3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
|
|
|
- Alžběta Urbanová
- před 8 lety
- Počet zobrazení:
Transkript
1 p
2 p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle mtdna a chromozomu Y 7) Další analýzy a co z toho vyplývá 8) Haploskupiny mtdna a chromozomu Y v ČR
3 Jak to, že máme společného předka? Hledání našeho společného předka každý máme dva společné předky v předchozí generaci = rodiče v další generaci 4 (babičky a dědečky, prarodiče) a v další již 8 (prababičky a pradědečky, praprarodiče), atd. počet předků v generaci n je tedy 2 n pokud bychom sledovali například 40. generaci (asi 1000 let zpátky), pak by předků bylo v této 40. generaci 2 40 = 1,09 x tolik lidí však ještě na planetě Zemi ani nežilo (dnes žije asi 7,0 x 10 9 )
4 ve skutečnosti, čím více jdeme zpátky, tím je pravděpodobnější, že nějací dva mí předci si byli příbuzní a tím se počet předků snižuje, např. místo 2 4 = 16, je to jen 14 čím jdeme více do historie, tím častěji budeme narážet na příbuzenská propojení a počet předků se tak bude dál a dál výrazněji snižovat
5 ovšem hledat příbuznost studiem všech předků v rodokmenu např. studiem jaderné DNA by bylo velmi složité každý jedinec dostane polovinu genetické informace od své matky a otce a stejně tak oni mají polovinu od své matky a otce atd.
6 proto k těmto studiím s výhodou využíváme mtdna, kterou každý jedinec získává jen od matky (modrá kolečka) a nebo chromozom Y, který dědí pouze synové od svých otců (žluté čtverečky) počet takto studovaných příbuzných se nám výrazně zmenší
7 Andy a Alice tak mají, co se týče mtdna, pouze jediného předka neboli jejich společným předkem je Diana, žijící před třemi generacemi obdobná situace u rozsáhlejšího rodokmene méně příbuzných jedinců
8 Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle mtdna a chromozomu Y 7) Další analýzy a co z toho vyplývá 8) Haploskupiny mtdna a chromozomu Y v ČR
9 Mitochondriální DNA (mtdna) člověka součástí mitochondrií kruhová molekula DNA o průměrné velikosti pb, dnes známe kompletní sekvenci 5-10 molekul v jedné organele, až mitochondrií v buňce 37 různých genů, dědí se jako celek neprobíhá rekombinace, mění se pouze procesem mutace vysoká mutační rychlost (až 10násobná oproti jaderné DNA) maternální dědičnost
10 maternální dědičnost výrazně zmenší počet studovaných příbuzných Hledání našeho společného předka
11 Andy a Alice, mají pouze jediného předka neboli jejich společným předkem je Diana, žijící před třemi generacemi v další generaci se mtdna tohoto společného předka bude předávat už pouze v rodokmenu Alice, protože Andy jako muž mtdna do dalších generací nepředává stejně tak se přenos zastaví, pokud Alice bude mít pouze syny
12 vzhledem k tomuto postupnému zanikání a předávání mtdna pouze přes ženskou linii, jsme nakonec schopni v daleké minulosti dospět až k jediné ženě společnému předku, jejíž mtdna se přenášela nejdéle pramáti Eva některé mtdna linie tak zanikají a jiné se přenáší dál a pomnožují díky tomu při pohledu ze současnosti do minulosti sledujeme tzv. koalescenci = = splývání linií mtdna systém zanikání a splývání mtdna linií ukazuje následující obrázek
13 současná generace 2 děti 25 % 1D+1C 25 % 2D 25 % 2C 25 % 1D+1C pravděpodobnost simulaci začínáme někde v minulosti se čtyřmi ženami Amandou, Alexis, Anne a Alicí každé se narodí dvě děti (C-chlapec, D-děvče) mtdna Ann zaniká má pouze chlapce u ostatních žen se přenáší do další generace, protože mají alespoň jedno děvče
14 současná generace v každé generaci se pak situace opakuje po šesti generacích vidíme, že se zachovala mtdna pouze Alice, jejíž potomci měli v každé generaci alespoň jednu dceru u Grace, Giny, Gabriely a Gertrudy nalezneme stejnou mtdna jakou měla Alice
15 současná generace zachovala se pouze mtdna Alice, přičemž pomocí mtdna můžeme zjistit, že nejbližším společným předkem Grace, Giny, Gabriely a Gertrudy byla Dawn (MRCA most recent common ancestor)
16 avšak to, že mají Grace, Gina, Gabriela a Gertruda stejnou mtdna jako Dawn nebo Alice není ve skutečnosti úplně pravda (ani být nemůže, protože pokud by přežila do současnosti mtdna jediné ženy, pak bychom dle tohoto postulátu měli všichni stejnou mtdna) ve skutečnosti se jejich mtdna navzájem liší např. mtdna Gertrudy a Gabriely jsou si podobnější než je jejich mtdna podobná k mtdna Grace to, čím se mtdna těchto žen liší, jsou náhodné mutace, které v ní vznikly jakmile se tedy objeví v mtdna nějaká mutace, tato je pak dále předávána v dané linii čím je tato linie starší, tím více mutací se v ní nahromadí
17 Kde v mtdna mutace hledáme: kódující region x kontrolní region HVR (regulace replikace mtdna, HVSI a HVSII) kódující region lze analyzovat pomocí RFLP, specifickou sadou restrikčních enzymů kontrolní region variabilita studována přímým sekvencováním
18 Co v mtdna zkoumáme: některé mutace se předávají na potomky vždy spolu = haplotyp = určují tedy dědičnou linii některé haplotypy jsou si podobnější (sdílejí stejné mutace) a ukazují na společného příbuzného předka tyto haplotypy se sdružují do tzv. haploskupin
19 Haplotypy x haploskupiny Hledání našeho společného předka Lze si to přiblížit na analogii se značkou aut: na světě existuje mnoho automobilek Škoda, Ford, Honda apod. každá vyrábí mnoho různých modelů např. Škodovka Fabia, Octavia; Honda Civic, Acord = tyto modely představují jednotlivé haplotypy některá tato auta různých značek jsou vybavena například klimatizací nebo GPS navigací (nesou podobné mutace) jsou si tedy v tomto podobnější a tvoří haploskupiny (např. malé vozy x limuzíny)
20 Haplotypy x haploskupiny Hledání našeho společného předka větší příbuznost menší příbuznost Haplotyp sestava mutací podobnější haplotypy sdílejí příbuznější jedinci Haploskupina příbuzné haplotypy sdílejí stejné nebo podobné sestavy mutací tvoří haploskupinu mají svůj geografický původ (vzhledem k původnímu předkovi)
21
22 Haplotypy x haploskupiny Hledání našeho společného předka na základě haplotypů a haploskupin jsme schopni určit, ke které linii daní jedinci náleží (hledáme příbuznost na základě rozdílů a podobností) = identifikace společného předka již víme - selekčně neutrální mutace v čase se hromadí konstantní rychlostí = molekulární hodiny podle počtu těchto mutací lze také odvodit stáří sledované linie, předka
23 Problémy: stanovit podobnost (příbuznost) mezi jedinci na základě rozdílů v sekvenci nemusí být vždy úplně jednoduché (viz následující příklad) ukážeme si na příkladu tří žen Alice, Betty a Cindy, u nichž známe část nějaké sekvence mtdna a víme, že se liší mezi sebou v jednom nukleotidu Alice (A) Betty (G) Cindy (A) existují tři možnosti (vývojové stromy), které ukazují, jak k těmto odlišnostem (mutaci) mohlo dojít
24 Varianta 1 Alice a Betty jsou si podobnější (příbuznější) než je každá z nich k Cindy
25 Varianta 2 Alice a Cindy jsou si podobnější (příbuznější) než je každá z nich k Betty
26 Varianta 3 Betty a Cindy jsou si podobnější (příbuznější) než je každá z nich k Alici
27 G Který z těchto stromů je nejpravděpodobněji správný? Alice i Cindy nesou obě stejný nukleotid A, zatímco Betty má ve stejném místě G první možností je, že v minulosti došlo k jedné mutační události, kdy se pravděpodobně A zaměnil za G nebo naopak G za A druhou možností je, že již oba nukleotidy jsou mutací nějakého jiného nukleotidu = dvě mutační události Ockhamova břitva budeme uvažovat jednu mutační událost
28 Který z těchto stromů je nejpravděpodobněji správný? abychom mohli konstruovat strom, musíme vědět, kdo byl společným předkem všech tří lidí, tak budeme vědět, který nukleotid je původní a který je mutovanou formou jak to můžeme zjistit nejvhodnější metodou je sekvencování nějaké jiné blízce příbuzné skupiny jedinců ( outgroup ) pro člověka je např. nejvhodnější šimpanz hypotetické srovnání se šimpanzí DNA nám odhalí, že původní je báze G v našich třech uvažovaných stromech tedy máme nastaveno, že společný předek všech tří žen měl ve své sekvenci G
29 Který z těchto stromů je nejpravděpodobněji správný? první strom Alice a Betty mají společného předka, který se odštěpil ze společného předka se Cindy víme, že původní byl v sekvenci G = muselo zde dojít ke dvěma odděleným mutačním událostem v obou případech mutoval G na A pro vysvětlení druhého stromu, kde mají Alice a Cindy společného předka, který vzniknul ze společného předka s Betty, nám postačí jediná mutační událost, která nastala u společného předka Alice a Cindy u třetího stromu opět potřebujeme dvě mutační události Ockhamova břitva nám říká, že nejpravděpodobnější je to jednodušší, tedy strom č. 2
30 takto však ve skutečnosti nepostupujeme, změna jediné báze v jedné sekvenci je málo informativní vycházíme z mnoha změn na různých místech a také u většího vzorku jedinců například pokud bychom analyzovali sekvenční rozdíl v jiné sekvenci, mohl by jako pravděpodobnější vyjít např. strom č. 1 pokud však budeme uvažovat více sekvencí, pak nám ale geometrickou řadou narůstá počet možných stromů, s nimiž musíme počítat celková analýza je velmi náročná a složitá např. máme 3 jedince a 3 odlišné sekvence (každá sekvence od jednoho jedince), pak jsou 3 možné stromy, ale při 4 odlišných sekvencích už je počet možných stromů 15 atd. Počet DNA sekvencí Počet možných stromů ,2 x ,8 x 10 76
31 většinou však analyzujeme stovky sekvencí počet možných stromů je pak astronomický např. Linda Vigilant a kol. analyzovali 135 mtdna sekvencí, jejichž rozdíly vedly až k možných stromů jakkoliv rozumně velký vzorek nám vždy bude generovat takový počet možných stromů, které nejsme schopni analyzovat ani pomocí moderní výpočetní techniky avšak byly vytvořeny programy, které umí vyhledat a pracovat se skupinou statisticky nejpravděpodobnějších stromů nejpoužívanějším je PAUP (Phylogenetic Analysis Using Parsimony, D. Swofford)
32 z pohledu analýzy dat je tu však ještě jeden problém vícenásobné substituce, kdy v jedné sekvenci proběhne více změn např. u společného předka máme sekvenci CAGT, ale následně došlo ke dvěma záměnám u jedné linie mutoval A na C a v jiné následné linii A na T = máme tady dvě následovnické linie se sekvencí CCGT a CTGT porovnáme-li tyto dvě linie, vypadá to jakoby šlo o jednu mutační událost C na T nebo T na C, ve skutečnosti však jde o dvě mutace ve stejném místě to může vést k podhodnocení výsledků PAUP a podobné programy však umí tento problém také vyřešit
33 po úspěšné konstrukci vývojového stromu se dále snažíme odpovědět na otázku: Kdy žil společný předek? na základě fylogenetického stromu dokážeme odhadnout počet genetických změn pro každou skupinu, při znalosti průměrné mutační rychlosti na generaci je pak možné lehce dopočítat počet generací ke společnému předku (viz dříve molekulární hodiny, zde aplikujeme uvnitř druhu) Ilustrační příklad: Zcela hypoteticky - víme, že průměrná sekvenční odlišnost mezi žijícími lidmi a kmenem vývojového stromu je 0,45 %. 0,45 %
34 Ilustrační příklad: Hledání našeho společného předka pokud je průměrná mutační rychlost 3 % za generací, což je 0,00006 % na generaci pak můžeme odhadnout dobu od společného předka jako 0,45 / 0,00006 = = generací budeme-li uvažovat 28 let na generaci, pak je výsledek x 28 = let zpátky výsledek však, jak vidíme, hodně záleží na správnosti hodnot, které do výpočtu vstupují (o tom bude řeč ještě později) Nyní se již můžeme podívat, na co přišli ve svém převratném článku Cann, Stoneking a Wilson (Nature, 1987, Mitochondrial DNA and human evolution )
35 Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle mtdna a chromozomu Y 7) Další analýzy a co z toho vyplývá 8) Haploskupiny mtdna a chromozomu Y v ČR
36 Co bylo zjištěno kde a kdy Eva žila Hledání našeho společného předka Cann a kol. analyzovali mtdna 147 lidí (145 izolováno z placenty z USA, Austrálie, Nová Guinea), jako zdroj kromě placenty lze využít také např. vlasy každý vzorek byl na základě informací příbuzných přidělen do jedné skupiny podle geografického původu - sub-saharská Afrika, kavkazsko (zahrnující Evropu, Severní Afriku a Střední východ), Východní Asie, Austrálie (původní obyvatelé) a Nová Guinea (původní obyvatelé) k analýze použili 12 restrikčních enzymů (průměrně připadlo 370 restrikčních míst na osobu) v některých případech poskytlo štěpení různých mtdna shodné výsledky tyto vzorky proto byly z analýzy vyloučeny = analyzováno tak bylo 133 jedinečných mtdna analýza pomocí PAUP, bez systému outgroup pro stanovení kmene stromu (nebyla k dispozici mtdna šimpanze)
37 Výsledek 1 Hledání našeho společného předka získaný strom obsahuje dva významné shluky: 1) zástupci se sub-saharským předkem 2) ostatní regiony včetně Afriky = s největší pravděpodobností byl společný předek z Afriky = druhé vysvětlení je méně pravděpodobnější a vyžadovalo by opakované oboustranné migrace mezi Afrikou a ostatními kontinenty společný africký předek je však v souladu jak s hypotézou nahrazení, tak i s multiregionálním modelem dávný africký předek (Homo erectus) je zřejmý v obou modelech avšak africký model počítá s ještě jedním mladším předkem oba základní modely se tedy liší názorem na to, jak a kdy k tomu došlo
38 Výsledek 2 Hledání našeho společného předka Cann a kol. se na základě odhadu četnosti mutací v mtdna a sledované variability pokusili o výpočet, kdy došlo k oddělení od tohoto společného předka vycházeli z rozdílů v četnostech mutací domorodých obyvatel Nové Guineje a Austrálie kalibrace pomocí archeologických dat, známe dobu, kdy předchůdci kolonizovali tyto světy vypočítanáčetnost je v průměru 2 4 % rozdílů za milion let (zhruba odpovídá četnosti změn vypočítané u řady druhů obratlovců) vypočítali, že společný předek žil před lety (2 % na milion let) až lety (při divergenci 4 % za milion let) = průměrný odhad tedy je let
39 mtdna moderních lidí pochází od společného předka, který žil v Africe před zhruba lety.? Co se stalo s mtdna předků, kteří v té době žili mimo Afriku? podle všeho byl H. erectus, neanderthalensis aj. zcela nahrazen moderním člověkem bez vzájemného křížení, jinak bychom nalezli nějaké starší stopy v mtdna (viz analýza archaické DNA )
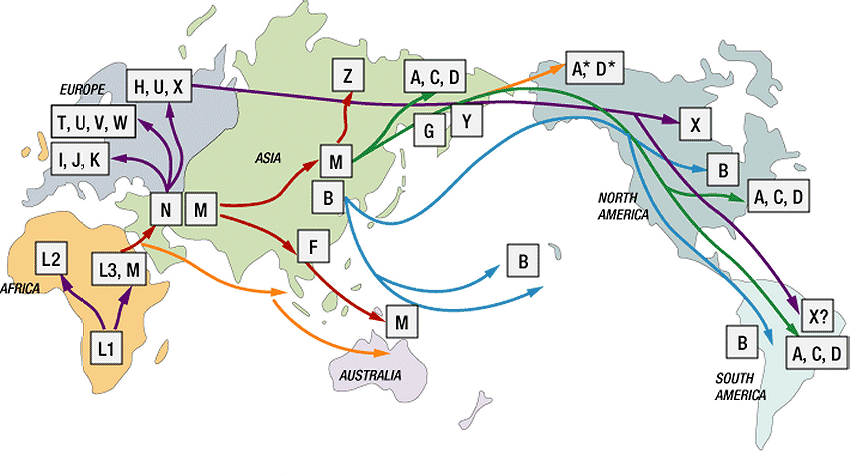
40 mtdna moderních lidí pochází od společného předka, který žil v Africe před zhruba lety. tento společný předek, žena, začala být nazývána Evou nebo mitochondriální Evou na základě spodobnění s biblickým původem lidí v Evě a Adamovi avšak pozor, ve skutečnosti neexistovala v té době v Africe pouze jediná Eva, ale těchto Ev, předchůdkyň bylo zcela logicky určitě několik stovek nebo tisíce z Afriky expandovala řada malých populací, avšak postupem času mtdna linie ostatních předchůdkyň zanikaly (neměly dcery, zanikaly populace apod.) až se do současnosti zachovala mtdna té jediné africké předchůdkyně (viz dříve recept na impalí polévku, nebo stejně jako zanikají ženská jména) každá z dalších (mladších) předchůdkyň nám zanechala část své mtdna v té naší = můžeme tak provést další dělení podle geografického původu (viz sedm dcer Eviných) a stopovat tak např. postupné osidlování Evropy těmito mladšími předchůdkyněmi současného člověka
41 3 v Africe (haploskupiny L1, L2 a L3) 7 v Asii (odvozené od společné předchůdkyně z Afriky označované jako M: A, B, C, D, E, F a G 9 v Evropě, z toho 7 tzv. hlavních (H - Helena, J Jasmina, K- Kateřina, T Tara, U Uršula, V Valda a X Xenie) 4 v Americe A, B, C, D (nedávné osídlení z Asie)
42
43 Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle mtdna a chromozomu Y 7) Další analýzy a co z toho vyplývá 8) Haploskupiny mtdna a chromozomu Y v ČR
44 Problém č. 1 Hledání našeho společného předka Analýza mtdna u nepůvodních obyvatel (osídlenců po migraci) např. studium mtdna Afroameričanů přesnost datování je ovlivněna rovnoměrným zastoupením sňatků žen Američanek s muži Afričany a sňatků žen Afričanek s Američany pak je i rovnoměrně zastoupená mtdna obou původních skupin můžeme sledovat nejen dávný původ mtdna v Africe, ale také modernější původ těchto prokřížených populací v Africe a Evropě lze tak vystopovat jejich původ
45 Problém č. 2 Hledání našeho společného předka Přesnost datování odhad mutační rychlosti první studie z roku 1987, Cann et al. kalibrace pomocí archeologických dat četnost mutací 2-4 % za milion let stáří Evy let, průměr je let Linda Vigilant et al., zkoumali nukleotidovou sekvenci v kontrolním regionu mtdna o délce pb získali velmi podobný strom jako Cann et al. dospěli k přesnější hodnotě mutační rychlosti, použili navíc šimpanzí mtdna pro stanovení kořene stromu ( outgroup ) zjistili 69,2 % odlišností ve studované šimpanzí a lidské sekvenci 17,3 % změn na milion let odhadnutá mutační rychlost od současného člověka ke společnému předku byla 2,87 % odlišností (v předchozí výpočty 2 až 4 %) tedy průměrná rychlost změn (kalibrace přes šimpanzí DNA) je 2,87 / 17,3 = 0,166 za milion let (předek se šimpanzem žil před 4 miliony let) až 2,87 / 11,5 = 0,2496 (předek se šimpanzem žil před 6 miliony let)
46 Problém č. 2 Hledání našeho společného předka Přesnost datování odhad mutační rychlosti vypočítané stáří společného předka moderního člověka se tak pohybovalo mezi až lety, s průměrem kolem let (x Cann et al. s průměrem kolem let) výsledky získané Lindou Vigilantovou s kol. byly tedy téměř shodné a systém datování i výsledky těchto analýz začaly být uznávány za platné současně je zřejmé, že analýzy mtdna podporují hypotézu o africkém původu moderního člověka, multiregionální model předpokládá předchůdce mnohem staršího (chyba není v datování) Ingman et al., 2000 studovali nejenom sekvence kontrolní oblasti (tvoří méně než 7 % mtdna) do studií zařadili i kódující oblast mtdna a pro potvrzení výsledků nově také jaderný genom v podoběčásti chromozomu X (oblast Xq13.3)
47 Ingman a kol (Nature) Závěr 1 o původu Hledání našeho společného předka na obrázku jsou výsledky analýzy pouze kódující části mtdna MRCA potvrzeny předchozí závěry o africkém původu Závěr 2 datování MRCA před až let, průměr let (odchylka je dána výrazně odlišnou mutační rychlostí u kontrolní a kódující oblasti) Závěr 3 chromozom X analýza sekvence chromozomu X (Xq13.3) / 3* = vs u mtdna potvrzuje výsledky získané u mtdna * u chromozomu X musíme počítat s 3x větší efektivní velikostí populace
48 Problém č. 2 Hledání našeho společného předka Přesnost datování variabilita mutační rychlosti (Christopher Wills, 1995) existuje například rozdíl v četnosti tranzicí (purin za purin, pyrimidin za pyrimidin) a transverzí (purin za pyrimidin a naopak) četnost tranzicí je mnohem vyšší než je četnost transverzí = četnost transverzí je pro časování vhodnější (rychlost transverzí je v průměru 3,32) použitím mutační rychlosti transverzí lze dospět k závěru, že mteva žila před až lety (zahrneme-li do odhadu 95% interval spolehlivosti, pak je rozmezí mezi až lety) Eva tak mohla žít mnohem dříve, což by mohlo být v částečném souladu s multiregionálním modelem. Při analýzách však bereme v úvahu všechny typy mutací, a to nejen substituce, ale také inzerce, delece, amplifikace = bereme v úvahu veškerý mutační tlak dříve prezentované výsledky jsou tedy reprezentativnější = námitku lze zamítnout
49 Problém č. 2 Hledání našeho společného předka Přesnost datování kalibrace molekulárních hodin původní metoda kalibrace podle fosilních nálezů 1 mutace na 1,0 miliardu bází za rok (kalibrace podle fosilií orangutana) Nové metody: na základě celogenomového mapování u novorozených dětí (např. islandská studie novorozenec má kolem 36 spontánních mutací, které nezdědil po rodičích) 1 mutace na 2,0 miliardy bází za rok (o polovinu pomalejší rychlost) rychlost mutací u mikrosatelitů 1 mutace na 1,2 miliardy až 1 mutace na 2,0 miliardy bází za rok rychlost mutací studiem rodokmenů 1,24 mutace na 1 milion bází za rok (3x rychlejší) Poznámka: kalibraci je potřeba provádět podle typu zvolených analýz a markerů mutační rychlost stanovená v krátkém časovém úseku (např. z rodokmenů) nemusí odpovídat té dlouhodobé (vliv purifikujícího výběru apod.)
50 Názory proti modelu Out of Africa Hledání našeho společného předka přestože řada prací i z nedávné doby dále potvrzuje získané výsledky, zmíníme si i názory proti Africkému původu člověka vycházející z mtdna 1) Výsledky mtdna vůbec nemusí svědčit o Africkém původu moderního člověka Otázka interpretace genetických stromů (Alan Templeton, 1993, 1997, 1998) získané genetické stromy nemusí odpovídat populačním stromům avšak řada studií až do současnosti ukazuje na vysokou korelaci mezi genetickými a populačními údaji = námitku lze zamítnout rozdělení mtdna na africkou a neafrickou nemusí odrážet reálné rozdělení populací (neboli geografická asociace tohoto rozdělení nemusí představovat ono následné nahrazení expandující africkou populací)
51 Názory proti modelu Out of Africa Hledání našeho společného předka A) odlišnost afrických populací od ostatních nemusí být důsledkem nahrazení, ale příčinou může být omezený genový tok v důsledku prostorové izolace jsou-li populace příliš vzdálené, neprobíhá genový tok a výsledek vypadá jako při nahrazení jedněch populací jinými Templeton 1997, Current Opinion in Genetics & Development B) dle Templetona neexistuje důkaz o expanzi africké populace mimo Afriku africké stopy přítomné ve všech populacích jsou spíše důsledkem stálého genového toku mezi geograficky blízkými oblastmi v uplynulých několika stech tisících letech (africké znaky se tak do těchto populací rozšířily)
52 Názory proti modelu Out of Africa Hledání našeho společného předka Templeton (2002) dále uvádí, že jeho nové analýzy genetických stromů (kombinuje až 10 různých znaků) ukazují na několik migrací z Afriky s neustálým genovým tokem první expanze proběhla před 1,7 miliony let Homo erectus druhá vlna před 800 až lety a) expanze Homo heidelbergensis (předek neandrtálce) nebo b) možná migrace jiného předka neandrtálce a moderního člověka třetí vlna před asi lety
53 Názory proti modelu Out of Africa Hledání našeho společného předka Templetonovy výsledky svědčící o opakovaném genovém toku (expanze z Afriky a následné křížení) v průběhu posledních 2 milionů let: podporují africké genetické kořeny = moderníčlověk se mohl rozšířit z Afriky, avšak nejen odtud = Eva mohla žít kdekoliv do jisté míry podporuje multiregionální model opakované migrace s genovým tokem vedou ke stejnému výsledku, tedy africkému původu, přestože je původ ve skutečnosti multiregionální plně však nepodporuje ani jeden ze dvou modelů uznáván jako další model Out of Africa again and again
54 Názory proti modelu Out of Africa Hledání našeho společného předka 2) Vliv velikosti populace (genetického driftu) na přesnost výpočtů: Př.: sledujeme 4 mtdna sekvence A, B, C a D, vzájemné genealogické sekvenční vazby mezi nimi jsou vyjádřeny jako CA1, CA2 a CA3 označují společné předky, místo koalescence (splynutí) většinou nás zajímá tzv. nejbližší společný předek (MRCA, Most Recent Common Ancestor), což je v našem případě CA3 množství rozdílů odráží rychlost splývání = = odhad stáří MRCA rychlost s jakou jednotlivé linie splývají je však ovlivněna také velikostí populace = splývání je rychlejší v menších populacích (jedinci jsou si tam podobnější než v populacích větších)
55 Názory proti modelu Out of Africa Hledání našeho společného předka 2) Vliv velikosti populace (genetického driftu) na přesnost výpočtů: Přepočet údajů o době objevení se Evy z původních dat kolektivu Cannové et al., (při 2% mutační rychlosti) zjištěná průměrná hodnota let byla podobná odhadu Cann et al. avšak po započítání vlivu genového posunu (velikosti populace) = dostáváme interval až let (oproti původnímu až , rozsah let) = dost velký rozsah ( let) je typický pro tento přístup, protože na variabilitu má právě významný vliv působící genový posun přepočet údajů Vigilantové et al až (4 miliony let od oddělení šimpanze) až (pro 6 milionů let) = tj let (oproti původnímu až )
56 Názory proti modelu Out of Africa Hledání našeho společného předka 2) Vliv velikosti populace (genetického driftu) na přesnost výpočtů: vliv genového posunu je nezanedbatelný na přesnost výpočtu avšak údaje jsou velmi daleko od afrického předka starého 2 miliony let I po korekcích na velikost populace zůstává podpora modelu nahrazení. Poznámka: vezmeme-li v úvahu i Templetonův model Out Africa again and again pak je po přepočtech výrazná podpora pro něj (druhá migrační vlna před až lety;??homo heidelbergensis??)
57 Názory proti modelu Out of Africa Hledání našeho společného předka 3) Vliv přírodního výběru pokud vycházíme z fenotypové variability, pak je nutné si uvědomit do jaké míry je podobnost mezi dvěma populacemi odrazem společného předka (původu) a do jaké míry je výsledkem konvergence v důsledku přírodního výběru budeme-li uvažovat o mutacích v mtdna jako o evolučně neutrálních, pak lze vliv přírodního výběru zanedbat v opačném případě: museli bychom vědět, jak velká část rozmanitosti žijících populací byla smazána působením přírodního výběru v neprospěch škodlivých mutací jinak by byly naše údaje podhodnoceny a společný předek by tak žil mnohem dříve než se nyní domníváme
58 Názory proti modelu Out of Africa Hledání našeho společného předka 4) Spolehlivost maternální dědičnosti mtdna: některé studie naznačují, že mtdna může být významnou měrou přenášena také od mužů a může zde probíhat také rekombinace (Shitara et al., 1998; Awadalla et al., 1999) pokud by to tak bylo, pak získané výsledky jsou velmi vratké avšak: ad.: Shitara et al. - testováno na mezidruhových hybridech myší - mtdna nebyla ve všech tkáních - v další generaci už pouze samičí mtdna = pravděpodobně jen důsledek mezidruhového křížení ad.: Awadalla et al. - později se objevuje několik článků, které závěry Awadalla et al. zpochybnily - chyby v datech, metodách analýzy a interpretační zmatky = dědičnost mtdna však asi bude ještě potřeba spolehlivě prověřit
59 V současné době probíhá tzv. Genographic Project ( odkaz Hledání našeho společného předka
60 cílem je vytvořit celosvětovou, rozsáhlou, veřejně přístupnou databázi mtdna u každého vzorku je přímým sekvencováním analyzována oblast HVS-I a dále 22 SNP v kódující oblasti projekt byl spuštěn začátkem roku 2005 na začátku se do něj přihlásilo přes dobrovolníků a více než také zaslalo svoji DNA pro analýzu mtdna nebo MSY (studium variability chromozomu Y), zatím převládají vzorky z USA a Západní Evropy (95 % databáze) v polovině roku 2007 (Behar et al., 2007) bylo analyzováno již různých mtdna spuštěn projekt 2.0 analyzuje se markerů v kompletní DNA umožňuje popsat vlastní dávnou historii mateřskou, otcovskou migrační vlny zjišťuje příměs neandrtálské a děnisovanské DNA analyzováno již téměř vzorků DNA
61 Výsledky poslední studie mtdna Hledání našeho společného předka Behar et al., 2012 Behar et al kompletních sekvencí mtdna (z toho je z dobrovolnické genealogické databáze Family Tree DNA = zejména západní euroasijský původ) jako outgroup poprvé použita mtdna Neandrtálce stáří společného předka moderního člověka přepočteno na ~ let
62 Celkový závěr mtdna lze použít jako bohatý zdroj genetických informací pro evoluční studie, avšak s nevýhodou studia pouze evoluce ženské části populací.
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
Genetické rozdíly mezi populacemi aneb něco o migracích a genovém toku. Genetické rozdíly mezi populacemi
 Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Jak to, že máme spole ného p edka?
 1) Jak to, že máme spole ného p edka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriálníeva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili sv t podle mtdna
1) Jak to, že máme spole ného p edka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriálníeva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili sv t podle mtdna
Paleogenetika člověka
 Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Kdo je náš předek?
Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Kdo je náš předek?
Jak měříme genetickou vzdálenost a co nám říká F ST
 Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Evoluce člověka a její modely
 Evoluce člověka a její modely Evoluce člověka a její modely 1) Jak dlouho jsme na Zemi 2) Evoluce rodu Homo z genetického pohledu a možné scénáře zajímá nás, co se dělo v posledních 2 milionech let a jak
Evoluce člověka a její modely Evoluce člověka a její modely 1) Jak dlouho jsme na Zemi 2) Evoluce rodu Homo z genetického pohledu a možné scénáře zajímá nás, co se dělo v posledních 2 milionech let a jak
Evoluce člověka a její modely
 a její modely a její modely 2) z genetického pohledu a možné scénáře zajímá nás, co se dělo v posledních 2 milionech let a jak se to dělo (evoluce rodu Homo) přestože je to velmi krátké období v historii
a její modely a její modely 2) z genetického pohledu a možné scénáře zajímá nás, co se dělo v posledních 2 milionech let a jak se to dělo (evoluce rodu Homo) přestože je to velmi krátké období v historii
Co víme o původu psa na základě analýzy DNA
 Co víme o původu psa na základě analýzy DNA RNDr. Pavel Lízal, Ph.D. Přírodovědecká fakulta MU Ústav experimentální biologie Oddělení genetiky a molekulární biologie lizal@sci.muni.cz 1) Hypotéza o domestikaci
Co víme o původu psa na základě analýzy DNA RNDr. Pavel Lízal, Ph.D. Přírodovědecká fakulta MU Ústav experimentální biologie Oddělení genetiky a molekulární biologie lizal@sci.muni.cz 1) Hypotéza o domestikaci
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Populační genetika III. Radka Reifová
 Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Analýza archaické DNA
 Analýza archaické DNA 1) Analýza adna člověka 2) Analýza první neandrtálské DNA 3) Analýza druhé neandrtálské DNA 4) Interpretace nalezených výsledků - Byli Neandrtálci odděleným druhem nebo byli odlišným
Analýza archaické DNA 1) Analýza adna člověka 2) Analýza první neandrtálské DNA 3) Analýza druhé neandrtálské DNA 4) Interpretace nalezených výsledků - Byli Neandrtálci odděleným druhem nebo byli odlišným
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Testování lidské identity
 Testování lidské identity Brno, 2009 J.M.Butler Forensic DNA Typing workshop, 2006 Bryan Sykes Sedm dcer Eviných, 2005 Využití testování lidské identity Řešení trestních činů shoda mezi podezřelým a stopou
Testování lidské identity Brno, 2009 J.M.Butler Forensic DNA Typing workshop, 2006 Bryan Sykes Sedm dcer Eviných, 2005 Využití testování lidské identity Řešení trestních činů shoda mezi podezřelým a stopou
Úvod do studia historie moderního člověka
 Paleogenetika člověka Úvod do studia historie moderního člověka 1) Studium historie člověka - před první analýzou DNA - po první analýze DNA 2) Kronika našeho druhu všichni naši předkové (rychlý náhled)
Paleogenetika člověka Úvod do studia historie moderního člověka 1) Studium historie člověka - před první analýzou DNA - po první analýze DNA 2) Kronika našeho druhu všichni naši předkové (rychlý náhled)
Kolik jsme měli předků?
 1) Velikost populace a demografická historie 2) Rozdílné pohledy na velikost populace 3) Odhad velikostí dávných populací - Ekologické odhady celkové velikosti populací - Genetické odhady efektivní velikosti
1) Velikost populace a demografická historie 2) Rozdílné pohledy na velikost populace 3) Odhad velikostí dávných populací - Ekologické odhady celkové velikosti populací - Genetické odhady efektivní velikosti
Coalesce spojit se, splynout, sloučit se. Didaktická simulace Coalescence = splynutí linií
 Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Co víme o původu psa na základě analýzy DNA
 Co víme o původu psa na základě analýzy DNA RNDr. Pavel Lízal, Ph.D. Přírodovědecká fakulta MU Ústav experimentální biologie Oddělení genetiky a molekulární biologie lizal@sci.muni.cz Přednáška představí
Co víme o původu psa na základě analýzy DNA RNDr. Pavel Lízal, Ph.D. Přírodovědecká fakulta MU Ústav experimentální biologie Oddělení genetiky a molekulární biologie lizal@sci.muni.cz Přednáška představí
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
1) Velikost populace a demografická historie. 1) Velikost populace a demografická historie. 1) Velikost populace a demografická historie
 Velikost populace a demografická historie. 1) Velikost populace a demografická historie. 1) Velikost populace a demografická historie") 1) Velikost populace a demografická historie 2) Rozdílné pohledy na velikost populace 3) Odhad velikostí dávných populací - Ekologické odhady celkové velikosti populací - Genetické odhady efektivní velikosti
1) Velikost populace a demografická historie 2) Rozdílné pohledy na velikost populace 3) Odhad velikostí dávných populací - Ekologické odhady celkové velikosti populací - Genetické odhady efektivní velikosti
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Zajímavost -mtdna a Y v R
 1) Jak to, že máme spole ného p edka v R si lze za poplatek nechat analyzovat svoji mtdna a chromozom Y konstrukce genetických strom a ur ení p íbuznosti k mtdna a Y liniím (nap. Forenzní DNA servis, nový
1) Jak to, že máme spole ného p edka v R si lze za poplatek nechat analyzovat svoji mtdna a chromozom Y konstrukce genetických strom a ur ení p íbuznosti k mtdna a Y liniím (nap. Forenzní DNA servis, nový
Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Bi5130 Základy práce s lidskou adna
 Mgr. et Mgr. Kristýna Brzobohatá brzobohata@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu 1) Modelování historických událostí a jejich limity 2) Teorie šíření moderního Homo
Mgr. et Mgr. Kristýna Brzobohatá brzobohata@sci.muni.cz Laboratoř biologické a molekulární antropologie, ÚEB, PřF, Mu 1) Modelování historických událostí a jejich limity 2) Teorie šíření moderního Homo
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,
: Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,") Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Projekt Genetika a příjmení a zapojení Klusáčků do něj
 Projekt Genetika a příjmení a zapojení Klusáčků do něj Písemnosti odeslané jmenovcům ze dne 10. září 2008: Vážený pane Klusáčku obracíme se na Vás touto cestou s nabídkou účasti v unikátním vědeckém projektu
Projekt Genetika a příjmení a zapojení Klusáčků do něj Písemnosti odeslané jmenovcům ze dne 10. září 2008: Vážený pane Klusáčku obracíme se na Vás touto cestou s nabídkou účasti v unikátním vědeckém projektu
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Příbuznost a inbreeding
 Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Metody studia historie populací. Metody studia historie populací. 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.
 Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.") 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva
1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské. doc. RNDr. Ivan Mazura, CSc.
 Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Analýza archaické DNA
 Analýza archaické DNA Analýza archaické DNA 1) Analýza adna člověka 2) Analýza první neandrtálské DNA 3) Analýza druhé neandrtálské DNA 4) Interpretace nalezených výsledků - Byli Neandrtálci odděleným
Analýza archaické DNA Analýza archaické DNA 1) Analýza adna člověka 2) Analýza první neandrtálské DNA 3) Analýza druhé neandrtálské DNA 4) Interpretace nalezených výsledků - Byli Neandrtálci odděleným
STATISTICKÉ ODHADY Odhady populačních charakteristik
 STATISTICKÉ ODHADY Odhady populačních charakteristik Jak stanovit charakteristiky rozložení sledované veličiny v základní populaci? Populaci většinou nemáme celou k dispozici, musíme se spokojit jen s
STATISTICKÉ ODHADY Odhady populačních charakteristik Jak stanovit charakteristiky rozložení sledované veličiny v základní populaci? Populaci většinou nemáme celou k dispozici, musíme se spokojit jen s
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Počet chromosomů v buňkách. Genom
 Počet chromosomů v buňkách V každé buňce těla je stejný počet chromosomů. Výjimkou jsou buňky pohlavní, v nich je počet chromosomů poloviční. Spojením pohlavních buněk vzniká zárodečná buňka s celistvým
Počet chromosomů v buňkách V každé buňce těla je stejný počet chromosomů. Výjimkou jsou buňky pohlavní, v nich je počet chromosomů poloviční. Spojením pohlavních buněk vzniká zárodečná buňka s celistvým
Migrace. 1) Jednosměrná migrace. 2) Obousměrná migrace. 3) Genový tok a historie populací. 4) Migrace a genetická odlišnost mezi populacemi
 Jednosměrná migrace. 2) Obousměrná migrace. 3) Genový tok a historie populací. 4) Migrace a genetická odlišnost mezi populacemi") P m p 0 1) Jednosměrná migrace 2) Obousměrná migrace 3) Genový tok a historie populací 4) Migrace a genetická odlišnost mezi populacemi 5) Odhady velikosti migrace 6) Přerušení izolace a Wahlundův princip
P m p 0 1) Jednosměrná migrace 2) Obousměrná migrace 3) Genový tok a historie populací 4) Migrace a genetická odlišnost mezi populacemi 5) Odhady velikosti migrace 6) Přerušení izolace a Wahlundův princip
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Předpověď plemenné hodnoty Něco málo z praxe. Zdeňka Veselá
 Předpověď plemenné hodnoty Něco málo z praxe Zdeňka Veselá vesela.zdenka@vuzv.cz Příprava datových souboru Databáze s výsledky užitkovosti jsou zpravidla obrovské soubory Např. kontrola užitkovosti masného
Předpověď plemenné hodnoty Něco málo z praxe Zdeňka Veselá vesela.zdenka@vuzv.cz Příprava datových souboru Databáze s výsledky užitkovosti jsou zpravidla obrovské soubory Např. kontrola užitkovosti masného
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
FYLOGEOGRAFIE A KOALESCENCE
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
Deoxyribonukleová kyselina (DNA)
") Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Osud jednotlivých kopií genů v populaci genové stromy
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
Základní pravidla dědičnosti
 Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Jana Vránová, 3.lékařská fakulta UK, Praha. Hypotézy o populacích
 Jana Vránová, 3.lékařská fakulta UK, Praha Hypotézy o populacích Příklad IQ test: Předpokládejme, že z nějakého důvodu ministerstvo školství věří, že studenti absolventi středních škol v Hradci Králové
Jana Vránová, 3.lékařská fakulta UK, Praha Hypotézy o populacích Příklad IQ test: Předpokládejme, že z nějakého důvodu ministerstvo školství věří, že studenti absolventi středních škol v Hradci Králové
Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
 Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Dědičnost vázaná na X chromosom
 12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
Cvičeníč. 10 Dědičnost a pohlaví. Mgr. Zbyněk Houdek
 Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
 Evoluční genetika KBI/GENE Mgr. Zbyněk Houdek Evoluční teorie Evoluční teorii vyslovil Ch. Darwin v díle O původu druhů (1859), kde ukazoval, že druhy se postupně měnily v dlouhých časových periodách.
Evoluční genetika KBI/GENE Mgr. Zbyněk Houdek Evoluční teorie Evoluční teorii vyslovil Ch. Darwin v díle O původu druhů (1859), kde ukazoval, že druhy se postupně měnily v dlouhých časových periodách.
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
676 + 4 + 100 + 196 + 0 + 484 + 196 + 324 + 64 + 324 = = 2368
 Příklad 1 Je třeba prověřit, zda lze na 5% hladině významnosti pokládat za prokázanou hypotézu, že střední doba výroby výlisku je 30 sekund. Přitom 10 náhodně vybraných výlisků bylo vyráběno celkem 540
Příklad 1 Je třeba prověřit, zda lze na 5% hladině významnosti pokládat za prokázanou hypotézu, že střední doba výroby výlisku je 30 sekund. Přitom 10 náhodně vybraných výlisků bylo vyráběno celkem 540
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
World of Plants Sources for Botanical Courses
 Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Projekční algoritmus. Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění. Jan Klíma
 Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Genetika pro začínající chovatele
 21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna
 Mendelova genetika v příkladech Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna Roman Longauer, Ústav zakládání a pěstění lesů, MENDELU
Mendelova genetika v příkladech Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna Roman Longauer, Ústav zakládání a pěstění lesů, MENDELU
Tok GI v buňce. Genetický polymorfizmus popis struktury populací. Organizace genetického materiálu. Definice polymorfismu
 Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Základní genetické pojmy
 Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
KBI / GENE Mgr. Zbyněk Houdek
 Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY
 GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
Základy genetiky populací
 Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Dědičnost a pohlaví. KBI/GENE Mgr. Zbyněk Houdek
 Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Paleolitická adna. Mitochondriální DNA Nukleární DNA Y Porovnání současné DNA (Paleogenetika, National Geographic )
") Paleolitická adna Paleolitická adna Mitochondriální DNA Nukleární DNA Y Porovnání současné DNA (Paleogenetika, National Geographic ) Genographic project https://genographic.nationalgeographic.com/ 1000genomes
Paleolitická adna Paleolitická adna Mitochondriální DNA Nukleární DNA Y Porovnání současné DNA (Paleogenetika, National Geographic ) Genographic project https://genographic.nationalgeographic.com/ 1000genomes
EVOLUCE ČLOVĚKA. úlohy k tématu + autorské řešení. Radka M. Dvořáková, Karolína Absolonová
 EVOLUCE ČLOVĚKA úlohy k tématu + autorské řešení Radka M. Dvořáková, Karolína Absolonová Následující úlohy mají sloužit jako inspirace pro školní práci s informacemi, které učitelé a studenti naleznou
EVOLUCE ČLOVĚKA úlohy k tématu + autorské řešení Radka M. Dvořáková, Karolína Absolonová Následující úlohy mají sloužit jako inspirace pro školní práci s informacemi, které učitelé a studenti naleznou
Heritabilita. Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability
 Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
Biologie - Oktáva, 4. ročník (přírodovědná větev)
") - Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
- Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Testování hypotéz. 1. vymezení základních pojmů 2. testování hypotéz o rozdílu průměrů 3. jednovýběrový t-test
 Testování hypotéz 1. vymezení základních pojmů 2. testování hypotéz o rozdílu průměrů 3. jednovýběrový t-test Testování hypotéz proces, kterým rozhodujeme, zda přijmeme nebo zamítneme nulovou hypotézu
Testování hypotéz 1. vymezení základních pojmů 2. testování hypotéz o rozdílu průměrů 3. jednovýběrový t-test Testování hypotéz proces, kterým rozhodujeme, zda přijmeme nebo zamítneme nulovou hypotézu
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií
 poznatků genetiky a evolucionistických teorií") Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
a) Sledovaný znak (nemoc) je podmíněn vždy jen jedním genem se dvěma alelami, mezi kterými je vztah úplné dominance.
 Sledovaný znak (nemoc) je podmíněn vždy jen jedním genem se dvěma alelami, mezi kterými je vztah úplné dominance.") GENEALOGIE (Genealogická metoda. Genealogické symboly. Rozbor rodokmenů. Základní typy dědičnosti.) ÚVOD Genealogie je základem genetického vyšetření člověka, jehož cílem je stanovení typu dědičnosti daného
GENEALOGIE (Genealogická metoda. Genealogické symboly. Rozbor rodokmenů. Základní typy dědičnosti.) ÚVOD Genealogie je základem genetického vyšetření člověka, jehož cílem je stanovení typu dědičnosti daného
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Biologie - Oktáva, 4. ročník (humanitní větev)
") - Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
- Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
Fylogeneze a diverzita obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
Systém a evoluce obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
Tabulka 1 Rizikové online zážitky v závislosti na místě přístupu k internetu N M SD Min Max. Přístup ve vlastním pokoji 10804 1,61 1,61 0,00 5,00
 Seminární úkol č. 4 Autoři: Klára Čapková (406803), Markéta Peschková (414906) Zdroj dat: EU Kids Online Survey Popis dat Analyzovaná data pocházejí z výzkumu online chování dětí z 25 evropských zemí.
Seminární úkol č. 4 Autoři: Klára Čapková (406803), Markéta Peschková (414906) Zdroj dat: EU Kids Online Survey Popis dat Analyzovaná data pocházejí z výzkumu online chování dětí z 25 evropských zemí.
Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce jak lze studovat evoluci a jak funguje mutace, přírodní výběr, genový posun a genový tok 3)
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce jak lze studovat evoluci a jak funguje mutace, přírodní výběr, genový posun a genový tok 3)
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou