Typy fylogenetických analýz
|
|
|
- Radomír Mareš
- před 6 lety
- Počet zobrazení:
Transkript

1 Typy fylogenetických analýz Distanční metody: Neighbor-Joining Minimum Evolultion UPGMA,... Maximum Likelihood Bayesian Inference Maximum Parsimony
 substituční modely odhady jednotlivých substitučních rychlostí pomocí Markovových modelů + frekvence výskytu")
2 Genetické distance, substituční modely pro výpočet fylogenetických analýz je nutné stanovit genetické (evoluční) distance mezi sekvencemi v případě molekulárních hodin je distance přímo úměrná času p-distance: prostý rozdíl sekvencí výrazné podhodnocení reálných distancí (saturace) substituční modely odhady jednotlivých substitučních rychlostí pomocí Markovových modelů + frekvence výskytu nukleotidů = Q matice
 Kimura (K80): jiné substituční rychlosti pro transice a transverze, shodné frekvence bází (nst=2) Hasegawa-Kishino-Yano (HKY): jiné")
3 Substituční modely Jukes-Cantor (JC69): během evoluce mají všechny nukleotidy stejnou pravděpodobnost substitucí i stejnou frekvenci výskytu bází (nst=1) Felsenstein (F81): nukleotidy mají stejnou pravděpodobnost substitucí ale jinou frekvenci výskytu bází (nst=1) Kimura (K80): jiné substituční rychlosti pro transice a transverze, shodné frekvence bází (nst=2) Hasegawa-Kishino-Yano (HKY): jiné substituční rychlosti pro transice a transverze, různé frekvence bází (nst=2) General time reverdible (GTR): pravděpodobnosti substitucí a frekvence bází jsou specifikovány pro každou možnost (nst=6)
: existence velkého množství nevariabilních")
: modeluje variabilitu v míře nukleotidových")
4 Substituční modely Gamma distribuce (Γ): modeluje variabilitu v míře nukleotidových substitucí na různých pozicích alignmentu. Většinou se model zjednodušuje do 4 α kategorií Proporce nevariabilních míst (I): existence velkého množství nevariabilních pozic negativně ovlivňuje odhad genetických distancí. Aplikace I modelu je např. důležitá při současné přítomnosti krátkých a dlouhých větví Kovarion (cov): modeluje variabilitu v míře nukleotidových substitucí v závislosti na fylogenetické pozici dané sekvence

5 Test topologie - bootstrapping výpočet stromů na základě nově generovaných alignmentů konstrukce majority-rule konsenzuálního stromu hodnoty bootstrapů je nutné zobrazit na topologii stromu zkonstruovaného na základě originálního alignmentu

 BioNJ: lepší přesnost topologie u vzdáleně")
6 Fylogenetické analýzy na základě distančních matic stromy se počítají na základě distancí, spočtených pro každou dvojici sekvencí aplikují se substituční modely vhodné pro rychlou analýzu velkého množství sekvencí (v řádu několika stovek až tisíc) Minimum Evolution (ME): hledá se strom o nejmenším součtu délek všech větví Neighbor Joining (NJ): heuristický algoritmus na rychlé nalezení ME stromu (začíná se u hvězdicovitého stromu) BioNJ: lepší přesnost topologie u vzdáleně příbuzných sekvencí
7 Maximum Likelihood (ML) hledání nejpravděpodobnějšího stromu odrážejícího evoluci sekvencí posuzování pravděpodobností nekonečně velkého množství stromů (různé topologie, délky větví, parametrů substitučních modelů,...) heuristické metody pro hledání struktury stromu: Nearest Neighbor Interchange (NNI) Subtree Pruning + Regrafting (SPR) Tree-Bisection + Reconnection (TBR)

, které ale mohou mít různou pravděpodobnost tzv. priors uniform priors stejná pravděpodobnost, žádné předpoklady (topologie) exponencial priors např.")
 lognormal priors např.")
8 Bayesova analýza (BI) tradiční otázka: pokud je v košíku stejně modrých a červených kuliček, jaká je pravděpodobnost, že si vytáhnu 3 modré a 3 červené kuličky? Bayesovská otázka: pokud si vytáhnu 3 modré a 3 červené kuličky, jaká je pravděpodobnost, že je v košíku stejně modrých a červených kuliček? podmíněná pravděpodobnost (posterior) existuje nekonečné množství předpokladů (víc modrých, víc červených,...), které ale mohou mít různou pravděpodobnost tzv. priors uniform priors stejná pravděpodobnost, žádné předpoklady (topologie) exponencial priors např. délky větví (likelihood je negativní exponenciální funkcí) dirichlet priors pravděpodobnosti oscilují okolo dané hodnoty (frekvence bází, substituční modely, I,...) lognormal priors např. kalibrace stromu fosilními daty priors se během analýzy mění na základě analyzovaných dat (alignment sekvencí), pomocí stochastických modelů získáme posteriorní pravděpodobnosti


9 Bayesova analýza (BI) Markov chain Monte Carlo (MCMC) sampling výpočet posteriorních pravděpodobností pomocí náhodných změn prior parametrů a ty buď zamítnout či přijmout na základě jejich pravděpodobností Metropolic coupling MCMC = (MC) 3 1 studený a 3 horké řetězce burn-in: odstranění iniciální fáze MCMC výsledná topologie: konsenzuální strom posteriorních topologií
10 Maximální parsimonie (MP) hledání stromu s co nejmenším počtem evolučních kroků heuristické prohledávání stromů stejné jako u ML: NNI, SPR, TBR Fitchův algoritmus: vážená parsimonie (wmp): MP výpočet parsimonního skóre: 1 pro substituci, 0 pro žádnou změnu wmp výpočet: každý typ substituce je vážen např. pomocí frekvence jeho výskytu (rescaled consistency index - méně častým mutacím je dáno vyšší skóre)
11 Long branch attraction Archezoa
12 Long branch attraction Archezoa
13 Long branch attraction
14 Long branch attraction
15 kontaminace, špatná identifikace,...
16 base composition bias sekvence jsou shlukovány podle velkého GC obsahu
17 použití špatných markerů
18 špatná bootstrapová podpora
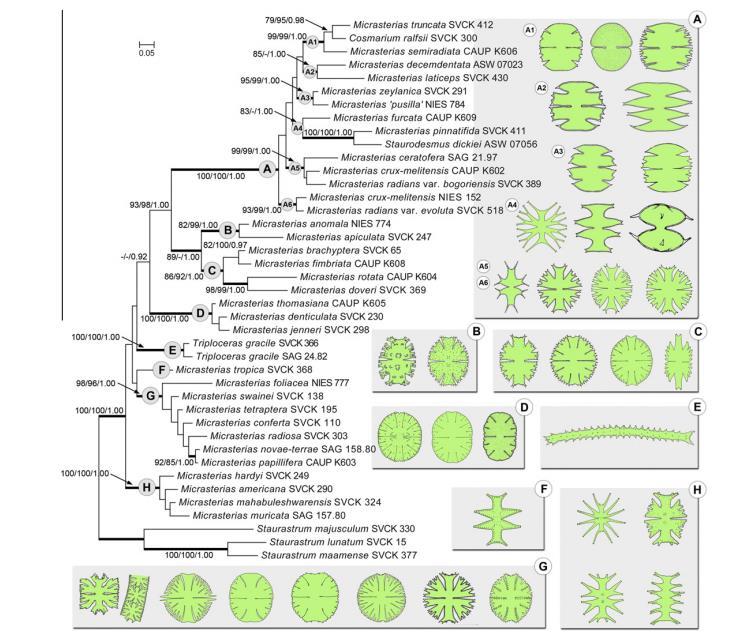
19

20 konflikt genů Cladonia
21 konflikt genů Cladonia

22 konflikt genů Xanthidium
23 Phylogeny at the level of populations and species Species are lineages A B C A B C >> Species phylogeny Population genetics: coalescence process (Wright-Fisher process)




24 Phylogeny at the level of populations and species Barriers to gene flow coalescence process : individual / allele copy
25 Phylogeny at the level of populations and species A B C Time T2 T1
26 Phylogeny at the level of populations and species A B C A B C T2 T1 deep coalescence = MRCA incomplete lineage sorting

27
28



29 NGS Environmentální sekvenování nyní více jak SSU rdna sekvencí v databázi GenBank Cuvelier et al




30 NGS




31 NGS L litter O organic A A horizon B B horizon P pine MH mixed hardwood

32 NGS

33 NGS Ekologická struktura linií 8 freshwater 31 marine 12 anoxic 2 deep-sea

34
35 Skryté pasti environmentálního sekvenování Objev 8 nových říší
36 Skryté pasti environmentálního sekvenování Nedostatečný taxon sampling


37 Skryté pasti environmentálního sekvenování Nedostatečný taxon sampling publikované určení sekvencí reanalyzované určení sekvencí


38 Skryté pasti environmentálního sekvenování Chimerické sekvence nedoběhnutá elongace krátké úseky DNA se stávají primery vznik chiméry
39 Lucking, 2014, pyrosequencing
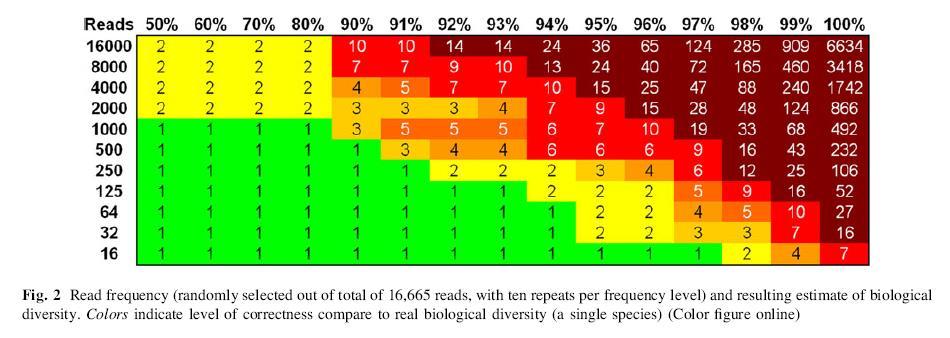
40
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,
: Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,") Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů)
") Počítačováčást 1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů) Pavel Munclinger, Petr Synek 2. fylogenetická
Počítačováčást 1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů) Pavel Munclinger, Petr Synek 2. fylogenetická
Modelování různé substituční rychlosti v různých pozicích alignmentu
 MOLEKULÁRNÍ TAXONOMIE - 7 (2018) Modelování různé substituční rychlosti v různých pozicích alignmentu U reálných dat lze očekávat, že různé pozice alignmentu budou substituovat s různou rychlostí aktivní
MOLEKULÁRNÍ TAXONOMIE - 7 (2018) Modelování různé substituční rychlosti v různých pozicích alignmentu U reálných dat lze očekávat, že různé pozice alignmentu budou substituovat s různou rychlostí aktivní
matematika v biologii: fylogenetika David Černý
 matematika v biologii: fylogenetika David Černý David Černý Úvod Linné (1735), Systema Naturae: živá příroda vykazuje hierarchické uspořádání Darwin (1859), On the Origin of Species: příčinou je společný
matematika v biologii: fylogenetika David Černý David Černý Úvod Linné (1735), Systema Naturae: živá příroda vykazuje hierarchické uspořádání Darwin (1859), On the Origin of Species: příčinou je společný
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Základy fylogenetiky a konstrukce fylogenetických stromů
 EKO/MEM Molekulární ekologie mikroorganismů Základy fylogenetiky a konstrukce fylogenetických stromů Iva Buriánková Katedra ekologie PřF UP Kde vyrostl první fylogenetický strom? Charles Darwin (1809 1882)
EKO/MEM Molekulární ekologie mikroorganismů Základy fylogenetiky a konstrukce fylogenetických stromů Iva Buriánková Katedra ekologie PřF UP Kde vyrostl první fylogenetický strom? Charles Darwin (1809 1882)
Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak)
") MOLEKULÁRNÍ TAXONOMIE - 6 (2015) Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak) Nyní si ukážeme obecný princip, jakým se obohacují substituční modely o parametry tak,
MOLEKULÁRNÍ TAXONOMIE - 6 (2015) Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak) Nyní si ukážeme obecný princip, jakým se obohacují substituční modely o parametry tak,
2. Maximální úspornost (Maximum Parsimony, MP)
") 2. Maximální úspornost (Maximum Parsimony, MP) Ze všech metod konstrukce fylogenetických stromů byly donedávna nejpoužívanější metody maximální úspornosti (parsimonie). Důvodem pro jejich mimořádnou oblibu
2. Maximální úspornost (Maximum Parsimony, MP) Ze všech metod konstrukce fylogenetických stromů byly donedávna nejpoužívanější metody maximální úspornosti (parsimonie). Důvodem pro jejich mimořádnou oblibu
MOLEKULÁRNÍ TAXONOMIE 9
 MOLEKULÁRNÍ TAXONOMIE 9 Zdokonalování substitučního modelu V předchozích přednáškách jsme si představili metodu maximum likelihood, která počítá s pravděpodobnostmi substitucí na větvích a topologiích.
MOLEKULÁRNÍ TAXONOMIE 9 Zdokonalování substitučního modelu V předchozích přednáškách jsme si představili metodu maximum likelihood, která počítá s pravděpodobnostmi substitucí na větvích a topologiích.
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
MOLEKULÁRNÍ TAXONOMIE 10
 MOLEKULÁRNÍ TAXONOMIE 10 Molekulární hodiny Skutečnost, že počet substitučních událostí vzrůstá s časem, vedla velmi brzy ke snahám využít sekvencí k datování stáří uzlů na fylogenetických stromech. Jako
MOLEKULÁRNÍ TAXONOMIE 10 Molekulární hodiny Skutečnost, že počet substitučních událostí vzrůstá s časem, vedla velmi brzy ke snahám využít sekvencí k datování stáří uzlů na fylogenetických stromech. Jako
Pravděpodobnost vs. Poměr šancí. Pravděpodobnostní algoritmy: Bayesova věta. Bayesova teorie rozhodování. Bayesova věta (teorém) Vzorec. ...
 Vzorec. ...") ravděpodobnostní algoritmy: Bayesova věta Fantasy is hardly an escape from reality. It is a way of understanding it. LLoyd Alexander ravděpodobnost vs. oměr šancí ravděpodobnost - poměr počtu jedinců surčitým
ravděpodobnostní algoritmy: Bayesova věta Fantasy is hardly an escape from reality. It is a way of understanding it. LLoyd Alexander ravděpodobnost vs. oměr šancí ravděpodobnost - poměr počtu jedinců surčitým
Pravděpodobnost, náhoda, kostky
 Pravděpodobnost, náhoda, kostky Radek Pelánek IV122, jaro 2015 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností krátké
Pravděpodobnost, náhoda, kostky Radek Pelánek IV122, jaro 2015 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností krátké
cluster clusters cluster cluster hierarchické klastrování: => strom je jedním z grafických znázornění hierarchického klastrování:
 Fylogenetika klastrovat na základě podobnosti lze cokoliv, vůbec nemusí jít o evoluční záležitosti (= lidstvo potřebuje popisovat a škatulkovat), je to obecný matematický základ pro analýzy dat, data mining,
Fylogenetika klastrovat na základě podobnosti lze cokoliv, vůbec nemusí jít o evoluční záležitosti (= lidstvo potřebuje popisovat a škatulkovat), je to obecný matematický základ pro analýzy dat, data mining,
Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany
: Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany") Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŢENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŢENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
STATISTICKÉ VYHODNOCENÍ FYLOGENEZE BIOLOGICKÝCH SEKVENCÍ
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Využití DNA sekvencování v
 Využití DNA sekvencování v taxonomii prokaryot Mgr. Pavla Holochová, doc. RNDr. Ivo Sedláček, CSc. Česká sbírka mikroorganismů Ústav experimentální biologie Přírodovědecká fakulta Masarykova univerzita,
Využití DNA sekvencování v taxonomii prokaryot Mgr. Pavla Holochová, doc. RNDr. Ivo Sedláček, CSc. Česká sbírka mikroorganismů Ústav experimentální biologie Přírodovědecká fakulta Masarykova univerzita,
BOOTSTRAPPINGOVÉ METODY VE FYLOGENETICE
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele)
") Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).
: RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).") Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Coalesce spojit se, splynout, sloučit se. Didaktická simulace Coalescence = splynutí linií
 Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Bayesovské metody. Mnohorozměrná analýza dat
 Mnohorozměrná analýza dat Podmíněná pravděpodobnost Definice: Uvažujme náhodné jevy A a B takové, že P(B) > 0. Podmíněnou pravěpodobností jevu A za podmínky, že nastal jev B, nazýváme podíl P(A B) P(A
Mnohorozměrná analýza dat Podmíněná pravděpodobnost Definice: Uvažujme náhodné jevy A a B takové, že P(B) > 0. Podmíněnou pravěpodobností jevu A za podmínky, že nastal jev B, nazýváme podíl P(A B) P(A
Úvod do optimalizace, metody hladké optimalizace
 Evropský sociální fond Investujeme do vaší budoucnosti Úvod do optimalizace, metody hladké optimalizace Matematika pro informatiky, FIT ČVUT Martin Holeňa, 13. týden LS 2010/2011 O čem to bude? Příklady
Evropský sociální fond Investujeme do vaší budoucnosti Úvod do optimalizace, metody hladké optimalizace Matematika pro informatiky, FIT ČVUT Martin Holeňa, 13. týden LS 2010/2011 O čem to bude? Příklady
Využití molekulárních markerů v systematice a populační biologii rostlin. 9. Sekvenování DNA II. nrdna, low-copy markery
 Využití molekulárních markerů v systematice a populační biologii rostlin 9. Sekvenování DNA II. nrdna, low-copy markery Jaderný genom mnoho genů je v mnoha kopiích (multiple-copy) problém s homologií nevíme,
Využití molekulárních markerů v systematice a populační biologii rostlin 9. Sekvenování DNA II. nrdna, low-copy markery Jaderný genom mnoho genů je v mnoha kopiích (multiple-copy) problém s homologií nevíme,
Jak se matematika poučila v biologii
 Jak se matematika poučila v biologii René Kalus IT4Innovations, VŠB TUO Role matematiky v (nejen) přírodních vědách Matematika inspirující a sloužící jazyk pro komunikaci s přírodou V 4 3 r 3 Matematika
Jak se matematika poučila v biologii René Kalus IT4Innovations, VŠB TUO Role matematiky v (nejen) přírodních vědách Matematika inspirující a sloužící jazyk pro komunikaci s přírodou V 4 3 r 3 Matematika
DISTANČNÍ METODY KONSTRUKCE FYLOGENETICKÝCH STROMŮ
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŢENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŢENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Pravděpodobnost, náhoda, kostky
 Pravděpodobnost, náhoda, kostky Radek Pelánek IV122 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností připomenutí, souvislosti
Pravděpodobnost, náhoda, kostky Radek Pelánek IV122 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností připomenutí, souvislosti
Evoluční algoritmy. Podmínka zastavení počet iterací kvalita nejlepšího jedince v populaci změna kvality nejlepšího jedince mezi iteracemi
 Evoluční algoritmy Použítí evoluční principů, založených na metodách optimalizace funkcí a umělé inteligenci, pro hledání řešení nějaké úlohy. Populace množina jedinců, potenciálních řešení Fitness function
Evoluční algoritmy Použítí evoluční principů, založených na metodách optimalizace funkcí a umělé inteligenci, pro hledání řešení nějaké úlohy. Populace množina jedinců, potenciálních řešení Fitness function
Cvičení 3. Posudek únosnosti ohýbaného prutu. Software FREET Simulace metodou Monte Carlo Simulace metodou LHS
 Spolehlivost a bezpečnost staveb, 4. ročník bakalářského studia (všechny obory) Cvičení 3 Posudek únosnosti ohýbaného prutu Software FREET Simulace metodou Monte Carlo Simulace metodou LHS Katedra stavební
Spolehlivost a bezpečnost staveb, 4. ročník bakalářského studia (všechny obory) Cvičení 3 Posudek únosnosti ohýbaného prutu Software FREET Simulace metodou Monte Carlo Simulace metodou LHS Katedra stavební
1. Číselné posloupnosti - Definice posloupnosti, základní vlastnosti, operace s posloupnostmi, limita posloupnosti, vlastnosti limit posloupností,
 KMA/SZZS1 Matematika 1. Číselné posloupnosti - Definice posloupnosti, základní vlastnosti, operace s posloupnostmi, limita posloupnosti, vlastnosti limit posloupností, operace s limitami. 2. Limita funkce
KMA/SZZS1 Matematika 1. Číselné posloupnosti - Definice posloupnosti, základní vlastnosti, operace s posloupnostmi, limita posloupnosti, vlastnosti limit posloupností, operace s limitami. 2. Limita funkce
Cvičení 9. Posudek únosnosti ohýbaného prutu metodou LHS v programu FREET. Software FREET Simulace metodou LHS
 Pravděpodobnostní posuzování konstrukcí 4. ročník bakalářského studia obor Konstrukce staveb Cvičení 9 Posudek únosnosti ohýbaného prutu metodou LHS v programu FREET Software FREET Simulace metodou LHS
Pravděpodobnostní posuzování konstrukcí 4. ročník bakalářského studia obor Konstrukce staveb Cvičení 9 Posudek únosnosti ohýbaného prutu metodou LHS v programu FREET Software FREET Simulace metodou LHS
Aplikace DNA markerů v mykologii a molekulárni taxonomii
 Mendelova genetika v příkladech Aplikace DNA markerů v mykologii a molekulárni taxonomii doc. RNDr. Michal Tomšovský, Ph.D., Ústav ochrany lesů a myslivosti, LDF MENDELU, Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Aplikace DNA markerů v mykologii a molekulárni taxonomii doc. RNDr. Michal Tomšovský, Ph.D., Ústav ochrany lesů a myslivosti, LDF MENDELU, Brno Tento projekt je spolufinancován
Systém a evoluce obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
Dobývání znalostí. Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze
 Dobývání znalostí Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze Dobývání znalostí Bayesovské modely Doc. RNDr. Iveta Mrázová, CSc.
Dobývání znalostí Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze Dobývání znalostí Bayesovské modely Doc. RNDr. Iveta Mrázová, CSc.
MOLEKULÁRNÍ METODY V EKOLOGII MIKROORGANIZMŮ
 MOLEKULÁRNÍ METODY V EKOLOGII MIKROORGANIZMŮ (EKO/MMEM) ZPRACOVÁNÍ DGGE VÝSTUPŮ A SEKVENAČNÍCH DAT Zpracování výstupů z denaturační gradientové gelové elektroforézy (DGGE) práce s programem Gel2k Program
MOLEKULÁRNÍ METODY V EKOLOGII MIKROORGANIZMŮ (EKO/MMEM) ZPRACOVÁNÍ DGGE VÝSTUPŮ A SEKVENAČNÍCH DAT Zpracování výstupů z denaturační gradientové gelové elektroforézy (DGGE) práce s programem Gel2k Program
NGS analýza dat. kroužek, Alena Musilová
 NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
Bioinformatika a výpočetní biologie. KFC/BIN VII. Fylogenetická analýza
 ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
Fylogeneze a diverzita obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
AFLP. protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech
 AFLP AFLP protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech AFLP protokoly kit Invitrogen bez použití kitu AmpliTaq Gold (ABI) možnost
AFLP AFLP protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech AFLP protokoly kit Invitrogen bez použití kitu AmpliTaq Gold (ABI) možnost
Umělá inteligence II
 Umělá inteligence II 11 http://ktiml.mff.cuni.cz/~bartak Roman Barták, KTIML roman.bartak@mff.cuni.cz Dnešní program! V reálném prostředí převládá neurčitost.! Neurčitost umíme zpracovávat pravděpodobnostními
Umělá inteligence II 11 http://ktiml.mff.cuni.cz/~bartak Roman Barták, KTIML roman.bartak@mff.cuni.cz Dnešní program! V reálném prostředí převládá neurčitost.! Neurčitost umíme zpracovávat pravděpodobnostními
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Studijní materiály pro bioinformatickou část ViBuChu. úloha II. Jan Komárek, Gabriel Demo
 Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague
 1 / 23 Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague 2 / 23 biologové často potřebují najít často se opakující sekvence DNA tyto sekvence bývají relativně krátké,
1 / 23 Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague 2 / 23 biologové často potřebují najít často se opakující sekvence DNA tyto sekvence bývají relativně krátké,
Akvizice dat. Dekonvoluce Registrace. zobrazení INVESTICE DO ROZVOJE VZDĚLÁVÁNÍ
 a analýza signálů v perfúzním zobrazení Ústav biomedicínského inženýrství FEKT, VUT v Brně 22. 5. 2009 INVESTICE DO ROZVOJE VZDĚLÁVÁNÍ Osnova Úvod 1 Úvod 2 3 4 5 Úvod diagnostika a průběh terapie nádorových
a analýza signálů v perfúzním zobrazení Ústav biomedicínského inženýrství FEKT, VUT v Brně 22. 5. 2009 INVESTICE DO ROZVOJE VZDĚLÁVÁNÍ Osnova Úvod 1 Úvod 2 3 4 5 Úvod diagnostika a průběh terapie nádorových
Markovovy modely v Bioinformatice
 Markovovy modely v Bioinformatice Outline Markovovy modely obecně Profilové HMM Další použití HMM v Bioinformatice Analýza biologických sekvencí Biologické sekvence: DNA,RNA,protein prim.str. Sekvenování
Markovovy modely v Bioinformatice Outline Markovovy modely obecně Profilové HMM Další použití HMM v Bioinformatice Analýza biologických sekvencí Biologické sekvence: DNA,RNA,protein prim.str. Sekvenování
Odborná skupina pro spolehlivost. Použití ordinálních a semikvantitativních postupů ve spolehlivosti. Jaroslav Zajíček
 Odborná skupina pro spolehlivost Použití ordinálních a semikvantitativních postupů ve spolehlivosti Jaroslav Zajíček Obsah 1. Úvod management rizika 2. Výskyt a analýza nekvantitativních postupů - matice
Odborná skupina pro spolehlivost Použití ordinálních a semikvantitativních postupů ve spolehlivosti Jaroslav Zajíček Obsah 1. Úvod management rizika 2. Výskyt a analýza nekvantitativních postupů - matice
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Markov Chain Monte Carlo. Jan Kracík.
 Markov Chain Monte Carlo Jan Kracík jan.kracik@vsb.cz Princip Monte Carlo integrace Cílem je (přibližný) výpočet integrálu I(g) = E f [g(x)] = g(x)f (x)dx. (1) Umíme-li generovat nezávislé vzorky x (1),
Markov Chain Monte Carlo Jan Kracík jan.kracik@vsb.cz Princip Monte Carlo integrace Cílem je (přibližný) výpočet integrálu I(g) = E f [g(x)] = g(x)f (x)dx. (1) Umíme-li generovat nezávislé vzorky x (1),
EVOLUČNÍ MODELY PRO VYHODNOCENÍ PŘÍBUZNOSTI ORGANISMŮ
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Vytváření fylogenetických stromů na základě alignmentů. Tomáš Novotný Jaroslav Knotek
 Vytváření fylogenetických stromů na základě alignmentů Tomáš Novotný Jaroslav Knotek Alignmenty - opakování Existují dvě základní varianty alignmentů: globální a lokální Globální: Hledáme nejlepší zarovnání
Vytváření fylogenetických stromů na základě alignmentů Tomáš Novotný Jaroslav Knotek Alignmenty - opakování Existují dvě základní varianty alignmentů: globální a lokální Globální: Hledáme nejlepší zarovnání
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).
: Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).") Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
SRE 03 - Statistické rozpoznávání
 SRE 03 - Statistické rozpoznávání vzorů II Lukáš Burget ÚPGM FIT VUT Brno, burget@fit.vutbr.cz FIT VUT Brno SRE 03 - Statistické rozpoznávání vzorů II Lukáš Burget, ÚPGM FIT VUT Brno, 2006/07 1/29 Opakování
SRE 03 - Statistické rozpoznávání vzorů II Lukáš Burget ÚPGM FIT VUT Brno, burget@fit.vutbr.cz FIT VUT Brno SRE 03 - Statistické rozpoznávání vzorů II Lukáš Burget, ÚPGM FIT VUT Brno, 2006/07 1/29 Opakování
Studijní program Matematika Obor Pravděpodobnost, matematická statistika a ekonometrie
 Studijní program Matematika Obor Pravděpodobnost, matematická statistika a ekonometrie Doporučené průběhy studia pro rok 2014/15 24. září 2014 Vysvětlivky: Tento dokument obsahuje několik alternativních
Studijní program Matematika Obor Pravděpodobnost, matematická statistika a ekonometrie Doporučené průběhy studia pro rok 2014/15 24. září 2014 Vysvětlivky: Tento dokument obsahuje několik alternativních
Evoluční genetika II. Radka Reifová
 Evoluční genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Evoluční genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Využití metod strojového učení v bioinformatice David Hoksza
 Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
FYLOGEOGRAFIE A KOALESCENCE
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
oddělení Inteligentní Datové Analýzy (IDA)
") Vytěžování dat Filip Železný Katedra počítačů oddělení Inteligentní Datové Analýzy (IDA) 22. září 2014 Filip Železný (ČVUT) Vytěžování dat 22. září 2014 1 / 25 Odhad rozdělení Úloha: Vstup: data D = {
Vytěžování dat Filip Železný Katedra počítačů oddělení Inteligentní Datové Analýzy (IDA) 22. září 2014 Filip Železný (ČVUT) Vytěžování dat 22. září 2014 1 / 25 Odhad rozdělení Úloha: Vstup: data D = {
Pokročilé neparametrické metody. Klára Kubošová
 Klára Kubošová Další typy stromů CHAID, PRIM, MARS CHAID - Chi-squared Automatic Interaction Detector G.V.Kass (1980) nebinární strom pro kategoriální proměnné. Jako kriteriální statistika pro větvení
Klára Kubošová Další typy stromů CHAID, PRIM, MARS CHAID - Chi-squared Automatic Interaction Detector G.V.Kass (1980) nebinární strom pro kategoriální proměnné. Jako kriteriální statistika pro větvení
STANOVENÍ SPOLEHLIVOSTI GEOTECHNICKÝCH KONSTRUKCÍ. J. Pruška, T. Parák
 STANOVENÍ SPOLEHLIVOSTI GEOTECHNICKÝCH KONSTRUKCÍ J. Pruška, T. Parák OBSAH: 1. Co je to spolehlivost, pravděpodobnost poruchy, riziko. 2. Deterministický a pravděpodobnostní přístup k řešení problémů.
STANOVENÍ SPOLEHLIVOSTI GEOTECHNICKÝCH KONSTRUKCÍ J. Pruška, T. Parák OBSAH: 1. Co je to spolehlivost, pravděpodobnost poruchy, riziko. 2. Deterministický a pravděpodobnostní přístup k řešení problémů.
a způsoby jejího popisu Ing. Michael Rost, Ph.D.
 Podmíněná pravděpodobnost, náhodná veličina a způsoby jejího popisu Ing. Michael Rost, Ph.D. Podmíněná pravděpodobnost Pokud je jev A vázán na uskutečnění jevu B, pak tento jev nazýváme jevem podmíněným
Podmíněná pravděpodobnost, náhodná veličina a způsoby jejího popisu Ing. Michael Rost, Ph.D. Podmíněná pravděpodobnost Pokud je jev A vázán na uskutečnění jevu B, pak tento jev nazýváme jevem podmíněným
Populačně-genetická data
 Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
 š Ě Á š Á š š š ú š ů ú š ú š š ž š Š ú š ž ž š ž ž ů ů ů ž š ň ž ž ň š ů Ž ú š ž ž š ň Č Č ž Ů Ž Č Í Ť Í ň ť Í Č Ě Í š ú š Š Č šť š ů š ň ž šť šť ž ž ň šť ž ž šť š ú š Š Č š ž Ž š ů ů ž ž ň š ž ž ú ž
š Ě Á š Á š š š ú š ů ú š ú š š ž š Š ú š ž ž š ž ž ů ů ů ž š ň ž ž ň š ů Ž ú š ž ž š ň Č Č ž Ů Ž Č Í Ť Í ň ť Í Č Ě Í š ú š Š Č šť š ů š ň ž šť šť ž ž ň šť ž ž šť š ú š Š Č š ž Ž š ů ů ž ž ň š ž ž ú ž
Strom života. Cíle. Stručná anotace
 Předmět: Doporučený ročník: Vazba na ŠVP: Biologie 1. ročník Úvod do taxonomie Cíle Studenti zařadí člověka do příslušných taxonů taxonomického systému. Studenti se seznámí s principem fylogenetického
Předmět: Doporučený ročník: Vazba na ŠVP: Biologie 1. ročník Úvod do taxonomie Cíle Studenti zařadí člověka do příslušných taxonů taxonomického systému. Studenti se seznámí s principem fylogenetického
Evoluce velikosti genomu v čeledi Costaceae Genome size evolution in Costaceae
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Botanika Bc. Alžběta Böhmová Evoluce velikosti genomu v čeledi Costaceae Genome size evolution in Costaceae Diplomová
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Botanika Bc. Alžběta Böhmová Evoluce velikosti genomu v čeledi Costaceae Genome size evolution in Costaceae Diplomová
Projekční algoritmus. Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění. Jan Klíma
 Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Polymorfimus DNA, kdy se jedinci nebo druhy liší v jedné nukleotidové záměně
 MOLEKULÁRNÍ TAXONOMIE - 5 (2015) Single nucleotide polymophism - SNP Polymorfimus DNA, kdy se jedinci nebo druhy liší v jedné nukleotidové záměně AAGCCTA AAGCTTA V tomto případě mluvíme o alelách C a T.
MOLEKULÁRNÍ TAXONOMIE - 5 (2015) Single nucleotide polymophism - SNP Polymorfimus DNA, kdy se jedinci nebo druhy liší v jedné nukleotidové záměně AAGCCTA AAGCTTA V tomto případě mluvíme o alelách C a T.
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Vyhledávání podobných sekvencí BLAST
 Vyhledávání podobných sekvencí BLAST Základní informace Následující text je součástí učebních textů předmětu Analýza sekvencí DNA a je určen hlavně pro studenty Matematické biologie. Může být ovšem přínosný
Vyhledávání podobných sekvencí BLAST Základní informace Následující text je součástí učebních textů předmětu Analýza sekvencí DNA a je určen hlavně pro studenty Matematické biologie. Může být ovšem přínosný
Vícerozměrné statistické metody
 Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Rizikové úseky silnic z pohledu dopravních nehod
 Rizikové úseky silnic z pohledu dopravních nehod Ing. Jan TESLA, Ing. Igor IVAN, Ph.D. INSTITUT GEOINFORMATIKY VYSOKÁ ŠKOLA BÁŇSKÁ TECHNICKÁ UNIVERZITA OSTRAVA Cíle projektu Zpracování dat o dopravních
Rizikové úseky silnic z pohledu dopravních nehod Ing. Jan TESLA, Ing. Igor IVAN, Ph.D. INSTITUT GEOINFORMATIKY VYSOKÁ ŠKOLA BÁŇSKÁ TECHNICKÁ UNIVERZITA OSTRAVA Cíle projektu Zpracování dat o dopravních
Vybrané partie z obrácených úloh. obrácených úloh (MG452P73)
") Vybrané partie z obrácených úloh obrácených úloh (MG452P73) Obsah přednášky Klasifikace obrácených úloh a základní pojmy Lineární inverzní problém, prostor parametrů a dat Gaussovy transformace, normální
Vybrané partie z obrácených úloh obrácených úloh (MG452P73) Obsah přednášky Klasifikace obrácených úloh a základní pojmy Lineární inverzní problém, prostor parametrů a dat Gaussovy transformace, normální
Úvod do kvantového počítání
 2. přednáška Katedra počítačů, Fakulta elektrotechnická České vysoké učení technické v Praze 17. března 2005 Opakování Část I Přehled z minulé hodiny Opakování Alternativní výpočetní modely Kvantové počítače
2. přednáška Katedra počítačů, Fakulta elektrotechnická České vysoké učení technické v Praze 17. března 2005 Opakování Část I Přehled z minulé hodiny Opakování Alternativní výpočetní modely Kvantové počítače
Klasifikace a rozpoznávání. Bayesovská rozhodovací teorie
 Klasifikace a rozpoznávání Bayesovská rozhodovací teorie Extrakce p íznaků Granáty Četnost Jablka Váha [dkg] Pravděpodobnosti - diskrétní p íznaky Uvažujme diskrétní p íznaky váhové kategorie Nechť tabulka
Klasifikace a rozpoznávání Bayesovská rozhodovací teorie Extrakce p íznaků Granáty Četnost Jablka Váha [dkg] Pravděpodobnosti - diskrétní p íznaky Uvažujme diskrétní p íznaky váhové kategorie Nechť tabulka
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
METODY VÍCENÁSOBNÉHO ZAROVNÁVÁNÍ NUKLEOTIDOVÝCH SEKVENCÍ
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Osud jednotlivých kopií genů v populaci genové stromy
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
Bayesovská klasifikace
 Bayesovská klasifikace založeno na Bayesově větě P(H E) = P(E H) P(H) P(E) použití pro klasifikaci: hypotéza s maximální aposteriorní pravděpodobností H MAP = H J právě když P(H J E) = max i P(E H i) P(H
Bayesovská klasifikace založeno na Bayesově větě P(H E) = P(E H) P(H) P(E) použití pro klasifikaci: hypotéza s maximální aposteriorní pravděpodobností H MAP = H J právě když P(H J E) = max i P(E H i) P(H
Monte Carlo Lokalizace. Martin Skalský
 Monte Carlo Lokalizace Martin Skalský Proč Lokalizace? Problém určení pozice robota a věcí kolem něj. (filtrování dat, state estimation) Je důležitá Knowledge about where things are is at the core of any
Monte Carlo Lokalizace Martin Skalský Proč Lokalizace? Problém určení pozice robota a věcí kolem něj. (filtrování dat, state estimation) Je důležitá Knowledge about where things are is at the core of any
4. Na obrázku je rozdělovací funkce (hustota pravděpodobnosti) náhodné veličiny X. Jakou hodnotu musí mít parametr k?
 náhodné veličiny X. Jakou hodnotu musí mít parametr k?") A 1. Stanovte pravděpodobnost, že náhodná veličina X nabyde hodnoty menší než 6: P( X 6). Veličina X má rozdělení se střední hodnotou 6 a směrodatnou odchylkou 5: N(6,5). a) 0 b) 1/3 c) ½ 2. Je možné,
A 1. Stanovte pravděpodobnost, že náhodná veličina X nabyde hodnoty menší než 6: P( X 6). Veličina X má rozdělení se střední hodnotou 6 a směrodatnou odchylkou 5: N(6,5). a) 0 b) 1/3 c) ½ 2. Je možné,
Molekulárně biologické metody princip, popis, výstupy
 & Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
& Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
Bioinformatika a výpočetní biologie KFC/BIN. I. Přehled
 Bioinformatika a výpočetní biologie KFC/BIN I. Přehled RNDr. Karel Berka, Ph.D. Univerzita Palackého v Olomouci Definice bioinformatiky (Molecular) bio informatics: bioinformatics is conceptualising biology
Bioinformatika a výpočetní biologie KFC/BIN I. Přehled RNDr. Karel Berka, Ph.D. Univerzita Palackého v Olomouci Definice bioinformatiky (Molecular) bio informatics: bioinformatics is conceptualising biology
Základy genomiky. I. Úvod do bioinformatiky. Jan Hejátko
 Základy genomiky I. Úvod do bioinformatiky Jan Hejátko Masarykova univerzita, Oddělení funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky I. Zdrojová literatura ke
Základy genomiky I. Úvod do bioinformatiky Jan Hejátko Masarykova univerzita, Oddělení funkční genomiky a proteomiky Laboratoř molekulární fyziologie rostlin Základy genomiky I. Zdrojová literatura ke
Inženýrská statistika pak představuje soubor postupů a aplikací teoretických principů v oblasti inženýrské činnosti.
 Přednáška č. 1 Úvod do statistiky a počtu pravděpodobnosti Statistika Statistika je věda a postup jak rozvíjet lidské znalosti použitím empirických dat. Je založena na matematické statistice, která je
Přednáška č. 1 Úvod do statistiky a počtu pravděpodobnosti Statistika Statistika je věda a postup jak rozvíjet lidské znalosti použitím empirických dat. Je založena na matematické statistice, která je
přetrénování = ztráta schopnosti generalizovat vlivem přílišného zaměření klasifikátorů na rozeznávání pouze konkrétních trénovacích dat
 Zkouška ISR 2013 přetrénování = ztráta schopnosti generalizovat vlivem přílišného zaměření klasifikátorů na rozeznávání pouze konkrétních trénovacích dat 1. Rozdílné principy u induktivního a deduktivního
Zkouška ISR 2013 přetrénování = ztráta schopnosti generalizovat vlivem přílišného zaměření klasifikátorů na rozeznávání pouze konkrétních trénovacích dat 1. Rozdílné principy u induktivního a deduktivního
Tok GI v buňce. Genetický polymorfizmus popis struktury populací. Organizace genetického materiálu. Definice polymorfismu
 Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Bioinformatika. hledání významu biologických dat. Marian Novotný. Friday, April 24, 15
 Bioinformatika hledání významu biologických dat Marian Novotný Bioinformatika sběr biologických dat archivace biologických dat organizace biologických dat interpretace biologických dat 2 Biologové sbírají
Bioinformatika hledání významu biologických dat Marian Novotný Bioinformatika sběr biologických dat archivace biologických dat organizace biologických dat interpretace biologických dat 2 Biologové sbírají
Zpětnovazební učení Michaela Walterová Jednoocí slepým,
 Zpětnovazební učení Michaela Walterová Jednoocí slepým, 17. 4. 2019 V minulých dílech jste viděli Tři paradigmata strojového učení: 1) Učení s učitelem (supervised learning) Trénovací data: vstup a požadovaný
Zpětnovazební učení Michaela Walterová Jednoocí slepým, 17. 4. 2019 V minulých dílech jste viděli Tři paradigmata strojového učení: 1) Učení s učitelem (supervised learning) Trénovací data: vstup a požadovaný
1 Tyto materiály byly vytvořeny za pomoci grantu FRVŠ číslo 1145/2004.
 Prostá regresní a korelační analýza 1 1 Tyto materiály byly vytvořeny za pomoci grantu FRVŠ číslo 1145/2004. Problematika závislosti V podstatě lze rozlišovat mezi závislostí nepodstatnou, čili náhodnou
Prostá regresní a korelační analýza 1 1 Tyto materiály byly vytvořeny za pomoci grantu FRVŠ číslo 1145/2004. Problematika závislosti V podstatě lze rozlišovat mezi závislostí nepodstatnou, čili náhodnou
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů
 Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?