EPIGENETICKÁ DĚDIČNOST
|
|
|
- Bohumila Martina Kadlecová
- před 6 lety
- Počet zobrazení:
Transkript

1 Epigenetika
2 EPIGENETICKÁ DĚDIČNOST Pojmenovaná britským biologem Conradem Waddingtonem r. 1940, Něco navíc, mimo v porovnání s mendelistickou genetikou, Umožňuje buňkám s identickým genotypem vznik odlišných fenotypů a přenos informace do dalších buněk, Epigenetické faktory ovlivňují fenotyp bez změny primární struktury genotypu, t.j. epigenetická informace není zakódovaná v sekvenci nukleotidů
3 Samotný epigenom je tvořen během buněčné diferenciace v embryonální fázi, kdy se z pluripotetních buněk vyvíjejí buňky specializované Diferenciační proces je ve změně exprese genu, aktivní geny se mohou vypnout a neaktivní geny zapnout, výsledkem je diferencovaná buňka. Tato buňka nese tzv. vzor genové exprese, který je dědičný a specifický pro daný buněčný typ. Předávání informace je podmíněno buněčnou pamětí. Studium epigenetické dědičnosti: EPIGENETIKA.
4 Epigenetická dědičnost a mendelistická dědičnost Neplatí Mendelova pravidla Neúplná penetrance přítomnost dominantní alely genu neznamená nutně a vždy jeho projev Variabilní expresivita Exprese genů může být různá v různých tkáních = mozaikový fenotyp Platí Mendelova pravidla Úplná penetrance přítomnost dominantní alely genu znamená jeho projev Stabilní expresivita Exprese genů je stejná pro daný typ tkáně
5 Příklad mozaikového fenotypu Za vznik zbarvení tzv. želvovinových koček je zodpovědná inaktivace chromozómu X vyvolaná dlouhou nekódující RNA jménem XIST.

6 Příklad mozaikového fenotypu Inaktivace chromozómu X s různou alelou má za následek tříbarevné zbarvení u koček 6
7 Spontánní a reverzibilní. Epigenetické procesy Epigenetické mechanismy zasahují na úrovni: A. transkripční aktivity genů: 1. DNA metylace (CpG metylace) 2. Modifikace histonů (acetylace a deacetylace histonů) B. post-transkripční aktivity genů: 1. RNA interference (RNAi)
8 DNA metylace Typ modifikace, která je děděna bez změny sekvence DNA. Stabilizuje kondenzovanou konformaci chromatinu a tak udržuje geny v inaktivním stavu. Má důležitou roli v genomovém imprintingu a inaktivaci chromozómu X. Podstatou metylace je přidání metylové skupiny na cytosin v CpG párech za vzniku 5 metylcytosinu za přítomnosti enzymu DNA metyltransferázy (DNMTázy). Některé DNMTázy metylují DNA de novo, ale většina metyluje jen nemetylované vlákno v hemimetylované DNA, tzv. dědičná metylace. CpG jsou seskupené do CpG ostrůvků, přítomné v oblasti promotoru, regulační oblast pro mnoho genů.
9 Metylace DNA se podílí na umlčování genů. Abnormální hypermetylace CpG v promotoru tumor-supresorových genů v maligním nádoru vede k utlumení transkripce tumorsupresorových genů. Cílená léčba nádorových onemocnění spočívá v inhibici DNA metyltransferáz a dosažení exprese tumor-supresorových genů.
10 Modifikace histonů Samotná struktura chromatinu je důležitá pro regulaci transkripce. Modifikace histonů vede k vytvoření transkripčně neaktivního heterochromatinu. Velký význam má NH 2 skupina lysinu, která má kladný charakter, proto se může vázat s negativně nabitými fosfáty v DNA. Ionovou vazbou se DNA v oblasti lysinu váže těsně s histony a tím dochází k blokování transkripce. Modifikace histonů a tedy také úroveň transkripce je regulovaná pomocí deacetyláz a acetyláz.
11 Mechanismus acetylace a deacetylace lysinu Účinkem deacetyláz dochází k tvorbě inaktivního chromatinu a tím zastavení transkripce genu. Acetylázy eliminují pozitivní náboj lysinů v histonech a tím se poruší těsná interakce DNA-histon. Transkripční faktory mají přístup k DNA a transkripce genu může začít. Mechanismus acetylace lysinu a vliv na vazbu histonu k DNA
12 RNA interference, RNAi (rušení RNA)
13 RNA interference Nedávno objasněný proces. Regulace transkripce a vnitrobuněčné exprese genu. Fragmenty dvoušroubovicové RNA (dsrna) interferují s exprimovanými geny. První experimenty cíleně využívající RNAi proběhly na začátku 90. let minulého století na rostlinných modelech, a brzy se začala využívat i pro studium dalších modelových organismů.
14 RNAi je vysoce účinným a specifickým procesem, který je aktivně vykonáván určitým buněčným mechanismem. Ačkoliv není detailně popsána, zdá se, že jakmile nalezne dvoušroubovicovou molekulu RNA, rozdělí ji na několik částí, oddělí od sebe jednotlivá vlákna dvoušroubovice, dále mechanismus pokračuje destrukcí dalších jednotlivých vláken RNA, které se ukáží být komplementární k nalezenému segmentu.
15 Životní cyklus mnoha virů zahrnuje fázi kdy je přítomná dvoušroubovicová RNA, takže je velmi pravděpodobné, že mechanismus RNAi je součástí obranného mechanismu před těmito viry. Znalost RNAi a budoucí praktické používání je slibné pro budoucí léčení dosud neléčitelných onemocnění již na molekulární bázi. Šlo by obdobu genové terapie, při které by však nedošlo k trvalé změně DNA pacienta, ale exprese nechtěného genu by byla znemožněna v následující fázi. Výsledky prvních testů na myších jsou slibné.
16 Význam některých pseudogenů jejich transkripty mohou vychytávat molekuly interagující RNA!
17 17
18 18

19 Mechanismus RNA interference dsrna je rozštěpena enzymem Dicer na molekuly malé interferující RNA (sirna). sirna se váže na nukleázový komplex a vzniká RISC (RNA induced silencing complex). RISC je aktivován rozpletením sirna duplexu a na principu komplementárního párování bazí může RISC zacílit na homologní transkript. Transkript je potom rozštěpen na 12 nukleotidové úseky. Mechanismus štěpení však není znám. Obrázek: Mechanismus RNAi, z olekylaer_metabolism/sm_in_vitro
20 RISC RNA-induced silencing complex je ribonukleoprotein, komplex několika proteinů a molekuly RNA. Komplementárně se váže na cílovou molekulu mrna a štěpí ji, případně alespoň blokuje translaci. Rozeznává se mirisc, který obsahuje navázanou mirna sirisc, jeho součástí je sirna.
21 RISC mirisc obvykle blokuje translaci, ale v případě perfektní komplementarity je možná i degradace mrna. mirna je kódována buňkou, ať už samostatnými geny pro mirna, oblastmi uvnitř intronů genů kódujících protein nebo geny pro jinou nekódující RNA. mirna reguluje genovou expresi tak, že zabraňuje translaci těch mrna, se kterými nedokonale páruje (příklad viz obrázek). Tento mechanismus se nazývá RNA silencing. sirisc typicky navozuje přímo degradaci cílové mrna, ale je možná i represe translace. sirna vzniká z dvouvláknové RNA především vnějšího původu (většinou dsrna viry).
22 RISC mimo RNA obsahuje zejména tři důležité proteiny, dicer, TRBP a Ago (argonaut), které jsou schopné i v nepřítomnosti vhodné RNA trimerizovat a vytvářet společně komplex. Dicer je ribonukleáza schopná štěpit dsrna (buď pre-mirna z níž vzniká mirna, nebo vhodnou dsrna z níž vzniká sirna) na krátké fragmenty. Jedno konkrétní vlákno takového krátkého fragmentu se následně spolu s proteiny TRBP a Ago stane součástí RISC komplexu. Ago, čili protein z argonautové superrodiny proteinů, obsahuje PIWI doménu, která je schopná přesně štěpit cílovou mrna a představuje tedy vlastní katalytický princip funkce RISC komplexu v degradaci RNA.
23
24 RNA interference Pokud dojde k destrukci mrna, neproběhne translace, nedojde k tvorbě genového produktu. Procesem RNAi dochází k umlčení genů na post-transkripční úrovni. Nejznámější studie na modelovém eukaryotickém organismu Caenorhabditis elegans.
25 Háďátko obecné (Caenorhabditis elegans) Volně žijící nepatogenní půdní helmint z kmene hlístic. Háďátko žije v půdě po celém světě a je významným modelovým organismem, jehož výzkum započal v roce Jedná se rovněž o první mnohobuněčný organismus, u něhož byl osekvenován kompletní genom. Rovněž byl poprvé u tohoto druhu prezentován fenomén RNA interference.
26 sirna small interfering RNA, short interfering RNA, silencing RNA Skupina dvouvláknových RNA o délce nt. Uplatňují se v RNA interferenci ovlivňují expresi určitého genu. Dále hrají roli i v dalších procesech souvisejících s RNA interferencí, jako je ochrana před viry. Pravděpodobně také ovlivňují prostorovou strukturu chromatinu.

27 sirna má typickou stavbu, skládá se obv. z 21 nt, je dvouvláknová, na jedné straně každého vlákna přečnívají dva nukleotidy, nespárované s nukleotidy vlákna druhého. Na 5' konci je fosfátová skupina, na 3' konci hydroxylová skupina. Tato struktura je určena enzymatickým účinkem proteinu dicer, ten je schopný nastříhat dlouhé dvouvláknové řetězce RNA, ale také small hairpin RNA na malé kousky.
28 sirna byly původně považovány za exogenní, tzn. jako molekuly vznikající rozštěpením například virové n. transpozonové RNA. Základní sirna vznikají rozštěpením těchto dlouhých a dvouvláknových molekul RNA. Dnes je však známo, že sirna mohou vznikat i přepisem částí genomu, např. v centromerických nebo repetitivních oblastech DNA. Některé sirna vznikají i štěpením určitých částí molekul mrna.
29 Funkce sirna V typickém případě, v kanonické sirna dráze, se sirna asociuje s proteinovým komplexem RISC a navádí ho ke konkrétnímu úseku mrna, jenž je s danou sirna plně komplementární. RISC pak katalyzuje přesné rozštěpení této cílové mrna. Díky tomu dochází k tzv. posttranskripčnímu umlčení, silencingu daného genu. Gen se sice trankribuje, ale jeho mrna je následně štěpena, bílkovinný produkt nevzniká. Byly však odhaleny i způsoby, jak sirna blokuje samotný přepis genu, a to mechanismy, jimiž navozuje vznik heterochromatinu, který není přepisován.
30 Kanonický mechanismus RNA interference, typický pro sirna. Žlutě je RISC komplex.
31 mirna Nekódující malé endogenní RNA. Negativně regulují expresi cílového genu: degradací mrna jež obsahuje komplementární sekvence ztížením translace změnou exprese cílového genu epigenetickými mechanismy jako např. metylace promotoru. Jsou běžně přítomné v eukar. buňkách. Jejich exprese je regulována časově specifickým a tkáňově typickým způsobem podobně u různých druhů. Tato evoluční konzervace mirna naznačuje, že hrají důležitou roli v různých biologických a buněčných procesech.
32 Funkce mirna spočívá v regulaci genů a jejich exprese. Molekuly mirna jsou komplementární k části jedné nebo několika konkrétních mrna. Živočišná mirna vykazuje komplementaritu obvykle k regionu 3 UTR (část mrna nekódující proteiny, ale vykonávající některé jiné regulační funkce vztahující se k dané molekule mrna). Rostlinná mirna je komplementární ke kódujícím regionům messenger RNA.
33 Když se spárují odpovídající řetězce mirna a mrna, je obvykle inhibována translace této mrna v protein. Někdy je namísto toho usnadněn rozklad molekuly mrna. V tomto případě zřejmě vznik dvouvláknové RNA navozuje v buňce proces podobný RNA interferenci způsobované sirna molekulami. mirna může zřejmě také zasáhnout DNA, která koresponduje s danou mrna, na níž se mirna navázala - v tomto případě fungují mirna spolu s proteiny, označovanými jako mirnp, microribonuclear proteins.
34 mirna Jednovláknové řetězce nekódující RNA o délce nt, podílí se na regulaci genové exprese. Vznikají transkripcí z genů v DNA, ale následně nedochází k translaci. Namísto toho se každý primární transkript mirna, pri-mirna, páruje s některými vlastními komplement. bázemi a nakonec se mění na plně funkční mirna. Tyto molekuly jsou částečně komplementární k určitým molekulám mrna v buňce a jsou schopné regulovat, snižovat produkci proteinů, které tyto mrna kódují. MiRNA se vyskytují zejména u rostlin a živočichů, ale vyvinula se u těchto skupin asi nezávisle.
35 Stačí částečná sekvenční shoda mezi mirna a mrna. Jedna mirna může regulovat více mrna. Jedna mrna může být regulována více mirna.
36 mirna Mohou působit jako důležité regulátory vývoje orgánů, diferenciace a proliferace buněk, apoptózy, buněčné smrti. Dále odpovědi na poranění a imunitní odpovědi na různé nemoci vč. rakoviny, diabetes, kardiovaskulárních chorob a infekcí. mirna mají původ v transkriptech, které se skládají a vytváří typické vlásenkové struktury. Jiné typy malých RNA se odvozují z mnohem delších vlásenek, důsledkem je vznik mnoha různých malých RNA (sirna), nebo z bimolekulárních RNA duplexů (sirna), nebo z prekurzorů bez jakékoliv dvoušroubovicové struktury (pirna, Piwi-interacting RNA.)
37 Vznik a úpravy mirna Geny kódující mirna jsou mnohem delší než finální mirna. Gen pro mirna je v jádře přepsán polymerázami do podoby asi 70 nt dlouhého řetězce pri-mirna s čepičkou na 5' konci a poly-a koncem 3 konci. První úpravy obstarává u živočichů proteinový microprocessor complex. Skládá se z nukleázy nazývané Drosha a proteinu Pasha, schopného vázat na sebe dsrna. Tento komplex mění pri-mirna na pre-mirna.
38 Vznik a úpravy mirna Následně pre-mirna vstupuje do cytoplazmy. Interaguje s endonukleázou naz. Dicer za vzniku zralého mirna duplexu. Ten je zabudován do mirnaindukovaného umlčujícího komplexu, mirna-induced silencing complex, mirisc, zde je jedno vlákno mirna duplexu degradováno (mirna*), druhé je zachováno. Spojením mirisc a zralé mirna je komplex aktivován. mirna zavede na cílovou mrna, ta je umlčena buď degradací, nebo represí translace. RISC je tedy schopen utlumovat expresi genů, jev známý jako RNA interference. U rostlin je celá kaskáda vzniku mirna mírně odlišná, není přítomen protein Drosha a jeho roli zastává Dicer.


39
40 mirna V lidském genomu se odhaduje ca 1000 mirna, společně řídí expresi až 60% genů!!! První mirna lin4 popsána u C. elegans v r Některé mirna geny jsou uvnitř intronu strukturního genu a jsou ko-transkribovány spolu s hostitelským genem. Většina mirna však sídlí v oblastech nekódujících proteiny a má vl. nezávislou transkripční jednotku vč. promotoru a terminátoru. Transkripce intronických mirna a většiny intergenových mirna pomocí polymerázy II. Transkripce je možná i polymerázou III.
41 Piwi-interacting RNA pirna Největší třída malých nekódujících RNA v živočišných buňkách. pirnas formují komplexy RNA - piwi proteiny. Tyto pirna komplexy jsou spojeny jak s epigenetickým tak post-transkripčním umlčováním genů (retrotranspozonů) a dalších genetických elementů v zárodečných buňkách, zejména ve spermatogenezi. Liší se od mirna velikostí (26-31 nt), nepřítomností sekvenční konzervace a vyšší komplexitou. Je nejasné, jak jsou pirnas tvořeny, ale možné způsoby byly navrženy. Biogeneze je odlišná od mirna, sirna. rasirna jsou poddruhem pirna.
42 pirnas identifikovány u obratlovců i bezobratlých. Ačkoliv se biogeneze a způsob účinku mezi druhy poněkud liší, řada procesů je konzervována. pirnas nemají jasné sekundární strukturní motivy. Délka nt a převaha 5 uridinu je společná pro pirna u obratl. i bezobr. pirna z C. elegans má 5 monofosfát a 3 modifikace, které působí jako blok 2 nebo 3 kyslíku, potvrzeno i u D.m., zebrafish, myši, krysy. Důvod modifikace není znám, snad zvyšuje pirna stabilitu. Pravděpodobně existuje několik set tisíc různých pirna u savců. Dosud odhaleno 50 tis. pirna u myši a 13 tis. u D.m.
43 pirnas je v klastrech v celém genomu, ty asi obsahují deset až několik tisíc pirnas a jejich velikost kolísá od 1 do několika set kb. Zatímco klastrování pirnas je mezi druhy silně konzervováno, sekvence nikoli. U D.m. a obratlovců jsou pirnas v oblastech bez protein kódujících genů, u C. elegans byly identifikovány uprostřed takových genů. U savců jsou pirna přítomny ve varlatech i vaječnících, ačkoli jsou potřebné jen u samců. U bezobratlých byly nalezeny v samčí i samičí zárodečné linii. Na buněčné úrovni byly pirna nalezeny v jádru, cytoplazmě. To naznačuje, že mají funkci v obou těchto oblastech a mají četné efekty.
44 Biogeneze pirna není zcela popsána. pirnas má významné rozdíly mezi vlákny, tedy je odvozena jen od jednoho vlákna DNA. To naznačuje, že je produktem dlouhých jednovláknových prekurzorových molekul. Možná existuje cesta k produkci pachytene pirnas: pirna prekurzory jsou transkribovány do pirna s tendencí cílit na 5 uridin. Dále existuje hypotéza Ping Pong mechanismu. Primární pirnas rozeznávají jejich komplementární cíle a umožní získat piwi proteiny. Výsledek štěpení transkriptu v místě deset nt od 5 konce primární pirna, což produkuje sekundární pirna. Ty jsou zacíleny na sekvence s adeninem na desáté pozici. Jelikož pirna účastná v ping pong cyklu směřuje svůj atak na transkripty transpozonů, ping pong cyklus působí pouze na úrovni transkripce. Jeden nebo oba tyto mechanismy působí u různých druhů. Např. C. elegans má pirnas, ale vůbec nepoužívá mechanismus ping pong. Značný počet pirnas u zebrafish a D.m. má adenin na 10. pozici. To se považuje za možný důkaz konzervovaného biosyntetického mechanismu napříč druhy. Znak ping pongu byly identifikovány u velmi primitivních organismů (houby, medúzy, polypy), to naznačuje existenci ping pong cyklu již u raných metazoí (živočichů).
45 Ping Pong Poprvé popsán u Drosophila. pirna asociovaná se 2 cytoplazmatickými piwiproteiny aubergin Aub a argonaut-3 Ago3 vykázala vysokou frekvenci sekvenční komplementarity přes přesně 10 nt na jejich 5 koncích. To je tzv. ping pong signatura, vyskytuje se i u pirna asociované s Mili a Miwi proteiny z myších varlat. Pravděpodobná funkce ping pongu u drozofily nebo myši není objasněna. Hypotéza je, že interakce mezi Aub a Ago3 dovoluje cyklické úpravy pirna, které jsou účelné pro zacílení na aktivní transpozonové sekvence. Aub pirna jsou antisense k transkriptům transpozičních elementů a jsou považovány za hlavní faktor při zacílení na škodlivé transkripty na principu komplementarity. Naopak, Ago3 pirna sekvence mají převážně sense orientaci k transkriptům transpozičních elementů, jsou odvozeny z produktů transpoziční mrna štěpené Aub. Jako taková Ago3 pirna nemá schopnost cílit přímo na transkripty transpozičních elementů.
46 Zřejmě Ago3 pirna řídí produkci pirna, které jsou nakládány na Aub prostřednictvím zacílení na nově exportované transkripty pirna klastrů. Některé důkazy ukazují na účinek Ago3 na produkci Aub pirna, zejména na základě testování pirna ve vaječnících drozofily. Molekulární mechanismus v základech ping pongu pravděpodobně obsahuje faktory spojené s pirna dráhou.
47 pirna Phasing Dráha pirna u Drosophila může být rozdělena do dvou větví: -cytoplazmatická, tj. Aub a Ago3, které řídí ping-pong mechanismus; -jaderná, týkající se ko-transkripčního umlčování genomových lokusů účinkem piwi v jádře. Štěpení provedené Aub a Ago3 spouští časované vložení pirna do Piwi. Časování začíná zacílením a štěpením komplementárního cíle buď proteinem Aub, nebo Ago3 asociovaného s pirna. Jakmile je rozštěpen, zacílený transkript je procesován mechanismem, který vyžaduje endonukleázu asociovanou s mitochondriemi, Zucchini, což vede k spojení Piwi proteinu se sekvenčními fragmenty transkriptu, na nějž je cíleno. Tímto způsobem Aub nebo Ago3 pirna sekvence štěpí komplementární cíl, který je potom nakrájen na periodické intervaly ca 27 nt, ty jsou poté sekvenčně spojeny s Piwi proteinem.
48 Proces působení pirna Jakmile je spojena s pirna, vstupuje Piwi do jádra zárodečných buněk a kotranskripčně umlčuje vznikající transkripty komplementární ke svému pirna průvodci. Aktuálně není známo, zda se fázování odehrává v jiných organismech.
49 Funkce Velká variabilita v sekvencích pirna a funkcích piwi u různých druhů je důvodem obtížného určení funkce pirnas. Podobně jako ostatní nekódující malé RNAs jsou pirna považovány za účastníky umlčování genů, specificky umlčování transpozonů. Většina pirnas je antisense k transpozonovým sekvencím, což ukazuje, že transpozony jsou jejich cílem. U savců se ukazuje, že aktivita pirnas v umlčování transpozonů je nejdůležitější během embryonálního vývoje a jak u C. elegans, tak u člověka jsou pirnas nezbytné pro spermatogenezi.
50 Umlčování RNA pirna má roli v umlčování RNA a to prostřednictvím utváření RISC (RNA induced silencing complex). pirnas interagují s piwi proteiny, které jsou součástí rodiny proteinů tzv. Argonautů. Ty jsou aktivní ve varlatech savců a jsou potřebné pro vývoj zárodečných buněk a kmenových buněk u bezobratlých. Tři podrodiny piwi proteinů, MIWI, MIWI2 s MILI, jsou nezbytné pro spermatogenezi u myší. pirnas směrují piwi proteiny na jejich transpozonové cíle. Pokles nebo chybějící exprese PIWI genů je korelována se zvyšující se expresí transpozonů. Transpozony mají vysoký potenciál poškozovat své hostitele a mutace v drahách pirna způsobují snížení fertility u D.m. Takové mutace však nemají podobný efekt u myši, to naznačuje redundance v pirna systému.
51 Umlčování RNA Dále, předpokládá se, že pirna a malé endogenní interferující RNA (endo-sirna) možná mají podobnou a dokonce redundantní funkci v kontrole transpozonů v savčích oocytech. pirna zřejmě mají vliv na některé metyltransferázy provádějící metylace, jež jsou vyžadovány pro rozpoznání a umlčení transpozonů, ale proces není dobře popsán.
52 Epigenetické efekty pirnas mohou být přenášeny maternálně, a podle výzkumu na D.m. mohou být účastny na maternálních epigenetických pochodech. Aktivita specifických pirnas v epigenetickém procesu rovněž vyžaduje interakce mezi piwi proteiny a HP1a a dalšími faktory. HP1a je konzervovaný eukaryontní chromosomální protein, asociovaný s pericentrickým heterochromatinem.
53 RNA interference je známá pouze u eukaryot. Řada bakterií a archeí má pro obranu proti bakteriofágům dalším parazitickým DNA elementům vyvinut funkční analog RNA interference - CRISPR systém. 53
54 Genomový imprinting (otisk) Imprintované geny jsou clusterovány na chromozomech, což naznačuje, že mechanismy imprintingu působí spíše v lokálních chromozómových doménách, než v jednotlivých genech. Je významným faktorem v embryonálním vývinu savců. Reverzibilní proces, specifická modifikace genů v parentální generaci vede k funkčním rozdílům mezi paternálními a maternálními genomy v diploidních buňkách potomstva. Jednoznačným důkazem geny, jejichž alely jsou aktivní jen pokud mají paternální nebo maternální původ, t.j. jejich exprese závisí výhradně na pohlaví rodiče, od kterého byla alela zděděna. Tento genetický jev vnáší rozpory do základních pravidel Mendelovské genetiky.
55 K vytvoření imprintu (t.j. informací o expresi nebo inaktivaci transkripce) dochází v průběhu meiotického dělení nebo tvorby gamet a jeho mechanismem je pravděpodobně metylace DNA. Genomický imprinting je výsledkem odlišného stupně metylace cytosinu na 5-metylcytosin u genu na maternálním - paternálním chromozomu. Změna metylace genu může nastat v zygotě, kdy dochází k novým vztahům mezi paternálními a maternálními chromozomy. Genomický imprinting může mít negativní i pozitivní účinky na eukaryotický organismus.
56 Negativní účinek genomového imprintingu Genomový imprinting má vliv na vznik různý genetických chorob. Např. Angelmanův syndrom Je to poškození v imprintované oblasti na 15. chromozomu člověka. Toto poškození je děděno od matky. Příznaky: hyperaktivita, absence řeči, problémy s přijímáním potravy. Prader-Williho syndrom Je to poškození ve stejné oblasti jako u Angelmanova syndromu, ale je zděděno od otce. Příznaky: nadměrný příjem jídla a slabý svalový tonus.
57 Genomický imprinting se uplatňuje u syndromů Angelman a Prader-Willi. Oba geny jsou lokalizovány na dlouhém raménku chromozómu č. 15, v oblasti 15q11-q13. Každý z těchto syndromů může být způsoben různými genetickými defekty: mikrodelecí, uniparentální disomií a defekty imprintingu. U Angelmanova syndromu se v 70% případů jedná o deleci maternálního 15q11-q13 (v této oblasti se gen UBE3A zodpovědný za AS vyskytuje). Otcovský gen je imprintován a nemůže se tedy exprimovat. Podstatou imprintingu je metylace bazí otcovského genu, který se nemůže funkčně projevit. Ve 3-5% případů se jedná o tzv. parentální disomie, to znamená., že oba geny jsou původu od jednoho rodiče, u Angelmannova syndromu otcovského, jedná se imprintované geny, nemohou se tedy projevit. Ve 2-5% případů se předpokládá mutace v imprintingovém centru a v 10-15% UBE3A mutace. U syndromu Prader Willi se jedná o velmi podobnou situaci, imprintovány jsou však alely maternální. V 70% se jedná o mikrodeleci, v tomto případě paternálního úseku 15 q11-q13, v 20-25% maternální disomie a ve 2-5% mutace imprintingového centra. Mikrodelece maternální u AS a paternální u PWS lze prokázat metodou FISH či molekulárně genetickými metodami. Parentální disomie lze prokázat pouze molekulárně geneticky. Bodové mutace v metylačním centru u nás zatím nejsme schopni diagnostikovat.
58 Uniparentální dizomie (UPD) je přítomnost dvou homologních chromozomů nebo jejich částí, které pocházejí od jednoho rodiče.

59
60 Mechanismus vzniku Angelmanova syndromu Imprintován je otcovský gen Del 15q11-q13 UPD IC mutace UBE3A mutace P M P P P M P M 70% 3-5% 2-5% 10-15% gen imprintován gen imprintován gen imprintován gen imprintován UPD - uniparentální disomie oba chromozomy od otce IC mutace imprintingového centra UBE3A mutace genu pro ubiquitin 3A Černě je vyznačen paternální chromozom, šedě maternální chromozom
61 Mechanismus vzniku Prader-Willi syndromu Imprintován je mateřský gen Del 15q11-q13 UPD IC mutace P M M M P M 70% 25-28% 2-5% gen imprintován gen imprintován gen imprintován UPD - uniparentální disomie oba chromozomy od matky IC mutace imprintingového centra Černě je vyznačen paternální chromozom, šedě maternální chromozom
62 Angelmanův syndrom nelze vyléčit, ale je možné zmírnit jeho projevy. Mezi typické známky AS patří opožděný psychomotorický vývoj, mentální retardace, problémy s koordinací pohybů a rovnováhou, hyperaktivita, poruchy spánku, poruchy pozornosti, epilepsie a neschopnost mluvit. Mají typický usměvavý výraz v obličeji, širokou opatrnou chůzi a jakékoliv vzrušení vyjadřují máváním rukama. Jsou velmi společenští, milují vodu a jakékoliv předměty vydávající zvuky nebo světlo. Jsou zvídaví, proto je zajímá dění v okolí a zkoumají vše co je na dosah. Mentální retardace je bohužel středně těžká až těžká. Během svého života dosáhnou mentálního věku maximálně na úrovni 3letého dítěte. Z tohoto důvodu nejsou schopni samostatného života a potřebují neustálý dozor a péči dospělé osoby. Díky své zvídavosti a lepší schopnosti soustředit se ve vyšším věku jsou schopni učit se neustále novým dovednostem.
63 Angelmanův syndrom patří mezi mikrodeleční syndromy. Onemocnění se diagnostikuje analýzou DNA, toto vysoce specializované vyšetření se v současnosti v ČR provádí na třech specializovaných pracovištích v Praze a Olomouci. Mikrodeleční syndromy jsou způsobeny delecí malých chromozomálních úseků, jde tedy o strukturní aberace. Angelmanův syndrom je způsoben delecí úseku 15q11-13 na maternálním 15. chromozomu (od matky), případně uniparentální disomií (zdvojení genetického materiálu otcovského chromosomu). Tyto změny jsou detekovatelné ca u 70 % případů dětí s AS. U 5-7% případů s AS je přítomná mutace tzv. UBE3A genu. Pokud dítě zdědí tuto mutaci od matky, onemocní Angelmanovým syndromem, pokud ji však zdědí od otce, tak nikoliv (tato mutace nemá detekovatelný účinek na dítě). V některých rodinách s touto mutací se AS může vyskytovat u více členů rodiny. U přibližně 15% případů dětí s AS se genetickým vyšetřením neprokáže žádná porucha, všechny testy jsou negativní, u cca 4 6% se popíše jiný typ genetického defektu. Poznání přesné genetické příčiny Angelmanova syndromu je velice důležité pro prenatální diagnostiku a pro genetické poradenství v rodině.
64 Praderův Williho syndrom (PWS) je vzácné genetické onemocnění charakteristické nezvladatelnou chutí k jídlu, malým vzrůstem, hypogonadismem a mírnou mentální retardací. Projevy PWS jsou způsobené poruchou funkce hypotalamu a liší se v závislosti na věku pacienta. PWS patří mezi patologie způsobené dysregulací imprintingu. Výskyt je sporadický. Prevalence je stejná u dívek i u chlapců.

65 Patnáctiletý chlapec s PWS Obezita, malý vzrůst, krátké ruce a nohy

66 Patnáctiletý chlapec, 41 letá žena s typickými obličejovými rysy při Praderově-Williho syndromu, protáhlý obličej, tenký horní ret, prominující nos.

67 Seznam imprintovaných transkripčních jednotek u člověka a myši
68
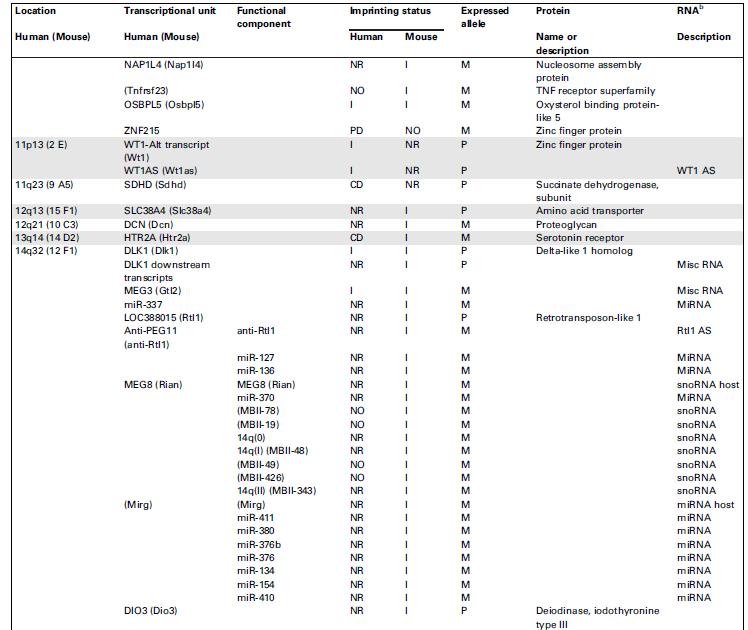
69

70

71
72
73 Hypotézy evolučního původu imprintingu Hypotéza genetického konfliktu Z důvodu většího počtu otců je potomstvo stejně příbuzné k matce, ale méně příbuzné k sobě navzájem. Matčin genetický zájem je sloužit všem stejně prostřednictvím distribuce zdrojů. Matka si může zachovat kontrolu inaktivací fetálních genů zesilujících růst. Otcův fitness (počet potomků) je však posílen, když jeho potomci dostanou co nejvíce zdrojů, třeba i na úkor polosourozenců a matky. Tomuto cíli slouží inaktivace fetálních růstových inhibitorů.
74 Hypotézy evolučního původu imprintingu Hypotéza vaječníkové časované bomby Spontánní vývoj neoplodněného vajíčka na vaječníku je formou ovariální trofoblastické nemoci, vlastně rakoviny. Inaktivace pouze maternální kopie časně účinkujícího růstového zesilovače takové riziko snižuje, stejně jako zvýšení aktivity jakéhokoliv inhibitoru růstu. Tato druhá změna může znamenat ztrátu plodu s příliš velkým počtem inhibitorů. Tato disbalance může být korigována snížením aktivity paternální kopie. Tzn. že předchozí i tato hypotéza předpokládají, že geny podílející se na růstu plodu jsou pravděpodobným cílem imprintingu. Zesilovače růstu mohou být zeslabovány maternálně, inhibitory růstu paternálně.
75 Hypotézy evolučního původu imprintingu Hypotéza pohlavně specifické selekce vázané na X chromozóm V časném vývoji inaktivují samice placentárních savců náhodně většinu jednoho X chromozómů. Samci mají jeden aktivní X chromozóm. To znamená, že změny v úrovni exprese genů na paternálním X budou postihovat pouze samice a změny na maternálním X budou postihovat samce více, než samice. Proto každý selekční tlak, který se liší mezi pohlavími, bude imprintingem zesilován. Zejména selekce na větší samce, obvyklá u savců, může být zesílena inaktivací maternálních inhibitorů růstu vázaných na X chromozóm a paternálních zesilovačů růstu rovněž vázaných na X.
76 Hypotézy evolučního původu imprintingu Hypotéza pohlavně antagonistické selekce Je rozšířením předchozí hypotézy na autozomální lokusy. Říká, že lokusy s různou úrovní optimální exprese u samců a samic mají pravděpodobnost být imprintovány. Imprinting bude preferován, pokud přínos pro potomky jednoho pohlaví převáží postižení u druhého. Navíc tato hypotéza říká, že lokusy mohou být imprintovány specificky podle pohlaví, např. mohou být maternálně inaktivovány u synů, ale ne u dcer.
77 77
78 Pozitivní účinek genomového imprintingu Mezi nejvýznamnější pozitivní účinek imprintingu patří dědičnost genu svalové hypertrofie u jehňat plemene Dorset. Tento gen se projevuje jen u heterozygotů a to pouze v tom případě, že jedinec zdědil Callipyge alelu od otce. To znamená, že dominantní alela zděděná od otce je aktivní, ale zděděná od matky je neaktivní.
79 Inaktivace X chromozomu Nejlepším příkladem imprintingu rozsáhlé oblasti genů je inaktivace chromozomu X homogametického pohlaví u savců. Důvodem inaktivace X chromozomu je kompenzace dávky genů lokalizovaných na X chromozomu. Samičí buňky obsahují dva pohlavní chromozomy XX, samčí XY. Shoda X a Y chromozomu je jen v krátké pseudoautozomální oblasti. Chromozom Y je zodpovědný za determinaci samčích pohlavních orgánů, zatímco na chromozomu X se nachází celá řada genů důležitých pro existenci organismu.
80 Inaktivace X chromozomu Inaktivace X chromozómu u samic placentárních savců je řízena Xist, jednou z nejdříve a nejlépe popsaných dlouhých nekódujících RNA (long ncrnas). Exprese Xist z později inaktivního X chromozómu a jeho následná inaktivace se děje během rané diferenciace kmenových buněk. Xist expression is followed by irreversible layers of chromatin modifications that include the loss of the histone (H3K9) acetylation and H3K4 methylation that are associated with active chromatin, and the induction of repressive chromatin modifications including H4 hypoacetylation, H3K27 trimethylation (Wutz 2007), H3K9 hypermethylation and H4K20 monomethylation as well as H2AK119 monoubiquitylation. These modifications coincide with the transcriptional silencing of the X-linked genes (Morey 2004). Xist RNA also localises the histone variant macroh2a to the inactive X chromosome (Costanzi 1998). There are additional ncrnas that are also present at the Xist loci, including an antisense transcript Tsix, which is expressed from the future active chromosome and able to repress Xist expression by the generation of endogenous sirna (Ogawa 2008). Together these ncrnas ensure that only one X-chromosome is active in female mammals.
81 Umlčení X chromozomu Inaktivace chromozomu X je náhodný jev, inaktivován je maternální nebo paternální chromozom. V dalších buněčných děleních se původně náhodná inaktivace zachovává. V buňkách je vždy aktivní jen jeden X chromozom. U homogametického pohlaví může dojít v určitých tkáních a orgánech k vzniku buněčného mosaicismu, heterozygotní genototyp X P X M. Inaktivovaný chromozom X je viditelný jako heterogametický komponent během interfáze v jádře buněk. Podle svého objevitele byl pojmenován jako Barrovo tělísko (sex chromatin, hetetochromatin, X chromatin ).
82 Proces inaktivace X chromozomu Umlčení X chromozomu nastává vlivem změny konformace chromatinu. Proces inaktivace je řízen genem XIST (X inactive specific transcript) v X inaktivačním centru (XIC) na chromozomu X. Inaktivované geny mají v oblasti promotoru lokalizovaný 5 metylcytosin a též u nich dochází k modifikaci histonů. Gen XIST kóduje nukleární RNA, která je považována za jeden z komponentů zabezpečujících inaktivaci chromozomu X. Inaktivovaný chromozom je charakteristický nízkou acetylací histonů H4 a silnou metylací CpG párů v promotorových oblastech genů.
83 Dlouhá nekódující RNA Long noncoding RNA lncrna
84 lncrna GWAS u hosp. zvířat založené na vysoce citlivém genotypování a sekvenování odhalily, že většina signálů, asociovaných s fenotypem komplexních vlastností, je v genomu lokalizována mimo známé oblasti kódující proteiny. Existuje celogenomově rozšířená transkripční aktivita. Genomy jsou templátem pro tisíce dlouhých nekódujících transkriptů lncrnas.
85 lncrna I když lncrnas nemají kódující funkci, u mnoha byla zjištěna funkční role v řadě biologických procesů. Tzn., že byla detekována nová regulační síť, součást komplexní strukturální organizace a funkce genomu. U hosp. zvířat je zatím komplexita transkriptomu málo popsána, např. ve srovnání s člověkem nebo myší.
86 lncrna trochu detailněji: Nekódující genomové varianty lokalizované v nepřekládaných a intronových obl. genů kódujících proteiny nebo v intergenových obl. mohou mít funkci v expresi specifické vlastnosti nebo fenotypu. To je možné, pokud jsou v regulační oblasti genomu. V poslední době se velmi zvyšuje počet nekódujících RNA (ncrna) transkriptů, odhalených NGS a dalšími novými technikami. Až 90% savčího genomu je transkribováno, v záv. na typu buňky, tkáně a vývojovém a fysiologickém stadiu.
87 Rozdělení nekódující RNA (ncrna): 1/ klasické nc typy, tj. trna, rrna; 2/ různé malé typy, tj. mirna, snrna, snorna, sirna, pirna; 3/ dlouhé nekódující RNA (lncrna) s délkou nad 200 nt. Třída pseudogenů napodobujících strukturní geny, které ale ztratily kódující funkci v důsledku nahromadění mutací se rovněž řadí do lncrna. lnccrnas mají jen slabý nebo žádný kódující potenciál, nízkou úroveň konzervace mezi druhy a nedostatek výrazných společných sekvenčních rysů nebo motivů. To brání jejich kategorizaci a funkční předpovědi.
88 Nicméně, mnohé lncrna sdílejí některé rysy s mrna kódující protein. Mohou být multiexonické, mohou být na 3 konci polyadenylovány a na 5 konci mohou mít čepičku. Mnohé obsahují v promotoru CpG ostrovy a mohou vykazovat komplex alternativních modelů sestřihu. Většina lncrnas má jasné příznaky transkripce RNA polymerázou II, s promotory značenými specifickými histonovými signály. Jsou diferenciálně exprimované v závislosti na typu buňky a tkáně, vývojovém a fyziologickém stadiu nebo stadiu choroby. Regulují transkripci způsobem specifickým pro konkrétní alelu nebo lokus. Úroveň exprese je významně nižší, než u strukturních genů.
89 Každá tkáň má svůj vlastní katalog specifických lncrna, které mohou přispívat k unikátní funkci dotyčné tkáně. Jsou účastny na regulaci řady normálních a patologických procesů.
90 Genomová lokalizace je jedním z kritérií, jak lncrnas rozlišit. Mohou být umístěny: 1/ intergenové genomické oblasti nepřekrývající se se známými lokusy kódujícími proteiny, např. long intergenic noncoding RNA, lincrna; 2/ intronové oblasti lokusů kódujících proteiny; 3/ protější vlákno vztažené k příslušnému transkriptu (antisense RNA); 4/ sousedství lokusu kódujícího protein UTR associated nebo promotor associated transkripty, označované jako pasrna ; 5/ extragenové enhancerové oblasti, erna.
91 lncrnas se účastní v široké škále biologických procesů. Působí jako velmi rozšířená součást sítě genové regulace. Vadná transkripce lncrna nebo mutace a dysregulace souvisí dle výzkumů s patogenezí různých chorob: růst nádorů; mentální a neurodegenerativní choroby; obezita a kardiovaskulární patologie. Dále souvisí s náchylností k infekci, s vývojovými a diferenciačními procesy.
92 lncrnas se účastní jako významná funkční součást v hlavních regulačních mechanismech genové exprese na úrovni: epigenetické regulace (modifikace chromatinu); modulace alelové exprese (genomový imprinting, inaktivace X chromozómu); transkripční a posttranskripční regulace genové exprese.
93 lncrnas působí prostřednictvím zacílení na urč. obl. genomu tím, že slouží jako funkční cis nebo trans regulační elementy, jako jsou promotory, zesilovače nebo insulátory a jako anti-sense interferenční elementy, často způsobem, který je specifický dle buňky, tkáně, vývojového stadia.
94 Insulátor je genetický vazebný element blokující interakci mezi enhancery a promotory. Předpokládá se, že insulátor musí být umístěn mezi enhancerem a promotorem, aby mohl inhibovat jejich pozdější interakce. tzn. že determinuje sadu genů, které může enhancer ovlivňovat. Potřeba insulátorů vzrůstá tam, kde sousední geny na chromozómu mají velmi odlišné vzory transkripce. Kritické je to, že indukční nebo represívní mechanismy jednoho genu neinterferují se sousedícím genem.
95 Insulátory klastrují na hranicích topologických asociačních domén (TADs) a možná mají roli v dělení genomu na chromosomální čtvrtě, tj. genomické oblasti, v nichž se realizuje regulace. Aktivita insulátorů zřejmě probíhá primárně prostřednictvím regulace 3D struktury DNA, je zprostředkovávaná proteiny vč. CTCF.
96 Mechanismus působení insulátorů Pravděpodobně mnoha mechanismy. Mnohé enhancery vytváří DNA loops, které jim umožňují těsnou fyzickou blízkost k promotorovým oblastem během aktivace transkripce. Insulátory možná podporují utváření DNA loops, které brání vzniku loops mezi promotorem a enhancerem. Bariérové insulátory zžejmě brání rozšíření heterochromatinu z umlčeného (silenced) genu ke genu, který je aktivně transkribován.
97 Dále, lncrnas mohou působit jako regulační lešení; průvodcové; návnady nebo signály. Mohou sloužit jako hostitelé mirnas a snornas.

98 Transkripční regulace navázání na transkripční faktor, navázání smyčky, vytvoření smyčky blokující transkripci transkripční koaktivace, koreprese Způsoby funkční regulace lncrna. Transkripční regulace: navázáním na transkripční faktor brání lncrna jeho napojení na cílový promotor a tím regulaci transkripce a genové exprese.
99 Post-transkripční regulace stabilita mrna, její rozklad napojení mirna, působí jako návnada alternativní sestřih aktivace a inhibice translace Post-transkripční regulace: soutěžení lncrna o mikrorna (navázání lncrna a mirna) brání tomu, aby se mirna vázala na mrna (vlevo). Nepřímá interakce s mrna (vpravo) reguluje (blokuje) translaci cílové mrna.

100 Epigenetická regulace chromatin, histonový ocas nebo modifikace DNA lncrna působí jako lovci, řetězy, průvodci, lešení Epigenetická regulace. lncrna interaguje s komplexy modifikujícími chromatin. Tím může působit jako lovec, řetěz, průvodce, lešení a může regulovat, reprimovat nebo zesilovat, genovou expresi.
101 Dosud byla většina lncrnas popsána ve velmi dobře prozkoumaných lidských a myších genomech. Většina lncrnas ještě musí být popsána, jejich biologická funkce vyžaduje upřesnění a potvrzení. Nejznámější a nejlépe popsané jsou XIST, HOTAIR, H19, KCNQ10T1, AIR. Genetická variabilita v nekódujících genomických oblastech vč. lncrnas je spojena s expresními rozdíly v různých lidských tkáních!
102 Pro identifikaci lncrnas je základní a hlavní problém rozlišení mezi protein kódujícími mrna, dlouhými nekódujícími transkripty a nespecifickým transkripčním smetím. Častým kritériem je délka ORF. Další strategií je hledat in silico translatované peptidové sekvence v databázích proteinů nebo proteinových domén. Byly vyvinuty i další bioinformatické postupy.
103 U skotu je prvním popsaným příkladem dlouhá intergenová RNA (lincrna) ovlivňující ontogenesi rohů skotu. U prasat např. ovlivňují lncrnas vývoj svalů, preimplantační procesy, graviditu, komunikaci endometrium-embryo. Dále buněčnou adhesi, signalizaci, aktivitu růstových faktorů, imunní systém, vývoj varlat, spermatogenesi.
104 U drůbeže např. ovlivňují odolnost proti Markově chorobě (Marek s disease virus, MDV). lncrna MHN má roli v růstu embryí a vývoji gonád. lncrna-agt má roli v přepínání exprese z embryonálního do adultního alpha D-globinového genu.
105 Dosud (podzim 2016) nejsou známé studie o lncrnas u dalších druhů, tj. koní, králíka, kachny, husy, krocana, křepelky.
106 Většina popsaných lncrnas se účastní biologických procesů, regulujících vývoj, diferenciaci, nebo jsou spojeny s buněčnou proliferací a smrtí. Díky poziční konzervaci lncrnas v genomu u různých obratlovců je možné, že regulace základních biologických procesů a systémů modulovaných lncrnas, tj. růst a vývoj, metabolismus, infekční odpověď, je rovněž sdílená (podobná) mezi různými druhy. lncrnas se účastní na regulaci a jemném vyladění metabolických a imunitních procesů.
107 Tzn., že podobná genová regulace základních fyziologických, endokrinních a imunologických procesů pomocí lncrnas zřejmě naznačuje mezidruhovou vazbu u příslušných fenotypových vlastností důležitých u hosp. zvířat.
108 lncrnas jsou spojeny s jadernými receptory, účastní se na vývoji mléčné žlázy, tukové tkáně, diferenciaci kosterní svaloviny, homeostázi glukózy, vzniku metabolických poruch. Existuje několik drah lncrnas, řídících vývoj a funkci metabolických tkání, např. lipidový metabolismus v játrech, kosterní svalovině a tukové tkáni.
109 Podílí se na lipidové homeostázi, na myogenezi a regeneraci svalů. U lidí je lncrna NBR2 indukována při energetickém stresu (glukózové deprivaci).
110 Jiné lncrnas vykazují vazbu mezi genovou expresí a imunitou. Je to nová třída molekul pro signalizaci vrozené a adaptivní imunity, regulátorů závislých na virech, resp. dokonce nová úroveň regulace genové exprese, která je odpovědná za utváření imunitní odpovědi během virové infekce a zánětu. lncrnas možná hrají kritickou roli v interakci s chromatinem a komplexy modifikujícími chromatin.
111 U hosp. zvířat jsme stále na počátku! Umožní nový pohled na metabolickou regulaci a také na fenotypovou varianci metabolických poruch, které nyní nelze vysvětlit strukturální nebo expresní variabilitou protein-kódujících transkriptů. Jsou možná regulačními molekulami, které představují molekulárně genetickou bázi identifikovaných QTL pro fenotypové vlastnosti a tedy přispívají k fenotypové varianci těchto vlastností prostřednictvím genetických a epigenetických mechanismů. V budoucnu zahrnutí do selekce prostřednictvím genomové selekce.
112 Děkuji za pozornost
EPIGENETICKÁ DĚDIČNOST
 Epigenetika EPIGENETICKÁ DĚDIČNOST Pojmenovaná britským biologem Conradem Waddingtonem r. 1940, Něco navíc, mimo v porovnání s mendelistickou genetikou, Umožňuje buňkám s identickým genotypem vznik odlišných
Epigenetika EPIGENETICKÁ DĚDIČNOST Pojmenovaná britským biologem Conradem Waddingtonem r. 1940, Něco navíc, mimo v porovnání s mendelistickou genetikou, Umožňuje buňkám s identickým genotypem vznik odlišných
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Dědičnost a pohlaví. KBI/GENE Mgr. Zbyněk Houdek
 Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Atestace z lékařské genetiky inovované otázky pro rok A) Molekulární genetika
 Molekulární genetika") Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
Deoxyribonukleová kyselina (DNA)
") Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Základy molekulární a buněčné biologie. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
RNA interference (RNAi)
") Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
Liběchov, 29. 11. 2013 RNA interference (RNAi) post-transkripční umlčení genové exprese přirozený mechanismus regulace genové exprese a genomové stability obranný antivirový mechanismus konzervovaný mechanismus
Genetika zvířat - MENDELU
 Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Genetika pohlaví genetická determinace pohlaví
 Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
GENETIKA. Dědičnost a pohlaví
 GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
MECHANIZMY EPIGENETICKÝCH PROCESŮ
 MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
DUM č. 10 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 10 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 26.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Procesy následující bezprostředně po transkripci.
projekt GML Brno Docens DUM č. 10 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 26.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Procesy následující bezprostředně po transkripci.
Molekulární mechanismy formování epigenomu
 lncrna a epigenom Molekulární mechanismy formování epigenomu Epigenetika = věda o stabilních genetických modifikacích, které vedou ke změně exprese a funkce genů beze změny sekvence DNA Epigenetické procesy
lncrna a epigenom Molekulární mechanismy formování epigenomu Epigenetika = věda o stabilních genetických modifikacích, které vedou ke změně exprese a funkce genů beze změny sekvence DNA Epigenetické procesy
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
Na rozdíl od genomiky se funkční genomika zaměřuje na dynamické procesy, jako je transkripce, translace, interakce protein - protein.
 FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
Glosář - Cestina. Odchylka počtu chromozomů v jádře buňky od normy. Např. 45 nebo 47 chromozomů místo obvyklých 46. Příkladem je trizomie 21
 Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
RNA molekuly. Analýza genové exprese pomocí cytometrických (a jiných) metod. Analýza exprese a funkce microrna. Úrovně regulace genové exprese
 metod. Analýza exprese a funkce microrna. Úrovně regulace genové exprese") Analýza genové exprese pomocí cytometrických (a jiných) metod Studium exprese a funkce microrna Eva Slabáková, Ph.D. Bi9393 Analytická cytometrie 12.11.2013 Oddělení cytokinetiky Biofyzikální ústav AVČR,
Analýza genové exprese pomocí cytometrických (a jiných) metod Studium exprese a funkce microrna Eva Slabáková, Ph.D. Bi9393 Analytická cytometrie 12.11.2013 Oddělení cytokinetiky Biofyzikální ústav AVČR,
Malé RNA a regulace genomu
 Malé RNA a regulace genomu Změna paradigmatu It appears that the genetic programming of humans and other complex organisms has been misunderstood for the past 50 years, due to the assumption that most
Malé RNA a regulace genomu Změna paradigmatu It appears that the genetic programming of humans and other complex organisms has been misunderstood for the past 50 years, due to the assumption that most
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
1. Definice a historie oboru molekulární medicína. 3. Základní laboratorní techniky v molekulární medicíně
 Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Dědičnost vázaná na X chromosom
 12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
The cell biology of rabies virus: using stealth to reach the brain
 The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
6. Nukleové kyseliny
 6. ukleové kyseliny ukleové kyseliny jsou spolu s proteiny základní a nezbytnou složkou živé hmoty. lavní jejich funkce je uchování genetické informace a její přenos do dceřinné buňky. ukleové kyseliny
6. ukleové kyseliny ukleové kyseliny jsou spolu s proteiny základní a nezbytnou složkou živé hmoty. lavní jejich funkce je uchování genetické informace a její přenos do dceřinné buňky. ukleové kyseliny
Základní pravidla dědičnosti
 Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Syntéza a postranskripční úpravy RNA
 Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Protinádorová imunita. Jiří Jelínek
 Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Cvičeníč. 10 Dědičnost a pohlaví. Mgr. Zbyněk Houdek
 Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc.
 Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Chromatin. Struktura a modifikace chromatinu. Chromatinové domény
 Chromatin Struktura a modifikace chromatinu Chromatinové domény 2 DNA konsensus 5 3 3 DNA DNA 4 RNA 5 ss RNA tvoří sekundární strukturu s ds vlásenkami ds forms 6 of nucleic acids Forma točivost bp/turn
Chromatin Struktura a modifikace chromatinu Chromatinové domény 2 DNA konsensus 5 3 3 DNA DNA 4 RNA 5 ss RNA tvoří sekundární strukturu s ds vlásenkami ds forms 6 of nucleic acids Forma točivost bp/turn
Syndrom fragilního X chromosomu (syndrom Martinův-Bellové) Antonín Bahelka, Tereza Bartošková, Josef Zemek, Patrik Gogol
 Antonín Bahelka, Tereza Bartošková, Josef Zemek, Patrik Gogol") Syndrom fragilního X chromosomu (syndrom Martinův-Bellové) Antonín Bahelka, Tereza Bartošková, Josef Zemek, Patrik Gogol 20.5.2015 Popis klinických příznaků, možnosti léčby Muži: střední až těžká mentální
Syndrom fragilního X chromosomu (syndrom Martinův-Bellové) Antonín Bahelka, Tereza Bartošková, Josef Zemek, Patrik Gogol 20.5.2015 Popis klinických příznaků, možnosti léčby Muži: střední až těžká mentální
NUKLEOVÉ KYSELINY. Složení nukleových kyselin. Typy nukleových kyselin:
 NUKLEOVÉ KYSELINY Deoxyribonukleová kyselina (DNA, odvozeno z anglického názvu deoxyribonucleic acid) Ribonukleová kyselina (RNA, odvozeno z anglického názvu ribonucleic acid) Definice a zařazení: Nukleové
NUKLEOVÉ KYSELINY Deoxyribonukleová kyselina (DNA, odvozeno z anglického názvu deoxyribonucleic acid) Ribonukleová kyselina (RNA, odvozeno z anglického názvu ribonucleic acid) Definice a zařazení: Nukleové
Epigenetické regulace. (Viz také speciální přednáška P. Svobody - Epigenetika)
") Epigenetické regulace (Viz také speciální přednáška P. Svobody - Epigenetika) Epigenetická dědičnost Epigenetická regulace genové exprese Definice: Heritabilní změna genové exprese beze změn DNA sekvence.
Epigenetické regulace (Viz také speciální přednáška P. Svobody - Epigenetika) Epigenetická dědičnost Epigenetická regulace genové exprese Definice: Heritabilní změna genové exprese beze změn DNA sekvence.
Potřebné genetické testy pro výzkum a jejich dostupnost, spolupráce s neurology Taťána Maříková. Parent projekt. Praha 19.2.2009
 Potřebné genetické testy pro výzkum a jejich dostupnost, spolupráce s neurology Taťána Maříková Parent projekt Praha 19.2.2009 Diagnostika MD její vývoj 1981-1986: zdokonalování diferenciální diagnostiky
Potřebné genetické testy pro výzkum a jejich dostupnost, spolupráce s neurology Taťána Maříková Parent projekt Praha 19.2.2009 Diagnostika MD její vývoj 1981-1986: zdokonalování diferenciální diagnostiky
GENETIKA. zkoumá dědičnost a proměnlivost organismů
 GENETIKA zkoumá dědičnost a proměnlivost organismů Dědičnost: schopnost organismů uchovávat informace o své struktuře a funkčních schopnostech a předávat je svým potomkům Proměnlivost (variabilita) je
GENETIKA zkoumá dědičnost a proměnlivost organismů Dědičnost: schopnost organismů uchovávat informace o své struktuře a funkčních schopnostech a předávat je svým potomkům Proměnlivost (variabilita) je
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Základy genetiky - Alelové a Genové interakce (Spolu)Působení genů Fenotypový
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Základy genetiky - Alelové a Genové interakce (Spolu)Působení genů Fenotypový
Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií
 poznatků genetiky a evolucionistických teorií") Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
Význam genetického vyšetření u pacientů s mentální retardací
 Význam genetického vyšetření u pacientů s mentální retardací Šantavá, A., Hyjánek, J., Čapková, P., Adamová, K., Vrtěl, R. Ústav lékařské genetiky a fetální medicíny FN a LF UP Olomouc Mentální retardace
Význam genetického vyšetření u pacientů s mentální retardací Šantavá, A., Hyjánek, J., Čapková, P., Adamová, K., Vrtěl, R. Ústav lékařské genetiky a fetální medicíny FN a LF UP Olomouc Mentální retardace
Molekulární procesy po fertilizacinormální či abnormální po ART?
 Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
Člověk a mikroby, jsme nyní odolnější? Jan Krejsek. Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové
 Člověk a mikroby, jsme nyní odolnější? Jan Krejsek Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové Jsme určeni genetickou dispozicí a životními podmínkami, které působí epigeneticky
Člověk a mikroby, jsme nyní odolnější? Jan Krejsek Ústav klinické imunologie a alergologie, FN a LF UK v Hradci Králové Jsme určeni genetickou dispozicí a životními podmínkami, které působí epigeneticky