Co jsou to enzymy? pozoruhodné chemické katalyzátory
|
|
|
- Iva Müllerová
- před 8 lety
- Počet zobrazení:
Transkript
1 Enzymy
2 Co jsou to enzymy? pozoruhodné chemické katalyzátory Vyšší reakční rychlost (6-12 řádů) Mírnější podmínky reakce (nižší teplota, atmosférický tlak, neutrální ph) Vyšší specifita reakce (specifické produkty, nejsou vedlejší produkty) Schopnost regulace (allosterická regulace, kovalentní modifikace, variabilita) Chemické Homogenní reaktanty a katalyzátor jsou v systému přítomny ve stejné fázi. Heterogenní nejčastěji je katalyzátor pevný a reaktanty jsou plynné. Tento typ katalýzy se často používá v průmyslu (odpadá problém oddělování katalyzátoru z reakční směsi).
: Jde o molekuly, které do reakce vstupují Produkt (P): Jde o molekuly, které")
3 Enzymy Proteiny (mohou být také RNA molekuly např. Ribosom), které slouží jako biologické katalyzátory. V popisu chemické reakce s katalyzátorem používáme pojmy: S + E ES P + E Substrát (S): Jde o molekuly, které do reakce vstupují Produkt (P): Jde o molekuly, které vznikají katalyzovanou reakcí Enzym (E) Komplex enzymu se substrátem (SE)
4 Enzym jako katalyzátor Katalyzátor je látka, která zvyšuje efektivitu chemické reakce. Katalyzátor z reakce vystupuje nezměněn. Ale v jejím průběhu vytváří energeticky méně stabilní molekuly - meziprodukty. Díky meziproduktům je energie potřebná pro vznik produktů rozdělena do více částí (s menší energetickou náročností). Energie potřebná k průběhu reakce se nazývá aktivační energie. Snížení aktivační energie umožňuje reakci za mírnějších podmínek (tzn. za nižší teploty, tlaku). Enzymy snižují aktivační bariéru. To je umožněno díky vzniku meziproduktů, ve kterých jsou rozrušeny staré vazby a naopak vznikají nové. Meziprodukt je značen jako komplex enzymsubstrát (ES).
5 Enzym jako katalyzátor Enzymy se od chemických katalyzátoru liší: Efektivitou (enzymatické reakce bývají mnohem účinnější než chemické) Např. je enzym Orotinmonofosfát-dekarboxyláza (enzym odbourávající CO2 z molekul, důležitý pro syntézu nukleových kyselin. Reakce samotná by zabrala miliony let, s enzymem je to otázkou milisekund). (OMP prekurzor pyrimidinových nukleotidů. Dekarboxylací orotidinmonofosfátu) Podmínkami (podmínky enzymatické reakce jsou dány okolním prostředí v organizmu) Aktivita enzymu je silně závislá na ph a teplotě prostředí. Vysoká specificita (enzymy často rozeznávají pouze jednu specifickou molekulu) Nezávislost na množství produktu (Chemická katalýza je přímo závislá na množství substrátu, tzn. kolik substrátu do reakce dáme, tolik bude produktu. Enzymy jsou často regulovány a tak může dojít k situaci, že i za přítomnosti substrátu nedojde k reakci).

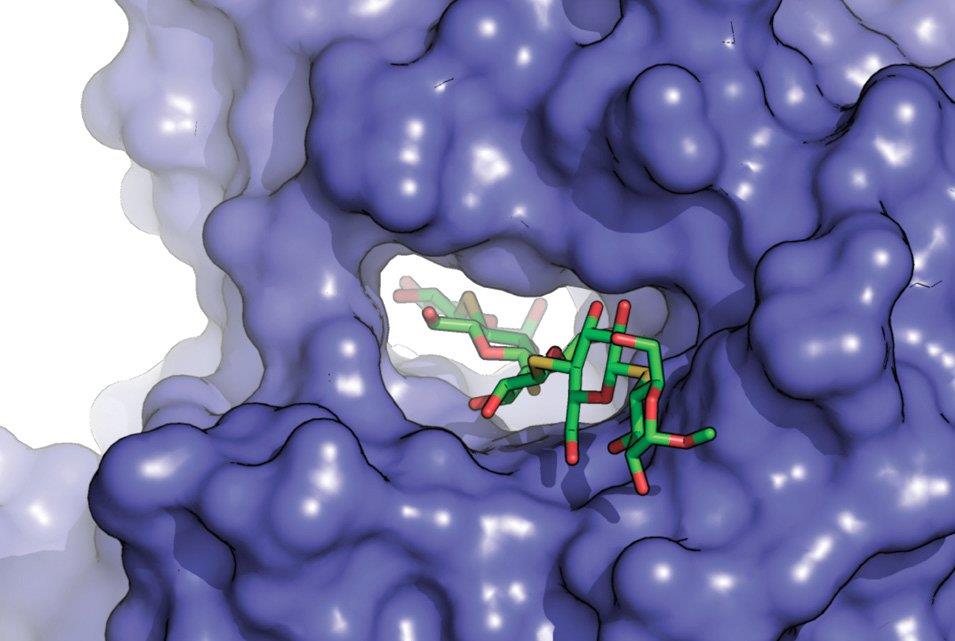
6 Enzymy - struktura Enzymy jsou většinou proteiny, které se mohou nacházet samostatně nebo fungují pouze v proteinových komplexech. Pro specificitu enzymové reakce je klíčová jeho 3D struktura. Správné uspořádání aminokyselin je důležité pro funkčnost tzv. aktivního místa. To lze rozdělit na katalytické a vazebné místo. Katalytické místo je oblast skládající se z několika aminokyselin, ve které dochází ke katalytické reakci. Katalytické místo je v proteinu obklopené vazebným místem. Vazebné místo funguje jako přistávací dráha pro substrát. Navíc prostorově substrát polohuje do správné pozice nutné pro katalýzu. Substrát a enzym musí být k sobě geometricky komplementární (jako zámek a klíč). Vazebné místo reguluje enzymovou aktivitu změnami afinity a polohování substrátu do katalytického místa.

7 Enzymy vazba substrátu Před enzymatickou reakcí je potřeba aby se do aktivního místa navázal substrát. Tomu napomáhá vazebné místo. Vazebné místo navíc zajišťuje specificitu enzymu - vazbu konkrétní molekuly, proteinu nebo skupiny proteinů. Specificita je zachována díky: Komplementárnímu tvaru vazebného místa a substrátu Rozložení elektrochemického náboje Hydrofobním nebo hydrofilním interakcím Model zámku a klíče Aktivní místo není strukturně rigidní a může se strukturně měnit po přijetí substrátu. V některých případech dochází navíc ke změně tvaru substrátu napomáhající katalytické reakci.
8 Enzymy aktivní místo Katalytická reakce může probíhat a) Stabilizací meziproduktu V místě proteinu díky blízkým AMK zbytkům dochází k výměně náboje s meziproduktem (který by byl velice nestabilní ve volném prostředí). b) Dočasnou interakcí se substrátem Enzym se stane příjemcem aktivní skupiny, umožňující snadnější přesun na jiné místo substrátu nebo jiný substrát c) Změna konformace molekuly Změnou struktury molekuly dochází také ke snížení energie potřebné pro vznik produktu.
9 Enzymy - kofaktory U některých enzymů se katalytické reakce neúčastí jenom zbytky aminokyselin, ale i jiné, pomocné molekuly - těmto molekulám říkáme kofaktory. Pokud je takový kofaktor vázán kovalentní vazbou k enzymu - nazýváme jej prostetickou skupinou. Pokud je vazba v katalytickém místě méně pevná jako v případě iontů kovů nebo organických molekul jedná se o koenzym. Mezi kofaktory patří například vitamíny rozpustné ve vodě (např. B1,B2, B6, B12, C atd.) Katalyticky aktivní dvojice kofaktoru a enzymu se nazývá holoenzym. Naopak enzym bez kofaktoru je apoenzym.
10 Enzymy - názvosloví Jak se v enzymech orientovat? Enzymy mají svoje specifické názvosloví. Název enzymu se skládá buďto z označení substrátu nebo z označení substrátu + typ dané katalytické reakce. Příponou názvu je - asa. Př. proteinasa, alkoholdehydrogenasa. V názvu tedy máme jak vstupní substrát, tak i typ reakce a můžeme usuzovat jaký produkt vznikne.
11 Pro podrobnější klasifikaci se využívá IUBMB (INTERNATIONAL UNION OF BIOCHEMISTRY AND MOLECULAR BIOLOGY) enzymová nomenklatura. V ní jsou enzymy rozděleny do 6 hlavních tříd podle typu reakce, kterou katalyzují. oxidoreduktasy (katalysují intermolekulární oxidačně-redukční reakce, např. ethanol + NAD+ acetaldehyd + NADH + H+); transferasy (přenos chemických skupin z molekuly donoru na akceptor, např. ATP + glukosa glukosa-6-fosfát + ADP); hydrolasy (štěpení vazeb vodou, např. sacharosa + H2O glukosa + fruktosa); lyasy (nejčastěji adice na dvojnou vazbu nebo eliminace za vzniku dvojné vazby, např. hydratace fumarátu v citrátovém cyklu -OOC-CH=CH-COO- + H2O -OOC-CH(OH)- CH2-COO-); isomerasy (isomerace, např. D-glukosa D-fruktosa nebo L-alanin D-alanin); ligasy (spojení dvou molekul, k němuž se dodává energii štěpením ATP nebo GTP, např. karboxylace pyruvátu na oxalacetát CH3-CO-COO- + CO2 + ATP + H2O - OOC-CH2-CO-COO- + ADP + Pi). např. EC znamená (EC) - Enzyme commision (1, hlavní třída) - oxidoreduktázy (v reakci dochází k výměně elektronů) (1, podtřída) - reakce probíhá na C-OH skupině (1, podskupina) - kofaktor - zde NAD nebo NADP (1, pořadové číslo) - určuje již konkrétní enzym - alkoholdehydrogenazu
12 Enzymová kinetika Enzymová kinetika je studium enzymatických reakcí. Studuje efektivitu a nejvhodnější podmínky pro enzymatickou reakci. Jaká je kinetika enzymatické reakce? Pro jeden substrát je rychlost enzymatické reakce definována rychlostí jakou se substrát váže na enzym a rychlostí jakou dochází k tvorbě produktu. Při nízké koncentraci substrátu vůči enzymu je reakce téměř přímo úměrná jeho koncentraci, protože nic nebrání katalýze. Druhým extrémem je vysoká koncentrace substrátu vůči enzymu. V takovém případě jsou aktivní místa enzymu zaplněny a nedochází k vazbě s přebytečným substrátem. V tomto případě je už celková rychlost reakce závislá na tom, jak rychle dokáže enzym zpracovat substrát na produkt - tím uvolní aktivní místo pro další substrát.
13 Dříve se podle pravidel Mezinárodní biochemické unie (IUB) z r užívalo pojmu enzymová jednotka (U) pro takové množství enzymu, které katalyzuje přeměnu 1 μmol substrátu za minutu za standardních podmínek (tj. nasycení enzymu substrátem, udaná teplota a ph). Podle doporučení IUB z r se však od pojmu množství enzymu upouští, protože tato definice se nehodí pro enzymy, které dosud nebyly izolovány. Namísto toho byly zavedeny pojmy: enzymová aktivita jako rychlost přeměny substrátu, která se dá přičíst na vrub enzymové katalýze; specifická aktivita jako enzymová aktivita vztažená za specifických podmínek ke hmotnosti (např. k určitému množství tkáně, séra, enzymového preparátu); molová aktivita jako enzymová aktivita vztažená za specifických podmínek k množství enzymové substance (byl-li enzym již izolován), které je zpravidla vyjádřeno v molech. Převod dosavadních enzymových jednotek (U) na kataly (kat) se řídí vztahem: 1 U = 1 μmol/min = 1/60 μmol/s = 1/60 μkat = 16,67 nkat Avšak ve starší literatuře se ještě s pojmem enzymové jednotky (U) setkáváme. U nás byly zavedeny kataly do klinicko-biochemické laboratorní praxe povinně a výsledky laboratorních vyšetřeni jsou takto vyjadřovány v séru nejčastěji jako μkat/l.

14 Graf ukazuje, že na začátku je rychlost reakce závislá na vazbě E se S, poté se enzym nasytí a poté přebírá důležitost krok přeměny ES na P.
15 Enzymová kinetika Enzymová kinetika je studium enzymatických reakcí. Studuje efektivitu a nejvhodnější podmínky pro enzymatickou reakci. Základní enzymatická reakce probíhá touto rovnicí: kde (E) je enzym, (S) substrát, (ES) komplex enzym-substrát, (P) produkt k 1, k 2 jsou rychlostní konstanty, ty popisují rychlost dané reakce. k -1 je rychlostní konstanta rozpadu komplexu ES. Rychlostní konstanta je rovna rychlosti reakce při jednotkových koncentracích výchozích látek. Za předpokladu, že žádný produkt neinteraguje zpětně s enzymem za vzniku ES, lze rychlost enzymatické reakce v (mol.s -1 ) definovat jako: V = k2 [ES]
16 Problém je, že ES je neměřitelný. Rychlost reakce se proto definuje podle koncentraci látek o kterých víme jejich koncentraci. Rozdělíme si vznik ES do dvou rovnic: a) vzniku produktu: [ES] = k1 [E] [S] b) rozpadu komplexu ES směrem k S nebo P: [ES] = (k-1 + k2) [ES] Za předpokladu, že je v reakci enzym plně nasycen tzn. Koncentrace ES je stále stejná: K1.[E].[S] = (k-1 + k2).[es] a dále
17 Koncentrace substrátu mezi nesaturovaným a saturovaným stavem enzymu je definována pomocí Michaelisovy konstanty K M konstanta poloviční saturace - ta se rovná koncentraci substrátu, při které došlo k polovičnímu nasycení enzymu substrátem a reakční rychlost a je rovna polovině maximální rychlosti. Z rovnice enzymatické katalýzy lze K M vyjádřit jako: Po dosazení do předchozí rovnice dostáváme:
18 Za předpokladu, že koncentrace E v reakci je mnohonásobně menší než S ([E] << [S]), je [ES] + [S] [S]. Koncentrace volného enzymu lze definovat jako rozdíl celkového enzymu a ES: [E] = [Eo] [ES] Po dosazení do rovnice dostáváme: Po vyjádření [ES] dostáváme: Víme už, že rychlost je definovaná jako: V = k2 [ES] Po dosazení dostaneme:
19 Je těžké experimentálně sledovat momentální rychlost enzymatické reakce. Proto se sleduje rychlost maximální Vmax. Vmax je definována rychlostí enzymatické reakce při plné saturaci enzymu (po ní je rychlost stabilní, lze ji pozorovat). Této rychlosti dosáhneme při [S] >>KM. Pak [S]/([S]+KM) 1 Vmax následně definujeme z předchozí rovnice: Následně dostáváme finální rovnici, ve které lze rychlost enzymatické rovnic získat měřením pozorovatelných veličin. Tato rovnice je známá jako Rovnice Michaelis-Mentenové:
20 Rovnice Michaelis-Mentenové Pro kinetiku enzymatické reakce s jedním substrátem. Tzn. Že do katalytického místa vstupuje pouze jedna molekula, která interaguje s enzymem. v 0 je počáteční rychlost enzymatické reakce, V max je limitní hodnota rychlosti při plném nasycení enzymu substrátem, [S] koncentrace substrátu, K M je Michaelisova konstanta. Pokud má enzym nízké K M, znamená to, že může dosahovat maximální rychlosti při nízké koncentraci substrátu. Enzymy s K M při M -1.s-1 prakticky každý kontakt se substrátem vede k přeměně na produkt.
21 Rovnice Michaelis-Mentenové Je třeba brát tuto rovnici pouze jako zjednodušenou. Nezohledňuje další faktory, které v reakci mohou nastat jako: inhibici reakce produktem zvratná reakce v meziproduktem nepopisuje allosterické chování proteinů (tedy že konformační změna proteinu může ovlivnit enzymatickou aktivitu) kooperativní interakce Navíc je platná za předpokladu, kdy: Koncentrace ES se mění mnohem pomaleji než koncentrace P a S. Koncentrace enzymu se v čase nemění.
22 Multisubstrátové enzymatické reakce Kinetika v takových reakcí navíc sleduje v jakém pořadí dochází k vazbě S na E. Bi-Bi reakce Oba substráty se vážou na enzym v jednom okamžiku a vyváří ternární EAB komplex. Může probíhat náhodným mechanismem - nezáleží na pořadí ve kterém se substráty navážou. V uspořádaném mechanismu je definováno a záleží na tom který substrát do aktivního místa přistupuje jako první, druhý atd. Bi-Bi mechanismus využívá například DNA Polymeráza.
23 Multisubstrátové enzymatické reakce Ping-Pong mechanismus enzymové reakce Enzym má dva stavy - základní a intermediární. V intermediárním stavu je enzym chemicky modifikován prvním substrátem. A - substrát umožňující přenos chemické skupina na enzym a po jejím vypuštění dochází k vazbě B, na který je chemické skupina z enzymu přenesena. Př. mohou být enzymy využívající kofaktory.
24 Kinetika allosterických enzymů allosterický charakterizovaný odlišným prostorovým sterickým uspořádáním Dochází ke kooperativní vazbě substrátu. Kinetika rozdílná oproti Michaelis-Mentenové. Struktura allosterických enzymů může být pozměněna vazbou allosterických efektorů do allosterického místa. Allosterické místo se nachází mimo aktivního místa. Efektory mohou změnit konformaci vazebného nebo aktivního místa enzymu vedoucí jak k aktivaci tak inhibici enzymatických procesů. Křivka závislosti koncentrace substrátu na rychlosti reakce je sigmoidní (jak známe např. z hemoglobinu). Kooperativní interakce jsou u enzymů běžné. Umožňuji regulovat jejich funkci na základě koncentrace substrátu nebo produktů. Například při pozitivní regulaci mohou být více citlivé na nízkou koncentraci S, v negativní naopak necitlivé.
25 Enzymová inhibice Inhibitory jsou molekuly, které redukují nebo blokují enzymatickou aktivitu. Inhibitor může bránit vazbě k aktivnímu místu enzymu nebo zabránit katalytické reakci. Naopak aktivátory enzymatickou aktivitu zvyšují. Inhibice může být vrátný nebo nevratný proces. Irreverzibilní inhibitory - vytváří kovalentní (pevnou) vazbu na inhibitor. Dochází k chemické modifikaci zbytků AMK potřebné pro enzymatickou aktivitu. Reverzibilní - vazba je nekovalentní (vodíkový můstek, hydrofobní interakce, iontová vazba). V kinetice inhibiční reakce sledujeme primárně specificitu inhibitoru a také jeho účinnost.
26 Inhibice enzymů působení mnoha léků je založeno na inhibici specifických enzymů metabolických drah tyto léky jsou analoga substrátů - mají podobnou strukturu jako substráty, ale nejsou enzymem přeměňovány (např. methotrexát, strukturní analog folátu /= koenzym/, soutěží s tímto koenzymem o vazbu do aktivního centra enzymu podílejícího se na syntéze thymidin monofosfátu pokles syntézy DNA vede k poklesu buněčného dělení nádorových buněk) Existují tři hlavní skupiny inhibitorů: 1) kompetitivní inhibitory strukturou podobné substrátu enzym nekatalyzuje přeměnu inhibitoru, inhibitor se pouze na enzym naváže váží se do vazebného místa substrátu (soutěží se substrátem o vazebné místo) zvýšením koncentrace substrátu lze inhibici potlačit Km se v přítomnosti inhibitoru zdánlivě zvyšuje V max se nemění 2) nekompetitivní inhibitory nebrání vazbě substrátu na enzym váží se mimo vazebné místo substrátu inhibici nelze potlačit zvýšením koncentrace substrátu, lze ji snížit pouze odstraněním inhibitoru dialýzou (inhibitor se neváže kovalentně) žádný z komplexů enzym-inhibitor (E-I ani E-I-S) není katalyticky aktivní snížení koncentrace aktivního enzymu _ pokles Vmax Km se nemění 3) akompetitivní inhibitory váží se pouze na komplex enzym-substrát dochází ke zdánlivé změně Km i Vmax
27 Neurotoxin je druh jedu, který působí na nervovou soustavu. Produkují ho jedové žlázy většiny korálovcovitých hadů žijících na souši (kobry, korálovci, mamby, ). Myotoxin je jed působící na svalovinu. V jedových žlázách ho mají všechny mořské druhy korálovcovitých hadů (vodnáři, vlnožilové) a některé suchozemské druhy korálovcovitých hadů žijících v Austrálii. Většina živočišných buněk má vysokou koncentraci K + uvnitř buněk a nízkou Na + ve vztahu k vnějšímu prostředí. Takový iontový gradient je generován specifickým transportním systémem, enzymem s názvem Na + - K + pumpa nebo také Na + - K + ATPasa. Hydrolýza ATP pumpou poskytuje energii potřebnou k aktivnímu transportu Na + ven z buňky a K + dovnitř, za účelem tvorby gradientu. Název Na + - K + ATPasa je používán proto, že k hydrolýze ATP dochází pouze za situace, kdy jsou oba ionty na pumpu vázány. Enzym vyžaduje přítomnost Mg 2+ iontů. Aktivní transport je má velmi významný fyziologický efekt. Na tvorbu gradientu se spotřebuje více než třetina vytvořeného ATP při odpočinku organismu. Gradient obou iontů kontroluje objem buňky, udržuje neurony a svalové buňky v elektricky excitovaném stavu a pohání aktivní transport sacharidů a aminokyselin.
28

29 Hemotoxin je druh jedu, který rozkládá krevní buňky uštknuté kořisti. Produkují ho jedové žlázy druhů hadů z čeledi zmijovitých (zmije, chřestýši, ploskolebci apod.).
30
31 Složky jedů ovlivňující hemokoagulaci Toxiny a enzymy ovlivňující funkci hemokoagulačního systému mají vliv na funkci krevních destiček i endotelu. Hemokoagulační aktivní toxiny mají na svědomí nejvíce fatálních případů intoxikace hadími jedy na světě. Klinicky se účinek projeví krvácením, avšak může dojít až k neztišenému krvácení z traumat, sliznic a do orgánů, smrt tedy končí poruchami typu diseminované intravaskulární koagulopatie. Tyto toxiny jsou charakteristické pro bojgy, zmije, korálovcovité a mořské hady. Hemostáza je ovlivněna zásahem toxinů do různých struktur hemokoagulačního systému. Toxiny mohou působit na tyto systémy: Krevní destičky Zásah toxinu je většinou do povrchních aktivačních struktur destiček. Enzymy typu fibrinogenáza, nukleotidáza, PLA2 mohou způsobit inhibici agregace krevních destiček. (Valenta 2008) Kontaktní cesta aktivace hemokoagulace Aktivace protrombinu Zvýšení trombinové aktivity je způsobena aktivací koagulačních faktorů, nebo přímým zásahem do molekuly protrombinu. Dochází ke krvácení typu diseminované intravaskulární koagulopatie. Další z typů enzymů, je enzym serinové protézy, účinkující se spoluúčastí fosfolipidů a Ca2+. Konvertující fibrinogen na fibrin Enzymy hadích jedů způsobují odštěpování fibrinopeptidů (fibrinopeptid A) z molekul fibrinogenu, a tak způsobují konverzi fibrinogenu na fibrin Degradující fibrinogen na fibrin Jedná se o enzymatické působení proteináz na fibrinogen nebo fibrin. Mohli bychom je také nazvat Fibrino(geno)lytické aktivity hadích jedů. Působením těchto látek vzniká zvýšené krvácení z ran a obraz hypofibrinogenémie nebo afibrinogenémie. Inaktivace antitrombinu Aktivace C proteinu
32 Reverzibilní Enzymové inhibice Kompetitivní reverzibilní inhibice Kde I je inhibitor. S a I se mohou vázat na enzym ve stejném čase. Inhibitor je často strukturálně velmi podobný substrátu, ale nedochází u něj k enzymatické reakci. I se tedy váže na vazebné (i aktivní) místo enzymu. O tyto místo tedy S a I soupeří.
33 Pokud zvýšíme koncentraci S, bude docházet k častějším interakcím mezi E a S a inhibiční účinek bude potlačen. Po inhibici kompetitivním inhibitorem dochází k zvýšení K M (Protože inhibitor obsazuje část molekul enzymu, pro vytvoření komplexu enzym-substrát je tedy k dispozici jen málo molekul volného enzymu. Z tohoto důvodu je reakce pomalá). V max se nemění.
34 Reverzibilní Enzymové inhibice Akompetivní reverzibilní inhibice Inhibitor se může vázat na jiné místo enzymu (a to pouze v případě, kdy je substrát vázaný na enzym). Po vazbě dochází k změně struktury enzymu vedoucí ke znemožnění enzymatické reakce. Zde se inhibitor váže pouze na ES. Pro inhibici tedy potřebuje přítomnost substrátu. S rostoucí koncentrací substrátu vede k zvýšené inhibice enzymu. Proto dochází ke snížení V max. Tím se snižuje se také i K M.
35 Reverzibilní Enzymové inhibice Smíšená reverzibilní inhibice Inhibitor se váže na aktivní místo enzymu nezávisle na tom, zdali je aktivní místo prázdné nebo je S v komplexu s enzymem. Nicméně má větší afinitu k jednomu nebo druhému stavu. Jedná se o mix kompetitivní a akompetitivní inhibice. Vazba inhibitoru ovlivňuje vazbu S (zvyšuje se K M ) a zároveň snižuje počet aktivních molekul enzymu (snížení V max ). Inhibiční efekt může být redukován, ale ne potlačen zvýšenou koncentrací S.
36 Reverzibilní Enzymové inhibice Nekompetivní reverzibilní inhibice Specifický případ smíšené enzymové inhibice. Inhibitor redukuje afinitu enzymu k substrátu, ale neovlivňuje jeho vazbu (S se tedy na enzym neváže). Inhibitor se podobně jako u akompetitivní inhibice váže na jiné místo proteinu a vyvolává strukturální změny v aktivním místě enzymu.
37 Tento typ inhibice je přímo závislý na koncentraci I, ne na koncentraci S (protože se Inhibitor váže na jiné místo enzymu a nesoupeří se substrátem). Takže když dojde k inhibici 50% enzymu, tak jak při nízké i vysoké koncentraci substrátu. Klesá V max, protože aktivita enzymu už je redukovaná a vysoká koncentrace S na ni nemá efekt. Ale K M zůstává stejná (mechanismus enzymatické reakce je stejný, ale s omezeným množstvím aktivních enzymů).
38 Vliv teploty na enzymatickou reakci Enzymy mohou být stabilní mezi teplotou 0 až 100 C (termofilní bakterie mají např. velmi stabilní enzymy, které jsou aktivní i při 110 C). V lidském těle jsou enzymy konstruovány na teplotu okolo 37 C. Enzymy jsou aktivní pouze v malém teplotním intervalu. Teplota, při kterém je enzym nejvíce aktivní - Teplotní optimim. To je umožněno ideální stabilitou proteinu a správnému uspořádání AMK v aktivním místě.
39 Vliv teploty na enzymatickou reakci Závislost reakce na teplotě je v oblasti teplotního optima definováno Arheniovou rovnicí, která definuje závislost jakékoliv chemické reakce na teplotě. S rostoucí teplotou získávají molekuly (S i E) větší kinetickou energii a narůstá pravděpodobnost ideální srážky mezi S a E. Při nízkých teplotách je sice enzym stabilní a není ovlivněna struktura proteinu, ale počet srážek je omezený. Při vyšších teplotách pak dochází k částečné nebo úplné denaturaci (rozpadu 3D struktury a tvorbě neuspořádaných proteinových klubek) a degradaci proteinu.
40 Vliv ph na enzymatickou reakci ph optimum závisí na funkci a prostředí ve kterém se protein vyskytuje. ph 7 - trypsin, enzymy v krvi ph 2 - pepsin ph 10 Arginasa Záleží na AMK zbytcích v aktivním místě, protože katalytická aktivita enzymů se odvíjí od toho zda jsou AMK ionizované. Také dochází k narušení elektrochemické interakce mezi vazebným místem a substrátem. Vysoké a nízké ph denaturuje proteinovou strukturu.

41 Enzymy Ing. Lívia Eiselleová, PhD.
, které slouží jako biologické katalyzátory Substrát (S) -")
 Komplex enzymu se")
42 Enzymy Proteiny (mohou být také RNA molekuly např. ribosom), které slouží jako biologické katalyzátory Substrát (S) - molekuly, které do reakce vstupují Produkt (P) - molekuly, které vznikají katalyzovanou reakcí Enzym (E) Komplex enzymu se substrátem (ES) E+S ES E+P
43 Na co slouží enzymy - vyšší reakční rychlost (6-12 řádů) - mírnější podmínky reakce (nižší teplota, atmosférický tlak, neutrální ph) - vyšší specifita reakce (specifické produkty, nejsou vedlejší produkty) - schopnost regulace (allosterická regulace, kovalentní modifikace, variabilita) Mohou být: - homogenní reaktanty a katalyzátor jsou v systému přítomny ve stejné fázi - heterogenní nejčastěji je katalyzátor pevný a reaktanty jsou plynné

44 Jaké známe enzymy

45 Replikační enzymy - podílejí sa na replikaci DNA

46 Restrikční enzymy - sekvenčne specifické endonukleázy
. Enzymy snižují aktivační bariéru - díky vzniku meziproduktů")
47 Enzym jako katalyzátor Katalyzátor - zvyšuje efektivitu chemické reakce - z reakce vystupuje nezměněn - v průběhu reakce vytváří energeticky méně stabilní molekuly meziprodukty Aktivační energie - energie potřebná k průběhu reakce - snížení aktivační energie umožňuje reakci za mírnějších podmínek (tzn. za nižší teploty, tlaku). Enzymy snižují aktivační bariéru - díky vzniku meziproduktů
 - podmínky (ph, teplota) - vysoká")

48 Enzymy se od chemických katalyzátorů liší - efektivita (enzymatické reakce bývají mnohem účinnější než chemické) - podmínky (ph, teplota) - vysoká specificita (enzymy často rozeznávají pouze jednu specifickou molekulu) - nezávislost na množství produktu (chemická katalýza je přímo závislá na množství substrátu)

49 Struktura enzymů - proteiny, které se mohou nacházet samostatně nebo v proteinových komplexech - pro specificitu enzymové reakce je klíčová jeho 3D struktura - správné uspořádání aminokyselin je důležité pro funkčnost tzv. aktivního místa Aktivní místo: Katalytické místo - oblast skládající se z několika aminokyselin, ve které dochází ke katalytické reakci Vazebné místo - funguje jako přistávací dráha pro substrát - prostorově substrát polohuje do správné pozice nutné pro katalýzu - reguluje enzymovou aktivitu změnami afinity a polohování substrátu do katalytického místa

50 Vazba substrátu - do aktivního místa (zajišťuje vazebné místo) - vazebné místo navíc zajišťuje specificitu enzymu Specificita je zachována díky: - komplementárnímu tvaru vazebného místa a substrátu - rozložení elektrochemického náboje - hydrofobním nebo hydrofilním interakcím Substrát a enzym musí být k sobě geometricky komplementární (model zámku a klíče)
51 Aktivní místo enzymu Katalytická reakce může probíhat a) stabilizací meziproduktu - v místě proteinu díky blízkým AMK zbytkům dochází k výměně náboje s meziproduktem (ve stejném prostředí by byl nestabilní) b) dočasnou interakcí se substrátem - enzym se stane příjemcem aktivní skupiny, umožňující snadnější přesun na jiné místo substrátu nebo jiný substrát c) změna konformace molekuly - změnou struktury molekuly dochází také ke snížení energie potřebné pro vznik produktu
 - pokud je kofaktor vázán kovalentní vazbou k enzymu - prostetická skupina - pokud je vazba v katalytickém místě méně pevná jako v případě iontů kovů nebo organických molekul jedná se o koenzym")
52 Enzymy - kofaktory - katalytické reakce se neúčastí jenom zbytky aminokyselin, ale i jiné molekuly kofaktory (např. vitamíny rozpustné v tucích, např. B1,B2, B6, B12, C atd.) - pokud je kofaktor vázán kovalentní vazbou k enzymu - prostetická skupina - pokud je vazba v katalytickém místě méně pevná jako v případě iontů kovů nebo organických molekul jedná se o koenzym Holoenzym - katalyticky aktivní dvojice kofaktoru a enzymu Apoenzym - enzym bez kofaktoru

53 Nazvosloví enzymů - enzymy mají svoje specifické názvosloví - název enzymu se skládá buďto z označení substrátu nebo z označení substrátu + typ dané katalytické reakce - příponou názvu je asa - např. proteinasa, alkoholdehydrogenasa, laktátdehydrogenasa
54 Klasifikace enzymů - podrobnější klasifikaci IUBMB (INTERNATIONAL UNION OF BIOCHEMISTRY AND MOLECULAR BIOLOGY) oxidoreduktasy (katalysují intermolekulární oxidačně-redukční reakce, např. ethanol + NAD+ acetaldehyd + NADH + H+) transferasy (přenos chemických skupin z molekuly donoru na akceptor, např. ATP + glukosa glukosa-6-fosfát + ADP) hydrolasy (štěpení vazeb vodou, např. sacharosa + H2O glukosa + fruktosa); lyasy (nejčastěji adice na dvojnou vazbu nebo eliminace za vzniku dvojné vazby, např. hydratace fumarátu v citrátovém cyklu -OOC-CH=CH-COO- + H2O -OOC-CH(OH)-CH2- COO-) isomerasy (isomerace, např. D-glukosa D-fruktosa nebo L-alanin D-alanin); ligasy (spojení dvou molekul, k němuž se dodává energii štěpením ATP nebo GTP, např. karboxylace pyruvátu na oxalacetát CH3-CO-COO- + CO2 + ATP + H2O -OOC-CH2-CO- COO- + ADP + Pi) např. EC znamená (EC) - Enzyme commision (1, hlavní třída) - oxidoreduktázy (v reakci dochází k výměně elektronů) (1, podtřída) - reakce probíhá na C-OH skupině (1, podskupina) - kofaktor - zde NAD nebo NADP (1, pořadové číslo) - určuje již konkrétní enzym - alkoholdehydrogenazu
55 Kinetika enzymů - studuje efektivitu a nejvhodnější podmínky pro enzymatickou reakci - rychlost enzymatické reakce definována rychlostí jakou se substrát váže na enzym a rychlostí jakou dochází k tvorbě produktu nízká koncentraci substrátu vůči enzymu - reakce téměř přímo úměrná jeho koncentraci vysoká koncentrace substrátu vůči enzymu - aktivní místa enzymu jsou zaplněny a nedochází k vazbě s přebytečným substrátem - celková rychlost reakce závislá na tom, jak rychle dokáže enzym zpracovat substrát na produkt
56 Kinetika enzymů - jednotky Dříve se podle pravidel Mezinárodní biochemické unie (IUB) z r užívalo pojmu enzymová jednotka (U) pro takové množství enzymu, které katalyzuje přeměnu 1 μmol substrátu za minutu za standardních podmínek Namísto toho byly zavedeny pojmy: enzymová aktivita - rychlost přeměny substrátu specifická aktivita - enzymová aktivita vztažená za specifických podmínek ke hmotnosti molová aktivita - enzymová aktivita vztažená za specifických podmínek k množství enzymovu vyjádřeno v molech Převod dosavadních enzymových jednotek (U) na kataly (kat) se řídí vztahem: 1 U = 1 μmol/min = 1/60 μmol/s = 1/60 μkat = 16,67 nkat U nás byly zavedeny kataly do klinicko-biochemické laboratorní praxe povinně a výsledky laboratorních vyšetřeni jsou takto vyjadřovány v séru nejčastěji jako μkat/l.
57 Enzymová kinetika - studium enzymatických reakcí: efektivita a nejvhodnější podmínky - základní enzymatická reakce se dá vyjádřit touto rovnicí: k1, k2 rychlostní konstanty, popisujú rychlost dané reakce k-1 rychlostní konstanta rozpadu komplexu ES Rychlostní konstanta je rovna rychlosti reakce při jednotkových koncentracích látek Za předpokladu, že žádny produkt neinteraguje zpětně s enzymem za vzniku ES, lze rychlost enzymatické reakce V (mol*s-1) definovat jako: V = k2 [ES]
 vzniku produktu: [ES] = k1 [E] [S] b) rozpadu komplexu ES směrem k S nebo P: [ES] = (k-1 + k2) [ES] Za předpokladu, že je v reakci enzym plně nasycen tzn.")
58 Enzymová kinetika Problém je, že ES je neměřitelný. Rychlost reakce se proto definuje podle koncentraci látek o kterých víme jejich koncentraci. Rozdělíme si vznik ES do dvou rovnic: a) vzniku produktu: [ES] = k1 [E] [S] b) rozpadu komplexu ES směrem k S nebo P: [ES] = (k-1 + k2) [ES] Za předpokladu, že je v reakci enzym plně nasycen tzn. koncentrace ES je stále stejná: K1.[E].[S] = (k-1 + k2).[es] a dále
59 Kinetika enzymů Michaelisova konstanta Koncentrace substrátu mezi nesaturovaným a saturovaným stavem enzymu je definována pomocí Michaelisovy konstanty K M - konstanta poloviční saturace - rovná se koncentraci substrátu, při které došlo k polovičnímu nasycení enzymu substrátem - reakční rychlost a je rovna polovině maximální rychlosti Z rovnice enzymatické katalýzy lze K M vyjádřit jako: Po dosazení do předchozí rovnice dostáváme:
60 Kinetika enzymů odvození rovnic Za předpokladu, že koncentrace E v reakci je mnohonásobně menší než S ([E] << [S]), je [ES] + [S] [S]. Koncentrace volného enzymu lze definovat jako rozdíl celkového enzymu a ES: [E] = [Eo] [ES] Po dosazení do rovnice dostáváme: Po vyjádření [ES] dostáváme: Víme už, že rychlost je definovaná jako: V = k2 [ES] Po dosazení dostaneme:
61 Kinetika enzymů odvození rovnic - experimentálně sledovat momentální rychlost enzymatické reakce je ťežké - proto se sleduje rychlost maximální Vmax - Vmax je definována rychlostí enzymatické reakce při plné saturaci enzymu (po ní je rychlost stabilní, lze ji pozorovat). Této rychlosti dosáhneme při [S] >>KM. Pak [S]/([S]+KM) 1 Vmax následně definujeme z předchozí rovnice: Následně dostáváme finální rovnici, ve které lze rychlost enzymatické rovnic získat měřením pozorovatelných veličin: rovnice Michaelis-Mentenové:
62 Rovnice Michaelis-Mentenové - platí pro kinetiku enzymatické reakce s jedním substrátem (do katalytického místa vstupuje ouze jedna molekula, která interaguje s enzymem) v 0 je počáteční rychlost enzymatické reakce V max je limitní hodnota rychlosti při plném nasycení enzymu substrátem [S] koncentrace substrátu K M je Michaelisova konstanta Pokud má enzym nízké K M, znamená to, že může dosahovat maximální rychlosti při nízké koncentraci substrátu Enzymy s K M při M -1.s-1 prakticky každý kontakt se substrátem vede k přeměně na produkt.
63 Rovnice Michaelis-Mentenové Nezohledňuje další faktory, které v reakci mohou nastat: - inhibice reakce produktem - zvratná reakce v meziproduktem - nepopisuje allosterické chování proteinů (tedy že konformační změna proteinu může ovlivnit enzymatickou aktivitu) - kooperativní interakce Navíc je platná za předpokladu, kdy: - koncentrace ES se mění mnohem pomaleji než koncentrace P a S - koncentrace enzymu se v čase nemění.
64 Multisubstrátové enzymatické reakce Kinetika v takových reakcí navíc sleduje v jakém pořadí dochází k vazbě S na E Bi-Bi reakce - oba substráty se vážou na enzym v jednom okamžiku a vyváří ternární EAB komplex - může probíhat náhodným mechanismem - nezáleží na pořadí ve kterém se substráty navážou Bi-Bi mechanismus využívá například DNA Polymeráza
65 Multisubstrátové enzymatické reakce Ping-Pong mechanismus enzymové reakce: Enzym má dva stavy: základní a intermediární (chemicky modifikován prvním substrátem) A - substrát umožňující přenos chemické skupina na enzym a po jejím vypuštění dochází k vazbě B, na který je chemické skupina z enzymu přenesena Př. mohou být enzymy využívající kofaktory
66 Kinetika allosterických enzymů - allosterický charakterizovaný odlišným prostorovým sterickým uspořádáním - dochází ke kooperativní vazbě substrátu - kinetika rozdílná oproti Michaelis-Mentenové - struktura allosterických enzymů může být pozměněna vazbou allosterických efektorů do allosterického místa (allosterické místo se nachází mimo aktivního místa) - efektory mohou změnit konformaci vazebného nebo aktivního místa enzymu vedoucí jak k aktivaci tak inhibici enzymatických procesů
 V")
67 Inhibitory enzymů Inhibitory - molekuly, které redukují nebo blokují enzymatickou aktivitu Aktivátory - enzymatickou aktivitu zvyšují Irreverzibilní inhibitory - vytváří kovalentní (pevnou) vazbu na inhibitor - dochází k chemické modifikaci zbytků AMK potřebné pro enzymatickou aktivitu Reverzibilní inhibitory - vazba je nekovalentní (vodíkový můstek, hydrofobní interakce, iontová vazba) V kinetice inhibiční reakce sledujeme primárně specificitu inhibitoru a také jeho účinnost.
 nekompetitivní inhibitory nebrání vazbě substrátu na enzym váží se mimo vazebné místo")
68 Inhibice enzymů 1) kompetitivní inhibitory strukturou podobné substrátu enzym nekatalyzuje přeměnu inhibitoru, inhibitor se pouze na enzym naváže váží se do vazebného místa substrátu zvýšením koncentrace substrátu lze inhibici potlačit Km se v přítomnosti inhibitoru zdánlivě zvyšuje V max se nemění 2) nekompetitivní inhibitory nebrání vazbě substrátu na enzym váží se mimo vazebné místo substrátu inhibici nelze potlačit zvýšením koncentrace substrátu, lze ji snížit pouze odstraněním inhibitoru dialýzou (inhibitor se neváže kovalentně) žádný z komplexů enzym-inhibitor (E-I ani E-I-S) není katalyticky aktivní snížení koncentrace aktivního enzymu _ pokles Vmax Km se nemění 3) akompetitivní inhibitory váží se pouze na komplex enzym-substrát dochází ke zdánlivé změně Km i Vmax
69 Reverzibilní enzymové inhibice Kompetitivní reverzibilní inhibice - S a I se mohou vázat na enzym ve stejném čase - inhibitor je často strukturálně velmi podobný substrátu, ale nedochází u něj k enzymatické reakci - I se tedy váže na vazebné (i aktivní) místo enzymu. O tyto místo S a I soupeří (kompetice)
70 - pokud zvýšíme koncentraci S, bude docházet k častějším interakcím mezi E a S - inhibiční účinek bude potlačen - dochází k zvýšení K M (inhibitor obsazuje část molekul enzymu, pro vytvoření komplexu enzym-substrát je tedy k dispozici jen málo molekul volného enzymu reakce je pomalá) - V max se nemění
71 Reverzibilní enzymové inhibice Akompetivní reverzibilní inhibice - inhibitor se může vázat na jiné místo enzymu (a to pouze v případě, kdy je substrát vázaný na enzym) - po vazbě dochází k změně struktury enzymu vedoucí ke znemožnění enzymatické reakce - inhibitor se váže pouze na ES - pro inhibici tedy potřebuje přítomnost substrátu - s rostoucí koncentrací substrátu vede k zvýšené inhibice enzymu - dochází ke snížení V max, tím se snižuje se také i K M.
72 Reverzibilní enzymové inhibice Smíšená reverzibilní inhibice - inhibitor se váže na aktivní místo enzymu nezávisle na tom, zdali je aktivní místo prázdné nebo je S v komplexu s enzymem - má větší afinitu k jednomu nebo druhému stav - mix kompetitivní a akompetitivní inhibice - vazba inhibitoru ovlivňuje vazbu S (zvyšuje se K M ) a zároveň snižuje počet aktivních molekul enzymu (snížení V max ) - inhibiční efekt může být redukován, ale ne potlačen zvýšenou koncentrací S
73 Reverzibilní enzymové inhibice Nekompetitivní reverzibilní inhibice - specifický případ smíšené enzymové inhibice - inhibitor redukuje afinitu enzymu k substrátu, ale neovlivňuje jeho vazbu (S se tedy na enzym neváže) - podobně jako u akompetitivní inhibice váže na jiné místo proteinu a vyvolává strukturální změny v aktivním místě enzymu.
74 - inhibice je přímo závislá na koncentraci I, ne na koncentraci S - když dojde k inhibici 50% enzymu, tak jak při nízké i vysoké koncentraci substrátu - V max klesá - aktivita enzymu už je redukovaná a vysoká koncentrace S na ni nemá efekt - K M zůstává stejná (mechanismus enzymatické reakce je stejný, ale s omezeným množstvím aktivních enzymů)

75
76
77 Vliv teploty na enzymatickou reakci - enzymy mohou být stabilní mezi teplotou 0 až 100 C - termofilní bakterie mají např. velmi stabilní enzymy, které jsou aktivní i při 110 C - v lidském těle jsou enzymy konstruovány na teplotu okolo 37 C - aktivní pouze v malém teplotním intervalu - teplota, při kterém je enzym nejvíce aktivní - teplotní optimum
78 Vliv teploty na enzymatickou reakci - závislost reakce na teplotě je v oblasti teplotního optima definováno Arheniovou rovnicí - s rostoucí teplotou získávají molekuly (S i E) větší kinetickou energii a narůstá pravděpodobnost ideální srážky mezi S a E - při nízkých teplotách je sice enzym stabilní a není ovlivněna struktura proteinu, ale počet srážek je omezený - při vyšších teplotách pak dochází k částečné nebo úplné denaturaci (rozpadu 3D struktury a tvorbě neuspořádaných proteinových klubek) a degradaci proteinu


79 Vliv ph na enzymatickou reakci - ph optimum závisí na funkci a prostředí, ve kterém se protein vyskytuje - záleží na AMK zbytcích v aktivním místě - katalytická aktivita enzymů se odvíjí od toho zda jsou AMK ionizované - vysoké a nízké ph denaturuje proteinovou strukturu ph 7 trypsin, enzymy v krvi ph2 pepsin ph10 -arginasa
80 Děkuji za pozornost Lívia Eiselleová
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Enzymologie. Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol Matej Kohutiar. akad. rok 2017/2018
 Enzymologie Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol Matej Kohutiar akad. rok 2017/2018 Osnova I. Základní principy enzymových reakcí II. Termodynamické a kinetické aspekty enzymové
Enzymologie Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol Matej Kohutiar akad. rok 2017/2018 Osnova I. Základní principy enzymových reakcí II. Termodynamické a kinetické aspekty enzymové
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. ENZYMY I úvod, názvosloví, rozdělení do tříd
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ENZYMY I úvod, názvosloví, rozdělení do tříd Úvod z řeckého EN ZYME (v kvasinkách) biologický katalyzátor, protein (RNA) liší se od chemických
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ENZYMY I úvod, názvosloví, rozdělení do tříd Úvod z řeckého EN ZYME (v kvasinkách) biologický katalyzátor, protein (RNA) liší se od chemických
Enzymologie. Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů.
 ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
Redoxní děj v neživých a živých soustavách
 Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Enzymy. aneb. Není umění dělat co tě baví, ale najít zalíbení v tom, co udělati musíš. Luboš Paznocht
 Enzymy aneb Není umění dělat co tě baví, ale najít zalíbení v tom, co udělati musíš. Luboš Paznocht Umožňují rychlý a koordinovaný průběh chemických přeměn v organismu Kinetika biochemických reakcí řád
Enzymy aneb Není umění dělat co tě baví, ale najít zalíbení v tom, co udělati musíš. Luboš Paznocht Umožňují rychlý a koordinovaný průběh chemických přeměn v organismu Kinetika biochemických reakcí řád
Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY
 Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
ENZYMY. Klasifikace enzymů
 ENZYMY Enzymy jsou bílkoviny, které katalyzují chemické reakce probíhající v živých organismech. Byly identifikovány tisíce enzymů, mnohé z nich byly izolovány čisté. Klasifikace enzymů Vzhledem k tomu,
ENZYMY Enzymy jsou bílkoviny, které katalyzují chemické reakce probíhající v živých organismech. Byly identifikovány tisíce enzymů, mnohé z nich byly izolovány čisté. Klasifikace enzymů Vzhledem k tomu,
>>> E A1 + E A2. . aktivační energie potřebná k reakci bez přítomnosti katalyzátoru E A E A1. energie potřebná ke vzniku enzym-substrátového komplexu
 Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Enzymy faktory ovlivňující jejich účinek
 Enzymy faktory ovlivňující jejich účinek Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek enzymy 10.8.2012 3. ročník čtyřletého G Faktory ovlivňující
Enzymy faktory ovlivňující jejich účinek Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek enzymy 10.8.2012 3. ročník čtyřletého G Faktory ovlivňující
Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996
 Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996 Šablona: III/2 č. materiálu: VY_32_INOVACE_CHE_419 Jméno autora: Třída/ročník: Mgr. Alena
Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996 Šablona: III/2 č. materiálu: VY_32_INOVACE_CHE_419 Jméno autora: Třída/ročník: Mgr. Alena
Aminokyseliny, proteiny, enzymologie
 Aminokyseliny, proteiny, enzymologie Aminokyseliny Co to je? Organické látky karboxylové kyseliny, které mají na sousedním uhlíku navázanou aminoskupinu Jak to vypadá? K čemu je to dobré? AK jsou stavební
Aminokyseliny, proteiny, enzymologie Aminokyseliny Co to je? Organické látky karboxylové kyseliny, které mají na sousedním uhlíku navázanou aminoskupinu Jak to vypadá? K čemu je to dobré? AK jsou stavební
Reakční kinetika. Nauka zabývající se rychlostí chemických reakcí a ovlivněním rychlosti těchto reakcí
 Nauka zabývající se rychlostí chemických reakcí a ovlivněním rychlosti těchto reakcí Vymezení pojmů : chemická reakce je děj, při kterém zanikají výchozí látky a vznikají látky nové reakční mechanismus
Nauka zabývající se rychlostí chemických reakcí a ovlivněním rychlosti těchto reakcí Vymezení pojmů : chemická reakce je děj, při kterém zanikají výchozí látky a vznikají látky nové reakční mechanismus
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
POLYPEPTIDY. Provitaminy = organické sloučeniny bez vitaminózního účinku, které se v živočišném těle mění působením ÚV záření nebo enzymů na vitaminy.
 POLYPEPTIDY Provitaminy = organické sloučeniny bez vitaminózního účinku, které se v živočišném těle mění působením ÚV záření nebo enzymů na vitaminy. Hormony = katalyzátory v živočišných organismech (jsou
POLYPEPTIDY Provitaminy = organické sloučeniny bez vitaminózního účinku, které se v živočišném těle mění působením ÚV záření nebo enzymů na vitaminy. Hormony = katalyzátory v živočišných organismech (jsou
Proč biokatalýza? Vyšší reakční rychlost Vyšší specificita reakce Mírnější reakční podmínky Možnost regulace
 Enzymy Proč biokatalýza? Vyšší reakční rychlost Vyšší specificita reakce Mírnější reakční podmínky Možnost regulace COO - - COO NH 2 OH - COO NH 2 - COO O OH - COO Chorismate mutase - OOC O OH - COO -
Enzymy Proč biokatalýza? Vyšší reakční rychlost Vyšší specificita reakce Mírnější reakční podmínky Možnost regulace COO - - COO NH 2 OH - COO NH 2 - COO O OH - COO Chorismate mutase - OOC O OH - COO -
Kinetika enzymově katalysovaných reakcí
 Kinetika enzymově katalysovaných reakcí Rychlost reakce aa + bb + c C + d D +... dn A d [ A] d [ B ] d [C ] v= = = = av d τ ad τ bd τ cd τ Počáteční rychlost reakce aa + bb + konc. c C + d D +... d [ A]
Kinetika enzymově katalysovaných reakcí Rychlost reakce aa + bb + c C + d D +... dn A d [ A] d [ B ] d [C ] v= = = = av d τ ad τ bd τ cd τ Počáteční rychlost reakce aa + bb + konc. c C + d D +... d [ A]
HISTORIE ENZYMOLOGIE
 ENZYMY HISTORIE ENZYMOLOGIE 1. Berzelius (18.stol.) v rostlinách i živočiších probíhají tisíce katalyzovaných reakcí FERMENTY fermentace (Fabrony) 2. W.Kühne en zýme = v kvasnicích enzymy 3. J. Sumner
ENZYMY HISTORIE ENZYMOLOGIE 1. Berzelius (18.stol.) v rostlinách i živočiších probíhají tisíce katalyzovaných reakcí FERMENTY fermentace (Fabrony) 2. W.Kühne en zýme = v kvasnicích enzymy 3. J. Sumner
Reakční kinetika enzymových reakcí
 Reakční kinetika enzymových reakcí studuje časový průběh enzymových reakcí za různých reakčních podmínek zabývá se faktory, které ovlivňují rychlost reakcí katalyzovaných enzymy - uvažujme monomolekulární
Reakční kinetika enzymových reakcí studuje časový průběh enzymových reakcí za různých reakčních podmínek zabývá se faktory, které ovlivňují rychlost reakcí katalyzovaných enzymy - uvažujme monomolekulární
ENZYMY A NUKLEOVÉ KYSELINY
 ENZYMY A NUKLEOVÉ KYSELINY Autor: Mgr. Stanislava Bubíková Datum (období) tvorby: 28. 3. 2013 Ročník: devátý Vzdělávací oblast: Člověk a příroda / Chemie / Organické sloučeniny 1 Anotace: Žáci se seznámí
ENZYMY A NUKLEOVÉ KYSELINY Autor: Mgr. Stanislava Bubíková Datum (období) tvorby: 28. 3. 2013 Ročník: devátý Vzdělávací oblast: Člověk a příroda / Chemie / Organické sloučeniny 1 Anotace: Žáci se seznámí
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Enzymy biokatalyzátory (6). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Enzymy biokatalyzátory (6). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
Chemická kinetika. Chemické změny probíhající na úrovni atomárně molekulové nazýváme reakční mechanismus.
 Chemická kinetika Chemická reakce: děj mezi jednotlivými atomy a molekulami, při kterých zanikají některé vazby v molekulách výchozích látek a jsou nahrazovány vazbami v molekulách nově vznikajících látek.
Chemická kinetika Chemická reakce: děj mezi jednotlivými atomy a molekulami, při kterých zanikají některé vazby v molekulách výchozích látek a jsou nahrazovány vazbami v molekulách nově vznikajících látek.
Enzymy charakteristika a katalytický účinek
 Enzymy charakteristika a katalytický účinek Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek enzymy 28.7.2012 3. ročník čtyřletého G Charakteristika
Enzymy charakteristika a katalytický účinek Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek enzymy 28.7.2012 3. ročník čtyřletého G Charakteristika
9. Chemické reakce Kinetika
 Základní pojmy Kinetické rovnice pro celistvé řády Katalýza Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti reakční mechanismus elementární reakce a molekularita reakce reakční rychlost
Základní pojmy Kinetické rovnice pro celistvé řády Katalýza Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti reakční mechanismus elementární reakce a molekularita reakce reakční rychlost
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV
 CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV a) Chemické složení a. biogenní prvky makrobiogenní nad 0,OO5% (C, O, N, H, S, P, Ca.) - mikrobiogenní pod 0,005%(Fe,Zn, Cu, Si ) b. voda 60 90% každého organismu - 90% příjem
CHEMICKÉ ZNAKY ŽIVÝCH SOUSTAV a) Chemické složení a. biogenní prvky makrobiogenní nad 0,OO5% (C, O, N, H, S, P, Ca.) - mikrobiogenní pod 0,005%(Fe,Zn, Cu, Si ) b. voda 60 90% každého organismu - 90% příjem
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
Sacharidy a polysacharidy (struktura a metabolismus)
") Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno:
 Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
Enzymy biologické katalyzátory. regulovatelnost účinnosti (aktivity) Platí o nich totéž co o chemických katalyzátorech, ale mají něco navíc:
 Platí o nich totéž co o chemických katalyzátorech, ale mají něco navíc:") Enzymy biologické katalyzátory Platí o nich totéž co o chemických katalyzátorech, ale mají něco navíc: účinné snížení aktivační energie specifita regulovatelnost účinnosti (aktivity) Regulace účinnosti
Enzymy biologické katalyzátory Platí o nich totéž co o chemických katalyzátorech, ale mají něco navíc: účinné snížení aktivační energie specifita regulovatelnost účinnosti (aktivity) Regulace účinnosti
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Historie. Pozor! né vždy jen bílkovinná část
 Enzymy a hormony Enzymy = biokatalyzátory jejich působení je umožněn souhrn chemických přeměn v organismu (metabolismus) jednoduché, složené bílkoviny globulární v porovnání s katalyzátory účinnější, netoxické,
Enzymy a hormony Enzymy = biokatalyzátory jejich působení je umožněn souhrn chemických přeměn v organismu (metabolismus) jednoduché, složené bílkoviny globulární v porovnání s katalyzátory účinnější, netoxické,
ENZYMY. Charakteristika enzymaticky katalyzovaných reakcí:
 ENZYMY Definice: Enzymy (biokatalyzátory) jsou jednoduché či složené makromolekulární bílkoviny s katalytickou aktivitou. Urychlují reakce v organismech tím, že snižují aktivační energii (Ea) potřebnou
ENZYMY Definice: Enzymy (biokatalyzátory) jsou jednoduché či složené makromolekulární bílkoviny s katalytickou aktivitou. Urychlují reakce v organismech tím, že snižují aktivační energii (Ea) potřebnou
Typy molekul, látek a jejich vazeb v organismech
 Typy molekul, látek a jejich vazeb v organismech Typy molekul, látek a jejich vazeb v organismech Organismy se skládají z molekul rozličných látek Jednotlivé látky si organismus vytváří sám z jiných látek,
Typy molekul, látek a jejich vazeb v organismech Typy molekul, látek a jejich vazeb v organismech Organismy se skládají z molekul rozličných látek Jednotlivé látky si organismus vytváří sám z jiných látek,
Stanovení vybraných enzymů. Roman Kanďár
 Stanovení vybraných enzymů Roman Kanďár Takže prvně malé opakování ENZYM Protein (RNA) s katalytickou aktivitou Protein (RNA) kofaktor (prosthetická skupina, koenzym) Jaký je vlastně rozdíl mezi prosthetickou
Stanovení vybraných enzymů Roman Kanďár Takže prvně malé opakování ENZYM Protein (RNA) s katalytickou aktivitou Protein (RNA) kofaktor (prosthetická skupina, koenzym) Jaký je vlastně rozdíl mezi prosthetickou
ENZYMOLOGIE. Pracovní sešit k přednáškám z biochemie pro studenty biologických kombinací ZDENĚK GLATZ
 EZYMLGIE Pracovní sešit k přednáškám z biochemie pro studenty biologických kombinací II ZDEĚK GLATZ 2004 Katalýza - Berzelius 1838 2 EZYMLGIE katalyzátor - látky urychlující chemické reakce - nemění rovnováhu
EZYMLGIE Pracovní sešit k přednáškám z biochemie pro studenty biologických kombinací II ZDEĚK GLATZ 2004 Katalýza - Berzelius 1838 2 EZYMLGIE katalyzátor - látky urychlující chemické reakce - nemění rovnováhu
Rychlost chemické reakce A B. time. rychlost = - [A] t. [B] t. rychlost = Reakční rychlost a stechiometrie A + B C; R C = R A = R B A + 2B 3C;
![Rychlost chemické reakce A B. time. rychlost = - [A] t. [B] t. rychlost = Reakční rychlost a stechiometrie A + B C; R C = R A = R B A + 2B 3C;](/thumbs/56/39180690.jpg "Rychlost chemické reakce A B. time. rychlost = - [A] t. [B] t. rychlost = Reakční rychlost a stechiometrie A + B C; R C = R A = R B A + 2B 3C;") Rychlost chemické reakce A B time rychlost = - [A] t rychlost = [B] t Reakční rychlost a stechiometrie A + B C; R C = R A = R B A + 2B 3C; 1 1 R A = RB = R 2 3 C Př.: Určete rychlost rozkladu HI v následující
Rychlost chemické reakce A B time rychlost = - [A] t rychlost = [B] t Reakční rychlost a stechiometrie A + B C; R C = R A = R B A + 2B 3C; 1 1 R A = RB = R 2 3 C Př.: Určete rychlost rozkladu HI v následující
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
13. Enzymy aktivační energie katalýza makroergické sloučeniny
 13. Enzymy Průběh chemických reakcí závisí též na schopnosti molekul přiblížit se dostatečně blízko a překonat repulsní energetickou bariéru. K tomu je zapotřebí energie typické pro každou reakci, tzv.
13. Enzymy Průběh chemických reakcí závisí též na schopnosti molekul přiblížit se dostatečně blízko a překonat repulsní energetickou bariéru. K tomu je zapotřebí energie typické pro každou reakci, tzv.
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
BIOKATALYZÁTORY I. ENZYMY
 BIOKATALYZÁTORY I. Obecné pojmy - opakování: Katalyzátory látky, které ovlivňují průběh katalyzované reakce a samy se přitom nemění. Dělíme je na: pozitivní (aktivátory) urychlující reakce negativní (inhibitory)
BIOKATALYZÁTORY I. Obecné pojmy - opakování: Katalyzátory látky, které ovlivňují průběh katalyzované reakce a samy se přitom nemění. Dělíme je na: pozitivní (aktivátory) urychlující reakce negativní (inhibitory)
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
První testový úkol aminokyseliny a jejich vlastnosti
 První testový úkol aminokyseliny a jejich vlastnosti Vysvětlete co znamená pojem α-aminokyselina Jaký je rozdíl mezi D a L řadou aminokyselin Kolik je základních stavebních aminokyselin a z čeho jsou odvozeny
První testový úkol aminokyseliny a jejich vlastnosti Vysvětlete co znamená pojem α-aminokyselina Jaký je rozdíl mezi D a L řadou aminokyselin Kolik je základních stavebních aminokyselin a z čeho jsou odvozeny
Farmakologie. -věda o lécích používaných v medicíně -studium účinku látek na fyziologické procesy -biochemie s jasným cílem
 Farmakologie -věda o lécích používaných v medicíně -studium účinku látek na fyziologické procesy -biochemie s jasným cílem Léky co v organismu ovlivňují? Většina léků působí přes vazbu na proteiny u nichž
Farmakologie -věda o lécích používaných v medicíně -studium účinku látek na fyziologické procesy -biochemie s jasným cílem Léky co v organismu ovlivňují? Většina léků působí přes vazbu na proteiny u nichž
Lékařská chemie a biochemie modelový vstupní test ke zkoušce
 Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Průvodka. CZ.1.07/1.5.00/ Zkvalitnění výuky prostřednictvím ICT. III/2 Inovace a zkvalitnění výuky prostřednictvím ICT
 Průvodka Číslo projektu Název projektu Číslo a název šablony klíčové aktivity CZ.1.07/1.5.00/34.0802 Zkvalitnění výuky prostřednictvím ICT III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Příjemce
Průvodka Číslo projektu Název projektu Číslo a název šablony klíčové aktivity CZ.1.07/1.5.00/34.0802 Zkvalitnění výuky prostřednictvím ICT III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Příjemce
16a. Makroergické sloučeniny
 16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
Enzymy. Prof. MUDr. Jiří Kraml, DrSc.
 Enzymy Prof. MUDr. Jiří Kraml, DrSc. ENZYMY JAKO HOMOGENNÍ BIOKATALYZÁTORY 1. Bílkovinná povaha ( + některé RNA-enzymy - ribozymy) 2. Větší účinnost (faktor minimálně 10 6 ) 3. Specifičnost - substrátová
Enzymy Prof. MUDr. Jiří Kraml, DrSc. ENZYMY JAKO HOMOGENNÍ BIOKATALYZÁTORY 1. Bílkovinná povaha ( + některé RNA-enzymy - ribozymy) 2. Větší účinnost (faktor minimálně 10 6 ) 3. Specifičnost - substrátová
Energie v chemických reakcích
 Energie v chemických reakcích Energetická bilance reakce CH 4 + Cl 2 = CH 3 Cl + HCl rozštěpení vazeb vznik nových vazeb V chemických reakcích dochází ke změně vazeb mezi atomy. Vazebná energie uvolnění
Energie v chemických reakcích Energetická bilance reakce CH 4 + Cl 2 = CH 3 Cl + HCl rozštěpení vazeb vznik nových vazeb V chemických reakcích dochází ke změně vazeb mezi atomy. Vazebná energie uvolnění
Gymnázium, Milevsko, Masarykova 183 Školní vzdělávací program (ŠVP) pro vyšší stupeň osmiletého studia a čtyřleté studium 4.
 pro vyšší stupeň osmiletého studia a čtyřleté studium 4.") Vyučovací předmět - Chemie Vzdělávací obor - Člověk a příroda Gymnázium, Milevsko, Masarykova 183 Školní vzdělávací program (ŠVP) pro vyšší stupeň osmiletého studia a čtyřleté studium 4. ročník - seminář
Vyučovací předmět - Chemie Vzdělávací obor - Člověk a příroda Gymnázium, Milevsko, Masarykova 183 Školní vzdělávací program (ŠVP) pro vyšší stupeň osmiletého studia a čtyřleté studium 4. ročník - seminář
Jana Fauknerová Matějčková
 Jana Fauknerová Matějčková glykosyltransferáza schopná syntetizovat řetězec prvních několika molekul glukosy jako základ nové molekuly glykogenu glykogenin tak slouží jako primer prvním krokem je navázání
Jana Fauknerová Matějčková glykosyltransferáza schopná syntetizovat řetězec prvních několika molekul glukosy jako základ nové molekuly glykogenu glykogenin tak slouží jako primer prvním krokem je navázání
BIOLOGICKÁ MEMBRÁNA Prokaryontní Eukaryontní KOMPARTMENTŮ
 BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Biochemie. ochrana životního prostředí analytická chemie chemická technologie Forma vzdělávání: Platnost: od 1. 9. 2009 do 31. 8.
 Studijní obor: Aplikovaná chemie Učební osnova předmětu Biochemie Zaměření: ochrana životního prostředí analytická chemie chemická technologie Forma vzdělávání: denní Celkový počet vyučovacích hodin za
Studijní obor: Aplikovaná chemie Učební osnova předmětu Biochemie Zaměření: ochrana životního prostředí analytická chemie chemická technologie Forma vzdělávání: denní Celkový počet vyučovacích hodin za
Enzymy: Struktura a mechanismus působení. Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1.LF UK
 Enzymy: Struktura a mechanismus působení Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1.LF UK 1 ENZYMY JAKO HOMOGENNÍ BIOKATALYZÁTORY 1. Bílkovinná povaha ( + některé RNA-enzymy - ribozymy) 2.
Enzymy: Struktura a mechanismus působení Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1.LF UK 1 ENZYMY JAKO HOMOGENNÍ BIOKATALYZÁTORY 1. Bílkovinná povaha ( + některé RNA-enzymy - ribozymy) 2.
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Fotosyntéza
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Chemie - 5. ročník. přesahy, vazby, mezipředmětové vztahy průřezová témata. očekávané výstupy RVP. témata / učivo. očekávané výstupy ŠVP.
 očekávané výstupy RVP témata / učivo Chemie - 5. ročník Žák: očekávané výstupy ŠVP přesahy, vazby, mezipředmětové vztahy průřezová témata 1.2., 2.1., 2.2., 2.4., 3.3. 1. Přeměny chemických soustav chemická
očekávané výstupy RVP témata / učivo Chemie - 5. ročník Žák: očekávané výstupy ŠVP přesahy, vazby, mezipředmětové vztahy průřezová témata 1.2., 2.1., 2.2., 2.4., 3.3. 1. Přeměny chemických soustav chemická
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Metabolismus sacharidů. VY_32_INOVACE_Ch0216.
 Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Lipidy a biomembrány
 5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
Enzymy (katalýza biochemických reakcí)
") Enzymy (katalýza biochemických reakcí) Enzymy (fermenty) Biokatalyzátory chemických reakcí (globulární proteiny) Ve velmi malých množstvích specificky urychlují průběh chemických reakcí tak, že snižují
Enzymy (katalýza biochemických reakcí) Enzymy (fermenty) Biokatalyzátory chemických reakcí (globulární proteiny) Ve velmi malých množstvích specificky urychlují průběh chemických reakcí tak, že snižují
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Opakování
 Slabé vazebné interakce Opakování Co je to atom? Opakování Opakování Co je to atom? Atom je nejmenší částice hmoty, chemicky dále nedělitelná. Skládá se z atomového jádra obsahujícího protony a neutrony
Slabé vazebné interakce Opakování Co je to atom? Opakování Opakování Co je to atom? Atom je nejmenší částice hmoty, chemicky dále nedělitelná. Skládá se z atomového jádra obsahujícího protony a neutrony
ZÁPADOČESKÁ UNIVERZITA V PLZNI
 ZÁPADOČESKÁ UNIVERZITA V PLZNI FAKULTA PEDAGOGICKÁ KATEDRA CHEMIE Enzymy ve výuce na vyšším stupni gymnázia DIPLOMOVÁ PRÁCE Jakub Král Učitelství pro střední školy, obor Ch - Ge Vedoucí práce: Mgr. Milan
ZÁPADOČESKÁ UNIVERZITA V PLZNI FAKULTA PEDAGOGICKÁ KATEDRA CHEMIE Enzymy ve výuce na vyšším stupni gymnázia DIPLOMOVÁ PRÁCE Jakub Král Učitelství pro střední školy, obor Ch - Ge Vedoucí práce: Mgr. Milan
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Glykolýza a neoglukogenese
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDr. Martin Vejražka, PhD. Tato prezentace je přístupnp stupná on-line CHE1.LF1.CUNI.CZ Prezentace Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha =
Regulace enzymové aktivity MUDr. Martin Vejražka, PhD. Tato prezentace je přístupnp stupná on-line CHE1.LF1.CUNI.CZ Prezentace Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha =
Oligobiogenní prvky bývají běžnou součástí organismů, ale v těle jich již podstatně méně (do 1%) než prvků makrobiogenních.
 než prvků makrobiogenních.") 1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty
 sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Biologie buňky. systém schopný udržovat se a rozmnožovat
 Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Produkce kyselin v metabolismu Těkavé: 15,000 mmol/den kyseliny uhličité, vyloučena plícemi jako CO 2 Netěkavé kyseliny (1 mmol/kg/den) jsou vyloučeny
 jsou vyloučeny") Vnitřní prostředí a acidobazická rovnováha 13.12.2004 Vnitřní prostředí Sestává z posuzování složení extracelulární tekutiny z hlediska izohydrie (= optimální koncentrace ph) izoionie (= optimální koncentrace
Vnitřní prostředí a acidobazická rovnováha 13.12.2004 Vnitřní prostředí Sestává z posuzování složení extracelulární tekutiny z hlediska izohydrie (= optimální koncentrace ph) izoionie (= optimální koncentrace
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
AMINOKYSELINY REAKCE
 CHEMIE POTRAVIN - cvičení AMINOKYSELINY REAKCE Milena Zachariášová (milena.zachariasova@vscht.cz) Ústav chemie a analýzy potravin, VŠCHT Praha REAKCE AMINOKYSELIN část 1 ELIMINAČNÍ REAKCE DEKARBOXYLACE
CHEMIE POTRAVIN - cvičení AMINOKYSELINY REAKCE Milena Zachariášová (milena.zachariasova@vscht.cz) Ústav chemie a analýzy potravin, VŠCHT Praha REAKCE AMINOKYSELIN část 1 ELIMINAČNÍ REAKCE DEKARBOXYLACE
V organismu se bílkoviny nedají nahradit žádnými jinými sloučeninami, jen jako zdroj energie je mohou nahradit sacharidy a lipidy.
 BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
TEST + ŘEŠENÍ. PÍSEMNÁ ČÁST PŘIJÍMACÍ ZKOUŠKY Z CHEMIE bakalářský studijní obor Bioorganická chemie 2010
 30 otázek maximum: 60 bodů TEST + ŘEŠEÍ PÍSEMÁ ČÁST PŘIJÍMACÍ ZKUŠKY Z CEMIE bakalářský studijní obor Bioorganická chemie 2010 1. apište názvy anorganických sloučenin: (4 body) 4 BaCr 4 kyselina peroxodusičná
30 otázek maximum: 60 bodů TEST + ŘEŠEÍ PÍSEMÁ ČÁST PŘIJÍMACÍ ZKUŠKY Z CEMIE bakalářský studijní obor Bioorganická chemie 2010 1. apište názvy anorganických sloučenin: (4 body) 4 BaCr 4 kyselina peroxodusičná
Metabolismus bílkovin. Václav Pelouch
 ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová
 Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
Pentosový cyklus. osudy glykogenu. Eva Benešová
 Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Úvod do studia organické chemie
 Úvod do studia organické chemie 1828... Wöhler... uměle připravil močovinu Organická chemie - chemie sloučenin uhlíku a vodíku, případně dalších prvků (O, N, X, P, S) Příčiny stability uhlíkových řetězců:
Úvod do studia organické chemie 1828... Wöhler... uměle připravil močovinu Organická chemie - chemie sloučenin uhlíku a vodíku, případně dalších prvků (O, N, X, P, S) Příčiny stability uhlíkových řetězců:
7. Enzymy. klasifikace, názvosloví a funkce
 7. Enzymy klasifikace, názvosloví a funkce Jsou to přírodní katalyzátory, živočišné i rostlinné Umožňují průběh biochemických reakcí Nachází se ve veškerých živých systémech Enzymy vykazují druhovou specifitu
7. Enzymy klasifikace, názvosloví a funkce Jsou to přírodní katalyzátory, živočišné i rostlinné Umožňují průběh biochemických reakcí Nachází se ve veškerých živých systémech Enzymy vykazují druhovou specifitu
Energetika a metabolismus buňky
 Předmět: KBB/BB1P Energetika a metabolismus buňky Cíl přednášky: seznámit posluchače s tím, jak buňky získávají energii k životu a jak s ní hospodaří Klíčová slova: energetika buňky, volná energie, enzymy,
Předmět: KBB/BB1P Energetika a metabolismus buňky Cíl přednášky: seznámit posluchače s tím, jak buňky získávají energii k životu a jak s ní hospodaří Klíčová slova: energetika buňky, volná energie, enzymy,
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Radiobiologický účinek záření. Helena Uhrová
 Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Chemická vazba. Molekula vodíku. Elektronová teorie. Oktetové pravidlo (Kossel, Lewis, 1916) Pevnost vazby vazebná energie.
 Pevnost vazby vazebná energie.") Elektronová teorie ktetové pravidlo (Kossel, Lewis, 1916) Chemická vazba sdílení 2 valenčních e - opačného spinu 2 atomy za vzniku stabilní elektronové konfigurace vzácného plynu Spojení atomů prvků v
Elektronová teorie ktetové pravidlo (Kossel, Lewis, 1916) Chemická vazba sdílení 2 valenčních e - opačného spinu 2 atomy za vzniku stabilní elektronové konfigurace vzácného plynu Spojení atomů prvků v