Genomika hospodářských zvířat
|
|
|
- Jindřich Pavlík
- před 6 lety
- Počet zobrazení:
Transkript
1 Genomika hospodářských zvířat 1
2 Mammalian genome ~3 x 10 9 base pairs Unique genes (protein-coding and RNA genes) ~1% Repetitive sequences ~60-65% Intergenic unique DNA ~35-40% Genes ~<5% Genes in more than one copy ~1% Repetitive sequences ~60-65% Protein-coding gene families ~3% RNA gene families ~1% Dispersed repeats Tandem repeats (satellites) Exons ~0,2% Introns ~4,8% Centromeric heterochromatin (satellites) ~10-15% Mini- and micro- (satellites) ~10-15% Transposons ~3% LINEs ~20% Endo-retrovirus ~10% SINE s ~15% 2
3 1. 3
4 Mapování lokusů pro QTL Quantitative Trait Loci 4
5 metodou MAS Markery Asistovaná Selekce 5
6 Markery asistovaná selekce MAS je založena na vazbě mezi markerem a QTL. 6
7 Vztah mezi markerem a QTL 7
8 Genetická vazba 8
9 Crossing-over není zcela náhodným procesem z hlediska toho, kde probíhá. Běžně nenastává uvnitř genů nebo jejich exonů. 9
10 Opakování: LOD skóre - statistická metoda pro detekci vazby z poměru dvou pravděpodobností - Log logaritmus - ODds šance pro převahu pravděpodobnosti - skóre je zde poměr skutečných nebo hypotetických rekombinant k nerekombinantám Dekadický logaritmus poměru pravděpodobností, šance, že pozorovaná část potomků vznikla jako výsledek genetické vazby k pravděpodobnosti, že vznikla náhodou.10
11 LOD skóre: lze určit existenci vazby a souč. její sílu výsledky segregace potomků z různých křížení lze jednoduše akumulovat k vyhodnocení stačí dvougenerační rodokmen (P a F1), v P nemusí být známa vazbová fáze pro důkaz vazby stačí minimální počet potomků potomci nemusí pocházet jen z dvojnásobných testovacích křížení, tj. AaBb x aabb, ale i z jiných typů, jednoduché nebo dvojnás. intercrossy AaBb x Aabb; AaBb x AaBb bez dominance či s dominancí v jedné nebo obou alelách. 11
12 Porovnávají se dvě alternativní hypotézy: H 1 vazba existuje, rekombinační zlomek v intervalu 0 < 1/2, přitom 0 znamená úplnou vazbu H 0 vazba neexistuje, volná kombinovatelnost vloh, rekombinační zlomek ½ V čitateli je hodnoceno konkrétní skóre ve prospěch existence vazby a ve jmenovateli je zvažováno skóre, jakoby vazba neexistovala. 12
13 LOD skóre Z log pravd. sekvence s danou hodnotou v azby pravd. vzniku sekvence bez vazby Výsledek Lod skóre z jednoho typu křížení (páření, rodiny) se sčítá s dalšími Z 1, Z 2, Z 3 SZ. 13
14 NR je počet nerekombinant v potomstvu R počet rekombinant 0.5 ve jmenovateli znamená, že jakékoliv alely, které jsou zcela volně komb., tzn. že jsou na různých chromozómech, mají 50% šanci rekombinace v důsledku nezávislého třídění θ je rekombinační zlomek, tj. podíl porodů v nichž proběhla rekombinace mezi sledovanými genetickými markery, tj. je roven R / (NR + R) 14
15 Je li LOD skóre > 3, vazba na autozomu je přítomna (pro vazbu na chrom. X stačí > 2), protože pravděpodobnost existence vazby je 1000x větší než pravděpodobnost nulové hypotézy. Je- li LOD skóre < než - 2, vazba není přítomna, je volná kombinovatelnost, vazba je 100x méně pravděpodobná než žádná vazba. Je-li vypočtená hodnota mezi -2 a 3, nelze učinit rozhodnutí, je nutné pokračovat v analýzách. 15
16 Příklad N = 632 počet potomků ze zpětného testovacího křížení (AaBb x aabb) = 29 počet rekombinantů = r = 603 počet nerekombinantů = N r d = rekombinantní zlomek = r/n = 29/632 = 0,046 LOD = log 10 (0, * (1-0,046) 603 / 0,5 632 ) = 242 vazba je přítomna 16
17 LOD score 17
18 Mapovací funkce 18

19 Rozdíly v délce vazbových map u samců a samic 19
20 Parciální genetická mapa 20
21 Mapování QTL u skotu (metoda vnuček) 21
22 Princip mapování QTL u skotu 22


 F 1 (3 x 19) F")
 F 2 (315) F")
23 Hohenheimské rodiny Struktura a velikost rodokmenů x x x Mating F 1 (2 x 28) F 1 (3 x 19) F 1 (2 x 21) F 2 (316) F 2 (315) F 2 (335) 23
24 Schéma vazby mezi markerem a QTL v F 2 rodině 24
25 Interval spolehlivosti (95% CI) CI 95 = 3000/(kNδ 2 ) 3000 = délka genomu v cm k = počet informativních rodičů na jedince (1 pro polo sourozence a zpětné páření, 2 pro F 2 populaci ) N = počet zvířat v experimentu Δ = velikost kontrastu (SD) Příklad: Máme rodinu polosourozenců (skot), N = 1000, efekt je roven 0,5 standardní odchylky pak CI 95 = 3000/(1 x 1000 x ) = 12 cm 25
26 Velikost intervalu spolehlivosti detekovaného QTL je nepřímo úměrná množství potomků v mapovací populaci a nepřímo úměrná druhé mocnině efektu QTL. 26
27 Velikost efektu QTL je kritická pro přesnost lokalizace. QTL s malým efektem mohou být lokalizovány pouze na chromozóm! QTL s velkým efektem mohou být v početné mapovací populaci lokalizovány s nejmenším intervalem spolehlivost 11 cm. Přesnost lokalizace QTL se středním efektem se pohybuje mezi výše uvedenými extrémy. 27
28 Faktory kritické pro přesnou lokalizaci 1) hustota markerů Čím více markerů, tím menší průměrný interval a vyšší rozlišení. 2) hustota crossoverů Pouze rekombinace poskytují mapovací informace. 3) detekovatelnost QTL Záleží na možnosti detekce genotypu QTL u daného jedince nebo na chromozómu. Zjištění polohy QTL vzhledem ke crossoveru vyžaduje znalost QTL alely vyskytující se na daném chromozómu. 4) Molekulární architektura QTL Mnoho QTL zahrnuje kombinovaný efekt více QTL. 28
29 Dosavadní výsledky vazbového mapování QTL Popsáno mnoho QTL. Ale popsáno pouze minimální množství genů (příčinných mutací)! Vět. jde o geny velkého účinku. Pravděpodobně existuje mnoho dalších genů malého účinku, jejich zjištění bude obtížné. Využití genomických informací získaných v rodinách je velmi omezené. 29
30 Pro mapování QTL existuje řada počítačových programů (Mapmaker/QTL, QTL Cartographer, Map Manager QT, QGene TM, MapQTL TM, PLABQTL, Multimapper, The QTL Cafe, Epistat). 30
31 QTL v IGF2 prasete IGF2-intron G je asociována s větším osvalením a menším množstvím tuku IGF2 je maternálně imprintovaný gen 31
32 32
 QTL")
33 Myostatin (GDF8) QTL 33
34 Ještě jednou: Markery asistovaná selekce MAS je založena na vazbě mezi markerem a QTL. 34
35 MAS nebyla nikdy masově implementována. Hl. důvodem nedostatek technologické efektivnosti. 35
36 Dílčí důvody: Mikrosatelity jsou v genomu distribuovány příliš řídce. Proto nejsou v dostatečně silné asociaci s cílovými geny. 36
37 Proto bylo nutné provést nákladné vazbové mapovací studie, schopné identifikovat genomické oblasti nazývané QTL s velkým (největším) efektem. Jelikož tyto oblasti jsou jen volně asociovány s markery, musely být připraveny komplikované šlechtitelské postupy, aby mohly být QTL použity ve více než jen jedné generaci. 37
38 Hlavním nedostatkem ale byly naivní předpoklady o genetickém pozadí komplexních vlastností. Ty mají většinou komplexní dědičnost s mnoha minorgeny, komplexní epistatické interakce, částečně ovlivněné epigenetickými mechanismy. Nebyl dostatečný počet QTL s dostatečně velkými efekty, které by se daly identifikovat ve vazbových studiích realistické velikosti. 38
39 MAS sice byla v několika šlechtitelských programech implementována, nestala se ale klíčovým elementem šlechtitelských programů. 39
40 Kandidátní geny a kriteria pro jejich výběr Kandidátní gen, kandidátní fyziologický gen Je součástí metabolických drah, vedoucích k manifestaci studovaného znaku. Poziční kandidátní gen Nachází se v dříve identifikované QTL oblasti. 40
41 Kandidátní geny a kriteria pro jejich výběr 1) znalost funkce genu 2) úloha genu ve vývoji dané vlastnosti 3) znalosti o vlivu genu na fenotyp u jiných druhů (myš, člověk) 4) exprese genu v dané tkáni a to i během vývoje (sval, tuk apod.) 5) gen se nachází v blízkosti QTL pro danou vlastnost (poziční kandidátní gen) 41
42 Standardy asociační analýzy používat práh signifikance pro mnohonásobné testování (viz dále) úplně popsat diverzitu haplotypů a strukturu existující ve studované populaci opakovat analýzu pozitivní asociace v nezávislé populaci zahrnout údaje o resekvenování odlišných haplotypů za účelem identifikace domnělých příčinných mutací Jinak se často pro vztah mezi genem a užitkovostí dělají předčasné nebo statisticky nepodložené závěry! 42
43 2. 43
44 Mapování QTL pomocí vazbové nerovnováhy Linkage Disequilibrium (LD) 44
45 Mapování QTL pomocí vazbové nerovnováhy založeno na existenci LD mezi QTL a markerem v náhodně se pářící populaci. Z efektu testovaného markeru na fenotypový znak usuzujeme na přítomnost a efekt QTL. 45
46 Linkage Disequilibrium Vazbová nerovnováha 46
47 Haplotyp je specifická kombinace alel vyskytujících se na témže chromozómu nebo segmentu chromozómu 47
48 SNP1 SNP2 SNP3 [A / G] [A / T] [C / G ] A A C A A G A T C A T G G A C G A G G T C G T G Počet haplotypů pro N SNP je 2 N tj. v našem případě 2 3 = 8 48
49 Vazba, markery ve vazbě: fyzická společná lokalizace markerů na témže chromozómu. Diplotyp je haplogenotyp, tj. pár haplotypů, jeden zděděn od otce, jeden od matky. 49
![SNP1 [ A / a ] SNP2 [ B / b ] Frekvence dominantní vlohy p(a) p(b) Frekvence recesívní vlohy p(a) p(b) Pro nezávisle segregující SNP](/docs-images/91/106230522/images/50-0.jpg "platí, že frekvence haplotypu p(ab) = p(a) x p(b) To platí při vazbové rovnováze.")
50 SNP1 [ A / a ] SNP2 [ B / b ] Frekvence dominantní vlohy p(a) p(b) Frekvence recesívní vlohy p(a) p(b) Pro nezávisle segregující SNP platí, že frekvence haplotypu p(ab) = p(a) x p(b) To platí při vazbové rovnováze. Vazbová nerovnováha Frekvence haplotypu p(ab) p(a) x p(b) 50
51 Vazbová nerovnováha Frekvence haplotypů neodpovídají frekvencím teoreticky stanoveným na základě alelových frekvencí. 51
52 Vazbová nerovnováha nenáhodné roztřídění alel ze 2 nebo více lokusů čím jsou markery blíže, tím silnější je LD, protože rekombinace bude probíhat s nižší frekvencí markery kosegregují uvnitř rodin a mezi rodinami 52
53 Vazbová rovnováha alely SNP2 B b alely SNP1 A a p(a)p(b) p(a)p(b) p(a)p(b) p(a)p(b) p(a) p(a) p(b) p(b) příklad: p(a)p(b) + p(a)p(b) = p(b) { p(a) + p(a)} = p(b) 53
54 SNP1 [ A / a ] SNP2 [ B / b ] Frekvence dominantní vlohy p(a) p(b) Frekvence recesívní vlohy p(a) p(b) Vazbová nerovnováha frekvence haplotypu p(ab) = p(a) p(b) + D Znaménko D je nahodilé, pokud nesrovnáváme hodnoty D mezi populacemi nebo různými studiemi. D je Lewontinův parametr LD (Lewontin 1960). 54
55 Měření LD Nejjednodušší míra Rozdíl mezi skutečnou a očekávanou frekvencí haplotypů D ij = P ij p i q j P ij frekvence haplotypu ij (i = alela i v lokusu 1; j = alela j v lokusu 2); p i a q j frekvence alely i v lokusu 1 a alely j v lokusu 2. Absolutní hodnota D ij ( D ij ) je identická pro oba haplotypy pro jakýkoliv dvoualelový lokus. D ij, obdobně jako poněkud složitěji počítané D jsou pro výše zmíněné účely nepoužitelné kvůli vysoké závislosti na frekvenci alel. 55
56 alely SNP1 Vazbová nerovnováha alely SNP2 B b A p(a)p(b)+d p(a)p(b)-d p(a) a p(a)p(b)-d p(a)p(b)+d p(a) p(b) p(b) Potom p(a)p(b)+d + p(a)p(b)-d =p(b){ p(a)+p(a)} = p(b) 56
57 Skutečně zjištěné frekvence: b B a 0,16 0,04 p(a)=0,20 Co je LD? A 0,14 0,66 p(b)=0,30 p(b)=0,70 p(a)=0,80 0 p(ab) p(a) p(b) p(ab) = p(a) p(b) + D 0,16 = 0,2 x 0,3 + D D = 0,1 Protože p(ab) = p(a)p(b) + D Bylo použito +D, zde D je +ve ale libovolně např. můžeme přeznačit alely A, B jako recesívní 57
58 Rozsah hodnot D (-ve až +ve) D má minimální a maximální hodnotu, která závisí na alelových frekvencích markerů. Jelikož frekvence haplotypů nemohou být ve: p(ab) = p(a)p(b) - D 0 p(ab) = p(a)p(b) - D 0 D p(a)p(b) D p(a)p(b) To nemůže platit zároveň, takže D min( p(a)p(b), p(a)p(b) ) p(ab) = p(a)p(b) + D 0 p(ab) = p(a)p(b) + D 0 D -p(a)p(b) D -p(a)p(b) To nemůže platit zároveň, takže D max( -p(a)p(b), -p(a)p(b) ) * Podobné rovnice pokud definujeme p(ab) = p(a)p(b) - D 58
59 Limity parametru vazbové nerovnováhy D Limity D jsou funkcí alelových frekvencí. Standardizace D přepočtem na podíl jeho maximální hodnoty pro dané alelové frekvence (D') D = D D max 59
60 D D = D / D max D max = min (p(a)p(b), p(a)p(b)) D < 0 D max = min (p(a)p(b), p(a)p(b)) D > 0 Opět, znaménko D závisí na definici. D = 1 nebo -1 pokud jeden z haplotypů p(a)p(b), p(a)p(b), p(a)p(b), p(a)p(b) = 0. To znamená, že je kompletní vazbová nerovnováha, tj. vyskytují se pouze 3 haplotypy. D =1 nebo -1 ukazuje, že mezi markery neproběhla rekombinace. Pozor na zřídkavé markery. Je možné, že marker není dost četný nebo není dost četný výběr, aby se vyskytl 4. haplotyp. 60
61 Interpretace D b B b B a 0,06 0,14 p(a)=0,20 a 0,2 0 p(a)=0,20 A 0,24 0,56 p(a)=0,80 A 0,1 0,7 P(A)=0,80 p(b)=0,30 p(b)=0,70 D = 0; D max nedefinováno četnosti haplotypů se rovnají příslušným součinům četností alel D = 1 úplná vazbová nerovnováha vyjádřená s použitím D - není rekombinace mezi markery - detekovány jen 3 haplotypy p(b)=0,30 p(b)=0,70 D = D max = 0,14; D = +1 p(a) = 0,2 p(b)= 0,3 61
62 Měření LD Nejlepší míra D ij 2 r 2 = p 1 p 2 q 1 q 2 r 2 v rozmezí 0 a 1 (0 = LD není, 1 kompletní LD). Také r 2 je částečně závislý na frekvenci alel, ale tato závislost je mnohem menší než u D ij a D. Lepší míra vazbové nerovnováhy pro korelace alel mezi markery, pro předpovídací schopnost alel SNP1 pro alely na SNP2. Užíván rozsáhle v asociačním testování při mapování genů pro choroby nebo fenotypů. 62
63 Interpretace r 2 b B b B a A 0,06 0,14 0,24 0,56 p(a)=0,20 p(a)=0,80 a A 0,2 0 0,1 0,7 p(a)=0,20 p(a)=0,80 p(b)=0,30 p(b)=0,70 p(b)=0,30 p(b)=0,70 D=0; D max nedefinováno D=D max =0,14; D = +1 r 2 = 0 r 2 = 0,58 r 2 1 korelace není úplná, i když D = 1 r 2 = 1 když D = 1 a p(a) = p(b) = 0,3 p(a) = 0,2 p(b) = 0,3 63
64 Interpretace r 2 p(a) = 0,3 p(b) = 0,3 Pouze 2 haplotypy r 2 = 1 korelace je úplná D =1 (méně než 4 haplotypy) p(a) = p(b) (= 0,3 v našem případě) r 2 =1 pokud je korelace mezi markery úplná a jeden genotyp umožňuje přesnou předpověď druhého. Jsou přítomny pouze 2 haplotypy. D = 1 > r 2 = 1 není rekombinace a markery musí mít stejné alelové frekvence. SNPs jsou podobně staré. důsledky nízké r 2 neznamená nutně vysokou rekombinaci rozdílné alelové frekvence 64
65 Obvyklé míry vazbové nerovnováhy -1 D 1 Rekombinace 0 r 2 1 Korelace Existují i jiné míry, ale obvykle se neužívají. 65
66 Vznik LD Nejsnáze pochopitelné, jsou-li markery fyzicky ve vazbě Vznik LD Mutace Efekt zakladatele Inbreeding, nenáhodné páření Selekce Křížení Migrace Průchod hrdlem láhve nebo populační stratifikace Epistatické interakce LD může být mezi markery, které nejsou ve vazbě! 66
67 SNP1 SNP1 SNP2 A B n=3 haplotypy A n=2 haplotypy A b Rekombinace a a B SNP1 SNP2 A B A a a b B b n=4 haplotypy 67
68 Disequilibrium (nerovnováha) gametické fáze je vhodnější termín. Nicméně používá se linkage disequilibrium, vazbová nerovnováha. 68
69 Hlavní faktory vytvářející LD v populaci mutace, selekce, křížení (migrace) Hlavním faktor degradující LD rekombinace LD mezi vzdálenými markery eroduje rychle, mezi blízkými markery (geny) pomalu. 69
70 Faktory ovlivňující vznik a degradaci LD v populaci IBD, Identity By Descends Úsek chromozómu se shodnými alelami (haplotyp), jaké měl předek, u kterého vznikla příčinná mutace ovlivňující variabilitu užitkového znaku. Je potomkům předáván bez rekombinace. 70
71 71
72 Faktory ovlivňující velikost a rozsah LD v populaci DŮLEŽITÉ Náhodný posun (random drift) U populací s relativně malým počtem jedinců ovlivňuje frekvence alel a holotypů. Čím je menší Ne populace tím větší je efekt driftu. 72
73 Faktory ovlivňující velikost a rozsah LD v populaci DŮLEŽITÉ Selekce Vliv selekce na LD závisí na směru, intenzitě a délce trvání selekce. Selekce redukuje genetickou variabilitu v následující generaci a produkuje negativní LD mezi lokusy. Když je selekce aplikovaná na určitý lokus, sousední lokusy, které jsou s tímto lokusem v LD budou mít zvýšenou LD. Tzv. hitchhiking efekt. 73
74 Faktory ovlivňující velikost a rozsah LD v populaci DŮLEŽITÉ Křížení Tvoří nové (falešné) LD mezi lokusy, které dříve nebyly v LD v původních populacích a mění rozsah LD pro lokusy které byly v LD v původních populacích. Tyto falešné LD se rychle ztrácejí v následujících generacích. 74
75 Faktory ovlivňující velikost a rozsah LD v populaci DŮLEŽITÉ Vztah mezi fyzickou a genetickou vzdáleností Procento rekombinací na jednotku fyzické vzdálenosti není v genomu rovnoměrné. V některých oblastech je frekvence rekombinací (θ) < 0.3cM/Mb, zatím co v jiných oblastech je to >3 cm/mb. Tyto rekombinační hot spots a cold spots, rekombinační džungle a rekombinační pouště korespondují s bloky s nízkým a vysokým LD. 75
76 Hlavní faktory ovlivňující LD u hospodářských zvířat DŮLEŽITÉ V populacích hospodářských zvířat je pro LD rozhodující velikost populace. Efektivní velikost populace je u většiny hospodářských zvířat malá, a proto je rozsah LD velký!!! LD způsobené křížením je malé při křížení plemen, která se liší pouze frekvencí alel. Toto LD vymizí po několika generacích. Významné mutace vznikly pravděpodobně před mnoha generacemi. Selekce je pravděpodobně významný zdroj LD, její efekt se pravděpodobně uplatňuje v okolí specifických genů a proto má relativně malý vliv na průměrnou LD v celém genomu. 76
77 Efektivní velikost populace N e je počet jedinců, který zapříčiní stejný přírůstek inbreedingu, jako kdyby se rozmnožovali jako ideální populace. Jinak: máme velkou populaci, která není ideální, např. je v ní větší počet samců. V ní je přírůstek inbreedingu stejný, jako v menší populaci, která je ideální. Dále je uveden výpočet pro stejný a nestejný poměr pohlaví. stejný počet samců a samic N e = 1 2 F N e N nestejný počet samců a samic N e = 4 N m N f N m + N f Tato metoda je založena na analýze poměru pohlaví, existuje více metod. 77

78 Závislost LD na efektivní velikosti populace prasat vazbová nerovnováha N e 78
79 Degradace LD Zejména rekombinací 79
80 Degradace LD během generací Úplná LD Mapová vzdálenost Není LD 80
81 Degradace LD V krátkých vzdálenostech ca bp rovněž může působit genová konverze. LD se rozkládá dlouho, generace interbreedingu. 81
82 Genová konverze (gene conversion) Typ genetické rekombinace, kdy během meiózy popřípadě mitózy dochází k náhradě určitého genu jeho jinou kopií. 82
83 Genová konverze se vztahuje k přenosu DNA sekvence mezi dvěma nesesterskými chromatidami během rekombinace. Na začátku jsou dvě molekuly DNA, donorová a akceptorová. Donorová molekula předá část svého řetězce molekule akceptorové. Na rozdíl od crossing-overu se změny odehrávají pouze na jednom vlákně akceptorové molekuly. 83
84 Rozdíl mezi crossing-overem a genovou konverzí U crossing-overu dochází ke změně na obou homologních úsecích. U genové konverze změna pouze na jednom vlákně DNA. 84

85 85
86 SNP1 SNP2 Pravděpodobnost, že rekombinace nastane = θ Pravděpodobnost, že rekombinace nenastane = 1-θ Počáteční LD mezi SNP1 a SNP2 je D 0 Po 1 generaci zbylá LD je Po t generacích D 1 = D 0 (1-θ) D t = D 0 (1- θ) t Pozn. jde o velmi jednoduchý model. Nepočítá s driftem měnícím v čase alelové frekvence. 86
87 Znázornění LD 87
88 D SNP ,0 0,8 0,6 0,2 0 SNP1 SNP2 SNP3 SNP4 SNP5 SNP6 Obvykle si nelámeme hlavu se znaménkem D 88

89 jednotlivé SNP hodnota p jednotlivé SNP 89
")
90 Haploview: TCN2 (r 2 ) 90
91 Co je ještě DŮLEŽITÉ 91
92 Inbreeding Inbreeding koeficient F je pravděpodobnost, že na náhodně vybraném lokusu v populaci je pár alel identic by descent, IBD. F se obvykle počítá z rodokmenu, pravděpodobnost že pár alel je IBD je odhadována statisticky. S použitím high-density SNP analýzy může být míra inbreedingu pro populaci určena, i když není rodokmen k dispozici. Skutečná hodnota inbreedingu pro jedince může být vypočtena z celogenomové sekvence. Dva postupy: 1/ testování markerů identických by state IBS 2/ runs of homozygosity ROH 92
93 Run of Homozygosity ROH ROH jsou úseky DNA s nepřerušenými úseky lokusů, které jsou homozygotní u jedince, ale polymorfní v populaci. Samozřejmě, to nemusí znamenat, že každý takový úsek zdědil jedinec od společného předka bez rekombinace. Nicméně, i když ROH může vzniknout z různých důvodů, předpokládá se, že hlavním důvodem je inbreeding. Pro jedince platí, že koeficient inbreedingu je podíl genomu, který je identical by descent, IBD, což lze určit z analýzy run of homozygosity, ROH. 93
94
95 Run of Homozygosity ROH Je vhodnou metodou pro hodnocení inbreedingu. Přesnější než výpočet z rodokmenu, protože odhady založené na genotypech ukazují skutečnou příbuznost mezi jedinci. Hodnocení inbreedingu pomocí genomového přístupu je zvl. vhodný, je-li neúplný rodokmen, nebo není dostatečně hluboký. Může se udělat u jakéhokoliv genotypovaného zvířete, i když nemá rodokmen. Lze provést také pro různé chromozómy. 95
96 Run of Homozygosity ROH U skotu byl ROH použit k analýze populační historie po nedávno proběhlé selekci, k odhadu koef. inb., ke studiu škodlivého vlivu inbreedingu na znaky ovlivňující ziskovost, k analýze a řízení nárůstu inbreedingu u genomově asistovaného šlechtitelského schematu. 96
97 Run of Homozygosity ROH Pro odhalování jsou vhodné FD čipy. Zjištěny vztahy mezi ROH a výskytem recesívních chorob. ROH mají vyšší riziko, že ponesou nežádoucí recesívní alely. Např. popsány ROH s vlivem na výskyt inf. chorob, mastitis, poruch pohybového aparátu, metabolických a reprodukčních poruch. Negativní asociace mezi přítomností ROH na BTA3, 7, 8, 12 a plodností dcer. Asociace mezi SCS a ROH na BTA 1, 3, 4, 5, 13, 21. Tj. negativní vliv inbrídingu. 97
98 Run of Homozygosity ROH Genomové úseky, které jsou cílem selekce, mají tendenci generovat dlouhé úseky ROH, s vysokou homozygotností kolem selektovaného lokusu. Riziko, že genomová selekce může vést k dlouhým homozygotním úsekům kolem oblastí s QTL. Selekce ovlivňuje model autozygotnosti v genomu. Brát v úvahu při ochraně genetické diverzity, zvl. u genových zdrojů. 98
99 Run of Homozygosity ROH Rekombinace narušuje dlouhé chromozomální úseky. Proto: dlouhé autozygotní úseky v individuálním genomu budou nalézány, má-li jedinec nedávného společného předka; kratší úseky budou nalézány, je-li společný předek více vzdálen. Čím delší jsou homozygotní úseky, tím je vzdálenost k proběhlému inbreedingu kratší. 99
100 Run of Homozygosity ROH Koeficienty inbreedingu založené na rodokmenech ignorují stochastické rozdíly v podílu genomu zděděného jako IBD relativně ke statistickému očekávání. Molekulárně a rodokmenově odhadnuté koeficienty inbreedingu se proto liší. Navíc, ne všechny ROH mají autozygotní původ. Malé ROH mohou vzniknout náhodně, ne jako IBD. Úseky kratší než 4 Mb mají nižší pravděpodobnost, že vznikly jako IBD. 100
101 Run of Homozygosity ROH 1000kb lze ca považovat za ekvivalentní 0,01 M. Mléčná plemena mají ROH delší a větší počet, než plemena kombinovaná a masná. To ukazuje na inbreeding v rel. nedávné době. Průměrný počet ROH na jedince, v závorce směrodatná odchylka Brown Holstein Marchigiana Piedmontese Simmental 101
102 Průměrná délka ROH u jednotlivých plemen v Mb Brown Holstein Marchigiana Piedm Simmental 102
103 Největší rozsah ROH zjištěn u BTA5 a BTA6. Simentál má krátkou průměrnou délku ROH, v genomu má více krátkých ROH, souvisí to s vyšší Ne. ROH lze měřit pomocí čipů, jako u dat na předešlých obr. U populací s vysokým LD a nedávným inbreedingem, např. holštýn, brown, stačí 54K čip. U populací s nízkým LD a starším inbreedingem je lepší high density čip. Ideální by bylo vycházet z celogenomové sekvence, což je otázka času (řádově roky). 103
104 ROH v Mb Počet generací ke společnému předku ,5 10 6,25 3,13 104
105 Dlouhé ROH jsou obohaceny o škodlivé mutace. Mnoho plemen pochází z bottlenecku. 105
106 Prasata: čínská mají nižší LD a méně ROH. Divoká prasata mají velký počet krátkých ROH, nemají dlouhé ROH. 106
107 Koně: malá diverzita a vysoký inbríding u plnokrevníka je důsledkem selekčního tlaku na výkonnost. Koně mají ve srov. s ost. druhy nejvyšší podíl genomu pokrytý ROH, nejvyšší počet ROH na zvíře, následují prasata. 107
108 S tím souvisí Inbrední deprese 108
109 Studie z poslední doby ukazují následující. Nebyl zjištěn efekt druhu, ale populace (plemene). Tj. deprese závisí více na frekvencích alel segregujících uvnitř plemen, než na fyziologických specifikách druhů. V některých studiích je vyšší deprese u produkčních znaků, než u fitness. U hosp. zvířat jakákoliv selektovaná vlastnost může být postižena inbrední depresí. Velká část genetické zátěže může být způsobena malým počtem alel s velkým škodlivým efektem. Existuje variace mezi liniemi, to je důležité pro šlechtění. 109
110 Velká část genomu, tj. velký počet genů, se podílí na inbrední depresi. Existuje částečná inbrední deprese. 110
111 Inbrední deprese Drůbež Produkční systémy jsou založeny téměř výhradně na křížení. Inbreedingová deprese může být z ekonomického hlediska pominuta. Mléčný skot Vět. čistokrevná plemena. Proto deprese musí být pod kontrolou. U ostatních druhů je situace intermediární. 111
112 Inbrední deprese U produkce mléka se odhaduje pokles 0,37% na 1% inbreedingu. U holštýna je roční přírůstek inbreedingu ca 0,12%, tj. za generaci 0,6% při generačním intervalu 5 let. Potom, je-li roční genetický zisk u holštýna ca 100 kg mléka na laktaci, tj. 1% současného fenotypového průměru 10 tis. kg. Inbrední deprese za rok potom odpovídá 0,044% fenotypového průměru, to je 4,4% genetického zisku. To sice není mnoho pro jednu vlastnost, ale počítáme-li s depresí i u ostatních vlastností, je ekonomický význam nepochybný. 112
113 Inbrední deprese Pomalý inbreeding zvyšuje počet generací, potřebných k očištění populace od genetické zátěže. Jedinci s inbredním původem mají nižší pravděpodobnost, že jsou přenašeči škodlivých alel! Staré škodlivé mutace jsou pravděpodobněji z populace vytlačeny ve srovnání s novými. Existují příklady populací, které se přirozeně zbavily genetické zátěže. Volně žijící populace Chillinghamského skotu, je životaschopná a plodná, ačkoliv je v důsledku inbreedingu téměř úplně homozygotní. 113
114 Inbrední deprese Inbrední jedinci jsou citliví na změny prostředí. ROH umožňuje rozlišit starý a nový inbreeding; lokální a genomwide inbreeding. Genomová selekce Umožní redukci inbreedingu. POZOR! Dosud se při genomové selekci inbreeding zvyšuje nutno napravit! 114
115 Faktory ovlivňující schopnost asociačního testu detekovat QTL testováním efektu jednotlivých markerů r 2 (tj. LD) mezi markerem a QTL. Velikost populace musí být zvýšena faktorem 1/r 2, aby bylo možno detekovat QTL ve srovnání s velikostí populace v případě, že byl QTL testován sám. Heritabilita vlastnost (h 2 ). Počet fenotypových záznamů (n). 115
116 Faktory ovlivňující schopnost asociačního testu detekovat QTL testováním efektu jednotlivých markerů Alelová frekvence méně často se vyskytující alely testovaného markeru (p). Průkaznost rychle klesá je li p <0.1. MAF < 0.1 (Minor Allele Frequency). Hladina průkaznosti (α) stanovená experimentátorem. 116
117 3. 117
118 Microarrays 118
119 Microarray Dvourozměrně uspořádané matrice, které obsahují vysokou hustotu (desítky tisíc až miliony) sond: DNA, cdna, PNA (peptidová nukleová kyselina), proteiny, protilátky, atd. Sondy obvykle ukotveny na chemicky upraveném skleněném nebo 119 plastovém podkladu.
120 Microarray Princip analýzy založen na selektivní hybridizaci imobilizovaných sond s cílovými biochemickými látkami nebo jejich fragmenty obsaženými v biologickém vzorku za použití nejčastěji fluorescenční nebo luminiscenční detekce. 120
121 Microarray Patří sem tedy více metod Společné je využití čipu s navázanými sondami - proteiny, DNA atd. 121
122 Microarray Microarrays, též bio-čipy, lze rozdělit dle hustoty sond a spotů - nízko, středně až vysoko hustotní, typu a účelu imobilizovaných molekul - protilátky, antigeny, bakteriální komponenty, sacharidy, DNA/cDNA, lipidy, oligonukleotidy, peptidy, proteiny, další malé molekuly, celé buňky a vzorky tkání, způsobu výroby, počtu možných aplikací biologických vzorků 122
123 DNA microarrays RNA aplikace - exprese RNA, exonů a krátkých řetězců, CGH comparative genomic hybridization, genotypizační nebo-li SNP (single nucleotide polymorphism), resekvenační, k analýze DNA metylací, populační čipy. 123
124 DNA microarrays Např. VIP čip (populační) pro novorozenecký screening, jedna analýza na jednom čipu, screening 1 tis. novorozenců pro 80 různých dědičných onemocnění najednou. Microarrays - ideální platforma pro genomové studie, rychlé zpracování velkého množství vzorků, přesná data a příznivé náklady. Klinické aplikace, např. genetická analýza v oblasti reprodukčního zdraví (prenatální, preimplantační a postnatální), onkologie (solidních nádorů a hematologických nádorů), transplantace. 124
125 VIP čip (populační) (Variation Identification Platform) využívá oproti původním DNA čipům, které jsou schopny testovat maximálně jednotky biologických vzorků, obrácené uspořádání volných sond a imobilizovaných vzorků, čímž se kapacita technologie diametrálně liší. Je možné testovat až biologických vzorků (jedinců populace) během jedné analýzy, každý v min. 10 lokusech. 125
126 Populační čipy umožňují ekonomicky provádět screening populace až desítek tisíc jedinců (od lidí až po mikroorganismy). Až desítky tisíc biologických vzorků na jednom čipu. 126
127 již nyní aplikace zahrnují VIP čip (populační) populační screening za účelem personalizované medicíny, racionální personální preskripce, korelaci genotypů u jedinců s děd. chorobou, např. mutantní geny u schizofreniků s následnou racionalizací preskripce, předpověď terapeutické účinnosti imunoglobulinové terapie u jednotlivých pacientů s relapsující remitující roztroušenou sklerózou (RRMS)7, autismus, diagnostika chorob, predikce onemocnění, neonatální screening, transplantační testy (HLA), forenzní využití, 127 parentální testy, genealogické testy.
128 VIP čip (populační) V oblasti veterinární a rostlinné se jedná zejména o populační screening za účelem šlechtění, nicméně do budoucna je pravděpodobné, že vznikne i v tomto odvětví poptávka po populačním screeningu za účelem genotypové diagnostiky. Technologie není druhově omezená. 128
129 Získání DNA Příklad postupu pro DNA microarray Kontrola kvality Celogenomová amplifikace Fragmentace Hybridisace Scan Softwarová analýza Data 129
130 Schéma klasické analýzy na čipu, tj. sonda je vázána, DNA vzorku je volná. 130

131 Funguje na principu alelově specifické hybridizace T C T C C 131
132 Jednou z možných aplikací je Expresní profil izolace mrna čištění vzorku (např. elektroforeticky) reverzní transkripce do cdna PCR označení vláken cdna fluorescenční barvou 132
133 Hybridizace, tj. navázání komplementárních sekvencí vzorku cdna a sond na čipu. Čtečka laserem excituje molekuly barvičky. Světlo je analyzováno. Výsledkem je informace o tom, jaké geny (info o tisících genů!) jsou exprimovány v příslušných buňkách (tkáni). Množství světla poskytuje informaci o relativním množství exprimovaného genu. Absolutní určení množství je komplikované. 133
134 Dvoukanálový experiment Směs dvou vzorků (např. ze zdravých a rakovinných buněk či z buněk jednoho druhu v různých podmínkách) označených různými barvivy. Výsledkem je relativní informace o rozdílu v expresi genů mezi vzorky. Info např. které geny jsou vlivem nemoci v buňkách podexprimované a které nadexprimované. 134
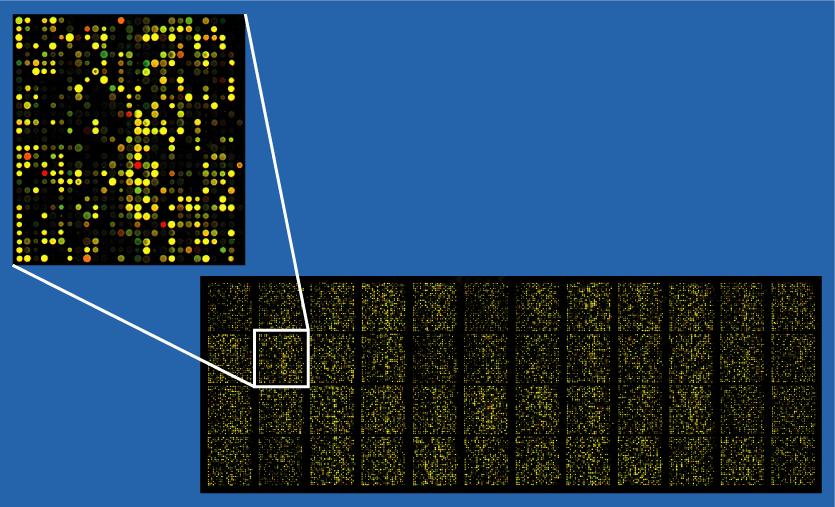
135 Dvoukanálový DNA čip 135
136 Microarrays a NGS: optimální je současné využití obou těchto platforem, které za současného state of the art vynikají vzájemnou výhodnou komplementaritou, a tato situace pravděpodobně potrvá několik příštích desetiletí. 136
137 Řada dalších aplikací. Nás zajímá zvl. identifikace alel SNP. Následné využití při genotypizaci, forenzní analýze, odhadu predispozice k chorobám, identifikaci mutací v zárodečných liniích, somatických mutací v nádorech. Pro naše účely je důležitá genotypizace alel SNP a následná analýza vazbové nerovnováhy. 137
138 Často se používají čipy fy Illumina (říjen 2016): Bovinní SNP čipy: BovineLD Genotyping BeadChip: SNPs BovineSNP50 v3 DNA Analysis BeadChip: SNPs BovineHD DNA Analysis Kit: SNPs Sekvenační technologie TruSeq Bovine Parentage Sequencing: panel 200 ISAGem doporučených SNPs pro parentitu plus 66 SNP asociovaných s recesivními dědičnými chorobami 138
139 Prase: PorcineSNP60 DNA Analysis Kit v2: 64,232 SNP 139

140 Vzdálenosti mezi SNP u BovineSNP50 140
141 Počet polymorfních lokusů BovineSNP50 BedChip u různých plemen skotu 141

142 Congenital muscular dystonia 1 - CMD1 142

143 143
144 4. 144
145 Genomová selekce 145
146 Genomová selekce Zásadním rozdílem mezi MAS a genomickou selekcí je, že MAS používá markery (SNP), které jsou statisticky významně asociovány se selektovanými znaky, zatím co genomická selekce používá celogenomový panel hustě vedle sebe umístěných markerů, takže se očekává, že všechny QTL jsou ve vazbové nerovnováze LD přinejmenším s jedním markerem. GS překonává problém s mnohonásobným testováním, tj. falešně pozitivní QTL a zachycení pouze fragmentu genetické proměnlivosti, protože všechny efekty jsou hodnoceny současně. 146
147 Genomová selekce Pracuje na základě rozdělení celého genomu do chromozómových úseků definovaných sousedními markery a následného sledování efektů těchto chromozómových úseků. 147
148 Genomová selekce Využívá předpokladu, že efekty chromozómových úseků budou shodné v celé populaci, protože markery jsou v LD s QTL mezi markery, mají stejnou vazbovou fázi! GS předpokládá, že všechny markery mohou být ve vazbě ke genu ovlivňujícím vlastnost. Koncentruje se na stanovení jejich efektu spíše než na testování významnosti. Proto hustota markerů musí být dostatečná, aby zajistila, že všechny QTL jsou v dostatečném LD s markery nebo haplotypy markerů. To umožňují SNP. 148
149 GPH může být interpretována dvěma ekvivalentními způsoby: výsledek kombinace odhadnutého efektu markerů a genotypu zvířete pro markery, výsledek odhadu BLUP, kde matrice příbuznosti mezi zvířaty v systému obsahuje genomové příbuznosti založené na tom, kolik markerů mají dvě zvířata společných, namísto rodokmenové příbuznosti. 149
150 Druhá definice je nádherně jednoduchá, zejména protože je nezávislá na znaku. Je mnohem snáze vysvětlitelná, stala se standartním způsobem vysvětlení. 150
151 Praktické použití genomové selekce 1. Odhad efektů chromozómových segmentů v referenční populaci. Zvířata referenční populace musí mít stanoveny genotypy markerů a mít záznamy o užitkovosti. 2. Předpověď GPH pro zvířata mimo referenční populaci, pro mladé býčky, kanečky atd. Tito kandidáti mají k dispozici pouze genotypy markerů, ale nemají k dispozici údaje o vlastní užitkovosti nebo užitkovosti potomstva. Formální kvantitativní genetika počítá s tím, že potomek zdědí polovinu vlastností od každého z rodičů. Ve skutečnosti to tak docela 151 neplatí.
152 Praktické použití genomové selekce Důležitá je genetická příbuznost, genetické propojení mezi referenční a selektovanou populací! Ref. populace musí být pravidelně aktualizována, aby byly odhady efektu markerů v souladu s novými generacemi šlechtěné populace. 152
153 Praktické použití genomové selekce Všechny SNP efekty jsou simultánně odhadovány v referenční populaci, která je genotypována a fenotypována s použitím statistického modelu (předpokládáme SNP): y i = m + X i1 x b 1 + X 2i x b X 50000i x b e i y i je fenotyp zvířete i; m průměr; X i1 genotyp zvířete i pro marker 1; e i residuální efekt 153
154 Praktické použití genomové selekce Předpokládáme, že efekty SNP mají normální rozdělení s průměrem 0 a konstantní variancí (která je celková genetická variance dělená ). Tedy metoda používá BLUP k odhadu SNP efektů, nazývá se někdy SNP-BLUP. 154
155 Praktické použití genomové selekce Genomová plemenná hodnota selekčního kandidáta j je v SNP-BLUP předpovídána jako: GEBV j = X 1j x 1 + X 2j x X 50000j x je odhad efektu SNP1, X 1j genotyp zvířete j pro SNP 1 155
156 Praktické použití genomové selekce Jinak: předpověď GPH pro zvířata bez údajů o vlastní užitkovosti se zakládá na sčítání efektů chromozómových úseků zjištěných v referenční populaci n GPH = Xgˆi i N počet chromozómových segmentů v genomu X i matrice, která přiřazuje zvířata k efektům markerů nebo haplotypů Xgˆi je vektor haplotypů v chromozómovém segmentu 156
157 Praktické použití genomové selekce Tzn. počítá se, jaký je efekt záměny alely v SNP. 157
158 Vlastní výpočet plemenné hodnoty se provádí např. metodou: BLUP-AM RR BLUP G BLUP ssg BLUP Blending ss G BLUP 158
159 V tradiční BLUP je PH odhadována s použitím fenotypů a rodokmenových příbuzností. Při použití metody GBLUP je GPH odhadována s použitím fenotypů a genomových příbuzností, které jsou založeny na celogenomových hustých markerových datech. 159
160 Genomická příbuznost mezi zvířaty 1 a 2 je počítána jako korelace mezi jejich SNP genotypy X j1 a X j2 přes všechny SNP j. GBLUP je tedy velmi podobná tradiční BLUP, ale rodokmenové příbuznosti jsou nahrazeny genomovými. Mohou být použity tradiční metody výpočtu a software. 160
161 Rodokmenová příbuznost sourozenců je 0,5 tedy by měli sdílet 50% společných alel. Ve skutečnosti mohou sdílet 40% nebo 60% společných alel. Tato odchylka může být detekována genotypováním markerů! GBLUP je přesnější, protože genomová příbuznost je přesnější, než rodokmenová. 161
162 Pro hosp. zvířata a příbuznost uvnitř plemene je SNP postačující. Příbuznost zvířat z různých plemen je malá, proto musí být použit větší počet SNP. 162
163 Statistický model pro GBLUP: y i = m + u i + e i u i je plemenná hodnota zvířete i GBLUP a SNP-BLUP jsou stejné, pokud definujeme u i jako: u i = X 1i x b 1 + X 2i x b X 50000i x b
164 GBLUP a SNP-BLUP mohou být z výpočetního hlediska velmi rozdílné. SNP-BLUP vyžaduje odhad efektů SNP, tj. řešení rovnic. GBLUP vyžaduje odhad N GPH a řešení N rovnic, N=počet zvířat. Protože počet zvířat je vět. menší než je GBLUP pro výpočetní důvody preferována. 164
165 Počet genotypovaných zvířat v budoucnu dramaticky naroste, možná že SNP-BLUP bude oblíbenější. Možná se stanou populárními i non-blup metody. 165
166 Nelineární metody genomové selekce Z biologického hlediska můžeme předpokládat, že některé SNP blízko genu mají efekt a mnohé jiné SNP nemají. Byla vyvinuta řada metod, zahrnujících informaci, že určitý podíl SNP π má efekt a podíl 1- π nemá. 166
167 Model: y i = m + I 1i x X 1i x b 1 + I 2i x X 2i x b I x X 50000i x b e i kde I j je indikátorová proměnná s hodnotami 0 nebo 1 indikující, zda SNP má nebo nemá efekt. 167
168 Apriorní distribuce SNP efektů užívaná v nelineárních metodách je biologicky logičtější než předpoklad, že všechny SNP mají efekt a všechny jsou velmi malé. V simulačních studiích nelineární metody jasně převyšují GBLUP. Ale: v reálných datech jsou nelineární metody poněkud lepší u některých vlastností, ne u všech. 168
169 Důvod: 1/ existuje mnoho genů ovlivňujících ekon. důležité vlastnosti, takže předpoklad, že všechny SNP mají efekt je přibližně správný. 2/ vazbová nerovnováha (ne-náhodná asociace mezi dvěma lokusy) zasahuje dlouhé úseky genomu v populacích zvířat, takže mnoho SNP je asociováno s genem. 3/ hustota SNP není dostatečná, takže každý QTL může být vysvětlen jedním SNP a je potřeba mnoho SNP, aby společně vysvětlily efekt QTL. 169
170 Zvyšování hustoty SNP Zvyšuje pravděpodobnost, že každý QTL má SNP v perfektní LD. Konečnou hustotou je nahrazení SNP genotypů úplnou genomovou sekvencí (WGS). Potom příčinné mutace budou v sekvenačních datech a tudíž genomová selekce bude působit přímo na mutace namísto toho, aby se spoléhala na LD mezi SNP a příčinnou mutací. Avšak tyto mutace jsou skryty mezi mnoha miliony SNP bez efektu. 170
171 Očekává se proto, že nelineární metody GS, které předpokládají, že mnoho SNP nemá efekt, budou u dat z celogenomového sekvenování (Whole Genome Sequencing) přesnější. 171
172 Imputace Na základě známých genotypů SNP zvířete je určen jeho haplotyp, protože stejný haplotyp byl pozorován u jiných zvířat. Tedy: chybějící genotyp může být přečten z genotypu těchto jiných zvířat se stejným haplotypem. Software: Beagle, Fimpute, Alphaimpute. 172
173 Imputace Může být použita v kombinaci s levnými SNP čipy. Klíčoví předci jsou genotypováni hustými ale drahými čipy, které identifikují haplotypy v populaci. Následně je velký počet potomků genotypován méně hustými levnými čipy, které ale mají dost SNP, aby odhalily, které haplotypy zvíře nese. Chybějící genotypy jsou dopočteny (imputovány). 173
174 Imputace Přesná je imputace u SNP s vysokou MAF (minor allele frequency). U SNP s nízkou MAF je přesnost malá. 174
175 Vícekroková GS 1/ jsou vypočteny pseudofenotypy pro genotypovaná zvířata, ty obsahují informace (záznamy) o jejich negenotypovaných příbuzných 2/ je provedena genomická předpověď s použitím pseudozáznamů (pseudofenotypů) a jejich genotypů 3/ tradiční plemenná hodnota a GPH jsou kombinovány do celkové PH. 175
176 Vícekroková GS Jinak: 1/ Máme genotypovaná zvířata, u kterých neznáme fenotyp. Ten dopočteme podle fenotypů jejich příbuzných. 2/ Následně provedeme odhad PH a GPH. 3/ Stanovíme celkovou PH. 176
177 Jednokroková GS, ssgblup Data jsou propočtena v jednom kroku. Zahrnuje všechny informace fenotypové, genomové, rodokmenové dohromady do jednoho modelu. Přechod od BLUP na GBLUP: nahrazujeme matrici rodokmenové příbuznosti matricí genomové příbuznosti. Udělat tam, kde je genomová příbuznost k dispozici. Kde není, nechat rodokmenovou příbuznost. U mléčného skotu je ssgblup o 0-2% přesnější než multistep. 177
178 Budoucnost ss odhad užívající nelineární statistickou metodu a data ze sekvenace! 178
179 Faktory ovlivňující přesnost GS Přesnost odhadu GPH závisí na několika parametrech. 1) Rozsah a velikost LD mezi markery a QTL. 2) Počet zvířat s fenotypy a genotypy v referenční populaci, ze které jsou efekty SNP odhadovány. 3) Příbuznost (genetické propojení) mezi referenční populací (která je genotypovaná a fenotypovaná) a populací s kandidátními zvířaty (která je genotypovaná, ale není fenotypovaná). 4) Dědivost (heritabilita - h 2 ) sledovaných znaků. 5) Distribuce QTL efektů. První tři faktory lze ovlivnit, druhé dva ne! 179
180 Heritabilita různých užitkových vlastnosti u černostrakatého skotu množství nadojeného mléka 0,30 množství mléčného tuku 0,30 množství mléčného tuku 0,30 procento tuku 0,58 procento bílkovin 0,51 celoživotní produkce mléka 0,15 celoživotní produkce tuku 0,15 celoživotní produkce bílkovin 0,14 délka produktivního života 0,13 počet somatických buněk v mléce 0,10 U populací chovaných ve špatných podmínkách jsou odhadovány nižší hodnoty. 180
181 Vztah mezi velikostí referenční populace, h 2 a přesností odhadu genomické plemenné hodnoty Počet genotypovaných a fenotypovaných zvířat požadovaných v referenční populaci pro dosažení žádoucí přesnosti odhadu GPH u zvířat, která jsou jen genotypována, ale ne fenotypována. N e = 100. Velikost referenční populace Přesnost odhadu GPH 0,75 0,50 Heritabilita vlastnosti 181

182 Velikost referenční populace a přesnost odhadu GPH s ohledem na efektivní velikost populace a heritabilitu vlastnosti Počet zvířat v referenční populaci Heritabilita vlastnosti Přesnost odhadu GPH u nefenotypovaných zvířat Počet zvířat v referenční populaci 182
183 Stav GS u různých druhů hospodářských zvířat 183
184 Srovnání komerčně nabízených metod čipové genotypizace SNP skotu (výběr) Produkt počet lokusů prům. mezera prům. MAF Illumina BovineSNP50 BovineHD Affymetrix GeneChip Bovine Mpapping 10K GeneChip Bovine Mpapping 25K Axiom BOS 1 Array Plate ,26 0,28 0,286 b 0,24 0,21-0,39 49,4 3, ,17 C a Molecular inversion probe. b Zahrnuje SNP z GeneChip Bovine Mapping 10K assay a markery z CSIRO, Australia. c SNP nebyly vybrány tak, aby byly rovnoměrně rozděleny, ale tak, aby byly rovnoměrně fyzicky rozděleny ve smyslu očekávané rekombinace mezi lokusy. Fyzické vzdálenosti mezi SNP jsou proto menší směrem k centromerám a telomerám chromozomů. 184 MAF Minor Allele Frequency.
185 První komerční čip v r Illumina BovineSNP50 BeadChip. Dostupné pro všechny důležité druhy hosp. zvířat, ovce, prase, kůň, kur, koza, pstruh, losos, atd. Počet SNP v první generaci čipů tis. V souč. (2015) je jen pro skot nabízeno 11 komerčních čipů od 3 největších firem, Illumina, Neogen-GeneSeek, Affymetrix. Dále se nabízí tzv. custom čipy. 185
186 Problém Standardizace - pojmenování SNP, - formátu vstupních a výstupních dat, - identifikace pozice SNP, tj. chromosom, posice páru basí Proto je obtížné srovnání výsledků z různých čipů a vzájemné využití dat pro analýzy. 186
187 Pozor při převodu plemenných hodnot mezi zeměmi. Př.: Korelace mezi zeměmi r = 0,85 Potom koef. determinace r 2 = 0,72 Spolehlivost odhadu býka v pův. zemi (např.) 0,80 Spolehlivost po převodu 0,80 x 0,72 = 0,58!!! 187
188 5. 188
189 Celogenomové asociační studie GWAS Genome-Wide Association Study 189

190 GWAS 190
191 Zavedením high-density microarray technologie pro detekci SNP se GWAS staly metodickým zlatým standardem pro identifikaci lokusů, které jsou příčinou komplexních chorob jak u lidí, tak u zvířat. 191
192 Rámcové schéma GWAS: - stažení surových dat z analýzy na čipech, kontrola kvality - detekce a oprava stratifikace populací, např. principal component analysis - vložení chybějících alel a genotypů; přípravný krok v řadě genetických analýz, protože většina modelů a softwarů neumí pracovat s chybějícími daty. Tzv. imputace. - testování asociací mezi jednotlivými SNP a kontinuálními nebo kategoriálními fenotypy 192
193 - celková analýza významnosti a oprava na hromadné testování - prezentace dat (grafy) - event. i metaanalýza, integrace dat z více GWAS 193
194 Ad vložení chybějících dat, imputace Klíčový krok ve většině GWAS. Provádí se i v genomové selekci. Vložení genotypů na SNPs, které nejsou na čipech, použitých ke studii. Tento proces značně zvyšuje počet SNPs, které mohou být testovány na asociaci, zvyšuje vypovídací sílu studie, ulehčuje metaanalýzu GWAS u různých populací. Imputace genotypu je provedena statistickými metodami, kombinujícími data GWAS s referenčním panelem haplotypů. Tyto metody využívají výhodu sdílení haplotypů mezi jedinci přes krátké úseky sekvencí k doplnění alel. 194

195 Graf znázorňuje několik silně asociovaných podezřelých lokusů. Každý bod představuje SNP, osa X je genomická lokalizace, osa Y úroveň asociace. Příklad ukazuje asociaci pro mikrocirkulaci, body nahoře indikují genetické varianty, častěji nalézané u jedinců konstrikcemi v malých cévách. 195
196 Na předešlém obr. je tzv. graf Manhattan. Typ rozptýleného grafu, zobrazuje dvě proměnné pro soubor dat. U GWAS jsou na ose X genomické koordináty (chromosomy), na ose Y záporné logaritmy P hodnot pro asociaci každého SNP. Každý bod znázorňuje SNP. Nejsilnější asociace mají nejnižší P hodnotu, např , jejich záporný logaritmus je nejvyšší,
197 Asociace SNP s odhadnutými plemennými hodnotami pro procento mléčného tuku u 2327 jedinců německého holštýna 197
198 Detailní pohled na dvě genomické oblasti uvnitř známých QTL pro obsah tuku v mléce u skotu 198

199 Charakteristika nejvýznamnějších SNP z 50K Illumina BeadChip a dalších polymorfismů u čtyř major QTL pro obsah mléčného tuku u německého holštýna 199

200 Schematický pohled na QTL oblast BTA5 zahrnující EPS8 200
201 Schematický pohled na QTL oblast BTA27 zahrnující GPAT4 201

202 Kombinovaný vliv čtyř identifikovaných QTL na BTA5, 14, 20 a 27 na odhadnuté plemenné hodnoty pro obsah mléčného tuku u německého holštýna 202
203 Rozdělení genetické variance pro obsah mléčného tuku do 30 chromozómů a čtyři identifikované QTL oblasti na BTA5, 14, 20 a 27 u německého holštýna 203
204 GWAS většinou identifikují varianty s malými efekty 204
205 Ilustrace metody případové studie GWAS. Je vyhodnocen počet alel každého SNP, v tomto případě pomocí χ 2 testu, s cílem identifikovat varianty asociované se znakem. Studie koronární arteriální choroby (CAD) z roku 2007, jedinci s alelou G na SNP1 205 (rs ) byli více zastoupeni ve skupině pacientů s CAD ve srovnání s kontrolou.
206 Regionální asociační graf, ukazuje SNPs v oblasti receptoru LDL a jejich asociaci k hladině LDL cholesterolu. Graf je podobný typu Manhattan, znázorňuje ale kratší úsek genomu. Struktura haplobloku je znázorněna barevnou škálou, stupeň asociace je na ose Y. Bod znázorňující SNP rs SNP má nejvyšší pozici na ose Y, protože 206 vysvětluje určitý podíl variance v hladině LDL cholesterolu.
207 Pozor na populační stratifikaci: Přítomnost systematických rozdílů v populaci ve frekvencích alel mezi subpopulacemi pravděpodobně v důsledku rozdílného původu (různých předků), zvl. v kontextu asociačních studií. Populační stratifikace je rovněž nazývána populační strukturou. Např. v případových studiích může být nalezena asociace jako důsledek populační struktury, ne v důsledku vazby k lokusu způsobujícímu chorobu. 207
208 SNPs asociovaných s chorobami jsou tisíce. První studie u lidí v roce 2005, tzv. ARMD, degenerace žluté skvrny. Koronární srdeční choroba, diabetes 1. typu, 2. typu, reumatoidní artritida, Crohnova choroba, bipolární porucha, hypertenze. Dva trendy v metodice. 1/ stále větší soubory, až 200 tis. lidí. Cílem spolehlivé odhalení rizikových SNPs s nízkým OR (odds ratio, zvyšují pravděpodobnost choroby málo) a nízkou alelickou frekvenci. 2/ analýza přesněji definovaných fenotypů, např. hladina krevních lipidů, proinsulinu a podobných biomarkerů. Ty jsou naz. intermediární fenotypy, jejich analýzy doporučovány jako vhodné ve funkčním výzkumu biomarkerů metabolomika! Ústřední bod diskuse o GWAS: většina variant SNP je asociována jen s malým zvýšením rizika choroby a mají jen malou prediktivní hodnotu. Medián OR pro rizikový SNP je 1,33, jen málo SNP má OR nad 3,0. SNP nevysvětlují velký podíl dědičné variability, známé z odhadů heritability, založených u lidí na monozygotních dvojčatech. 208
209 Příklady provedených celogenomových asociačních analýz u skotu Všechny studie používaly Illumina BovSNP50 array (není-li uvedeno jinak). Studie jsou seskupeny volně podle vlastností skupin. 209
210 VLASTNOST ZDROJ PLEMENO POČET ZV. POČET VÝZN. SNP Somatic cell score Strillaci et al. (2014) Valdostana Red Pied 171 Plodnost samic Khatkar et al. (2014) různá Plodnost samic McDaneld et al. (2014) různá 210
211 VLASTNOST ZDROJ PLEMENO POČET ZV. POČET VÝZ. SNP Konverze krmiva Serao et al. (2013) Angus, Simmental 1321 Porodní hmotnost Utsunomiya et al. (2013) Nellore 654 býků Celoživotní počet telat na krávu Sasaki et al. (2013) Japanese black
212 ZNAK, VLASTNOST ZDROJ PLEMENO POČET VZORKŮ POČET VÝZNAM. SNPs KONTROLA V NEZÁVISLÉ POPULACI KOMENTÁŘ produkce mléka, ukazatele plodnosti Pryce et al. (2010a) Holštýn, Jersey (napříč 6 znaky) Ano, 544 SNPs u Holštýna, 159 u Jersey Testovány jednotlivé SNPs, stejně jako varhap perzistence laktace Pryce et al. (2010b) Holštýn, Jersey , při P<0,005 Ano odolnost vůči tepelnému stresu, schopnost dojit při nízké úrovni krmení Hayes et al. (2010) Holštýn, Jersey Ano produkce mléka, PSM Kolbehdari et al. (2009) Holštýn 28 Ne 1536 SNPs produkce mléka - interakce s úrovní krmení Lillehamme r et al. (2009) Holštýn 384 Ne 9918 SNPs 212
213 ZNAK, VLASTNOST ZDROJ PLEMENO POČET VZORKŮ POČET VÝZN. SNPs KONTROLA V NEZÁVISLÉ POPULACI KOMENTÁŘ produkce mléka Mai et al. (2010) Jersey Ne konformace zmasilost, funkční znaky Kolbehdari et al. (2008) Holštýn Ne 1036 SNPs v intronech plodnost, věk při nástupu puberty Fortes et al. (2010) tropická plemena skotu ne Použit nový multivarietní postup plodnost býků Feugang et al. (2009) Holštýn 20 býků vynikající plodnosti 97 Ano, 4 nejvíce signifikantní SNPs se testovaly u 210 býků s mimořádnou plodností, dva signifikantní, související s integrin beta 5 proteinem SNPs plodnost samic Sahana et al. (2010) Holštýn Ne 213
214 ZNAK, VLASTNOST ZDROJ PLEMENO POČET VZORKŮ POČET VÝZN. SNPs KONTROLA V NEZÁVISLÉ POPULACI KOMENTÁŘ plodnost, míra oplození, oplození, blastocysty Huang et al. (2010) Holštýn 27 Ne, ale souhrnné výsledky potvrzeny individuální genotypizací. První využití souhrnné DNA anylýzy pomocí GWAS u skotu mrtvě narozená mláďata, dystokie Olsen et al. (2010) Norská červinka Ano 17,343 SNPs snadnost telení Pausch. et al. (2010) Fleckvieh 1800 Ne dvojče Kim et al. (2009) Holštýn Ano, Birman et al. (2010). 55 SNPs ověřeno v nezávávislém souboru dat 921 holštýnských býků. Poslední set 18 SNPs vysvětluje 34% variabilitu ve výskytu dvojčat. složení mastných kyselin v hovězím mase Uemoto et al. (2010) Japonský černý skot 160 mimořádných zvířat 32 Ne 214
215 ZNAK, VLASTNOST ZDROJ PLEMENO POČET VZORKŮ POČET VÝZN. SNPs KONTROLA V NEZÁVISLÉ POPULACI KOMENTÁŘ odolnost vůči Johnově chorobě (posuzováno dle odolnosti vůči M. aviumpoddr. paratuberculosis - MAP infekce) Zanella et al. (2010) Holštýn 90 plus, 16 pozit., 25 kontrolních 5 SNPs Ne MAP - status Settles et al. (2009) Holštýn Ne MAP - status (ELISA) Pant et al. (2010) Holštýn 232 zvířat se známým MAP statusem 12 genomic. úseků Ne serologicky pozitivní (či neg.) na MAP (ELISA) Minnozi et al. (2010) Holštýn 483 MAP pozit., 483 negat. (ELISA) 4 úseky Ano, ověřeno 5 SNPs. vnímavost k BSE Murdoch et al. (2010) Holštýn 481 polosourozenců, 149 BSE pozit., 184 kont. Vzorků 27 SNPs Ne Použito 3072 SNPs 215
216 ZNAK, VLASTNOST ZDROJ PLEMENO POČET VZORKŮ růst Snelling et. al (2010) Angus, Charolais, Gelbvieh, Hereford, Limousin, Red Angus, Simentál POČET VÝZN. SNPs KONTROLA V NEZÁVISLÉ POPULACI SNPs Ne KOMENTÁŘ využití krmiva Barendse et al. (2007) Angus, Brahman, Belmont, Hereford, S. Gert., Shorthorn 189 mimořádných zvířat 161 SNPs při P<0,01 Ano využití krmiva Sherman et al. (2010) Angus, Charolais, Alberta (hybrid) při P<0,01 Ne Použito 2633 SNPs odolnost k tick burden Turner et al. (2010) Jersey, Holštýn, Aussie Red, Sahiwal, Illawara, Shorthorn 189 mimořádných zvířat 27 při P<0,05 Ne Použito 7397 SNPs. Signifikantní SNPs blízko u Integrin alpha 11 genu v navaz. studii. 216
217 6. 217
218 Ještě pár poznámek 218
219 Customer chips 219

220 220

221 221

222 222

223 223

224 224

225 225

226 226

227 227

228 228
Genomika hospodářských zvířat
 Genomika hospodářských zvířat 1 1. 2 Mapování QTL pomocí vazbové nerovnováhy Linkage Disequilibrium (LD) 3 Mapování QTL pomocí vazbové nerovnováhy založeno na existenci LD mezi QTL a markerem v náhodně
Genomika hospodářských zvířat 1 1. 2 Mapování QTL pomocí vazbové nerovnováhy Linkage Disequilibrium (LD) 3 Mapování QTL pomocí vazbové nerovnováhy založeno na existenci LD mezi QTL a markerem v náhodně
Genomika hospodářských zvířat
 Genomika hospodářských zvířat 1 Mammalian genome ~3 x 10 9 base pairs Unique genes (protein-coding and RNA genes) ~1% Repetitive sequences ~60-65% Intergenic unique DNA ~35-40% Genes ~
Genomika hospodářských zvířat 1 Mammalian genome ~3 x 10 9 base pairs Unique genes (protein-coding and RNA genes) ~1% Repetitive sequences ~60-65% Intergenic unique DNA ~35-40% Genes ~
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Genetika kvantitativních znaků. - principy, vlastnosti a aplikace statistiky
 Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Základy genetiky populací
 Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Thomas Hunt Morgan ( ) americký genetik a embryolog pokusy s octomilkou (D. melanogaster)
 americký genetik a embryolog pokusy s octomilkou (D. melanogaster)") Vazba vloh Thomas Hunt Morgan (1866 1945) americký genetik a embryolog pokusy s octomilkou (D. melanogaster) Morganova pravidla 1. geny jsou na chromozómu uspořádány lineárně za sebou 2. počet vazbových
Vazba vloh Thomas Hunt Morgan (1866 1945) americký genetik a embryolog pokusy s octomilkou (D. melanogaster) Morganova pravidla 1. geny jsou na chromozómu uspořádány lineárně za sebou 2. počet vazbových
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Příbuznost a inbreeding
 Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Konzervační genetika INBREEDING. Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.
 Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Aktuality ke genotypování a ověřování původů dle SNP. Josef Kučera
 Aktuality ke genotypování a ověřování původů dle SNP Josef Kučera Obsah igenetika Aktuální stav genotypování Ověřování původů (parentage verification) Dohledání původu (parentage discovery) Ukázky problémových
Aktuality ke genotypování a ověřování původů dle SNP Josef Kučera Obsah igenetika Aktuální stav genotypování Ověřování původů (parentage verification) Dohledání původu (parentage discovery) Ukázky problémových
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
KBI / GENE Mgr. Zbyněk Houdek
 Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Zdeňka Veselá Tel.: Výzkumný ústav živočišné výroby, v.v.i.
 BIOTECHNOLOGICKÉ METODY VE ŠLECHTĚNÍ HOSPODÁŘSKÝCH ZVÍŘAT Zdeňka Veselá vesela.zdenka@vuzv.cz Tel.: 267009571 Výzkumný ústav živočišné výroby, v.v.i. Co je šlechtění? Soustavné zlepšování genetických schopností
BIOTECHNOLOGICKÉ METODY VE ŠLECHTĚNÍ HOSPODÁŘSKÝCH ZVÍŘAT Zdeňka Veselá vesela.zdenka@vuzv.cz Tel.: 267009571 Výzkumný ústav živočišné výroby, v.v.i. Co je šlechtění? Soustavné zlepšování genetických schopností
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Cvičení č. 8. KBI/GENE Mgr. Zbyněk Houdek
 Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Experiment s dlouhodobou selekcí krav na ukazatele produkce a zdravotního stavu v Norsku Ing. Pavel Bucek, Českomoravská společnost chovatelů, a.s.
 Experiment s dlouhodobou selekcí krav na ukazatele produkce a zdravotního stavu v Norsku Ing. Pavel Bucek, Českomoravská společnost chovatelů, a.s. Z chovatelské praxe a z celé řady vědeckých experimentů
Experiment s dlouhodobou selekcí krav na ukazatele produkce a zdravotního stavu v Norsku Ing. Pavel Bucek, Českomoravská společnost chovatelů, a.s. Z chovatelské praxe a z celé řady vědeckých experimentů
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Z D E Ň K A V E S E L Á, V E S E L A. Z D E N K V U Z V. C Z
 HYBRIDIZACE ZD E Ň K A V E S E LÁ, V E S E LA. ZD E N K A @ V U Z V. C Z DVĚ METODY DOCÍLENÍ ŠLECHTITELSKÉHO POKROKU 1. Využití genetické proměnlivosti mezi jedinci uvnitř populace Selekce Záměrné připařování
HYBRIDIZACE ZD E Ň K A V E S E LÁ, V E S E LA. ZD E N K A @ V U Z V. C Z DVĚ METODY DOCÍLENÍ ŠLECHTITELSKÉHO POKROKU 1. Využití genetické proměnlivosti mezi jedinci uvnitř populace Selekce Záměrné připařování
Genetické markery. pro masnou produkci. Mgr. Jan Říha. Výzkumný ústav pro chov skotu, s.r.o.
 Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Hodnocení plemenných + chovných + užitkových prasat
 Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
Genomika hospodářských zvířat
 Genomika hospodářských zvířat QTL pro rezistenci a/nebo citlivost vůči leishmanióze a jiným infekcím u myší QTL pro hybridní sterilitu samců u myší White et al. 2011, Genetics Jemné genetické mapování
Genomika hospodářských zvířat QTL pro rezistenci a/nebo citlivost vůči leishmanióze a jiným infekcím u myší QTL pro hybridní sterilitu samců u myší White et al. 2011, Genetics Jemné genetické mapování
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY
 GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
1. Definice a historie oboru molekulární medicína. 3. Základní laboratorní techniky v molekulární medicíně
 Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Nauka o dědičnosti a proměnlivosti
 Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Předpověď plemenné hodnoty Něco málo z praxe. Zdeňka Veselá
 Předpověď plemenné hodnoty Něco málo z praxe Zdeňka Veselá vesela.zdenka@vuzv.cz Příprava datových souboru Databáze s výsledky užitkovosti jsou zpravidla obrovské soubory Např. kontrola užitkovosti masného
Předpověď plemenné hodnoty Něco málo z praxe Zdeňka Veselá vesela.zdenka@vuzv.cz Příprava datových souboru Databáze s výsledky užitkovosti jsou zpravidla obrovské soubory Např. kontrola užitkovosti masného
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
Úvod do nonhla-dq genetiky celiakie
 Úvod do nonhla-dq genetiky celiakie František Mrázek HLA laboratoř, Ústav Imunologie LF UP a FN Olomouc Celiakie - časté chronické zánětlivé onemocnění tenkého střeva s autoimunitní a systémovou složkou
Úvod do nonhla-dq genetiky celiakie František Mrázek HLA laboratoř, Ústav Imunologie LF UP a FN Olomouc Celiakie - časté chronické zánětlivé onemocnění tenkého střeva s autoimunitní a systémovou složkou
Heritabilita. Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability
 Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Výzkumné centrum genomiky a proteomiky. Ústav experimentální medicíny AV ČR, v.v.i.
 Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
8.2 PLEMENÁŘSKÁ PRÁCE V CHOVU SKOTU
 8.2 PLEMENÁŘSKÁ PRÁCE V CHOVU SKOTU A. SELEKCE SKOTU Zákon č. 154/2000Sb O šlechtění,plemenitbě a evidenci hospodářských zvířat ( plemenářský zákon ) Organizace zabývající se šlechtěním: Plemo a.s., ČMSCH
8.2 PLEMENÁŘSKÁ PRÁCE V CHOVU SKOTU A. SELEKCE SKOTU Zákon č. 154/2000Sb O šlechtění,plemenitbě a evidenci hospodářských zvířat ( plemenářský zákon ) Organizace zabývající se šlechtěním: Plemo a.s., ČMSCH
MTS spol. s r.o. NEJLEPŠÍ GENETIKA NEJLEPŠÍ KNOW-HOW Už více jak 20 let
 MTS spol. s r.o. NEJLEPŠÍ GENETIKA NEJLEPŠÍ KNOW-HOW Už více jak 20 let Mezníky před zavedením do praxe. Beckman a Soller (1983) - genetické markery bude moţné vyuţít v praxi plemenářské práce Lande a
MTS spol. s r.o. NEJLEPŠÍ GENETIKA NEJLEPŠÍ KNOW-HOW Už více jak 20 let Mezníky před zavedením do praxe. Beckman a Soller (1983) - genetické markery bude moţné vyuţít v praxi plemenářské práce Lande a
Genotypování markerů užitkovosti a zdraví u skotu
 Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
Mezinárodní odborný seminář Využití chovatelských dat onemocnění skotu pro management stád, šlechtění a pro racionální užívání antimikrobik. Genotypování markerů užitkovosti a zdraví u skotu Jitka Kyseľová
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Selekce. Zdeňka Veselá
 Selekce Zdeňka Veselá vesela.zdenka@vuzv.cz Selekce Cílem změna genetické struktury populace 1. Přírodní 2. Umělá Selekce 1. Direkcionální (směrovaná) Nejčastější Výsledkem je posun střední hodnoty populace
Selekce Zdeňka Veselá vesela.zdenka@vuzv.cz Selekce Cílem změna genetické struktury populace 1. Přírodní 2. Umělá Selekce 1. Direkcionální (směrovaná) Nejčastější Výsledkem je posun střední hodnoty populace
NGS analýza dat. kroužek, Alena Musilová
 NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
NGS analýza dat kroužek, 16.12.2016 Alena Musilová Typy NGS experimentů Název Materiál Cílí na..? Cíl experimentu? amplikon DNA malý počet vybraných genů hledání variant exom DNA všechny geny hledání
Popis modelu pro odhady PH mléčné užitkovosti
 Popis modelu pro odhady PH mléčné užitkovosti Zvířata zařazená do hodnocení V modelu plemene H jsou hodnoceny krávy s podílem krve H nebo 75% a výše. V modelu plemene C jsou hodnoceny krávy s podílem krve
Popis modelu pro odhady PH mléčné užitkovosti Zvířata zařazená do hodnocení V modelu plemene H jsou hodnoceny krávy s podílem krve H nebo 75% a výše. V modelu plemene C jsou hodnoceny krávy s podílem krve
FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ
 FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ SARKOIDÓZOU: POTVRZENÍ VÝSLEDKŮ CELOGENOMOVÉ ASOCIAČNÍ STUDIE. Sťahelová A. 1, Mrázek F. 1, Kriegová E. 1, Hutyrová B. 2, Kubištová Z. 1, Kolek V.
FUNKČNÍ VARIANTA GENU ANXA11 SNIŽUJE RIZIKO ONEMOCNĚNÍ SARKOIDÓZOU: POTVRZENÍ VÝSLEDKŮ CELOGENOMOVÉ ASOCIAČNÍ STUDIE. Sťahelová A. 1, Mrázek F. 1, Kriegová E. 1, Hutyrová B. 2, Kubištová Z. 1, Kolek V.
2 Inkompatibilita v systému Rhesus. Upraveno z A.D.A.M.'s health encyclopedia
 2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK
 ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
Biologie a genetika, BSP, LS7 2014/2015, Ivan Literák
 Biologie a genetika, BSP, LS7 2014/2015, Ivan Literák KVANTITATIVNÍ GENETIKA dědičnost kvantitativních znaků ZNAKY KVALITATIVNÍ: gen znak barva hrachu: žlutá zelená (i komplikovaněji penetrace, epresivita,
Biologie a genetika, BSP, LS7 2014/2015, Ivan Literák KVANTITATIVNÍ GENETIKA dědičnost kvantitativních znaků ZNAKY KVALITATIVNÍ: gen znak barva hrachu: žlutá zelená (i komplikovaněji penetrace, epresivita,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele)
") Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Genetika přehled zkouškových otázek:
 Genetika přehled zkouškových otázek: 1) Uveďte Mendelovy zákony (pravidla) dědičnosti, podmínky platnosti Mendelových zákonů. 2) Popište genetický zápis (mendelistický čtverec) monohybridního křížení u
Genetika přehled zkouškových otázek: 1) Uveďte Mendelovy zákony (pravidla) dědičnosti, podmínky platnosti Mendelových zákonů. 2) Popište genetický zápis (mendelistický čtverec) monohybridního křížení u
Jak měříme genetickou vzdálenost a co nám říká F ST
 Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Jana Vránová, 3. lékařská fakulta UK
 Jana Vránová, 3. lékařská fakulta UK Vznikají při zkoumání vztahů kvalitativních resp. diskrétních znaků Jedná se o analogii s korelační analýzou spojitých znaků Přitom předpokládáme, že každý prvek populace
Jana Vránová, 3. lékařská fakulta UK Vznikají při zkoumání vztahů kvalitativních resp. diskrétních znaků Jedná se o analogii s korelační analýzou spojitých znaků Přitom předpokládáme, že každý prvek populace
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE
 ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
ÚVOD DO TRANSPLANTAČNÍ IMUNOLOGIE Základní funkce imunitního systému Chrání integritu organizmu proti škodlivinám zevního a vnitřního původu: chrání organizmus proti patogenním mikroorganizmům a jejich
Proč nový systém odhadu plemenných hodnot?
 Proč nový systém odhadu plemenných hodnot? Faktory ovlivňující užitkovost Chovatel Výživa Prostředí Užitkovost Genetická výbava 5-15% Zisk chovatele Spotřebitel Hodnocení zvířat 1900 KU 1920 vývoj metod
Proč nový systém odhadu plemenných hodnot? Faktory ovlivňující užitkovost Chovatel Výživa Prostředí Užitkovost Genetická výbava 5-15% Zisk chovatele Spotřebitel Hodnocení zvířat 1900 KU 1920 vývoj metod
- Definice inbreedingu a jeho teorie
 Negativní důsledky inbrední deprese v chovu skotu Ing. Jiří Bezdíček, Ph.D. Výzkumný ústav pro chov skotu, s.r.o., Rapotín 26. listopadu 2009 - Definice inbreedingu a jeho teorie - Proč je inbreeding v
Negativní důsledky inbrední deprese v chovu skotu Ing. Jiří Bezdíček, Ph.D. Výzkumný ústav pro chov skotu, s.r.o., Rapotín 26. listopadu 2009 - Definice inbreedingu a jeho teorie - Proč je inbreeding v
Zdeňka Veselá Tel.: Výzkumný ústav živočišné výroby, v.v.i.
 ZÁKLADNÍ ZÁSADY ŠLECHTĚNÍ Zdeňka Veselá vesela.zdenka@vuzv.cz Tel.: 267009571 Výzkumný ústav živočišné výroby, v.v.i. Význam šlechtění Nejefektivnější činnost chovatele Soustavné zlepšování genetických
ZÁKLADNÍ ZÁSADY ŠLECHTĚNÍ Zdeňka Veselá vesela.zdenka@vuzv.cz Tel.: 267009571 Výzkumný ústav živočišné výroby, v.v.i. Význam šlechtění Nejefektivnější činnost chovatele Soustavné zlepšování genetických
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Selekční indexy v praxi. Josef Kučera
 Selekční indexy v praxi Josef Kučera Selekce Cílem selekce je výběr zvířat k produkci potomstva pro obměnu stáda nebo v celé populaci k produkci další generace zvířat na všech úrovních šlechdtelského programu
Selekční indexy v praxi Josef Kučera Selekce Cílem selekce je výběr zvířat k produkci potomstva pro obměnu stáda nebo v celé populaci k produkci další generace zvířat na všech úrovních šlechdtelského programu
Základní pravidla dědičnosti
 Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Základy biostatistiky II. Veřejné zdravotnictví 3.LF UK - II
 Základy biostatistiky II Veřejné zdravotnictví 3.LF UK - II Teoretické rozložení-matematické modely rozložení Naměřená data Výběrové rozložení Teoretické rozložení 1 e 2 x 2 Teoretické rozložení-matematické
Základy biostatistiky II Veřejné zdravotnictví 3.LF UK - II Teoretické rozložení-matematické modely rozložení Naměřená data Výběrové rozložení Teoretické rozložení 1 e 2 x 2 Teoretické rozložení-matematické
Genetika vzácných druhů zuzmun
 Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Informační a znalostní systémy
 Informační a znalostní systémy Teorie pravděpodobnosti není v podstatě nic jiného než vyjádření obecného povědomí počítáním. P. S. de Laplace Pravděpodobnost a relativní četnost Pokusy, výsledky nejsou
Informační a znalostní systémy Teorie pravděpodobnosti není v podstatě nic jiného než vyjádření obecného povědomí počítáním. P. S. de Laplace Pravděpodobnost a relativní četnost Pokusy, výsledky nejsou
Vrozené vývojové vady, genetika
 UNIVERZITA KARLOVA V PRAZE Fakulta tělesné výchovy a sportu Vrozené vývojové vady, genetika studijní opora pro kombinovanou formu studia Aplikovaná tělesná výchova a sport Doc.MUDr. Eva Kohlíková, CSc.
UNIVERZITA KARLOVA V PRAZE Fakulta tělesné výchovy a sportu Vrozené vývojové vady, genetika studijní opora pro kombinovanou formu studia Aplikovaná tělesná výchova a sport Doc.MUDr. Eva Kohlíková, CSc.
INDUKTIVNÍ STATISTIKA
 10. SEMINÁŘ INDUKTIVNÍ STATISTIKA 3. HODNOCENÍ ZÁVISLOSTÍ HODNOCENÍ ZÁVISLOSTÍ KVALITATIVNÍ VELIČINY - Vychází se z kombinační (kontingenční) tabulky, která je výsledkem třídění druhého stupně KVANTITATIVNÍ
10. SEMINÁŘ INDUKTIVNÍ STATISTIKA 3. HODNOCENÍ ZÁVISLOSTÍ HODNOCENÍ ZÁVISLOSTÍ KVALITATIVNÍ VELIČINY - Vychází se z kombinační (kontingenční) tabulky, která je výsledkem třídění druhého stupně KVANTITATIVNÍ
ší šířen VAZEBNÁ ANALÝZA Vazba genů
 VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
VAZEBNÁ ANALÝZA Vazba genů Americký genetik Thomas Morgan při genetických pokusech s octomilkami (Drosophila melanogaster) popsal zákonitosti o umístění genů na chromosomech, které existují až do současnosti
Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).
: RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).") Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
VYBRANÉ GENETICKÉ ÚLOHY II.
 VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které
VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které
Hodnocení plemenných + chovných + užitkových prasat
 Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
Hodnocení plemenných + chovných + užitkových prasat Metodické pokyny SCHP Hodnocení plemenných prasat Cíl hodnocení stanovit předpoklad využití zvířat v plemenitbě k dalšímu šlechtění populace k masovému
Selekční efekt. Úvod do šlechtění zvířat 1
 Selekční efekt Úvod do šlechtění zvířat 1 Dědičnost tělesné výšky u lidí ( F. Galton 1800-1911) Generac Odchylky od průměru populace e Rodičů -6,0-4,5-3,0-1,5 0 +1,5 +3,0 +4,5 +5,0 Potomků -4,0-2,5-1,5-1,0
Selekční efekt Úvod do šlechtění zvířat 1 Dědičnost tělesné výšky u lidí ( F. Galton 1800-1911) Generac Odchylky od průměru populace e Rodičů -6,0-4,5-3,0-1,5 0 +1,5 +3,0 +4,5 +5,0 Potomků -4,0-2,5-1,5-1,0
II. ročník, zimní semestr 1. týden OPAKOVÁNÍ. Úvod do POPULAČNÍ GENETIKY
 II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
II. ročník, zimní semestr 1. týden 6.10. - 10.10.2008 OPAKOVÁNÍ Úvod do POPULAČNÍ GENETIKY 1 Informace o výuce (vývěska) 2 - nahrazování (zcela výjimečně) - podmínky udělení zápočtu (docházka, prospěch
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Aplikace molekulárně- genetických dat ve šlechtění. Humpolíček Petr
 Aplikace molekulárně- genetických dat ve šlechtění Humpolíček Petr Co vás čeká Tradiční šlechtitelské postupy Typy molek-gen. dat Přístupy k aplikaci molek-gen. dat Srovnání přístupů Šlechtění na odolnost
Aplikace molekulárně- genetických dat ve šlechtění Humpolíček Petr Co vás čeká Tradiční šlechtitelské postupy Typy molek-gen. dat Přístupy k aplikaci molek-gen. dat Srovnání přístupů Šlechtění na odolnost
JEDNOVÝBĚROVÉ TESTY. Komentované řešení pomocí programu Statistica
 JEDNOVÝBĚROVÉ TESTY Komentované řešení pomocí programu Statistica Vstupní data Data umístěná v excelovském souboru překopírujeme do tabulky ve Statistice a pojmenujeme proměnné, viz prezentace k tématu
JEDNOVÝBĚROVÉ TESTY Komentované řešení pomocí programu Statistica Vstupní data Data umístěná v excelovském souboru překopírujeme do tabulky ve Statistice a pojmenujeme proměnné, viz prezentace k tématu
Hardy-Weinbergův zákon - cvičení
 Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
GENETIKA Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální dědičnost
 Polygenní dědičnost Multifaktoriální dědičnost") GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ. I. Šubrt Společnost lékařské genetiky ČLS JEP
 EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
Možnosti selekce na zlepšenou konverzi krmiva u ovcí
 Možnosti selekce na zlepšenou konverzi krmiva u ovcí Ing. Pavel Bucek, Českomoravská společnost chovatelů, a. s. Příspěvek byl publikován v upravené podobě v časopise Farmář Z praxe a celé řady výzkumných
Možnosti selekce na zlepšenou konverzi krmiva u ovcí Ing. Pavel Bucek, Českomoravská společnost chovatelů, a. s. Příspěvek byl publikován v upravené podobě v časopise Farmář Z praxe a celé řady výzkumných
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Popis modelu pro odhady PH mléčné užitkovosti
 Plemdat, s.r.o. 2.4.206 Popis modelu pro odhady P mléčné užitkovosti vířata zařazená do hodnocení V modelu plemene jsou hodnoceny krávy s podílem krve nebo 75% a výše. Krávy s podílem krve masného plemene
Plemdat, s.r.o. 2.4.206 Popis modelu pro odhady P mléčné užitkovosti vířata zařazená do hodnocení V modelu plemene jsou hodnoceny krávy s podílem krve nebo 75% a výše. Krávy s podílem krve masného plemene
4. Genetické základy šlechtění hospodářských zvířat
 část 3. (rough draft version) 4. Genetické základy šlechtění hospodářských zvířat Obecné principy postupu šlechtění Šlechtění je ekonomicky výhodnější než prostá produkce živočišných produktů ať v krátkodobém
část 3. (rough draft version) 4. Genetické základy šlechtění hospodářských zvířat Obecné principy postupu šlechtění Šlechtění je ekonomicky výhodnější než prostá produkce živočišných produktů ať v krátkodobém