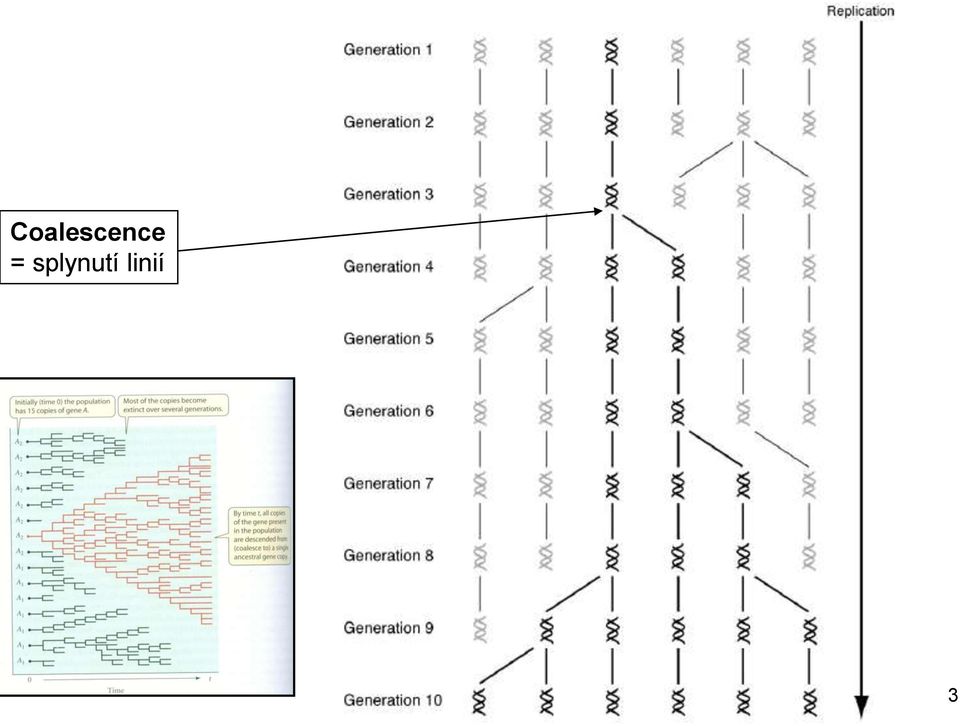
Coalescence = splynutí linií
|
|
|
- Martina Moravcová
- před 9 lety
- Počet zobrazení:
Transkript
1 Koalescence 1

2 2
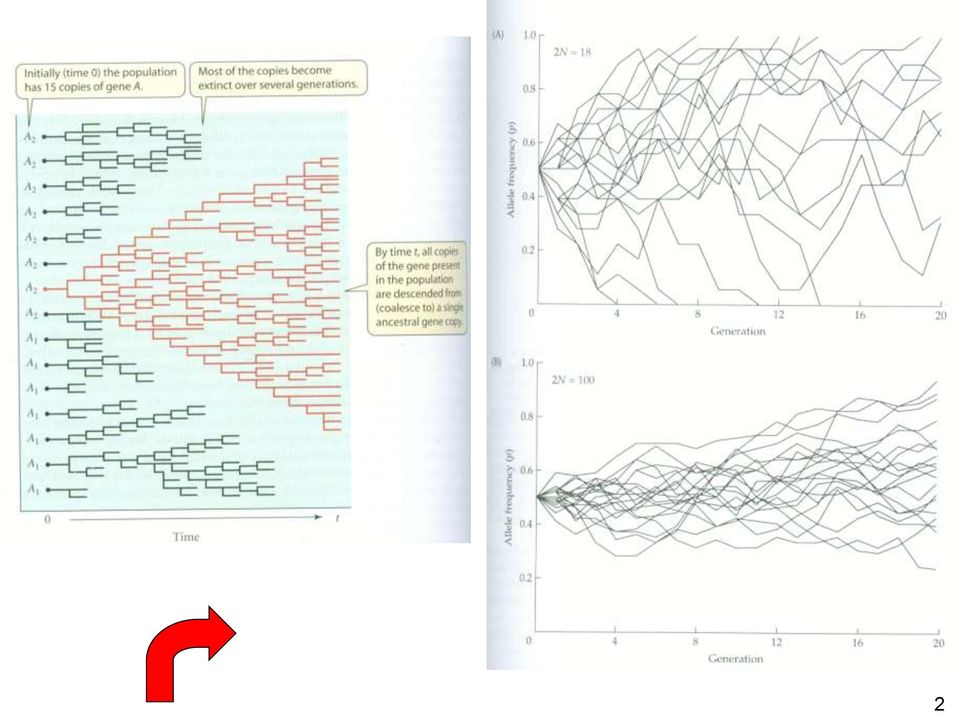
3 Coalescence = splynutí linií 3
4 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme opačně v čase než u klasických modelů populační genetiky (Wright-Fisher model) 4

5 Wright-Fisher model čas Sewall Wright Sir Ronald Fisher 5

6 Wright-Fisher model čas Sewall Wright Sir Ronald Fisher 6

7 Wright-Fisher model čas Sewall Wright Sir Ronald Fisher 7

8 Wright-Fisher model čas Sewall Wright Sir Ronald Fisher 8

9 Teorie koalescence čas koalescence = splynutí linií 9

10 Teorie koalescence čas koalescence = splynutí linií 10

11 Teorie koalescence čas koalescence = splynutí linií 11

12 Teorie koalescence čas koalescence = splynutí linií 12


13 13

14 14

15 Genové genealogie (gene trees) Popisují cestu jakou se jednotlivé geny dědily z generace na generaci Geny, které nejsou ve vazbě, mají jiné genealogie genový strom 15

16 Genové genealogie Geny, které mají společného předka Identical by descent Nejbližší společný předek vybraných linií Most Recent Common Ancestor (MRCA) Coalescence = splynutí linií v MRCA Zpět v čase MRCA 16
 Coalescence = splynutí linií v MRCA Zpět v")
17 MRCA - příklad Stolař z Rawengisburgu Jan Montclinger Anna Skobrtalin (vdova Škobrtalová) * Josef Munclinger 17
 * 11.2.")
18 Jan MRCA Josef František Jan Josef František Method Josef Václav František Jan Ignác Antonín Method Miloslav Josef Richard Josef Jindřích Miloslav Method Václav Karel Ladislav 18

19 Meč a přeslice Rodokmen odpovídá historii chromosomu Y (pokud ovšem ženy nebyly nevěrné) Rodokmen přes matky (a pro mtdna) by byl úplně jiný! A jiný bude pro jiné geny 19

20 Mitochondriální Eva Adam pro chromosom Y 20

21 Přetrvání ancestrálního polymorfismu (species tree gene tree) 21
22 Přetrvání ancestrálního polymorfismu (species tree gene tree) 22
23 Chr 19 23
24 čas Pravděpodobnost koalescence v předchozí generaci 1/N mtdna 24
25 Koalescence čas mtdna, Y Lineage sorting přežije jen jedna linie sekvence však nejsou identické díky mutacím! Koalescence splynutí současných linií v MRCA Most Recent Common Ancestor Čas k MCRA ~4N e generací N e
26 N (N e ) může být různé pro různé geny mtdna jaderný lokus 26
27 mtdna, Y (cpdna) versus jaderné geny Jaderné geny: složitější osud, větší N e (4x) delší doba pro lineage sorting jiné výsledky u nedávno oddělených populací mtdna dospěje rychleji do reciproční monofylie 27
28 Y a N e N e je čtvrtinová vzhledem k autosomům, srovnatelná s mtdna Větší náchylnost k driftu Rozdíl ve varianci reprodukčního úspěchu mužů (samců) a žen (samic) nižší N e, nedávné koalescence 28
29 29
30 Max. divergence člověk/šimpanz 30
31 mutace Gene copies (ač mají společného předka) se liší mutacemi Ө expected number of mutations separating two gene copies Ө = 4Nμ μ mutační rychlost Závislost na N Malé populace méně variability (kratší čas koalescence, tedy k MRCA málo času na mutace) 31
32 Lze odhadnout frekvenční spektrum mutací! 32
33 Gene tree většinou neznáme Haplotype tree toto zrekonstruujeme 33
34 Nezakořeněné stromy, Haplotype network, síť haplotypů (často nevíme, co bylo dřív) Toto jsou sice stromy, ale bežně se jim říká sítě. Předpokládaný nikoliv však pozorovaný haplotyp Konkrétní zjištěný haplotyp Velikost kroužku = počet jedinců s tímto haplotypem 34

35 Různé stromy, různé významy, různá interpretace 35
36 mtdna a Mitochondriální Eva Větve stromu Afrika jedna z afrických větví - zbytek světa MRCA pro mtdna až 250 tisíc let MRCA s neandrtálci 465 tisíc let ( ) Severní Afrika zpětně osídlena z Levantu (před tisíci let) Eve OR Steve? předpoklady neutrality a striktně maternální dědičnosti 36
37 Phylogeography Geomys pinetis (pytlonoš) Avise et al maximum parsimony network 37
 je zásadní i mutační")
38 Pro výsledné haplotypové stromy (sítě) je zásadní i mutační rychlost 38
39 Rutilus frisii Plotice perleťová 39
40 Jaderné lokusy složitá a delší historie, často ale méně mutací 40
41 Rekombinace mění genealogie podél chromosomu Vystopovat společného předka pro autosomální lokusy je obtížné, protože rekombinují. Rekombinace mění genealogie. Na autosomech mají geny, které nejsou ve vazbě různé genealogie. Čím blíže jsou u sebe na chromosomu, tím mají podobnější genealogii. Jak rychle se genealogie podél genomu mění záleží na míře vazebné nerovnováhy (u druhů s velkou Ne a vysokou mírou rekombinace mohou být nerekobinující oblasti velmi krátké několik desítek až stovek bp, tj. méně než délka genu!) 41
42 K odhadu MRCA stačí malý vzorek Pravděpodobnost zachycení MRCA celé populace vzorkem o n jedincích: (n 1)/(n + 1) n = 3 0,5 n = 20 > 0,9 Versus Klasická populační (frekventistická) genetika Odhad frekvence velký vzorek 42
43 Koalescence Náhodné genealogické stromy Takto můžou vypadat genealogie vzniklé náhodným slučováním linií. Různé lokusy představují nezávislé replikáty společné historie. Čím víc nezávislých lokusů studujeme, tím lepší odhady o historii populace dostaneme. 43
44 Coalescent Koalescenční teorie Interpretace času ke společnému předku alel pomocí evolučních procesů Procesy: drift, migrace, selekce, změna populační velikosti 44
45 Odhad času k MRCA K čemu to teda vlastně je dobré? Pozorované odchylky (hodnoty parametrů) biologicky zajímavé děje (demografie, selekce, migrace) Konkrétní štěpení stromů nás vlastně ani nezajímá (rozdíl oproti fylogenetickým metodám) Detekce: Historie šíření druhů, migrace Demografických událostí (expanze, bottlenecky) Selekce Problém: Selekce i demografické události podobný signál 45
46 Rozdíl mezi fylogenetickým a populačně genetickým (koalescenčním) přístupem Data, strom, mutační model = strom Data, strom, mutační model, parametry 46
47 Detekce demografických událostí (expanze) 47
48 Zvonci v Evropě 48
49 49
50 50
51 Bayesian Skyline Plot Změny velikostí populací Bison priscus 51
52 Odhady migrace 52
53 Model Isolation with Migration Rozlišení ancestrálních polymorfismů a migrací Velikosti populací Čas divergence Migrace Programy IM, IMa, IMa2 53
54 54
55 Zbadáno 48 genů Population size Migration 55
56 Positivní selekce Simulováno 10 populací Nová mutace N je 100 Pouze drift Fitness - heterozygota pro novou alelu o 10 % vyšší - homozygota o 20 % vyšší 56

57 Selective sweep, hitchhiking 57

58 Positive selection čas neutral positive Výhodná varianta Rychlé šíření Společný předek nedávno selective sweep 58
59 Background selection Odstraňování škodlivých mutací Zánik linií (i těch, co nesou v blízkosti neutrální nebo slabě výhodné mutace) Snížení N E rychlejší fixace Kratší čas ke společnému předkovi neutral background 59
60 Role heterozygotů Against heterozygotes (heterozygote disadvantage, underdominance), nestabilní homozygotes (heterozygote advantage, overdominance), stabilní frekvence alel odpovídá w homozygotů underdominance overdominance 60
61 Balancing selection Overdominance je jen jedním z příkladů Balancing selection = selekce udržující polymorfismus Jsou i jiné příčiny Často Frequency-dependent selection The fitness of a genotype depends on the genotype frequencies in the population. Inverse frequency - dependent selection the rarer a phenotype the greater its fitness 61

62 Balancing selection čas neutral balancing Selekce na udržení více variant Overdominance, frekvenční závislost Společný předek variant hlouběji v minulosti Extrémní příklad mezidruhové polymorfismy, např. MHC 62
63 Trans-species polymorfismus: Microtus Arvicola Clethrionomys 7-8 miliónů let MHC: stejné nebo podobné alely byly nalezeny u různých druhů když působí selekce, tak se výhodné sekvence udrží i přes období divergence druhů 63

64 Selekce a variabilita Positivní i negativní snižují variabilitu Balancing zvyšuje variabilitu udržení více linií delší čas ke společnému předku více času na mutace 64
65 Zásadní problém Vliv selekce často těžko odlišitelný od jiných vlivů.????????????????????????????????????? Vidím výsledek: Selekce Demografické změny Populační struktury Migrace????????????????????????????????????? Pozor na analýzy založené na malém počtu genů. 65
Coalesce spojit se, splynout, sloučit se. Didaktická simulace Coalescence = splynutí linií
 Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
FYLOGEOGRAFIE A KOALESCENCE
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
Osud jednotlivých kopií genů v populaci genové stromy
 FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
FYLOGEOGRAFIE A KOALESCENCE A T T T T G G G C C A C T G Koalescence Osud jednotlivých kopií genů v populaci genové stromy Species trees vs. gene trees: gen A Species trees vs. gene trees: gen B Fylogenetické
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika III. Radka Reifová
 Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Úvod do populační genetiky
 Úvod do populační genetiky 1 Termíny Gene Locus (loci) Allele alternative forms of a gene Genome single copy of all of the genes Diploid - 2 alely pro každý gen (výjimka Chr Y, cpdna a mtdna) 2 genomy
Úvod do populační genetiky 1 Termíny Gene Locus (loci) Allele alternative forms of a gene Genome single copy of all of the genes Diploid - 2 alely pro každý gen (výjimka Chr Y, cpdna a mtdna) 2 genomy
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Opakování z minula. Ideální populace (Fischer Wright model) Hardy-Weinberg principle odchylky: inbreeding, asortativní křížení, pop.
 Hardy-Weinberg principle odchylky: inbreeding, asortativní křížení, pop.") Opakování z minula Ideální populace (Fischer Wright model) Hardy-Weinberg principle odchylky: inbreeding, asortativní křížení, pop. struktura Drift (náhoda) Effective population size 1 Selection Mutace
Opakování z minula Ideální populace (Fischer Wright model) Hardy-Weinberg principle odchylky: inbreeding, asortativní křížení, pop. struktura Drift (náhoda) Effective population size 1 Selection Mutace
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Frekvence alel 40... C C... H 10... CC... Q. frekvence p alely C... (2 x 150 + 40)/400 =0.85. frekvence q alely C... (2 x 10 + 40)/400 =0.
/400 =0.85. frekvence q alely C... (2 x 10 + 40)/400 =0.") Frekvence alel Př.1 U nocenky podmiňuje neúplně dominantní alela C tmavožluté zbarvení květu, alela C bílé; heterozygoti jsou světle žlutí. V populaci bylo celkem u vyšetřených 200 rostlin pozorováno:
Frekvence alel Př.1 U nocenky podmiňuje neúplně dominantní alela C tmavožluté zbarvení květu, alela C bílé; heterozygoti jsou světle žlutí. V populaci bylo celkem u vyšetřených 200 rostlin pozorováno:
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
Evoluční (populační) genetika Radka Reifová
 genetika Radka Reifová") Evoluční (populační) genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Evoluční genetika Obor zabývající se genetickou variabilitou v populacích a procesy,
Evoluční (populační) genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Evoluční genetika Obor zabývající se genetickou variabilitou v populacích a procesy,
Typologická koncepce druhu
 Speciace Co je to druh? Nebudu zde ani probírat různé definice pojmu druh. Žádná z nich až dosud neuspokojila všechny přírodovědce, ale každý přírodovědec zhruba ví, co míní tím, když mluví o druhu. (Charles
Speciace Co je to druh? Nebudu zde ani probírat různé definice pojmu druh. Žádná z nich až dosud neuspokojila všechny přírodovědce, ale každý přírodovědec zhruba ví, co míní tím, když mluví o druhu. (Charles
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Opakování z minula Za předpokladu neutrality Genetická diverzita v rámci druhu θ = 4N e μ Genetická divergence mezi druhy D = 2μt Proč se substituční rychlost liší u
Populační genetika II Radka Reifová Opakování z minula Za předpokladu neutrality Genetická diverzita v rámci druhu θ = 4N e μ Genetická divergence mezi druhy D = 2μt Proč se substituční rychlost liší u
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).
: RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).") Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele)
") Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Název: Genetické zákonitosti v populacích
 Název: Genetické zákonitosti v populacích Výukové materiály Autor: PaedDr. Pavel Svoboda Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: biologie, statistika,
Název: Genetické zákonitosti v populacích Výukové materiály Autor: PaedDr. Pavel Svoboda Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: biologie, statistika,
1.3.1 Kruhový pohyb. Předpoklady: 1105
 .. Kruhový pohyb Předpoklady: 05 Předměty kolem nás se pohybují různými způsoby. Nejde pouze o přímočaré nebo křivočaré posuvné pohyby. Velmi často se předměty otáčí (a některé se přitom pohybují zároveň
.. Kruhový pohyb Předpoklady: 05 Předměty kolem nás se pohybují různými způsoby. Nejde pouze o přímočaré nebo křivočaré posuvné pohyby. Velmi často se předměty otáčí (a některé se přitom pohybují zároveň
= oplození mezi biologicky příbuznými jedinci
 = oplození mezi biologicky příbuznými jedinci Jestliže každý z nás má 2 rodiče, pak má 4 prarodiče, 8 praprarodičů... obecně 2 n předků tj. po 10 generacích 2 10 = 1024, po 30 generacích = 1 073 741 824
= oplození mezi biologicky příbuznými jedinci Jestliže každý z nás má 2 rodiče, pak má 4 prarodiče, 8 praprarodičů... obecně 2 n předků tj. po 10 generacích 2 10 = 1024, po 30 generacích = 1 073 741 824
Nondisjunkce v II. meiotickém dělení zygota
 2. semestr, 1. výukový týden OPAKOVÁNÍ str. 1 OPAKOVÁNÍ VYBRANÉ PŘÍKLADY letního semestru: 1. u Downova a Klinefelterova syndromu, 2. Hodnocení karyotypu s aberací, 3. Mono- a dihybridismus, 4. Vazba genů
2. semestr, 1. výukový týden OPAKOVÁNÍ str. 1 OPAKOVÁNÍ VYBRANÉ PŘÍKLADY letního semestru: 1. u Downova a Klinefelterova syndromu, 2. Hodnocení karyotypu s aberací, 3. Mono- a dihybridismus, 4. Vazba genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat
 Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Genetické markery ve studiu genetické diverzity v populacích hospodářských zvířat Bakalářská
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Genetické markery ve studiu genetické diverzity v populacích hospodářských zvířat Bakalářská
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Genetika vzácných druhů zuzmun
 Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
GENETIKA Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální dědičnost
 Polygenní dědičnost Multifaktoriální dědičnost") GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Konzervační genetika INBREEDING. Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.
 Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
2.8.9 Parametrické rovnice a nerovnice s absolutní hodnotou
 .8.9 Parametrické rovnice a nerovnice s absolutní hodnotou Předpoklady: 0,, 806 Pedagogická poznámka: Opět si napíšeme na začátku hodiny na tabuli jednotlivé kroky postupu při řešení rovnic (nerovnic)
.8.9 Parametrické rovnice a nerovnice s absolutní hodnotou Předpoklady: 0,, 806 Pedagogická poznámka: Opět si napíšeme na začátku hodiny na tabuli jednotlivé kroky postupu při řešení rovnic (nerovnic)
Základy genetiky populací
 Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
SPOLEHLIVOST KONSTRUKCÍ & TEORIE SPOLEHLIVOSTI část 2: Statistika a pravděpodobnost
 SPOLEHLIVOST KONSTRUKCÍ & TEORIE SPOLEHLIVOSTI část 2: Statistika a pravděpodobnost Drahomír Novák Jan Eliáš 2012 Spolehlivost konstrukcí, Drahomír Novák & Jan Eliáš 1 část 2 Statistika a pravděpodobnost
SPOLEHLIVOST KONSTRUKCÍ & TEORIE SPOLEHLIVOSTI část 2: Statistika a pravděpodobnost Drahomír Novák Jan Eliáš 2012 Spolehlivost konstrukcí, Drahomír Novák & Jan Eliáš 1 část 2 Statistika a pravděpodobnost
Tok GI v buňce. Genetický polymorfizmus popis struktury populací. Organizace genetického materiálu. Definice polymorfismu
 Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA
 učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
Příbuznost a inbreeding
 Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Genetika populací. KBI / GENE Mgr. Zbyněk Houdek
 Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH
 RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání
RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
4.6.6 Složený sériový RLC obvod střídavého proudu
 4.6.6 Složený sériový LC obvod střídavého proudu Předpoklady: 41, 4605 Minulá hodina: odpor i induktance omezují proud ve střídavém obvodu, nemůžeme je však sčítat normálně, ale musíme použít Pythagorovu
4.6.6 Složený sériový LC obvod střídavého proudu Předpoklady: 41, 4605 Minulá hodina: odpor i induktance omezují proud ve střídavém obvodu, nemůžeme je však sčítat normálně, ale musíme použít Pythagorovu
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Tepelná výměna. výměna tepla může probíhat vedením (kondukce), sáláním (radiace) nebo prouděním (konvekce).
, sáláním (radiace) nebo prouděním (konvekce).") Tepelná výměna tepelná výměna je termodynamický děj, při kterém dochází k samovolné výměně tepla mezi dvěma tělesy s různou teplotou. Tepelná výměna vždy probíhá tak, že teplejší těleso předává svou vnitřní
Tepelná výměna tepelná výměna je termodynamický děj, při kterém dochází k samovolné výměně tepla mezi dvěma tělesy s různou teplotou. Tepelná výměna vždy probíhá tak, že teplejší těleso předává svou vnitřní
Genetika pohlaví genetická determinace pohlaví
 Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
4.2.7 Voltampérová charakteristika rezistoru a žárovky
 4.2.7 Voltampérová charakteristika rezistoru a žárovky Předpoklady: 4205 Pedagogická poznámka: Tuto hodinu učím jako běžnou jednohodinovku s celou třídou. Některé dvojice stihnou naměřit více odporů. Voltampérová
4.2.7 Voltampérová charakteristika rezistoru a žárovky Předpoklady: 4205 Pedagogická poznámka: Tuto hodinu učím jako běžnou jednohodinovku s celou třídou. Některé dvojice stihnou naměřit více odporů. Voltampérová
Testování lidské identity
 Testování lidské identity Brno, 2009 J.M.Butler Forensic DNA Typing workshop, 2006 Bryan Sykes Sedm dcer Eviných, 2005 Využití testování lidské identity Řešení trestních činů shoda mezi podezřelým a stopou
Testování lidské identity Brno, 2009 J.M.Butler Forensic DNA Typing workshop, 2006 Bryan Sykes Sedm dcer Eviných, 2005 Využití testování lidské identity Řešení trestních činů shoda mezi podezřelým a stopou
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Genetika kvantitativních znaků. - principy, vlastnosti a aplikace statistiky
 Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
( ) 2.4.4 Kreslení grafů funkcí metodou dělení definičního oboru I. Předpoklady: 2401, 2208
 2.4.4 Kreslení grafů funkcí metodou dělení definičního oboru I. Předpoklady: 2401, 2208") .. Kreslení grafů funkcí metodou dělení definičního oboru I Předpoklady: 01, 08 Opakování: Pokud jsme při řešení nerovnic potřebovali vynásobit nerovnici výrazem, nemohli jsme postupovat pro všechna čísla
.. Kreslení grafů funkcí metodou dělení definičního oboru I Předpoklady: 01, 08 Opakování: Pokud jsme při řešení nerovnic potřebovali vynásobit nerovnici výrazem, nemohli jsme postupovat pro všechna čísla
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Úvod do obecné genetiky
 Úvod do obecné genetiky GENETIKA studuje zákonitosti dědičnosti a proměnlivosti živých organismů GENETIKA dědičnost - schopnost uchovávat soubor dědičných informací a předávat je nezměněný potomkům GENETIKA
Úvod do obecné genetiky GENETIKA studuje zákonitosti dědičnosti a proměnlivosti živých organismů GENETIKA dědičnost - schopnost uchovávat soubor dědičných informací a předávat je nezměněný potomkům GENETIKA
{ } 9.1.9 Kombinace II. Předpoklady: 9108. =. Vypiš všechny dvoučlenné kombinace sestavené z těchto pěti prvků. Urči počet kombinací pomocí vzorce.
 9.1.9 Kombinace II Předpoklady: 9108 Př. 1: Je dána pěti prvková množina: M { a; b; c; d; e} =. Vypiš všechny dvoučlenné kombinace sestavené z těchto pěti prvků. Urči počet kombinací pomocí vzorce. Vypisujeme
9.1.9 Kombinace II Předpoklady: 9108 Př. 1: Je dána pěti prvková množina: M { a; b; c; d; e} =. Vypiš všechny dvoučlenné kombinace sestavené z těchto pěti prvků. Urči počet kombinací pomocí vzorce. Vypisujeme
Genetické markery. pro masnou produkci. Mgr. Jan Říha. Výzkumný ústav pro chov skotu, s.r.o.
 Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Vypracované otázky z genetiky
 Vypracované otázky z genetiky 2015/2016 Dana Hatoňová 1. Základní zákony genetiky 2. Dihybridismus 3. Aditivní model polygenní dědičnosti 4. Interakce nealelních genů 5. Genová vazba 6. Genotyp a jeho
Vypracované otázky z genetiky 2015/2016 Dana Hatoňová 1. Základní zákony genetiky 2. Dihybridismus 3. Aditivní model polygenní dědičnosti 4. Interakce nealelních genů 5. Genová vazba 6. Genotyp a jeho
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Pojem plemeno je používán pro rasy, které vznikly záměrnou činností člověka, např. plemena hospodářských zvířat.
 POPULAČNÍ GENETIKA Populační genetika se zabývá genetickými zákonitostmi v definovaných souborech jedinců téhož druhu. Genetické vztahy uvnitř populace jsou komplikované, a proto se v populační genetice
POPULAČNÍ GENETIKA Populační genetika se zabývá genetickými zákonitostmi v definovaných souborech jedinců téhož druhu. Genetické vztahy uvnitř populace jsou komplikované, a proto se v populační genetice
2. Věková a vzdělanostní homogamie párů žijících v manželství a v nesezdaných soužitích
 2. Věková a vzdělanostní homogamie párů žijících v manželství a v nesezdaných soužitích Díky datům o domácnostech ze sčítání lidu je možné zkoumat homogamii partnerů nejen u manželských párů, ale také
2. Věková a vzdělanostní homogamie párů žijících v manželství a v nesezdaných soužitích Díky datům o domácnostech ze sčítání lidu je možné zkoumat homogamii partnerů nejen u manželských párů, ale také
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
3.2.4 Podobnost trojúhelníků II
 3..4 odobnost trojúhelníků II ředpoklady: 33 ř. 1: Na obrázku jsou nakresleny podobné trojúhelníky. Zapiš jejich podobnost (aby bylo zřejmé, který vrchol prvního trojúhelníku odpovídá vrcholu druhého trojúhelníku).
3..4 odobnost trojúhelníků II ředpoklady: 33 ř. 1: Na obrázku jsou nakresleny podobné trojúhelníky. Zapiš jejich podobnost (aby bylo zřejmé, který vrchol prvního trojúhelníku odpovídá vrcholu druhého trojúhelníku).
Důsledky selekce v populaci - cvičení
 Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).
: Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).") Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích
 Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Populačně-genetická data
 Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
Genetika populací. kvalitativních znaků
 Genetika populací kvalitativních znaků Úroveň studia genetických procesů Molekulární - struktura a funkce nukleových kyselin Buněčná buněčné struktury s významem pro genetiku, genetické procesy na buněčné
Genetika populací kvalitativních znaků Úroveň studia genetických procesů Molekulární - struktura a funkce nukleových kyselin Buněčná buněčné struktury s významem pro genetiku, genetické procesy na buněčné
Hardy-Weinbergův zákon - cvičení
 Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Příklady z populační genetiky lesních dřevin
 Obecná genetika Příklady z populační genetiky lesních dřevin Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem a Státním
Obecná genetika Příklady z populační genetiky lesních dřevin Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem a Státním
1. 21.2.2012 Klinická genetika genetické poradenství MUDr. Renata Gaillyová, Ph.D.
 Plán výuky jarní semestr 2011/2012 LF ošetřovatelství, porodní asistentka presenční forma Velká posluchárna, Komenského náměstí 2 Úterý 10:20-12:00 sudé týdny (první týden je sudý) 1. 21.2.2012 Klinická
Plán výuky jarní semestr 2011/2012 LF ošetřovatelství, porodní asistentka presenční forma Velká posluchárna, Komenského náměstí 2 Úterý 10:20-12:00 sudé týdny (první týden je sudý) 1. 21.2.2012 Klinická
Umělá inteligence. Příklady využití umělé inteligence : I. konstrukce adaptivních systémů pro řízení technologických procesů
 Umělá inteligence Pod pojmem umělá inteligence obvykle rozumíme snahu nahradit procesy realizované lidským myšlením pomocí prostředků automatizace a výpočetní techniky. Příklady využití umělé inteligence
Umělá inteligence Pod pojmem umělá inteligence obvykle rozumíme snahu nahradit procesy realizované lidským myšlením pomocí prostředků automatizace a výpočetní techniky. Příklady využití umělé inteligence
Efektivní velikost populace Wright (1931)
") Efektivní velikost populace Wright (1931) - velikost populace z genetického hlediska nemusí být rovna censu, někteří jedinci mohou zanechat potomků více, jiní se rozmnožování nezúčastní vůbec - to má dopad
Efektivní velikost populace Wright (1931) - velikost populace z genetického hlediska nemusí být rovna censu, někteří jedinci mohou zanechat potomků více, jiní se rozmnožování nezúčastní vůbec - to má dopad
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Euro a stabilizační role měnové politiky. 95. Žofínské fórum Euro s otazníky? V Česku v představách, na Slovensku realita Praha, 13.
 Euro a stabilizační role měnové politiky Zdeněk k TůmaT 95. Žofínské fórum Euro s otazníky? V Česku v představách, na Slovensku realita Praha, 13. listopadu 2008 Co nás spojuje a v čem se lišíme Režim
Euro a stabilizační role měnové politiky Zdeněk k TůmaT 95. Žofínské fórum Euro s otazníky? V Česku v představách, na Slovensku realita Praha, 13. listopadu 2008 Co nás spojuje a v čem se lišíme Režim
Typy selekce. Positive Balancing Background
 SELEKCE Typy selekce Positive Balancing Background Positive selection neutral positive Výhodná varianta Rychlé šíření Společný předek nedávno selective sweep Selective sweep, hitchhiking Detekce selekce
SELEKCE Typy selekce Positive Balancing Background Positive selection neutral positive Výhodná varianta Rychlé šíření Společný předek nedávno selective sweep Selective sweep, hitchhiking Detekce selekce
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
II. ročník, zimní semestr 1. týden OPAKOVÁNÍ. Úvod do POPULAČNÍ GENETIKY
 II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Atestace z lékařské genetiky inovované otázky pro rok A) Molekulární genetika
 Molekulární genetika") Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
Atestace z lékařské genetiky inovované otázky pro rok 2017 A) Molekulární genetika 1. Struktura lidského genu, nomenklatura genů, databáze týkající se klinického dopadu variace v jednotlivých genech. 2.
Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK
 ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
Počet chromosomů v buňkách. Genom
 Počet chromosomů v buňkách V každé buňce těla je stejný počet chromosomů. Výjimkou jsou buňky pohlavní, v nich je počet chromosomů poloviční. Spojením pohlavních buněk vzniká zárodečná buňka s celistvým
Počet chromosomů v buňkách V každé buňce těla je stejný počet chromosomů. Výjimkou jsou buňky pohlavní, v nich je počet chromosomů poloviční. Spojením pohlavních buněk vzniká zárodečná buňka s celistvým
Podobně, jako letadlo může řídit počítač, mohou nastat ale situace, kdy bez zásahu člověka by nového modelu je potřeba zásah člověka.
 1 Podobně, jako letadlo může řídit počítač, mohou nastat ale situace, kdy bez zásahu člověka by letadlo havarovalo. Stejně tak např. v roboti dokážou kompletně vyrobit nové auto, ale k vytvoření nového
1 Podobně, jako letadlo může řídit počítač, mohou nastat ale situace, kdy bez zásahu člověka by letadlo havarovalo. Stejně tak např. v roboti dokážou kompletně vyrobit nové auto, ale k vytvoření nového
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Molekulární podstata adaptací a detekce selekce na molekulární úrovni
 Molekulární podstata adaptací a detekce selekce na molekulární úrovni Adaptace Metody detekce selekce na molekulární úrovni Netřeba znát předem fenotyp. Lze detekovat i selekci, která působila v minulosti.
Molekulární podstata adaptací a detekce selekce na molekulární úrovni Adaptace Metody detekce selekce na molekulární úrovni Netřeba znát předem fenotyp. Lze detekovat i selekci, která působila v minulosti.