Metody průkazu tvorby biofilmu u lékařsky významných mikrobů
|
|
|
- Daniel Jaroš
- před 6 lety
- Počet zobrazení:
Transkript
1 MASARYKOVA UNIVERZITA V BRNĚ LÉKAŘSKÁ FAKULTA MIKROBIOLGICKÝ ÚSTAV Metody průkazu tvorby biofilmu u lékařsky významných mikrobů (Dizertační práce) Brno 2007 Ing. Veronika Holá
2 Poděkování Ráda bych na tomto místě poděkovala všem, kteří se větší či menší měrou stali podporou při vzniku této dizertační práce. Na prvním místě musím poděkovat svému školiteli prof. MUDr. Miroslavu Votavovi, CSc., za jeho vedení a cenné podněty k celé mé práci. Ráda bych též poděkovala kolegům MUDr. Filipu Růžičkovi a MUDr. Renatě Tejkalové za vytvoření tvůrčí atmosféry na pracovišti a dobrou spolupráci. Dále bych ráda poděkovala laborantce Zuzce Krškové za její pomoc s časově náročným zpracováním některých pokusů a za její svědomitý přístup k práci. Děkuji též doc. Srdjanu Stepanovicovi (Univerzita Bělehrad) za dobrou a příjemnou spolupráci. V neposlední řadě bych ráda poděkovala též manželovi Vladimírovi za přínosné diskuze o řešených problémech a teplé večeře v nejnáročnějším čase psaní dizertační práce. Můj dík patří také grantovým agenturám, které se svou finanční podporou zasloužily o zdarné řešení výzkumných úkolů - Interní grantové agentuře Ministerstva zdravotnictví ČR (granty IGA MZ a IGA MZ ), Fondu rozvoje vysokých škol (granty podpory tvůrčí práce studentů , a ) a grantu Institutu Danone ( )
3 Summary Chronic infections caused by biofilm-forming bacteria represent serious medical problem nowadays. Their higher prevalence is associated with more frequent use of artificial implants and medical devices, more frequent invasive manipulation and higher number of immunocompromised patients. The problem is much bigger on the ICUs, where the number of immunocompromised patients is even higher. The artificial surface of used implants facilitates adhesion of bacteria, which than form biofilm. For the prevention and treatment of all types on infections we have to know and understand souces and vectors of the transmission of the infectious agents as well as their strategy of survival in both outer and inner environment. This work focuses on the study of biofilm problematics and application of theoretical knowledge into clinical practice. Studies were performed on bacterial strains isolated from human clinical material and from swabs from the environment of the St. Anna Teaching Hospital and Teaching Hospital Brno. The dissertation thesis is a set of 9 papers published, accepted for publication and sent for peer review. Results of all papers are commented and discussed. The papers included in the dissertation thesis can be divided into three thematic fields: I. Theoretical papers summarizing present knowledge about bacterial biofilms on the basis of our work and work of other authors (3 papers); II. Study of culture conditions of bacterial biofilms (2 papers); III. Study of clinical significance of biofilm-positive bacteria (4 papers). The presented studies were finantially supported by the grants of Internal Grant Agency of the Ministry of Health, Czech Republic (grants No. IGA MZ and IGA MZ ), Grant Agency of the Ministry of Schollarship (FRVŠ), Czech Republic (grants No , and ) and grant of the Danone Institute, Czech Republic ( )
4 Obsah 1. ÚVOD A CÍL 6 2. LITERÁRNÍ PŘEHLED ŘEŠENÉ PROBLEMATIKY Co je biofilm Vznik biofilmu Struktura biofilmu Možnosti průkazu biofilmu Christensenova zkumavková metoda Christensenova metoda v mikrotiračích destičkách Mikroskopické metody Kultivace na agaru s kongo červení Průkaz genů zodpovědných za tvorbu biofilmu Další metody detekce a studia biofilmu Kvantifikace biofilmpozitivních mikrobů v klinickém materiálu Makiho rolovací metoda Kvantitativní vyšetření katétrů pomocí sonifikace Rezistence buněk v biofilmu k antimikrobiálním látkám Snížený průnik látek do biofilmové vrstvy pomalý růst v biofilmu a heterogenita mikrobiální populace Vznik tzv. perzistorů Univerzální odpověď na stres Vznik biofilmového fenotypu Další mechanizmy uplatňující se v biofilmové rezistenci PUBLIKACE ZAŘAZENÉ DO DIZERTAČNÍ PRÁCE KOMENTÁŘE K PŘEDKLÁDANÝM PRACÍM Quantification of biofilm in microtiter plate: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci 4.2 Biofilmy a antimikrobiální látky Biofilm dutiny ústní Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis
5 4.5 Dynamic of Staphylococcus epidermidis biofilm formation in relation to nutrition, temperature and time 4.6 Biofilm detection and the clinical significance of Staphylococcus epidermidis isolates 4.7 Rozdíly v citlivosti k antibiotikům u biofilmpozitivních a biofilmnegativních kmenů Staphylococcus epidermidis izolovaných z hemokultur 4.8 Stanovení citlivosti k antibiotikům u biofilmpozitivních forem mikroorganizmů 4.9 Selective pressure of higher sanitation standards increases relative number of biofil-positive microbes on the intensive care unit ZÁVĚR LITERATURA PŘEDKLÁDANÉ PRÁCE
6 1. Úvod a cíl Bakteriální biofilmy jsou povlaky bakterií, které se mohou tvořit prakticky na všem, od zubů a protetických náhrad kloubů, srdečních chlopní, či žilních katétrů až po vodovodní sítě, odpadní potrubí a kýly lodí. Celosvětově způsobují bakteriální biofilmy svým růstem a korozí materiálů ekonomické škody vyčíslované na miliardy dolarů. Nemenší problémy a stejně rozsáhlé škody mohou být bakteriálním biofilmům přičteny v oblasti lidského zdraví, hygieny a medicíny. Svými vlastnostmi se bakterie rostoucí v biofilmech zásadně liší od stejných kmenů planktonicky rostoucích bakterií. V medicíně se biofilmy pro svou schopnost adherence k povrchům implantovaných chirurgických náhrad a zařízení či k poškozeným tkáním mohou stát příčinou přetrvávajících a těžko léčitelných infekcí a schopnost růstu bakterií v biofilmu je též spojována s přirozenou rezistencí těchto biofilmů k účinkům antibiotik. Bakterie v biofilmu unikají obranným schopnostem hostitele a odolávají antimikrobiální terapii. Pro prevenci a léčbu všech typů infekčních onemocnění je primárním úkolem poznání a pochopení zdrojů a vektorů přenosu infekčních agens a také strategií jejich přežívání ve vnějším i vnitřním prostředí. Předkládaná práce je zaměřena na studium problematiky biofilmů a aplikaci teoretických poznatků o biofilmech do klinické praxe. Studie byly prováděny na kmenech bakterií izolovaných z humánního klinického materiálu či ze stěrů a seškrabů z prostředí klinik FN u svaté Anny v Brně a FN Brno, jak je uvedeno v jednotlivých publikacích. Předkládaná dizertační práce je koncipována jako soubor prací již publikovaných, přijatých do tisku nebo zaslaných k recenznímu řízení. Každá práce je opatřena stručným komentářem dosažených výsledků a diskuzí. Publikace zahrnuté v dizertační práci lze rozdělit do tří tématických okruhů: 1) Teoretické práce shrnující současné poznatky o bakteriálních biofilmech na základě našich prací i na základě prací dalších autorů (3 práce); 2) Studium růstových podmínek bakteriálních biofilmů a jejich dynamiky (2 práce); 3) Studium klinického významu biofilmpozitivních bakterií (4 práce)
7 2. Literární přehled řešené problematiky 2.1 CO JE BIOFILM Termínem biofilm se rozumí výsledek adherence mikrobů na různé povrchy, ať již umělé či nativní. Na nich vznikají mnohovrstevné shluky mikrobů, mikrokolonie, obklopené vrstvou polymerního extracelulárního materiálu. Mikroby rostoucí v podobě biofilmu se vyznačují zvýšenou odolností vůči obranným mechanizmům hostitele a schopnost růstu bakterií v biofilmu je též spojována s přirozenou rezistencí těchto biofilmů k účinkům antibiotik, což je z hlediska klinické mikrobiologie považováno za důležitý faktor virulence společný všem biofilmům (Davey et al., 2000). Biofilmy se tak díky svým schopnostem adherence k povrchům implantovaných chirurgických náhrad a zařízení či k poškozeným tkáním mohou stát příčinou perzistentních infekcí. Mezi nemoci, jež jsou všeobecně spojovány s tvorbou biofilmů, patří periodontitida, osteomyelitida či chronické pulmonální infekce, zejména cystická fibróza. Velmi snadno se biofilmy vytvářejí na různých katétrech např. močových, hemodialyzačních a cévních, dále na srdečních implantátech, jako jsou kardiostimulátory a umělé srdeční chlopně, na cévních náhradách, na umělých kloubních náhradách a kloubních hlavicích, tracheálních cévkách apod. (Stewart et al., 2001). Zvláště závažné jsou infekce krevního oběhu, především katétrové sepse. Odhaduje se, že biofilmy jsou spojeny s 65 % nozokomiálních infekcí (Licking E., 1999). V klinické mikrobiologii se problematikou tvorby biofilmu zabýváme zejména ve dvou případech, a to posuzujeme-li faktory virulence daného kmene mikroba a tedy i jeho případný klinický význam nebo hodnotíme-li citlivost daného kmene mikroba k antimikrobiálním látkám a volíme možnosti terapie. Kmen, který je schopen tvořit biofilm, je považován za virulentnější, a to zejména z již výše zmíněných důvodů infekce způsobené biofilmpozitivními kmeny jsou perzistentní, dlouhotrvající a obtížně léčitelné. Na základě tohoto faktoru virulence můžeme mimo jiné lépe rozlišit mezi klinicky relevantním izolátem z hemokultury nebo její pouhou kontaminací, způsobenou např. nesprávným způsobem odběru. Většina autorů se shoduje v tvrzení, že biofilm tvoří častěji ty izoláty z hemokultur, které jsou považovány za klinicky relevantní (Arciola et al., 2002; de Nicolas et al., 1995; Růžička et al., 2004). Při hodnocení možností efektivní terapie biofilmových infekcí je potřeba znát citlivost mikroba v biofilmu k antimikrobiálním látkám. Při stanovení antibiotické citlivosti - 7 -
8 biofilmpozitivních kmenů se používá pojem minimální biofilm eradikující koncentrace antibiotika, tedy MBEC, a minimální biofilm inhibující koncentrace, tedy MBIC (Bardouniotis et al., 2001; Ceri et al., 1999). Minimální biofilm eradikující koncentrace je taková koncentrace antimikrobiálních látek, která zcela likviduje buňky v biofilmu, a minimální biofilm inhibující koncentrace je taková koncentrace antimikrobiálních látek, která zastavuje jejich růst a množení (Ceri et al., 2001). MBEC a MBIC jsou vůči hodnotám minimální inhibiční koncentrace (MIC) a minimální baktericidní koncentrace (MBC) stanoveným konvenčním postupem na planktonické populaci vyšší až o čtyři řády (Costerton, 1999; Holá et al., 2004a). Například podle Ceriho et al. (1999) je potřeba použít u ampicilinu více než 500násobnou dávku ve srovnání s MIC, aby se o tři řády snížilo množství buněk E. coli v biofilmu, a Williams et al. (1997) zjistili, že ke snížení počtu buněk S. aureus v biofilmu o tři řády je potřeba více než desetinásobek MBC (minimální baktericidní koncentrace) vankomycinu. Přestože se tomuto tématu věnuje celosvětově mnoho autorů, neexistuje zatím bohužel standardní metodika stanovení MBEC a MBIC Vznik biofilmu Tvorbu mikrobiálního biofilmu lze rozdělit do několika základních fází do fáze atrakce buněk, jejich adheze, akumulace a jejich disperze. Vzniku biofilmu však obvykle nejprve předchází vznik pelikuly, tj. vrstvičky iontů a molekul biologického původu, které se na základě fyzikálních interakcí naváží na čistý povrch. Pelikula vzniká v prostředí bohatém na organické látky (krevní řečiště, dutina ústní, odpadní vody apod.) během několika málo minut, v organicky chudších prostředích vzniká v řádu několika hodin (Costerton et al., 1995). Pelikula mění povrchové vlastností substrátu a v prostředí chudém na živiny je navíc též jejich zdrojem. V krevním řečišti se na tvorbě pelikuly podílí různé bílkoviny, např. albuminy, fibronektin, ionty Na +, K +, glykoproteiny či další makromolekuly. Pelikula usnadňuje vazbu bakterií. Přítomnost takového povrchu zjistí volně plovoucí bakterie během dalších několika málo minut (fáze atrakce) a buď náhodně či s pomocí receptorů se usazují (Costerton, 1999; Stoodley et al., 2002). Tím dochází k fázi adheze a později fázi akumulace. Počáteční vazba je ovlivněna elektrickými náboji vznikajícími mezi povrchem a bakteriemi (tvorba vodíkových můstků), Van der Waalsovými silami a elektrostatickou přitažlivostí povrchu a bakterie. Později bakterie pevně adherují za vzniku specifických vazeb. Tyto vazby mají povahu chemickou, nebo se jedná o vazby mezi receptorem na povrchu bakterie a adhezinem na kolonizovaném povrchu, které jsou specifické pro každý konkrétní druh bakterií
9 K pevně adherovaným bakteriím koadherují další bakteriální buňky, čímž vznikají mikrokolonie (Costerton et al., 1999; Mack et al., 2004), dochází k jejich rozrůstání dělením i další koagregací. Usazené buňky produkují polymerní exoprodukty, zejména polysacharidové povahy, které se stávají vedle bakterií jednou ze základních stavebních složek biofilmové vrstvy. Do této vrstvy se dále mohou vázat i bílkoviny a buňky z těla hostitele. Během několika hodin až dní vzniká trojrozměrná struktura zralého biofilmu (obr. 1) s houbovitými strukturami bakterií a exopolysacharidů a s dutinami a kanálky umožňujícími přísun živin i do hlubších vrstev biofilmu (podrobněji viz kapitola 2.1.2). Na nativních površích Obr. 1 Model struktury zralého biofilmu s proudící kapalinou. VH lidského těla (dutina ústní, střevní sliznice, vagina) se tvoří rozsáhlá konsorcia různých druhů bakterií. K bakteriálnímu biofilmu mohou dále přisedat i různé druhy dalších mikroorganizmů (améby, kvasinky apod.). Při kolonizaci umělých povrchů se jedná většinou o biofilm jednodruhový, výjimku tvoři biofilm močových katétrů. Po dosažení určitě buněčné hustoty se ze zralého biofilmu začínají oddělovat buď jednotlivé buňky, nebo celé jejich shluky i s částmi biofilmu, dochází k fázi disperze. Buňky se uvolňují zpět do prostředí a mohou kolonizovat další povrchy vhodné pro růst biofilmu. Tím v biofilmu vzniká dynamická rovnováha, která je regulována dostupností živin, podmínkami vnějšího prostředí, ale i mikroby samotnými, např. systémem quorum-sensing (Davies et al., 1998; O'Toole et al., 2000; Stoodley et al., 2002) či produkcí látek, které degradují polymerní struktury biofilmu a napomáhají tak uvolnění vlastních bakteriálních buněk z jeho struktury (Davey et al., 2000). Systém quorum sensing je hlavní regulační a komunikační prostředek bakteriálního společenstva (Stoodley et al., 2002; Davies et al., 1998). Umožňuje bakteriím reagovat na jejich počet v určitém prostoru, o čemž svědčí i termín quorum sensing, tedy vnímání množství. Quorum sensing systém je založen na produkci signálních molekul a jejich transportu vně buněk, kde jsou signální molekuly rozpoznány receptory okolních bakteriálních buněk, čímž mohou ovlivnit jejich chování. Vnímání populační hustoty je - 9 -

10 bakteriemi využíváno jako podnět pro regulaci celé řady buněčných funkcí (Singh et al., 2000; Vuong et al., 2000) Struktura biofilmu Struktura biofilmu je podmíněna vlastnostmi prostředí (ph, hydrodynamické síly aj.), v němž se mikroorganizmy nacházejí, přítomností a dostupností živin a kyslíku a přítomnými druhy bakterií. Díky široké škále prostředí, s nimiž se mikroby mohou setkat a ve kterých mohou tvořit biofilm, existuje řada různých forem a struktur biofilmu (Donlan et al., 2002). Základní model biofilmu, který byl popsán při studiu metanogenních bakterií v reaktorech s pevným ložem a biofilmů ve vodovodních potrubích, platí s menšími obměnami pro všechny biofilmy a je znázorněn na obr. 1 (viz výše). Tento modelový biofilm je popisován jako vysoce hydratovaná porézní struktura, která obsahuje shluky mikrobiálních buněk a která je protkána dutinami a kanálky (Robinson et al., 1984; Donlan, 2001). V dutinách a kanálcích protéká tekutina, která přináší živiny a odnáší zplodiny a metabolické produkty. Díky tomu, že v kanálcích proudí tekutina mnohem pomaleji, než v prostředí samotném, mohou bakterie vytěžit maximum živin i z jinak velmi chudých prostředí (Danese et al., 2001). Popsaná porézní struktura biofilmu s dutinami, kanálky a houbovitými útvary je též dobře patrná na mikrofotografii řezu katétru, na němž jsme prokázali biofilm Candida parapsilosis (obr. 2). Extracelulární hmota a voda tvoří celkem asi 98 % objemu bakteriální biofilmové struktury. Kromě polysacharidové matrix obsahuje zpravidla i další organické a anorganické složky získané z vnějšího prostředí a v biofilmu plní ochrannou funkci. Bakteriální buňky jsou díky tomu chráněny před vnějšími vlivy, jako je např. vysychání či účinky UV záření, před predátory (např. amébami v přírodním prostředí, ale i v dutině ústní), imunitním systémem hostitele a před průnikem antibiotik a dezinfekčních prostředků (Raad, 1998; Schierholz et al., 2001). V prostředích s vysokým obsahem živin, jako jsou například odpadní vody či papírenské a potravinářské výroby, jsou biofilmy mnohem kompaktnější a silnější a do matrix biofilmu je zakomponováno mnoho zbytků a živin z vnějšího prostředí. V oligotrofních prostředích je biofilm spíše mozaikovité a nízké struktury. Je-li prostředí úživnější, biofilm Obr. 2 Příčný řez biofilmem Candida parapsilosis (a) kanálek uvnitř biofilmové vrstvy, (b) mikrokolonie, VH
11 získává porézní strukturu s dutinami a kanálky a se shluky mikrobiálních buněk, jak byla popsána výše. Biofilm v lidském těle tvoří obecně komplexnější struktury, než můžeme nalézt v přírodě, a i zde je do biofilmové vrstvy zakomponováno množství látek z vnějšího prostředí (různé makromolekuly, polysacharidy, bílkovinné struktury aj.) (Donlan et al., 2002)
12 2.2 MOŽNOSTI PRŮKAZU BIOFILMU K průkazu tvorby biofilmu se používá jednak metod fenotypových, jednak genotypových. Fenotypové metody jsou založeny na kultivaci mikroba a poté na obarvení vzniklého biofilmu na vnitřním povrchu zkumavky či důlku mikrotitrační destičky (Christensenova metoda), na mikroskopickém pozorování obarvené vrstvy biofilmu např. na povrchu katétru, na posouzení vzhledu kolonií na půdách s kongočervení, nebo na posouzení povrchových vlastností mikroba. Genetické metody se zakládají na průkazu genů pro tvorbu biofilmu, např. na průkazu ica-operonu u S. epidermidis (Frebourg et al., 2000) a na studiu exprese konkrétních genů za konkrétních podmínek (Fitzpatrick et al., 2002) Christensenova zkumavková metoda Christensenova zkumavková metoda je již klasická metoda zaměřená na průkaz schopnosti příslušného mikroba tvořit biofilm. Princip je založen na kultivaci testovaného mikroba ve zkumavce s tryptonosojovým bujonem (TSB) (Christensen et al., 1982) či mozkosrdcovou infuzí (BHI) (Mulder et al., 1998; Knobloch et al., 2002a) a následném obarvení vzniklé biofilmové vrstvy. Při průkazu biofilmu u kvasinek používají někteří autoři též kvasničné médium s dusíkem (yeast nitrogen base) (Hawser et al., 1994; Hawser et al., 1998). Často se využívá i suplementace média glukózou, protože glukóza zvyšuje schopnost tvorby biofilmu (Deighton et al., 1990; Mulder et al., 1998; Dobinsky et al., 2003; Mathur et al., 2006). Kultivační médium se inokuluje zpravidla mikrobiální suspenzí o hustotě 1 dle McFarlandovy zákalové stupnice v množství 100 µl do 2 ml kultivačního média, tj. tak, aby výsledná hustota mikrobiálních buněk v kultivačním médiu odpovídala buněk/ml (Stepanovic et al., 2003). Bylo prokázáno, že s rostoucí hustotou inokula roste i denzita biofilmové vrstvy, proto by mělo být používáno standardizované inokulum (Deighton et al., 2001; Stepanovic et al., 2003). Délka inkubace se pohybuje zpravidla v rozmezí 24 až 48 hodin, v závislosti na druhu mikroba a růstových podmínkách. Teplota inkubace je u bakterií obvykle 37 C, u kvasinek se používá teplota nižší, 30 C (Stepanovic et al., 2000). Po inkubaci jsou zkumavky vyprázdněny, třikrát promyty fyziologickým roztokem či fyziologickým roztokem s fosfátovým pufrem (ph 7,4), aby se vyplavily neadherované buňky, a fixovány sušením či fixačními roztoky (Baldassarri et al., 1993). Na vnitřním povrchu zkumavky se potom barvením prokazuje vytvořená biofilmová vrstva. Adherovaná vrstva biofilmu se barví 20 minut krystalovou violetí, safraninem, karbolfuchsinem či jinými

13 barvivy (Christensen et al., 1982; Knobloch et al., 2002a; Moretro et al., 2003; Tormo et al., 2005; Mulder et al., 1998). Nejčastěji je využíváno barvení krystalovou violetí. Jako biofilmpozitivní jsou hodnoceny kmeny s dobře patrnou obarvenou vrstvou v celé části zkumavky původně naplněné kultivačním médiem. Jako biofilmnegativní jsou hodnoceny Obr. 3 Christensenova zkumavková metoda, biofilmnegativní a biofilmpozitivní kmen, VH kmeny nevykazující žádnou obarvenou vrstvu na stěnách zkumavky, případně jen proužek v místě původní hladiny média, viz obr. 3. Pro lepší interpretaci získaných výsledků se pro hodnocení využívá ordinální stupnice 0 3 (0 netvoří biofilm; 1 slabý producent biofilmu; 2 středně silný producent biofilmu; 3 silný producent biofilmu). Christensenovu zkumavkovou metodu lze provádět ve skleněných i v plastových zkumavkách (například polystyrenových či polypropylenových). Při použití různých materiálů je však nutno mít na zřeteli, že různé druhy mikrobů adherují k různým povrchům odlišně, pravděpodobně kvůli rozdílným fyzikálně-chemickým vlastnostem jejich povrchů, jejich hydrofilitě či hydrofobicitě, přítomnosti elektrického náboje apod. (Kennedy et al., 2004; Kiers et al., 2001; Holá et al., 2004c). Nevýhodami Christensenovy zkumavkové metody jsou nemožnost přesně kvantifikovat nárůst biofilmu a značná subjektivita hodnocení výsledku. Výhodou je naopak jednoduchost jejího provedení a nenáročnost k laboratornímu vybavení. Díky těmto výhodám se Christensenova zkumavková metoda řadí mezi metody dobře využitelné v rutinní laboratorní praxi běžné laboratoře klinické mikrobiologie Christensenova metoda v mikrotitračních destičkách Poměrně často se používá modifikované Christensenovy zkumavkové metody, která je založena na kultivaci testovaného mikroba v jamkách mikrotitrační destičky a hodnocení jeho nárůstu na stěnách těchto jamek (Christensen et al., 1985). Kultivace probíhá opět v TSB či BHI za stejných kultivačních podmínek jako v případě Christensenovy zkumavkové metody. Množství inokula se zmenšuje úměrně kultivačnímu objemu jamek mikrotitrační destičky tak, aby výsledná hustota bakteriální suspenze bylo opět buněk/ml. Po inkubaci, fixaci a následném obarvení je síla biofilmového nárůstu hodnocena spektrofotometricky (obr. 4). Christensen et al. (1985) měřili spektrofotometricky sílu biofilmové vrstvy vytvořené na dně jamky mikrotitrační destičky. Přesnější je však
, kdy se použité barvivo v jamkách rozpustí etanolem, jehož zabarvení se pak odečítá spektrofotometrem při vhodných vlnových délkách, tj. 595 620 nm.")
14 modifikace tohoto postupu dle Stepanovice (Stepanovic et al., 2000), kdy se použité barvivo v jamkách rozpustí etanolem, jehož zabarvení se pak odečítá spektrofotometrem při vhodných vlnových délkách, tj nm. Výsledkem je hodnota optické denzity (OD) každého testovaného důlku, která odráží akumulaci barviva v biofilmu nejen na dně důlku, ale i na jeho stěnách. OD jednotlivých důlků se potom porovnává s tzv. cut off value, která se vypočítá na základě OD důlků negativní kontroly. Kmeny, jejichž OD je vyšší než cut off value, jsou považovány za biofilmpozitivní a kmeny, jejichž OD je nižší než cut off value, se považují za biofilmnegativní. Obr. 4 Christensenova metoda v mikrotitračních destičkách, VH Průkaz biofilmpozitivních mikrobů v mikrotitračních destičkách umožňuje kvantifikaci tvorby biofilmu a řeší problém subjektivního hodnocení nárůstu biofilmu. Další nespornou výhodou je možnost kultivace mikroba paralelně ve více jamkách a následné získání průměrné hodnoty OD daného mikroba, což dále zpřesňuje celé hodnocení Mikroskopické metody Při studiu biofilmů se využívají transmisní i rastrovací elektronová mikroskopie (TEM, SEM), konfokální rastrovací laserová mikroskopie (CSLM), mikroskopie atomárních sil (AFM), fluorescenční mikroskopie i běžná světelná mikroskopie (Lawrence et al., 1999; Surman et al., 1996; Palmer et al., 1999). U většiny mikroskopických technik je nutno vzorek předem připravit barvením, fixací, zmražením, dehydratací, řezáním apod. To u tak vysoce hydratovaných struktur, jakými jsou mikrobiální biofilmy, značně ovlivňuje strukturu biofilmové matrix a tedy i výsledný mikroskopický obraz (Stewart et al., 1995). Při mikroskopii atomárních sil není nutná dehydratace vzorku, a proto nedochází k deformacím biofilmu jako u výše uváděných metod. Vlivem pohybu vody kolem senzoru jsou však takto získané fotografie často rozostřené (Bremer et al., 1992). Konfokální rastrová laserová mikroskopie poskytuje dobrý a ostrý obraz, a to bez nutnosti fixace a řezání vzorků. Pomocí optických řezů je CSLM schopna znázornit i struktury ukryté pod povrchem biofilmu a jejich následným poskládáním pak 3D-strukturu (Zaura-Arite et al., 2001; Takenaka et al., 2001). Fluorescenční mikroskopie je co do přístrojového vybavení oproti předchozím uvedeným metodám výrazně levnější. S úspěchem se dá využít např. při studiu prostorového uspořádání jednotlivých složek biofilmu, např. buněk (obr. 5) barvením akridinovou oranží či

.")
15 ethidiumbromidem (Holá et al., 2004d; Yu et al., 1994), nebo naopak polysacharidové matrix barvením kalkofluorem (Stoodley et al., 1998). Pomocí akridinové oranži je rovněž možné vizualizovat heterogenitu bakteriální populace uvnitř biofilmu. Výsledek barvení akridinovou oranží je totiž ovlivněn relativním poměrem RNA/DNA a vede ke vzniku odlišných výsledných barev. To umožňuje rozlišit rychle rostoucí buňky od buněk rostoucích pomalu (Wentland et al., 1996). Buňky s vysokým relativním obsahem RNA se barví do oranžova až oranžovočervena, buňky s nízkým relativním obsahem RNA se barví do zelena až žlutozelena. Mikroskopické metody využívané v běžné světelné mikroskopii jsou založeny na průkazu biofilmu pomocí barvení. Zpracovávat lze tímto postupem přímo klinický materiál Obr. 6 - Oddělování biofilmu od katétru vinou řezání parafinových bločků, VH (v případě biofilmu tedy nejčastěji katétry). Katétr a případný biofilmový nárůst na něm je nejprve fixován (např. ethanolem či Bouinonou tekutinou) (Baldassarri et al., 1993) a obarven a poté nařezán mikrotomem. Výsledné řezy jsou mikroskopicky pozorovány a vyhodnoceny. Protože deparafinace řezů je problematická a celkově méně komfortní, osvědčila se nám lépe než zalévání vzorku do parafínu (obr. 6) technika řezání zmražených katétrů na mikrotomu zmrazovacím. Na takto zpracovaných vzorcích katétru se nám podařilo prokázat biofilmový nárůst kvasinek již v den doručení klinického materiálu, tj. minimálně 24 hodin před pozitivním kultivačním nálezem (obr. 2). Pro laboratorní diagnostiku biofilmpozitivních mikrobů by mohla být výhodou rychlost této metody, kdy výsledek mikroskopie může být klinikovi sdělen již za několik hodin po zaslání katétru. Nevýhodou je nutnost zhotovení velkého množství řezů a prohlédnutí velkého množství zorných polí, zejména u větších katétrů. Další nevýhodou jsou nároky kladené na přístrojové vybavení pracoviště (mikrotom). Obr. 5 Barvení biofilmové vrstvy akridinovou oranží, červeně jsou obarveny buňky rostoucí, zeleně buňky v růstu stagnující, VH Mikroskopickou metodou, použitelnou však jen omezeně a u průhledných materálů, je mikroskopie celého katétru, případně kontaktní čočky, po jejich obarvení (Inglis et al., 1995;

16 Berra et al., 2004). U této metody je však problematické použití imerze a imerzního objektivu, takže výsledné použitelné zvětšení není pro mikroskopii bakteriálního biofilmu příliš vhodné, postačuje však na mikroskopii prvoků v biofilmu, např. améb (Beattie et al., 2006; Simmons et al., 1998) Kultivace na agaru s kongo červení Rozlišení biofilmpozitivních a biofilmnegativních stafylokoků podle charakteristického růstu na agaru s kongo červení bylo poprvé popsáno Freemanem (1989). Agar s kongo červení je agar připravený z mozkosrdcové infuze, 5 % sacharózy a kongo červeně. Nárůst a vzhled kolonií se hodnotí po 24 a 48 hodinách kultivace. Jako biofilmpozitivní jsou hodnoceny kmeny s černými suchými koloniemi s matným povrchem, případně s koloniemi černohnědými s výraznou precipitační zónou v okolí. Jako biofilmnegativní jsou hodnoceny kmeny s koloniemi lesklými, hladkými a červenými až červenohnědými (obr. 7). Nevýhodou této metody je velmi výrazná subjektivita hodnocení. Tato subjektivita je dle našich výsledků vyšší než u Christensenovy zkumavkové metody (Růžička et al., 2006a; Obr. 7 Kolonie Staphylococcus epidermidis na agaru s kongo červení, černé, suché kolonie biofilmpozitivní kmen, červené, hladké kolonie biofilmnegativní kmeny, VH Růžička et al., 2004). Tato subjektivita hodnocení je dále podtržena tím, že vzhled kolonií na agaru se značně liší i u stejného kmene testovaného na různých šaržích agaru s kongo červení. Další nevýhodou je její nízká specifita a velké množství falešně negativních či falešně pozitivních výsledků, na čemž se shoduje řada autorů (Cramton et al., 2001; Deighton et al., 1992) a což odpovídá i našim zkušenostem. Z tohoto důvodu se většina autorů přiklání k tomu, že hodnota tohoto testu je pouze orientační a stanovení biofilmpozitivity či biofilmnegativity kmene by mělo být podloženo ještě některým z dalších testů. Agar s kongo červení byl použit na rozlišení biofilmpozitivních a biofilmnegativních kmenů i u gramnegativních bakterií (Jain et al., 2006). Biofilmpozitivní gramnegativní bakterie na něm mají tvořit hladké krémově zbarvené kolonie, biofilmnegativní bakterie pak kolonie bezbarvé. Množství falešně negativních a falešně pozitivních výsledků však tuto metodu pro gramnegativní bakterie činí stejně rozporuplnou, jako pro stafylokoky
17 2.2.5 Průkaz genů zodpovědných za tvorbu biofilmu Tvorba biofilmu je u složitý proces, na němž se podílí mnoha genů (Beenken et al., 2004; Beloin et al., 2005; Bollinger et al., 2001). U některých mikrobů lze v tomto procesu prokázat geny, jež hrají určitou klíčovou úlohu a které pak můžeme pomocí PCR prokázat. U většiny mikrobů tomu tak však bohužel není a exprimace jednoho genu může být nahrazena exprimací genu se stejným či podobným účinkem (Cheung et al., 2001). V literatuře se uvádí množství genů popsaných v souvislosti s tvorbou biofilmu pro jednotlivé rody či druhy bakterií. U stafylokoků byly takto popsány např. geny icaa, icab, icad a icac, kódující intercelulární adhezin (Frebourg et al., 2000); saa, kódující slime-associated antigen (Ammendolia et al., 1999); atl, atlc, atle a aas, kódující stafylokokové autolyziny (Allignet et al., 2001); bap, kódující biofilm-associated protein (Cucarella et al., 2001), u pseudomonád PvrR kódující zvýšenou rezistenci (Drenkard et al., 2002), u escherichií CsgBAC kódující produkci celulózy a zároveň jeden z kroků tvorby biofilmu (Da Re et al., 2006; Brombacher et al., 2006) apod. Další a neméně významnou komplikací genetických metod průkazu tvorby biofilmu je fakt, že prokázaná přítomnost genu zodpovědného např. za tvorbu biofilmu nám ještě neindikuje fakt, zda je tento gen skutečně exprimován a testovaný kmen tedy biofilm skutečně tvoří. Přes tyto své nevýhody je průkaz genů zodpovědných za tvorbu biofilmu metodou s velkým potenciálem využití. Průkaz genů zodpovědných za tvorbu biofilmu se používá např. u stafylokoků, u kterých existuje pro tvorbu biofilmu klíčový soubor genů, jejichž přítomnost lze prokázat. Tento soubor genů je souhrnně označován jako ica operon a obsahuje geny icaa, icad, icab, icac a dále represivní gen icar. Sekvence primerů pro detekci ica operonu byla popsána Frebourgem et al. (2000). Průkaz těchto genů u klinických izolátů je v současné době využíván spíše jako jedno z kritérií indikujících klinickou významnost izolátu (Arciola et al., 2001; Arciola et al., 2002; Cramton et al., 1999). U ostatních mikroorganizmů nebyly prokázány geny s tak klíčovou úlohou při tvorbě biofilmu, jako je tomu u stafylokoků, a genetické metody jsou využívány spíše k výzkumným účelům (Branchini et al., 1994; Beloin et al., 2005; Burne et al., 1997; Vianney et al., 2005)
18 2.2.6 Další metody detekce a studia biofilmu Měření mikrobiální aktivity Mikrobiální biofilm je relativně slabou, ale hustou vrstvou jednotlivých buněk, což ztěžuje možnosti detekce jednotlivých procesů probíhajících v tomto prostoru. Na velmi malém prostoru totiž probíhá současně mnoho procesů, z nichž některé se doplňují či překrývají a některé působí naopak protichůdně (nitrifikace a denitrifikace, oxidace a redukce sirných sloučenin apod.). Pro měření mikrobiální aktivity se používá buď mikrosenzorů, nebo chemických reakcí. Každý z těchto přístupů se využívá k jinému účelu. Mikrosenzory se zpravidla používají pro měření hladin chemických látek a jejich změn in situ. V současné době jsou k dispozici mikrosenzory pro měření ph či hladin O 2, N 2 O, NH 4, NO3, S, H2 S, - NO 2, CH4, CO 2 aj. (de Beer et al., 1994; de Beer et al., 1997). Hlavním předpokladem použití mikrosenzorů je prostorové oddělení jednotlivých probíhajících procesů (de Beer et al., 1999). Chemické reakce se využívají zpravidla na průkaz enzymatické aktivity konkrétního enzymu jako průkazu živých a životaschopných buněk. Zjišťuje se tak například účinek antimikrobiálních látek na buňky v biofilmu. Nejjednodušší takovou reakcí je změna hodnoty ph a tím změna barvy kolorimetrického média (Holá et al., 2004a), dalším příkladem může být redukce tetrazoliových solí na formazan apod. (Hawser et al., 1994; Hawser et al., 1995; Hawser et al., 1998). Enzymatické chemické reakce lze též využít ke kvantifikaci schopnosti tvorby biofilmu (Dunne, Jr. et al., 1987; Butterfield et al., 2002). Využití povrchových vlastností pro průkaz tvorby biofilmu Povrchové vlastnosti mikrobů lze studovat pomocí mnoha chemicko-analytických technik. Pravděpodobně nejznámější a nejvíce prostudovanou je Matrix-Assisted Laser Desorption/Ionisation, tedy MALDI. Při studiu biofilmu se MALDI využívá zejména k získání informací o složení extracelulární substance (Leone et al., 2006), při studiu syntézy bílkovin v biofilmových a planktonických kulturách (Hefford et al., 2005; Tremoulet et al., 2002) apod. Byly též popsány možnosti využití MALDI pro rychlou identifikaci mikroorganizmů např. ke kontrole čistoty kultur v bioreaktorech (Wunschel et al., 2005). Další možností studia povrchových vlastností buněk jsou elektroforetické metody, z kapilárních např. izoelektrická fokusace (CIEF), která využívá rozdílů v povrchovém náboji amfoterních částic, tedy i mikrobiálních buněk, čímž umožňuje jejich separaci pomocí izoelektrického bodu (pi). Stanovením pi lze bakteriální buňky nejen identifikovat (Horká et al., 2006; Horká et al., 2005), ale lze ji využít i pro odlišení biofilmpozitivních a
19 biofilmnegativních kmenů S. epidermidis (Růžička et al., 2006b). Značnou nevýhodou této metody je její přístrojová náročnost, kvůli které ji v současné době nelze řadit mezi rutinně laboratorně využitelné metody
20 2.3 KVANTIFIKACE BIOFILMPOZITIVNÍCH MIKROBŮ V KLINICKÉM MATERIÁLU Znalost kvantity mikroba v klinickém materiálu je v mnoha případech důležitým vodítkem pro hodnocení závažnosti mikrobiologického nálezu. Proto by se kvantitativní či alespoň semikvantitativní metody průkazu mikrobů měly v klinicko-mikrobiologické laboratoři používat vždy, kdy má kvantifikace z mikrobiologického hlediska smysl a je realizovatelná. Je nutné tedy zhodnotit, zda u konkrétního klinického materiálu převažují důvody pro kvantifikaci, či proti ní. Kromě případné finanční náročnosti a náročnost zpracování pro laboratorní personál je nutné vzít v úvahu též klinickou závažnost jednotlivých materiálů (jinak budeme posuzovat výtěr z nosu a jinak centrální venózní katétr) a také fakt, že pozitivní kultivační nález v některých materiálech je dosti závažný již sám o sobě a není nutné mikroba nutno kvantifikovat (a to i z toho důvodu, že daný klinický materiál je za normální situace sterilní, jako např. likvor). Nejčastějšími biofilmovými infekcemi jsou infekce krevního řečiště, tedy kolonizace katétrů biofilmpozitivními mikroby (Donlan et al., 2002; Raad, 1998), dále pak infekce močových katétrů a dalších umělých povrchů (kloubní hlavice, elektrody srdeční či mozkové, stenty apod). Přímá kultivace katétru má hodnotu pouze potvrzovací a neříká nám nic o původním množství mikroba na katétru. To s sebou nese též vysoké riziko chybné interpretace, pokud byl implantát pouze kontaminován. Znalost kvantity vykultivovaného mikroba nám umožňuje nejen posoudit závažnost infekce, ale i odlišit případnou kontaminaci. Mezi metody používané v laboratorní praxi k průkazu biofilmu na katétru se řadí semikvantitativní metoda dle Makiho (1977) a kvantitativní kultivační metoda dle Bruna (1987) Makiho rolovací metoda Makiho metoda je založena na průkazu nárůstu biofilmu na katétru jeho rolováním po povrchu agaru a následným počítáním CFU (colony-forming units). Její nevýhodu je nemožnost průkazu nárůstu biofilmu na vnitřním povrchu katétru, nepřesná kvantifikace způsobená rozdílnou silou adheze různých mikroorganizmů k povrchu katétru a poměrně složitá manipulace s katétrem, zvyšující riziko kontaminace (Sanger et al., 1989). Přesto nám poskytuje alespoň přibližné vodítko k rozlišení klinicky relevantního nálezu mikroba na

21 katétru od pravděpodobné kontaminace (např. z kůže pacienta, rukama ošetřujícího personálu apod.). Provedení je jednoduché a nevyžaduje žádné zvláštní vybavení. Sterilní pinzetou se katétr přenese na krevní agar, po kterém se krouživými pohyby roluje a válí tak, aby se na agar otiskl celý povrch katétru. Zvláštní pozornost by se měla věnovat špičce katétru, odkud se dá mikrob vykultivovat nejsnáze, tvoří-li biofilm v lumen katétru. Po 24hodinové kultivaci při 37 C se hodnotí množství narostlých kolonií. Nález méně než 15 kolonií svědčí spíše pro kontaminaci katétru, nález více než 15 kolonií lze považovat za klinicky signifikantní. Při katétrových infekcích krevního řečiště bývá nárůst mikroba obvykle masívní. Obr. 8 Rolování katétru po agaru, výsledná kultivace (S. epidermidis), VH Kvantitativní vyšetření katétrů pomocí sonifikace Brun-Buissonova metoda je založena na sonifikaci katétru ve fyziologickém roztoku, který je poté inokulován na agar ke kultivaci a odečtení CFU. Výhodou oproti Makiho metodě je záchyt mikrobů i z lumen katétru a přesnější kvantifikace. Nevýhodou je větší pracnost této metody a potřeba sonikátoru. V literatuře se uvádí několik různých modifikací a postupů sonifikace katétrů. Mezi sebou se liší zejména délkou sonifikace a použitím vortexace nebo třepání jako dalšího pomocného faktoru k rozrušení biofilmové vrstvy (Kobayashi et al., 2006; McDowell et al., 2005; Dworniczek et al., 2003; Sheehan et al., 2004; Nguyen et al., 2002; Cozzaglio et al., 1997). Obsah zkumavky s katétrem (v případě příliš dlouhého katétru je vhodné jej přísně asepticky nastříhat na kousky dlouhé asi 1 cm, aby došlo snáze k vyplavení mikroorganizmů i z lumen katétru) je vhodné promíchat pomocí vortexu či třepačky ještě před započetím sonifikace. Poté se zkumavka vloží do ultrazvukové lázně a sonifikuje cca 30 minut s průběžným protřepáváním pomocí vortexu či třepačky. Po sonifikaci se vyočkuje předem stanovený objem bujónu na krevní agar (ideálně 0,01 objemu bujónu ve zkumavce). Pro lepší a rychlejší identifikaci narostlých mikrobů je možno bujón vyočkovat i na selektivní či selektivně-diagnostické půdy (Sabouroudův agar, krevní agar s přídavkem NaCl, Endův agar). Kultivují se inokulované misky i sonifikovaný bujón s katétrem. Po 24hodinové kultivaci při 37 C se hodnotí množství narostlých kolonií dle tab. 1. Počet kolonií vyrostlých na agaru
22 odpovídá počtu mikrobů v 0,01 původního objemu bujónu, počet mikrobů uvolněných z katétru je tedy stokrát větší. Tabulka 1 interpretační kritéria pro kvantifikaci mikrobů na katétru Počet kolonií na KA Zákal Hodnocení 0 kolonií Ne STERILNÍ 0 kolonií Ano < 100 CFU 1 10 kolonií Ano tis. CFU kolonií Ano 1 tis. 10 tis.cfu > 100 kolonií Ano > 10 tis. CFU Legenda: Počet kolonií na KA - počet kolonií (CFU) narostlých na krevním agaru; Zákal přítomnost či nepřítomnost zákalu v bujónu; CFU colony-forming units V případě, že bujón s katétrem vykazuje známky nárůstu mikroorganizmů (zákal), ale na miskách nenarostou bakterie, hodnotí se množství jako < 100 CFU a bujón se vyočkuje, aby bylo možno zjistit druh mikroba. Takové množství mikroba se u saprofytických druhů bakterií (např. Staphylococcus epidermidis) považuje za kontaminaci, u obligátních patogenů, jako jsou Staphylococcus aureus či Streptococcus pyogenes je hodnocení nálezu jiné
23 2.4 REZISTENCE BUNĚK V BIOFILMU K ANTIMIKROBIÁLNÍM LÁTKÁM Problematice biofilmové rezistence k antimikrobiálním látkám se podrobně věnuje jedna z předkládaných souhrnných prací, proto pro podrobnější informace k této problematice odkazuji na přílohu č. 2 této dizertační práce. Mikrobiální buňky žijící ve formě biofilmu jsou mnohonásobně rezistentnější k antimikrobiálním látkám, než planktonické buňky stejného kmene (Stewart et al., 2001; Donlan et al., 2002). Zvýšená rezistence biofilmů k biocidům byla prokázána v mnoha studiích in vitro a je významnou komplikací u infekcí implantátů a cizích těles in vivo. Mnohé studie in vitro dokázaly, že účinnost biocidů je u biofilmových buněk oproti buňkám planktonickým nižší o jeden až čtyři řády (Costerton, 1999; Holá et al., 2004b; Ceri et al., 1999 a další). Biofilmová rezistence se projevuje několika základními mechanizmy, které se liší v závislosti na druhu biofilmpozitivního mikroba a aplikované antimikrobiální látky. Rozdíl je též v tom, jaké mechanizmy se projevují u biofilmu v počáteční fázi růstu a jaké u plně vyzrálého a starého biofilmu. Uvádí se řada vysvětlení vyšší rezistence biofilmových forem mikrobů oproti formám planktonickým. Mezi nejvýznamnější a nejvíce prostudované mechanizmy biofilmové rezistence patří: fyzikální a chemické bariéry bránící penetraci látek do biofilmové vrstvy snížená rychlost růstu buněk v biofilmu aktivace univerzální stresové odpovědi (general stress response) vznik určité rezistentní skupiny buněk, tzv. perzistorů vznik biofilm-specifického fenotypu a exprese pravděpodobných genů rezistence. Všechny tyto mechanizmy se v biofilmu spíše vzájemně doplňují, než působí jednotlivě. Ve vícedruhovém biofilmu je pak problematika biofilmové rezistence ještě mnohem složitější Snížený průnik látek do biofilmové vrstvy Biofilmová vrstva může jako penetrační bariéra působit dvěma způsoby jako určité síto, nepropouštějící větší molekuly, nebo jako donor volných vazebných míst. Mechanizmus síta mikrobiálním biofilmům poskytuje též účinnou ochranu proti velkým molekulám imunitního systému hostitele, jako jsou např. antimikrobiální proteiny lysozymy a
24 komplement (Bjarnsholt et al., 2005; Donlan et al., 2002; Leid et al., 2005). Poměrně silně hydratované polyanionické polysacharidy a glykoproteiny biofilmové vrstvy působí jako výměníky iontů a adsorbují kationty antimikrobiálních látek (Wagman et al., 1975). Ve většině případů však polysacharidová bariéra postup antimikrobiálních látek pouze zpomalí. Příkladem látek s takto zpomaleným průnikem, které sice pronikají biofilmem, ale s určitou časovou prodlevou, mohou být fluorované chinolony (Ishida et al., 1998; Anderl et al., 2000; Shigeta et al., 1997; Vrany et al., 1997). I tato časová prodleva však může zvýšit efektivitu obranných mechanizmů bakteriální buňky, např. enzymatické degradace pronikající antimikrobiální látky Pomalý růst biofilmu a heterogenita mikrobiální populace Při nasycení určitého prostředí bakteriemi, nebo při úbytku živin dochází v bakteriální populaci ke zpomalení růstu. Zpomalení rychlosti růstu buněk je obecně provázeno zvýšením jejich rezistence k těm antibiotikům, která působí na rostoucí bakterie (Tuomanen et al., 1986; Cozens et al., 1986). Některé studie prokázaly, že růstová rychlost mikrobů, např. S. epidermidis, v biofilmu silně ovlivňuje jeho citlivost čím vyšší je rychlost buněčného růstu, tím rychleji jsou buňky inaktivovány ciprofloxacinem (Duguid et al., 1992) i rifampicinem (Zheng et al., 2002). Ve vyzrálých biofilmech je možné pozorovat komplexní trojrozměrnou strukturu s mnoha odlišnými typy mikroprostředí, která se liší např. rozdílnou osmolaritou, přísunem živin či buněčnou hustotou (Fux et al., 2005). Gradienty živin, různé odpadní produkty a četné signalizační faktory, to vše přispívá k této heterogenitě uvnitř biofilmu. Proto se nutně nabízí předpoklad, že mikrobiální populace v biofilmu je heterogenní a že jednotlivé buňky či jejich populace žijí za trochu odlišných podmínek než zbývající buňky ve stejném biofilmu, a tedy i rostou jinou rychlostí Vznik tzv. perzistorů V roce 1944 publikoval Bigger v The Lancet článek o svých výzkumech na penicilinu (Schierholz et al., 2001). Jedním z publikovaných poznatků bylo, že při expozici stafylokokové kultury penicilinu nedošlo k úplné eradikaci bakteriální kultury, ale přežila malá část buněk, které Bigger vhodně nazval perzistoři. Bigger odhadl počet perzistorů v bakteriální populaci na 10-6 a prokázal též, že kultivací a množením těchto perzistorů vzniká populace nerozeznatelná od původní populace podobně citlivá k inhibici růstu penicilinem
25 se stejným procentem nových perzistorů. Keren et al. (2004) zopakovali na základě Biggerova článku pokusy s E. coli a účinkem ampicilinu a došli ke stejným výsledkům přežívající perzistoři vytvoří znovu populaci buněk se stejnou citlivostí k ampicilinu jako byla původní populace a jsou tedy pouze fenotypovou variantou původního kmene. Při studiu biofilmových buněk a planktonických buněk ve stacionární fázi růstu prokázali Spoering et al. (2001), že biofilmové buňky a planktonické buňky stacionární fáze mají podobnou rezistenci k antibiotikům a že tato rezistence je u obou studovaných skupin závislá zejména na přítomnosti perzistorů. I další studie prokázaly přítomnost perzistorů v biofilmu, např. Brooun et al. (2000) či Drenkard et al. (2002) Univerzální odpověď na stres Bakterie odpovídají na nedostatek živin a na jiné environmentální stresy syntézou σ faktorů. Tyto σ faktory regulují transkripci genů, jejichž produkty zmírňují účinky stresu. Syntézou produktů těchto genů tedy bakterie odpovídají na působení stresových faktorů a tím se u nich aktivuje mechanizmus univerzální odpovědi na působení stresových faktorů. Ten je založen např. na tom, že mění permeabilitu buněčné membrány a brání tak dalšímu působení těchto faktorů na buňku (Liu et al., 2000) Vznik biofilmového fenotypu Již před patnácti lety se objevila v lékařské mikrobiologii teorie, že primární adheze bakterií k povrchu může vést k indukci či represi genů, které mohou dále ovlivnit velké množství fyziologických odpovědí (Otto et al., 2002). DNA microarray analýzy ukázaly, že genová exprese se mezi biofilmovými a planktonickými buňkami u Bacillus subtilis liší o 6 %, a u P. aeruginosa o 1 % (Stanley et al., 2003; Whiteley et al., 2001). U S. epidermidis se odlišná genová exprese projevuje mj. expresí sara (staphylococcal accessory regulator) a icaadbc operonu (Beenken et al., 2004; Cramton et al., 1999). Indukce biofilm-specifického genotypu minimálně u subpopulace biofilmových buněk může vést k exprimaci aktivních mechanizmů potlačujících účinky antimikrobiálních látek (Gilbert et al., 1997; Maira-Litran et al., 2000a; Maira-Litran et al., 2000b; Cochran et al., 2000)
26 2.4.6 Další mechanizmy uplatňující se v biofilmové rezistenci Quorum sensing Role systému quorum sensing v biofilmové rezistenci k antimikrobiálním látkám není dosud zcela jasná. Předpoklad, že systém quorum sensing hraje v biofilmové rezistenci svou roli, podporuje fakt, že rezistentní biofilmové buňky mohou svou citlivost k antimikrobiálním látkám velmi rychle znovu nabýt, jsou-li přeneseny do čerstvého média, které nejen poskytuje živiny (a tím umožňuje rychlejší růst), ale snižuje také koncentraci ochranných mezibuněčných signalizačních molekul (Anderl et al., 2003). Stáří a vyzrálost biofilmové vrstvy Svou roli hraje také stáří a zralost biofilmové vrstvy. Monzon et al. (2002) ve své studii prokázali, že rezistence S. epidermidis k vankomycinu roste s jeho stářím (studovali biofilm 6 48 hodin starý). Tomu odpovídají i klinické zkušenosti, kdy v ortopedii odstranění infikovaného ložiska s ponecháním protézy je úspěšnější v případě časných infekcí, než v případě pozdních pooperačních infekcí, které měly čas se určitou dobu vyvíjet (Patel, 2005). Těsný kontakt buněk a horizontální přenos genetické informace V biofilmu se díky užšímu kontaktu buněk mohou snáze šířit plazmidy a extrachromozomální úseky DNA. Právě plazmidy a transpozony kódují rezistenci k řadě antimikrobiálních látek. Reisner et al. (2006) prokázali přenos plazmidů konjugací mezi různými gramnegativními bakteriemi i v biofilmu. Četnosti tohoto horizontálního přenosu genetické informace byly několikařádově vyšší právě v biofilmové kultuře než v planktonické kultuře stejného mikroba. Ke stejným výsledkům došli i ostatní autoři (Roberts et al., 1999; Hausner et al., 1999; Christensen et al., 1998). Buněčné mechanizmy rezistence Buněčné mechanizmy bakteriální rezistence nejsou sice specifikem biofilmpozitivních bakterií, ale i tyto mechanizmy rezistence biofilmpozitivní mikroby využívají. Fenotypová adaptace je určité přizpůsobení se bakterie na změněné metabolické pochody. Genetická modifikace bývá podmíněna buď chromozomálně, tj. modifikací genu na chromozomu, nebo převzetím genetického materiálu od rezistentních buněk, např. pomocí plazmidů, a její mechanizmy můžeme rozdělit na změnu cílové molekuly, impermeabilitu buněčné stěny a použití aktivních pump, intrinsic rezistenci či enzymatickou inaktivaci
27 3. Publikace zařazené do dizertační práce * Teoretické práce shrnující současné poznatky o bakteriálních biofilmech na základě našich prací i na základě prací dalších autorů: 1) Quantification of biofilm in microtiter-plate: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci; S. Stepanović, D. Vuković, V. Holá, G. Di Bonaventura, S. Djukić, I. Ćirković, F. Růžička; Acta Pathologica, Microbiologica et Immunologica Scandinavica (accepted for publication). 2) Biofilmy a antimikrobiální látky; Holá V., Toršová V., Gallo J.; in Mikrobiální biofilmy; M. Rulík, V. Holá, F. Růžička, M. Votava (eds.); (in press; předpokládané datum vydání jaro 2007). 3) Biofilm dutiny ústní; Holá V., Kaprálová S.; in Mikrobiální biofilmy; M. Rulík, V. Holá, F. Růžička, M. Votava (eds.); (in press; předpokládané datum vydání jaro 2007). * Studium růstových podmínek bakteriálních biofilmů a jejich dynamiky: 4) Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis; Holá V., Růžička F., Votava M., 2004; Microbiologica; vol. 27; 3: ) Dynamic of Staphylococcus epidermidis biofilm formation in relation to nutrition, temperature and time; Holá V., Růžička F., Votava M., 2006; Scripta Medica (Brno) 79; 3: * Studium klinického významu biofilmpozitivních bakterií: 6) Biofilm detection and the clinical significance of Staphylococcus epidermidis isolates; Růžička F., Holá V., Votava M., Tejkalová R., Horváth R., Heroldová M., Woznicová V., 2004; Folia Microbiologica; 49; 5: ) Rozdíly v citlivosti k antibiotikům u biofilmpozitivních a biofilmnegativních kmenů Staphylococcus epidermidis izolovaných z hemokultur; Holá V., Růžička F., Votava M. 2004; Epidemiologie, mikrobiologie, imunologie; 53; 2: ) Stanovení citlivosti k antibiotikům u biofilmpozitivních forem mikroorganizmů; Holá V., Růžička F., Tejkalová R., Votava M., 2004; Klinická mikrobiologie a infekční lékařství; 10; 5: ) Selective pressure of higher sanitation standards increases relative number of biofilmpositive microbes on the intensive care unit; V. Holá, L. Horáková, F. Růžička, M. Trnková, M. Votava; Infection Control and Hospital Epidemiology (sent for publication)
28 4. Komentáře k předkládaným pracím 4.1 Quantification of biofilm in microtiter-plate: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci Práce je souhrnným přehledem zkušeností několika pracovišť (Institute of Microbiology and Immunology, School of Medicine, Belgrade, Serbia; Mikrobiologický ústav Lékařské fakulty MU a Fakultní nemocnice u sv. Anny v Brně; Laboratory of Clinical Microbiology, Aging Research Center Ce.S.I., G. D Annunzio University of Chieti-Pescara, Italy) s kvantitativním hodnocením produkce biofilmu u stafylokoků a zároveň rozsáhlou analýzou metod hodnocení kvantity biofilmového nárůstu pravděpodobně nejčastěji používanou metodou, tedy Christensenovou metodou s mikrotitračními destičkami, které jsou dosud publikovány v odborné literatuře. Cílem této souhrnné práce je shrnout zkušenosti a poskytnout podrobný popis všech kroků při kvantifikaci nárůstu biofilmu v mikrotitračních destičkách. Tyto informace totiž v dostupné literatuře často chybí, nebo jsou podány vágně a nekonkrétně, případně si zcela odporují, což ztěžuje porovnatelnost a interpretovatelnost získaných výsledků mezi jednotlivými laboratořemi a výzkumnými centry. V práci jsme se zároveň upozornili na nejkritičtější místa a body celého postupu hodnocení tvorby biofilmu. Uchování bakteriálních kmenů pro testování biofilmu, inokulum Uchování mikrobiálních kmenů je nezbytnou součástí výzkumu. Bakterie mohou být uchovány po dobu 1 3 let při teplotě -20 C a po dobu 1 10 let při teplotě -70 C. V tekutém dusíku je možné uchovávat zamražené bakterie až 30 let (de Paoli, 2005). Ačkoliv uchovávané bakterie obvykle nepozbývají svých faktorů virulence, bylo prokázáno, že uchování a rozmražení může ovlivnit jejich životaschopnost a stabilitu jejich antigenních, molekulárních a biochemických vlastností (de Paoli, 2005). Proto může po rozmražení čistá kultura stafylokoků obsahovat směs fenotypů, které se liší rovněž v jejich schopnosti tvorby biofilmu (tzv. fázové varianty). Fázové varianty jsou geneticky identické buňky s různým fenotypovým projevem. Tyto fázové varianty se mohou rovněž lišit ve schopnosti tvorby biofilmu (Ziebuhr et al., 1999; Ziebuhr et al., 2001; Handke et al., 2004 a další). Na základě kultivace kmene na agaru s kongo červení lze z takto vzniklé kultury odlišit fázové varianty, oddělit je a podrobit dalšímu testování (Deighton et al., 2001)
29 Stanovení kvantity biofilmu je závislé nejen na hustotě použitého inokula (Deighton et al., 2001; Stepanovic et al., 2003), ale i na tom, zda bylo inokulum připraveno z pevného či tekutého média. Bylo prokázáno, že stafylokoky pěstované na pevném či v tekutém médiu se od sebe liší svými povrchovými vlastnostmi (Cheung et al., 1988; Kiers et al., 2001). Média a mikrotitrační destičky pro kultivaci biofilmu Složení média je velmi významným faktorem, který ovlivňuje schopnost tvorby biofilmu u bakterií in vitro (Rohde et al., 2001; Holá et al., 2004c; Kennedy et al., 2004). V mnoha studiích bylo prokázáno, že pro interpretaci výsledků u slabě biofilmpozitivních kmenů je mozkosrdcová infuze (BHI) lepší, než tryptono-sojový bujón (TSB) (Knobloch et al., 2002a; Holá et al., 2004c; Stepanovic et al., 2003; Mathur et al., 2006). TSB je však médium, které je při kultivaci stafylokokových biofilmů užíváno stejně často (Deighton et al., 1990; Stepanovic et al., 2000; Cramton et al., 2001 a další). Suplementace média glukózou zvyšuje schopnost stafylokoků tvořit biofilm (Christensen et al., 1985; Mulder et al., 1998), proto se často používá TSB či BHI suplementované 0.25% - 4% glukózy (Gelosia et al., 2001; Fitzpatrick et al., 2005 a další). Pro kultivaci biofilmu by měly být používány sterilní polystyrenové mikrotitrační destičky pro tkáňové kultury. Některé kmeny tvoří biofilm pouze na destičkách pro tkáňové kultury a mnoho kmenů vykazuje nižší tvorbu biofilmu na destičkách neupravených pro tkáňové kultury (Christensen et al., 1985; Kennedy et al., 2004). Smývání a odlupování biofilmové vrstvy je v mikrotitračních destičkách pro tkáňové kultury též nižší. Kultivace biofilmu, promývání, fixace Každý kmen by měl být kultivován minimálně ve třech jamkách. U negativní kontroly je vhodná kultivace ve více jamkách, obvykle v šesti. Vzhledem k tomu, že síla biofilmové vrstvy je závislá na inkubačním čase, je doba inkubace důležitá (Deighton et al., 2001; Holá et al., 2006). Doba inkubace je nejčastěji 24 hodin při teplotě 37 C (Deighton et al., 2001; Mack et al., 2001), někteří autoři inkubují mikrotitrační destičku přes noc (Allignet et al., 1999; Cramton et al., 2001; Fowler, Jr. et al., 2001) či 18 hodin (Fox et al., 2005; Tormo et al., 2005; Arciola et al., 2006), někteří hodin, a někteří autoři používají prodlouženou kultivaci 48 hodin (Deighton et al., 2001; Ozerdem et al., 2003). Domníváme se, že dostatečná doba kultivace je 18 hodin. Prodloužená inkubace může vést dokonce k nižším hodnotám optické denzity (Holá et al., 2006). Další velmi důležitý krok je promývání biofilmu, při kterém se z jamky odstraňují všechny neadherované buňky. Zároveň je třeba zachovat integritu vykultivované biofilmové
30 vrstvy a nerozrušit ji příliš razantním promýváním. Nedokonalé promytí může vést k falešně pozitivním výsledkům díky ulpívajícím planktonickým buňkám, které budou následně fixovány a obarveny, zatímco příliš hrubé a intenzívní promytí může vést k přílišnému poškození biofilmové vrstvy a tedy naopak k falešně negativním výsledkům. Proto je potřeba věnovat pozornost nejen vlastnímu provedení promytí, ale i počtu jeho opakování. Na rozdíl od různých autorů (Cramton et al., 2001; Deighton et al., 2001; Heilmann et al., 1996) se domníváme, že dvojnásobné promývání není dostatečně účinné. Lze však doporučit troj- (Perez-Giraldo et al., 1995; Vasudevan et al., 2003) či čtyřnásobné promytí (Mulder et al., 1998; Allignet et al., 1999; Arciola et al., 2006). Adherované bakterie by měly být fixovány teplým vzduchem. Lze též využít fixace alkoholem (po dobu 20 minut, poté nechat mikrotitrační destičku uschnout), která je téměř stejně účinná, jako teplý vzduch (Baldassarri et al., 1993). Barvení, měření a interpretace výsledků, kontrola kvality K barvení vrstvy bakterií adherovaných ke dnu a k bokům jamek mikrotitrační destičky se používá nejčastěji krystalová violeť. Přestože krystalová violeť barví pouze stafylokokové buňky a ne extracelulární matrix biofilmu, lze využít jejího barvícího účinku, protože všechny neadherované (tedy planktonické) buňky byly v předchozím kroku promýváním odstraněny. Krystalovou violetí jsou tedy obarvené pouze buňky, které jsou ve vrstvě adherovaného biofilmu. Optická denzita každé jamky by měla být měřena spektrofotometricky při 570 nm. Problém jednobodového měření OD většiny spektrofotometrů lze vyřešit rozpuštěním navázaného barviva do ethanolu, což nám umožňuje sice nepřímé, ale přesné měření síly tvorby biofilmu daného kmene. Na základě stanovené mezní hodnoty, tzv. cut off value můžeme interperetovat získané výsledky. Tato mezní hodnota nám oddělí na základě získaných OD biofilmpozitivní kmeny od kmenů biofilmnegativních. Některými autory je mezní hodnota počítána jako součet OD důlku se sterilním médiem a trojnásobku jeho standardní odchylky (Christensen et al., 1985), součet OD důlku s biofilmnegativním kmenem a trojnásobku jeho standardní odchylky (Mulder et al., 1998; Fox et al., 2005), další používají předem definované hodnoty, např. 0,12 (Deighton et al., 2001; Gelosia et al., 2001; Ziebuhr et al., 2001), 0,1 (Mack et al., 2001; Dobinsky et al., 2003), či 0,2 (Moretro et al., 2003). Někteří autoři též použili ke stanovení hodnoty cut off biofilmpozitivní kontrolní kmen (Allignet et al., 1999) a někteří využili ke stanovení hodnoy cut off dvojnásobku OD biofilmnegativního kmene (Fowler, Jr. et al.,
31 2001). Podle našich zkušeností hodnotě cut off odpovídá hodnota součtu průměru OD důlků se sterilním médiem a trojnásobku jejich standardní odchylky. Pro kontrolu postupů doporučujeme 4 biofilmpozitivní kmeny S. epidermidis ze sbírky ATCC (ATCC 35981, ATCC 35982, ATCC a ATCC 35984). Nejčastěji je používán kmen S. epidermidis ATCC 35984, silně biofilmpozitivní kmen (Christensen et al., 1985). V dalších sbírkách mikroorganizmů jsou další biofilmpozitivní kmeny, např. S. epidermidis CCM 7221, uložený v České sbírce mikroorganizmů, který je rovněž silně biofilmpozitivní (Růžička et al., 2004). Dle našich zkušeností lze tuto metodiku s případnými drobnými modifikacemi použít i na další druhy mikrobů (testovány byly kvasinky rodu Candida, enterokoky, gramnegativní nefermentující bakterie, zejm. rodů Pseudomonas, Acinetobacter, Stenotrophomonas a Burkholderia aj.)
32 4.2 Biofilmy a antimikrobiální látky Předkládaná práce je kapitolou z monografie Mikrobiální biofilmy. Kniha Mikrobiální biofilmy je souhrnnou prací předkládající problematiku mikrobiálních biofilmů v přírodním i humánním prostředí české odborné veřejnosti. Kapitola Biofilmy a antimikrobiální látky předkládá současný pohled na problematiku biofilmové rezistence k antibiotikům. * * * Schopnost mikrobiálních buněk v biofilmu přežívat při několikařádově vyšších koncentracích antimikrobiálních látek než přežívají planktonické buňky stejného kmene mikroba byla v této práci zmíněna již několikrát a je obecně uznávaným faktem. Bakterie v biofilmové vrstvě jsou mnoha různými mechanizmy chráněny před účinky antibiotik a imunitního systému, a proto snadno přežívají konvenční antibiotickou terapii. Antibiotika podávaná v koncentracích MIC totiž likvidují jen planktonicky žijící část bakteriální populace a bakterie rostoucí v biofilmové vrstvě zůstávají životaschopné. In vivo mohou tedy takto administrovaná antibiotika potlačit symptomy infekce likvidací volně plovoucích bakterií uvolněných z přisedlé populace, ale selhávají při likvidaci bakteriálních buněk dosud usazených v matrix biofilmu. Po ukončení antimikrobiální léčby může biofilm sloužit jako ložisko a vyvolat tak recidivu infekce. Infekce spojené s biofilmem obvykle přetrvávají, dokud není kolonizovaný povrch chirurgicky odstraněn (Stewart et al., 2001). Minimální inhibiční koncentrace (MIC) a minimální baktericidní koncentrace (MBC) antibiotik se výrazně liší od koncentrací antibiotik, které jsou schopné účinně zasáhnout buňky uvnitř biofilmu, tzv. MBEC, tedy minimální biofilm eradikující koncentrace, nebo alespoň MBIC (minimální biofilm inhibiční koncentrace). Minimální biofilm eradikující koncentrace je taková koncentrace antimikrobiálních látek, která zcela likviduje buňky v biofilmu a minimální biofilm inhibující koncentrace je taková koncentrace antimikrobiálních látek, která zastavuje jejich růst a množení. Problémem antibiotické terapie biofilmových infekcí je, že hodnoty MBEC i MBIC výrazně přesahují tzv. break-pointy, tedy maximální koncentrace antibiotik dosažitelné v séru pacienta a tyto hodnoty tedy nelze terapeuticky využít. Slibné se zdají kombinace některých antibiotik s dalšími látkami, např. lysostaphinem (Walencka et al., 2005; Walencka et al., 2006), či kombinace antibiotik s dalšími postupy, např. působením ultrazvuku (Ensing et al., 2006; Ensing et al., 2005), fotodynamické postupy (kombinace fotosenzitivního účinku polyethyleniminu a chlorinu a působení laserové diody o vlnové délce 660nm) (Garcez et al.,
33 2006) a také nová antibiotika, např. quinpristin-dalfopristin (Trampuz et al., 2006; Halliman et al., 2004). Stále tedy platí, že v současné době mohou být biofilmové infekce cizích těles a implantátů léčeny pouze vyjmutím infikovaného implantátu. Takový zákrok však znamená nejen vyšší finanční náklady na léčbu, ale i vyšší zátěž pro pacienta samotného. Kromě toho jsou zákroky spojené s odstraněním těchto materiálů, ať již srdečních chlopní, kloubních náhrad či tunelizovaných katétrů, spojeny s vyšším rizikem řady dalších komplikací (Kuhn et al., 2002), proto se u pacientů s kritickým stavem využívá udržovací terapie, kdy administrace konvenčních dávek antibiotické terapie potlačuje symptomy infekce způsobené planktonickými buňkami mikroba a s vyjmutím či výměnou katétru se čeká do té doby, dokud není pacientův stav alespoň natolik stabilizován, aby výměna implantátu nebyla zbytečným rizikem
34 4.3 Biofilm dutiny ústní Předkládaná práce je kapitolou z monografie Mikrobiální biofilmy. Kniha Mikrobiální biofilmy je souhrnnou prací předkládající problematiku mikrobiálních biofilmů v přírodním i humánním prostředí české odborné veřejnosti. Kapitola Biofilm dutiny ústní předkládá současný pohled na problematiku biofilmu v dutině ústní, tedy zejména zubního plaku z mikrobiologického i stomatologického hlediska. * * * Stomatologie je vůbec první oblastí humánní mikrobiologie, kde lékařští mikrobiologové přijali možnost, že zubní plak, neboli biofilm, je druhým přirozeným způsobem života bakterií a že by bakterie a bakteriální společenstva mohla žít přisedle a tvořit určité nánosy (plaky, biofilmy), a že tedy bakterie nemusí žít a existovat jen ve formě volně plovoucích, tedy planktonických buněk. Dutina ústní je z hlediska mikrobiální kolonizace a růstu ideálním prostředím. Svými vlastnostmi totiž bakteriím zajišťuje ideální teplotu a dostatečnou vlhkost. Přesto však zdaleka ne všechny mikroorganizmy, které se dostanou do dutiny ústní, ji jsou schopny kolonizovat. Dutina ústní je díky svým vlastnostem ekologicky odlišná od všech dalších povrchů těla, čímž se výběr mikrobů schopných kolonizace a perzistence omezuje na ty, které jsou vybaveny potřebnými faktory virulence. V rámci dutiny ústní existuje navíc i další stratifikace a rozrůzněnost prostředí, z nichž každé bude svými biologickými vlastnostmi podporovat růst určitých mikrobů a určité mikrobiální komunity. V dutině ústní se nalézá mnoho biotopů s výrazně odlišnými podmínkami pro růst mikrobů rty, bukální povrchy a měkké patro, supragingivální hladký povrch zubů, dásňové žlábky a subgingivální povrch zubů, povrch jazyka apod. Široké množství těchto biotopů umožňuje koexistenci mnoha bakteriálních druhů. Udává se, že z dutiny ústní lze vykultivovat více než 400 různých druhů bakterií a další stovky nekultivovatelných bakterií lze rozlišit genetickými metodami (Marsh, 2004). Mikrobiální biofilm se v dutině ústní tvoří na měkkých i tvrdých tkáních. Složení biofilmu se na těchto místech výrazně liší, a to jak mikrobiálním zastoupením, tak složením extracelulární substance. Zcela jasně nám pro mikrobiální kolonizaci budou poskytovat odlišná prostředí povrchy sliznic, např. rty, vnitřní strana tváří (bukální sliznice), měkké patro či jazyk a povrchy zubů s částmi subgingiválními a supragingiválními, zde pak s plochami koronárními a okluzálními s fisurami. Vlastnosti některých z těchto prostředí se u jedince mění s věkem. Ekologické podmínky v dutině ústní se též mění při výměně prvního za trvalý
35 chrup, při extrakci zubů, při následném používání dentálních protéz a pomůcek a samozřejmě i při každém zubním zákroku (Marsh, 2006). Dočasné fluktuace ve stabilitě orálního ekosystému mohou být indukovány též frekvencí a typem požívané stravy, změnami v toku slin či případnou antibiotickou léčbou. Zubní plak je tvořen z 80 % z vody a 20 % ze sušiny, z toho asi 1/3 tvoří mikrobiální buňky a 2/3 extracelulární substance. Extracelulární substance je tvořena zejména organickými látkami, polysacharidy a proteiny, ale i anorganickými látkami, zejména sloučeninami vápníku a fosfáty. Tato extracelulární substance představuje nárazníkový systém plaku. Základní mechanizmy, uplatňující se při tvorbě zubního biofilmu, jsou stejné jako mechanizmy tvorby biofilmů jednodruhových, které se uplatňují v jiných, převážně sterilních částech lidského těla (infekce krevního řečiště, močového traktu, kontaktních očních čoček apod.). Stejně jako tam, i zde dochází k úpravě povrchu a vzniku pelikuly, dále pak nasedání primárních kolonizátorů, koadhezi a koagregaci dalších kolonizátorů sekundárních a terciárních. U jednodruhových biofilmů ke koadhezi a koagregaci dochází mezi jednotlivými kmeny stejného druhu, v dutině ústní dochází k interakcím mezidruhovým. V dutině ústní dochází později ke vzniku zubního kamene. Pelikula je vrstva proteinů navázaných na povrch zubu. Vazbami mezi jednotlivými sloučeninami v pelikule může dojít ke změně jejich konformace, což může mít za následek ovlivnění vazebných míst pro mikroorganizmy. Tímto způsobem se mění např. molekulární struktura glukanů, naváží-li se na povrch zubu zároveň i glukosyltransferázy (Kopec et al., 2002). Složení pelikuly tedy výsledně ovlivňuje druhovou strukturu primárních kolonizátorů (Marsh, 2004). Méně než v jiných biofilmech se u bakterií biofilmu dutiny ústní projevují po adhezi na povrch zubů změny v expresi či represi různých genů. Je to dáno tím, že mikroflóra dutiny ústní je, na rozdíl od volně žijících planktonických buněk jiných mikrobů, na schopnosti tvorby biofilmu a na životě v biofilmové vrstvě existenčně závislá (Burne et al., 1999; Svensater et al., 2001). Přesto se některé geny ve větší míře v biofilmové vrstvě uplatňují, jde například o geny spojené se syntézou glukanů a fruktanů, jak bylo prokázáno u Streptococcus mutans (Li et al., 2001). V zubním biofilmu vznikají díky typickému a v lidském těle unikátnímu složení typické struktury, které jsou v jiných biofilmech neobvyklé, jako jsou tzv. kukuřičné klasy a rozety (Kolenbrander et al., 2000) (viz obr. 9)
36 Zubní plak funguje jako skutečné mikrobiální společenstvo, ve kterém interakce přítomných druhů vedou k výrazně vyšší metabolické účinnosti, než by byla sumární metabolická účinnost daná součtem metabolické účinnosti jednotlivých přítomných druhů. Mikrobiální biofilm zubního plaku toho dosahuje právě díky své výrazné druhové rozrůzněnosti. Obr. 9 Struktura kukuřičných klasů typická pro biofilm dutiny ústní (převzato z Kolebrander et al., 2000)
. Pro testování těchto koncentrací je nutné vypěstovat dostatečně homogenní biofilm o definované kvantitě.")
37 4.4 Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis Metodika stanovení citlivosti biofilmpozitivních mikrobů k antibiotikům, tedy stanovení MBEC a MBIC, nebyla dosud uspokojivě vyřešena (Costerton et al., 1999; Raad et al., 2007). Pro testování těchto koncentrací je nutné vypěstovat dostatečně homogenní biofilm o definované kvantitě. Postupy popisované v odborné literatuře jsou mnohdy časově a přístrojově náročné a často též zvyšují riziko kontaminace biofilmové kultury z vnějšího prostředí (Ceri et al., 1999; Ceri et al., 2001). Používané metody a přístroje (např. modiffied Robbins device) (Everaert et al., 1998; Al Fattani et al., 2006) rovněž nedovolují použití mikrodilučních metod, čímž narůstá finanční náročnost celého vyšetření a také časová náročnost testování citlivosti určitého kmene. Aby bylo možno napěstovat dostatečně homogenní biofilmovou vrstvu o definované kvantitě, bylo potřeba nejprve splnit základní podmínku vzniku biofilmu přítomnost vhodného adhezního povrchu. V této práci jsme se zaměřili na testování různých povrchových úprav polystyrenu a jejich vliv na adhezi stafylokoků. Výběr a testování plastů, testované kmeny Polystyren jsme k tomuto testování zvolili na základě předchozích studií provedených v rámci grantového projektu FRVŠ Stafylokokový biofilm a jeho rezistence vůči antibiotikům. V rámci zmíněného projektu jsme mj. testovali různé materiály a hodnotili je z hlediska adherence a homogenity narostlého biofilmu. Testované materiály jsou spolu se sumárními výsledky tohoto testování uvedeny v tabulce 2. Homogenita a kvantita narostlého biofilmu byla hodnocena pomocí tříčlenné stupnice (2 silný a homogenní nárůst biofilmu; 1 slabý či nehomogenní nárůst biofilmu; 0 žádný nárůst biofilmu). Protože žádný z materiálů neodpovídal ideálně našim představám o homogenním a silném nárůstu biofilmu, přistoupili jsme k povrchové úpravě. K té jsme, vzhledem k výhodným vlastnostem (cenová dostupnost, snadné čištění, tvarová stálost, čirost a nebarevnost materiálu), vybrali jeden z typů polystyrenu krasten 171. Pro účely dalších studií byl krasten 171 používán ve tvaru destiček, které jsou protitvarem klasické mikrotitrační destičky, Obr. 10 destička s hroty pro kultivaci biofilmu, VH
38 tj. víčkem destičky s kolíky, jež se volně zasouvají, do jamek mikrotitrační destičky. Svým tvarem umožňují tyto destičky kultivaci biofilmu na všech 96 kolících zároveň, čímž zrychlují a zjednodušují práci s biofilmy (viz obr. 10). Do studie bylo zařazeno 40 kmenů S. epidermidis izolovaných z hemokultur, 20 biofilmpozitivních a 20 biofilmnegativních, a to fenotypovými (Christensenova zkumavková metoda, růst na agaru s kongo červení) i genetickými metodami (průkaz ica operonu). Povrchová úprava polystyrenu etanolem Abychom vyloučili možnost, že biofilm na destičky s hroty neadheruje kvůli povrchovým nečistotám a olejům z výrobního procesu, očistili jsme destičky ethanolem. Hroty destiček byly ponořeny na pět minut do 75% denaturovaného etanolu. Poté byly hroty osušeny na vzduchu. Povrchová úprava sulfonací Ke zvýšení hydrofility povrchu byla použita sulfonace kyselinou sírovou. Sulfonace snižuje vazbou sulfonové skupiny styren hydrofobicitu povrchové vrstvy (Kučera et al., 1996), aniž by se měnila struktura polystyrenu (viz obr. 11). Hroty destiček byly sulfonovány ponořením na 30 minut do 96% H 2 SO 4, poté byly hroty pečlivě omyty sterilní redestilovanou vodou a osušeny. Obr Sulfonace polystyrenu kyselinou sírovou Povrchová úprava chemickou abrazí Hladký povrch polystyrenu lze narušit působením koncentrovaného acetonu. Působení koncentrovaného acetonu jemně narušuje strukturu povrchové vrstvy polystyrenu při zachování jejích fyzikálně-chemických vlastností. Dochází ke vzniku mikroskopických skulinek a dutinek. Hroty destiček byly na dobu 5 sekund ponořeny do koncentrovaného acetonu. Polystyrenový povrch byl poté omyt sterilní redestilovanou vodou a osušen
.")
39 Výsledky a shrnutí Homogenita a kvantita narostlého biofilmu byla opět hodnocena dle výše zmíněné tříčlenné stupnice (viz obr. 12). Data byla hodnocena generalizovanými lineárními modely statistického programu S-plus, verze 4.0 (Lucent Technologies Inc., 1988). K transformaci ordinálních dat byla využita funkce logit link a pro vyhodnocení testu byl použit Akaike index (AIC). Akaike index je index pro hodnocení relativní shody srovnávaných modelů, čím nižší je hodnota AIC, tím lépe odpovídají skutečná data hodnocenému modelu (Hsu et al., 1998). Všechny testy byly hodnoceny na hladině pravděpodobnosti p = 0,05. (a) (b) (c) Obr. 12 Nárůst biofilmu na hrotech destičky (a) hrot se silným a homogenním nárůstem; (b) hrot se slabším a nehomogenním nárůstem; (c) hrot bez nárůstu biofilmu, VH Při hodnocení plastových povrchů z hlediska jejich použitelnosti na kultivaci biofilmu byl kladen důraz na homogenitu a kvantitu nárůstu biofilmu. Biofilm se tvořil na všech materiálech kromě polyuretanu. Na mnoha materiálech se sice vytvořila biofilmová vrstva, ale ta se snadno oddělovala a smývala, což není pro studium biofilmu a jeho vlastností vhodné. Tabulka 2 Sumarizované výsledky testování plastových materiálů homogenita Krasten 171 (polystyren) ++ Krasten 127 (polystyren) ++ Krasten 336 (polystyren) ++ kvantita ++ / / / +++ Kuličky skleněné Korálky keramické Polyvinylchlorid High-Density Polyetylen Low-Density Polyetylen Polypropylen ++ Polyamid alkalický + + / Polytan 44/90 ShA (polyuretan) / +++ Polytan 44/80 ShA (polyuretan) 0 0 Polytan 44/65 ShA (polyuretan) 0 0 Silikon + + Polyetylen-tereftalát ++ Polymetylmetakrylát / ++ + / ++ Teflon + 0 / + Nerezová ocel nehodnotitelné nehodnotitelné Chrom-nikl 18/10 nehodnotitelné nehodnotitelné Legenda: ordinární škála pro hodnocení homogenity a kvantity nárůstu biofilmu
40 Nejsilnější nárůst biofilmu se tvořil na polystyrenu (všechny použité typy), polyvinylchloridu, sklu, HD-polyetylenu a polypropylenu. Výsledky testování materiálů jsou shrnuty v tabulce 2. Výsledky testování povrchové úpravy krastenu 171 jsou souhrnně znázorněny v grafech v příloze 7.4. V tabulce 3 je shrnuto statistické hodnocení povrchových úprav krastenu 171 generalizovanými lineárními modely. Lepšího nárůstu bylo dosaženo na polystyrenu sulfonovaném. Stejného výsledku bylo dosaženo v obou sledovaných médiích, BHI i TSB. Pro sledování vlastností biofilmu nelze doporučit pěstování biofilmu na polystyrenu bez povrchové úpravy. Tabulka 3 Výsledky statistického hodnocení povrchových úprav polystyrenu ICA ~ 1 Sum of Sq RSS AIC Nulový model Hydrofilizace poly-l-lysinem Mikrotitrační destičky pro tkáňové kultury Povrchová úprava etanolem Povrchová úprava chemickou abrazí Sulfonace kyselinou chromsírovou Sulfonace kyselinou sírovou Legenda: Sum of Sq součet čtverců; RSS součet residuálních čtverců; AIC Akaike index (Akaike information criterion) V době, kdy předkládaný článek byl již zařazen do tisku, jsme testovali též další povrchové úpravy tvrzeného polystyrenu, a to úpravu plazmovým výbojem ve vakuu a hydrofilizaci poly-l-lysinem. Plazmový výboj ve vakuu není bohužel pro výzkumné účely volně k dispozici a lze tedy využít pouze komerčně dodávaných polystyrenových pomůcek, které jsou tímto postupem upraveny. Úprava povrchu navázáním bílkovin (fibronektin) či polypeptidů ( poly-l-lysin, poly-l-glutamát apod.) se pro zvýšení adheze buněk používá ve tkáňových kulturách (McAteer et al., 2002). Využili jsme uvedené metodiky a destičky s hroty jsme na 120 minut vystavili působení poly-l-lysinu o koncentraci 20µg/ml při teplotě 37 C a poté osušili. Obě tyto metody, úprava plazmovým výbojem i hydrofilizace poly-llysinem, poskytují stejné výsledky jako sulfonace, jejich výhodou oproti sulfonaci je snadnější úprava povrchu polystyrenu (v případě poly-l-lysinu) a komerční dostupnost již hotového výrobku (v případě mikrotitračních destiček upravených plazmovým výbojem ve vakuu)
41 4.5 Dynamic of Staphylococcus epidermidis biofilm formation in relation to nutrition, temperature and time Znalost dynamiky tvorby biofilmu je nezbytná pro pochopení procesů probíhajících v biofilmové vrstvě. Je publikováno mnoho prací popisujících některou z vlastností biofilmu či biofilmpozitivních bakterií, ale autoři těchto prací se neshodují v podmínkách, jež jsou pro tvorbu biofilmu příhodné (Stepanovic et al., 2001; Deighton et al., 1990; Gelosia et al., 2001; Dunne, Jr. et al., 1993). V publikovaných pracích se autoři shodují v jednom, a to kultivační teplotě, 37 C. Ostatní podmínky, jako např. přítomnost živin a čas kultivace se u řady autorů liší. Cílem této práce bylo zjistit rozdíly ve tvorbě biofilmu za různých podmínek a stanovit minimální čas potřebný k vytvoření vyzrálé a homogenní struktury biofilmu. Studii jsme prováděli pouze s biofilmpozitivními kmeny druhu S. epidermidis (22 klinických izolátů a biofilmpozitivní kmen CCM 7221). Klinické izoláty byly izolovány z hemokultur, katétrů či chlopní pacientů s klinickými i laboratorními markery sepse. Biofilmpozitivita jednotlivých kmenů byla stanovena fenotypově Christensenovou zkumavkovou metodou (Christensen et al., 1982) i geneticky (průkazem ica operonu) dle Frebourga (2000). K vyloučení falešně negativních výsledků způsobených inhibicí PCR reakce byla zároveň amplifikována sekvence specifická pro S. epidermidis s primery SE705-1 a SE705-2 (Martineau et al., 1996) a výslednou délkou amplifikačního produktu 124 bp. Tato reakce zároveň sloužila k potvrzení správné determinace S. epidermidis. Studii jsme prováděli za aerobních podmínek v mozkosrdcové infuzi (BHI) v mikrotitračních destičkách pro tkáňové kultury (GAMA Group a.s., Trhové Sviny, CZ). Za stejných výchozích podmínek (hustota inokula, jeho zředění, rychlost zpracování apod.) jsme testovali tvorbu biofilmu při dvou odlišných teplotách, a to 25 C a 37 C. Při obou těchto teplotách jsme testovali různé koncentrace glukózy v BHI médiu (0 %, 4 % a 9 %). Abychom získali průměrné hodnoty OD, byly všechny kmeny kultivovány ve čtyřech jamkách. Každou celou hodinu byly určené jamky vyprázdněny, promyty pufrovaným fyziologickým roztokem (ph 7,4) a fixovány sušením (Baldassarri et al., 1993). Studie pokrývala prvních 48 hodin růstu biofilmové vrstvy. Získaná data byla statisticky vyhodnocena multifaktorovou analýzou variance (MANOVA). Výsledky jsou znázorněny v grafech 1 a 2 v příloze 7.5. Dynamika růstu biofilmu se liší jak z pohledu testovaných teplot, tak z pohledu testovaných živných médií (suplementací glukózou). Všechny testované kmeny rostly lépe při teplotě 37 C. Také nutričně bohatší
42 médium podporovalo růst biofilmu. Pro získání vyzrálé struktury biofilmu se zdá být dostačující kultivace hodin při teplotě 37 C. Model růstu biofilmové vrstvy testovaných kmenů S. epidermidis může být zjednodušené popsán následujícími charakteristikami: první známky bakteriální adheze byly viditelné po 2-4 hodinách kultivace, první homogenní, i když velmi slabá biofilmová vrstva byla patrná po pěti hodinách, po sedmi hodinách byla tato vrstva cca 3-krát tlustší. Po hodinách byla biofilmová vrstva pravděpodobně vyzrálá, změny v její tloušťce již po této době nebyly tak výrazné. Po kultivaci delší, než hodin se začaly odlupovat části biofilmové vrstvy, a to i při sebejemnějším promývání. Biofilmová vrstva v tomto kultivačním čase přestala být homogenní. Při teplotě 25 C bylo množství vytvořeného biofilmu cca 4krát nižší a rozdíly v množství biofilmu mezi jednotlivými živnými médii nebyly tak výrazné a při statistickém zpracování nesignifikantní (Multifaktorová ANOVA, program R: A Language and Environment for Statistical Computing 2.1.1; p=0.32) (R Development Core Team, 2005). Tvorba biofilmové vrstvy byla pomalá a růstová křivka téměř lineární. Suplementace BHI glukózou tvorbu biofilmu při této teplotě neovlivnila (p = 0.05). Při teplotě 37 C naopak suplementace média glukózou vytváří signifikantní rozdíly. Při této teplotě můžeme též pozorovat několik fází vzniku biofilmové vrstvy. Po adhezi jednotlivých buněk či jejich shluků nastává fáze exponenciálního růstu, která trvá cca 12 hodin. Potom můžeme pozorovat zpomalení bakteriálního růstu. V této době předpokládáme, že bylo dosaženo ekvilibria vyzrálého biofilmu. V rámci dynamické rovnováhy neroste, nebo roste pouze nevýrazně, množství biofilmu. Někteří autoři předpokládají, že v této době dochází v horní vrstvě biofilmu k oddělování jednotlivých buněk či jejich shluků a biofilmová vrstva uvolňuje planktonické buňky, které mohou kolonizovat další pevné povrchy (Costerton, 1999). Poté, co je prostředí nasyceno buňkami a živiny vyčerpány, biofilmová vrstva se začíná rozpadat. To může být způsobeno vznikem resting cells (Lewis, 2001) či strádáním způsobeným nedostatkem živin, sníženou perfuzí v biofilmové vrstvě, nízkým ph prostředí, poklesem parciálního tlaku kyslíku či akumulací toxických zplodin bakteriálního metabolizmu. Těmito změnami vysvětlujeme oddělování velkých částí biofilmové vrstvy od pevného povrchu pozorovatelné po 34 hodinách kultivace. Z výsledků naší práce vyplývá, že růstové optimum u stafylokoků není ovlivněno tvorbou biofilmu a schopnost tvorby biofilmu stafylokokům nenapomáhá v obraně před nepohodou způsobenou odlišnými kultivačními teplotami. Teplotní optimum pro tvorbu biofilmu je tedy 37 C, při nižších teplotách je množství biofilmu za stejný časový úsek nižší
43 Při této teplotě je biofilm vyzrálý po hodinách a při kultivaci delší než 34 hodin se biofilmová vrstva v jamkách mikrotitrační destičky začíná odlučovat. Proto by pro studium vlastností biofilmu, např. testování citlivosti vůči antibiotikům měla být dostatečná doba kultivace hodin v závislosti na suplementaci živného média glukózou, ale ne delší, než 48 hodin
44 4.6 Biofilm detection and the clinical significance of Staphylococcus epidermidis isolates Staphylococcus epidermidis je hlavní součástí běžné mikroflóry kůže člověka. V posledních desetiletích je však i u S. epidermidis, stejně jako u ostatních koagulázanegativních stafylokoků, stále více diskutována též jeho schopnost způsobovat infekce krevního řečiště (IKŘ). Nozokomiální infekce způsobené koagulázanegativními stafylokoky jsou spojeny s umělými materiály a chirurgickými pomůckami přítomnými v těle pacienta. Jedná se zejména o různé katétry, shunty, umělé srdeční chlopně, pacemakery apod. (Costerton, 1999). Koagulázanegativní stafylokoky jsou zodpovědné za téměř 40 % infekcí krevního řečiště spojených s katétry (Sherertz et al., 1990). Problémem v současné době stále zůstává rozlišení S. epidermidis jako etiologického agens IKŘ od pouhé kontaminaty, saprofytického kmene S. epidermidis, který se mohl do odebírané hemokultury dostat např. při jejím nesprávném odběru. Jedním z faktorů, které by nám měly při interpretaci mikrobiologického nálezu S. epidermidis v hemokultuře či hemokulturách pomoci, je schopnost daného izolátu tvořit biofilm. Tento faktor virulence je častěji prokazován u relevantních izolátů z hemokultur, než u jejich kontaminant z kůže (de Nicolas et al., 1995; von Eiff et al., 2001). Pro detekci schopnosti tvorby biofilmu se využívají dva přístupy fenotypový a genetický. Nejpoužívanějšími fenotypovými metodami jsou Christensenova zkumavková metoda, nebo její modifikace v mikrotitračních destičkách a kultivace na agaru s kongo červení. Christensenova zkumavková metoda odhaluje schopnost mikroba adherovat k pevným povrchům a růst ve vrstvě biofilmu (Christensen et al., 1982), zatímco kultivace na agaru s kongo červení detekuje produkci extracelulárního polysacharidu (Freeman et al., 1989). Genetické metody jsou založeny na detekci genů zodpovědných za tvorbu biofilmu, v případě stafylokoků tedy konkrétně genů ica operonu (Frebourg et al., 2000). V předkládané práci byly na základě dvou definovaných souborů kmenů S. epidermidis srovnány všechny výše zmíněné metody detekce tvorby biofilmu průkaz ica operonu, Christensenova zkumavková metoda a kultivace na agaru s kongo červení z hlediska jejich vypovídací schopnosti. První skupina kmenů S. epidermidis zahrnovala kmeny izolované z hemokultur pacientů s klinickými i laboratorními markery sepse, tedy klinicky relevantní kmeny. Druhá skupina kmenů zahrnovala kmeny S. epidermidis izolované z kůže zdravých dobrovolníků
45 Ze 468 kmenů S. epidermidis izolovaných z hemokultur pacientů různých oddělení FN u sv. Anny v Brně byly do první skupiny byly zařazeny pouze ty kmeny (147), které splňovaly definované podmínky indikující IKŘ (Paterson et al., 2000), tj. opakovanou izolaci stejného kmene z několika hemokultur jednoho pacienta, jeden či více klinických příznaků sepse u tohoto pacienta (tělesná teplota nad 38 C či pod 36 C, tachykardie vyšší než 1,5 Hz, tj. více než 90 úderů srdce za minutu, respirační rychlost větší než 0,33 Hz, tj. více než 20 vdechů za minutu, parciální tlak CO 2 4,3 kpa, hypotenze vyšší než 12 kpa, tj. méně než 90 mmhg) a zároveň jeden či více laboratorních markerů systémové zánětlivé odpovědi (leukocytóza vyšší než 12x10 9 /l či leukopenie nižší než 4x10 9 /l, zvýšená hladina CRP, prokalcytoninu, vyšší sedimentace apod.). Kmeny zahrnuté do druhé skupiny byly izolovány z kůže loketní jamky zdravých dobrovolníků. Těchto kmenů bylo izolováno tolik, aby testované skupiny byly stejně velké (147). Přítomnost ica operonu byla prokázána u 92 (62,6 %) ze 147 kmenů izolovaných z hemokultur a u 44 (29,9 %) ze 147 kmenů izolovaných z kůže. Christensenovou zkumavkovou metodou bylo mezi izoláty z hemokultur prokázáno 57 (40,1 %) silně pozitivních a 22 (15 %) slabě pozitivních kmenů, mezi izoláty z kůže pak 22 (14,9 %) silně pozitivních a 11 (7.5 %) slabě pozitivních kmenů. Metodou kultivace na agaru s kongo červení bylo mezi izoláty z hemokultur prokázáno 37 (25,2 %) silně pozitivních a 27 (18,4 %) slabě pozitivních kmenů, mezi izoláty z kůže pak 18 (11,6 %) silně pozitivních a 13 (8.8 %) slabě pozitivních kmenů. Výsledky jsou shrnuty v tabulce 4. Tabulka 4 Percentuální zastoupení kmenů s přítomným ica operonem a schopností tvorby biofilmu hemokultury kožní kmeny Tvorba biofilmu CRA (%) CTT (%) ICA (%) CRA (%) CTT (%) ICA (%) negativní 83 (56.4) 68 (46.3) 55 (37.4) 116 (78.9) 114 (77.6) 103 (70.1) slabě pozitivní 27 (18.4) 22 (15.0) 13 (8.8) 11 (7.5) 92 (62.6) silně pozitivní 37 (25.2) 57 (38.7) 18 (12.3) 22 (14.9) 44 (29.9) celkem pozitivní 64 (43.5) 79 (53.7) 92 (62.6) 31 (21.1) 33 (22.4) 44 (29.9) Legenda: CRA kultivace na agaru s kongo červení; CTT Christensenova zkumavková metoda, ICA přítomnost ica operonu Ze statistického porovnání všech tří metod vyplývá, že Christensenova zkumavková metoda, stejně jako detekce ica operonu korelují lépe s klinickou významností kmene a jsou tedy metodami s vyšší vypovídací hodnotou, než metoda kultivace na agaru s kongo červení (p = 0,00022). Výsledky statistického zpracování jsou uvedeny v tabulce 5. Detekce ica operonu je metodou rychlou, dobře hodnotitelnou, a proto exaktní. Nevýhodou této metody je fakt, že vypovídá pouze o přítomnosti genetické informace v genomu, a tedy potenciální možnosti testovaného kmene tvořit biofilm. Pomocí této metody nejsme schopni zhodnotit,
46 zda je schopnost tvorby biofilmu exprimována a zda se tedy fenotypově projeví (O'Gara et al., 2001). Metody průkazu fenotypové exprese tvorby biofilmu se zaměřují na aktuální stav exprese ica operonu u daného kmene, a tedy přesněji vypovídají o jeho klinickém významu. Na druhou stranu jejich hodnocení je obtížné a je tedy zatíženo poměrně velkou subjektivní chybou. Další jejich nevýhodou je závislost výsledků získaných těmito metodami na konkrétních kultivačních podmínkách, což zvyšuje nestandardnost výsledků. Určitým řešením je současná kombinace genetických a fenotypových metod, tedy vyšetření testovaného kmene jak na přítomnost ica operonu, tak Christensenovou zkumavkovou metodou. Tabulka 5 Výsledky statistických analýz Sum of Sq AIC F <zero model> CRA x 10-8 CTT ICA x 10-7 Legenda: CRA kultivace na agaru s kongo červení; CTT Christensenova zkumavková metoda, ICA přítomnost ica operonu; Sum of Sq součet čtverců; AIC Akaike index (Akaike information criterion); F pravděpodobnost Využití schopnosti tvorby biofilmu jako rozlišujícího znaku mezi klinicky významnými izoláty z hemokultur a kontaminantami je obtížné zejména kvůli vysoké prevalenci kmenů s přítomností ica operonu mezi kožními izoláty. I v tom nám však může pomoci kombinace genetických a fenotypových vyšetřovacích metod. Kmeny, v jejichž genomu je přítomen ica operon a které jsou schopny tvořit biofilm, představují klinicky nejvýznamnější kmeny, které způsobují IKŘ nejčastěji (Mulder et al., 1998; Frebourg et al., 2000; Vogel et al., 2000; Vandecasteele et al., 2003). Kmeny, které netvoří biofilm ani v jejich genomu není přítomen ica operon představují kmeny, které zřídka kolonizují umělé povrchy v krevním řečišti a rovněž zřídka způsobují IKŘ. Takové kmeny mohou být považovány za klinicky nevýznamné. Kmeny, které mají v genomu přítomen ica operon by měly být považovány za kmeny potenciálně nebezpečné, i když fenotypově schopnost tvorby biofilmu nevykazují. Exprese ica operonu u nich může být totiž indukována např. stresem vnějšího prostředí (Fitzpatrick et al., 2002), což může znenadání zvýšit jejich klinický význam
47 4.7 Rozdíly v citlivosti k antibiotikům u biofilmpozitivních a biofilmnegativních kmenů Staphylococcus epidermidis izolovaných z hemokultur Je známo, že bakterie rostoucí ve formě biofilmu bývají až o několik řádů odolnější k působení antibiotik než bakterie planktonické, a proto se ke stanovení jejich citlivosti nedá použít konvenční technika stanovení MIC (Donlan, 2000; Costerton et al., 1995). Vzhledem k tomu, že pro stanovení minimální biofilm eradikující koncentrace antibiotik zatím neexistují mezinárodně schválené postupy, může se klinik v současné době spoléhat pouze na stanovení MIC a případně znalost schopnosti tvorby biofilmu u konkrétního mikrobiálního kmene. Cílem předkládané práce bylo zjistit, zda hodnota MIC stanovená konvenčními postupy vypovídá něco o rezistenci biofilmpozitivních kmenů k antibiotikům a zda je mezi biofilmpozitivními a biofilmnegativními kmeny rozdíl v MIC jednotlivých antibiotik. Do studie byla zahrnuto 86 kmenů S. epidermidis opakovaně izolovaných z hemokultur pacientů se známkami sepse. U všech kmenů byla stanovena schopnost tvorby biofilmu genetickými (průkaz přítomnosti ica operonu) i fenotypovými metodami (Christensenova zkumavková metoda, metoda kultivace na agaru s kongo červení). Kmeny byly na základě schopnosti tvorby biofilmu rozděleny do dvou skupin. Do první skupiny byly zařazeny kmeny ve všech testovaných metodách pozitivní, tedy kmeny jasně biofilmpozitivní (43), do druhé skupiny byly zařazeny kmeny ve všech testovaných metodách negativní, tedy kmeny jasně biofilmnegativní (43). Minimální inhibiční koncentrace antibiotik byla stanovována mikrodiluční metodou s komerčně dodávanými destičkami (Urbášková, 1998). Data byla hodnocena dvouvýběrovými t-testy (Statistica for Windows, 1999). Graf 1 v příloze 7.7 znázorňuje průměrné hodnoty MIC biofilmpozitivních a biofilmnegativních kmenů k daným antibiotikům. U kmenů biofilmpozitivních jsou patrné vyšší průměrné hodnoty MIC. V tabulce 6 jsou zaznamenány nejčastěji zachycené hodnoty MIC u obou skupin kmenů (medián). Z tabulky i z grafu jsou patrné významné rozdíly v citlivosti mezi srovnávanými skupinami. Statisticky významný rozdíl v citlivosti mezi kmeny biofilmpozitivními a kmeny biofilmnegativními byl nalezen u oxacilinu, tetracyklinu, kotrimoxazolu a gentamicinu (P < 0,05). Vůči teikoplaninu a vankomycinu byla zaznamenána 100% citlivost v obou testovaných skupinách. Přestože je hodnota MIC stanovena pro planktonickou formu růstu, byl prokázán rozdíl v rezistenci kmenů S. epidermidis biofilmpozitivních oproti kmenům biofilmnegativním. Můžeme tedy říci, že biofilmpozitivní kmeny jsou i v planktonické formě rezistentnější než kmeny biofilmnegativní
48 Tabulka 6 - Nejčastěji zachycené hodnoty MIC Biofilmnegativní kmeny Biofilmpozitivní kmeny Průměr Medián SE Průměr Medián SE PEN OXA CMP TET COT ERY CLI CIP GEN TEI VAN Legenda: PEN penicilin; OXA oxacilin; CMP chloramfenikol; TET tetracyklin; COT kotrimoxazol; ERY erytromycin; CLI klindamycin; CIP ciprofloxacin; GEN gentamicin; TEI teikoplanin; VAN vankomycin; SE střední chyba průměru Vyšší MIC biofilmpozitivních kmenů lze vysvětlit vlivem slabé slizové vrstvy v okolí buněk a tvorbou mikrokolonií. Tato ochranná vrstva je však (narozdíl od silné vrstvy vyzrálého biofilmu) u samostatných planktonických buněk poměrně slabá. To spolu s absencí dalších faktorů, které se uplatňují až ve zformovaném biofilmu (zpomalení růstu uvnitř biofilmové vrstvy, výrazné změny vnitřního prostředí včetně acidifikace a nedostatku kyslíku apod.), vede k pozorovanému, ale relativně malému rozdílu mezi MIC biofilmpozitivních a biofilmnegativních kmenů, na rozdíl od studií, které sledovaly minimální biofilm inhibující koncentraci (MBIC) u již plně vyvinutého a zralého biofilmu (Monzon et al., 2002; Knobloch et al., 2002b) a další. Vazba molekul antibiotik na záporně nabité chemické struktury obsažené ve slizové vrstvě může být odpovědná za rozdíly v rezistenci ke gentamicinu. Lze též předpokládat, že již v rámci mikrokolonií může docházet k malým změnám fyzikálně-chemických podmínek, což může rezistenci vůči gentamicinu umocňovat. Změna fyzikálně-chemických podmínek může mít vliv i na tetracyklin. Také koncentrace enzymů degradujících antibiotika v mikrokoloniích i v těsném okolí bakteriální buňky vlivem slizové vrstvy může zvyšovat rezistenci bakterií, např. k oxacilinu. Rozdíly v hodnotách MIC biofilmpozitivních a biofilmnegativních kmenů korespondují s výsledky prací jiných autorů (Amorena et al., 1999; Souli et al., 1998; Knobloch et al., 2002b) a poukazují na nutnost hodnotit výsledky MIC opatrně a s ohledem na zvýšenou rezistenci biofilmpozitivních kmenů. Přestože podle stanovení MIC standardním způsobem se některé kmeny mohou zdát citlivé, podmínky in vivo jsou dosti odlišné a biofilmpozitivní kmeny mohou standardně používané koncentrace antibiotik přežívat
49 4.8 Stanovení citlivosti k antibiotikům u biofilmpozitivních forem mikroorganizmů Nozokomiální infekce způsobované biofilmpozitivními mikroorganizmy představují závažný terapeutický problém. Mikroorganizmy jsou v biofilmu chráněny proti nepříznivým vlivům zevního prostředí včetně účinku antibiotik. Je známo, že hodnoty minimálních inhibičních koncentrací (MIC) stanovené pro planktonickou formu neodpovídají skutečným koncentracím antibiotik potřebným k likvidaci bakterií v biofilmu. Minimální biofilm inhibující koncentrace (MBIC) a minimální biofilm eradikující koncentrace (MBEC) nemají žádný přímý či dobře odvoditelný vztah k MIC či MBC, a tudíž při jejich stanovování nelze z MIC a MBC vycházet (Ceri et al., 1999; Ceri et al., 2001 a další). Tento fakt velmi komplikuje léčbu biofilmových infekcí, protože v rutinní mikrobiologické praxi je u mikrobů rutinně stanovována pouze MIC daného antibiotika. Rozdíly mezi hodnotami MIC a MBEC se liší nejen mezi jednotlivými antibiotiky a jednotlivými bakteriálními druhy, ale i mezi jednotlivými kmeny stejného druhu mikroba. Hodnota MBEC konkrétního antibiotika může být u jednoho testovaného druhu může být vyšší v rozmezí koncentrací deseti- až tisícinásobných ve srovnání s MIC či MBC (Costerton, 1999; Mah et al., 2001). Z klinického hlediska představuje vyšší rezistence biofilmových forem bakterií závažný problém a je příčinou častého selhávání antimikrobiální terapie těchto infekcí. Cílem této práce bylo navržení postupu stanovení minimální biofilm inhibující koncentrace a minimální biofilm eradikující koncentrace a porovnání těchto hodnot s hodnotami MIC. Ve studii bylo použito 45 biofilmpozitivních kmenů S. epidermidis izolovaných z hemokultur. U všech těchto kmenů byla biofilmpozitivita vyšetřena jak metodami genetickými (průkaz ica operonu), tak metodami fenotypovými (Christensenova zkumavková metoda, kultivace na agaru s kongo červení). Do studie byly zahrnuty pouze kmeny silně biofilmpozitivní všemi testovanými metodami. Pro studium biofilmu a jeho rezistencí k antibiotikům byly opět použity polystyrenové destičky s hroty. Na destičkách s hroty byla vypěstována vrstva biofilmu a tento biofilm byl vystaven působení logaritmicky ředěných antibiotik. Koncentrace použitých antibiotik jsou uvedeny v tabulce 7. Pro stanovení MBEC bylo použito kolorimetrické médium, ve kterém na základě bakteriálního metabolizmu a tedy změny ph dochází ke změně barvy. Přeživší bakteriální buňky díky tomu mohou být detekovány snáze a dříve, než při odečítání zákalu či sedimentu
50 Tabulka 7 Použité koncentrace antibiotik Mikrotitrační destička se standardními koncentracemi antibiotik PEN OXA AMS CMP TET COT ERY CLI CIP GEN TEI VAN KR Mikrotitrační destička se zvýšenými koncentracemi antibiotik PEN OXA AMS CMP TET COT ERY CLI CIP GEN TEI VAN KR Legenda: PEN penicilin; OXA oxacilin; AMS ampicilin-sulbaktam; CMP chloramfenikol; TET tetracyklin; COT kotrimoxazol; ERY erytromycin; CLI klindamycin; CIP ciprofloxacin; GEN gentamicin; TEI teikoplanin; VAN vankomycin, KR kontrola růstu. Minimální biofilm inhibující koncentrace antibiotik, MBIC, hodnotíme analogicky k MIC, je to koncentrace antibiotika, při které v jamce mikrotitrační destičky již není patrný zákal ani sediment bakteriálních buněk. Minimální biofilm eradikující koncentrace, MBEC, byla hodnocena na základě změny barvy kolorimetrického média. Je to taková koncentrace antibiotika, při které již nedochází k růstu mikroba, a tedy ani ke změně barvy kolorimetrického média, bez ohledu na absenci zákalu či sedimentu na dně jamek (obr. 13). V tabulce 8 jsou uvedeny průměrné a střední hodnoty MIC, MBIC a MBEC. Rozdíly mezi hodnotou MIC a hodnotami ovlivňujícími buňky uvnitř biofilmu jsou statisticky významné (p = 0,01). Graf 1 v příloze 7.8 ilustruje rozdíly v hodnotách MIC, MBIC a MBEC v závislosti na logaritmickém ředění antibiotik. Zjištěné hodnoty MBEC naznačují, že běžně užívaných koncentrací testovaných antibiotik nelze v monoterapii pro účinnou eradikaci biofilmu použít. Teoreticky může být tato rezistence vysvětlena mechanizmy biofilmové rezistence, jak byly diskutovány v předchozích kapitolách. Pouhé stanovení MIC tedy ke zjištění koncentrace antibiotika, která

51 je schopna mikrobiální buňky v biofilmu zlikvidovat, nestačí. Je rovněž zřejmé, že koncentrace antibiotik schopných účinně zasáhnout biofilm vysoce přesahují hladiny antibiotik dosažitelných v séru pacienta. Uvedená fakta jsou důvodem častého selhání terapie biofilmových infekcí, je-li založená pouze na znalosti MIC (Costerton, 1999). Antibiotická terapie založená na znalosti MIC může vést u těchto infekcí k potlačení klinických příznaků vzniklých uvolněním bakterií z biofilmu, ale selhává při likvidaci bakteriálních buněk dosud usazených v matrix biofilmu. Po ukončení antimikrobiální léčby pak může vrstva biofilmu sloužit jako ložisko a vyvolat tak recidivu infekce. Je známo, že infekce spojené s biofilmem i Obr. 12 Hodnocení MBEC pomocí kolorimetrického média - původní barva média je červená, metabolickou aktivitou přežívajících bakterií se mění na žlutou; Antibiotika rozplněna v následujícím pořadí: PEN, OXA, AMS, CMP, TET, COT, ERY, CLI, CIP, GEN, TEI, VAN; vysvětlivky zkratek antibiotik viz tab. 7, VH přes antimikrobiální terapii obvykle přetrvávají, dokud není ložisko odstraněno z těla pacienta (Stewart et al., 2001). Tabulka 8 Průměrné hodnoty minimální inhibiční koncentrace (MIC), minimální biofilm inhibující koncentrace (MBIC) a minimální biofilm eradikující koncentrace (MBEC) u 45 kmenů Staphylococcus epidermidis izolovaných z hemokultur při sepsi MIC MBIC MBEC průměr medián průměr medián průměr medián PEN 2, OXA 11, AMS 5,6 2 34, CMP 8, , TET 6,68 0,5 25, COT 16, ERY 14, CLI 9, CIP 9, GEN 10,4 0,25 80, TEI 2, , VAN 1,64 2 7, Legenda: PEN penicilin; OXA oxacilin; AMS ampicilin-sulbaktam; CMP chloramfenikol; TET tetracyklin; COT kotrimoxazol; ERY erytromycin; CLI klindamycin; CIP ciprofloxacin; GEN gentamicin; TEI teikoplanin; VAN vankomycin
52 4.9 Selective pressure of higher sanitation standards increases relative number of biofilm-positive microbes on the intensive care unit Je známo, že schopnost tvorby biofilmu je významným faktorem virulence, který je přítomen u mnoha kmenů mnoha bakteriálních druhů. Schopnost tvorby biofilmu vykazuje určité procento bakterií izolovaných z vnějšího prostředí, pro bakterie kolonizující např. kanyly či jiné plastové povrchy v těle pacienta je tento faktor virulence klíčovým, a proto se mezi kmeny, jež jsou považovány za etiologická agens takto vzniklých onemocnění, vyskytuje schopnost tvorby biofilmu výrazně častěji, než u mikrobů izolovaných z vnějšího prostředí či u mikrobů náhodně kontaminujících vzorky odebrané k mikrobiologickému zpracování (Růžička et al., 2004; Freeman et al., 1989). Zvláště významnou roli mohou hrát biofilmpozitivní mikroby u infekcí imunokompromitovaných pacientů, jejichž oslabený imunitní systém nemůže tělu poskytnout dostatečnou obranu (Bals et al., 1999). Přítomnost biofilmpozitivních bakterií na odděleních s těmito pacienty tak může být příčinou mnoha problémů. Za hlavní zdroj nozokomiálních patogenů v nemocničních zařízeních se obvykle považují pacienti a personál (Akyol et al., 2006), méně pozornosti se věnuje prostředí těchto zařízení, které je však důležitým rezervoárem mikrobů (Macias et al., 2005; Seenivasan et al., 2005). Cílem předkládané práce bylo porovnat mikrobiální kontaminaci prostředí aseptických boxů pro pacienty po allogenních transplantacích, tedy jednotek s vyššími nároky na čistotu prostředí, s prostředím standardní lůžkové části stejné kliniky. Prostředí srovnávaných jednotek Aseptické boxy (AB) Interní hemato-onkologické kliniky jsou konstruovány tak, aby poskytovaly aseptické prostředí pacientům po allogenních transplantacích. AB se skládá z pokoje pro pacienta s koupelnou a sociálním zařízením. Tento pokoj je od chodby kliniky oddělen uzavřenou předsíní. Celý pokoj včetně stěn a stropu je vyroben z materiálů s omyvatelnou povrchovou úpravou, kterou je možné dezinfikovat. Klimatizace AB má vlastní okruh. Každý AB je vybaven sterilními pracovními ochrannými pomůckami, do kterých se převléká všechen personál při vstupu do předsíně AB (plášť, rukavice, rouška, obuv). Všechny předpisy jsou řešeny tak, aby se co možná nejvíce minimalizoval kontakt imunosuprimovaného pacienta mikrobiální kontaminací z vnějšího prostředí. Tomu odpovídá i čištění a dezinfekce jednotlivých AB. Pro každý AB jsou určeny oddělené čistící pomůcky,
53 které jsou skladovány v předsíni konkrétního AB. Dezinfekční látky používané při čištění boxů se mění každé dva týdny. Sprchová růžice je ponořena do dezinfekčního roztoku celou dobu, kdy ji pacient nepoužívá. Všechna vyústění odpadů jsou denně dezinfikována. Používané dezinfekční prostředky se svými účinnými látkami řadí do několika skupin (preparáty na bázi fenolu, chloru, aldehydů, alkoholů s kvartérními amoniovými solemi aj.). Poté, co pacient opustí AB, jsou všechny povrchy tohoto boxu dvakrát omyty roztokem detergentů a dezinfekčních látek, ponechány uschnout a vydezinfikovány sprejem. Poté je AB připraven pro dalšího pacienta. Oddělení nemocniční hygieny sleduje pravidelně prostředí AB. Výsledky kontrolních mikrobiologických stěrů jsou téměř vždy sterilní, pouze ojediněle se vyskytují koagulázanegativní stafylokoky či P. aeruginosa v zanedbatelných počtech. Standardní lůžková část (SHU) stejné kliniky je dělena na vícelůžkové pokoje se sociálním zázemím společným pro celé oddělení. Vzhledem k tomu, že jsme se již při odběru vzorků zaměřovali na biofilmpozitivní mikroby, prováděli jsme odběr jako rázný seškrab povrchu sterilním skalpelem. Tuto odběrovou metodu jsme zvolili na základě našich předchozích zkušeností (Holá et al., unpubl. data). Místa odběru vzorků byla: umyvadlo (5 x AB; 5 x SHU), vnitřní a vnější povrchy vodovodního kohoutku (5 x AB; 5 x SHU), dlaždičky v okolí umyvadla (5 x AB; 5 x SHU) a vnitřní a vnější povrch sprchové růžice (5 x AB; 5 x SHU). Z 60 vyšetřených vzorků bylo pouze 11 sterilních (18,3 %). Graf 1 v příloze 7.9 ukazuje počty izolovaných bakterií v jednotlivých vzorcích. Celkem jsme izolovali 116 bakteriálních kmenů, z nichž většinu tvořily gramnegativní bakterie (Pseudomonas aeruginosa, Stenotrophomonas maltophilia, Klebsiella pneumoniae, Serratia marcescens, Enterobacter aerogenes, Proteus mirabilis, Acinetobacter baumanii-calceolyticus aj.). Z grampozitivních bakterií jsme izolovali zejména Staphylococcus epidermidis, Micrococcus luteus a Bacillus sp. Nejčastěji izolovanými druhy byly Pseudomonas aeruginosa a Stenotrophomonas maltophilia. Z SHU bylo izolováno 61,2 % bakterií (71 kmenů), z AB pak 38, 8 % (45 kmenů). Souhrnné počty CFU jsou uvedeny v tabulce 9, druhové spektrum izolovaných bakterií pak v tabulce 10. Tabulka 9 Počty CFU izolovaných kmenů Počet CFU SHU (71 kmenů) AB (55 kmenů) < (14.1%) 5 (9.0%) (22.5%) 20 (36.4%) (36.6%) 17 (30.9%) > (26.8%) 13 (23.6%) Legenda: CFU colony-forming units; SHU standardní lůžková část; AB aseptické boxy
54 Schopnost tvorby biofilmu byla prokázána u 71,2 % kmenů izolovaných z AB a 34,5 % kmenů izolovaných z SHU. Rozložení kvantitativně vyjádřené schopnosti tvorby biofilmu na obou odděleních je patrné z grafu 2 v příloze 7.9. Tabulka 10 Bakteriální druhy izolované ze standardní lůžkové části a z aseptických boxů SHU AB G+ Staphylococcus auricularis - Staphylococcus epidermidis Staphylococcus epidermidis Staphylococcus lentus - Micrococcus luteus - Micrococcus spp. Micrococcus spp. Kocuria varians Kocuria varians Kytococcus sedentarius Kytococcus sedentarius Bacillus spp. - G- Klebsiella pneumoniae Klebsiella pneumoniae Serratia marcescens - Enterobacter aerogenes Enterobacter aerogenes Entertobacter spp. - Proteus mirabilis - GNFB Acinetobacter johnsonii - skupina A. baumanii-calceolyticus skupina A. baumanii-calceolyticus Comamonas terrigena - Moraxella canis - Pseudomonas aeruginosa Pseudomonas aeruginosa - Stenotrophomonas maltophilia - Stenotrophomonas sp. Legenda: SHU standardní lůžková část; AB aseptické boxy; G+ - grampozitivní bakterie; G- - gramnegativní bakterie;, GNFB gramnegativní nefermentující bakterie Rozdíly v počtech izolovaných mikrobů je pravděpodobně způsobeno rozdílným způsobem čištění těchto dvou oddělení. Na standardním lůžkovém oddělení není nutno eradikovat všechny mikroby z prostředí, ale pouze snížit jejich počet na únosnou míru. V aseptických boxech je, vzhledem k přítomnosti imunokompromitovaných pacientů, nutno dosáhnout vyšší úrovně čistoty prostředí, ideálně by prováděné postupy měly zaručit vznik aseptického prostředí. V této souvislosti se jako velmi problematická jeví přítomnost gramnegativních nefermentujících bakterií, zejm. P. aeruginosa ve vzorcích z vnitřních povrchů sprchových růžic a vodovodních kohoutků. Je známo, že P. aeruginosa dokáže v nutričně chudých prostředích přežívat a dokonce se i množit, ale její přítomnost ve sprchových růžicích kontinuálně ponořených do roztoku dezinfekčního prostředku (AB) je alarmující. Druhové spektrum bakterií získaných z AB je výrazně užší, než spektrum bakterií získaných z SHU, stejně jako počty CFU jednotlivých bakterií. Vyšší nároky na čistotu
55 prostředí však mohou být paradoxně příčinou výrazně vyššího percentuálního zastoupení biofilmpozitivních bakterií ve vzorcích z AB. Celkové nižší počty mikrobů ve vzorcích z AB v porovnání se vzorky z SHU jsou způsobeny důkladnějšími sanitačními postupy. Biofilmnegativní bakterie mohou být totiž těmito čistícími postupy eradikovány, či jejich počty mohou být velmi výrazně sníženy, zatímco biofilmpozitivní bakterie přežívají. Proto je schopnost tvorby biofilmu pro bakterie v těchto podmínkách výhodným faktorem ovlivňujícím jejich přežití v prostředí. Ačkoliv mnoho druhů izolovaných z nemocničního prostředí je považováno na nepatogenní, případně nízce patogenní, přítomnost faktorů virulence, jako je například schopnost tvorby biofilmu, zvyšuje jejich klinický význam, a to zejména u imunokompromitovaných pacientů či pacientů s dlouhodobě zavedenými umělými implantáty
56 5. Závěr Schopnost tvorby biofilmu je významným faktorem virulence, který se projevuje v různé míře u různých mikrobů. Biofilmpozitivní mikroby jsou schopny kolonizovat širokou škálu prostředí, a to jak přírodních, tak umělých. Infekce způsobené biofilmpozitivními mikroby jsou celosvětově významným problémem a problematika jejich rezistence k antimikrobiálním látkám a dezinfekčním činidlům je stále aktuálním tématem. Abychom byli schopni léčit infekční onemocnění či zabránit jejich šíření, musíme v prvé řadě poznat a pochopit zdroje a vektory přenosu mikroorganizmů a stejně tak strategie jejich přežití uvnitř i vně hostitele. Předložené práce se věnovaly studiu povrchových vlastností adhezních materiálů, studiu dynamiky tvorby biofilmu za různých podmínek a také studiu účinku antimikrobiálních látek na biofilm a rezistence biofilmpozitivních mikroorganizmů k antimikrobiálním látkám. Další z předkládaných prací se pak věnovala hygienickoepidemiologickým souvislostem schopnosti tvorby biofilmu u mikrobů v nemocničním prostředí. Na základě studia povrchových vlastností adhezních materiálů jsme získali vhodný povrch pro kultivaci a studium biofilmů in vitro, což je důležité právě pro sledování dynamiky procesů probíhajících v biofilmu. Dynamika růstu biofilmu a studium jeho fyziologie jsou zase nezbytnými předpoklady pro pochopení biofilmové rezistence k antimikrobiálním látkám. Sledování rezistence biofilmpozitivních mikrobů a minimálních biofilm eradikujících koncentrací je důležité nejen pro znalost hodnot MBEC samotných, ale též pro možnost lépe a podrobněji informovat širokou lékařskou veřejnost o tomto závažném problému. Studium epidemiologie by zase mělo pomoci v poznání skutečných šancí a možností biofilmpozitivních mikrobů v našich nemocnicích, protože boj s nimi se rozhodně ještě neskončil. Nelze než souhlasit se sirem Alexandrem Flemingem, který v roce 1945 na přednášce v Londýně prohlásil: Ať je nadšení pro penicilin sebevětší, musíme mít na paměti, že lidské tělo a mikroby mají zvláštní způsob, jak reagovat na nové činitele, s nimiž přijdou do styku. Příroda se nepodrobuje snadno a kdykoli člověk nalezl nový prostředek, odpověděla s větší či menší prudkostí a vyrobila protiprostředek, jímž se pokouší neutralizovat nebo zneškodnit dobré účinky nové látky. Prozatím nevíme, jaké komplikace vyvstanou z ustavičného používání penicilinu. Ale ať přijdou kdykoli, lékařská věda bude proti nim muset rázně bojovat
57 6. Literatura 1. Akyol, A., Ulusoy, H., Ozen, I., 2006: Handwashing: a simple, economical and effective method for preventing nosocomial infections in intensive care units; Journal of Hospital Infection; 62; 4: p Al Fattani, M. A., Douglas, L. J., 2006: Biofilm matrix of Candida albicans and Candida tropicalis: chemical composition and role in drug resistance; J. Med. Microbiol.; 55; Pt 8: p Allignet, J., Galdbart, J. O., Morvan, A., Dyke, K. G., Vaudaux, P., Aubert, S., Desplaces, N., el Solh, N., 1999: Tracking adhesion factors in Staphylococcus caprae strains responsible for human bone infections following implantation of orthopaedic material; Microbiology; 145; Pt 8: p Allignet, J., Aubert, S., Dyke, K. G., el Solh, N., 2001: Staphylococcus caprae strains carry determinants known to be involved in pathogenicity: a gene encoding an autolysin-binding fibronectin and the ica operon involved in biofilm formation; Infection and Immunity; 69; 2: p Ammendolia, M. G., Di Rosa, R., Montanaro, L., Arciola, C. R., Baldassarri, L., 1999: Slime production and expression of the slime-associated antigen by staphylococcal clinical isolates; Journal of Clinical Microbiology; 37; 10: p Amorena, B., Gracia, E., Monzon, M., Leiva, J., Oteiza, C., Perez, M., Alabart, J. L., Hernandez-Yago, J., 1999: Antibiotic susceptibility assay for Staphylococcus aureus in biofilms developed in vitro; Journal of Antimicrobial Chemotherapy; 44; 1: p Anderl, J. N., Franklin, M. J., Stewart, P. S., 2000: Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin; Antimicrobial Agents and Chemotherapy; 44; 7: p Anderl, J. N., Zahller, J., Roe, F., Stewart, P. S., 2003: Role of nutrient limitation and stationary-phase existence in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin; Antimicrob. Agents Chemother.; 47; 4: p Arciola, C. R., Baldassarri, L., Montanaro, L., 2001: Presence of icaa and icad genes and slime production in a collection of staphylococcal strains from catheter-associated infections; Journal of Clinical Microbiology; 39; 6: p Arciola, C. R., Baldassarri, L., Montanaro, L., 2002: In catheter infections by Staphylococcus epidermidis the intercellular adhesion (ica) locus is a molecular marker of the virulent slime-producing strains; J. Biomed. Mater. Res.; 59; 3: p Arciola, C. R., Campoccia, D., Baldassarri, L., Donati, M. E., Pirini, V., Gamberini, S., Montanaro, L., 2006: Detection of biofilm formation in Staphylococcus epidermidis from implant infections. Comparison of a PCR-method that recognizes the presence of ica genes with two classic phenotypic methods; J. Biomed. Mater. Res. A; 76; 2: p Baldassarri, L., Simpson, W. A., Donelli, G., Christensen, G. D., 1993: Variable fixation of staphylococcal slime by different histochemical fixatives; Eur. J. Clin. Microbiol. Infect. Dis.; 12; 11: p Bals, R., Weiner, D. J., Wilson, J. M., 1999: The innate immune system in cystic fibrosis lung disease; J. Clin. Invest.; 103; 3: p
58 14. Bardouniotis, E., Huddleston, W., Ceri, H., Olson, M. E., 2001: Characterization of biofilm growth and biocide susceptibility testing of Mycobacterium phlei using the MBEC assay system; FEMS Microbiology Letters; 203; 2: p Beattie, T. K., Tomlinson, A., McFadyen, A. K., 2006: Attachment of Acanthamoeba to first- and second-generation silicone hydrogel contact lenses; Ophthalmology; 113; 1: p Beenken, K. E., Dunman, P. M., McAleese, F., Macapagal, D., Murphy, E., Projan, S. J., Blevins, J. S., Smeltzer, M. S., 2004: Global gene expression in Staphylococcus aureus biofilms; J. Bacteriol.; 186; 14: p Beloin, C., Ghigo, J. M., 2005: Finding gene-expression patterns in bacterial biofilms; Trends Microbiol.; 13; 1: p Berra, L., De Marchi, L., Yu, Z. X., Laquerriere, P., Baccarelli, A., Kolobow, T., 2004: Endotracheal tubes coated with antiseptics decrease bacterial colonization of the ventilator circuits, lungs, and endotracheal tube; Anesthesiology; 100; 6: p Bjarnsholt, T., Jensen, P. O., Burmolle, M., Hentzer, M., Haagensen, J. A., Hougen, H. P., Calum, H., Madsen, K. G., Moser, C., Molin, S., Hoiby, N., Givskov, M., 2005: Pseudomonas aeruginosa tolerance to tobramycin, hydrogen peroxide and polymorphonuclear leukocytes is quorum-sensing dependent; Microbiology; 151; Pt 2: p Bollinger, N., Hassett, D. J., Iglewski, B. H., Costerton, J. W., McDermott, T. R., 2001: Gene expression in Pseudomonas aeruginosa: evidence of iron override effects on quorum sensing and biofilm-specific gene regulation; J. Bacteriol.; 183; 6: p Branchini, M. L., Pfaller, M. A., Rhine-Chalberg, J., Frempong, T., Isenberg, H. D., 1994: Genotypic variation and slime production among blood and catheter isolates of Candida parapsilosis; Journal of Clinical Microbiology; 32; 2: p Bremer, P. J., Geesey, G. G., Drake, B., 1992: Atomic force microscopy examination of the topography of a hydrated bacterial biofilm on a copper surface; Current Microbiology; 24; 4: p Brombacher, E., Baratto, A., Dorel, C., Landini, P., 2006: Gene expression regulation by the Curli activator CsgD protein: modulation of cellulose biosynthesis and control of negative determinants for microbial adhesion; J. Bacteriol.; 188; 6: p Brooun, A., Liu, S., Lewis, K., 2000: A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms; Antimicrob. Agents Chemother.; 44; 3: p Brun, B. C., Rauss, A., Legrand, P., 1987: Semiquantitative culture of catheter tips; Journal of Clinical Microbiology; 25; 7: p Burne, R. A., Chen, Y. Y., Penders, J. E., 1997: Analysis of gene expression in Streptococcus mutans in biofilms in vitro; Adv. Dent. Res.; 11; 1: p Burne, R. A., Quivey, R. G., Jr., Marquis, R. E., 1999: Physiologic homeostasis and stress responses in oral biofilms; Methods Enzymol.; 310: p Butterfield, P. W., Bargmeyer, A. M., Camper, A. K., Biederman, J. A., 2002: Modified enzyme activity assay to determine biofilm biomass; J. Microbiol. Methods; 50: p Ceri, H., Olson, M. E., Stremick, C., Read, R. R., Morck, D., Buret, A., 1999: The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms; J. Clin. Microbiol.; 37; 6 : p
59 30. Ceri, H., Olson, M., Morck, D., Storey, D., Read, R., Buret, A., Olson, B., 2001: The MBEC Assay System: multiple equivalent biofilms for antibiotic and biocide susceptibility testing; Methods Enzymol.; 337: p Cheung, A. L., Fischetti, V. A., 1988: Variation in the expression of cell wall proteins of Staphylococcus aureus grown on solid and liquid media; Infection and Immunity; 56; 5: p Cheung, A. L., Schmidt, K., Bateman, B., Manna, A. C., 2001: SarS, a SarA homolog repressible by agr, is an activator of protein A synthesis in Staphylococcus aureus; Infection and Immunity; 69; 4: p Christensen, G. D., Simpson, W. A., Bisno, A. L., Beachey, E. H., 1982: Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces; Infection and Immunity; 37; 1: p Christensen, G. D., Simpson, W. A., Younger, J. J., Baddour, L. M., Barrett, F. F., Melton, D. M., Beachey, E. H., 1985: Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices; Journal of Clinical Microbiology; 22; 6: p Christensen, B. B., Sternberg, C., Andersen, J. B., Eberl, L., Moller, S., Givskov, M., Molin, S., 1998: Establishment of new genetic traits in a microbial biofilm community; Appl. Environ. Microbiol.; 64; 6: p Cochran, W. L., McFeters, G. A., Stewart, P. S., 2000: Reduced susceptibility of thin Pseudomonas aeruginosa biofilms to hydrogen peroxide and monochloramine; J. Appl. Microbiol.; 88; 1: p Costerton, J. W., Lewandowski, Z., Caldwell, D. E., Korber, D. R., Lappin-Scott, H. M., 1995: Microbial biofilms; Annu. Rev. Microbiol.; 49: p Costerton, J. W., 1999: Introduction to biofilm; International Journal of Antimicrobial Agents; 11; 3-4: p Costerton, J. W., Stewart, P. S., Greenberg, E. P., 1999: Bacterial biofilms: a common cause of persistent infections; Science; 284; 5418 : p Cozens, R. M., Tuomanen, E., Tosch, W., Zak, O., Suter, J., Tomasz, A., 1986: Evaluation of the bactericidal activity of beta-lactam antibiotics on slowly growing bacteria cultured in the chemostat; Antimicrobial Agents and Chemotherapy; 29; 5: p Cozzaglio, L., Bozzetti, F., Bonfanti, G., Viola, G., 1997: Sonication: a useful adjunct to the microbial assessment of central venous catheters; Nutrition; 13; 1: p Cramton, S. E., Gerke, C., Schnell, N. F., Nichols, W. W., Gotz, F., 1999: The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation; Infection and Immunity; 67; 10: p Cramton, S. E., Gerke, C., Gotz, F., 2001: In vitro methods to study staphylococcal biofilm formation; Methods Enzymol.; 336: p Cucarella, C., Solano, C., Valle, J., Amorena, B., Lasa, I., Penades, J. R., 2001: Bap, a Staphylococcus aureus surface protein involved in biofilm formation; Journal of Bacteriology; 183; 9: p Da Re, S., Ghigo, J. M., 2006: A CsgD-independent pathway for cellulose production and biofilm formation in Escherichia coli; J.Bacteriol.; 188; 8: p Danese, P. N., Pratt, L. A., Kolter, R., 2001: Biofilm formation as a developmental process; Methods Enzymol.; 336: p
60 47. Davey, M. E., O'Toole, G. A., 2000: Microbial biofilms: from ecology to molecular genetics; Microbiol. Mol. Biol. Rev.; 64; 4: p Davies, D. G., Parsek, M. R., Pearson, J. P., Iglewski, B. H., Costerton, J. W., Greenberg, E. P., 1998: The involvement of cell-to-cell signals in the development of a bacterial biofilm; Science; 280; 5361: p de Beer, D., Srinivasan, R., Stewart, P. S., 1994: Direct measurement of chlorine penetration into biofilms during disinfection; Appl. Environ. Microbiol.; 60; 12: p de Beer, D., Schramm, A., Santegoeds, C. M., Kuhl, M., 1997: A Nitrite Microsensor for Profiling Environmental Biofilms; Appl. Environ. Microbiol.; 63; 3: p de Beer, Dirk and Stoodley, Paul, 1999: Microbial biofilms; [online]; Heidelberg; Springer Verlag; [cited ]; last modified ; available at: de Nicolas, M. M., Vindel, A., offez-nieto, J. A., 1995: Epidemiological typing of clinically significant strains of coagulase-negative staphylococci; Journal of Hospital Infection; 29; 1: p de Paoli, P., 2005: Bio-banking in microbiology: from sample collection to epidemiology, diagnosis and research; FEMS Microbiol. Rev.; 29 ; 5: p Deighton, M. A., Balkau, B., 1990: Adherence measured by microtiter assay as a virulence marker for Staphylococcus epidermidis infections; Journal of Clinical Microbiology; 28; 11: p Deighton, M. A., Capstick, J., Borland, R., 1992: A study of phenotypic variation of Staphylococcus epidermidis using Congo red agar; Epidemiology and Infection ; 109; 3: p Deighton, M. A., Capstick, J., Domalewski, E., van Nguyen, T., 2001: Methods for studying biofilms produced by Staphylococcus epidermidis; Methods Enzymol.; 336: p Dobinsky, S., Kiel, K., Rohde, H., Bartscht, K., Knobloch, J. K., Horstkotte, M. A., Mack, D., 2003: Glucose-related dissociation between icaadbc transcription and biofilm expression by Staphylococcus epidermidis: evidence for an additional factor required for polysaccharide intercellular adhesin synthesis; J. Bacteriol.; 185; 9: p Donlan, R. M., 2000: Role of biofilms in antimicrobial resistance; ASAIO J.; 46; 6: p. S47-S Donlan, R. M., 2001: Biofilm formation: a clinically relevant microbiological process; Clin. Infect. Dis.; 33; 8: p Donlan, R. M., Costerton, J. W., 2002: Biofilms: survival mechanisms of clinically relevant microorganisms; Clin. Microbiol. Rev.; 15; 2: p Drenkard, E., Ausubel, F. M., 2002: Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation; Nature; 416; 6882: p Duguid, I. G., Evans, E., Brown, M. R., Gilbert, P., 1992: Growth-rate-independent killing by ciprofloxacin of biofilm-derived Staphylococcus epidermidis; evidence for cell-cycle dependency; Journal of Antimicrobial Chemotherapy; 30; 6: p Dunne, W. M., Jr., Sheth, N. K., Franson, T. R., 1987: Quantitative epifluorescence assay of adherence of coagulase-negative staphylococci; Journal of Clinical Microbiology; 25; 4: p
61 64. Dunne, W. M., Jr., Mason, E. O., Jr., Kaplan, S. L., 1993: Diffusion of rifampin and vancomycin through a Staphylococcus epidermidis biofilm; Antimicrobial Agents and Chemotherapy; 37; 12: p Dworniczek, E., Kuzko, K., Mroz, E., Wojciech, L., Adamski, R., Sobieszczanska, B., Seniuk, A., 2003: Virulence factors and in vitro adherence of Enterococcus strains to urinary catheters; Folia Microbiol. (Praha); 48; 5: p Ensing, G. T., Roeder, B. L., Nelson, J. L., van Horn, J. R., van der Mei, H. C., Busscher, H. J., Pitt, W. G., 2005: Effect of pulsed ultrasound in combination with gentamicin on bacterial viability in biofilms on bone cements in vivo; J. Appl. Microbiol.; 99; 3: p Ensing, G. T., Neut, D., van Horn, J. R., van der Mei, H. C., Busscher, H. J., 2006: The combination of ultrasound with antibiotics released from bone cement decreases the viability of planktonic and biofilm bacteria: an in vitro study with clinical strains; J. Antimicrob. Chemother.; 58; 6: p Everaert, E. P., Belt-Gritter, B., van der Mei, H. C., Busscher, H. J., Verkerke, G. J., Dijk, F., Mahieu, H. F., Reitsma, A., 1998: In vitro and in vivo microbial adhesion and growth on argon plasma-treated silicone rubber voice prostheses; J. Mater. Sci. Mater. Med.; 9; 3: p Fitzpatrick, F., Humphreys, H., Smyth, E., Kennedy, C. A., O'Gara, J. P., 2002: Environmental regulation of biofilm formation in intensive care unit isolates of Staphylococcus epidermidis; J. Hosp. Infect.; 52; 3: p Fitzpatrick, F., Humphreys, H., O'Gara, J. P., 2005: Evidence for icaadbc-independent biofilm development mechanism in methicillin-resistant Staphylococcus aureus clinical isolates; J. Clin. Microbiol.; 43; 4: p Fowler, V. G., Jr., Fey, P. D., Reller, L. B., Chamis, A. L., Corey, G. R., Rupp, M. E., 2001: The intercellular adhesin locus ica is present in clinical isolates of Staphylococcus aureus from bacteremic patients with infected and uninfected prosthetic joints; Med. Microbiol. Immunol. (Berl); 189; 3: p Fox, L. K., Zadoks, R. N., Gaskins, C. T., 2005: Biofilm production by Staphylococcus aureus associated with intramammary infection; Vet. Microbiol.; 107; 3-4: p Frebourg, N. B., Lefebvre, S., Baert, S., Lemeland, J. F., 2000: PCR-Based assay for discrimination between invasive and contaminating Staphylococcus epidermidis strains; Journal of Clinical Microbiology; 38; 2: p Freeman, D. J., Falkiner, F. R., Keane, C. T., 1989: New method for detecting slime production by coagulase negative staphylococci; J. Clin. Pathol.; 42; 8: p Fux, C. A., Costerton, J. W., Stewart, P. S., Stoodley, P., 2005: Survival strategies of infectious biofilms; Trends Microbiol.; 13; 1: p Garcez, A. S., Ribeiro, M. S., Tegos, G. P., Nunez, S. C., Jorge, A. O., Hamblin, M. R., 2006: Antimicrobial photodynamic therapy combined with conventional endodontic treatment to eliminate root canal biofilm infection; Lasers Surg. Med.; 39; 1: p Gelosia, A., Baldassarri, L., Deighton, M., van Nguyen, T., 2001: Phenotypic and genotypic markers of Staphylococcus epidermidis virulence; Clin. Microbiol. Infect.; 7; 4: p Gilbert, P., Das, J., Foley, I., 1997: Biofilm susceptibility to antimicrobials; Adv. Dent. Res.; 11; 1: p Halliman, D. G., Ahearn, D. G., 2004: Relative susceptibilities to vancomycin and quinupristin-dalfopristin of adhered and planktonic vancomycin-resistant and
62 vancomycin-susceptible coagulase-negative staphylococci; Curr. Microbiol.; 48; 3: p Handke, L. D., Conlon, K. M., Slater, S. R., Elbaruni, S., Fitzpatrick, F., Humphreys, H., Giles, W. P., Rupp, M. E., Fey, P. D., O'Gara, J. P., 2004: Genetic and phenotypic analysis of biofilm phenotypic variation in multiple Staphylococcus epidermidis isolates; J. Med. Microbiol.; 53; Pt 5: p Hausner, M., Wuertz, S., 1999: High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis; Appl. Environ. Microbiol.; 65; 8: p Hawser, S. P., Douglas, L. J., 1994: Biofilm formation by Candida species on the surface of catheter materials in vitro; Infection and Immunity; 62; 3: p Hawser, S. P., Douglas, L. J., 1995: Resistance of Candida albicans biofilms to antifungal agents in vitro; Antimicrobial Agents and Chemotherapy; 39; 9: p Hawser, S. P., Norris, H., Jessup, C. J., Ghannoum, M. A., 1998: Comparison of a 2,3- bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)carbonyl]-2h-t etrazolium hydroxide (XTT) colorimetric method with the standardized National Committee for Clinical Laboratory Standards method of testing clinical yeast isolates for susceptibility to antifungal agents; Journal of Clinical Microbiology; 36; 5: p Hefford, M. A., D'Aoust, S., Cyr, T. D., Austin, J. W., Sanders, G., Kheradpir, E., Kalmokoff, M. L., 2005: Proteomic and microscopic analysis of biofilms formed by Listeria monocytogenes 568; Can. J. Microbiol.; 51; 3: p Heilmann, C., Schweitzer, O., Gerke, C., Vanittanakom, N., Mack, D., Gotz, F., 1996: Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis; Molecular Microbiology ; 20; 5: p Holá, V., Růžička, F., Tejkalova, R., Votava, M., 2004a: Stanovení citlivosti k antibiotikům u biofilmpozitivních forem mikroorganizmů; Klin. Mikrobiol. Infekc. Lek.; 10; 5: p Holá, V., Růžička, F., Votava, M., 2004b: Rozdíly v citlivosti k antibiotikům u biofilmpozitivních a biofilmnegativních kmenů Staphylococcus epidermidis izolovaných z hemokultur; Epidemiol. Mikrobiol. Imunol.; 53; 2: p Holá, V., Růžička, F., Votava, M., 2004c: Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis; New Microbiol.; 27; 3: p Holá, V., Růžička, F., Votava, M., 2004d: Využití mikroskopických technik k průkazu a při studiu dynamiky růstu biofilmu; p Holá, V., Růžička, F., Votava, M., 2006: Dynamics of Staphylococcus epidermidis biofilm formation in relation to nutrition, temperature and time; Scripta Medica (Brno); 79; 3: p Horká, M., Růžička, F., Holá, V., Šlais, K., 2005: Dynamic modification of microorganisms by pyrenebutanoate for fluorometric detection in capillary zone electrophoresis; Electrophoresis; 26; 3: p Horká, M., Růžička, F., Holá, V., Šlais, K., 2006: Capillary isoelectric focusing of microorganisms in the ph range 2-5 in a dynamically modified FS capillary with UV detection; Anal. Bioanal. Chem.; 385; 5: p Hsu, J. C., Nelson, B., 1998: Multiple comparisons in the general linear model; Journal of Computational and Graphical Statistics; 7; 1: p
63 95. Inglis, T. J., Lim, T. M., Ng, M. L., Tang, E. K., Hui, K. P., 1995: Structural features of tracheal tube biofilm formed during prolonged mechanical ventilation; Chest; 108; 4: p Ishida, H., Ishida, Y., Kurosaka, Y., Otani, T., Sato, K., Kobayashi, H., 1998: In vitro and in vivo activities of levofloxacin against biofilm-producing Pseudomonas aeruginosa; Antimicrobial Agents and Chemotherapy; 42; 7: p Jain, S., Chen, J., 2006: Antibiotic resistance profiles and cell surface components of Salmonellae; J. Food Prot.; 69; 5: p Kennedy, C. A., O'Gara, J. P., 2004: Contribution of culture media and chemical properties of polystyrene tissue culture plates to biofilm development by Staphylococcus aureus; J. Med. Microbiol.; 53; Pt 11: p Keren, I., Shah, D., Spoering, A., Kaldalu, N., Lewis, K., 2004: Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli; J. Bacteriol.; 186; 24: p Kiers, P. J., Bos, R., van der Mei, H. C., Busscher, H. J., 2001: The electrophoretic softness of the surface of Staphylococcus epidermidis cells grown in a liquid medium and on a solid agar; Microbiology.; 147; Pt 3: p Knobloch, J. K., Horstkotte, M. A., Rohde, H., Mack, D., 2002a: Evaluation of different detection methods of biofilm formation in Staphylococcus aureus; Med. Microbiol. Immunol. (Berl); 191; 2: p Knobloch, J. K., Von Osten, H., Horstkotte, M. A., Rohde, H., Mack, D., 2002b: Minimal attachment killing (MAK): a versatile method for susceptibility testing of attached biofilm-positive and -negative Staphylococcus epidermidis; Med. Microbiol. Immunol. (Berl); 191; 2: p Kobayashi, N., Bauer, T. W., Tuohy, M. J., Fujishiro, T., Procop, G. W., 2006: Brief Ultrasonication Improves Detection of Biofilm-formative Bacteria Around a Metal Implant; Clin. Orthop. Relat. Res.; 457: Kolenbrander, P. E., Palmer, R. J., Jr., Rickard, A. H., Jakubovics, N. S., Chalmers, N. I., Diaz, P. I., 2000: Bacterial interactions and successions during plaque development; Periodontol.; 42: p Kopec, L. K., Vacca Smith, A. M., Wunder, D., Ng-Evans, L., Bowen, W. H., 2002: Influence of antibody on the structure of glucans; Caries Res.; 36; 2: p Kučera, F., Jancar, J., 1996: Preliminary study of sulfonation of polystyrene by homogeneous and heterogeneous reaction; Chemical Papers-Chemicke Zvesti; 50; 4: p Kuhn, D. M., George, T., Chandra, J., Mukherjee, P. K., Ghannoum, M. A., 2002: Antifungal susceptibility of Candida biofilms: unique efficacy of amphotericin B lipid formulations and echinocandins; Antimicrobial Agents and Chemotherapy; 46; 6: p Lawrence, J. R., Neu, T. R., 1999: Confocal laser scanning microscopy for analysis of microbial biofilms; Methods Enzymol.; 310: ; p Leid, J. G., Willson, C. J., Shirtliff, M. E., Hassett, D. J., Parsek, M. R., Jeffers, A. K., 2005: The exopolysaccharide alginate protects Pseudomonas aeruginosa biofilm bacteria from IFN-gamma-mediated macrophage killing; J. Immunol.; 175; 11: p
64 110. Leone, S., Molinaro, A., Alfieri, F., Cafaro, V., Lanzetta, R., Di Donato, A., Parrilli, M., 2006: The biofilm matrix of Pseudomonas sp. OX1 grown on phenol is mainly constituted by alginate oligosaccharides; Carbohydr. Res.; 341; 14: p Lewis, K., 2001: Riddle of biofilm resistance; Antimicrob. Agents Chemother.; 45; 4: p Li, Y., Burne, R. A., 2001: Regulation of the gtfbc and ftf genes of Streptococcus mutans in biofilms in response to ph and carbohydrate; Microbiology; 147; Pt 10: p Licking E., 1999: Getting a grip on bacterial slime; Bussiness Week; 13; 9: p Liu, X., Ng, C., Ferenci, T., 2000: Global adaptations resulting from high population densities in Escherichia coli cultures; Journal of Bacteriology; 182; 15: p Lucent Technologies Inc., 1988: S-Plus 4.0 Release 2 for Windows [CD ROM]; version 4.0; Halcyon; MathSoft, Inc Macias, A. E., Ponce-de-Leon, S., 2005: Infection control: old problems and new challenges ; Arch. Med. Res.; 36; 6: p Mack, D., Bartscht, K., Fischer, C., Rohde, H., de Grahl, C., Dobinsky, S., Horstkotte, M. A., Kiel, K., Knobloch, J. K., 2001: Genetic and biochemical analysis of Staphylococcus epidermidis biofilm accumulation; Methods Enzymol.; p Mack, D., Becker, P., Chatterjee, I., Dobinsky, S., Knobloch, J. K., Peters, G., Rohde, H., Herrmann, M., 2004: Mechanisms of biofilm formation in Staphylococcus epidermidis and Staphylococcus aureus: functional molecules, regulatory circuits, and adaptive responses; Int. J. Med. Microbiol.; 294; 2-3: p Mah, T. F., O'toole, G. A., 2001: Mechanisms of biofilm resistance to antimicrobial agents; Trends Microbiol.; 9 ; 1: p Maira-Litran, T., Allison, D. G., Gilbert, P., 2000a: An evaluation of the potential of the multiple antibiotic resistance operon (mar) and the multidrug efflux pump acrab to moderate resistance towards ciprofloxacin in Escherichia coli biofilms; J. Antimicrob. Chemother.; 45; 6: p Maira-Litran, T., Allison, D. G., Gilbert, P., 2000b: Expression of the multiple antibiotic resistance operon (mar) during growth of Escherichia coli as a biofilm; J. Appl. Microbiol.; 88; 2: p Maki, D. G., Weise, C. E., Sarafin, H. W., 1977: A semiquantitative culture method for identifying intravenous-catheter-related infection; N. Engl. J. Med.; 296; 23: p Marsh, P. D., 2004: Dental plaque as a microbial biofilm; Caries Res.; 38; 3: p Marsh, P. D., 2006: Dental plaque as a biofilm and a microbial community - implications for health and disease; BMC Oral Health; 6 Suppl 1: p. S Martineau, F., Picard, F. J., Roy, P. H., Ouellette, M., Bergeron, M. G., 1996: Speciesspecific and ubiquitous DNA-based assays for rapid identification of Staphylococcus epidermidis; Journal of Clinical Microbiology; 34; 12: p Mathur, T., Singhal, S., Khan, S., Upadhyay, D. J., Fatma, T., Rattan, A., 2006: Detection of biofilm formation among the clinical isolates of Staphylococci: an evaluation of three different screening methods; Indian J. Med. Microbiol.; 24; 1: p McAteer, J., Davies, J. M., 2002: The culture of attached cells; in Davies, J. M.; Basic Cell Culture: a Practical Approach; Oxford; Oxford University Press; p
65 128. McDowell, A., Patrick, S., 2005: Evaluation of nonculture methods for the detection of prosthetic hip biofilms; Clin. Orthop. Relat. Res.; 437: p Monzon, M., Oteiza, C., Leiva, J., Lamata, M., Amorena, B., 2002: Biofilm testing of Staphylococcus epidermidis clinical isolates: low performance of vancomycin in relation to other antibiotics; Diagn. Microbiol. Infect. Dis.; 44; 4: p Moretro, T., Hermansen, L., Holck, A. L., Sidhu, M. S., Rudi, K., Langsrud, S., 2003: Biofilm formation and the presence of the intercellular adhesion locus ica among staphylococci from food and food processing environments; Appl.Environ.Microbiol.; 69; 9: p Mulder, J. G., Degener, J. E., 1998: Slime-producing properties of coagulase-negative staphylococci isolated from blood cultures; Clin. Microbiol. Infect.; 4; 12: p Nguyen, L. L., Nelson, C. L., Saccente, M., Smeltzer, M. S., Wassell, D. L., McLaren, S. G., 2002: Detecting bacterial colonization of implanted orthopaedic devices by ultrasonication; Clin. Orthop. Relat. Res.; 403: p O'Gara, J. P., Humphreys, H., 2001: Staphylococcus epidermidis biofilms: importance and implications; J. Med. Microbiol.; 50; 7: p O'Toole, G., Kaplan, H. B., Kolter, R., 2000: Biofilm formation as microbial development; Annu. Rev. Microbiol.; 54: p Otto, K., Silhavy, T. J., 2002: Surface sensing and adhesion of Escherichia coli controlled by the Cpx-signaling pathway; Proc. Natl. Acad. Sci. USA; 99; 4: p Ozerdem, A. N., Elci, S., Atmaca, S., Akbayin, H., Gul, K., 2003: The effects of magnesium, calcium and EDTA on slime production by Staphylococcus epidermidis strains; Folia Microbiol. (Praha); 48; 5: p Palmer, R. J., Jr., Sternberg, C., 1999: Modern microscopy in biofilm research: confocal microscopy and other approaches; Curr. Opin. Biotechnol.; 10; 3: p Patel, R., 2005: Biofilms and antimicrobial resistance; Clin. Orthop. Relat. Res.; 437: p Paterson, R. L., Webster, N. R., 2000: Sepsis and the systemic inflammatory response syndrome; J. R. Coll. Surg. Edinb.; 45; 3: p Perez-Giraldo, C., Rodriguez-Benito, A., Moran, F. J., Hurtado, C., Blanco, M. T., Gomez-Garcia, A. C., 1995: Influence of the incubation atmosphere on the production of slime by Staphylococcus epidermidis; Eur. J. Clin. Microbiol. Infect. Dis.; 14; 4: p R Development Core Team, 2005: R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; Vienna, Austria; ISBN Raad, I., 1998: Intravascular-catheter-related infections; Lancet; 351; 9106: p Raad, I., Hanna, H., Dvorak, T., Chaiban, G., Hachem, R., 2007: Optimal antimicrobial catheter lock solution, using different combinations of minocycline, EDTA, and 25- percent ethanol, rapidly eradicates organisms embedded in biofilm; Antimicrob. Agents Chemother.; 51; 1: p Reisner, A., Holler, B. M., Molin, S., Zechner, E. L., 2006: Synergistic effects in mixed Escherichia coli biofilms: conjugative plasmid transfer drives biofilm expansion; J. Bacteriol.; 188; 10 : p Roberts, A. P., Pratten, J., Wilson, M., Mullany, P., 1999: Transfer of a conjugative transposon, Tn5397 in a model oral biofilm; FEMS Microbiol. Lett.; 177; 1: p
66 146. Robinson, R. W., Akin, D. E., Nordstedt, R. A., Thomas, M. V., Aldrich, H. C., 1984: Light and Electron Microscopic Examinations of Methane-Producing Biofilms from Anaerobic Fixed-Bed Reactors; Appl. Environ. Microbiol.; 48; 1: p Rohde, H., Knobloch, J. K., Horstkotte, M. A., Mack, D., 2001: Correlation of Staphylococcus aureus icaadbc genotype and biofilm expression phenotype; Journal of Clinical Microbiology; 39; 12: p Růžička, F., Holá, V., Votava, M., Tejkalová, R., Horvath, R., Heroldová, M., Woznicová, V., 2004: Biofilm detection and the clinical significance of Staphylococcus epidermidis isolates; Folia Microbiol. (Praha); 49; 5: p Růžička, F., Holá, V., Votava, M., 2006a: Možnosti průkazu tvorby biofilmu v rutinní mikrobiologické praxi; Epidemiol. Mikrobiol. Imunol.; 55; 1: p Růžička, F., Horka, M., Holá, V., Votava, M., 2006b: Capillary Isoelectric Focusing - Useful tool for detection of the biofilm formation in Staphylococcus epidermidis; J. Microbiol. Methods; 68: Sanger, J. R., Sheth, N. K., Franson, T. R., 1989: Adherence of microorganisms to breast prostheses: an in vitro study; Ann. Plast. Surg.; 22; 4: p Schierholz, J. M., Beuth, J., 2001: Implant infections: a haven for opportunistic bacteria; Journal of Hospital Infection; 49; 2: p Seenivasan, M. H., Yu, V. L., Muder, R. R., 2005: Legionnaires' disease in long-term care facilities: overview and proposed solutions; J. Am. Geriatr. Soc.; 53; 5: p Sheehan, E., McKenna, J., Mulhall, K. J., Marks, P., McCormack, D., 2004: Adhesion of Staphylococcus to orthopaedic metals, an in vivo study; J. Orthop. Res.; 22; 1: p Sherertz, R. J., Raad, I. I., Belani, A., Koo, L. C., Rand, K. H., Pickett, D. L., Straub, S. A., Fauerbach, L. L., 1990: Three-year experience with sonicated vascular catheter cultures in a clinical microbiology laboratory; Journal of Clinical Microbiology; 28; 1: p Shigeta, M., Komatsuzawa, H., Sugai, M., Suginaka, H., Usui, T., 1997: Effect of the growth rate of Pseudomonas aeruginosa biofilms on the susceptibility to antimicrobial agents; Chemotherapy; 43; 2: p Simmons, P. A., Tomlinson, A., Seal, D. V., 1998: The role of Pseudomonas aeruginosa biofilm in the attachment of Acanthamoeba to four types of hydrogel contact lens materials; Optom. Vis. Sci.; 75; 12: p Singh, P. K., Schaefer, A. L., Parsek, M. R., Moninger, T. O., Welsh, M. J., Greenberg, E. P., 2000: Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms; Nature; 407; 6805: p Souli, M., Giamarellou, H., 1998: Effects of slime produced by clinical isolates of coagulase-negative staphylococci on activities of various antimicrobial agents; Antimicrob. Agents Chemother.; 42; 4: p Spoering, A. L., Lewis, K., 2001: Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials; Journal of Bacteriology; 183; 23: p Stanley, N. R., Britton, R. A., Grossman, A. D., Lazazzera, B. A., 2003: Identification of catabolite repression as a physiological regulator of biofilm formation by Bacillus subtilis by use of DNA microarrays; J. Bacteriol.; 185; 6: p
67 162. Stepanovic, S., Vukovic, D., Dakic, I., Savic, B., Svabic-Vlahovic, M., 2000: A modified microtiter-plate test for quantification of staphylococcal biofilm formation; J. Microbiol. Methods; 40; 2: p Stepanovic, S., Vukovic, D., Jezek, P., Pavlovic, M., Svabic-Vlahovic, M., 2001: Influence of dynamic conditions on biofilm formation by staphylococci; Eur. J. Clin. Microbiol. Infect. Dis.; 20; 7: p Stepanovic, S., Djukic, N., Opavski, N., Djukic, S., 2003: Significance of inoculum size in biofilm formation by staphylococci; New Microbiol.; 26; 1: p Stewart, P. S., Murga, R., Srinivasan, R., De Beer, D., 1995: Biofilm structural heterogeneity visualized by three microscopic methods; Water Research; 29; 8: p Stewart, P. S., Costerton, J. W., 2001: Antibiotic resistance of bacteria in biofilms; Lancet; 358; 9276: p Stoodley, P., Lewandowski, Z., Boyle, J. D., Lappin-Scott, H. M., 1998: Oscillation characteristics of biofilm streamers in turbulent flowing water as related to drag and pressure drop; Biotechnol. Bioeng.; 57; 5: p Stoodley, P., Sauer, K., Davies, D. G., Costerton, J. W., 2002: Biofilms as complex differentiated communities; Annu. Rev. Microbiol.; 56: Surman, S. B., Walker, J. T., Goddard, D. T., Morton, L. H. G., Keevil, C. W., Weaver, W., Skinner, A., Hanson, K., Caldwell, D., Kurtz, J., 1996: Comparison of microscope techniques for the examination of biofilms; Journal of Microbiological Methods; 25; 1: p Svensater, G., Welin, J., Wilkins, J. C., Beighton, D., Hamilton, I. R., 2001: Protein expression by planktonic and biofilm cells of Streptococcus mutans; FEMS Microbiol. Lett.; 205; 1: p Takenaka, S., Iwaku, M., Hoshino, E., 2001: Artificial Pseudomonas aeruginosa biofilms and confocal laser scanning microscopic analysis; J. Infect. Chemother.; 7; 2: p Tormo, M. A., Marti, M., Valle, J., Manna, A. C., Cheung, A. L., Lasa, I., Penades, J. R., 2005: SarA is an essential positive regulator of Staphylococcus epidermidis biofilm development; J. Bacteriol.; 187; 7: p Trampuz, A., Widmer, A. F., 2006: Infections associated with orthopedic implants; Curr. Opin. Infect. Dis.; 19; 4: p Tremoulet, F., Duche, O., Namane, A., Martinie, B., Labadie, J. C., 2002: A proteomic study of Escherichia coli O157:H7 NCTC cultivated in biofilm or in planktonic growth mode; FEMS Microbiol. Lett.; 215; 1: p Tuomanen, E., Cozens, R., Tosch, W., Zak, O., Tomasz, A., 1986: The rate of killing of Escherichia coli by beta-lactam antibiotics is strictly proportional to the rate of bacterial growth; Journal of General Microbiology; 132; 5: p Urbášková, P., 1998: Rezistence bakterií k antibiotikům. Vybrané metody; Praha; TRIOS; 100 pp Vandecasteele, S. J., Peetermans, W. E., Merckx, R., Rijnders, B. J., Van Eldere, J., 2003: Reliability of the ica, aap and atle genes in the discrimination between invasive, colonizing and contaminant Staphylococcus epidermidis isolates in the diagnosis of catheter-related infections; Clin. Microbiol. Infect.; 9; 2: p
68 178. Vasudevan, P., Nair, M. K., Annamalai, T., Venkitanarayanan, K. S., 2003: Phenotypic and genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for biofilm formation; Vet. Microbiol.; 92; 1-2: p Vianney, A., Jubelin, G., Renault, S., Dorel, C., Lejeune, P., Lazzaroni, J. C., 2005: Escherichia coli tol and rcs genes participate in the complex network affecting curli synthesis; Microbiology; 151; Pt 7: p Vogel, L., Sloos, J. H., Spaargaren, J., Suiker, I., Dijkshoorn, L., 2000: Biofilm production by Staphylococcus epidermidis isolates associated with catheter related bacteremia; Diagn. Microbiol. Infect. Dis.; 36; 2: p von Eiff, C., Proctor, R. A., Peters, G., 2001: Coagulase-negative staphylococci. Pathogens have major role in nosocomial infections; Postgraduate Medicine; 110; 4: p , Vrany, J. D., Stewart, P. S., Suci, P. A., 1997: Comparison of recalcitrance to ciprofloxacin and levofloxacin exhibited by Pseudomonas aeruginosa bofilms displaying rapid-transport characteristics; Antimicrobial Agents and Chemotherapy; 41; 6: p Vuong, C., Saenz, H. L., Gotz, F., Otto, M., 2000: Impact of the agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus; Journal of Infectious Diseases; 182; 6: p Wagman, G. H., Bailey, J. V., Weinstein, M. J., 1975: Binding of aminoglycoside antibiotics to filtration materials; Antimicrobial Agents and Chemotherapy; 7; 3: p Walencka, E., Sadowska, B., Rozalska, S., Hryniewicz, W., Rozalska, B., 2005: Lysostaphin as a potential therapeutic agent for staphylococcal biofilm eradication; Pol. J. Microbiol.; 54; 3: p Walencka, E., Sadowska, B., Rozalska, S., Hryniewicz, W., Rozalska, B., 2006: Staphylococcus aureus biofilm as a target for single or repeated doses of oxacillin, vancomycin, linezolid and/or lysostaphin; Folia Microbiol. (Praha); 51; 5: p Wentland, E. J., Stewart, P. S., Huang, C. T., McFeters, G. A., 1996: Spatial variations in growth rate within Klebsiella pneumoniae colonies and biofilm; Biotechnol. Prog.; 12; 3: p Whiteley, M., Bangera, M. G., Bumgarner, R. E., Parsek, M. R., Teitzel, G. M., Lory, S., Greenberg, E. P., 2001: Gene expression in Pseudomonas aeruginosa biofilms; Nature; 413; 6858: p Williams, I., Venables, W. A., Lloyd, D., Paul, F., Critchley, I., 1997: The effects of adherence to silicone surfaces on antibiotic susceptibility in Staphylococcus aureus; Microbiology; 143; Pt 7: p Wunschel, D. S., Hill, E. A., Mclean, J. S., Jarman, K., Gorby, Y. A., Valentine, N., Wahl, K., 2005: Effects of varied ph, growth rate and temperature using controlled fermentation and batch culture on matrix assisted laser desorption/ionization whole cell protein fingerprints; J. Microbiol. Methods; 62; 3: p Yu, F. P., McFeters, G. A., 1994: Rapid in situ assessment of physiological activities in bacterial biofilms using fluorescent probes; J. Microbiol. Methods; 20: p Zaura-Arite, E., van Marle, J., ten Cate, J. M., 2001: Conofocal microscopy study of undisturbed and chlorhexidine-treated dental biofilm; J.Dent.Res.; 80; 5: p Zheng, Z., Stewart, P. S., 2002: Penetration of rifampin through Staphylococcus epidermidis biofilms; Antimicrobial Agents and Chemotherapy; 46; 3: p
69 194. Ziebuhr, W., Krimmer, V., Rachid, S., Lossner, I., Gotz, F., Hacker, J., 1999: A novel mechanism of phase variation of virulence in Staphylococcus epidermidis: evidence for control of the polysaccharide intercellular adhesin synthesis by alternating insertion and excision of the insertion sequence element IS256; Mol. Microbiol.; 32; 2: p Ziebuhr, W., Loessner, I., Krimmer, V., Hacker, J., 2001: Methods to detect and analyze phenotypic variation in biofilm-forming Staphylococci; Methods Enzymol.; 336: p
70 7. Předkládané práce
71 7.1 S. Stepanović, D. Vuković, V. Holá, G. Di Bonaventura, S. Djukić, I. Ćirković, F. Růžička (in press): Quantification of biofilm in microtiter-plate: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci; Acta Pathologica, Microbiologica et Immunologica Scandinavica (accepted for publication).
72 1 Quantification of biofilm in microtiter-plate: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci Running title: Quantification of biofilm in microtiter-plate Srdjan Stepanović, 1 Dragana Vuković, 1 Veronika Hola, 2 Giovanni Di Bonaventura, 3 Slobodanka Djukić, 1 Ivana Ćirković, 1 Filip Ruzicka 2 Department of Bacteriology, Institute of Microbiology and Immunology, School of Medicine, Belgrade, Serbia, 1 Institute for Microbiology, Faculty of Medicine, Masaryk University, Brno, Czech Republic, 2 and Laboratory of Clinical Microbiology, Aging Research Center Ce.S.I., G. D Annunzio University of Chieti-Pescara, Chieti, Italy 3 Summary The purpose of this paper is to provide details of all steps involved in the quantification of biofilm formation in microtiter-plate. The presented protocol incorporates information on assessment of biofilm production by staphylococci, gained both by direct experience as well as by analysis of methods for assaying biofilm production available in the literature. The presented data should make quantification of biofilm formation in microtiter-plate simple more reliable and comparable among different laboratories. Key words: biofilm; microtiter-plate; quantification; Staphylococcus Abbreviations: TSB - tryptic soy broth TSA - tryptic soy agar BHI - brain heart infusion broth OD - optical density Introduction Biofilm has been defined as an assemblage of microbial cells that is associated with a surface and enclosed in an extracellular matrix of principally polysaccharide material [Donlan, 2002]. Biofilm-associated organisms are fundamentally different from populations of suspended cells. Bacteria in biofilm display elevated resistance to antibiotics, disinfectants, as well as to host immune system clearance [Donlan, 2002; Donlan & Costerton, 2002]. The importance of biofilm has been well recognized in medical, environmental, and industrial contexts, and the study of microbial biofilms has received significant attention over the last decades. A number of methods have been developed for cultivation and quantification of biofilms [Deighton et al., 2001; Harraghy et al., 2006], but no standardized protocol for assessment of biofilm formation by different bacterial species has been established so far. Quantification of biofilms started with a method based on the cultivation of biofilm on the wall of a test tube and subsequent detection by stain for biofilm recognition [Christensen et al., 1982]. Later, the wells of microtiter-plate, as culture vessel, were used, and the results were measured
73 2 spectrophotometrically [Christensen et al., 1985]. Currently a number of different methods are used such as tube test, microtiter-plate test, radiolabeling, microscopy, etc. [Harraghy et al., 2006]. Nevertheless, the microtiter-plate method remains among the most frequently used assays for investigation of biofilm, and meanwhile a number of its modifications have been developed for the in vitro cultivation and quantification of bacterial biofilms. Staphylococci are the most common causative agents of medical device-associated infections. The crucial step in the pathogenesis of these infections is the formation of stable biofilm on the surface of the implanted biomedical device [Mack et al., 2004]. The ability to produce an adherent multilayered biofilm on implanted biomaterials is considered as an important virulence factor of staphylococci [Mack et al., 2004]. Biofilm production by staphylococci has been extensively investigated, particularly in S. epidermidis and S. aureus. The purpose of this paper is to present in a step-by-step manner all phases in quantification of biofilm formation by staphylococci in microtiter-plate, and to present the most important variations of the method. Testing conditions 1. Storage of bacterial strains for biofilm testing Preservation is important in the maintenance of bacteria for research applications. Freeze drying (i.e. lyophilization) and, more frequently, cryopreservation are used for bacteria isolates archiving [De Paoli, 2005]. For cryopreservation staphylococci are inoculated onto appropriate non-inhibitory medium (e.g. tryptic soy agar (TSA), blood agar) and incubated overnight aerobically at C. After verifying the purity of the strains, one colony is emulsified in appropriate broth (e.g. tryptic soy broth (TSB), brain heart infusion broth (BHI)). Following overnight incubation at C, the broth is vortexed and poured in suitable tube, and previously sterilized glycerol is aseptically added to reach final concentration of 10-15%. Alternatively, few colonies from the overnight incubated agar medium are emuslified in appropriate broth supplemented with 10-15% of glycerol to obtain suspension greater than 2 on the McFarland scale, and after labeling, immediately freezed [CA-SFM, 1996]. Bacteria can be stored at -20 C for one to three years, at -70 C for one to ten years, while freezing in liquid nitrogen preserves bacteria for up to 30 years [De Paoli, 2005]. Storage at -20 C should be used as a temporary measure only. After storage, a rapid thawing of cultures and a quick transfer of bacteria to an appropriate growth medium is recommended [De Paoli, 2005]. Experimental infections using stored bacteria have shown that virulence properties are usually maintained after lyophilization or freezing at -70 C [De Paoli, 2005]. However, after storage and thawing it was shown that some fastidious bacteria may pose problems in terms of viability and stability of their antigenic, molecular and biochemical properties [De Paoli, 2005]. After retrival from storage, the pure culture of staphylococci may contain a mixture of phenotypes that differ in their ability to form biofilm (see below for phase variations). 2. Inoculum
74 3 Prior to inoculation tested strains may be cultivated either in a broth or on a solid medium. Incubation of staphylococci in a liquid medium is more frequently used. The strain is transferred from the stock culture onto appropriate non-inhibitory medium (i.e. TSA or blood agar) and incubated overnight aerobically at C. After verifying purity of the strain, three to four well-isolated identical colonies are suspended in 5 ml of TSB, and incubated without shaking for 18 h ± 30 min [Deighton et al., 2001]. After incubation, the stationary phase culture is vortexed and therafter diluted 1:2 to 1:1000 [Christensen et al., 1985; Baldassarri et al., 1993; Heilmann et al., 1996; Mulder & Degener, 1998; Allignet et al., 1999; Baldassarri et al., 2001; Cramton et al., 2001; Fowler et al., 2001; Mack et al., 2001; Ziebuhr et al., 2001; Moretro et al., 2003; Vasudevan et al., 2003; Tormo et al., 2005; Arciola et al., 2006; Mathur et al., 2006], but most frequently 1:100, in medium for bioflim cultivation (TSB supplemented with glucose, see below). The diluted bacteria are vortexed and then inoculated into a microtiter-plate (200 µl per well). Another approach, which is prefered in laboratories participating in this study, is to transfer the strain from the stock culture, prior to assaying for biofilm production, onto TSA, Mueller Hinton agar, or blood agar and to incubate overnight to 24 h aerobically at C. After verifying purity of the strain, one to few colonies with identical morphology are suspended into sterile distilled water or physiological saline. The turbidity of the bacterial suspension is adjusted to match turbidity comparable to that of the 0.5 McFarland standard (~10 8 CFU/ml). It is preferable to perform this step by using a photometric device. The obtained suspension should be vortexed for at least 1 minute. Although both above noted protocoles are acceptable, it is noteworthy that staphylococci grown on a solid medium differ in the expression of cell-associated molecules from staphylococci grown in a liquid medium [Cheung et al., 1988; Kiers et al., 2001]. Probably all staphylococci possess capsule or slime layers which are regarded as soft, polyelectrolyte layers surrounding the bacteria [Kiers et al., 2001]. The presence of such a layer decreases the energy barrier due to electrostatic repulsion in the interaction of the organisms with negatively charged substrata and, thus, plays an important role in their adhesion [Kiers et al., 2001]. The cell surface softness of S. epidermidis strains grown on agar medium increased sometimes by a factor of two, as comapared to the cell surface softness for the strains grown in liquid medium [Kiers et al., 2001]. Therefore, preparation of the inoculum from broth or agar growing staphylococci may directly influence the biofilm produced by these bacteria, since adhesion is the first step in biofilm formation. It should be noted that infecting bacteria are often surface-associated and their cell surface can, therefore, be expected to be more similar to that of bacteria grown on a solid medium than to that found in organisms grown in liquid media [Schleifer & Kroppenstedt, 1990]. Hence, the inoculum prepared from bacteria grown on agar probably more closely resembles in vivo situation, than that prepared from bacteria grown in broth. Irrespective of the method used for preparation of the inoculum, the inoculum size should be carefully determined. It has been shown that the size of the inoculum considerably influence the amount of biofilm produced, i.e. biofilm density increases with increasing initial inoculum [Deighton et al., 2001; Stepanovic et al., 2003c]. Thus, standardized inoculum should be used. However, in most studies dealing with biofilm quantification, the exact size of the inoculum has not been determined. We consider that an initial inoculum of ~10 8 CFU/ml is most suitable. The same inoculum was adopted as a standard in the well-known reference procedure for antimicrobial susceptibility testing, and procedures for obtainig such inoculum are well described [CA-SFM, 1996]. Before final dilution into a fresh broth
75 4 medium, the turbidity of the bacterial suspension should be adjusted to obtain turbidity optically comparable to that of the 0.5 McFarland standard (~10 8 CFU/ml). Subsequent 1:100 dilution of this suspension will result in the final testing inoculum of ~10 6 CFU. Another point of interest in preparing inocula is to avoid inoculation of preexisting cell clusters [Deighton et al., 2001], since they may lead to false positive results. Therefore, prepared cell suspensions must be vortexed. Alternatively, preexisting cell clusters in the testing suspensions may be broken up with a syringe fitted with a 23-gauge needle and brief vortexing [Deighton et al., 2001]. Still, some investigators consider that vortexing may not be sufficient to break up all preexisting cell clusters if the suspension is made from bacteria grown in standard TSB medium or bacteria grown on a solid medium [Mack et al., 2001]. Biofilm formation by staphylococci is enhanced in the presence of glucose and incubation of bacteria in a broth which provides optimal conditions for biofilm formation, i.e. TSB (standard TSB medium commonly contains 0.25% glucose), may give a bias within the test due to possible presence of preexisting cell clusters. Accordingly, the inoculated bacteria should optimaly be precultured in a broth which does not support biofilm formation by a given bacterial species. TSB lacking glucose is a broth which generaly does not support biofilm formation by staphylococci [Christensen et al., 1985; Dobinsky et al., 2003; Mulder & Degener, 1998]. Thus, it was recommended that staphylococci should be incubated in TSB medium without glucose or other sugars [Mack et al., 2001], and diluted in TSB with glucose (see below). However, standard TSB which contains glucose was used in great majority of previous studies and, moreover, availability of TSB without glucose may pose a problem for some laboratories. Although we are aware of limitations of vortexing, we propose it as an acceptable solution for elimination of preexisting cell clusters. When preparing the inocula, it is also important to be aware that phase variations involving biofilm formation are seen in staphylococci [Christensen et al., 1990; Ziebuhr et al., 1997; Ziebuhr et al., 1999; Deighton et al., 2001; Ziebuhr et al., 2001; Handke et al., 2004]. This means that upon subcultivation of single colony of a biofilm-forming strain biofilmnegative variants may occur [Ziebuhr et al., 1999; Ziebuhr et al., 2001]. These variations have been noted after serial passages or after retrieval from storage at -70 C [Ziebuhr et al., 1999; Deighton et al., 2001]. Presence of phase variations in staphylococci is easily achieved by observing colonial morphology on Congo red agar [Ziebuhr et al., 1997; Ziebuhr et al., 1999; Deighton et al., 2001; Ziebuhr et al., 2001; Handke et al., 2004]. If more than one colonial morphotype is present, the predominant one should be selected for further testing [Deighton et al., 2001]. Working cultures may be stored on appropriate medium (i.e. TSA or blood agar) at 4 C, but due to the phase variations, the number of subcultures made before testing must be kept to an absolute minimum [Deighton et al., 2001]. 3. Medium for biofilm cultivation Although a clearcut recommendation for single medium appropriate for testing biofilm formation by different staphylococcal species is difficult, our choice, based upon direct experience and the overall data, would be TSB (which commonly contains 0.25% glucose) supplemented with additional 1% glucose. Composition of the medium is probably the most important factor influencing the ability of bacteria to produce biofilm in in vitro conditions [Deighton et al., 2001; Mack et al., 2001; Rohde et al., 2001; Knobloch et al., 2002; Stepanovic et al., 2003b; Hola et al., 2004; Kennedy and O'Gara, 2004; Mathur et al., 2006]. This has been investigated in a number
76 5 of studies and the results showed BHI being usually better than TSB [Knobloch et al., 2002; Hola et al., 2004; Stepanovic et al., 2003b; Kennedy and O'Gara, 2004; Mathur et al., 2006], although it has also been shown that some strains of a given staphylococcal species produce more biofilm quantitites in TSB, while others in BHI [Knobloch et al., 2002]. Nevertheless, TSB is the medium most frequently used to grow staphylococcal biofilms [Christensen et al., 1985; Deighton and Balkau, 1990; Baldassarri et al., 1993; Perez- Giraldo et al., 1995; Heilmann et al., 1996; Ziebuhr et al., 1997; Mulder & Degener, 1998; Allignet et al., 1999; Stepanovic et al., 2000; Baldassarri et al., 2001; Deighton et al., 2001; Cramton et al., 2001; Mack et al., 2001; Rohde et al., 2001; Ziebuhr et al., 2001; Vasudevan et al., 2003]. Supplementation of the medium with glucose increases ability of staphylococci to form biofilm [Christensen et al., 1985; Deighton and Balkau, 1990; Mack et al., 1992; Mulder & Degener, 1998; Baldassarri et al., 2001; Knobloch et al., 2002; Dobinsky et al., 2003; Mathur et al., 2006]. Therefore, liquid medium (TSB or BHI) supplemented with additional 0.25% - 4% glucose [Baldassarri et al., 2001; Cramton et al., 2001; Deighton et al., 2001; Fowler et al., 2001; Gelosia et al., 2001; Knobloch et al., 2002; Hola et al., 2004; Hola et al., 2006; Fitzpatrick et al., 2005; Mathur et al., 2006], was frequently used. Subinhibitory antibiotic concentrations, ethanol, and NaCl also stimulate the production of biofilm [Knobloch et al., 2001; Cho et al., 2002; Fitzpatrick et al., 2002] and, accordingly, were also used as supplements to a basic medium. In addition, some investigators showed that TSB without additional glucose may not be useful for the detection of biofilm formation by S. aureus [Baldassarri et al., 2001; Knobloch et al., 2002]. To identify biofilm positive S. aureus isolates, usage of at least two media, TSB + 1% glucose and BHI + 2% glucose + 2% sucrose, was recommended [Knobloch et al., 2002]. Finally, it should be noted that significant variation in quantities of biofilm produced were detected by using the TSB medium manufactured by different suppliers [Mack et al., 2000; Mack et al., 2001; Rohde et al., 2001], as well as by using different lots of the same medium [Mack et al., 2000; Mack et al., 2001]. 4. Microtiter-plate The plates for biofilm cultivation should be sterile flat-bottomed 96-well polystyrene tissue culture treated microtiter-plate with a lid. The process of biofilm formation begins with initial attachment of bacteria to a surface which is affected by a number of factors, including the characteristics of a surface to which bacteria adhere. There is a number of different microtiter-plate types designed for different purposes. Microtiter-plate prepared for tissue culture implies that its surface was specially treated to allow an excellent cell attachment and proliferation. Some strains produce biofilm only on tissue culture plates, and many strains show weaker biofilm-positivity on non-tissue culture microtiter-plates [Christensen et al., 1985; Kennedy and O'Gara, 2004]. In addition, disruption and falling off in pieces of biofilm layer during washing procedure is lower in tissue culture-treated microtiter-plates. The bottom of microtiter-plate my be U-shape, V-shape or flat. Although in rare occasions U- bottomed microtiter-plates were used for biofilm quantification [Vasudevan wt al., 2003; Fox et al., 2005], flat-bottomed polystyrene microtiter-plates were most frequently used.
77 6 It is important to be aware that tissue culture treated plates supplied by different manufacturers may not provide identical conditions for biofilm cultivation and quantification [Stepanovic et al., 2003a], although the difference was not noted by all authors [Christensen et al., 1985]. Still, in order to keep assay conditions constant, it is recommendable to use the plates supplied by the same manufacturer. 5. Cultivation of biofilm (A1) If inoculum is prepared from bacteria grown in a broth, then the culture is diluted 1:100 in TSB supplemented with 1% glucose, and 200 µl is poured into the well. The negative control wells contain broth only; 200 µl of TSB supplemented with 1% glucose per well. (A2) If inoculum is prepared from bacteria cultivated on agar plates, then the wells of microtiter-plate are filled with 180 µl of TSB supplemented with 1% glucose. Thereafter, the quantity of 20 µl of previously prepared bacterial suspensions are added into each well. The negative control wells contain broth only; 200 µl of TSB supplemented with 1% glucose per well. To reduce the possibility of contamination, this step should be performed in a biological safety cabinet. Since phenotypic expression of biofilm formation is highly susceptible to various in vitro conditions, in order to minimize errors and provide reliable analysis of the data obtained, it is esential to perform testing of each strain at least in triplicate (three wells per strain). In addition, each test should be carried out three times. Six wells should be used for negative control. Up to 30 strains may be tested per one 96-well microtiter-plate. (B) The inoculated plate should be covered with the lid and incubated aerobically for 24 h ± 30 min at C under static conditions. Some investigators incubate microtiter-plates overnight [Allignet et al., 1999; Cramton et al., 2001; Fowler et al., 2001; Ziebuhr et al., 2001], or for 18 h [Baldassarri et al., 2001; Deighton et al., 2001; Fox et al., 2005; Tormo et al., 2005; Arciola et al., 2006], some for h [Deighton et al., 2001; Mack et al., 2001], while some authors use prolonged incubation of 48 h [Deighton et al., 2001; Ozerdem Akpolat et al., 2003]. The duration of incubation is important since the density of biofilm is dependent on incubation time [Deighton et al., 2001; Hola et al., 2006]. We recommend the incubation for 24 h which was used in great majority of the biofilm studies [Stepanovic et al., 2000; Deighton et al., 2001; Moretro et al., 2003; Vasudevan et al., 2003; Kennedy and O'Gara, 2004; Mathur et al., 2006]. Besides the incubation time, other factors like atmospheric conditions, temperature, dynamic/static conditions, etc. also influence the biofilm production by tested bacteria [Barker et al., 1990; Perez-Giraldo et al., 1995; Deighton et al., 2001; Stepanovic et al., 2001; Stepanovic et al., 2003d; Svabic-Vlahovic et al., 2003; Gotz, 2002]. 6. Washing After incubation, the contents of the wells are decanted into a discard container. Each well is washed three times with 300 µl of sterile phosphate buffered saline (PBS; ph 7.2) by using an appropriate micropipette. PBS should previously be warmed at the room temperature.
78 7 Following every washing step, the wells are emptied by flicking the plates. Prior to fixation of the biofilm, the plates should be drained in an inverted position. Washing of biofilm is another step of utmost importance since it is supposed to remove all nonadherent cells while simultaneously providing preservation of the biofilm integrity. There are two critical points to be addressed within the recommended washing protocol: first, number of washings and, second, the technique used for washing. It is clear that insufficient washing may lead to false positive results while, on the other side, excessive washing may lead to false negatives. Different protocols comprise two [Cramton et al., 2001; Deighton et al., 2001; Heilmann et al., 1996; Fowler et al., 2001], three [Perez-Giraldo et al., 1995; Stepanovic et al., 2000; Baldassarri et al., 2001; Deighton et al., 2001; Vasudevan et al., 2003; Kennedy and O'Gara, 2004], or four washing steps [Christensen et al., 1985; Mulder & Degener, 1998; Allignet et al., 1999; Deighton et al., 2001; Mack et al., 2001; Arciola et al., 2006]. We believe that two-step washing protocole is not fully effective while both three- and four-step protocoles are acceptable. The three-step washing protocole is applied in laboratories participating in this survey As far as the washing technique is concerned, there is an obvious variety in the methodologies applied. Some investigators use a washing roboter, which is able to wash very gently, but may lead to false positives. Other authors propose washing of biofilm by immersion of plates into large containers of water or PBS, since they consider that usage of pipettes as well as mechanical plate washers may lead to the disruption of the biofilm [Deighton et al., 2001]. We found that careful pipetting does not compromise integrity of the biofilm and recommend washing by using micropipettes and empting by flicking as simple and effective method. It should be noted that only turning the microtiter-plate upside down will not empty the wells because of capillary and intermolecular forces. It is necessary to splash the content out of microtiter-plate which, however, may result in formation of aerosol and possibile contamination of environment. Therefore, it should be carried out with great care. Irespective of the techique used, biofilm integrity should be carefully monitored during washing, and all wells with clearly disrupted biofilm layer should be excluded from further calculations. It is advisable to measure the growth of tested strains in microtiter-plate prior to washing. The strain may fail to grow or the growth may be reduced due to a variety of possible factors and/or technical errors. As the consequence, no biofilm at all or less quantities of biofilm will be produced. There are several ways to measure the bacterial growth, but the easiest way to do this is to measure the turbidity in a wells using microtiter-plate reader and 492 nm [] or 600 nm [Moretro et al., 2003] filter. It is important to take measurement of turbidity without previous shaking of microtiter-plate, to ensure that the integrity of the biofilm is not disturbed. 7. Fixation After the washing, the remaining attached bacteria should be heat fixed by exposing them to hot air at 60 C for 60 minutes.
79 8 It was shown that Bouin's reagent is the most reliable biofilm fixative, followed by air drying (1 h at 60 C) [Baldassarri et al., 1993]. However, air drying appears to be the method of choice, due to safety concerns regarding Bouin's reagent which contains explosive chemicals [Baldassarri et al., 1993]. As an alternative to air drying, fixation by methanol can also be used since it proved to be almost as effective as Bouin's reagent [Baldassarri et al., 1993]. Following fixation with 150 µl of methanol for 20 minutes, the microtiter-plates should be emptied by simple flicking, and left to air dry overnight in inverted position at room temperature. 8. Staining Two methods were used for staining and subsequent quantification of biofilm. In the first one, originaly described by Christensen et al. [Christensen et al., 1985], only the biofilm formed on the botom of well was measured. The modified Christensen's method includes resolubilization of the dye and measures the biofilm formed both on the botom and walls of the well. For the reasons described below, we recommend the second approach. (A) The adherent biofilm layer formed in each microtiter-plate well is stained with 150 µl of crystal violet used for Gram staining (2% Hucker crystal violet) for 15 min at room temperature. After the staining, the stain should be aspirated with a pipette and the excess stain should be rinsed off by placing the microtiter-plate under running tap water. The washing is continued until the washings are free of the stain. Crystal violet stains bacterial cells but not the slimy material [Christensen et al., 1985]. To demonstrate the presence of slimy materials, alcian blue, which selectively stains acid mucopolysaccharides, can be used instead of crystal violet [Christensen et al., 1985]. Other dyes, like 0.1%-1% safranin [Christensen et al., 1985; Heilmann et al., 1996; Mulder & Degener, 1998; Deighton et al., 2001; Cramton et al., 2001; Moretro et al., 2003; Ozerdem Akpolat et al., 2003; Tormo et al., 2005] or trypan blue [Christensen et al., 1985; Mulder & Degener, 1998], were also used for the staining. However, the crystal violet was more frequently used stain [Christensen et al., 1985; Baldassarri et al., 1993; Ziebuhr et al., 1997; Allignet et al., 1999; Stepanovic et al., 2000; Baldassarri et al., 2001; Fowler et al., 2001; Deighton et al., 2001; Gelosia et al., 2001; Mack et al., 2001; Ziebuhr et al., 2001; Dobinsky et al., 2003; Vasudevan et al., 2003; Kennedy and O'Gara, 2004; Arciola et al., 2006]. Although the crystal violet stains only staphylococcal cells and not the slimy material, it is acceptable to use it, because former washing steps wash off all non-adherent cells, so the resting adherent cells will be the only stained. In some experiments the biofilm was stained for only 30 s (usually when safranin was used as the dye) [], in others for one min [Deighton et al., 2001], five min [Stepanovic et al., 2000], 10 min [Ziebuhr et al., 1997], or 15 min [Vasudevan et al., 2003]. We consider that staining for 15 min is the most apropriate, since the dye has to penetrate through biofilm. (B) After the microplate is air dried at room temperature, the dye bound to the cells should be resolubilized i.e. eluted from attached cells with 150 µl of 95% ethanol per well. Ethanol should be gently added and, thereafter, the microtiter-plate covered with the lid (to minimize evaporation) should be left at room temperature for at least one hour without shaking. The addition of ethanol enables indirect measuring of bacteria attached both to the bottom and walls of the wells. Some strains produce deposit at the liquid air interface, but this
80 9 deposit is not considered to be indicative of biofilm production [Christensen et al., 1985]. Although final volume of liquid per well is 200 µl, in order to exclude the deposit at the liquid air interface, only 150 µl of crystal violet and 150 µl of ethanol are used for staining and resolubilization of the dye, respectively. For the same reason, ethanol should be gently added to the wells, and shaking of the microtiter-plate to speed up the process of resolubilization is prohibited. Alternatively, 33% glacial acetic acid [Stepanovic et al., 2000] or methanol can also achieve resolubilization of the dye. 9. Measurement of results The optical density (OD) of each well stained with crystal violet is measured at 570 nm using a microtiter-plate reader. The problem of common OD readers is that they measure the OD only at one point in the middle of the well. Thus, if thickness of the biofilm at that point significantly differs from the rest of the well, the measurement will not be accurate. However, homogeneous resolubilization of the dye bound to the bacterial cells in biofilm layer achieved by the recommended protocol enables indirect, but precise measurement of the biofilm production. In some experiments, the product of decolorization was transferred into new microtiter-plate for the OD measurement [Beenken et al., 2003]. Although this costly and time-consuming procedure may be beneficial, we consider that it is not of crucial importance for the accuracy of the results. 10. Interpretation of the results The interpretation of obtained results requires definition of the cut-off value which separates biofilm producing from no-biofilm producing strains. Some investigators calculate it on the basis of three standard deviations above that of uninoculated medium (negative control) [Christensen et al., 1985], or above that of biofilm negative control strain [Mulder & Degener, 1998; Fox et al., 2005]. Other investigators use predefined values for the cut-off point, 0.12 [Deighton et al., 2001; Gelosia et al., 2001; Ziebuhr et al., 2001], 0.1 [Mack et al., 2001; Dobinsky et al., 2003], or 0.2 [Moretro et al., 2003]. In some experiments the result of positive control strain was used as the starting point for calculation [Allignet et al., 1999], while in other a strain was considered positive if its OD was two times that of the negative control strain [Fowler et al., 2001]. Our recommendation is that obtained results should be averaged and expressed as numbers. The averaged OD values should be calculated for all tested strains and negative controls, since all tests are performed in triplicate and repeated for three times. Second, the cut-off value (ODc) should be established. It is defined as the three standard deviations (SD) above the mean OD of the negative control: ODc = average OD of negative control + (3 SD of negative control). Final OD value of a tested strain is expressed as averaged OD value of the strain reduced by ODc value (OD = average OD of a strain - ODc). If a negative value is obtained, it should be presented as zero. For the purposes of easier interpretation of the results, strains may be classified into the following categories [Stepanovic et al., 2000]: no biofilm producer (0), weak biofilm
81 10 producer (+ or 1), moderate biofilm producer (++ or 2) and strong biofilm producer (+++ or 3) based upon the previously calculated OD values: OD ODc = no biofilm producer, ODc < OD 2 ODc = weak biofilm producer, 2 ODc < OD 4 ODc = moderate biofilm producer, 4 ODc < OD = strong biofilm producer. We consider that the important advantage of the described guidelines for interpretation of the results is the use of negative control as the starting point for all calculations. Application of positive or negative control strains, as the reference for calculations, may lead to certain inconsistencies due to numerous above noted factors which influence the biofilm production. 11. Quality control Four strains deposited by Christensen et al. [Christensen et al., 1985] in ATCC collection (ATCC 35981, ATCC 35982, ATCC and ATCC 35984) may be used as the control strains for quality assurance of the test. The most frequently used strain is S. epidermidis ATCC (designated by Christensen et al. as RP62A), which is a strong biofilm producer. There are other strong biofilm producing strains, such as S. epidermidis CCM 7221 deposited in the Czech Collection of Microorganisms, which also may be used as the control strains. Conclusions The presented protocol incorporated sublimed knowledge on assessment of biofilm production by staphylococci gained both by direct experience in the field of biofilm research in laboratories participating in this survey as well as by comprehensive analysis of methods for assaying biofilm production by staphylococci available in the literature. The study aimed to provide details of all steps involved in quantification of biofilm formation by staphylococci, which are quite frequently lacking in the literature, and to identify the most important problems met through performance of the assay. These should make the procedure more easily applicable, which is of particular importance for the ones newly starting in this field of research. The recognition of single medium suitable for quantification of biofilm production by all staphylococci and clear-cut recommendations for washing procedure are the two most critical points. It is quite clear that complete standardization of the procedure for quantification of biofilm production by staphylococci, in particular the two above noted points, requires further scientific effort. An important shortcoming of this as well as all other in vitro methods for biofilm measurement is that they do not exactly reflect situations in vivo. Guidelines for extrapolation of the data obtained under ideally optimized in vitro testing conditions, which provide maximal biofilm formation, to an actual in vivo situation remain to be established. References Fitzpatrick et al., 2002 Fitzpatrick F, Humphreys H, Smyth E, Kennedy CA, O'Gara JP.: Environmental regulation of biofilm formation in intensive care unit isolates of Staphylococcus epidermidis. J Hosp Infect Nov;52(3):212-8.
82 11 Hola et al., 2006 Hola V, Ruzicka F, Votava M: Dynamic of biofilm formation in relation to nutrition, temperature and time. Scripta Medica 2006 Sept;79(5): Allignet et al., 1999 Allignet J, Galdbart JO, Morvan A, Dyke KG, Vaudaux P, Aubert S, Desplaces N, el Solh N. Tracking adhesion factors in Staphylococcus caprae strains responsible for human bone infections following implantation of orthopaedic material. Microbiology Aug;145 ( Pt 8): Fowler et al., 2001 Fowler VG Jr, Fey PD, Reller LB, Chamis AL, Corey GR, Rupp ME. The intercellular adhesin locus ica is present in clinical isolates of Staphylococcus aureus from bacteremic patients with infected and uninfected prosthetic joints. Med Microbiol Immunol (Berl) Apr;189(3): Kiers et al., 2001 Kiers PJ, Bos R, van der Mei HC, Busscher HJ. The electrophoretic softness of the surface of Staphylococcus epidermidis cells grown in a liquid medium and on a solid agar. Microbiology Mar;147(Pt 3): Ozerdem Akpolat et al., 2003 Ozerdem Akpolat N, Elci S, Atmaca S, Akbayin H, Gul K. The effects of magnesium, calcium and EDTA on slime production by Staphylococcus epidermidis strains. Folia Microbiol (Praha). 2003;48(5): Gotz, 2002 Gotz F. Staphylococcus and biofilms. Mol Microbiol Mar;43(6): Hubalek, 2003 Hubalek Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003;46: Gelosia et al., 2001 Gelosia A, Baldassarri L, Deighton M, van Nguyen T. Phenotypic and genotypic markers of Staphylococcus epidermidis virulence. Clin Microbiol Infect Apr;7(4): Vasudevan et al., 2003 Vasudevan P, Nair MK, Annamalai T, Venkitanarayanan KS. Phenotypic and genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for biofilm formation. Vet Microbiol Mar 20;92(1-2): Mack et al., 1992 Mack D, Siemssen N, Laufs R. Parallel induction by glucose of adherence and a polysaccharide antigen specific for plastic-adherent Staphylococcus epidermidis: evidence for functional relation to intercellular adhesion. Infect Immun May;60(5): Ziebuhr et al., 2001
83 12 Ziebuhr W, Loessner I, Krimmer V, Hacker J. Methods to detect and analyze phenotypic variation in biofilm-forming Staphylococci. Methods Enzymol. 2001;336: Deighton et al., 2001 Deighton MA, Capstick J, Domalewski E, van Nguyen T. Methods for studying biofilms produced by Staphylococcus epidermidis. Methods Enzymol. 2001;336: Mack et al., 2004 Mack, D., Becker, P., Chatterjee, I., Dobinsky, S., Knobloch, J.K., Peters, G., Rohde, H., Herrmann, M., Mechanisms of biofilm formation in Staphylococcus epidermidis and Staphylococcus aureus: functional molecules, regulatory circuits, and adaptive responses. Int. J. Med. Microbiol. 294, van Griethuysen et al., 2005 van Griethuysen A, van Loo I, van Belkum A, Vandenbroucke-Grauls C, Wannet W, van Keulen P, Kluytmans J. Loss of the meca gene during storage of methicillin-resistant Staphylococcus aureus strains. J Clin Microbiol 2005;43: Cramton et al., 2001 Cramton SE, Gerke C, Gotz F. In vitro methods to study staphylococcal biofilm formation. Methods Enzymol. 2001;336: Mack et al., 2000 Mack D, Rohde H, Dobinsky S, Riedewald J, Nedelmann M, Knobloch JK, Elsner HA, Feucht HH. Identification of three essential regulatory gene loci governing expression of Staphylococcus epidermidis polysaccharide intercellular adhesin and biofilm formation. Infect Immun Jul;68(7): Dobinsky et al., 2003 Dobinsky S, Kiel K, Rohde H, Bartscht K, Knobloch JK, Horstkotte MA, Mack D. Glucoserelated dissociation between icaadbc transcription and biofilm expression by Staphylococcus epidermidis: evidence for an additional factor required for polysaccharide intercellular adhesin synthesis. J Bacteriol 2003;185: Handke et al., 2004 Handke LD, Conlon KM, Slater SR, Elbaruni S, Fitzpatrick F, Humphreys H, Giles WP, Rupp ME, Fey PD, O'Gara JP. Genetic and phenotypic analysis of biofilm phenotypic variation in multiple Staphylococcus epidermidis isolates. J Med Microbiol 2004;53: Mack et al., 2001 Mack D, Bartscht K, Fischer C, Rohde H, de Grahl C, Dobinsky S, Horstkotte MA, Kiel K, Knobloch JK. Genetic and biochemical analysis of Staphylococcus epidermidis biofilm accumulation. Methods Enzymol. 2001;336: Fitzpatrick et al., 2005 Fitzpatrick F, Humphreys H, O'Gara JP. Evidence for icaadbc-independent biofilm development mechanism in methicillin-resistant Staphylococcus aureus clinical isolates. J Clin Microbiol Apr;43(4): Knobloch et al., 2002
84 13 Knobloch JK, Horstkotte MA, Rohde H, Mack D. Evaluation of different detection methods of biofilm formation in Staphylococcus aureus. Med Microbiol Immunol (Berl) Oct;191(2): Kennedy and O'Gara, 2004 Kennedy CA, O'Gara JP. Contribution of culture media and chemical properties of polystyrene tissue culture plates to biofilm development by Staphylococcus aureus. J Med Microbiol 2004;53: Stepanovic et al., 2003b Stepanovic S, Dakic I, Opavski N, Jezek P, Ranin L. Influence of the growth medium composition on biofilm formation by Staphylococcus sciuri. Ann Microbiol 2003b; 53: Stepanovic et al., 2000 Stepanovic, S., Vukovic, D., Dakic, I., Savic, B., Svabic-Vlahovic, M., A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods. 40, Stepanovic et al., 2001 Stepanovic S, Vukovic D, Jezek P, Pavlovic M, Svabic-Vlahovic M. Influence of dynamic conditions on biofilm formation by staphylococci. Eur J Clin Microbiol Infect Dis Jul;20(7): Stepanovic et al., 2003d Stepanovic S, Djukic N, Djordjevic V, Djukic S. Influence of the incubation atmosphere on the production of biofilm by staphylococci. Clin Microbiol Infect. 2003d Sep;9(9): Stepanovic et al., 2003a Stepanovic, S., Cirkovic, I., Mijac, V., Svabic-Vlahovic, M., 2003a. Influence of the incubation temperature, atmosphere and dynamic conditions on biofilm formation by Salmonella spp. Food Microbiol. 20, Stepanovic et al., 2003c Stepanovic S, Djukic N, Opavski N, Djukic S. Significance of inoculum size in biofilm formation by staphylococci. New Microbiolc Jan;26(1): Svabic-Vlahovic et al., 2003 Svabic-Vlahovic, M., Dakic, I., Vukovic, D., Stepanovic, S., Biofilm production by Staphylococcus epidermidis at 37 C and 32 C. 13th European Congres of Clinical Microbiology and Infectious Duseases, Glasgow, United Kingdom, May 10-13, Clin. Microbiol. Infect. 9(suppl. 1 - CD), P926. Baldassarri et al., 2001 Baldassarri L, Bertuccini L, Ammendolia MG, Arciola CR, Montanaro L. Effect of iron limitation on slime production by Staphylococcus aureus. Eur J Clin Microbiol Infect Dis May;20(5): Ziebuhr et al., 1997
85 14 Ziebuhr W, Heilmann C, Gotz F, Meyer P, Wilms K, Straube E, Hacker J. Detection of the intercellular adhesion gene cluster (ica) and phase variation in Staphylococcus epidermidis blood culture strains and mucosal isolates. Infect Immun Mar;65(3): Cho et al., 2002 Cho SH, Naber K, Hacker J, Ziebuhr W. Detection of the icaadbc gene cluster and biofilm formation in Staphylococcus epidermidis isolates from catheter-related urinary tract infections. Int J Antimicrob Agents Jun;19(6): Arciola et al., 2006 Arciola CR, Campoccia D, Baldassarri L, Donati ME, Pirini V, Gamberini S, Montanaro L. Detection of biofilm formation in Staphylococcus epidermidis from implant infections. Comparison of a PCR-method that recognizes the presence of ica genes with two classic phenotypic methods. J Biomed Mater Res A Feb;76(2): Mathur et al., 2006 Mathur T, Singhal S, Khan S, Upadhyay DJ, Fatma T, Rattan A. Detection of biofilm formation among the clinical isolates of Staphylococci: an evaluation of three different screening methods. Indian J Med Microbiol Jan;24(1):25-9. Rohde et al., 2001 Rohde H, Knobloch JK, Horstkotte MA, Mack D. Correlation of biofilm expression types of Staphylococcus epidermidis with polysaccharide intercellular adhesin synthesis: evidence for involvement of icaadbc genotype-independent factors. Med Microbiol Immunol (Berl) 2001;190: Heilmann et al., 1996 Heilmann C, Gerke C, Perdreau-Remington F, Gotz F. Characterization of Tn917 insertion mutants of Staphylococcus epidermidis affected in biofilm formation. Infect Immun Jan;64(1): Fux et al., 2005 Fux CA, Shirtliff M, Stoodley P, Costerton JW. Can laboratory reference strains mirror "realworld" pathogenesis? Trends Microbiol Feb;13(2): Fox et al., 2005 Fox LK, Zadoks RN, Gaskins CT. Biofilm production by Staphylococcus aureus associated with intramammary infection. Vet Microbiol May 20;107(3-4): Knobloch et al., 2001 Knobloch JK, Bartscht K, Sabottke A, Rohde H, Feucht HH, Mack D. Biofilm formation by Staphylococcus epidermidis depends on functional RsbU, an activator of the sigb operon: differential activation mechanisms due to ethanol and salt stress. J Bacteriol Apr;183(8): Moretro et al., 2003 Moretro T, Hermansen L, Holck AL, Sidhu MS, Rudi K, Langsrud S. Biofilm formation and the presence of the intercellular adhesion locus ica among staphylococci from food and food processing environments. Appl Environ Microbiol Sep;69(9):
86 15 De Paoli, 2005 De Paoli P. Biobanking in microbiology: From sample collection to epidemiology, diagnosis and research. FEMS Microbiol Rev 2005;29: Tormo et al., 2005 Tormo MA, Knecht E, Gotz F, Lasa I, Penades JR. Bap-dependent biofilm formation by pathogenic species of Staphylococcus: evidence of horizontal gene transfer? Microbiology Jul;151(Pt 7): Harraghy et al., 2006 Harraghy N, Seiler S, Jacobs K, Hannig M, Menger MD, Herrmann M. Advances in in vitro and in vivo models for studying the staphylococcal factors involved in implant infections. Int J Artif Organs Apr;29(4): Christensen et al., 1982 Christensen GD, Simpson WA, Bisno AL, Beachey EH. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect Immun 1982;37: Christensen et al., 1985 Christensen GD, Simpson WA, Younger JJ, Baddour LM, Barrett FF, Melton DM, Beachey EH. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. J Clin Microbiol 1985;22: Ziebuhr et al., 1999 Ziebuhr W, Krimmer V, Rachid S, Lossner I, Gotz F, Hacker J. A novel mechanism of phase variation of virulence in Staphylococcus epidermidis: evidence for control of the polysaccharide intercellular adhesin synthesis by alternating insertion and excision of the insertion sequence element IS256. Mol Microbiol Apr;32(2): Schleifer & Kroppenstedt, 1990 Schleifer KH, Kroppenstedt RM. Chemical and molecular classification of staphylococci. Soc Appl Bacteriol Symp Ser. 1990;19:9S-24S. Baldassarri et al., 1993 Baldassarri L, Simpson WA, Donelli G, Christensen GD. Variable fixation of staphylococcal slime by different histochemical fixatives. Eur J Clin Microbiol Infect Dis 1993;12: CA-SFM, 1996 Courvalin P, Soussy C-J (Eds.) Report of the Comité de l'antibiogramme de la Société Française de Microbiologie. Clin Microbiol Infect 1996;2:(Supplement 1). Mulder & Degener, 1998 Mulder JG, Degener JE. Slime-producing properties of coagulase-negative staphylococci isolated from blood cultures. Clin Microbiol Infect Feb;4(12): Deighton and Balkau, 1990 Deighton MA, Balkau B. Adherence measured by microtiter assay as a virulence marker for Staphylococcus epidermidis infections. J Clin Microbiol 1990;28:
87 16 Christensen et al., 1990 Christensen GD, Baddour LM, Madison BM, Parisi JT, Abraham SN, Hasty DL, Lowrance JH, Josephs JA, Simpson WA. Colonial morphology of staphylococci on Memphis agar: phase variation of slime production, resistance to beta-lactam antibiotics, and virulence. J Infect Dis 1990;161: Perez-Giraldo et al., 1995 Perez-Giraldo C, Rodriguez-Benito A, Moran FJ, Hurtado C, Blanco MT, Gomez-Garcia AC. Influence of the incubation atmosphere on the production of slime by Staphylococcus epidermidis. Eur J Clin Microbiol Infect Dis Apr;14(4): Hola et al., 2004 Hola V, Ruzicka F, Votava M. Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis. New Microbiol 2004;27: Beenken et al., 2003 Beenken KE, Blevins JS, Smeltzer MS. Mutation of sara in Staphylococcus aureus limits biofilm formation. Infect Immun Jul;71(7): Donlan, 2002 Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis 2002;8: Donlan & Costerton, 2002 Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev 2002;15: Barker et al., 1990 Barker LP, Simpson WA, Christensen GD. Differential production of slime under aerobic and anaerobic conditions. J Clin Microbiol Nov;28(11): Cheung et al., 1988 Cheung AL, Fischetti VA. Variation in the expression of cell wall proteins of Staphylococcus aureus grown on solid and liquid media. Infect Immun May;56(5):
88 7.2 Holá V., Toršová V., Gallo J.: Biofilmy a antimikrobiální látky; in M. Rulík, V. Holá, F. Růžička, M. Votava (eds.); Mikrobiální biofilmy; (in press; předpokládané datum vydání jaro 2007).
89 Mikrobiální biofilmy 1 9. Biofilmy a antimikrobiální látky 9. Biofilmy a antimikrobiální látky 9.1 Mechanizmy rezistence biofilmu k antimikrobiálním látkám Nemožnost průniku látek do biofilmové vrstvy Zábrana difúze Vazba na složky biofilmu Pomalý růst biofilmu a heterogenita mikrobiální populace Pomalý růst biofilmu Heterogenita mikrobiální populace Univerzální odpověď na stres (general stress response) Vznik tzv. perzistorů Vznik biofilmového fenotypu Fenotyp rezistence spojený s primární adhezí Fenotyp rezistence spojený s produkcí MDR pump Další mechanizmy uplatňující se v biofilmové rezistenci Quorum sensing Úzká vazba mezi bakteriálními buňkami v biofilmu Problematika biofilmové rezistence u kvasinek Programovaná buněčná smrt Buněčné mechanizmy bakteriální rezistence Závěr 9.2 Možnosti ovlivnění biofilmové rezistence vnějšími vlivy Inhibice mikrobiální adherence Inhibice quorum sensing (mezibuněčné komunikace) Bioakustický efekt Bioelektrický efekt Bioradiační efekt Degradace extracelulární matrix 9.3 Současné možnosti léčby biofilmových infekcí Biofilmy a antimikrobiální látky Mikrobiální buňky žijící ve formě biofilmu jsou mnohonásobně rezistentnější k biocidům než planktonické formy stejného kmene. Vzhledem k vysoké rezistenci biofilmů k antimikrobiálním látkám mohou být biofilmové infekce cizích těles a implantátů často léčeny pouze vyjmutím infikovaného implantátu, což znamená nejen vyšší finanční náklady na léčbu, ale i vyšší zátěž pro pacienta samotného. Kromě toho jsou zákroky spojené s odstraněním těchto materiálů, ať již srdečních chlopní, kloubních náhrad či tunelizovaných katétrů, spojeny s vyšším rizikem řady dalších komplikací (Kuhn et al. 2002a). Zvýšená rezistence biofilmů k biocidům byla prokázána v mnoha studiích in vitro a je významnou komplikací u infekcí implantátů a cizích těles in vivo. Mnohé studie in vitro dokázaly, že účinnost biocidů je u biofilmových buněk oproti buňkám planktonickým nižší o jeden až čtyři řády (Costerton et al. 1999; Hola et al. 2004), např. podle práce Ceri et al ho et al. (1999) je potřeba více než 500-násobnou dávku oproti MIC (minimální inhibiční koncentrace) ampicilinu, aby se množství buněk v biofilmu E. coli snížilo o tři řády. Williams et al et al. (1997) zjistili, že ke snížení počtu buněk v biofilmu S. aureus o tři řády je potřeba více než desetinásobek MBC (minimální baktericidní koncentrace) vankomycinu. Stejně tak planktonické buňky P. aeruginosa jsou 15-krát citlivější k tobramycinu, než jejich biofilmová forma (Hoyle et al. 1992). Mnoho autorů též popsalo selhání konvenční terapie in vivo pacientů s infekcemi spojovanými právě s biofilmy a biofilmpozitivními mikroby (Wu et al. 2003; Neut et al. 2005). Biofilm je pro bakterie významným faktorem umožňujícím přežití a množení. Vlastnosti biofilmu a biofilmpozitivních mikrobů vysvětlují podstatu recidivy biofilmových
90 Mikrobiální biofilmy 2 9. Biofilmy a antimikrobiální látky infekcí. Vyzrálá vrstva biofilmu poskytuje bakteriálním buňkám ochranu před účinky vnějšího prostředí, antimikrobiálními látkami a imunitním systémem hostitele. Z tohoto vyzrálého biofilmu se uvolňují jednotlivé buňky či mikrokolonie a ty jsou primárně zodpovědné za manifestaci onemocnění, protože vyvolávají imunitní odpověď hostitele. Při léčbě infekce antimikrobiálními látkami se sice daří eliminovat většinu planktonických a část biofilmových buněk, menší část buněk však přežívá i vysoké koncentrace antimikrobiálních látek. Obdobně si i imunitní systém hostitele s těmito buňkami dokáže poradit v planktonické podobě, ale nedokáže zničit buňky usazené v biofilmové vrstvě. Po skončení léčby antimikrobiálními látkami se přeživší bakterie mohou začít znovu množit a výsledkem je recidiva infekce (Spoering& Lewis 2001). Vznik biofilmové rezistence není dosud zcela objasněn, v nedávných studiích však bylo použita řada modelových systémů, aby se zjistilo jak a proč jsou biofilmy tak rezistentní k antimikrobiálním látkám. S rostoucí závažností nozokomiálních infekcí způsobovaných biofilmpozitivními mikroby se studiu efektů antimikrobiálních látek na tyto mikrobiální komunity věnuje více pozornosti. 9.1 Mechanizmy rezistence biofilmu k antimikrobiálním látkám Biofilmová rezistence se projevuje mnoha mechanizmy, které se mohou lišit dle druhu mikroba tvořícího biofilm a také látky, která je vůči tomuto mikrobu aplikovaná. Jiné mechanizmy biofilmové rezistence se projevují též u biofilmu v počáteční fázi růstu a jiné u plně vyzrálého a starého biofilmu. Je publikováno mnoho teorií vysvětlujících, proč jsou bakteriální buňky v biofilmu rezistentnější, než jejich planktonické formy. Mezi nejvýznamnější a nejvíce prostudované mechanizmy biofilmové rezistence patří: fyzikální a chemické bariéry bránící penetraci látek do biofilmové vrstvy snížená rychlost růstu buněk v biofilmu aktivace univerzální odpovědi na stres (general stress response) vznik určité rezistentní skupiny buněk, tzv. perzistorů vznik biofilm-specifického fenotypu a exprese pravděpodobných genů rezistence. Všechny tyto mechanizmy mohou v biofilmu působit jednotlivě nebo se mohou vzájemně doplňovat. Ve vícedruhovém biofilmu je samozřejmě problematika biofilmové rezistence ještě mnohem složitější Nemožnost průniku látek do biofilmové vrstvy Prvním mechanizmem rezistence, který se ve spojení s biofilmem a biofilmovou vrstvou přímo nabízí, je nemožnost průniku látek do biofilmové vrstvy. Biofilmová vrstva může jako penetrační bariéra působit dvěma způsoby jako určité síto, nebo jako donor volných vazebných míst. Zpomalení difúze snižuje koncentraci antibakteriálních látek, které se dostávají do biofilmové vrstvy, což ve spojení s přítomnými degradujícími enzymy, jako jsou například β-laktamázy, kataláza apod., umožňuje neutralizovat či výrazně snížit vliv difundujících antibakteriálních látek. Taková synergie mezi zpomalenou difůzí a degradací antibiotik byla prokázána např. u β-laktamových antibiotik a biofilmů P. aeruginosa produkujících β-laktamázy (Giwercman et al. 1991). Synergický efekt mezi zpomalenou difůzí a degradací pomocí enzymů byl též přesvědčivě prokázán pomocí matematických modelů založených na těchto pozorováních (Stewart 1996).
91 Mikrobiální biofilmy 3 9. Biofilmy a antimikrobiální látky Zábrana difúze Funkce síta byla prokázána u určitých složek biofilmu grampozitivních i gramnegativních bakterií. U grampozitivních bakterií prokázali Souli& Giamarellou 1998 et al. (1998), že s využitím exopolysacharidu izolovaného z biofilmu S. epidermidis lze snížit citlivost Bacillus subtilis k mnoha antimikrobiálním látkám. Vliv tohoto exopolysacharidu na antimikrobiální látky se však lišil citlivost B. subtilis ke glykopeptidům (vankomycinu a teikoplaninu, tedy látkám s poměrně velkou molekulou) byla výrazně nižší, ale citlivost k dalším látkám, např. rifampicinu, klindamycinu a makrolidům zůstala na stejné úrovni, nebo byla snížena jen málo. Ke stejnému jevu dochází i u gramnegativních bakterií, a to zejména díky přítomnosti alginátu. Hatch et al. (1998) prokázali, že 2% suspenze alginátu, což je významná složka biofilmu P. aeruginosa, inhibovala difůzi gentamicinu a tobramycinu a tento efekt byl opět zvrácen použitím alginát-lyázy. Další autoři zkoumali difůzi některých antibiotik (ceftazidimu, cefsulodinu, piperacilinu, gentamicinu a tobramycinu) přes syntetický a přírodně produkovaný alginátový gel a zjistili, že β-laktamová antibiotika difundují do matrix rychleji, než např. aminoglykosidy (Coquet et al. 1998; Cochran et al. 2000a). Tento mechanizmus rezistence mikrobiálním biofilmům poskytuje též účinnou ochranu proti velkým molekulám imunitního systému hostitele, jako jsou např. antimikrobiální proteiny lysozymy a komplement. Difúzní bariéra je též pravděpodobně účinná i proti menším antimikrobiálním peptidům mnohým defenzinům a jejich analogům ( Martinez& Casadevall 2006; Fedtke et al. 2004). Vazba na složky biofilmu Biofilmová vrstva a její složky na sebe též mohou vázat určité látky na základě fyzikálních či chemických interakcí. Relativně silně hydratované polyanionické gelové polysacharidy a glykoproteiny biofilmové vrstvy působí jako výměníky iontů a adsorbují kationty antimikrobiálních látek, dokud nejsou všechna jejich vazebná místa nasycena (Wagman et al. 1975, as cited in Schierholz& Beuth 2001). Vazba na složky biofilmu se může projevovat buď reakcí látky s konkrétní složkou biofilmu (chemické interakce), nebo vazbou na konkrétní složku biofilmu (fyzikální interakce). Projevují se zde např. vazby vodíkovými můstky, Van der Waalsovy síly, vazby dipólové apod. Ačkoliv na základě matematických modelů by se dalo předpokládat, že mnoho antimikrobiálních látek by touto bariérou nemělo být ovlivněno, četné studie dokázaly pravý opak. Například u chloru, jenž je součástí běžně používaných dezinfekčních látek, se zjistilo, že koncentrace ve spodních vrstvách dvoudruhového biofilmu Klebsiella pneumoniae a P. aeruginosa nedosáhla více, než 20 % své koncentrace v okolním médiu (de Beer et al. 1994) a penetrační profil chloru odpovídal penetračnímu profilu substrátu konzumovaného v matrix biofilmu. Suci et al et al. (1994) pomocí infračervené mikroskopie dokázali, že poměr transportu antibiotika ciprofloxacinu k pevnému povrchu byl výrazně nižší u povrchu kolonizovaného biofilmem P. aeruginosa ve srovnání se sterilním povrchem. Transport ciprofloxacinu v médiu ke sterilnímu povrchu trval jen 40 sekund, k transportu skrz biofilmovou vrstvu potřeboval ciprofloxacin 21 minut, z čehož autoři usuzují, že ciprofloxacin se váže na některé složky biofilmu. Ke stejným výsledkům došli i další autoři, a to s řadou dalších antimikrobiálních látek, např. s piperacilinem u biofilmu P. aeruginosa (Hoyle et al. 1992). Podle dalších autorů je negativně nabitý exopolysacharid též velmi účinnou ochranou buněk před pozitivně nabitými aminoglykosidy (Hoyle et al. 1990; Shigeta et al. 1997). Zjistilo se také, že aminoglykosidy se zpočátku váží na alginát, ale jejich difúze se zrychlí po minutové prodlevě. Tato prodleva se přičítá právě času potřebnému k nasycení volných vazebných míst v biofilmu. V pokuse se stejným designem, kde byl
92 Mikrobiální biofilmy 4 9. Biofilmy a antimikrobiální látky zkoumán vliv biofilmu S. epidermidis na difůzi rifampicinu a vankomycinu, však došli autoři k právě opačným výsledkům a průnik rifampicinu a vankomycinu by podle nich neměl být ovlivněn vazbou na složky biofilmu (Dunne, Jr. et al. 1993). Na druhou stranu se můžeme setkat s tím, že účinek glykopeptidových antibiotik na biofilmpozitivní kmeny bakterií není příliš vysoký (Mathur et al. 2005; Konig et al. 2001). Samozřejmě zde mohou hrát svou roli ještě i další mechanizmy biofilmové rezistence. Ve většině případů však polysacharidová bariéra postup antimikrobiálních látek pouze zpomalí. Příkladem takovýchto látek mohou být fluorované chinolony, které pronikají biofilmem velmi dobře (Ishida et al. 1998; Anderl et al. 2000; Vrany et al. 1997), i když s určitou časovou prodlevou. Jak již však bylo zmíněno výše, i tato časová prodleva může hrát významnou roli při enzymatické degradaci antimikrobiálních látek. Svědčí o tom i studie provedené s difůzí peroxidu vodíku skrz silnou vrstvu biofilmu, jenž byl vytvořen normálním biofilmpozitivním kmenem P. aeruginosa a kmenem mutantním, kterému chyběl jeden z hlavních genů pro katalázu (enzym neutralizující peroxid vodíku), kata, které prokázaly, že bez přítomnosti tohoto enzymu peroxid vodíku skrz vrstvu biofilmu difunduje snáze a rychleji (Stewart et al. 2000). Tyto výsledky napovídají, že v silných biofilmech jsou buňky P. aeruginosa chráněny před účinky peroxidu vodíku též katalázou-zprostředkovaným rozkladem této sloučeniny. Stejné výsledky vyplynuly i ze studií chlornanu (Brown et al., 1988). U ampicilinu bylo prokázáno, že nebyl schopen difundovat skrz biofilm K. pneumoniae a že za tento jev byla pravděpodobně zodpovědná tvorba β-laktamáz, protože přes biofilm mutanta neschopného tvořit β-laktamázy ampicilin difundoval (Anderl et al. 2000). Na druhou stranu byly i buňky tohoto mutanta v biofilmu rezistentní k působení ampicilinu, což napovídá, že i k této rezistenci napomohly opět i jiné mechanizmy. V rámci stejné studie byl sledován i ciprofloxacin, který difundoval biofilmem stejně jako ampicilin, nicméně i jeho eradikační schopnosti selhaly, přestože zde to nemůže být přisuzováno vlivu β-laktamáz. Mnoha autory byly též pozorovány významné rozdíly v difůzi antimikrobiálních látek skrz různě silnou vrstvu biofilmu. I když skrz slabou vrstvu biofilmu pronikají látky např. peroxid vodíku lépe, než skrz silnou, i buňky v tomto slabém biofilmu jsou ve srovnání s planktonickými buňkami rezistentnější (Stewart et al. 2000). Exopolysacharidová matrix (nebo jiné složky biofilmu) tedy netvoří neproniknutelnou bariéru vůči difůzi antimikrobiálních agens a je tedy nutno vzít v potaz i další mechanizmy, které mohou bakteriím v biofilmu pomoci přežít tlak těchto látek. Pro určité sloučeniny však exopolysacharidová matrix tvoří počáteční bariéru, která minimálně zpomalí jejich průnik Pomalý růst biofilmu a heterogenita mikrobiální populace Pomalý růst biofilmu Začnou-li bakteriální buňky v kultuře strádat a hladovět, nebo chybí-li jim byť jedna konkrétní živina, jejich růst se zpomalí. Tento přechod z exponenciálního růstu k růstu pomalému či žádnému je obecně provázen zvýšením rezistence k antibiotikům (Tuomanen et al., 1986a; 1986b), a to i v planktonických kulturách. Zpomalený růst buněk byl pozorován též ve zralých biofilmech (Brown et al., 1988; Wetland et al., 1996). Protože předpokládáme, že buňky rostoucí v biofilmu zakusí nějakou formu strádání či limitace živinami, předpokládáme též, že tato fyziologická změna může mít za následek zvýšenou rezistenci biofilmu k antimikrobiálním látkám. Těmto předpokladům nasvědčují i studie publikované skupinou vědců z Manchesterské univerzity, kteří studovali efekt pomalého růstu bakterií za kontrolovaných podmínek v planktonických kulturách a v biofilmu u P. aeruginosa, E. coli a S. epidermidis (Evans et al. 1991; Duguid et al. 1992a; Duguid et al. 1992b). Došli k závěru,
93 Mikrobiální biofilmy 5 9. Biofilmy a antimikrobiální látky že citlivost obou forem, planktonické i biofilmové, k ciprofloxacinu a tobramycinu se zvyšuje s rostoucí rychlostí růstu. To podporuje právě předpoklad, že pomalá růstová rychlost biofilmových buněk je chrání před účinkem antibiotik. U P. aeruginosa v nízkých růstových rychlostech byly planktonické i biofilmové buňky stejně rezistentní k ciprofloxacinu, ale jakmile se zvýšila růstová rychlost buněk v obou kulturách, planktonické buňky byly mnohem citlivější, než buňky biofilmové (Evans et al. 1991). I výsledky dalších autorů odpovídají této teorii (Spoering& Lewis 2001), i když i u Evansova experimentu vidíme, že se i zde dostává ke slovu některý z dalších mechanizmů biofilmové rezistence. K obdobným výsledkům došli i Desai et al et al. (1998) při studiu biofilmů Burkholderia cepacia. Porovnáním rezistence planktonické a biofilmové kultury v různých fázích exponenciálního i stacionárního růstu došli autoři k závěru, že vlivem zpomalení růstu ve stacionární fázi se v obou kulturách zvýšila rezistence buněk k antimikrobiálním látkám, přičemž u biofilmové kultury byla rezistence 15-krát vyšší, než u stacionární fáze planktonických buněk. Výsledky opět naznačují, že se v biofilmové kultuře projevil i jiný mechanizmus rezistence a že zpomalení růstu je jen jedním z mnoha mechanizmů ovlivňujících tuto rezistenci. Navíc, mnohé studie prokázaly, že zpomalený růst neovlivňuje citlivost vůči všem antibiotikům například Brooun et al et al. (2000) prokázali, že rezistence biofilmu P. aeruginosa k tetracyklinům je ovlivněna zpomaleným růstem, tento efekt však nepozorovali u tobramycinu. Evans et al et al. (1990) se zaměřili na růstovou rychlost buněk v biofilmu a její vztah k biofilmové rezistenci s využitím technik používaných při práci s buněčnými kulturami. Zjistili, že ty buňky v biofilmu E. coli, které rostly nejpomaleji, byly také nejvíce rezistentní k antibiotiku cetrimidu a při růstových rychlostech vyšších, než 0,3/hod byly planktonické buňky a buňky biofilmové stejně citlivé k antibiotikům. Jiná studie ukázala, že růstové rychlosti biofilmu S. epidermidis silně ovlivňují jeho citlivost čím vyšší je rychlost buněčného růstu, tím rychleji jsou buňky inaktivovány ciprofloxacinem (DuGuid et al., 1990). I vliv rifampicinu na bakteriální buňky v biofilmu a jejich přežívání odpovídá hypotéze, že zpomalení růstu bakterií v biofilmové vrstvě je zodpovědné za výrazné zvýšení rezistence těchto buněk (Zheng& Stewart 2002). Heterogenita mikrobiální populace Při studiu biofilmu za přísně definovaných podmínek růstu se nutně nabízí předpoklad, že jakákoliv buňka v biofilmu žije za trochu odlišných podmínek, než zbývající buňky ve stejném biofilmu, a proto též roste jinou růstovou rychlostí. Gradienty živin, různé odpadní produkty a četné signalizační faktory, to vše přispívá k této heterogenitě uvnitř biofilmu. Ve vyzrálých biofilmech můžeme sledovat komplexní trojrozměrnou strukturu s mnoha odlišnými typy mikroprostředí. Tato mikroprostředí se liší např. rozdílnou osmolaritou, přísunem živin a buněčnou hustotou. Díky této heterogenitě můžeme v biofilmové populaci nalézt buňky s mnoha odlišnými projevy jejich fenotypu, jediný, univerzální biofilmový fenotyp tedy neexistuje (Fux et al. 2005). Toto tvrzení podpírá i studie Sufya et al. (2003), která ukázala, že růstové rychlosti daného mikroba se mohou překvapivě lišit i v rámci identických mikroprostředí. Například průměrný generační čas jednotlivých klonů získaných z pozdní logaritmické kultury se výrazně lišil 500 a 45 minut. Heterogenitu biofilmu a rozdíly uvnitř biofilmové vrstvy je možné vizualizovat barvicími technikami pomocí akridinové oranži. Barvení akridinovou oranží využívá její vazby na nukleové kyseliny a na základě relativního poměru RNA DNA a jejich odlišné výsledné barvy rozlišuje rychle rostoucí buňky od buněk rostoucích pomalu (Wetland et al., 1996). Buňky s vysokým relativním obsahem RNA se barví do oranžova až oranžovočervena, buňky s nízkým relativním obsahem RNA se barví do zelena až žlutozelena. Když se tato
94 Mikrobiální biofilmy 6 9. Biofilmy a antimikrobiální látky barvící technika použije na týden starý biofilm, zbarví se oranžově rozhraní biofilmu s tekutinou a okrajové zóny biofilmu a zeleně střed biofilmu. Tato heterogenita v růstu buněk v biofilmu byla rovněž prokázána pomocí technik sledujících syntézu proteinů a respirační aktivitu (Huang et al. 1998; Xu et al. 2000). Někteří autoři se též zaměřili na studium heterogenity fyziologické aktivity, která je odpovědí na účinek antimikrobiálních látek. Huang et al et al. (1995) sledovali například vzorec respirační aktivity biofilmu K. pneumoniae jako odpověď na působení chloraminu. Zjistili, že buňky nejblíže k rozhraní biofilm vodná fáze ztratily aktivitu jako první, tedy jako první podlehly účinkům chloraminu. K podobným výsledkům dospěli i jiní autoři při studiu dalších antimikrobiálních látek (např. Korber et al. 1994). Tyto studie nám ukazují, že odpověď jednotlivých buněk v biofilmu na působení antimikrobiálních látek se může výrazně lišit (a také se výrazně liší) v závislosti na pozici konkrétní buňky v biofilmové komunitě. Vzhledem k tomu, že tyto rozdíly mezi jednotlivými buňkami mohou být velmi výrazné, měl by být tento fakt brán v úvahu při léčbě biofilmových infekcí a likvidaci biofilmů obecně Univerzální stresová odpověď (general stress response) Bakterie odpovídají na nedostatek živin a na jiné environmentální stresy syntézou σ faktorů. Tyto σ faktory, které jsou např. u gramnegativních bakterií kontrolovány regulonem rpos, regulují transkripci genů, jejichž produkty zmírňují účinky stresu. Syntézou produktů těchto genů tedy bakterie odpovídají na působení stresových faktorů a tím se u nich aktivuje mechanizmus univerzální odpovědi na působení stresových faktorů. Ten je založen např. na tom, že mění permeabilitu buněčné membrány a brání tak dalšímu působení stresových faktorů na buňku (Liu et al., 2000). Studiem biofilmů E. coli, tvořených kmeny s genem rpos a bez něj, zjistili Adams& McLean 1999 et al. (1999), že biofilmy E. coli s regulonem rpos mají vyšší denzitu a vyšší množství životaschopných mikroorganizmů. Závislost vyššího množství životaschopných buněk na buněčné hustotě zkoumané populace byla prokázána i v biofilmech Rhizobium sp., na kterých se s těmito experimenty začínalo (Thorne et al., 1999). Vzhledem k tomu, že regulon rpos je aktivován v průběhu pomalého růstu tohoto organizmu, zdá se, že podmínky, které vyvolávají zpomalení bakteriálního růstu, jako např. nedostatek živin či produkce toxických metabolitů, podporují vznik biofilmu. Nedostatek živin a vyšší koncentrace toxických metabolitů mohou být velmi výrazné v hlubších vrstvách biofilmu. Tresse et al et al. (2003) zjistili, že buňky E. coli pěstované na agarových plotnách byly více rezistentní k aminoglykosidům, pokud se při kultivaci snížila tenze kyslíku. Předpokládají, že k tomuto jevu došlo kvůli snížené absorpci antibiotika buňkami, které postrádaly kyslík (a byly tedy vystaveny určitému stresu). Teorie, že zpomalený růst některých buněk uvnitř biofilmu není způsoben nedostatkem živin jako takovým, ale je univerzální odpovědí na stres, která je vyvolána růstem v biofilmu (Brown& Barker 1999), je zajímavá, protože odpověď organizmu na stres, ať se jedná o bakterii nebo člověka, vede k určitým fyziologickým změnám. Funkcí těchto fyziologických změn je ochrana buňky před jakýmkoliv stresem z vnějšího prostředí, takže buňky jsou potom schopny chránit se např. před škodlivými vlivy tepelného šoku, změn ph prostředí a před mnoha chemickými látkami (Hengge-Aronis, 1996), tedy i antibiotiky. Ústředním regulátorem této odpovědi je u gramnegativních bakterií σ faktor RpoS. Předpokládá se, že RpoS je indukován vysokou buněčnou hustotou; buňky rostoucí v těchto vysokých hustotách vypadají, že prošly působením jakéhosi stresu a tedy i zmobilizovaly svou univerzální stresovou odpověď. Důkazem pro tento předpoklad je produkce trehalózy (což je osmoprotektant) a katalázy těmito buňkami (Liu et al., 2000).
95 Mikrobiální biofilmy 7 9. Biofilmy a antimikrobiální látky Ve chvíli, kdy buňky v biofilmu pocítí vysokou buněčnou hustotu, můžeme logicky předpokládat, že tyto buňky budou exprimovat RpoS. Tomu odpovídají i výsledky další studie, ve které autoři prokázali pomocí RT-PCR, že ve sputu pacientů s cystickou fibrózou s chronickou biofilmovou infekcí P. aeruginosa je přítomná rpos mrna (Foley et al., 1999). Důležitým poznatkem i to, že buňky E. coli, kterým chybí regulon rpos, nejsou schopny tvořit normální biofilm, zatímco planktonické buňky nejsou očividně absencí σ faktoru nijak ovlivněny (Adams& McLean 1999). Další odpovědí bakteriálních buněk na působení stresu je ztráta kultivovatelnosti. Dlouhodobé strádání indukuje za běžných podmínek tuto ztrátu, přičemž buňky zůstávají metabolicky aktivní a strukturálně nepoškozené (Rayner et al. 1998) Vznik tzv. perzistorů V roce 1944 publikoval Bigger v The Lancet článek o svých výzkumech na nově zavedeném penicilinu, ve kterém zmínil dva důležité poznatky (Bigger, 1944). Prvním z nich bylo, že penicilin je spíše baktericidní než bakteriostatické antibiotikum, což odporovalo převažujícím názorům té doby o jeho bakteriostatičnosti. Druhým poznatkem bylo, že vystavením stafylokokové kultury penicilinu nedošlo ke sterilizaci této kultury, ale přežila malá část buněk, které Bigger vhodně nazval perzistoři. Bigger odhadl počet perzistorů v bakteriální populaci na V této pionýrské, ale často přehlížené a na dlouhý čas zapomenuté studii Bigger zvažoval dvě hypotézy přežití perzistorů zda je rezistence perzistorů k inhibičnímu účinku penicilinu na jejich růst dána dědičně, nebo zda jsou perzistoři jen určité varianty původního kmene, které mají stejnou citlivost k inhibičnímu účinku penicilinu na jejich růst jako hlavní část bakteriální populace, ale jsou necitliví k zabití penicilinem. Bigger též ukázal, že z kultivace těchto perzistorů vzniká populace nerozeznatelná od původní populace jsou podobně citliví k inhibici růstu penicilinem a produkují nové perzistory. Keren et al et al. (2004) zopakovali na základě Biggerova článku pokusy s E. coli a účinkem ampicilinu a došli ke stejným výsledkům přežívající perzistoři vytvoří znovu populaci buněk se stejnou citlivostí k ampicilinu jako byla původní populace a jsou tedy pouze fenotypovou variantou původního kmene. Perzistoři se vyskytují i v planktonické populaci, i když v množstvích několikařádově nižších, než v biofilmu. Dokladem tohoto může být studie Ahbyho et al. (1994), ve které sledovali působení rostoucích koncentrací ciprofloxacinu a imipenemu na planktonické buňky E. coli. Obě antibiotika snížila koncentraci přežívajících mikrobů o dva až tři řády již v relativně nízkých koncentracích, zatímco zbylé přeživší buňky přežívaly další nárůst koncentrací antibiotik i přes dosažené extrémní hodnoty. Ke stejným výsledkům došli Muli& Struthers 1998 et al. (1998) u biofilmu Lactobacillus acidophilus s amoxicilinem a klindamycinem a u Gardnerella vaginalis s erytromycinem a metronidazolem. Spoering& Lewis 2001 et al. (2001) studovali planktonické buňky ve stacionární fázi růstu a biofilmové buňky a prokázali, že biofilmové buňky a planktonické buňky stacionární fáze mají podobnou rezistenci k antibiotikům a že tato rezistence je u obou studovaných skupin závislá zejména na přítomnosti perzistorů. I další studie prokázaly přítomnost perzistorů v biofilmu, např. Brooun et al et al. (2000) či Drenkard& Ausubel 2002 et al. (2002) s biofilmem P. aeruginosa. Jejich data nasvědčují tomu, že většina buněk v biofilmu je k antibiotikům citlivých stejně jako planktonické buňky, a že za přežívání populace je zodpovědná frakce perzistorů. Perzistující buňky jsou přítomny jak v biofilmech, tak v planktonických kulturách. V planktonické populaci buněk se vyskytuje 0,001 % buněk, které přežívají MIC, tedy i zde je malé procento perzistorů. V kultuře planktonických buněk ve stacionární fázi je procento těchto perzistorů
96 Mikrobiální biofilmy 8 9. Biofilmy a antimikrobiální látky vyšší (cca 1 %) a stejně tak v biofilmu (0,1 %) (Spoering& Lewis 2001). Vyšší percentuální zastoupení perzistorů ve stacionární fázi planktonického růstu, než v biofilmu, vysvětlují autoři tím, že ve stacionární fázi buňky nerostou, kdežto v biofilmu ano, i když velmi pomalu (Spoering& Lewis 2001; Keren et al., J Bacteriol. 2005). Tuto negativní korelaci přítomnosti perzistorů a metabolické aktivity potvrdili i ve svých dalších studiích. Zjistili, že se perzistoři z populace stacionární fáze rychle ztratí, když populaci rozředí čerstvým médiem (Lewis 2005). Zdá se, že čím méně bakteriální kultura roste a množí se, tím více perzistorů se v této populaci vyskytuje. Protože závislost procenta perzistorů na růstové fázi může být i quorum-sensing fenoménem, zaměřili se Keren et al. (J Bacteriol. 2005) i na tuto možnost. Udělali jednoduchý standardní test pro potvrzení quorum-sensing signálů v kultuře do kultury buněk v logaritmické fázi růstu přidali médium použité předtím pro kultivaci stacionární kultury (tedy médium s případnými signálními molekulami) a měřili procento perzistorů. Vzhledem k tomu, že přídavek tohoto média procento perzistorů v logaritmické fázi nezvýšil ani u E. coli ani u P. aeruginosa, nasvědčuje všechno tomu, že quorum-sensing nemá s přítomností perzistorů nic společného (Lewis 2005). Na základě těchto pozorování se většina autorů přiklání k závěru, že tvorba perzistorů je závislá na stupni metabolické aktivity. Podle studie Drenkard& Ausubel 2002 a et al. (2002) u perzistorů dochází k dočasným fenotypovým změnám, jež jsou spojovány se schopností bakterií tvořit biofilm (např. vysoká povrchová hydrofobicita, která vede ke zvýšenému přichycování buněk a vzniku biofilmu s vyšším množstvím biomasy). Tito autoři též identifikovali regulační protein, který reguluje změnu mezi rezistentní a citlivou formou P. aeruginosa. Ukázalo se, že tento regulační protein reguluje kromě rezistence k antibiotikům také tvorbu biofilmu, což nasvědčuje tomu, že fenotypová změna a biofilmová rezistence mohou být vzájemně úzce propojeny. Fenotypová změna spojená se zvýšenou rezistencí byla popsána i u několika dalších bakteriálních druhů, reverzibilní přechod z citlivé do rezistentní fenotypové formy byl popsán u kmenů S. aureus vystavených působení gentamicinu (Massey et al., 2001). Navíc, výskyt fenotypové změny, vedoucí ke zvýšenému shlukování a tvorbě biofilmu, je shodně spojován s rezistencí k chloru, kyselinám, oxidačnímu a osmotickému stresu u řady bakterií, naznačujíc, že změny ve fenotypové charakteristice spojené se zvýšenou tvorbou biofilmu jsou důležité ve vzrůstající rezistenci k environmentálním stresům (Anriany et al. 2001; Wai et al. 1998; Yildiz& Schoolnik 1999 et al., 1999). Není důvod předpokládat, že by některý bakteriální druh tuto schopnost neměl. I když je existence perzistorů velmi dobře popsána, zatím se stále lépe ví spíše to, co perzistoři nejsou, než co skutečně jsou. Perzistoři nejsou mutanti a nevznikají žádnou genetickou mutací. Lze též říci, že perzistoři nejsou žádnou speciální fází buněčného cyklu, i když i tyto teorie se objevily (Black et al., 1994; Falla et al., 1998). Při četnosti výskytu perzistorů v planktonické populaci 10-6 by tato fáze buněčného cyklu totiž u buňky trvala 1,8 milisekundy (jako generační doba je bráno v úvahu 30 minut, vzhledem k tomu, že v jednom okamžiku zachytíme v populaci pouze 10-6 bakterií-perzistorů, vycházela by délka trvání této fáze buněčného cyklu právě na 1,8 milisekundy). Vzhledem k rychlosti, s jakou probíhají buněčné děje, by to znamenalo obrovskou časovou synchronizaci všech procesů, i jen pro jednoduché vypnutí enzymatické reakce (Lewis 2001). Perzistoři rovněž nejsou speciální buňky v dormantním stavu bez růstu. Perzistoři by mohli být defektními bakteriemi, které ztratily schopnost programované buněčné smrti (Lewis 2001; Spoering& Lewis 2001) (viz kapitolka Programovaná buněčná smrt). Tato hypotéza předpokládá, že antimikrobiální látky nejsou přímo zodpovědné za bakteriální smrt, ale že způsobí buněčné poškození, které nepřímo vyvolá programovanou buněčnou smrt, stejně jako u mnohobuněčných organizmů vyvolá poškození buňky různými toxiny apoptózu (Spoering& Lewis 2001). Pokud by tomu tak skutečně bylo, bylo by stále možno pomocí
97 Mikrobiální biofilmy 9 9. Biofilmy a antimikrobiální látky antibiotik zasáhnout celou bakteriální biofilmovou populaci. Lewis jde však ve svém předpokladu dál a navrhuje, že perzistoři jsou buňky mají defektním programem pro buněčnou smrt. Tuto hypotézu podkládá mj. existencí mutací, které ovlivňují schopnost bakterií přežít působení antimikrobiálních látek bez molekulárních změn rezistence (exprimace genů hip, mar apod.). Podle další teorie jsou perzistoři určitým typem altruistických buněk, které se zříkají svého buněčného dělení ve prospěch celku. Proto, aby buněčná kultura mohla přežít, potřebuje se nejen hodně množit, ale musí mít i zálohu v buňkách, které ji nahradí v případě nečekané katastrofy (Lewis 2005; Lewis, 1998). Jiní autoři jdou ještě dále a odlišují od sebe pouhé fenotypové varianty a perzistory. Zásadní rozdíl mezi perzistory a fenotypovými variantami vidí v tom, že perzistoři jednoduše přežívají pokusy o eradikaci antimikrobiálními látkami, zatímco fenotypové varianty jsou pouze schopné růst za zvýšených koncentrací antimikrobiálních látek (Drenkard 2003) Vznik biofilmového fenotypu Již před patnácti lety se objevila v lékařské mikrobiologii teorie, že primární adheze bakterií k povrchu může vést k indukci či represi genů, které mohou dále ovlivnit velké množství fyziologických odpovědí (Dagostino et al., 1991). DNA microarray analýzy ukázaly, že genová exprese se mezi biofilmovými a planktonickými buňkami u Bacillus subtilis liší o 6 %, a u P. aeruginosa o 1 % (Stanley et al. 2003; Whiteley et al. 2001). U S. epidermidis se odlišná genová exprese projevuje mj. expresí sara (staphylococcal accessory regulator) a icaadbc operonu (Beenken et al. 2004; Cramton et al. 1999). Další teorií mechanizmu biofilmové rezistence je tedy možnost indukce biofilmspecifického genotypu u určité subpopulace buněk. Tento specifický genotyp pak může vést k exprimaci aktivních mechanizmů potlačujících účinky antimikrobiálních látek (Maira-Litran et al. 2000b; Maira-Litran et al. 2000a; Cochran et al. 2000b). Předpokládá se, že buňky po přisednutí k povrchu začínají exprimovat nějaký typ biofilmového fenotypu a mnoho autorů se zaměřilo na identifikaci genů, které jsou aktivovány či potlačeny v biofilmu ve porovnání s planktonickými buňkami (Kuchma& O'toole 2000; Fitzpatrick et al. 2006; Drenkard& Ausubel 2002 a další). Je možné, že fenotyp zvýšené rezistence k antimikrobiálním agens exprimují ve větší či menší míře všechny buňky v biofilmu, nebo naopak pouze určitá část těchto buněk. Tento rezistentní fenotyp může být indukován nedostatkem živin, určitým typem stresu, vysokou hustotou bakteriálních buněk nebo kombinací těchto a mnohých dalších faktorů. Molekulární mechanizmy této biofilmové rezistence se mohou vztahovat k potlačení bakteriálního růstu uvnitř biofilmu, k fyzikálně-chemickým interakcím matrix biofilmu s určitými antibiotiky (vazba dipól-dipól, tvorba komplexních sloučenin apod.) (Wagman et al. 1975, as cited in Schierholz& Beuth 2001), mohou nějakým způsobem ovlivnit v bakteriální buňce cílové místo antimikrobiálního působení látky atd. Fenotyp rezistence spojený s primární adhezí Bakterie detekují blízkost pevného povrchu a již v tu chvíli začínají pozitivně regulovat produkci extracelulárních polysacharidů a velmi rychle po primární adhezi pak mění svou citlivost k antimikrobiálním látkám (Ashby et al. 1994; Das et al. 1998). Das et al et al. (1998) prokázali, že citlivost P. aeruginosa a S. aureus k řadě různých antimikrobiálních látek se změnila velmi rychle po buněčné adhezi a počátku tvorby biofilmu. V některých případech prokázali tří až pětinásobný vzestup rezistence okamžitě po přisednutí. Vzhledem k rychlé fenotypové změně se za mechanizmus této rezistence považuje spíše zábrana penetrace či přítomnost odtokových pump, než změna receptoru bakteriální buňky, který je cílovým místem působení antimikrobiální látky (Gilbert et al. 2002). V další studii Fujiwara et al et al. (1998) prokázali, že po jednohodinové inkubaci se MBC
98 Mikrobiální biofilmy Biofilmy a antimikrobiální látky adherovaných buněk P. aeruginosa, Serratia marcescens a Proteus mirabilis signifikantně zvýšila. Hladina zvýšené rezistence, kterou lze pozorovat okamžitě po přisednutí bakterií k pevnému povrchu, ale ještě před počátkem tvorby biofilmové vrstvy, je obecně mnohem nižší, než můžeme pozorovat ve vyzrálých biofilmech. Davies et al et al. (1998) publikovali, že tato změna v citlivosti by mohla být zprostředkována akumulací HSL-like signálních molekul. V tom případě by se zde naskytla eventuální možnost podání HSL antagonistů, jako jsou například furanony. Fenotyp rezistence spojený s produkcí MDR pump Současné práce se zaměřily na identifikaci genů, které mohou přispívat k fenotypu této zvýšené rezistence. Prvním z těchto mechanizmů rezistence, který by mohl ovlivňovat biofilmovou rezistenci jsou tzv. širokospektré odtokové pumpy (multidrug efflux pumps). Ty mohou vyloučit z buňky i chemicky nepříbuzné antimikrobiální látky. Exprese mar operonu u E. coli vede k fenotypu širokospektré rezistence (Mah& O'toole 2001). Zaměříme-li se na otázku, zda tento známý MDR (Multi Drug Resistance) systém se účastní biofilmové rezistence k antimikrobiálním látkám, výsledky výzkumů tuto teorii nepodporují. Exprese operonu mar je v biofilmové kultuře mnohem nižší, než v kultuře stacionární fáze planktonických buněk (Maira-Litran et al. 2000a). Stejní autoři se zaměřili též na testování mutantů s chybějícími lokusy mar a acrab, aby zjistili, zda je biofilmová rezistence E. coli k ciprofloxacinu ovlivněná ztrátou těchto lokusů (Maira-Litran et al. 2000b). Ztráta genů mar a acrab neovlivnila nijak negativně biofilmy E. coli, ale na druhou stranu určitý stupeň exprese acrab poskytoval částečnou ochranu proti účinkům ciprofloxacinu. Ačkoliv tyto výsledky naznačují že pozitivní regulace genů pro MDR systém specificky neovlivnila rezistenci biofilmu E. coli k antimikrobiálním látkám, nevylučují možnost případné existence jiných systémů MDR pump, které by byly indukovány jako odpověď na růst ve formě biofilmu. Například u P. aeruginosa jsou dosud známé tři druhy MDR pump a u dalších několika systémů, kódovaných v genomu P. aeruginosa, se funkce MDR pump předpokládá. Brooun et al et al. (2000) prokázali význam jedné z těchto pump, MexAB-OprM, v biofilmové rezistenci k ofloxacinu, když přišli na to, že při nízkých koncentracích ofloxacinu byly biofilmy bez této MDR pumpy citlivější k tomuto antibiotiku a naopak. Pro další chinolon, ciprofloxacin, se však nepodařilo prokázat žádný rozdíl. Proto odpověď na otázku, jak velkou roli hrají MDR pumpy v biofilmové rezistenci zůstává pořád nejasná. Další mechanismus rezistence, který může být indukován biofilmovými buňkami, je změna membránových proteinů či jejich složení jako odpověď na působení antimikrobiálních látek. Tato změna vede ke snížené permeabilitě buňky k těmto látkám. Například mutace v genech ompb a ompf (regulátorové geny kódující porinové proteiny OmpF a OmpC ve vnější membráně) zvyšuje rezistenci E. coli k β-laktamovým antibiotikům (Jaffe et al., 1982). Mutanty, které nemají porinový protein OmpF, jsou rezistentnější k chloramfenikolu a tetracyklinu (Pugsley et al., 1978). Navíc, u buněk kultivovaných při nedostatku živin, tedy určitým způsobem strádajících, je relativní poměr hlavních porinů E. coli, OmpC a OmpF, změněn ve prospěch produkce menšího porinu, OmpC (Liu et al., 1998). Tyto výsledky podporují předpoklad, že změna exprese porinů ovlivňuje skutečnou rezistenci bakterií k antimikrobiálním látkám. Navíc bylo prokázáno, že exprese ompc a tří dalších osmoticky regulovaných genů byla zvýšená u biofilmových bakterií ve srovnání s planktonickými buňkami (Prigent-Combaret& Lejeune 1999), což nasvědčuje tomu, že bakterie v biofilmu žijí skutečně v prostředí s vyšším osmotickým stresem. Z toho důvodu též mohou environmentální podmínky v biofilmu vést ke změně buněčných povrchů, což chrání bakterie před zhoubným účinkem antimikrobiálních látek.
99 Mikrobiální biofilmy Biofilmy a antimikrobiální látky Další mechanizmy uplatňující se v biofilmové rezistenci Quorum sensing Role quorum sensing systému v biofilmové rezistenci k antimikrobiálním látkám není dosud zcela jasná. Práce, kterou publikovali Davies et al et al. (1998), prokázala, že mutant P. aeruginosa s absencí lasr-lasi quorum sensing systému nebyl schopen vytvořit biofilm s normální strukturou. Tito autoři též prokázali, že lasi mutantní biofilmy byly abnormálně citlivé k působení SDS. Na druhou stranu Brooun et al et al. (2000) neprokázali mezi normálními buňkami a mutanty s defektním quorum sensing systémem v rezistenci k detergentům a antibiotikům žádný rozdíl. Skutečnou roli quorum sensing systému v biofilmové rezistenci nám navíc komplikuje fakt, že σ faktor RpoS hraje určitou roli i v regulaci quorum sensing systému (Whiteley et al., 2000). Teorii, že quorum sensing hraje určitou úlohu i v biofilmové rezistenci podporuje i fakt, že rezistentní biofilmové buňky mohou svou citlivost k antimikrobiálním látkám velmi rychle znovu nabýt, jsou-li přeneseny do čerstvého média, které nejen poskytuje živiny (a tím umožňuje rychlejší růst), ale snižuje také koncentraci ochranných mezibuněčných signalizačních molekul (Anderl et al. 2003). Tato reverzibilita rovněž dokazuje, že biofilmová rezistence k antimikrobiálním látkám je spíše fenotypového typu, než výsledek genetických změn. Stáří biofilmu a jeho vliv na rezistenci Biofilmová rezistence k antimikrobiálním látkám začíná ve fázi přisednutí na pevný povrch a se stářím biofilmu se zvětšuje (Knobloch et al. 2002). Např. Monzon et al et al. (2002) ve své studii prokázali, že rezistence S. epidermidis k vankomycinu roste s jeho stářím (studovali biofilm v rozmezí 6 48 hodin starý). Tomuto pozorování odpovídají i klinické zkušenosti, kdy v ortopedii odstranění infikovaného ložiska s ponecháním protézy je úspěšnější v případě časných infekcí, než v případě infekce pozdní pooperační, které měly čas se určitou dobu vyvíjet (Patel 2005). Anwar et al et al. (1992) zjistili, že starší (desetidenní) biofilm P. aeruginosa byl signifikantně rezistentnější k tobramycinu a piperacilinu, než mladší (dvoudenní) biofilm. Dávka 500 µg tobramycinu s 5 µg piperacilinu v 1 ml kompletně inaktivovala jak planktonické buňky, tak buňky mladého (dvoudenního) biofilmu. U buněk ve starším, desetidenním, biofilmu byl při stejné dávce antibiotik jejich počet zredukován cca o 20 %. Podobné výsledky získali i další autoři (Amorena et al. 1999; Desai et al. 1998). Úzká vazba mezi bakteriálními buňkami v biofilmu V biofilmu se mohou díky užšímu kontaktu buněk snáze šířit plazmidy a extrachromozomální úseky DNA. Vzhledem k tomu, že právě plazmidy a transpozony kódují rezistenci k řadě antimikrobiálních látek (na plazmidech již byly prokázány geny rezistence ke všem v současnosti užívaným antibiotikům). Je již dlouho známo, že mnoho bakteriálních druhů je schopno sdílet své plazmidy s jinými bakteriálními druhy. Ehlers et al. (1999) prokázali přenos plazmidů konjugací mezi různými gramnegativními bakteriemi i v biofilmu. Četnosti tohoto horizontálního přenosu genetické informace byly několikařádově vyšší právě v biofilmové kultuře, než v kultuře planktonické stejného mikroba. Ke stejným výsledkům došli i ostatní autoři (Roberts et al. 1999; Hausner& Wuertz 1999; Christensen et al. 1998). Důvodem této zvýšené četnosti přenosu genetické informace může být například vyšší pravděpodobnost kontaktu mezi jednotlivými buňkami nebo zanedbatelný efekt střižných sil,
100 Mikrobiální biofilmy Biofilmy a antimikrobiální látky které jinak v planktonické kultuře znamenají zrušení kontaktu mezi buňkami nebo poškození pili potřebných pro konjugaci (Donlan 2001). Další případy zvýšené komunikace mezi buňkami v biofilmu jsou známy z klinické praxe Hoiby 2002 (2002) prokázala zvýšenou četnost mutací u P. aeruginosa v plicích při cystické fibróze. Hypermutantní kmeny P. aeruginosa zde dosahovaly až 36% zastoupení. Vzhledem k tomu, že P. aeruginosa se ve sputu pacientů s cystickou fibrózou může nacházet až v množstvích CFU/ml napomáhá tento úzký kontakt možnostem spontánních mutací. Programovaná buněčná smrt Další hypotézou o zvýšené rezistenci biofilmů je programovaná buněčná smrt a potenciál poškozených bakteriálních buněk podrobit se apoptóze nebo programované buněčné smrti (Gilbert et al. 2002). Lewis 2001 (2001) navrhl, že smrt buněk, která následuje po účinku antimikrobiálních látek, není důsledkem přímého účinku látky, ale programovaného sebevražedného mechanizmu, programované buněčné smrti. Jak již bylo zmíněno výše, určitá subpopulace buněk, perzistorů, dokáže přežít koncentrace antimikrobiálních látek mnohem vyšší, než je MBC a tyto koncentrace pak dosahují extrémních hodnot i tisícinásobku MIC apod. Tyto buňky však po následné kultivaci nevykazují zvýšenou rezistenci k antimikrobiálním látkám. Proto někteří autoři předpokládají, že tyto buňky jsou defektními mutanty, které nejsou schopny podstoupit programovanou buněčnou smrt (Brooun et al. 2000; Lewis 2001). Takové buňky mohou napomáhat i rezistenci biofilmové. Problematika biofilmové rezistence u kvasinek Kvasinky rostoucí ve formě biofilmu jsou, stejně jako bakterie, také rezistentnější k antimykotikům, ačkoliv mechanizmy této zvýšené rezistence jsou zatím stále neznámé. Kvasinkový biofilm totiž obsahuje velmi málo exopolysacharidové matrix (Hawser& Douglas 1995), která by buňkám ukrytým uvnitř mohla poskytovat určitou ochranu. Kvasinkový biofilm je rezistentní vůči všem běžným antimykotikům (flukonazol, nystatin, flucytosin, amfotericin B apod.). Stejně tak se udává rezistence i vůči novým triazolovým antimykotikům (vorikonazol a ravukonazol). Určitou nadějí se dle nedávných výzkumů zdají být liposomální formule amfotericinu a echinokandiny (Kuhn et al. 2002b), což by vůči invazivním kvasinkovým infekcím znamenalo výraznou výhodu. Buněčné mechanizmy bakteriální rezistence Buněčné mechanizmy bakteriální rezistence nejsou sice specifikem biofilmpozitivních bakterií, ale vzhledem k tomu, že i tyto mechanizmy rezistence biofilmpozitivní mikroby využívají, patří se o nich alespoň v krátkosti zmínit. K bližšímu studiu může sloužit např. Votava (2005), Lochman (1999), Hynie (2001). Mechanizmy buněčné rezistence lze také rozdělit na fenotypovou adaptaci a změnu geneticky podmíněnou. Fenotypová adaptace je určité přizpůsobení se bakterie na změněné metabolické pochody. Může k tomu docházet např. potlačením různých represorů genové transkripce s následným uvolněním nových metabolických pochodů. Spadá sem například produkce enzymu penicilinázy v přítomnosti penicilinu v prostředí. Jsou to změny částečné a přechodné a dají se zvládnout buď vyšší dávkou antibiotika, nebo jeho úplným vysazením, čímž se obnoví původní metabolické pochody. Genetická modifikace bývá podmíněna buď chromozomálně, tj. modifikací genu na chromozomu, nebo převzetím genetického materiálu od rezistentních buněk, např. pomocí plazmidů a její mechanizmy můžeme rozdělit do následujících typů: změna cílové molekuly
101 Mikrobiální biofilmy Biofilmy a antimikrobiální látky impermeabilita buněčné stěny a použití aktivních pump intrinsic rezistence enzymatická inaktivace Změna cílové molekuly Většina antibiotik působí na metabolické pochody tím, že se vážou na proteiny bakteriální buňky. Ke vzniku rezistence postačuje změnit cílovou molekulu, aby se nemohlo navázat antibiotikum. Tento mechanizmus rezistence je známý např. u rezistence na makrolidy, linkosamidy, ale i β-laktamy. Impermeabilita buněčné stěny a použití aktivních pump Antibiotika se transportují dovnitř buňky aktivním transportem, tedy pomocí bakteriálních přenašečů. Změnou molekul tohoto přenašeče vniká rezistence k antibiotikům. Další možností zabránění aktivnímu transportu dovnitř buňky je snížení produkce ATP (aktivní transport je energeticky velmi náročný proces). Impermeabilita buněčné stěny se uplatňuje např. u aminoglykosidů, tetracyklinu či chinolonů. Použití aktivních pump bylo popsáno již v kapitole Fenotyp rezistence spojený s produkcí MDR pump. Aktivní odtokové pumpy mohou vyloučit z buňky i chemicky nepříbuzné látky. Intrinsic rezistence Bakterie může mít pro jednu funkci dva enzymy. Pokud je jeden z nich citlivý k působení antibiotika, a tedy může být pro poškozen, bakterie místo původního může využít enzym náhradní, který je kódovaný plazmidicky. Enzymatická inaktivace Enzymatická inaktivace je nejobvyklejším mechanizmem přenosné rezistence. Rezistentní kmeny produkují enzymy, které buď štěpí molekuly antibiotika a činí je neúčinnými, nebo antibiotika substituují a obsazují jim vazebná místa na cílových molekulách. Patří sem enzymy rozkládající β-laktamy různé β-laktamázy (penicilinázy, cefalosporinázy karbapenemázy aj.), enzymy modifikující aminoglykosidy např. N-acetyltransferázy, O-fosfotransferázy, O-nukleotidyltransferázy apod Závěr Je více než zřejmé, že k biofilmové rezistenci k antimikrobiálním látkám přispívá mnoho faktorů a velmi záleží na tom, o který bakteriální druh a o které antibiotikum se jedná, abychom mohli alespoň předpokládat, které mechanizmy ta která bakterie využije. Environmentální heterogenita, existující v biofilmové vrstvě, též vede ke vzniku heterogenních buněčných populací, jenž pak mohou vykazovat různé stupně rezistence k různým antimikrobiálním látkám. Tak například buňky nejblíže k povrchu biofilmu mohou být chráněny vrstvou polysacharidové matrix a produkcí enzymů. Buňky rostoucí ve střední vrstvě biofilmu mohou růst pomalu a být chráněny právě svým pomalým růstem a dále tou vnější vrstvou mikrobů. A populace buněk rostoucích ve spodní vrstvě biofilmu může vykazovat biofilm-specifický fenotyp rezistence indukovaný konkrétními stresovými faktory vnějšího prostředí, které tyto buňky obklopuje.
102 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 9.2 Možnosti ovlivnění biofilmové rezistence vnějšími vlivy Vzhledem k problémům, jež nám způsobuje rezistence biofilmů k antimikrobiálním látkám, zkoušejí se i jiné varianty boje s mikrobiálními biofilmy. Jak bylo popsáno v předchozí části, koncentrace antibiotik, které jsou schopné eradikovat biofilm, jsou o několik řádů vyšší, než koncentrace MIC a jsou tedy v humánní medicíně prakticky nevyužitelné. Testovány byly následující strategie: narušení mikrobiální adherence inhibice systému quorum sensing degradace extracelulární matrix bioakustický efekt bioelektrický efekt bioradiační efekt kombinace antibiotik Inhibice systému quorum sensing Regulace vzniku biofilmu využitím mezibuněčné bakteriální komunikace (např. systém quorum sensing) nám poskytuje nový potenciální cíl v boji s biofilmy. Biofilmy vytvořené mutantem P. aeruginosa, který nemá schopnost produkovat signály quorum sensing, jsou slabší s blíže umístěnými buňkami, mají méně viditelné extracelulární matrix a jsou citlivější ke kanamycinu, než jsou biofilmy vytvořené původním kmenem (Shih& Huang 2002). Mezi látky, které inhibují produkci signálů quorum sensing patří např. furanony, RNAIII-inhibující peptid (RNAIII-inhibiting peptide, RIP) aj. Syntetické deriváty přírodních furanonových složek, které se našly v mořském prostředí, působí jako silní antagonisté quorum sensing systému u P. aeruginosa a zvyšují in vitro citlivost biofilmu P. aeruginosa k antimikrobiálním látkám, jako např. k tobramycinu. Bylo též bylo dokázáno, že furanony zvyšují clearance P. aeruginosa v myším modelu pulmonární infekce (Hentzer et al. 2003). RIP, tedy peptid inhibující RNAIII je heptapeptid, který narušuje tvorbu biofilmu u S. aureus a S. epidermidis podle všeho též narušením mechanizmů quorum sensing (Balaban et al. 2003). RNAIII-inhibující peptid působí synergicky s konvenčními antimikrobiálními látkami a bylo in vivo prokázáno, že eliminuje transplantační infekce způsobené S. epidermidis. Na modelech biofilmů Staphylococcus aureus a Candida dubliniensis byla rovněž studována synergie mezi farnesolem, což je signální molekula quorum sensing, a některými antibiotiky a antimykotiky (Jabra-Rizk et al. 2006b; Jabra-Rizk et al. 2006a) Inhibice mikrobiální adherence Furanony jsou rovněž antagonistou HSL a HSL-like signálních molekul, které začínají bakterie produkovat podle některých autorů téměř okamžitě po adhezi na pevný povrch. K produkci těchto signálních molekul dochází ještě před začátkem tvorby biofilmové vrstvy a nárůst rezistence pozorovaný okamžitě po bakteriální adhezi na povrch je mnohem nižší, než nárůst rezistence pozorovaný u vyzrálých biofilmů. Tito autoři se též domnívají. že tento nárůst rezistence by mohl být zprostředkován právě akumulací HSL-like signálních molekul (Davies et al. 1998). Pomocí furanonů bychom tedy mohli působit i tomto ranném stádiu vzniku biofilmu.
103 Mikrobiální biofilmy Biofilmy a antimikrobiální látky Bioakustický efekt Aplikace ultrazvuku v hladinách bez inhibiční či baktericidní aktivity (67kHz) zvyšuje inhibiční a baktericidní účinky gentamicinu proti planktonickým kulturám P. aeruginosa a E. coli (Pitt et al. 1994). Bylo dokonce zjištěno, že se stářím kultury se bakterie stávají rezistentnější k působení samotného gentamicinu, použití ultrazvuku však dokáže tuto rezistenci potlačit. Pitt& Ross 2003 et al. (2003) též ukázali, že nízkofrekvenční (70 khz) a nízkoenergetický (< 2 W/cm 2 ) ultrazvuk zvyšuje růstovou rychlost S. epidermidis, P. aeruginosa a E. coli adherovaných k polyetylenovému (PE) povrchu v porovnání s růstem bez působení ultrazvuku, a že ultrazvuk zvyšoval planktonický růst S. epidermidis a jiných planktonických bakterií. Ultrazvuk rovněž zvyšuje transport gentamicinu skrz biofilmovou vrstvu (Carmen et al. 2004a). Rediske et al. 1999; Rediske et al et al., (1999; 2000) subkutánně implantovali PE disky s kulturou E. coli na záda králíka a použili nízkofrekvenční (28,48 khz) a nízkoenergetický (300 mw/cm 2 ) ultrazvuk po 24 hodin spolu se systémovým podáváním gentamicinu a bez něj. Zatímco samotná expozice ultrazvuku nezpůsobila v životaschopnosti bakterií výrazné rozdíly, v přítomnosti gentamicinu došlo k podstatnému snížení vitality. Bohužel bylo zároveň pozorováno poškození kožní tkáně obou sledovaných králíků. Podobná studie byla provedena s PE disky porostlými biofilmem S. epidermidis, ultrazvukem a systémovým podáváním vankomycinu (Carmen et al. 2004b). Biofilm S. epidermidis odpovídal nejlépe na kombinaci ultrazvuku a vankomycinu, ale ve srovnání s E. coli a léčbou gentamicinem bylo zapotřebí delšího působení obou složek Bioelektrický efekt Šance na eradikaci biofilmpozitivních bakterií zvyšuje též, v porovnání s antibiotikem samotným, použití slabého elektrického proudu kombinovaného s antimikrobiálními látkami. Bioelektrický efekt umožnil snížit vysoké koncentrace aminoglykosidů, tetracyklinů či chinolonů, nezbytné k eradikaci biofilmu až k hladinám, které odpovídají likvidaci planktonických buněk stejných druhů (Caubet et al. 2004; Pickering et al. 2003; Wattanakaroon& Stewart 2000). Mechanizmus účinku bioelektrického efektu ale není dosud kompletně vysvětlen a rovněž nebyl dokázán in vivo (Costerton et al. 1994; Stewart et al. 1999) Bioradiační efekt Dalším způsobem, kterým je možné zničit bakterie uvnitř biofilmu, je radioaktivní ozáření. Princip akce je jednoduchý a atraktivní, předpokládá se totiž, že po cíleném doručení lokálního zářiče k biofilmu dojde k jeho zničení v důsledku kontrolované radiační expozice. Nedávno byla publikována studie, která hodnotila in vitro efektivitu α-zářiče (Bismut 213 ). Ten byl navázán na monoklonální protilátky tak, aby došlo ke specifické vazbě na polysacharidy biofilmu (Martinez et al. 2006). Na modelu s Cryptococcus neoformans došlo k 50% redukci metabolické aktivity biofilmu. Pozoruhodné bylo i to, že se nepodařilo prokázat snížení metabolické aktivity biofilmu při použití β- či γ-zářičů, což by mohlo souviset s ochrannými vlastnostmi matrix biofilmu Degradace extracelulární matrix Lysostaphin, glycylglycin-endopeptidáza, specificky štěpí pentaglycinové můstky nalézající se ve stafylokokovém peptidoglykanu a rychle zabíjí planktonické buňky S. aureus i v nízkých koncentracích (MIC 90 se pohybuje v rozmezí 0,001 0,064 µg/ml) a S.
104 Mikrobiální biofilmy Biofilmy a antimikrobiální látky epidermidis v koncentracích o něco vyšších (MIC 90 se pohybuje v rozmezí 12,5 64 µg/ml) (Wu et al. 2003). Wu et al et al. (2003) ve své studii rovněž prokázali, že lysostaphin rozrušoval extracelulární matrix biofilmu S. aureus in vitro na plastových a skleněných površích. Eradikoval však pouze lysostaphin-citlivé kmeny S. aureus, a to v koncentracích tak nízkých jako je 1 µg/ml. Lysostaphin však nedokázal eradikovat lysostaphin-rezistentní buňky S. aureus. Vyšší koncentrace lysostaphinu rozrušovaly také biofilmy S. epidermidis. Podobně působí i další složky lidských sekretů, jako například laktoferrin, který blokuje tvorbu biofilmu u P. aeruginosa (Singh et al. 2002). Účinným nástrojem k degradaci biofilmu by mohly být i další enzymy. Bylo například zjištěno, že kombinace streptokinázy a streptodornázy jsou účinné proti biofilmu S. aureus a P. aeruginosa (Nemoto et al. 2003; Nemoto et al. 2000). Jiní výzkumníci testovali antibiofilmové aktivity oxidoreduktázy nebo komplexní směsi polysacharidy-hydrolyzujících enzymů (Johansen et al. 1997). Hatch et al. (1998) prokázali, že použití alginát-lyázy umožňuje zvýšenou difúzi aminoglykosidů přes alginát biofilmu P. aeruginosa. Allicin, produkt rostlin rodu Allium, je sloučenina s obsahem síry, která se v rostlinách tvoří v malých množstvích enzymatickým působením allinázy na alliin. In vitro se podařilo prokázat, že je aktivní proti S. epidermidis a C. albicans a rovněž snižuje tvorbu biofilmu u S. epidermidis a C. albicans (Perez-Giraldo et al. 2003; Shuford et al. 2005). Literatura ke kapitole Biofilmy a ATB 1. Adams JL, McLean RJ (1999): Impact of rpos deletion on Escherichia coli biofilms. Appl.Environ.Microbiol. 65: Amorena B, Gracia E, Monzon M, Leiva J, Oteiza C, Perez M, Alabart JL, Hernandez- Yago J (1999): Antibiotic susceptibility assay for Staphylococcus aureus in biofilms developed in vitro. Journal of Antimicrobial Chemotherapy 44: Anderl JN, Franklin MJ, Stewart PS (2000): Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob.Agents Chemother. 44: Anderl JN, Zahller J, Roe F, Stewart PS (2003): Role of nutrient limitation and stationary-phase existence in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob.Agents Chemother. 47: Anriany YA, Weiner RM, Johnson JA, de Rezende CE, Joseph SW (2001): Salmonella enterica serovar Typhimurium DT104 displays a rugose phenotype. Appl.Environ.Microbiol. 67: Anwar H, Strap JL, Chen K, Costerton JW (1992): Dynamic interactions of biofilms of mucoid Pseudomonas aeruginosa with tobramycin and piperacillin. Antimicrob.Agents Chemother. 36: Ashby MJ, Neale JE, Knott SJ, Critchley IA (1994): Effect of antibiotics on nongrowing planktonic cells and biofilms of Escherichia coli. J.Antimicrob.Chemother. 33: Balaban N, Giacometti A, Cirioni O, Gov Y, Ghiselli R, Mocchegiani F, Viticchi C, Del Prete MS, Saba V, Scalise G, Dell'Acqua G (2003): Use of the quorum-sensing inhibitor RNAIII-inhibiting peptide to prevent biofilm formation in vivo by drug-resistant Staphylococcus epidermidis. J.Infect.Dis. 187: Beenken KE, Dunman PM, McAleese F, Macapagal D, Murphy E, Projan SJ, Blevins JS, Smeltzer MS (2004): Global gene expression in Staphylococcus aureus biofilms. J.Bacteriol. 186:
105 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 10. Brooun A, Liu S, Lewis K (2000): A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob.Agents Chemother. 44: Brown MR, Barker J (1999): Unexplored reservoirs of pathogenic bacteria: protozoa and biofilms. Trends Microbiol. 7: Carmen JC, Nelson JL, Beckstead BL, Runyan CM, Robison RA, Schaalje GB, Pitt WG (2004a): Ultrasonic-enhanced gentamicin transport through colony biofilms of Pseudomonas aeruginosa and Escherichia coli. J.Infect.Chemother. 10: Carmen JC, Roeder BL, Nelson JL, Beckstead BL, Runyan CM, Schaalje GB, Robison RA, Pitt WG (2004b): Ultrasonically enhanced vancomycin activity against Staphylococcus epidermidis biofilms in vivo. J.Biomater.Appl. 18 : Caubet R, Pedarros-Caubet F, Chu M, Freye E, de Belem RM, Moreau JM, Ellison WJ (2004): A radio frequency electric current enhances antibiotic efficacy against bacterial biofilms. Antimicrob.Agents Chemother. 48: Ceri H, Olson ME, Stremick C, Read RR, Morck D, Buret A (1999): The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J.Clin.Microbiol. 37: Christensen BB, Sternberg C, Andersen JB, Molin OS (1998): In situ detection of gene transfer in a model biofilm engaged in degradation of benzyl alcohol. APMIS Suppl. 84:25-8.: Cochran WL, McFeters GA, Stewart PS (2000a): Reduced susceptibility of thin Pseudomonas aeruginosa biofilms to hydrogen peroxide and monochloramine. J.Appl.Microbiol. 88: Cochran WL, Suh SJ, McFeters GA, Stewart PS (2000b): Role of RpoS and AlgT in Pseudomonas aeruginosa biofilm resistance to hydrogen peroxide and monochloramine. J.Appl.Microbiol. 88: Coquet L, Junter GA, Jouenne T (1998): Resistance of artificial biofilms of Pseudomonas aeruginosa to imipenem and tobramycin. J.Antimicrob.Chemother. 42: Costerton JW, Ellis B, Lam K, Johnson F, Khoury AE (1994): Mechanism of electrical enhancement of efficacy of antibiotics in killing biofilm bacteria. Antimicrob.Agents Chemother. 38: Costerton JW, Stewart PS, Greenberg EP (1999): Bacterial biofilms: a common cause of persistent infections. Science 284: Cramton SE, Gerke C, Schnell NF, Nichols WW, Gotz F (1999): The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect.Immun. 67: Das JR, Bhakoo M, Jones MV, Gilbert P (1998): Changes in the biocide susceptibility of Staphylococcus epidermidis and Escherichia coli cells associated with rapid attachment to plastic surfaces. J.Appl.Microbiol. 84: Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP (1998): The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280: de Beer D, Srinivasan R, Stewart PS (1994): Direct measurement of chlorine penetration into biofilms during disinfection. Appl.Environ.Microbiol. 60: Desai M, Buhler T, Weller PH, Brown MR (1998): Increasing resistance of planktonic and biofilm cultures of Burkholderia cepacia to ciprofloxacin and ceftazidime during exponential growth. J.Antimicrob.Chemother. 42: Donlan RM (2001): Biofilms and device-associated infections. Emerg.Infect.Dis. 7:
106 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 28. Drenkard E (2003): Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes.Infect. 5: Drenkard E, Ausubel FM (2002): Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature. 416: Duguid IG, Evans E, Brown MR, Gilbert P (1992a): Effect of biofilm culture upon the susceptibility of Staphylococcus epidermidis to tobramycin. J.Antimicrob.Chemother. 30: Duguid IG, Evans E, Brown MR, Gilbert P (1992b): Growth-rate-independent killing by ciprofloxacin of biofilm-derived Staphylococcus epidermidis; evidence for cell-cycle dependency. J.Antimicrob.Chemother. 30: Dunne WM, Jr., Mason EO, Jr., Kaplan SL (1993): Diffusion of rifampin and vancomycin through a Staphylococcus epidermidis biofilm. Antimicrobial Agents and Chemotherapy 37: Evans DJ, Allison DG, Brown MR, Gilbert P (1990): Effect of growth-rate on resistance of gram-negative biofilms to cetrimide. J.Antimicrob.Chemother. 26: Evans DJ, Allison DG, Brown MR, Gilbert P (1991): Susceptibility of Pseudomonas aeruginosa and Escherichia coli biofilms towards ciprofloxacin: effect of specific growth rate. J.Antimicrob.Chemother. 27: Fedtke I, Gotz F, Peschel A (2004): Bacterial evasion of innate host defenses--the Staphylococcus aureus lesson. Int.J.Med.Microbiol. 294: Fitzpatrick F, Humphreys H, O'Gara JP (2006): Environmental regulation of biofilm development in methicillin-resistant and methicillin-susceptible Staphylococcus aureus clinical isolates. J.Hosp.Infect. 62: Fujiwara S, Miyake Y, Usui T, Suginaka H (1998): Effect of adherence on antimicrobial susceptibility of Pseudomonas aeruginosa, Serratia marcescens, and Proteus mirabilis. Hiroshima J.Med.Sci. 47: Fux CA, Costerton JW, Stewart PS, Stoodley P (2005): Survival strategies of infectious biofilms. Trends Microbiol. 13: Gilbert P, Allison DG, McBain AJ (2002): Biofilms in vitro and in vivo: do singular mechanisms imply cross-resistance? J.Appl.Microbiol. 92 Suppl:98S-110S.: 98S-110S. 40. Giwercman B, Jensen ET, Hoiby N, Kharazmi A, Costerton JW (1991): Induction of beta-lactamase production in Pseudomonas aeruginosa biofilm. Antimicrob.Agents Chemother. 35: Hausner M, Wuertz S (1999): High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis. Appl.Environ.Microbiol. 65: Hawser SP, Douglas LJ (1995): Resistance of Candida albicans biofilms to antifungal agents in vitro. Antimicrobial Agents and Chemotherapy 39: Hentzer M, Eberl L, Nielsen J, Givskov M (2003): Quorum sensing : a novel target for the treatment of biofilm infections. BioDrugs. 17: Hoiby N (2002): Understanding bacterial biofilms in patients with cystic fibrosis: current and innovative approaches to potential therapies. J.Cyst.Fibros. 1: Hola V, Ruzicka F, Tejkalova R, Votava M (2004): [Determination of sensitivity of biofilm-positive forms of microorganisms to antibiotics.]. Klin.Mikrobiol.Infekc.Lek. 10: Hoyle BD, Alcantara J, Costerton JW (1992): Pseudomonas aeruginosa biofilm as a diffusion barrier to piperacillin. Antimicrob.Agents Chemother. 36: Hoyle BD, Jass J, Costerton JW (1990): The biofilm glycocalyx as a resistance factor. J.Antimicrob.Chemother. 26: 1-5.
107 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 48. Huang CT, Xu KD, McFeters GA, Stewart PS (1998): Spatial patterns of alkaline phosphatase expression within bacterial colonies and biofilms in response to phosphate starvation. Appl.Environ.Microbiol. 64: Huang CT, Yu FP, McFeters GA, Stewart PS (1995): Nonuniform spatial patterns of respiratory activity within biofilms during disinfection. Appl.Environ.Microbiol. 61: Ishida H, Ishida Y, Kurosaka Y, Otani T, Sato K, Kobayashi H (1998): In vitro and in vivo activities of levofloxacin against biofilm-producing Pseudomonas aeruginosa. Antimicrob.Agents Chemother. 42: Jabra-Rizk MA, Meiller TF, James CE, Shirtliff ME (2006a): Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob.Agents Chemother. 50: Jabra-Rizk MA, Shirtliff M, James C, Meiller T (2006b): Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 6: Johansen C, Falholt P, Gram L (1997): Enzymatic removal and disinfection of bacterial biofilms. Appl.Environ.Microbiol. 63: Keren I, Shah D, Spoering A, Kaldalu N, Lewis K (2004): Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J.Bacteriol. 186: Knobloch JK, Von Osten H, Horstkotte MA, Rohde H, Mack D (2002): Minimal attachment killing (MAK): a versatile method for susceptibility testing of attached biofilm-positive and -negative Staphylococcus epidermidis. Med.Microbiol.Immunol.(Berl). 191: Konig C, Schwank S, Blaser J (2001): Factors compromising antibiotic activity against biofilms of Staphylococcus epidermidis. Eur.J.Clin.Microbiol.Infect.Dis. 20: Korber DR, James GA, Costerton JW (1994): Evaluation of Fleroxacin Activity against Established Pseudomonas fluorescens Biofilms. Appl.Environ.Microbiol. 60: Kuchma SL, O'toole GA (2000): Surface-induced and biofilm-induced changes in gene expression. Curr.Opin.Biotechnol. 11: Kuhn DM, Chandra J, Mukherjee PK, Ghannoum MA (2002a): Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infect.Immun. 70: Kuhn DM, George T, Chandra J, Mukherjee PK, Ghannoum MA (2002b): Antifungal susceptibility of Candida biofilms: unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrobial Agents and Chemotherapy 46: Lewis K (2001): Riddle of biofilm resistance. Antimicrob.Agents Chemother. 45: Lewis K (2005): Persister cells and the riddle of biofilm survival. Biochemistry (Mosc.). 70: Mah TF, O'toole GA (2001): Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 9: Maira-Litran T, Allison DG, Gilbert P (2000a): An evaluation of the potential of the multiple antibiotic resistance operon (mar) and the multidrug efflux pump acrab to moderate resistance towards ciprofloxacin in Escherichia coli biofilms. J.Antimicrob.Chemother. 45: Maira-Litran T, Allison DG, Gilbert P (2000b): Expression of the multiple antibiotic resistance operon (mar) during growth of Escherichia coli as a biofilm. J.Appl.Microbiol. 88:
108 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 66. Martinez LR, Bryan RA, Apostolidis C, Morgenstern A, Casadevall A, Dadachova E (2006): Antibody-guided alpha radiation effectively damages fungal biofilms. Antimicrob.Agents Chemother. 50: Martinez LR, Casadevall A (2006): Susceptibility of Cryptococcus neoformans biofilms to antifungal agents in vitro. Antimicrob.Agents Chemother. 50: Mathur T, Singhal S, Khan S, Upadhyay D, Fatma T, Rattan A (2005): Adverse effect of staphylococci slime on in vitro activity of glycopeptides. Jpn.J.Infect.Dis. 58: Monzon M, Oteiza C, Leiva J, Lamata M, Amorena B (2002): Biofilm testing of Staphylococcus epidermidis clinical isolates: low performance of vancomycin in relation to other antibiotics. Diagn.Microbiol.Infect.Dis. 44: Muli F, Struthers JK (1998): Use of a continuous-culture biofilm system to study the antimicrobial susceptibilities of Gardnerella vaginalis and Lactobacillus acidophilus. Antimicrob.Agents Chemother. 42: Nemoto K, Hirota K, Murakami K, Taniguti K, Murata H, Viducic D, Miyake Y (2003): Effect of Varidase (streptodornase) on biofilm formed by Pseudomonas aeruginosa. Chemotherapy. 49: Nemoto K, Hirota K, Ono T, Murakami K, Murakami K, Nagao D, Miyake Y (2000): Effect of Varidase (streptokinase) on biofilm formed by Staphylococcus aureus. Chemotherapy. 46: Neut D, Hendriks JG, van Horn JR, van der Mei HC, Busscher HJ (2005): Pseudomonas aeruginosa biofilm formation and slime excretion on antibiotic-loaded bone cement. Acta Orthop. 76: Patel R (2005): Biofilms and antimicrobial resistance. Clin.Orthop.Relat Res Perez-Giraldo C, Cruz-Villalon G, Sanchez-Silos R, Martinez-Rubio R, Blanco MT, Gomez-Garcia AC (2003): In vitro activity of allicin against Staphylococcus epidermidis and influence of subinhibitory concentrations on biofilm formation. J.Appl.Microbiol. 95: Pickering SA, Bayston R, Scammell BE (2003): Electromagnetic augmentation of antibiotic efficacy in infection of orthopaedic implants. J.Bone Joint Surg.Br. 85: Pitt WG, McBride MO, Lunceford JK, Roper RJ, Sagers RD (1994): Ultrasonic enhancement of antibiotic action on gram-negative bacteria. Antimicrob.Agents Chemother. 38: Pitt WG, Ross SA (2003): Ultrasound increases the rate of bacterial cell growth. Biotechnol.Prog. 19: Prigent-Combaret C, Lejeune P (1999): Monitoring gene expression in biofilms. Methods Enzymol. 310:56-79.: Rayner MG, Zhang Y, Gorry MC, Chen Y, Post JC, Ehrlich GD (1998): Evidence of bacterial metabolic activity in culture-negative otitis media with effusion. JAMA. 279: Rediske AM, Roeder BL, Brown MK, Nelson JL, Robison RL, Draper DO, Schaalje GB, Robison RA, Pitt WG (1999): Ultrasonic enhancement of antibiotic action on Escherichia coli biofilms: an in vivo model. Antimicrob.Agents Chemother. 43: Rediske AM, Roeder BL, Nelson JL, Robison RL, Schaalje GB, Robison RA, Pitt WG (2000): Pulsed ultrasound enhances the killing of Escherichia coli biofilms by aminoglycoside antibiotics in vivo. Antimicrob.Agents Chemother. 44: Roberts AP, Pratten J, Wilson M, Mullany P (1999): Transfer of a conjugative transposon, Tn5397 in a model oral biofilm. FEMS Microbiol.Lett. 177:
109 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 84. Schierholz JM, Beuth J (2001): Implant infections: a haven for opportunistic bacteria. Journal of Hospital Infection 49: Shigeta M, Tanaka G, Komatsuzawa H, Sugai M, Suginaka H, Usui T (1997): Permeation of antimicrobial agents through Pseudomonas aeruginosa biofilms: a simple method. Chemotherapy. 43: Shih PC, Huang CT (2002): Effects of quorum-sensing deficiency on Pseudomonas aeruginosa biofilm formation and antibiotic resistance. J.Antimicrob.Chemother. 49: Shuford JA, Steckelberg JM, Patel R (2005): Effects of fresh garlic extract on Candida albicans biofilms. Antimicrob.Agents Chemother. 49: Singh PK, Parsek MR, Greenberg EP, Welsh MJ (2002): A component of innate immunity prevents bacterial biofilm development. Nature. 417: Souli M, Giamarellou H (1998): Effects of slime produced by clinical isolates of coagulase-negative staphylococci on activities of various antimicrobial agents. Antimicrob.Agents Chemother. 42: Spoering AL, Lewis K (2001): Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J.Bacteriol. 183: Stanley NR, Britton RA, Grossman AD, Lazazzera BA (2003): Identification of catabolite repression as a physiological regulator of biofilm formation by Bacillus subtilis by use of DNA microarrays. J.Bacteriol. 185: Stewart PS (1996): Theoretical aspects of antibiotic diffusion into microbial biofilms. Antimicrobial Agents and Chemotherapy 40: Stewart PS, Roe F, Rayner J, Elkins JG, Lewandowski Z, Ochsner UA, Hassett DJ (2000): Effect of catalase on hydrogen peroxide penetration into Pseudomonas aeruginosa biofilms. Appl.Environ.Microbiol. 66: Stewart PS, Wattanakaroon W, Goodrum L, Fortun SM, McLeod BR (1999): Electrolytic generation of oxygen partially explains electrical enhancement of tobramycin efficacy against Pseudomonas aeruginosa biofilm. Antimicrob.Agents Chemother. 43: Suci PA, Mittelman MW, Yu FP, Geesey GG (1994): Investigation of ciprofloxacin penetration into Pseudomonas aeruginosa biofilms. Antimicrob.Agents Chemother. 38: Tresse O, Lescob S, Rho D (2003): Dynamics of living and dead bacterial cells within a mixed-species biofilm during toluene degradation in a biotrickling filter. J.Appl.Microbiol. 94: Vrany JD, Stewart PS, Suci PA (1997): Comparison of recalcitrance to ciprofloxacin and levofloxacin exhibited by Pseudomonas aeruginosa bofilms displaying rapidtransport characteristics. Antimicrob.Agents Chemother. 41: Wagman GH, Bailey JV, Weinstein MJ (1975): Binding of aminoglycoside antibiotics to filtration materials. Antimicrobial Agents and Chemotherapy 7: Wai SN, Mizunoe Y, Takade A, Kawabata SI, Yoshida SI (1998): Vibrio cholerae O1 strain TSI-4 produces the exopolysaccharide materials that determine colony morphology, stress resistance, and biofilm formation. Appl.Environ.Microbiol. 64: Wattanakaroon W, Stewart PS (2000): Electrical enhancement of Streptococcus gordonii biofilm killing by gentamicin. Arch.Oral Biol. 45: Whiteley M, Bangera MG, Bumgarner RE, Parsek MR, Teitzel GM, Lory S, Greenberg EP (2001): Gene expression in Pseudomonas aeruginosa biofilms. Nature. 413:
110 Mikrobiální biofilmy Biofilmy a antimikrobiální látky 102. Williams I, Venables WA, Lloyd D, Paul F, Critchley I (1997): The effects of adherence to silicone surfaces on antibiotic susceptibility in Staphylococcus aureus. Microbiology. 143: Wu JA, Kusuma C, Mond JJ, Kokai-Kun JF (2003): Lysostaphin disrupts Staphylococcus aureus and Staphylococcus epidermidis biofilms on artificial surfaces. Antimicrob.Agents Chemother. 47: Xu KD, McFeters GA, Stewart PS (2000): Biofilm resistance to antimicrobial agents. Microbiology. 146: Yildiz FH, Schoolnik GK (1999): Vibrio cholerae O1 El Tor: identification of a gene cluster required for the rugose colony type, exopolysaccharide production, chlorine resistance, and biofilm formation. Proc.Natl.Acad.Sci.U.S.A. 96: Zheng Z, Stewart PS (2002): Penetration of rifampin through Staphylococcus epidermidis biofilms. Antimicrob.Agents Chemother. 46:
111 7.3 Holá V., Kaprálová S.: Biofilm dutiny ústní; in M. Rulík, V. Holá, F. Růžička, M. Votava (eds.); Mikrobiální biofilmy; (in press; předpokládané datum vydání jaro 2007).
112 Mikrobiální biofilmy 1 4. Biofilm dutiny ústní 4. Biofilm dutiny ústní 4.1 Charakteristika orálního biofilmu dle lokalizace Rty Bukální povrchy a měkké patro Jazyk Supragingivální zubní plak Subgingivální zubní plak Koronární a fisurální zubní plak 4.2 Vznik a zrání zubního biofilmu 4.3 Druhové složení zubního biofilmu Složení rezidentní mikroflóry Vliv umístění na druhové složení zubního biofilmu Vliv živin na druhové složení zubního biofilmu 4.4 Mikrobiální interakce v zubním biofilmu Mezidruhové interakce Sukcese v zubním plaku Vznik zubního kamene 4.5 Mikrobiální rozdíly ve zdraví a v nemoci 4.6 Mikrobiální etiologie zubního kazu a parodontopatií Zubní kaz Vztah mezi zubním plakem a zubním kazem Parodontopatie Implantáty 4.7 Patogenní determinanty kariogenních bakterií 4.8 Fokální infekce 4.9 Detekce a likvidace plaku Detekce plaku Odstranění plaku 4.10 Antimikrobiální léčba Biofilm dutiny ústní Stomatologie je vůbec první oblastí humánní mikrobiologie, kde lékařští mikrobiologové přijali možnost, že zubní plak, neboli biofilm, je druhým přirozeným způsobem života bakterií a že by bakterie a bakteriální společenstva mohla žít přisedle a tvořit určité nánosy (plaky, biofilmy), a že tedy bakterie nemusí žít a existovat jen ve formě planktonických buněk. Zubní mikrobiální povlak plak je získaný měkký povlak pokrývající jak přirozený tak arteficielní povrch zubu a gingivy, který se skládá z organických a anorganických složek a různorodé komunity mikrobiálního biofilmu, jenž se nachází na zubním povrchu. Zubní plak rezidentní mikroflóry (tj. mikroflóry hostiteli vlastní, endogenní) se na zubech vytváří přirozeně a je důležitou součástí obranných mechanizmů hostitele, protože pomáhá bránit kolonizaci zubní skloviny exogenními mikroorganizmy (jde o tzv. colonisation resistance) (Marsh 2004). Na druhou stranu je však neošetřovaný zubní plak také hlavním etiologickým faktorem parodontopatií a rozhodujícím faktorem vzniku zubního kazu (Dřízhal & Slezák 1993; Kilian et al. 1996). Procesem kalcifikace přechází zubní plak v zubní kámen. 4.1 Dutina ústní jako mikrobiální biotop Zdaleka ne všechny mikroorganizmy, které se dostanou do dutiny ústní, ji jsou schopny kolonizovat. Dutina ústní je díky svým vlastnostem ekologicky odlišná od všech dalších povrchů těla, čímž se výběr mikrobů schopných kolonizace a perzistence omezuje. V rámci dutiny ústní existuje navíc i další stratifikace a rozrůzněnost prostředí, z nichž každé
113 Mikrobiální biofilmy 2 4. Biofilm dutiny ústní bude svými biologickými vlastnostmi podporovat růst určitých mikrobů a určité mikrobiální komunity. Mikrobiální biofilm se v dutině ústní tvoří na měkkých i tvrdých tkáních. Složení biofilmu se na těchto místech výrazně liší, a to jak mikrobiálním zastoupením, tak složením extracelulární substance. Zcela jasně nám pro mikrobiální kolonizaci budou poskytovat odlišná prostředí povrchy sliznic, např. rty, vnitřní strana tváří (bukální sliznice), měkké patro či jazyk a povrchy zubů s částmi subgingiválními a supragingiválními, zde pak s plochami koronárními a okluzálními (s fissurami). Vlastnosti některých z těchto prostředí se u jedince mění s věkem. Ekologické podmínky v dutině ústní se též mění při výměně prvního za trvalý chrup, při extrakci zubů, při následném používání dentálních protéz a pomůcek a samozřejmě i při každém zubním zákroku (Marsh & Martin 2001). Dočasné fluktuace ve stabilitě orálního ekosystému mohou být indukovány též frekvencí a typem požívané stravy, změnami v toku slin či případnou antibiotickou léčbou (Rudney 2000; Rudney et al. 2003) Rty Mikrobiální biomasa je na rtech omezovaná odlupováním svrchní vrstvy epitelií a částečně také odlišným prostředím rty jsou nejsušší částí dutiny ústní s nejlepším přísunem kyslíku. Proto zde převládají odolné mikroby, zejména grampozitivní koky, např. rodu Streptococcus. Z okolní pokožky se na rty může dostat i přirozená kožní mikroflóra, např. koagulázanegativní stafylokoky Bukální povrchy a měkké patro I na těchto površích je mikrobiální biomasa omezená odlupováním svrchní vrstvy epitelií. Přítomnost keratinizovaných a nekeratinizovaných částí pak ovlivňuje prostorové uspořádání mikrobiální flóry a případnou přítomnost či absenci konkrétních bakteriálních druhů. Prostředí bukálních sliznic i měkkého patra je dobře zásobeno živinami a je dostatečně aerované. Proto se na těchto sliznicích se vyskytují aerobní a fakultativně anaerobní zástupci běžné mikroflóry dutiny ústní. Jedná se zejména o zástupce rodů Streptococcus (viridující streptokoky), Neisseria, Haemophillus, Lactobacillus aj. Biofilmová vrstva vytvořená na těchto površích je slabší než zubní plak a je výrazně ovlivňována samočistícími schopnostmi dutiny ústní Jazyk Jazyk má vysoce zbrázděný a papilární povrch s nízkým redoxním potenciálem. Z toho důvodu slouží jako rezervoár anaerobních mikrobů. To hraje významnou roli též v patogenezi onemocnění parodontu, kde tyto anaerobní mikroby hrají významnou roli. Jedná se zejména o rody Veillonella, Fusobacterium, Bacteroides aj Supragingivální zubní plak Semipermeabilní pelikula, tvořená především různými proteiny ze slin, která se naváže na vápenaté a fosfátové ionty apatitu, obsaženého v tvrdých zubních tkáních, zvlhčuje zub a chrání ho při žvýkání před abrazí. Během 24 hodin nasedají převážně grampozitivní streptokoky a aktinomycety, v následujících dnech nasedají gramnegativní koky, ale i grampozitivní a gramnegativní tyčky a vláknité mikroorganizmy. V biofilmu 7 14 dní starém převažují tyčky a filamenta. Plak začne zvětšovat svůj objem a dochází k pomnožení gramnegativních anaerobů. Za 3 týdny převažují vláknité gramnegativní mikroorganizmy

114 Mikrobiální biofilmy 3 4. Biofilm dutiny ústní obklopené grampozitivními koky a vznikají typické útvary, např. tzv. kukuřičný klas (obr. 4.2). Uvnitř plaku přežívají anaerobní a fakultativně anaerobní mikroorganizmy a nahrazují postupně mikroorganizmy aerobní. Vyzrálý plak se skládá z hustě naskládaných buněk (60 70 % objemu), které jsou umístěny v matrix biofilmu Rosan & Lamont Subgingivální zubní plak Díky své lokalizaci má subgingivální plak, tedy plak nacházející se pod okrajem dásní, v dásňovém žlábku, v pravém či nepravém parodontálním chobotu, odlišnou topografii chybí zde antimikrobiální substance přítomné např. ve slinách. Na povrchu zubního kořene se nachází různě silná vrstvička pevně lpícího strukturovaného plaku, který je složením podobný supragingiválnímu plaku, i když zde mohou dosahovat vyšších četností anaerobní mikroorganizmy. Tento plak, který může kalcifikací přejít i v subgingivální zubní kámen, označujeme jako adherující plak. Subgingivální zubní kámen usnadňuje adhezi dalších druhů mikroorganizmů a stává se tak dalším retenčním místem pro jejich usazování. K měkkým tkáním chobotu přiléhá vrstva plaku tvořená téměř výhradně anaerobními mikroorganizmy (zejména gramnegativními tyčkami a spirochetami). V období progrese, tedy akutního zánětu, tato vrstva mohutní a vzniká tzv. plak neadherující či plovoucí. Složení subgingiválního plaku je u zdravého parodontu, při gingivitidě a při parodontitidách různé. Obr. 4.1 Rozložení subgingiválního plaku Koronární a fisurální zubní plak Plak na hladkém povrchu zubní korunky se vyskytuje se zejména v mezizubních prostorech a v gingivální třetině korunky. Vždy zde můžeme nalézt zástupce rodu Streptococcus, Veillonella a Actinomyces, řidčeji též zástupce rodu Lactobacillus. Fisurální zubní plak, neboli plak v rýhách a jamkách zubů, pokrývá sklovinu v místech, kde nejčastěji začíná zubní kaz. V intaktních fisurách převažují grampozitivní koky a tyčky nad gramnegativními koky a tyčkami. Laktobacilů a kvasinek je zde velmi málo a fusiformní i filamentózní organizmy prakticky chybí (Kilian et al. 1996; Hellwig et al. 2003). 4.2 Vznik a zrání orálního biofilmu Základní mechanizmy, uplatňující se při tvorbě zubního biofilmu, jsou stejné jako mechanizmy tvorby biofilmů jednodruhových, které se uplatňují v jiných, převážně sterilních částech lidského těla. Stejně jako tam, i zde dochází nejprve k úpravě povrchu různými proteiny (albumin, imunoglobuliny, různé fosfoproteiny, glykoproteiny apod.) a vzniku pelikuly. Je známo, že vazbami mezi jednotlivými sloučeninami nacházejícími se v této povrchové vrstvě může dojít ke změně konformace těchto sloučenin, což může výsledně ovlivnit vazebná místa pro mikroorganizmy. Tímto způsobem se mění např. molekulární struktura glukanů, naváží-li se na povrch zubu zároveň i glukosyltransferázy (Vacca-Smith et al. 1996; Kopec
115 Mikrobiální biofilmy 4 4. Biofilm dutiny ústní et al. 2001). Složení povrchové vrstvy pak výsledně velmi ovlivňuje druhovou strukturu primárních kolonizátorů (Marsh 2004). Pelikula svou tloušťkou zřídka přesáhne 1 µm. Na tuto vrstvu nasedají primární kolonizátoři, kteří si pomáhají nespecifickými interakcemi povrchu mikrobiální buňky právě s touto vrstvou (Van der Waalsovy síly apod.) (Busscher & van der Mei 1997). Primární kolonizaci zubního povrchu dále napomáhají síly krátkého dosahu, zejména elektrostatické síly, a také bakteriální adheziny (Lamont & Jenkinson 2000). Primární kolonizace se účastní především fakultativně anaerobní bakterie, zejména streptokoky, např. Streptococcus mutans, S. sanguis a S. mitis. Přidávají se další mikrobiální druhy, které buď kolonizují přímo dosud neobsazená místa zubního povrchu, nebo dochází k jejich koadhezi a koagregaci s již přisedlými kolonizátory primárními. S rostoucí tloušťkou zubního plaku vzniká v hlubších vrstvách anaerobní prostředí, což umožňuje kolonizaci převážně anaerobním bakteriím. Účastní se zde rody Veillonella, Neiserria, Actinomyces, Corynebacterium, Bacteroides apod. V pozdější fázi se též přidávají filamenty. Bakterie ústní mikroflóry mají na svém buněčném povrchu obvykle více, než jen jeden typ adhezinů a mohou se tedy podílet na mnohočetných interakcích, a to jak s hostitelskými molekulami, tak s obdobnými receptory na povrchu jiných bakteriálních buněk (Marsh 2004), čímž vzniká různorodé společenstvo vícedruhového biofilmu. V tomto stádiu se též výrazně projevují specifické interakce mezi bakteriálními adheziny a receptory, což vede ke zvýšení druhové diverzity biofilmu a zejména ke vzniku morfologických struktur v jiných biofilmech neobvyklých, jako jsou tzv. kukuřičné klasy a rozety (Kolenbrander et al. 2000) (viz obr. 4.2). Obr. 4.2 Struktura kukuřičných klasů typická pro biofilm dutiny ústní (převzato z Kolebrander et al. 2000) Komplex zubního plaku se obohacuje o spirochety a fusiformní bakterie. Při dalším zrání vícedruhového zubního biofilmu dochází díky produkci glukanů, levanů a dalších polymerů k pevnější adhezi mikrobů k zubní sklovině a později též ke kalcifikaci biofilmové struktury a tím ke vzniku zubního kamene (Dřízhal & Slezák 1993; Kilian et al. 1996; Mutschelknauss et al. 2002).
116 Mikrobiální biofilmy 5 4. Biofilm dutiny ústní Méně než v jiných biofilmech se u bakterií biofilmu dutiny ústní projevují po adhezi na povrch zubů změny v expresi či represi různých genů. Je to dáno tím, že mikroflóra dutiny ústní je, na rozdíl od volně žijících planktonických buněk jiných mikrobů, na schopnosti tvorby biofilmu a na životě v biofilmové vrstvě existenčně závislá (Burne et al. 1999; Svensater et al. 2001). Přesto se některé geny ve větší míře v biofilmové vrstvě uplatňují, jde například o geny spojené se syntézou glukanů a fruktanů, jak bylo prokázáno u Streptococcus mutans (Li et al. 2001). Pro mikroflóru dutiny ústní je naopak výhodnější vzájemně se shlukovat. Toto shlukování se u planktonických bakterií nazývá koagregace, pokud je jedna z interagujících bakterií přisedlá, používá se pojem koadheze. Koagregace a koadheze jsou u mikrofóry dutiny ústní častější než kdekoliv jinde a koadheze je významným faktorem usnadňujícím kolonizaci zubních povrchů. Například Streptococcus sanguinis usnadňuje adhezi Actinomyces naeslundii, která sama o sobě schopnost adheze vykazuje velmi nízkou (Ganeshkumar et al. 1991). Zubní plak je tvořen z 80 % z vody a 20 % ze sušiny, z toho asi 1/3 tvoří mikrobiální buňky a 2/3 extracelulární substance. Extracelulární substance je tvořena zejména organickými látkami, polysacharidy a proteiny, ale i anorganickými látkami, zejména sloučeninami vápníku a fosfáty. Tato extracelulární substance představuje nárazníkový systém plaku. Zubní plak je nutno odlišit od pigmentace zubů a přítomnosti neorganizovaných hmot. Zabarvení a pigmentace zubů jsou způsobeny např. produkty dehtových zplodin při kouření či přítomností barviv v potravě, např. v kávě, čaji, červeném víně a medikamentech. Neorganizované hmoty, tzv. materia alba, jsou nánosy ze zbytků potravy, odloučených epitelií a bakterií. Materia alba je šedobílé až žlutavé barvy, usazuje se na zubech či plaku a vyskytuje se na špatně čistitelných místech. Tato hmota se dá odstranit pouhým energickým výplachem (Kilian et al.1996). 4.3 Druhové složení zubního biofilmu Složení rezidentní mikroflóry Rezidentní mikroflóra zubního plaku je velmi různorodá a liší se též podle jeho stáří. Udává se, že tvorby biofilmu v ústní dutině se může účastnit více než 500 mikrobiálních druhů, z nichž je jen cca 60 % kultivovatelných. Částečné znalosti o nekultivovatelných bakteriálních druzích jsme získali na základě molekulárně-biologických přístupů (Paster et al. 2000; Suzuki et al a další). Druhů, které se tvorby zubního biofilmu účastní nejčastěji, je cca 200 (nejčastěji zastoupené rody jsou uvedeny v tab. 4.2). Prvními bakteriemi, které kolonizují zubní povrch, jsou viridující streptokoky a grampozitivní tyčky, např. aktinomycety. Z aktinomycet je nejčastějším zástupcem Actinomyces naeslundii. Mezi streptokoky převažují během prvních čtyř hodin vzniku zubního plaku druhy Streptococcus mitis, S. sanguinis a S. oralis, tvoří % kultivovatelných streptokoků (Nyvad et Kilian 1990). Za pozdější kolonizátory jsou v zubním plaku považovány bakteriální rody Fusobacterium, Porphyromonas, Prevotella, Veillonella, Capnocytophaga a Actinobacillus (Lu et al. 2002). Přítomnost širokého spektra mikrobů a jejich produktů ovlivňuje podmínky ve vytvořeném biofilmu a tím i v ústní dutině. Důležitou složkou mikroflóry dutiny ústní, a tedy i zubního biofilmu, jsou streptokoky skupiny mutans. Do této skupiny se řadí v současné době tři sérotypy S. mutans, dva sérotypy S. sobrinus, S. criceti, S. ratti a S. downei (Banas & Vickerman 2003).
117 Mikrobiální biofilmy 6 4. Biofilm dutiny ústní Tab. 4.2 Mikrobiální rody vyskytující se nejčastěji v zubním plaku Fakultativně anaerobní Striktně anaerobní Fakultativně anaerobní Grampozitivní Gramnegativní Striktně anaerobní Koky Streptococcus Peptostreptococcus Neisseria Veillonella Micrococcus Peptococcus Moraxella Bacteroides Tyčky Actinomyces Actinomyces Actinobacillus Fusobacterium Bacterionema Arachnia Capnocytophaga Wolinella Rothia Eubacterium Eikenella Leptotrichia Nocardia Propionibacterium Haemophilus Selenomonas Lactobacillus Bifidobacterium Campylobacter Corynebacterium Prevotella Porphyromonas Spirochety a další mikroorganismy Treponema Candida Entamoeba gingivalis Mycoplasma Saccharomyces Trichomonas tenax Prostředí biofilmové vrstvy je výrazně vertikálně stratifikováno, do hlubších vrstev proniká méně živin, je zde nižší ph, nižší parciální tlak kyslíku apod. Heterogenitě prostředí rovněž napomáhá přítomnost mnoha produktů jak bakteriálního, tak hostitelského původu. Takto výrazně heterogenní prostředí biofilmové vrstvy zřetelně napomáhá koexistenci druhů s tak odlišnými nároky na okolní prostředí, že by v homogenním prostředí spolu koexistovat nemohly. Proto dosahují absolutní počty bakteriálních druhů zastoupených v zubním biofilmu tak vysokých hodnot Vliv umístění na druhové složení zubního biofilmu Zubní plak se nachází zejména na chráněných površích s minimálním průtokem slin či sulkulární tekutiny. Jedná se zejména o aproximální plošky zubů, které jsou z toho důvodu nejvíce ohroženy mikrobiální kolonizací. Environmentální podmínky na povrchu zubů nejsou samozřejmě uniformní, naopak v rámci ústní dutiny nacházíme jak rozdíly ve stupni účinku čistících sil ústní dutiny (nejnižší jsou přirozeně opět na aproximálních ploškách zubů, tj. v mezizubních prostorách), tak v gradientu mnoha biologických a chemických faktorů, které ovlivňují rezidentní mikroflóru (dostatek živin, parciální tlak kyslíku, redoxní potenciál prostředí atd.). Tyto rozdíly se projevují ve změnách složení mikrobiálního společenstva, a to zejména na místech tak výrazně odlišných, jako jsou gingivální sulky (dásňové žlábky), aproximální plošky zubů, hladké povrchy či fisury (rýhy) okluzní části zubů (viz obrázek 4.3). Na okluzních ploškách zubů je fisurální plak například více ovlivněn slinami, než plak na jiných ploškách zubů, na plak v dásňových sulcích má zase větší vliv sulkulární tekutina. V gingiválních sulcích je také nižší redoxní potenciál a ty jsou díky tomu kolonizovány vyšším množstvím anaerobních bakterií, zejména proteolytických druhů, které získávají klíčové živiny a růstové faktory zejména z katabolizmu glykoproteinů přítomných v sulkulární tekutině.
118 Mikrobiální biofilmy 7 4. Biofilm dutiny ústní Studie plaku, vyvíjejícího se v hlubokých parodontálních chobotech ukázaly, že nejhlubší zóny jsou kolonizovány zejména spirochétami a gramnegativními bakteriemi, zatímco mělčí vrstvy těchto chobotů jsou osídleny zejména grampozitivními koky (Wecke et al. 2000). Obr. 4.3 Lokalizace zubního biofilmu na zubech Vliv živin na druhové složení zubního biofilmu Převládá názor, že mikrobiální diverzita zubního biofilmu je způsobena spíše endogenně poskytovanými živinami než rozdíly v přijímané stravě. Ze slin bakterie pro svůj růst získávají aminokyseliny, peptidy, proteiny a glykoproteiny (muciny). Sulkulární tekutina je séru podobná tekutina, která omývá dásňové krčky a jejíž průtok je výrazně zvýšen v průběhu zánětlivého procesu v ústní dutině. Kromě složek hostitelova imunitního systému (IgG, IgA, IgM, neutrofilů, makrofágů, komplementu či B a T lymfocytů) poskytuje též peptidy, proteiny a glykoproteiny, transferriny, haptoglobin apod., kteréžto makromolekuly mohou být nutričně využity rezidentní mikroflórou dásňových sulků. Navíc k těmto endogenním zdrojům živin bakterie využívají i živiny nacházející se ve stravě. Jediná složka stravy, která výrazně ovlivňuje složení biofilmu, jsou fermentovatelné cukry. Ty mohou být buď katabolizovány na kyseliny, nebo přeměněny na řadu extracelulárních či intracelulárních polysacharidů. Kyseliny, jako např. kyselina mléčná, dočasně sníží ph v zubním plaku, což vede k demineralizaci zubní skloviny a při častějším opakování tohoto jevu až ke vzniku zubního kazu. Opakované snižování ph též narušuje rovnováhu mikrobiální flóry zdravé dutiny ústní, protože potlačí růst druhů přirozené mikroflóry, které jsou ke sníženému ph citlivé. Sacharóza může být též metabolizována na řadu rozpustných i nerozpustných fruktanů a glukanů (Walker et al. 1987). Rozpustné fruktany slouží jako zdroj živin dalších bakterií v biofilmové vrstvě, nerozpustné glukany upevňují adhezi některých druhů mikrobů k zubnímu povrchu. Další složky stravy, které mohou ovlivnit ekologii zubního plaku, i když v mnohem menší míře než fermentovatelné cukry, jsou mléčné produkty a náhražky sacharózy, tedy tzv. umělá sladidla. Mléčné proteiny a polypeptidy se mohou adsorbovat na zubní sklovinu a snižovat tak bakteriální adhezi (Olsson et al. 1992; Reynolds et al. 1983). Mléčné výrobky

119 Mikrobiální biofilmy 8 4. Biofilm dutiny ústní mohou také, díky uvolňování fosforečnanu vápenatého, podporovat remineralizaci zubní skloviny. Některé z používaných náhražek sacharózy působí antimikrobiálně (Grenby et al. 1986; Mäkinen 1989), například dlouhodobé používání xylitolu může snížit percentuální zastoupení streptokoků skupiny mutans v zubním plaku (Mäkinen 1989). 4.4 Mikrobiální interakce v zubním biofilmu Mezidruhové interakce V biofilmu, jakým je zubní plak, jsou mikroorganizmy ve vzájemné těsné blízkosti a v důsledku toho mezi nimi dochází k interakcím. Tyto interakce mohou být pro jednu či více zúčastněných populací výhodné, nebo mohou být naopak antagonistické. Mikrobiální metabolizmus uvnitř plaku produkuje řadu faktorů, ovlivňujících růst jiných druhů, včetně spotřebování základních živin se souběžnou produkcí a akumulací toxických či inhibujících vedlejších produktů. Tyto gradienty vedou k tomu, že se uvnitř zubního plaku vyvine vertikální a horizontální stratifikace. Taková heterogenita prostředí umožňuje mikroorganizmům s široce se lišícími požadavky na růst a zajišťuje koexistenci druhů, které by byly inkompatibilní v homogenním prostředí. Ve společenstvu zubního biofilmu dochází k mnoha antagonickým a synergickým biochemickým interakcím (Marsh 1999), což může zvýšit efektivitu metabolických řetězců, zvláště pokud se bakterie nacházejí v těsném kontaktu. Přežívání obligátních anaerobů v aerobním prostředí ústní dutiny může být například usnadněno těsnou koadhezí s druhy bakterií využívajícími kyslík (Bradshaw et al. 1998). Pozitivní interakce zahrnují např. společné působení dvou či více mikrobiálních druhů vedoucí k rozkladu hostitelských makromolekul, které jednotlivé mikrobiální druhy nejsou schopné katabolizovat (např. mucin), dále vznik potravních řetězců, kdy vedlejší či odpadní produkty jednoho mikrobiálního druhu jsou Obr. 4.4 Časově-prostorový model bakteriální kolonizace dutiny ústní ilustruje rozpoznání receptorů, přítomných substrátem pro druh další (např. v pelikule časnými kolonizátory a koagregaci mezi časnými rozklad sacharózy různými druhy kolonizátory, fusobakteriemi a pozdními kolonizátory. bakterií na laktát a následná Všechny znázorněné koagregace byly prokázány in vitro. utilizace laktátu druhy rodu Veillonella) (Kolebrander et al. 2003). Dalším projevem pozitivních interakcí je např. koagregace, tedy shlukování bakterií,
120 Mikrobiální biofilmy 9 4. Biofilm dutiny ústní které usnadňuje pozdním kolonizátorům přisednutí k zubnímu povrchu a začlenění do mikrobiálního společenstva biofilmu (Rosan & Lamont 2000). Antagonistické interakce zahrnují produkci inhibujících složek, např. bakteriocinů, peroxidu vodíku či organických kyselin Sukcese v zubním plaku Časní kolonizátoři zubních povrchů jsou zejména neisserie a viridující streptokoky. Růst a metabolizmus těchto pionýrských druhů mění lokální environmentální podmínky (např. redoxní potenciál, ph, dostupnost živin, možnost koagregace s dalšími mikroby) a tím umožňuje kolonizaci i choulostivějším a náročnějším mikrobům. Například obligátní anaerobi bývají v zubním plaku pozdními kolonizátory, protože jsou schopny růstu jen za nízkého redoxního potenciálu a za nepřítomnosti kyslíku. Heterogenní a koloniální typ struktur byly pozorovány v plaku z hladkých zubních povrchů. Heterogenní typ je spojován s palisádovými regiony, kde tyčky a koky se objevují upravené do paralelních, podélných a pravoúhelných tvarů vůči povrchu zubní skloviny. Byly pozorovány také mikrokolonie pravděpodobně jednotlivých populací. Navíc je popisována též horizontální rozvrstvenost. Raná stádia vývoje zubního biofilmu vedou ke zhuštěné vrstvě limitovaného počtu bakteriálních druhů. Po 7 14 dnech se vytvoří větší vrstvy biofilmu, který vykazuje sice méně orientovaný růst, ale vyšší morfologickou diverzitu. Tato vrstevnatost je přičítána přímo bakteriální sukcesi (viz níže). Ve vyzrálém zubním plaku jsou pak bakterie v přímém kontaktu se zubní sklovinou protože enzymaticky rozložily původní bílkovinný povlak, jež se na zubu vytvořil Vznik zubního kamene Zubní kámen je složen z minerálních, ale i organických složek. Množství minerálních složek se pohybuje od 37 % u zubního kamene supragingiválního do 58 % u kamene subgingiválního. Dále jsou v zubním kameni přítomny proteiny, v malé míře i lipidy a další látky (Jin & Yip 2002). Zubní kámen je vždy pokryt slabou a rozvolněnou vrstvou mikroorganizmů. Na supragingiválním zubním kameni je tato vrstva tvořena převážně z vláknitých mikroorganizmů. Pod vrstvou mikroorganizmů jsou kalcifikující oblasti, které mají vrstevnatý vzhled. Supragingivální zubní kámen je heterogenní díky přítomnosti nekalcifikovaných vrstev. Tyto nekalcifikované vrstvy jsou obvykle nepravidelné a dodávají supragingiválnímu kameni vzhled vrstevnatosti. Vnitřní nepravidelně mineralizovaná vrstva se skládá z jehlicovitých a sloupcovitých krystalů (Mutschelknauss 2002). K mineralizaci zubního plaku dochází v řadě jednotlivých ložisek (Friskopp 1983), ze kterých se zubní kámen mineralizací šíří postupně dále. Charakteristický vzhled zubního kamene je uveden v barevné příloze na obr Je známo, že anorganická část zubního kamene se skládá zejména z hydroxyapatitu [Ca 10 (PO 4 ) 6 (OH) 2 ], ale i dalších vápenatých fosforečnanů [Ca 8 (PO 4 ) 4 (HPO 4 ) 2 5HO], [Ca 10 (HPO 4 )(PO 4 ) 6 ]. Nejdůležitějšími ionty pro tvorbu zubního kamene jsou Ca 2+ a PO 4 3-, oba se vyskytují ve větší či menší míře ve slinách. Přesycení slin těmito ionty může vést tedy k mineralizaci. Mineralizace závisí na součinu rozpustnosti a stupni nasycení slin danými solemi. Ačkoliv tvorba zubního kamene byla prokázána i u gnotobiotických zvířat, mineralizaci zubního plaku podporuje i metabolizmus mikrobů. Některé mikrobiální druhy jsou naopak spojovány s absencí zubního kamene. Takovým druhem je například
121 Mikrobiální biofilmy Biofilm dutiny ústní Actinobacillus actinomycetemcomitans, který je proto považován za antagonistu bakterií vytvářejících plak a působících kalcifikace (Listgarten 2000). Největší množství supragingiválního zubního kamene je přítomno na jazykové straně dolních předních zubů a jeho množství klesá směrem ke třetím molárům. Na horní čelisti se supragingivální zubní kámen tvoří nejčastěji na bukální straně prvních molárů. U subgingiválního zubního kamene je závislost na místě jeho tvorby méně výrazná (Corbett et al. 1998). 4.5 Mikrobiální rozdíly ve zdraví a v nemoci Plak se na zubech vytváří zcela přirozeně a přináší hostiteli výhodu tím, že poskytuje kolonizační rezistenci (patogenní mikrobi se nemají kde uchytit). Již jednou vyvinutá mikrobiální flóra v zubním plaku zůstává relativně stabilní navzdory mnoha negativním vnějším vlivům. Tato stabilita, nebo také mikrobiální homeostáza, není způsobena metabolickou netečností rezidentní mikroflóry, ale je výsledkem dynamické rovnováhy ustavené mezi jednotlivými členy tohoto společenstva (Sbordone & Bortolaia 2003). Příležitostně se mikrobiální homeostáza poruší a vznikne mikrobiální nerovnováha, která je predisponujícím faktorem daného místa k onemocnění. Například opakovaný příjem fermentovatelných cukrů v potravě vedek častému snížování ph v zubním plaku, což inhibuje růst mnoha druhů spojených se zdravou ústní mikroflórou a zdravými zuby a selektuje vysoce acidogenní a acidotolerantní druhy, jako např. streptokoky skupiny mutans a laktobacily, kteréžto obě skupiny jsou spojovány se vznikem zubního kazu. V onemocnění parodontu je posun ve složení mikroflóry zubního plaku směrem k více proteolytickým gramnegativním anaerobním společenstvím, které mohou vyvolat poškození tkání, a to buď nepřímo cestou vedlejších efektů zánětlivé odpovědi hostitele, nebo přímo produkcí proteáz, cytotoxinů a dalších faktorů virulence. Složení mikroflóry se mění v závislosti na věku jedince. Při narození je ústní dutina prakticky sterilní, ke kontaminaci dochází postupně, nejčastěji přenosem od osob o dítě pečujících, zejména od matky. Ústní dutina nabízí zpočátku ke kolonizaci pouze epitel, proto flóra v ranném dětství není příliš pestrá. Jako první kolonizátor se objevuje Streptococcus salivarius. Laktobacily bývají kultivovány stabilně až po 2. roce života. Počátek prořezávání zubů znamená v osídlování dutiny ústní radikální změnu a do popředí se dostávají právě mikroorganizmy schopné adherovat na zubní sklovinu. V období puberty přibývá kmenů potenciálně patogenních pro parodont, což zřejmě souvisí s produkcí hormonů. Obraz mikrobiální flory podléhá i dalším vlivům, jako jsou např. stravovací návyky, ústní hygiena apod. (Kilian et al. 1996). U jedinců trpících onemocněním diabetes mellitus se můžeme se změnami mikrobiálního složení setkat jak v případě gingivitid, kdy se do popředí dostávají zejména rody Actinomyces a Streptococcus, ale i Veillonella parvula a Fusobacterium nucleatum, tak v případě parodontitid, kdy převažují zejména rod Capnocytophaga, Actinobacillus actinomycetemcomitans, anaerobní tyčky a spirochety. Při těhotenské gingivitidě není množství mikrobiálního biofilmu zvýšeno, ale pozorujeme změny v jeho složení, kdy nacházíme zejména P. intermedia, P. melaninogenica a Bacteroides sp. (Hellwig et al. 2003). 4.6 Mikrobiální etiologie zubního kazu a parodontopatií Zubní kaz Zubní kaz je lokalizovaná destrukce zubních tkání kyselinou produkovanou bakteriálním rozkladem fermentovatelných cukrů. Jednou z hlavních obtíží v určení
122 Mikrobiální biofilmy Biofilm dutiny ústní bakteriální etiologie zubního kazu je fakt, že zubní kaz se vyvíjí na místech s existující a rozrůzněnou rezidentní mikroflórou, takže je obtížné dojít k závěru, který bakteriální druh je etiologickým agens. Důkazy, že se mikroorganizmy podílejí na etiologii zubních kazů máme ze studií na zvířatech i z humánních klinických studií. Studie na gnotobiotických zvířatech ukázaly, že kaz může být způsobován určitými bakteriemi, zejména streptokoky skupiny mutans (např. Streptococcus mutans a S. sobrinus), ale pouze když jsou tato zvířata krmena kariogenní stravou, tj. stravou s vysokým obsahem sacharózy. Tyto studie též ukázaly možnost přenosu těchto streptokoků mezi jednotlivými zvířaty a také to, že pomocí antimikrobiálních látek a vakcinace můžeme dosáhnout ochrany. Modely s primáty potvrdily roli streptokoků skupiny mutans, možnost vakcinace a také pasivní imunizace, kdy mohou být protilátky aplikovány přímo v dutině ústní. Průřezové a průběžné studie řady skupin pacientů ukázaly silný vztah mezi přítomností streptokoků skupiny mutans a počátečními stádii zubního kazu. Tato korelace je nejsilnější pro agresivní fisurový kaz, u kazu aproximálních povrchů je tento vztah slabší, možná také z toho důvodu, že tyto povrchy jsou hůře přístupné jak pro odebírání vzorků zubního plaku, tak pro diagnostiku počátečních poškození zubní skloviny. Vznik zubního kazu lze na druhou stranu diagnostikovat na mnoha místech bez zjevné přítomnosti streptokoků skupiny mutans a tyto bakterie mohou být přítomny ve vysokých počtech na místech bez jakékoliv známky zubního kazu a demineralizace, což naznačuje, že etiologie zubního kazu není dosud zcela dořešena (Kolenbrander et al. 1999; Kolenbrander et al. 2000). Mikrobiální flóra se mění dlouhou dobu před vznikem viditelného poškození zubní skloviny. Nejprve v místě kazivé léze ubývají zástupci rodu Veillonella a S. sanguis a přibývají kvasinky rodu Candida, aktinomycety a zejména laktobacily a Streptococcus mutans. S. mutans se výrazně množí již asi 4 8 týdnů před vznikem viditelného poškození zubní skloviny, kdežto laktobacily převažují až asi 6 týdnů po jeho vzniku. Množství S. mutans po celou tuto dobu strmě narůstá. U počínajícího kazu cementu jsou jako zastoupeny aktinomycety, S. sanguis, S. mutans, S. mitis a dále zástupci rodů Veillonella, Cytophaga a Capnocytophaga. Při průniku kazu do dentinu tvoří více než 90 % mikroflóry grampozitivní mikroorganizmy, vysoký je podíl laktobacilů, podíl S. mutans naopak klesá. Přítomná je choulostivá mikroflóra, z grampozitivních zástupců např. Actinomyces naeslundii, A. odontolyticus, Propionibacterium sp. a Eubacterium sp., z gramnegativních zástupců pak Fusobacterium sp., Capnocytophaga sp. a Veillonella sp. Mikroby ponechané v dentinu ošetřených zubů mohou přežívat i více než dva roky. Kaz kořenových povrchů byl původně spojován s přítomností zástupců rodu Actinomyces, ale nedávné studie naznačují etiologii kazu podobnou kazu sklovinovému, tedy streptokoky skupiny mutans a laktobacily s možnou další rolí A. naeslundii. Infikovaný kořenový kanálek se může vyskytovat v zubech s otevřenou i uzavřenou dřeňovou dutinou. V případě otevřeného kořenového kanálku je osídlení jeho povrchu podobné mikroflóře dutiny ústní. Uzavřený kořenový kanálek je osídlen převážně anaerobními mikroorganizmy. V období akutizace zánětu převažují proteolytické mikroorganizmy. Při chronických stavech se pomnožují spíše glykolytické druhy bakterií. Při ústupu infekce opět vzrůstá počet laktobacilů a streptokoků (Ležovič et al. 1991). V etiologii apikálních granulomů, tedy granulomů nasedajících na kořenový systém zubu, hrají důležitou roli zejména Enterococcus faecalis, Veillonella sp., Fusobacterium sp., Prevotella sp. a Porphyromonas sp. (Hellwig et al. 2003).
123 Mikrobiální biofilmy Biofilm dutiny ústní Vztah mezi zubním plakem a zubním kazem Je několik různých pohledů, kterými se můžeme dívat na vztah mezi zubním plakem a zubním kazem. Od toho se budou samozřejmě odvíjet i naše kroky při studiu možností léčby zubního kazu. Z mnoha bakterií, jež se vyskytují v zubním plaku, vyvolává kaz jen pouze velmi malá skupina druhů. V tomto případě by bylo možné využít specifických a cílených protilátek. Plak je mikrobiální komunita, onemocnění zubním kazem vyplývá z výsledku interakcí mezi všemi zainteresovanými druhy a tedy prevencí by mohla být kontrola tvorby zubního plaku. Nemoc vyplývá ze změn rovnovážného stavu s přítomností rezidentní mikroflóry. Potenciálně kariogenní bakterie mohou být přítomny i u zdravých jedinců na zdravých zubech, ale v množstvích, která nejsou klinicky významná. Onemocnění by mohlo být kontrolováno nejen zacílením na domnělé patogeny, ale také ovlivněním faktorů zodpovědných za posun v mikroflóře negativním směrem Parodontopatie Bakteriální plak je rovněž příčinou parodontopatií, tedy onemocnění dásně a závěsného aparátu zubu. Destrukce parodontální tkáně může být výsledkem reakce hostitelského organizmu na mikrobiální faktory. V patogenitě plaku ve vztahu k parodontu hraje roli jeho objem, složení a metabolizmus. Důležitou úlohu při vzniku parodontopatií hraje individuální stav imunitního systému (Kilian et al.1996). Ve zdravém dásňovém sulku převažují z 85 % grampozitivní mikroorganizmy, zejména S. sanguis a S. mitis a aktinomycety - A. naeslundi a A. viscosus. S přibývajícím zánětem dásní dochází k posunu směrem ke gramnegativním tyčkám a spirochetám. U prostého zánětu dásní gingivitidy nacházíme v sulku třikrát více gramnegativních bakterií oproti bakteriím grampozitivním. Zvyšuje se podíl A. viscosus, dále najdeme Fusobacterium nucleatum, Prevotella melaninogenica, P. intermedia, rod Haemophilus a jiné gramnegativní mikroorganizmy. Jedná-li se o gingivitidu podmíněnou hormonálně (např. puberta, gravidita, hormonální léčba), dochází ke zmnožení anaerobních bakterií hojně se nachází např. P. intermedia. Subgingivální aktivní choboty vykazují složení mikroflóry podobné jako rychle progredující formy parodontitidy, tedy dominují tedy zejména P. gingivalis, B. forsythus a A. actinomycetemcomitans. V neaktivních chobotech je biofilm tvořen smíšenou flórou, která obsahuje zvýšené množství anaerobů a spirochet. Změny složení mikroflóry plaku u rychle progredující parodontitidy jsou podobné jako u aktivních chobotů. Podíl spirochet narůstá až přes 45 %. Dále bývají často identifikovány Porphyromonas gingivalis, Prevotella intermedia, Bacteroides forsythus, Wolinella recta, rod Capnocytophaga a Fusobacterium nucleatum. U mladších pacientů bývá často zastoupenm též Actinobacillus actinomycetemcomitans. Vina za těžké destrukce parodontu bývá přisuzována především P. gingivalis. Refrakterní parodontitida odolná, rezistentní na terapii je charakterizována výskytem komplexního spektra bakterií, především gramnegativních anaerobních tyček a spirochet. Z grampozitivních bakterií bývá prokázán např. Streptococcus intermedius (Hellwig et al. 2003; Mutschelknauss 2002). U akutní nekrotizující ulcerózní gingivitidy (ANUG, Plaut-Vincentova gingivitis) se mikrobiologicky jedná o fusospirochetovou infekci, vyvolanou především F. nucleatum, P. gingivalis, P. intermedia, B. vincenti, B. fusiformis.
124 Mikrobiální biofilmy Biofilm dutiny ústní Mikrobiální flóra implantátů Většina moderních dentálních implantátů se vhojuje osseointegrací (jedná se o tzv. funkční ankylózu). Kost přirůstá těsně k povrchu implantátu nebo se s ním dokonce chemicky spojí (biointegrace). Na povrchu implantátu není tedy v ideálním případě žádný biofilm a implantát zůstává sterilní. Dojde-li však k poruchám vhojování implantátu, např. ke tvorbě chobotů, může u takto obnažených či neosifikovaných implantátů dojít k infekci, periimplantitidě, která svým charakterem odpovídá parodontitidě (Takemura et al. 2004). V takovém případě jsou v infekčním ložisku zastoupeny spirochety a další anaerobní mikroflóra (Mutschelknauss et al. 2002), lze prokázat např. A. actinomycetemcomitans, P. gingivalis a P. intermedia. Obecně se zde nachází je zvýšený vyšší podíl grampozitivních bakterií (Hellwig et al. 2003). U vyjímatelných zubních náhrad se mohou díky nedostatečné ústní hygieně či hygieně zubních náhrad přemnožit kvasinky, zejména rodu Candida, které potom způsobují vleklé a obtížně léčitelné infekce dásní a měkkého patra. Kvasinky ve své invazívní podobě tvoří místo klasických kvasinkových buněk pseudohyfy, tedy nepravá vlákna, která se stávají součástí biofilmové struktury. U zubní náhrady dochází nejen k tvorbě kvasinkového biofilmu na povrchu této náhrady, ale může dojít i k prorůstání kvasinkových pseudohyf skrze plastový materiál zubní náhrady, což potom celou zubní náhradu nevratně znehodnocuje (Quirynen et al. 2002). 4.7 Patogenní determinanty kariogenních bakterií Kariogenní potenciál plaku je dán složením mikrobiální flóry a jejím metabolizmem. Rozhodující význam má acidogenní potenciál plaku, acidotolerance některých bakteriálních druhů v zubním plaku a schopnost produkovat extracelulární a intracelulární polysacharidy. Acidogenita plaku většina mikrobiálních druhů plaku může metabolizovat uhlovodíky na organické kyseliny. Acidogenní bakterie jsou tedy v zubním plaku přítomny vždy. Činností těchto bakterií vzrůstá kyselost zubního plaku a dochází pozvolna k poklesu ph a k demineralizaci zubní skloviny. Zejména S. mutans tvoří organické kyseliny z uhlovodíků obsažených ve stravě více a rychleji než ostatní streptokoky a aktinomycety. Je tomu tak zejména díky tzv. absorpčnímu systému cukr-fosfotransferáza, což je vysoce afinitní proces urychlující transport cukrů ze stravy. Vysoké rychlosti glykolýzy (acidogenity) mohou vést ke konečnému ph nižšímu než 4,5 v pouhých několika minutách. Streptokoky skupiny mutans mají těchto transportních systémů více. Konečným produktem metabolizmu acidogenních bakterií je směs organických kyselin (např. kyselina mléčná, octová), jejichž vzájemný poměr a koncentrace mají pro úroveň demineralizace podstatný význam. Metabolizmus acidogenních bakterií lze ztlumit přítomností fluoridových iontů v plaku. Tolerance a schopnost růstu při nízkém ph růst mnoha bakterií nalézaných na zdravé zubní sklovině (např. S. sanguis je v růstu inhibován ph nižším než 5,5, zatímco toto ph je pro kariogenní streptokoky optimální). Streptokoky skupiny mutans vykazují při nižším ph nezměněnou životaschopnost, což ve spojení s jejich schopností rozkládat uhlovodíky na organické kyseliny tvoří začarovaný kruh přítomnost mutans streptokoků znamená vyšší produkci kyselin a tedy snížení ph snížení ph znamená úbytek přirozené mikroflóry a posun přežívající mikroflóry směrem k acidotolerantním streptokokům skupiny mutans jejich vyšší přítomnost vede opět k vyšší produkci organicných kyselin atd. Syntéza extracelulární polysacharidové matrix tyto polymery napomáhají tvořit matrix zubního plaku. Glukosyltransferázy mění sacharózu na rozpustné a nerozpustné glukany, které upevňují bakteriální adhezi. S. mutans též produkuje specifický, vysoce nerozpustný polymer (mutan). Fruktosyltransferázy mění sacharózu na fruktany, které jsou rozpustné a mohou být využity bakteriemi v zubním plaku jako zdroj živin.
125 Mikrobiální biofilmy Biofilm dutiny ústní Intracelulární syntéza polysacharidů může být využita během nedostatku živin a katabolizována na kyselinu, pokud nejsou k dispozici cukry ze stravy. Mezi aktivitou zubního kazu a počtem bakterií produkujících intracelulární polysacharidy byl prokázán pozitivní vztah. Bakterie, které tvoří intracelulární polysacharidy patří mezi tzv. acidurické, což znamená, že jsou schopné tvořit kyseliny i bez přísunu exogenních cukrů. 4.8 Fokální infekce Dentální fokální infekce je tedy ovlivnění vzdálených tkání a orgánů infekčním zánětem (Li et al., 2000). Nejčastější onemocnění tkání dutiny ústní jsou způsobena dlouhodobou perzistencí bakteriálního povlaku (např. zubní kaz, onemocnění parodontu, chronická periodontitis). Mikroflóra dutiny ústní může vzdálené tkáně ovlivňovat třemi různými způsoby: Vznikem metastatické infekce k té může dojít při disseminaci mikroba krevním řečištěm (dochází k bakteriémii). Při chirurgických zákrocích v dutině ústní dochází k průniku velkého množství bakterií do krevního oběhu. Bakteriémie vzniká i při běžné extrakci zubů, při odstraňování zubního kamene, ale i při nevinném žvýkání žvýkačky či tužší stravy, irigaci při ústní hygieně apod. Imunitní systém hostitele si ve valné většině případů s přítomností bakterií v krevním řečišti poradí, problémy může tato dočasná bakteriémie způsobit pouze u disponovaných jedinců. Následkem bakteriémie může být u disponovaných jedinců např. vznik endokarditidy či perikarditidy (Li et al., 2000). K tomuto onemocnění disponují některé typy vrozených srdečních vad, přítomnost umělé srdeční chlopně apod. Viridující streptokoky, které jsou běžnou součástí mikroflóry dutiny ústní způsobují subakutní až chronické formy endokarditid a perikarditid, stejně jako např. H. parainfluenze, další zástupce běžné ústní mikroflóry (Hellwig et al. 2003; Pávek et al. 1991). Vznikem metastatického poškození uvolněním exotoxinů, tedy produktů bakteriálního metabolizmu toxického charakteru, do krevního oběhu. Působením toxických metabolitů některých zástupců běžné flóry dutiny ústní může dojít ke vzniku či progresi ateromatóznich plátů v cévní stěně a výsledkem může být ischemická choroba srdeční, angina pectoris či infarkt myokardu (Li et al., 2000). Vznikem metastatického zánětu imunitní reakcí protilátek se specifickým antigenem dochází k tvorbě imunokomplexů, které se hromadí ve vysoce vaskularizovaných orgánech např. glomerulech ledvin či ciliárním tělese oka. K tvorbě imunokomplexů může též dojít až v cílové tkáni, či může vlivem zkřížené reakce protilátek dojít ke vzniku např. revmatické horečky. Možnost vzniku fokálních infekcí nebyla dosud ani potvrzena ani vyloučena. Stav, který se jako následek dentální fokální infekce popisuje nejvíce a nejčastěji, je infekční endokarditida. Intenzivně se řeší vztah mezi parodontitidou a rozvojem či vznikem aterosklerózy. Nezbytný je v této oblasti další výzkum (Závodský & Bednář 2005). 4.9 Detekce a likvidace plaku Detekce plaku Rozhodujícím kritériem hodnocení úrovně ústní hygieny je rozsah a množství plaku, hodnocení lokalizace a rozsahu plaku poskytuje představu o úrovni ústní hygieny konkrétního jedince. Prostou aspekcí nelze zubní plak hodnotit, jeho hodnocení je nutno provádět spolehlivou a objektivní metodou.
126 Mikrobiální biofilmy Biofilm dutiny ústní Detekce plaku je možná dvojím způsobem vizuálně či hmatem. Při hodnocení nárůstu zubního plaku hmatem je stomatologickou sondou seškrabáván nános plaku, přičemž se zrakem hodnotí množství a rozsah plaku. K vizuálnímu hodnocení, tj. obarvení plaku speciálním činidlem, jsou nezbytné detekční roztoky, barvící selektivně zubní plak. Používané indikátory plaku musí být netoxické, nesmí alergizovat, nesmí barvit slinu, ústní sliznici ani výplňové materiály. K nejčastěji užívaným činidlům patří 5% roztok erythrosinu B, 2% roztok malachitové zeleni, 2% roztok genciánové violeti, hněď Bismarcova B a další. Zubní plak obarvený roztokem erythosinu B je uveden v barevné příloze na obr Vlastní posouzení úrovně ústní hygieny umožňují hygienické indexy, založené na prvním způsobu detekce. V ordinacích zubního lékaře se můžeme setkat s OHI-S indexem, který je již spíše obsolentní, dále tzv. API index, jenž hodnotí přítomnost plaku v aproximálním prostoru, hodnocení se vyjadřuje znaménky + a -. Běžně užívaný je plakový index PlI, hodnotící přítomnost plaku číselně 0 3 dle množství (Kilian et al.1996) Odstranění plaku Odstranění zubního plaku je možné pouze mechanicky, a to čištěním pomocí kartáčku a dalších pomůcek domácí ústní hygieny. Tam, kde selhávají či nestačí mechanické prostředky nastupují prostředky chemické, které musí splňovat následující požadavky (Kilian et al. 1996): zvyšování odolnosti tvrdých zubních tkání (přísunem fluoridových iontů z preparátů) zamezení tvorby zubního plaku likvidace již vytvořeného zubního plaku (např. abrazivní složky zubních past) usnadnění jeho mechanického odstranění. Z roztoků se v současné době se používá zejména chlorhexidin, sanguinarin a triclosan. Chlorhexidin je bis-biguanid, povrchově aktivní látka ze skupiny kationických tenzidů. Byl syntetizován ve 40. letech minulého století, v 60. letech byl pak objeven jeho protiplakový účinek. Chlorhexidin je antiseptikum se širokým spektrem antimikrobiálních účinků, působí dobře na grampozitivní bakterie, méně na gramnegativní bakterie, má účinky antimykotické i antivirové. Doporučuje se podávání 0,12% roztoku 5-10, max. však 14 dní. V akutních stavech se doporučuje koncentrace vyšší, a to 0,2% roztok. Nevýhodou je hořká chuť a četné vedlejší účinky, zvýrazněné zejména u vyšších koncentrací (poruchy chuti, černání jazyka, zubů i výplní), tyto změny jsou však reverzibilní a po ukončení terapie se vše navrací k normálu (Slezák 2003; Heinz 2000). Chlorhexidin je součástí roztoků ústní hygieny, gelů a zubních past. Sanguinarin (Sa) je kvartérní alkaloid isochinolinového typu, zdrojem je rostlina Sanguinaria canadensis. Kromě účinků antimikrobiálních má i účinky antiflogistické. Nežádoucí účinky jsou mírnější než u chlorhexidinu, udává se prakticky jen hnědavé zabarvení zubů. Obvyklá koncentrace je 0,045 0,05 %. Sanguinarin je součástí zubních past, roztoků a gelů. Triclosan je fenol, který vykazuje antimikrobiální aktivitu se širokým spektrem. Je účinný proti široké škále gramnegativních i grampozitivních bakterií, má antimykotické účinky. Triclosan je prostředkem, který nemá při klinickém použití vedlejší nežádoucí účinky. Minimální inhibiční koncentrace pro patogeny dutiny ústní je 3 µg/ml. Triclosan je součástí zubních past a ústních vod (Kilian et al. 1996). Úloha zubních past spočívá v odstranění zubního plaku a zbytků potravy z dutiny ústní. Zubní pasta se skládá z mnoha složek. Nejdůležitější složkou zubních past jsou
127 Mikrobiální biofilmy Biofilm dutiny ústní abrazivní látky. Podíl těchto látek tvoří až 60 % celkového objemu zubní pasty. Abraziva neboli brusné částice, zajišťují mechanické odstranění plaku a pigmentů a současně leští povrch skloviny. Jejich velikost se pohybuje mezi 0,1 1,0µm. K nejběžněji užívaným abrazivům patří křemík, oxid hlinitý, dále pak různé uhličitany a fosfáty. Aktivní, nebo též fakultativní složky zubních past mohou zvyšují odolnost zubní skloviny, snižují citlivost krčků a současně snižují tvorbu mikrobiálního povlaku (fluoridy, triclosan, chlorhexidin aj.). Důležitou složkou zubních past jsou zvlhčující složky, tedy jsou látky zabraňující tvrdnutí pasty a jejímu vysychání, např. glycerin či sorbitol. Pěnící složka či detergenty, jsou látky, které snižují povrchové napětí. Mají též slabý antibakteriální účinek. Nejčastěji užívanou látkou je laurylsulfát sodný. Konzervační látky zajišťují stabilitu zubních past, chuťové přísady, barviva a sladidla zpříjemňují jejich používání. Aby nedošlo k oddělení jednotlivých součástí zubní pasty, používají se stabilizátory (gely, tužidla). Vzájemný poměr složek v různých zubních pastách je samozřejmě odlišný (Kilian et al. 1996; Mutschelknauss et al. 2002) Antimikrobiální léčba Terapie parodontu spočívá v eliminaci infekce parodontu, která vychází ze zubního biofilmu. Běžně užívaným lokálním antiseptikem je chlorhexidin, nejčastěji ve formě gelu či roztoku (Stelzel 2003). V poslední době se můžeme setkat s metodou tzv. kontrolovaného uvolňování chlorhexidinu. Jedná se o použití tzv. PerioChip, kdy se čep napuštěný chlorhexidinem aplikuje do parodontálního chobotu, který jinak nereaguje na běžnou léčbu. Chlorhexidin se z čepu pozvolna uvolňuje do prostoru s mikrobiální infekcí, účinná koncentrace chlorhexidinu byla ve slinách prokázána ještě 24 hodin po aplikaci. Čep je resorbovatelný, základem je želatinová matrice a aplikaci lze opakovat dle potřeby (Harnack et al. 2005). Chlorhexidin má své uplatnění i v terapii silně karies-rezistentních pacientů. Pro pacienty, kteří i přes fluoridovou profylaxi vykazují nadále silnou aktivitu zubního kazu, existují dvě možnosti ošetřování. Jednak omezení přísunu uhlohydrátů pomocí přísné diety, nebo eliminace kariogenních bakterií pomocí lokálních orálních antiseptik. Klinicky se vyzkoušelo použití 1% chlorhexidinového gelu, který se aplikuje na zuby pomocí individuálně zhotovené lisované dlahy. Takováto dlaha se ambulantně aplikuje po dobu jednoho týdne vždy asi na 10 minut denně, podmínkou jsou čerstvě vyčištěné zuby. Publikovány jsou též případy použití laku s vysokou koncentrací chlorhexidinu, jenž se aplikuje po dobu minut na povrch zubů. Podmínkou je suché pole, tedy zamezení přístupu slin, pomocí vatových válečků (Ruppert & Shlagenhauf 2004). Při zvláště vleklém průběhu onemocnění či celkově velmi špatném zdravotním stavu pacienta se uchylujeme k systémové antimikrobiální léčbě. Na základě stanovení citlivosti patogenů k antibiotikům volíme vhodnou terapii jednotlivými antibiotiky či jejich kombinací. K nejužívanějším antibiotikům v zubním lékařství patří metronidazol, klindamycin, amoxicillin (monoterapie nebo v kombinaci s metronidazolem), doxycyklin a tetracyklin (Smola et al. 2001).
128 Mikrobiální biofilmy Biofilm dutiny ústní Literatura 1. Banas JA, Vickerman MM (2003): Glucan-binding proteins of the oral streptococci. Crit Rev.Oral Biol.Med. 14: Bradshaw DJ, Marsh PD, Watson GK, Allison C (1998): Role of Fusobacterium nucleatum and coaggregation in anaerobe survival in planktonic and biofilm oral microbial communities during aeration. Infect.Immun. 66: Burne RA, Quivey RG, Jr., Marquis RE (1999): Physiologic homeostasis and stress responses in oral biofilms. Methods Enzymol. 310: : Busscher HJ, van der Mei HC (1997): Physico-chemical interactions in initial microbial adhesion and relevance for biofilm formation. Adv.Dent.Res. 11: Jin Y, Yip HK (2002): Supragingival calculus: formation and control. Crit Rev.Oral Biol.Med. 13: Kolenbrander PE, Andersen RN, Kazmerzak K, Wu R, Palmer RJ, Jr. (1999): Spatial organization of oral bacteria in biofilms. Methods Enzymol. 310: : Kolebrander PE, Lerud RF, Blehert DS, England PG, Foster JS, Palmer RJ (2003): The role of coaggregation in oral biofilm formation. in: Lens P, Moran AP, Mahony T, Stoodley P, O Flaherty V (2003): Biofilms in Medicine, Industry and Environmental Biotechnology. IWA Publishing, London, Kolenbrander PE, Palmer RJ, Jr., Rickard AH, Jakubovics NS, Chalmers NI, Diaz PI (2000): Bacterial interactions and successions during plaque development. Periodontol. 2006;42:47-79.: Lamont RJ, Jenkinson HF (2000): Subgingival colonization by Porphyromonas gingivalis. Oral Microbiol.Immunol. 15: Li YH, Lau PC, Lee JH, Ellen RP, Cvitkovitch DG (2001): Natural genetic transformation of Streptococcus mutans growing in biofilms. J.Bacteriol. 183: Listgarten MA (2000): The structure of dental plaque. Periodontol. %1994 Jun;5:52-65.: Marsh PD (1999): Microbiologic aspects of dental plaque and dental caries. Dent.Clin.North Am. 43: 599-5vi. 13. Marsh PD (2004): Dental plaque as a microbial biofilm. Caries Res. 38: Olsson J, van der HY, Holmberg K (1992): Plaque formation in vivo and bacterial attachment in vitro on permanently hydrophobic and hydrophilic surfaces. Caries Res. 26: Paster BJ, Olsen I, Aas JA, Dewhirst FE (2000): The breadth of bacterial diversity in the human periodontal pocket and other oral sites. Periodontol. %2006;42:80-7.: Rosan B, Lamont RJ (2000): Dental plaque formation. Microbes.Infect. 2: Rudney JD (2000): Saliva and dental plaque. Adv.Dent.Res. 14:29-39.: Rudney JD, Pan Y, Chen R (2003): Streptococcal diversity in oral biofilms with respect to salivary function. Arch.Oral Biol. 48: Sbordone L, Bortolaia C (2003): Oral microbial biofilms and plaque-related diseases: microbial communities and their role in the shift from oral health to disease. Clin.Oral Investig. 7: Suzuki N, Yoshida A, Nakano Y (2005): Quantitative analysis of multi-species oral biofilms by TaqMan Real-Time PCR. Clin.Med.Res. 3: Svensater G, Welin J, Wilkins JC, Beighton D, Hamilton IR (2001): Protein expression by planktonic and biofilm cells of Streptococcus mutans. FEMS Microbiol.Lett. 205:
129 Mikrobiální biofilmy Biofilm dutiny ústní 22. Takemura N, Noiri Y, Ehara A, Kawahara T, Noguchi N, Ebisu S (2004): Single species biofilm-forming ability of root canal isolates on gutta-percha points. Eur.J.Oral Sci. 112: Wecke J, Kersten T, Madela K, Moter A, Gobel UB, Friedmann A, Bernimoulin J (2000): A novel technique for monitoring the development of bacterial biofilms in human periodontal pockets. FEMS Microbiol.Lett. 191: Kolebrander PE, Lerud RF, Blehert DS, England PG, Foster JS, Palmer RJ (2003): The role of coaggregation in oral biofilm formation. in: Lens P, Moran AP, Mahony T, Stoodley P, O Flaherty V (2003): Biofilms in Medicine, Industry and Environmental Biotechnology. IWA Publishing, London,
130 7.4 Holá V., Růžička F., Votava M., 2004: Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis; Microbiologica; vol. 27; 3:
131 Impact of surface coating on the adherence of slime producing and nonproducing Staphylococcus epidermidis Veronika Holá, Filip Růžička, Miroslav Votava Institute for Microbiology, Faculty of Medicine, Masaryk University, Pekařská 53, Brno, Czech Republic Postal address: Institute for Microbiology LF MU a FN USA Pekařská Brno Czech Republic Running title: Impact of surface coating on the adherence Keywords: Staphylococcus; epidermidis; biofilm; polystyrene; growth conditions; adhesion; ica-operone Corresponding author: Veronika Holá, Institute for Microbiology, Faculty of Medicine, Masaryk University, Pekařská 53, Brno, Czech Republic; phone: , fax: ; veronika.hola@fnusa.cz 1
132 Abstract The ability of Staphylococcus epidermidis to grow in the form of a biofilm not only facilitates its persistence in the host, but also allows it to survive at antibiotic concentrations several orders higher than the Minimum Inhibitory Concentration (MIC). We evaluated different surface treatments of hardened polystyrene in order to develop a model system for growth of S. epidermidis as a biofilm. We assayed for biofilm growth of S. epidermidis clinical isolates on unmodified polystyrene, on polystyrene modified by chemical abrasion and on polystyrene modified by sulfonation, using either Tryptic Soya Broth or Brain Heart Infusion as a growth medium. We concluded that sulfonated polystyrene and Brain Heart Infusion provided the best growth system for predicting the ability of a clinical isolate to form biofilm (Akaike value ). Using this method, biofilm formation was detected in 14 (70 %) of ica-positive strains and negative in 16 (80 %) of ica-negative strains. Short communication Coagulase-negative staphylococci and particularly Staphylococcus epidermidis are the most frequent causes of nosocomial infections, especially catheter- and medical device-related sepsis. Many pathogenic strains of S. epidermidis are slime producers and can grow in a biofilm form. Several methods describe how to determine the ability of bacteria to produce slime and grow in a biofilm (Christensen et al., 1982; Christensen et al., 1985; Mulder et al., 1998; Stepanovic et al., 2000; An et al., 2001 and others); however, for the study of antimicrobial resistance under routine laboratory conditions, these methods are difficult to use, time-consuming, or completely inappropriate. For example, Ceri et al. (1999) described a method for detection of biofilm growth and evaluation of antimicrobial resistance using a device known as the Calgary Biofilm Device. The device seems to be convenient for the assessment of the antimicrobial resistance, but the method is not available for routine laboratory use. The aim of our study was to evaluate the applicability of different surface 2
133 treatments of hardened-polystyrene-pegged plates for biofilm formation and for subsequent assessment of minimum biofilm inhibitory concentration of antibiotics (MBIC) in routine laboratory conditions. Forty Staphylococcus epidermidis strains isolated from blood cultures of patients with bacteraemia were used in this study. Isolates were identified to the species level by the STAPHYtest System (Lachema, Brno, Czech Republic), with confirmation by PCR detection of S. epidermidis-specific DNA (Martineau et al, 1996). Bacterial cultures were grown at 37 C overnight on Mueller-Hinton agar (MHA, Oxoid, Basingstoke, UK). Isolates were classified into two groups based on the presence or absence of ica sequences following PCR amplification (Frebourg et al., 2000). All isolates (20 ica-positive and 20 ica-negative) were examined phenotypically by assessing their growth on Congo Red agar (CRA) (Himedia, Mumbai, India) as described by Freeman et al. (1989) and by using a modification of the Christensen tube method (Christensen et al., 1982), in which Brain Heart Infusion (BHI) was substituted for Tryptic Soya Broth (TSB). For evaluation of bacterial growth, we used plates of hardened polystyrene designed with pegs to match the wells of a standard 96-well microtiter plate. We tested biofilm growth in both TSB (Oxoid, Basingstoke, UK) and BHI (Himedia, Mumbai, India) media on unmodified pegs, with the surface cleaned from all contaminants and chemicals before testing, and on pegs modified by chemical abrasion with acetone or by sulfonation using 96% sulphuric acid. Abrasion of hardened polystyrene by means of acetone slightly impairs and erodes the surface layer, resulting in the formation of small cavities. Sulfonation of hardened polystyrene decreases the hydrophobicity of the polystyrene surface layer (Kucera and Jancar, 1996) without changing the structure. To prepare biofilm cultures, we used fresh overnight cultures in MHA to inoculate the selected medium to a density of 0.5 on the McFarland scale. The medium was aliquoted into 3
134 microtiter plates (150 µl/well), and the polystyrene plates were fitted over the microtiter plates, with pegs submerged into the wells. The plates were incubated for 48 hours at 37 C under constant agitation (25 rpm) on an orbital shaker (Tool MIX 30K, Tool s.r.o., Prague, Czech Republic). After incubation, the pegged plates were gently washed with sterile physiological saline and adherent biofilm was stained with crystal violet. We evaluated the homogeneity and quantity of the biofilm using the following 3-point scale: 0 non-adherent; 1 - weak or non-homogenous biofilm layer; 2 - strong and homogenous biofilm layer. Data were analyzed by the General Linear Models using S-plus, 4.0 (MathSoft Inc., 1997), using the logit link function for reaching data transformation. Test evaluation was performed using the Akaike information criterion (AIC), which assesses the relative fit of competing models. A lower AIC value indicates a closer fit of the real data to the model (Hsu and Nelson, 1998). All tests were examined at the level equivalent to P values < Figures 1 and 2 show the results of all tests evaluating slime production in the icapositive and ica-negative strains, respectively, including the tube test with TSB broth, the tube test with BHI broth, the CRA method, and the pegged-plates with sulfonated pegs, with abraded pegs, and with unmodified pegs. None of the 20 ica-positive strains tested was positive under all conditions of any method, but none was completely negative with any method. All ica-positive strains produced a weak or non-homogenous biofilm layer with one or more methods. Of the 20 ica-negative strains tested, 8 were negative. The remaining 12 strains produced a weak or non-homogenous biofilm layer with one or more methods. Discordant positive results were seen in one strain grown in BHI on plates with acetone-abraded pegs and in another strain grown in BHI on plates with unmodified pegs. The highest predicative values for both positive and negative production of slime were obtained using BHI broth with sulfonated pegged plates and using BHI broth in the standard tube test (AIC value of for both methods). Using sulfonated pegged plates and BHI, we 4
135 correctly identified 70% (14/20) of the ica-positive strains and 80% (16/20) of the icanegative strains. Coagulase-negative species of the genus Staphylococcus were initially regarded as non-pathogenic, but are now frequently isolated not only as causative agents of clinical infections in humans, but also as causative agents of many sub-clinical and chronic infections (Costerton et al., 1999). Pathogenicity factors, especially the ability to form biofilm, help the bacteria survive in the host. Consequently, the ability to determine whether a strain can form a biofilm and to understand and dissect the process under routine laboratory conditions is clinically important. Our investigation was based on whether clinical isolates were positive or negative for ica sequences as determined by PCR analysis. This classification appears to predict biofilm growth better than studies analyzing only ica-positive wild strains and icapositive controls, without any ica-negative controls. Our study suggests that more reliable growth results are obtained with a more nutrientrich medium such as BHI, irrespective of the underlying material. The growth tests performed in TSB, including the tube test and the various tests with pegged plates, gave much worse results than those performed in BHI, with AIC values ranging from for tests in TSB, compared to AIC values ranging from for tests in BHI. In this respect our results contradict those of Christensen et al. (Christensen et al., 1982; Christensen et al., 1985), although they support the results obtained by others (Mulder and Degener, 1998; Knobloch et al., 2002). Of the materials we tested, the one most suitable for supporting biofilm growth was sulfonated polystyrene, which was effective with both in BHI medium and in TSB medium. We cannot recommend the use of the pegged plate without surface modification. When we used plates with unmodified polystyrene, the adherent layer was easily disrupted by washing, making unmodified polystyrene less suitable as a support material than either sulfonated 5
136 polystyrene or polystyrene modified by chemical abrasion. In summary, we recommend the use of a sulfonated pegged plate with BHI medium for biofilm growth of slime-producing strains of S. epidermidis. Acknowledgements The study was supported by the grant IGA from Ministry of Health, Czech Republic and by the fund FRVŠ from Ministry of Education, Czech Republic. The authors are indebted to Barbara J. Rutledge of Corvin Resources, Inc., for revising the language of the manuscript. Literature cited AN, Y.H., MCGLOHORN, J.B., BEDNARSKI, B.K., MARTIN, K.L., AND FRIEDMAN, R.J. (2001). An open channel flow chamber for characterizing biofilm formation on biomaterial surfaces. Methods Enzymology, 337, CERI, H., OLSON, M.E., STREMICK, C., READ, R.R., MORCK, D., AND BURET, A. (1999). The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. Journal of Clinical Microbiology, 37, CHRISTENSEN, G.D., SIMPSON, W.A., BISNO, A.L., AND BEACHEY, E.H. (1982). Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infection and Immunity, 37, CHRISTENSEN, G.D., SIMPSON, W.A., YOUNGER, J.J., BADDOUR, L.M., BARRETT, F.F., MELTON, D.M., BEACHEY, E.H. (1985). Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. Journal of Clinical Microbiology, 22,
137 COSTERTON, J.W., STEWART, P.S., AND GREENBERG, E.P. (1999). Bacterial biofilms: a common cause of persistent infections. Science, 284, FREBOURG, N.B., LEFEBVRE, S., BAERT, S., AND LEMELAND, J.F. (2000). PCR-Based assay for discrimination between invasive and contaminating Staphylococcus epidermidis strains. Journal of Clinical Microbiology, 38, FREEMAN, D.J., FALKINER, F.R., KEANE, C.T. (1989). New method for detecting slime production by coagulase negative staphylococci. Journal of Clinical Pathology, 42, HSU, J.C., AND NELSON, B. (1998). Multiple comparisons in the general linear model. Journal of Computational and Graphical Statistics, 7, KNOBLOCH, J.K., HORSTKOTTE, M.A., ROHDE, H., MACK, D. (2002). Evaluation of different detection methods of biofilm formation in Staphylococcus aureus. Medical Microbiology and Immunology (Berl), 191, KUCERA, F., AND JANCAR, J. (1996). Preliminary study of sulfonation of polystyrene by homogeneous and heterogeneous reaction. Chemical Papers-Chemicke Zvesti, 50, MAH, T.F., AND O'TOOLE, G.A. (2001) Mechanisms of biofilm resistance to antimicrobial agents. Trends in Microbiology, 9, MARTINEAU, F., PICARD, F.J., ROY, P.H., OUELLETTE, M. AND BERGERON, M.G. (1996). Species-specific and ubiquitous DNA-based assays for rapid identification of Staphylococcus epidermidis. Journal of Clinical Microbiology, 34, MULDER, J.G., DEGENER, J.E. (1998). Slime-producing properties of coagulase-negative staphylococci isolated from blood cultures. Clinical Microbiology and Infection, 4, PEREZ-GIRALDO, C., RODRIGUEZ-BENITO, A., MORAN, F.J., HURTADO, C., BLANCO, M.T. AND GOMEZ-GARCIA, A.C. (1995). Influence of the incubation atmosphere on the production of slime by Staphylococcus epidermidis. European Journal of Clinical Microbiology and Infectious Diseases, 14,
138 STEPANOVIC, S., VUKOVIC, D., DAKIC, I., SAVIC, B., AND SVABIC-VLAHOVIC, M. (2000). A modified microtiter-plate test for quantification of staphylococcal biofilm formation. Journal of Microbiological Methods, 40,
139 Figure 1 Ica-positive strains 100% 90% 80% 70% 60% 50% negative slightly positive 40% 30% 20% 10% 0% CRA Ch-TSB Ch-BHI pegs- TSB 4 10 pegs- BHI 3 pegs A-TSB 12 pegs A-BHI 5 3 pegs S-TSB 14 pegs S-BHI positive 9
140 Figure 2 Ica-negative strains 100% 90% 80% 70% 60% 50% negative slightly positive 40% positive 30% 20% 10% 0% 3 2 CRA Ch-TSB Ch-BHI pegs- TSB pegs- BHI pegs A-TSB pegs A-BHI 1 pegs S-TSB 4 pegs S-BHI 10
141 Figure 1 Results of all tests for the determination of slime production in investigated ica-positive strains in percentage terms. Key to the methods compared CRA Congo Red Agar method, Ch-TSB - tube test with TSB, Ch-BHI - tube test with BHI, pegs-tsb unadapted pegs in TSB, pegs-bhi unadapted pegs in BHI, pegs A-TSB - abraded pegs in TSB, pegs A-BHI abraded pegs in BHI, pegs S-TSB - sulfonated pegs in TSB, pegs S-BHI - sulfonated pegs in BHI Figure 2 Results of all tests for the determination of slime production in investigated ica-negative strains in percentage terms. Key to the methods compared CRA Congo Red Agar method, Ch-TSB - tube test with TSB, Ch-BHI - tube test with BHI, pegs-tsb unadapted pegs in TSB, pegs-bhi unadapted pegs in BHI, pegs A-TSB - abraded pegs in TSB, pegs A-BHI abraded pegs in BHI, pegs S-TSB - sulfonated pegs in TSB, pegs S-BHI - sulfonated pegs in BHI 11
142 7.5 Holá V., Růžička F., Votava M., 2006: Dynamic of Staphylococcus epidermidis biofilm formation in relation to nutrition, temperature and time; Scripta Medica (Brno) 79; 3:
143
144
145
146
147
148
149 7.6 Růžička F., Holá V., Votava M., Tejkalová R., Horváth R., Heroldová M., Woznicová V., 2004: Biofilm detection and the clinical significance of Staphylococcus epidermidis isolates; Folia Microbiologica; 49; 5:
150
151
152
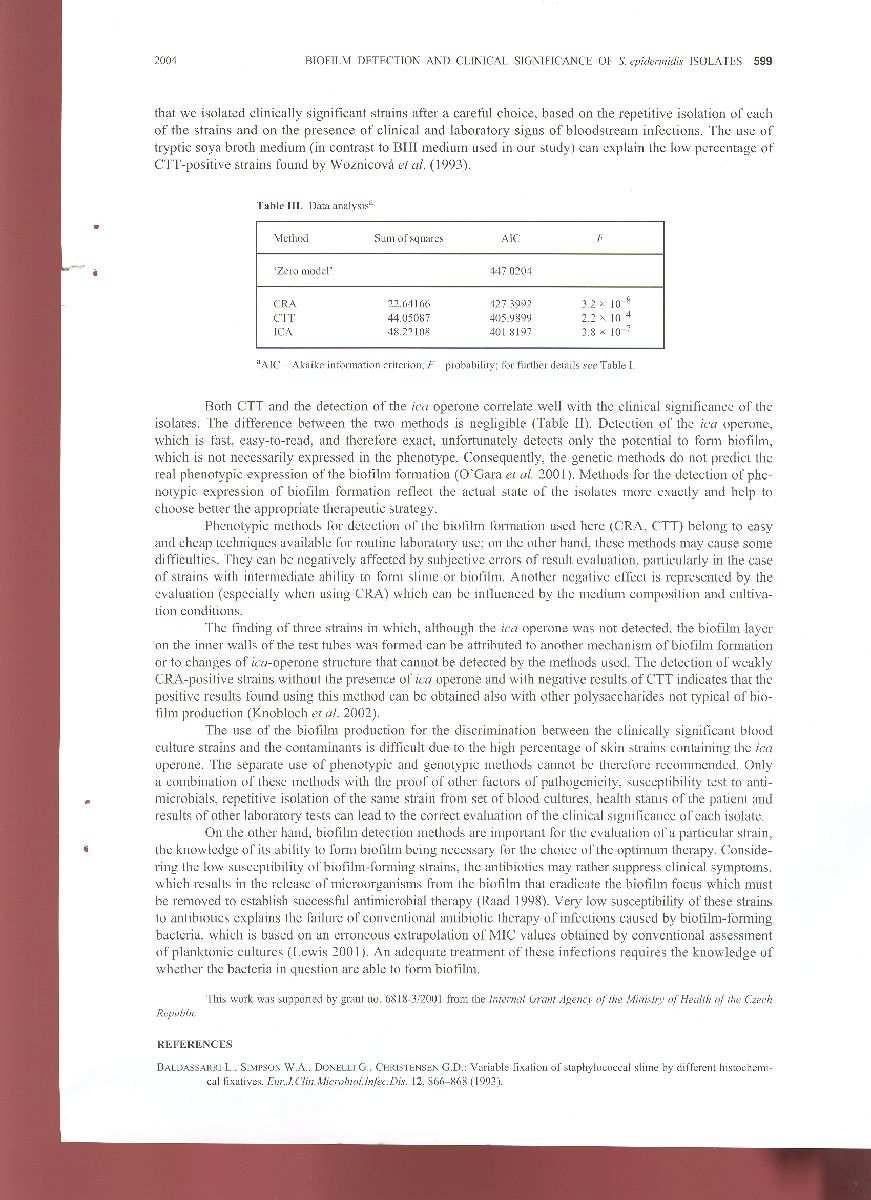
153
154
155 7.7 Holá V., Růžička F., Votava M. 2004: Rozdíly v citlivosti k antibiotikům u biofilmpozitivních a biofilmnegativních kmenů Staphylococcus epidermidis izolovaných z hemokultur; Epidemiologie, mikrobiologie, imunologie; 53; 2:
156

157
158

159
160 7.8 Holá V., Růžička F., Tejkalová R., Votava M., 2004: Stanovení citlivosti k antibiotikům u biofilmpozitivních forem mikroorganizmů; Klinická mikrobiologie a infekční lékařství; 10; 5:

161

162
tvoří tvoř biofilm biofilm tvoří biof tvoř ilm tvoří biof tvoř ilm tvoří biofilm tvoř
 Tvorba biofilmu u mikrobů izolovaných z klinického materiálu MUDr. Černohorská Lenka, Ph.D. Kdo tvoří biofilm (bakterie, viry, kvasinky, 1druh, více druhů, multibakteriální komunity) Kde se tvoří biofilm
Tvorba biofilmu u mikrobů izolovaných z klinického materiálu MUDr. Černohorská Lenka, Ph.D. Kdo tvoří biofilm (bakterie, viry, kvasinky, 1druh, více druhů, multibakteriální komunity) Kde se tvoří biofilm
Biofilm a jeho význam
 Biofilm a jeho význam Filip Růžička Mikrobiologický ústav LF MU a FN u sv. Anny v Brně Přednáška pro PrF 2016 HISTORIE 1676 Antony van Leeuwehoek sledoval bakterie v zubním povlaku Od dob Pasteura a Kocha
Biofilm a jeho význam Filip Růžička Mikrobiologický ústav LF MU a FN u sv. Anny v Brně Přednáška pro PrF 2016 HISTORIE 1676 Antony van Leeuwehoek sledoval bakterie v zubním povlaku Od dob Pasteura a Kocha
Biofilm a jeho význam
 Biofilm a jeho význam Filip Růžička Mikrobiologický ústav LF MU a FN u sv. Anny v Brně Přednáška pro II. r. ZL 2012 HISTORIE 1676 Antony van Leeuwehoek sledoval bakterie v zubním povlaku Od dob Pasteura
Biofilm a jeho význam Filip Růžička Mikrobiologický ústav LF MU a FN u sv. Anny v Brně Přednáška pro II. r. ZL 2012 HISTORIE 1676 Antony van Leeuwehoek sledoval bakterie v zubním povlaku Od dob Pasteura
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva
 Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
ANALÝZA VÝSKYTU JEDNOTLIVÝCH BAKTERIÁLNÍCH DRUHŮ NA POVRCHU CENTRÁLNÍCH ŽILNÍCH KATÉTRŮ Autor: Mudroch T.
 ANALÝZA VÝSKYTU JEDNOTLIVÝCH BAKTERIÁLNÍCH DRUHŮ NA POVRCHU CENTRÁLNÍCH ŽILNÍCH KATÉTRŮ Autor: Mudroch T. Úvod Počet nákaz vzniklých v souvislosti s pobytem v nemocnici (tzv. nozokominální infekce) a způsobených
ANALÝZA VÝSKYTU JEDNOTLIVÝCH BAKTERIÁLNÍCH DRUHŮ NA POVRCHU CENTRÁLNÍCH ŽILNÍCH KATÉTRŮ Autor: Mudroch T. Úvod Počet nákaz vzniklých v souvislosti s pobytem v nemocnici (tzv. nozokominální infekce) a způsobených
DIAGNOSTIKA INFEKČNÍCH CHOROB KULTIVACE V LABORATORNÍCH PODMÍNKÁCH
 STŘEDNÍ ZDRAVOTNICKÁ ŠKOLA A VYŠŠÍ ODBORNÁ ŠKOLA ZDRAVOTNICKÁ ŽĎÁR NAD SÁZAVOU DIAGNOSTIKA INFEKČNÍCH CHOROB KULTIVACE V LABORATORNÍCH PODMÍNKÁCH MGR. IVA COUFALOVÁ DIAGNOSTIKA INFEKČNÍCH CHOROB KULTIVACE
STŘEDNÍ ZDRAVOTNICKÁ ŠKOLA A VYŠŠÍ ODBORNÁ ŠKOLA ZDRAVOTNICKÁ ŽĎÁR NAD SÁZAVOU DIAGNOSTIKA INFEKČNÍCH CHOROB KULTIVACE V LABORATORNÍCH PODMÍNKÁCH MGR. IVA COUFALOVÁ DIAGNOSTIKA INFEKČNÍCH CHOROB KULTIVACE
VÍME VŠE O PREVENCI NEMOCNIČNÍ INFEKCÍ SPOJENÝCH S KATETRIZACÍ CENTRÁLNÍ ŽÍLY?
 VÍME VŠE O PREVENCI NEMOCNIČNÍ INFEKCÍ SPOJENÝCH S KATETRIZACÍ CENTRÁLNÍ ŽÍLY? Jarmila Kohoutová Oddělení nemocniční hygieny FNOL VI. Markův den, Korunní pevnůstka Olomouc, 12.11.2015 KRITÉRIA CDC laboratorně
VÍME VŠE O PREVENCI NEMOCNIČNÍ INFEKCÍ SPOJENÝCH S KATETRIZACÍ CENTRÁLNÍ ŽÍLY? Jarmila Kohoutová Oddělení nemocniční hygieny FNOL VI. Markův den, Korunní pevnůstka Olomouc, 12.11.2015 KRITÉRIA CDC laboratorně
Prevence ventilátoro Prevence pneumonie
 Prevence ventilátorové pneumonie nové koncepty Renata Pařízková Klinika anesteziologie, resuscitace a intenzivní medicíny Univerzita Karlova v Praze, Lékařská fakulta v Hradci Králové Fakultní nemocnice
Prevence ventilátorové pneumonie nové koncepty Renata Pařízková Klinika anesteziologie, resuscitace a intenzivní medicíny Univerzita Karlova v Praze, Lékařská fakulta v Hradci Králové Fakultní nemocnice
Odběr a transport biologického materiálu do mikrobiologické laboratoře. Jana Juránková OKM FN Brno
 Odběr a transport biologického materiálu do mikrobiologické laboratoře Jana Juránková OKM FN Brno Správný výsledek mikrobiologického vyšetření začíná správným odběrem biologického materiálu a jeho správným
Odběr a transport biologického materiálu do mikrobiologické laboratoře Jana Juránková OKM FN Brno Správný výsledek mikrobiologického vyšetření začíná správným odběrem biologického materiálu a jeho správným
Funkční vzorek 5454/2017. Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních. látek u Pseudomonas spp.
 Funkční vzorek 5454/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Pseudomonas spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
Funkční vzorek 5454/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Pseudomonas spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
Streptokoky - průkaz fenotypu rezistence k antibiotikům ze skupiny makrolidů, linkosamidů a streptograminub
 Streptokoky - průkaz fenotypu rezistence k antibiotikům ze skupiny makrolidů, linkosamidů a streptograminub Revize 27. 1. 2014 A. Klinické vzorky pro vyšetření citlivosti Citlivost k antibiotikům (včetně
Streptokoky - průkaz fenotypu rezistence k antibiotikům ze skupiny makrolidů, linkosamidů a streptograminub Revize 27. 1. 2014 A. Klinické vzorky pro vyšetření citlivosti Citlivost k antibiotikům (včetně
MTI Cvičení č. 2 Pasážování buněk / Jana Horáková
 MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
Diagnostické metody v lékařské mikrobiologii
 Diagnostické metody v lékařské mikrobiologii Výuková prezentace z: Lékařské mikrobiologie Jan Smíšek ÚLM 3. LF UK 2009 Princip identifikace Soubor znaků s rozdílnou diskriminační hodnotou Základní problémy
Diagnostické metody v lékařské mikrobiologii Výuková prezentace z: Lékařské mikrobiologie Jan Smíšek ÚLM 3. LF UK 2009 Princip identifikace Soubor znaků s rozdílnou diskriminační hodnotou Základní problémy
NÁVRH A PŘÍPRAVA PEPTIDŮ A LIPOPEPTIDŮ S ANTIMIKROBIÁLNÍM ÚČINKEM A STUDIUM JEJICH BIOLOGICKÝCH VLASTNOSTÍ
 NÁVRH A PŘÍPRAVA PEPTIDŮ A LIPOPEPTIDŮ S ANTIMIKROBIÁLNÍM ÚČINKEM A STUDIUM JEJICH BIOLOGICKÝCH VLASTNOSTÍ A. Macůrková R. Ježek P. Lovecká V. Spiwok P. Ulbrich T. Macek Antimikrobiální peptidy přírodní
NÁVRH A PŘÍPRAVA PEPTIDŮ A LIPOPEPTIDŮ S ANTIMIKROBIÁLNÍM ÚČINKEM A STUDIUM JEJICH BIOLOGICKÝCH VLASTNOSTÍ A. Macůrková R. Ježek P. Lovecká V. Spiwok P. Ulbrich T. Macek Antimikrobiální peptidy přírodní
Mnohobuněčné kvasinky
 Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Vliv chemických desinfekčních metod na bakteriální biofilm
 MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMENTÁLNÍ BIOLOGIE Vliv chemických desinfekčních metod na bakteriální biofilm Bakalářská práce Markéta Dopitová VEDOUCÍ PRÁCE: doc. MUDr. Filip Růžička,
MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV EXPERIMENTÁLNÍ BIOLOGIE Vliv chemických desinfekčních metod na bakteriální biofilm Bakalářská práce Markéta Dopitová VEDOUCÍ PRÁCE: doc. MUDr. Filip Růžička,
Funkční vzorek 5452/2017. Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních. látek u gramnegativních bakterií II.
 Funkční vzorek 5452/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u gramnegativních bakterií II. řada Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Funkční vzorek 5452/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u gramnegativních bakterií II. řada Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Funkční vzorek 5456/2017. Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních. látek u Enterococcus spp.
 Funkční vzorek 5456/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Enterococcus spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
Funkční vzorek 5456/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Enterococcus spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
Nové technologie v mikrobiologické laboratoři, aneb jak ovlivnit čas k získání klinicky relevantního výsledku
 B I O M E D I C AL Nové technologie v mikrobiologické laboratoři, aneb jak ovlivnit čas k získání klinicky relevantního výsledku Jaroslav Hrabák CHARLES UNIVERSITY Obsah prezentace Historie, vývoj a trendy
B I O M E D I C AL Nové technologie v mikrobiologické laboratoři, aneb jak ovlivnit čas k získání klinicky relevantního výsledku Jaroslav Hrabák CHARLES UNIVERSITY Obsah prezentace Historie, vývoj a trendy
Tisková zpráva závěrečné zprávy projektu
 Tisková zpráva závěrečné zprávy projektu QJ1210119 Vývoj a výroba veterinárních setů pro stanovení minimálních inhibičních koncentrací antimikrobiálních látek standardizovanou mikrodiluční metodou a nový
Tisková zpráva závěrečné zprávy projektu QJ1210119 Vývoj a výroba veterinárních setů pro stanovení minimálních inhibičních koncentrací antimikrobiálních látek standardizovanou mikrodiluční metodou a nový
Ověřená technologie 4782/2017
 Ověřená technologie 4782/2017 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Enterococcus spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Ověřená technologie 4782/2017 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Enterococcus spp. Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Lactobacillus brevis kazit pivo
 Genetický základ schopnosti Lactobacillus brevis kazit pivo Mgr. Dagmar Matoulková, Mikrobiologie VÚPS Ing. Karel Sigler, DrSc., Mikrobiologický ústav AVČR 23. pivovarskosladařské dny, České Budějovice,
Genetický základ schopnosti Lactobacillus brevis kazit pivo Mgr. Dagmar Matoulková, Mikrobiologie VÚPS Ing. Karel Sigler, DrSc., Mikrobiologický ústav AVČR 23. pivovarskosladařské dny, České Budějovice,
Rezistence patogenů vůči antimikrobialním látkám. Martin Hruška Jan Dlouhý
 Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
Využití antibakteriálních testů v textilním průmyslu Mgr. Irena Šlamborová, Ph.D.
 Využití antibakteriálních testů v textilním průmyslu Mgr. Irena Šlamborová, Ph.D. Fakulta Přírodovědně-humanitní a pedagogická, katedra chemie OBSAH: 1. Stavba a fyziologie bakterií. 2. Kultivace bakterií,
Využití antibakteriálních testů v textilním průmyslu Mgr. Irena Šlamborová, Ph.D. Fakulta Přírodovědně-humanitní a pedagogická, katedra chemie OBSAH: 1. Stavba a fyziologie bakterií. 2. Kultivace bakterií,
Enterotoxiny Staphylococcus aureus. Jana Kotschwarová Andrea Koťová
 Enterotoxiny Staphylococcus aureus Jana Kotschwarová Andrea Koťová Obsah Charakteristika Staphylococcus aureus Vlastnosti Faktory virulence Enterotoxiny Patogeneze Výskyt Metody stanovení Prevence výskytu
Enterotoxiny Staphylococcus aureus Jana Kotschwarová Andrea Koťová Obsah Charakteristika Staphylococcus aureus Vlastnosti Faktory virulence Enterotoxiny Patogeneze Výskyt Metody stanovení Prevence výskytu
Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči. Marek Protuš
 Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči Marek Protuš Sepsis - 3 Sepse je definovaná jako život ohrožující orgánová dysfunkce způsobená dysregulovanou odpovědí
Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči Marek Protuš Sepsis - 3 Sepse je definovaná jako život ohrožující orgánová dysfunkce způsobená dysregulovanou odpovědí
Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči. Marek Protuš
 Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči Marek Protuš Sepsis - 3 Sepse je definovaná jako život ohrožující orgánová dysfunkce způsobená dysregulovanou odpovědí
Využití molekulárně-biologických postupů a multimarkerových strategií v intenzívní péči Marek Protuš Sepsis - 3 Sepse je definovaná jako život ohrožující orgánová dysfunkce způsobená dysregulovanou odpovědí
Odběry vzorků u pacientů s podezřením na infekci krevního řečiště
 Odběry vzorků u pacientů s podezřením na infekci krevního řečiště Eva Krejčí Centrum klinických laboratoří, Zdravotní ústav se sídlem v Ostravě Lékařská fakulta Ostravské univerzity Infekce krevního řečiště
Odběry vzorků u pacientů s podezřením na infekci krevního řečiště Eva Krejčí Centrum klinických laboratoří, Zdravotní ústav se sídlem v Ostravě Lékařská fakulta Ostravské univerzity Infekce krevního řečiště
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Funkční vzorek 5474/2017
 Funkční vzorek 5474/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u bakteriálních anaerobních patogenů hospodářských zvířat MVDr. Kateřina Nedbalcová, Ph.D. 1 MVDr.
Funkční vzorek 5474/2017 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u bakteriálních anaerobních patogenů hospodářských zvířat MVDr. Kateřina Nedbalcová, Ph.D. 1 MVDr.
5. Bioreaktory. Schematicky jsou jednotlivé typy bioreaktorů znázorněny na obr. 5.1. Nejpoužívanějšími bioreaktory jsou míchací tanky.
 5. Bioreaktory Bioreaktor (fermentor) je nejdůležitější částí výrobní linky biotechnologického procesu. Jde o nádobu různého objemu, ve které probíhá biologický proces. Dochází zde k růstu buněk a tvorbě
5. Bioreaktory Bioreaktor (fermentor) je nejdůležitější částí výrobní linky biotechnologického procesu. Jde o nádobu různého objemu, ve které probíhá biologický proces. Dochází zde k růstu buněk a tvorbě
INTERPRETACE VÝSLEDKŮ CITLIVOSTI NA ANTIBIOTIKA. Milan Kolář Ústav mikrobiologie Fakultní nemocnice a LF UP v Olomouci
 INTERPRETACE VÝSLEDKŮ CITLIVOSTI NA ANTIBIOTIKA Milan Kolář Ústav mikrobiologie Fakultní nemocnice a LF UP v Olomouci Současná medicína je charakteristická svým multidisciplinárním přístupem k řešení mnoha
INTERPRETACE VÝSLEDKŮ CITLIVOSTI NA ANTIBIOTIKA Milan Kolář Ústav mikrobiologie Fakultní nemocnice a LF UP v Olomouci Současná medicína je charakteristická svým multidisciplinárním přístupem k řešení mnoha
Imunologické metody fagocytóza, stanovení oxidačního metabolismu, chemiluminiscence, baktericidní test
 Imunologické metody fagocytóza, stanovení oxidačního metabolismu, chemiluminiscence, baktericidní test Fagocytóza Jeden z evolučně nejstarších obranných mechanismů Součást přirozené imunity Schopnost buněk
Imunologické metody fagocytóza, stanovení oxidačního metabolismu, chemiluminiscence, baktericidní test Fagocytóza Jeden z evolučně nejstarších obranných mechanismů Součást přirozené imunity Schopnost buněk
Nové technologie v mikrobiologické diagnostice a jejich přínos pro pacienty v intenzivní péči
 B I O M E D I C AL Nové technologie v mikrobiologické diagnostice a jejich přínos pro pacienty v intenzivní péči Jaroslav Hrabák CHARLES UNIVERSITY IN PRAGUE Obsah prezentace ČSIM 2016 Mikrobiologická
B I O M E D I C AL Nové technologie v mikrobiologické diagnostice a jejich přínos pro pacienty v intenzivní péči Jaroslav Hrabák CHARLES UNIVERSITY IN PRAGUE Obsah prezentace ČSIM 2016 Mikrobiologická
Funkční vzorek 4595/2018. Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních. látek u Streptococcus suis
 Funkční vzorek 4595/2018 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Streptococcus suis Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
Funkční vzorek 4595/2018 Set ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Streptococcus suis Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního lékařství,
NA ANTIBIOTIKA NA ČOV
 Vodárenská biologie 2018, 6. 2. 2018 DETEKCE GENŮ REZISTENCE NA ANTIBIOTIKA NA ČOV TESTOVÁNÍ METODIKY Dana Vejmelková Kristýna Časarová Eva Proksová Jana Říhová Ambrožová PROBLEMATIKA REZISTENCE NA ANTIBIOTIKA
Vodárenská biologie 2018, 6. 2. 2018 DETEKCE GENŮ REZISTENCE NA ANTIBIOTIKA NA ČOV TESTOVÁNÍ METODIKY Dana Vejmelková Kristýna Časarová Eva Proksová Jana Říhová Ambrožová PROBLEMATIKA REZISTENCE NA ANTIBIOTIKA
Aplikace molekulárně biologických postupů v časné detekci sepse
 Aplikace molekulárně biologických postupů v časné detekci sepse Mgr. Jana Ždychová, Ph.D. IKEM PLM - LLG Sepse je častou příčinou úmrtí během hospitalizace. Včasné nasazení odpovídající ATB terapie je
Aplikace molekulárně biologických postupů v časné detekci sepse Mgr. Jana Ždychová, Ph.D. IKEM PLM - LLG Sepse je častou příčinou úmrtí během hospitalizace. Včasné nasazení odpovídající ATB terapie je
VÝVOJ REZISTENCE BAKTERIÁLNÍCH PATOGENŮ
 LABORATORNÍ LISTY Listopad č.29/2016 2016 Vážené kolegyně a kolegové, v tomto čísle našich Laboratorních listů Vám předkládáme statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme testovali
LABORATORNÍ LISTY Listopad č.29/2016 2016 Vážené kolegyně a kolegové, v tomto čísle našich Laboratorních listů Vám předkládáme statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme testovali
ONLINE BIOSENZORY PŘI HLEDÁNÍ KONTAMINACE PITNÉ VODY
 ONLINE BIOSENZORY PŘI HLEDÁNÍ KONTAMINACE PITNÉ VODY Ing. Jana Zuzáková Ing. Jana Zuzáková, Doc. RNDr. Jana Říhová Ambrožová, PhD., Ing. Dana Vejmelková, PhD., Ing. Roman Effenberg, RNDr. Miroslav Ledvina
ONLINE BIOSENZORY PŘI HLEDÁNÍ KONTAMINACE PITNÉ VODY Ing. Jana Zuzáková Ing. Jana Zuzáková, Doc. RNDr. Jana Říhová Ambrožová, PhD., Ing. Dana Vejmelková, PhD., Ing. Roman Effenberg, RNDr. Miroslav Ledvina
ANTIBIOTICKÉ DISKY Antibiotické disky pro testování citlivosti
 ANTIBIOTICKÉ DISKY Antibiotické disky pro testování citlivosti POPIS ANTIBIOTICKÉ DISKY jsou papírové disky se speciálními vlastnostmi, které jsou impregnované antibiotiky a používají se pro testy citlivosti
ANTIBIOTICKÉ DISKY Antibiotické disky pro testování citlivosti POPIS ANTIBIOTICKÉ DISKY jsou papírové disky se speciálními vlastnostmi, které jsou impregnované antibiotiky a používají se pro testy citlivosti
Mikrobiologická diagnostika infekcí CŽK
 Mikrobiologická diagnostika infekcí CŽK Otázky spojené s diagnostikou katétrových sepsí Filip Růžička Mikrobiologický ústav Lékařské fakulty Masarykovy univerzity a Fakultní nemocnice u sv. Anny, Brno
Mikrobiologická diagnostika infekcí CŽK Otázky spojené s diagnostikou katétrových sepsí Filip Růžička Mikrobiologický ústav Lékařské fakulty Masarykovy univerzity a Fakultní nemocnice u sv. Anny, Brno
Ověřená technologie 4797/2017
 Ověřená technologie 4797/2017 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u bakteriálních anaerobních patogenů hospodářských zvířat Autoři: MVDr. Kateřina Nedbalcová,
Ověřená technologie 4797/2017 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u bakteriálních anaerobních patogenů hospodářských zvířat Autoři: MVDr. Kateřina Nedbalcová,
Odběr a transport biologického materiálu do mikrobiologické laboratoře. Jana Juránková OKM FN Brno
 Odběr a transport biologického materiálu do mikrobiologické laboratoře Jana Juránková OKM FN Brno Specifika odběrů pro bakteriologická vyšetření Správný výsledek mikrobiologického vyšetření začíná správným
Odběr a transport biologického materiálu do mikrobiologické laboratoře Jana Juránková OKM FN Brno Specifika odběrů pro bakteriologická vyšetření Správný výsledek mikrobiologického vyšetření začíná správným
Výskyt multirezistentních bakteriálních kmenů produkujících betalaktamázy
 Výskyt multirezistentních bakteriálních kmenů produkujících betalaktamázy Jaromíra Kratochvílová, Veronika Janásová 19. mezinárodní konference nemocniční epidemiologie a hygiena Mikulov 17. 18. dubna 2012
Výskyt multirezistentních bakteriálních kmenů produkujících betalaktamázy Jaromíra Kratochvílová, Veronika Janásová 19. mezinárodní konference nemocniční epidemiologie a hygiena Mikulov 17. 18. dubna 2012
M. Laichmanová NOVINKY V NABÍDCE KONTROLNÍCH KMENŮ ČESKÉ SBÍRKY MIKROORGANISMŮ
 M. Laichmanová NOVINKY V NABÍDCE KONTROLNÍCH KMENŮ ČESKÉ SBÍRKY MIKROORGANISMŮ držitel certifikátu Kompletní nabídku referenčních mikroorganizmů tvoří 157 kontrolních kmenů bakterií vláknitých hub kvasinek
M. Laichmanová NOVINKY V NABÍDCE KONTROLNÍCH KMENŮ ČESKÉ SBÍRKY MIKROORGANISMŮ držitel certifikátu Kompletní nabídku referenčních mikroorganizmů tvoří 157 kontrolních kmenů bakterií vláknitých hub kvasinek
PREGRADUÁLNÍ VZDĚLÁVÁNÍ V LÉKAŘSKÉ MIKROBIOLOGII
 PREGRADUÁLNÍ VZDĚLÁVÁNÍ V LÉKAŘSKÉ MIKROBIOLOGII Milan Kolář Lékařská fakulta UP v Olomouci ZÁVĚRY Z PŘEDCHÁZEJÍCÍCH SETKÁNÍ Výuka lékařské mikrobiologie patří k nezbytným předpokladům pro výuku klinických
PREGRADUÁLNÍ VZDĚLÁVÁNÍ V LÉKAŘSKÉ MIKROBIOLOGII Milan Kolář Lékařská fakulta UP v Olomouci ZÁVĚRY Z PŘEDCHÁZEJÍCÍCH SETKÁNÍ Výuka lékařské mikrobiologie patří k nezbytným předpokladům pro výuku klinických
Diagnostická laboratoř Praha. Laboratorní příručka Příloha č.1. Přehled vyšetření prováděných v Laboratoři klinické mikrobiologie
 Strana č: 1 Kultivační vyšetření výtěrů z horních cest dýchacích Kultivační vyšetření materiálu z dolních cest dýchacích (mimo TBC) B01/LKM B06/LKM B02/LKM Kultivační semikvantitativní vyšetření moče Typ
Strana č: 1 Kultivační vyšetření výtěrů z horních cest dýchacích Kultivační vyšetření materiálu z dolních cest dýchacích (mimo TBC) B01/LKM B06/LKM B02/LKM Kultivační semikvantitativní vyšetření moče Typ
Přímé stanovení celkového počtu buněk kvasinek pomocí Bürkerovy komůrky Provedení vitálního testu
 Přímé stanovení celkového počtu buněk kvasinek pomocí Bürkerovy komůrky Provedení vitálního testu Otázky k zamyšlení: Bude se jednat o přímé nebo nepřímé stanovení počtu buněk? Stanovujeme počet živých
Přímé stanovení celkového počtu buněk kvasinek pomocí Bürkerovy komůrky Provedení vitálního testu Otázky k zamyšlení: Bude se jednat o přímé nebo nepřímé stanovení počtu buněk? Stanovujeme počet živých
VÝVOJ REZISTENCE BAKTERIÁLNÍCH PATOGENŮ
 LABORATORNÍ LISTY Srpen č.27/2015 2015 Vážené kolegyně a kolegové, v tomto čísle našich Laboratorních listů Vám předkládáme statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme testovali standardní
LABORATORNÍ LISTY Srpen č.27/2015 2015 Vážené kolegyně a kolegové, v tomto čísle našich Laboratorních listů Vám předkládáme statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme testovali standardní
LRR/BUBCV CVIČENÍ Z BUNĚČNÉ BIOLOGIE 3. TESTY ŽIVOTASCHOPNOSTI A POČÍTÁNÍ BUNĚK
 LRR/BUBCV CVIČEÍ Z BUĚČÉ BILGIE 3. TESTY ŽIVTASCHPSTI A PČÍTÁÍ BUĚK TERETICKÝ ÚVD: Při práci s buňkami je jedním ze základních sledovaných parametrů stanovení jejich životaschopnosti (viability). Tímto
LRR/BUBCV CVIČEÍ Z BUĚČÉ BILGIE 3. TESTY ŽIVTASCHPSTI A PČÍTÁÍ BUĚK TERETICKÝ ÚVD: Při práci s buňkami je jedním ze základních sledovaných parametrů stanovení jejich životaschopnosti (viability). Tímto
Téma: Testy životaschopnosti a Počítání buněk
 LRR/BUBV vičení z buněčné biologie Úloha č. 3 Téma: Testy životaschopnosti a Počítání Úvod: Při práci s buňkami je jedním ze základních sledovaných parametrů stanovení jejich životaschopnosti (viability).
LRR/BUBV vičení z buněčné biologie Úloha č. 3 Téma: Testy životaschopnosti a Počítání Úvod: Při práci s buňkami je jedním ze základních sledovaných parametrů stanovení jejich životaschopnosti (viability).
Laboratorní testování na přítomnost koliformních bakterií, psychrotrofních a termorezistentních mikroorganismů a sporotvorných anaerobních bakterií
 Laboratorní testování na přítomnost koliformních bakterií, psychrotrofních a termorezistentních mikroorganismů a sporotvorných anaerobních bakterií Ing. Pavel Kopunecz, ČMSCH a.s. Přehled metod hodnocení
Laboratorní testování na přítomnost koliformních bakterií, psychrotrofních a termorezistentních mikroorganismů a sporotvorných anaerobních bakterií Ing. Pavel Kopunecz, ČMSCH a.s. Přehled metod hodnocení
CITLIVOST A REZISTENCE MIKROORGANISMŮ K ANTIBIOTIKŮM
 CITLIVOST A REZISTENCE MIKROORGANISMŮ K ANTIBIOTIKŮM Mgr. Sylva Janovská, Ph.D. Mgr. Eva Slehová Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný
CITLIVOST A REZISTENCE MIKROORGANISMŮ K ANTIBIOTIKŮM Mgr. Sylva Janovská, Ph.D. Mgr. Eva Slehová Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný
Molekulárně biologické metody v mikrobiologii. Mgr. Martina Sittová Jaro 2014
 Molekulárně biologické metody v mikrobiologii Mgr. Martina Sittová Jaro 2014 Harmonogram 1. den Izolace DNA 2. den Měření koncentrace DNA spektrofotometricky, real-time PCR 3. den Elektroforéza Molekulární
Molekulárně biologické metody v mikrobiologii Mgr. Martina Sittová Jaro 2014 Harmonogram 1. den Izolace DNA 2. den Měření koncentrace DNA spektrofotometricky, real-time PCR 3. den Elektroforéza Molekulární
Tvorba biofilmu potravinářsky významnými bakteriemi. Martina Hrubanová
 Tvorba biofilmu potravinářsky významnými bakteriemi Martina Hrubanová Bakalářská práce 2011 UTB ve Zlíně, Fakulta technologická 2 UTB ve Zlíně, Fakulta technologická 3 UTB ve Zlíně, Fakulta technologická
Tvorba biofilmu potravinářsky významnými bakteriemi Martina Hrubanová Bakalářská práce 2011 UTB ve Zlíně, Fakulta technologická 2 UTB ve Zlíně, Fakulta technologická 3 UTB ve Zlíně, Fakulta technologická
Lékařská mikrobiologie II
 Publikováno z 2. lékařská fakulta Univerzity Karlovy (https://www.lf2.cuni.cz) LF2 > Lékařská mikrobiologie II Lékařská mikrobiologie II Napsal uživatel Marie Havlová dne 8. Březen 2013-0:00. Sylabus praktických
Publikováno z 2. lékařská fakulta Univerzity Karlovy (https://www.lf2.cuni.cz) LF2 > Lékařská mikrobiologie II Lékařská mikrobiologie II Napsal uživatel Marie Havlová dne 8. Březen 2013-0:00. Sylabus praktických
Člověk a mikroby, jsme nyní odolnější? Jan Krejsek. Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové
 Člověk a mikroby, jsme nyní odolnější? Jan Krejsek Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové Jsme určeni genetickou dispozicí a životními podmínkami, které působí epigeneticky
Člověk a mikroby, jsme nyní odolnější? Jan Krejsek Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové Jsme určeni genetickou dispozicí a životními podmínkami, které působí epigeneticky
EHK 544 Bakteriologická diagnostika-vyhodnocení
 Zprávy CEM (SZÚ, Praha) 2007; 16(12): 555 557. EHK 544 Bakteriologická diagnostika-vyhodnocení Helena Žemličková, Marina Maixnerová, Jana Zavadilová, Petr Petráš, Pavla Urbášková CELKOVÉ HODNOCENÍ IDENTIFIKACÍ
Zprávy CEM (SZÚ, Praha) 2007; 16(12): 555 557. EHK 544 Bakteriologická diagnostika-vyhodnocení Helena Žemličková, Marina Maixnerová, Jana Zavadilová, Petr Petráš, Pavla Urbášková CELKOVÉ HODNOCENÍ IDENTIFIKACÍ
Kultivační metody stanovení mikroorganismů
 Kultivační metody stanovení mikroorganismů Základní rozdělení půd Syntetická, definovaná media, jednoduché sloučeniny, známé sloţení Komplexní media, vycházejí z ţivočišných nebo rostlinných tkání a pletiv,
Kultivační metody stanovení mikroorganismů Základní rozdělení půd Syntetická, definovaná media, jednoduché sloučeniny, známé sloţení Komplexní media, vycházejí z ţivočišných nebo rostlinných tkání a pletiv,
MIKROSKOPIE JAKO NÁSTROJ STUDIA MIKROORGANISMŮ
 Mikroskopické techniky MIKROSKOPIE JAKO NÁSTROJ STUDIA MIKROORGANISMŮ Slouží k vizualizaci mikroorganismů Antoni van Leeuwenhoek (1632-1723) Čočka zvětšující 300x Různé druhy mikroskopů, které se liší
Mikroskopické techniky MIKROSKOPIE JAKO NÁSTROJ STUDIA MIKROORGANISMŮ Slouží k vizualizaci mikroorganismů Antoni van Leeuwenhoek (1632-1723) Čočka zvětšující 300x Různé druhy mikroskopů, které se liší
Optimalizace metody PCR pro její využití na vzorky KONTAMINOVANÝCH PITNÝCH VOD
 Optimalizace metody PCR pro její využití na vzorky KONTAMINOVANÝCH PITNÝCH VOD Dana Vejmelková, Milan Šída, Kateřina Jarošová, Jana Říhová Ambrožová VODÁRENSKÁ BIOLOGIE, 1. 2. 2017 ÚVOD Sledované parametry,
Optimalizace metody PCR pro její využití na vzorky KONTAMINOVANÝCH PITNÝCH VOD Dana Vejmelková, Milan Šída, Kateřina Jarošová, Jana Říhová Ambrožová VODÁRENSKÁ BIOLOGIE, 1. 2. 2017 ÚVOD Sledované parametry,
ANTIBIOTICKÁ LÉČBA OFF-LABEL Z POHLEDU MIKROBIOLOGA. Milan Kolář Ústav mikrobiologie FNOL a LF UP v Olomouci
 ANTIBIOTICKÁ LÉČBA OFF-LABEL Z POHLEDU MIKROBIOLOGA Milan Kolář Ústav mikrobiologie FNOL a LF UP v Olomouci Septický pacient s nozokomiální pneumonií Endosekret: KLPN + ESCO Terapie: PPT + GEN INTERPRETACE?
ANTIBIOTICKÁ LÉČBA OFF-LABEL Z POHLEDU MIKROBIOLOGA Milan Kolář Ústav mikrobiologie FNOL a LF UP v Olomouci Septický pacient s nozokomiální pneumonií Endosekret: KLPN + ESCO Terapie: PPT + GEN INTERPRETACE?
Ukázky: CCM katalog Anaerostat + generátor anaerobní atmosféry Plastové kličky Termostat se třepačkou. Očkovánía kultivace
 Ukázky: CCM katalog Anaerostat + generátor anaerobní atmosféry Plastové kličky Termostat se třepačkou Očkovánía kultivace Bakteriálnídruh = jasněvymezenáskupina navzájem příbuzných kmenů, zahrnujících
Ukázky: CCM katalog Anaerostat + generátor anaerobní atmosféry Plastové kličky Termostat se třepačkou Očkovánía kultivace Bakteriálnídruh = jasněvymezenáskupina navzájem příbuzných kmenů, zahrnujících
Fakultní nemocnice Brno Laboratoře Oddělení klinické mikrobiologie Jihlavská 20, Brno
 Vyšetření: 802 - Lékařská mikrobiologie, 804 Lékařská parazitologie, 805 Lékařská virologie, 822 Lékařská mykologie 1. Vyšetření mikroorganismů z klinického materiálu a z kultur mikroskopicky - barvený
Vyšetření: 802 - Lékařská mikrobiologie, 804 Lékařská parazitologie, 805 Lékařská virologie, 822 Lékařská mykologie 1. Vyšetření mikroorganismů z klinického materiálu a z kultur mikroskopicky - barvený
Stav rezistence bakteriálních patogenů v Karlovarském kraji.
 Stav rezistence bakteriálních patogenů v Karlovarském kraji. Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní diskovou
Stav rezistence bakteriálních patogenů v Karlovarském kraji. Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní diskovou
SeptiFast. rychlá detekce sepse. Olga Bálková Roche s.r.o., Diagnostics Division
 SeptiFast rychlá detekce sepse Olga Bálková Roche s.r.o., Diagnostics Division Požadavky kliniků Rychlá detekce a identifikace patogenů způsobujících sepsi Rychlé výsledky Výsledky do několika hodin Jednoznačné
SeptiFast rychlá detekce sepse Olga Bálková Roche s.r.o., Diagnostics Division Požadavky kliniků Rychlá detekce a identifikace patogenů způsobujících sepsi Rychlé výsledky Výsledky do několika hodin Jednoznačné
Stav rezistence bakteriálních patogenů v Karlovarském kraji.
 Stav rezistence bakteriálních patogenů v Karlovarském kraji. Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní diskovou
Stav rezistence bakteriálních patogenů v Karlovarském kraji. Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní diskovou
LABORATOŘ MIKROBIOLOGIE II ROD STREPTOCOCCUS ROD ENTEROCOCCUS
 LABORATOŘ MIKROBIOLOGIE II ROD STREPTOCOCCUS ROD ENTEROCOCCUS Petra Lysková STREPTOCOCCUS PYOGENES antigenní struktura a faktory virulence vázané na buňku [1] STREPTOCOCCUS PYOGENES mimobuněčné produkty
LABORATOŘ MIKROBIOLOGIE II ROD STREPTOCOCCUS ROD ENTEROCOCCUS Petra Lysková STREPTOCOCCUS PYOGENES antigenní struktura a faktory virulence vázané na buňku [1] STREPTOCOCCUS PYOGENES mimobuněčné produkty
Protokol č. 7 Pozorování živých a mrtvých buněk kvasinek Vitální test
 Protokol č. 7 Pozorování živých a mrtvých buněk kvasinek Vitální test Cíl cvičení: Bude se jednat o přímé nebo nepřímé stanovení počtu buněk? Stanovujeme počet živých nebo mrtvých buněk? Jak odlišíme živé
Protokol č. 7 Pozorování živých a mrtvých buněk kvasinek Vitální test Cíl cvičení: Bude se jednat o přímé nebo nepřímé stanovení počtu buněk? Stanovujeme počet živých nebo mrtvých buněk? Jak odlišíme živé
ROZDĚLENÍ ANTIMIKROBIÁLNÍCH LÁTEK, VYŠETŘOVÁNÍ CITLIVOSTI. M.Hanslianová Antibiotické středisko OKM FN Brno
 ROZDĚLENÍ ANTIMIKROBIÁLNÍCH LÁTEK, VYŠETŘOVÁNÍ CITLIVOSTI M.Hanslianová Antibiotické středisko OKM FN Brno Antimikrobiální látky I. Antibiotika= léčiva používaná k profylaxi a terapii infekčních onemocnění
ROZDĚLENÍ ANTIMIKROBIÁLNÍCH LÁTEK, VYŠETŘOVÁNÍ CITLIVOSTI M.Hanslianová Antibiotické středisko OKM FN Brno Antimikrobiální látky I. Antibiotika= léčiva používaná k profylaxi a terapii infekčních onemocnění
Způsoby potlačení tvorby biofilmů event. jejich degradace. Doležalová Fehérová 2015/2016
 Způsoby potlačení tvorby biofilmů event. jejich degradace Doležalová Fehérová 2015/2016 Biofilmy Přisedlé společenstvo mikroorganismů Tvořeny buňkami produkujícími extracelulární polymerní látky Extracelulární
Způsoby potlačení tvorby biofilmů event. jejich degradace Doležalová Fehérová 2015/2016 Biofilmy Přisedlé společenstvo mikroorganismů Tvořeny buňkami produkujícími extracelulární polymerní látky Extracelulární
Jednotné pracovní postupy zkoušení krmiv STANOVENÍ OBSAHU KVASINEK RODU SACCHAROMYCES
 Národní referenční laboratoř Strana 1 STANOVENÍ OBSAHU KVASINEK RODU SACCHAROMYCES 1 Rozsah a účel Metodika slouží ke stanovení počtu probiotických kvasinek v doplňkových látkách, premixech a krmivech.
Národní referenční laboratoř Strana 1 STANOVENÍ OBSAHU KVASINEK RODU SACCHAROMYCES 1 Rozsah a účel Metodika slouží ke stanovení počtu probiotických kvasinek v doplňkových látkách, premixech a krmivech.
LABORATORNÍ LISTY Vážené kolegyně a kolegové,
 LABORATORNÍ LISTY Červen Vážené kolegyně a kolegové, VÝVOJ REZISTENCE BAKTERIÁLNÍCH PATOGENŮ Horní cesty dýchací Streptococcus pyogenes č. 19/2018 2018 v tomto čísle našich laboratorních listů Vám předkládáme
LABORATORNÍ LISTY Červen Vážené kolegyně a kolegové, VÝVOJ REZISTENCE BAKTERIÁLNÍCH PATOGENŮ Horní cesty dýchací Streptococcus pyogenes č. 19/2018 2018 v tomto čísle našich laboratorních listů Vám předkládáme
VYUŽITÍ SCREENINGOVÝCH MIKROBIOLOGICKÝCH TESTERŮ HACH LANGE
 VYUŽITÍ SCREENINGOVÝCH MIKROBIOLOGICKÝCH TESTERŮ HACH LANGE Při své práci se pravidelně setkávám s produkty společnosti HACH LANGE s.r.o. Mikrobiologické testy pro stanovení přítomnosti či nepřítomnosti
VYUŽITÍ SCREENINGOVÝCH MIKROBIOLOGICKÝCH TESTERŮ HACH LANGE Při své práci se pravidelně setkávám s produkty společnosti HACH LANGE s.r.o. Mikrobiologické testy pro stanovení přítomnosti či nepřítomnosti
Návod k odečítání. Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti. Verze 3.0 Duben 2013
 Návod k odečítání Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti Verze 3.0 Duben 2013 2 Modifikace obrázkového návodu k odečítání EUCAST Verze Verze 3.0 Duben 2013 Verze 2.0 Květen
Návod k odečítání Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti Verze 3.0 Duben 2013 2 Modifikace obrázkového návodu k odečítání EUCAST Verze Verze 3.0 Duben 2013 Verze 2.0 Květen
Interpretace výsledků bakteriologických vyšetření
 Interpretace výsledků bakteriologických vyšetření Veškeré nálezy vyšetření je nutno hodnotit vzhledem k diagnóze, věku, zánětlivým parametrům, klinickému stavu pacienta, ev. k dalším důležitým anamnestickým
Interpretace výsledků bakteriologických vyšetření Veškeré nálezy vyšetření je nutno hodnotit vzhledem k diagnóze, věku, zánětlivým parametrům, klinickému stavu pacienta, ev. k dalším důležitým anamnestickým
Odbornost pro revize COPAL
 Odbornost pro revize COPAL ODBORNOST. ZÁVAZEK. ODPOVĚDNOST. Kompetence v revizní artroplastice Účinně kombinovat technické znalosti, progresivní a nekonvenční myšlení to jsou zásady, podle nichž společnost
Odbornost pro revize COPAL ODBORNOST. ZÁVAZEK. ODPOVĚDNOST. Kompetence v revizní artroplastice Účinně kombinovat technické znalosti, progresivní a nekonvenční myšlení to jsou zásady, podle nichž společnost
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 KLINICKÁ MIKROBIOLOGIE Klinická mikrobiologie je medicínský obor - pojítko mezi
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 KLINICKÁ MIKROBIOLOGIE Klinická mikrobiologie je medicínský obor - pojítko mezi
Izolace nukleových kyselin
 Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Hlavní mechanismy rezistence. In vitro testování citlivosti a rezistence. Základy antimikrobiální terapie VSAT081 Týden 4 Ondřej Zahradníček
 Hlavní mechanismy rezistence. In vitro testování citlivosti a rezistence. Základy antimikrobiální terapie VSAT081 Týden 4 Ondřej Zahradníček Rezistence mikrobů na antimikrobiální látky Primární rezistence:
Hlavní mechanismy rezistence. In vitro testování citlivosti a rezistence. Základy antimikrobiální terapie VSAT081 Týden 4 Ondřej Zahradníček Rezistence mikrobů na antimikrobiální látky Primární rezistence:
Látka toxická pro mikroorganismy a vyšší živočichy i v nízké koncentraci. Do prostředí se dostává: Používá se například:
 Látka toxická pro mikroorganismy a vyšší živočichy i v nízké koncentraci. Do prostředí se dostává: při rozkladu organických zbytků lesních požárech většina má průmyslový původ Používá se například: při
Látka toxická pro mikroorganismy a vyšší živočichy i v nízké koncentraci. Do prostředí se dostává: při rozkladu organických zbytků lesních požárech většina má průmyslový původ Používá se například: při
LABORATORNÍ STUDIE ANTIMIKROBNÍ AKTIVITY CHLOROVÝCH PŘÍPRAVKŮ NA BÁZI DICHLORIZOKYANURANU SODNÉHO
 LABORATORNÍ STUDIE ANTIMIKROBNÍ AKTIVITY CHLOROVÝCH PŘÍPRAVKŮ NA BÁZI DICHLORIZOKYANURANU SODNÉHO Doc. Ing. Marie Hartmanová, CSc. Vojenská lékařská akademie JEP, Hradec Králové, Česká republika Mezi klasické,
LABORATORNÍ STUDIE ANTIMIKROBNÍ AKTIVITY CHLOROVÝCH PŘÍPRAVKŮ NA BÁZI DICHLORIZOKYANURANU SODNÉHO Doc. Ing. Marie Hartmanová, CSc. Vojenská lékařská akademie JEP, Hradec Králové, Česká republika Mezi klasické,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ELEKTROMIGRAČNÍ METODY
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ELEKTROMIGRAČNÍ METODY ELEKTROFORÉZA K čemu to je? kritérium čistoty preparátu stanovení molekulové hmotnosti makromolekul stanovení izoelektrického
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ELEKTROMIGRAČNÍ METODY ELEKTROFORÉZA K čemu to je? kritérium čistoty preparátu stanovení molekulové hmotnosti makromolekul stanovení izoelektrického
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
VYUŽITÍ PRŮTOKOVÉ CYTOMETRIE PRO DETEKCI ÚČINNOSTI FILTRACE BAKTERIÍ V PROCESECH ČIŠTĚNÍ ODPADNÍCH VOD
 VYUŽITÍ PRŮTOKOVÉ CYTOMETRIE PRO DETEKCI ÚČINNOSTI FILTRACE BAKTERIÍ V PROCESECH ČIŠTĚNÍ ODPADNÍCH VOD P. Mikula a*, J. Lev b,c, L. Kalhotka b, M. Holba a,c, D. Kimmer d, B. Maršálek a, M. Vítězová b a)
VYUŽITÍ PRŮTOKOVÉ CYTOMETRIE PRO DETEKCI ÚČINNOSTI FILTRACE BAKTERIÍ V PROCESECH ČIŠTĚNÍ ODPADNÍCH VOD P. Mikula a*, J. Lev b,c, L. Kalhotka b, M. Holba a,c, D. Kimmer d, B. Maršálek a, M. Vítězová b a)
Ověřená technologie 4903/2018
 Ověřená technologie 4903/2018 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Streptococcus suis Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Ověřená technologie 4903/2018 Výroba setu ke stanovení minimálních inhibičních koncentrací antimikrobiálních látek u Streptococcus suis Autoři: MVDr. Kateřina Nedbalcová, Ph.D., Výzkumný ústav veterinárního
Návod k odečítání Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti
 Návod k odečítání Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti Verze 5.0 Leden 2017 Změny proti předchozí verzi (v 4.0) Obrázek Změna 3 17 Objasnění týkající se odečítání zón a
Návod k odečítání Disková difuzní metoda EUCAST pro vyšetřování antibiotické citlivosti Verze 5.0 Leden 2017 Změny proti předchozí verzi (v 4.0) Obrázek Změna 3 17 Objasnění týkající se odečítání zón a
Antibiotická profylaxe v gynekologické operativě
 Antibiotická profylaxe v gynekologické operativě J. MAŠATA Gynekologicko porodnická klinika VFN a 1. LF UK, Praha Definice infekce Kolonizace (colonisation) běžná přítomnost bakterie Kontaminace (contamination)
Antibiotická profylaxe v gynekologické operativě J. MAŠATA Gynekologicko porodnická klinika VFN a 1. LF UK, Praha Definice infekce Kolonizace (colonisation) běžná přítomnost bakterie Kontaminace (contamination)
analýza dat a interpretace výsledků
 Genetická transformace bakterií III analýza dat a interpretace výsledků Předmět: Biologie ŠVP: Prokaryotní organismy, genetika Doporučený věk žáků: 16-18 let Doba trvání: 45 minut Specifické cíle: analyzovat
Genetická transformace bakterií III analýza dat a interpretace výsledků Předmět: Biologie ŠVP: Prokaryotní organismy, genetika Doporučený věk žáků: 16-18 let Doba trvání: 45 minut Specifické cíle: analyzovat
prokaryotní Znaky prokaryoty
 prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
Moderní metody stanovení mikroorganismů
 Moderní metody stanovení mikroorganismů Mgr. Martin Polách NOACK ČR, spol. s r.o. Legislativní požadavky EU na mikrobiologické parametry potravin Závazným předpisem je nařízení 1441 / 2007 / ES, pozměňující
Moderní metody stanovení mikroorganismů Mgr. Martin Polách NOACK ČR, spol. s r.o. Legislativní požadavky EU na mikrobiologické parametry potravin Závazným předpisem je nařízení 1441 / 2007 / ES, pozměňující
European Committee on Antimicrobial Susceptibility Testing
 European Committee on Antimicrobial Susceptibility Testing Breakpointy průměrů zón pro rychlé vyšetření antimikrobní citlivosti přímo z hemokultivačních lahviček (RAST) Verze 1.0, platná od 28-11-2018
European Committee on Antimicrobial Susceptibility Testing Breakpointy průměrů zón pro rychlé vyšetření antimikrobní citlivosti přímo z hemokultivačních lahviček (RAST) Verze 1.0, platná od 28-11-2018
ANTIBIOTICKÉ LÉČBY V INTENZIVNÍ MEDICÍNĚ
 PRINCIPY ANTIBIOTICKÉ LÉČBY V INTENZIVNÍ MEDICÍNĚ Milan Kolář Ústav mikrobiologie Fakultní nemocnice Olomouc Lékařská fakulta Univerzity Palackého v Olomouci Bakteriální infekce byly, jsou a s velkou pravděpodobností
PRINCIPY ANTIBIOTICKÉ LÉČBY V INTENZIVNÍ MEDICÍNĚ Milan Kolář Ústav mikrobiologie Fakultní nemocnice Olomouc Lékařská fakulta Univerzity Palackého v Olomouci Bakteriální infekce byly, jsou a s velkou pravděpodobností
Automatizace v klinické mikrobiologii
 Automatizace v klinické mikrobiologii 1Jaroslav Hrabák, 2Pavel Dřevínek 1Ústav mikrobiologie, Biomedicínské centrum, Lékařská fakulta a Fakultní nemocnice v Plzni, Univerzita Karlova 2Ústav lékařské mikrobiologie,
Automatizace v klinické mikrobiologii 1Jaroslav Hrabák, 2Pavel Dřevínek 1Ústav mikrobiologie, Biomedicínské centrum, Lékařská fakulta a Fakultní nemocnice v Plzni, Univerzita Karlova 2Ústav lékařské mikrobiologie,
Stav rezistence bakteriálních patogenů v Karlovarském kraji za rok 2009
 Stav rezistence bakteriálních patogenů v Karlovarském kraji za rok 9 Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní
Stav rezistence bakteriálních patogenů v Karlovarském kraji za rok 9 Vážení kolegové, předkládáme vám k prostudování statistiku rezistencí hlavních bakteriálních patogenů. Kmeny jsme otestovali standardní
POH O L H E L D E U D U M
 SEPSE Z POHLEDU MIKROBIOLOGA Milan Kolář Ústav mikrobiologie FNOL a LF UP v Olomouci K významným problémům současné medicíny patří bezesporu septické stavy z důvodu vysoké morbidity, mortality a současně
SEPSE Z POHLEDU MIKROBIOLOGA Milan Kolář Ústav mikrobiologie FNOL a LF UP v Olomouci K významným problémům současné medicíny patří bezesporu septické stavy z důvodu vysoké morbidity, mortality a současně
Perorální bakteriální. u alergických pacientů. Jaroslav Bystroň Ingrid Richterová
 Perorální bakteriální imunomodulátory u alergických pacientů Jaroslav Bystroň Ingrid Richterová Imunitní systém Fyziologické obranné bariéry Nemají imunologickou povahu První etáží imunitní reakce jsou:
Perorální bakteriální imunomodulátory u alergických pacientů Jaroslav Bystroň Ingrid Richterová Imunitní systém Fyziologické obranné bariéry Nemají imunologickou povahu První etáží imunitní reakce jsou:
Poslední trendy krevních odběrů. Mgr. Tomáš Grim Mgr. Zuzana Kučerová
 Poslední trendy krevních odběrů Mgr. Tomáš Grim Mgr. Zuzana Kučerová Laboratorní metody Účel: Diagnostický Sledování průběhu nemoci Monitorování léčby Určování prognózy onemocnění Preventivní či screeningové
Poslední trendy krevních odběrů Mgr. Tomáš Grim Mgr. Zuzana Kučerová Laboratorní metody Účel: Diagnostický Sledování průběhu nemoci Monitorování léčby Určování prognózy onemocnění Preventivní či screeningové
ZDRAVOTNÍ NEZÁVADNOST
 ZDRAVOTNÍ NEZÁVADNOST POTRAVIN Tvorba biofilmů mikroorganismy problémy v potravinářských technologiích Michaela Vintrová Lucie Tomešová OBSAH Co je to biofilm? Složení biofilmu Bakteriální biofilmy Tvorba
ZDRAVOTNÍ NEZÁVADNOST POTRAVIN Tvorba biofilmů mikroorganismy problémy v potravinářských technologiích Michaela Vintrová Lucie Tomešová OBSAH Co je to biofilm? Složení biofilmu Bakteriální biofilmy Tvorba
1. Definice a historie oboru molekulární medicína. 3. Základní laboratorní techniky v molekulární medicíně
 Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.