Minerální výživa rostlin. příjem, asimilace a funkce minerálních prvků
|
|
|
- Aneta Žáková
- před 8 lety
- Počet zobrazení:
Transkript


1 Minerální výživa rostlin příjem, asimilace a funkce minerálních prvků
2 Minerální výživa rostlin příjem, asimilace a funkce minerálních prvků Kriteria nezbytnosti prvků pro rostliny Klasifikace minerálních živin Příjem minerálních živin a transport rostlinou Vlastnosti půdy, iontová výměnná kapacita Příjem kořenem Transport přes membránu transportéry (přenašeče) makrobiogenní minerální prvky N metabolismus dusíku S, P, K, Ca, Mg, mikrobiogenní minerální prvky Cl, Fe, B, Mn, Na, Zn, Cu, Ni, Mo Benefiční prvky
3 Minerální výživa rostlin Rostliny jsou: Autotrofní tvorba organických sloučenin z anorganických Plně dostupné C, H, O2 - CO2, H2O Ostatní prvky (převážně) z půdy soil miners - půdní výživa (kořeny, mykorhizní symbióza)




4 Historie studia minerální výživy rostlin Jan Baptista van Helmont ( ) Člověk když jí, tak roste. Ale z čeho roste strom? Z vody. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková


5 Historie studia minerální výživy rostlin Justus von Liebig ( ) r spis Organická chemie a její použití v zemědělství a fyziologii minerální teorie Živinami všech rostlin jsou látky nerostné. Chlévský hnůj, lidské a zvířecí výkaly nepůsobí svými součástkami ústrojnými na život rostlin, nýbrž nepřímo zplodinami svého rozkladu, tedy přeměnou svého uhlíku v kysličník uhličitý a dusíku v amoniak nebo kyselinu dusičnou. Ústrojný hnůj složený z částí nebo zbytků rostlin a zvířat dá se nahradit neústrojnými sloučeninami, v něž se v půdě rozpadá. Ctirad Růžička, Úroda 10/2003 Převzato: dr. E. Tylová


6 Historie studia minerální výživy rostlin Justus von Liebig ( ) Liebigův zákon minima: Rostliny jsou životně závislé na tom prvku, který je v jejich životním prostředí obsažen nejméně Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková
 Studoval vztahy mezi výnosy rostlin a množstvím živin přidávaných do půdy a vyjádřil je matematickými rovnicemi 8-1")
7 Historie studia minerální výživy rostlin A. Mitscherlich ( ) Studoval vztahy mezi výnosy rostlin a množstvím živin přidávaných do půdy a vyjádřil je matematickými rovnicemi 8-1 Yield (y, t ha ) 7 A (A-y) y y=a-(a-a).e a Fertilizers (x, t ha ) -k.x 0.30

 na obsahu živin v pletivu")
8 Závislost růstu (produkce biomasy, výnosu, rychlosti procesu apod.) na obsahu živin v pletivu deficience dostupnost toxicita 95%
9 Nástup aplikace minerálních hnojiv - superfosfát (poč. 19. st) kosti, koprolity a fosfátové horniny - draselná hnojiva dusíkatá hnojiva Ca(NO3) NH (Haber a Bosch syntéza amoniaku: N2+H2 při 200 C a zvýšeném tlaku)
10 Minerální prvky v rostlinách Stanovení obsahu v rostlinách Sušina: sušení čerstvé biomasy (1-2 dny) při C. Spálení sušiny při teplotě nad 600 C Analýza popele (popeloviny) a plynných zplodin zplodiny. Takto v rostlinách zjištěno více než 60 prvků včetně zlata, olova, rtuti, arsenu a uranu. Které z nalezených prvků jsou však pro rostliny nezbytné? Nutno zjistit exprimentálně!


11 Minerální prvky v rostlinách Hydroponie 1. Obecný význam: 2. Požadavky: fyziologie rostlin, zahradnictví koncentrace živin, vyrovnanost poměru, dostupnost O2, ph Živné roztoky: Knop Hoagland Taiz a Zeiger 2002
 Prvek nemůže být nahrazen jiným prvkem (deficit vyvolaný nepřítomností jednoho prvku nemůže být odstraněn dodáním jiného prvku) (3) Daný prvek má svou funkci v")
12 Minerální prvky v rostlinách Kriteria nezbytnosti prvků pro rostliny (1) Bez daného prvku rostlina neukončí svůj životní cyklus. Justus von Liebig: zákon minima (2) Prvek nemůže být nahrazen jiným prvkem (deficit vyvolaný nepřítomností jednoho prvku nemůže být odstraněn dodáním jiného prvku) (3) Daný prvek má svou funkci v metabolismu rostliny (nezávisle na chemických, biologických či jiných podmínkách v půdě). (Arnon, Stout, 1939)
13 Minerální prvky v rostlinách - klasifikace prvky biogenní obecně všechny prvky, které tvoří živé organismy prvky organických látek C, H, O prvky minerální výživy rostlin 1. esenciální = nezbytné = nenahraditelné = funkčně nezastupitelné makrobiogenní minerální prvky 1000 mg (1 g) v 1 kg sušiny N, K, Ca, Mg,P,S, mikrobiogenní minerální prvky 100 mg (0,1 g) v 1 kg sušiny 2. prvky benefiční prospěšné Stimulují růst, požadované některými druhy, např.: Na, Si, Se
 získané z půdy Mikroprvky (ppm) relativní počet atomů vztaženo k počtu atomů")
14 Minerální prvky v rostlinách - klasifikace Prvek chemická značka koncentrace v sušině (%, ppm) získané z vody nebo CO2 získané z půdy Makroprvky (%) získané z půdy Mikroprvky (ppm) relativní počet atomů vztaženo k počtu atomů molybdenu
15 Minerální prvky v rostlinách - klasifikace prvky biogenní makrobiogenní minerální prvky prvky organických látek C, H, O 1000 mg (1 g) v 1 kg sušiny 1, % sušiny 1,4 % sušiny Uhlík 45 Kyslík 45 Vodík 6 1,2 1 0,8 0,6 0,4 N..1,5 K 1,0 Ca.0,5 Mg.0,2 P.0,2 S..0,1 Si 0,1 0,2 0 N Uhlík Kyslík Vodík mikrobiogenní minerální prvky 100 mg (0,1 g) v 1 kg sušiny Epstein 1972, K Ca Mg P S Si ppm sušiny Cl 100 Fe 100 B.60 Mn.50 Na Zn Cu.. 6 Ni...0,1 Mo....0,1 Cl Fe B Mn Na Zn Cu Ni Mo
16 Minerální prvky v rostlinách - klasifikace podle biochemické funkce 4 skupiny: Skupina 11- části organických uhlíkatých sloučenin N- aminokyseliny, amidy, proteiny, nukleové kyseliny, nukleotidy, koenzymy, hexosaminy atd. S cystein, cystin, methionin-proteiny, mastné kyseliny,koenyzm A, thiamin, glutathion, biotin atd. Skupina 2- významné při ukládání energie nebo ve strukturální integritě P- fosfáty cukrů, aminokyseliny, nukleotidy, koenzymy, fosfolipidy. Klíčová úloha v reakcích zahrnujících ATP. Si- amorfní křemík v buněčné stěně-pevnost a elastiicta buněčných stěn. B- manitol, manan-buněčná stěna- prodlužování buněk, metabolismus nukleových kyselin Skupina 33- zůstávající v podobě iontů (volných nebo vázaných K- kofaktor více než 40 enzymů, základní kation udržující buněčný turgor a buněčnou elektroneutralitu. Ca- střední lamela buněčné stěny, kofaktor enzymů hydrolýzy ATP a fosfolipidů, druhý posel v metabolické regulaci. Mg-chlorofyl, přenos fosfátů Cl- fotosyntetické reakce vývoj kyslíku Mn-aktivita dehydrogenáz, dekarboxyláz, kináz, oxidáz a peroxidáz, vývoj kyslíku Na- regenerace fosfoenol pyruvátu u C4 a CAM rostlin Skupina 44- účastnící se redoxredox- reakcí reakcí-- přenosu elektronů Fe-cytochromy a nehemové proteiny pro fotosyntézu, dýchání a fixaci N 2 Zn-alkohol dehydrogenáza, karbon anhydráza atd Cu- kofaktor dalších enzymů, plastocyanin Ni- ureáza Mo- nitrogenáza, nitrát reduktáza atd. Podle Taiz and Zeiger 2002
17 Příjem minerálních látek rostlinou = vstup prvků do biosféry
18 Příjem minerálních živin a transport rostlinou Vlastnosti půdy, iontová výměnná kapacita příjem možný celým povrchem těla nejvíce kořeny z půdy

 organické částice voda vzduch Brady a Weil")
19 Příjem minerálních živin a transport rostlinou Vlastnosti půdy, iontová výměnná kapacita Půda: 4 složky: minerální částice: kameny a štěrk písek (částice o velikosti 2-0,05mm) prachové částice (0,05-0,002mm) jílové částice (méně než 0,002mm) organické částice voda vzduch Brady a Weil 2002


20 Příjem minerálních živin a transport rostlinou Vlastnosti půdy, iontová výměnná kapacita Fyzikální a chemické vlastnosti půdy: ph ph a- aktivita růstu kořenů, mikrobiální Růst kořene optimum ph Kyselé podmínky zvětrávání K, Ca, Mg, Mn
21 Příjem minerálních živin a transport rostlinou Vlastnosti půdy, iontová výměnná kapacita Půda: Minerální živiny v půdě ve třech stavech: a) Vázané. V tomto stavu nemohou být rostlinami přímo využity. Jsou součástí hornin, zvětrávání, působením samotné rostliny (vylučováním vodíkových iontů kořeny). b) Výměnné. Jsou to hlavně kationty iontově vázané na povrchu půdních koloidů, tj. jílových a humusových částic. Snadno uvolněny výměnou za jiné kationty, např. za kationt vodíkový produkovaný při dýchání kořenů c) Rozpuštěné. Živiny přítomné v půdním roztoku, které rostlina může přímo využít.

22 Příjem minerálních živin a transport rostlinou Vlastnosti půdy půdy,, iontová výměnná kapacita adsorpce kationtů na negativně nabitý povrch půdních částic dýchání kořenů CO2 H2CO3 HCO3- + H+ HCO3- výměna za anionty v půdním roztoku na povrchu kořene H+ výměna za kationty na povrchu půdních částic, koloidů




23 Příjem minerálních živin a transport rostlinou Vlastnosti půdy půdy,, iontová výměnná kapacita Koloidy disperzní soustavy obsahující částice, které: jsou dostatečně malé aby zůstaly v suspenzi příliš velké na to, aby byly rozpuštěny a tvořily pravý roztok mají velký specifický povrch schopnost povrchově vázat kationty
24 příjem minerálních látek rostlinami příjem možný celým povrchem těla nejvíce kořeny z půdy zóna s kořenovými vlásky (mykorhiza) Základní mechanismy příjmu živin přijímány převážně jako ionty anionty, kationty z půdního roztoku
25 příjem minerálních látek Příjem listy -minerální látky v plynné formě: prostřednictvím průduchů -(NH3, SO2, NO2), -možný je i opačný proces únik plynných forem z listu do atmosféry (NH3, H2S a další těkavé sirné sloučeniny) -minerální látky v kapalné formě: difuzí přes kutikulu (prostřednictvím hydrofilních pórů)...hnojení na list (železo, měď, Mn, dusík)
26 příjem minerálních látek kořeny Faktory kořenového systému, které ovlivňují příjem iontů kořenem: - velikost a architektura kořenového systému - morfologie kořene (průměr, kořenové vlásky) - kapacita pro příjem živin na jednotku délky nebo plochy kořene - schopnost uvolňování látek, které ovlivňují chemické vlastnosti rhizosféry - mykorrhiza - aktuální požadavek rostliny - transport živin přes plazmatickou membránu

 Převzato od")
27 příjem minerálních látek rostlinami Morfologie kořenového systému: různé typy kořen hlavní a kořeny postranní Allorhizie (nahosemenné a dvouděložné) kořeny Svazčité Homorhizie (jednoděložné) Převzato od Dr. Votrubové

 životnost")
28 příjem minerálních látek rostlinami kořenové vlásky vznikají apikálním růstem buněk rhizodermis hustota kolem 100 na mm2 délka um (někdy až 1000 um) životnost několik dní


29 příjem minerálních látek buněčná stěna je pro minerální látky propustná cesty : apoplast symplast bariéra plazmatická membrána (Casparyho proužky) suberin hydrofobní síťovitý polymer mastných kyselin, alkoholů s dlouhým řetězcem a fenolických látek
 Or entire root (corn) Draslík, nitrát, iont amonný, fosfát All locations of root")

30 příjem minerálních látek rostlinami Kořen absorbuje různé ionty v různých oblastech Vápník - Ca Apical region Železo Apical region (barley) Or entire root (corn) Draslík, nitrát, iont amonný, fosfát All locations of root surface - In corn, elongation zone has max K accumulation and nitrate absorption In corn and rice, root apex absorbs ammonium faster than the elongation zone does In several species, root hairs are the most active phosphate absorbers

31 příjem minerálních látek kořeny Růst kořenů do živinami bohatých oblastí půdy Vliv selektivní přítomnosti živin na tvorbu kořenového systému
32 Základní způsoby (resp. mechanismy) transportu těchto i dalších iontů a molekul v rostlinách jsou: -Hromadný tok roztoku -Prostá difuze - Transport prostřednictvím membránových bílkovin
33 Základní způsoby (resp. mechanismy) transportu těchto i dalších iontů a molekul v rostlinách jsou: -Hromadný tok roztoku transport na dlouhé vzdálenosti (viz Rostlina a voda). vázán na vodivá pletiva. Do nadzemních částí převážně transpiračním proudem nebo kořenovým vztlakem v xylemu, dálkový transport proti směru transpiračního proudu probíhá ve floemu, Mnohé minerální ionty tak v rostlinách cirkulují. Např. fosfát
34 Základní způsoby (resp. mechanismy) transportu těchto i dalších iontů a molekul v rostlinách jsou: - Hromadný tok roztoku - Prostá difuze - Transport prostřednictvím membránových bílkovin
35 Základní způsoby (resp. mechanismy) transportu těchto i dalších iontů a molekul v rostlinách jsou: - Prostá difuze 1. Fickův zákon (viz Rostlina a voda). flux permeability areas of cell membrane coefficient (cm/s) Spontánně, směr je určen gradientem chemického potenciálu (viz dále) Rostlinné membrány jsou pro velkou většinu biologicky významných molekul a iontů prakticky neprostupné. Čím jsou difundující částice větší a polárnější, tím je jejich membránová permeabilita menší. Jen několik biologicky významných solutů je však nepolárních a jen tři (02 CO2 a NH4+) pronikají lipidní dvouvrstvou difuzí.
36 Základní způsoby (resp. mechanismy) transportu těchto i dalších iontů a molekul v rostlinách jsou: -Hromadný tok roztoku -Prostá difuze - Transport prostřednictvím membránových bílkovin
37 Transport látek přes membránu prostřednictvím membránových bílkovin
38 Transport minerálních látek přes membrány membrány selektivně propustné bariéry množství volné energie schopné konat práci = transportovat látku přes membránu je dáno: koncentrací dané látky tlakem nábojem hodnota i (chemický potenciál) integruje působení těchto faktorů Gibbsova volná energie a chemický potenciál
39 Transport minerálních látek přes membrány Gibbsova volná energie a chemický potenciál 1. hlavní věta termodynamiky o zachování energie (energie vesmíru je konstantní 2. Hlavní věta termodynamiky o produkci entropie (entropie vesmíru vzrůstá Pro biologické procesy, kdy stejný tlak změnu vnitřní energie du možno nahradit H (enthalpie) Reační enthalpie dh vyjadřuje přeměnu energie v reakci Část enthalpie schopná konat práci = Gibbsova volná energie dg = dh TdS kde dh - změna enthalpie, T - absolutní teplota, ds změna entropie Změna Gibbsovy volné energie je ta část reakční enthalpie, která ve spontánně probíhajícím procesu může být maximálně přeměněna v práci
40 Transport minerálních látek přes membrány množství volné energie schopné konat práci = transportovat látku přes membránu je dáno: koncentrací dané látky tlakem nábojem hodnota i (chemický potenciál) integruje působení těchto faktorů = Gibbsova volná energie na mol určitého druhu látky (i) u iontů hraje důležitou roli náboj potenciál elektrochemický i vyjadřuje se jako napětí v mv nebo jako energie v kj. mol-1
41 Transport minerálních látek přes membrány množství volné energie schopné konat práci = transportovat látku přes membránu je dáno: koncentrací dané látky tlakem nábojem hodnota i (chemický potenciál) integruje působení těchto faktorů Pohyb látek přes membránu: hnací silou je jµrozdíl chemického potenciálu dané látky na opačných stranách membrány u iontů hraje důležitou roli náboj potenciál elektrochemický i vyjadřuje se jako napětí v mv nebo jako energie v kj. mol-1
42 Transport minerálních látek přes membrány složka: i chemický potenciál iontu i = koncentrační tlaková i0 + R. T. ln Ci + PVi + chemický potenciál iontu i za standardních podmínek koncentrační složka iontu i tlaková složka iontu i elektrická zi. F. E složka elektrického potenciálu iontu i (vliv náboje) R = univerzální plynová konstanta (8,314 J.mol -1.K-1) T = teplota v kelvinech ln Ci = přirozený logaritmus koncentrace iontu i P = tlak (rozdíl tlaku na iont v soustavě a tlakem atmosférickým = hydrostatický tlak v buňce) Vi = parciální molální objem iontu i zi = elektrostatický náboj (K+ = +1; Cl - = -1) F = Faradayova konstanta (96484 C. mol-1; C = coulomb, J/ V.mol) E = celkový elektrický potenciál (napětí) prostředí (V = volt)
 probíhá samovolně po spádu gradientu chemického potenciálu Rovnováha pokud")

43 Transport minerálních látek přes membrány chemický potenciál v kompartmentu A chemický potenciál v kompartmentu A Popis situace Pasivní transport (difuse) probíhá samovolně po spádu gradientu chemického potenciálu Rovnováha pokud neprobíhá aktivní transport stabilní stav Aktivní transport probíhá proti spádu gradientu chemického potenciálu G nutný k přesunu 1 molu látky z kompartmentu A do B = rozdílu Reakce musí být spřažena s procesem jehož G je negativnější než ( )
44 Transport minerálních látek přes membrány hybná síla transportu iontu do buňky je dána rozdílem elektrochemického potenciálu iontu v apoplastu a v cytosolu K+ = K+A - K+C < 0 možný pasivní transport 0 nutno dodat energii = 0 stejný počet iontů se přenáší oběma směry rozdíl elektrických potenciálů na opačných stranách membrány může udržet transport určitého iontu v dynamické rovnováze i při značném rozdílu koncentrace iontu tento potenciál se nazývá potenciál Nernstův (EN) CiA RT ENi = EA EC = zif ln CiC
 CiA RT ENi = EA EC = zif Po nahrazení konstant číselnými hodnotami můžeme Nernstovu rovnici pro jednomocný kationt a 25 C dále zjednodušena")
45 Transport minerálních látek přes membrány rozdíl elektrických potenciálů na opačných stranách membrány může udržet transport určitého iontu v dynamické rovnováze i při značném rozdílu koncentrace iontu tento potenciál se nazývá potenciál Nernstův (EN) CiA RT ENi = EA EC = zif Po nahrazení konstant číselnými hodnotami můžeme Nernstovu rovnici pro jednomocný kationt a 25 C dále zjednodušena takto Z rovnice vyplývá, že desetinásobný rozdíl v koncentraci iontu (mezi oběma kompartmenty) odpovídá Nernstovu potenciálu 59 mv, v případě dvojmocného iontu 29,5 mv. Tj. membránový potenciál 59 mv udrží (resp. vyváží) desetinásobný koncentrační gradient jednomocného iontu transportovaného difuzí. ln CiC
46 Transport minerálních látek přes membrány rozdíl elektrických potenciálů na opačných stranách membrány může udržet transport určitého iontu v dynamické rovnováze i při značném rozdílu koncentrace iontu tento potenciál se nazývá potenciál Nernstův (EN) CiA RT ENi = EA EC = zif ln CiC
47 Transport minerálních látek přes membrány biologické membrány oddělují prostředí s velmi rozdílnými elektrickými potenciály = potenciál membránový (Vm) Taiz and Zeiger 2002

48 Transport minerálních látek přes membrány biologické membrány oddělují prostředí s velmi rozdílnými elektrickými potenciály = potenciál membránový (Vm) vzniká komplexním působením komponent membrány i obou prostředí negativní náboj je na straně cytosolu vzniká aktivním transportem protonů z cytosolu protonovými pumpami rozdílná koncentrace protonů na membráně = protonmotorická síla (pmf -proton motive force) v jednotkách elektrického napětí (dělěno F) -200 až -300 mv obvykle se udává -268 mv Taiz and Zeiger 2002
49 Transport minerálních látek přes membrány Membránové potenciály vznikají třemi způsoby: 1. Difúze: anionty a kationty určitého elektrolytu membránou rozdílnými rychlostmi (v důsledku rozdílných koeficientů permeability). 2. Donnanova rovnováha - působení fixovaných iontů: Většina cytosolických bílkovin nese při fyziologickém ph 7,5 záporné náboje (na karboxylech RCOO- aminokyselinových zbytků), které jsou vyvažovány hlavně ionty K+. Vnitrobuněčná koncentrace K+ je ve srovnání s koncentrací vnější vysoká. Draselné ionty budou tedy volně difundovat prostřednictvím draslíkových kanálů z buňky, ale současně budou přitahovány zpět do buňky nedifuzibilními, v buňce fixovanými anionty. Ve výsledné rovnováze bude síla koncentračního gradientu ženoucího K+ z buňky vyvažována silou elektrického gradientu táhnoucího draslíkové ionty zpět do buňky. Anionty budou odpuzovány převahou vnitřních fixovaných záporných nábojů a přitahovány vnějšími kladnými náboji. Takovéto rozdělení imobilních (záporných) a mobilních (kladných a záporných) nábojů vede ke vzniku tzv. Donnanova potenciálu. 3. Vm vzniká hlavně aktivním vylučováním (pumpováním) vodíkových iontů z vnitřního cytoplasmatického do vnějšího buněčného prostoru (zde jako vnějšek platí také obsah vakuoly). K tomu dochází činností membránových enzymů - protonové pumpy. V dynamické rovnováze je kontinuální činnost protonových pump kompenzována tokem pozitivních nábojů do cytosolu nebo negativních z cytosolu. Rovnováha mezi vysokou permeabilitou K+ a činností protonových pump v membránách je obvykle hlavním faktorem, který stabilizuje V m rostlinných membrán (viz dále Kanály pro K+). Za určitých podmínek může přechodně nastat značné zvýšení permeability membrány pro anionty nebo pro Ca2+. To obvykle vyvolá přechodné vychýlení V m k pozitivním hodnotám neboli depolarisaci membrány: anionty vytékají z cytosolu nebo ionty Ca2+ do něho vtékají. Skripta Luštinec a Žárský
 se membrána hyperpolarizuje importem kationtů")

50 membránový potenciál plazmalemy asi -150 mv tonoplastu -20 mv až -90 mv činností pump (H+ a Ca2+ATPáz) se membrána hyperpolarizuje importem kationtů do cytosolu a exportem aniontů do apoplastu se membrána depolarizuje rychlost transportu dané látky ovlivněna propustností = permeabilitou membránytaiz and Zeiger 2002
51 transportu látek přes membránu slouží pumpy přenašeče kanály

 Taiz and")
52 transportu látek přes membránu slouží pumpy přenašeče kanály Kanál Přenašeč Pumpa prostá difúze Pasivní transport (ve směru gradientu elktrochemického potenciálu) Primární aktivní transport (proti směru gradientu elektrochemického potenciálu) Taiz and Zeiger 2002
53 transportu látek přes membránu slouží pumpy přenašeče kanály Pumpy: membránové bílkoviny enzymového charakteru, hydrolysují ATP nebo pyrofosfát a pomocí uvolněné chemické energie transportují ionty vodíkové (H+), vápenaté (Ca2+) a sodné (Na+) jakož i některé látky organické z jedné strany membrány na druhou. Charakterizovány jsou jako elektrogenní a elektroneutrální. Činností elektrogenních pump dochází k čistému pohybu náboje přes membránu, elektroneutrální pumpy takovýto pohyb náboje nezpůsobují. Elektrogenním transportem kationtů z vnitřního do vnějšího buněčného prostoru se vytváří negativní membránový potenciál - vnitřní prostor se stává elektronegativní Pumpy pohánějí aktivní membránový transport klasifikovaný jako primární. Skripta Luštinec a Žárský
54 transportu látek přes membránu slouží pumpy pumpy přenášejí iont nebo molekulu do míst s jeho vyšší koncentrací nutno dodat energii získává se z ATP nebo PPi primární aktivní transport H+ pumpy lokalizovány na celé řadě membrán (plazmalema, tonoplast, ER) H+ATPázy typ P v plazmatické membráně (10 transmembránových domén, u Arabidopsis 10 genů) 1 ATP 1 H+ typ V v tonoplastu (část V0 v tonoplastu, V1 v cytosolu, 1 ATP 2H+ do vakuoly) typ F v chloroplastech. mitochondriích H+PPázy štěpí PPi (pyrofosfát) mladá vyvíjející se pletiva, 16 transmembránových domén Ca2+ATPázy udržují nízkou hladinu Ca2+ v cytosolu, k transportu je třeba ATP a protonmotorická síla Vakuolární H+-pyrofosfatasa - Protony uvolněné při hydrolyse anorganického pyrofosfátu (PP) pumpuje do vakuoly a do Golgiho cisteren. Je specifická pro rostliny. ABC-transportéry (ATP-binding casette) v tonoplastu, přenos látek do vakuoly transportují různé sekundární metabolity, např. flavonoidy, produkty rozkladu chlorofylu a některá xenobiotika

 H+ATPase - plasma membrane-type protonpumping ATPase (Ptype ATPase)")
55 transportu látek přes membránu slouží pumpy H+ATPase-proton pumps are the major factor in the membrane potential of most plant cells antiport symport Uncharged solute (ex. sugars) H+ATPase - plasma membrane-type protonpumping ATPase (Ptype ATPase) H+ATPase - tonoplasttype proton-pumping ATPase (V-type ATPase) Převzato Žárský a Kulich
 Hydrolysis of ATP at the binding site is thought to change the conformation of the enzyme, thereby exposing the H+-binding site to the")

56 transportu látek přes membránu slouží pumpy H+ATPázy typ P v plazmatické membráně (10 transmembránových domén, u Arabidopsis 10 genů) 1 ATP 1 H+ Plasma membrane type proton-pumping ATPase P-type ATPase) Hydrolysis of ATP at the binding site is thought to change the conformation of the enzyme, thereby exposing the H+-binding site to the outside of the membrane where the H+ is released. vnitřní straně plasmalemy hydrolyzuje ATP a H+ přenáší (pumpuje) na vnější stranu membrány. Vzniklá protonmotorická síla pohání různé přenašeče a prostřednictvím V m podporuje transport iontů přes kanály. The ATP-binding site of P-type ATPase is an aspartic acid residue (D). Převzato Žárský a Kulich
57 transportu látek přes membránu slouží pumpy H+ATPázy typ P v plazmatické membráně Plasma membrane-type proton-pumping ATPase (P-type ATPase) inhibited by VO3-, stimulated by K+, insensitive to NO3-. VO3- is competing with phosphate for phosphate binding site, so there is phosphoryl transfer between ATP and P-type ATPase. Převzato Žárský a Kulich
58 transportu látek přes membránu slouží pumpy H+ATPázy typ P v plazmatické membráně P-H+-ATPasy mají další funkce: a) Vlivem auxinu se více exprimují a okyselují buněčnou stěnu (na ph 5,5 až 5,0) a tím umožňují expansní růst buněk (viz Fytohormony). b) Odčerpávají přebytečné vodíkové ionty z cytosolu a přispívají ke stabilitě cytosolického ph, jeho obvyklá hodnota je 7,0-7,5. c) Podílejí se na schopnosti kořenů okyselovat půdu. (K tomu dochází i respirací kořenů.) Převzato Žárský a Kulich
 vakuolární H+")

59 transportu látek přes membránu slouží pumpy H+ATPázy typ V v tonoplastu tonoplast-type proton-pumping ATPase (V-type ATPase) vakuolární H+ ATPáza protonová pumpa V-ATPaza v tonoplastu V1 katalytická část pro štěpení ATP V0 trasmembránový komplex pro přenos protonů Převzato Žárský a Kulich
 - Pumpuje protony do vakuloy,")
 -")
 indicate that it does not form complex with")
60 transportu látek přes membránu slouží pumpy H+ATPázy typ V v tonoplastu tonoplast-type proton-pumping ATPase (V-type ATPase) - Pumpuje protony do vakuloy, udržuje vakuolu více pozitivní než cytosol, takže anionty (Cl-) mohou do vakuoly. - strukturálně se více podobá F-typu ATPases (chloroplast and mitochondria) - inhibited by NO3-, it is insensitive to VO3-, oligomycin, azide. The insensitivity of vanadate (VO3-) indicate that it does not form complex with phosphate. Převzato Žárský a Kulich

61 transportu látek přes membránu slouží pumpy Ca2+-ATPases Plasma membranes, chloroplast envelope, ER, and vacuolar membrane also contain calcium-pumping ATPases (Ca2+ATPase). Ca2+-ATPase couple the hydrolysis of ATP with the translocation of Ca2+ across the membrane. In the case of plasma membrane, the calcium is pumped out of the cytosol to keep [Ca2+] low so phosphate won t precipitate and the Ca2+-dependent signaling pathways operating properly. [Ca2+]i signal attenuation Převzato Žárský a Kulich
62 transportu látek přes membránu slouží přenašeče přenašeče při přenosu mění konformaci přenos aktivní nebo pasivní membránové transportní bílkoviny katalysující pohyb anorganických iontů a jednoduchých organických molekul přes membránu. Spřažení sestupného (exergonického) toku protonů se vzestupným (endergonickým) tokem solutů. Tok solutu katalysovaný přenašečem ve stejném směru jako tok H+ se nazývá symport. Tok solutu katalysovaný přenašečem proti toku H+ se nazývá antiport. Při něm dochází k výměně solutů za protony. Při symportu i antiportu se protonmotorická síla převádí v elektrochemický potenciál transportovaných solutů. Membránový transport katalysovaný symportovými a antiportovými přenašeči (symportery a antiportery) probíhá díky cirkulaci protonů membránou: sekundární aktivní transport. Některé přenašeče katalyzují i pasivní membránový transport, který bývá označován jako usnadněná difuze (faciliated diffusion), ačkoliv se difuzi podobá jen v tom, že transport látek probíhá po spádu elektrochemického potenciálu bez spotřeby další energie. Luštinec a Žárský
 Taiz and")
63 transportu látek přes membránu slouží přenašeče přenašeče při přenosu mění konformaci přenos aktivní nebo pasivní schéma sekundárního aktivního transportu přenašečem (symport) Taiz and Zeiger 2002
64 transportu látek přes membránu slouží přenašeče přenašeče při přenosu mění konformaci přenos aktivní nebo pasivní Charakteristické znaky: Sekundární aktivní transport membránovými přenašeči má tyto charakteristické znaky: (1) Je poháněn metabolickou energií v podobě protonmotorické síly. (2) Je selektivní -substrátově vysoce specifický. (3) Může probíhat proti koncentračnímu spádu. (4) Vyznačuje se saturační kinetikou, tj. hyperbolickým vztahem mezi rychlostí transportu a koncentrací transportovaného solutu (obr. 33). Tento vztah může být v mnoha případech vyjádřen rovnicí Michaelise a Mentenové. Obr. 33. Aktivní transport a difuze solutu v závislosti na jeho koncentraci. Při velmi nízkých koncentracích se rychlost aktivního transportu zvyšuje proporcionálně se stoupající koncentrací, zatímco při vysokých koncentracích se postupně zpomaluje, až dosáhne maximální rychlosti (Vmax ). Luštinec a Žárský Koncentrace solutu


65 transportu látek přes membránu slouží přenašeče přenašeče Kinetika transportu : difuze vs. přenašeče Taiz and Zeiger 2002


66 transportu látek přes membránu slouží přenašeče sekundární aktivní transport A např. cukr B např. Na+ Taiz and Zeiger 2002
67 transportu látek přes membránu slouží kanály kanály usnadňují pasivní spontánní přenos látek přes membránu zůstávají otevřeny různě dlouhou dobu, spontánně se zavírají různá míra selektivity vlastní přenos látky není spojen se změnou konformace transportního proteinu (otvírání a zavírání ano) Hnací silou pohybu iontů kanálem je gradient jejich elektrochemického potenciálu. Průchodnost kanálů je regulována mechanismem fungujícím jako molekulární branka, která se na podnět určitých signálů otvírá a zavírá. podněty: změna membránového potenciálu, vápenaté ionty, ph, světlo nebo fytohormon Čtvereční mikrometr membrány obsahuje několik set až několik tisíc pump, ale jen I až 10 kanálů. Tok iontů kanálem je tak silný, že lze měřit jeho elektrickou vodivost a stanovit jeho transportní kapacitu. Takové měření lze provést metodou "patch clamp". Kanály se střídavě otvírají a zavírají

68 transportu látek přes membránu slouží kanály kanály - Low affinity K+ channels Kanály pro K + Vnitrobuněčná koncentrace draselných iontů, ovlivňující mnohé životně důležité funkce rostlinné buňky, je regulována především prostřednictvím zvláštních draslíkových tzv. rektifikačních kanálů, které jsou aktivovány určitými membránovými potenciály. (Za nedostatku draslíku v půdě se zvyšuje také aktivita přenašečů draslíku.) V plasmalemě existují kanály pro pohyb draselných iontů směrem do cytoplasmy (tzv. přítokové rektifikační kanály, inward rectifiers) a kanály pro pohyb směrem z cytoplasmy (tzv. výtokové rektifikační kanály, outward rectifiers). Kanály pro K + After depolarization K+ are released to outside. After hyperpolarization K+ are released to inside. Převzato Žárský a Kulich



69 transportu látek přes membránu slouží kanály kanály - Low affinity K+ channels Taiz and Zeiger 2002

70 transportu látek přes membránu slouží kanály kanály Tok iontů kanálem je tak silný, že lze měřit jeho elektrickou vodivost a stanovit jeho transportní kapacitu. Takové měření lze provést metodou "patch clamp". Kanály se střídavě otvírají a zavírají měření s celým protoplastem měření s vyjmutým políčkem plasmalemy Měření elektrického proudu procházejícího iontovými kanály pomocí metody "patch clamp". Skleněná pipeta o průměrem špičky asi 1 mm, obsahující elektrodu a elektrodovou kapalinu, je uvedena ve styk s membránou izolovaného protoplastu (viz kap. o izolovaných protoplastech) nebo buněčné organely (např. vakuoly). Slabým sáním při lne membrána k ústí pipety a její část (patch) může být z protoplastu odtržena. Koncová část pipety má funkci jakési svorky nebo držáku (clamp). Proud procházející iontovými kanály přítomnými v takto izolovaném kousku membrány může být po příslušné amplifikaci měřen. Převzato: Luštinec a Žárský

71 Transport látek přes membránu pumpy přenašeče kanály Pavlová 2005 Přednáška Fyziologie rostlin MB130P MB130P1 14 Katedra experimentální biologie rostlin, Albrechtová, 2011
72 transportu látek přes membránu slouží pumpy přenašeče kanály rychlost transportu kanály přenášejí 106 až 108 iontů za sekundu přenašeče přenášejí 103 iontů za sekundu pumpy přenášejí 102 iontů za sekundu


73 Taiz and Zeiger 2002
74 Příjem minerálních živin a jejich asimilace a využití rostlinou
75 DUSÍK

76 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě
77 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě
78 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě
79 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě
80 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě Nitrosomonas Nitrobacter
81 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě Nitrosomonas Nitrobacter
82 Cyklus dusíku (terestrický) dusík N schéma koloběhu dusíku v biosféře a v půdě Rhizobium Frankia Anabena Nitrosomonas Nitrobacter


83 Dusík - N -nejdůležitější minerální živina (2-5% SH) -Je potřeba v největším množství -Dostupnost dusíku ovlivňuje Řadu fyziologických procesů: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy a rovnováhu metabolismu +N - transport látek v rostlině - obsah prvků a metabolitů v rostlině -N

 - akumulace anthocyaninů obiloviny (purpurové zbarvení")
84 Dusík - N Výskyt v těle rostlin aminokyseliny, proteiny, nukleové kyseliny, porfyriny, kofaktory, fytohormony, alkaloidy Projevy deficience: -zpomalení růstu -chloróza- žloutnutí a opad starších listů - snadno pohyblivý mezi částmi rostliny, recyklace -zdřevnatělé stonky - (přebytek polysacharidů, C:N poměr) - akumulace anthocyaninů obiloviny (purpurové zbarvení listů, stonků)-rajče,

85 Dusík - N Projevy deficience: převzato z přednášky M. Srby Masožravé rostliny -zpomalení růstu -chloróza- žloutnutí a opad starších listů - snadno pohyblivý mezi částmi rostliny, recyklace -zdřevnatělé stonky - (přebytek polysacharidů, C:N poměr) - akumulace anthocyaninů obiloviny (purpurové zbarvení listů, stonků)-rajče,
86 Příjem dusíku NH4+ rychle metabolizován v cytoplazmě (je toxický ve vyšší koncentraci ) vázán na kyselinu glutamovou glutamin více než jeden přenašeč v membráně různá afinita NO3- symport s 2H+ redukce rovnou v kořeni nebo transport a redukce v mezofylu uložení do vakuol transportní systémy s nízkou a vysokou afinitou k NO3-

87 Příjem dusíku Dva typy transportních systémů: s nízkou a vysokou afinitou k NO3-
 Kde?")
")
 X Kde?")
88 Asimilace N - nitrát ue.edu/rhodcv/hort6 40c/nuptake/NR30. gif Nitrátreduktáza (NR) Kde? - cytoplasma, vnější membrána plastidů NO3- + NAD(P)H+ H+ + 2e- NO2- + NAD(P) + H2O Regulace NR: aktivace NO3-, světlem, sacharidy, cytokininy (transkripce, translace) posttranslační modifikace (reverzibilní fosforylace = inaktivace) Nitritreduktáza (Ni NiR) X Kde? chloroplasty (plastidy v kořenech) NO2- + 6Fdred + 8H+ + 6e- NH4+ + 6Fdox + 2H2O Regulace NiR: aktivace NO3-, světlem, inhibice asparagin, glutamin


89 Asimilace N - nitrát - první fáze -redukce NO3-, nitrátreduktáza v cytosolu N+5 N+3 - druhá fáze, nitritreduktáza v plastidu N+3 N-3 Model spřažení elektronového transportu v primární fázi fotosyntézy a redukce dusitanu přes redukovaný ferredoxin
 plastidová (regulace světlem a sacharidy, lokál.")
90 Asimilace N ionty amonné Zabudování NH4+ Glutaminsyntáza (GS) cytoplazmatická (dominantní forma, v kořenech, transport) plastidová (regulace světlem a sacharidy, lokál. spotřeba) Glutamátsyntáza (GOGAT) plastidy
 mitochondrie a chloroplasty Hlavní funkce:")
91 Asimilace N ionty amonné Zabudování NH4+ Alternativní dráha glutamátdehydrogenáza (GDH) mitochondrie a chloroplasty Hlavní funkce: realokace dusíku

92 Asimilace N ionty amonné sumárně asimilace NH4+ NH4+ + glutamát + ATP NH4 ++ glutaminsyntetáza GS glutamin + ADP + Pi glutamátdehydrogenáza 2-oxoglutarát + NAD(P)H GDH glutamát + NAD(P)+ + H2O glutamin + oxoglutarát + NADH glutamátsyntáza GOGAT 2 glutamát + NAD+ asparaginsyntetáza glutamin + aspartát + ATP glutamát + asparagin + AMP + PPi AS aminotransferázy = transaminázy




93 Asimilace N ionty amonné sumárně ve vzorcích
 Syntéza N-bohatých AMK Amidy-")

94 Asimilace N - biosyntéza AMK Transaminace Př.: Aspartátaminotransferáza (aminotransferázy) Kde? - téměř všude ( v cytoplasmě, chloroplastech, mitochondriích, glyoxysomech, peroxisomech) Syntéza N-bohatých AMK Amidy- glutamin, asparagin Asparaginsyntáza (AS) cytoplasma Hlavní funkce asparaginu: důležitá transportní a zásobní forma AMK Hlavní funkce aspartátu: -malát-aspartátový člunek přenáší red. ekviv. z mitochondrií a chloroplastů, účast na metabolismu C4 rostlin v mezofylu, kofaktor vitaminu B6




95 Asimilace N - biosyntéza AMK AMK se sírou glykolýza transaminace v peroxisómu viz fotorespirace syntéza šikimátovou cestou společné s alkaloidy, ligninem viz sekund. metabolity asimilace NH4+

 plasmatická")
96 Asimilace N lokalizace v rostlině Redukce nitrátu: - probíhá v nadzemní i podzemní části (v závislosti na rostlinném druhu a aktuálních ekologických podmínkách) přenašeč nitrátu NO H+ + 8e- NH3 + 2H2O + OH(mocenství N se mění z +5 na 3) plasmatická membrána


97 Asimilace N lokalizace v rostlině Redukce nitrátu závisí na množství přijatého NO3- list Nízký příjem - asimilace v kořeni Zvýšený příjem transport do nadzemních částí - druhově specifické mírné pásmo redukce převážně v kořeni, - tropické a subtropické druhy převažuje redukce v nadzení části kořen



98 formy N v xylému u různých druhů rostlin Druhově specifické allantoin kyselina allantoová
 eukalypty NH4 + citlivé: rajče, brambor, fazol, ječmen pionýrské dřeviny (topol) - toxicita amonného iontu růst, distribuce biomasy příjem")
99 Preference rostlin pro příjem N Nitrát versus amonný iont: - dostupnost v prostředí x preference rostliny NH4 + tolerantní: rýže, ostřice borůvky, brusinky, vřes klimaxové jehličnany (smrk, tsuga) některé krytosemenné dřeviny mírného pásma (dub, buk, habr) eukalypty NH4 + citlivé: rajče, brambor, fazol, ječmen pionýrské dřeviny (topol) - toxicita amonného iontu růst, distribuce biomasy příjem ostatních živin transport látek v rostlině oslabení protonového gradientu!



100 Energetická bilance asimilace N 25% veškeré energie
101 Proč nevyužívají rostliny molekulární dusík (N2), když v atmosféře je ho 78 %? N N
102 Fixace vzdušného dusíku fixace vzdušného dusíku symbióza s prokaryotickými organizmy Přednáška B130P14: Fyziologie rostlin Katedra fyziologie rostlin, UK PřF, doc. Albrechtová
 aktinomycety Frankia (symbióza např.")
103 Fixace vzdušného dusíku fixace vzdušného dusíku symbióza s prokaryotickými organizmy nodulující: bakterie Rhizobium, Sinorhizobium, Bradyrhizobium, Azotobacter (čel. Fabaceae) aktinomycety Frankia (symbióza např. s Alnus, 60 druhů rostlin z 9 čeledí) nenodulující: sinice Anabena (symbióza s Azolla), Nostoc (houby, mechorosty, cykasy,) bakterie - Acetobacter diazotrophicus v apoplastu stonku cukrové třtiny Přednáška B130P14: Fyziologie rostlin Katedra fyziologie rostlin, UK PřF, doc. Albrechtová
 protein syntetizován hostitelskou rostlinou hemová prostetická skupina produkována bakterií")
104 Fixace vzdušného dusíku N ATP + 8 e- +8 H+ nitrogenáza 2 NH3 + H ADP + 16 Pi nitrogenáza inhibována O2 a vznikajícím NH3 leghemoglobin protein s hemem reguluje přístup O2-vysoká afinita k O2 (10x vyšší afinita ke kyslíku než u hemoglobinu červených krvinek) protein syntetizován hostitelskou rostlinou hemová prostetická skupina produkována bakterií Zdroj: Pavlová
 kořeny vylučují látky (flavonoidy) ke kořeni kladná chemotaxe bakterií Vazba na produkt genů bakterie nod nodulujících genů aktivace exprese")

105 Fixace vzdušného dusíku vznik kořenových hlízek nutná exprese specifických genů v obou organismech rostlina: nodulinové geny (Nod) bakterie: nodulační geny (nod) kořeny vylučují látky (flavonoidy) ke kořeni kladná chemotaxe bakterií Vazba na produkt genů bakterie nod nodulujících genů aktivace exprese dalších Kontakt s kořenovým vláskem, depolarizace plasmatické membrány, hydrolýza buněčné stěny tvorba infekčního vlákna a další exprese nodulinových bílkovin, morfogeneze hlízek Přednáška B130P14: Fyziologie rostlin Katedra fyziologie rostlin, UK PřF, doc.albrechtová
")
106 Fixace vzdušného dusíku vznik kořenových hlízek Kladná chemotaxe bakterií Vylučování flavonoidů kořenem Kontakt s kořenovým vláskem Meristematický základ nodulu (hlízky)
 napojení")


107 Fixace vzdušného dusíku vznik kořenových hlízek Meristematický základ nodulu (hlízky) napojení na xylém Přednáška B130P14: Fyziologie rostlin. Vchlípení plasm. membrány, tvorba infekčního vlákna 2007 Katedra fyziologie rostlin, UK PřF, doc.albrechtová




108 Fixace vzdušného dusíku vznik kořenových hlízek peribakteriální membrána z plazmalemy nodulu (rostliny), také bakteriální proteiny Symbiozom ohraničený PBM Bakteroid forma aktivní fixace N2 symbióza hlízkových bakterií s rostlinami čeledi Fabaceae Upraveno: Pavlová Přednáška B130P14: Fyziologie rostlin Katedra fyziologie rostlin, UK PřF, doc.albrechtová



109 symbióza hlízkových bakterií s rostlinami čeledi Fabaceae kůra hlízky s cévními svazky Hlízky růžové zbarvení je dáno leghemoglobinem Credit: Jennifer Dean infikované buňky neinfikované zralá hlízka sklerenchymatická pochva hlízky
110 SÍRA
111 síra přijímána jako SO42- - symport s 3 H+ redukována na S2- nebo vázána jako sulfát (sulfurylové skupiny) O II O- S OII O redukce je energeticky náročná thiolová skupina SH (cystein) - slouží vazbě dalších ligandů vazby v rámci molekuly proteinu (sulfátové můstky; Fe-S centra) cystein, methionon, KoA, vitaminy (thiamin B1)

112 síra glutathion - tripeptid = k. glutamová-cystein-glycin váže a tím inaktivuje endogenní toxiny i exogenní škodlivé látky vazbu katalyzují glutathion-s-transferázy transport komplexu do vakuoly glutathion je stavebním kamenem fytochelatinů (glutamát-cystein)2 až 11- glycin fytochelatiny vážou a tím inaktivují těžké kovy
113 asimilace síry přímá asimilace potřeba ATP redukce redukce je náročná na redukovaný ferredoxin probíhá v chloroplastech (plastidech)
114 DRASLÍK

115 draslík - K+ přijímán jako kation neváže se do stabilních struktur důležitý pro osmotické poměry buněk reverzibilní změny objemu buněk např. při pohybech průduchy, pohyby listů ovlivňuje hydrataci proteinů změny konformace vliv na aktivitu enzymů Buchanan 2000
116 draslík - K+ Příjem větší počet transportních proteinů vysokoafinitní - využívá se protonmotorická síla (symport s H+) nízkoafinitní příjem kanály v rostlině pohyblivý iont nedostatek se projevuje sníženým růstem a světlými skvrnami na starších listech
 - cytosolu a v chloroplastech koncentrace K+ vysoká (80 až 200 mm) - funkčně nezastupitelný jiným kladně jednomocným prvkem (Na+) - buněčný růst - regulace otevřenosti průduchů (regulace")
117 draslík - K+ Význam draslíku v rostlině: -účast na tvorbě membránového potenciálu (hlavní cytosolový kationt) -do jisté míry můž být zastoupen jinými kationty (Na+, Mg2+, Ca2+) -osmotikum (regulace turgoru) - cytosolu a v chloroplastech koncentrace K+ vysoká (80 až 200 mm) - funkčně nezastupitelný jiným kladně jednomocným prvkem (Na+) - buněčný růst - regulace otevřenosti průduchů (regulace rychlosti transpirace) - pohyby rostlin - vyrovnávání náboje v buňce, v xylému - účast v proteosyntéze, ve fotosyntéze, floémovém transportu - ovlivňuje odolnost rostliny vůči stresu



118 draslík - K+ Deficience draslíku: - žloutnutí listů, okrajová nekróza - pokles syntézy proteinů, enzymatické aktivity - snížená odolnost vůči suchu, mrazu, houbovým patogenům K deficientní rajče K deficientní kukuřice
119 VÁPNÍK
 volný a pohyblivý v apoplastu transport z cytosolu do apoplastu zajišťují Ca2+ATPázy vstup do cytosolu kanály difúze")
120 vápník Ca2+ distribuován xylémem redistribuce ze starších částí rostliny do mladších je velmi omezená vázaný vápníkové můstky v buněčné stěně a střední lamele ovlivňuje soudržnost lipidů v membránách stabilizace membrán (interakce s negativními náboji na povrchu membrány) volný a pohyblivý v apoplastu transport z cytosolu do apoplastu zajišťují Ca2+ATPázy vstup do cytosolu kanály difúze cytoplazmou je pomalá tvorba gradientů


121 vápník Ca2+ Příjem vápníku: - pasivní příjem - aktivní čerpání z cytoplazmy (Ca2+ pumpa)
122 vápník Ca2+ transport do apoplastu zajišťují Ca2+ATPázy vstup do cytosolu zajišťují kanály větší počet transportních proteinů reakce na různé signály plazmatická membrána kanály reagují na podněty mechanické změny membránového potenciálu tonoplast kanály reagují na membránový potenciál vazbu ligandů (např. IP3 inositoltrifosfát) difúze cytoplazmou je pomalá tvorba gradientů v buňce v cytoplazmě tvoří komplex s kalmodulinem vazba na proteiny modifikace konformace u rostlin přímá aktivace proteinů (kinázy, fosfatázy) kalmodulinová struktura na C-konci proteinů inhibuje kinázovou aktivitu N-konce vazba Ca2+ na C-konec inhibici zruší

123 vápník Ca2+ kalmodulin Ca2+ - druhý posel při přenosu signálů!!! zesílení signálu přijatého primárním receptorem



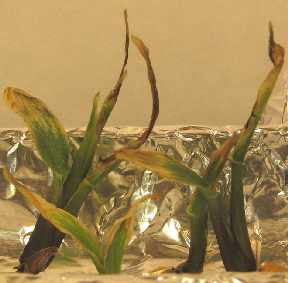

124 vápník Ca2+ Deficience vápníku: -poškození meristemů, -poruchy růstu vegetačních vrcholů
125 HOŘČÍK
126 hořčík Mg2+ - transportován xylémem, redistribuován floémem zásobní forma v semenech sůl kyseliny fytové kyselina fytová - hořčíkové můstky zajišťují správné funkční prostorové uspořádání ve strukturách stálých i dynamických (enzymatické reakce) aktivuje enzymy (např. Rubisco, RNApolymerázy, fosforylační reakce) složka chlorofylů zajišťuje soudržnost podjednotek ribozomů - proteosyntéza nedostatek chlorotické skvrny na listech
 - význam pro aktivitu Rubisca - význam pro transport sacharidů")
 zvýšení koncentrace Mg2+ (vyrovnávání elektrického gradientu vzniklého")
127 hořčík Mg2+ význam ve fotosyntéze: - součást chlorofylu (6-25% celk. obsahu Mg) - význam pro aktivitu Rubisca - význam pro transport sacharidů Aktivace Rubisco Signál: zvýšení ph (ve stromatu!) zvýšení koncentrace Mg2+ (vyrovnávání elektrického gradientu vzniklého přenosem H+ při fotochemii) karbamylace inaktivní aktivní






128 hořčík Mg2+ Deficience hořčíku: - intervenózní chlorózy - pokles rychlosti fotosyntézy, přesto hromadění nestrukturních sacharidů v listech - snížený růst
129 FOSFOR
130 fosfor - P přijímán jako fosfátový anion H2PO4- neredukuje se koncentrace v půdním roztoku 1 µm a nižší v cytosolu 5 až 10 mm rozdíl tří řádů a více význam endomykorhizy pro příjem fosfátu v rhizodermis přenašeč s vysokou afinitou k fosfátovému iontu - často limitující minerální živina O OH P HO O
131 fosfor fosfor -- PP Strategie rostliny k získávání fosfátu: - Zvyšování aktivity vysokoafinitního transportního systému na plazmalemě - Mykorhizní asociace - Změny architektury kořenového systému (délka, větvení) - Zvyšování hustoty kořenových vlásků a jejich délky - Uvolňování organických kyselin a protonů za účelem solubilizace fosforu - Vylučování fosfatáz za účelem uvolňování organicky vázaného fosforu


132 Změny architektury kořenového systému (délka, větvení) - Zvyšování hustoty kořenových vlásků a jejich délky fosfor - P proteoidní kořeny Banksia prionotes, Proteaceae, J-Z Austrálie dauciformní kořeny koř. vlášení Tetraria sp., Cyperaceae, J. Afrika
133 fosfor - P O OH P HO O v rostlině je volný (Pi) nebo vázaný (n. kyseliny, fosfolipidy) asimilován v kořeni i v nadzemní části, ukládán do vakuoly energetický metabolizmus vznik ATP (fotosyntéza, dýchání) ATP, GTP, CTP, UTP, TTP nukleové kyseliny kinázy, fosfatázy, mutázy význam fosforylace změna vnitřní energie aktivita enzymů (např. CDK) záporný pól změna konformace fosforylované molekuly často spojená se změnou lokalizace v buňce (např. transport do jádra) přenos signálů!!! (kinázové kaskády)
134 mikrobiogenní minerální prvky
135 chlór Clvšudypřítomný prvek osmoticky aktivní, v rostlině pohyblivý substituce organických látek (kys. chloroctová) stabilizuje komplex rozkládající vodu (asociován s PSII) významný s K+ při otevírání a zavírání průduchů - obsah v rostlině mg/g SH, skutečný požadavek kolem 0,2-0,4 mg/g SH - deficience při obsahu méně než 0,1-5,7mg/g SH, nad 500 mg/g SH toxicita
 Deficience")


136 chlór Cl- Toxicita chlóru: - rozšířená (aridní oblasti) Deficience chlóru: - snížení růstu - vadnutí listů, intervenózní chloróza - potlačení rozvoje laterálních kořenů - halofytní x nehalofytní druhy - u rostlin podél silnic


137 mikrobiogenní minerální prvky kovy Brady a Weil 2002 rozpětí mezi deficiencí, optimálním zásobením a toxickými kocentracemi mnohdy velmi malé

 -100 mg/g SH, deficience při")


138 železo Fe v půdě jako Fe3+ obtížně rozpustný, tvoří cheláty s organickými látkami (složky humusu, organické kyseliny) -100 mg/g SH, deficience při obsahu méně než mg/g SH, -nad 500 mg/g SH toxicita - v rostlině součástí různých ligandů - účast v oxidoredukčních reakcích v buňce, oxidační stavy Fe(II), Fe(III)
139 železo Fe příjem kořeny vylučují fenolické látky (např. kyselinu kávovou) tvoří komplexy s Fe3+ ionty Fe3+chelátreduktáza na vnější straně membrány rhizodermis - potřeba NAD(P)H Fe se uvolní z komplexu jako Fe2+ do cytosolu při nedostatku Fe se tvoří bohatě větvený kořenový systém čel. Poaceae vylučování fytosideroforů = fytometaloforů, které tvoří s Fe komplexy komplex je transportován přes membránu, v cytosolu je Fe3+ uvolněn fytosiderofor kyselina avenová transport xylémem Fe není v rostlině redistribuován
140 železo Fe Význam železa v rostlině: - význam v přenosu redukčních ekvivalentů v oxido-redukčních reakcích -součást proteinů zajišťujících přenos redukčních ekvivalentů 1. hemové proteiny cytochromy (chloroplasty, mitochondrie) nitrátreduktáza kataláza, peroxidáza (ligniny) 2. Fe-S proteiny ferredoxin SOD (superoxiddismutáza) Rieskeho bílkovina v cytochromu b6f (světelná fáze fotosyntézy) nitritreduktása, sulfitreduktáza akonitása (Krebsův cyklus)

141 Cytochromový komplex b6f důležití hráči při přenosu e- cytochrom b6 Rieskeho protein FeFe-S cytochrom f


142 železo Fe Ferritin zásobní forma Fe v rostlinách, živočiších, bakteriích i houbách - stroma plastidů - proteinový komplex s Fe lokalizovaným uvnitř fytoferritin zásobní forma v chloroplastech, Semenech jádro 5 až 6 tisíc Fe3+ (oxidy, fosfáty), proteinový obal
 slunečnice nedostatek snížená tvorba RNA - narušená polarita buněk - desintegrace")

143 bór +B -B přijímán jako H3BO3 transport xylémem a redistribuce v rostlině strukturní úloha v buněčné stěně váže se na OH- hemicelulóz a fenolických látek (lignin) slunečnice nedostatek snížená tvorba RNA - narušená polarita buněk - desintegrace stonkových meristémů - tvorba dutin ve stonku metabolismu cukrů bylo prokázáno, že se vliv bóru projevuje v transportu glukozy z mladých orgánů do orgánů reprodukčních. Ryant 2003
 - pohyblivost Mn v rostlině malá Význam manganu v rostlině: - význam v")
 komplex vyvíjející kyslík - aktivace enzymů")
144 mangan - příjem ve formě Mn2+, dostupnost ovlivněna ph a redoxními podmínkami (podobně jako Fe) - pohyblivost Mn v rostlině malá Význam manganu v rostlině: - význam v oxido-redukčních reakcích - přímá součást některých enzymů: Superoxiddismutáza (likvidace aktivních forem kyslíku) komplex vyvíjející kyslík - aktivace enzymů toxicita deficience
145 zinek přijímán v komplexech s organickými kyselinami vždy Zn2+ - funkce strukturní zinkové prsty transkripnční faktory enzymy alkoholdehydrogenáza, aldoláza, CuZnSOD stabilita ribozomů integrita membrán
, často")


146 zinek deficience - v přírodě častá (srážkami bohatá stanoviště, vápenité půdy), často společně s deficiencí Fe - zkracování internodií, zmenšování listů - chlorózy mladých listů Toxicita zinku: - kontaminant antropogenního znečištění - zastavení elongace kořenů, chlorózy listů



147 měď +Cu Deficience mědi: - poruchy tvorby reproduktivních orgánů - zasychání špiček listů - pokles lignifikace - sterilita pylu -Cu Deformace klasu ječmene při silném nedostatku mědi

 nemodré proteiny peroxidázy, oxidují mono- a difenoly proteiny obsahující více atomů")
148 měď vázána v komplexech s organickými látkami v půdě i při transportu v rostlině Význam mědi v rostlině: -význam v oxido-redukčních reakcích - 3 skupiny enzymů obsahujících měď: modré proteiny nemají oxidázovou aktivitu, účast v přenosu e- (např. plastocyanin) nemodré proteiny peroxidázy, oxidují mono- a difenoly proteiny obsahující více atomů mědi oxidázy (např. Askorbátoxidáza nebo difenoloxidáza)



149 nikl strukturní složka ureázy enzym důležitý v metabolizmu dusíku (štěpí močovinu = ureu; puriny allantoát, allantoin urea HN4+ + CO2) - požadavek rostliny nízký 0,005 µg/g SH - tolerance je charakteristická u rostlin na serpentinitech - schopnost hyperakumulace, komplexace, sekvestrace tařice zední Ni až 2,5% DW Berkheya coddii
 nitrátreduktáza, sulfitreduktáza")

150 molybden MoO2- - molybdenan Mo+6 rozpustný strukturní i katalytický kofaktor oxidačně redukčních enzymů (Mo+4, Mo+5) nitrátreduktáza, sulfitreduktáza nitrogenáza u prokaryotických symbiotických organizmů - 0,1 µg/g SH, toxicita nad 5 µg/g SH, deficience pod 0,03 µg/g SH Poškození růžice květáku (deficit Mo)
 Toxicita sodíku: Atriplex vesicaria - rozšířená (aridní oblasti) - souvislost se salinitou halofytní x nehalofytní")
151 Benefiční prvky Na, Si, Co, Se, - pozitivní vliv na růst rostlin - zmírnění negativního působení jiného prvku - esenciální jen pro některé druhy SODÍK - esenciální pro některé druhy (Atriplex vesicaria, některé C4 zástupci čeledí Chenopodiaceae, Amaranthaceae, Cyperaceae) Toxicita sodíku: Atriplex vesicaria - rozšířená (aridní oblasti) - souvislost se salinitou halofytní x nehalofytní druhy
4 14-20 mg Si/l, dostupnost závislá na ph 10-15% DW přesličky, vodní")
 1-3% DW suchozemští zástupci trav včetně většiny obilnin, některé dvouděložné méně")
152 Benefiční prvky Na, Si, Co, Se, KŘEMÍK - v půdním roztoku jako kys. křemičitá Si(OH) mg Si/l, dostupnost závislá na ph 10-15% DW přesličky, vodní zástupci trav (např. rýže) 1-3% DW suchozemští zástupci trav včetně většiny obilnin, některé dvouděložné méně než 0,5% DW většina dvouděložných - esencialitu obtížné prokázat (přeslička, rýže, rajče, sója, okurka)
153 Benefiční prvky Na, Si, Co, Se, - pozitivní vliv na růst rostlin - zmírnění negativního působení jiného prvku - esenciální jen pro některé druhy KOBALT - intenzita příjmu a distribuce v rostlině závislá na druhu - požadavek především u rostlin se symbiotickou fixací N projevuje se jako deficience N


154 Benefiční prvky Na, Si, Co, Se, - pozitivní vliv na růst rostlin - zmírnění negativního působení jiného prvku - esenciální jen pro některé druhy SELEN - rozdíly v příjmu i toleranci k vyšší koncentraci Se v substrátu akumulátoři x neakumulátoři - toxicita nad µg Se g-1 SH, kozinec

 - rostliny tolerantní k Al - čajovník akumulátoři - 1000 µg Al/g SH jehličnany - 25-500 µg/g SH čajovník")
155 Toxické prvky Al, Cd, Pb, Cr, Hg HLINÍK Působení hliníku na rostliny: benefiční vliv - VÝJIMEČNĚ toxický efekt -reakce rostlin na přítomnost Al3+ -Al3+ blokuje transportní ka-nály pro Ca2+ a K+ v plazmatické membráně - narušení struktury a dlouživého růstu kořene - citlivé druhy (pšenice, ječmen, čirok) - středně citlivé druhy (kukuřice, hrách, žito) - rostliny tolerantní k Al - čajovník akumulátoři µg Al/g SH jehličnany µg/g SH čajovník akumulátor Al
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
6. Mikroelementy a benefiční prvky. 7. Toxické prvky Al a těžké kovy, mechanismy účinku, obranné mechanismy rostlin
 1. Základní úvod do problematiky Historie studia minerální výživy rostlin, obecné mechanismy příjmu minerálních živin, transportní procesy na membránách. 2. Příjem minerálních živin kořeny rostlin a jejich
1. Základní úvod do problematiky Historie studia minerální výživy rostlin, obecné mechanismy příjmu minerálních živin, transportní procesy na membránách. 2. Příjem minerálních živin kořeny rostlin a jejich
Mineráln. příjem, asimilace a funkce mineráln. lních prvků
 Mineráln lní výživa rostlin příjem, asimilace a funkce mineráln lních prvků 1. Historie studia minerální výživy rostlin, 2. Kriteria nezbytnosti prvků pro rostliny 3. Klasifikace minerálních živin 4. Příjem
Mineráln lní výživa rostlin příjem, asimilace a funkce mineráln lních prvků 1. Historie studia minerální výživy rostlin, 2. Kriteria nezbytnosti prvků pro rostliny 3. Klasifikace minerálních živin 4. Příjem
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch
 Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
- Cesta GS GOGAT - Cesta GDH
 Buchanan 2000 Asimilace amonného iontu: - Cesta GS GOGAT - Cesta GDH Buchanan 2000 GS (glutaminsyntetáza, EC 6.3.1.2) - oktamerní protein o velikosti 350-400 kda, tvořený 8 téměř identickými podjednotkami
Buchanan 2000 Asimilace amonného iontu: - Cesta GS GOGAT - Cesta GDH Buchanan 2000 GS (glutaminsyntetáza, EC 6.3.1.2) - oktamerní protein o velikosti 350-400 kda, tvořený 8 téměř identickými podjednotkami
DOKONČENÍ PŘÍJEM ŽIVIN
 DOKONČENÍ PŘÍJEM ŽIVIN Aktivní příjem = příjem vyžadující energii, dodává ji ATP (energie k regeneraci nosičů) Pasivní příjem = příjem na základě elektrochemického potenciálu (ve vnitřním prostoru převažuje
DOKONČENÍ PŘÍJEM ŽIVIN Aktivní příjem = příjem vyžadující energii, dodává ji ATP (energie k regeneraci nosičů) Pasivní příjem = příjem na základě elektrochemického potenciálu (ve vnitřním prostoru převažuje
Mikroelementy Chlór Bór Železo Mangan Zinek Měď Molybden Nikl
 Prvek Chemický symbol Koncentrace v sušině (µg/g) Koncentrace v čerstvé biomase Makroelementy Dusík Draslík Vápník Hořčík Fosfor Síra N K Ca Mg P S 15 000 10 000 5 000 2 000 2 000 1 000 71,4 mm 17 mm 8,3
Prvek Chemický symbol Koncentrace v sušině (µg/g) Koncentrace v čerstvé biomase Makroelementy Dusík Draslík Vápník Hořčík Fosfor Síra N K Ca Mg P S 15 000 10 000 5 000 2 000 2 000 1 000 71,4 mm 17 mm 8,3
= prvky, které rostlina přijímá jen ve stopovém množství, o to více jsou ale pro ni důležité
 9. Mikroprvky = prvky, které rostlina přijímá jen ve stopovém množství, o to více jsou ale pro ni důležité Mangan Mn - Mnoho různých oxidačních stavů (II a IV nejvíce) - Velikost iontu je podobná Mg a
9. Mikroprvky = prvky, které rostlina přijímá jen ve stopovém množství, o to více jsou ale pro ni důležité Mangan Mn - Mnoho různých oxidačních stavů (II a IV nejvíce) - Velikost iontu je podobná Mg a
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Srovnání obsahů makro- a mikroživin v biomase rostlin
 Srovnání obsahů makro- a mikroživin v biomase rostlin Mangan Příjem, funkce v rostlině, projevy nedostatku Formy Manganu v půdě a rostlinách Mnoho různých oxidačních stavů (II a IV nejčast.) Velikost iontu
Srovnání obsahů makro- a mikroživin v biomase rostlin Mangan Příjem, funkce v rostlině, projevy nedostatku Formy Manganu v půdě a rostlinách Mnoho různých oxidačních stavů (II a IV nejčast.) Velikost iontu
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
MINERÁLNÍ VÝŽIVA ROSTLIN. Minerální živiny Koloběh živin Mechanizmy transportu minerálních živin v rostlině Funkce jednotlivých živin
 MINERÁLNÍ VÝŽIVA ROSTLIN Minerální živiny Koloběh živin Mechanizmy transportu minerálních živin v rostlině Funkce jednotlivých živin Minerální živina prvek, při jehož nedostatku přestávají rostliny růst
MINERÁLNÍ VÝŽIVA ROSTLIN Minerální živiny Koloběh živin Mechanizmy transportu minerálních živin v rostlině Funkce jednotlivých živin Minerální živina prvek, při jehož nedostatku přestávají rostliny růst
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Transport živin do rostliny. Radiální a xylémový transport. Mimokořenová výživa rostlin.
 Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
Úvod do biologie rostlin Transport látek TRANSPORT. Krátké, střední, dlouhé vzdálenosti
 Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Teoretický úvod: MINERÁLNÍ VÝŽIVA. Praktikum fyziologie rostlin. MINERÁLNÍ VÝŽIVA - teoretický úvod 1
 Teoretický úvod: MINERÁLNÍ VÝŽIVA Praktikum fyziologie rostlin MINERÁLNÍ VÝŽIVA - teoretický úvod 1 Teoretický úvod: MINERÁLNÍ VÝŽIVA Vedle prvků, které tvoří organické látky C, H a O funkční struktury
Teoretický úvod: MINERÁLNÍ VÝŽIVA Praktikum fyziologie rostlin MINERÁLNÍ VÝŽIVA - teoretický úvod 1 Teoretický úvod: MINERÁLNÍ VÝŽIVA Vedle prvků, které tvoří organické látky C, H a O funkční struktury
Minerální a organická výživa rostlin
 Minerální a organická výživa rostlin Marie Hronková Přírodovědecká fakulta JčU hronkova@umbr.cas.cz Složení nejdůležitějších stavebních prvků rostlinného těla- prvky esenciální= nezbytné = nenahraditelné
Minerální a organická výživa rostlin Marie Hronková Přírodovědecká fakulta JčU hronkova@umbr.cas.cz Složení nejdůležitějších stavebních prvků rostlinného těla- prvky esenciální= nezbytné = nenahraditelné
Fyziologie rostlin. 8. Minerální výživa rostlin část 3. Ca, Mg a mikroelementy. Alena Dostálová, Ph.D.
 Fyziologie rostlin 8. Minerální výživa rostlin část 3. Ca, Mg a mikroelementy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Min. výživa rostl. Ca, Mg, mikroelementy - vápník,
Fyziologie rostlin 8. Minerální výživa rostlin část 3. Ca, Mg a mikroelementy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Min. výživa rostl. Ca, Mg, mikroelementy - vápník,
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
BIOLOGICKÁ MEMBRÁNA Prokaryontní Eukaryontní KOMPARTMENTŮ
 BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA
 TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty.
 o značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty. Podobné složení živých organismů Rostlina má celkově více cukrů Mezidruhové rozdíly u rostlin Živočichové
o značné množství druhů a odrůd zeleniny ovocné dřeviny okrasné dřeviny květiny travní porosty. Podobné složení živých organismů Rostlina má celkově více cukrů Mezidruhové rozdíly u rostlin Živočichové
FOTOSYNTÉZA Správná odpověď:
 FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
1- Úvod do fotosyntézy
 1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
Rostlinná cytologie. Přednášející: RNDr. Jindřiška Fišerová, Ph.D. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Odborná škola výroby a služeb, Plzeň, Vejprnická 56, Plzeň. Číslo materiálu 19. Bc. Lenka Radová. Vytvořeno dne
 Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
Membránové potenciály
 Membránové potenciály Vznik a podstata membránového potenciálu vzniká v důsledku nerovnoměrného rozdělení fyziologických iontů po obou stranách membrány nestejná propustnost membrány pro různé ionty různá
Membránové potenciály Vznik a podstata membránového potenciálu vzniká v důsledku nerovnoměrného rozdělení fyziologických iontů po obou stranách membrány nestejná propustnost membrány pro různé ionty různá
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
FOTOSYNTÉZA. Princip, jednotlivé fáze
 FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů
 Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů") MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
12-Fotosyntéza FRVŠ 1647/2012
 C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
Fyziologie buňky. RNDr. Zdeňka Chocholoušková, Ph.D.
 Fyziologie buňky RNDr. Zdeňka Chocholoušková, Ph.D. Přeměna látek v buňce = metabolismus Výměna látek mezi buňkou a prostředím Buňka = otevřený systém probíhá výměna látek i energií s prostředím Některé
Fyziologie buňky RNDr. Zdeňka Chocholoušková, Ph.D. Přeměna látek v buňce = metabolismus Výměna látek mezi buňkou a prostředím Buňka = otevřený systém probíhá výměna látek i energií s prostředím Některé
Vyjádření fotosyntézy základními rovnicemi
 FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP. Šárka Poláková
 STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Jaro 2010 Kateřina Slavíčková
 Jaro 2010 Kateřina Slavíčková Biogenní prvky Organismy se liší od anorganického okolí mimo jiné i složením prvků. Některé prvky, které jsou v zemské kůře zastoupeny hojně (např. hliník), organismus buď
Jaro 2010 Kateřina Slavíčková Biogenní prvky Organismy se liší od anorganického okolí mimo jiné i složením prvků. Některé prvky, které jsou v zemské kůře zastoupeny hojně (např. hliník), organismus buď
5. Lipidy a biomembrány
 5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim
 Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník
Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník
Úvod do biochemie. Vypracoval: RNDr. Milan Zimpl, Ph.D.
 Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
05 Biogeochemické cykly
 05 Biogeochemické cykly Ekologie Ing. Lucie Kochánková, Ph.D. Prvky hlavními - biogenními prvky: C, H, O, N, S a P v menších množstvích prvky: Fe, Na, K, Ca, Cl atd. ve stopových množstvích I, Se atd.
05 Biogeochemické cykly Ekologie Ing. Lucie Kochánková, Ph.D. Prvky hlavními - biogenními prvky: C, H, O, N, S a P v menších množstvích prvky: Fe, Na, K, Ca, Cl atd. ve stopových množstvích I, Se atd.
jungle kompletní výživa rostlin Nahlédnutí pod pokličku indabox pro všechny typy pěstebních systémů /mírně odborné pojednání MEDICAL QUALITY GROWIN
 /mírně odborné pojednání kompletní výživa rostlin pro všechny typy pěstebních systémů JungleInDaBox je třísložkový komplex minerálního základu a synergicky působících biologických doplňků. Vysoká efektivita
/mírně odborné pojednání kompletní výživa rostlin pro všechny typy pěstebních systémů JungleInDaBox je třísložkový komplex minerálního základu a synergicky působících biologických doplňků. Vysoká efektivita
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
Fosfor: Projevy deficience P: - 0,2-0,5 % SH rostlin. - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1
 Fosfor: Buchanan 2000-0,2-0,5 % SH rostlin - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1 µm) - toxicita velmi zřídka nad 1% SH (např. Cajanus cajan při 0,3-0,5%, Vigna
Fosfor: Buchanan 2000-0,2-0,5 % SH rostlin - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1 µm) - toxicita velmi zřídka nad 1% SH (např. Cajanus cajan při 0,3-0,5%, Vigna
Předmět: KBB/BB1P; KBB/BUBIO
 Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Energie fotonů je předávána molekulám chlorofylu A, který se zachyceným fotonem excituje (uvolní se energeticky bohatý elektron).
.") Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
Anorganické látky v buňkách - seminář. Petr Tůma některé slidy převzaty od V. Kvasnicové
 Anorganické látky v buňkách - seminář Petr Tůma některé slidy převzaty od V. Kvasnicové Zastoupení prvků v přírodě anorganická hmota kyslík (O) 50% křemík (Si) 25% hliník (Al) 7% železo (Fe) 5% vápník
Anorganické látky v buňkách - seminář Petr Tůma některé slidy převzaty od V. Kvasnicové Zastoupení prvků v přírodě anorganická hmota kyslík (O) 50% křemík (Si) 25% hliník (Al) 7% železo (Fe) 5% vápník
>>> E A1 + E A2. . aktivační energie potřebná k reakci bez přítomnosti katalyzátoru E A E A1. energie potřebná ke vzniku enzym-substrátového komplexu
 Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Průduchy regulace příjmu CO 2
 Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Fotosyntéza (2/34) = fotosyntetická asimilace
 = fotosyntetická asimilace") Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
MitoSeminář II: Trochu výpočtů v bioenergetice. Souhrn. MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK
 MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
Obsah 5. Obsah. Úvod... 9
 Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
Obsah 5 Obsah Úvod... 9 1. Základy výživy rostlin... 11 1.1 Rostlinné živiny... 11 1.2 Příjem živin rostlinami... 12 1.3 Projevy nedostatku a nadbytku živin... 14 1.3.1 Dusík... 14 1.3.2 Fosfor... 14 1.3.3
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Typy molekul, látek a jejich vazeb v organismech
 Typy molekul, látek a jejich vazeb v organismech Typy molekul, látek a jejich vazeb v organismech Organismy se skládají z molekul rozličných látek Jednotlivé látky si organismus vytváří sám z jiných látek,
Typy molekul, látek a jejich vazeb v organismech Typy molekul, látek a jejich vazeb v organismech Organismy se skládají z molekul rozličných látek Jednotlivé látky si organismus vytváří sám z jiných látek,
Sekunda (2 hodiny týdně) Chemické látky a jejich vlastnosti Směsi a jejich dělení Voda, vzduch
 Chemické látky a jejich vlastnosti Směsi a jejich dělení Voda, vzduch") Sekunda (2 hodiny týdně) Chemické látky a jejich vlastnosti Směsi a jejich dělení Voda, vzduch Atom, složení a struktura Chemické prvky-názvosloví, slučivost Chemické sloučeniny, molekuly Chemická vazba
Sekunda (2 hodiny týdně) Chemické látky a jejich vlastnosti Směsi a jejich dělení Voda, vzduch Atom, složení a struktura Chemické prvky-názvosloví, slučivost Chemické sloučeniny, molekuly Chemická vazba
FOTOSYNTÉZA. soubor chemických reakcí,, probíhaj v rostlinách a sinicích. z CO2 a vody jediný zdroj kyslíku ku pro život na Zemi
 Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
 Život rostlin (i ostatních organismů) je neoddělitelně spjat s vodou stálou a nenahraditelnou složkou rostlinného těla. první rostliny vznikly ve vodním prostředí, kde velmi dlouho probíhala jejich evoluce;
Život rostlin (i ostatních organismů) je neoddělitelně spjat s vodou stálou a nenahraditelnou složkou rostlinného těla. první rostliny vznikly ve vodním prostředí, kde velmi dlouho probíhala jejich evoluce;
DEKOMPOZICE, CYKLY LÁTEK, TOKY ENERGIÍ
 DEKOMPOZICE, CYKLY LÁTEK, TOKY ENERGIÍ Vše souvisí se vším Živou hmotu tvoří 3 hlavní organické složky: Bílkoviny, cukry, tuky Syntézu zajišťuje cca 20 biogenních prvků Nejdůležitější C, O, N, H, P tzv.
DEKOMPOZICE, CYKLY LÁTEK, TOKY ENERGIÍ Vše souvisí se vším Živou hmotu tvoří 3 hlavní organické složky: Bílkoviny, cukry, tuky Syntézu zajišťuje cca 20 biogenních prvků Nejdůležitější C, O, N, H, P tzv.
Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová
 autoi: E. Tylová, L. Moravcová") Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová Takto vypadají kontrolní, kultivované v roztoku obsahujícím všechny živiny. Pokud se vaše rostlinka vizuáln liší, kliknte
Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová Takto vypadají kontrolní, kultivované v roztoku obsahujícím všechny živiny. Pokud se vaše rostlinka vizuáln liší, kliknte
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Odběr rostlinami. Amonný N (NH 4 )
") Složka N do půdy N z půdy Spady Export Atmosférický dusík Minerální hnojiva Stájová hnojiva Fixace N Organický dusík Rostlinné zbytky Amonný N + (NH 4 ) Odběr rostlinami Volatilizace Nitrátový N - (NO
Složka N do půdy N z půdy Spady Export Atmosférický dusík Minerální hnojiva Stájová hnojiva Fixace N Organický dusík Rostlinné zbytky Amonný N + (NH 4 ) Odběr rostlinami Volatilizace Nitrátový N - (NO
VODNÍ REŽIM ROSTLIN. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1
 VODNÍ REŽIM ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1 Význam vody pro rostlinu: Rozpouštědlo, transport látek. Účastní se fotosyntézy a dýchání. Termoregulační
VODNÍ REŽIM ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_06_BI1 Význam vody pro rostlinu: Rozpouštědlo, transport látek. Účastní se fotosyntézy a dýchání. Termoregulační
Energetický metabolismus rostlin
 Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
Stavba dřeva. Základy cytologie. přednáška
 Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Vodní režim rostlin. Obsah vody, RWC, vodní potenciál a jeho komponenty: Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy,
 Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
14. Fyziologie rostlin - fotosyntéza, respirace
 14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
Buňka. Buňka (cellula) základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách
 základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách") Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Úvod do biologie rostlin Úvod PŘEHLED UČIVA
 Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
12. Elektrochemie základní pojmy
 Důležité veličiny Elektroda, článek Potenciometrie Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Důležité veličiny proud I (ampér - A) náboj Q (coulomb - C) Q t 0 I dt napětí, potenciál
Důležité veličiny Elektroda, článek Potenciometrie Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Důležité veličiny proud I (ampér - A) náboj Q (coulomb - C) Q t 0 I dt napětí, potenciál
Transport v rostlinách. Kateřina Schwarzerová Olga Votrubová
 Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Ukázky z pracovních listů z biochemie pro SŠ A ÚVOD
 Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Sylabus pro předmět Biochemie pro jakost
 Sylabus pro předmět Biochemie pro jakost Kód předmětu: BCHJ Název v jazyce výuky: Biochemie pro Jakost Název česky: Biochemie pro Jakost Název anglicky: Biochemistry Počet přidělených ECTS kreditů: 6 Forma
Sylabus pro předmět Biochemie pro jakost Kód předmětu: BCHJ Název v jazyce výuky: Biochemie pro Jakost Název česky: Biochemie pro Jakost Název anglicky: Biochemistry Počet přidělených ECTS kreditů: 6 Forma
Vápník. Deficience vápníku: - 0,4-1,5% DW. - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM)
") Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení
 Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení Ročník 1.
Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení Ročník 1.
V organismu se bílkoviny nedají nahradit žádnými jinými sloučeninami, jen jako zdroj energie je mohou nahradit sacharidy a lipidy.
 BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
Chlór. Deficience chlóru: - snížení růstu. - obsah v rostlině mg/g SH, skutečný požadavek kolem 0,2-0,4 mg/g SH
 Chlór - obsah v rostlině 2-200 mg/g SH, skutečný požadavek kolem 0,2-0,4 mg/g SH - deficience při obsahu méně než 0,1-5,7mg/g SH, nad 500 µg/g SH toxicita - spolu s K + hlavní osmotikum buňky Deficience
Chlór - obsah v rostlině 2-200 mg/g SH, skutečný požadavek kolem 0,2-0,4 mg/g SH - deficience při obsahu méně než 0,1-5,7mg/g SH, nad 500 µg/g SH toxicita - spolu s K + hlavní osmotikum buňky Deficience
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Přednáška 6: Respirace u rostlin
 Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Lékařská chemie a biochemie modelový vstupní test ke zkoušce
 Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
B4, 2007/2008, I. Literák
 B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované