Molekulární biologie člověka :
|
|
|
- Lenka Vávrová
- před 6 lety
- Počet zobrazení:
Transkript
1 Molekulární biologie člověka : Mutace a nestability v lidském genomu Dynamické mutace, repetitivní DNA Multigenové rodiny Marie Vojtíšková OGMB, MU Brno 2007
2 Lidský genom kompletní sekvence 3miliard párů bází genů jaderné DNA 37 genů mitochondriální DNA (10 3 kopií) poznání sekvence LG geny obsahují cca 3% celkové DNA (TESIMESENAVASISPOLUPRACI) Aplikace studia lidského genomu (LG)
3 % kompletní sekvence LG vzhledem k jednotlivým chromosomům Science 296, 2002, 1600

4 DNA uchování genetické informace Posloupnost čtyř bazí A, G, T, C
5 Reasociační kinetiky lidské DNA prokázaly tři skupiny pakujících se sekvencí: 60 % velmi nízký počet kopií 30% středně repetitivní 10% vysoce repetitivní Metody: DNA sekvencování DNA hybridizace PCR
6
7 non B struktury DNA Závislost na sekvenci repetice

8 Struktura tří vláken DNA Intermolekulární triplex Vazba třetího vlákna do velkého žlábku stabilní triplex Homopurin. hompyrimidinová sekvence Uplatnění v humánní medicině? Možnost aktivace nebo potlačení exprese genů LG :
9 Mutace LG- změny v genotypu Četnost spontánních mutací nízká, pravděpodobnost 10-7 Normální karyotyp člověka 46 chromosomů (XX/XY)/buňka (gamety 23 chromosomů) Mutace chromosomové (duplikace, delece, inverse, translokace, fragmentace, isochromosom, ring chromosom) Mutace genomové,polyplodie Mutace v genech, informace v genech, mutace na úrovni nukleotidů (inzerce, delece, substituce) dynamické mutace v závislosti na sekvenci buňky zárodečná linie - vrozené mutace maligní linie buněk získané a neopravené mutace, vedoucí k neoplastické transformaci mrna reflekce exprese genů fyziol./patol. protein / funkce - fyziol. /patol.
10 Poškození DNA na úrovni somatických buněk: Intracelulární -inkorporace chybného nukleotidu při replikaci -náhodné chemické změny DNA v buňce -reaktivní produkty metabolismu, kyslíkové radikály - viry- inkorporace do genomu infikové buňky - mobilní elementy (transpozony a retrotranspozony Extracelulární - chemické látky, mutagenní vlivy vnějšího prostředí - ionizační záření, UV záření Cílené - terapeutické působení cytostatik
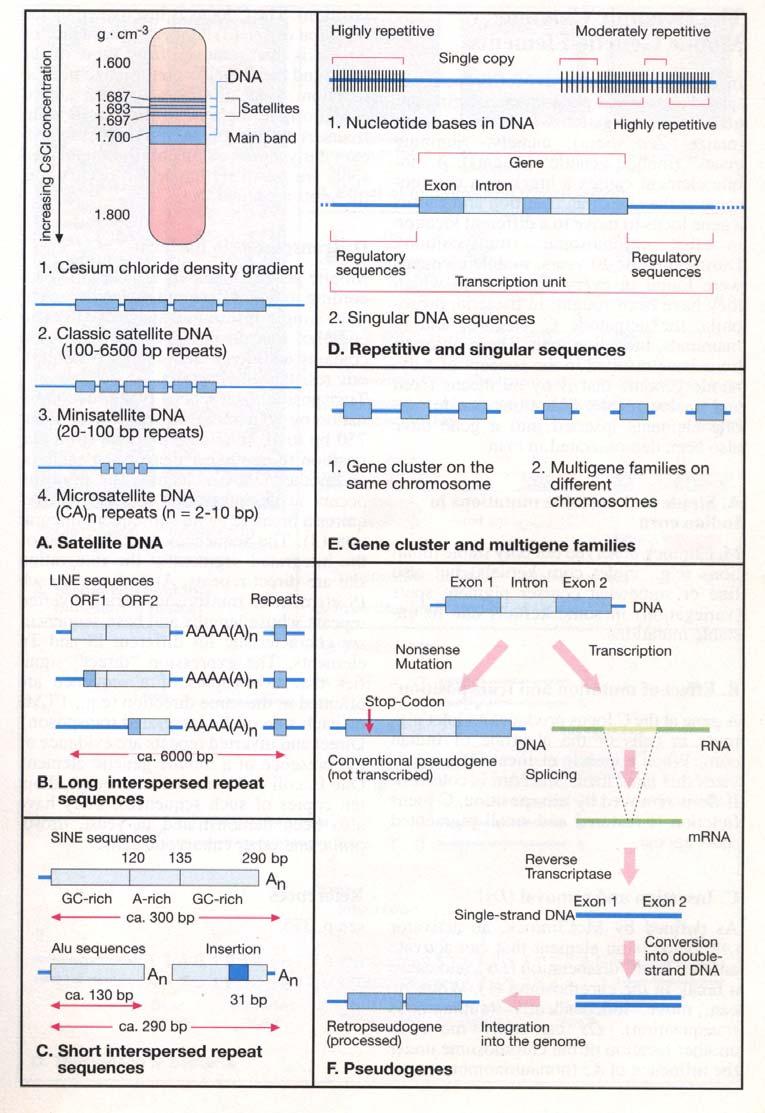
11 Repetitivní DNA DNA eukaryot a také člověka obsahuje značný podíl nekódujících sekvencí. Tak jako kódující DNA i nekódující může být unikátní anebo se může nacházet v genomu ve více identických nebo podobných kopiích. Sekvence DNA s vysokým množstvím kopií se nazývají repetitivní sekvence. Pokud jsou kopie sekvenčního motivu v blocích, v řadě za sebou, hovoříme o tandemových repeticích, od nich odlišujeme repetitivní sekvence rozptýlené v genomu jako jednotlivé kopie (rozptýlené repetice - anglicky interspersed repeats). Podstata rozptýlených repetic - transpozony Většina rozptýlených repetic vzniká procesem transpozice, což je "skákání" segmentu DNA na jiné místo genomu. Rozlišujeme v podstatě dva typy transpozibilních elementů DNA, neboli transpozonů: DNA transpozony a retrotranspozony. Hlavní skupiny rozptýlených repetic se schopností transpozice : LINE (long interspersed repeats), př. L1; LTR-retrotranspozon; 1,2kb -gen pro transpozásu je obklopen ITR, duplikace cílového místa; SINE ( short interspread repeat) př. 282 bp dlouhý Alu element
12 DNA transpozony DNA transpozony jsou v lidském genomu považovány za inaktivní, díky akumulaci mutací v průběhu fylogeneze obratlovců, a tak můžeme najít pouze jejich evolučně staré zbytky, neboli "fosilie". Nicméně aktivní transpozon odvozený z lidských fosilních elementů může být "vyroben" s použitím informací získaných z lidského genomu i genomu ostatních obratlovců. Jedním z příkladů je transpozon "Sleeping Beauty" (Šípková Růženka), který by se mohl např. stát základem další generace genové terapie, díky více specifickému místu integrace, než je tomu např. u retrovirů. Jak funguje typický DNA transpozon? Jádrem je sekvence kódující enzym transpozázu. Tento enzym se váže k oběma koncům repetitivního elementu, které jsou tvořeny invertovanými repeticemi. Tyto invertované konce si tedy mohou "vyměnit" řetězce a stabilizovat tak strukturu stopka-klička, nezbytnou pro aktivitu transpozázy. Transpozáza pak vyštěpí transpozon a liguje takto vzniklé volné konce chromozomální DNA. [Téměř shodný mechanismus je činný během maturace genů pro imunoglobuliny (V-D-J rekombinace) a TCR (T-cell receptor, receptor T-lymfocytu) při vyštěpení mezilehlých sekvencí. Je zajímavé, že enzym katalyzující tuto reakci (skládá se z dvou podjednotek RAG1 a RAG2) se skutečně pravděpodobně vyvinul z transpozázy.]. Uvolněný komplex transpozon-transpozáza se váže na specifický sekvenční motiv jinde v genomu, transpozáza štěpí hostitelskou DNA a liguje transpozon na nové místo. Takto se transpozon pohybuje mechanismem vyjmout-vložit (cut and paste) a počet kopií zůstává stabilní
13 Retrotranspozony Retrotranspozony jsou v lidském genomu mnohem důležitějšími transpozibilními elementy. Zaprvé jsou daleko hojnější, přímo tvoří nejméně 45% lidského genomu (odhady se různí, ale mnoho výzkumníků věří, že by to mělo být více, neboť starobylé retrotranspozony které byly inaktivovány, divergovaly díky mutacím tak, že jsou již nerozeznatelné). Zadruhé jsou retrotranspozony v lidském genomu stále aktivní. Pro "skákání" vyžadují buněčné RNA polymerázy (II nebo III), kterými jsou přepsány do RNA, zatímco původní kopie zůstává na svém místě. RNA kopie podléhá reverzní transkripci do DNA, která je vložena do genomu na nové místo. Tyto elementy tedy expandují (co do množství) mechanismem duplikace (kopírovat-vložit, copy and paste). Jak je dále popsánu pro L1 retrotranspozon, proces retrotranspozice je náchylný k různorodým chybám, a tak jsou nově vzniklé kopie většinou inaktivovány delecemi nebo bodovými mutacemi. Protože je většina kopií inaktivní, další expanze dané rodiny retrotranspozonů je řízena několika aktivními úplnými elementy. Avšak i když by později během fylogeneze došlo ke ztrátě všech aktivních elementů, genom může být doslova přeplněn fosilními členy dané rodiny sekvencí. Retrotranspozony mohou být dále klasifikovány jako autonomní nebo neautonomní. Autonomní retrotranspozony kódují proteiny nezbytné k jejich transpozici, ačkoli pro úspěšné "skákání" jsou také závislé na hostitelových RNA polymerázách a enzymech opravujících DNA. Neautonomní retrotranspozony nekódují proteiny a musí tak zneužít enzymy jiného transpozonu aby byly schopné transpozice.
14 LTR retrotranspozony - endogenní retroviry Endogenní retroviry, také nazývané LTR retrotranspozony, připomínají svým složením proviry skutečných retrovirů - obsahují LTR (long terminal repeats, dlouhé terminální repetice) a geny gag, pol, env a prt, ale alespoň jeden z genů nezbytných pro sestavení infekčních virových částic je mutován nebo chybí, zvláště se to týká genu env. Proto semohou endogenní retroviry pohybovat pouze uvnitř buněk, jinak je jejich životní cyklus podobný infekčním retrovirům, jako je HIV. Ačkoli jsou endogenní retroviry aktivní u mnoha savců, včetně šimpanze, lidský genom v současné době obsahuje pouze fosilie endogenních retrovirů (mutované a neschopné transpozice), které zaplňují asi 8% genomu. Intaktní endogenní retroviry jsou dlouhé 7-9 kb, ale stejně jako u L1 retrotranspozonu (viz dále) mnoho z nich je zkrácených, zejména na 5 konci. Často také můžeme najít pouze samostatné LTR, jako výsledek integrace retroviru a následné intrachromozomální rekombinace mezi oběma LTR nebo nerovnoměrné rekombinace dvou homologních chromozomů vedoucí k deleci kódující části retroviru.
15 Non-LTR retrotranspozony LINE LINE (long interspersed nuclear elements = dlouhé rozptýlené jaderné elementy) jsou autonomní retrotranspozony. Tvoří asi 21% lidského genomu. Aktivní elementy patří k nejhojnější rodině LINE-1 neboli L1, která sama o sobě zahrnuje 17% genomu. Ze zhruba půl milionu kopií L1 v našem genomu, skoro má úplnou velikost a asi 100 je stále schopno retrotranspozice. Aktivní L1 element je dlouhý asi 6 kb a obsahuje dva otevřené čtecí rámce (open reading frames), ORF1 a ORF2. 5 UTR (untranslated region, nepřekládaná oblast) funguje také jako promotor, 3 UTR obsahuje signál k polyadenylaci. Funkce ORF1 není jasná, známo je jen, že se váže na L1 mrna. ORF2 obsahuje doménu s aktivitou reverzní transkriptázy a endonukleázovou doménu a je enzymem zodpovědným za integraci. Životní cyklus L1 začíná transkripcí L1 DNA buněčnou RNA polymerázou II a standardní maturací v mrna molekulu. L1 mrna je transportována do cytoplazmy, kde je syntetizován protein ORF1. Pak je translace reiniciována na "vnitřním místě pro vstup ribozomu" (internal ribosomal entry site, IRES) (proces to nekanonický a tím neefektivní u eukaryot, a tak jen část L1 mrna molekul získá svůj protein ORF2). Oba proteiny se po své translaci neprodleně váží na L1 mrna. Tento komplex protein-mrna je transportován do jádra. ORF2 štěpí chromosomální DNA v cílovém místě (cílové místo není úplně specifické jak je tomu např. v případě restrikčních endonukleáz, ale je zde určitá preference pro sekvence bohaté A a T, místo štěpení je přibližně TT/AAAA). Štěpení DNA je nerovnoměrné (vytváří se kohezní konce). Volná 3 OH skupina na jedné straně štěpené DNA molekuly je užita reverzní transkriptázou proteinu ORF2 k zahájení syntézy prvního řetězce cdna (target primed reverse transcription, reverzní transkripce s cílovou sekvencí jako primerem).
16 Detailní mechanismus syntézy druhého řetězce cdna je stále předmětem diskuse, proces však končí stabilní integrací dvouvláknové L1 DNA na novém místě genomu. Díky stupňovitému zlomu cílové DNA vyrobenému endonukleázou transpozonu je integrovaný L1 element obklopen duplikací cílového místa o velikosti 7-20 párů bazí. Reverzní transkriptáza je většinou neschopna ukončit syntézu prvního řetězce, což vede ke zkrácení nové kopie na 5 konci. Reverzní transkriptáza také nemá 3-5 exonukleázovou aktivitu a tak často zavádí do nové kopie bodové mutace. Je zajímavé, že L1 mrna je exprimována zejména v meiotických a postmeiotických spermatocytech, zvyšujíc tak potentiál L1 pro expanzi (kopie introdukované to zárodečné linie jsou na rozdíl od nových somatických integrací dědičné).
17 Neautonomní retrotranspozony SINE SINE (short interspersed nuclear elements = krátké rozptýlené jaderné elementy) jsou typicky kratší než 500 bp a nemají žádný kódující potenciál. Hlavní rodinou SINE u člověka jsou Alu elementy (jméno je odvozeno od jejich objevu spojenému s párem konzervovaných restrikčních míst pro endonukleázu AluI). Více než 1 milión Alu elementů tvoří asi 11% lidského genomu. Alu elementy sdílí konsenzus 282 bp který je příbuzný a byl patrně odvozen z RNA podjednotky SRP (zvané 7SL RNA). SRP (signal recognition particle = částice rozpoznávající signál) je ribonukleoproteinový komplex, který rozpoznává signální peptid, váže se na něj a přemístí komplex ribozom-mrna-nascentní peptid ke kanálu endoplazmatického retikula (ER), skrz nějž je nascetní peptid translokován do lumen ER nebo integrován v membráně ER. Alu jsou, stejně jako gen pro 7SL RNA transkribovány RNA polymerázou III. Alu RNA váže dva proteiny SRP (9 a 14). Pravděpodobně se tak může Alu vázat na ribozom a díky svému "ocasu" bohatému na adenin také (pokud ribozom zrovna zpracovává LINE-1 mrna) na nascentní protein ORF2 a zneužít ORF2 k reverzní trnaskripci a integraci vlastní RNA a nikoli LINE-1.
18 Funkce transpozonů Z bezprostředního pohledu nemají transpozony žádnou důležitou funkci v buňce - hovoříse o"starém harampádí" - odpadní DNA (junk DNA); nebo o sobecké DNA, neboť se transpozony propagují na úkor buněčných energetických zdrojů. Z širšího úhlu pohledu může být mobilita retrotranspozonů důležitá pro plasticitu genomu. Příležitostná inzerce do genu může vyřadit gen z funkce a způsobit dědičné onemocnění. LTR a LINE elementy mohou také měnit genovou expresi, pokud se inzerují do blízkosti nějakého genu, neboť LTR a LINE 5 UTR mají silnou promotorovou aktivitu v obou směrech. Protože má LINE-1 retrotranspozon relativně slabý polyadenylační signál, stává se, že RNA polymeráza II se skrz něj pročte, a tak připojí k L1 mrna i následující sekvenci, která podlehne reverzní transkripci a přesunu na nové místo. Tak může být LINE-1 vektorem pro mobilitu samostatně nemobilních sekvencí. Navíc jsou retrotransponované kopie L1 často zkrácené na 5 konci, a tak se mobilizovaná DNA (která je na 3 konci) může dostat na nové místo i beze zbytků L1 sekvence. To může mít význam hlavně pro mobilitu menších DNA fragmentů -např. k výměně exonů mezi geny.
19 Retrotransposice L1 může dokonce vyústit v delece a inverze. Zřídka je normální buněčná mrna předmětem reverzní transkripce a transpozice enzymem z L1 nebo z jiného retrotranspozonu. V tomto případě dochází k duplikaci genu. Nová kopie se nazývá "procesovaný pseudogen" (processed pseudogene), neboť je odvozena ze zralé "zpracované" mrna bez intronů, a je obvykle nefunkční, díky chybějícímu promotoru. Zřídka však může procesovaný pseudogen přijmout novou funkci pod selekčním tlakem. Velmi známý příklad je gen pro podjednotku E1alfa pyruvátdehydrogenázy. Tento gen (PDHA1) leží u placentálních savců na chromozomu X. Ale exprese mnoha genů na chromozomu X je v průběhu spermatogeneze zastavena, včetně PDHA1, ačkoli jeho produkt je nezbytný pro funkci všech buněk. Tato chybějící funkce byla očividně zachráněna retrotranspozicí: na chromozomu 4 se nachází velmi podobný gen PDHA2, ale tento gen postrádá introny - a to je pro procesované pseudogeny typické. Vysoce exprimované "provozní" (housekeeping) geny mají samozřejmě větší pravděpodobnost retrotranspozice. Nacházíme tak mnoho procesovaných pseudogenů pro ribozomální proteiny, glykolytické enzymy, beta-aktin, a podobně. Procesované pseudogeny by neměly být zaměňovány za druhou kategorii "obyčejných" pseudogenů, které vznikají duplikací genomické DNA (např.pseudogeny ve skupině genů pro hemoglobin) a zachovávají proto původní strukturu (exony, introny, promotor...i když s porušenou funkcí
20 Bylo objeveno několik genů přímo odvozených z retrotranspozonů. Poslední přídavek je gen Peg10 (paternally expressed 10, paternálně exprimovaný gen 10), odvozený z LTR retrotranspozonu z rodiny Ty3/gypsy (velmi podobný retrotranspozon byl nalezen v aktivní formě u ryby fugu {Takifugu rubripes}). Peg10 je nezbytný pro vývoj placenty u myší a stejnou funkci bude mít pravděpodobně u člověka. Jiné příklady zahrnují geny pro syncytin, odvozené z endogenních retrovirů z rodiny HERV-W. Produkty těchto genů jsou důležité pro vytvoření syncytia z buněk trofoblastu, mechanismus fůze membrán připomíná vstup retroviru do buňky. I neaktivní repetitivní elementy zvětšují plasticitu genomu tím, že podporují mezichromozomový nerovnoměrný crosing-over nebo intrachromozomovou rekombinaci V neposlední řadě se uvažuje o tom, že by transpozony mohly mít nějakou reálnou fyziologickou funkci, např. proto, že jejich exprese je obecně zvýšena během stresové odpovědi. Ale různé hypotézy, které mohou být koncipovány na základě takových pozorování jsou v současné době nepotvrzené.
21 Tandemové repetice Tandemové repetice jsou tvořeny za sebou jdoucími identickými a nebo téměř identickými jednotkami. Tolik se však různí v délce jednotky repetice i celé repetice, že je jakákoli klasifikace neuspokojivá, a je nutno ji brát "cum grano salis". Největší repetice, které mají dendenci být složeny z relativně dlouhých jednotek se nazývají satelity. Jméno satelity je pochází z centrifugace DNA v hustotních gradientech. Nejprve, během konvenční izolace DNA, je tato předmětem namáhání smykem (shear stress), s výslednou fragmentací DNA (in vivo obsahuje jeden chromosom v G1 fázi 1 molekulu DNA). Tyto fragmenty mohou být centrifugovány v hustotních gradientech tak, že molekuly DNA obsazují v gradientu místa se stejnou hustotou prostředí jako má molekula DNA. Většina DNA vytvoří jednotný "proužek". Ale fragmenty DNA se signifikantně odlišným obsahem CG/AT, způsobeným např. rozsáhlými monotónními repeticemi vytvoří méně intenzivní přídavné "satelitní" proužky. Označení satelitní DNA bylo později rozšířeno a zahrnuje i podobně repetitivní sekvence, které však nevytváří tyto satelitní proužky. Primární jednotky repetice u satelitů jsou různorodé, od GGAAT u satelitu 2 a 3 až po 171 bp u alfa satelitu. Ale tyto primární jednotky jsou často degenerované, s určitými nepravidelnostmi. Tyto nepravidelnosti se mohou periodicky opakovat a tak tvořit sekundární jednotky. Satelitní DNA je hojná v oblasti centromer a konstitutivního heterochromatinu. Přestože je lidský genom považován za úplně sestavený, oblasti centromer a heterochromatin obsahující satelitní sekvence nejsou zahrnuty, neboť sekvenování takových oblastí je z různých důvodů problematické (absence patřičných restrikčních míst, obtížné sekvenování, téměř nemožné sestavení jednotlivých sekvencí do tzv. kontigu apod.). Z mnoha satelitů nacházených v oblasti centromer, tvoří rodina alfa satelitu (s primární jednotkou dlouhou 171 bp) pravděpodobně funkční jádro centromery, neboť je důležitá pro "poskládání" kinetochory během buněčného dělení (některé proteiny kinetochory se váží na alfa satelit v centromeře a tím zahajují sestavování kinetochory). Funkce ostatních satelitů je neznámá, jsou považovány obvykle za odpadní (junk) DNA.
22 Minisatelity jsou kratší tandemové repetice, v rozsahu kilobazí, které se více vyskytují v subtelomerických oblastech chromozomů. Jsou obvykle vysoce polymorfní co do počtu opakování jednotky repetice (mnoho alel v populaci) a mohou být použity jako genetické markery - VNTR (variable number of tandem repeats = variabilní množství tandemových repetic). VNTR jsou často příliš dlouhé pro amplifikaci pomocí PCR a jsou tudíž typicky stanovovány pomocí Southernova blotu (a jejich obliba tudíž klesá). Někdy se uvažuje o tom, že by některé minisatelity mohly mít regulační funkce, jako např. VNTR v promotoru inzulinového genu, kde byla různá délka VNTR asociována s různými typy diabetu. Telomery lidských chromozomů, Tvořené několika kilobazemi hexamerové repetice TTAGGG patří rozsahem také k minisatelitům, i když vznikají specifickým mechanismem - pomocí enzymu telomerázy. Telomeráza je složena z bílkovinné podjednotky a z RNA podjednotky obsahující sekvenci komplementární k TTAGGG, která slouží jako templát pro elongaci telomery (bílkovinná podjednotka je příbuzná reverzním trankriptázám non-ltr retrotranspozonů). Nicméně se mohou telomery elongovat i pasivně, mechanismem nerovnoměrného crossing-overu, např. v nádorových buňkách. (Výzkum struktury telomer - tetraplexy) Možná by stálo za to na tomto místě znovu připomenout, že sekvence lidsého genomu zahrnuje euchromatické úseky, ohraničené proximálně (ale nezahrnující) centromerou a pericentromerickým heterochromatinem a distálně telomery, které také, spolu se subtelomerickými oblastmi, nejsou obsaženy
23 Microsatelity jsou zpravidla tvořeny opakováním 1-5 bp, s množstvím opakování zřídka překračujícím stovky. Nejčastější jsou dinukleotidové repetice, ze kterých převažuje typ (CA)n. Mikrosatelity jsou v genomu velice časté, vysoce polymorfní a jsou často používány jako genetické markery (příklady mikrosatelitů jako genetických markerů jsou pro genetickém mapování)
24
25 Nemoci způsobené expanzí trinukleotidových repetic Pokud jsou uvnitř nebo v blízkosti genů, mohou mít mikrosatelity, resp. jejich různá délka, závažné důsledky, např. v heterogenní skupině monogenních nemocí podmíněných expanzí trinukleotidových repetic. Nejznámějším příkladem je Huntingtonova chorea, fatální neurologické onemocnění s nástupem v dospělosti, projevující se jako demence s extrapyramidovou poruchou motoriky. V genu pro huntingtin je repetitivní sekvence (CAG)n, která kóduje úsek bílkoviny tvořený zbytky glutaminu (polyglutaminový úsek, polyglutamine tract). Za normálních okolností mají lidé méně než 20 trinukleotidů CAG a tedy i glutaminů v huntingtinu, kde tyto tvořídůležitou doménu pro interakce s jinými proteiny. Pokud se však mutací toto množství zvětší nad 30 glutaminů, protein nepracuje správně (jak přesně je předmětem rozsáhlého výzkumu ) s výsledným progresivním odumíráním neuronů v nucleus caudatus. U jiného onemocnění, myotonické dytrofie (svalová dystrofie se svalovou slabostí provázenou paradoxně zvýšeným svalovým tonem) se nachází patologická expanze trinukleotidu CTG v 3 nepřekládané oblasti genu DMPK (dystrophia myotonica protein kinase). Mutantní mrna má sama o sobě patogenní potenciál, škodí pravděpodobně sekvestrací různých trankripčních faktorů.
26 Nový mutační mechanismus - expanze trinukleotidů - Genomová nestabilita spojená s opakováním trinukleotidů, (tetra -, penta.. dodekanukleotidů) Počet dosud známých chorob spojený s expanzí cca 15 Rozdílná fyziologická a patologická hladina počtu opakování
27 Společný prvek : na příslušném místě příslušného lokusu existuje tandemová repetice tripletu ( tetra.. penta ---) bází, přičemž zvýšení aktuálního počtu repeticí vede ke vzniku choroby patologie spojené s expanzí trinukleotidů: a) dědičné b) genetická nestabilita v somatických nádorových buňkách ( poruchy reparačních mechanismů - NER, reparace chybného párování c) genetická nestabilita v somatických buňkách v závislosti na věku
28 Charakteristický rys dynamických mutací - prodlužování nestabilní repetice během mezigeneračních transmisí Exprese patologie dynamických mutací pouze u člověka
29 Unstable CTG repeats in the DM family I. I /EXP /100 13/870 Family pedigree. Numbers of CTG repeats on both alleles are shown. EXP denoted long range pathological CTG repeats determined by TP- PCR. Individuals III/1, I/3,4 are with DM phenotype. Mother II/2 is healthy. Arrows represented investigated member of the family.
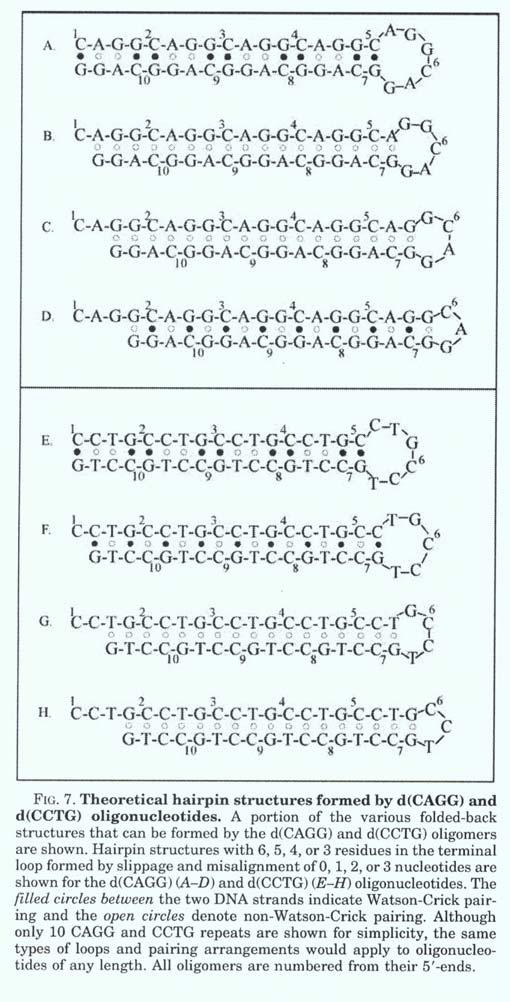
30 Molekulární příčiny genetické nestability dynamických mutací: Trinukleotidové repetice - specifická podskupina STRmikrosatelitů, vysoká frekvence (1/ kb) v lidském genomu Sekvenční homogennost, symetrie repetitivních úseků - tandemový motiv [NNN] n, n = počet opakování tvorba non B struktur dvoušroubovice DNA: např. vlásenky ( hairpins), lokální posun ( klouzání) řetězců Hoogsteenovo párování - triplexy, tetraplexy
31 Mutace způsobené expanzí trinukleotidových repeticí Expanze trinukleotidových repeticí představuje nový mutační mechanismus, jehož kauzální role v oblasti výlučně lidských chorob se dotýká stále se rozšiřujícího počtu onemocnění, jejichž společným znakem je primární zasažení nervové tkáně. Podle typu sekvence trinukleotidů na patologické alele mohou být choroby rozděleny na skupiny s expanzí v kodónu (CAG) n pro glutamin, choroby polyglutaminového traktu a na skupinu chorob s velkými expanzemi v nekódujících oblastech genů ( SyFraX, Myotonická dystrofie, Friedreichova ataxie). Původ mutací expandujících trinukleotidů dosud není spolehlivě vysvětlen. Bylo prokázáno, že nepřerušovaná sekvence CAG repetic je více náchylná k expanzi, než sekvence se vsuvkou CAT.
32
33 Model chybného párování sklouznutím řetězce (SSM) během DNA replikace v repetitivních sekvencích může způsobit zejména v dlouhých sekvencích inzerci až expanzi alely vedoucí k patologii nebo deleci v dceřiném řetězci v závislosti na tom, na kterém řetězci se bublina chybného párování nachází. Je-li bublina na dceřiném řetězci, tak dojde k inzerci, jeli na rodičovském, tak dochází k deleci. Normální alely vykazují určitou variabilitu v počtu opakování repetic ve fyziologickém rozhraní.
34
35 Fyziologické zastoupení alel (CTG) n v genu pro myotonickou dystrofii (19q13.3)
36
37 FraX (CGG)n aug FMR1 gene transcription Absence of FMR 1 protein Disruption of RNA processing in brain and testes
38 DM (CUG)n 5 AUG stop 3 Altered expression of RNA CUG-binding proteins Altered processing of CUG containing mrnas in skeletal muscle, heart, brain, and testes
39 Metody pro detekci expandujících tripletů (CTG) n Algoritmus DNA diagnostiky 1. krok: PCR (P1,P2) alely pro n< krok:tp PCR (P1, P3,P4CTG/P2,P3,P4CAG pro n>100, bez omezení 3. krok: XL PCR (+ 7-deaza-dGTP) pro n>100 n= ( ) Časový faktor: l den 4. krok: Southern blott/p5b1.4 EcoRI/BglI pro n > Časový faktor: l týden
40 TP PCR PCR metoda pro detekci dlouhých opakujících se CAG (CTG) sekvencí metoda využívá současně jak specifický značkující primer pro CAG (CTG) repetici tak dvojici primerů syntetizujících z několikanásobných míst v mezích rozsahu opakování Výsledkem TP syntézy je žebříček různě dlouhých fragmentů, lišících se o jeden trinukleotid Separace a detekce : Genomový analyzátor na principu kapilární elektroforézy a detekce LIF, PAGE a barvení AgNO 3
41 PCR P1,P2 (fragment 75pb pro n=5) Alela cca 100 CTG Alela > cca 100 CTG P1 CTG P2 P1 P2 PCR-ELFO Alela 1 Alela 2 P1 P1 PCR-ELFO P2 Alela 1 non PCR P2 Alela 2 Alela 2 Alela 1 Alela 1
42 Princip TP PCR Počáteční hybridizace P4 CTG P3 není homologní k LG
43
44
45 Zastoupení alel v souboru vyšetřených jedinců normál MD CTG n>100
46 Myotonická dystrofie typu 2 DM2 (3q21), expanze (CCTG) n v intronu 1 ( ZNF9) genu protein zinkového prstu Normální alela počet opakování tetranukleotidu do n = 30 Patologická alela - expanze n = ( ) CCTG

47
48
49

50
51 Replikace v tetranukleotidových opakujících se sekvencích (CCTG) : disociace řetězců - následná syntéza opoždˇujícího řetězce pomocí Ókazakiho fragmentů. Sekundární pseudostruktury - hlavní příčína genetické nestability TR vznik expanzí nebo delecí
52 Příčiny nestability dynamických mutací v několikanásobně se opakujících sekvencích tri, tetra, penta. nukleotidů : replikace, rekombinace, reparace Klouzání ( slippage) v repeticích vytváří non B- DNA struktury
53 Multigenové rodiny: Skupina sekvenčně příbuzných genů, které mají společný evoluční původ a stejnou biologickou funkci př. Geny kodující polypeptidové řetězce hemoglobinu Pseudogeny: nepřesné kopie strukturních genů, které vznikly asi nepřesnou duplikací nebo mutací, kterou se původně aktivní gen v genové rodině inaktivoval Tandemové genové repetice, jsou geny nebo skupiny genů opakující se bezprostředně za sebou, tedy v tandemu Geny uvnitř skupiny mají vždy stejné pořadí
54 Tandemovým způsobem se opakují: Geny přepisované do 5S rrna (četnost repetice u člověka 250) Geny přepisované do trna (četnost repetice 10 až 100 pro jednotlivý druh trna u člověka Geny ko dující histony (četnost repetice 20 pro skupinu genů ko dujících všech 5 histonů Biologický význam těchto tandemových genových repeticí spočívá v tom, že jedna kopie každého z uvedených genů by nestačila krýt spotřebu jeho produktů buňkou. V buňce musí být dostatečná zásoba 5S rrna a trna pro realizaci translace a histonů pro syntézu histonového oktameru při replikaci.
55 Př. Multigenových rodin Hlavní histokompatibilní komplex (HLA Human Leucocyte Antigen ) na chromozomu 6 Biologickou funkcí HLA proteinů je prezentace antigenních peptidů T lymfocytům spuštění komplexní imunitní odpovědi proti patogenu. Obecná koncepce imunitní odpovědi připravenost na jakýkoliv typ infekce. HLA geny 1. třídy: HLA-A, HLA-B, HLA-C HLA geny 2. třídy: HLA-DR, HLA-DQ, HLA-DP Alelická bohatost HLA genů, heterozygotní jedinec schopnost čelit většímu počtu infekcí, heterozygotní výhodnost.
56 Struktura HLA genové rodiny, počet cca 20 genů ( lokus 6p21.3). velikost 2 Mb. Exprese 6 genů (viz výše), další 4 jsou konvenční pseudogeny a 7 representuje nefunkční geny nebo fragmenty genů
57 Genové rodiny kódující produkty s velkými konzervovanými doménami. Výskyt homologie v silně konzervovaných oblastech genů, často jsou kódovány proteiny s významnou funkcí ve vývoji. Konzervovaný sekvenční motiv obsahují geny PAX (390 pb) nebo homeoboxy (180 pb). Homeotické proteiny se váží na specifické sekvence DNA a regulují expresi genů na úrovni transkripce. Řada genů, které kódují transkripční faktory, obsahuje homeobox. HMG proteiny.
58 Genové rodiny kódující produkty s krátkými konzervovanými motivy aminokyselin jsou charakterizovány obecnou funkcí. Např. Genová rodina s DEAD boxem obsahuje několik rozdílných genů s RNA helikázovou funkcí s krátkým charakteristickým motivem aminokyselin DEAD box = Asp-Glu-Ala-Asp. Dalším typem je WD repetitivní genová rodina ( tandemově se opakující tryptophan aspartate) s pravěpodobnou regulační funkcí.
59 Genové superrodiny Charakteristickým rysem je nízká sekvenční homologie velkých segmentů, ale v obecném smyslu funkční příbuznost a podobnost ve struktuře domén. Př. Členové Ig imunoglobulinové superrodiny jsou povrchové proteiny s podobným typem struktury domén (S-S disulfidové můstky)
60 Rozptýlené genové repetice, gen nebo genová rodina, jejichž kopie se vyskytují na různých místech haploidního genomu. Např. některé geny přepisované do trna
61 Nemendelovská dědičnost Některá onemocnění a znaky, za které zodpovídají variace jednotlivých genů, nesledují do značné míry pravidla přenosu do dalších generací, která platí pro dědičnost "klasických monogenních onemocnění, ať už autozomálně či gonozomálně determinovaných. Proto se tato různorodá skupina někdy vyděluje pod pojmem tzv. nemendelovské dědičnosti (angl.non-mendelian inheritance). Řadíme sem především mitochondriální dědičnost, nestabilitu repetitivních sekvencí a genomický imprinting. Zcela zvláštní kapitolou je epigenetika. Naopak se do této skupiny obvykle nepočítají jevy, které "komplikují" monogenní dědičnost, např. penetrance, expresivita znaku atd.

62

63
64 1. Mitochondrie a mitochondriální dědičnost a) Mitochondriální DNA Ačkoli je podstatná část lidského genomu uložena v jaderné DNA, 37 strukturních genů je kódováno v cirkulárním mitochondriálním genomu, který je tvořen páry bazí DNA (označované jako mtdna). Třetina těchto genů kóduje podjednotky komplexů respiračního řetězce zodpovědného za produkci ATP v rámci tzv. oxidačně-fosforylačního (OXPHOS) systému (13 polypeptidů, zbylých 74 je kódováno jaderným genomem), zbylé pak zodpovídají za tvorbu specifických molekul RNA potřebných pro syntézu proteinů (22 transferových RNA a dvě ribosomální RNA). V cytoplazmě je přítomno několik set až tisíc kopií mtdna, když v každé mitochondrii je přítomna řada molekul mtdna. O homoplazmii mluvíme, když je přítomna jen jedna homogenní populace mtdna (odpovídá stavu u drtivé většiny "normálních" jedinců). Frekvence mutací je u mitochondrií 10-20x vyšší než u jaderné DNA, což vede k heteroplazmii, tedy stavu, kdy je přítomna smíšená populace s více variantami mtdna. Mnohé vlastnosti mitochondriální DNA podporují teorii o původu mitochondrií coby endosymbiontických bakterií proto-eukaryotických buněk před cca 1,5 miliardami let. Mezi nejmarkantnější z nich určitě patří cirkulární organizace genomu, absence histonů, absence intronů či diskrétní počátky replikace. Navíc se mitochondrie obratlovců i dalších organismů se vyznačují odchylkami od univerzálního genetického kódu - kodón UGA není terminační, ale kóduje aminokyselinu tryptofan, naopak kodóny AGA a AGG nekódují leucin, ale ukončují translaci a nakonec kodón AUA místo leucinu kóduje metionin. Biologicky jsou asi nejbližšími příbuznými Rickettsie (mj. původci skvrnitého tyfu), což jsou obligátní intracelulární parazité. Předpokládá se, že Rickettsie a mitochondrie mají společného předka, který prodělal přechod od autonomní existence k endosymbióze. Mitochondriální DNA skutčně obsahuje jen jednu významnou "nekódující" sekvenci, takzvanou D-smyčku (z angl. D-loop), což je krátký úsek mtdna, ve kterém je těžký řetězec vytěsňován fragmentem DNA ( nukleotidů), komplementárním k řetězci lehkému (a v tomto úseku má tedy mitochondrie třířetězcovou DNA). Zde je počátek replikace tzv. těžkého řetězce (označovaného jako H z angl. heavy), zmiňovaný fragment funguje jako primer pro začátek replikace. Počátek replikace lehkého řetězce (L z angl. light) je umístěn mimo Dkličku, asi ve 2/3 mt DNA. Transkripty obou řetězců musí být rozštěpeny, aby se mohly uvolnit funkční RNA (rrna, trna a mrna).
65 b) Mitochondriální dědičnost Z hlediska genetiky je zásadní fakt, že mtdna je předávána další generaci výhradně matkou (matroklinní dědičnost), když po oplodnění jsou zachovány pouze mitochondrie lidského vajíčka. To patrně není pouhým důsledkem nepoměru počtu mitochondrií lidského oocytu (cca ) a spermie (50-70), ale předpokládá se aktivní proces, který po oplození zlikviduje mitochondrie paternálního původu. Tomu odpovídá i typický maternální přenos chorob způsobených mutacemi mtdna v rodokmenu (Obr.1.). Pokud je heteroplazmická mutace zděděna nebo k ní dojde v časných fázích embryogeneze, normální i mutovaná varianta jsou náhodně předávány při buněčném dělení dceřinným buňkám (mitotická i meiotická segregace). Distribuce a zastoupení mutované mtdna v jednotlivých orgánech jsou proto patrně závislé na čase a vzniku mutace a rovněž na typu postižené buňky.
66 c) Onemocnění s mitochondriální dědičností Je třeba si uvědomit, že zdaleka ne veškerá onemocnění způsobená dysfunkcí mitochondrií mají mitochondriální typ dědičnosti. Pokud je defektní protein, jehož některé podjednotky kódují geny jaderného genomu a jiné zase přímo mtdna, může být dědičnost zcela medndelistická. Čtyři základní charakteristiky dědičných onemocnění, u kterých je třeba diferenciálně diagnosticky uvažovat o mitochondriální dědičnosti: 1. Maternální typ dědičnosti (viz výše) 2. Díky mitotické a meiotické segregaci nacházíme různý stupeň postižení v různých tkáních a variabilní projevy u potomků v jedné mateřské linii, což souvisí s existencí tzv. prahového efektu, což je pozorování, že pro manifestaci dysfunkce je nutné, aby byl překročen určitý poměr mitochondrií s mutantní a normální mtdna (ve většině případů 60-90%). 3. Projevy poruchy OXPHOS systému. Dědičná mitochondriální onemocnění většinou postihují tkáně s vysokými nároky na přísun energie, jako jsou neuromuskulární či centrální nervový systém. Mezi běžně pozorované symptomy chorob s mitochondriální dědičností patří encefalopatie, ataxie, spasticita, (kardio)myopatie, hluchota nebo diabetes mellitus. 4. Rozdíl oproti gonozomálně dominantní dědičnosti vázané na chromozóm X je v absenci přenosu mitochondriálně vázaného onemocnění na potomstvo a signifikantní převahu postižených žen u X-vázané dominantní dědičnosti.
Genetické haraburdí - repetitivní DNA
 Genetické haraburdí - repetitivní DNA Repetitivní DNA DNA eukaryot a také člověka obsahuje značný podíl nekódujících sekvencí. Tak jako kódující DNA i nekódující může být unikátní anebo se může nacházet
Genetické haraburdí - repetitivní DNA Repetitivní DNA DNA eukaryot a také člověka obsahuje značný podíl nekódujících sekvencí. Tak jako kódující DNA i nekódující může být unikátní anebo se může nacházet
Nemendelovská dědičnost
 Nemendelovská dědičnost Některá onemocnění a znaky, za které zodpovídají variace jednotlivých genů, nesledují do značné míry pravidla přenosu do dalších generací, která platí pro dědičnost "klasických"
Nemendelovská dědičnost Některá onemocnění a znaky, za které zodpovídají variace jednotlivých genů, nesledují do značné míry pravidla přenosu do dalších generací, která platí pro dědičnost "klasických"
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Struktura a organizace genomů
 CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Základy molekulární a buněčné biologie. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin.
 Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin. Nukleové base - purinové a pyrimidinové Ribonukleosidy - base + ribosa Deoxyribonukleosidy base + 2 - deoxyribosa Nukleotidy,
Struktura, vlastnosti a funkce nukleových kyselin, DNA v jádře, chromatin. Nukleové base - purinové a pyrimidinové Ribonukleosidy - base + ribosa Deoxyribonukleosidy base + 2 - deoxyribosa Nukleotidy,
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
DMPK (ZNF9) V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M
 V DIFERENCOVANÝCH. Z, Kroupová I, Falk M* M") FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
FISH ANALÝZA m-rna DMPK (ZNF9) V DIFERENCOVANÝCH TKÁNÍCH PACIENT IENTŮ S MYOTONICKOU DYSTROFI FIÍ Lukáš Z, Kroupová I, Falk M* M Ústav patologie FN Brno *Biofyzikáln lní ústav AVČR R Brno Definice MD Myotonická
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
Genetika zvířat - MENDELU
 Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Genetika. Genetika. Nauka o dědid. dičnosti a proměnlivosti. molekulárn. rní buněk organismů populací
 Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE
 ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
Molekulární genetika II zimní semestr 4. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc.
 Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Molekulárn. rní genetika
 Molekulárn rní genetika Centráln lní dogma molekulárn rní biologie cesta přenosu genetické informace: DNA RNA proteiny výjimkou reverzní transkripce retrovirů: RNA DNA Chemie nukleových kyselin dusíkaté
Molekulárn rní genetika Centráln lní dogma molekulárn rní biologie cesta přenosu genetické informace: DNA RNA proteiny výjimkou reverzní transkripce retrovirů: RNA DNA Chemie nukleových kyselin dusíkaté
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie.
 Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie. Připravila L.Fajkusová Online Mendelian Inheritance in Man: #229300 FRIEDREICH ATAXIA 1; FRDA *606829 FRDA GENE; FRDA Popis onemocnění
Návrh směrnic pro správnou laboratorní diagnostiku Friedreichovy ataxie. Připravila L.Fajkusová Online Mendelian Inheritance in Man: #229300 FRIEDREICH ATAXIA 1; FRDA *606829 FRDA GENE; FRDA Popis onemocnění
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
B6, 2007/2008, I. Literák
 B6, 2007/2008, I. Literák REPLIKACE GENETICKÉ INFORMACE REPLIKACE GENETICKÉ INFORMACE život závisí na stabilním uchovávání a předávání genetické informace v buňce jsou mechanismy pro: přesné kopírování
B6, 2007/2008, I. Literák REPLIKACE GENETICKÉ INFORMACE REPLIKACE GENETICKÉ INFORMACE život závisí na stabilním uchovávání a předávání genetické informace v buňce jsou mechanismy pro: přesné kopírování
Syntéza a postranskripční úpravy RNA
 Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Molekulární základ dědičnosti
 Molekulární základ dědičnosti Dědičná informace je zakódována v deoxyribonukleové kyselině, která je uložena v jádře buňky v chromozómech. Zlomovým objevem pro další rozvoj molekulární genetiky bylo odhalení
Molekulární základ dědičnosti Dědičná informace je zakódována v deoxyribonukleové kyselině, která je uložena v jádře buňky v chromozómech. Zlomovým objevem pro další rozvoj molekulární genetiky bylo odhalení
TRANSLACE - SYNTÉZA BÍLKOVIN
 TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
MOBILNÍ GENETICKÉ ELEMENTY. Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 MOBILNÍ GENETICKÉ ELEMENTY Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. Demerec (1937) popsal nestabilní mutace u D. melanogaster B. McClintocková (1902-1992, Nobelova cena 1983) ukázala ve
MOBILNÍ GENETICKÉ ELEMENTY Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. Demerec (1937) popsal nestabilní mutace u D. melanogaster B. McClintocková (1902-1992, Nobelova cena 1983) ukázala ve
Souhrnný test - genetika
 Souhrnný test - genetika 1. Molekuly DNA a RNA se shodují v tom, že a) jsou nositelé genetické informace, b) jsou tvořeny dvěma polynukleotidovými řetězci,, c) jsou tvořeny řetězci vzájemně spojených nukleotidů,
Souhrnný test - genetika 1. Molekuly DNA a RNA se shodují v tom, že a) jsou nositelé genetické informace, b) jsou tvořeny dvěma polynukleotidovými řetězci,, c) jsou tvořeny řetězci vzájemně spojených nukleotidů,
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Translace (druhý krok genové exprese)
") Translace (druhý krok genové exprese) Od RN k proteinu Milada Roštejnská Helena Klímová 1 enetický kód trn minoacyl-trn-synthetasa Translace probíhá na ribosomech Iniciace translace Elongace translace
Translace (druhý krok genové exprese) Od RN k proteinu Milada Roštejnská Helena Klímová 1 enetický kód trn minoacyl-trn-synthetasa Translace probíhá na ribosomech Iniciace translace Elongace translace
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
GENETIKA. Dědičnost a pohlaví
 GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
The cell biology of rabies virus: using stealth to reach the brain
 The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
Genetický kód. Jakmile vznikne funkční mrna, informace v ní obsažená může být ihned použita pro syntézu proteinu.
 Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Proměnlivost organismu. Mgr. Aleš RUDA
 Proměnlivost organismu Mgr. Aleš RUDA Faktory variability organismů Vnitřní = faktory vedoucí k proměnlivosti genotypu Vnější = faktory prostředí Příčiny proměnlivosti děje probíhající při meioze segregace
Proměnlivost organismu Mgr. Aleš RUDA Faktory variability organismů Vnitřní = faktory vedoucí k proměnlivosti genotypu Vnější = faktory prostředí Příčiny proměnlivosti děje probíhající při meioze segregace
Mutace jako změna genetické informace a zdroj genetické variability
 Obecná genetika Mutace jako změna genetické informace a zdroj genetické variability Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt
Obecná genetika Mutace jako změna genetické informace a zdroj genetické variability Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt
Buňky, tkáně, orgány, soustavy
 Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
b) Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?
 Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?") 1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
Molekulární genetika: Základní stavební jednotkou nukleových kyselin jsou nukleotidy, které jsou tvořeny
 Otázka: Molekulární genetika, genetika buněk Předmět: Biologie Přidal(a): jeti52 Molekulární genetika: Do roku 1953 nebylo přesně známa podstata genetické informace, genů, dědičnosti,.. V roce 1953 Watson
Otázka: Molekulární genetika, genetika buněk Předmět: Biologie Přidal(a): jeti52 Molekulární genetika: Do roku 1953 nebylo přesně známa podstata genetické informace, genů, dědičnosti,.. V roce 1953 Watson
Huntingtonova choroba
 Huntingtonova choroba Renata Gaillyová OLG FN Brno Huntingtonova choroba je dědičné neurodegenerativní onemocnění mozku, které postihuje jedince obojího pohlaví příznaky se obvykle začínají objevovat mezi
Huntingtonova choroba Renata Gaillyová OLG FN Brno Huntingtonova choroba je dědičné neurodegenerativní onemocnění mozku, které postihuje jedince obojího pohlaví příznaky se obvykle začínají objevovat mezi
Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK
 ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
A. chromozómy jsou rozděleny na 2 chromatidy spojené jen v místě centromery. B. vlákna dělícího vřeténka jsou připojena k chromozómům
 Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Genetika - maturitní otázka z biologie (2)
") Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):
Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):
ší šířen VAZEBNÁ ANALÝZA Vazba genů
 VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
Systém HLA a prezentace antigenu. Ústav imunologie UK 2.LF a FN Motol
 Systém HLA a prezentace antigenu Ústav imunologie UK 2.LF a FN Motol Struktura a funkce HLA historie struktura HLA genů a molekul funkce HLA molekul nomenklatura HLA systému HLA asociace s nemocemi prezentace
Systém HLA a prezentace antigenu Ústav imunologie UK 2.LF a FN Motol Struktura a funkce HLA historie struktura HLA genů a molekul funkce HLA molekul nomenklatura HLA systému HLA asociace s nemocemi prezentace