulatum), rozsivky a sinice (vazači vzdušného N 2 ). Význam: obohacují půdu o organické částice, tvorbou slizu ovlivňují strukturu půdy, obohacují ji o kyslík. b) Aerofytické sinice a řasy (petrofytické a epifytické) Snášejí dlouhá období vyschnutí. Rostou na povrchu skal (sinice Stigonema - inkoustové skvrny, Trentepohlia aurea - červené povlaky), na kmenech stromů (Apatococcus tvoří zelené povlaky s dalšími druhy), na vlhkých pařezech, mezi mechy, na vytrvalých plodnicích hub sinice, rozsivky i zelené řasy. Aeroplankton - převážně klidová stádia sinic a řas, která jsou větrem unášena na velké vzdálenosti. 18
.")
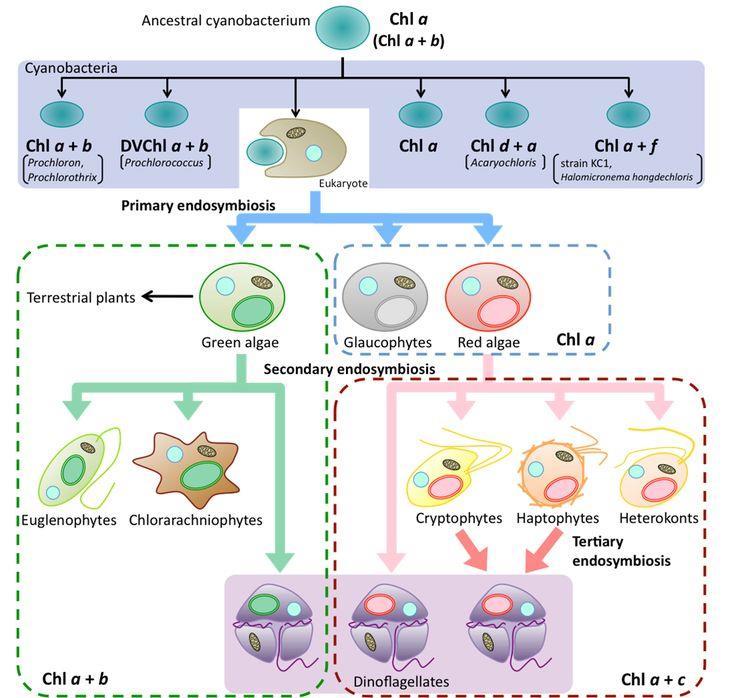
21 c) Kryofyta (kryoseston ) - řasy na sněhu - mají krátké vegetační období, vyskytují se v horách na tajících sněhových polích (Chlamydomonas nivalis). Jsou adaptovány na nízké teploty, na rychlé přechody ze zmrzlého do vegetačního stavu, ale i na silné záření (v buňkách ochranné pigmenty). SYMBIOTICKÉ VZTAHY SINIC A ŘAS a) Lichenismus fotobiont a mykobiont vzájemná kontrola obou složek. b) Endofyti v mezibuněčných prostorách některých vodních rostlin (Ceratophyllum, Lemna), játrovek, nahosemenných (Cykas), kapradin (Azolla) byly zjištěny jako endosymbionti zelené řasy, běžně žijí řasy v hyalocystách rašeliníků, autonomní autotrofní orgány i u některých bezobratlých (cyanely sinice, zoochlorely - zelené řasy, zooxantely - obrněnky, kryptomonády). c) Nejdůležitější úrovní symbióz jsou vnitrobuněčné symbiózy (endosymbiózy). Jejich původ sahá k počátku vývoje eukaryotických buněk. Dlouhá souběžná evoluce autotrofního symbionta a heterotrofní hostitelské buňky vedla ke ztrátě samostatnosti symbionta. Došlo ke značné redukci genomu endosymbionta. Mnohé z chybějících genů se objevily v jaderné DNA původně hostitelské buňky, jiné se zcela ztratily. Přestože byl původ plastidů z předků sinic potvrzen, zůstávají jisté nejasnosti: Jak se předkové chloroplastů dostali do buněk ruduch a chlorofyt, když nemají fagocytózu, která je důležitým předpokladem endosymbiotických asociací? Proběhla endosymbióza v evoluci opakovaně? Fylogenomické studie prokazují, že všechny plastidy recentních druhů mají stejný primární původ v jediné endosymbióze sinice (jedinou známou výjimkou je kořenonožec Paulinella chromatophora), a to z příbuzenstva recentních sinic rodu Pseudanabaena. Primární endosymbióza endosymbiontem byla sinice glaukofyta, ruduchy, zelené řasy (2 membrány na povrchu plastidu). Glaucophyta mají plastidy silně připomínající sinice, tzv. cyanely, ty jsou ztraceným článkem mezi sinicemi a plastidy. Sekundární endosymbióza endosymbiontem byla eukaryotní buňka ruduch či zelených řas - skrytěnky, chlorarachniofyta, eugleny, heterokontophyta (3-4 membrány na povrchu plastidu, spojení s jadernou blánou pomocí cysterny ER existuje u heterokontofyt, v prostoru mezi membránami u primitivních skupin nalezen nukleomorf, skrytěnky, chlorarachniophyta, tj. značně redukované jádro endosymbionta). V další evoluci došlo ke ztrátě nukleomorfu, zřejmě poté, co všechny potřebné plastidové geny byly přeneseny do jádra. Sekundární symbióza proběhla v evoluci opakovaně, což vysvětluje rozmanitost plastidů i rozmanitost řas. Terciální endosymbióza původní plastid byl nahrazen jiným endosymbiontem, např. skrytěnkou (mnohé obrněnky). 19

22 20
23 ÚLOHA ŘAS V BIOGEOCHEMICKÉM KOLOBĚHU LÁTEK Studium geologie, paleoklimatologie, plynů v ledovcových dutinách a dálkový průzkum Země poskytuje poznatky o změnách klimatu, horotvorných procesech a koloběhu látek, na nichž se podílely řasy: - vznik kyslíkové atmosféry, - koloběh C, tvorba vápenců a fosilních paliv (koloidní uhlík), - tvorba křemičitanových sedimentů (rozsivky), - koloběh S. ŘASY JAKO PRODUCENTI LÁTEK NEGATIVNĚ OVLIVŇUJÍCÍCH PŘÍRODNÍ PROSTŘEDÍ Sinice a řasy často zvýrazňují negativní vlivy zhoršujícího se životního prostředí. Narůstající eutrofizace moří a pevninských vod umožňuje vysokou produktivitu sinic a řas (vodní květy, vegetační zákaly, nárosty na přehradních hrázích, památkách narušují povrch kyselinou uhličitou, vznikající při respiraci), tím se kvalita vody dále zhoršuje, neboť je ovlivňována látkami uvolňovanými z živých (toxiny), ale zejména z odumírajících buněk. Invazivní mořské druhy znehodnocují pláže, porůstají trupy lodí (zelená řasa Caulerpa, Codium, některé ruduchy). Chaluhy produkují organobromidy a DMS, způsobující kyselé deště a redukci ozonosféry. skupina biotop společenstvo působení sinice kontinentální vody, moře fytoplankton, vodní květy cyanotoxiny obrněnky moře fytoplankton, "red tide" toxiny zlativky sladké vody fytoplankton zápach pitné vody po rybím tuku chaluhy moře bentos litorálu, sublitorálu produkce organobromidů a DMS kyselé deště, redukce ozonosféry chloromonády moře fytoplankton, "red tide" toxické pro ryby haptofyta moře, brakické toxické pro bezobratlé i ryby, produkce fytoplankton vody DMS kyselé deště zelené řasy moře bentos litorálu invazní druhy, znehodnocení pláží VYUŽITÍ ŘAS V BIOTECHNOLOGIÍCH Využívány již dlouhou dobu v různých oblastech lidské činnosti. Dálný východ sinice i řasy jsou tradiční součástí pokrmů (Porphyra pěstována od r. 1670), též v Británii, J a S Americe. Zejména v severských oblastech slouží jako krmivo, topivo, surovina pro výrobu potaše, sody, jódu, hnojiva. Alginizace rýžových polí sinice z řádu Nostocales vážou vzdušný dusík Velkoobjemové kultury mikroskopických řas využití pro výživu lidí (nepříznivý je vysoký obsah NK a některých alergizujících látek) a potravu mořských měkkýšů, kteří jsou následně součástí lidské stravy. Extrakce látek z biomasy řas: agar, karagen, algináty, fykocyanin, β-karoten. Botryococcus braunii nebo rozsivky - potenciální zdroj uhlovodíků příprava látky podobné ropě. 21
24 HOUBY A JIM PODOBNÉ ORGANISMY JAKO BIOLOGICKÁ SKUPINA Mykologie se věnuje studiu vlastních hub (říše Opisthokonta, incl. Fungi), ale i některých skupin z říší Excavata, Rhizaria, Amoebozoa a Chromalveolata, které bývají označovány jako Pseudofungi (Pseudomycota). V širším slova smyslu je termín houby používán pro heterogenní polyfyletickou skupinu eukaryotních, primárně heterotrofních stélkatých organismů s osmotrofní výživou (výjimkou jsou lišejníky díky přítomnosti fotobionta). Produktem metabolismu je glykogen, v buňkách jsou lysozomy, i některé metabolické pochody v buňkách hub probíhají spíše podobně jako v živočišných, než v rostlinných buňkách. Houby jsou destruenty, výživou jsou přímo či nepřímo odkázané na autotrofy. Získávají živiny jako paraziti, popř. hyperparaziti, a to obligátní nebo fakultativní, saprofyti nebo symbionti (mykorhiza, lichenismus). Další znaky, kterými lze houby odlišit od řas či rostlin, neplatí absolutně pro všechny skupiny organismů řazené k houbám: chitinová BS, stélka tvořená vlákny (hyfami), která mohou tvořit podhoubí (mycelium), přítomnost dikaryofáze v ŽC, tvorba plodnic, posteriorní (opistokontní) postavení bičíků. Vznik a vývoj houbových oddělení není dosud zcela objasněn. Dřívější fylogenetické spekulace byly založeny převážně na morfologických podobnostech, jako např. Sachsova teorie o společném původu ruduch a vřeckovýtrusných hub z r Současné teorie vycházejí z fyziologických, biochemických a molekulárně-biologických charakteristik. Z vývojového hlediska představují houby a jim podobné organismy fylogeneticky vzdálené a nepříbuzné skupiny. Hlenky jsou polyfyletickou skupinou, která se vyvinula pravděpodobně z heterotrofních amébovitých prvoků (Rhizopoda). Plasmodiophoromycota má rovněž společný původ s některými prvoky (Proteomyxa) a jejich morfologická podobnost s hlenkami je spíše otázkou konvergentního vývoje. Oddělení Peronosporomycota (celulózní BS, stavba bičíků, syntéza lysinu přes kyselinu diaminopimelovou) jsou pravděpodobně heterotrofní větví pravěkých organismů se sifonální stélkou, paralelní autotrofní větev je představována rodem Vaucheria (Tribophyceae, Heterokontophyta). Oddělení Chytridiomycota je obecně považováno za výchozí vývojovou skupinu dnešních vlastních hub (Fungi). Oddělení Ascomycota a Basidiomycota jsou odvozována od řádu Glomerales (Glomeromycota). 22
25 DOMÉNA (NADŘÍŠE): PROKARYA ŘÍŠE: BACTERIA Impérium s jedinou říší bakterie (Bacteria) obsahuje jednobuněčné nebo vláknité organismy s jednoduchou stavbou buňky. Chybí jádro a cytoskelet, interní membrány se vyskytují jen u některých skupin (tylakoidy). Protoplast pokrývá buněčná stěna na základě barvení podle Grama se dělí bakterie podle stavby BS na grampozitivní (na povrchu protoplastu 1 plazmatická membrána a silná BS) a gramnegativní (2 PM a mezi nimi pevná a pružná BS, barvivo se etanolem vypláchne). DNA v podobě kruhové molekuly (nukleoid, nukleoplazmatická oblast) a v podobě plazmidů. Mezi prokaryoty mají svůj původ organismy, které se staly donory plastidů a mitochondrií eukaryotických buněk. Purpurové bakterie a chlorobakterie mají jednoduchý fotosyntetický aparát založený na bakteriochlorofylu uloženém v plazmatické membráně, donorem elektronů při fotosyntéze, která probíhá v anaerobním prostředí, je sirovodík. Anoxygenní fotosyntéza purpurových bakterií je považována za předchůdce oxygenní fotosyntézy. Oddělení CYANOPHYTA (CYANOBACTERIA) SINICE Český název sinice pochází z termínu sinný = modrý, to je v podstatě překlad latinského názvu, z řeckého cyanos = modrý. Sinice jsou výjimečnou skupinou bakterií, obsahují chlorofyl ve specializovaných membránách, tylakoidech, na jejichž povrchu jsou umístěny fykobilizomy, světlosběrné antény, které umožňují optimální využití světla. Jsou funkční oba fotosystémy, PS II je spojen s fotolýzou vody, kde jako vedlejší produkt je uvolňován kyslík. Oxygenní fotosyntéza se později rozšířila na veškeré fotoautotrofní organismy a zásadním způsobem ovlivnila podmínky na Zemi. Sinice byly na počátku vývoje, vedoucímu ke vzniku kyslíkové atmosféry. Sinice také byly hlavními donory chloroplastů pro heterotrofní eukaryotní hostitele, kteří dali vzniknout všem autotrofním eukaryotům. Sinice jsou autotrofní prokaryotické organismy s fotosyntézou rostlinného typu (oxygenní fototrofní baktérie) s jednobuněčnou nebo vláknitou stélkou, žijí jednotlivě nebo v koloniích. V jejich buňkách chybí jádro, chloroplasty a mitochondrie, nemají cytoskelet ani bičíky. DNA je obsažena v nukleoplazmatické oblasti (centroplazmě), podobné nukleoidu bakterií a v plazmidech. Četné sinice mají schopnost fixovat plynný dusík a redukovat jej na amonné soli. Rozmnožují se dělením buněk a fragmentací vláken. Pohlavní proces (rekombinace a transformace) znám u jednobuněčných druhů. Jsou všeobecně rozšířené ve vodním prostředí, žijí i v biotopech s extrémní teplotou, salinitou či ph. Ochotně vstupují do symbiotických interakcí. Jsou nejstaršími fotoautotrofy na Zemi, paleontologické nálezy jsou staré až 3,2 mld let. Stavba buňky Buněčná stěna - sinice patří mezi gramnegativní baktérie. BS je vícevrstevná, vnější povrch pokrývá vrstva slizu, často vrstevnatého či zbarveného (z lipopolysacharidů, tzv. glykokalyx, má fibrilární strukturu), někdy tvoří nápadný obal pochvu. Další složkou je dvojice lipoproteinových membrán (vnější a vnitřní membrána), mezi nimi je pevná vrstva z peptidoglykanů, hlavní složkou je murein. Je citlivá k penicilinu, odbourá ji lysozym. Kontraktilní bílkovinná filamenta v blízkosti mureinové vrstvy BS umožňují spolu se slizem pohyb sinic. 23
 membrána obklopuje protoplast, zajišťuje aktivní transport iontů a exkreci látek, zahajuje oddělování dceřiných protoplastů při buněčném dělení, je sídlem dýchacích enzymů a")
. Histoproteiny chybějí. Ribozómy sinic mají sedimentační konstantu 70 S. Tylakoidy (fotosyntetické membrány) mají tvar plochých měchýřků.")
26 Plazmatická (vnitřní) membrána obklopuje protoplast, zajišťuje aktivní transport iontů a exkreci látek, zahajuje oddělování dceřiných protoplastů při buněčném dělení, je sídlem dýchacích enzymů a souvisí s ní vývoj tylakoidů. Protoplast - v živé buňce lze rozlišit periferní chromatoplazmu s tylakoidy a centroplazmu (nukleoplazmatickou oblast). Je zde umístěn jediný kompaktní genofor (kruhová molekula DNA). Histoproteiny chybějí. Ribozómy sinic mají sedimentační konstantu 70 S. Tylakoidy (fotosyntetické membrány) mají tvar plochých měchýřků. Fotosyntetické pigmenty chlorofyl a, a+b, a+c, a+d, ß-karoten a několik xantofylů jsou uloženy v membráně tylakoidů. Zjištění všech typů chlorofylů u sinic je fylogeneticky významné. Fykobiliny (modrý c-fykocyanin a červený c-fykoerytrin a modrý allofykocyanin) jsou umístěny ve fykobilisomech na povrchu tylakoidů a tvoří světlosběrné antény. 24
 chybějí tylakoidy, fikobilizomy sedí na povrchu plazmatické membrány")
27 Poté, co zástupci oddělení Prochlorophyta byli zařazeni mezi sinice, rozlišují se 3 typy fotosyntetického aparátu: 1) chybějí tylakoidy, fikobilizomy sedí na povrchu plazmatické membrány primitivní stav, příbuznost s purpurovými a chloro- bakteriemi, 2) četné tylakoidy s fikobilizomy, 3) dřívější oddělení Prochlorophyta - fykobilizomy chybějí, pigmenty (chlorofyl a+b) uložené v tylakoidech, tylakoidy srostlé po 2 4. Poměr červeného a modrého pigmentu určuje výslednou barvu sinicové buňky (zelená, modrá, šedá či červená). Barva je sice pro určité druhy charakteristická, ale mají schopnost chromatické adaptace (změna zastoupení jednotlivých typů fykobilinů), která je významná především v podmínkách přezáření, nedostatku světla nebo při častých změnách intenzity světla, umožňuje sinicím žít při světelných poměrech, které nesnesou ostatní fotosyntetizující organismy. S fotosyntézou souvisí přítomnost karboxyzómů, submikroskopických tělísek tvaru mnohostěnu. Obsahují enzym ribuloso-1,5-bifosfát karboxylázu/oxidázu (RuBisCo), který zajišťuje fixaci CO 2 v Calvinově cyklu. Analogickou strukturou u eukaryotických řas je pyrenoid, u druhů bez pyrenoidu sídlí RuBisCo v chloroplastech nebo cytosolu. Hlavní zásobní látka je sinicový škrob, svými vlastnostmi je podobný glykogenu hub a živočichů. Zvláštností sinic je uchovávat polypeptidy - cyanofycinová zrnka - jako zásobní energetické zdroje (polymer kys. asparagové a argininu), slouží především jako zásoba dusíku. Zásobní význam mají též polyfosfátové granule volutin, využívána v době nedostatku fosforečnanů (např. přezimování). Některé sinice produkují polysacharid podobný škrobu. Výraznou strukturou planktonních sinic jsou plynové vezikuly, měchýřky válcovitého tvaru, agregované v rozsáhlých aerotopech, dříve nesprávně nazývaných plynné vakuoly. Jejich stěna je propustná pro plyny rozpuštěné ve vodě. Jsou jedinou známou plynem zaplněnou strukturou v živých buňkách, jsou velmi stabilní, snižují specifickou hmotnost buněk a umožňují jim vznášet se ve vodě. Sinice je mohou tvořit a dezorganizovat v závislosti na abiotických faktorech prostředí a tím regulovat svoji polohu ve vodním sloupci Specializace buněk a fixace dusíku Kromě vegetativních buněk se u vláknitých sinic vyskytují heterocyty a artrospory. Při fixaci dusíku se střetávají dva zdánlivě neslučitelné procesy: fixace N a fotosyntéza, která vede k uvolňování O 2, ten je toxický pro nitrogenázu. Proto se u sinic vyvinuly strategie prostorové a časové separace fotosyntézy a fixace N 2. Ke studiu: 25

28 Heterocyty u některých vláknitých sinic, tlustostěnné buňky, vznikající z vegetativních buněk především v podmínkách dusíkového hladovění. V nich probíhá fixace plynného dusíku a pomocí nitrogenázy redukce na amonné sloučeniny. V heterocytech jsou vytvářeny anaerobní podmínky, aby nedošlo k inaktivaci nitrogenázy. Diazocyty např. Trichodesmium, primitivní diferenciace buněk, která není patrná v optickém mikroskopu a odehrává se výhradně na biochemické úrovni. Časová separace u několika rodů kokálních i vláknitých sinic, den x noc. Artrospory (akinety) vznikají u sinic z jedné nebo splynutím více vegetativních buněk, mají tlustou stěnu, bohatou zásobu živin. Mají význam pro přežití nepříznivých podmínek, po určité době z nich vyklíčí nová vlákna. - upraveno Cyanotoxiny Biologicky aktivní sekundární metabolity: neurotoxiny, anatoxiny a hepatotoxiny působí smrtelné otravy, cytotoxiny smrtelné otravy nepůsobí. Jsou méně jedovaté než bakteriální botulin nebo toxin tetanu, ale jsou srovnatelné s hadími jedy (kobra). Jsou toxičtější než rostlinné alkaloidy (kurare, strychnin). Následkem kontaktu s vodním květem jsou kožní alergie, záněty spojivek, atp. K otravám dochází u skotu napájeného kontaminovanou vodou. Působí komplikace při výrobě pitné vody nutné nákladné technologie. Některé mohou být využity jako cytotoxická antibiotika s protinádorovými účinky. Ke studiu: více o toxinech u sinic na Dělení buněk, větvení vláken, rozmnožování Dělení buněk sinic - prostým zaškrcením od obvodu ke středu buňky. Rovinu dělení určuje kruhové vchlípení plazmatické membrány, souběžně vrůstá mezi dceřiné protoplasty BS, tak vzniká příčná přehrádka. Slizová pochva se dělení neúčastní. Dělení na 2 dceřiné buňky, popř. ve 2 až 3 rovinách kolonie. Vláknité stélky jednoduché nebo větvené. Nerozvětvená řada vegetativních buněk, spojených plazmatickými vlákny tvoří trichom, je-li trichom uložen ve slizové pochvě, celek tvoří vlákno. Nepravé a pravé větvení. Rozmnožování jednobuněčné sinice - dělením buněk a fragmentací kolonií. Vláknité pomocí hormogonií - několikabuněčných částí mateřského vlákna. Chamaesiphon se rozmnožuje exocyty (exosporami), Chroococcidiopsis pomocí beocytů (uvolnění po rozplynutí mateřské BS). Pohlavní proces transformace přenos plazmidu z donora do recipienta, prokázán pouze laboratorně, ale je zřejmě rozšířen v přírodě (vysvětlení morfologické a fyziologické variability sinic). Variabilita sinic Výrazným znakem sinic je fenoplasticita, proměnlivost znaků vlivem vnějších podmínek, má adaptační charakter. Neobyčejná přizpůsobivost prostředí je výsledkem účinných opravných 26
29 mechanismů DNA a genového transferu. Výsledkem je, že současné sinice jsou geneticky odlišné od morfologicky podobných sinic fosilních. Sinice mají zvláštní typ vývoje, nazývaný stagnující evoluce. Ekologie sinic Jsou všudypřítomné: sladké vody i moře, povrchové vrstvy půdy i vlhké skály, endoliti, často vstupují do mutualistických vztahů s jinými organismy. Vodní květ vzniká při přemnožení planktonních sinic ve vodách s nadbytkem dusičnanových a fosforečnanových živin. V našich rybnících se vodní květ vyskytuje nejčastěji v létě, často tvoří mohutnou biomasu, např. Microcystis, Aphanizomenon, Anabaena. Pobyt u hladiny umožňují aerotopy. Problémy působí hlavně ve vodárenských nádržích cyanotoxiny, hnilobné procesy při rozkladu biomasy vyčerpání kyslíku, další produkce toxických látek. Populace sinic kontrolují cyanofágy, pokusy s umělým infikováním vodního květu zatím nebyly efektivní. Rudé moře má jméno dle vodního květu červené sinice Trichodesmium erythreum. Díky výrazným adaptačním mechanismům, souvisejícím s geologickým stářím sinic, osidlují sinice i stanoviště s extrémními podmínkami. Chromatická adaptace umožňuje optimálně využívat zelenou a červenou část spektra, a to změnou poměru fykocyaninu a fykoerytrinu ve fykobilizómech. I v anaerobních podmínkách, např. na dně vodních nádrží, probíhá fotosyntéza, při níž jako alternativní zdroj elektronů slouží sirovodík (anoxygenní fotoautotrofie). Termální sinice - v horkých pramenech s teplotou vody vyšší než 70 C. Termální vody bývají i silně mineralizované, při fotosyntéze sinic se sníží koncentrace oxidu uhličitého a rozpuštěné soli se vysrážejí v podobě minerálů, např. travertinu, tato činnost se nazývá biomineralizace. Sinice žijí též v prostředí s extrémními hodnotami ph (od 0,05 po 13,5), snášejí prudké změny teplot (termální sinice snesou zmrazení na -180 C) i rychlou ztrátu vody (po 100 letech v herbáři klíčivé spory). Žijí i v polárních oblastech, kde není konkurence jiných autotrofů, mezi krystaly ledu (Grónsko), na dně antarktických jezer povlaky sinic až 90 cm vysoké (nejmohutnější, jaké byly na Zemi nalezeny). Má se za to, že neobyčejná přizpůsobivost prostředí je umožněna přenosem genetické informace pomocí plazmidů a cyanofágů. Ochrana proti UV záření zastíněná místa, endolitické druhy, pigmenty karoteny, xantofyly (scytonemin, gloeocapsin). Pozoruhodnou skupinou jsou symbiotické sinice. Vývojově nejvýznamnější je symbiotický původ chloroplastů, které jsou přímo nebo zprostředkovaně odvozeny od sinic. Sinice tvoří asociace s houbami (lišejníky): Gloeocapsa, Chroococcus, Nostoc, Stigonema - 8 % druhů lišejníků, s játrovkami (Blasia) a hlevíky (Anthoceros), s kapradinami (Azolla) a se semennými (Gunnera, Cykas). Geosiphon pyriforme (Glomeromycota) je příkladem endocyanózy, má význam z hlediska evoluce mykorrhiz. Cyanely jsou primitivní plastidy s mureinovou BS a redukovaným genomem. Žijí např. v buňkách prvoků, glaukofyt. Geologické stáří a vývoj sinic Podle fosilních nálezů (stromatolitů) se objevily sinice v prekambriu, před 3 až 2,5 mld let. Stromatolity jsou nejstarší známé fosilie, víceméně hřibovité útvary, které vznikají usazováním především uhličitanu vápenatého v pochvách sinic. Před 2 mld let byly sinice dominující skupinou na Zemi (2,5 0,6 mld let věk sinic ). Možní předci jsou anaerobní fotosyntetické bakterie (bakteriochlorofyl je strukturálně příbuzný chlorofylu a). Sinice, u nichž se vyvinul fotosystém II, měly významný podíl na vzniku kyslíkaté atmosféry Země. 27
30 Hospodářský význam sinic Vazači plynného dusíku - zúrodňování rýžových polí. Biotechnologie: vysoký obsah proteinů v sušině (60-70%, u zelených řas např. r. Chlorella pouze 12-28%), obsah vitamínů. Problémem je vysoký podíl NK, které v lidském těle metabolizují na kyselinu močovou, ta se ukládá do kloubů a vyvolává dnu. Možnost využití fikobiliproteinů (netoxická barviva) v biomedicíně místo radionuklidů. Vodní květy naopak znesnadňují technické, rekreační, rybářské a zejména vodohospodářské využití vodních nádrží. Některé druhy produkují jedovaté cyanotoxiny. Výrazné problémy jsou spojené s poklesem koncentrace kyslíku a hnilobnými procesy při náhlém úhynu vodního květu. Systém (umělý - podle stavby stélky a přítomnosti heterocytů): Řád Chroococcales jednobuněčné sinice tvaru koků nebo tyček žijící jednotlivě nebo v koloniích, Synechococcus jednotlivě žijící buňky, Microcystis kolonie nepravidelného tvaru, vodní květ, produkuje hepatotoxiny, Merismopedia deskovité kolonie, Chroococcus slizové kolonie, Gloeocapsa kulovité buňky obklopené vrstevnatým slizem, v němž se dělí, Chamaesiphon přisedle žijící válcovité polární buňky, množí se exosporami, Prochloron - jednobuněčný symbiont v zažívacím traktu mořských sumek, Prochlorococcus - jednobuněčný, dominující složka oceanického pikoplanktonu v letním období, až 30-50% primární produkce oceánů. Řád Oscillatoriales vláknité sinice bez heterocytů a akinet, tvoří hormogonie, Oscillatoria, Limnothrix, Phormidium, Spirulina a Arthrospira - šroubovitě vinuté trichomy, výživové doplňky, Trichodesmium - Rudé moře. Řád Nostocales mají vlákna přímá či s nepravým větvením, tvoří heterocyty a akinety, výskyt jednotlivě či v koloniích, řada z nich je součástí vodního květu a producenty toxinů - Anabaena, Aphanizomenon; Nostoc ve vodě i na povrchu půdy, častý symbiont, v Číně prodáván jako drahá zelenina. Řád Stigonematales - pravé větvení vláken, Stigonema (Pozn.: nepravé větvení při přerušení vlákna, pravé větvení změna roviny dělení buňky). 28


31 DOMÉNA (NADŘÍŠE): EUKARYA ŘÍŠE: EXCAVATA Excavata jsou zpravidla jednobuněčné organismy s centrálním cytostomem (buněčná ústa) ve tvaru rýhy. V něm kmitá bičík, který svým pohybem přihání drobné organismy. Druhý bičík směřuje dopředu a slouží k pohybu. Mnozí exkaváti však tyto znaky druhotně ztratili, např. přichází o cytostóm, jeden bičík, atp. Oddělení EUGLENOPHYTA (EUGLENOZOA) - KRÁSNOOČKA Jednobuněční, zpravidla volně žijící bičíkovci. Buňku pokrývá pelikula uložená pod plazmatickou membránou a složená ze šroubovitě vinutých do sebe zapadajících bílkovinných proužků. Někteří zástupci mají hnědě zbarvenou schránku - loriku. Pohyb zajišťují bičíky, které vyrůstají z lahvicovité nádržky (ampuly), na předním konci buňky. Fotosyntetické pigmenty: chlorofyl a+b, ß-karoten, xantofyly (diadinoxanthin). Povrch chloroplastu pokrývají 3 membrány, 3-12 tylakoidů tvoří lamelu. Chloroplasty vznikly sekundární symbiózou, kde endosymbiontem byla zelená řasa. Zásobní látka: paramylon (chemicky příbuzný s chrysolaminaranem heterokontofyt). Autotrofní druhy vyžadují pro svůj růst organické zdroje N, C a vitamíny (výživa mixotrofní a auxotrofní). Druhy bez chloroplastů se živí osmotickým vstřebáváním roztoků nebo pohlcováním prvoků a bakterií (fagotrofie). Dravé druhy jsou vybaveny faryngeálním aparátem, který vysouvají z buňky a vbodávají do buňky (kořisti), kterou následně vysají. Stavba buňky Krásnoočko štíhlé se často používá jako modelový organismus, dobře se pěstuje v kulturách. Buňky mají proměnlivý tvar. V klidu jsou vřetenovité, při podráždění vakovité či široce oválné. Proměny tvaru umožňuje pružná pelikula. Pod pelikulou jsou mukocysty, produkující sliz, u fagotrofních druhů vymrštitelné trichocysty. beled.jpg, Bičíky (1-2) - zajišťují pohyb krásnooček, vyrůstají z ampuly na předním konci buňky, resp. z bazálních tělísek umístěných v plazmě pod nádržkou. Původní jsou 2 bičíky, např. u r. Euglena jeden zakrní a zůstává uvnitř nádržky. Bičík na příčném průřezu je oválný, mírné zaškrcení jej dělí na 2 části: jedna obsahuje axonema s obvyklou strukturou /(9x2)+2/, ve druhé je 29
32 paraflagelární lišta (má parakrystalickou strukturu), hlavní složka fotoreceptoru, který spolu se stigmatem řídí fototaktické reakce buňky. Povrch bičíku je pokryt porostem mastigonemat. Ampula je typickou strukturou krásnooček, skládá se z rozšířené vakovité části, úzkého kanálku a ústí, do ní se uvolňuje sliz tvořený v diktyozómu a vyprazdňují se pulsující vakuoly (osmoregulace), ústí do ní faryngeální aparát. Stigma - uloženo v plazmě na úrovni ampuly, nesouvisí s chloroplasty (extraplastidiální). Sestává z četných pigmentových granulí, pohybuje se po povrchu ampuly a tím zakrývá část paraflagelární lišty v bičíku, sídlo fotoreceptoru. Jádro - má velké jadérko, i v interfázi zůstávají chromozómy v kondenzovaném stavu. Chloroplasty obsahují pyrenoid. Tylakoidy se vyvinou pouze při růstu krásnoočka na světle, ve tmě po 150 hod. mizí a zůstanou pouze struktury připomínající proplastidy (apochlorie). Za určitých podmínek (vysoká teplota kultivace, UV záření, antibiotika) je možné zbavit eugleny chloroplastů zcela (apoplastie), to neohrozí jejich životaschopnost, pokud zajistíme současně heterotrofní výživu. V plazmě některých krásnooček (Euglena sanquinea) se tvoří karotenoid astaxanthin, který překrývá zelenou barvu chloroplastu. Některé druhy (Trachelomonas) vylučují na svém povrchu hnědě zbarvenou schránku, základ schránky tvoří sliz, který je mineralizovaný sloučeninami manganu a železa. Na povrchu jsou bradavky, osténky... Rozmnožování podélným dělením buněk za pohybu či v palmeloidním stádiu (kolonie obklopené slizem). Výskyt v přírodě téměř ve všech sladkovodních biotopech s čistou i znečištěnou vodou i v brakické vodě. Způsob výživy určuje výskyt ve vodách znečištěných či značně eutrofizovaných (význam při čištění odpadních vod). Některé druhy patří k indikátorům znečištěných vod, Euglena viridis vegetuje i v polysaprobní vodě a je jedním z mála fotosyntetizujících organismů, který se v takovém prostředí udrží. Jiný druh, E. mutabilis žije naopak v extrémně kyselé vodě. Fylogenetické vztahy Krásnoočka zaujímají izolované postavení mezi ostatními řasami. Některé cytologické a biochemické znaky je sbližují s říší rostlinnou, jiné s živočišnou (AAA-dráha syntézy lyzinu - jako u hub - se zřejmě vyvinula velmi brzy u heterotrofních eukaryí, které byly i předky euglen). Původní jsou pravděpodobně fagotrofní eugleny, při vývoji fototrofních druhů proběhla endosymbióza zřejmě jen jednou. Zástupci Eutreptia - pokládá se za vývojového předka euglen, má 2 přibližně stejné bičíky Peranema, Entosiphon - bezbarví bičíkovci s faryngeálním aparátem řád: Euglenales - zelení nebo bezbarví bičíkovci s jedním bičíkem vyrůstajícím z buňky, druhý je zakrnělý, buňka má pevný nebo proměnlivý tvar, Euglena E. gracilis - terčkovité chloroplasty, E. viridis - páskovité chloroplasty uspořádané do hvězdice, E. spirogyra - tuhá, výrazně ornamentovaná pelikula, E. sanquinea - astaxanthin, tvořívá cihlově červenou neustonickou blanku ve znečištěných rybnících a tůních, Phacus - ploché buňky Trachelomonas hnědé až černé loriky s druhově specifickým tvarem hrdla, jímž prochází jediný bičík Colacium - rod poněkud výjimečný svou životní strategií, buňky bez bičíku žijí přisedle na povrchu drobných vodních bezobratlých (korýši, vířníci), vzácněji i na větších pohyblivých řasách (Volvox, Trachelomonas). Na povrch hostitele je Colacium přichyceno slizovou stopkou nebo 30

33 terčíkem, dceřiné buňky, které vzniknou po dělení, zůstávají na hostiteli mateřské buňky spojené do kolonie. Pohyblivé bičíkaté formy se vytvářejí jen výjimečně a hlavním účelem je transport na jiného hostitele. Oddělení ACRASIOMYCOTA - AKRASIE, BUNĚČNÉ HLENKY Základní typ stélky je haploidní bezblanná myxaméba s 1 jádrem - válcovitá, s lalokovitými pseudopodii obvykle bez subpseudopodií (jemné výběžky na koncích pseudopodií). Bičíkatá stádia (myxomonády) většinou chybějí. Výživa myxaméb holozoicky (pohlcování baktérií, kvasinek, spor hub apod.), rozmnožování dělením, nepříznivé období v podobě mikrocyst či sférocyst. Po intenzivní trofické a reprodukční fázi na úrovni myxaméb dojde ke shlukování (agregaci) myxaméb vznikají mnohobuněčné shluky - pseudoplazmodia, z nich vznikají sorokarpy (stopka, sorofor nese na vrcholu sorogen, z něhož vzniká sorus spor, BS spor je chitinózní, bez celulózy). Výskyt a ekologie: krátkodobě žijící organismy, hojné v půdě (zemědělské, lesní), na borce stromů, na tlejícím organickém materiálu, hrají určitou roli v biologii půdy. Druhy pěstované v kulturách slouží pro studium morfogeneze a buněčného dělení. Systém: malá skupina améboidních organismů, které připomínají některými znaky dictyostelidy nebo i vzdáleněji ostatní skupiny hlenek. Acrasis rosea - nejprozkoumanější druh, běžný v půdě a na organickém materiálu, pěstování na agarových půdách s přidáním kvasinek a baktérií jako zdroje potravy, oranžovorůžová barva myxomonád a pseudoplasmodií, sorokarp rozvětvený, spory v jednoduchých nebo rozvětvených řetízkách /Fig-1-Amoebae-of-Heterolobosea-A-Acrasis-rosea-B-Fumarolamoeba-ceborucoi-C.png 31
 a panožky - kořenovité retikulopodie, tenké filopodie nebo axopodie. Nenacházíme u nich široké panožky (lobopodie) jako mají měňavky.")
34 ŘÍŠE: RHIZARIA Morfologicky i ekologicky to velmi rozmanitá skupina, zahrnující zástupce bývalých polyfyletických kmenů bičíkovců i kořenonožců. Většina rhizarií má dva bičíky (i když u některých zástupců chybí) a panožky - kořenovité retikulopodie, tenké filopodie nebo axopodie. Nenacházíme u nich široké panožky (lobopodie) jako mají měňavky. Rhizaria jsou většinou heterotrofní organismy bez chloroplastů, žijí volně, mřížovci jsou součástí mořského planktonu, dírkonošci tvoří benthos. Patří sem i druhy parazitické (např. nádorovky) nebo fotosyntetisující (Chlorarachniophyta, Paulinella chromatophora). Oddělení CHLORARACHNIOPHYTA (RETICULOPHYLOSA) Donedávna jediným zástupcem je Chlorarachnion reptans, popsaný v r z kultury mořských řas z Kanárských ostrovů (dodnes druhově velmi chudá, ale teoreticky významná skupina). Vegetativní stélkou je filoplazmodium, v němž jsou jednojaderné buňky spojeny tenkými plazmatickými vlákny. Fotosyntetické pigmenty: chlorofyly a+b, zásobní látka podobná chrysolaminaranu, výživa částečně autotrofní, převažuje fagotrofní (živí se rozsivkami). Chloroplast obalují 4 membrány, mezi dvojicemi membrán je poměrně široký periplastidiální prostor, 3 tylakoidy tvoří lamely. V chloroplastu je přítomen nukleomorf, pozůstatek jádra původního eukaryotického fototrofního endosymbionta. Chlorarachniophyta jsou důležitým důkazem vzniku autotrofní buňky cestou laterálního přenosu chloroplastu, při kterém se uplatnila fagocytóza. Oddělení PLASMODIOPHOROMYCOTA - PLAZMODIOFORY, NÁDOROVKY Ekonomicky významná skupina vysoce specializovaných biotrofně parazitických organismů, obligátní endoparazité cévnatých rostlin, ale i parožnatek, chromist a oomycetů. Podstatnou fází životního cyklu je beztvará, nahá, mnohojaderná masa protoplazmy parazita uvnitř buněk hostitele, tzv. paraplazmodium (na rozdíl od plazmodia hlenek nevzniká splýváním menších plazmodií, ale dělením jader mladých, 1-jaderných plazmodií, není schopno 32
35 améboidního pohybu a výživa je osmotrofní). Myxaméby chybějí, připomínají je mladá, jednojaderná plazmodia uvnitř hostitele, která jsou schopna přesunout se améboidně do sousedních buněk, dělením jader z nich vznikají mnohojaderná paraplazmodia. Zoospory jsou neoblaněné, mají 2 heterokontní akrokontní bičíky, infikují další hostitelské rostliny. BS sporangií a cyst obsahuje chitin a další látky. Životní cyklus: na příkladu nádorovky kapustové Plasmodiophora brassicae. Způsobuje nádory a deformace na kořenech napadených rostlin (hypertrofie: abnormální zvětšení a hyperplázie: patologické zmnožení buněk). Buňky nádorů zaplňují cysty (odpočívající spory), které po rozpadu napadených pletiv přezimují v půdě. Za příznivých podmínek (vlhko, chemotaxe: přítomnost hostitele) vyklíčí primární zoospora, pohybuje se pomocí bičíků, přichytí se na kořenový vlásek hostitele, proděraví stěnu hostitelské buňky a pronikne do ní (zvláštní organela v podobě trubice s trnem). Mladé jednojaderné paraplazmodium se dělí a napadá sousední buňky, později dochází k dělení jader a vzniku mnohojaderných haploidních primárních sporangiálních paraplazmodií. V létě se paraplazmodium rozdělí na tenkostěnná sporangia, z nichž se uvolní sekundární zoospory do půdy a působí další infekce. Některé sekundární zoospory v půdě splývají (gametogamie) a vzniklé dvoujaderné zoospory po proniknutí do kořenů hostitele jsou základem pro vznik cystogenního paraplazmodia, které je zprvu dvoujaderné, poté mnohojaderné - to působí vznik hypertrofií a hyperplázií. Zde karyogamie a meiosa - tlustostěnné cysty. Paraplasmodia jsou tedy dvou typů: sporangiogenní (haploidní) vznik tenkostěnných sporangií (gametangií) se zoosporami (gametami), cystogenní (diploidní) po R! se mění v sorus tlustostěnných odpočívajících spor (cyst). 33
36 Výskyt, význam, zástupci: parazité na některých řasách (Vaucheria, Chara), oomycetech (Saprolegnia, Pythium) i na cévnatých rostlinách. Nejzávažnější je výskyt na pěstovaných cévnatých rostlinách: Plasmodiophora brassicae - nádorovitost košťálové zeleniny, u nás i ve světě způsobuje značné výnosové ztráty jak na brukvovité zelenině, tak i na krmných a olejnatých plodinách, hlavně na řepce olejné, 5-8 let nepěstovat na pozemku brukvovité rostliny, může pomoci vápnění půdy či některé fungicidy. Spongospora subterranea - působí prašnou strupovitost bramborových hlíz, přísně karanténní choroba, díky čemuž je u nás poměrně vzácná. Polymyxa betae v kořenech cukrovky ale i jednoděložných rostlin včetně obilnin, vektor rostlinných virů. ŘÍŠE: AMOEBOZOA Amoebozoa či česky měňavkovci je skupina pojmenovaná podle toho, že se k ní řadí velké množství organismů amébovitého (měňavkovitého) typu. Pro Amoebozoa jsou typické panožky, tedy organely pohybu. Ten je způsobován v podstatě polymerizací a depolymerizací aktinu. Panožky jsou vždy lobopodiového či filopodiového typu. Pokud tyto organismy mají mitochondrie, pak vždy s tubulárními kristami. Je pravděpodobné, že společný předek Amoebozoa měl jeden bičík; proti tomu však mluví skutečnost, že většina měňavkovců má dnes bičíky dva. Oddělení HLENKY MYXOMYCOTA (MYCETOZOA) Trofickou fází jsou myxaméby (MA, ploché s vláknitými pseudopodii a subpseudopodii), myxomonády (MM) nebo mnohojaderné slizovité útvary: pseudoplazmodia či plazmodia, klidovým stádiem mikrocysty či sklerocia. Holozoická (fagocytóza) nebo osmotrofní výživa, popř. parazitismus. Reprodukční fáze tvorba mikroskopických i makroskopických plodniček (sorokarpů a sporokarpů). Sorokarpy jsou bez buněčné stěny a vznikají z pseudoplasmodií, sporokarpy již mají stěnu, vznikají z pravých plazmodií. Spory jednobuněčné, většinou s celulózní buněčnou stěnou. Oddělení je rozčleněno na 3 třídy: Protosteliomycetes, Dictyosteliomycetes, Myxomycetes. De Bary (1884) objevil životní cyklus hlenek a usoudil, že nejsou příbuzné s houbami, ale s prvoky. Zajímavé je, že v současných systémech, kde chybí říše prvoků, patří hlenky do skupiny Amoebozoa, která je řazena do Unikonta opět spolu s houbami, živočichy aj. TŘÍDA: PROTOSTELIDY PROTOSTELIOMYCETES Nejjednodušší mikroskopické hlenky, jejichž trofické stádium tvoří myxaméby nebo myxomonády s 1-2 akrokontními (vrcholovými, anteriorními) bičíky, splýváním myxaméb vznikají mnohojaderná plazmodia. Sporokarpy obsahují 1-4(-8) spory. Životní cyklus: U nejjednodušších typů (Nematostelium) ze spory vyklíčí 1 myxaméba, která během kulminační fáze vytvoří stopkatý sporokarp. Ten se oblaní a uvolní 1 sporu... U r. Ceratiomyxa při klíčení spory vznikne 1-2 bičíkatá myxomonáda, která přechází v myxamébu. MA splývají v mnohojaderné plazmodium, zde se předpokládá kopulace MA, a tedy vznik 2n-plazmodia. Plazmodium zbělá, vytvoří povlak bělavých sloupečků či parůžků až 10 mm vysokých. Na nich vznikají po meiozi na jemných stopečkách jednobuněčné spory (jednosporová sporangia), zprvu jednojaderné, po mitóze vznikne 4-8 buněčný útvar, z každé buňky se uvolní MM. 34
,")
37 Výskyt a ekologie: v půdě, na rozkládajícím se organickém materiálu i borce stromů. Tvoří podstatnou složku dekompozitních společenstev. Zástupci: Protostelium mycophaga - jeden z nejběžnějších druhů na nejrůznějším organickém materiálu Ceratiomyxa válečkovka, exosporická hlenka s relativně velkými sporokarpy (až několik cm), střídáním jaderných fází, pohlavním rozmnožováním, nejběžnější C. fruticulosa - velmi hojný druh na tlejícím dřevě TŘÍDA: DIKTYOSTELIDY DICTYOSTELIOMYCETES MM se netvoří, trofické stádium tvoří jednojaderné haploidní MA živící se bakteriemi. Agregací MA vzniká pseudoplazmodium, spory se tvoří na mnohobuněčných sorokarpech, které jsou makroskopické (až 1 cm). Mnoho shodných znaků s tř. Acrasiomycetes, ale mají odlišnou stavbu pseudopodií, více vyvinuté sorokarpy s celulózní stěnu spor. Výskyt, význam, zástupci: druhy půdní i koprofilní, význam v biologii půdy Dictyostelium - dosud nejprostudovanější zástupce, modelový organismus v genetických nebo biochemických studiích Pohlavní rozmnožování je vzácné: dvě buňky fúzují a zvětšují se v obří buňku (makrocysta) kanibalisticky požírajíce sousední améby. Obří buňka se podrobuje meiotickým a mitotickým dělením, která dávají vznik novým haploidním amébám. Vegetativní část ŽC: haploidní buňky se uvolní ze spory, žijí jako améby živící se holozoicky, vytváří tenká filopodia (pseudopodia), za nepříznivých podmínek vytvoří mikrocystu, z níž se opět uvolní MA. Reprodukují se asexuálně mitotickými děleními. Když vyčerpají potravu, améby přítomné v určité oblasti se seskupí v určitém bodě do agregátu pomocí chemotaktických signálů emitovaných buňkami uprostřed agregátu, v nich se tvoří atraktant akrasin (cyklický AMP), chemotakticky láká další MA k agregačnímu centru. Vzniká pseudoplazmodium, které získá tvar slimáka (slug), pohybuje se na vhodnější místo a vytvoří plodničku (fruiting body). MA tvoří stopečku, po které další MA putují na vrcholek - sorokarp (spory mají celulózní BS). 35
38 Rod Coenonia s jediným druhem C. denticulata byl popsán 1884 z tlejících bobových lusků. Od té doby se jej nepodařilo znovu nalézt, přestože byla vypsána cena karton (12 lahví) dobré whisky TŘÍDA: VLASTNÍ HLENKY MYXOMYCETES Trofickou fázi tvoří haploidní jednojaderné MA a MM (2 hladké, apikální heterokontní bičíky), přechod obou typů může být reverzibilní, a diploidní, mnohojaderné, pohybující se plazmodium. Obě stadia se množí dělením a živí holozoicky. 3 základní typy plazmodií (specifické vlastnosti, které nenajdeme u žádného jiného oddělení Protist: rytmické proudění plazmy, synchronizované dělení jader, splývání plazmodií): protoplazmodium - nejprimitivnější typ, mikroskopické, protoplazma se pomalu pohybuje, dává vznik vždy jen 1 sporokarpu, afanoplazmodium - větší (lupa), jemné, tvořené průhlednými, síťovitě uspořádanými žilkami a cytoplazmou, která je obklopuje, proudění protoplazmy rychlé, rytmické, dá vznik více sporokarpům, faneroplazmodium - makroskopické, žilnaté, i barevné (červená, žlutá, černá, atp. flavony či karotenoidy, ph substrátu, přítomnost pohlcených organismů) protoplazma silně zrnitá, tvořena želatinózní a tekutou částí, rytmické proudění protoplazmy, vznik většího počtu sporokarpů. Pohyb plazmodia je negativně fototaktický (do tmy), pozitivně hydrotaktický (do vlhka) a směrem k potravě. Plazmodia se mohou změnit na sklerocium (tvrdý, nepravidelný, nebuněčný útvar, vzniká za nepříznivých podmínek) či sporokarp (fruktifikační orgán, který představuje haploidní reprodukční fázi životního cyklu). 3 typy sporokarpů: sporangium - vzniká z celých protoplazmodií či z částí jiných typů plazmodií, stopkaté či přisedlé, aethalium - vzniká z větších částí plazmodií sloučením řady sporangií, velké, polštářovité, nestopkaté (pseudoaethalium - stěny jednotlivých sporangií uvnitř aethalia zřetelné), plazmodiokarp - vzniká z velkých částí síťovitého plazmodia, zachovává si siťovitě žilnatý tvar. Uvnitř sporokarpu kapilicium (vlákna jednotlivá různě strukturovaná nebo jejich sítě, nebuněčná struktura, vyloučeno z vakuol) nebo pseudokapilicium (nepravidelné niťovité či destičkovité útvary mezi sporami), popř. kolumela (tlustší centrální, ze stopky vybíhající útvar) a spory (celulózní BS), různá barva a povrch. Obal sporokarpu: peridie, je tvořena exkrecí plazmy, často inkrustována uhličitanem vápenatým, proto je křehká a drobivá. Životní cyklus: haplo-diplontní, 2n-fáze převažuje, ze spory se uvolní 1-2(-8) haploidní MA nebo MM, MA se mění v MM, holozoická výživa, dělení, MM i MA mohou tvořit cysty (nepříznivé podmínky), opačně pohlavně laděné MM či MA kopulují - diploidní MA (zygota) - mitotická dělení - vícejaderné 2n-plazmodium - splývání plazmodií - výsledek: plazmodium s několika tisíci 2n-jader (bělavé i pestré, obsahuje bílkovinu (aktin a myozin) schopnou stahování a roztahování, tvorba sklerocií), při přechodu do reprodukční fáze se tvoří na povrchu blanka (hypothalus), z ní potom vyrůstají sporokarpy, uvnitř vzniká kapilicium a dochází k meiotickému dělení jader - vznik spor, pomocí hygroskopických pohybů kapilicia jsou spory vymršťovány z plodniček, klíčivost i několik desítek let. Pro systematické zařazení a určování hlenek má význam morfologie sporokarpu, kapilicia a zbarvení a ornamentika výtrusů. 36
39 Výskyt a ekologie: kosmopoliti, vazba na vlhčí lesní biotopy (u nás pozdní jaro až ranný podzim) - odumřelé rostliny, tlející dřevo, listí, borka, půda, význam v přírodě není zcela objasněn. Zástupci: Echinostelium lunatum - nejmenší vlastní hlenky, protoplazmodia Tubifera ferruginosa - hojný druh, několik cm velká rezavá aethalia ze vzpřímených válcovitých sporangií Lycogala epidendrum - kulovitá aethalia (1 cm), hojná na tlejícím dřevě (korálově červená, zralá: šedá) Reticularia (Enteridium) lycoperdon - aethalia až 8 cm, stříbřitá peridie Physarum - stopkatá sporangia či plazmodiokarpy Leocarpus fragilis - stopkatá elipsoidní sporangia 2-4 mm (žlutá, pak višňová) Fuligo septica - aethalia až 20 cm, žlutá, bělavá, naoranžovělá Arcyria - druhy na tlejícím dřevě, stopkatá válcovitá sporangia Hemitrichia serpula - plasmodiokarp Trichia varia - hojný druh v lese Stemonitis - až 2 cm dlouhá válcovitá sporangia na dlouhých stopkách těsně nahloučená vedle sebe, S. fusca - častý na dřevě Comatricha nigra - staré větévky, zem, kulovitá sporangia Cribraria - síťovitý rozpad peridie ŘÍŠE: CHROMALVEOLATA Rozsáhlá říše obsahující heterotrofní I autotrofní organismy. Bičíkovci, zoospory a planogamety mají zpravidla 2 bičíky, alespoň jeden z nich je pleuronematický (péřitý, porostlý 2 řadami trubicovitých mastigonem). 37
, ocellus atd. Jádro - dinokaryon - obsahuje téměř trvale kondensované chromozomy bez histoproteinů.")
40 Oddělení DINOPHYTA (DINOFLAGELLATA, DINOZOA) - OBRNĚNKY Převážně mořští bičíkovci se 2 bičíky a obalem buňky (amfiesma - théka). Mají velmi mnoho specifických rysů: pancíř, dinokaryon, pusuly (specializované potravní vakuoly), ocellus atd. Jádro - dinokaryon - obsahuje téměř trvale kondensované chromozomy bez histoproteinů. Asi 50% druhů obsahuje chloroplasty získané sekundární nebo terciární endosymbiózou, ostatní druhy jsou heterotrofní. Kombinace fotosyntetických pigmentů: chlorofyly a + c, karotenoid peridinin. Zásobní látka: škrob. Autotrofní výživa doplňována lovem či příjmem organických látek a vitamínů. Heterotrofní obrněnky jsou fagotrofové, saprofyté (roztoky) i parazité. Významné jsou symbiotické interakce obrněnek. Mnohé mořské obrněnky produkují nebezpečné toxiny. Stavba buňky Převládají bičíkovci. Tvar buněk: zploštělé, jiné nesouměrné, dorziventrálně stavěné. Buňka je při pohledu ze strany rozdělena příčnou rýhou (cingulum) na 2 nestejné části, horní epikón (epitéka) a spodní hypokón (hypotéka). Na břišní straně probíhá krátká podélná rýha (sulcus). Obal buňky: amfiesma, vnější povrch amfiesmy tvoří plazmatická membrána, pod ní je uložena vezikulární vrstva z plochých měchýřků. U některých obrněnek nemají vezikuly pevný obsah a buňky se chovají jako bezblanné (Gymnodinium, Noctiluca). Jiné obrněnky (Ceratium, Peridinium) mají ve vezikulech celulózní destičky, které tvoří pevný pancíř. Destičky mají určitý tvar a polohu v pancíři, jejich počet se s růstem buňky nemění, pouze dorůstají na okraji. Tvar, poloha a počet destiček je pro jednotlivé druhy charakteristický. Destičky a BS zygospor obsahují acetorezistentní vrstvu, dinosporin, která umožňuje zachování obrněnek jako mikrofosilií. Buňky mohou starou théku svléci (ekdyzis) a vytvořit si novou bičíky (na povrchu 2 řady bičíkového vlášení - mastigonemat) vyrůstají buď na předním konci buňky (desmokontní) nebo v místech, kde se stýkají rýhy (dinokontní). Kratší z nich se vlní v prsténcové rýze, má význam pro víření vody, přísun potravy, delší bičík - pro pohyb. Jádro je nápadně velké, dobře patrné v živých buňkách, nepřítomnost histoproteinů, které u jiných eukaryot obalují vlákna DNA, je považována za primitivní znak, sbližující obrněnky s prokaryoty (mezokaryon). Průběh mitózy je rovněž zvláštní, zůstává zachována jaderná blána, mitotické vřeténko je mimojaderné. 38
41 Chloroplasty většinou žlutohnědé, na povrchu 3 membrány, 3 tylakoidy tvoří lamelu. Vznik sekundární či terciální endosymbiózou, popř. kleptoplastidy (chloroplasty z kořisti, přežívají až několik měsíců a zůstávají fotosynteticky aktivní). Zásobní látka: škrob, lipidy, volutin. Světločivná organela - u některých přítomna struktura podobná stigmatu, nejdokonalejší je ocellus u r. Warnowia (připomíná jednoduché oko). Výživa je hetero- nebo mixotrofní, existují různé mechanismy získání potravy (fagopod trubička, kterou po vbodnutí do kořisti je vysáta cytoplazma, palium závoj membránová struktura, která překryje kořist a obsahuje trávicí enzymy, které buňku rychle stráví. K dalšímu zpracování potravy slouží pusuly. Mezi destičkami pancíře jsou umístěny trichocysty, při podráždění vystřelují tenká vlákna. U některých mořských obrněnek (Noctiluca) je známa bioluminiscence, v organelách scintilonách přítomny látky luciferin a luciferáza, které světélkování způsobují. Dělení buněk a rozmnožování Rovina dělení probíhá šikmo k podélné ose buňky. Druhy s pancířem - nestejnoměrné rozdělení, každá buňka si dotvoří chybějící část. Kapsální, kokální a vláknité druhy tvoří zoospory podobné buňkám r. Gymnodinium. U některých obrněnek znám pohlavní proces izo- a anizogamie. Zajímavá je tvorba dvou typů zygot pohyblivé planozygoty, která vypadá jako velká vegetativní buňka a nepohyblivé hypnozygoty, která je dormantním stádiem (fosilní = hystichosféry). Symbiózy obrněnek S mořskými bezobratlými: zooxantely jsou pro korálové polypy hlavním zdrojem živin, poskytují svým hostitelům až 50 % fotosyntetických produktů. Výskyt v přírodě S rozsivkami jsou obrněnky hlavní součástí mořského fytoplanktonu. Při přemnožení v pobřežních eutrofních vodách tvoří červenohnědé zbarvení vody (red tide, rudý příliv). Některé produkují toxiny, např. saxitoxin (je 1000x účinnější než kyanid a 50x více než kurare). Podle působení se toxiny dělí na paralyzující, neurotoxické, hepatotoxické, cytotoxické. Problém s těmito toxiny mají všechny přímořské státy, ročně onemocní asi 50 tis. osob. Stačí vdechování vodního aerosolu s obrněnkami nebo se dostávají do potravního řetězce prostřednictvím herbivorů. U nás tvoří plankton stojatých vod a rašelinných tůní. 39
42 Geologické stáří Velmi stará skupina: hystrichosféry (fosilní hypnozygoty) z prekamria, asi 600 mil. let. Zástupci Prorocentrum - moře a brakická voda, pancíř z levé a pravé misky, produkce paralytického jedu venerupinu (otravy ryb, jejich požitím i lidí) Dinophysis - mořský plankton, kožovitý límeček na předním konci buňky Ceratium - epikón s dlouhým výběžkem, hypokón s 2-3 výběžky, hlavně teplá moře, u nás běžný C. hirundinella, živí se drobnými planktonty, které zachycuje panožkami, vyrůstajícími z břišní strany buňky Peridinium - oválné, z břišní strany zploštělé buňky, pokryté pancířem (epikon tvoří 14, hypokon 7 destiček) Gymnodinium - nahé buňky v moři, rybnících, rašelinných tůňkách Noctiluca - až 1,2 mm velké, kulovité, nahé buňky s ohebným přívěskem, který pomáhá při lovu potravy Blastodinium kokální obrněnka, parazituje ve střevech korýšů, ale přesto buňky obsahují chloroplasty Pfisteria piscicida bývá označována jako číhající predátor a způsobuje rozsáhlé úhyny ryb. Má složitý životní cyklus s mnoha stádii. Nejsou-li v blízkosti ryby, živí se drobným fytoplanktonem. Rozpozná chemický signál vysílaný rybami, vytvoří velké množství zoospor, které produkují prudký jed, ichtyotoxin, ten ryby znehybní, usmrtí a vyvolá rozpad kožního epitelu. V místech otevřených ran se zoospory přichytí, dělí se a pohlavně rozmnožují. Pak se změní v cysty a čekají na další příležitost. Oddělení HETEROKONTOPHYTA (CHROMOPHYTA) Oddělení spojující dosti heterogenní skupinu tříd (jednobuněčné mikroskopické, ale i desítky metrů velké pletivné stélky) s následujícími společnými znaky: - bičíkaté buňky mají dva heterokontní bičíky lišící se délkou, strukturou mastigonem i funkcí, mastigonema je glykoproteinová a je složena ze 3 subjednotek: kotevní struktury, trubicovité části a 1 nebo několika terminálních přívěsků - chromatofory jsou kryty 4 membránami - tylakoidy srostlé po 3; po obvodu chloroplastu probíhá u většiny druhů věncová lamela, která odpovídá prstencovitému tvaru nukleoidu, v plastidu je často stigma a pyrenoid - fotosyntetická barviva jsou chlorofyl a+c, β-karoten, pro většinu tříd je charakteristická přítomnost xantofylu fukoxantinu, způsobujícího hnědavé zbarvení chromatoforů - zásobní látky jsou chrysolaminaran (β-1,3-glukan), dále olej a volutin (polyfosfátová zrna), nikdy škrob! Oddělení je v současnosti členěno na 9 tříd: Chrysophyceae (zlativky), Synurophyceae, Dictyochophyceae, Pelagophyceae, Phaeothamniophyceae, Phaeophyceae (chaluhy), Xanthophyceae (různobrvky), Eustigmatophyceae a Bacillariophyceae (rozsivky). TŘÍDA: ZLATIVKY CHRYSOPHYCEAE Malá, dobře definovaná skupina vyskytující se převážně ve sladkých vodách. Převažují jednotliví nebo koloniální bičíkovci s nahými buňkami. Povrch buňky kryje periplast. Skládá se z plazmatické membrány a systému podpůrných 40
, buňky nemnohých zástupců kryty křemitými šupinami. http://www.sinicearasy.cz/sites/default/files/chrysophyceae.")

43 mikrotubulů, což umožňuje průběžně měnit tvar buňky, někdy třeba i vytvářet panožky k lovu. Periplast může být překryt lorikou z celulózních mikrofibril (popř. i chitin; prostor mezi fibrilami vyplňuje amorfní hmota), buňky nemnohých zástupců kryty křemitými šupinami. Bičíky: ukotvení zajišťují 4 mikrotubulární kořeny, bazální tělísko hlavního bičíku je spojeno s povrchem jádra pomocí příčně pruhovaného kontraktilního vlákna (rhizoplastu). K pohybu slouží delší bičík se 2 řadami mastigonem. Kratší bičík jen s 1 řadou mastigonem, může být zakrnělý, je spojován s pohybem za světlem, v jeho bazální části je fotoreceptor. Výrazný je diktyosom (syntetická a exkreční funkce - sliz). Na tvorbě slizu se podílejí i mukocysty. U druhů žijících v hypotonickém prostředí vyvinuty pulsující vakuoly, zajišťující odčerpávání přebytečné vody (a odpadu metabolismu) z buňky (osmoregulační funkce). Chromatofory hnědé, často přítomen pyrenoid a stigma. Výživa je mixotrofní (osmotrofní nebo fagotrofní) nebo auxotrofní, u apoplastických druhů heterotrofní. Zlativky mají značnou spotřebu fosforečnanů - při dostatku je hromadí, při nedostatku pak využívají, teprve tehdy se kompetičně prosadí proti jiným řasám. Některé druhy mají vymrštitelné diskobolocysty (vymrštění způsobí prudký pohyb buňky opačným směrem). Buňky některých bičíkovců (chrysomonád) obývají schránky (loriky) z celulózních mikrofibril (může být i chitin); prostor mezi fibrilami vyplňuje amorfní hmota; buňky jiných zástupců kryty křemitými šupinami. Rozmnožování dělením buněk, vzácně hologamie, přečkávání nepříznivého období v podobě endogenních stomatocyst. V buňce se vytvoří kulovitý váček, vstupním otvorem do něj vejde plazma, organely a zásobní látky, stěna zkřemení, v otvoru se utvoří zátka; při klíčení se rozpustí zátka a z cysty vyrejdí bičíkovci. Výskyt: žijí převážně ve sladkých, ale i brakických i mořských vodách, upřednostňují čistší a měkkou vodu (nízký obsah Ca), ponejvíce planktonní, při přemnožení zlativek má voda zápach rybího tuku. Zástupci: bičíkovci s nahými buňkami: Uroglena planktonní kolonie 1-2 mm, Ochromonas jednotlivé buňky, s nálevkovitými lorikami: Dinobryon mnohobuněčné, volně plovoucí kolonie, Epipyxis přisedle na vláknitých řasách 41
44 křemité šupiny: Paraphysomonas jednotlivě žijící buňky bez chloroplastu, kruhové šupiny s ostnem, fagotrofie, hojný sladkovodní druh kapsální stélka: Hydrurus - větvené slizové kolonie na kamenech v horských potocích TŘÍDA: SYNUROPHYCEAE Hlavní rozdíly oproti předchozí třídě: povrch delšího bičíku s organickými šupinami, kratší je hladký, chybí stigma, rozdílná poloha bazálních tělísek bičíků, 2 mikrotubulární kořeny, žijí pouze ve sladkých vodách, buňka kryta křemitými šupinami, které mají druhově specifický tvar. Mallomonas - jednotliví bičíkovci, šupiny s ostnem, Synura - koloniální typy, obojí se vyskytují v planktonu našich rybníků. TŘÍDA: DICTYOCHOPHYCEAE Převážně mořské nanoplanktonní organismy. V periferní plazmě se tvoří tentakuly (panožky s vnitřním svazkem mikrotubulů). Patří sem křemití bičíkovci neboli silikoflageláti - pro jejich buňky je charakteristická vnitřní křemitá kostra různých tvarů - od jednoduchého kruhu po složené tvary (hvězdičky); tyto známy již z křídových usazenin. TŘÍDA: HNĚDÉ ŘASY FUCOPHYCEAE (PHAEOPHYCEAE) Až na malé výjimky mořské řasy. V souvislém pásu porůstají skalnatá pobřeží a sestupují do hloubky až 100 m. Nejvýznamnější skupina mořských makrofyt z hlediska produkce organické hmoty. Stélky vždy mnohobuněčné, přirostlé k podkladu: vláknité, heterotrichální i pletivné, mikroskopické i obří rozměry (Macrocystis až 60 m). Rozměrům stélky odpovídá její vnější a vnitřní diferenciace. Chaluhy jsou organismy se značnými nároky na světlo. Kombinace pigmentů: chlorofyl a+c, β-karoten, fukoxantin a violaxantin. Zásobní látky chrysolaminaran, manitol a olej jsou uloženy v plazmě nebo ve vakuolách. Stavba stélky a látky produkované chaluhami Stélka chaluh se nazývá stichoblast. Největších rozměrů a složitosti dosahují sporofyty zástupců ř. Laminariales: rhizoidy, kauloid, fyloidy (rhizoidy a kauloid mohou být až 10 let staré, fyloidy se každý rok obnovují). Stélka vykazuje též vnitřní morfologickou a funkční diferenciaci: pletivo krycí (epidermis), asimilační a v kauloidu vodivé pletivo a vnitřní dřeň (medula), na průřezu kauloidem jsou patrné periodické přírůstky. Nadnášení ve vodě pomocí dutých měchýřů naplněných plynem (u Fucus vesiculosus párové, u Sargassum kulovité na krátké stopce, u Nereocystis obří plovací měchýř nese četné dlouhé fyloidy, podobné měchýře používali Indiáni jako nádoby na vodu). Hnědá barva stélek určována především polyfenoly - feotaninem a florotaninem (bakteriostatické účinky, odpuzuje býložravce), který se hromadí v drobných vakuolách - fysodách. Buněčná stěna tvořena amorfní složkou, alginovými kyselinami (které vyplňují i mezibuněčné prostory) a fibrilární celulózní složkou. Vodorozpustné algináty (jednomocné soli) mají povahu gelu, jsou hlavní složkou slizu (nazývá se fukoidan). Velká produkce slizu snižuje možnost mechanického poškození a brání vyschnutí při odlivu. Dimetylsulfopropionát chaluhy produkují jako látku vyrovnávající osmotickou nerovnováhu, ten se postupně odpaří jako dimetylsulfid jeho vylučováním do prostředí se chaluhy významně podílejí na koloběhu S v atmosféře i vodě. 42
.")
45 Podílí se na tvorbě mraků, v nichž se mění na oxid siřičitý, nad pevninou pak vznikají kyselé deště. Chaluhy produkují také prchavé organobromidy, s nimi spojována destrukce ozonové vrstvy nad Arktidou (spolu s freony). V životním cyklu je častá rodozměna: Ectocarpales izomorfická (vývojově primitivní), Laminariales - heteromorfická, dominuje mohutný mnoholetý sporofyt, Fucales - gametofyt jako samostatná rostlina chybí, gamety vznikají v konceptakulech na receptakulech na sporofytu. Pohlavní rozmnožování - izogamie, anizogamie, oogamie. Při oplození se uplatňují feromony, chemotakticky lákají spermatozoidy k vaječným buňkám. Nepohlavní rozmnožování zoospory (zoidy, na S 2n, na G n), fragmentace stélek (Sargassum). Výskyt v přírodě Žijí v mořské i brakické vodě, ze sladkých vod jsou známy 4 rody. V moři obývají úzký pruh pobřeží - litorál a sublitorál, dominují v chladných mořích. Tvoří spolu s ruduchami a zelenými řasami vertikálně členěná společenstva. Hloubka, do které chaluhy sestupují, závisí na intenzitě světla. Složení společenstev ovlivňováno geografickou polohou: S část Atlantského pobřeží Fucus, Ascophyllum (litorál), Laminaria (sublitorál). Severní pobřeží Tichého oceánu: Macrocystis, Nereocystis. Sargasové moře - jméno dle řasy Sargassum, původně bentické řasy se adaptovaly na vznášení, rozmnožují se pouze fragmentací stélky. Hospodářský význam Odedávna využívány na palivo, hnojivo, výrobu sody (Na 2 CO 3 ), potaše (K 2 CO 3 ), jódu, krmivo ovcí. V Asii a Rusku - potravina ( morskaja kapusta Laminaria), zdroj vitamínů, bílkovin, stopových prvků a dalších cenných látek pro zdraví. Pěstování několika rodů (marikultury) naočkované mladé sporofyty na nylonových lanech. Zdravotnický výzkum - látky brzdící růst rakovinných nádorů. Algináty - soli alginových kyselin (soli s Na + a K + rozpustné, s Ca 2+ a Mg 2+ nerozpustné) - potravinářství, farmaceutický, textilní, papírenský průmysl, zubní lékařství otisky, alginátová vlákna v chirurgii. Algináty jsou pro člověka nestravitelné, ale vyvolávají pocit sytosti: přípravky na hubnutí. Imobilizace kvasinek aj. biotechnologicky využívaných mikroorganismů (velký kvasný povrch díky zabudování do alginátových gelů). Systém: (14 řádů, zde 3) 43
46 Řád: Ectocarpales - heterotrichální stélka, izomorfická rodozměna, feromon ektokarpen s vůní džinu, řád byl považován za nejprimitivnější, dnes spíše za odvozený, Ectocarpus Řád: Laminariales - obsahuje druhy s největšími stélkami (sporofyty), rhizoidy, kauloid a fyloidy, růst pomocí dělivého pletiva, meristodermu, na rozhraní kauloidu a fyloidu, diferencovaná pletiva, heteromorfická rodozměna, oogamie, v zimě na S se tvoří zoidangia, v nich meioza, ze zoidů vyrůstají a gametofyty, feromón lamoxiren: na G až spermatoziodů, jejichž vypuzení trvá 0,3 s (reakce nastane, když povrch spermatozoidu zasáhne 10 molekul feromonu za sekundu), ze G vyroste trs sporofytů Laminaria saccharina - celistvý fyloid, L. digitata - fyloid roztřepený na pruhy, lesy laminárií v sublitorálu, 1-2,5 m vysoké Nereocystis jednoleté, ale největší mořské rostliny, kauloid 10 m, plovací měchýř velikosti fotbalového míče, z něho kštice fyloidů, celková délka stélky až 40 m Macrocystis váha až 300 kg, kauloid m, na něm kopinaté fyloidy s kulovitými měchýřky, bazální část stélky s rhizoidy je víceletá (až 30 let), každý rok vyrůstají nové kauloidy s fyloidy (denní přírůstek až 30 cm) Řád: Fucales - bez rodozměny, pohlavní buňky v konceptakulech na receptakulech na sporofytu, oogamie Fucus vesiculosus - asi 40 cm, vidličnatě větvená stélka, páskovitá, s dvojicemi plovacích měchýřků, víceletá, konce větví s bradavičnatými receptakuly, na jejich povrchu ústí dutinek - konceptákulí, feromon fukoseraten, oplození na dně mimo konceptakulum, dvoudomý Ascophyllum nodosum - válcovitá stélka, v místech plov. měchýřů zbytnělá Sargassum (bentické a pelagické druhy) S. fluitans a S. natans - Sargasové moře, S. filipendula přisedlé Lithoderma fluviatile - jako jediná chaluha byla nalezena i u nás, vlákna rostoucí kolmo na podklad tvoří na kamenech v potocích a řekách husté hnědé nárosty, hlavně v horách, ale kdysi byla nalezena i ve Vltavě a Sázavě v okolí Štěchovic TŘÍDA: RŮZNOBRVKY XANTOPHYCEAE (TRIBOPHYCEAE) Preferovaný název Xanthophyceae charakterizuje nejnápadnější vlastnost skupiny xanthos, z řečtiny žlutý, žlutozelený podle převládající barvy buněk způsobené kombinací pigmentů (chybí fukoxantin). V této třídě jsou téměř všechny organizační stupně stélky od monádoidní po trubicovitou. Převládají kokální, vláknité a sifonální druhy. Mnoho společných znaků se zlativkami, ale chybí fukoxantin. BS celulózní, amorfní složka tvořená alginovými kyselinami a hemicelulózou. Monadoidní různobrvky a zoospory mají 2 heterokontní bičíky, z nichž delší je péřitý. U r. Vaucheria při nepohlavním rozmnožování vzniká mnohojaderná synzoospora s četnými páry bičíků na celém povrchu, jsou téměř stejně dlouhé a mají zřejmě hladký povrch. Pohlavní proces - oogamie. Kromě zoospor vznikají při nepohlavním rozmnožování též aplanospory. U některých druhů jsou v životním cyklu oba typy výtrusů. V přírodě jsou různobrvky častou, ale nepříliš významnou součástí společenstev. Vyskytují se ve sladkovodních biotopech i v moři. Z vývojového hlediska je pozoruhodný paralelismus ve tvaru stélky mezi různobrvkami a zelenými řasami (např. Characiopsis x Chlamydopodium, Tribonema x Microspora) odlišení na základě reakce s Lugolovým roztokem. Zástupci 44
47 Myxochloris - rhizopodová stélka, z fylogenetického hlediska významný rod, připomíná plazmodium hlenek (vývojová paralela), žije v hyalocytech rašeliníků (u nás v posledních letech nezjištěna) Characiopsis kokální, podobá se zelené řase Characium (Chlamydopodium), přisedá na různé podklady pomocí tenkých stonků Tribonema vláknité stélky, v čistých, mírně kyselých tůních, nevětvená vlákna, BS ze 2 válcovitých dílů, které mají uprostřed kruhovou přehrádku, při pozorování v mikroskopu mají tvar ležícího písmene H (nepravá vlákna) Botrydium granulatum - sifonální, mnohojaderná stélka (1-2 mm) vyrůstá na vlhké půdě na březích řek a rybníků, v půdě se uchycuje pomocí rozvětvených bezbarvých rhizoidů, nadzemní vakovitá část s chloroplasty Vaucheria (posypanka) - zvláštní místo mezi různobrvkami, oogamie, trubicovitá, bohatě větvená stélka, přirůstající k podkladu bezbarvými rhizoidy, žije na vlhké půdě na dně potoků a mělkých tůní. Sifonální stélka byla srovnávána s hyfami hub tř. Oomycetes, podobnost též v průběhu mitózy, úvahy o vývojové příbuznosti. TŘÍDA: EUSTIGMATOPHYCEAE Jednobuněčné řasy s odlišnou stavbou zoospor: pouze jeden péřitý bičík (pleuronematický s tripartitními mastigonemy), ale přítomno i druhé bazální tělísko (2. bičík zřejmě během vývoje redukován). Chybí věncová lamela. Stigma mimo plastid. Sladkovodní biotopy, půda, méně v mořích. TŘÍDA: ROZSIVKY BACILLARIOPHYCEAE (DIATOMAE) Rozsivky jsou patrně druhově nejpočetnější skupinou řas, v přírodě velmi rozšířené. Jsou to jednobuněčné kokální řasy žijící samostatně nebo v koloniích, jejich buňka je uzavřena do dvoudílné buněčné stěny (frustuly), která má tvar kruhové krabičky s radiální souměrností (centrické rozsivky) nebo podlouhlé krabičky s bilaterální souměrností (penátní rozsivky). Stavební materiál frustuly je amorfní polymer oxidu křemičitého, blízký opálu, vnější i vnitřní povrch frustuly pokrývá ochranná polysacharidová vrstva (diatotepin). V buňkách je jediné diploidní jádro. Rozmnožování - dělením buněk, vede ke zmenšování rozměru buňky, původní rozměry se obnoví následkem pohlavního procesu. Rozsivky jsou jedinou skupinou organismů naprosto závislou na přítomnosti rozpustných forem oxidu křemičitého v prostředí. Jsou významnými primárními producenty, vytvářejí 25 % veškeré biomasy produkované rostlinami (obrovská produkce např. na spodní straně arktických a antarktických ledových ker). Populace rozsivek ovlivňují svou aktivitou řadu biochemických procesů na planetě. Stavba buněčné stěny - frustuly Křemitá frustula, hlavní znak rozsivek, je složená ze 2 přibližně stejných ték, zapadajících do sebe jako dno a víko krabičky. Vrchní epitéka, spodní hypotéka. Plocha misky - valva (valvární pohled), boční pás - pleura (pleurální pohled). Pásy se překrývají a tvoří bok frustuly. Na vnitřní ploše pleury bývají křemité výběžky do nitra buňky (septa, Tabellaria). Valva má složitou druhově specifickou strukturu, sestávající z pravidelně uspořádaných systémů pórů, komůrek a žeber. U centrických rozsivek je paprsčitě uspořádaná. Penátní rozsivky mají 2 souměrné systémy oddělené osovým polem (střední žebro), které je buď celistvé (dříve označováno pseudoraphe), nebo bývá proraženo štěrbinovou raphe. Raphe má význam pro 45

48 pohyb buňky, který se projevuje klouzáním po povrchu podkladů. Mechanismus pohybu: stahy kontraktilních filamentů v povrchové plazmě jsou převáděny na podklad pomocí vylučovaného slizu. Štěrbinová raphe - probíhá ve směru podélné osy buňky, uprostřed je ztlustlina, centrální nodulus, na obou koncích - polární noduly. Na příčném průřezu má štěrbinová raphe tvar ležatého písmene V. Kanálková raphe - tvar septované trubice, umístěné blízko okraje valvy (eventuálně v odsazených kýlech), spojené otvory s vnitřním prostorem buňky. Stavba protoplastu Protoplast obsahuje velkou vakuolu, která reguluje osmotické poměry v buňce, další organely jsou v nástěnné vrstvě plazmy (centrické r.) nebo jádro ve středu buňky a 2 vakuoly po stranách (penátní r.). Chloroplasty centrické r.: několik, penátní r.: 1-2. Diktyozómy se podílejí na stavbě frustuly a produkují sliz. Dělení buněk a pohlavní rozmnožování Rozmnožování - dělením buňky na 2 dceřiné, každá dceřiná buňka zdědí polovinu BS, která je epitékou, nově se tvoří hypotéka, to vede k postupnému zmenšování buněk. Při tvorbě nové valvy dochází k intenzivnímu příjmu kyseliny křemičité z prostředí. Po dosažení kriticky malých rozměrů buňky dojde k pohlavnímu procesu nebo buňky odumírají. Pohlavní proces - u centrických - oogamie (vaječná buňka a 4 spermatozoidy s 1 péřitým bičíkem). U penátních izo- či anizogamie, kopulace protoplastů 2 dceřiných buněk (po meiotickém dělení). Zygota roste v auxosporu, během jejího růstu se vytvoří iniciální buňka, která má obě téky nové a zahajuje posloupnost nových generací vegetativních buněk. Pohlavnímu procesu předchází meiotické dělení. Výskyt v přírodě a hospodářský význam Rozsivky - nejrozšířenější skupina řas, podílejí se čtvrtinou na celkové primární produkci, osidlují nejrůznější biotopy: hlavně vodní prostředí, ale i povrchová vrstva půdy, vlhké skály, termální vody, aeroplankton. Většinou kosmopolitní rozšíření, ale i endemické druhy (Ochrid: Cyclotella fottii). Tvoří druhově bohatá společenstva planktonu a perifytonu. U nás hlavně na jaře a na podzim. V mořích převažují centrické (po vyčerpání křemíku klesají ve shlucích do hlubin, mořský sníh, odkud se za příznivých podmínek opět vynoří nebo klesnou až ke dnu), ve 46
49 sladkých vodách penátní druhy. 1 druh je toxický (potrava slávky jedlé, otravy lidí neurotoxinem), některé druhy jsou potravou chytridiomycetů. Význam pro vodohospodářskou praxi - složení společenstev citlivě reaguje na znečištění odpadními vodami (sledování trofie a saprobity vody), tomu napomáhá relativně snadná určitelnost jednotlivých druhů. Využití diatomární analýzy v paleobotanice. Jako forenzmí nástroj v kriminalistice: testuje se přítomnost rozsivek v tělech utonulých. Rozsivky byly nalezeny v kostní dřeni a srovnávány s druhy zjištěnými v předpokládaném místě utonutí. Oceánologie - stenotermní rozsivky, mapování teplých a studených proudů. Biotechnologie - získání olejů. Sedimentující schránky tvoří mocné vrstvy na dně moří a jezer vznik křemeliny (diatomit). Zřejmě první průmyslové využití diatomitu představovala výroba dynamitu (A. Nobel), dnes jako filtrační materiál, ve farmacii jako absorbční materiál, izolace Struktura frustuly se používá k testování optických mikroskopů (struktura Pleurosigma angulatum musí být zřetelná již při použití 40x zvětšujícího objektivu, pro silnější objektivy se užívá druhů Surirella gemma a Amphipleura pellucida). (Pozn.: trofie: množství živin v prostředí, eutrofizace: zvyšování trofie, tj. úživnosti vody, saprobita: množství organických látek, které jsou schopny rozkladu, tj. stupeň znečištění vody) Geologické stáří Spolehlivé nálezy z druhohor, raná křída, 120 mil let. Nejstarší - centrické. V příznivých podmínkách vznikaly vrstvy sedimentované horniny diatomitu (kvartérní sedimenty). Metamorfózou vzniká porcelanit a rohovec. Systém: podle souměrnosti misek a přítomnosti raphe dělení na 3 podtřídy: Coscinodiscophycideae, Fragillariophycidae, Bacillariophycidae PODTŘÍDA: COSCINODISCOPHYCIDEAE (BÝVALÝ ŘÁD CENTRALES) Centrické rozsivky s kruhovými, eliptickými, trojúhelníkovitými či polygonálními miskami s radiální souměrností, vždy bez raphe a středního žebra. Ooogamie, mořský a sladkovodní plankton. Stephanodiscus -jednobuněčné, krátce válcovité, jarní plankton Melosira - dlouhé vláknité kolonie, sliz spojuje plochu misek, plankton, perifyton Coscinodiscus - mořské, ploché kruhové misky Triceratium - trojúhelníkovité misky Chaetoceras valvy nesou 2 nebo více ostnů, ty spojují buňky do řetízkovitých kolonií, nejhojnější mořská planktonní rozsivka PODTŘÍDA: FRAGILLARIOPHYCIDAE (ARAPHIDINEAE) Bilaterálně souměrné misky se středním žebrem, raphe chybí. Tabellaria - plankton i perifyton, silně zkřemenělá vnitřní septa Asterionella - hvězdicovité kolonie Fragilaria, Synedra, Diatoma PODTŘÍDA: BACILLARIOPHYCIDAE (MONO- A BIRAPHIDINEAE) Bilaterálně souměrné frustuly se štěrbinovým či kanálkovým raphe na jedné či obou miskách - možnost klouzavého pohybu. Některé druhy přisedlé. Eunotia, Cocconeis, Achnantes, Navicula, Pinnularia, Cymbella, Pleurosigma, Nitzschia, Surirella 47


50 TŘÍDA: CHLOROMONÁDY RAPHIDOPHYCEAE (CHLOROMONADOPHYCEAE) Malá skupina bičíkovců s dorziventrální stavbou buňky. Břišní strana lehce zploštělá, zde vyrůstají dva nestejně dlouhé bičíky, z nichž jeden je péřitý. Buňky pokryty periplastem (zpevněná plazmatická membrána), pod ním jsou vymrštitelná tělíska, trichocysty a sliz produkující mukocysty. Vnitřní plazma obsahuje jádro s čepičkou z diktyozómů, vnější plazma obsahuje vakuoly a jasně zelené chloroplasty, v nichž byl nalezen nukleomorf. Buňky jsou citlivé na změny prostředí, snadno praskají. V přírodě vzácné, jejich výskyt je periodický, v současnosti hojnější, bývá spojován s acidifikací vod: čisté rybníky a tůně s mírně kyselou reakcí vody (Vacuolaria, Gonyostomum), moře (Fibrocapsa). Oddělení HAPTOPHYTA (PRYMNESIOPHYTA) Relativně malá skupina co do počtu druhů, ale zejména mořští zástupci mají značný ekologický význam. Bičíkovci, kapsální a krátce vláknité řasy (zde bičíkovci jsou stádiem životního cyklu). Výživa mixotrofní. Monofyletická skupina, donorem plastidu zřejmě ruducha. Bičíkový aparát: 2 bičíky a haptonema. Povrch bičíků je hladký nebo s velmi jemným vlášením místo tubulárních mastigonem. Haptonema je útvar podobný bičíku, na průřezu z 5 7 mikrotubulů obklopených cisternou endoplazmatického retikula, která zřejmě obsahuje toxiny. Neslouží k pohybu, povrch hladký, může být dlouhá, kratičká či kontraktilní, uplatňuje se při přesunu potravních částic do buňky nebo k uchycení na hostiteli. 2 skupiny: 1) zástupci mající na povrchu buňky polysacharidové šupiny (celulózní), 2) vápenatí bičíkovci mající na povrchu buňky zvápenatělé šupiny (kokolity) z CaCO 3. Kalcifikace probíhá současně s fotosyntézou, a proto je závislá na množství světla. Při plném světle, kdy probíhá intenzivní fotosyntéza, zvýší se ph vody z 8 na 8,3, tehdy je ve vodě nadbytek hydrogenuhličitanových iontů a kalcifikace probíhá dle rovnice: 2 HCO Ca 2+ CaCO 3 + CO 2 + H 2 O čím vyšší je intenzita fotosyntézy, tím intenzívnější je kalcifikace kokolitů a buňka získává více CO 2 pro fotosyntézu. Odumřelé buňky a nadbytečné kokolity přispívají k tvorbě mořského sněhu a propadávají na dno oceánu. Tvar kokolitů je druhově specifický, velikost 1-3 μm, jako mikrofosilie mají význam ve stratigrafii (srovnání sedimentů stejného stáří) a jako bioindikátory 48
51 při hledání ropných ložisek. Haptophyta mají zásadní význam pro fungování globálního ekosystému (cyklus C a S) sedimenty kalcitu, vazba CO 2, produkce dimetylsulfoxidu k zajištění osmotické rovnováhy buňky (únik do atmosféry, kyselé deště). Výskyt: v moři, brakické vodě, sladkovodní zástupci jsou ojedinělí. Řády Prymnesiales a Phaeocystales - 2 bičíky a haptonema, na povrchu těla a bičíků polysacharidové šupiny, vznikající v diktyozómu Chrysochromulina - v nanoplanktonu našich rybníků Prymnesium - v brakických jezerech, při přemnožení produkuje toxiny nebezpečné pro ryby, odpovídá za hromadný úhyn ryb na lososích farmách v Norsku v r , frekvence výskytu toxických vodních zákalů vzrostla s eutrofizací vody Phaeocystis - buňky tvoří slizové kolonie až několik cm velké, žije v jižních i severních mořích, slizem ucpává rybářské sítě, znehodnocuje pláže, působí též úhyn ryb, zřejmě ucpáním žaber, nikoli toxiny; při fotosyntéze vzniká dimetylsulfid, který dodává rybám zápach shnilé kapusty (nepříznivý vliv na tah sleďů) Řád: Coccolitophoridales tzv. vápenatí bičíkovci, na povrchu těla mají zvápenatělé šupiny kokolity, odpovídají za podstatnou část převodu C a Ca mezi atmosférou a oceanickými sedimenty dna (eliminace CO 2 v povrchových vodách a syntéza CaCO 3 ). Už v geologické minulosti vytvořili rozsáhlé sedimenty křídy a vápence. Tvoří 25% celkového ročního vertikálního transportu uhlíku do hlubin oceánu. Emiliania huxleyi tvoří vegetační zákaly (mléčné) v S Atlantiku o ploše km 2 a hloubce 60 m, odraz světla od této plochy zvyšuje albedo a to má ochlazující vliv na místní klima. Hymenomonas roseola - druh, který žije v rašelinných tůních i u nás Oddělení CRYPTOPHYTA - SKRYTĚNKY, KRYPTOMONÁDY Malá, dobře definovaná skupina bičíkovců žijících v kontinentálních vodách i v mořích. Chloroplast (s nukleomorfem) vznikl sekundární endosymbiózou, donorem byla jednobuněčná ruducha. Skrytěnky jsou fotoheterotrofní bičíkovci se 2 bičíky s podílem osmotrofní a fagotrofní výživy. Buňka je dorziventrálně stavěná. Povrch pokrývá periplast, sestávající z vnější plazmatické membrány a tenkých hexagonálních destiček bílkovinné povahy pod ní. Mezi destičkami jsou drobné trichocysty. Na břišní straně ústí vakovitý jícen, směřující šikmo do buňky, na stěnách jícnu jsou ejektozómy, vymrštitelná tělíska (obranné nebo únikové struktury: vymrštění vyvolá pohyb buňky v opačném směru). Na předním konci buňky - 2 bičíky téměř stejně dlouhé, delší se dvěma řadami vlášení, kratší má pouze jednu řadu. Specifickou strukturou bičíkového aparátu je rhizostyl (podélně spojené mikrotubuly), který směřuje od bazálních tělísek k zadnímu konci buňky, jeho funkcí je zřejmě upevnění bičíkového aparátu v buňce. Chloroplasty mají na povrchu 4 membrány, tylakoidy srostlé po 2. Pigmenty: chlorofyly a+c, α- a ß-karoten, xantofyly (alloxantin), fykobiliny (krypto-)fykoerytrin a (krypto-)fykocyanin (umístěné ve vnitřním prostoru tylakoidů, neexistují fykobilisomy). V periplastidálním prostoru nukleomorf. Zásobní látka: škrob (i u heterotrofních druhů). Kryptomonády žijí v moři i ve sladkých vodách. V našich podmínkách tvoří 1. maximum biomasy v rybnících záhy zjara. Jsou to chladnomilné organismy, ve studených a hlubokých vodách tvoří podstatnou složku fytoplanktonu. V antarktických jezerech trvale pokrytých ledem tvoří v létě až 70% biomasy planktonu. Tvoří jev zvaný hloubkové maximum chlorofylu - na 49


52 hranicích oxické a anoxické zóny se shromažďují fotosyntetické organismy, kterým nevadí nízká hladina světla, protože mají pigmenty schopné ji využít, v této zóně je méně konkurentů i predátorů než na hladině a živin je dostatek. Některé jsou toxické. Cryptomonas nejhojnější sladkovodní rod, hnědé chloroplasty Rhodomonas červenohnědý (převládá fykoerytrin) Chroomonas modrozelený (převládá fykocyanin) Chilomonas - apochlorický, ztráta pigmentů je druhotná zastíněná lesní jezírka a rašelinné tůně HETEROTROFNÍ SKUPINY ŘÍŠE CHROMALVEOLATA Primární heterotrofie je zřejmá u oddělení Labyrinthulomycota. Předpoklad, že u oddělení Peronosporomycota a Hyphochytriomycota došlo k druhotné ztrátě chloroplastu, není prokazatelně doložen. Dnes převládá názor, že i tyto skupiny jsou primárně heterotrofní a vyvíjely se paralelně s autotrofními. Zásobní látka mykolaminaran. 1-2 bičíky, jeden se dvěma řadami tubulárních mastigonem. Oddělení LABYRINTHULOMYCOTA - LABYRINTULY, VODNÍ HLENKY Malá skupina nepříliš dobře probádaných organismů vázaných na mořské biotopy. Zástupci jsou saprotrofové i parazité řas a vyšších rostlin. Dříve řazeni mezi hlenky, prvoky, houby i řasy. Trofickou fázi tvoří buď pohyblivé vřetenovité či elipsoidální buňky nebo nepohyblivé kulovité stélky. Charakteristickým znakem je vytváření ektoplazmatických výběžků, které se spojují v síťovité útvary. Jsou produkovány botrozomy (sagenogeny), specifickými organelami na povrchu buněk. Oddělení HYPHOCHYTRIOMYCOTA Nepočetná skupina převážně půdních či sladkovodních saprofytů a parazitů, morfologicky podobná zástupcům odd. Chytridiomycota, ale v ultrastrukturních a biochemických znacích podobná odd. Peronosporomycota. 50
53 Oddělení PERONOSPOROMYCOTA (OOMYCOTA) - OOMYCETY, ŘASOVKY Oomycety jsou dosti početná a významná skupina vodních a suchozemských organismů, které jsou saprotrofní nebo fakultativně i obligátně parazitické. Primitivní, obligátně parazitické druhy mají stélky endobiotické (žijící uvnitř těla hostitele), intra- nebo intercelulární. Jejich stélka je eukarpická (pouze část stélky se přemění v rozmnožovací orgán) a monocentrická (ze stélky vzniká 1 sporangium), ojediněle i holokarpická (celá stélka se přemění v rozmnožovací orgán). U pokročilejších typů tvoří stélku vláknité, větvené, nepřehrádkované (coenocytické, mnohojaderné) mycelium (eukarpické polycentrické stélky), přehrádky oddělují pouze rozmnožovací orgány (sporangia a gametangia). Intracelulární endobiotičtí parazité mají amorfní stélky bez BS. Vláknitá mycelia mají stěny tvořené celulózou a dalšími polysacharidy. Zásobní látkou je β-1,3-polyglukan nazývaný mykolaminaran, podobný chrysolaminaranu, ne glykogenu. Od hub se liší i cestou syntézy lysinu přes DAP, houby ji syntetizují přes AAA, hlavním sterolem je fukosterol (u hub je to ergosterol). Nepohlavní rozmnožování je velmi rozmanité. Zoospory: většina zástupců tvoří 1 typ - pleurokontní (sekundární zoospory, ledvinité, s bičíky umístěnými laterálně), takové druhy označujeme jako monomorfní, jev, tj. tvorbu 1 pohyblivého stádia jako monoplanetismus. Někteří zástupci tvoří nejprve akrokontní zoospory (primární, hruškovité, bičíky v přední části buňky) a po encystaci dávají vznik sekundárním zoosporám, druhy dimorfní, diplanetismus. Ty mohou opět encystovat (u nejpokročilejších typů) a vyrejdí opět sekundární zoospory (polyplanetismus) nebo vyklíčí aplanospory (aplanetismus). U suchozemských parazitických druhů (řád Peronosporales) dochází k redukci výskytu zoospor ve prospěch tvorby jednosporových sporangií, která jsou schopna klíčit hyfou. Pohlavní rozmnožování: oogametangiogamie - nedochází k tvorbě volných gamet, ale samčí jádra po kontaktu anteridia s oogoniem přecházejí kopulačními kanálky do oogonia. Anteridium kyjovité, na speciální anteridiální větvi a je hormonálně přitahováno k oogoniu. Oogonia jsou kulovitá či oválná, obsahují 1 nebo více oosfér. Oospora (zygota) vyklíčí v klíční hyfu, na jejímž konci se buď vytvoří zoosporangium nebo se hyfa rozvětví a vyrůstá v novou diploidní stélku. Životní cyklus je u většiny zástupců diplobiontický, k meióze dochází při tvorbě gametangií. Vývojové trendy v oddělení Peronosporomycota představují obecný směr evoluce, a to přechod z vodního prostředí na souš, ten je doprovázen: 1) potlačením tvorby zoospor ve prospěch jednosporových sporangií, klíčících hyfou (od diplanetismu k aplanetismu) 2) přechodem od saprotrofismu k biotrofnímu parazitismu (ztráta některých biosyntéz) až specializace na hostitele a orgány (organotropie). Původ: pravděpodobně apoplastická větev pravěkých chromalveolat se sifonální stélkou (třída Xanthophyceae). Odlišnosti od ostatních skupin chromalveolat: nepřítomnost pohyblivých (volných) samčích gamet (proto oogametangiogamie). Ekologie - dvě základní skupiny: 1) Vodní saprotrofní i parazitické organizmy. Saprotrofní na mrtvém hmyzu, bezobratlých i obratlovcích. Parazité na korýších, vířnících, vodním hmyzu i rybách. 2) Suchozemské typy především biotrofní parazité cévnatých rostlin, ale i půdní saprotrofní zástupci, případně původci hnilob kořenových částí rostlin. 51


54 Obr.: Životní cyklus dimorfního vodního zástupce r. Saprolegnia a rostlinného parazita Plasmopara. Hospodářský význam: několik významných fytopatogenních druhů, též původci skládkových chorob kořenové zeleniny, dosud nevyužité možnosti druhů, parazitujících na komárech resp. jejich larvách. Systém a zástupci: nejdůležitější rody jsou zastoupeny v řádech Peronosporales, Pythiales a Saprolegniales. Řád: Peronosporales sporangiofory charakteristického tvaru, oogonia s jedinou oosférou, organotropie, zástupci jsou obligátní parazité suchozemských rostlin. Symptom napadení: olejové skvrny na líci napadených listů, na rubu bělavý povlak sporangioforů, vyrůstajících z průduchů Peronospora (vřetenatka) větvené sporangiofory se zašpičatělými koncovými větévkami, nejznámější je druh P. brassicae v. kapustová, napadající brukvovité rostliny a působící např. hnilobu hlávek zelí, dále P. destructor na cibuli nebo P. tabacina parazitující na tabáku Pseudoperonospora (vřetenatka) má dva důležité druhy: P. humuli je nebezpečný parazit chmele, do Evropy se dostala z Dálného východu zhruba před 100 lety, a dále druh P. cubensis (v. okurková) parazitující na rostlinách tykvovitých Plasmopara (vřetenatka), sporangiofory větvené s tupě zakončenými větvemi, P. umbeliferarum je velmi hojný parazit na listech miříkovitých (např. na Aegopodium podagraria). Ekonomicky významným druhem je P. viticola (v. révová), působící nekrózy na listech révy 52

55 vinné (zavlečení do Evropy z Ameriky s podnoži odolnými vůči révokazu, ochrana měďnatými přípravky, např. tzv. bordeauxskou jíchou ). Životní cyklus je diplobiontní, zoosporangia roznášena větrem a vodou, v rose či po dešti vyklíčí na další hostitelské rostlině sekundární zoospory, ty klíčí v hyfu, která průduchem pronikne do listu (za určitých podmínek, vyšší teplota, nižší vlhkost, klíčí zoosporangia přímo hyfou), mycelium se rozrůstá a tvoří nové sporangiofory k šíření nákazy, pohlavní proces v listech během podzimu (redukční dělení při tvorbě oosfér a samčích jader v anteridiu), tlustostěnné oospory jsou obnaženy po zetlení listů a s kapkami deště vymršťovány na listy, klíčí kratším vláknem na konci se zoosporangii Bremia lactucae je nebezpečným parazitem semenáčků a mladých rostlinek hlávkového salátu Albugo napadá laskavcovité, hvězdnicovité, brukvovité, A. candida (plíseň bělostná) je častá na kokošce aj., polštářovitá ložiska pod pokožkou hostitele, krátké nahuštěné sporangiofory s řetízky zoosporangií: Řád: Pythiales sporangia se netvoří na specializovaných hyfách, sporangiofory se příliš neliší od vegetativních hyf, jedna oosféra v oogoniu, půdní a vodní saprofyti a fakultativní či obligátní paraziti Phytophtora s nejvýznamnějším druhem P. infestans (plíseň bramborová), který napadá listy brambor, ale i rajčata. Listy žloutnou, nekrotizují a postupně usychají celé nadzemní části. Za vlhkého a chladného počasí se infekce rychle šíří a přechází i na bramborové hlízy. Do Evropy choroba zavlečena v polovině 19. století. V letech následkem neúrody brambor umřel v Irsku až 1 mil lidí, zároveň masové vystěhovalectví do Ameriky. Ochrana pomocí fungicidů a šlechtěním resistentních odrůd. P. parasitica napadá rajská jablka. Pythium zahrnuje převážně půdní saprotrofní druhy, ale další druhy způsobují i padání klíčních rostlin (P. debaryanum, P. ultimum) nebo hniloby kořenů, včetně hnilob skladované kořenové zeleniny. P. oligandrum hyperparazit, využití v kosmetice, biologické ochraně rostlin (Polyversum), stavebnictví. Řád: Saprolegniales - zástupci jsou většinou saprofyté na rostlinných i živočišných zbytcích ve sladkých vodách, vzácněji paraziti řas, drobných vodních živočichů, někteří žijí v půdě či parazitují v kořenech kulturních rostlin. Dimofrismus, polyplanetismus, aplanetismus, oogonia obsahují více oosfér. Saprolegnia (hnilobytka) - druh S. parasitica může parazitovat na rybách (včetně akvarijních), jikrách, žábách i vodním hmyzu, S. monoica je hojný saprotrof na mrtvém hmyzu, rostlinném detritu ve vodě, ale i v půdě. Některé druhy rodu Aphanomyces (hnileček) jsou parazité rostlin, včetně kulturních (hniloby kořenů cukrovky, brambor, ředkviček), popř. některých živočichů i řas. A. cochlioides působí hnilobu kořenů řepy cukrovky, A. euteiches napadá kořeny vikvovitých a brambor, A. astaci 53
56 původce tzv. račího moru. Ke studiu: ŘÍŠE: OPISTHOKONTA (incl. FUNGI) Houby jsou eukaryotické heterotrofní (výživa absorpční, osmotrofní, tedy heterotrofní, a to nikoli fagotrofní) stélkaté organismy (lišejníky díky autotrofnímu fotobiontu sekundárně autotrofní), které se rozmnožují výtrusy (sporami). V buněčné stěně většinou převládá chitin, nemají plastidy, chybí fotosyntetické pigmenty, jsou přítomny karoteny a xantofyly odlišné od rostlinných, dále melaniny, fenoly, produktem metabolismu je zpravidla glykogen. Důležitým znakem, kterým se liší houby od ostatních organismů, je syntéza AK lyzinu. Živočichové ji syntetizovat nedovedou. Houby ji syntetizují přes α-aminoadipovou kyselinu (AAA dráha syntézy), všechny ostatní organismy přes diaminopimelovou kyselinu (DAP dráha). Tyto dvě metabolické cesty jsou zcela rozdílné, což svědčí o tom, že se houby vyvinuly nezávisle, a to v dávných dobách historického vývoje. Vegetativní stélka: u nejjednodušších typů jednobuněčná (nevláknitá), u pokročilejších typů vláknitá, ale neseptovaná (coenocyt), u většiny pak stélku tvoří septované, vláknité mycelium s více- dvou- či jednojadernými úseky. U kvasinek je stélka dimorfická (hyfy i kulovité buňky tvořící pučivé pseudomycelium). V určitých stádiích tvoří hyfy nepravá pletiva (plektenchym, pseudoparenchym): rhizomorfy pevné myceliální svazky, plodnice, stromata kompaktní myceliální struktury, v nichž se tvoří plodnice, sklerocia přetrvání nepříznivých podmínek. Výživa (heterotrofní, absorpční): houby odkázány přímo či nepřímo na autotrofní organismy (mají v ekosystémech úlohu dekompozitorů), buď jako paraziti (živiny z živých buněk rostlin či živočichů), hyperparaziti (paraziti na houbách), a to závazní (obligátní) nebo příležitostní (fakultativní) nebo jako saprofyti (výživa z odumřelých těl rostlin či živočichů), popř. jako symbionti (mykorhiza, lichenismus). Časté přechody: saproparazitismus - parazitické houby, přežívající na odumřelém hostiteli. Produkce extracelulárních enzymů: rozklad vysokomolekulárních látek tak, aby je bylo možno absorbovat přes buněčnou stěnu. Biotrofní paraziti nezpůsobují přímé odumírání pletiv nebo hostitelských rostlin, protože jsou schopni se vyživovat pouze z živých buněk (sněti, rzi ). Nekrotrofní paraziti působí odumírání napadených pletiv a část jejich životního cyklu se tedy odehrává na odumřelém substrátu, je již vlastně saprotrofní. Mykorhiza vyskytuje se u 95% rostlin, není pouze u rostlin vodních a ruderálních. Mycelium, zvyšuje rostlině příjem živin nejen tím, že intenzivně kolonizuje půdu, ale i tím, že aktivní systémy buněčných membrán hub jsou schopné hromadit látky proti koncentračnímu gradientu. Houba též pomáhá rostlině v příjmu vody. Rostlina zásobuje houbu monosacharidy. Ektomykorhiza houbová vlákna obalují drobné kořínky rostliny, na jejichž povrchu vytvářejí kompaktní plášť. Asi 2000 druhů rostlin, hlavně dřeviny: smrky, duby, buky, jedle, borovice, břízy Hlavně bazidiomycety (2000 druhů). Mezi druhem houby a rostliny dochází často k úzké specializaci (klouzek modřínový). Endomykorhiza houbová vlákna pronikají do mezibuněčných prostor i do buněk, kde vytvářejí charakteristické útvary vezikulo-arbuskulární m. (vezikuly, arbuskuly), erikoidní m. (klubíčka). Známa asi u 1000 druhů rostlin, ale předpokládá se až u druhů, mezi něž patří řada zemědělsky významných plodin, př. orchideje, trávy, ale i olše, jasany. Asi 130 druhů hub, převážně glomeromycota. 54

57 Nepohlavní rozmnožování: u řady druhů dominantním způsobem rozmnožování, nepohlavní rozmnožovací cyklus může proběhnout několikrát během sezóny, vznikne velké množství jedinců (důležité u parazitů). Anamorfa, mitosporická houba: houba se rozmnožuje nepohlavně. Teleomorfa, meiosporická houba: pohlavní stádium. Obě stádia popisována dříve často jako samostatné druhy, mnohé druhy známy dosud jen jako anamorfy, jedná se o tzv. mitotickou holomorfu, druh se vyskytuje pouze v konidiálním stádiu. Meiotická holomorfa, druhy, u nichž je známo jen pohlavní rozmnožování. Pleomorfická holomorfa, houba, která má oba typy rozmnožování. Holomorfa = celá houba, tj. jedinec určitého druhu s jeho dosud známým reprodukčním vybavením. Pojmenování holomorfy podle jména teleomorfy. Zoospory pouze u Chytridiomycota, 1 bičík, hladký, bez mastigonemat, u ostatních oddělení aplanospory. Sporangiospory - vznikají endogenně ve sporangiích (Mucoromycotina). Častěji se tvoří konidie - na specializovaných hyfách (konidioforech) z konidiogenní buňky. 2 základní typy vzniku konidií (konidiogeneze): a) thalický (artrický) - konidie vzniká z již existující buňky hyfy, která se rozdělí přehrádkami a rozpadne na jednobuněčné části, holoblastický typ (účast všech vrstev BS) enteroblastický (vnější stěna BS se na tvorbě konidie neopdílí), např. arthrospory a též chlamydospory (tlustostěnné klidové spory vznikající interkalárně či terminálně a jsou jedno- či vícebuněčné) b) blastický - konidie vzniká vypučením z konidiogenní buňky, např. porospory - vyhřeznou z póru a následně vytvoří BS, fialospory - vznikají ve fialidách, lahvicovitých buňkách s ústím v apikální části, anelospory - přepážka oddělující nově vzniklou konidii je protrhávána následně vznikající konidií, zbytky protržených stěn tvoří límečky u ústí konidiogenní buňky. Konidiofory se nacházejí izolovaně na různých místech na myceliu nebo se seskupují v určité útvary (konidiomata): 55
58 sporodochium (ložisko) - poduškovitý útvar hyf, na němž jsou hustě palisádovitě uspořádány konidiofory (vždy na povrchu hostitele) acervulus (klubíčko) - útvar podobný předešlému, je však ponořený v pletivu hostitele, alespoň pod kutikulou a v době zralosti vystoupí na povrch koremie - svazek dlouhých, vzpřímených, navzájem slepených konidioforů, které se na vrcholku sromkovitě větví pyknida - kulovitý či lahvicovitý útvar s ostiolem (ústím), na povrchu hostitele i ponořený, vnitřek vystlán konidiofory, konidie bývají spojeny slizovitou hmotou a vytlačovány ven jako slizová pentlice Pohlavní rozmnožování: spojeno se změnou ploidie, vždy musí dojít k plazmogamii, karyogamii a meióze. Zvláštností většiny hub je oddálení plazmo- a karyogamie (dikaryofáze, dikaryon - 2 kompatibilní jádra ve společné plazmě, konjugované - současně probíhající - mitózy). Typy pohlavního procesu u hub velmi různorodé, nejčastější jsou: a) gametogamie Chytridiomycota, izo-, anizo- i oogamie b) gametangiogamie - izo-, anizo- i oogametangiogamie, častá, Mucoromycotina, Ascomycota c) gameto-gametangiogamie - spermatizace, častá, Ascomycota d) somatogamie - hlavně hyfogamie, Basidiomycota, vzácně hologamie, Saccharomycotina e) gameto-somatogamie - spermatizace u Uredinales U hub se setkáváme se všemi dosud známými životními cykly, i když pro většinu je charakteristická redukce diploidní fáze na 1 buňku (zygota, zygospora, askus, bazidie). Životní cyklus převážně haploidní nebo dikaryotický, vzácně i diploidní. Druhy jednodomé, dvoudomé i pohlavně nediferencované. Druhy homo- i heterothalické. Haplobionti Chytridiomycota, Mucoromycotina, mikrocyklické Uredinales. Diplobionti - vzácní - některé druhy řádu Saccharomycetales. Haplo-diplobionti vzácní, např. izomorfická rodozměna u Saccharomyces cerevisiiae, heteromorfická u Protomyces. Haplo-dikaryonti - prakticky všichni zástupci Pezizomycotina, rzi. Dikaryonti - téměř všichni zástupci Basidiomycota. Výskyt: všudypřítomné, rozšíření globální, dosud popsáno kolem druhů hub, odhadovaných až 1,5 milionu. Výrazně převažuje množství mikroskopických druhů hub nad makroskopickými. Význam v přírodě (dekompozitoři) i pro člověka je obrovský. Odedávna houby využívány ke kynutí těsta, kvašení piva, vína, konzumaci plodnic. V průběhu 20. století rozvoj biotechnologií: antibiotika i další léčiva, enzymy, kyselina citrónová, vitamíny, biologická ochrana rostlin, rozklad látek znečišťujících životní prostředí. Naopak parazitické houby snižují úrodu, saprofyti znehodnocují skladované potraviny a krmiva, některé houby způsobují onemocnění živočichů i člověka. Původ: není dosud spolehlivě vysvětlen (např. askomycety od ruduch, Sachs 1874, Demoulin 1975), dříve jako polyfyletická skupina (houby v původním širokém pojetí). Vznik z předků prvoků (pravděpodobně společný původ s trubénkami - Choanoflagellata). Pokročilejší skupiny pravděpodobně vznikly z předků chytridií před cca 550 miliony let, k oddělení hub vřeckovýtrusných a stopkovýtrusných mohlo dojít přibližně před 450 miliony let. Nejstarší 56

59 prokázané fosilní doklady pocházejí z pozdního siluru (paleozoikum, cca 420 milionů let, tedy společně s prvním hmyzem a rhyniofyty). Náplň této skupiny doznala v posledních desetiletích velkých změn, především převedením některých zástupců, které tradičně studuje mykologie, do říší Excavata, Amoebozoa, Rhizaria, Chromalveolata. Současné členění na 7 oddělení a 4 pododdělení neurčitého zařazení (Hibbett et al. 2007): CHYTRIDIOMYCOTA jednobuněčné či cenocytické stélky, bičíkatá stádia přítomna, dikaryofáze chybí BLASTOCLADIOMYCOTA - nejprimitivnější skupina hub s rodozměnou, často značně diferencovaná stélka se rhizoidální částí NEOCALLIMASTIGOMYCOTA - vysoce specializovaní anaerobní obyvatelé zažívacího traktu někt. býložravců MICROSPORIDIOMYCOTA obligátní parazité bez buněčné stěny, spory s chitinózní stěnou GLOMEROMYCOTA - coenocytické mycelium s chitinem v buněčné stěně, hlavní endomykorhizní houby SKUPINA HUB SPÁJIVÝCH ZYGOMYCOTA coenocytické mycelium, bičíkaté formy a dikaryofáze se nevyskytují. Dnes není tato skupina uznávána na úrovni oddělení, ale byla rozdělena na 4 pododdělení neurč. zařazení: Mucoromycotina, Entomophthoromycotina, Zoopagomycotina a Kicxellomycotina. ASCOMYCOTA mycelium přehrádkované, chybějí bičíkatá stádia, dikaryofáze přítomna, endogenní meiospory BASIDIOMYCOTA mycelium přehrádkované, chybějí bičíkatá stádia, výrazná dikaryofáze, exogenní meiospory 57
60 Oddělení CHYTRIDIOMYCOTA CHYTRIDIOMYCETY Zřejmě vývojově nejstarší (nejpůvodnější) skupina říše Fungi, vykazující některé primitivní znaky, se kterými jsme se setkali u předcházejících skupin. Dnešní zástupci mohou mít společný původ s trubénkami (Choanoflagellata) Vývojově vyšší skupiny jsou odvozovány od prapředků chytridiomycetů: centrum tzv. bazálních linií hub (Rozella). Odlišnost od ostatních skupin říše Fungi: přítomnost pohyblivých zoospor. Stélka jednobuněčná (holokarpní, popř. eukarpní) s rhizoidy či rhizomyceliem, nejodvozenější je stélka tvořená cenocytickým myceliem, přehrádky oddělují pouze reprodukční orgány, BS chitinové. Rozmnožování - nepohlavní pomocí jednobičíkatých (bičík není péřitý) opistokontních zoospor, zoosporangia operkulátní (otvírají se víčkem) či inoperkulátní (otvírání většinou štěrbinou). Pohlavní: u jednotlivých skupin dosti rozdílné, u některých rodů existuje dikaryotická fáze. Výskyt: primárně vodní organismy, hojné i v půdě, některé jsou obligátními parazity jiných chytridiomycetů, cévnatých rostlin, hmyzu, mnohé jsou saprofyty. Význam: parazité kulturních rostlin (Synchytrium endobioticum na bramborách, Olpidium brassicae na rostlinách brukvovitých). Z hlediska fylogeneze nutno zmínit nové znaky: oogamie, poprvé a naposledy u heterotrofů (Monoblepharis), operkulátní typy sporangií (Chytridium, Chytriomyces). Systém zahrnuje 2 třídy: Monoblepharidomycetes (odvozenější) a Chytridiomycetes (původnější). Zástupci: Synchytrium (rakovinec), všechny druhy jsou parazity cévnatých rostlin, S. endobioticum (r. bramborový) způsobuje na hlízách v místě oček bujení pletiv a vznik tmavých bradavčitých nádorů, výsledně hnilobu celé hlízy, napadá i stonky, listy, květy, rakovina brambor je přísně sledovaná fytokaranténní choroba (ohlášení, zákaz obhospodařování, po určitou dobu zákaz vstupu na pozemek, zničit všechny zástupce čel. Solanaceae v okolí). Zajímavost: rakovina se nevyskytuje v Andách, v domovině brambor, v Evropě se asi přenesla na brambory z planě rostoucích hostitelů (Solanum nigrum, S. dulcamara). Pohlavní proces je izogamie, stélka je primitivní, endobiotická a holokarpická. Výskyt hlavně ve vlhčích chladnějších oblastech. Díky velmi odolným trvalým sporangiím může v půdě přežívat i více než 20 let. Šíření patogena nemocnými nebo kontaminovanými hlízami, rostlinnými zbytky, zamořenou půdou ulpívající na mechanizaci, odpadními produkty ze zpracujícího průmyslu atp. Ochranná opatření: likvidace trvalých sporangií chemickými přípravky není možná především z ekologického hlediska. Doporučují se preventivní opatření: více než tříletý odstup v pěstování brambor na pozemku, používání pouze zdravé sadby z nenapadených oblastí, pěstování rezistentních odrůd. Životní cyklus haplobiontický: z odpočívajících sporangií vyklíčí na jaře jednobičíkaté zoospory, které se dotknou hlízy v místě očka, zatáhnou bičík a obklopí se buněčnou stěnou, po proniknutí do buněk hostitele protoplast roste a obklopí se dvojitým žlutohnědým obalem, vnikne tak letní výtrus - prosorus, v okolních buňkách dochází k neorganizovanému dělení a vzniku nádoru. Prosorus klíčí, probíhá jeho dělení a vznikne 4-9 částí, tj. sorus sporangií. Stěna hostitele praskne, sporangia se uvolní a vyrejdí zoospor, které se pohybují v půdní vodě a šíří infekci dále. Za suchého počasí se zoospory chovají jako gamety, kopulují a vzniklá zygota infikuje hostitele. V buňce hostitele vznikne odpočívající (trvalé) sporangium, které přetrvá zimu. 58
61 S. anemonae - na sasankách, S. mercurialis - na bažance, S. taraxaci na Asteraceae Rhizophydium pollinis-pini roste na pylových zrnech borovic tlejících ve vodě, často nalezen v kalužích v době dozrávaní pylu, stélka monocentrická, eukarpní, epibiotická Olpidium - parazité řas, hub a cévnatých rostlin, O. brassicae - působí padání klíčních rostlin brukvovitých (nekróza hypokotylu) Batrachochytrium - jediný chytridiomycet napadající obratlovce, onemocnění pokožky žab chytridiomykóza. Od konce 80. let minulého století vyhynulo asi 120 druhů obojživelníků, řadu z nich má na svědomí houba Batrachochytrium dendrobatidis. O její rozšíření se významně přičinil člověk - roznášel nevědomky chorobu po světě s nakaženými africkými drápatkami Xenopus laevis. Houba se živí keratinem v pokožce, kůže žab zasažená houbou nedokáže transportovat ionty, klesá koncentrace Na a K v krvi až je rovnováha iontů v těle narušena natolik, že srdce žab přestane bít. 59
 Oddělení BLASTOCLADIOMYCOTA Houby s rodozměnou (Allomyces a Blastocladiella), střídání hostitelů (Coelomomyces), objekty pro studium")
62 Druhy rodu Chytridium napadají řasy, monocentrická, eukarpní, epibiotická stélka, operkulátní zoosporangia Monoblepharis (ze třídy Monoblepharidomycetes) - pohlavní rozmnožování oogamie, druh běžný na organickém materiálu ve vodě (dřevo, bylinné zbytky) Oddělení BLASTOCLADIOMYCOTA Houby s rodozměnou (Allomyces a Blastocladiella), střídání hostitelů (Coelomomyces), objekty pro studium morfogeneze, cytologie, genetiky (Allomyces, Blastocladiella), biologický boj (Coelomomyces), fytopatologicky významné druhy (Physoderma). Coelomomyces - parazité v coelomových dutinách larev komárů rodů Culex a Anopheles, u jednoho druhu zjištěno střídání hostitelů (komár a buchanka) Allomyces - saprotrofové, ve vlhkých půdách tropických a subtropických oblastí, často pěstován v laboratořích Blastocladiella - saprotrofové, na zbytcích hmyzu ve vodě nebo žijí v půdě Physoderma alfalfae - nádory na kořenech a stoncích vojtěšky, rakovina vojtěšky Physoderma maydis - hnědá skvrnitost kukuřice Oddělení NEOCALLIMASTIGOMYCOTA Neocallimastix, Orpinomyces, Anaeromyces obligátní anaerobní saprobionti, rozkladači zbytků potravy v zažívacím traktu mnoha druhů velkých býložravců, s bakteriemi jsou primárními rozkladači vlákniny. Vytvářejí monocentrickou nebo polycentrickou mnohojadernou stélku. Rozmnožují se zoosporami nesoucími většinou 4 bičíky, vzácně je bičíků více. Životní cyklus je nedostatečně znám. Ekonomicky vysoce významná skupina, má také velký biotechnologický potenciál (enzymy). Oddělení MICROSPORIDIOMYCOTA MIKROSPORIDIE Malá skupina vysoce specializovaných jednobuněčných, druhotně zjednodušených, obligátních intracelulárních parazitů živočichů s charakteristicky utvářenými sporami. Morfologicky výrazně odlišné organizmy od ostatních jednobuněčných zástupců hub. Původně považované za kmen prvoků, příslušnost k houbám byla zjištěna na základě molekulárněbiologických znaků. Spory mikrosporidií, které jsou schopny přežívat mimo hostitele, mají v buněčné stěně chitin, buňky nemají mitochondrie. Spory mikrosporidií obsahují: 1 2 jádra, 3 specifické organely, které nemají ani prvoci ani jiné houby: pólové trubicovité vlákno (spirálně stočené podél vnitřní stěny spory), polaroplast (sloupec membrán a měchýřků), útvar podobný vakuole. 60

 a N. ceranae jsou významní parazité včel, Nosema bombycis způsobuje hynutí larev bource morušového.")
63 Vegetativní stádium: obsah spory se dostane do hostitelské buňky injekčním mechanismem v podobě primitivní buňky (sporoplazma), která vyrůstá v útvar meront, ten se uvnitř buněk hostitele množí, infikuje další buňky a vytvářejí se spory s chitinózní stěnou. Spora se aktivuje chemickým podrážděním v zažívacím traktu hostitele a vystřelí pólové vlákno do buňky. Obsah spory (sporoplazma) tak přejde kanálkem ve vláknu tlakem vakuoly do hostitele. Výskyt, význam, zástupci: parazité hlavně hmyzu, ryb, ale i obratlovců včetně člověka. Nosema apis (hmyzomorka včelí, je původcem včelí úplavice, nosematózy) a N. ceranae jsou významní parazité včel, Nosema bombycis způsobuje hynutí larev bource morušového. Encephalitozoon a Enterocytozoon napadají ptáky, savce i člověka (při snížení imunity např. oslabením virem HIV). Oddělení GLOMEROMYCOTA Zástupci vytvářejí s mnoha druhy rostlin vesikulo-arbuskulární, resp. arbuskulární mykorhizu (VAM, AM, endomykorhiza), u většiny kulturních rostlin - zásobování hostitele fosforem, 80% všech známých endomykorhiz. Coenocytická stélka, nepohlavní rozmnožování pomocí chlamydospor (mnohojaderné, jednotlivé nebo obalené hyfami ve sporokarpu). Výskyt: ve všech obdělávaných půdách, pod povrchem půdy, na kořenech rostlin, na povrchu půdy. Rody Glomus, Gigaspora. Průmyslové využití: např. Symbivit (6 druhů rodu Glomus) pro zlepšení zdravotního stavu pěstovaných rostlin. Geosiphon pyriforme - endosymbióza s Nostoc punctiforme - jediný známý případ endocyanózy u hub, význam z hlediska evoluce mykorhiz. Stélka v podobě zduřelého vlákna, v přírodě nacházen velmi zřídka, na zemi, zatím cca 6 lokalit v Evropě. SKUPINA HUB SPÁJIVÝCH ZYGOMYCOTA Polyfyletická skupina, charakterizovaná především dvěma znaky: typem pohlavního rozmnožování (gametangiogamie), jehož výsledkem je zygosporangium s jedinou zygosporou a tvorbou rozsáhlého, převážně nepřehrádkovaného mycelia, u některých vývojově pokročilejších zástupců už ale dochází i k tvorbě sept ve vegetativním myceliu. Bičíkaté formy a dikaryofáze se nevyskytují. Dnes není tato skupina uznávána na úrovni oddělení, ale byla rozdělena na 4 pododdělení neurčitého zařazení: Mucoromycotina, Entomophthoromycotina, Zoopagomycotina, Kicxellomycotina. 61
64 Pododdělení MUCOROMYCOTINA Nejznámější, nejčetnější a největší skupina spájivých hub. Vegetativní mycelium je dobře vyvinuté a rozsáhlé, většinou nepřehrádkované (coenocytické), větvené, mnohojaderné, haploidní. Lze rozlišit rhizoidy a stolony. BS je chitinózní. Septa se tvoří při oddělování rozmnožovacích orgánů. Často jde o velmi rychle rostoucí organismy. Nepohlavní rozmnožování pomocí endogenních nepohyblivých sporangiospor, které se tvoří ve sporangiích na koncích specializovaných hyf (sporangioforů). Sporangiofor je ve sporangiu zakončen jako tzv. kolumela (střední sloupek). Po rozpadu stěny zralého sporangia zůstává její malá část kolem sporangioforu jako tzv. límeček. Vývojový trend směřuje od velkých sporangií s mnoha sporangiosporami k menším sporangiím s malým počtem sporangiospor (merosporangia) až k jednosporovým sporangiím (sporangioly). Pohlavní rozmnožování je izo- nebo anizogametangiogamie. Gametangia na stejných či různých myceliích (druhy homo- a heterothalické). Mnohojaderná gametangia jsou nesena rozšířeným koncem hyfy (suspensorem) a oddělena septem. Po konjugaci se vytvoří zygosporangium, obsahující jedinou zygosporu. Kolem zygosporangia se může vytvářet hyfový obal (ochranná vlákna, vyrůstající ze suspenzorů), snad jako vývojový základ plodnic (nejpokročilejší u Endogonaceae). V zygospoře dochází redukčnímu dělení a po klidovém období klíčí hyfa, nesoucí klíční sporangium. Životní cyklus je tedy haplobiontický. Význam: produkce významných metabolitů (výroba kyseliny mléčné (Rhizopus stolonifer, R. arrhizus, R. oryzae), kyseliny fumarové (Rhizopus stolonifer), citronové, šťavelové apod., sojová omáčka). Naopak: znehodnocování především sladkých potravin (ovoce, džemy, pečivo). V humánní medicíně mají význam druhy, působící plicní nebo kožní a podkožní zygomykózy, v ojedinělých případech i napadení sliznic nebo mozkových plen, gastrointestinální infekce (Rhizopus oryzae, R. microsporus, R. azygosporus, Mucor ramosissimus, Absidia corymbifera, Conidiobolus coronatus). Evoluční trendy: absence bičíkatých stádií, redukce počtu sporangiospor ve sporangiu, Endogone - ektomykorhizní nebo saprotrofní, pravděpodobní předkové vřeckovýtrusých hub (přehrádkování mycelia, oddálení plazmogamie a karyogamie, sporokarp). 62
65 Výskyt: v půdě, na ovoci, na rostlinných zbytcích, na výkalech (Pilobolus, Mucor mucedo), na kloboukatých houbách. Mohou působit onemocnění obratlovců včetně člověka - zygomykózy. Mucor - jednotlivé sporangiofory, mnohosporová sporangia, půdní saprofyté, M. mucedo (plíseň hlavičková), na koňském trusu a tlejících organických substrátech Rhizopus - stolony a svazečky konidioforů, zkvašování cukrů, řada druhů produkuje pro člověka významné sloučeniny: kys. fumarová, mléčná, citronová, šťavelová, výroba orientálních potravin, R. stolonifer (kropidlovec černavý), působí hniloby skladovaného ovoce Zygorhynchus - rozdílně velká gametangia, v půdě Syzygites - vidličnatě větvené sporangiofory, porůstá plodnice stopkovýtrusých hub (Boletus, Russula, Lactarius) Pilobolus (měchomršt) koprofilní, v subsporangiálním vaku se hromadí voda, zvýšení tlaku, prasknutí, houba se rozšiřuje odmrštěním celého sporangia ve směru největší světelné intenzity Choanephora cucurbitarum - hniloba a odumírání květů a mladých plodů tykví, okurek, paprik Syncephalastrum výskyt v půdě, oportunní patogen člověka (dermatomykózy a onychomykózy), merosporangia (pozn.: oportunní patogeny vyvolávají onemocnění většinou jen u hostitele s významnou poruchou imunity) Pododdělení ENTOMOPHTHOROMYCOTINA Některé druhy parazitují na rostlinách, jiné na bezobratlých (na hmyzu - Entomophthora, Conidiobolus, jiní na vířnících, želvuškách, hlísticích, houbách). Mycelium je coenocytické jen v mládí. Tendence k rozpadu hyf na tzv. hyfová tělíska. Tvoří se tzv. blastokonidie, což je terminální sporangium, které se po dopadu na nevhodný substrát může rozpadat na sekundární konidie. Entomophthora muscae (hmyzomorka muší, působí muší mor) - sporangium na povrchu mouchy vyklíčí ve vlákno, proroste do hostitele, rozrůstá se a rozpadá na hyfová tělíska, postupně enzymatickou činností houby rozrušeny všechny orgánové soustavy, moucha usedne a uhyne, hyfová tělíska prorostou ven a vytvoří konidiofory, lepkavá sporangia jsou odmršťována až 2 cm kolem mouchy Conidiobolus - fakultativní parazité hmyzu či hub, některé druhy jsou patogenní pro člověka - onemocnění podkoží Oddělení ASCOMYCOTA - VŘECKOVÝTRUSÉ HOUBY Největší skupina říše Fungi, 50% všech hub (kolem rodů a druhů). Velká morfologická i ekologická rozmanitost a tedy i rozsáhlá terminologie. Do této skupiny náleží většina lichenizovaných hub. Společný znak: tvorba vřecek (asci, ascus), u většiny zástupců je vřecko jedinou diploidní buňkou v cyklu, je meiosporangiem, dochází v něm k meiozi a diferenciaci endogenně vznikajících askospor (nejčastěji 8). Charakteristická je pro mnoho zástupců přítomnost dikaryofáze ve formě askogenních hyf. Převažující typ ŽC haplodikaryotický. Nepohlavní část životního cyklu = anamorfa, pohlavní část životního cyklu = teleomorfa, organismus jako celek = holomorfa. Morfologické, časové a lokální oddělení anamorfy a teleomorfy přináší důsledky taxonomické (dvě i více různých jmen). Systematické zařazení založeno na teleomorfě, pojmenování holomorfy podle jména teleomorfy. Pleomorfní holomorfa v ŽC je pohl. i nepohl. rozmnožování Meiotická holomorfa v ŽC je pouze pohl. rozmnožování 63
66 Mitotická holomorfa - v ŽC je pouze nepohl. rozmnožování Synanamorfy v ŽC 2 různé typy nepohlavního rozmnožování Deuteromycety (Fungi imperfecti) umělá skupina pro houby nepohlavně se rozmnožující Systematické uspořádání vřeckovýtrusých hub prošlo v minulosti velkými změnami a bylo založeno na kombinaci typu plodnice a typu vřecka. V současné době na základě především molekulárních a ultrastrukturálních znaků je oddělení členěno na 3 pododdělení (Taphrinomycotina, Saccharomycotina, Pezizomycotina) a celkem 16 (4, 1, 11) tříd. Uvedený systém není možno považovat za finální, lze očekávat další změny a upřesnění podle nově získaných informací. Pododdělení TAPHRINOMYCOTINA Zahrnuje vývojově původní, nejstarší typy, skupina bazálních askomycetů. Morfologicky a ekologicky nesourodá skupina, jejíž zástupci byli v minulosti na základě nefylogenetických znaků řazeni do různých taxonomických skupin. Chitin v BS většinou chybí nebo je v nepatrném množství. Anamorfy mají kvasinkovitý charakter, je přítomno jednak haploidní, jednak dikaryotické (Taphrina) nebo diploidní (Protomyces) mycelium. Třídy: Neolectomycetes, Pneumocystidomycetes, Schizosaccharomycetes, Taphrinomycetes TŘÍDA: NEOLECTOMYCETES Saprotrofové, malá monotypická skupina (pouze rod Neolecta), bazální malá skupina pro pododdělení Pezizomycotina a Saccharomycotina. Pravděpodobně důležitá vývojově. Neolecta jediný rod z celého poddodělení taphrinomycotina, který tvoří plodnice. TŘÍDA: PNEUMOCYSTIDOMYCETES Monotypická skupina s jediným rodem Pneumocystis, který byl v minulosti považován za prvoka. Nemyceliální stélka. Běžný obyvatel plicního epitelu savců včetně člověka, v případě silného oslabení imunity se přemnoží, vyplní plicní alveoly a dochází až ke smrtelnému zánětu plic (pneumocystóza). Objev lidské pneumocystózy je významným přínosem české vědy. Prof. Otto Jírovec s kolegy identifikovali v r Pneumocystis (dnes P. jiroveci, syn. P. carinii) jako původce epidemicky se vyskytujícího onemocnění kojenců (nedonošenců a oslabených dětí). V současnosti je výskyt choroby u kojenců u nás ojedinělý. TŘÍDA: TAPHRINOMYCETES Patří sem dva biotrofně parazitické řády. Řád: Protomycetales Parazité cévnatých rostlin s intercelulárním septovaným mnohojaderným diploidním myceliem schopným pučení. Na myceliu se tvoří tlustostěnné askogenní buňky (chlamydospory), které po přezimování vytvoří mnohojaderný měchýřek, který je považován za primitivní vřecko (někdy nazývané synaskus ). Po meioze vzniká velký počet haploidních askospor. Askospory vytvářejí saprofyticky žijící kvasinkovitá stádia, potom kopulují a dávají vznik diploidnímu vegetativnímu myceliu, schopnému infikovat hostitele. Nejznámějším druhem je Protomyces macrosporus (prvohouba velkovýtrusná), parazitující na řapících a čepelích listů bršlice (Aegopodium) i dalších rodech z čeledi Daucaceae, na nichž vytváří charakteristické drobné nádorky. 64
. Netvoří se plodnice, pohlavní rozmnožování: somatogamie, nepohlavní rozmnožování: blastogeneze, životní cyklus haplodikaryotický. Organotropnost, fytopatologický význam.")
67 Řád: Taphrinales Biotrofní vysoce specifičtí parazité dřevin, dimorfizmus stélek (dikaryotické vegetativní paraziticky žijící mycelium a schopnost tvořit i kvasinkovitou fázi, která žije saprofyticky). Netvoří se plodnice, pohlavní rozmnožování: somatogamie, nepohlavní rozmnožování: blastogeneze, životní cyklus haplodikaryotický. Organotropnost, fytopatologický význam. Důvody zařazení do samostatné systematické jednotky vysoké hodnoty: dikaryotická vegetativní fáze, nevytvářejí se askogenní hyfy, vřecka vznikají odlišným způsobem, schopnost tvorby haploidní kvasinkovité fáze. Taphrina deformans (kadeřavka broskvoňová) - životní cyklus: askospory se množí pučením, oddělují blastokonidie nebo dají vznik pučivému pseudomyceliu, které žije na povrchu hostitele. Z některé blastospory vyklíčí vláknité mycelium, jehož jádro se rozdělí (nebo blastospory kopulují), tak vznikne dikaryotické mycelium, které infikuje pletivo hostitele, rozrůstá se subkutikulárně nebo intercelulárně a vytváří nádorky. Některé buňky se zvětšují a vytvoří souvislou vrstvu chlamydospor (askogenních buněk). Zde dojde ke karyogamii, původní kulovité buňky se prodlužují a prorážejí kutikulu hostitele. Dochází k rozdělení buňky na 2 dceřiné, bazální (stopková) degeneruje a terminální se mění ve vřecko. Ve vřecku dojde k meiozi a následné mitóze a vzniká 8 askospor, které se množí pučením. Jediný rod Taphrina (anamorfa Lalaria). Zástupci jsou vysoce specifičtí parazité dřevin: Rosaceae, Salicaceae, Betulaceae, Corylaceae, Fagaceae, Ulmaceae, Aceraceae. Pletiva hostitele neumrtvují, ale podněcují k tvorbě hypertrofií a hyperplázií: hypertrofická kadeřavost listů (T. deformans, T. tosquinetii, T. populina), zmnožení větví a tvorba čarověníků (T. betulina), deformace plodů (T. pruni). T. cerasi (čarověníky a kadeřavost listů třešní a višní), T. deformans (kadeřavost listů broskvoní, do Evropy zavlečena z Číny, koncem 19. stol. již zde hojná), T. betulina (čarověníky na břízách), T. tosquinetii (listy olší), T. pruni (původce choroby, zvané puchrovitost švestek, 65
68 plody nedozrávají, hnědnou), T. amentorum (na květenství olše), T. johansonii (květenství osiky), Taphrina padi (deformace plodů střemchy - Prunus padus a třešně ptačí - Prunus avium) TŘÍDA: SCHIZOSACCHAROMYCETES Malá skupina, 1-buněčná stélka, morfologická podobnost s pravými kvasinkami (Saccharomycotina), od nichž se liší molekulárně genetickými znaky. Druh Schizosaccharomyces pombe se využívá ke zkvašování prosa při přípravě tzv. afrického piva pombe, ale i v některých biotechnol. procesech (např. biologický rozklad nafty), genetických a molekulárně-biologických studiích Pododdělení SACCHAROMYCOTINA (HEMIASCOMYCOTINA) Jediná třída s jediným řádem Saccharomycetales (Endomycetales), tzv. pravé kvasinky. Vegetativní stélka je nejčastěji ve formě jednotlivých pučivých buněk, některé druhy vytvářejí tzv. pučivé pseudomycelium, vzácně se vyskytuje i vláknité přehrádkované mycelium schopné tvořit blastospory. Buněčná stěna je (ultrastrukturálně) jednovrstevná (na rozdíl od skupiny Pezizomycotina, kde je dvouvrstevná) a je tvořena polysaridy typu β-glukan a β-mannan, chitin byl zjištěn pouze u některých typů. Nepohlavní rozmnožování tvorba dceřiných buněk (lze je nazývat blastospory, ale jsou to v podstatě konidie) pučením. Po odpadnutí dceřiných blastospor zůstávají na povrchu mateřské buňky zřetelné jizvy. Při nepohlavním rozmnožování může docházet i k prostému dělení buněk, u vláknitých typů se někdy tvoří arthrospory. Pohlavní rozmnožování nejčastěji somatogamie, tedy kopulace dvou vegetativních buněk chovajících se jako gamety. Vzniká diploidní zygota, která se (většinou) mění přímo ve vřecko. Životní cyklus může být haplobiontní, haplo-diplobiontní nebo diplobiontní. Výskyt a význam: převažují saprotrofní druhy na částech rostlin s vysokým obsahem cukrů (plody, nektaria, mízotoky), ale i na dřevě a v půdě. Schopnosti zkvašovat cukr na etanol a CO 2 se využívá v potravinářském průmyslu (výroba piva, vína, čajů), ale vzhledem k různým biosyntetickým vlastnostem mají zástupci využití i ve farmaceutickém a chemickém průmyslu. První využití kvasinek je dokumentováno již kolem r př. n. l. Systém a nejdůležitější zástupci: Saccharomyces tvoří jednotlivé oválné pučící buňky, nejznámějším druhem je S. cerevisiae (kvasinka pivní), která zkvašuje cukry (glukózu, maltózu, galaktózu) na alkohol, pekařství, výroba piva. Druh S. ellipsoideus (kvasinka vinná) se používá při výrobě vína. Jde o poměrně vysoce vyšlechtěný druh, původně izolovaný z plodů a květů révy vinné. Druh S. fragilis se používá při zkvašování mléčných výrobků. Druhy rodu Dipodascus (anamorfa Geotrichum) izolovány z mízotoku dřevin i z půdy, saprofyti, ale i oportunní patogeny člověka, Ascoidea z exsudátů rostlin a Endomyces z půdy, odpadních vod a mléčných produktů, se vyznačují tvorbou vláknitého mycelia. Pododdělení: PEZIZOMYCOTINA (ASCOMYCOTINA) Předchozí dvě pododdělení se vyznačovala: 1) primitivními vřecky s jednoduchou stěnou bez jakéhokoliv složitějšího otevíracího aparátu; 2) dikaryofáze buď nepřítomna, nebo měla jinou podobu než u skupiny Pezizomycotina; 3) naprostá většina zástupců nevytvářela plodnice. Naproti tomu u pododdělení Pezizomycotina: 66
69 1) vřecka, která vznikají na askogenních hyfách, můžeme na základě funkční struktury stěny a otvíracího aparátu dělit na prototunikántí, unitunikátní a bitunikátní; 2) dikaryotická fáze je přítomna ve formě askogenních hyf s konjugovanými mitózami, askogenní hyfy rostou ze samičího gametangia (askogonu) do oblasti výtrusorodé vrstvy (hymenia, thecia), životní cyklus haplo-dikaryotický; 3) vytvářejí se různé typy plodnic. Vegetativní stélka: tvořena vláknitým, větveným, dobře vyvinutým myceliem. Hyfy s přehrádkami s jednoduchými póry, v blízkosti póru jsou Woroninova tělíska, úseky jednojaderné i vícejaderné. Úplné přehrádky oddělují gametangia. Pozn.: segmentované mycelium představuje dokonalejší adaptaci k životu v terestrických podmínkách: je mechanicky pevnější než sifonální hyfa. Při poranění segmentu jsou sousední segmenty chráněny tak, že Woroninova tělíska ucpou póry v septu a zabrání úniku protoplasmy. Tvorba plektenchymatických pletiv v souvislosti s vývojem plodnic, stromat, sklerocií. BS dvouvrstevné, podstatnou složkou je chitin. Vřecko je vysoce specializované sporangium, z genetického pohledu jde o meiosporangium. V mladém vřecku dochází ke karyogamii, mladé vřecko je tedy zygotou, jedinou diploidní buňkou v životním cyklu. Ihned dochází k meioze, následné mitóze a výsledkem je 8 haploidních jader, po vytvoření buněčné stěny 8 endogenních spor, nazývaných askospory. Askospory jsou tedy endogenní meiospory. Ultrastrukturálně je stěna vřecka vícevrstevná, funkčně (tj. dle způsobu otvírání) rozlišujeme vřecka prototunikátní, unitunikátní nebo bitunikátní. Důležitým znakem je typ otvíracího aparátu vřecka (askoapikální aparát: AAA), barvitelnost stěny nebo otvíracího aparátu jodem (značí se J+: amyloidní nebo J-). a) Protunikátní (prototunikátní) vřecko má tenkou jednovrstevnou stěnu bez otvíracího aparátu, spory uvolňovány rozpadem nebo zeslizovatěním stěny vřecka. b) Unitunikátní vřecko má dvouvrstevnou stěnu členěnou v exoaskus a endoaskus. Obě vrstvy jsou tenké, navzájem spojené a při otvírání vřecka fungují jako 1 celek. Spory uvolňovány pórem či štěrbinou na vrcholu vřecka (inoperkulátní) nebo otvorem po odpadnutí víčka (operkulátní). Turgorem jsou vymršťovány na značnou vzdálenost. c) Bitunikátní (fissitunikátní) vřecko má dvouvrstevnou stěnu, před dozráním spor praskne exoaskus, endoaskus vyhřezne a prodlouží se ještě o délku vřecka, potom dojde k vymršťování spor. Pouze inoperkulátní. Životní cyklus a rozmnožování Životní cyklus je haplodikaryotický, haploidní je vegetativní mycelium a plodnice, dikaryotické jsou pouze askogenní hyfy, diploidní zygota. Pohlavní proces je nejčastěji gametangiogamie nebo gameto-gametangiogamie, kdy donorem samčího jádra je spermacie (samčí aplanogameta vznikající na spermacioforech nebo ve spermogoniích), popř. konidie, která ale neztrácí svou původní funkci v nepohlavním rozmnožování. Nepohlavní rozmnožování je hojné. Důležitým znakem je způsob vzniku konidií (konidiogeneze). Gametangiogamie (gametangie) - samčí větev je tvořená nosnou hyfou a válcovitým až kyjovitým anteridiem. Samičí větev je často mnohobuněčná a spirálně zkroucená, nese 1 nebo více askogonů, které jsou kulovité a nesou vláknité výrůstky - trichogyny. Gametangia jsou většinou mnohojaderná, někdy jednojaderná. Po kontaktu anteridia s trichogynem přechází do askogonu plazma a jádra z anteridia. Individuální vývoj: haplo-dikaryonti s výraznou haploidní a krátkou dikaryotickou fází: klíčením askospory vzniká haploidní mycelium, rozrůstá se, dělí se jádra, vznikají přehrádky, probíhá nepohlavní rozmnožování. V určitém období se založí gametangia, samčí jádra a plazma přejdou trichogynem do askogonu, dojde k plazmogamii a mísení, posléze párování jader. Na 67
. Pro většinu askomycetů je typické hákování hyf, terminální buňka se zakřivuje a vyvine se ve vřecko.")
70 askogonu vznikají výrůstky, které se stávají základem askogenních hyf, přecházejí do nich dvojice jader a tvoří se zde dikaryotické buňky. Askogenní hyfy přestanou růst na úrovni výtrusorodé vrstvy (hymenia, thecia). Pro většinu askomycetů je typické hákování hyf, terminální buňka se zakřivuje a vyvine se ve vřecko. V mladém vřecku dochází ke karyogamii. Následující meiozou a mitózou vzniká obvykle 8 haploidních jader (askospor). Plodnice (askomata, askokarpy) jsou tvořeny nepravými pletivy, obsahují pohlavní orgány, askogenní hyfy a vřecka. Stěna plodnice je peridie. Vřecka v plodnici mohou být neuspořádána nebo tvoří výtrusorodou vrstvu (hymenium, thecium). Sterilní hyfy v plodnici jsou např. perifýzy a parafýzy. Parafýzy - jsou protáhlé, někdy větvené hyfy, vyrůstající z báze plodnice a končící na úrovni hymenia. U některých rodů jejich vrcholky srůstají nad hymeniem v pevnou vrstvu (epithecium). Perifýzy - krátké vláskovité hyfy kolem ústí plodnice. Vývoj plodnic může probíhat dvěma způsoby: 1) askohymeniální ontogeneze: nejdříve dojde k pohlavnímu procesu a spolu s růstem askogenních hyf se vytvářejí i stěny plodnice. Tímto způsobem vznikají plodnice typu kleistothecium, apothecium a perithecium s prototunikátními nebo unitunikátními vřecky, 2) askolokulární ontogeneze: na myceliu se nejdříve vytvoří základ plodnice ve formě stromatického pseudoparenchymatického útvaru, ve kterém se diferencují gametangia, askogenní hyfy a vřecka vrůstají do sekundárně vytvořené lyzigenní dutiny (lokulu). Vlastní stěna plodnice se tedy netvoří, výsledná plodnice se nazývá askostroma a vřecka spojená s tímto vývojem jsou funkčně bitunikátní. 68

71 Typy plodnic: Kleistothecium - všestranně uzavřená plodnice, otvírá se zeslizovatěním či rozpadem stěny, uvnitř s nepravidelně rozptýlenými prototunikátními vřecky. Skupina s tímto typem plodnic se dříve nazývala PLECTOMYCETES. Apothecium otevřená miskovitá plodnice, někdy se stopkou, s výtrusorodou vrstvou (hymeniem) a unitunikátními operkulátními nebo inoperkulátními vřecky. Parafýzy mohou tvořit epithecium, pod hymeniem vrstva hustě propletených hyf (hypothecium), okraj je excipulum. Skupina s tímto typem plodnic se dříve nazývala DISCOMYCETES. Perithecium lahvicovitá plodnice s malým, determinovaným otvorem (ostiolem), vřecka unitunikátní, inoperkulátní, para- i perifýzy přítomny. Skupina s touto plodnicí se v minulosti nazývala PYRENOMYCETES. Další typy plodnic vřeckovýtrusých hub: askohymeniální vývoj protothecium (seskupení vřecek obalené spletí hyf), gymnothecium (přechod mezi proto- a kleistotheciem, primitivní stěna není tvořená pseudoparenchymem), tuberothecium (uzavření apothecia, uvnitř jsou komůrky a lišty) askolokulární vývoj - myriothecium (polštářovitá uzavřená plodnice s nepravidelně uspořádanými dutinami, v každé dutině 1 vřecko), pseudothecium (unilokulární askostroma, askolokulární plodnice s 1 dutinou), pseudoperithecium (podobné peritheciu s 1 nebo více dutinami, uspořádaná vřecka), pseudoapothecium (askolokulární paralela apothecia), hysterothecium (štěrbinovitě protáhlé apothecium, otvírá se podélnou štěrbinou). Výskyt a význam: všeobecně rozšířené houby, saprofyti či obligátní nebo fakultativní paraziti rostlin a živočichů, několik zástupců je mykorhizními houbami, asi třetina jsou lichenizované houby. Řada druhů je průmyslově využívána: zrání sýrů, produkce antibiotik, dalších léčiv, drog, konzumují se plodnice některých druhů. Systém: vnitřní systematika této velké skupiny je dosud neustálená. V minulosti se její zástupci členili na základě typů vřecek a plodnic na praktické skupiny - Plectomycetes (prototunikátní vřecka, kleistothecium), Discomycetes (unitunikátní vřecka, apothecium) a Pyrenomycetes (unitunikátní vřecka, perithecium). Po objevu askohymeniálního a askolokulárního typu vývoje plodnice se často používalo základní členění na skupiny - Ascohymenomycetes (askohymeniální vývoj a prototunikátní nebo funkčně unitunikátní vřecka) a Ascoloculomycetes (askolokulární typ ontogeneze a funkčně bitunikátní vřecka). V současné době na základě molekulárních znaků a integrace anamorf je skupina členěna na zhruba 9-12 tříd s několika desítkami řádů (kolem 40). Třídy jsou vymezeny na úrovni 69
72 morfologických, ontogenetických, ale především molekulárních znaků. Zde následuje výběr pouze několika reprezentativních tříd, řádů a zástupců. třída řád typ vřecka typ askomatu EUROTIOMYCETES Eurotiales p pr, kl h vývoj askomatu lichenizace poznámka EUROTIOMYCETES Onygenales p pr, kl h mazaedium PEZIZOMYCETES Pezizales u p, tu h operkulátní LEOTIOMYCETES Erysiphales u kl (? = pe) h LEOTIOMYCETES Helotiales u ap h LEOTIOMYCETES Rhytismatales u psap, hy (l) LECANOROMYCETES Gyalectales u ap h ano LECANOROMYCETES Lecanorales (u, b) ap h ano LECANOROMYCETES Pertusariales (u, b) ap h ano SORDARIOMYCETES Hypocreales u pe, kl h SORDARIOMYCETES Ophiostomatales p pe h SORDARIOMYCETES Xylariales psu, u pe h DOTHIDEOMYCETES Dothideales b pspe, my, CHAETOTHYRIOMYCETES Verrucariales b pspe, pe?h ano typ vřecka b bitunikátní p psb psp psu u protunikátní pseudobitunikátní pseudoprotunikátní pseudounitunikátní unitunikátní typ askomatu ap apothecium hy kl my pe pr psap pspe th tu hysterothecium kleistothecium myriothecium perithecium protothecium pseudoapothecium pseudoperithecium thyriothecium tuberothecium vývoj askomatu hy askohymeniální l askolokulární th l barevné stroma Řád: Eurotiales Plodnice typu gymnothecium (peridie je tvořena pouze nesouvislou sítí větvených hyf, např. u rodu Talaromyces) nebo kleistothecium (peridie pseudoparenchymatická, jedno- či vícevrstevná, např. u rodu Eurotium). Vřecka protunikátní, nejčastěji kulovitá, tenkostěnná, ve zralosti se rozplývající. 70
73 Nepohlavní rozmnožování v životním cyklu většinou převládá, konidiogeneze nejčastěji blastická, konidie jednobuněčné, fialospory uvolňované v řetízcích z lahvicovitých fialid, uspořádání fialid na konidioforu je typické pro jednotlivé anamorfní rody, z nich nejdůležitější jsou Penicillium, Aspergillus nebo Paecilomyces. Ekologie a význam: hojný výskyt s globálním rozšířením destruenti rostlinných i živočišných substrátů, produkce sekundárních metabolitů (toxinů). Využití v potravinářském průmyslu a medicíně (produkce antibiotik ale i nebezpečných toxinů, působení konidií jako alergenů), ekonomický význam při rozkladu uskladněných produktů, potravin a krmiv. Eurotium (anamorfa Aspergillus) v sušených potravinách a krmivech, půdní druhy. Řada druhů působí choroby zvířat i lidí - aspergilózy (A. fumigatus, A. flavus, A. niger, mnohé druhy patří k producentům mykotoxinů (karcinogenní aflatoxin B 1: A. flavus, sterigmatocystin: A. versicolor), produkce kyseliny citronové: A. niger, další druhy se využívají ke zkvašování sojových bobů, rýže, fermentaci kávy... Eupenicillium (anamorfa Penicillium) druhy půdní, osmofilní i termofilní. Konidiofory tvoří štětičku, nesou přeslenitě uspořádané metuly na nich přeslenitě uspořádané fialidy, mezi nejvýznamnější patří druhy produkující antibiotika: P. chrysogenum, P. griseofulvum, jiné se uplatňují při zrání sýrů: P. roquefortii, P. camembertii, další druhy jsou patogenní: P. italicum, P. digitatum, P. commune, P. expansum aj. Řád: Ophiostomatales Askomata jsou kulovitá či hruškovitá tmavě zbarvená perithecia, 1 mm velká, na vrcholku vytažená v dlouhé rostrum s ostiolem, vřecka protunikátní. Saprofyté nebo nepříliš specializovaní parazité kořenů, popř. vysoce specifičtí, obvykle nekrotrofní parazité stromů. Patří sem i druhy žijící v symbiózách s kůrovci ( ambrosiové houby ). Jsou roznášeny kůrovci, vegetují ve vodivých drahách stromů, způsobují jejich ucpání a tím usychání hostitele, navíc produkují toxiny. Onemocnění stromů se označují tracheomykózy: usychání konců větví nebo vrcholků stromů, žloutnutí a opad listů a pomalé odumírání celého stromu: Ophiostoma, Ceratocystis (anamorfy Graphium, Sporothrix, Chalara). Na jilmech O. ulmi, na dubech C. fagacearum a C. fimbriata. Řád: Erysiphales - padlí Mycelium porůstá povrch hostitele (extramatrikální mycelium), vytváří typický pomoučněný vzhled, hyfy pronikají haustorii do buněk hostitele. Nepohlavní rozmnožování: arthrospory na krátkých vzpřímených vláknech tvoří řetízky tzv. oidií. Askomata uzavřená, makroskopicky viditelná, hnědá až černá, jsou opatřená přívěsnými vlákny (apendixy), jejichž délka i tvar jsou rodově charakteristické. Plodnice považovány za kleistothecium, popř. primitivní erysiphální perithecium. Vřecka kulovitá či vejčitá, unitunikátní, tlustostěnná. Obligátní, vysoce specifičtí parazité cévnatých rostlin. Rozšiřování konidiemi, popř. plodnicemi na peří ptáků (apendixy). Ochrana rostlin: mechanické odstraňování napadených částí, výběr odolných kultivarů, preparáty obsahující síru. Podosphaera leucotricha, padlí jabloňové, anamorfa Oidium farinosum Sphaerotheca mors-uvae, americké padlí angreštové, S. pannosa, padlí růžové Erysiphe cichoraceum, padlí čekankové, hlavně na druzích z čel. Asteraceae, ale též na okurkách, tykvích aj., E. heraclei napadá druhy z čel. Daucaceae Microsphaera alphitoides, padlí dubové, plodnice nepříliš časté, většinou se tvoří anamorfa Oidium quercinum, M. grossularia, evropské padlí angreštové, není tak nebezpečné jako americké padlí 71
74 Blumeria graminis, padlí obilní, napadá jednoděložné rostliny Uncinula necator - padlí révy vinné, schéma životního cyklu: Řád: Pezizales (inkl. Tuberales) Saprotrofní operkulátní diskomycety (operculum je víčko). Askomata poměrně velká, přisedlá i stopkatá apothecia, miskovitá, kalichovitá, zprohýbaná, až 10 či 20 cm velká. Z ontogenetického hlediska mohou být plodnice gymnokarpní (od počátku otevřené), hemiangiokarpní (zpočátku uzavřené, později otevřené), angiokarpní (trvale uzavřené, otevírají se teprve tlakem zralých vřecek). Zástupci rodu Tuber a některé další rody mají tuberothecium, uzavřenou plodnici s komůrkami a lištami, která vzniká zprohýbáním a druhotným uzavřením apothecia, je hypogeická, hlízovitá, až přes 10 cm v průměru, povrch kryje peridie, uvnitř je gleba, výtrusorodá část. Vřecka unitunikátní. Nepohlavní rozmnožování méně rozšířené než u ostatních skupin askomycetů (u některých rodů není známo). Saprotrofní druhy půdní (často mykorhizní), koprofilní, antrakofilní, na mrtvém dřevě, na rostlinném opadu. Význam mají především druhy s jedlými plodnicemi (Tuber, Morchella, Verpa), naopak jedovaté jsou druhy r. Gyromitra. Sarcoscypha coccinea, ohňovec šarlatový, zářivě červené terče zjara nebo pozdě na podzim na trouchnivějících větvích listnáčů Aleuria aurantia, mísenka oranžová, velké oranžové plodnice na zemi, zvláště na lesních cestách Scutellinia scutellata, kosmatka štítovitá, ohnivě červená apothecia 5-10 mm, na okraji černě brvitá, na lesní půdě i trouchnivém dřevě Peziza badia, velká hnědá apothecia, hojně na písčité lesní půdě Rhizina undulata, kořenitka nadmutá, nepravidelné, ploché, hnědé, bíle olemované plodnice, až 7 cm velké, přirostlé četnými silnými vlákny k podkladu, na kořenech jehličnanů Morchella, smrž, kulovitý či kuželovitý klobouk s žebrovitými lištnami srostlý s třeněm, jedlé 72
75 Gyromitra, ucháč, mohutný, laločnatě zprohýbaný klobouk (apothecium), na dutém třeni, G. esculenta, u. obecný, zjara v písčitých borech, G. gigas, u. obrovský, až 30 cm široký, okrově hnědý klobouk, jedovaté Verpa, kačenka, zvoncovitý klobouk na delším třeni, V. bohemica, k. česká, zjara ve vlhkých hájích, jedlá Tuber, lanýž, podzemní, hlízovité, černé plodnice s kuželovitými bradavkami, se silně aromatickou dužninou, T. aestivum, l. letní, u nás vzácně v teplých oblastech na vápnité půdě, T. melanosporus pěstován ve Francii, sběr pomocí psů, ale i elektrických přístrojů, i k výrobě léků a voňavek Řád: Leotiales Inoperkulátní diskomycety, velmi početný řád. Nepohlavní rozmnožování časté: blastokonidie v pyknidách či acervulech. Apothecia plochá či miskovitá, stopkatá či přisedlá, od několika mm do 4 cm, často živě zbarvená, někdy vznikají na sklerociích či stromatech. Vřecka unitunikátní. Půdní a dřevní saprofyté, specializovaní parazité rostlin, lichenizované houby. Mollisia, terčenka, vosková či masitá apothecia, saprofyt na dřevě či rostlinných zbytcích Drepanopeziza ribis parazituje na větvích rybízu, anamorfa Gloeosporidiella ribis napadá listy (antraknóza rybízu) Chlorosplenium, zelenitka, zelená hniloba tlejícího dřeva, není hnilobou, ale jen zabarvuje dřevo kyselinou xylochlorovou Lachnellula willkommii, brvenka modřínová, 1-3 mm široká, oranžová, na okraji bíle brvitá apothecia. Výskyt souvisí se závažnou chorobou (nádory, úhyn stromů), rakovinou modřínů, houba je jedním z původců choroby nebo napadá nádory až sekundárně Dasyscyphus, chlupáček, apothecia zevně hustě chlupatá, na větévkách, stéblech trav a ležících listech Botryotinia fuckeliana, anamorfa Botrytis cinerea napadá listy a plody vinné révy, plody usychají nebo se pokrývají svazky konidioforů Monilinia, anamorfa Monilia fructigena působí moniliovou hnilobu plodů jabloní a hrušní, koncentricky uspořádané polštářky bělavých, postupně hnědnoucích konidií, napadený plod je prorůstán myceliem houby a stává se pseudosclerociem, u napadených jablek se někdy neobjevují konidiofory, ale napadený plod zčerná ( černá hniloba ) Mitrula paludosa, čapulka bažinná, kyjovité žluté až oranžové plodnice s bílou stopkou, na tlejících rostlinných zbytcích ve vodě Baeomyces, malohubka, lichenizovaná houba, fotobiont zelená řasa Coccomyxa, B. roseus, m. růžová, růžová stopkatá apothecia, na vlhké zemi, B. rufus, m. plšivková, hnědá apothecia Řád: Rhytismatales Askokarpy jsou pseudoapothecia (askolokulární paralela apothecia) či hysterothecia (štěrbinovitě protáhlé pseudoapothecium, otvírá se štěrbinou, hymenium se za vlhka široce obnažuje, za sucha uzavírá). Vřecka unitunikátní, inoperkulátní. Zástupci převážně parazité na listech či jehlicích. Lophodermium pinastri, sypavka borová, jeden z původců choroby zvané jehlosyp Colpoma quercinum, štěrbinatka dubová, saprofyt na dubových větvích Rhytisma acerinum, svraštělka javorová, parazit na listech, bioindikátor (obr.: stromata na listu s hysterothecii a vřecky) 73

76 Řád: Lecanorales (inkl. Peltigerales, Pertusariales, Teloschistales) Druhově nejpočetnější řád diskomycetů, patří sem nejméně polovina (7 500) všech známých druhů lichenizovaných hub. Všechny typy lišejníkových stélek, apothecia plochého diskovitého tvaru, někdy stopkatá. Kromě lišejníků sem patří několik druhů, které parazitují na lišejnících. Collema, huspeník, rosolovitá, téměř černá homeomerická stélka, kterou tvoří slizovitá tuhá hmota sinice r. Nostoc protkaná houbovými hyfami, většinou basifilní druhy Parmelia, terčovka, lupenitá stélka: P. saxatilis, P. sulcata, P. conspersa Pseudevernia furfuracea, lupenitá stélka s černou spodní kůrou, množství isidií, epifyt Hypogymnia physodes, terčovka bublinatá, lupenitá stélka, rtovité sorály, epifyt Cetraria, pukléřka, keříčkovitá nebo lupenitá stélka, u nás i v severské tundře běžná p. islandská (C. islandica), součást plicních čajů (species pulmonares) Usnea, provazovka, epifyt mizející vlivem znečištění ovzduší, keříčkovitá ( vousovitá )stélka Cladonia, dutohlávka, dimorfická stélka: lupenité přízemní šupiny a vystoupavé duté kmínky (podecia), C. pyxidata, C. coniocraea, keříčkovitá stélka bez přízemních šupin: C. rangiferina, C. arbuscula Umbilicaria, pupkovka, mohutná, lupenitá, v jednom místě k substrátu přirostlá stélka, U. hirsuta, U. (Lasallia) pustulata Candelariella, Lecidea, Pertusaria, druhy se stélkou korovitou Lecanora, misnička, korovitá stélka, výrazně toxitolerantní je L. conizaeoides Peltigera, havnatka, nápadná velká lupenitá stélka s výraznými vztyčenými apothecii Lobaria pulmonaria, důlkatec plicní, náš největší lupenitý lišejník, epifyt, silně ohrožený druh Xanthoria, terčovník, žlutá až oranžová lupenitá stélka, X. parietina, terčovník zední, nejčastěji na stromech u cest, drobnější podobné stélky má rod Caloplaca Rhizocarpon geographicum epilitický lišejník s korovitou stélkou a černými apothecii lecideového typu Řád: Hypocreales (inkl. Clavicipitales) Nepohlavní rozmnožování převažuje, nejčastější jsou fialospory. Někdy se tvoří sklerocia. Plodnice: živě zbarvená perithecia. Stromata též živě zbarvená, někdy připomínají plodnice stopkovýtrusých hub. Vřecka unitunikátní, inoperkulátní. Saprofyté na odumřelých rostlinných zbytcích (dřevě) nebo parazité rostlin, hub i larev hmyzu. Význam: někteří zástupci působí 74

77 onemocnění rostlin, Claviceps purpurea - alkaloidy, farmaceutika, C. paspali - LSD (v tropech pěstována na r. Paspalum). Claviceps purpurea, paličkovice nachová, parazit v semenících žita a jiných trav životní cyklus: askospory zachycené na bliznách prorostou do semeníku a vytvoří pseudostroma, v němž jsou komůrky vystlané konidiofory s konidiemi plovoucími v medovici (sfaceliové stádium dle anamorfy Sphacelia segetum), konidie jsou roznášeny hmyzem, napadený semeník se změní ve fialové sklerocium, námel, který vypadne z klasu a po přezimování na něm vyrostou stromata s bělavou stopkou a nachovou paličkou, palička obsahuje perithecia, niťovité askospory jsou vystřelovány z vřecek a unášeny větrem na další rostliny, byla-li sklerocia rozemleta do mouky, působila otravy (ergotismus) alkaloidy ergotaminové a ergotoxinové řady, dnes se námel uměle pěstuje pro výrobu léků proti krvácení Epichloe typhina, obalka stéblová, plíseň dusivá, bělavá stromata obalující stébla trav Hypomyces, nedohub, parazit askomycetů a bazidiomycetů (na rodech Elaphomyces, Boletus, Lactarius) Cordyceps, housenice, parazit na larvách hmyzu Nectria cinnabarina, rážovka ruměnná, rumělková perithecia na větvích listnatých stromů, častější je anamorfa Tubercularia vulgaris, hlívenka obecná, oranžové polštářky konidioforů N. galligena, spolupůvodce nektriové rakoviny na jabloních Řády: Xylariales a Diatrypales Typické stromatické tvrdohouby (pyrenomycety). Vřecka unitunikátní, inoperkulátní. Zástupci jsou saprofyty na dřevě či rostlinných zbytcích. Xylaria, dřevnatka, kyjovitá černá, několik cm vysoká stromata na pařezech či tlejícím dřevě listnáčů: X. polymorpha a X. hypoxylon Ustulina, spálenka, rozlitá černá stromata, U. deusta, s. skořepnatá, může napadat i živé stromy Diatrype, korovitka, černá, ploše rozložená nebo diskovitá stromata na listnáčích Hypoxylon, dřevomor, polokulovitá hnědá stromata (obr.: stroma a průřez stromatem s perithecii) 75

78 Řád: Dothideales Patří sem většina bitunikátních askomycetů. Vývoj plodnice askolokulárního typu: pseudoperithecia či jejich modifikace. Druhově velmi početný řád, široké spektrum substrátů, hlavně saprofyté a parazité rostlin. Některé rody tvoří tzv. černě, žijící saprofyticky na sladkých výměšcích hmyzu a rostlin na povrchu listů. Mohou pronikat haustorii do epidermálních buněk hostitele a přejít k parazitismu, ale v zásadě škodí zastiňováním asimilační plochy listů souvislým povlakem černého mycelia. Mycosphaerella, anamorfy způsobují skvrnitosti listů, M. tassiana má široké spektrum hostitelů, anamorfa Cladosporium herbarum působí chorobu označovanou jako čerň obilná, další druhy škodí na cukrovce, jahodníku, květáku, třešních... Herpotrichia nigra, plsťovka černá, obrůstá myceliem větve a jehličí smrků a kosodřeviny v místech, kde dlouho leží sníh Druhy rodu Venturia způsobují strupovitost ovoce, askospory na jaře infikují listy a mladé větévky, pod kutikulou se rozrůstá černé mycelium, tvořící konidiofory, konidie šíří nákazu na listy i plody, na podzim proroste mycelium do hloubky pletiv a vytvoří se gametangia a pseudoperithecia, V. pirina, anamorfa Fusicladium pirinum, strupovitost hrušní, V. inaequalis (F. pomi), strupovitost jabloní, V. cerasi (F. cerasi), napadá třešně a broskvoně Pseudogymnoascus destructans (anamorfa Geomyces destructans) způsobuje syndrom bílého nosu, choroba pojmenovaná podle bílých povlaků tvořících se zejména v obličejové části zimujících netopýrů, houba roste i na uších a křídlech zimujících netopýrů Řád: Verrucariales Lichenizované houby s korovitou, obvykle endolitickou stélkou, vzácně s lupenitou nebo nepatrně keříčkovitou. Plodnice jsou pseudoperithecia nebo perithecia, přisedlá či ponořená. Vřecka bitunikátní. Verrucaria, korovité lišejníky s endolitickou stélkou na vápencích, několik druhů je vodních Dermatocarpon fluviatile, lupenitá stélka, na kamenech v potocích s čistou vodou Oddělení BASIDIOMYCOTA - HOUBY STOPKOVÝTRUSÉ Velká, morfologicky i ekologicky značně diverzifikovaná skupina, zřejmě paralelní vývoj s oddělením Ascomycota. Základní společný znak: karyogamie a meióza probíhá v buňce zvané bazidie (meiosporangium), haploidní bazidiospory se tvoří exogenně na sterigmatech (stopečkách). Další společné znaky: vegetativní stélka tvořena vláknitým, dobře vyvinutým myceliem, které je přehrádkované a přehrádky opatřeny doliopóry (tvar soudku, obě strany póru kryje polokulovitá, membránami tvořená čepička, tzv. parentozóm). U zástupců řádů Uredinales a Ustilaginales pouze jednoduché póry. 76
, převažující životní fáze většiny zástupců, konjugované mitózy, které probíhají v buňkách, jsou většinou spojeny s tvorbou přezek, může existovat")
 plodnic, dikaryotické.")
79 Primární mycelium je jednojaderné, vzniká klíčením bazidiospory, je krátkého trvání. Sekundární mycelium dikaryotické (heterokaryotické), převažující životní fáze většiny zástupců, konjugované mitózy, které probíhají v buňkách, jsou většinou spojeny s tvorbou přezek, může existovat léta bez vytvoření plodnic. K vytvoření plodnic jsou třeba specifické podmínky (substrát, klima aj.). Terciální mycelium tvoří specializovaná pletiva (pseudoparenchym, plektenchym) plodnic, dikaryotické. Pohlavní rozmnožování: somatogamie (setkání 2 geneticky si odpovídajících, kompatibilních mycelií nebo bazidiospor), gameto-somatogamie (Uredinales), oddálení plasmogamie a karyogamie, probazidie (orgán karyogamie) a metabazidie (meióza), dle polohy vřeténka při první meiotické fázi bazidie chiastická nebo stichická. Holobazidie (nedělená, většinou chiastická) a fragmobazidie (přehrádkovaná). Typ bazidie (furkátní, kulovitá ) je typický pro většinu řádů. Bazidie může vyrůstat přímo na myceliu, popř. z klidových spor (teliospor, chlamydospor), nejčastěji jsou uspořádány ve výtrusorodé vrstvě (hymeniu) na povrchu nebo uvnitř plodnic (v glebě). Bazidie u rouškatých hub vyrůstají ze subhymenia, vrstvy buněk pod hymeniem (výtrusorodá vrstva), které vyrůstá na tramě (pletivo plodnice nesoucí bazidie), mezi bazidiemi jsou sterilní buňky (cystidy, jejich fce je sekreční, ochranná, zásobní). Na vrcholku nebo po stranách bazidie vyrůstají stopečky sterigmata (4). Haploidní jádra vycestují z bazidií, obklopí se plazmou a BS a vzniknou bazidiospory (pohlavní spory, tedy meiospory, exospory). Z hymenomycetoidních bazidií jsou spory aktivně odmršťovány (balistospory), z gastroidních bazidií se uvolňují pasivně (statismospory). Bazidiospory jsou různého tvaru, barvy, velikosti, ornamentace. Důležitým znakem je barvitelnost jejich stěny (cyanofilní - barví se anilinovou modří intenzívně modře, amyloidní jódem modře až fialově, dextrinoidní jódem hnědočerveně). Klíčení bazidiospory: hyfou primárního mycela nebo vznikne kvasinkovitá buňka schopná pučení, sekundární spora, popř. mikrokonidie. Asi 90 % druhů je heterothalických. Životní cyklus je dikaryotický, diploidní je pouze mladá bazidie (probazidie), haploidní fáze omezena na bazidiospory a primární mycelium. 77
 hyfy generativní (tenkostěnné, větvené a septované); b) hyfy skeletové (tlustostěnné, nevětvené, neseptované, s")
, pletivo monomitické (pouze generativní hyfy), trimitické (hyfy generativní, skeletové i vazbové)")
80 Plodnice (bazidiomata, bazidiokarpy) jsou na rozdíl od askomycetů není jejich vznik přímo spojen s pohlavním procesem. Plodnice jsou tvořeny plektenchymem, který může obsahovat tři různé typy hyf: a) hyfy generativní (tenkostěnné, větvené a septované); b) hyfy skeletové (tlustostěnné, nevětvené, neseptované, s širokým lumen); c) hyfy vazbové (také tlustostěnné, nevětvené, téměř plné). Podle přítomnosti určitého typu hyf rozlišujeme pletivo dimitické (generativní a skeletové hyfy), pletivo monomitické (pouze generativní hyfy), trimitické (hyfy generativní, skeletové i vazbové) Hymenofor: část plodnice, která nese hymenium (lupenatý, rourkatý, lištovitý, ostnitý nebo i hladký). Habituelní typy hymeniálních plodnic: 1. pilothecium je plodnice více či méně zřetelně dělená na třeň a klobouk, jednoletá, hymenofor na spodní straně klobouku (agarikoidní, kanthareloidní, pleurotoidní, cyfeloidní, resupinátní, hericiodní ) 2. holothecium nemá vytvořen klobouk a třeň, hymenium povléká celý povrch plodnice (klavariodní, ramarioidní, tremelloidní) 3. krustothecium je víceletá i jednoletá, postupně se vyvíjející (koncentricky vrstvená) plodnice, někdy členěná na klobouk a třeň (hydnelloidní, ganodermatoidní, fomitoidní, stereoidní, korticioidní) 4. plodnice geastrálního typu. Plodnice mají mnohdy obaly: velum (u rouškatých hub) a peridie (u břichatek). Velum (plachetka) může obalovat celé primordium (základ) plodnice (velum universale) a zanechává potom pochvu na třeni a bradavky na klobouku nebo zakrývá pouze hymenofor (velum partiale) a zanechává prstenec nebo pavučinu (kortinu). 78
81 Z hlediska ontogeneze se rozlišují gymnokarpní plodnice (hymenium od počátku otevřené), angiokarpní (hymenium se obnaží po dozrání spor), hemiangiokarpní (raná stádia plodnic v obalech, které zanikají a výtrusy zrají již obnažené). POZOR, DŮLEŽITÉ! Rozdíly mezi houbami vřeckovýtrusými a stopkovýtrusými: 1) pohlavní proces (gametangiogamie x somatogamie); 2) rozsah dikaryotické fáze (askogenní hyfy x sekundární vegetativní mycelium); 3) poměr jaderných fází v životním cyklu (haplodikaryonti x dikaryonti); 4) charakter meiospor (endospory x exospory); 5) vznik plodnice (přímá x nepřímá návaznost na pohlavní proces); 6) ultrastruktura stěny hyf (dvouvrstevná x vícevrstevná); 7) typ pórů v septech hyf (jednoduché póry x dolipóry). Nepohlavní rozmnožování: existuje, ale je nenápadné. Ekologie a význam: saprotrofyti a biotrofní parazité se značným fytopatologickým významem (Urediniomycetes a Ustilaginomycetes), zčásti symbionti (mykorhiza, lichenizovaných hub je nepatrné množství, symbiózy s termity a blanokřídlými). Řada druhů ze skupiny rouškatých hub rozrušuje celulózu či lignin (dřevokazné houby), mohou napadat i živé stromy. Nejnebezpečnější Serpula lacrymans, dřevomorka (hnědá kostkovitá hniloba opracovaného dřeva ve vlhkých prostorách). Jedlé druhy stopkovýtrusých hub: malá výživnost a energetická hodnota, poměrně vysoké % esenciálních AK, minerálních látek, látky snižující hladinu cholesterolu a ovlivňující krevní tlak, předností je i nízká stravitelnost hub, jež způsobuje větší peristaltiku střev. Houbaření je i rekreací. Cytostatické účinky mají i látky izolované z makromycetů: betulin (Piptoporus betulinus), lentinan (šiitake, Lentinus edodes), flamulin (Flammulina velutipes), kalvacin (Langermannia gigantea) Smrtelně jedovaté houby u nás: Amanita phalloides, A. verna, A. virosa, A. pantherina, Entoloma lividum, Inocybe patoulliardi (erubescens), Cortinarius orellanus aj. Ke kultovním účelům v některých oblastech využívány halucinogenní houby, např. r. Psilocybe Systém: V minulosti značně neustálený vzhledem k neujasněnému významu jednotlivých znaků. Ještě nedávno základní členění dle typu bazidie a schopnosti tvorby kvasinkovitých stádií a sekundárních spor na dvě skupiny: Homobasidiomycetes (Holobasidiomycetes) a Heterobasidiomycetes (Phragmobasidiomycetes). Dnes na základě molekulárních znaků členění na tři pododdělení: Pucciniomycotina, Ustilaginomycotina a Agaricomycotina: Pododdělení Agaricomycotina se dále člení na tři třídy: Tremellomycetes - dělená, vícebuněčná bazidie, Dacrymycetes nedělená dichotomicky větvená bazidie, Agaricomycetes - nedělená, jednobuněčná bazidie. Oddělení je členěno na více než 30 řádů, zahrnuje kolem rodů a kolem druhů. Výzkum na molekulární úrovni stále pokračuje, lze očekávat, že vnitřní systematika bazidiomycetů ještě projde změnami. 79
, doliopóry chybějí. Plodnice většinou chybějí, jsou-li vytvořeny, pak jsou morfologicky málo diferencované a nenápadné.")
82 PODODDĚLENÍ PUCCINIOMYCOTINA Monofyletická, ale morfologicky heterogenní, skupina vymezená na základě biochemických (přítomnost cukrů, koenzymů) a ultrastrukturálních (póry, šupinky) znaků. Dimorfická stélka (haploidní kvasinkovitá a dikaryotická vláknitá), doliopóry chybějí. Plodnice většinou chybějí, jsou-li vytvořeny, pak jsou morfologicky málo diferencované a nenápadné. 95% zástupců jsou rzi. Zde pouze vybrané řády. Řád: Septobasidiales Malá, ale fylogeneticky významná skupina organismů, které tvoří zvláštní typ symbiózy s červci přecházející až v parazitismus. Houba přijímá od několika jedinců výživné látky, které hmyz saje z hostitelské rostliny, za to poskytuje ostatním červcům ochranu před ptáky a parazitickými blanokřídlými. Řád: Uredinales rzi Rzi jsou dnes považovány za nejprimitivnější typy bazidiomycetů: relativně dlouhá haploidní fáze, odlišný typ oplození (gameto-somatogamie), absence doliopórů a přezek, obligátní, vysoce specializovaný parazitismus, převaha nepohlavního rozmnožování různými typy spor. Obligátně biotrofní parazité rostlin. Netvoří plodnice, ale ložiska výtrusů. Mycelium v mezibuněčných prostorách pletiv hostitele, do jejichž buněk vysílá haustoria. Napadené buňky zpravidla nejsou usmrcovány, přítomnost parazita může vést ke vzniku hypertrofií a hyperplázií (nádory, čarověníky). Řada druhů působí velké škody na kulturních rostlinách, nejúčinnější prostředek proti rzím je vyšlechtění odolných odrůd. Životní cyklus haplodikaryotický: bazidiospóry klíčí v haploidní mycelium, na něm se tvoří pohlavní orgány (spermogonia, označení 0) v nichž se tvoří spermacie; pohlavním procesem 80
; teliospory představují probazidie (karyogamie), po karyogamii a následné meiozi vyklíčí z teliospory příčně dělená bazidie se 4 haploidními bazidiosporami (označované")
83 vzniká dikaryotické mycelium, na němž se zakládají postupně ložiska nepohlavních výtrusů: jarní (aecia s aeciosporami, označení I), letní (uredia s urediosporami, označení II) a zimní (telia s teliosporami, označení III); teliospory představují probazidie (karyogamie), po karyogamii a následné meiozi vyklíčí z teliospory příčně dělená bazidie se 4 haploidními bazidiosporami (označované také jako sporidie, označení IV). Druhy makrocyklické (0, I, II, III, IV), u mnoha druhů mohou 1 3 typy ložisek chybět: brachycyklické (netvoří aecia), druhy mikrocyklické (např. III+IV). Druhy heteroecické, dvoubytné - střídají hostitele, druhy autoecické, jednobytné - beze změny hostitele. Hostitelská specializace. Životní cyklus dvoubytné (heteroecické) rzi na příkladu rzi travní, Puccinia graminis. Haploidní fáze vázána na mezihostitele (dřišťál), bazidiospora vyklíčí v hyfu, která vniká do tkání hostitele, a rozrůstá se v intercelulární monokaryotické mycelium, na líci listu vznikne skupinka drobných kuželovitých bradaviček (spermogonií), polokulovitých ložisek typu acervulů či pyknid. Spermogonia tvoří krátké hyfy, odškrcující na koncích množství spermacií, a též nektar, který je spolu se spermaciemi vytlačován ven a hromadí se u ústí spermogonií. Sladká šťáva nektaru láká hmyz, který rozšiřuje spermacie. Z ústí spermogonií vyčnívají přijímací hyfy, na něž se dostanou opačně pohlavně laděné spermacie. Jádro spermacie přejde do přijímací hyfy a posouvá se do mycelia pod spermogoniem, kde vznikne dikaryotická buňka. Dikaryotická buňka dává vznik prášilkám (aeciím), které mají tvar pohárků na spodní straně listu (jednobuněčné dikaryotické aeciospory, jarní výtrusy). Aeciospory slouží k přenosu nákazy na druhého hostitele (popř. infikují téhož hostitele), kde se vyvíjí dikaryotické mycelium, dávající vznik urediím s uredosporami (letní výtrusy). Ty jsou většinou oranžové a slouží k šíření nákazy na druhém hostiteli (trávy). Před dozráním hostitelské rostliny se na místech, kde se tvořila uredia, vytvoří telia s tmavými zimními výtrusy (teliosporami, teleutosporami). Teliospory představují probazidie, v nichž dochází ke karyogamii, posléze každá buňka teliospory klíčí bazidií, do ní se přesune diploidní jádro, dojde k meioze a vzniknou 4 haploidní buňky se 4 bazidiosporami (sporidiemi). 81
84 Puccinia má dvoubuněčné teliospory, druh Puccinia graminis, subsp. graminis na obilninách, subsp. graminicola na planě rostoucích travách, heteroecický druh, mezihostitel je dřišťál; P. persistens var. triticina je rez pšeničná, mezihostitel je žluťucha, P. coronata, rez ovesná, mezihostitel je řešetlák nebo krušina, P. punctiformis tvoří vonná spermogonia na pcháči rolním (monoecická) Uromyces má jednobuněčné teliospory, druh U. pisi, rez hrachová, na pryšci a vikvovitých Phragmidium s vícebuněčnými teliosporami, druhy časté na čeledi Rosaceae další důležité rody: Tranzschelia, Melampsora, Coleosporium, Cronartium Cronartium ribicola, rez vejmutovková, aecia na borovicích s 5-ti jehlicemi, uredia a telia na rybízu a angreštu PODODDĚLENÍ USTILAGINOMYCOTINA Biotrofní parazité rostlin (převážně krytosemenných); netvoří plodnice, ale ložiska (sory, sorus) chlamydospor. Dimorfní stélka: haploidní fáze kvasinkovitá, dikaryotická fáze vláknitá. Životní cyklus je dikaryotický, haploidní fáze je silně potlačena. Somatickou kopulací spor nebo primárních mycelií vzniká parazitické dikaryotické mycelium, rozrůstající se v mezibuněčných prostorách pletiv hostitele; napadené orgány rostlin se mění v sorus vyplněný masou tlustostěnných chlamydospor (označovaných též jako teliospory), v nichž dojde ke karyogamii, a po klidové fázi se tvoří promycel (bazidie). Zde dochází k redukčnímu dělení a tvorbě haploidních bazidiospor (označovaných též jako sporidie). Řád Ustilaginales sněti prašné Obligátní parazité s vysokou specifitou a organotropností. Boj proti napadení pomocí rtuťnatých a měďnatých přípravků, také vyšlechtěním rezistentního osiva. Ustilago segetum, prašná sněť ovesno-ječná, U. tritici, p. s. pšenično-ječná, U. mayidis, p. s. kukuřičná, U. violacea, teliospory v prašnících hvozdíkovitých Řád Exobasidiales Vysoce specifičtí parazité rostlin z čeledí Ericaceae, Empetraceae, Theaceae. Exobasidium vaccinii je známým druhem na listech brusnic, kde vyvolává deformace (miskovité nádory z jedné strany červené, z druhé bílé) nebo čarověníky větviček. Řád Tilletiales - sněti mazlavé Obligátní parazité krytosemenných rostlin, hlavně čeledí Poaceae, Ranunculaceae a Asteraceae. Vytvářejí mazlavá ložiska často páchnoucí zbytky hyf po vytvoření teliospor nebo deformace. Na pšenici a žitu parazitují druhy Tilletia caries, T. controversa. PODODDĚLENÍ AGARICOMYCOTINA Velká skupina se značnou morfologickou i ekologickou diverzitou. Většina zástupců tvoří plodnice. Do tohoto pododdělení náleží několik skupin veřejnosti známých hub: většina mykorhizních hub (tzv. makromycety), dále houby chorošovité a houby břichatkovité (gasteromycety). Systém: Tremellomycetes - dělená, vícebuněčná bazidie (většinou chiastická fragmobazidie), Dacrymycetes nedělená dichotomicky větvená bazidie, Agaricomycetes - nedělená, jednobuněčná chiastická bazidie. 82
85 TŘÍDA DACRYMYCETES Řád Dacrymycetales Plodnice polokulovité nebo parůžkovité typu holothecium, žluté až oranžové (ß-karoten), voskovité až rosolovité konzistence, vidlicovitá bazidie. Saprotrofové na dřevě. Předpokládá se, že to je bazální větev bazidiomycetů. Dacrymyces, kropilka, bochníčkovitá holothecia, D. stillatus, na řezné ploše pařezů. Calocera, krásnorůžek, holothecia připomínající kuřátka, na jehličnanech C. viscosa TŘÍDA TREMELLOMYCETES Bazidie přehrádkované nebo nepřehrádkované. Nápadné jsou dlouhé a tenké výběžky z buněk bazidií nesoucí bazidiospory. Řád: Auriculariales Plodnice rosolovité až chrupavčité konzistence. Saprofyté, popř. parazité rostlin. Hirneola auricula-judae, ucho jidášovo, mrtvé dřevo listnáčů, především bezu černého a akátu Auricularia, ušičko (boltcovitka), některé druhy pěstovány v Asii jako tzv. černé houby Exidia, černorosol, hymenium na svrchní straně plodnice, E. plana, č. bukový, až 10 cm velká zprohýbaná holothecia Pseudohydnum gelatinosum, rosolozub huspenitý, mléčně bílé, bokem přirostlé krustothecium s ostnitým hymenoforem, na zemi, pařezech Řád Tremellales Většinou se tvoří bazidiomata, hymenium povléká celý povrch plodnice nebo jen jeho spodní či svrchní část. Konzistence slizovitá či rosolovitá. Tremella, rosolovka, laločnatá, mozkovitě zprohýbaná holothecia. T. mesenterica, r. mozkovitá, na suchých větvích listnáčů, T. foliacea, r. listovitá, na pařezech dubů a buků T. fuciformis je často využívaná v tradiční asijské kuchyni do dezertů, polévek atd., přisuzují se jí i léčivé účinky (prevence proti kardiovaskulárním, imunologickým a neurologickým chorobám a proti nepříznivým efektům při radioterapii). Také se pěstuje, suší a dováží do Evropy do pleťových krémů jako tzv. botanická kyselina hyaluronová. Řád Filobasidiales Filobasidiella neoformans je teleomorfou patogenu Cryptococcus neoformans, způsobujícího kryptokokózu, zdrojem nákazy je holubí nebo netopýří trus, inhalací se dostává do těla, nebezpečí pro lidi s oslabenou imunitou (např. AIDS). Řád Tulasnellales Plodnice jsou korticioidní krustothecia povlékající substrát. Saprofyté a nespecializovaní parazité na dřevě a listech. Tulasnella - vločkovitá až rosolovitá plodnice na nrtvém dřevě TŘÍDA AGARICOMYCETES Současné vnitřní členění třídy, postavené na molekulárních, nikoli morfologických znacích, je zatím velmi obtížně interpretovatelné. Systematika skupiny je v prudkém vývoji poznání a bude v blízké budoucnosti jistě podléhat změnám. Proto se zde přidržíme tradičního pojetí, které zahrnuje 3 skupiny hub a plně vyhovuje potřebám našeho kurzu. Analýza znaků na molekulární úrovni i mikromorfologie plodnic ukazují, že se jedná o nefylogenetické skupiny, jejichž zástupci náleží do několika vývojových linií přirozeného systému stopkovýtrusných hub. 83
86 Zařazení vybraných zástupců do přirozeného systému: BASIDIOMYCOTA, pododdělení AGARICOMYCOTINA, třída AGARICOMYCETES, vybraní zástupci řád HOUBY NELUPENATÉ (CHOROŠE) BŘICHATKY HOUBY ROUŠKATÉ Polyporales Polyporus, Laetiporus, Trametes, Piptoporus, Pycnoporus, Fomitopsis, Fomes, Daedalea, Ganoderma Sparasis Agaricales Fistulina, Schizophyllum Tricholoma, Pleurotus, Clitocybe, čeledi Omphalina, Armillaria, LYCOPERDACEAE, Marasmius, Amanita, Agaricus, NIDULARIACEAE, Lepiota, Coprinus, Entoloma, TULOSTOMACEAE Cortinarius, Inocybe Boletales Serpula Scleroderma Boletus, Tylopilus, Strobilomyces, Xerocomus, Suillus, Paxillus Russulales Heterobasidion, Stereum Russula, Lactarius, Hericium Hymenochaetales Phelinus Cantharellales Cantarellus, Hydnum Phallales čeleď PHALLACEAE, rody Geastrum, Sphaerobolus Clavariadelphus, Ramaria Umělé, nefylogenetické skupiny: a) Aphyllophorales (houby nelupenaté) umělá skupina, založená na morfologii plodnic a hymenoforu. Převážně lignikolní chorošovité houby v nejširším smyslu s gymnokarpními plodnicemi. b) Gasteromycetes (houby břichatkovité) tradiční umělá skupina, založená na morfologii plodnice (angiokarpní plodnice s peridií a glebou), hymenium trvale skryto uvnitř plodnice a s tím související typ basidií, které nemají aktivní uvolňování spor (statismospory). Terestrické epigeické i hypogeické plodnice, saprotrofní i ektomykorhizní typy, rozšíření kosmopolitní, ale časté v teplých a suchých oblastech. Polyfyletická skupina, jejíž zástupci se opět vyskytují v několika vývojových liniích hub stopkovýtrusých. Integrace gasteromycetů do přirozeného systému vede k výrazným změnám v dosavadním chápání některých tradičních skupin. c) Hymenomycetes (houby rouškaté) tradiční umělá skupina, stojící v protikladu ke skupině předcházející. Plodnice gymno- nebo hemiangiokarpní, hymenium ve zralosti vždy otevřené, bazidie s aktivním uvolňováním spor (balistospory). Opět polyfyletická skupina, do které byly zahrnovány jak houby nelupenaté, tak běžné skupiny kloboukatých bazidiomycetů. Skupina hub nelupenatých ( Aphyllophorales ) Plodnice jsou většinou víceletá krustothecia, vzácněji jednoletá nebo přezimující pilothecia, konzistence masovitá, kožovitá či dřevovitá. Hymenofor většinou rourkatý či lamelovitý. Dřevní saprofyté nebo saproparazité, mnohé druhy značně rezistentní k vysušení, mohou růst na extrémních stanovištích (dřevěné ploty apod.). Některé druhy obligátně parazitické. Polyporus, choroš, jednoleté, často přezimující plodnice s třeněm, P. brumalis, ch. zimní, P. squamosus, ch. šupinatý, oba druhy na pařezech a kmenech listnatých stromů Fistulina hepatica, pstřeň dubový, jazykovitý, červený, šťavnatý klobouk s postranním třeněm, na živých dubových kmenech, jedlý 84
87 Laetiporus sulphureus, sírovec žlutooranžový, na živých kmenech listnáčů, jedlý Piptoporus betulinus, březovník obecný, světlý kopytovitý klobouk bokem přirostlý, pouze na břízách, napadá i živé jedince Ganoderma, lesklokorka, ploché plodnice, G. applanatum, l. ploská, G. lucidum, l. lesklá Fomitopsis pinicola, troudník pásovaný, z nejhojnějších druhů stejně jako Fomes fomentarius, troudnatec kopytovitý Heterobasidion annosus, kořenovník vrstevnatý, rozrušuje centrální část kmenů smrků (střední válec), šíří se kořeny, významný škůdce monokultur Trametes, outkovka, řada hojných druhů, rourkatý hymenofor Daedalea, síťkovec, labyrintický hymenofor, nejhojnější: D. quercina, D. confragosa Stereum, pevník, bokem přirostlé plodnice, často částečně resupinátní, hladký hymenofor, mnoho druhů Serpula lacrymans, dřevomorka domácí, ve vlhkých nevětraných prostorách, rozkládá celulózu až na vodu a CO 2, čímž si vytváří podmínky (vlhko) pro další šíření Skupina hub rouškatých ( Hymenomycetes ) 1. Plodnice je hemiangiokarpní pilothecium, vždy jednoletá. Hymenofor lupenitý. Obaly plodnice časté. Saprofyti, slabí parazité či mykorrhizní houby. Řada druhů jedlých, řada jedovatých. Pleurotus ostreatus, hlíva ústřičná, postranní třeň, saprofyt na pařezech, komerčně pěstována Tricholoma, čirůvka (havelka, zelánka, mýdlová), Calocybe gambosa (č. májovka), Lepista nuda (č. fialová) Armillaria mellea (václavka obecná), rhizomorfy působí značné škody ve smrčinách Omphalina (kalichovka), lichenizovaná houba, její mycelium je napojeno na řase Marasmius, špička (obecná, žíněná, česneková) Amanita, muchomůrka, jedlá A. rubescens (m. růžovka), jedovaté či smrtelně jedovaté jsou: A. pantherina (m. tygrovaná), A. muscaria (m. červená), A. phalloides (m. zelená, hlízovitá), A. verna (m. jarní), A. virosa (m. jízlivá) Agaricus, žampión, pečárka, tmavé výtrusy, A. bisporus, uměle pěstován a patří k nejprodávanějším houbám, hojně sbírány: A. arvensis (ž. ovčí), A. campestris (ž. polní), A. sylvaticus (ž. lesní), jedovatý A. xanthodermus (ž. zápašný) páchne fenolem Lepiota, bedla (vysoká, červenající) Cortinarius, pavučinec, mnoho druhů, řada jedovatých Inocybe patouillardii, vláknice Patouillardova a Entoloma sinuatum, závojenka olovová, obě smrtelně jedovaté Coprinus, hnojník, plodnice se v době zralosti mění v mazlavou hmotu, v mládí jedlé (obsahuje coprin, pozor na alkohol) Psilocybe, lysohlávka, halucinogenní účinky 2. Plodnice hemiangiokarpní či gymnokarpní pilothecium, hymenofor rourkatý či lupenitý, velum partiale u řady druhů přítomno. Převážně mykorrhizní houby, často s úzkou vazbou na 1 nebo několik druhů dřevin. Strobilomyces, šiškovec, šupinatý černohnědý klobouk Tylopilus felleus, hřib žlučník, Boletinus cavipes, hřibovec dutonohý 85
88 Boletus, hřib: B. edulis, h. smrkový, B. parasiticus, h. cizopasný (na plodnicích pestřeců), B. satanas, h. satan..., B. (Leccinum), kozák, křemenáč, B. (Xerocomus) badius, h. hnědý, B. (X.) subtomentosus, h. plstnatý, B. (X.) chrysenteron, h. žlutomasý, B. (Suillus), klouzek Paxillus, čechratka 3. Hemiangiokarpní pilothecia, hymenofor lupenitý. Trama kruchá, obsahuje kulovité buňky (sférocysty) a mléčnice. Převážně mykorrhizní houby. Russula, holubinka, mléčnice bez mléka, u nás přes 100 druhů jedlých i nejedlých Lactarius, ryzec, na řezu či lomu roní latex, též jedlé i nejedlé druhy 4. Gymnokarpní pilothecia či holothecia. Půdní mykorrhizní houby, saprofyté, popř. fakultativní parazité, některé druhy jedlé. Cantharellus cibarius, liška obecná, nálevkovitá žlutá plodnice s lištovitým hymenoforem Craterellus, stroček, hnědočerná nálevkovitá plodnice, jedlý Ramaria, kuřátka, bohatě keříčkovitě větvená holothecia, některá jedlá Clavariadelphus pistillaris, kyj Herkulův, 20 cm vysoké oranžové kyjovité plodnice Sparassis crispa, kotrč kadeřavý, téměř kulovité plodnice, bohatě keříčkovité, nejčastěji na borovicích, velmi chutný Skupina hub břichatkovitých ( Gasteromycetes ) 1. Plodnice hypogeická nebo epigeická kulovitého či hlavičkovitého tvaru. Exoperidie brzy zaniká, endoperidie tuhá, vytrvalá. Otevírá se většinou na vrcholu. U některých rodů zůstává spodní část gleby sterilní (subgleba). Asi 200 převážně saprofytických druhů, mnohé v mládí jedlé. Lycoperdon, pýchavka, nejhojnější na zemi v lesích je L. perlatum, p. obecná Calvatia, pýchavka Langermannia gigantea, p. obrovská (vatovec o.), na pastvinách, plodnice vážící i několik kg Bovista, prášivka, na rozdíl od předešlých chybí subgleba 2. Plodnice subhypogeická kulovitého tvaru s vícevrstevnou okrovkou. Exoperidie třívrstevná, vnější vrstva kožovitá, střední tvořená hustě propletenými hyfami, vnitřní parenchymatická. Tlakem vnitřních vrstev hvězdicovitě praská kožovitá vrstva exoperidie, cípy se stáčejí dolů a vynášejí glebu krytou endoperidií nad povrch. Asi 50 druhů, saprofytických, mnohé teplomilné. Geastrum, hvězdovka 3. Plodnice okrouhle hlízovitá, epigeická nebo subhypogeická. Peridie tuhá, otvírá se nepravidelným roztržením na vrcholku. Asi 40 převážně saprofytických druhů, některé mykorhizní. Scleroderma citrinum, pestřec obecný, na písčité půdě v borech. Ve větším množství toxický Pisolithus arhizus, měcháč písečný, významná mykorhizní houba na písčitých půdách 4. Plodnice je epigeická, kulovitá či pohárkovitá, zakrytá blankou (epifragmou). Peridie 1-6vrstevná, otvírá se prasknutím epifragmy, takže plodnice nabývá tvar číše či nálevky. Gleba obsahuje, uzavřené komůrky vystlané hymeniem. Zralá gleba se rozpadá na jedno až více čočkovitých tělísek (peridiol), která jsou přichycena ke stěně peridie slizovitou hmotou nebo provazci (funiculi). Za deště jsou pecičky vymršťovány a pomocí provazce přichycovány např. na stébla trav. Asi 50 druhů, převážně saprofyti na mrtvém dřevě i mezi trávou. Nidularia, hnízdovka, peridioly ve slizovité tekutině Crucibulum, pohárovka, odpadávající epifragma, na tlejících rostlinných zbytcích Cyathus, číšenka, hustě chlupaté plodnice na tlejícím dřevě 86
89 Sphaerobolus, hrachovec, jediná peridiola vymršťovaná až 1 m daleko 5. Plodnice se zakládá hypogeicky na silném, vícevrstevném myceliálním provazci. Peridie obvykle vícevrstevná, otvírá se rozpadem nebo tlakem receptakula. Komůrky gleby jsou vyplněny mazlavou páchnoucí masou spor. Gleba u výše organizovaných skupin obsahuje receptakulum (houbovitý pseudoparenchymatický útvar, který při dozrání proráží slizový obal a peridii a vynáší spory nad úroveň substrátu, je bílé, žluté či červené). Vůně a barva receptakula vábí hmyz, který rozšiřuje spory. Asi 50 druhů, většinou pozemní saprofyté, většina druhů tropických. Phallus impudicus, hadovka smrdutá, duté bílé receptakulum se zvonovitým kloboukem, pokrytým zelenošedou masou s výtrusy, mladé uzavřené plodnice jsou jedlé Mutinus caninus, psivka obecná, 2-4 cm vysoké receptakulum Clathrus ruber, mřížovka červená, kulovité, ohnivě červené, mřížovité receptakulum, původní v J Evropě, ojedinělě i u nás Anthurus archeri, květnatec Archerův, u nás ojediněle, zavlečen z Australské oblasti, receptakulum z 5-8 oranžově červených ramen Pomocné oddělení DEUTEROMYCOTA (FUNGI IMPERFECTI) - DEUTEROMYCETY, HOUBY NEDOKONALÉ, KONIDIÁLNÍ HOUBY Umělá pomocná skupina, vytvořená pro nedokonale známé houby, tzn. pro houby známé pouze v anamorfní fázi. S výjimkou skupiny Blastomycetes je stélka všech druhů tvořena vláknitým, přehrádkovaným, větveným myceliem. Díky ultrastruktuře hyf předpokládáme, že převážná většina druhů této skupiny představuje anamorfy vřeckovýtrusých hub. Rozmnožování: pomocí různých typů konidií na hyfách či na speciálních konidioforech. Genetická proměnlivost je zajišťována pomocí parasexuálního procesu (nevznikají noví jedinci, jen rekombinovaná DNA), kdy dochází k plazmogamii, karyogamii i haploidizaci (heterokaryosa - dotyk dvou geneticky odlišných mycelií a přechod jader, mutace). Výskyt a význam: 30 % všech známých druhů hub, nejčastěji v půdě nebo na rostlinných substrátech, mnohé jsou významnými rostlinnými parazity, některé rody hyperparazity, parazité živočichů a člověka jsou vzácnější (tropy a subtropy), v posledních letech vlivem oslabujícího se imunitního systému lidí vzrůstá význam fakultativních patogenů, vyvolávajících sekundární mykózy. Systém: pomocné třídy tvořeny na základě především morfologických charakteristik konidioforů a konidií, popř. způsobu konidiogeneze. POMOCNÁ TŘÍDA: BLASTOMYCETES Skupina zahrnující nesporogenní kvasinky, které mohou být anamorfami askomycetů i bazidiomycetů. Patří sem rody, vytvářející většinou jednobuněčnou kvasinkovitou stélku, někdy pučivé pseudomycelium, příležitostně vláknité mycelium. Zástupci jsou saprofyté na různých substrátech, mnohé druhy způsobují kvašení, některé druhy jsou parazité živočichů a člověka (dermatomykózy i napadení vnitřních orgánů) Torulopsis, mnohé druhy jsou anamorfami pravých kvasinek Candida, pseudomycelium nebo pravé mycelium, C. albicans způsobuje onemocnění kůže, sliznic, plic i centrálního nervového systému POMOCNÁ TŘÍDA: HYPHOMYCETES Nejpočetnější skupina konidiálních hub, zahrnující zejména saprofyty, které se účastní rozkladu biomasy v souvislosti s její mineralizací. Patří sem řada patogenů rostlin, živočichů i 87
90 člověka. Konidie vznikají přímo na myceliu nebo na konidioforech, které se seskupují v koremie či sporodochia. Bývá sem řazena i řada rodů, které mají známé teleomorfy: Penicillium, Aspergillus, Monilia, Oidium, Fusarium Acremonium strictum, běžný půdní druh, produkuje antibiotikum cephalosporin Cercospora, skvrnatička, parazit na listech cukrovky, krmné řepy, celeru, špenátu, mrkve Cladosporium, Alternaria, na rostlinných zbytcích v půdě, často působí i onemocnění kulturních plodin, konidie těchto rodů ve velké míře přítomny v ovzduší a způsobují s konidiemi dalších rodů alergická onemocnění dýchacích cest POMOCNÁ TŘÍDA: COELOMYCETES Vytvářejí ohraničená (acervuli) či uzavřená (pyknidy) konidiomata. Hyfy mohou tvořit stromatické útvary. Mnohé rody významné z fytopatologického hlediska. U některých známy teleomorfy: Phomopsis, Melanconium aj. Phoma herbarum je hojným saprofytem na stoncích bylin POMOCNÁ TŘÍDA: AGONOMYCETES Sterilní mycelia bez fruktifikačního stádia. Sclerotium, sklerociové útvary, které mohou patřit vřeckovýtrusým i stopkovýtrusým houbám Rhizomorpha, zřejmě anamorfa k bazidiomycetům Cystocoleus, Racodium, lichenizované typy, kde fotobiontem je zelená řasa Trentepohlia LICHENES LIŠEJNÍKY - umělá, ekologická nikoli taxonomická skupina Komplexní organismy, složené ze složky houbové (mykobiont), ke které se vztahuje jméno lišejníku a řasové, resp. sinicové (fotobiont, a to fykobiont nebo cyanobiont), která má své vlastní jméno. Převládající složkou je většinou mykobiont, který obvykle určuje morfologický tvar stélky, vzácněji jsou obě složky v rovnováze (Collema) nebo je tvar stélky určován vláknitým fotobiontem (Cystocoleus, Ephebe). Spojení obou složek je velmi těsné, bývalo označováno za symbiózu, vztah je však komplikovanější. Složitost vztahů obou partnerů (mutualismus, helotismus, komenzalismus) je někdy prezentována jako kontrolovaný parazitismus, míra kontroly je měřítkem fylogenetického stáří lišejníku. Ne každé soužití řasy a houby je lišejníkem. Řada definic, nejpřijatelnější je tato: Houbová složka je ve své výživě obligátně vázána na určitou řasu či sinici a tvoří s ní morfologickofyziologickou jednotku. Tvorba specifických sloučenin a specifických struktur nepohlavního rozmnožování není nutnou podmínkou existence lišejníku. Má se za to, že fotobiont dodává organismu organické látky, o mykobiontu se soudí, že dodává vodu a anorganické látky. K lišejníkům nepatří náhodně rostoucí houby spolu s řasami, saprofyticky či paraziticky rostoucí houby na řasách. Mykobiont: převážně askomycety, lišejníky tvoří samostatné rody, čeledi, popř. řády (Lecanorales), rody zahrnující lichenizované i nelichenizované typy jsou vzácné, méně jsou mykobiontem bazidiomycety (Agaricomycetidae), lichenizace je zde na primitivní úrovni, většina lichenizovaných typů bazidiomycetů patří do společných rodů s nelichenizovanými typy (např. Omphalina, dnes Lichenomphalia). Fotobiont (celkem pouze asi 100 druhů): 8 % případů sinice (Chroococcus, Gloeocapsa, Nostoc, Scytonema, Stigonema), ve většině případů zelené řasy (23 rodů, Trebouxia - polovina druhů lišejníků, Coccomyxa, Trentepohlia), výjimečně u několika druhů r. Verrucaria různobrvka a chaluha. Na rozdíl od mykobiontů patří většina fotobiontů k samostatně se vyskytujícím druhům sinic a řas. 88
91 Cefalodia: specifické výrůstky na stélce lišejníku, vzniklé, když se jeden mykobiont setká se dvěma kompatibilními fotobionty (př. Peltigera aphthosa: Coccomyxa a Nostoc). Anatomická stavba: homeomerická stélka - obě složky ve stélce rovnoměrně rozptýleny (morfologicky je stélka vláknitá či rosolovitá) heteromerická stélka - fotobiont lokalizován převážně v řasové vrstvě (z morfologického hlediska stélka korovitá, keříčkovitá či lupenitá) Stavba heteromerické stélky: korová vrstva - isodiametrické nebo válcovité buňky mykobionta tvoří pseudoparenchymatickou tkáň, popř. tvořena volně uloženými buňkami mykobionta nebo může chybět, bývá vytvořena na svrchní, někdy i spodní straně stélky, v kůře některých lišejníků bývají na spodní či svrchní straně ztenčeniny (pseudocyfely) či na spodní straně stélky průlomy (cyfely), které mají význam při výměně plynů a ve vodním režimu řasová vrstva - gonidiová - tvořena oběma složkami dřeňová vrstva medulla - tvořena vzájemně propletenými dlouhými hyfami houby Morfologie: vegetativní stélka vykazuje velkou variabilitu, je však u téhož druhu více méně konstantní: keříčkovitá (frutikózní) stélka: k substrátu přisedá úzkou částí, negativně geotropicky rostoucí lišejníky a tzv. vousovité druhy, radiální stavba lupenitá (foliózní) stélka: ploše rozložená stélka, přirůstá k podkladu v 1 nebo několika místech, lze ji oddělit od podkladu, laločnaté okraje, spodní strana s kůrou tvoří obvykle rhiziny, příchytná vlákna korovitá (krustózní) stélka: celou spodní stranou vrostlá či přirostlá k substrátu, nemá spodní kůru, přechod k lupenité tvoří efigurátní stélka dimorfická stélka: lupenitá či korovitá přízemní část (thallus horizontalis) a keříčkovitá vystoupavá část (thallus verticalis, podetium), Baeomyces, Cladonia vláknitá stélka: vlákno fotobionta opletené hyfami mykobionta rosolovitá stélka: může se podobat keříčovitým, lupenitým či korovitým typům, má rosolovitou konzistenci a fotobiontem je hlavně sinice leprariovitá stélka: práškovitá, morfologicky nediferencovaná Růst: velmi pomalý (některé korovité lišejníky 1 mm za 20 let, stáří některých stélek r. Rhizocarpon je odhadováno na let), využití při určování stáří staveb nebo ledovcových morén (lichenimetrie), příčina extrémní, živinami chudá a suchá stanoviště Rozmnožování: při odděleném rozmnožování obou složek, má-li vzniknout opět lišejníková stélka, musí dojít znovu k setkání obou komponent, pouze nepohlavní rozmnožování zajišťuje tvorbu diaspor obsahujících obě složky: fragmentace stélky: nejjednodušší typ, regenerace úlomků (Cladonia, Parmelia), někdy se celá stélka rozpadne na drobné části (Diploschistes scruposus) izidie: vzniknou protržením korové vrstvy stélky za vzniku lupenovitých (fylidie), papilovitých, korálovitých výrůstků, které stavbou odpovídají stavbě stélky, jsou pokryty korovou vrstvou, u některých lišejníků neslouží k rozmnožování, ale ke zvětšení povrchu stélky soredia: malé útvary, tvořené buňkami fotobionta, opletenými hyfami mykobionta, rozrušením kůry se uvolní (podoba práškovité masy), někdy k jejich uvolňování slouží diferencovaná místa - sorály, soredia vznikají na celém povrchu stélky (Cladonia fimbriata), popř. celá stélka 89
92 představuje rozptýlená soredia (Lepraria), sorály mohou mít různý tvar: bodové, hlavičkové, štěrbinovité, okrajové... Pohlavní rozmnožování: pouze u mykobionta (v ojedinělých případech se v plodnicích mykobinta vyskytují buňky fotobionta, tzv. hymeniální řasy, které garantují společné rozšiřování a vznik nových stélek) Strategie nalezení partnera sporami houby není ještě spolehlivě vysvětlena u druhů, kde fotobiontem není řasa běžně se vyskytující v přírodě (Trebouxia), klíčící mykobiont může získat řasu paraziticky z již existujících stélek nebo ze soredií, popř. vytváří volné asociace s volně žijícími řasami, než potká odpovídajícího fotobionta Fyziologie Díky spojení 2 nepříbuzných organismů vzniká nová fyziologická jednotka (vodní režim přežití období sucha v anabióze, hromadění radionuklidů 10x více než rostliny indikace zamořených oblastí, lišejníkové sloučeniny) Z produktů lišejníků patří mezi nejzajímavější tzv. lišejníkové sloučeniny, čili sekundární produkty metabolismu, dříve označované jako lišejníkové kyseliny. Jedná se o produkty vyvolané soužitím obou partnerů ve stélce, samostatně kultivovaný mykobiont nebo fotobiont není schopen jejich syntézy. V současnosti je známo asi 700 těchto látek, které jsou většinou specifické pro lišejníky, takže se u jiných organismů vyskytují jen ojediněle (parietin Aspergillus, kys. polyporová Polyporus ). Z toho je asi 100 kyselin, většinou slabě fenolického typu, další jsou alifatické až různě složité aromatické látky s nejrůznějšími radikály. Většinou jsou bezbarvé, některé mají funkci pigmentů (Xanthoria oranžovou, Rhizocarpon žlutou, ) a působí jako filtr slunečního světla pro fotobionty. O dalším významu existuje řada teorií, slouží jako zásobní uhlíkaté látky, jsou antiherbivorní (hořká chuť), antibiotické, pomáhají lišejníkům v kompetici o prostor a živiny s baktériemi, mikroskopickými houbami, usnadňují narušení substrátu. Některé látky jsou dokonce toxické pro savce (kyselina vulpinová). Někteří autoři se domnívají, že lišejníkové látky jsou pro lišejníky bezvýznamnými sekundárními produkty metabolismu. K jejich zjišťování je vypracována řada mikrochemických reakcí, fluorescenční analýza, zjišťování v UV světle, papírová chromatografie. Dnes není možný popis taxonu bez znalosti chemického složení lišejníku. Obsahu látek se využívá i k determinaci (barevné reakce s KOH, chlorovým vápnem a parafenylendiaminem). Některé z těchto látek lze získat extrakcí ze stélek a jejich následnou rekrystalizací. Výskyt a význam Lišejníky osidlují od pólů k rovníku především extrémní stanoviště, kde je snížená konkurence ostatních skupin organismů, zde působí jako půdotvorní činitelé, některé druhy jsou citlivými indikátory vzdušného znečištění či stanovištních podmínek. Rostou na kamenech a skalách (epilithické a endolithické druhy), na borce stromů (epi- či endofloedické), na zemi (epigeické), v tropech i na listech (foliikolní), některé jsou parazitické, naopak parazity na lišejnících jsou zejména nelichenizované typy z rodů, čeledí a řádů lichenizovaných hub. Antibiotického působení lišejníků (lišejníkových látek) se využívá proti bakteriím a některým houbám. V lidovém léčitelství byly používány dříve některé druhy na základě pověr souvisejících s podobou stélek s některými orgány (Lobaria: plíce, Usnea: růst vlasů), Cetraria islandica je součástí plicních čajů. Některé druhy lokálně využívány jako potrava či krmivo, velký význam mají jako potrava zvěře, zvláště sobů v tundře a mnohých bezobratlých. Jedovaté lišejníky: deriváty kyseliny vulpinové (Letharia /Vulpiaria/ vulpina - jed proti vlkům, Cetraria pinastri), výroba barviv (Roccella - lakmus), parfémů (Evernia prunastri a Pseudevernia furfuracea - nosiče vonné esence), výroba hřbitovních věnců (Cladonia stellaris). 90
93 Systém: Asi druhů, 21% všech hub, náležejí převážně k vřeckovýtrusným houbám (Ascolichenes), v malé míře ke stopkovýtrusným houbám (Basidiolichenes). ASCOMYCOTA, pododělení PEZIZOMYCOTINA Řády obsahující jen některé lichenizované zástupce: Ostropales, Helotiales, Dothideales, Lecanidiales, řády obsahující pouze lichenizované typy: Gyalectales, Lecanorales, Lichinales, Arthoniales, Pyrenulales, Verrucariales. TŘÍDA LEOTIOMYCETES Helotiales: Baeomyces, malohubka, fotobiont zelená řasa Coccomyxa TŘÍDA LECANOROMYCETES Gyalectales: Graphis, čárnička, čárkovitě protáhlá hysterothecia, na borce listnáčů (Trentepohlia), Diploschistes vyklenutá apothecia, epilit Lecanorales: druhově nejpočetnější řád diskomycetů, patří sem nejméně polovina všech známých druhů lichenizovaných hub mnoho rodů (Cladonia, Usnea, Parmelia, Physcia, Cetraria, Lecanora, Lecidea, Rhizocarpon, Xanthoria, Peltigera a další) Lichinales: Ephebe vláknité povlaky na skalách (Stigonema) TŘÍDA CHAETOTHYRIOMYCETES Verrucariales: pseudoperithecia nebo perithecia, Verrucaria, endolitická stélka, na vápencích, několik druhů je vodních, Dermatocarpon, lupenitá stélka, na kamenech v potocích s čistou vodou BASIDIOMYCOTA, pododělení AGARICOMYCOTINA TŘÍDA AGARICOMYCETES Všechny známé lichenizované bazidiomycety patří do několika čeledí: Thelephoraceae, Corticiaceae, Hygrophoraceae, Clavariaceae a Tricholomataceae. Omphalina (Lichenomfalia) plodnice pilothecium je v kontaktu na bázi třeně s fotobiontem v granulích nebo šupinách Cora fotobiont tvoří zřetelnou vrstvu uvnitř plodnice Multiclavula plodnice vyrůstají z tenkého povlaku fotobionta na substrátu ŘÍŠE: ROSTLINY PLANTAE Rozsáhlá říše obsahující převážně fotoautotrofní mikroskopické a makroskopické organismy s různým stupněm diferenciace stélky (thalus) nebo rostlinného těla (cormus). Chloroplasty pokrývá dvojice povrchových membrán (vznikly cestou primární endosymbiózy, jejich donorem byla ve všech případech jednobuněčná sinice). 2 podříše Biliphytae a Viridiplantae jsou sesterské skupiny, z nichž každá má monofyletický původ. PODŘÍŠE: BILIPHYTAE Sinicový původ chloroplastů je zřejmý, u glaukofyt se jedná o cyanely (organely pokryté peptidoglykanovým obalem, podobným buněčné stěně sinic), ruduchy mají pravé chloroplasty pokryté dvojicí membrán. Chloroplasty obsahují jednotlivé tylakoidy, fotosyntetické pigmenty: chlorofyl a a fykobiliproteiny ve fykobilizomech. Zásobní látka: škrob. Nové uspořádání říše Plantae na základě výsledků molekulárních studií (Saunders a Hommersand, 2004) má 3 podříše: Biliphytae (s odd. Glaucophyta), Rhodoplantae (s odd. Rhodophyta se 3 pododěleními a odd. Cyanidiophyta, kam byly zařazeny jednobuněčné ruduchy žijící v extrémních životních podmínkách) a Viridiplantae. 91
94 ODDĚLENÍ: GLAUCOPHYTA (GLAUCOCYSTOPHYTA) Několik fotoautotrofních druhů s eukaryotní stavbou buňky. Společným znakem jsou cyanely, organely pokryté peptidoglykanovým obalem, podobným buněčné stěně sinic, tylakoidy nesou fykobilizomy. Oddělení je přechodným stavem mezi prokaryotními sinicemi a eukaryoty s chloroplasty. 3 rody Cyanophora, Gloeochaete, Glaucocystis jsou jednobuněčné, monádoidní, kapsální a kokální řasy. Glaucocystis žije v tůních s mírně kyselou reakcí vody v koloniích spojených zbytky buněčných stěn, protoplast obsahuje pulzující vakuoly, dvojici velmi krátkých bičíků a četné cyanely. ODDĚLENÍ: RUDUCHY RHODOPHYTA Ruduchy jsou fotoautotrofní eukaryotní rostliny s jednobuněčnou, vláknitou, často bohatě větvenou nebo ploše listovitou stélkou. Chybějí bičíkatá stádia v životních cyklech všech známých ruduch. Zvláštností ruduch je schopnost fúze protoplastů a prodlužovací růst. Buňky osního vlákna některých ruduch (Lemanea, Ceramium) se mohou až 100x prodloužit a 14000x zvětšit svůj objem. Když některá buňka vlákna odumře, buňky nad ní a pod ní se rozdělí, prodlouží a splynou, čímž se vlákno opraví. U jednoho druhu zjištěn specifický glykoprotein (rhodomorfin), fungující jako hormon při opravě poranění. Stavba buňky Buněčná stěna - polysacharidová, složená z amorfních vysoce hydrofilních polygalaktanů, které bobtnají a rozpouštějí se v horké vodě, nejdůležitější jsou agar a karagén. V amorfní hmotě jsou uloženy mikrofibrily celulózy, které zajišťují mechanickou pevnost stěny. Chloroplast s dvojicí membrán na povrchu, tylakoidy jednotlivé, na jejich povrchu jsou fykobilisomy (fykobiliproteiny: fykocyanin, fykoerytrin, allofykocyanin). Kombinace fotosyntetických pigmentů: chlorofyl a, (d), dále α- a ß- karoten, zeaxantin a lutein. Výsledná barva ruduch je výsledkem kombinace pigmentů. Červená barva stélky je považována za typickou, vyjadřuje převahu fykoerytrinu. Chromatická adaptace chybí. Zásobní látka: florideový škrob uložený v cytoplazmě. Rozmnožování a rodozměna. Nepohlavní rozmnožování aplanosporami, jimiž se rozmnožují gametofyty i sporofyty (n nebo 2n monospory). Pohlavní rozmnožování - oogamie - vaječná buňka (karpogon) je oplozema spermacií. U některých zástupců má karpogon protáhlý trichogyn, na němž se zachytí spermacie. Po karyogamii a plazmogamii prodělává zygota další vývoj, který má řadu obměn a jeho výsledkem jsou diploidní karpospory. Obvykle ze zygoty vyrůstá několik sporogenních vláken (gonimoblastů), ta splynou s podpůrnými (auxiliárními) buňkami a pak teprve se začnou tvořit karpospory. Z karpospor vyroste diploidní tetrasporofyt, v tetrasporangiích po meiose se tvoří haploidní tetraspory. Z nich vyroste nový gametofyt. (Rodozměna se dvěma typy sporofytu: karposporofyt a tetrasporofyt). Díky obrovskému množství karpospor i nepohlavnímu rozmnožování S i G mají ruduchy velkou reprodukční kapacitu, kterou jsou schopné vyrovnávat predační tlak. 92
, ubývá jich od rovníku k pólům.")
95 Výskyt v přírodě Většinou mořské řasy, dominantní skupina mořských makrofyt (více druhů než ostatních dohromady), ubývá jich od rovníku k pólům. Rostou v litorálu a sublitorálu, díky možnosti využívat modrozelené spektrum světla dosahují do značných hloubek (v průzračných tropických oceánech až 180 m). Corallina a Lithothamnion - inkrustace uhličitanem vápenatým, tvoří rhodolity, kulovité útvary na mořském dně, které velmi pomalu rostou. Geologické stáří Bangiomorpha pubescens z kanadské Arktidy, 1,2 mld let, je 1. nálezem mnohobuněčného eukaryotního organismu. Hlavní vývojové linie - z kambria (600 mil let). Ruduchy byly považovány za nejstarší eukaryota (absence bičíků), proto byly řazeny na počátek systému řas. Podle současných poznatků vznikly ruduchy ve stejné linii, z níž se oddělily zelené řasy a mnohobuněční živočichové (ztráta bičíků je sekundární, mnohé zelené rostliny a živočichové bičíky mají, druhotná ztráta též u Zygnematophyceae, u Chlorella, většiny cévnatých rostlin, u hub - Fungi). Hospodářský význam ruduch Potrava pro lidi (Japonsko, Čína a Korea, i umělé pěstování - Porphyra), krmivo pro zvířata, hnojivo, surovina pro výrobu bioplynu. Nejvýznamnějším produktem - agar - Gelidium, Gracillaria aj., tvoří až 40% sušiny stélky, získává se extrakcí v horké vodě (90 C) - koloidní roztok, gelóza, po ochlazení na 35 C ztuhne v průzračný gel: potravinářské rosoly, bakteriologické půdy. Karagén - Chondrus, Gigartina - tužidlo v mlékárenství, ve farmacii, textilní a kožedělný průmysl. 93
96 TŘÍDA: CYANIDIOPHYCEAE Jednobuněčné ruduchy. Výživa mixotrofní. Extrémní životní podmínky (Cyanidium - extrémně kyselé termální půdy ve vulkanických oblastech, Yellowston, Vesuv...) TŘÍDA: BANGIOPHYCEAE Zpravidla mikroskopické, jednobuněčné nebo vláknité řasy. Výjimkou je Porphyra, jejíž stélka je ploše listovitá, ale tvořená 1 vrstvou buněk. Jeden centrální chloroplast s pyrenoidem. Porphyridium jednobuněčné ruduchy ve sladké vodě a půdě, modelový organismus pro studium fotosyntézy Bangia - vláknitá nevětvená stélka, v řekách a mořském litorálu Porphyra - listovitá stélka je G, 15 až 150 cm, S - mikroskopický, vláknitý, na povrchu lastur či kamenech, gametofyty jednoleté, sporofyty mnoholeté (konchocelidové stádium, podobnost s ruduchou Conchocelis), heteromorfickou rodozměnu u Porphyra objevila Kathleen D. Baker, tím umožnila produkci porfyry v hodnotě mnoha mld dolarů ročně, proto má v Japonsku park a sochu, je oslavována jako Matka moře, zřejmě jediný oslavovaný algolog na světě, 3-4 sklizně v roce, v Japonsku nori, v Číně známa od 6. století, nutriční hodnota dána obsahem lehce stravitelných aminokyselin a řady vitamínů (dieta založená na konzumaci porfyry omezuje výskyt některých zhoubných nádorů) TŘÍDA: FLORIDEOPHYCEAE Mnohobuněčné, makroskopické stélky heterotrichálního nebo pletivného typu. Vlákna stélky jsou jednoosá (uniaxiální) či mnohoosá (multiaxiální), osní vlákna pokrývá korová vrstva z 94
97 drobných buněk. V buňkách četné nástěnné drobné chloroplasty bez pyrenoidu, karpogon s trichogynem. Žijí převážně v mořích. 50 rodů je sladkovodních. Lemanea - nevětvené asi 10 cm vysoké stélky na kamenech v horských potocích a řekách, podobné tmavým pravidelně zaškrcovaným žíním (G), boční větévky srůstají v souvislou kůru, karpospory uvnitř, sporofyt mikroskopický Batrachospermum - mírně zastíněné kameny v potocích a rašelinných tůňkách, stélka s výrazně přeslenitou stavbou ponořená do slizu Corallina, Lithothamnion inkrustované stélky v mořích tvoří kostru korálových útesů Hildebrandia - v čistých potocích, stélky přirostlé celou plochou ke kamenům, nedají se oddělit Ceramium - jemná keřovitá stélka v litorálu moří, dichotomicky větvená, klešťovitě zahnuté vrcholy Gelidium, Euchema, Chondrus, Ahnfeltia, Rhodymenia - využití na agar, karagen, potraviny PODŘÍŠE: ZELENÉ ROSTLINY VIRIDIPLANTAE (CHLOROBIONTA) Intenzívním studiem fylogeneze zelených rostlin v minulých desetiletích, aplikací ultrastukturálních a molekulárních metod (srovnání sekvencí aminokyselin - aktin, Rubisko - a nukleotidů 18 S rrna), se potvrdil monofyletický původ podříše zelené rostliny. Vývojová linie zelených rostlin vznikla před 1,5 mld let, suchozemské rostliny (úspěšně se adaptovaly v měnících se biogeochemických podmínkách Země a bezkonkurenčně obsadily její povrch) se odštěpily pravděpodobně před 700 mil (resp mil.) let. Morfogeneze suchozemských rostlin je spojena s řadou adaptačních inovací: jsou zpravidla větší a těžší, mají větší mechanickou odolnost (parožnatky cm, sekvojovce m, průměr kmene 8 m). Názvy vývojová linie a vývojová větev označují skupiny, u nichž zatím nelze určit stupeň taxonomické hodnoty (koncovka ae naznačuje dočasné řešení). Systém: 1. vývojová linie: Chlorophytae - zelené řasy oddělení Chlorophyta 2. vývojová linie: Streptophytae 2a. vývojová větev Charophytae oddělení Charophyta 2b. vývojová větev Bryophytae mechorosty oddělení Anthocerotophyta, Marchantiophyta, Bryophyta 2c. vývojová větev Cormophytae - cévnaté rostliny VL zelené řasy s oddělením Chlorofyta jsou řasy se zastoupením stélek všech typů, kromě volně žijící rhizopodové. Mitóza je zpravidla uzavřená. Buněčná stěna se zakládá mezi dceřinými buňkami jako mikrotubulární systém zvaný fykoplast. Většina druhů žije ve vodním prostředí. Mezi zelenými řasami a streptofyty není přímá vývojová návaznost, zelené řasy jsou považovány za slepou vývojovou linii. VL streptofyta obsahuje vývojové větve parožnatky, mechorosty a cévnaté rostliny. Mitóza je zpravidla otevřená. Oddělení dceřiných buněk zajišťuje mikrotubulární systém fragmoplast. Parožnatky jsou považovány za vývojové předky vyšších rostlin, ale jejich ekologie připomíná spíše zelené řasy. Jejich životní cyklus je haplontní, zygota je jedinou diploidní 95
98 buňkou. Mechorosty mají heteromorfickou rodozměnu s převahou gametofytu. V životním cyklu cévnatých rostlin převládá většinou víceletý diploidní sporofyt (kormus s kořeny, stonkem a listy). Předky cévnatých rostlin jsou zřejmě méně specializované třídy parožnatek (např. štětinatky). VÝVOJOVÁ LINIE: ZELENÉ ŘASY CHLOROPHYTAE ODDĚLENÍ: ZELENÉ ŘASY CHLOROPHYTA Jedna z největších skupin řas se všechny typy stélek kromě rhizopodové. Chloroplasty obsahují chlorofyl a+b, α- a ß-karoten, xantofyly: lutein, neoxanthin, violaxanthin, zeaxanthin aj. Na povrchu je dvojice obalných membrán, tylakoidy po 2-6 v lamelách. Pyrenoid je součástí chloroplastu, povrch bývá pokryt škrobovou čepičkou. Obsahuje aktivní enzym RuBisCO, který váže oxid uhličitý v temné fázi fotosyntézy. Zásobní látka: škrob ukládá se na povrchu pyrenoidu nebo v chloroplastech a polyfosfátová zrnka (volutin). Důležité znaky v systematice zelených řas: 1. sekvence nukleotidů v 18 S rrna, 2. morfologie stélek, povrch buněk atd., 3. způsob rozmnožování, 4. postavení bazálních tělísek bičíků tzv. absolutní konfigurace. Pozn.: termín absolutní konfigurace naznačuje, že tomuto znaku byla v nedávné minulosti přisuzována naprostá nadřazenost nad všemi ostatními znaky a celá systematika zelených řas byla dělána podle něj (měla navrch submikroskopická stavba buněk). Ale ne všechny skupiny, vykazující stejné postavení bičíků, jsou doopravdy monofyletické (Pleurastrophyceae) a naopak, v jedné skupině se může vyskytnout dvojí typ orientace bazálních tělísek. Z toho plyne poučení, že absolutní řešení neexistuje Bičíky a bičíkový aparát. Zpravidla 2 (nebo 4), stejně dlouhé, s jemným vlášením (čtyři bičíky má jen několik málo rodů bičíkovců a zooidy vláknitých zelených řas). Stefanokontní zoospory - věnec bičíků (u r. Oedogonium). Z kinetozomu vyrůstají bičíkové kořeny 3 typů: - mikrotubulární kořeny (nekontraktilní), ze 2, 4 i více srostlých mikrotubulů, v určité vzdálenosti se rozplétají a stávají se součástí cytoskeletu - příčně pruhovaná kontraktilní spojovací vlákna, zajišťují pohyb bičíků a tedy i celé buňky - rhizoplast, příčně pruhované kontraktilní vlákno od kinetozomu k povrchu jádra, kde se větví, určuje polohu jádra v buňce Bičíkový aparát je považován za konzervativní buněčnou strukturu, která během fylogeneze eukaryot prodělávala malé změny. Proto je jeho popis důležitým znakem při popisu oddělení a tříd zelených řas a primitivních streptofytů. Pro charakteristiku tříd je důležité vzájemné postavení kinetozomů a mikrotubulárních kořenů při pohledu od vrcholu buňky k jejímu zadnímu konci. Charakteristické je křížové uspořádání bazálních tělísek, které se vyznačuje 180º rotační symetrií. Z každého bazálního tělíska vyrůstají 2 mikrotubulární kořeny, z nichž jeden obsahuje 2 srostlé mikrotubuly, druhý se skládá ze 4 a více mikrotubulů. Při popisu vzájemné polohy bazálních tělísek si pomáháme hodinovým ciferníkem. Existují 4 typy postavení BT bičíků: A. bičíky jsou umístěny asymetricky na boku. Jejich mikrotubulární zakotvení je velmi nesymetrické jeden kořen je o mnoho silnější než druhý (Charophyceae, Charophyta). B. bičíky jsou umístěny symetricky na předním konci buňky. B1: V příčném řezu na TEM při pohledu shora na apikální konec bky jsou jejich bazální tělíska umístěna přesně naproti tzv. direct opposite, DO (část Chlorophyceae). B2: V příčném řezu na TEM jsou jejich bazální tělíska vůči sobě posunuta ve směru hodinových ručiček clock-wise position, CW. Také bývá toto postavení charakterizováno pomocí čísel z ciferníku hodin, tedy jako 1/7 (část Chlorophyceae). 96
99 B3: V příčném řezu na TEM jsou jejich bazální tělíska vůči sobě posunutá proti směru hodinových ručiček counter-clockwise position, CCW, nebo též 11/5 (Trebouxiophyceae, Ulvophyceae). Mitóza, cytokineze. Stejně jako struktura bičíkového aparátu jsou i jaderné a buněčné dělení konzervativní svou strukturou i průběhem jednotlivých fází, proto poskytujívýznamné informace o fylogenezi. Vegetativní buňky jsou jednojaderné (např. v kokální nebo vláknité stélce) i vícejaderné (cenoblastická, sifonální a sifonokladální stélka). Průběh mitózy odlišný u jednotlivých skupin zelených řas: Nejprimitivnější je uzavřená vnitrojaderná pleuromitóza. Při ní se jaderná membrána nerozpouští, uvnitř jádra vzniknou dvě vřeténka svírající nejprve ostrý úhel. Nevytváří se ekvatoriální destička. Vyskytuje se u Prasinophyceae Většina zelených řas má ortomitózu to znamená, že je vytvořeno bipolární vřeténko od pólu k pólu, a v metafázi jsou chromozómy uspořádány v ekvatoriální destičce. Existují dva typy ortomitózy, podle stupně rozpadu jaderné membrány: uzavřená (nebo částečně otevřená) ortomitóza jaderná blána zůstává zachována (nebo se na pólech otevřou polární okénka) a otevřená ortomitóza - klasický typ, kdy se jaderná membrána rozpadá u zelených řas není vyvinuta, vyskytuje se u oddělení Charophyta. Po mitóze následuje cytokineze (dělení cytoplasmy). Hranice dceřiných protoplastů určují mikrotubulární systémy 2 typů, fykoplast a fragmoplast, výsledkem je vytvoření přehrádky (nové buněčné stěny) mezi dceřinými buňkami. Fykoplast není odvozen od mitotického vřeténka, je to nový systém, který je kolmý na osu původního vřeténka, následně se tvoří septum, které odděluje protoplasty dceřiných buněk. Fragmoplast - systém, využívající mikrotubulů mitotického vřeténka, tvoří se na konci telofáze, vzniká buněčná destička, jako základ budoucí příčné přehrádky - progresivní struktura (Coleochaetacea, Charophyceae, dále mechorosty a cévnaté rostliny). Cytokineze bez účasti fykoplastu nebo fragmoplastu je spojená s oboustranným zaškrcením buňky, dělicí rýha vrůstá v ekvatoriální rovině od obvodu ke středu buňky. Buněčné stěny mají různé složení, většina zelených řas má stěny polysacharidové ve formě mikrofibril (celulóza) + amorfní složka (hemicelulóza, pektin). Některé řasy (resp. BS) obsahují sporopolenin vrstva odolných biopolymerů na povrchu buňky. Zástupci třídy Prasinophyceae mají na povrchu polysacharidové submikroskopické šupiny, u r. Chlamydomonas je glykoproteinová chlamys (plášť). Životní cykly zelených řas zpravidla haploidní, jedinou diploidní buňkou je zygota, při jejím klíčení probíhá meióza. Haplo-diplontní ŽC ve třídách Ulvophyceae, Cladophorophyceae, Bryopsidophyceae, Dasycladophyceae. Nepohlavní rozmnožování mitospory (n: G, 2n: S), zoospory v zoosporangiu, aplanospory, hemiaplanospory (chybí bičík, mají stigma, pulsující vakuoly), autospory (po opuštění mateřské BS mají tvar jako mateřská buňka). Všechno jsou to endogenní spory, při opouštění mateřské BS se uplatňují autolyziny. Zoospory a autospory často tvoří kolonie spojené slizem či zbytky mateřských stěn. Zvláštním typem kolonie je cenobium (jedna generace buněk, počet buněk 2 n, pravidelné uspořádání, morfologické a funkční specializace buněk, rozmnožování dceřinými cenobii (Chlamydophyceae, Chlorophyceae). Pohlavní rozmnožování druhy jednopohlavné (diecické) a oboupohlavné (monoecické), homo- a heterothalické, časté jsou feromony. Izogamie, anizogamie, oogamie. 97
100 Systém v minulých letech značně změněn, dříve na základě organizačních stupňů stélek, dnes na základě submikroskopické stavby bičíkového aparátu, mechanismů mitózy a cytokineze. Molekulární fylogeneze na základě analýzy sekvence 18S rrna. 8 tříd: Prasinophyceae, Ulvophyceae, Cladophorophyceae (žabovlasovité), Bryopsidophyceae (trubicovky), Dasycladophyceae, Trentepohliophyceae, Trebouxiophyceae, Chlorophyceae (zelenivky). TŘÍDA: PRASINOPHYCEAE (MICROMONADOPHYCEAE) Volně žijící bičíkovci, kapsální, méně kokální řasy, povrch buňky a bičíků pokrývá 1 i více vrstev submikroskopických polysacharidových šupin. Bičíky (1 8) vyrůstají z mělké prohlubně na předním konci buňky. Třída je heterogenní skupinou, která zřejmě představuje vývojový základ zelených řas a streptofyt. U jednotlivých rodů se liší orientace bazálních tělísek i bičíkových kořenů i průběh mitózy. Výskyt: mořské i sladkovodní, planktonní i přisedle žijící řasy (Pyramimonas, Prasinocladus). Některé druhy patří mezi nejhojnější mořské planktonní, resp. pikoplantonní řasy. TŘÍDA: CLADOPHOROPHYCEAE - ŽABOVLASOVITÉ Řasy se sifonokladální stélkou. CCW orientace bazálních tělísek bičíků zoidů. Mitóza uzavřená. ŽC haplo-diplontní, izomorfní rodozměna, gamety se dvěma bičíky, zoospory 4- bičíkaté. Cladophora (žabí vlas) patří mezi nejrozšířenější řasy, druhy rostou v moři, brakických i sladkých vodách. U nás všeobecně rozšířená Cladophora glomerata na kamenech v proudící eutrofní vodě (vlákna bohatě větvená, na omak drsná). TŘÍDA: BRYOPSIDOPHYCEAE - TRUBICOVKY Vegetativní stélka je sifonální mnohojaderná, tzv. cenocyt (tvar rozvětvené trubice). Dvě skupiny trubicovek: jedny mají pouze chloroplasty (homoplastické druhy: Bryopsis, Codium), druhé mají kromě chloroplastů i amyloplasty (v nich jsou zásobní polysacharidy, heteroplastické druhy: Caulerpa, Halimeda). Mitóza uzavřená. Specifické xantofyly sifonein a sifonoxantin. Životní cyklus haplo-diplontní, izogamie. Zoidy s CCW orientací bazálních tělísek. Gametofyt makroskopický, diploidním stádiem je pouze zygota, která se může množit stefanokontními zoosporami. Rostou pouze v teplých mořích. Trubicovky jsou vystaveny značnému predačnímu tlaku, ale mají efektivní opravný systém. Bryopsis - v litorálu evropských moří, cm vysoká rozvětvená stélka, souměrné postranní větévky Codium - bohatě větvená keřovitá, bochánkovitá až kulovitá stélka Caulerpa - tropická a subtropická moře, některé druhy obsahují pro mořské živočichy toxický terpen kaulerpenyen, přízemní stolony s rhizoidy a fotosyntetizující fyloidy (ve Středozemním moři je invazní C. taxifolia, původně pěstovaná v Monaku v mořském akváriu) Halimeda ploché vějířovité segmenty stélky, takže zvápenatělé (inkrustace aragonitem) stélky připomínají kaktus Opuntia, významný budovatel korálových útesů v tropických mořích, holokarpní ŽC celá stélka se přemění v gamety, po jejich vyrejdění zbude jen prázdná kalcifikovaná stélka 98
101 TŘÍDA: ULVOPHYCEAE Převážně mořské řasy s jednobuněčnou, vláknitou, sifonální, sifonokladální nebo ploše listovitou stélkou. Volně žijící bičíkovci chybějí. Zoidy (zoospory, gamety se 4 nebo 2 bičíky) a CCW orientací bazálních tělísek. Mitóza uzavřená. Fykoplast se netvoří, v buněčných přepážkách chybějí plazmodesmy (nepravá vlákna). Haplo-diplontní nebo haplontní ŽC. Pohlavní proces je izo- nebo anizogamie. Ulva lactuca (porost locikový) - ploše listovitá zkadeřená stélka, tvořená 2 vrstvami parenchymatických buněk, asi cm vysoká, v litorálu moří na kamenech, někdy se upravuje jako salát. Nepohlavní rozmnožování fragmentací stélky a čtyřbičíkatými zoosporami. Heterothalismus, anizogamie, izomorfická rodozměna. Gamety se dvěma bičíky mají univerzální fci, pokud nedojde ke kopulaci, vyrostou z nich gametofyty. Ulothrix (kadeřnatka) - v moři i sladkých vodách, kosmopolit. Trichální stélka (přirůstá k podkladu bazální buňkou bez chloroplastů), heterothalismus, zoospory se 4, gamety se 2 bičíky, izogamie, planozygota, zygota, sporofyt (kodiolové stádium) klíčí 4-bičíkatými zoosporami. Ulothrix zonata na kamenech v čistých potocích, řekách, jezerech. Enteromorpha - trubicovitá stélka z 1 vrstvy buněk, mořské pobřeží, též v tůních na slancových půdách. U nás v silně eutrofizovaných vodách (chloridy ze silnic či odpadních vod). 99
, které je jedinou diploidní buňkou v ŽC.")
102 TŘÍDA: DASYCLADOPHYCEAE Stélka (jediná sifonální buňka) měří několik cm, sestává z hlavní vzpřímené osy, která nese krátké, někdy srůstající boční přesleny větévek (gametangií). Inkrustace CaCO 3. K podkladu přirůstá systémem rhizoidů. CCW orientace bazálních tělísek bičíků zoidů. Zygota má velké diploidní jádro (až 100 μm), které je jedinou diploidní buňkou v ŽC. Jádro roste se zygotou, během vývoje prodělává meiotické dělení. Geologické stáří - největší druhové bohatství v křídě, významné horninotvorné rostliny. Acetabularia - na kamenech v litorálu Středozemního moře, 5-10 cm, v době rozmnožování připomíná malý deštníček, složitý životní cyklus, objekt pro karyologické a cytologické studie a_07_500x395_simplelifecycle.jpg TŘÍDA: TRENTEPOHLIOPHYCEAE Aerofytické řasy mírného až tropického pásu s heterotrichálními stélkami. Ze spleti vláken pokrývajících substrát vyrůstají vzpřímené větvičky nesoucí sporangia. Zoospory se 4, gamety se 2 bičíky. Trentepohlia - druhy dokonale adaptovány na růst v terestrických podmínkách, v buňkách jsou hnědé nebo červenohnědé pigmenty (směs sekundárních karotenoidů a polyhydroxyalkoholů) - možná jako ochrana proti UV. Zoosporangia na koncích vzpřímených větévek, ve zralosti se ulamují a mohou být unášena vzdušnými proudy. Gametangia na poléhavých vláknech, kde kondenzující kapky vody vytvářejí podmínky pro kopulaci. T. iolithus - fialková vůně, na kamenech při horských potocích, T. aurea - oranžově červené povlaky na skalách a zídkách, T. umbrina - hnědá, na borce stromů. 100
103 TŘÍDA: TREBOUXIOPHYCEAE (PLEURASTROPHYCEAE) Kokální nebo vláknité řasy, CCW orientace bazálních tělísek, uzavřená mitóza. Terestrické a aerofytické nebo sladkovodní řasy, fotobionti lišejníků. Pleurastrum, jak se zjistilo, patří do třídy Chlorophyceae. Trebouxia - četné druhy rostou ve stélkách lišejníků (fykobiont), některé rostou epifyticky Chlorella - zelenivka - biotechnologicky významná řasa, pěstuje se ve velkoobjemových kulturách, biomasa se používá jako přírodní léčivo, do krmných směsí..., Ch. vulgaris - plankton tůní Oocystis - plankton stojatých vod Microthamnion- dichotomicky či trichotomicky větvené keříky, větve mají oblé konce, mírně kyselé rašelinné tůňky Prasiola na vlhké půdě, zkadeřená lupenitá jednovrstevná stélka, buňky tvoří políčka po TŘÍDA: ZELENIVKY CHLOROPHYCEAE Druhově bohatá třída, volně žijící jednotliví nebo koloniální bičíkovci, kapsální (gleomorfní), kokální a vláknité řasy. Bičíky v počtu 2, 4 i více, jsou bez mastigonemat a stejně dlouhé. Mitóza je částečně otevřená, bazální tělíska bičíků mají CW, popř. DO orientaci, během cytokineze se vytvoří fykoplast. BS se u jednotlivých řádů liší: může chybět (nazí bičíkovci), u řádů Chlamydomonadales a Volvocales je glykoproteinová (chlamys), pevná a pružná, důležitou složkou je AK hydroxyprolin nebo u velké skupiny kokálních řas polysacharidová s chemicky rezistentní povrchovou vrstvou tvořenou sporopoleninem (oxidativní polymer sekundárních karotenoidů, který odolá i horké acetolýze, což je dlouhodobý var ve směsi 9 dílů anhydridu kyseliny octové a 1 dílu konc. kyseliny sírové), patří k nejodolnějším látkám v rostlinné říši a umožňuje bezvadnou fosilizaci. Některé jednobuněčné nebo vláknité zelenivky tvoří kolonie spojené slizem či zbytky mateřských stěn. Zvláštním typem kolonie je cenobium (jedna generace buněk, počet buněk 2 n, pravidelné uspořádání, někdy i morfologické, resp. funkční specializace buněk, rozmnožování dceřinými cenobii. Nepohlavní rozmnožování mitospory (n: G, 2n: S), zoospory v zoosporangiu, aplanospory (přeskočily stádium zoospory), hemiaplanospory (chybí bičík, mají stigma, pulsující vakuoly), autospory (po opuštění mateřské BS mají tvar jako mateřská buňka). Autospory na rozdíl od aplanospor nemají bičíkatá stádia. Všechno jsou to endogenní spory, při opouštění mateřské BS se uplatňují autolyziny. Bezblanní bičíkovci - schizotomie, podélné dělení. Oblaněné jednobuněčné řasy schizogonie - rozmnožovací či dceřiné buňky vznikají uvnitř mateřské buněčné stěny. Pohlavní rozmnožování druhy jednopohlavní a oboupohlavní, homo- a heterothalické, časté jsou feromony. Izogamie, anizogamie, oogamie. ŽC haplontní. Zástupci v 9 řádech, převážně sladkovodní, některé tvoří v rybnících zelený vegetační zákal (primární producenti), mnohé druhy mají biotechnologické využití (Scenedesmus). Řád: Chlamydomonadales - bičíkovci Chlamydomonas (pláštěnka) - je běžný bičíkovec stojatých a pomalu tekoucích vod a svrchní vrstvy půdy, dobře se pěstuje na umělých půdách, buňky mají kapkovitý tvar, chlamys tvoří na 101
, po vypuštění jímky lze nalézt na stěnách nádrže červenohnědé akinety Řád: Volvocales")
104 apikálním konci ztlustlinu (papilu), kterou prostupují bičíky, jejichž pohyb je při plavání synchronní Haematococcus typické jsou paprsčité plazmatické výběžky vyrůstající z protoplastu, častý obyvatel umělých vodních nádrží, jeho vegetační zbarvení je zprvu zelené, později červenohnědé (astaxanthin), po vypuštění jímky lze nalézt na stěnách nádrže červenohnědé akinety Řád: Volvocales cenobiální bičíkovci Volvox (váleč) - kulovitá či lehce oválná cenobia, buňky podobné chlamydomonádě v jedné vrstvě na obvodu cenobia ve slizových komůrkách (šestibokých nebo hvězdicovitých), ven vystupují dvojice bičíků, vnitřek cenobia zaplněn slizem tvorba dceřiných cenobií: ze specializovaných vegetativních buněk intenzívním dělením vznikne kulovitý útvar z 64 a více buněk, jejich bičíky směřují dovnitř, v konečném stádiu vývoje dojde k inverzi kolonie (vývoj dceřiného cenobia připomíná vývoj zárodku mnohobuněčných organismů morula, blastula, gastrula), po roztržení mateřského cenobia dceřiná cenobia vyrejdí ven, pohlavní proces: oogamie, cenobia jednodomá či dvoudomá, feromon feroforin se tvoří v anteridiích: V. globator - 0,8 mm, až 2000 buněk, rybníky a tůně Řád: Tetrasporales kapsální řasy Tetraspora - řasa s kapsální stélkou žijící ve slizu, ve vegetativním stavu nepohyblivá, horské potoky, okrajové buňky kolonie mají dvojice dlouhých pseudocilií (nepohyblivých bičíků, u nichž chybí centrální dvojice mikrotubulů) 102
105 Řád: Chlorococcales kokální řasy - molekulární taxonomie potvrzuje heterogenní povahu mnoha rodů, často dochází k přeřazení do třídy Trebouxiophyceae Chlorococcum povrchová vrstva půdy, sladkovodní biotopy Chlamydopodium (Characium) kokální, přisedá na ponořených rostlinách v tůních četné druhy kokálních zelenivek dominují v letním a podzimním planktonu, kde jsou hlavními primárními producenty: Kirchneriella, Coelastrum, Ankistrodesmus, Tetraedron, Scenedesmus Scenedesmus popsáno velké množství druhů, mnohé taxony jsou však ekomorfami (fenoplasticita variabilita založená na ekologických, popř. kultivačních podmínkách), cenobia tvoří 4,8,16 buněk v jedné či dvou řadách, rozmnožování autocenobii, nejběžnější S. quadricauda (= Desmodesmus communis), S. obliquus a S. acuminatus - často pěstování ve velkých objemech pro biotechnologické využití Řád: Sphaeropleales- mnohojaderné buňky, tzv. cenoblasty, resp. cenocyty Protosiphon - vakovité buňky dosahující velikosti až 1,5 mm, na vlhké půdě na okrajích vodních nádrží Pediastrum - buňky srostlé v plochých hvězdicovitých cenobiích, rybníky a tůně Hydrodictyon vodní síť - může zcela zaplňovat menší eutrofní tůně, buňky až 1 cm dlouhé spojeny v síť s šestibokými oky Sphaeroplea nevětvená vlákna, periodicky vysychavé tůně a vlhká půda Řád: Microsporales trichální stélky (nepravá vlákna) Microspora quadrata čisté, mírně kyselé vody, trichální stélka s nevětvenými nepravými vlákny, buňky s dvoudílnou BS, složenou ze 2 válcovitých prstenců, které do sebe zapadají v polovině buňky Řád: Oedogoniales - vláknité řasy s nepravými vlákny větvenými či nevětvenými, buňky i vlákna mají polární stavbu, BS ze 2 různě dlouhých dílů, nad srůstem obou částí se po každém dělení vytvoří charakteristický límeček: Nepohlavní rozmnožování: stefanokontní zoospory, na předním konci s věncem 40 párů bičíků. Pohlavní proces: oogamie s řadou zvláštností ve vývinu samčích gametangií. Rozlišujeme dvě skupiny druhů: 1. makrandrické - anteridia se vyvíjejí přímo z vegetativních buněk, v každém anteridiu vznikají 2 stefanokontní spermatozoidy, které po vyrejdění oplodní vaječnou buňku v oogoniu. 2. nanandrické - z vegetativních buněk vznikají androsporangia, v nich se vyvíjí 1 androspora s věncem bičíků, ta vyrejdí a přisedne na vlákno v blízkosti oogonia, zde vyroste v krátké vlákno (2-3 buňky), ve vrcholové buňce se tvoří 2 spermatozoidy. Oogonia produkují feromony (circein), otvírají se otvorem (mikropyle). Oedogonium - čapkoblanka - nevětvená vlákna, na ponořených větévkách v tůních 103

106 Bulbochaete - rovněž epifyticky na vodních rostlinách, vlákna větvená, boční větévky vyrůstají z cibulovitě ztlustlé bazální buňky a končí dlouhým chlupem on+the+same+filaments,+this+pecies+is+homothallic..jpg Řád: Chaetophorales - řasy s heterotrichální stélkou přirůstající k podkladu bazální buňkou, stélka bývá bohatě větvená, často rozlišena vlákna s podpůrnou funkcí a s funkcí asimilační. Apathococcus (zrněnka) jednobuněčná hojná aerofytická řasa, tvoří nepravidelné shluky buněk, zelené povlaky na kůře stromů či skalách, pravděpodobně dojde k přeřazení do tř. Trebouxiophyceae Draparnaldia - hlavní osní vlákno a bohatě větvená boční vlákna, uložena v řídkém slizu, mělké tůňky a kaluže s čistou vodou Chaetophora - rozvětvená vlákna tvoří 0,5-2 cm velké kolonie v pevném slizu, v tůních na ponořených rostlinách Fritschiella - nalezena v Indii, přizpůsobená růstu na vlhké půdě, bezbarvá plazivá vlákna pod povrchem půdy a zelená vzpřímená vlákna, dříve považována za možného předka suchozemských rostlin VÝVOJOVÁ LINIE: STREPTOFYTY STREPTOPHYTAE Obsahuje vývojové větve parožnatky, mechorosty a vyšší rostliny. Společným znakem je průběh jaderného a buněčného dělení: mitóza je zpravidla otevřená, oddělení dceřiných protoplastů zajišťuje fragmoplast. V životním cyklu mnohých druhů jsou bičíkaté buňky (zoospory, spermatozoidy), jejich stavba je podobná u parožnatek, mechorostů, některých kapraďorostů (2 bičíky) u jiných kapraďorostů a některých nahosemenných (cykasy až 20 bičíků). U spájivek, některých nahosemenných a všech krytosemenných rostlin bičíkatá stádia chybějí. Povrch zoidů (často i jejich bičíků) je pokryt drobnými polysacharidovými šupinami. Anatomická a funkční diferenciace rostlinného těla souvisí s postupnou adaptací na suchozemský způsob života. Evoluční tendence potlačování gametofytu ve prospěch sporofytu. 104
107 VÝVOJOVÁ VĚTEV: CHAROPHYTAE Je z fylogenetického hlediska výchozí pro zelené rostliny vývojových větví Bryophytae a Tracheophytae. ODDĚLENÍ: CHAROPHYTA Jednobuněčné a vláknité řasy a parožnatky jsou převážně sladkovodní, půdní a aerofytické řasy. Parožnatky jsou považovány za vývojové předky vyšších rostlin, jejich ekologie připomíná spíše zelené řasy. Životní cyklus je haplontní, zygota je jedinou diploidní buňkou. Fylogeneticky významná je stavba bičíkového aparátu. Bičíky vyrůstají pod vrcholem asymetrické buňky a směřují do strany. Bičíkový aparát je asymetrický vzhledem k ose buňky (unilaterální). Kinetozom (bazální tělísko) jednoho bičíku má mikrotubulární kořen ze 60 do plochy srostlých mikrotubulů (mnohovrstevná struktura, multilayered structure MLS). TŘÍDA: MESOSTIGMATOPHYCEAE Jedna z nejstarších linií zelených řas, snad společný předek vývojových linií Chlorophyta a Streptophyta. TŘÍDA: COLEOCHAETOPHYCEAE Coleochaete (štětinatka) - plochá terčíkovitá stélka, přirůstající spodní plochou k vodním rostlinám, vegetativní buňky mají po jednom chloroplastu, chloroplast se otáčí 1x za 2 minuty kolem své osy. Některé buňky vybíhají v dlouhou štětinu. Pohl. proces oogamie, lahvicovitá oogonia s dlouhým trichogynem, po oplození vznikne sporokarp (oospora s pseudoparenchymatickou vrstvou na povrchu), který přezimuje. Na jaře vyrejdí ze sporokarpu zoospory, které vyrostou v novou stélku. Životní cyklus štětinatky je podkladem koleochetové hypotézy vzniku mechorostů, je považován za klíčový v evoluci streptofyt. TŘÍDA: PAROŽNATKY CHAROPHYCEAE Makroskopické vodní rostliny, vzhledem připomínající přesličky, vzpřímená stélka dosahuje výšky 5 až 90 cm. Osu parožnatky tvoří dlouhé buňky článkové (internodia) a krátké uzlinové (nodi), z nich vyrůstají přesleny bočních větví. K podkladu přirůstají rhizoidy. Růst stélky je vrcholový, uskutečňuje se dělením 1 buňky, která odčleňuje vždy 1 buňku článkovou a 1 uzlinovou. Uzlinová se dále dělí a dá vznik a) bočním větévkám, b) korové vrstvě u r. Chara, c) gametangiím, d) obalu oogonia. Článková buňka (mladá je jednojaderná) pouze roste do délky, plně vyvinutá je několik cm dlouhá, 2-3 mm široká a je mnohojaderná. U většiny parožnatek jsou článkové buňky obaleny obalem z korových buněk. Buněčná stěna často inkrustovaná uhličitanem vápenatým (tvorba travertinů). Rozmnožování: fragmentací stélky, odnožováním a rozmnožovacími tělísky mezi rhizoidy. Pohlavní proces: oogamie, gametangia v paždí bočních větévek, druhy jedno- či dvoudomé. Oranžová kulovitá anteridia, na povrchu s 8 vypouklými štítky, ze středu štítku dovnitř směřuje kyjovitá buňka, z jejího konce vyrůstají spermatogenní vlákna, v jejich buňkách se po jednom vyvíjejí spermatozoidy (spirální tvar, 2 bičíky, na povrchu se šupinami). Oogonia oválná, v mládí zelená, oosféra obklopena několika spirálně vinutými buňkami. Tento obal (sporostegium slouží k výživě oogonia) je na vrcholu ukončen korunkou z 5 či 10 buněk. Po oplození srůstá stěna sporostegia se stěnou zygoty a vzniká sporokarp (černý). Klíčení po období klidu, mladá rostlinka je prochara: ze 2 vláken, jedno pozitivně geotropické - rhizoidy, druhé vzpřímené. 105


108 Parožnatky rostou ve stojatých či mírně tekoucích vodách, v řadě lokalit mají významný podíl na vzniku travertinu. V mořích chybějí, vyskytují se ve sladkých a brakických vodách. Fosilní zygospory gyrogonity. Chara - 5-buněčná korunka, články s korovou vrstvou Nitella - 10-buněčná korunka, korová vrstva často chybí, větve nesoucí gametangia se opakovaně větví TŘÍDA: ZYGNEMATOPHYCEAE (CONJUGATOPHYCEAE) - SPÁJIVKY Jednobuněčné nebo vláknité řasy. Buňky mají celistvou nebo dvoudílnou BS. Obsahují 1-2 chloroplasty a 1 jádro. V životním cyklu chybějí jakákoliv bičíkatá stádia. Hlavní způsob rozmnožování je vegetativní dělení. Pohlavní proces - spájení, konjugace - jako gamety vystupují celé protoplasty, schopné améboidního pohybu v prostoru, jež vymezují obě kopulující buňky. Ke styku gametů dochází v kopulačním kanálku či ve slizovém obalu, výsledkem je tlustostěnná zygospora. Životní cyklus je haplontní, jedinou 2n buňkou je zygospora. Žijí pouze ve sladkých vodách, hojné jsou v rašelinných tůních s nízkým ph. Systém a zástupci Řád Zygnematales - jařmatkotvaré jednobuněčné řasy nebo nevětvená vlákná se stejnocennými válcovitými buňkami. Na jejich povrchu je vrstva slizu. V buňce je 1 nebo více chloroplastů charakteristického tvaru (vodítko při určování rodů). Jádro v plazmatickém vaku ve středu buňky, větší část středového prostoru vyplňuje vakuola. Rozmnožování: fragmentací vlákna, ke spájení dochází koncem jara či začátkem léta. Chomáče vláken vystupují na hladinu. Typy konjugace: žebříková (skalariformní), nejčastější - kopulační kanálky se tvoří mezi dvěma různými vlákny, laterální - kopulují protoplasty sousedních buněk vlákna. 106
FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN
 DISTANČNÍ OPORY PRO KOMBINOVANÉ STUDIUM BIOLOGIE FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN Lenka Němcová UNIVERZITA JANA EVANGELISTY PURKYNĚ PŘÍRODOVĚDECKÁ FAKULTA katedra biologie Ústí nad Labem 2006 Cíl předmětu:
DISTANČNÍ OPORY PRO KOMBINOVANÉ STUDIUM BIOLOGIE FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN Lenka Němcová UNIVERZITA JANA EVANGELISTY PURKYNĚ PŘÍRODOVĚDECKÁ FAKULTA katedra biologie Ústí nad Labem 2006 Cíl předmětu:
Botanika bezcévných rostlin pro učitele 5. praktické cvičení
 Botanika bezcévných rostlin pro učitele 5. praktické cvičení ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae Chlamydomonas
Botanika bezcévných rostlin pro učitele 5. praktické cvičení ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae Chlamydomonas
Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae
Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae
Škola: Gymnázium, Brno, Slovanské náměstí 7 III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN
 Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů
 Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae ŘÁD: Bangiales Porphyra (HB) TŘÍDA: Florideophyceae ŘÁD: Gelidiales Gelidium (HB) ŘÁD: Gigartinales
Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae ŘÁD: Bangiales Porphyra (HB) TŘÍDA: Florideophyceae ŘÁD: Gelidiales Gelidium (HB) ŘÁD: Gigartinales
FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN
 Univerzita J. E. Purkyně v Ústí nad Labem Přírodovědecká fakulta FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN KBI/P301, KBI/K301 Milan Gryndler Lenka Němcová Ústí nad Labem 2013 0 Obor: Klíčová slova: biologie
Univerzita J. E. Purkyně v Ústí nad Labem Přírodovědecká fakulta FYLOGENEZE A SYSTÉM NIŽŠÍCH ROSTLIN KBI/P301, KBI/K301 Milan Gryndler Lenka Němcová Ústí nad Labem 2013 0 Obor: Klíčová slova: biologie
BOTANIKA BEZCÉVNÝCH ROSTLIN. pro odbornou biologii B120P76I, 3/2, Z + Zk, 6 kreditů
 BOTANIKA BEZCÉVNÝCH ROSTLIN pro odbornou biologii B120P76I, 3/2, Z + Zk, 6 kreditů Verze 2010 BOTANIKA BEZCÉVNÝCH ROSTLIN pro odbornou biologii B120P76I, 3/2, Z + Zk, 6 kreditů Karel Prášil prasil@natur.cuni.cz
BOTANIKA BEZCÉVNÝCH ROSTLIN pro odbornou biologii B120P76I, 3/2, Z + Zk, 6 kreditů Verze 2010 BOTANIKA BEZCÉVNÝCH ROSTLIN pro odbornou biologii B120P76I, 3/2, Z + Zk, 6 kreditů Karel Prášil prasil@natur.cuni.cz
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
ŘÍŠE (REGNUM): ROSTLINY (PLANTAE) Podříše (Subregnum): Nižší rostliny řasy (Thallobionta)
: ROSTLINY (PLANTAE) Podříše (Subregnum): Nižší rostliny řasy (Thallobionta)") ŘÍŠE (REGNUM): ROSTLINY (PLANTAE) Charakteristickým znakem všech jaderných organismů je eukaryotická buňka. Jaderné organismy jsou rozdělené do tří říší: rostliny, houby a živočichové. Charakteristickým
ŘÍŠE (REGNUM): ROSTLINY (PLANTAE) Charakteristickým znakem všech jaderných organismů je eukaryotická buňka. Jaderné organismy jsou rozdělené do tří říší: rostliny, houby a živočichové. Charakteristickým
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Glaucophyta TŘÍDA: Glaucophyceae Glaucocystis (PP) ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae Porphyridium
Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Glaucophyta TŘÍDA: Glaucophyceae Glaucocystis (PP) ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae Porphyridium
NIŽŠÍ ROSTLINY - řasy
 Maturitní téma č.8 NIŽŠÍ ROSTLINY - řasy CHARAKTERISTICKÉ ZNAKY ŘAS Rostlinnou říši dělíme na dvě základní skupiny - nižší rostliny (Algobionta) a vyšší rostliny (Cormobionta). Mezi nižší rostliny řadíme
Maturitní téma č.8 NIŽŠÍ ROSTLINY - řasy CHARAKTERISTICKÉ ZNAKY ŘAS Rostlinnou říši dělíme na dvě základní skupiny - nižší rostliny (Algobionta) a vyšší rostliny (Cormobionta). Mezi nižší rostliny řadíme
Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů
 Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta TŘÍDA: Dinophyceae ŘÁD: Gonyaulacales Ceratium (TP) ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta
Botanika - bezcévné rostliny 2. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta TŘÍDA: Dinophyceae ŘÁD: Gonyaulacales Ceratium (TP) ŘÍŠE: Protozoa ODDĚLENÍ: Dinophyta
ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY
 ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY Řasy (dříve nesprávně označovány jako podříše nižších rostlin v rámci rostlinné říše) představují velmi různorodou skupinu organismů od několika mikrometrů velkých
ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY Řasy (dříve nesprávně označovány jako podříše nižších rostlin v rámci rostlinné říše) představují velmi různorodou skupinu organismů od několika mikrometrů velkých
Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.
 Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016 Kapitoly z biologie sinic a řas Petr Hašler Katedra
Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016 Kapitoly z biologie sinic a řas Petr Hašler Katedra
Barbora Chattová. Fylogeneze a diverzita řas a hub: 2. přednáška Euglenophyta, Dinophyta, Cryptophyta
 Barbora Chattová Fylogeneze a diverzita řas a hub: 2. přednáška Euglenophyta, Dinophyta, Cryptophyta Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé řasy) Rhodophyta
Barbora Chattová Fylogeneze a diverzita řas a hub: 2. přednáška Euglenophyta, Dinophyta, Cryptophyta Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé řasy) Rhodophyta
Otázka: Nižší rostliny. Předmět: Biologie. Přidal(a): Evka NIŽŠÍ ROSTLINY= PROTOBIONTA
: Evka NIŽŠÍ ROSTLINY= PROTOBIONTA") Otázka: Nižší rostliny Předmět: Biologie Přidal(a): Evka NIŽŠÍ ROSTLINY= PROTOBIONTA oddělení Ruduchy, Zelené řasy a Mechorosty eukaryotické autotrofní organismy tělo tvoří jednobuněčná nebo mnohobuněčná
Otázka: Nižší rostliny Předmět: Biologie Přidal(a): Evka NIŽŠÍ ROSTLINY= PROTOBIONTA oddělení Ruduchy, Zelené řasy a Mechorosty eukaryotické autotrofní organismy tělo tvoří jednobuněčná nebo mnohobuněčná
ORGANISMY A SYSTÉM ŘASY A MECHOROSTY
 Zelené řasy Zelené řasy je významné oddělení jednobuněčných i mnohobuněčných stélkatých zelených rostlin. Představují blízké příbuzné vyšších rostlin, které se z jedné linie zelených řas vyvinuly. Stavba-
Zelené řasy Zelené řasy je významné oddělení jednobuněčných i mnohobuněčných stélkatých zelených rostlin. Představují blízké příbuzné vyšších rostlin, které se z jedné linie zelených řas vyvinuly. Stavba-
Botanika bezcévných rostlin pro učitele 1. praktické cvičení
 Botanika bezcévných rostlin pro učitele 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Botanika bezcévných rostlin pro učitele 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
TŘÍDA: TREBOUXIOPHYCEAE
 TREBOUXIOPHYCEAE TŘÍDA: TREBOUXIOPHYCEAE Tato třída je zatím nejméně jasnou skupinou v novém systému zelených řas. Podle fragmentárních poznatků jsou do třídy Pleurastrophyceae řazeny kokální a vláknité
TREBOUXIOPHYCEAE TŘÍDA: TREBOUXIOPHYCEAE Tato třída je zatím nejméně jasnou skupinou v novém systému zelených řas. Podle fragmentárních poznatků jsou do třídy Pleurastrophyceae řazeny kokální a vláknité
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ
 Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíle praktického cvičení z Botaniky bezcévných rostlin: - na konkrétním materiálu se seznámit s reprezentativními zástupci
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíle praktického cvičení z Botaniky bezcévných rostlin: - na konkrétním materiálu se seznámit s reprezentativními zástupci
Systematická biologie,její minulý a současný vývoj
 Systematická biologie,její minulý a současný vývoj Systematická biologie - věda o rozmanitosti organismů. Jejím úkolem je vytvořit informační soustavu, která poskytuje srovnatelné údaje o všech známých
Systematická biologie,její minulý a současný vývoj Systematická biologie - věda o rozmanitosti organismů. Jejím úkolem je vytvořit informační soustavu, která poskytuje srovnatelné údaje o všech známých
Biologie - Kvinta, 1. ročník
 - Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
- Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
trubicovitá pletivná vláknitá, větvená vláknitá
 ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
Část I. Fylogeneze a systém niţších rostlin Cíl předmětu:
 Část I. Fylogeneze a systém niţších rostlin Cíl předmětu: Velmi stručně se seznámit s morfologií, systémem, ekologií, historickým vývojem a významem organismů, které tradičně studují vědecké obory algologie,
Část I. Fylogeneze a systém niţších rostlin Cíl předmětu: Velmi stručně se seznámit s morfologií, systémem, ekologií, historickým vývojem a významem organismů, které tradičně studují vědecké obory algologie,
Třída: XANTHOPHYCEAE
 HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
Lesnická botanika speciální přednáška 3
 Lesnická botanika speciální přednáška 3 Řasy Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Fylogenetický strom www.sinicearasy.cz Glaucocystis
Lesnická botanika speciální přednáška 3 Řasy Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Fylogenetický strom www.sinicearasy.cz Glaucocystis
Saprofité-rozklad org. zbytků Paraziticky- mykosy... Symbioticky- s cévnatými rostlinami(mykorhiza)- 95% rostlinných druhů, rostlina poskytuje
- 95% rostlinných druhů, rostlina poskytuje") Otázka: Houby a nižší rostliny Předmět: Biologie Přidal(a): LenkaKrchova Houby fungia Samostatná říše- napůl živočich a rostlina Eukaryotické heterotrofní organismy, které se rozmnožují výtrusy. Tělo se
Otázka: Houby a nižší rostliny Předmět: Biologie Přidal(a): LenkaKrchova Houby fungia Samostatná říše- napůl živočich a rostlina Eukaryotické heterotrofní organismy, které se rozmnožují výtrusy. Tělo se
Botanika bezcévných rostlin pro učitele 3. praktické cvičení
 Botanika bezcévných rostlin pro učitele 3. praktické cvičení ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Phaeophyceae ŘÁD: Dictyotales Dictyota (HB) Padina (HB) ŘÁD: Fucales Fucus
Botanika bezcévných rostlin pro učitele 3. praktické cvičení ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Phaeophyceae ŘÁD: Dictyotales Dictyota (HB) Padina (HB) ŘÁD: Fucales Fucus
URČOVÁNÍ SINIC A ŘAS Jak na to? Logicky a jednoduše stačí se pozorně v klidu dívat a srovnávat
 URČOVÁNÍ SINIC A ŘAS Jak na to? Logicky a jednoduše stačí se pozorně v klidu dívat a srovnávat RNDr. Lenka Šejnohová, Ph.D. Mikrobiologický ústav Akademie věd ČR, Třeboň www.alga.cz eustigmatos@gmail.com
URČOVÁNÍ SINIC A ŘAS Jak na to? Logicky a jednoduše stačí se pozorně v klidu dívat a srovnávat RNDr. Lenka Šejnohová, Ph.D. Mikrobiologický ústav Akademie věd ČR, Třeboň www.alga.cz eustigmatos@gmail.com
Buňka. Kristýna Obhlídalová 7.A
 Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
Vyšší rostliny Embryophyta. Milan Štech, PřF JU
 Vyšší rostliny Embryophyta Milan Štech, PřF JU = suchozemské rostliny Embryophyta * mechorosty * cévnaté rostliny * jejich společní předci/přímí předchůdci Vznik chloroplastu klíčová událost na cestě k
Vyšší rostliny Embryophyta Milan Štech, PřF JU = suchozemské rostliny Embryophyta * mechorosty * cévnaté rostliny * jejich společní předci/přímí předchůdci Vznik chloroplastu klíčová událost na cestě k
Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus TŘÍDA: Synurophyceae Synura
Botanika bezcévných rostlin 3. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Chromista (Chromalveolata) ODDĚLENÍ: Heterokontophyta TŘÍDA: Chrysophyceae Hydrurus foetidus TŘÍDA: Synurophyceae Synura
TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají
 a neustále se ukazuje, že do ní spadají") TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají nové a nové rody. Stavbou stélek a rozmnožováním i ekologií
TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají nové a nové rody. Stavbou stélek a rozmnožováním i ekologií
Barbora Chattová. Fylogeneze a diverzita rostlin: řasy a sinice
 Barbora Chattová Fylogeneze a diverzita rostlin: řasy a sinice Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé řasy) Rhodophyta (ruduchy) Chlorophyta (zelené řasy)
Barbora Chattová Fylogeneze a diverzita rostlin: řasy a sinice Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé řasy) Rhodophyta (ruduchy) Chlorophyta (zelené řasy)
 Evoluce (nejen) rostlinné buňky Martin Potocký laboratoř buněčné biologie ÚEB AV ČR, v.v.i. potocky@ueb.cas.cz http://www.ueb.cas.cz Evoluce rostlinné buňky Vznik a evoluce eukaryotních organismů strom
Evoluce (nejen) rostlinné buňky Martin Potocký laboratoř buněčné biologie ÚEB AV ČR, v.v.i. potocky@ueb.cas.cz http://www.ueb.cas.cz Evoluce rostlinné buňky Vznik a evoluce eukaryotních organismů strom
Heterokontní (oddělení Heterokontophyta), skrytěnky (Cryptophyta), obrněnky (Dinophyta), krásnoočka (Euglenophyta)
, skrytěnky (Cryptophyta), obrněnky (Dinophyta), krásnoočka (Euglenophyta)") Heterokontní (oddělení Heterokontophyta), skrytěnky (Cryptophyta), obrněnky (Dinophyta), krásnoočka (Euglenophyta) Oddělení Heterokontophyta Pleuronematický bičík (pohybový) Akronematický bičík Chromatofory
Heterokontní (oddělení Heterokontophyta), skrytěnky (Cryptophyta), obrněnky (Dinophyta), krásnoočka (Euglenophyta) Oddělení Heterokontophyta Pleuronematický bičík (pohybový) Akronematický bičík Chromatofory
Barbora Chattová. Fylogeneze a diverzita rostlin 1. přednáška Cyanobacteria, Euglenophyta, Dinophyta, Cryptophyta
 Barbora Chattová Fylogeneze a diverzita rostlin 1. přednáška Cyanobacteria, Euglenophyta, Dinophyta, Cryptophyta Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé
Barbora Chattová Fylogeneze a diverzita rostlin 1. přednáška Cyanobacteria, Euglenophyta, Dinophyta, Cryptophyta Euglenophyta (krásnoočka) Cryptophyta (skrytěnky) Dinophyta (obrněnky) Chromophyta (hnědé
Heterokontní řasy a ruduchy (oddělení Heterokontophyta, Rhodophyta)
") Heterokontní řasy a ruduchy (oddělení Heterokontophyta, Rhodophyta) Oddělení Heterokontophyta Pleuronematický bičík (pohybový) Akronematický bičík Chromatofory se 4 membránami Chlorofyl a, c Fukoxantin,
Heterokontní řasy a ruduchy (oddělení Heterokontophyta, Rhodophyta) Oddělení Heterokontophyta Pleuronematický bičík (pohybový) Akronematický bičík Chromatofory se 4 membránami Chlorofyl a, c Fukoxantin,
chlorofyl a chlorofyl c
 Systém a evoluce řas HEN Řasy = Algae různé autotrofní fotosyntetizující vodní organismy, které nemusí být navzájem příliš příbuzné Fylogeneticky jde o nesourodou skupinu Systematicky jsou řazeny do dvou
Systém a evoluce řas HEN Řasy = Algae různé autotrofní fotosyntetizující vodní organismy, které nemusí být navzájem příliš příbuzné Fylogeneticky jde o nesourodou skupinu Systematicky jsou řazeny do dvou
Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů "říše" Plantae; podříše Viridiplantae Oddělení: Chlorophyta; třída: Ulvophyceae a) Ulva b) Cladophora c) Anadyomene d) Valonia
Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů "říše" Plantae; podříše Viridiplantae Oddělení: Chlorophyta; třída: Ulvophyceae a) Ulva b) Cladophora c) Anadyomene d) Valonia
Botanika bezcévných rostlin 1. praktické cvičení
 Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Aplikované vědy. Hraniční obory o ţivotě
 BIOLOGICKÉ VĚDY Podle zkoumaného organismu Mikrobiologie (viry, bakterie) Mykologie (houby) Botanika (rostliny) Zoologie (zvířata) Antropologie (člověk) Hydrobiologie (vodní organismy) Pedologie (půda)
BIOLOGICKÉ VĚDY Podle zkoumaného organismu Mikrobiologie (viry, bakterie) Mykologie (houby) Botanika (rostliny) Zoologie (zvířata) Antropologie (člověk) Hydrobiologie (vodní organismy) Pedologie (půda)
Maturitní témata - BIOLOGIE 2018
 Maturitní témata - BIOLOGIE 2018 1. Obecná biologie; vznik a vývoj života Biologie a její vývoj a význam, obecná charakteristika organismů, přehled živých soustav (taxonomie), Linného taxony, binomická
Maturitní témata - BIOLOGIE 2018 1. Obecná biologie; vznik a vývoj života Biologie a její vývoj a význam, obecná charakteristika organismů, přehled živých soustav (taxonomie), Linného taxony, binomická
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Třída: RAPHIDOPHYCEAE
 HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají
 a neustále se ukazuje, že do ní spadají") TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají nové a nové rody. Stavbou stélek a rozmnožováním i ekologií
TŘÍDA: TREBOUXIOPHYCEAE Tato třída začala být uznávána teprve nedávno (původně pod názvem Pleurastrophyceae) a neustále se ukazuje, že do ní spadají nové a nové rody. Stavbou stélek a rozmnožováním i ekologií
Buňka buňka je základní stavební a funkční jednotka živých organismů
 Buňka - buňka je základní stavební a funkční jednotka živých organismů - je pozorovatelná pouze pod mikroskopem - na Zemi existuje několik typů buněk: 1. buňky bez jádra (prokaryotní buňky)- bakterie a
Buňka - buňka je základní stavební a funkční jednotka živých organismů - je pozorovatelná pouze pod mikroskopem - na Zemi existuje několik typů buněk: 1. buňky bez jádra (prokaryotní buňky)- bakterie a
PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY
 PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY 2010 Ing. Andrea Sikorová, Ph.D. 1 Problémy životního prostředí - organismy V této kapitole se dozvíte: Co je to organismus. Z čeho se organismus skládá. Jak se dělí
PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY 2010 Ing. Andrea Sikorová, Ph.D. 1 Problémy životního prostředí - organismy V této kapitole se dozvíte: Co je to organismus. Z čeho se organismus skládá. Jak se dělí
Obecná charakteristika hub
 Fyziologie hub Prvá část: Charakteristiku hub na základě výživy Ekologická charakteristika výživy hub Chemické zdroje výživy hub Druhá část Fyziologie růstu a rozmnožování Způsoby stanovení růstu, způsoby
Fyziologie hub Prvá část: Charakteristiku hub na základě výživy Ekologická charakteristika výživy hub Chemické zdroje výživy hub Druhá část Fyziologie růstu a rozmnožování Způsoby stanovení růstu, způsoby
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka
 primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
Buňka. Autor: Mgr. Jitka Mašková Datum: Gymnázium, Třeboň, Na Sadech 308
 Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
Maturitní témata Biologie MZ 2017
 Maturitní témata Biologie MZ 2017 1. Buňka - stavba a funkce buněčných struktur - typy buněk - prokaryotní buňka - eukaryotní buňka - rozdíl mezi rostlinnou a živočišnou buňkou - buněčný cyklus - mitóza
Maturitní témata Biologie MZ 2017 1. Buňka - stavba a funkce buněčných struktur - typy buněk - prokaryotní buňka - eukaryotní buňka - rozdíl mezi rostlinnou a živočišnou buňkou - buněčný cyklus - mitóza
Zemědělská botanika. joza@zf.jcu.czzf.jcu.cz
 Zemědělská botanika Vít Joza joza@zf.jcu.czzf.jcu.cz Biologický systém (část 1) říše: Protozoa (prvoci) oddělení: Myxomycota (hlenky) Euglenophyta (krásnoočka, eugleny) Dinophyta (obrněnky) říše: Chromista
Zemědělská botanika Vít Joza joza@zf.jcu.czzf.jcu.cz Biologický systém (část 1) říše: Protozoa (prvoci) oddělení: Myxomycota (hlenky) Euglenophyta (krásnoočka, eugleny) Dinophyta (obrněnky) říše: Chromista
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota)
") Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota) Sinice celková charakteristika Sinice jsou velmi drobné a velmi jednoduché autotrofní prokaryotické organizmy. Jsou evolučně nesmírně staré a jsou schopné
Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota) Sinice celková charakteristika Sinice jsou velmi drobné a velmi jednoduché autotrofní prokaryotické organizmy. Jsou evolučně nesmírně staré a jsou schopné
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
- význam: ochranná funkce, dodává buňce tvar. jádro = karyon, je vyplněné karyoplazmou ( polotekutá tekutina )
") Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
Třída: RAPHIDOPHYCEAE
 HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
HYDROBOTANIKA Třída: RAPHIDOPHYCEAE Celková charakteristika Malá skupina jednobuněčných bičíkovců zahrnuje jen devět rodů. Jejich buňky jsou poměrně velké (až 100 µm). Žijí jak ve sladké vodě, tak i v
Zelené řasy (Chlorophyta) Milan Dundr
 Milan Dundr") Zelené řasy (Chlorophyta) Milan Dundr Barviva barviva v chloroplastech: chlorofyl a chlorofyl b -karoten xantofyly Chlorofyl a, chlorofyl b zelené řasy zásobní látka: škrob často pyrenoid bílkovinné tělísko
Zelené řasy (Chlorophyta) Milan Dundr Barviva barviva v chloroplastech: chlorofyl a chlorofyl b -karoten xantofyly Chlorofyl a, chlorofyl b zelené řasy zásobní látka: škrob často pyrenoid bílkovinné tělísko
Typy stélek. Rozdělení řas, charakteristické znaky hlavních skupin a jejich systematické zařazení; ekologický význam, řasy jako symbiotické organismy.
 Otázka: Řasy Předmět: Biologie Přidal(a): Don Typy stélek. Rozdělení řas, charakteristické znaky hlavních skupin a jejich systematické zařazení; ekologický význam, řasy jako symbiotické organismy. Řasy
Otázka: Řasy Předmět: Biologie Přidal(a): Don Typy stélek. Rozdělení řas, charakteristické znaky hlavních skupin a jejich systematické zařazení; ekologický význam, řasy jako symbiotické organismy. Řasy
Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky.
 Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky. Materiál je plně funkční pouze s použitím internetu. základní projevy života
Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky. Materiál je plně funkční pouze s použitím internetu. základní projevy života
Číslo a název projektu Číslo a název šablony
 Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva
 a kokální stélkou. Pohyb zajišťují dva") CHLAMYDOPHYCEAE TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva až čtyři stejně dlouhé bičíky vyrůstající
CHLAMYDOPHYCEAE TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva až čtyři stejně dlouhé bičíky vyrůstající
PRAPRVOCI A PRVOCI Vojtěch Maša, 2009
 PRAPRVOCI A PRVOCI Vojtěch Maša, 2009 Opakování Prokarytotické organismy Opakování Prokaryotické organismy Nemají jádro, ale jen 1 chromozóm neoddělený od cytoplazmy membránou Patří sem archea, bakterie
PRAPRVOCI A PRVOCI Vojtěch Maša, 2009 Opakování Prokarytotické organismy Opakování Prokaryotické organismy Nemají jádro, ale jen 1 chromozóm neoddělený od cytoplazmy membránou Patří sem archea, bakterie
-1-slovné názvosloví rod (latinsky velkým písmenem; česky malým písmenem)
") Otázka: Nižší rostliny Předmět: Biologie Přidal(a): Peta NIŽŠÍ ROSTLINY A MECHOROSTY SYSTÉM, CHARAKTERISTIKA A VÝZNAM KLASIFIKACE ROSTLIN -18. st. C. LINNÉ vytvořil 1. smysluplný systém živých organismů,
Otázka: Nižší rostliny Předmět: Biologie Přidal(a): Peta NIŽŠÍ ROSTLINY A MECHOROSTY SYSTÉM, CHARAKTERISTIKA A VÝZNAM KLASIFIKACE ROSTLIN -18. st. C. LINNÉ vytvořil 1. smysluplný systém živých organismů,
Buňka. Buňka (cellula) základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách
 základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách") Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Diverzita autotrofních protist
 Diverzita autotrofních protist Po stopách evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Protista Eukaryotické organismy s jednoduchou organizací stélky Dlouhá evoluční historie Protista
Diverzita autotrofních protist Po stopách evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Protista Eukaryotické organismy s jednoduchou organizací stélky Dlouhá evoluční historie Protista
Oddělení: CRYPTOPHYTA
 HYDROBOTANIKA Oddělení: CRYPTOPHYTA Celková charakteristika Jsou to volně žijící bičíkovci, mořští i sladkovodní, důležití jsou zejména pro plankton jarních stojatých vod. Oddělení: CRYPTOPHYTA Stavba
HYDROBOTANIKA Oddělení: CRYPTOPHYTA Celková charakteristika Jsou to volně žijící bičíkovci, mořští i sladkovodní, důležití jsou zejména pro plankton jarních stojatých vod. Oddělení: CRYPTOPHYTA Stavba
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
= primitivní vyšší rostliny, primárně suchozemské. pravděpodobně se vyvinuly z řas řádu Charales nemají pravé cévní svazky
 MECHOROSTY = primitivní vyšší rostliny, primárně suchozemské první fosilie ze svrchního devonu (před 360 mil. let) pravděpodobně se vyvinuly z řas řádu Charales nemají pravé cévní svazky -odkázány na kapilární
MECHOROSTY = primitivní vyšší rostliny, primárně suchozemské první fosilie ze svrchního devonu (před 360 mil. let) pravděpodobně se vyvinuly z řas řádu Charales nemají pravé cévní svazky -odkázány na kapilární
Botanika bezcévných rostlin 2. praktické cvičení. Přehled pozorovaných objektů
 Botanika bezcévných rostlin 2. praktické cvičení ŘÍŠE: Amoebozoa ODDĚLENÍ: Myxomycota TŘÍDA: Myxomycetes Přehled pozorovaných objektů Myxomycetes trofická fáze (OP) ŘÁD: Liceales Lycogala (HB) ŘÁD: Physarales
Botanika bezcévných rostlin 2. praktické cvičení ŘÍŠE: Amoebozoa ODDĚLENÍ: Myxomycota TŘÍDA: Myxomycetes Přehled pozorovaných objektů Myxomycetes trofická fáze (OP) ŘÁD: Liceales Lycogala (HB) ŘÁD: Physarales
Algologie ve výuce biologie a přírodopisu
 Algologie ve výuce biologie a přírodopisu Petra Vágnerová, Plzeň Algologie je věda, která zkoumá sinice a řasy. Tyto skupiny nejsou v rámci biologie a přírodopisu probírány jako jeden tematický celek,
Algologie ve výuce biologie a přírodopisu Petra Vágnerová, Plzeň Algologie je věda, která zkoumá sinice a řasy. Tyto skupiny nejsou v rámci biologie a přírodopisu probírány jako jeden tematický celek,
prokaryotní Znaky prokaryoty
 prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
Název: ŘASY Autor: PaedDr. Ludmila Pipková
 Název: ŘASY Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie, geografie Ročník: 2. a 3. (1. ročník vyššího gymnázia)
Název: ŘASY Autor: PaedDr. Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie, geografie Ročník: 2. a 3. (1. ročník vyššího gymnázia)
Impérium: Eukarya. Oddělení: Dinophyta (Dinoflagellata) - obrněnky. Oddělení: Euglenophyta krásnoočka, eugleny. Oddělení: Cryptophyta skrytěnky
 - obrněnky. Oddělení: Euglenophyta krásnoočka, eugleny. Oddělení: Cryptophyta skrytěnky") Systém sinic a řas Impérium: Prokarya Říše: Bacteria Oddělení: Cyanophyta (Cyanobacteria) sinice Třída : Cyanophyceae- sinice Řád: Chroococcales Řád: Oscillatoriales Řád: Nostocales Impérium: Eukarya Říše:
Systém sinic a řas Impérium: Prokarya Říše: Bacteria Oddělení: Cyanophyta (Cyanobacteria) sinice Třída : Cyanophyceae- sinice Řád: Chroococcales Řád: Oscillatoriales Řád: Nostocales Impérium: Eukarya Říše:
Vzdělávací obsah vyučovacího předmětu
 Vzdělávací obsah vyučovacího předmětu Přírodopis 6. ročník Zpracovala: RNDr. Šárka Semorádová Obecná biologie rozliší základní projevy a podmínky života, orientuje se v daném přehledu vývoje organismů
Vzdělávací obsah vyučovacího předmětu Přírodopis 6. ročník Zpracovala: RNDr. Šárka Semorádová Obecná biologie rozliší základní projevy a podmínky života, orientuje se v daném přehledu vývoje organismů
ZELENÉ ŘASY (CHLOROPHYTA)
") ZELENÉ ŘASY (CHLOROPHYTA) metodický list Zelené řasy lze rozdělit dle tříd, toto rozdělení je ale pro žáky málo srozumitelné a navíc systém není jednotný. Volila jsem proto rozdělení podle typů stélky.
ZELENÉ ŘASY (CHLOROPHYTA) metodický list Zelené řasy lze rozdělit dle tříd, toto rozdělení je ale pro žáky málo srozumitelné a navíc systém není jednotný. Volila jsem proto rozdělení podle typů stélky.
Stavba dřeva. Základy cytologie. přednáška
 Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
World of Plants Sources for Botanical Courses
 Botanika 2 Prokarya Impérium Prokarya (Prokaryota) jediná říše Bacteria (bakterie) jednobuněčné organismy stélky jednobuněčné nebo vláknité žijí jednotlivě nebo v koloniích patří k nejstarším a nejrozšířenějším
Botanika 2 Prokarya Impérium Prokarya (Prokaryota) jediná říše Bacteria (bakterie) jednobuněčné organismy stélky jednobuněčné nebo vláknité žijí jednotlivě nebo v koloniích patří k nejstarším a nejrozšířenějším
Jaro 2010 Kateřina Slavíčková
 Jaro 2010 Kateřina Slavíčková Obsah: 1. Biologické vědy. 2. Chemie a fyzika v biologii koloběh látek a tok energie. 3. Buňka, tkáně, pletiva, orgány, orgánové soustavy, organismus. 4. Metabolismus. 5.
Jaro 2010 Kateřina Slavíčková Obsah: 1. Biologické vědy. 2. Chemie a fyzika v biologii koloběh látek a tok energie. 3. Buňka, tkáně, pletiva, orgány, orgánové soustavy, organismus. 4. Metabolismus. 5.
Mgr. Šárka Bidmanová, Ph.D.
 Mgr. Šárka Bidmanová, Ph.D. Loschmidtovy laboratoře, Ústav experimentální biologie Přírodovědecká fakulta, Masarykova univerzita 77580@mail.muni.cz 1. Úvod do studia mikrobiologie 2. Archea 3. Bakterie
Mgr. Šárka Bidmanová, Ph.D. Loschmidtovy laboratoře, Ústav experimentální biologie Přírodovědecká fakulta, Masarykova univerzita 77580@mail.muni.cz 1. Úvod do studia mikrobiologie 2. Archea 3. Bakterie
Evoluce primárních producentů. Pavel Škaloud katedra botaniky PřF UK
 Evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Primární produkce Více jak polovinu primární produkce mají na svědomí řasy Sinice a řasy Carl Linné (1758) Sinice a řasy Sinice a řasy
Evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Primární produkce Více jak polovinu primární produkce mají na svědomí řasy Sinice a řasy Carl Linné (1758) Sinice a řasy Sinice a řasy
H O U B Y. (Fungi, Mycota) B. Voženílková
 B. Voženílková") H O U B Y (Fungi, Mycota) B. Voženílková Charakteristické rysy hub Houby mají ze všech původců rostlinných chorob největší význam. Ve středoevropských podmínkách je jimi vyvoláno asi 82-84 % všech ekonomicky
H O U B Y (Fungi, Mycota) B. Voženílková Charakteristické rysy hub Houby mají ze všech původců rostlinných chorob největší význam. Ve středoevropských podmínkách je jimi vyvoláno asi 82-84 % všech ekonomicky
Po stopách rané evoluce primárních producentů. Pavel Škaloud katedra botaniky PřF UK
 Po stopách rané evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Protista Eukaryotické organismy s jednoduchou organizací stélky Dlouhá evoluční historie Protista Protista Historicky
Po stopách rané evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Protista Eukaryotické organismy s jednoduchou organizací stélky Dlouhá evoluční historie Protista Protista Historicky
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162
 ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
Základy buněčné biologie
 Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota)
") Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota) Sinice celková charakteristika Sinice jsou velmi drobné a velmi jednoduché autotrofní prokaryotické organizmy. Jsou evolučně nesmírně staré a jsou schopné
Sinice Cyanobacteria (Cyanophyta, Cyanoprokaryota) Sinice celková charakteristika Sinice jsou velmi drobné a velmi jednoduché autotrofní prokaryotické organizmy. Jsou evolučně nesmírně staré a jsou schopné
základní přehled organismů
 základní přehled organismů Všechny tyto organismy mají podobný chemický základ Doména Archaea Tato doména nebyla rozpoznána až do konce 70. let minulého století Co se týče morfologie, neliší se archeální
základní přehled organismů Všechny tyto organismy mají podobný chemický základ Doména Archaea Tato doména nebyla rozpoznána až do konce 70. let minulého století Co se týče morfologie, neliší se archeální
RNDr. Bohuslav Uher, Ph.D.
 Masarykova univerzita, Přírodovědecká fakulta, Ústav botaniky a zoologie Chromista - Heterokontophyta Phaeophyceae & Plantae - Glaucophyta, Rhodophyta 4. přednáška z cyklu přednášek předmětu Bi1090 Systém
Masarykova univerzita, Přírodovědecká fakulta, Ústav botaniky a zoologie Chromista - Heterokontophyta Phaeophyceae & Plantae - Glaucophyta, Rhodophyta 4. přednáška z cyklu přednášek předmětu Bi1090 Systém
Od sinice k první kytce...
 Od sinice k první kytce... Po stopách evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Primární produkce Více jak polovinu primární produkce mají na svědomí řasy Primární producenti
Od sinice k první kytce... Po stopách evoluce primárních producentů Pavel Škaloud katedra botaniky PřF UK Primární produkce Více jak polovinu primární produkce mají na svědomí řasy Primární producenti
základní přehled organismů
 základní přehled organismů Doména Archaea Tato doména nebyla rozpoznána až do konce 70. let minulého století Co se týče morfologie, neliší se archeální buňky od buněk bakteriálních Rozdíly jsou biochemické
základní přehled organismů Doména Archaea Tato doména nebyla rozpoznána až do konce 70. let minulého století Co se týče morfologie, neliší se archeální buňky od buněk bakteriálních Rozdíly jsou biochemické
plodnice většinou makroskopický útvar vyrůstají za příznivých podmínek z podhoubí a sloužící k rozšíření výtrusů (jen u vyšších hub)
") Otázka: Houby Předmět: Biologie Přidal(a): cathrinefirth CHARAKTERISTIKA početná a různorodá skupina znaky rostlin (nepohyblivost) i znaky živočichů (heterotrofní výživa org. látky, zásobní látka glykogen)
Otázka: Houby Předmět: Biologie Přidal(a): cathrinefirth CHARAKTERISTIKA početná a různorodá skupina znaky rostlin (nepohyblivost) i znaky živočichů (heterotrofní výživa org. látky, zásobní látka glykogen)
ŠVP Gymnázium Ostrava-Zábřeh. 4.8.10. Seminář a cvičení z biologie
 4.8.10. Seminář a cvičení z biologie Volitelný předmět Seminář a cvičení z biologie je koncipován jako předmět, který vychází ze vzdělávací oblasti Člověk a příroda Rámcového vzdělávacího programu pro
4.8.10. Seminář a cvičení z biologie Volitelný předmět Seminář a cvičení z biologie je koncipován jako předmět, který vychází ze vzdělávací oblasti Člověk a příroda Rámcového vzdělávacího programu pro
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Biologická klasifikace rostlin BOT/BIKR Petr Hašler Katedra botaniky PřF UP Nižší rostlina-vymezení pojmu Tělo rostliny je tvořeno
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Biologická klasifikace rostlin BOT/BIKR Petr Hašler Katedra botaniky PřF UP Nižší rostlina-vymezení pojmu Tělo rostliny je tvořeno
ÚVOD MECHOROSTY. Ivana Lipnerová
 ÚVOD MECHOROSTY Ivana Lipnerová ÚVOD - ORGANIZACE pokyny v informačním e-mailu zápočet max 2 absence protokoly morfologický herbář do 31. 5. 2013! http://www.botanickafotogalerie.cz/ zásady bezpečné práce
ÚVOD MECHOROSTY Ivana Lipnerová ÚVOD - ORGANIZACE pokyny v informačním e-mailu zápočet max 2 absence protokoly morfologický herbář do 31. 5. 2013! http://www.botanickafotogalerie.cz/ zásady bezpečné práce