Molekulární genetika zvířat 5 Antonín Stratil. Česká zemědělská univerzita v Praze
|
|
|
- Leoš Svoboda
- před 6 lety
- Počet zobrazení:
Transkript
1 Molekulární genetika zvířat 5 Antonín Stratil Česká zemědělská univerzita v Praze
2 Osnova Kandidátní geny Příčinné geny Gen RYR1 prasete Gen IGF2 prasete Gen PRKAG3 prasete Gen MSTN skotu Defekt adherence leukocytů Komplexní vertebrální malformace Lokus CLPG u ovcí Genetické aspekty produkce masa Genetické aspekty produkce mléka Gen DGAT1 skotu Efekty genotypů mléčných proteinů Gen NHEJ1 u psů Genomický imprinting Inaktivace X chromozómu Zjišťování pohlaví u zvířat Geny pro reprodukci (BMPR1B; BMP15; QTL) Priony. BSE Imunogenetika Imunita - Imunoglobuliny Domény imunoglobulinových řetězců Komplexní geny - Hlavní histokompatibilitní komplex - Krevněskupinové antigeny
3 Mapování QTL Celogenomová analýza Není potřebná znalost funkce Testování velkého počtu márkrů Stanovit vysoké prahy statistické významnosti Neidentifikuje přímo kandidátní nebo příčinný gen
4 Kandidátní geny Výhody: Není potřeba testovat příliš mnoho márkrů (testuje se jeden gen) Identifikace příčinné alely umožňuje přímé a jednoduché využití v selekci
5 Kandidátní geny Nevýhody: Vyžaduje dobrou znalost biologie Mnoho genů je neznámých Asociace mezi určitou alelou a užitkovým znakem nedokazuje příčinný vztah
6
7
8 Příklady kandidátních genů u prasete Gen Název Asociace ESR1 Estrogenový receptor 1 Reprodukce PIT1 Hypofyzární transkripční faktor Růst a reprodukce MYOG Myogenní transkripční faktor Myogenesa; produkce masa MYOD1 Myogenní transkripční faktor Myogenesa; produkce masa MYF5 Myogenní transkripční faktor Myogenesa; produkce masa MYF6 Myogenní transkripční faktor Myogenesa; produkce masa LEP Leptin Žravost, obezita, reprodukce LEPR Receptor pro leptin Reprodukce FABP3 Protein vázající mastné kyseliny, Intramuskulární tuk srdeční forma A-FABP Adipocytární protein vázající Intramuskulární tuk mastné kyseliny GH1 Růstový hormon Růst a masná užitkovost SPP1 Osteopontin Reprodukce
9 Kandidátní geny asociované se znaky reprodukce u prasat
10 Asociační studie ESR1 (estrogen receptor 1) (Rothschild et al., 1996) Prasnice s 50% podílem Meishana: první vrhy - navíc 2,3 selete všechny vrhy - navíc 1,5 selete pozitivní efekt alely B (homozygoti) metoda: RFLP - PvuII Prasnice Large White homozygoti BB - 1 sele navíc V dalších pracích (např. Short et al., 1997) efekt alely B byl 0,3 1,4 selete (na kopii alely B).
11 V jiných pracích efekt alely B nebyl, nebo byla výhodnější alela A (např. Goliášová a Wolf, 2004) První vrhy AA prasnice o 0,5 selete více než prasnice BB Všechny vrhy AA prasnice o 0,25 selete více než prasnice BB nejedná se zřejmě o příčinnou mutaci, ale o genetickou vazbu, mezi ESR a příčinným genem není těsná vazba, vyskytující se rekombinace mohou ovlivnit výsledek asociace, úloha genetického pozadí na expresi?
12 Příklady příčinných genů Druh Znak Gen Prase Skot Ovce Kůň Kur Maligní hyperthermie Zmasilost Obsah glykogenu ve svalech Vnímavost na infekci E. coli Hypercholesterolemie Biosyntéza vitaminu C Dominantní bílé zbarvení Zbarvení Svalová hypertrofie Křehkost masa Defekt adherence leukocytů (BLAD) Komplexní vertebrální malformace (CVM) Deficience uridinmonofosfátsyntázy (DUMPS) Deficience kyselé alfa-glukozidázy (Pompeho choroba) Zápach mléka Zbarvení srsti Svalová hypertrofie (callipyge) Plodnost, ovulace (booroola) Plodnost, ovulace (inverdale) Hyperkalemická periodická paralýza (HYPP) Těžká kombinovaná imunodeficience (SCID) Zbarvení Bílé zbarvení megacolon Albinismus Zbarvení peří Dominantní bílé zbarvení RYR1 (CRC) IGF2 PRKAG3 FUT1 LDLR GULO KIT MC1R MSTN CAPN1 CD18 SLC35A3 UMPS AAG FMO3 MC1R CLPG BMPR1B BMP15 SCN4A DNA-PK MC1R ASIP MATP EDNRB TYR MC1R PMEL17
13
14 Gen RYR1 (HAL; CRC1) prasete (kanál uvolňující vápník v sarkoplazmatickém retikulu - CRC) SSC6; bodová mutace záměna jedné báze (1843C>T) autozomální recesivní dědičnost citlivosti k halotanu 1966 objev maligní hyperthermie u prasat (Hall et al.) citlivost k halothanu asociace Hal s kvalitou masa a PSS dědičnost citlivosti k halothanu průkaz genetické vazby lokusu HAL s geny PHI, EAH, EAS, PGD a A1BG haplotypování stanovení genotypu v HAL lokusu pomocí vázaných genů lokalizace na chromozómu 6 prasete průkaz homologie mezi segmentem chromozómu 19 člověka, kde je lokalizovaný gen RYR1, a segmentem chromozómu 6 prasete rozdíly v peptidových mapách proteinu RYR1 mezi prasaty náchylných k maligní hyperthermii a normálními 1991 identifikace mutace (C T) v genu RYR1 prasete, v poloze 1843 (CC homozygot NN; TT homozygot nn = citlivost ke stresu) záměna aminokyseliny (argininu cysteinem) (Fujii et al.) Testování: PCR-RFLP (134 bp fragment se štěpí HhaI: alela C (N) bp; alela T (n) 134 bp Pleiotropní efekt na komplexní znaky zvířata nn mají vyšší podíl libového masa
15 Vliv genotypů v genu RYR1 (HAL) na znaky masné užitkovosti a kvality masa u prasat
16

17 Gen IGF2 prasete (Inzulin-like growth factor 2) Paternálně exprimovaný QTL na p raménku chromozómu 2; 15 30% fenotypové variability podílu masitých částí a 10 20% fenotypové variability výšky hřbetního tuku. Příčina: mutace v intronu 3 genu IGF2 (3072G>A); regulační oblast)
18 Genetická a fyzická mapa oblasti SSC15 vedoucí k identifikaci genu PRKAG3 (ovlivňuje obsah svalového glykogenu; autozomální dominantní dědičnost vysokého obsahu; vysoký obsah zhoršená výtěžnost a kvalita masa) (PRKAG3 - gama podjednotka AMP-aktivované proteinkinázy) (1. část) (A) (B) Vazbová mapa se vzdálenostmi v centimorganech (cm) Radiační hybridní mapa se vzdálenostmi v centirayích je ukázáno pořadí orientačních markerů a je naznačena poloha dalších lokusů. (C) Vybrané klony z 2,5 Mbp BAC kontigu na prasečím chromozómu 15 ukazující lokalizaci genů a genetických markerů.
19 Genetická a fyzická mapa oblasti chromozómu SSC15 vedoucí k identifikaci genu PRKAG3 (2. část) Vazbová nerovnováha mezi RN (vysoký obsah glykogenu) a alelami markerů zjišťovaná pomocí programu DISMULT na náhodném vzorku 91 prasat plemene švédský hampshire
20 Porovnání sekvencí v různých genotypech PRKAG3 Dominantní mutace RN - = vysoký obsah svalového glykogenu (kodon 200R>Q; báze G>A)
21 Gen MSTN u skotu Myostatin; growth and differentiation factor 8; GDF8 (negativní regulátor růstu svalů) delece 11 bp v exonu chromozóm 2; autozomální recesivní dědičnost dvojitého osvalení heterozygoti mají zvětšený m. longissimus dorsi a snížené množství podkožního a intramuskulárního tuku, a porody probíhají normálně 1997 identifikována delece 11 bp v exonu 3 u plemene belgické modrobílé důsledek: posun fáze translace, předčasné ukončení a eliminace celé aktivní oblasti molekuly proteinu důsledek pro fenotyp: výskyt dvojitého osvalení vyšší masná produkce (o 20%) (hyperplazie i hypertrofie) problémy: obtížné porody (dystocie; většinou císařským řezem), snížená plodnost, vyšší úhyn telat, zvýšená citlivost ke stresu
22 Mutace v genu MSTN (myostatin) skotu způsobující dvojité osvalení Mutace v MSTN genu způsobující dvojité osvalení byly nalezeny u následujících plemen skotu: Belgian Blue (belgické modrobílé), Limousine, Parthenaise, Asturiana, Rubea Gallega, Gasconne, Piedmontese, Maine-Anjou, Charolais, Limousine a Marchigiana. Blonde d Aquitane typicky nemají mutace v MSTN neidentifikované faktory pro hypertrofii?

23 Plemeno belgické modrobílé
24 Bovine leukocyte adhesion deficiency (BLAD) (porucha adhese leukocytů skotu) u plemene holštýn Gen: ITGB2 (CD18) (chromozóm 1) produkt: beta-2 subjednotka integrinu Příčinná mutace: 383A>G (v proteinu 128D>G) BLAD je autozomální recesivní genetická choroba. Neutrofily ztratily schopnost přilnout k endotelu a prostupovat do tkáně. Projev onemocnění: Opakované bakteriální infekce, špatné hojení ran a mnoho dalších klinických příznaků (zápal plic, průjmová onemocnění, zakrslý vzrůst, zvýšený počet neutrofilů aj.) Telata obyčejně hynou během 2-4 měsíců po narození. Heterozygoti jsou klinicky normální; postiženi jsou recesivní homozygoti. BLAD byl identifikován v 80. letech. Příčinná mutace byla odhalena v roce 1992 (Schuster et al.). Původ defektu sahá k býkovi Osborndale Ivanhoe (otec holštýnského plemene), který byl široce využíván. Jedním z potomků je i Carlin-M Ivanhoe Bell, který je přenašečem defektní alely BLAD, a rovněž defektní alely pro CVM. (v USA byl výskyt nositelů mutace 15% u býků v AI a 8% u krav) Testování: PCR-RFLP Podobný defekt se vyskytuje i u člověka (LAD) a u psů (CLAD)

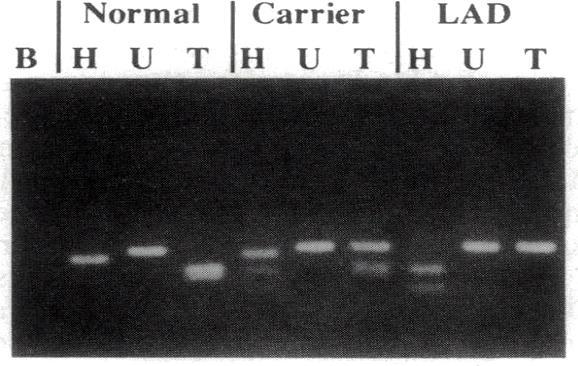
25 PCR-RFLP BLAD
26 Komplexní vertebrální malformace (CVM) holštýnského skotu Gen: SLC35A3 (chromozóm 3) (solute carrier family 35, member A3) produkt: UDP-N-acetylglukosamin transporter Substituce: G T (v proteinu V180F) Autozomální defekt s recesivní dědičností. Mnohočetné malformace u telat proporcionální dwarfismus, symetrická artrogryposa předních, někdy i zadních končetin, znetvoření obratlů krční a hrudní části páteře, případně i malformace srdce. Mutace (v homozygotním stavu) způsobuje prenatální úhyn telat nebo se telata rodí předčasně a často mrtvá. Zvířata s jednou defektní alelou jsou zcela normální, jsou však přenašeči. Postiženi jsou recesivní homozygoti. Býk Carlin-M Ivanhoe Bell přenašeč CVM i BLAD. DNA testy se provádějí v mnoha laboratořích. V ČR k mezi plemennými býky odhaleno 9,2% přenašečů.


27 Telata postižená CVM (komplexní vertebrální malformace)
28 Deficiency of uridine monophosphate synthase (DUMPS) u plemene holštýn Gen: UMPS (chromozóm 1) produkt: uridine monophosphate synthase) bodová mutace v kodonu 405 C T Autozomální recesivní dědičnost. Nepravidelná syntéza DNA a RNA. Úhyn plodu kolem 40. dne embryonálního vývoje. Test: PCR-RFLP; AvaI Citrullinemie Gen: ASS Produkt: argininosuccinate synthetase deficiency bodová mutace v exonu 5 Autozomální recesivní dědičnost. Nedostatek enzymu, který se uplatňuje při tvorbě močoviny. Projevuje se vyšší tvorbou čpavku, který vede k nepravidelnostem v centrálním nervovém systému novorozených telat (úhyn během 3 5 dnů po narození. Test: PCR-RFLP (AvaI)
29 Ceník služeb poskytovaných Laboratoří imunogenetiky Název služby Cena za jednotku v Kč 1. Stanovení genotypu krevních skupin skotu 800,00 2. Stanovení genotypu mikrosatelitů skotu 800,00 4. Stanovení genotypu mikrosatelitů koní 900,00 5. Stanovení genotypu mikrosatelitů prasat 600,00 6. Stanovení genotypu mikrosatelitů koz 650,00 7. Stanovení genotypu mikrosatelitů ovcí 650,00 8. Stanovení genotypu mikrosatelitů psů 800,00 9. Stanovení genotypu BLAD Stanovení genotypu RYR1 (MHS stres) 400, Stanovení genotypu CAS (kappa kasein) 400, Stanovení genotypu RED 900, Stanovení genotypu CVM 1500, Stanovení genotypu SCID 350, Stanovení genotypu dvojího osvalení 550, Ověření správnosti původu* - použití genotypu již testovaného zvířete skot krevní skupiny skot mikrosatelity koně mikrosatelity prasata mikrosatelity kozy a ovce mikrosatelity psi mikrosatelity 60,00 85,00 100,00 65,00 50,00 100, Výpis stanoveného genetického typu z databáze 500, Vydání mezinárodní karty genetického typu z databáze 150, Vystavení kopie protokolu 500, Fragmentová analýza z produktu PCR dodaného zákazníkem 400, Stanovení genotypu exotických zvířat 950,00
30 Lokus CLPG u ovcí (kontrolní element na velkou vzdálenost) (long-range control element LRCE záměna A - G 1983 (farmář v Oklahomě) beran s výrazně vyvinutým svalstvem (Solid Gold) dědičnost: maternální imprinting*, jmenovitě paternální polární superdominance (hypertrofie jen u heterozygotů, kteří zdědili mutovanou alelu od otce) maso má horší kvalitu je tuhé lokus CLPG (callipyge pěkný zadek) mapován na distální konec chromozómu identifikace mutace sekvenováním 210 kb oblasti mezi dvěma pozičními kandidátními geny DLK1 a GTL2 u inbredního berana, heterozygota v CLPG lokusu (Freking et al.) CLPG mutace je záměna jedné báze (A G) v předpokládaném LRCE (kontrolní element na velkou vzdálenost) * imprinting alely určitého genu od otce a matky se liší v expresi. Může to být způsobeno rozdíly v metylaci DNA.
31 Lokus CLPG u ovcí Fig. 2. The gene content in the vicinity of the HSA14q32 imprinted domain (from data in [21]). Bi-allelically expressed genes are represented in green, genes expressed from the paternal allele in red, and genes expressed from the maternal allele in blue. Genes whose imprinting status is unknown are shown in gray. Putative genes reported by [21] are shown in black. The genes are sorted according to the direction of transcription ( + versus strand). The approximate position of the CLPG mutation is marked by the yellow asterisk. Genes whose expression is known to be influenced by the CLPG mutation are marked by the orange arrows [25]. The effect of the CLPG mutation on the expression of the other genes in the region is being tested.
32 Gen DGAT1 u skotu (Acyl-CoA:diacylglycerolacyltransferáza 1) Enzym katalyzuje poslední stupeň v syntéze triglyceridů Gen je lokalizován na centromerickém konci bovinního chromozómu 14 - v DNA - záměna AA GC v exonu 8 - v proteinu - záměna lyzinu (K) alaninem (A) v pozici 232 Nositelé alely K (dojnice holštýnsko-fríského skotu, Nový Zéland) mají o 5,76 kg více tuku v mléce, o 2,45 kg méně proteinu a o 134 l méně mléka než nositelé alely A. 1998; zjištěn QTL pro obsah mléčného tuku v centromerické oblasti bovinního chromozómu ; interval zúžen na 3,8 cm identifikace mutace 232K>A a zjištěn rozdíl v efektu alel použitím expresního vektoru, do kterého byla ligována cdna DGAT1 a měřením enzymatické aktivity syntetizovaného rekombinantního DGAT1 byla prokázána vyšší aktivita u alely 232K

33 Gen DGAT1 skotu Mutace AA GC v exonu 8 (v proteinu je K232A)

34
. Projevuje se v lehčí a těžší formě. V těžší formě dojde až k úplnému oslepnutí.")
35 Gen NHEJ1 psa (nonhomologous end-joining factor 1) Mutace v tomto genu způsobuje chorobu anomálie oka kolií (Collie eye anomaly; CEA). Dědičné onemocnění nedostatečný vývoj cévnatky (vrstva v zadní části oka). Projevuje se v lehčí a těžší formě. V těžší formě dojde až k úplnému oslepnutí. CEA se dědí autozomálně recesívně nemoc se projeví u jedinců, kteří mají mutované obě alely (recesívní homozygoti). U heterozygotů (jedna alela normální, druhá mutovaná) se nemoc neprojeví. Příčinná mutace: Delece bp v NHEJ1 genu v intronu 5 (podle Ensembl); Tento intron má celkovou velikost ~67 kb. Delece je blíže exonu 6 (460 bází od exonu 6). Sekvence inzerce obsahuje vysoce konzervovanou doménu, na kterou se váže několik proteinů důležitých pro vývoj. Testování: PCR pomocí dvou párů primerů (dvoustupňová PCR): 1. pár primerů (F17 a R17) je navržený ze sekvence inzerce; tato sekvence u postižených (A) psů chybí, a fragment se neamplifikuje. 2. pár primerů (F20 a R23) je navržený ze sekvencí před delecí a za delecí. Amplifikuje fragment, který má deleci, a neamplifikuje fragment s inzercí (je příliš dlouhý na PCR amplifikaci).
36 An example of (comprehensive) survey of DNA polymorphisms and associations with production traits Ibeagha-Awemu et al., Mamm. Genome (2008) 19: (7 pages total)
 for CMD2 in cattle Celogenomové asociační mapování")
37 Association mapping (using SNPs) and identification of causative gene (SLC6A5) for CMD2 in cattle Celogenomové asociační mapování
38 Příčinné geny u ovcí BMPR1B - bone morphogenetic protein receptor 1B (gen je na chromozómu 6) Mutace v genu mění aminokyselinu (Gln249Arg). Důsledek vyšší počet folikulů a vyšší počet ovulovaných oocytů = vyšší počet jehňat ve vrhu u plemene Booroola (až 10 jehňat). (gen se rovněž označuje Booroola gen nebo FecB). (Mulsant et al., 2001) BMP15 - bone morphogenetic protein 15 (také FecX nebo Inverdale) je lokalizovaný na X chromozómu. Mutace zvyšuje počet ovulovaných oocytů o jeden u heterozygotních ovcí, ale u homozygotů s mutovanou alelou způsobuje sterilitu. (Galloway et al., 2000)
39 Genetické aspekty produkce masa - Prenatální myogeneze - Postnatální růst svalů (hypertrofie) - Inverzní vztah mezi počtem svalových vláken a jejich velikostí - Regulační systémy: * polygenní dědičnost * transkripční faktory pro vývoj svalů (MRF): MYOD1, MYF5, MYOG, MEF2 * ostatní transkripční faktory: NFAT proteiny (aktivace řízená kalcineurinem), buněčné onkogeny * četné další růstové faktory, steroidní hormony, další hormony ovlivňující metabolizmus * výživa
40 Faktory ovlivňující počet svalových vláken, jejich velikost a složení - Druh - Pohlaví - Selekce (genetická variabilita/heritabilita) - Plemeno - Výživa * prenatální výživa * postnatální výživa - Fyzická aktivita - Látky stimulující růst * růstový hormon * beta adrenergní agonisté (např. clenbuterol) * estrogeny a androgeny - Vliv genů velkého účinku: * Gen RYR1 (HAL) (u prasete) * Gen MSTN (u skotu) * Gen CLPG (u ovce) * Gen IGF2 (u prasete) * Gen PRKAG3 (u prasete)
41 Genomický imprinting Genomický imprinting představuje epigenetickou* modifikaci genů, jejímž důsledkem je monoalelická exprese. U savců je tento termín používán pro monoalelickou expresi nebo inaktivaci buď mateřské nebo otcovské alely určitého lokusu. Imprintovaná mutovaná alela se bude jevit jako recesivní, když bude zděděna od jednoho z rodičů, protože bude inaktivní (a tudíž bez projevu), zatímco druhá aktivní alela, zděděná od druhého rodiče, se bude jevit jako dominantní. Jedním z typů epigenetických modifikací u savců je metylace cytosinu v poloze 5, která se nejčastěji objevuje v CpG dinukleotidech. Tento typ DNA modifikace je velmi významný, neboť metylované sekvence DNA jsou transkripčně inaktivní. Odhady počtů imprintovaných genů u savců se pohybují v rozmezí od 100 do více než 200. Některé z nich jsou ukázány v tabulce. Imprinting se vyskytuje také u hmyzu a vyšších rostlin. *Epigenetické modifikace ovlivňují fenotyp aniž by měnily genotyp. Zahrnují změny ve vlastnostech buňky, které jsou děděny, ale nedochází ke změně genetické informace. Příklady: imprinting, inaktivace X chromozómu, poziční efekt, reprogramování, maternální efekty, modifikace histonů a heterochromatinu, aj.

42 Schéma znázorňující podstatu a účinek epigenetických mechanismů Histone deacetylation
43 Genomický imprinting Dva příklady hypotetického imprintovaného genu zodpovědného za barvu pokožky (VLEVO) V tomto příkladu je gen pro pigment maternálně imprintovaný (mateřská) alela je inaktivována). Spojením pohlavních buněk muže, který má alelu pro pigment, a ženy, která má alelu pro nevytváření pigmentu se narodí děti, které mají pouze fenotyp s pigmentem. V tomto příkladu je mateřská alela imprintována a inaktivována u potomků. Tudíž jedinou aktivně exprimovanou alelou je otcova alela pro pigment, která není imprintována u potomků. (VPRAVO) V tomto příkladu je gen pro pigment paternálně imprintovaný (otcovská alela je inaktivována). Spojením pohlavních buněk muže, který má alelu pro pigment a ženy, která má alelu pro nevytváření pigmentu se narodí děti, které mají fenotyp bez pigmentu. V tomto příkladu je imprintována otcovská alela a je inaktivována u potomků. Tudíž jedinou exprimovanou alelou je mateřská alela pro nevytváření pigmentu, která není imprintována u potomků.
44 Některé imprintované geny savců (celkem je imprintovaných asi 150 genů) Imprintované geny Igf2 Igf2r H19 Snrpn Ins1 and 2 U2afbp-rs Znf127 Par1 Par5 Xisf Mas IPW WT1 Mash-2 p57(kip2) Peg1/MEST ApoE Pk3 Pp/c Aktivní alela (P= otcovská; M = mateřská P M M P P P P P P P P P M M M P P P P
45 Inaktivace X chromozómu U samic je jeden z X chromozómů (získaný buď od matky nebo od otce) (náhodně) inaktivován v časném embryonálním vývoji, s inaktivovaným stejným chromozómem v buňkách, které vzniknou dělením této buňky. Jedná se o epigenetickou modifikaci - je to dědičná změna v genové funkci, aniž by došlo ke změně DNA Tento jev se také označuje jako lyonizace (podle anglické genetičky Mary Lyon). Barrovo tělísko kondenzovaný heterochromatin v procesu lyonizace hemizygot gen u diploidního organismu jen v jednom exempláři kompenzace dávky genu mechanizmus, který upravuje aktivitu genů lokalizovaných na X chromozómu u samců a samic tak, aby se vyrovnal rozdíl mezi haploidním stavem u samců a diploidním stavem u samic
46 Zjišťování pohlaví u zvířat - u embryí - v kriminalistice - rutinní genotypování - PCR amplifikace DNA fragmentu specifického pro Y chromozóm (gen SRY) Na Y chromozómu savců je gen SRY (sex determining region on chromosome Y), který je zodpovědný za zahájení vývoje samčího pohlaví. Gen SRY je lokalizován na p raménku Y chromozómu, směrem 3 od pseudoautozomální oblasti 1 (PAR1). PCR amplifikací fragmentu genu SRY je možno identifikovat samce. Jestliže je výsledek amplifikace negativní, jedinec je samice, ale nevylučuje to možnost chyby v experimentálním postupu, která způsobí stejný negativní výsledek. Pro spolehlivé stanovení je nutno do experimentu zahrnout i nějaký další gen, který je přítomen jak u samců, tak u samic, a ten musí být amplifikován ve stejné zkumavce jako pozitivní kontrola. - PCR amplifikace fragmentu genu AMEL (amelogenin), který je na X i Y chromozómu, ale tyto dvě formy se liší četnými inzercemi/delecemi, takže je možno rozlišit fragmenty specifické pro X a Y chromozóm, na základě rozdílné velikosti fragmentů, použitím elektroforézy v agarózovém gelu. (např. u člověka, skotu, ovce, jelena).
47 Srovnání sekvencí DNA fragmentů genu amelogeninu (AMEL) z X a Y chromozómu ovce a jelena. (.) ins/del

48 Rozdíly ve fragmentech genu AMEL u samců a samic ovce a jelena (gen AMEL je přítomen jak na X, tak na Y chromozómu, ale liší se ve velikosti to je způsobeno mnoha inzercemi/delecemi)
49 Priony (S.B. Prusiner, 1982) Základ prionové teorie Nervové buňky savců obsahují tzv. prionové bílkoviny (v angl. literatuře je častá zkratka PrP, případně PrP C pro zdravou buněčnou bílkovinu). Má se zato, že tyto prionové bílkoviny hrají významnou úlohu při odpočinku a spánku. Prion (PrP Sc ) představuje vadnou formu této normální bílkoviny, od které se odlišuje rozdílnou konformací (prostorovým uspořádáním). Důsledkem této změny konformace je mimořádná odolnost vůči různým fyzikálním vlivům, prakticky absolutní odolnost proti štěpným enzymům odklízejícím vadné bílkoviny a schopnost navazovat se na zdravé formy prionových bílkovin a konvertovat je na svoji vadnou formu. Následkem je, že se v buňce hromadí rostoucí shluky propojených vadných molekul prionové bílkoviny, kterých se buňka nedokáže zbavit, posléze je zcela zaplněna a uhyne. Předchozí část teorie je obecně přijímána. Navazující část je sice přijímána většinou odborníků, nicméně ne absolutně. Ta předpokládá možnost přenosu prionových onemocnění potravou nejen mezi jedinci jednoho druhu (což je též obecně přijímáno v případě nemoci kuru se to má za dostatečně podložené), ale i mezi jedinci různých druhů, zejména pak se zdůrazňuje přenos ovce (scrapie) kráva (bovinní spongiformní encefalopatie) a kráva (BSE) člověk (nová varianta CJD). Zde mají někteří vědci výhrady a nepovažují tuto část teorie za dostatečně prokázanou. K přenosům mezi druhy existuje několik předpokládaných obecností: šance na přenos mezi druhy je přímo úměrná podobnosti prionových bílkovin obou druhů největší šance na přenos skýtá konzumace mozku či jiných vysoce inervovaných tkání masožravci jsou obecně odolnější proti přenosu než býložravci.
50 Bovinní spongiformní encefalopatie Bovinní spongiformní encefalopatie (BSE, lidově nemoc šílených krav) je prionové onemocnění postihující skot, které se objevilo na konci 20. století. Předpokládá se, že by mohlo být bovinní variantou scrapie ovcí (svrbivka) a že požití tkání obsahující priony by mohlo u člověka vyvolávat tzv. novou variantu Creutzfeldt-Jacobovy choroby (nvcjd = New variant of Creutzfeldt-Jacob Disease). Jméno nemoc šílených krav získalo toto onemocnění proto, že v pozdních stadiích vyvolává u krav neobvyklé chování. Odborné jméno poukazuje na to, že jde o onemocnění skotu (bovinní), které se projevuje pórovatěním mozku, který pak připomíná v pozdějších stadiích houbu (spongiformní = houbotvárné; encefalopatie = nezánětlivé onemocnění mozku). Nemoc má velmi dlouhou inkubační dobu u krav a inkubační doba nové varianty CJD u člověka se odhaduje ještě delší (až přes deset let), z pochopitelných důvodů se tedy nepodařilo dokázat na základě nějakého konkrétního případu, že přenos na člověka je možný. Zatímco o přenosu nemoci mezi dobytčaty jednoho druhu (skrmováním nedostatečně tepelně zpracované masokostní moučky) se přiliš nepochybuje, myšlenka mezidruhového přenosu: ovce (scrapie ovcí) skot (BSE) člověk (nvcjd) je předmětem kontroverzí, byť ji většina vědců i vlád bere v rámci předběžné opatrnosti v potaz. Vychází se přitom z různých logických úvah a ze statistických analýz, z nichž některé ukazují na pozitivní korelaci mezi počtem nemocných kusů dobytka a počtem lidí trpících CJD. Kvůli této nemoci byla na přelomu 20. a 21. století zavedena rozsáhlá restriktivní opatření a byl vybit nespočet krav. V ČR bylo jen několik případů a to hlavně proto, že české kafilerie upravovaly masokostní moučku při 400 C. Při těchto teplotách se prion BSE ničí. Předpisy EU ale požadovaly nižší teploty, při kterých nedocházelo k destrukci bílkovinných prionů.

51 Normální a abnormální konformace prionů (vlevo normální, vpravo abnormální) The figure at right shows a model of two conformations of prion; on the left is the known, normal conformation of the structured C-terminal region of PrP C. The N-terminal region is not shown here for having a flexible structure in aqueous solution. The structured domain shown is mainly made of three spirals called alpha helices (pink), with two short 'flat' regions of beta sheet (β sheet) structure (green). On the right is a proposed model of how the abnormal prion might look. Although the exact 3D structure of PrP Sc is not known, there is increased β sheet content (green arrows) in the diseased form of the molecule. These β sheets are thought to lead to amyloid aggregation. Many prion protein transformations are characterized by increased ß-sheet content (depicted by green arrows). In this picture, the left image is the normal Prnp protein. After isomerization, note that the right-side image contains considerably more ß-sheet content. These ß-sheets often constitute an electric dipole and can lead to amyloid aggregation.


52 Předpokládaný mechanismus vzniku a množení abnormálních prionů
53
54

55 Schéma molekuly imunoglobulinu (tetramer sestávající ze dvou těžkých a dvou lehkých řetězců)
56
57
58
59
60
61
62
63

64 Konstrukce variabilních oblastí lehkého a těžkého řetězce z genových segmentů (subgenů)

65 Počty funkčních genových segmentů (subgenů) pro variabilní oblasti těžkých a lehkých řetězců lidského imunoglobulinu

66 Genomická organizace genových segmentů pro těžké a lehké řetězce imunoglobulinů člověka
67 Lokalizace genového klastru IGH na lidském chromozómu 14q32.3
68 Organizace genového klastru IGH na lidském chromozómu 14q32.33 (1. část)

69 Organizace genového klastru IGH na lidském chromozómu 14q32.33 (2. část)
70 Genomická mapa SLA komplexu (MHC komplex prasete) Each locus is annotated according to type, orientation, and position within the SLA. The tiling path of the sequenced BACs is shown on the top with overlaps shown in black. Below this, the distribution of repeats and C + G content across the region is shown. RNA genes identified using Rfam and CpG islands are also depicted. Segment 1 illustrates the section of the HLA class I region that is absent in pig. Segment 2 illustrates the HLA class III RCCX module absent in pig. Segment 3 highlights the porcine-specific BTNL cluster surrounding the centromere. Segment 4 illustrates the section of the HLA class II region containing the DP loci that is absent from pig.
71 Fyzická mapa SLA komplexu (prase) Black boxes: loci containing MHC-related sequences. White boxes: loci without MHC-related sequences. From the long arm to the short arm of chromosome 7, the order of the regions is class II (II), class III (III) and class I (I).

72 Struktura molekuly MHC třídy I člověka

73 Struktura molekuly MHC třídy II člověka
74 GMO Geneticky modifikovaný organismus (GMO) je takový organismus, jehož dědičný materiál byl změněn genetickou modifikací, t. j. cílenou změnou dědičného materiálu způsobem, kterého se nedosáhne přirozeně např. křížením, šlechtěním. Patří sem např. mutace, produkce transgenních zvířat, aj. GMO, do kterého byl metodami genového inženýrství vnesen gen jiného druhu, se označuje jako transgenní organismus. Přenos genů do živočišných buněk Živočišné somatické buňky mají omezený vývojový potenciál a za normálních okolností nemohou být využity pro produkci transgenních zvířat. Transformace zárodečných buněk vyžaduje přenos do pluripotentních nebo totipotentních buněk, jako vajíčka, časná embrya nebo izolované zárodečné buňky (vyjímečně transformace embryonální kmenové buňky nebo přenos somatických jader do enukleovaných oocytů). Změna genotypu vneseným transgenem může být přechodná nebo stálá, v závislosti na typu vektoru a osudu vneseného genetického materiálu.
75 Čtyři strategie přenosu genů do živočišných buněk: - přímý přenos fyzikální transfekcí (mikroinjekce, elektroporace, ultrazvuk nebo bombardování částicemi). Mikroinjekce injekce DNA do několika buněk v buněčné kultuře, do oocytů nebo buněk časného embrya. DNA může být injikována přímo do tkání nebo do krve (kde je rychle degradována). Elektroporace: Po vystavení buněk krátkému elektrickému pulzu se v buněčné membráně vytvoří póry velikosti nanometrů. Těmito póry vstupuje do buněk DNA, která je transportována do jádra. Podobný mechanismus se uplatňuje při ultrazvukové transfekci. Bombardování částicemi: Malé kovové částice, na jejichž povrchu je DNA, se vystřelují (např. vysokým tlakem plynu nebo elektrickým nábojem) do cílové tkáně. - chemicky zprostředkovaná transfekce. Koprecipitát DNA/fosforečnan vápenatý přilne k buňce a následně je internalizován (endocytózou). DNA se pak uvolní a část pronikne do jádra a může se exprimovat. Nízká účinnost. Transfekce při použití rozpustných komplexů polykationtových sloučenin (polyplexy), které vytvářejí spontánní elektrostatické interakce s DNA (např. polyaminy pozitivní náboj). Lipofekce: Použití směsi kationtových a neutrálních lipidů, které mohou vytvářet stabilní komplexy s DNA. Ty interagují s buněčnou membránou a následně se DNA dostává do buňky endocytózou. - přenos genů pomocí virů (adenoviry, baculoviry, herpesvirus, retroviry aj.) - přenos genů pomocí bakterií (bactofection): (zejména u rostlin Agrobacterium tumefaciens). Tato bakterie byla použita pro přenos DNA i u člověka. Bakterie pronikne do hostitelské živočišné buňky, podlehne lýze a uvolní se plazmidová DNA. Plazmidová DNA se pak dostane do jádra, kde se inkorporuje do hostitelského genomu a je exprimována.
76 Dvě stádia transfekce: - vnesení DNA do buňky - inkorporace do jádra a integrace do hostitelského chromozómu Většina transfektovaných buněk neintegruje cizí DNA. DNA se nachází v jádře v extrachromozómovém stavu, a když neobsahuje začátek replikace, který je funkční v hostitelské buňce, přežívá jen krátkodobě a je postupně degradována (přechodná, dočasná transformace; transient transformation). Jen v malé části transfektovaných buněk se DNA zabuduje do genomu a vytvoří nový genetický lokus, který se bude dědit (stabilní transformace). Vytváření a vlastnosti konstruktu (pro vysokou expresi transgenu) - kromě kódující sekvence je nutno připojit: - silný konstitutivní promotor - někdy je nutné včlenit intron (zejména při vytváření transgenních zvířat) - včlenit polyadenylační signál a připojit několik set adenosinových zbytků (poly(a)konec) - odstranit zbytečné nekódující sekvence (5 a 3 UTR) - optimalizovat sekvenci v blízkosti iniciačního místa pro translaci - inkorporovat signál pro cílovou organelu (např. sekvenci pro signální peptid) Selekční marker poskytuje vlastnost, která umožňuje transformovaným buňkám přežít v selekčních podmínkách. Reportérový gen je součástí konstruktu. Kóduje produkt, který může být prokázán při použití jednoduchého a levného testu. Testy pro zjišťování aktivity reportérového genu jsou kvantitativní, takže mohou být použity pro zjišťování účinnosti transformace (např. bioluminiscenční markery luciferáza nebo zelený fluoreskující protein).
77
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
Genetické markery. pro masnou produkci. Mgr. Jan Říha. Výzkumný ústav pro chov skotu, s.r.o.
 Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
Genotypování markerů užitkovosti a zdraví u skotu
 Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
Základní genetické pojmy
 Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Glosář - Cestina. Odchylka počtu chromozomů v jádře buňky od normy. Např. 45 nebo 47 chromozomů místo obvyklých 46. Příkladem je trizomie 21
 Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Glosář - Cestina alely aneuploidie asistovaná reprodukce autozomálně dominantní autozomálně recesivní BRCA chromozom chromozomová aberace cytogenetický laborant de novo Různé formy genu, které se nacházejí
Molekulární genetika II zimní semestr 4. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
VARIABILITY OF THE PORCINE MYOD1 GENE VARIABILITA GENU MYOD1 U PRASAT
 Verner J., Kuciel J. VARIABILITY OF THE PORCINE MYOD1 GENE VARIABILITA GENU MYOD1 U PRASAT Ústav genetiky, Agronomická fakulta, Mendelova zemědělská a lesnická univerzita v Brně, Zemědělská 1, 613 00,
Verner J., Kuciel J. VARIABILITY OF THE PORCINE MYOD1 GENE VARIABILITA GENU MYOD1 U PRASAT Ústav genetiky, Agronomická fakulta, Mendelova zemědělská a lesnická univerzita v Brně, Zemědělská 1, 613 00,
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Elektronoptický snímek viru mozaikové choroby tabáku. Mozaiková choroba tabáku. Schéma viru mozaikové choroby tabáku
 Obecná virologie Viry lat. virus šťáva, jed, v lékařské terminologii infekční činitel 1879 1882: první pokusný přenos virového onemocnění (mozaiková choroba tabáku) 1898: první pokusný přenos živočišného
Obecná virologie Viry lat. virus šťáva, jed, v lékařské terminologii infekční činitel 1879 1882: první pokusný přenos virového onemocnění (mozaiková choroba tabáku) 1898: první pokusný přenos živočišného
Deoxyribonukleová kyselina (DNA)
") Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Gonosomální dědičnost
 Gonosomální dědičnost Praktické cvičení č.12 Jaro 2016 Aneta Kohutová aneta.baumeisterova@gmail.com Biologický ústav Lékařská fakulta Masarykova univerzita Kamenice 5, 625 00 Brno Cíle cvičení Student:
Gonosomální dědičnost Praktické cvičení č.12 Jaro 2016 Aneta Kohutová aneta.baumeisterova@gmail.com Biologický ústav Lékařská fakulta Masarykova univerzita Kamenice 5, 625 00 Brno Cíle cvičení Student:
Konzervační genetika INBREEDING. Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.
 Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
GENETIKA. Dědičnost a pohlaví
 GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
GENETIKA Dědičnost a pohlaví Chromozómové určení pohlaví Dvoudomé rostliny a gonochoristé (živočichové odděleného pohlaví) mají pohlaví určeno dědičně chromozómovou výbavou jedince = dvojicí pohlavních
Dědičnost a pohlaví. KBI/GENE Mgr. Zbyněk Houdek
 Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162
 Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 ZŠ Určeno pro Sekce Předmět Téma / kapitola Prameny 8. třída (pro 3. 9. třídy)
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 ZŠ Určeno pro Sekce Předmět Téma / kapitola Prameny 8. třída (pro 3. 9. třídy)
Buňky, tkáně, orgány, soustavy
 Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Cvičeníč. 10 Dědičnost a pohlaví. Mgr. Zbyněk Houdek
 Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Genetika pohlaví genetická determinace pohlaví
 Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ
 FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ SARKOIDÓZOU: POTVRZENÍ VÝSLEDKŮ CELOGENOMOVÉ ASOCIAČNÍ STUDIE. Sťahelová A. 1, Mrázek F. 1, Kriegová E. 1, Hutyrová B. 2, Kubištová Z. 1, Kolek V.
FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ SARKOIDÓZOU: POTVRZENÍ VÝSLEDKŮ CELOGENOMOVÉ ASOCIAČNÍ STUDIE. Sťahelová A. 1, Mrázek F. 1, Kriegová E. 1, Hutyrová B. 2, Kubištová Z. 1, Kolek V.
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE
 ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Genetika zvířat - MENDELU
 Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Dědičnost vázaná na X chromosom
 12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
12 Dědičnost vázaná na X chromosom EuroGentest - Volně přístupné webové stránky s informacemi o genetickém vyšetření (v angličtině). www.eurogentest.org Orphanet - Volně přístupné webové stránky s informacemi
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Genová etiologie nemocí
 Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Dr. Ing. Tomáš. Ústav morfologie, fyziologie a genetiky zvířat MZLU v Brně. mendelu.cz. urban@mendelu. Genetika 1. C-hodnota 3,43 3,93 2,34 3,50 3,24
 Genetika skotu, ovcí a koz Dr. Ing. Tomáš Urban Ústav morfologie, fyziologie a genetiky zvířat MZLU v Brně urban@mendelu mendelu.cz Genetika 1 Biodiverzitní charakteristiky Druh Skot Bos taurus Ovce Ovis
Genetika skotu, ovcí a koz Dr. Ing. Tomáš Urban Ústav morfologie, fyziologie a genetiky zvířat MZLU v Brně urban@mendelu mendelu.cz Genetika 1 Biodiverzitní charakteristiky Druh Skot Bos taurus Ovce Ovis
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Hodnocení plemenných + chovných + užitkových prasat
 Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
ší šířen VAZEBNÁ ANALÝZA Vazba genů
 VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
Molekulární genetika, mutace. Mendelismus
 Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
GENETIKA V MYSLIVOSTI
 GENETIKA V MYSLIVOSTI Historie genetiky V r. 1865 publikoval Johann Gregor Mendel výsledky svých pokusů s hrachem v časopisu Brněnského přírodovědeckého spolku, kde formuloval principy přenosu vlastností
GENETIKA V MYSLIVOSTI Historie genetiky V r. 1865 publikoval Johann Gregor Mendel výsledky svých pokusů s hrachem v časopisu Brněnského přírodovědeckého spolku, kde formuloval principy přenosu vlastností
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
polymorfní = vícetvarý, mnohotvárný
 Genetický polymorfismus s Řeckyy morphos = tvar polymorfní = vícetvarý, mnohotvárný Genetický polymorfismus je tedy označení pro výskyt téhož znaku ve více tvarech, formách, přičemž tato mnohotvárnost
Genetický polymorfismus s Řeckyy morphos = tvar polymorfní = vícetvarý, mnohotvárný Genetický polymorfismus je tedy označení pro výskyt téhož znaku ve více tvarech, formách, přičemž tato mnohotvárnost
Působení genů. Gen. Znak
 Genové interakce Působení genů Gen Znak Dědičnost Potomek získává predispozice k vlastnostem z rodičovské buňky nebo organismu. Vlastnosti přenášené do další generace nemusí být zcela totožné s vlastnostmi
Genové interakce Působení genů Gen Znak Dědičnost Potomek získává predispozice k vlastnostem z rodičovské buňky nebo organismu. Vlastnosti přenášené do další generace nemusí být zcela totožné s vlastnostmi
PhD. České Budějovice
 PhD. České Budějovice Sledování a využívání poznatků o genetické biodiverzitě mezi populacemi hospodářských zvířat Dvořák Josef prof. Genetiky živočichů Ústavu genetiky MZLU v Brně Pro seminář doktorského
PhD. České Budějovice Sledování a využívání poznatků o genetické biodiverzitě mezi populacemi hospodářských zvířat Dvořák Josef prof. Genetiky živočichů Ústavu genetiky MZLU v Brně Pro seminář doktorského
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
rodokmeny vazby mezi členy rodiny + popis pro konkrétní sledovaný znak využití Mendelových zákonů v lékařství genetické konzultace o možném výskytu
 Genealogie Monogenní dědičnost rodokmeny vazby mezi členy rodiny + popis pro konkrétní sledovaný znak využití Mendelových zákonů v lékařství genetické konzultace o možném výskytu onemocnění v rodině Genealogické
Genealogie Monogenní dědičnost rodokmeny vazby mezi členy rodiny + popis pro konkrétní sledovaný znak využití Mendelových zákonů v lékařství genetické konzultace o možném výskytu onemocnění v rodině Genealogické
GENETIKA 1. Úvod do světa dědičnosti. Historie
 GENETIKA 1. Úvod do světa dědičnosti Historie Základní informace Genetika = věda zabývající se dědičností a proměnlivostí živých soustav sleduje variabilitu (=rozdílnost) a přenos druhových a dědičných
GENETIKA 1. Úvod do světa dědičnosti Historie Základní informace Genetika = věda zabývající se dědičností a proměnlivostí živých soustav sleduje variabilitu (=rozdílnost) a přenos druhových a dědičných
Výukový materiál zpracován v rámci operačního projektu. EU peníze školám. Registrační číslo projektu: CZ.1.07/1.5.00/34.0512
 Výukový materiál zpracován v rámci operačního projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0512 Střední škola ekonomiky, obchodu a služeb SČMSD Benešov, s.r.o. ZDRAVOVĚDA Genetika
Výukový materiál zpracován v rámci operačního projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0512 Střední škola ekonomiky, obchodu a služeb SČMSD Benešov, s.r.o. ZDRAVOVĚDA Genetika
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Genetika kvantitativních znaků. - principy, vlastnosti a aplikace statistiky
 Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Mutace genu pro Connexin 26 jako významná příčina nedoslýchavosti
 Mutace genu pro Connexin 26 jako významná příčina nedoslýchavosti Petr Lesný 1, Pavel Seeman 2, Daniel Groh 1 1 ORL klinika UK 2. LF a FN Motol Subkatedra dětské ORL IPVZ Přednosta doc. MUDr. Zdeněk Kabelka
Mutace genu pro Connexin 26 jako významná příčina nedoslýchavosti Petr Lesný 1, Pavel Seeman 2, Daniel Groh 1 1 ORL klinika UK 2. LF a FN Motol Subkatedra dětské ORL IPVZ Přednosta doc. MUDr. Zdeněk Kabelka
VYBRANÉ GENETICKÉ ÚLOHY II.
 VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které
VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které