hormony a) Auxiny b) Brasinosteroidy c) Ethylen d) Kyselina abscisová (ABA) e) Kyselina salicylová (SA) f) Kyselina jasmonová (JA)
|
|
|
- Aneta Černá
- před 6 lety
- Počet zobrazení:
Transkript
 Auxiny b) Brasinosteroidy c)")
 f) Kyselina")
1 2015 3) Regulace růstu a vývoje rostlin hormony 1 a) Auxiny b) Brasinosteroidy c) Ethylen d) Kyselina abscisová (ABA) e) Kyselina salicylová (SA) f) Kyselina jasmonová (JA)


2 2 Vývoj organismu regulují signály (faktory) Receptor R Vnitřní genetická info metabolismus hormony Vnější světlo teplota vlhkost plyny gravitace Signál Receptor Zesilovač Reakce (např. exprese genů)
 i parakrinní (účinky v místě syntézy) Nemají přesná místa syntézy a")
 a přenos signálu v buňce (systém druhých poslů, protein kinasová kaskáda, regulace aktivity")
3 3 Historii hormonů začíná Julius von Sachs ( ), studoval v Praze. Endokrinní (přenos do jiných částí rostliny) i parakrinní (účinky v místě syntézy) Nemají přesná místa syntézy a účinku Slabší koncentrační závislost účinku Menší specificita účinku mechanismus účinku ale obdobný Receptor (vazebné místo) a přenos signálu v buňce (systém druhých poslů, protein kinasová kaskáda, regulace aktivity enzymů, genové exprese apod.) PřF UK, Praha


4 a) Auxiny Fototropismus (1880) Ch. Darwin ~ 180 minut 4 Auxinový gradient Zakřivení koleoptile Auxin stimuluje růst buněk více na zastíněné než na ozářené straně koleoptile => zakřivení růstu

 Tento experiment ukazující stimulaci růstu auxinem byl použit jako základ pro")
5 5 Ve 30. letech byl izolován auxin a bylo ukázáno, že stimuluje růst.. Úhel zakřivení je úměrný množství auxinu v bloku Akumulace auxinu ze špiček koleoptile do agarových bloků...a ukázal, že materiál akumulovaný v agarových blocích je látka, která stimuluje růst. IAA - Kyselina indol-3-octová (Indole-3-acetic acid) Tento experiment ukazující stimulaci růstu auxinem byl použit jako základ pro purifikaci auxinu American Society of Plant Biologists Redrawn from Went, F.W. (1935) Auxin, the plant growth-hormone. Bot. Rev. 1:


6 Auxiny důležité rostlinné hormony zapojené v širokém spektru růstových a vývojových procesů: 6 embryogeneze IAA prodlužování stonku apikální dominance 4-Cl-IAA PAA foto- a gravitropismus tvorba laterálních kořenů Buněčná úroveň: buněčné dělení expanze buněk diferenciace buněk

7 7 Biosyntéza auxinu IAA Indolová dráha Tryptofanová dráha Hlavním místem biosyntézy jsou mladá dělící se pletiva a meristémy.
8 Konjugáty 8 Kovalentně vázaná IAA (hl. semena a zásobní orgány) netransportovatelné inaktivní formy auxinu (A) Reverzibilní IAA-myo-inositol, IAA-glukóza, IAA-AMK, methylester-iaa (B) Ireverzibilní IAA-aspartát, IAA-glutamát Degradace: oxidace IAA (substrát IAA i ireverzibilní konjugáty)
9 Transport auxinu 9 Polární (bazipetální) z apexu, koleoptile, řapíky; v kořeni převládá akropetální (není ovlivněn orientací) Buněčný orientovaný (převládá), méně apoplastický, xylémový i floémový transport Inhibitory transportu NPA: kys. 1-N-naftylftalamová TIBA: kys. 2,3,5-triiododbenzoová NOA: kys. 1-naftoxyoctová quercetin, genistein
 - Aktivní permeása - AUX1 přenašeč")
 WT")
10 Příjem (influx) Výdej (efflux) 10 - Pasivní (difúze IAAH, lipofilní) - Aktivní permeása - AUX1 přenašeč (symport 2H + /IAA - ) - Aktivní (přenašeče: PIN proteiny, P-glykoproteiny (ATP-závislý přenašeč) WT pin1 mutant
, váže na sebe H + a stává se elektricky neutrálním (IAAH). AUX1 2.")

. 5.")
11 11 Buněčné základy polárního transportu auxinu John Raven and Mary Helen Goldsmith 1. V cytoplazmě (ph 7) se auxin vyskytuje v ionizované formě. Když se v této formě dostane do prostředí buněčné stěny (ph 5), váže na sebe H + a stává se elektricky neutrálním (IAAH). AUX1 2. Jako relativně malá a neutrální molekula je auxin schopen procházet přes plazmatickou membránu dovnitř buňky. PIN1 3. Když se neutrální auxin dostane dovnitř buňky s ph 7, ionizuje se. V této formě se auxin po určitou dobu akumuluje v buňce, protože v ionizované formě má menší schopnost prostupovat plazmatickou membránou než stejně velké neutrální molekuly. 4. Uvolněný H + je transportován protonovou pumpou do apoplastu (prostor buněčné stěny). 5. Anionty auxinu se dostávájí z buňky pomocí membránového proteinového přenašeče. Protonová pumpa udržuje transport auxinu tím, že generuje membránový potenciál, který favorizuje transport aniontů auxinu ven z buňky


12 Inhibitory transportu blokují sekreci PIN1 proteinu 12 Akumulace PIN1 na plazmatické membráně Akumulace PIN1 v jádře NPA: kys. 1-N-naftylftalamová TIBA: kys. 2,3,5-triiododbenzoová NOA: kys. 1-naftoxyoctová quercetin, genistein
 u Arabidopsis Kepinski and Leyser 2005")
 První intracelulární auxinový")
13 13 Signální dráhy auxinů Auxinový receptor TIR1 Intracelulární receptor TIR1 (Transport Inhibitor Response 1) u Arabidopsis Kepinski and Leyser 2005 Dharmasiri et al Jádro Auxin se váže v jádře přímo k TIR1 1) První intracelulární auxinový receptor 2) Receptor zprostředkující transkripční reakce k auxinu TIR1 A TIR1 A
. To vede k degradaci substrátového proteinu a tak k uvolnění exprese auxinem-indukovaných genů.")
14 F-box zprostředkuje transkripční reakce k auxinu 14 TIR1 TIR1 A A TIR1 A Degradace substrátu Jádro 2010 American Society of Plant Biologists Vazba auxinu k TIR1 umožňuje vazbu substrátového proteinu ( = represoru transkripce auxinem-indukovaných genů). To vede k degradaci substrátového proteinu a tak k uvolnění exprese auxinem-indukovaných genů. Auxin funguje jako lepidlo umožňující spojení TIR1 se substrátem

15 Substrát je poly-ubiquitinován a směřován do 26S proteazomu, kde je degradován. 15 Ub Ubiquitin Jádro Ub UbUb E1 Ubiquitin-activující enzym 26S Proteazom E1 E2 F-box Ubiquitin-conjugující enzym F-box E3 ubiquitin-ligázy Cílový protein A F-box ASK1 Ub Ub CUL1 Ub E2 RBX1 E1 Ub Ub Ubiquitin Ubiquitinace - nezbytný regulační mechanizmus v rostlinných i živočišných buňkách
16 16 Signalizace prostřednictvím jaderného receptoru TIR1 Ub UbUb 26S Proteasome E1 A TIR1 auxin F-box podjednotka E3-ubiquitin ligázy; auxinový receptor A TIR1 ASK1 Ub Ub CUL1 Ub E2 RBX1 E1 Ub Ub ARF represor transkripce auxinem-indukovaných genů transkripční faktor (aktivátor exprese genů) Auxinem-indukované geny obsahují AuxRE = auxin responsivní element TIR1 A TIR1 A A TIR1 A [Auxin] A A A A A ARF AuxRE Auxin inducible gene de-repression ARF AuxRE e.g. AUX/IAA, SAUR, GH3, etc. Plant development
 Čtyřnásobný mutant stále funkční rostlina Receptory TIR1 a AFB nejsou nepostradatelné Col-0 tir1 afb1 afb2 afb3")

17 Signální dráha receptoru TIR1 důležitá, ale ne jediná, dráha pro funkci auxinu 17 Homology TIR1: AFB1, AFB2, AFB3 stejná funkce jako TIR1 Avšak: 1) Čtyřnásobný mutant stále funkční rostlina Receptory TIR1 a AFB nejsou nepostradatelné Col-0 tir1 afb1 afb2 afb3 2) Dharmasiri et al. 2005b Auxin indukuje prodlužování buněk s lag fází 8-15 min => rychlá reakce vylučující zapojení TIR1, tedy transkripci genů či syntézu proteinů Auxin funguje prostřednictvím signální dráhy s jiným receptorem Receptor ABP1 Steffens and Lüthen 2000
18 18 Auxinový receptor ABP identifikace ABP1 (Auxin-Binding Protein 1) v izolovaných membránách z buněk kukuřičné koleoptile; vazba radioaktivního auxinu v membráně 1985 izolace proteinu ABP1 z kukuřice; velikost 22 kd Konec 80. let klonování ABP1 a struktura ABP1 Auxin-binding doména Pravděpodobná auxin-binding doména N Box a Box b Box c KDEL C Auxin se váže k plazmamembránové frakci? ALE Protein ABP1 nemá transmembránovou doménu Navržen model Docking proteinu
19 Transport ABP1 z endoplazmatického retikula do apoplastu 19 Většina ABP1 je lokalizována v endoplazmatickém retikuku (ER). Malá část ABP1 je pomocí vezikul sekretovába zatím neznámým mechanizmem do apoplastu a přichycena na plazmatickou membránu (Plm). Sekrece ABP1 byla dokázána v různých typech buněčných kultur. ABP1 se pravidelně nalézá i v kultivačním médiu. Brefeldin - inhibitor vesikulárního transportu - inhibuje sekreci ABP1 do kultivačního media.

20 Teorie kyselého růstu, protonová pumpa a draslíkové kanály 20 Auxin Exkrece H + do apoplastu Snížení ph v apoplastu Rayle D and Cleland R (1970) Aktivace enzymů (expansinů) rozvolňujících buněčnou stěnu apoplast cytozol H H + + H + H + H + IAA ATP H + -ATPáza ADP? ABP1 Docking protein Plazmatická membrána Christian M et al. 2006, upraveno Vazba auxinu k ABP1 Stimulace aktivity H + -ATPázy Hyperpolarizace membrány Acidifikace apoplastu
21 Podmínkou růstu buňky je turgor. Auxin sám nezvyšuje turgorový tlak. 21 Akumulace H + v apoplastu Kompenzace náboje v cytosolu K + kanály transportující K + dovnitř buňky apoplast cytosol ATP H + K+ K + H + -ATPáza ADP H + H + IAA ABP1 Docking protein? H + H + Plazmatická membrána Christian M et al. 2006, upraveno K + kanál K + kanál K + K + K + Akvaporin H 2 O H 2 O Akumulace K + v cytosolu Transport H 2 0 do buňky Turgor RŮST Přítomnost K + : podmínka udržení trvalé acidifikace a trvalého růstu
22 22 b) Brasinosteroidy Rostliny 1 miliarda let Živočichové Hormony - polyhydroxylované steroidní molekuly BR a steroidy kontrolují: - regulaci genové exprese - buněčné dělení - expanzi buněk - diferenciaci buněk - PCD - homeostázu
 - inhibuje prodlužování kořene (vysoké koncentrace) - stimuluje klíčení semen a růst listů HO Brasinolid OH OH Prodlužování 2.")

23 brasinolid (BL) koncový produkt biosyntetické dráhy - stimuluje prodlužování stonku - stimuluje prodlužování kořene (nízké koncentrace) - inhibuje prodlužování kořene (vysoké koncentrace) - stimuluje klíčení semen a růst listů HO Brasinolid OH OH Prodlužování 2. internodu stonku fazole HO H O O Brasinosteroidy a auxiny mají synergické účinky. Stimulace: - tvorby laterálních kořenů - prodlužování stonku - růstu pylové láčky - ohýbaní listů a epinastie - aktivity protonové pumpy - diferenciace xylému Ohýbání listů Auxin rychlý účinek Brasinolid pomalý účinek
24 24 Použití brasinosteroidů v zemědělství: - zvyšování výnosů - stimulace rezistence ke stresům Výskyt: - ve všech testovaných pletivech - nejvíce v apikálních částech stonků Brasinosteroidy nejsou pravděpodobně transportovány na dlouhé vzdálenosti

25 25 Genetický přístup fyziologická, biochemická a molekulární charakterizace mutantů Selekce mutantů s redukovanou biosyntézou BR (rajče, hrách, rýže, Arabidopsis) Výrazný fenotyp mutantů; Arabidopsis pleiotropický efekt mutací Tma: krátký vzrůst, tlustý hypokotyl, otevřené zvětšené dělohy, přítomnost primárních listových pupenů bri1 WT cpd WT Světlo: trpasličí vzrůst, tmavozelená barva, pylová sterilita, zpožděná senescence chloroplastů a listů, změny v reakci ke světlu. det2 det2 WT WT


26 26 Aplikace exogenních BR vede k normalizaci mutantních fenotypů CL - campesterol CT - cathasterone TE - teasterone DT - dehydroteasterone TY - typhasterol CS - castasterone BL - brassinolide
27 Biosyntéza brasinosteroidů 27 Kontrolní mechanizmy regulující hladinu BR: - regulace biosyntézy - inaktivace BR - zpětná vazba od signální dráhy Dvě biosyntetické dráhy BR ranná C-6 oxidace pozdní C-6 oxidace Rostliny Campesterol BR Živočichové Cholesterol Steroidy Klíčové kroky syntézy BR a steroidů jsou vysoce konzervované přítomnost 5 -reduktázy
28 28
 WT brd2 BR-insensitivní mutanti jsou podobní mutantům s defektem v syntéze BR.")
29 Signální dráhy brasinosteroidů 29 Mutovaná populace rostlin Identifikace BR-insensitivních mutantů 0 BL Vysoká konc. BR BR-insensitivní mutanti rýže Mutant bri1 (brassinosteroid-insensitive1) WT brd2 BR-insensitivní mutanti jsou podobní mutantům s defektem v syntéze BR. Jejich fenotyp však nelze změnit exogenními BR.



30 30 Laboratoř J. Chory identifikace dalších trpasličích alel bri1 Plně funkční BRI1 je pozitivním regulátorem signálních drah BR Další geny této skupiny: BRL1, BRL2 a BRL3 BRL1 a BRL3 se specificky exprimují ve vaskulárním systému => mutace brl1 a brl3 vedou k abnormálnímu poměru v diferenciaci floému a xylému. Trojitý mutant bri1brl1brl3 - zesílený trpasličí vzrůst a vaskulární fenotyp bri1 bri1brl1brl3 Vazebné studie: vysoká specificita vazby BL k BRI1 proteinu BRI1 je receptorem brasinosteroidů WT brl1 brl3
31 N-terminal signal peptide Cysteine pair Struktura receptoru BRI1 Putative leucine zipper amino acid island LRR22 Leucine-rich repeats (LRR) BL-binding site Transmembrane region PM BRI1 kóduje leucin-rich repeat (LRR) transmembránovou receptor-like kinázu (RLK) (LRR-RLK) (poziční klonování) Lokalizace: plazma membrána (exprese BRI1-GFP) Ser/Thr kinase domain (KD) C-terminal tail Juxtamembrane domain Cytoplazma
 P P C-terminal tail P P Hothorn M et al.")
32 Fosforylace receptoru BRI1 Analýza mutantů s defekty v BRI1 Identifikace domén nutných pro přenos signálu 32 BL-binding site 70-amino acid island LRR22 Juxtamembrane domain P P Ser/Thr kinase domain (KD) P P C-terminal tail P P Hothorn M et al. (2011) Nature 474:
33 Buněčná stěna Přítomnost BL = vazba BL k receptoru 33 Cytozol Plazm. membrána Aktivovaný BRI1/BAK1 heterodimer inhibuje aktivitu BIN2 kinázy prostřednictvím BSK (Brassinosteroid-Signaling Kinases) a BSU1 (bri1 SUpressor 1) + Přímá aktivita BSU1 Akumulace defosforylované formy BES1 a BZR1 v jádře BES1 (bri1-ems-suppressor 1) BZR1 (brassinazole-resistant 1) BES1 a BZR1 transkripční faktory BR-indukovaných genů; krátká životnost; degradace 26S proteasome Jádro Represe nebo aktivace BR-responzivních genů Represe nebo aktivace BR biosyntetických genů Aktivace či potlačení genové exprese
34 Buněčná stěna 34 Plazm. membrána Absence BR: Fosforylace proteinů BES1 a BZR1 konstitutivně lokalizovaných v jádře. Cytozol Fosforylované BES1 a BZR1 se nemohou vázat k DNA a jsou degradovány BR-indukované geny nejsou exprimovány Jádro Represe nebo aktivace BR-responzivních genů Represe nebo aktivace BR biosyntetických genů
 identifikoval ethylen chemicky jako produkt rostlinného metabolismu. Triple response.")
35 35 c) Ethylen Plynný hormon D. Neljubov (1901) inhibice etiolovaných rostlin svítiplynem; identifikoval ethylen-triple response. H. Cousins 1910 zjistil, že ethylen je přírodní látka plyn skladovaných plodů pomerančů R. Gane (1934) identifikoval ethylen chemicky jako produkt rostlinného metabolismu. Triple response. Dlouho nebyl považován za rostlinný hormon jeho efekty zprostředkovány auxinem, až po objevu plynové chromatografie (1958), rozpoznán jako fytohormon.
36 Struktura, biosyntéza a měření ethylenu 36 Hořlavý plyn, snadno oxidovatelný a hydrolizovatelný! Ethylen je produkován zejména zrajícími plody, při senescenci, poranění a fyziologických stresech, ale i houbami, kvasinkami a bakteriem (E. coli). Koncentrace ethylenu se pohybuje v ml/l.


37 Fyziologické funkce ethylenu 37 Ethylen stimuluje zrání některých plodů, tzv. klimakterických: banány, avokado, jablka, rajčata never ripe WT Neklimakterické plody: citrusy, ananas, vodní meloun,..

38 38 Vliv ethylenu na listovou epinastii + ethylen Epinastické listy rajčete = ohnuté směrem dolů Buňky vrchní části řapíku rostou rychleji než spodní buňky Interakce auxin-ethylen auxin indukuje produkci ethylenu



39 39 Koncentrace ethylenu roste ve stresovaných rostlinách např. po zaplavení aerenchym V ponořených rostlinách dochází v kořenech k hypoxii (nedostatku kyslíku). Hypoxie v kořenech stimuluje tvorbu etylénu. Etylén zvyšuje cytozolickou koncentraci Ca 2+, což indukuje smrt některých buněk v kortexu - vzniká aerenchym. Transport ACC z kořenů, které jsou v anaerobních podmínkách.

40 40 Triple response = trojitá odezva u etiolovaných rostlin 1. Ztloustnutí hypokotylu 2. Zkrácení hypokotylu 3. Vytvoření háčku

41 41 Ethylen indukuje tvorbu adventivních kořenů a kořenových vlásků
")


42 Ethylen urychluje senescenci a opad listů u etr1 nebo ein2 mutantů (receptorový mutanti) prodlužuje délku života rostlin o 30% 42 WT Auxin Auxin etr1 mutant defekt v receptoru pro ethylen Citlivost k ethylenu Citlivost k ethylenu Produkce ethylenu Při opadu listů antagonismus s auxinem

43 43 Ethylen má značné praktické využití v zemědělství: opad plodů, květů, dozrávání, synchronizace kvetení Ethephon - 2-choroethylfosfonová kyselina Ethephon = syntetická látka, která se v rostlině mění metabolismem na ethylen

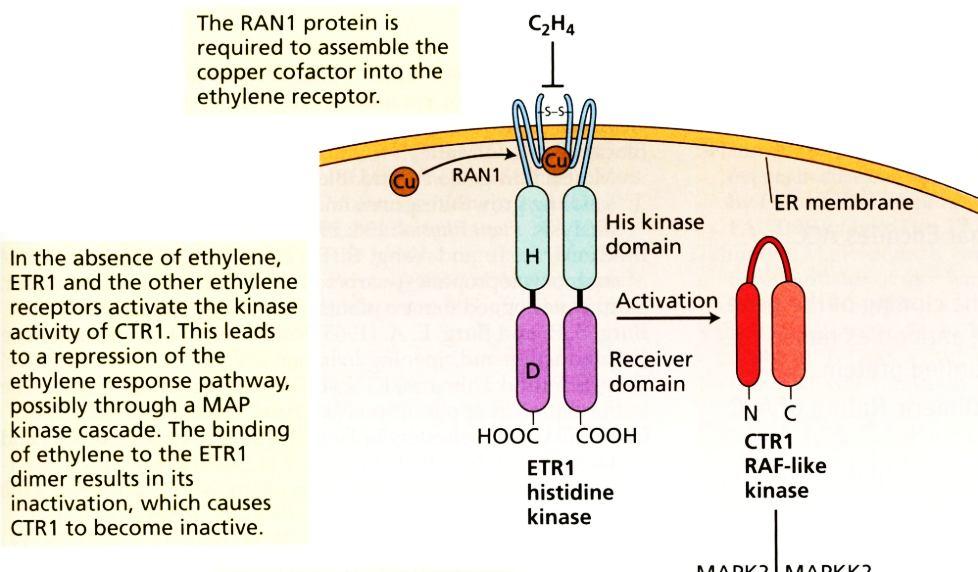
44 44 Signální dráhy ethylenu Izolace mutanta etr1 necitlivého k ethylenu. Receptor ethylenu


45 45 Za nepřítomnosti ethylenu jsou receptory na membráně endoplazmatického retikula aktivní a blokují signální dráhu. Vazba ethylenu k receptorům vede k jejich zablokování. Tím se uvolní signální dráhy a dochází k indukcím reakcí k ethylenu.
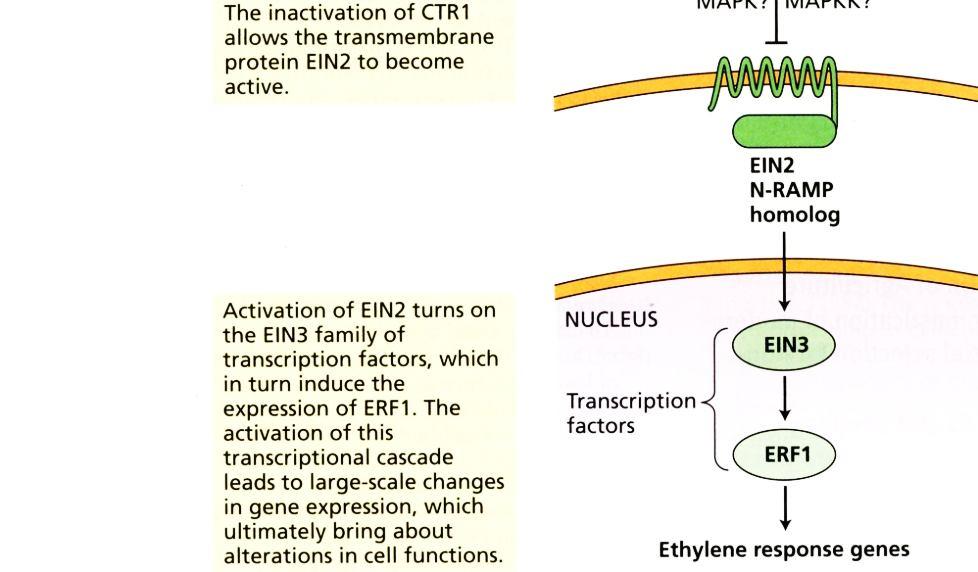
46 46

47 47 d) Kyselina abscisová (ABA) Kyselina abscisová (ABA) (dříve zvaná abscisin II nebo dormin) je inhibiční fytohormon. Zpomaluje růst rostlin, připravuje rostlinu na období vegetačního klidu. Strukturně patří mezi seskviterpeny. ABA má řadu isomerů aktivní a přírodní je pouze S-(cis)-forma
48 48 ABA - historie Frederickem Addicottem při hledání látek, odpovědných za opadávání plodů bavlníku - abscisin I a abscisin II (dnes ABA) Ve stejné době: Philip Wareing - studoval procesy spojené s vegetačním klidem u dřevin a tak tuto látku pojmenoval dormin (od pojmu dormance, období vegetačního klidu u pupenů). Van Stevenincke - studoval opadávání květů a lusků vlčího bobu
 Kyselina 4 -dihydrofazeová (DPA) ABA-aldehyd (C 15 ) Kyselina abscisová (C 15 )")
49 PLASTID isopentenyl difosfát (IPP) Biosyntéza a metabolismus ABA 49 β-karoten Zeaxantin (C 40 ) Stres Trans-violaxantin (C 40 ) Trans-neoxantin 9 cis-neoxantin (C 40 ) CYTOZOL Inaktivace ABA oxidativními procesy Enzym CYP707A Kyselina fazeová Xantoxin (C 15 ) Kyselina 4 -dihydrofazeová (DPA) ABA-aldehyd (C 15 ) Kyselina abscisová (C 15 ) (ABA)

50 Fyziologická funkce ABA 50 ABA inhibuje předčasné klíčení - viviparii Prokázáno u ABA-deficientních mutantů (vp2, vp5, vp7, vp9, vp14) dochází k parciální obnově fenotypu po aplikaci ABA. ABA reguluje akumulaci zásobních proteinů a proteinů zodpovědných za vysušení semen (proteiny LEA late-embryogenesisabundant)
51 51 Dormance semen Klid (dormance) semene je způsoben zejména přítomností ABA v semenných obalech. 2 typy semenné dormance: a) embryo dormance řízena ABA z kotyledonů (líska, jasan, broskev), po odstranění kotyledonů normální růst b) obalem řízená dormance rostliny z aridních oblastí se silným obalem (vojtěška)
 podobě ABAH. Vodní stres: ph xylémového roztoku roste na 7.")
52 ABA uzavírá průduchy (stomata) v závislosti na vodním stresu 52 Zavodněné rostliny: ph xylémového roztoku se pohybuje kolem 6.3 => pohlcování ABA mezofylovými buňkami, protože ABA se nachází v nedisociované (protonované) podobě ABAH. Vodní stres: ph xylémového roztoku roste na 7.2 => stresem indukovaná alkalizace apoplastu upřednostňuje tvorbu disociované formy ABA, ABA -. Dehydratace současně vede k okyselení cytozolu => ABA - se dostává do mezofylových buněk špatně => více ABA - se dostává ke svěracím buňkám, kde se váže na membránový receptor.

53 Vodní stres: Mechanizmus působení ABA ve svěracích buňkách ABA se váže na receptor. 2. Vazba ABA indukuje proud Ca 2+ do buňky. 3. Zvyšuje se intracelulární konc. Ca 2+ a indukuje se dále proud Ca 2+ z vakuoly. 4. ABA stimuluje indukci NO a cadpr. 5. ABA zvyšuje hladinu IP 3 prostřednictvím G proteinu. 6. IP 3 stimuluje proud Ca 2+ z vakuoly. 7. Vysoká hladina Ca 2+ blokuje proud K + do buňky. 8. Vysoká hladina Ca 2+ stimuluje proud Cl - z buňky a způsobuje depolarizaci membrány. 9. Vysoká hladina Ca 2+ blokuje protonovou pumpu, zesiluje se depolarizace. 10. Depolarizace membrány aktivuje proud K + z buňky. 11. K + a Cl - proudí z vakuoly do cytosolu. ROS - Reactive oxygen species IP 3 - Inositol-trifosfát cadpr cyclic ADP-ribose NO nitric oxide (oxid dusnatý) PLC phospholipase C) PLD phospholipase D) PA phosphatidic acid)
 ABA stimuluje růst kořenů")

54 54 Při nízkém vodním potenciálu (ψ W ; nedostatek vody) ABA stimuluje růst kořenů a inhibuje růst výhonů (vp mutant) Při nedostatku vody inhibice tvorby ethylenu v kořenech
55 55 ABA podporuje listovou senescenci nezávisle na ethylenu a auxinu
 Terčem proteinů PP2C je skupina proteinů")
 je aktivní a defosforyluje SNRK2. SNRK2 kináza je tak inaktivována.")
56 Signální dráhy ABA 56 ABA receptory: PYrabactin Resistance 1 (PYR1) (homology PYL a PYR-Like). Vazba ABA k PYR1 vede k inhibici fosfatáz typu 2C (PhosPhatase type 2C, PP2C) Terčem proteinů PP2C je skupina proteinů zvaných SNF1-podobná protein kináza 2 (SNF1-Related protein Kinase 2, SNRK2). Absence ABA: Fosfatáza PP2C (ABI1, ABI2) je aktivní a defosforyluje SNRK2. SNRK2 kináza je tak inaktivována. Přítomnost ABA: Fosfatáza PP2C je inhibována a SNRK kinázy jsou aktivní.
 Aktivace pomalých aniontových kanálů Inhibice K")
57 PYR = PYrabactin Resistance PYL = PYR-Like 57 RCARs = Regulatory Component of ABA Receptors Receptory ABA Fosfatázy - ABA PYR/PYL/RCARs PP2Cs SnRK2s Kinázy ABA? + ABA PYR/PYL/RCARs PP2Cs SnRK2s P ABFs P SLAC1 P KAT1 P Sean Cutler Aktivace ABA-zprostředkované exprese genů (v jádře) Aktivace pomalých aniontových kanálů Inhibice K + kanálů

 Jméno podle Spiraea")
58 e) Kyselina salicylová (SA) 58 Kyselina salicylová Kyselina acetylsalicylová Derivát SA Aspirin Jméno od Salix alba (Vrba bílá) Jméno podle Spiraea (Tavolník)


 V r.")
 V r.")
59 Historie SA 59 Hippocrates psal od použití vrby k odstranění bolesti ~ před 2400 lety Neošetřená strana Strana ošetřená SA V r White ukázal, že ošetření listu aspirinem nebo SA je spojeno s jeho rezistencí k tabákovému viru mozaiky tabáku (TMV) V r bylo ukázáno, že endogenní SA je zodpovědná za produkci tepla v květech kaly (Arum) V r bylo zjištěno, že SA funguje v rostlinách jako endogenní signál při obraných reakcích
60 60 Biosyntéza a metabolismus SA PAL Kyselina chorismová 2 syntetické dráhy SA ICS1 Kyselina trans-skořicová 1. Kys. trans skořicová 2. Kys. chorismová


61 V mnoha rostlinách je gen pro ICS1 upregulován po napadení patogenem. 61 Exprese genu ICS1 se zvyšuje po napadení patogenem ICS1 PR1 PR1 je SA-indukovaný gen exprimovaný po napadení pathogenem Erysiphe houba vyvolávající onemocnění zvané padlí Gram-negativní bakterie




62 62 Fyziologické funkce SA Reakce k patogenům - lokální a hypersensitivní reakce - systemická získaná resistence Interakce Další funkce - abiotický stres - termogeneze
63 63 Signál (např. patogen, UV, teplo, vývojové podněty Zvýšená syntéza SA Exprese patogenindukovaných genů OBRANA Reakce ke stresům Vývojové reakce Hlavním účinkem SA u rostlin je schopnost měnit expresi genů.
64 64 Patern-triggered immunity (PTI) PRR Pathogen-associated molecular patterns (PAMPs) jsou konzervované molekuly jako flagellin nebo chitin. SA Tyto molekuly jsou rozpoznány extracelulárním receptorem PRRs (pattern recognition receptors) Obranné reakce Rozpoznání PAMP spouští syntézu SA a indukci obranných genů = pattern-triggered immunity (PTI)

65 Effector-triggered immunity (ETI) 65 Některé patogeny překonávají PTI tím, že do buňky vypouští efektory. Buňka vytváří R-proteiny, které rozpoznávají efektory a spouští tak zvýšenou reakci. SA SA Obranné reakce Obranné reakce Efektorem-spouštěná imunita
, zesílení buněčné stěny, indukci PR genů a buněčnou smrt.")
66 66 Lokální reakce hypersensitivní reakce Rozpoznání patogenu spouští lokální hypersensitivní reakci, která zahrnuje oxidativní výbuch (oxidative burst), zesílení buněčné stěny, indukci PR genů a buněčnou smrt. Zpevnění buněčné stěny Akumulace ROS SA Buněčná smrt, indukce PR genů

67 Hypersensitivní reakce (HR) uzavře patogena v hrobě mrtvých buněk 67 HR zabíjí infikované buňky a buňky okolní a tak pomáhá bránit rozšiřování patogena.
")
 po inokulaci TMV. Experiment demonstruje systemickou signalizaci.")

68 Systemická získaná resistence (systemic acquired resistance; SAR) 68 Popsána u listů tabáku infikovaných virem tabákové mozaiky (TMV - Tobacco Mosaic Virus) Tato rostlina nebyla preinokulována TMV a ukazuje velké oblasti léze (poškození) po inokulaci TMV. Experiment demonstruje systemickou signalizaci. Tato rostlina byla pre-inokulována TMV na třech spodních listech. Po 7 dnech byly inokulovány TMV tři vrchní listy. Tyto vrchní listy ukazují zvýšenou rezistenci k TMV.

")

69 Termogeneze 69 SA Aminy + Zápach Teplo Látka indukující termogenezi byla poprvé nazvána calorigen. Na konci 80. let bylo zjištěno, že jde právě o SA. Aplikace SA na rostliny kaly vede k indukci termogeneze. Masařka na květu Helicodiceros muscivorus Zmijovec titánský (áronova hůl) (Amorphophallus titanum) Některé rostliny čeledi Aracea páchnou jako hnijící maso a lákají hmyz. Květy produkují teplo, které udržuje zápach a napodobuje tak rozkládající se tělo.
 Plant Cell.")
70 Signální dráhy SA 70 SA SA indukuje enormní transkripční reakce PR PR geny jsou skupinou tzv. pathogenesis related genů. Jsou zapojeny v obraných reakcích rostlin Ward ER et al. (1991) Plant Cell. 3:
71 71 Mnoho genů reaguje k SA: rané geny zesilují signál Rané ICS1 EDS1 PAD4 NPR1 Několik WRKYs (transkripční faktory) Rozeznání patogena EDS1 / PAD4 ICS1 Pozdní PR-1 PR-5 BGL2 NIMIN1 ATNUDT6 FRK1 Několik WRKYs (transkripční faktory) SA NPR1 Transkripční reakce Van den Burg HA, Takken FLW (2009) Trends Plant Sci 14:
 Delaney TP et al.")
72 72 Wild-type npr1 (aka nim1) NPR1 (NONEXPRESSOR OF PATHOGENESIS- RELATED GENES1) je nutný v obranných reakcích a je klíčový v SA signalizaci. Rostliny byly ošetřeny SA. Po 3 dnech je rostlina ochromena pathogenem (Hyaloperonospora arabidopsidis) Delaney TP et al. (1995) PNAS 92:


 Cell 88: 57-63 Cao H et al.")
73 NPR1 je nezbytný a dostatečný pro downstream signalizaci a obranu rostliny 73 Loss-of-function mutant: více citlivý Wild-type npr1 SA Gain-of-function mutant: rezistentní Pseudomonas syringae infection NPR1 PR gene expression Bgl2-GUS (PR gene) expression Resistance Cao H et al. (1997) Cell 88: Cao H et al. (1998) PNAS 95:

74 NPR1, NPR3 a NPR4 jsou receptory SA 74 V roce 2012 dvě skupiny vědců nezávisle na sobě publikovaly závěr, že NPR jsou receptory SA. NPR1 je regulován přímo vazbou SA a/nebo NPR3/4 váže SA a reguluje NPR1 Fu ZQ et al. (2012) Nature 486:
 Cell Rep.")
75 NPR1 je aktivován vazbou SA 75 NPR1 váže SA, dochází ke spuštění konformačních změn, které uvolňují vlastní C-terminální aktivační doménu z inhibice a dochází ke spuštění transkripce. Wu Y et al. (2012) Cell Rep. 1:
![NPR3 a NPR4 regulují proteolytickou likvidaci NPR1 76 Neinfikovaná rostlina Infikovaná rostlina LOW [SA]; some NPR1 accumulates VERY HIGH](/docs-images/91/106500167/images/76-0.jpg "[SA]; NPR1 degraded, cell death occurs Intermediate [SA]; some NPR1 accumulates and activates defenses Fu ZQ et al.")
76 NPR3 a NPR4 regulují proteolytickou likvidaci NPR1 76 Neinfikovaná rostlina Infikovaná rostlina LOW [SA]; some NPR1 accumulates VERY HIGH [SA]; NPR1 degraded, cell death occurs Intermediate [SA]; some NPR1 accumulates and activates defenses Fu ZQ et al. (2012) Nature 486:
77 77 Jaderný protein NPR1 aktivovaný SA stimuluje transkripci genů obranných reakcí NO SA + SA NPR1 -WRKYs -TGAs +WRKYs +TGAs +TGAs Exprese obranných genů je potlačena negativními regulátory WRKY. Exprese obranných genů je stimulována pozitivními regulátory WRKY.
78 78 WRKYs je velká skupina transkripčních faktorů Arabidopsis: 74 TF WRKY Rýže: 90 TF WRKY Genes induced by WRKYs include NPR1 and ICS1 signal amplification! -WRKY +WRKY ICS1 SA NPR1 NPR1 +WRKY Mnoho TF WRKY je zapojeno v obranné signalizaci. Některé stimulují, některé potlačují transkripci.

79 f) Kyselina jasmonová (JA) 79 V roce 1962 byl z jasmínu bílého (Jasminum grandiflorium) izolován metyl jasmonát Metyl jasmonát voní jako jiné estery jde o dominantní vůni jasmínových květů. CH 2 COOCH 3 O Metyl jasmonát
80 Metyl jasmonát se tvoří z kyseliny jasmonové (jasmínové) (JA) 80 Jasmonic acid (JA) JA-Isoleucine (JA-Ile) OH (3R,7S)-jasmonic acid a.k.a. (+)-7-iso-jasmonic acid JA-methyl transferase (JMT) Methyl jasmonate (MeJA) JA-Ile OCH 3 Jasmonoyl izoleucin (JA-Ile) je nejúčinějším jasmonátem. MeJA je transportovatelná forma jasmonátů.
 Syntéza jasmonátů probíhá v plastidech, peroxizomech a cytoplazmě.")
81 Biosyntéza JA DAD1, DGL, GLA1 = lipolytické ezymy 81 Prekurzory jasmonátů jsou odvozeny od membránových lipidů volných mastných kyselin. LOX OPDA = 12-oxophytodienoic acid OPR3 Transport OPDA do peroxizomu JA-methyl transferase (JMT) Syntéza jasmonátů probíhá v plastidech, peroxizomech a cytoplazmě. β-oxidace proces, který vede k odstranění uhlíkových skupin (OPDA 18 skupin, JA 12 skupin). JA může být konjugována s aminokyselinami pomocí enzymu JAR1 (jasmonic acid-amido synthetase).



82 Fyziologické funkce JA 82 Produkce MeJA je kontrolována vývojově a indukována poraněním Exprese JMT dosahuje maxima při praskání prašníků JMT - JA-methyl transferase Zvýšená produkce MeJA vede k ochraně proti nekrotrofickým patogenům JMT je indukována poraněním nebo MeJA


83 Jasmonáty se podílejí na obraně rostlin k hmyzu a nekrotrofům. 83 Hmyz a nekrotrofy spouští produkci jasmonátů. Biotrofy spouští produkci salicylátů. Jasmonates Salicylates Transkripční reakce


84 84 Jasmonáty přispívají ke kontrole vývoje a růstu Samčí a samičí reprodukční vývoj Tvorba trichomů Kontrola buněčného cyklu, senescence a vzájemných interakcí.

 JAZ Z MYC2 BD AD J SCF COI1 JA- Ile MYC2 BD AD Gene expression SCF =")
85 Signální dráhy JA 85 JA-Ile se váže ke co-receptoru COI1-JAZ (součást SCF komplexu) Dochází k ubiquitinaci a degradaci proteinů JAZ Uvolňuje se transkripční aktivace transkripčních faktorů MYC2 MYC2 aktivuje expresi obranných genů (kódující např. proteiny toxické pro škůdce) JAZ Z MYC2 BD AD J SCF COI1 JA- Ile MYC2 BD AD Gene expression SCF = SKP1, Cullin, and an F-box-containing protein

86 86 COI1 je F-box protein velice podobný auxinovému receptoru TIR1. Tato podobnost naznačuje princip fungování systému.

87 87 JA receptor COI byl identifikován pomocí Arabidopsis mutanta coi1, který je necitlivý ke coronatinu a MeJA WT coi1 WT coi1 MeJA coi1 WT Control COR
 indukuje obranné látky v kultivovaných buňkách kalifornského máku.")
88 88 Coronatin je bakteriální látka, které napodobuje účinky jasmonátů. MeJA Coronatin (COR) je toxin produkovaný některými patogeními bakteriemi. Coronatin napodobuje účinky jasmonátu a strukturně se podobá JA-Ile. COR Control MeJA nebo coronatin (COR) indukuje obranné látky v kultivovaných buňkách kalifornského máku. Coronatine (3R,7S)-JA-Ile Eschscholzia californica Sluncovka kalifornská



89 89 Za přítomnosti jasmonátů proteiny JAZ podléhají rychlé degradaci. Control JAZ-GFP + jasmonate Stabilita proteinů byla testována u transgenních rostlin exprimujících JAZ-GFP nebo JAZ-GUS konstrukty.

90 Rostlinám exprimujícím stabilizované proteiny JAZ chybí obrana proti hmyzu. 90 Rostliny se stabilizovanými proteiny JAZ mají defekt v obranných reakcích. Je to vidět na růstu housenek, které se těmito rostlinami živily.

91 91 Proteiny JAZ obsahují konzervovanou doménu Jas a doménu ZIM/TIFY. Doména Jas usnadňuje interakci JAZ s proteiny COI1 a MYC2. Doména ZIM (také zvaná TIFY) je dimerizační doména sloužící k interakci s dalšími JAZ proteiny a proteinem NINJA.



92 92 Doména ZIM proteinu JAZ umožňuje interakci s proteinem NINJA JAZ NINJA = Novel INteractor of JAZ Z NINJA J NINJA interaguje s TOPLESS (TPL)

93 93 Při absenci hormonů protein TOPLESS potlačuje transkripci. Aux/IAA ARF BD III/IV III/IV EAR TPL JAZ MYC2 AD BD Z J NINJA EAR TPL TOPLESS se váže přímo k EAR doméně represorů Aux/IAA. TOPLESS se váže nepřímo k JAZ prostřednictvím NINJA.
94 Jasmonátová signální dráha 93 JAZ Z NINJA EAR TPL 2. JAZ je ubiquitinován a degradován ve 26S proteazomu. MYC2 BD AD J JA-Ile SCF COI1 JA- Ile 1.JA-Ile se váže k SCF COI1 a proteinu JAZ 3. Degradace represoru umožňuje aktivaci transkripce transkripčními faktory MYC2. MYC2 BD AD
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
3) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Průduchy regulace příjmu CO 2
 Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů
 Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů") MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce)
") Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce) Úkol: Sepište krátký rukopis vědeckého původního článku na téma "Směrovaný transport auxinu přes plazmatickou membránu hraje úlohu
Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce) Úkol: Sepište krátký rukopis vědeckého původního článku na téma "Směrovaný transport auxinu přes plazmatickou membránu hraje úlohu
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
CZ.1.07/1.1.00/
 Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
10) Reakce rostlin k abiotickému stresu
 Reakce rostlin k abiotickému stresu") 1 2015 10) Reakce rostlin k abiotickému stresu f) Tepelný stres a tepelný šok g) Kyslíkový deficit Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing Ashraf M et al. (2005) Abiotic Stresses.
1 2015 10) Reakce rostlin k abiotickému stresu f) Tepelný stres a tepelný šok g) Kyslíkový deficit Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing Ashraf M et al. (2005) Abiotic Stresses.
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů
 1 2018 Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů a) Brasinosteroidy Arabidopsis b) Podobné účinky brasinosteroidů a steroidů c) Ecdysteroidy Drosophila Martin Fellner Laboratoř
1 2018 Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů a) Brasinosteroidy Arabidopsis b) Podobné účinky brasinosteroidů a steroidů c) Ecdysteroidy Drosophila Martin Fellner Laboratoř
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů
 1 Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů a) Brasinosteroidy Arabidopsis b) Podobné účinky brasinosteroidů a steroidů c) Ecdysteroidy Drosophila Martin Fellner 2015 2 Vývoj organismu
1 Přenos signálu brasinosteroidů v rostlinách a steroidů u živočichů a) Brasinosteroidy Arabidopsis b) Podobné účinky brasinosteroidů a steroidů c) Ecdysteroidy Drosophila Martin Fellner 2015 2 Vývoj organismu
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Autophagie a imunitní odpověd. Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha
 Autophagie a imunitní odpověd Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha Ostrava, 29. ledna 2019 Historie Nobel Prize 2016 Yoshinori Ōsumi https:p//nobeltpizrog/utplodss/2l018//06/ohsuiillchtul
Autophagie a imunitní odpověd Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha Ostrava, 29. ledna 2019 Historie Nobel Prize 2016 Yoshinori Ōsumi https:p//nobeltpizrog/utplodss/2l018//06/ohsuiillchtul
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
I rostliny mají hormony!... co a jak s auxinem. Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha
 I rostliny mají hormony!... co a jak s auxinem Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha Rostliny a tvarová (a vývojová) různorodost Reakce na okolní prostředí Reakce na nepříznivé podmínky
I rostliny mají hormony!... co a jak s auxinem Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha Rostliny a tvarová (a vývojová) různorodost Reakce na okolní prostředí Reakce na nepříznivé podmínky
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
BIOLOGICKÁ MEMBRÁNA Prokaryontní Eukaryontní KOMPARTMENTŮ
 BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
2012/2013. Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol.
 2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
Joanne Chory (1955) VORF-4.
 VORF-4.") VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Regulace metabolizmu lipidů
 Regulace metabolizmu lipidů Principy regulace A) krátkodobé (odpověď s - min): Dostupnost substrátu Alosterické interakce Kovalentní modifikace (fosforylace/defosforylace) B) Dlouhodobé (odpověď hod -
Regulace metabolizmu lipidů Principy regulace A) krátkodobé (odpověď s - min): Dostupnost substrátu Alosterické interakce Kovalentní modifikace (fosforylace/defosforylace) B) Dlouhodobé (odpověď hod -
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy
 3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy Co to jsou hormony? termín hormon poprvé použitý v medicíně před 100 lety, pochází z řečtiny ηορμονε = stimulovat (ale nezahrnuje
3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy Co to jsou hormony? termín hormon poprvé použitý v medicíně před 100 lety, pochází z řečtiny ηορμονε = stimulovat (ale nezahrnuje
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky Reakce průduchů na světlo
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA
 TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 2 1 2017 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gunawardena,
2 1 2017 7) Senescence a programová smrt buňky (PCD) a) Typy buněčné smrti b) PCD v životním cyklu rostlin c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gunawardena,
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Protinádorová imunita. Jiří Jelínek
 Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.)
") Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
AMPK AMP) Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze
 Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze") AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
VORF Winslow Russel Briggs ( )
") VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Antigeny. Hlavní histokompatibilitní komplex a prezentace antigenu
 Antigeny Hlavní histokompatibilitní komplex a prezentace antigenu Antigeny Antigeny: kompletní (imunogen) - imunogennost - specificita nekompletní (hapten) - specificita antigenní determinanty (epitopy)
Antigeny Hlavní histokompatibilitní komplex a prezentace antigenu Antigeny Antigeny: kompletní (imunogen) - imunogennost - specificita nekompletní (hapten) - specificita antigenní determinanty (epitopy)
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
6) Reakce rostlin k abiotickým stresům
 Reakce rostlin k abiotickým stresům") MBRO2 1 2018 6) Reakce rostlin k abiotickým stresům f) Tepelný stres a tepelný šok g) Kyslíkový deficit Nejnovější review: Obecné: Flood PJ, Hancock AM (2017) Current Opinion in Plant Biology 36: 88-94
MBRO2 1 2018 6) Reakce rostlin k abiotickým stresům f) Tepelný stres a tepelný šok g) Kyslíkový deficit Nejnovější review: Obecné: Flood PJ, Hancock AM (2017) Current Opinion in Plant Biology 36: 88-94
Propojení metabolických drah. Alice Skoumalová
 Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
8. Polysacharidy, glykoproteiny a proteoglykany
 Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Úvod do biologie rostlin Úvod PŘEHLED UČIVA
 Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Teoretický úvod: FOTOTROPISMUS. Praktikum fyziologie rostlin
 Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů