Univerzita Karlova v Praze. Přírodovědecká fakulta. Studijní program: Biologie. Studijní obor: Ekologická a evoluční biologie
|
|
|
- Tadeáš Kolář
- před 5 lety
- Počet zobrazení:
Transkript

1 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Ekologická a evoluční biologie Martin Pošmourný Vliv vysychání na streptofytní řasy mechanismy stresové odolnosti The effect of desiccation on streptophyte algae - mechanisms of stress resistance Bakalářská práce Školitel: RNDr. Martina Pichrtová Ph.D. Praha
2 PROHLÁŠENÍ: Prohlašuji, ţe jsem závěrečnou práci zpracoval samostatně a ţe jsem uvedl všechny pouţité informační zdroje a literaturu. Tato práce ani její podstatná část nebyla předloţena k získání jiného nebo stejného akademického titulu. V Praze Martin Pošmourný 2
3 PODĚKOVÁNÍ Za vţdy ochotnou a odbornou pomoc při zpracování této práce a hlavně za milou trpělivost velice děkuji RNDr. Martině Pichrtové Ph.D. Své rodině děkuji za podporu při studiu. A veliký dík patří také Františku Brezíkovi, bez kterého bych tuto práci nikdy nedokončil. 3
4 Obsah: Úvod... 6 Streptofytní řasy - obecná charakteristika Nedostatek vody v buňce Důsledky vysychání Strukturální změny Fyziologické změny Dvě roviny odolnosti proti nedostatku vody Význam stresové odolnosti streptofytních řas Ekologický: půdní krusty - významný producent biomasy a fixátor dusíku a uhlíku, pionýrské sukcesní stádium, půdotvorný efekt Evoluční: kolonizace souše, vznik evolučních novinek Stresová odolnost jednotlivých skupin streptofytních řas Chlorokybophyceae Klebsormidiophyceae Charophyceae Coleochaetophyceae Zygnematophyceae Shrnutí Seznam použité literatury Obrazová příloha
5 ABSTRAKT V této bakalářské práci jsem se věnoval odolnosti streptofytních řas proti vysychání. Přesto, ţe se této oblasti doposud věnovalo jen pár lidí, v posledních letech bylo na toto téma publikováno značné mnoţství prací. Byly zjištěny zajímavé informace a objeveny nové skutečnosti. Výzkum pokračuje stále dál a tak by bylo uţitečné se poohlédnout, na co všechno se přišlo. Věřím, ţe porozumění tomuto fenoménu je klíčové pro pochopení některých událostí ve vývoji přírody a uvědomění si, jak houţevnatý dokáţe ţivot být na samých hranicích svého moţného výskytu. Pokusil jsem se utřídit současné vědomosti o mechanismech stresové odolnosti streptofytních řas a doufám, ţe získaný přehled mi do budoucna pomůţe lépe se v dané problematice orientovat. Doposud bylo pozorováno několik přístupů k obraně proti vysychání. Prevence vysušení, adaptace na nedostatek vody a tolerance úplného vyschnutí. Mezi preventivní způsoby obrany patří vytváření shluků buněk, kobercovitých porostů, či sekrece slizového obalu. Jako adaptaci na nedostatek vody řasy vyvinuly sloţitější odpovědi v podobě změn ultrastruktury, či regulaci fyziologických procesů. Např. Klebsormidium je schopné během půlhodiny nastartovat výrobu značného mnoţství kalózy a zabudovat ji do buněčné stěny. Tím získá potřebnou pruţnost, aby v důsledku rapidního úbytku vody nedošlo k mechanickému poškození jeho ultrastruktury. Bylo zjištěno, ţe vysychání urychluje proces fotoinhibice. Výsledky výzkumů naznačují, ţe některé odpovědi na stresové podmínky jsou druhově specifické a ţe si streptofyta své předpoklady pro dobytí souše nesou stále s sebou. In this thesis I dealt streptophyta algae resistance against desiccation. Even though the area previously devoted only a few people. Considerable amount of work in recent years has been published on the subject. They were found interesting information and discovered new facts. Research continues on and on, and it would be useful to look at what was observed. I believe that understanding this phenomenon is the key to understanding some of the events in the evolution of nature and realizing how tough life can be on the very border of its possible occurrence. I tried to sort out the current knowledge about the mechanisms of stress resistance streptophyta algae and hope that I obtained an overview will help me understand better this issues. So far, it has been observed several approaches to defend against drying. Preventing drying, adaptation to water shortage and tolerance to desiccation. Among the preventive methods of defense include creating clusters of cells, multi-layered mats or secretion mucilage. As an adaptation to the lack of water algae evolved more complex answers in the form of changes in ultrastructure, or regulation of physiological processes. Klebsormidium is capable of half an hour to start the production of significant quantities callose and incorporate it into the cell wall. This will give the necessary flexibility, that due to rapid loss of water, avoid mechanical damage to the ultrastructure. It was found that drying accelerates photoinhibition. The research results suggest that some responses to stress conditions are species specific and that streptofyta its assumptions for the conquest of the land still carry with them. 5
6 Klíčová slova: Streptofytní řasy, vysychání, stresová odolnost, tolerance k vyschnutí, buněčná stěna, akinety, mechanismy obrany Úvod Alpinské ledovce, dočasná řečiště, chladné pláně ale i čerstvě odkrytá syrová půda. Všechna tato stanoviště mají něco společného, jsou tu drsné podmínky k ţivotu v podobě málo dostupné vody, silného slunečního záření a vysoké, ale i nízké teploty. Tyto lokality jsou v mnoha směrech extrémní. A přesto mají společnou ještě alespoň jednu věc. Často se na nich vyskytuje skupina organismů, které jsou schopny takové podmínky přeţít. Dokonce tam dokáţí ţít trvale. Patří mezi ně i streptofytní řasy, organismy, které jsou nejbliţšími příbuznými dávného předka, který jako první kolonizoval pevninu. Přechod na souš musel klást na pionýrský druh obrovské nároky, a přesto se dokázal s tak velkým stresem vyrovnat. Předpoklady pro odolnost proti takovému stresu po něm pravděpodobně zdědili i jeho dnešní nejbliţší příbuzní. Právě proto je zajímavé se jejich stresovou odolností, potaţmo mechanismy, které jim takovou odolnost propůjčují zabývat. Pomůţe nám to osvětlit jednu z nejdůleţitějších událostí v historii přírody. 6
7 1 Streptofytní řasy - obecná charakteristika Streptofyta jsou vývojová větev linieviridiplantae, pod kterou patří vyšší rostliny a streptofytní řasy, neboli Charophyta. Charophyta jsou tedy parafyletickou skupinou, neboť mají společného předka, ale nezahrnují všechny jeho potomky, tedy vyšší rostliny. Charakteristická je pro ně přítomnost fragmoplastu, mikrotubulární struktury vymezující prostor, kde se mateřská buňka rozdělí na dvě dceřiné. Mikrotubulární charakter fragmoplastu umoţňuje dopravu materiálu na stavbu střední lamely, která svými kvalitami nemá daleko od plnohodnotné buněčné stěny. Vznik fragmoplastu zřejmě umoţňuje lepší regulaci dělení v rámci poměrně rigidní buněčné stěny a je tak dobrým předpokladem pro mnohobuněčnost. Streptofytní řasy jsou jednobuněčné i mnohobuněčné stélkaté zelené rostliny. Jejich buňky obsahují především chlorofyl a a b. Obsahují i další pigmenty ale ty většinou chloroplasty nepřekrývají, takţe jsou streptofytní řasy převáţně zelené. Tylakoidy tvoří grana. Zásobní látkou je většinou škrob. Streptofytní řasy jsou primárně sladkovodní a jsou povaţovány za nejbliţší příbuzné prvních rostlin kolonizujících souš. (Jan de Vries et al. 2016) Druhou vývojovou větví linie Viridiplantae jsou chlorophyta. Chlorophyta byly původně na rozdíl od streptofyt mořskými organismy, ale později se i v jejich řadách vyvinuly sladkovodní druhy. Od streptofyt se liší třeba tím, ţe tylakoidy netvoří grana, ale srůstají v lamely. ( Na následujícím fylogenetickém stromě můţeme vidět současnou představu o příbuzenských vztazích v rámci skupin Chlorophyta a Streptophyta.(Obr.1.) 7
8 2 Nedostatek vody v buňce Přítomnost vody je nutnou podmínkou pro ţivot. Její nedostatek značně stěţuje ţivot všem organismům na planetě. V přírodě je nedostatek vody pro streptofytní řasy způsobován mnoha příčinami. Tou asi nejběţnější je nadměrný odpar způsobený buď vysokými teplotami, nebo nízkou vzdušnou vlhkostí. Další příčinou je nedostatek sráţek. Hned několika způsoby můţe způsobit nedostatek vody mráz. Tání či zamrzání vody ve vyšších polohách můţe ovlivnit průtok vody, potaţmo výšku hladiny potoků v polohách niţších. Pak platí prostý fakt, ţe zamrzlá voda je pro buňku nedostupná. A třetí cestou jak mráz ovlivní vodní reţim buňky je ta, ţe s postupně zamrzající vodou roste koncentrace rozpuštěných látek ve vodě zatím nezamrzlé, čímţ stoupá její osmotický potenciál. (B.Fuller 2004) Jako poslední příčinu nedostatku vody uvedu salinitu, ta můţe být způsobena nadměrným odparem, zamrzáním nebo lokálními podmínkami na stanovišti. Nedostatek vody jako stres bychom tedy mohly rozdělit do dvou základních kategorií. Dlouhodobější-sezónní nedostatek, který většinou nastupuje pozvolna, ale trvá déle. A krátkodobý-nárazový nedostatek způsobený nenadálou událostí - poklesem vodní hladiny, rychlou změnou počasí či náhlou změnou lokálních podmínek, která můţe být zapříčiněna celou paletou událostí. Mezi příčiny krátkodobého nedostatku vody můţeme zařadit také periodické změny teploty, osvitu či výšky hladiny vody během dne. Odlišné parametry těchto dvou tipů stresové zátěţe kladou na řasy odlišné nároky a vedou ke vzniku odlišných odpovědí a rozvoji jiných strategií. 8
9 3 Důsledky vysychání Pro správné porozumění mechanismům stresové odolnosti streptofytních řas bychom se měli nejprve podívat, jaké následky nedostatek vody, potaţmo vysychání, v buňce má. 3.1 Strukturální změny Voda v buňce plní hned několik funkcí a její nedostatek vede ke změnám v ultrastruktuře. (Karsten 2012) Povaţovat ji pouze za rozpouštědlo by bylo chybou. Přítomnost vody je například nezbytná pro správnou konformaci a funkčnost většiny membrán a biomolekul. (Holzinger 2013) Pokud mnoţství vody v cytoplazmě výrazně poklesne, začne docházet k agregaci proteinů, dojde ke vzniku kovalentních vazeb a k denaturaci proteinů. (Agrawal 2009) Postupně dojde k narušení integrity organel a k zániku buňky. Ať uţ řízenou cestou apoptózy, nebo dojde k nekróze (F. Kaplan 2011,2012, Holzinger, Karsten 2013, Pichrtová 2014) Integritu organel dokáţe váţným způsobem narušit i zamrzání. A to hned dvěma způsoby. Jeden z efektů zamrzání vody je zvyšování koncentrace roztoku v okolí buňky, čímţ roste osmotický tlak v buňce, který můţe vyústit v poškození organel. Druhý efekt je mechanické poškození buněčných struktur v důsledku tvorby krystalků ledu přímo uvnitř buňky. (B.Fuller 2004, Elster et al. 2008) Často pozorované zahuštění cytoplazmy různými cukry, polyalkoholy a oligosacharidy (F.Kaplan 2012) stejně jako zvýšení koncentrace osmoticky aktivních látek můţe souviset se snahou sníţit teplotu tání potaţmo zamrzání a zabránit tvorbě krystalků ledu. Popřípadě díky vyššímu osmotickému potenciálu zadrţet déle vodu a zajistit hydrataci některých strukturních proteinů, či membrán, pro něţ je, jak uţ bylo řečeno, hydratace nutnou podmínkou pro udrţení třídimenzionální struktury, potaţmo jejich funkčnosti. (Karsten 2010) 9
10 Při vysychání také dochází k těţkému poškození F-aktinových vláken. (Holzinger et al. 2011) Rozpad tubulárního aparátu však můţeme chápat jako adaptaci na vysušení. Právě ztráta rigidity, kterou buňce tubulární aparát propůjčuje, umoţňuje buňce spolu s pruţností buněčných stěn a přepáţek reagovat na ztrátu vody celkovým smrštěním buňky. Tím se buňka vyvaruje protrţení protoplastu a dalším mechanickým poškozením. Nabízí se otázka, zda je moţné oddělit od sebe jevy probíhající v buňce následkem vysychání na prosté následky nedostatku vody a na funkční adaptace. Pravděpodobně ţádná takto ostrá hranice neexistuje. V průběhu evoluce se jednotlivé nutné reakce, zaloţené na fyzikální a chemické podstatě buněčných komponent, zapojily do více či méně sloţitých reakcí buňky na stres. Nám tak nezbývá, neţ chápat celý proces komplexně, a uvědomit si, ţe v buňce souvisí všechno se vším a stresová odolnost je výsledkem celé řady různě provázaných pochodů a reakcí, které na konci vedou k přeţití buňky, či nikoliv. Příkladem sloţitosti těchto dějů můţe být fotoinhibice, které se budu věnovat v další kapitole. 3.2 Fyziologické změny Veškeré fyziologické pochody v buňce jsou ovlivněny její celkovou kondicí. Pokud nejsou podmínky ideální, má to vliv na celkovou kondici buňky a následně to ovlivní fyziologické pochody v ní probíhající. Například pokles vzdušné vlhkosti sniţuje schopnost příjmu CO 2. (Bertsch 1966) Oxid uhličitý je pro fotoautotrofní organismy zdrojem uhlíku, který tvoří převáţnou část biomasy buňky. Pokles vzdušné vlhkosti tak nejen buňku vysušuje, ale navíc můţe zhoršit přístup ke zdroji základního stavebního kamene pro většinu syntetických procesů v buňce. Mezi tyto procesy patří také syntéza nových D1 proteinů pro fotosystém II. Funkce fotosystému II je úzce spjata s calvinovým cyklem. Pokud tedy dojde k omezení příjmu CO 2 sníţí se spotřeba ATP potaţmo NADPH. Nadbytek NADPH vede logicky k nedostatku NADP +. NADP + je příjemcem e -. 10
11 Pokud nemá buňka dostatečné mnoţství NADP + které by absorbovalo elektrony, tyto elektrony dále reagují za vzniku volných kyslíkových radikálů označovaných jako ROS. Mezi ROS patří i peroxid vodíku H 2 O 2, který působí inhibičně na syntézu prekurzorů proteinu D1. V důsledku vysychání tedy dochází k tvorbě volných kyslíkových radikálů, kterou nazýváme oxidativní stres a pro buňku je neţádoucí. Oxidativní stres vede k celé řadě poškození buněčných struktur a to včetně DNA. (Ron Mittler 2002) Výkon fotosystému II je daný rovnováhou mezi mnoţstvím světlem poškozených D1 proteinů a mnoţstvím těchto proteinů nově syntetizovaných. S poklesem mnoţství funkčních D1 proteinů se dále zpomaluje fotosyntéza, čímţ opět klesá mnoţství fixovaného CO 2, tato zpětnovazebná reakce vede nakonec k tomu, ţe fotosyntéza se poměrně rychle zastaví, dojde k tzv. fotoinhibici. Jde tedy o jakýsi regulační systém, který tlumí fotosyntézu v momentě, kdy mnoţství jejím prostřednictvím přijímané energie přesáhne schopnost buňky takové mnoţství energie zpracovat. Jako je tomu například při nedostatku vody. (Obr.2.) (Takahashi and Murata 2008) Je však třeba míti na zřeteli, ţe k fotoinhibici můţe dojít z mnoha dalších důvodů a mnoha jinými způsoby. Omezení fotosyntézy jako následek vysychání byl ukázán v pokusu na Klebsormidiu dissectum, kde byl měřen kvantový výtěţek fotosystému II během vysychání a během následné re-hydratace. Experiment byl rozdělen na několik částí. V první byla zkoumána odpověď na náhlé vysušení a následnou re-hydrataci, 3h vysoušení a 3h rehydratace. V další části schopnost regenerace fotosystému po týdenním a třítýdenním vysoušení při různých úrovních vzdušné vlhkosti (100%, 55%, 5%). Ve všech případech došlo k omezení či úplné inhibici fotosyntézy s různými parametry následné rekonvalescence. (Karsten, Holzinger 2011) 11
12 3.3 Dvě roviny odolnosti proti nedostatku vody Můţeme, a měli bychom, se snaţit rozlišit mezi dvěma typy tolerance k nedostatku vody. Tolerance k dehydrataci, tj. situaci, kdy se buňkám nedostává vody v dostatečném mnoţství a ty se snaţí jejím dalším ztrátám v maximální míře zabránit a k vysušení tj. úplné ztrátě vody. Organismy tolerantní k dehydrataci se snaţí za kaţdou cenu udrţet vnitřní homeostázi. Snaţí se tedy vyuţít kaţdé příleţitosti, aby se ztrátě vody vyhnuli. V přírodě to vypadá tak, ţe ze svých vláken formují mnohovrstevnaté koberce, kde blíţe povrchu nacházející se vrstvy chrání spodní vrstvy před světlem a brání nadměrnému odparu (Holzinger 2013). Podobně fungují i půdní krusty (Š.Chmelár 2011) Řasy si také vytváří slizové obaly a tlusté buněčné stěny. (K.Herburger 2015, A.Holzinger 2011, M.Pichrtová 2014) Vyvinuly se i funkční fyziologické odpovědí na úrovni regulace exprese jednotlivých genů. Které přispívají k udrţení homeostáze (P.Mulo 2012). Velice zajímavá je schopnost Coleochaete naprosto změnit svůj fenotyp, pokud je vystavena aeroterestrickým podmínkám. (Linda E. Graham, 2012). Elegantním způsobem jak se vyhnout ztrátě vody během nepříznivých podmínek je tvorba odolných stádií (oospory, pre-akinety, akinety), která dokáţí dlouhá období nedostatku vody přečkat. (M.O.Morison 1985) Tolerance k vysušení naopak znamená, ţe organismus je schopný vyschnout na úroveň 10% obsahu vody, ale i méně, a po re-hydrataci opět obnovit své fyziologické funkce. (Oliver et al., 2010) Tato schopnost není záleţitostí pouze řas. Tolerantní k vyschnutí jsou lišejníky, mechy (Roser et al. 1992), vyšší rostliny tzv. resurection plants. (G.Bianchi 1991), ale i ţivočichové např. ţelvušky, které přeţijí pokles vnitřního obsahu vody aţ na 2% původní hodnoty. (L.Rebecchi 2007, J.C.Wright 1988) 12
13 4 Význam stresové odolnosti streptofytních řas 4.1 Ekologický: půdní krusty - významný producent biomasy a fixátor dusíku a uhlíku, pionýrské sukcesní stádium, půdotvorný efekt Ekologický význam stresové odolnosti streptofytních řas spočívá v tom, ţe tato jim umoţňuje přeţít i na územích, která svými tvrdými podmínkami brání ve výskytu ostatním rostlinám. Tato houţevnatost jim dává nejen velice důleţitou roli v osidlování nových stanovišť, ale i významnou úlohu coby primárních producentů na takovýchto lokalitách. Řasy spolu se sinicemi, houbami a heterotrofními bakteriemi vytváří heterogenní společenstva, takzvané půdní krusty. (U. Karsten, A. Holzinger 2012, Š. Chmelár 2014) Tento typ agregace jim přináší mnohé výhody v podobě stabilnějšího vodního reţimu, vzájemného stínění a tedy ochrany proti UV záření či koloběhu prvků a metabolitů. Vzájemný vztah všech členů těchto společenstev můţeme označit za synergismus. (Malachová 2004) Další ekologický význam půdních krust, potaţmo tedy stresové odolnosti členů těchto společenstev patřících mezi streptofytní řasy, se zakládá na zpevnění nejsvrchnějších vrstev půd a tedy v omezení eroze. (Wynn-Williams 1990) Nezanedbatelným přínosem půdních krust coby raného sukcesního stádia, nebo s ohledem k lokálním podmínkám, jako stádia finálního, je fixace uhlíku - celosvětově 6% a dusíku - celosvětově 40%. Přesto, ţe jsou tato čísla významná, půdní krusty zůstávají dále velice málo prozkoumanými společenstvy. (Elster, Kováčik 2008) Ekologicky významná je také schopnost tvořit odolná vegetatvivní stádia, která se mohou šířit větrem a umoţňují šíření na nová stanoviště. (Holzinger 2009, Pichrtová 2014) 13
14 4.2 Evoluční: kolonizace souše, vznik evolučních novinek Přechod rostlin na souš je bezpochyby jednou z nejdůleţitějších událostí v evoluci ţivota na zemi. V kontextu posledních studií, dle kterých jsou nejbliţšími příbuznými vyšších rostlin právě streptofytní řasy (S. Wodniok 2011, Turmel et al. 2007), jeví se prozkoumání jejich ochranných mechanismů jako klíčové pro pochopení, způsobu jakým probíhala kolonizace souše rostlinami. Jako krásný příklad toho, ţe výzkumy na tomto poli mohou přinést světlo do dějů probíhajících před miliony let a přiblíţit nám dosud tajemný způsob přechodu rostlin na souš, nám poslouţí zajímavý objev, který učinila Linda E. Graham v roce 2012 při pokusu na Coleochaete. (Linda E. Graham, 2012) Pokus spočíval v kultivaci dvou druhů Coleochaete, která je povaţována za obligátně vodní řasu, ve vlhkých aeroterestrických podmínkách na minerálním médiu a na křemičitém písku. Takto kultivované Coleochaete orbicularis a scutata vykázaly na obou podkladech morfologii naprosto odlišnou od své vodní formy. V aeroterestrických podmínkách tvořily mnohovrstevné shluky buněk, jakési hromádky. Které na svém povrchu nesly jednovrstevné mnohobuněčné laloky. Obdoby primitivních lístečků s chemicky odolnými buněčnými stěnami. Celá tato struktura se nápadně podobá mikrofosilním záznamům záhadného předka z období paleozoika. Je tedy moţné, ţe právě za těmito fosilními nálezy se skrývají streptofytní předchůdci dnešních suchozemských rostlin. Na obrázku 3. můţete vidět rozdíl v morfologiích Coleochaete kultivované v aquatických a terestrických podmínkách. (Linda E. Graham 2012) Z uvedeného by mohlo vyplývat, ţe z evolučního hlediska byla významnější schopnost avoidance, neboli snaha uniknout vyschnutí, neţ tolerance k úplnému vyschnutí. Právě avoidance, jakoţto sloţitější řešení stresu z dehydratace, přináší vznik nových komplexnějších struktur odlišných funkcí. Ty mohli dále podléhat evolučnímu tlaku a být tak zdrojem evolučních novinek a mohli dát postupně vzniknout úplně novým formám ţivota. 14
15 5 Stresová odolnost jednotlivých skupin streptofytních řas V této kapitole se budu věnovat jednotlivým skupinám streptofytních řas, které se vţdy pokusím lehce charakterizovat. Následně uvedu mechanismy, které vyvinuli během evoluce v boji proti vysychání. 5.1 Chlorokybophyceae Charakteristika Chlorokybophyceae Zahrnují zatím jediný rod, Chlorokybus. Patří mezi jedny z morfologicky nejjednodušších streptofytních řas. Vyskytuje se ve vlhkých terestrických podmínkách a v půdě. Tato řasa je typická vytvářením malých sarcionoidních balíčků buněk, které jsou spojeny a obaleny slizovou pochvou. (Škaloud 2009) (Obr.4.) Rozmnoţují se asexuálně pomocí pohyblivých zoospor. ( Mechanismy stresové odolnosti u Chlorokybophyceae Právě tvorbu těchto balíčků a vytváření slizové pochvy můţeme povaţovat za způsob ochrany před vysycháním. (Rogers et al.,1980) Avšak ţádné studie, které by se zabývaly odolností rodu Chlorkybus před vysycháním zatím nebyly vypracovány. Pro konkrétnější závěry tak chybí potřebná data. 15
16 5.2 Klebsormidiophyceae Charakteristika rodu Klebsormidiophyceae Klebsormidium je celosvětově rozšířený rod s poměrně širokou ekologickou valencí. (D.Ryšánek a kol. 2015, Herburger 2015) Můţeme ho nalézt jak v řekách, potocích či rašeliništích, tak i na takových lokacích jako kůra stromů, skály, omítky nebo v půdách. Je pro něj typická vláknitá stélka tvořená buňkami s nástěnným chloroplastem a pyrenoidem. ( Mechanismy stresové odolnosti u Klebsormidiophyceae Klebsormidium Klebsormidium je pravděpodobně díky své kosmopolitnosti (U.Karsten, A. Holzinger 2012) jednou z nejtestovanějších řas v oblasti odolnosti vůči vysychání. Byla na něm provedena celá řada experimentů, které dobře dokumentují jeho schopnost přizpůsobit se těm nejdrsnějším podmínkám. Pro Klebsormidia je typická tvorba mnohovrstevnatých kobercovitých porostů. (Karsten, Holzinger 2012, Karsten et al. 2013) Takováto agregace umoţňuje ochranu spodnějších vrstev před nadměrným odparem vody a slunečním zářením. Tomu, ţe je Klebsormidium na takovouto strategii dobře adaptováno nasvědčují i poměrně nízké nároky na intenzitu světla pro efektivní fotosyntézu. Asi nejnápadnější reakcí na vysychání u Klebsormidia je scvrknutí buněk. A to výrazné, K. crenulatum se na příčné ose dokáţe smrštit z téměř 9,5µm na 5,9µm, coţ je zhruba 60% původní šířky. (Obr.5) (Holzinger 2011) K. dissectum se smrští dokonce i podélně. Po jednodenním vysoušení při 5% vzdušné vlhkosti se K. dissectum smrštilo v podélné ose na 65% a v příčné na 46% hodnot naměřených u kontrolního vzorku. Schopnost takového smrštění je u Klebsormidií dána zvláštním sloţením buněčných stěn a přepáţek. Ty neobsahují tolik celulózy a glykosidických vazeb jako tomu je u jiných embryofyt. Naopak obsahují větší mnoţství kalózy, coţ vysvětluje jejich obrovskou pruţnost. (Holzinger 2013) 16
17 Ţe jde v případě obsahu kalózy o jednoznačnou adaptaci na vysoušení ukazují nejnovější studie. Ty porovnávaly změnu mnoţství kalózy v buněčných stěnách u Zygnemy a Klebsormidia během vysoušecího experimentu. Ukázalo se, ţe Klebsormidium je schopné reagovat na vysoušení okamţitou syntézou kalózy. Zatímco u Zygnemy nic takového pozorováno nebylo. K. crenulatum vykázalo po 210 minutách více jak 200% přírůstek mnoţství kalózy. (Obr.6.) (Herburger, Holzinger 2015) Smrštění buňky v takovém rozsahu by ale nebylo moţné, pokud by zůstal zachovaný cytoskelet. Jeho rozpad byl také pozorován. (Obr.7.) (Holzinger et al. 2011) Nerozpadá se jen cytoskelet, rozpadají se i celá vlákna na jednotlivé buňky. Výhoda rozpadu na jednotlivé buňky je nejspíš ve snadnějším zapojení jednotlivých buněk do půdní krusty, která můţe poskytnout vhodný úkryt před prouděním vzduchu či osvitem. (Karsten 2012) Jakkoliv je takováto flexibilita úctyhodná a umoţní Klebsormidiím přeţít náhlou ztrátu vody, pro přeţití sezónních období nedostatku by to nestačilo. Přesto dokáţe Klebsormidium vydrţet celé týdny a měsíce. Vyvinulo totiţ celou řadu fyziologických adaptací. Mezi takové adaptace jistě patří vysoký osmotický potenciál cytosolu. Ten je v porovnání s ostatními zelenými řasami nezvykle vysoký. K plazmolýze u K. crenulatum došlo aţ při koncentraci roztoku sorbitolu 800mM. A u K. nittens při koncentraci 600mM. K tak silnému osmotickému potenciálu přispívá i, při vysychání zvýšená, produkce nízkomolekulárních sacharidů a oligosacharidů jako jsou seróza, glukóza, fruktóza, xylóza, galaktóza, manóza, inositol, glycerol, mannitol a sorbitol. Tak silný osmotický potenciál však nemůţe být dán pouze obsahem těchto cukrů, neboť těch je po vysušení pouhých 1,2% sušiny. Co je 17
18 příčinou takto silného osmotického potenciálu by mělo být předmětem dalších studií. (Kaplan 2012) Vysoký osmotický potenciál má také K. rivulare, které po 4 týdením vysoušení vykazuje aţ 350% nárůst osmolitů. Po této době došlo také k tvorbě tzv. akinet. Buněk, které mají aţ čtyřnásobně ztlustlé buněčné stěny, vysoký obsah škrobových zrn a zásobu lipidů. Akinety mají redukované vakuoly a menší mnoţství chlorofylu. Jsou schopné přečkat dlouhá nepříznivá období, po jejichţ uplynutí z nich opět vyrostou vláknité stélky. (M. Morison 1985, Nagao 2008) Interfilum Další pokusy byly provedeny na Interfilu, které je dalším ze zástupců Klebsormidiophyceae. Všechny testované kmeny vykázaly postupný pokles měřeného kvantového výtěţku fotosyntézy s následnou inhibicí. Lišily se však v časech nástupu těchto jevů. Velký rozdíl byl mezi kmeny vyskytujícími se v podobě solitérních buněk a kmeny tvořícími agregované shluky. U samostatně se vyskytujících buněk došlo k inhibici poměrně záhy. Zatímco shluky buněk odolávaly déle. To ukazuje, ţe agregace buněk je efektivní obranou proti vysychání. (Karsten. 2014) Hormidiella, Entransia Další méně běţní zástupci klebsormidiophyceae. Těmto rodům dosud příliš velká pozornost věnována nebyla. Je to nejspíše tím, ţe se nevyskytují tak často jako dva předešlé. Přesto i tyto řasy byly podrobeny výzkumu efektivity fotosyntézy při různých stresových zátěţích. Oba rody jsou dobře přizpůsobeny podmínkám, ve kterých se vyskytují. 18
19 Entransia je řasa vyskytující se v šeru limnetických zón. Z toho také pramení větší efektivita vyuţití světla a naopak méně pruţná reakce na přebytek světla. Rekonvalescence Entransie po vysušení a následné re-hydrataci byla sice horší, neţ u sesterské Hormidielly, ale lepší, neţ u jiných sladkovodních řas. Hormidiella je řasa vyskytující se v půdách vysychajících stanovišť. Jako taková je tedy citlivější na změny osvitu a teploty a reaguje na ně pruţněji, tj. podobně jako ostatní aeroterestrické řasy. V porovnání s Entransií ji vysušení ovlivnilo méně. Po rehydrataci vykazovala fotosyntéza stejné hodnoty jako před vysušením. (Herburger 2015) 19
20 5.3 Charophyceae Charakteristika rodu Charophyceae Zástupci řadící se do řádu charophyceae, česky Parožnatky ţijí většinou v čistých stojatých a především sladkovodních vodách. Jejich stélka patří k těm nejsloţitějším ze streptopfytních řas, je pletivná a dělená na články a uzliny. K podkladu je přichycená tzv. rhyzoidy. Růst zajišťuje jedna vrcholová buňka tzv. terminála, která odděluje vţdy jednu článkovou buňku a buňku uzliny. Článková buňka roste pouze do délky a bývá dlouhá i několik cm. Buňka uzliny se dále dělí a vytváří tak boční větvě v přeslenu. Buněčná stěna je často inkrustována vápníkem. Charophyceae se rozmnoţují pohlavně i nepohlavně. Pohlavní rozmnoţování je oogamní. Nepohlavní rozmnoţování je umoţněno fragmentací stélky, odnoţováním či přítomností speciálních tělísek, která se nacházejí mezi rhyzoidy. ( Mechanismy stresové odolnosti u Charophyceae Jediné odolné stádium v ţivotním cyklu Parožnatek je oospora. Ta je často inkrustována uhličitanem vápenatým. Takto inkrustované schránky jsou velice estetické a jsou nazývány gyrogonity. (Obr.8.) Jsou uzpůsobeny k tomu, aby přeţily anoxické prostředí na dně jezera, vysušení i pozření ptákem, jehoţ trusem jsou následně šířeny. Oospory jsou schopné přeţít v sedimentu několik let a v příznivých podmínkách opět vyklíčit. 20
21 5.4 Coleochaetophyceae Charakteristika rodu Coleochaetophyceae Coleochaete je vláknitá řasa jejíţ stélka je tvořena heterotrichálními vlákny, která se sdruţují v disk. Rozmnoţují se pohlavně i nepohlavně. K nepohlavnímu rozmnoţování dochází fragmentací stélky, nebo zoosporami. Pohlavní rozmnoţování je oogamie. Při pohlavním rozmnoţování vzniká zygospora, která je tvořena zygotou obklopenou sterilními vlákny. ( Mechanismy stresové odolnosti u Coleochaetophyceae Z těchto vláken vznikne časem korovitá vrstva zajišťující ochranu před vnějším stresem. Coleochaete tvoří tzv. algaenan, sporopoleninu podobnou látku, která chrání zygosporu před UV zářením a vyschnutím. (Delwiche et al. 1989) Coleochaete vystavená aeroterestrickým podmínkám, v laboratoři například na agarovém médiu, nebo křemičitém písku, vykazuje morfologii naprosto odlišnou od stélky, kterou tvoří ve vodě. Místo diskovité stélky tvořené jednou vrstvou buněk, vzniklou stočením bohatě větvených vláken, vytvoří mnohovrstevnaté shluky buněk. Tyto hromádky mají na svém povrchu laloky tvořené jednou vrstvou buněk připomínající primitivní lístečky. (Obr.9.) Buňky této aeroterestricé formy mají ztlustlé, chemicky odolné buněčné stěny. (L.Graham 2012) 21
22 5.5 Zygnematophyceae Charakteristika rodu Zygnematophyceae Třída Zygnematophyceae je čistě sladkovodní skupina kokálních a vláknitých řas. Dělíme je na Zygnematales a Desmidiales, jejichţ zástupce nalezneme spíše ve vodách s niţším ph. Zygnematales tvoří většinou nevětvenou vláknitou stélku, jejíţ fragmentace slouţí k nepohlavnímu rozmnoţování. Buňky ve vláknu jsou odděleny septem, jehoţ středový otvor je zakryt fragmoplastem. Pohlavní rozmnoţování se nazývá konjugace. Ke konjugaci můţe dojít mezi dvěma buňkami různých vláken (tzv.skalariformní konjugace), nebo mezi sousedními buňkami téhoţ vlákna (konjugace laterální). Při konjugaci vystupují jako gamety protoplasty sterilních buněk, které se amébovitě pohybují ve vymezeném prostoru. Po jejich splynutí dochází ke karyogamii, tím vznikne odolná zygospora obalená tlustou buněčnou stěnou. Po přečkání nepříznivých podmínek dochází v zygospoře k meiotickému dělení a vyklíčení. Desmidiales jsou jednobuněčné řasy, ţijící samostatně či v koloniích. Jejich buňky jsou uprostřed rozděleny zářezem na dvě tzv. semicely. Nepohlavní rozmnoţování probíhá rozestoupením semicel, v prostoru vzniklém mezi nimi dojde k rozdělení jádra. Dceřiná jádra se oddělí septem a nově vzniklé buňky si následně dotvoří druhou semicelu. Pohlavní rozmnoţování je opět konjugace. Dvě buňky se k sobě přiblíţí, obklopí se slizovým obalem. Následně se poloviny buněk rozestoupí a dojde ke splynutí protoplastů. Vzniklá zygota se opět obalí a vzniká odolná zygospora. ( 22
23 Mechanismy stresové odolnosti u Zygnematophyceae Zygnematales Podobně jako někteří zástupci Klebsormidiophyceae tak i mezi Zygnematophyceae jsou řasy, které vytváří mnohovrstevné kobercovité nárosty, kde svrchní vrstvy stíní vrstvy spodní a vytváří jim tak přívětivější podmínky k ţivotu. O tom, ţe je takové řešení účinné svědčí měření na Spyrogiře, které provedli Berri a Lembi. Při svém experimentu měřili úroveň záření dopadajícího na svrchní stínící vrstvu a poté úroveň záření pod touto vrstvou. Úroveň záření v zastíněné oblasti byla 30x niţší. Kdyţ byl poté měřen výkon fotosyntézy, byl naměřen vyšší v zastíněné oblasti. Z toho plyne, ţe tato nízká úroveň záření je pro průběh fotosyntézy optimální. (Berri a Lembi 2000) Zygnema je taktéţ adaptována na nízkou úroveň záření a i ona vytváří kobercovité porosty. (Herburger et al. 2015) Zygogonium ericetorum dokonce vytváří speciální typ buněk bohatších na pigment, které se vyskytují ve svrchních vrstvách a lépe tak absorbují sluneční záření, čímţ chrání nejen sebe, ale také účinněji stíní spodní vrstvy. (A. Holzinger et al. 2010) Výše zmiňovaná strategie bývá označována jako tzv. self-protection. Další způsob jak se bránit uschnutí, který můţeme pozorovat u Zygnematophyceae je přečkání nepříznivých podmínek v podobě nějakého odolného vytrvalého stádia. Často je to nějakým způsobem specializovaná buňka. U Zygnematophyceae jich je hned několik. První z nich je zygospora, která vzniká pohlavním rozmnoţováním. Buněčná stěna zygospory má tři vrstvy, přičemţ pro stresovou odolnost je klíčová prostřední vrstva obsahující algaenan, coţ je sporopoleninu podobná látka zajišťující odolnost proti vyschnutí a UV záření. (L.Graham 2012) Další specializované buňky, které však nevznikají sexuálním rozmnoţováním, ale z vegetativních buněk jsou pre-akinety. (Obr.11) Charakteristické jsou sekundárně ztlustlou buněčnou stěnou, zvýšeným mnoţstvím škrobových zrn a lipidů, redukovaným chloroplastem. (McLean 1971, Holzinger et al. 2011, Herburger et al. 2015) Vznik pre-akinet není indukován ani tak nedostatkem vody jako spíše nedostatkem dusíku. S rostoucím stářím populace přibývá i pre-akinet, coţ můţe být dáno i postupným vyčerpáváním zdrojů. Pokud jsou pre-akinety při své genezy vystaveny 23
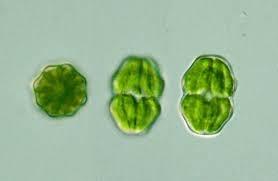
24 mírnému vysychání, jsou schopny se na takovou zátěţ adaptovat. Takzvaně se otuţí. Otuţilé pre-akinety jsou pak schopny snést intenzivní vysoušení při 10% relativní vzdušné vlhkosti. Pre-akinety jsou pro přeţití Zygnemy v drsných podmínkách klíčové, neboť zygospory v těţkých podmínkách vznikají jen zřídka. Výhodou pre-akinet je, ţe nejde o spící stádia, takţe mohou v případě návratu příznivých podmínek nastartovat své fyziologické funkce a dát vzniknout novým vláknům. Po rozpadu vláken mohou být také roznášeny větrem, či ptáky, coţ napomáhá šíření. (Pichrtová 2014) Desmidiales O schopnosti desmidiales čelit vysychání toho dosud příliš napsáno nebylo. I přesto jsou některé takové případy evidovány. Konkrétně Staurastrum habeebense, jehoţ vegetativní stádia mají ztlustlé buněčné stěny, příznivý poměr mezi plochou buňky a jejím oběmem a produkují sliz. (Obr.10) Tak dokáţí přeţít bez vody i několik měsíců. (J. Šťastný 2008) 24
25 6 Shrnutí U streptofytních řas bylo pozorováno velké mnoţství adaptací a mechanismů, které řasám umoţňují vyrovnat se s mnohdy drsným prostředím v místě jejich výskytu. Veškeré pozorované mechanismy stresové odolnosti bych rozdělil do tří základních skupin. V první skupině je prevence, snaha organismu vyhnout se ztrátě vody. Tak činí například agregací buněk a tvorbou slizového sekretu. Nebo vzájemným stíněním, ať uţ jde jen o shluk jednobuněčných řas nebo tvorbu kobercových nárostů řas vláknitých, kde vrchní vrstva stíní vrstvy spodní. Často pozorované sekundární ztlustnutí buněčné stěny je také preventivním mechanismem ochrany proti vyschnutí. Zapojení buněk do půdní krusty, či rozpad vlákna na jednotlivé buňky usnadní ukrytí do stínu. Do mechanismů preventivních bych zahrnul i tvorbu odolných stádií, jako jsou zygospory, oospory, akinety, či pre-akinety. Druhá skupina mechanismů se dá nazvat adaptace na vysychání. Mezi tyto mechanismy patří třeba schopnost Klebsormidia velice rychle zareagovat na ztrátu vody produkcí kalózy, která zajistí pruţnost buněčných stěn a umoţní tak buňce smrštění potaţmo zachování vnitřních struktur nepoškozených. Patří sem také produkce osmoticky aktivních látek, které zvýší osmotický potenciál vnitřního prostředí buňky a tím v něm udrţí větší mnoţství vody, která zabrání denaturaci biomolekul a membrán. Schopnost regulovat úroveň fotosyntézy zcela jistě přispívá udrţení stabilního vodního reţimu buňky a brání poškození buněčného aparátu kyslíkovými radikály. Posledním mechanismem jsou opravy a následná rekonvalescence. Mnoţství nutných oprav závisí na tvrdosti podmínek, kterým byla řasa vystavena, a na účinnosti obranných mechanismů vyjmenovaných výše. Čím lépe je řasa adaptována, ať uţ svou evoluční historií, či jen tím, ţe se otuţila během mírnějšího stresu, tím rychleji se zotavuje. Jinak strukturovaný přehled příčin vysychání a moţných odpovědí na ně nám poskytne (Obr.12) z jednoho staršího review. 25
26 7 Seznam použité literatury 1. S.Wodniok, H.Brinkmann, G.Glockner, A.J.Heidel, H.Philippe, M.Melkonian a B.Becker (2011) Origin o fland plants: Do conjugating algae hold the key? BMC Evolutionary Biology 2. L.E.Graham, P.Arancibia-Avila, W.A.Taylor, P.K.Strother a M.E.Cook (2012) Aeroterrestrial Coleochaete (Streptophyta, Coleochaetales) Models early plant adaptation to land, American Journal of Botany 99(1): Š.Chmelár (2011) Mikrobiální půdní aktivity v půdách spontánně vznikajících na obnaţené hornině, Diplomová práce 4. David D. Wynn-Wiliams (1990) Microbial colonization processes in antarctic fellfield soils-an experimental overview, Proc. NIPR Symp. Polar Biology, 3, K.Malachová (2005) Ekologie mikroorganizmů. Ostrava: Ostravská univerzita, 86 s. 6. J.Elster, P.Degma, L.Kováčik, L.Valentová, K.Šrámková a A.B.Pereira (2008) Freezing and desiccation resistence in the filamentous green alga Klebsormidium from the Antarctic, Arctic and Slovakia, Biologia 63/6: U.Karsten, A.Holzinger (2012) Light, temperature, and desiccation effects on photosyntetic aktivity, and drought-induced ultrastructural changes in The green alga Klebsormidium dissectum (Streptophyta) from a high alpine soil crust 8. A.Bertsch (1966) CO2 Gaswechsel der Grunalge Apatococcus lobatus, Planta 70, A.Holzinger (2009) Desiccation tolerance on green algae: Implications of Physiological adaptation and structural requirements 10. Jan de Vries et al. (2016) Streptophyte Terrestrialization in Light of Plastid Evolution 11. Rogers,C.E., Mattox, K. R., and Stewart, K. D. (1980) The zoospore of Chloroky- bus atmophyticus, a charophyte with sarcinoid growth habit. Am.J.Bot. 67: Ryšánek, D., Hrčková, K. and Škaloud, P. (2015), Global ubiquity and local endemism of free-living terrestrial protists: phylogeographic assessment of the streptophyte alga Klebsormidium. Environ Microbiol, 17: Mulo, P., Sakurai, I., and Aro, E. M. (2012). Strategies for psba gene expression in cyanobacteria, green algae and higher plants: from tran- scription to PSII repair. Biochim. Biophys. Acta 1817, Pichrtová (2014) Stress resistance of polar hydro-terrestrial algae Zygnema spp. (Zygnematophyceae, Streptophyta) 15. D. J. Roser, D.R. Melick, H.U. Ling and R.D. Seppelt (1992) Australian Polyol and sugar content of terrestrial plants from continental Antarctica. Antarctic Sci F.Kaplan, L.Lewis, J.Wastian et al. (2011) Plasmolysis effects and osmotic potential of two phylogenetically distinct alpine strains of Klebsormidium (Streptophyta). 17. B.Fuller (2004) Cryoprotectants : The essential antifreezes to protect life in the frozen state 26
27 18. L.Rebecchi, T Altiero, L Guidetti (2007) Anhydrobiosis: the extreme limit of desiccation tolerance 19. F.Kaplan, L.Lewis, K.Herburger (2012) Osmotic stress in Arctic and Antarctic strains of the green alga Zygnema (Zygnematales, Streptophyta): Effects on photosynthesis and ultrastructure. 20. A.Holzinger, U.Karsten (2013) Desiccation stress and tolerance in green algae:consequences for ultrastructure, physiological and molecular mechanisms 21. Holzinger A., Lütz C., and Karsten U. (2011). Desiccation stress causes structural and ultra-structural alterations in the aeroterrestrial green alga Klebsormidium crenulatum (Klebsormidiophyceae, Streptophyta) isolated from an alpine soil crust. J. Phycol. 47, Elster, Degma, Kováčik (2008), Freezing and desiccation injury resistance in the filamentous green alga Klebsormidium from the Antarctic, Arctic and Slovakia, Biologia 63/6: S.Agrawal (2009) Factors affecting spore germination in algae - review. Folia Microbiol. 54 (4), U.Karsten, C. Lütz. A. Holzinger (2010) Ecophysiological performance of the aeroterrestrial green alga Klebsormidium crenulatum (Charophyceae, Streptophyta) isolated from an Alpine soil crust with an emphasis on desiccation stress 25. Ron Mittler (2002) Oxidative stress, antioxidants and stress tolerance, Trends in Plant Science, Volume 7, Issue 9, K.Herburger, A. Holzinger (2015) Localization and Quantification of Callose in the Streptophyte Green Algae Zygnema and Klebsormidium: Correlation with Desiccation Tolerance 27. M. Nagao, K. Matsui a M. Uemura (2008), Klebsormidium flaccidum, a charophycean green alga, exhibits cold acclimation that is closely associated with compatible solute accumulation and ultrastructural changes. Plant, Cell & Environment, 31: U. Karsten, K. Herburger, A. Holzinger (2014) Dehydration, temperature, and light tolerance in members of the aeroterrestrial green algal genus Interfilum (Streptophyta) from biogeographically different temperate soils 29. K. Herburger, A. Holzinger, U. Karsten (2015) Entransia and Hormidiella, sister lineages of Klebsormidium (Streptophyta), respond differently to light, temperature, and desiccation stress 30. Delwiche, Graham, Thomson (1989) Lignin-like compounds and sporopollenin in Coleochaete, an algal model for land plant ancestry 31. Škaloud (2009) Species composition and diversity of aero-terrestrial algae and cyanobacteria of the Boreč Hill ventaroles, Fottea 9(1): U. Karsten, F. Rindi (2010) Ecophysiological performance of an urban strain of the aeroterrestrial green alga Klebsormidium sp. (Klebsormidiales, Klebsormidiophyceae) 27
28 33. Berri a Lembi (2000) Effects of temperature and irradiance on the seasonal variation of a Spirogyra (Chlorophyta) population in a Midwestern Lake (U.S.A.) Journal of Phycology 36(5): K. Herburger, L. Lewis, A. Holzinger (2015) Photosynthetic efficiency, desiccation tolerance and ultrastructure in two phylogenetically distinct strains of alpine Zygnema sp. (Zygnematophyceae, Streptophyta): Role of pre-akinete formation, Protoplasma 35. A. Holzinger, A. Tschnaikner, D. Remias (2010) Cytoarchitecture of the desiccationtolerant green alga Zygogonium ericetorum. Protoplasma (2010) 243: Obecné charakteristiky ze stránek, 28

 29")
29 8 Obrazová příloha Obr. 1 (Leliaert et al. 2012) Obr. 2 (S.Takahashi 2008) 29


30 Obr. 3 (L.Graham 2012) Obr. 4 Chlorokybus atmophyticus (foto Pavel Škaloud, 30
 algal filaments observed in immersion oil in the")
 (Holzinger et al. 2011) Obr.")
31 Obr. 5 (c) One-day desiccated (5% relative air humidity [RH]) algal filaments observed in immersion oil in the light microscope; note the shrinkage of the cell diameter (arrows) (Holzinger et al. 2011) Obr. 6 (Herburger 2015) 31
 Obr.")

32 Obr. 7 (Holzinger et al. 2011) Obr. 8 Gyrogonity ( 32

33 Obr. 9 (L.Graham 2012) Obr. 10 Staurastrum habeebense ( 33

34 Obr. 11(Herburger 2015) 34

35 Obr. 12 (Holzinger 2013) 35
ZYGNEMATOPHYCEAE spájivky
 ZYGNEMATOPHYCEAE spájivky Zvláštní způsob pohlavního rozmnožování, spájení neboli konjugace, dal název této třídě oddělení Chlorophyta. Při spájení se mění celé protoplasty buněk v gamety a kopulují párovitě.
ZYGNEMATOPHYCEAE spájivky Zvláštní způsob pohlavního rozmnožování, spájení neboli konjugace, dal název této třídě oddělení Chlorophyta. Při spájení se mění celé protoplasty buněk v gamety a kopulují párovitě.
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu
 TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu
 TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
TŘÍDA: ZYGNEMATOPHYCEAE Sladkovodní skupina vzhledově velmi estetických řas, jednobuněčných nebo vláknitých. V žádné fázi svého životního cyklu netvoří bičíkatá stádia. Mají specifický způsob pohlavního
Kde v systému se nacházíme? Impérium: Eukaryota Říše: Rostliny - Plantae Podříše: Viridiplantae - zelené rostliny
 Leliaert et al. 2011 Kde v systému se nacházíme? Impérium: Eukaryota Říše: Rostliny - Plantae Podříše: Viridiplantae - zelené rostliny dvě vývojové linie - Chlorophytae a Streptophytae mechorosty a cévnaté
Leliaert et al. 2011 Kde v systému se nacházíme? Impérium: Eukaryota Říše: Rostliny - Plantae Podříše: Viridiplantae - zelené rostliny dvě vývojové linie - Chlorophytae a Streptophytae mechorosty a cévnaté
Barbora Chattová. Fylogeneze a diverzita řas a hub: 5. přednáška Charophyta
 Barbora Chattová Fylogeneze a diverzita řas a hub: 5. přednáška Charophyta Oddělení Charophyta, třídy Mesostigmatophyceae Klebsormidiophyceae Coleochaetophyceae Charophyceae Zygnematophyceae Vývojová větev
Barbora Chattová Fylogeneze a diverzita řas a hub: 5. přednáška Charophyta Oddělení Charophyta, třídy Mesostigmatophyceae Klebsormidiophyceae Coleochaetophyceae Charophyceae Zygnematophyceae Vývojová větev
Kde v systému se nacházíme?
 Kde v systému se nacházíme? Impérium: Eukaryota Říše: Rostliny - Plantae Podříše: Viridiplantae - zelené rostliny vývojová linie - Streptophytae Třída: Charophyceae - parožnatky Chara, parožnatka Tolypella
Kde v systému se nacházíme? Impérium: Eukaryota Říše: Rostliny - Plantae Podříše: Viridiplantae - zelené rostliny vývojová linie - Streptophytae Třída: Charophyceae - parožnatky Chara, parožnatka Tolypella
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA. CHLOROPHYTA zelené řasy
 HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
HYDROBOTANIKA CHLOROPHYTA zelené řasy ODDĚLENÍ: CHLOROPHYTA - zelené řasy Zelené řasy jsou velice široká skupina. Její příslušníci mají všechny druhy stélek, jen rhizopodiální typ se vyskytuje pouze jako
Botanika bezcévných rostlin 1. praktické cvičení
 Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů "říše" Plantae; podříše Viridiplantae Oddělení: Chlorophyta; třída: Ulvophyceae a) Ulva b) Cladophora c) Anadyomene d) Valonia
Botanika bezcévných rostlin 6. praktické cvičení Přehled pozorovaných objektů "říše" Plantae; podříše Viridiplantae Oddělení: Chlorophyta; třída: Ulvophyceae a) Ulva b) Cladophora c) Anadyomene d) Valonia
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162
 ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
ZŠ Určeno pro Sekce Předmět Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 Téma / kapitola Dělnická 6. 7. třídy ZŠ základní
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
ZELENÉ ŘASY (CHLOROPHYTA)
") ZELENÉ ŘASY (CHLOROPHYTA) metodický list Zelené řasy lze rozdělit dle tříd, toto rozdělení je ale pro žáky málo srozumitelné a navíc systém není jednotný. Volila jsem proto rozdělení podle typů stélky.
ZELENÉ ŘASY (CHLOROPHYTA) metodický list Zelené řasy lze rozdělit dle tříd, toto rozdělení je ale pro žáky málo srozumitelné a navíc systém není jednotný. Volila jsem proto rozdělení podle typů stélky.
Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů
 Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae ŘÁD: Bangiales Porphyra (HB) TŘÍDA: Florideophyceae ŘÁD: Gelidiales Gelidium (HB) ŘÁD: Gigartinales
Botanika - bezcévné rostliny 6. praktikum Přehled pozorovaných objektů ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae ŘÁD: Bangiales Porphyra (HB) TŘÍDA: Florideophyceae ŘÁD: Gelidiales Gelidium (HB) ŘÁD: Gigartinales
Botanika bezcévných rostlin pro učitele 1. praktické cvičení
 Botanika bezcévných rostlin pro učitele 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
Botanika bezcévných rostlin pro učitele 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíl praktického cvičení: na konkrétním materiálu se seznámit s reprezentativními zástupci nejdůležitějších systematických
ZYGNEMATOPHYCEAE spájivky
 ZYGNEMATOPHYCEAE spájivky TŘÍDA: ZYGNEMATOPHYCEAE - spájivé řasy, spájivky Zvláštní způsob pohlavního rozmnožování, spájení neboli konjugace, dal název této třídě oddělení Chlorophyta. Při spájení se mění
ZYGNEMATOPHYCEAE spájivky TŘÍDA: ZYGNEMATOPHYCEAE - spájivé řasy, spájivky Zvláštní způsob pohlavního rozmnožování, spájení neboli konjugace, dal název této třídě oddělení Chlorophyta. Při spájení se mění
Třída: Chlorophyceae - Chlamydomonadales
 Leliaert et al. 2011 Třída: Chlorophyceae Relationships among the five monophyletic groups of the Chlorophyceae as inferred from 44 chloroplast protein-coding genes of 11 chlorophytes and nine streptophytes.
Leliaert et al. 2011 Třída: Chlorophyceae Relationships among the five monophyletic groups of the Chlorophyceae as inferred from 44 chloroplast protein-coding genes of 11 chlorophytes and nine streptophytes.
Buňka buňka je základní stavební a funkční jednotka živých organismů
 Buňka - buňka je základní stavební a funkční jednotka živých organismů - je pozorovatelná pouze pod mikroskopem - na Zemi existuje několik typů buněk: 1. buňky bez jádra (prokaryotní buňky)- bakterie a
Buňka - buňka je základní stavební a funkční jednotka živých organismů - je pozorovatelná pouze pod mikroskopem - na Zemi existuje několik typů buněk: 1. buňky bez jádra (prokaryotní buňky)- bakterie a
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
ORGANISMY A SYSTÉM ŘASY A MECHOROSTY
 Zelené řasy Zelené řasy je významné oddělení jednobuněčných i mnohobuněčných stélkatých zelených rostlin. Představují blízké příbuzné vyšších rostlin, které se z jedné linie zelených řas vyvinuly. Stavba-
Zelené řasy Zelené řasy je významné oddělení jednobuněčných i mnohobuněčných stélkatých zelených rostlin. Představují blízké příbuzné vyšších rostlin, které se z jedné linie zelených řas vyvinuly. Stavba-
Číslo a název projektu Číslo a název šablony
 Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Fotosyntéza (2/34) = fotosyntetická asimilace
 = fotosyntetická asimilace") Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Glaucophyta TŘÍDA: Glaucophyceae Glaucocystis (PP) ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae Porphyridium
Botanika bezcévných rostlin 9. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Glaucophyta TŘÍDA: Glaucophyceae Glaucocystis (PP) ODDĚLENÍ: Rhodophyta TŘÍDA: Bangiophyceae Porphyridium
Botanika bezcévných rostlin pro učitele 5. praktické cvičení
 Botanika bezcévných rostlin pro učitele 5. praktické cvičení ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae Chlamydomonas
Botanika bezcévných rostlin pro učitele 5. praktické cvičení ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae Chlamydomonas
Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů
 Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae
Botanika bezcévných rostlin 10. praktické cvičení Přehled pozorovaných objektů ŘÍŠE: Plantae ODDĚLENÍ: Chlorophyta TŘÍDA: Trebouxiophyceae Chlorella (PP) Trebouxia (PP) Stichococcus (PP) TŘÍDA: Chlorophyceae
Třída: XANTHOPHYCEAE
 HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
HYDROBOTANIKA Třída: XANTHOPHYCEAE Celková chrakteristika Asi 600 druhů v 90 rodech. Dříve byla tato skupina nazývána Heterokontae, neboli různobrvky, dnes bývá nazývána v některých moderních textech Tribophyceae
Buňka. Autor: Mgr. Jitka Mašková Datum: Gymnázium, Třeboň, Na Sadech 308
 Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie
 synekologie b) autekologie c) demekologie") 1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Fluorescence chlorofylu
 Pro připomenutí Fluorescence chlorofylu Princip Fotochemické a nefotochemické zhášení fluorescence Excitace chlorofylu: plantphys.info Analýza zhášení (quenching analysis) Temnostní adaptace Kautského
Pro připomenutí Fluorescence chlorofylu Princip Fotochemické a nefotochemické zhášení fluorescence Excitace chlorofylu: plantphys.info Analýza zhášení (quenching analysis) Temnostní adaptace Kautského
Název: Fotosyntéza, buněčné dýchání
 Název: Fotosyntéza, buněčné dýchání Výukové materiály Autor: Mgr. Blanka Machová Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: Biologie, chemie Ročník: 2. Tematický
Název: Fotosyntéza, buněčné dýchání Výukové materiály Autor: Mgr. Blanka Machová Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: Biologie, chemie Ročník: 2. Tematický
Zelené řasy (Chlorophyta) Milan Dundr
 Milan Dundr") Zelené řasy (Chlorophyta) Milan Dundr Barviva barviva v chloroplastech: chlorofyl a chlorofyl b -karoten xantofyly Chlorofyl a, chlorofyl b zelené řasy zásobní látka: škrob často pyrenoid bílkovinné tělísko
Zelené řasy (Chlorophyta) Milan Dundr Barviva barviva v chloroplastech: chlorofyl a chlorofyl b -karoten xantofyly Chlorofyl a, chlorofyl b zelené řasy zásobní látka: škrob často pyrenoid bílkovinné tělísko
Biologie - Kvinta, 1. ročník
 - Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
- Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY
 ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY Řasy (dříve nesprávně označovány jako podříše nižších rostlin v rámci rostlinné říše) představují velmi různorodou skupinu organismů od několika mikrometrů velkých
ŘASY PRACOVNÍ LIST PRO STŘEDNÍ ŠKOLY Řasy (dříve nesprávně označovány jako podříše nižších rostlin v rámci rostlinné říše) představují velmi různorodou skupinu organismů od několika mikrometrů velkých
1- Úvod do fotosyntézy
 1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
Vylepšování fotosyntézy
 Vylepšování fotosyntézy Využití fotosyntézy potraviny energie (paliva) Obojího bude podle predikcí potřebovat lidstvo čím dál tím víc. Energetické využití fotosyntézy potřeba nahrazení fosilních paliv
Vylepšování fotosyntézy Využití fotosyntézy potraviny energie (paliva) Obojího bude podle predikcí potřebovat lidstvo čím dál tím víc. Energetické využití fotosyntézy potřeba nahrazení fosilních paliv
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka
 primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
primární producenti: řasy, sinice, vodní rostliny konkurence o zdroje mikrobiální smyčka přirozená jezera (ledovcová, tektonická, ) tůně rybníky přehradní nádrže umělé tůně (lomy, pískovny) Dělení stojatých
Třída Xanthophyceae tváří se jak zelené ale jsou hnědé
 Třída Xanthophyceae tváří se jak zelené ale jsou hnědé RNDr. Lenka Šejnohová, Ph.D. Oddělení experimentální fykologie a ekotoxikologie Botanický ústav Akademie věd ČR, Brno Centrum pro cyanobakterie a
Třída Xanthophyceae tváří se jak zelené ale jsou hnědé RNDr. Lenka Šejnohová, Ph.D. Oddělení experimentální fykologie a ekotoxikologie Botanický ústav Akademie věd ČR, Brno Centrum pro cyanobakterie a
OBSAH 1 ÚVOD... 7. 1.1 Výrobek a materiál... 7 1.2 Přehled a klasifikace materiálů pro výrobu... 8 2 ZDROJE DŘEVA... 13
 OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
OBSAH 1 ÚVOD................................................. 7 1.1 Výrobek a materiál........................................ 7 1.2 Přehled a klasifikace materiálů pro výrobu..................... 8 2
Základy biologie a ekologie VZNIK A VÝVOJ ŽIVOTA
 Základy biologie a ekologie VZNIK A VÝVOJ ŽIVOTA Výsledky vzdělávání Učivo Ţák Základy biologie charakterizuje názory na vznik a vývoj vznik a vývoj ţivota na Zemi ţivota na Zemi, porovná délku vývoje
Základy biologie a ekologie VZNIK A VÝVOJ ŽIVOTA Výsledky vzdělávání Učivo Ţák Základy biologie charakterizuje názory na vznik a vývoj vznik a vývoj ţivota na Zemi ţivota na Zemi, porovná délku vývoje
Řasy. Zvyšování kvality výuky v přírodních a technických oblastech CZ.1.07/1.128/02.0055. (laboratorní práce) Označení: EU-Inovace-Př-6-04
 Označení: EU-Inovace-Př-6-04") Zvyšování kvality výuky v přírodních a technických oblastech CZ.1.07/1.128/02.0055 Řasy (laboratorní práce) Označení: EU-Inovace-Př-6-04 Předmět: přírodopis Cílová skupina: 6. třída Autor: Mgr. Miroslav
Zvyšování kvality výuky v přírodních a technických oblastech CZ.1.07/1.128/02.0055 Řasy (laboratorní práce) Označení: EU-Inovace-Př-6-04 Předmět: přírodopis Cílová skupina: 6. třída Autor: Mgr. Miroslav
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Buňka. Kristýna Obhlídalová 7.A
 Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
VY_32_INOVACE_02.08 1/12 3.2.02.8 Sinice, lišejníky, řasy Sinice modrozelené organismy
 1/12 3.2.02.8 Sinice modrozelené organismy cíl - popsat stavbu těla sinic - vyjmenovat příklad sinic - chápat nebezpečí sinic pro člověka - patří k nejstarším organismům na Zemi - stavba podobná bakterii,
1/12 3.2.02.8 Sinice modrozelené organismy cíl - popsat stavbu těla sinic - vyjmenovat příklad sinic - chápat nebezpečí sinic pro člověka - patří k nejstarším organismům na Zemi - stavba podobná bakterii,
Ekosystém. tok energie toky prvků biogeochemické cykly
 Ekosystém tok energie toky prvků biogeochemické cykly Ekosystém se sestává z abiotického prostředí a biotické složky (společenstva) a jejich vzájemných interakcí. Ekosystém si geograficky můžeme definovat
Ekosystém tok energie toky prvků biogeochemické cykly Ekosystém se sestává z abiotického prostředí a biotické složky (společenstva) a jejich vzájemných interakcí. Ekosystém si geograficky můžeme definovat
KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc.
 KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc. Úvod do problematiky Fytoplankton=hlavní producent biomasy, na kterém
KYSLÍKOVÉ DEFICITY - PROJEV NESTABILITY RYBNIČNÍHO EKOSYSTÉMU? Ing. Ivana Beděrková Ing. Zdeňka Benedová doc. RNDr. Libor Pechar, CSc. Úvod do problematiky Fytoplankton=hlavní producent biomasy, na kterém
Vyšší rostliny Embryophyta. Milan Štech, PřF JU
 Vyšší rostliny Embryophyta Milan Štech, PřF JU = suchozemské rostliny Embryophyta * mechorosty * cévnaté rostliny * jejich společní předci/přímí předchůdci Vznik chloroplastu klíčová událost na cestě k
Vyšší rostliny Embryophyta Milan Štech, PřF JU = suchozemské rostliny Embryophyta * mechorosty * cévnaté rostliny * jejich společní předci/přímí předchůdci Vznik chloroplastu klíčová událost na cestě k
- pro učitele - na procvičení a upevnění probírané látky - prezentace
 Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
BYTOVÝ DŮM: Mimoňská , Praha 9
 BYTOVÝ DŮM: Mimoňská 633-643, Praha 9 Počet stran: 10 Vypracovala: Ing. Michaela Jakubičková Str. 1 17.10.2017 Vypracovala: Ing. Michaela Jakubičková Obsah 1. ÚVOD... 3 1.1 PŘÍČINA RŮSTU ŘAS A PLÍSNÍ...
BYTOVÝ DŮM: Mimoňská 633-643, Praha 9 Počet stran: 10 Vypracovala: Ing. Michaela Jakubičková Str. 1 17.10.2017 Vypracovala: Ing. Michaela Jakubičková Obsah 1. ÚVOD... 3 1.1 PŘÍČINA RŮSTU ŘAS A PLÍSNÍ...
LES Ročník: 6. Vzdělávací oblast.: Člověk a příroda Vzdělávací obor: Přírodopis
 VY_52_INOVACE_04 LES Ročník: 6. Vzdělávací oblast.: Člověk a příroda Vzdělávací obor: Přírodopis Základní škola a Mateřská škola Nikolčice, příspěvková organizace Petr Chalupný Anotace Materiál obsahuje
VY_52_INOVACE_04 LES Ročník: 6. Vzdělávací oblast.: Člověk a příroda Vzdělávací obor: Přírodopis Základní škola a Mateřská škola Nikolčice, příspěvková organizace Petr Chalupný Anotace Materiál obsahuje
Buňka cytologie. Buňka. Autor: Katka www.nasprtej.cz Téma: buňka stavba Ročník: 1.
 Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Mnohobuněčné kvasinky
 Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Vodní režim rostlin. Obsah vody, RWC, vodní potenciál a jeho komponenty: Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy,
 Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Rozmnožování hub. Typy hniloby dřeva. Hlenky. Mechy. Lišejníky. Řasy
 Rozmnožování hub Ostatní organizmy Dřevokazné houby - stopkovýtrusné Rozmnožování organizmů, které se řadí k houbám, je velmi variabilní a značně složité. Stopkovýtrusné houby, které jsou i níže uvedené
Rozmnožování hub Ostatní organizmy Dřevokazné houby - stopkovýtrusné Rozmnožování organizmů, které se řadí k houbám, je velmi variabilní a značně složité. Stopkovýtrusné houby, které jsou i níže uvedené
Fotosyntéza Ekofyziologie. Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni
 Fotosyntéza Ekofyziologie Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Fyziologické a ekologické aspekty fotosyntézy vliv stresů a proměnného prostředí na fotosyntézu; mechanismy
Fotosyntéza Ekofyziologie Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Fyziologické a ekologické aspekty fotosyntézy vliv stresů a proměnného prostředí na fotosyntézu; mechanismy
FOTOSYNTÉZA. Princip, jednotlivé fáze
 FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje. 26.2.2010 Mgr.
 Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje 26.2.2010 Mgr. Petra Siřínková ABIOTICKÉ PODMÍNKY ŽIVOTA SLUNEČNÍ ZÁŘENÍ TEPLO VZDUCH VODA PŮDA SLUNEČNÍ
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje 26.2.2010 Mgr. Petra Siřínková ABIOTICKÉ PODMÍNKY ŽIVOTA SLUNEČNÍ ZÁŘENÍ TEPLO VZDUCH VODA PŮDA SLUNEČNÍ
Číslo materiálu: VY 32 INOVACE 18/08
 Číslo materiálu: Název materiálu: PÍSEMNÉ OPKOVCÍ TESTY 1.pololetí Číslo projektu: CZ.1.07/1.4.00/21.1486 Zpracovala: Marcela Kováříková 1. Co je symbioza. Jaký je rozdíl mezi symbiozou a parazitismem.
Číslo materiálu: Název materiálu: PÍSEMNÉ OPKOVCÍ TESTY 1.pololetí Číslo projektu: CZ.1.07/1.4.00/21.1486 Zpracovala: Marcela Kováříková 1. Co je symbioza. Jaký je rozdíl mezi symbiozou a parazitismem.
Základy buněčné biologie
 Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
EKOLOGICKÉ PRINCIPY A ZÁKONITOSTI
 EKOLOGICKÉ PRINCIPY A ZÁKONITOSTI Alena Ševců Centrum pro nanomateriály, pokročilé technologie a inovace, alena.sevcu@tul.cz, tel. 485 353 786 Obsah: Ekologie jako věda. Základní ekologické pojmy a principy.
EKOLOGICKÉ PRINCIPY A ZÁKONITOSTI Alena Ševců Centrum pro nanomateriály, pokročilé technologie a inovace, alena.sevcu@tul.cz, tel. 485 353 786 Obsah: Ekologie jako věda. Základní ekologické pojmy a principy.
a) zkonzumují za život velké množství jedinců, avšak nespotřebují jedince celého, nezpůsobují jeho smrt, i když mu svou aktivitou škodí
 zkonzumují za život velké množství jedinců, avšak nespotřebují jedince celého, nezpůsobují jeho smrt, i když mu svou aktivitou škodí") 1. Praví predátoři: a) zkonzumují za život velké množství jedinců, avšak nespotřebují jedince celého, nezpůsobují jeho smrt, i když mu svou aktivitou škodí b) konzumují část kořisti, kořist zpravidla neusmrtí,
1. Praví predátoři: a) zkonzumují za život velké množství jedinců, avšak nespotřebují jedince celého, nezpůsobují jeho smrt, i když mu svou aktivitou škodí b) konzumují část kořisti, kořist zpravidla neusmrtí,
PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY
 PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY 2010 Ing. Andrea Sikorová, Ph.D. 1 Problémy životního prostředí - organismy V této kapitole se dozvíte: Co je to organismus. Z čeho se organismus skládá. Jak se dělí
PROBLÉMY ŽIVOTNÍHO PROSTŘEDÍ ORGANISMY 2010 Ing. Andrea Sikorová, Ph.D. 1 Problémy životního prostředí - organismy V této kapitole se dozvíte: Co je to organismus. Z čeho se organismus skládá. Jak se dělí
FOTOSYNTÉZA Správná odpověď:
 FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva
 a kokální stélkou. Pohyb zajišťují dva") CHLAMYDOPHYCEAE TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva až čtyři stejně dlouhé bičíky vyrůstající
CHLAMYDOPHYCEAE TŘÍDA: CHLAMYDOPHYCEAE Třída obsahuje jednotlivě žijící i cenobiální bičíkovce, řasy s kapsální (gleomorfní) a kokální stélkou. Pohyb zajišťují dva až čtyři stejně dlouhé bičíky vyrůstající
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková
 Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ
 Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíle praktického cvičení z Botaniky bezcévných rostlin: - na konkrétním materiálu se seznámit s reprezentativními zástupci
Botanika bezcévných rostlin 1. praktické cvičení INFORMACE O ORGANIZACI CVIČENÍ cíle praktického cvičení z Botaniky bezcévných rostlin: - na konkrétním materiálu se seznámit s reprezentativními zástupci
Zemědělství a ochrana přírody a krajiny. Roman Scharf odbor obecné ochrany přírody a krajiny MŢP
 Zemědělství a ochrana přírody a krajiny Roman Scharf odbor obecné ochrany přírody a krajiny MŢP Vývoj krajiny ve vztahu k zemědělství Do konce 19. století rozvoj osídlení včetně horských a podhorských
Zemědělství a ochrana přírody a krajiny Roman Scharf odbor obecné ochrany přírody a krajiny MŢP Vývoj krajiny ve vztahu k zemědělství Do konce 19. století rozvoj osídlení včetně horských a podhorských
Pedagogická činnost pro jmenovací řízení
 Pedagogická činnost pro jmenovací řízení Jiří Neustupa Pedagogická činnost Bakalářský studijní program Biologie (studijní obory Biologie, Ekologická a evoluční biologie, Molekulární biologie a biochemie
Pedagogická činnost pro jmenovací řízení Jiří Neustupa Pedagogická činnost Bakalářský studijní program Biologie (studijní obory Biologie, Ekologická a evoluční biologie, Molekulární biologie a biochemie
prokaryotní Znaky prokaryoty
 prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
aneb Fluorescence chlorofylu jako indikátor stresu
 Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
Vyjádření fotosyntézy základními rovnicemi
 FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
Fotosyntéza. Ondřej Prášil
 Fotosyntéza 2 Ondřej Prášil prasil@alga.cz 384-340430 Obsah přednášky membrány a organely světlo termodynamika historie Fotosyntetické membrány Electron tomography Cells contain ~100 chlorosomes appressed
Fotosyntéza 2 Ondřej Prášil prasil@alga.cz 384-340430 Obsah přednášky membrány a organely světlo termodynamika historie Fotosyntetické membrány Electron tomography Cells contain ~100 chlorosomes appressed
VY_32_INOVACE_ / Prvoci Prvoci jednobuněční živočichové
 1/7 3.2.02.9 jednobuněční živočichové cíl - popsat stavbu, tvar, pohyb, výskyt a rozmnožování prvoků - uvést zástupce - jednobuněční živočichové, tvoří je jedna buňka, která vykonává všechny životní funkce
1/7 3.2.02.9 jednobuněční živočichové cíl - popsat stavbu, tvar, pohyb, výskyt a rozmnožování prvoků - uvést zástupce - jednobuněční živočichové, tvoří je jedna buňka, která vykonává všechny životní funkce
Buňka. základní stavební jednotka organismů
 Buňka základní stavební jednotka organismů Buňka Buňka je základní stavební a funkční jednotka těl organizmů. Toto se netýká virů (z lat. virus jed, je drobný vnitrobuněčný cizopasník nacházející se na
Buňka základní stavební jednotka organismů Buňka Buňka je základní stavební a funkční jednotka těl organizmů. Toto se netýká virů (z lat. virus jed, je drobný vnitrobuněčný cizopasník nacházející se na
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
Využití zásoby živin a primární produkce v eutrofních rybnících
 Využití zásoby živin a primární produkce v eutrofních rybnících Libor Pechar a kolektiv Jihočeská Univerzita v Českých Budějovicích Zemědělská fakulta, Laboratoř aplikované ekologie a ENKI o.p.s., Třeboň
Využití zásoby živin a primární produkce v eutrofních rybnících Libor Pechar a kolektiv Jihočeská Univerzita v Českých Budějovicích Zemědělská fakulta, Laboratoř aplikované ekologie a ENKI o.p.s., Třeboň
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
VESMÍR. za počátek vesmíru považujeme velký třesk před 13,7 miliardami let. dochází k obrovskému uvolnění energie, která se rozpíná
 VESMÍR za počátek vesmíru považujeme velký třesk před 13,7 miliardami let dochází k obrovskému uvolnění energie, která se rozpíná vznikají první atomy, jako první se tvoří atomy vodíku HVĚZDY první hvězdy
VESMÍR za počátek vesmíru považujeme velký třesk před 13,7 miliardami let dochází k obrovskému uvolnění energie, která se rozpíná vznikají první atomy, jako první se tvoří atomy vodíku HVĚZDY první hvězdy
Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky.
 Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky. Materiál je plně funkční pouze s použitím internetu. základní projevy života
Anotace: Materiál je určen k výuce přírodopisu v 6. ročníku ZŠ. Seznamuje žáky se základní stavbou rostlinné a živočišné buňky. Materiál je plně funkční pouze s použitím internetu. základní projevy života
EFFECT OF CADMIUM ON TOBACCO CELL SUSPENSION BY-2
 EFFECT OF CADMIUM ON TOBACCO CELL SUSPENSION BY-2 Štěpán Z., Klemš M., Zítka O., Havel L. Department of Plant Biology, Faculty of Agronomy, Mendel University in Brno, Zemědělská 1, 613 00 Brno, Czech Republic
EFFECT OF CADMIUM ON TOBACCO CELL SUSPENSION BY-2 Štěpán Z., Klemš M., Zítka O., Havel L. Department of Plant Biology, Faculty of Agronomy, Mendel University in Brno, Zemědělská 1, 613 00 Brno, Czech Republic
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
1. Ekologie zabývající se studiem společenstev se nazývá a) autekologie b) demekologie c) synekologie
 autekologie b) demekologie c) synekologie") 1. Ekologie zabývající se studiem společenstev se nazývá a) autekologie b) demekologie c) synekologie 2. Obor ekologie lesa se zabývá zejména: a) vzájemnými vztahy organismů s prostředím a mezi sebou b)
1. Ekologie zabývající se studiem společenstev se nazývá a) autekologie b) demekologie c) synekologie 2. Obor ekologie lesa se zabývá zejména: a) vzájemnými vztahy organismů s prostředím a mezi sebou b)
Pražská vysoká škola psychosociálních studií
 Pražská vysoká škola psychosociálních studií Stres u mužů v pomáhajících profesích a jeho zvládání pomocí extrémních sportů Martin Balej Vedoucí práce: MUDr. Olga Dostálová, CSc. Praha 2014 Prague college
Pražská vysoká škola psychosociálních studií Stres u mužů v pomáhajících profesích a jeho zvládání pomocí extrémních sportů Martin Balej Vedoucí práce: MUDr. Olga Dostálová, CSc. Praha 2014 Prague college
Saprofité-rozklad org. zbytků Paraziticky- mykosy... Symbioticky- s cévnatými rostlinami(mykorhiza)- 95% rostlinných druhů, rostlina poskytuje
- 95% rostlinných druhů, rostlina poskytuje") Otázka: Houby a nižší rostliny Předmět: Biologie Přidal(a): LenkaKrchova Houby fungia Samostatná říše- napůl živočich a rostlina Eukaryotické heterotrofní organismy, které se rozmnožují výtrusy. Tělo se
Otázka: Houby a nižší rostliny Předmět: Biologie Přidal(a): LenkaKrchova Houby fungia Samostatná říše- napůl živočich a rostlina Eukaryotické heterotrofní organismy, které se rozmnožují výtrusy. Tělo se
Třída: Chlorophyceae - Chlamydomonadales
 Leliaert et al. 2011 Třída: Chlorophyceae Relationships among the five monophyletic groups of the Chlorophyceae as inferred from 44 chloroplast protein-coding genes of 11 chlorophytes and nine streptophytes.
Leliaert et al. 2011 Třída: Chlorophyceae Relationships among the five monophyletic groups of the Chlorophyceae as inferred from 44 chloroplast protein-coding genes of 11 chlorophytes and nine streptophytes.
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.)
") CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
Ekologie a její obory, vztahy mezi organismy a prostředím
 Variace 1 Ekologie a její obory, vztahy mezi organismy a prostředím Autor: Mgr. Jaromír JUŘEK Kopírování a jakékoliv další využití výukového materiálu je povoleno pouze s uvedením odkazu na www.jarjurek.cz.
Variace 1 Ekologie a její obory, vztahy mezi organismy a prostředím Autor: Mgr. Jaromír JUŘEK Kopírování a jakékoliv další využití výukového materiálu je povoleno pouze s uvedením odkazu na www.jarjurek.cz.
BIOLOGIE OCEÁNŮ A MOŘÍ
 BIOLOGIE OCEÁNŮ A MOŘÍ 1. ekologické faktory prostředí světlo salinita, hustota, tlak teplota obsah rozpuštěných látek a plynů 2 1.1 sluneční světlo ubývání světla do hloubky odraz světla od vodní hladiny,
BIOLOGIE OCEÁNŮ A MOŘÍ 1. ekologické faktory prostředí světlo salinita, hustota, tlak teplota obsah rozpuštěných látek a plynů 2 1.1 sluneční světlo ubývání světla do hloubky odraz světla od vodní hladiny,
trubicovitá pletivná vláknitá, větvená vláknitá
 ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
ŘASY METODICKÝ LIST PRO UČITELE (STŘEDNÍ ŠKOLY) řešení doplňující otázky/úkolu z pracovního listu doplňující informace k tomu, co žáci uvidí v mikroskopu a je vhodné je na to upozornit doplňující informace,
Jsme zelené a kulaté, ale přesto nejsme Chlor(ella)ococcales kdo jsme? tváří se jak zelené ale jsou hnědé. RNDr. Lenka Šejnohová, Ph.D.
ococcales kdo jsme? tváří se jak zelené ale jsou hnědé. RNDr. Lenka Šejnohová, Ph.D.") Děkuji za pozorn Jsme zelené a kulaté, ale přesto nejsme Chlor(ella)ococcales kdo jsme? tváří se jak zelené ale jsou hnědé RNDr. Lenka Šejnohová, Ph.D. Mikrobiologický ústav AV ČR, Třeboň Botanický ústav
Děkuji za pozorn Jsme zelené a kulaté, ale přesto nejsme Chlor(ella)ococcales kdo jsme? tváří se jak zelené ale jsou hnědé RNDr. Lenka Šejnohová, Ph.D. Mikrobiologický ústav AV ČR, Třeboň Botanický ústav
Téma: MORFOLOGIE ŢIVOČIŠNÝCH BUNĚK
 Téma: MORFOLOGIE ŢIVOČIŠNÝCH BUNĚK ŢIVÉ SOUSTAVY Nebuňečné (priony, viroidy, viry) Buněčné (jedno- i mnohobuněčné organismy) PROKARYOTICKÝ TYP BUNĚK 1-10 µm Archebakterie Eubakterie (bakterie a sinice)
Téma: MORFOLOGIE ŢIVOČIŠNÝCH BUNĚK ŢIVÉ SOUSTAVY Nebuňečné (priony, viroidy, viry) Buněčné (jedno- i mnohobuněčné organismy) PROKARYOTICKÝ TYP BUNĚK 1-10 µm Archebakterie Eubakterie (bakterie a sinice)
 Evoluce (nejen) rostlinné buňky Martin Potocký laboratoř buněčné biologie ÚEB AV ČR, v.v.i. potocky@ueb.cas.cz http://www.ueb.cas.cz Evoluce rostlinné buňky Vznik a evoluce eukaryotních organismů strom
Evoluce (nejen) rostlinné buňky Martin Potocký laboratoř buněčné biologie ÚEB AV ČR, v.v.i. potocky@ueb.cas.cz http://www.ueb.cas.cz Evoluce rostlinné buňky Vznik a evoluce eukaryotních organismů strom
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody
 VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody Čtvrté skupenství vody: Hexagonální voda: Na univerzitě ve Washingtonu bylo objeveno čtvrté skupenství vody, což může vysvětlit
VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody Čtvrté skupenství vody: Hexagonální voda: Na univerzitě ve Washingtonu bylo objeveno čtvrté skupenství vody, což může vysvětlit
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo