Charles University, Prague Faculty of Science. Ph.D. Thesis Adriana Jelínková
|
|
|
- Přemysl Vacek
- před 5 lety
- Počet zobrazení:
Transkript
1 Charles University, Prague Faculty of Science Ph.D. Thesis 2010 Adriana Jelínková
2

3 Cytoskeleton and endocytosis, a dynamic system for the localization of auxin efflux carriers A dissertation presented to the Department of Experimental Plant Biology, Faculty of Science Charles University in partial fulfilment of the requirements for the PhD degree Adriana Jelínková Charles University Prague 2010
4
5 SUPERVISOR Jan Petrášek, Ph.D. Institute of Experimental Botany ASCR, Prague, Czech Republic Department of Plant Physiology, Faculty of Science, Charles University, Prague, Czech Republic CO-SUPERVISOR Ass. Prof. Dr. Eva Zaţímalová Institute of Experimental Botany ASCR, Prague, Czech Republic Department of Plant Physiology, Faculty of Science, Charles University, Prague, Czech Republic This is to certify that this thesis is not a subject of any other defending procedure. It contains set of original results, some of them have been published in the international peer reviewed scientific journals. Prague Adriana Jelínková On behalf of the co-authors of the papers published, I hereby confirm the agreement with inclusion of the papers below into the dissertation thesis of Adriana Jelínková. The papers were produced as a team work and the particular contribution of Adriana Jelínková is specified at the beginning of relevant result blocks of the thesis. Prague Jan Petrášek
6
7 Acknowledgements Mé srdečné díky patří J.P. za vše co se mého vědeckého působení týče. Velmi děkuji E.Z. a také celému laboratornímu týmu (K.M., M.P., J.Ţ., D.S., P.S., P.K., S.S., M.Č., M.L., K.H., P.K.,M.K., L.K. a ) za podporu, pomoc a pochopení. Gracias por todo C. de la T., S. de la E., M., A.C., P.A., J.G., E.A. y R.S. Thaknk you D.M. S láskou děkuji rodině. This work was supported by several local and international grant projects: Grant Agency of Charles University (grant number Grant Agency of the Academy of Sciences of theczech Republic (grant number KJB ) Grant from the Ministry of Education, Youth and Sports of the Czech Republic (LC06034) European Fund for Regional Development, the Operational Programme Prague- Competitiveness, project n: CZ.216/3.1.00/21159.
8
9 Contents Abstract 1 Abstrakt 3 1 Introduction Polarity in plants Auxin and the polarity of plants Auxin action Auxin as a morphogen Auxin transport Mechanisms of protein targeting via vesicle transport Vesicle trafficking processes in plants Endosomal space and recycling of the plasma membrane proteins Dynamics of the auxin transporters in the endosomal space Cytoskeleton and endomembrane traffic interactions in the formation of auxin gradients Cytoskeleton, a dynamic network of proteins Microtubules Actin filaments Examples of MTs and AFs interactions Cytoskeleton in the vesicle trafficking Cytoskeleton in the targeting of auxin transporters 19 2 Objectives and outlines 23 3 Results Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Addresing the role of cytoskeleton in the endocytosis of PIN1 auxin efflux carrier in BY-2 cells Probing plant membranes with FM dyes: tracking, dragging or blocking? Cytoskeletal proteins mediating targeted deposition of auxin transporters 51 4 Materials and Methods 71 5A Conclusions and Prospects 75 5B Výsledky a závěry 77 6 References 79 Addenda 89 Curriculum vitae 91
10
11 Abstract Although the participation membrane trafficking processes in the positioning of auxin carriers that direct auxin flow is studied already several years, there is still a significant lack of the information on the mechanism of cytoskeleton action in these processes. Auxin efflux carriers undergo constitutive cycling in the endosomal space of plant cell and this process is crucial in their dynamic positioning to various domains in the plasma membrane. This process constitutes the important background for the transduction of developmental cues leading to the directional auxin transport. One of the most frequent experimental approaches to study this issue includes the usage of inhibitors of membrane trafficking processes and cytoskeleton dynamics. Indeed, the disruption of targeting of auxin efflux carriers to the plasma membrane and reduction of the auxin efflux has been reported for both the inhibitors actin cytoskeleton and membrane trafficking. In this thesis, it is demonstrated using tobacco BY-2 cells that short treatment with brefeldin A (BFA), the inhibitor of anterograde protein trafficking, induced remodelling of actin filaments around BFA-induced endomembrane compartments. This suggests the connection between actin filaments and endomembrane dynamics. To dissect the requirement of cytoskeleton and endocytosis for the trafficking of auxin efflux carriers, spectrum of inhibitors of protein trafficking was used and their effect on FM 4-64 endocytic tracer uptake and auxin efflux carrier PIN1 followed in BY-2. After the disruption of both actin and microtubular cytoskeleton, endocytosis of FM 4-64 has been blocked. Moreover, the importance of actin cytoskeleton for the trafficking processes in the endosomal space was uncovered. During these observations, the unexpected effect of FM 4-64 in-vivo endocytosis marker on the distribution of plasma membrane proteins was characterized. To obtain comprehensive in planta data on the role of cytoskeleton in the processes of protein targeting and auxin transport, a microscopical phenotypic screen of all available Arabidopsis thaliana mutants in the cytoskeletal proteins and their important interactors was initiated. The aim is to identify molecular players linking the cytoskeleton, endocytosis and targeting of auxin efflux carriers. In the screen, the rate of endocytosis is tested by FM 4-64 uptake in the root epidermal and cortex cells and concurrently the localization of PIN1 and PIN2 is determined using indirect immunofluorescence staining. In this thesis, the first part of the screen focused on the actin cytoskeleton is presented. 1
12 2
13 Abstrakt Ačkoliv je zapojení endomembránového systému v procesech cíleného umísťování auxinových přenašečů směrujících tok auxinu studováno jiţ řadu let, není role cytoskeletu v těchto procesech zcela zřejmá. Přenašeče auxinu podléhají konstitutivnímu cyklování v endosomálním prostoru rostlinných buněk. Tento proces je klíčový pro jejich depozici do různých domén plazmatické membrány a představuje důleţitý základ pro přenos vývojových signálů zprostředkovaných směrovaným tokem auxinu. Mezi nejčastější experimentální přístupy pro studium této problematiky patří vyuţití inhibitorů dynamiky endomembránového systému a cytoskeletu. V několika studiích byl skutečně popsán jejich inhibiční účinek na distribuci auxinových přenašečů do oblasti plazmatické membrány a následné sníţení transportu auxinu z buněk. V této disertační práci je demonstrováno, ţe v tabákových buňkách BY-2 dochází po krátkodobém působení inhibitoru anterográdního transportu brefeldinu A (BFA) k přestavbě aktinového cytoskeletu a jeho akumulace okolo endomembránových shluků vyvolaných působením BFA, coţ naznačuje propojení mezi aktinovým cytoskeletem a endomembránovým systémem. K prohloubení znalostí o účasti cytoskeletu a endocytózy v procesech cyklování auxinových přenašečů bylo testováno spektrum inhibitorů endomembránové dynamiky v buněčných liniích tabáku BY-2. Byl sledován vliv těchto inhibitorů na endocytózu barvy FM 4-64 a na lokalizaci přenašeče auxinu ven z buňky PIN1. Po narušení aktinového nebo mikrotubulárního cytoskeletu došlo k inhibici endocytózy FM Důleţitost aktinového cytoskeletu byla navíc prokázána i pro dynamiku membránového transportu v endosomálním prostoru buněk. Během těchto experimentů byl pozorován a následně charakterizován neočekávaný účinek barvy FM 4-64 na umístění proteinů v plazmatické membráně. K získání ucelenější představy o roli cytoskeletu v procesech umísťování auxinových přenašečů a transportu auxinu v celistvé rostlině byla zahájena fenotypová studie všech dostupných mutantních linií v genech kódujících cytoskeletální a asociované proteinů v huseníčku (Arabidopsis thaliana). Cílem této studie je identifikovat proteiny propojující dynamiku cytoskeletu a váčkového transportu se zvláštním ohledem na procesy polárního umísťování auxinových přenašečů. Testována je rychlost endocytózy FM 4-64 v pokoţkových buňkách a v buňkách primární kůry kořene spolu s lokalizací proteinů PIN1 a PIN2 v buňkách středního válce značených pomocí nepřímé imunofluorescence. V této disertační práci je prezentována první část této studie zaměřená na aktinový cytoskelet. 3
14 4
15 1 Introduction 1.1 Polarity in plants Similar as in other multicellular organisms, the processes of cell division, growth and differentiation determine the character of three-dimensional formation of plant body. The polarity on the multicellular level is mimicked by the polarity of its individual cells. In turn, the polarity of single cell reflects the asymmetrical distribution of its intracellular components (reviewed in Feraru and Friml 2008). Thanks to the sessile life strategy of plants and the absence of cell motility, the orientation of cell division and expansion is crucial in the organogenesis and the formation of their overall habitus. The unique symmetrical and asymmetrical shapes are established in early embryogenesis and display plasticity through the ontogenesis. During the plant body plan formation, centres of symmetry and polarity axes are assigned through the perception of diverse intracellular and extracellular signals. The plant body is highly polarized in the shoot-root axis. This feature of the plant body was for the first time introduced by Hermann von Vöchting in 1878, where was shown, that pieces of willow (Salix species) stems form roots and shoots at the corresponding ends irrespective of the gravity vector (reviewed in Mohr and Schopfer, 1995). In many higher plants the asymmetrical (and thus polar) organization appears as early as in the non-random distribution of cytoplasmic components of the unfertilized egg cell, which is followed by fertilization and formation of asymmetrical zygote. During embryonic development asymmetrical division of zygote establishes the apical-basal axis. Followed by other sets of divisions the pattern of future plant body is recognizable as three major domains, shoot apex with cotyledons, hypocotyl and primary root. During postembryonic development the organogenesis of stem, leaves, roots and reproductive organs is accomplished (Goldberg et al. 1994). All these different forms of plant body arising by symmetrical and asymmetrical cell growth and division are based on the gradients of signalling molecules within the single cell, cell to cell communication and specific developmental and environmental signals. In the favorite model of Arabidopsis thaliana there are several situations, where the phenomenon of polarity establishment and maintenance might be studied. These are tip growth of pollen tube and root hairs, diffuse elongation growth of hypocotyl cells root cells, planar polarity initiation and maintenance in the positioning of epidermal cells forming trichomes, stomatal guard cells or root hairs and formation of cells with complex shapes such lobed pavement epidermal leaf cells (reviewed by Grebe et al. 2001, Boutte et al. 2007, Yang 2008). 5
16 1.2 Auxin and the polarity of plants Auxin action The auxin, represented by its prevalent form indole-3-acetic acid (IAA), is the key endogenous plant hormone regulating many aspects of plant growth and development. Auxin controls and moderates cell expansion, division and changes in developmental fate depending on cell position within the plant body (reviewed by Teale et al. 2006, Benjamins and Sheres 2008). It induces both fast responses without gene expression involved and specific gene expression. Fast nongenomic effects (generally changes in the flux of ions across plasma membrane) are seemingly connected with the action AUXIN BINDING PROTEIN 1 (ABP1) at the plasma membrane (Dahlke et al. 2010), other well described non-genomic effect is the influence of auxin on the endocytosis (see chapter 1.3.3). ABP1 was discovered as a protein with high affinity binding to auxin (Venis and Napier 1995). It is the glycoprotein with no transmembrane domain, located at the plasma membrane and in the lumen of the endoplasmic reticulum (Henderson 1997). The fact that ABP1 is ubiquitous in green plants and that the Arabidopsis abp1 null mutant is lethal at the embryonic stage prompts its high importance for plant development (Chen 2001). ABP1 can bind auxin in the extracellular space thus triggering hyperpolarization of the plasma membrane (Napier 2002). At the genomic level, auxin regulates transcription by rapidly modulating levels of Auxin/indole-3-acetic acid (Aux/IAA) transcription factors (repressors) throughout various stages of plant development. IAA binds to its binding protein TRANSPORT INHIBITOR RESPONSE 1 (TIR1), the F-box subunit of the ubiquitin ligase complex Skp-Cdc53-F-Box- Transport Inhibitor Response 1 (SCF TIR1 ) and thus stabilizes the interaction between TIR1 and Aux/IAA substrates (Dharmasiri et al. 2005, Kepinski and Leyser 2005). Based on the determination of TIR1 crystal structure it seems that IAA does not induce conformational changes but rather functions as a molecular glue between Aux/IAA and TIR1 (Tan et al. 2007). This interaction promotes Aux/IAA ubiquitination and subsequent proteasomemediated degradation and release of transcription factors form Auxin Response Factors (ARFs) family. Auxin-specific gene expression follows. In another words auxin concentration determines ARF activity by modifying the level of AUX/IAA protein. The network of auxin activity is very diverse and complex due to the existence of extensive gene families of three major components, TIR1 and other AUXIN SIGNALLING F-BOX (AFB) receptors, Aux/IAA and ARFs transcriptional factors, their expression patterns, post-translational modifications and protein-protein interactions (reviewed by Paciorek and Friml 2006, Lokerse and Weijers 2009, Kieffer et al. 2010), see also Fig. 1. 6

17 Figure 1 The scheme of auxin signalling. The scheme is reproduced from Paciorek and Friml Auxin as a morphogen In 1952 Turing introduced the term morphogen as the hypothetical substance, whose concentration gradients govern the patterning of tissues throught development. He showed on the model of regeneration of Hydra and a pattern of whorled leaves that chemical substances (morphogens), could self-organize into spatial patterns, starting from homogenous distributions. Later on, on a French flag model was demonstrated that morphogen is secreted from a group of source cells and forms a gradient of concentration in the target tissue. Different target genes are expressed at different distances to the source, hence generating a spatial pattern of gene expression (Wolpert 1969). The impact of morphogen on target cells should be direct, with no intermediate signals, throught activating of gene expression (Wolpert 1998). Animals use these concentration gradients of signals termed morphogens for tissue patterning, but whether plants also use them still remains unclear. Since auxin contributes to plant development, differentiation and organ shaping by its spatiotemporal uneven distribution and its action is direct on single cells by activating gene expression in dose-dependent manner (reviewed by Bhalerao and Bennett 2003, Vanneste and Friml 2009, Petrášek and Friml 2009, Jönsson and Krupinski 2010), it might be considered as a plant morphogen. The signal generated by the actual auxin concentration during tissue patterning, cell faith determination and rapid tropical responses is transduced at both genomic and nongenomic levels. The morphogenic action of auxin gradients become apparent during the embryogenesis, where directional distribution of auxin leads to the determination of apical-basal axis and 7
18 subsequent growth and development of primary growth poles, where future organs are formed (Friml et al. 2002a; Friml et al. 2003b). During the post-embrynic development auxin concentration gradients are crucial for the patterning processes in both shoot and root apices and leaves as well as for the initiation and development of lateral organs (Tanaka et al. 2006). Direct IAA measurements and use of auxin-responsive reporter genes in Arabidopsis thaliana determined auxin maxima in root tips, developing embryos, lateral root and leaf primordia (Sabatini et al. 1999, Friml et al. 2002, Benková et al. 2003). One of the very few model systems, where auxin morphogenic action might be attributed to the exact concentration of auxin, is developing xylem in wood-forming tissues of trees. By method of mass spectrometry concentration peaks of IAA were defined in cambial meristems with descending gradient towards the edges of developing xylem, towards the phloem (Uggla et al. 1996, Kramer 2002, Kramer 2006). Auxin acts here as a morphogen in terms of organizing the cells in zones of cell division, expansion and secondary cell wall formation. Here, the distribution and gradient of auxin determines the xylem cell fate. Other example of tissue patterning via auxin gradients is the formation of lateral roots (Dubrovsky et al. 2008), leaf primordium formation during phylotaxis (Reinhardt et al. 2003) or floral development (Aloni et al. 2006). Recently, valuable review on the morphogenic action of auxin addressing the morphogenetic trigger of auxin during de novo formation organogenesis has been published (Benková et al. 2009) Auxin transport The establishment of the effective auxin levels in the particular cell or group of cells always involve its biosynthesis, conjugation, degradation and utilization by gene expression machinery and transport (Fig. 1). This complexity underlines the plasticity of plant body in reaction to specific environmental and developmental signals. Auxin is synthesized predominantly in meristematic tissues (shoot, root, young leaves and flowers) and translocated on longer distances in the symplast of phloem elements. Moreover, it is transported from cell-to-cell by the mechanism involving diffusion and carrier-mediated transport of auxin across plasma membrane. This transport is crucial for the fast directional, developmentally important establishment of auxin gradients. For short-distance transport, plants developed a unique, coordinated transport directly from cell to cell. This type of the transport is polar, due to the uneven distribution of auxin transport proteins within a plasma membrane (Friml 2010). The introduction of the chemiosmotic theory in the mid 1970s (Rubery and Sheldrake 1974, Raven 1975, Goldsmith 1977) explained the polarity of the cell-to-cell auxin transport. 8
19 The idea is, that auxin can exist in two forms, depending on the ph of the medium. In the acidic extracellular space (ph 5.5) part of the auxin exists in its protonated, electrically neutral IAAH form and thus can freely diffuse the cytoplasm. In the cytoplasm (ph 7), is auxin present in its anion form (IAA ) and due to its disability to pass the cell membrane, the IAA - anions are trapped there. Although the majority of IAA is transported passively inside, active auxin influx through carriers also contributes to the uptake IAA into cells. AUX1 amino acid permease and its three related genes LAX1 have been identified in Arabidopsis thaliana as a symporters of IAA - anion across the plasma membrane (Bennett et al. 1996). As mentioned above in the alkaline enviroment of cytosol (ph 7) are IAA - anions trapped. Their efflux was explained by the hypothetical existence of auxin efflux carriers Up to now, several proteins were shown to be involved in the auxin efflux. The first characterized auxin efflux carrier was member of PIN family PIN1 from Arabidopsis thaliana (named by pinformed mutant) (Galweiler et al. 1998). The phenotype of this mutant might be phenocopied by the auxin efflux inhibitor N-(1- naphthyl)-phtalamic acid (NPA) treatment (Okada et al. 1991). To this day, there have been described eight members of this family (PIN1-PIN8) (Zaţímalová et al. 2007; Křeček et al. 2010). Other proteins involved in auxin transport are ATP-Binding Cassette type B (ABCBs) transporters also known as multidrug resistant proteins or phosho (P)-glycoproteins (MDR- PGPs) (Noh et al. 2001, Titapiwatanakun and Murphy 2008). All of the PIN auxin efflux carriers have 6-10 transmembrane domains separated by hydrophilic loop (Muler et al. 1998) and they vary in their pattern of expression and subcellular localization. In the root meristem PIN1, PIN2, PIN3, PIN4 and PIN7 are localized at the plasma membrane of various cell types generating together the auxin fountain coming form the inner tissues to the quiescent centre and back through the outer tissues PIN1, PIN3 and PIN7 are located basally (towards he root apex) in the stele cells, delivering auxin towards the quiescent center (Friml et al. 2002, Blilou et al. 2005). PIN4 and PIN3 are localized laterally achieving auxin flow from the columella to the epidermis and lateral root cap (Friml et al. 2003). PIN2 is located on the apical side of epidermal cells, delivering auxin upwards to the elongating zone of the root (Müller et al. 1998, Friml et al. 2003). The localization of PINs is directly regulated by PINOID (PID) serine-threonine protein kinase (Bennet et al. 1995). At the shoot apex there was proposed that PID assigns apical or basal localization of the PIN protein (Friml et al. 2004) by its direct phosphorylation by PID or dephosporylation PP2A (protein phosphatase 2A) (Michniewicz et al. 2007). However, recently published data suggest that PINOID-independent PIN phosphorylation at one single 9
20 site is adequate to change the PIN polarity and thus direct auxin transport (Zhang et al, 2010). The so-called short PINs (with reduced middle hydrophilic loop) PIN5, PIN6 and PIN8 are localized on the endoplasmatic reticulum, possibly maintaining intracelluar pool of active auxin (Mravec et al. 2009). The asymmetry of PIN proteins localization within the cell directs auxin flow and is reflected in establishment of auxin gradients within the root apex. Indeed the localization and targeting of PINs (and proteins in general) at its place of action is enabled due to cooperation of endomembrane transport system with the cytoskeleton. 1.3 Mechanisms of protein targeting via vesicle transport Vesicle trafficking processes in plants The general information about a mechanism of vesicle trafficking is largely based on the studies of the mammalian and yeast cells. However, in contrast to other eukaryotes, protein trafficking apparatus in plants often contains higher number of isoformes of some components and differs in the organization and dynamics of the endomembrane space that might reflect different life strategy of the plants (Fig. 2) (reviewed by Hwang and Robinson 2009). In general, nascent polypeptides synthesized at the rough endoplasmic reticulum (ER) are translocated into the lumen of the endoplasmic reticulum. Here, depending on the sequencespecific signals, they could be integrated into the ER membrane or they could stay in the ER lumen and be subjected to various postranslational modifications with subsequent secretion towards the cell periphery. The continuous flow of membrane vesicles connects ER, Golgi apparatus (GA), trans-golgi network (TGN), endosomal space, prevacuolar and vacuolar compartments and plasma membrane. Interestingly, depending on the purpose, the cargo carried by these vesicles in the post-golgi and endosomal space could be found both inside of them as well as being incorporated in their membranes. With respect to the orientation of the vesicle transport, main directions are anterograde (towards plasma membrane) and retrograde (towards endoplasmic reticulum). The transport between all of these compartments appears to involve budding and fusion of transport vesicles. Budding of vesicles from the donor membrane is accomplished by vesicle coat proteins. They are responsible for packing of the specific cargo and directing it into its destination. Clathrin coats are responsible for trafficking between plasma membrane and trans-golgi network. The COPII (coat complex II) protein coat is formed by vesicles budding from ER and is destined for downstream components of the secretory pathway. Vesicles that recycle from GA to ER, involved in retrograde transport in GA (trans to cis direction) are formed by COPI coats. For the budding of the vesicles at the donor compartment and formation of protein coats are responsible small guanosine triphosphatase GTPases, namely Adenosyl Ribosylation Factors (ARFs), Secretion-associated 10
 and GAPs (GTPase-Activating Proteins) (reviewed by Hwang and Robinson 2009).")
21 and Ras-related (SAR) and Secretory (SEC) proteins. The switch of GTPase between its active and inactive form is regulated by GEFs (Guanine Exchange Factor) and GAPs (GTPase-Activating Proteins) (reviewed by Hwang and Robinson 2009). As long as the coat protein is present, vesicles cannot fuse with the target compartment. Vesicles are transported along the cytoskeleton to the target membrane, where they are recognized by the tethering factors with subsequent release of their coat proteins by ATPases or GAPs (GTPaseactivating proteins). After docking, vesicles fuse with the help of Soluble N-ethylmaleimidesensitive factor Attachment protein Receptors (SNARe) protein complex. Vesicle containing v-snare protein is recognized by t-snare complex of the target membrane (Hay and Sheller 1997, Sanderfoot and Raikhel 1999). Figure 2 The scheme of trafficking pathways of the proteins in plants. CCV- clathrin coated vesicle, RE - recycling endosome, TGN/EE trans-golgi network/early endosome, PVC/MVB - pre-vacuolar compartment/multivesicular body, G -Golgi apparatus, V -vacuole, ER - endoplasmatic reticulum, N- nucleus..reproduced from Hwang and Robinson (2009). The above-mentioned machinery is ubiquitinous in all eukaryotes. However, in plants there exist more isoforms of the major regulators of vesicle-mediated protein transport. Moreover, there is the absence of the intermediate compartment between GA and ER (reviewed by Hwang and Robinson 2009). Golgi stack in higher plants appear to have the ability to form de novo at the ER exit sites (ERES) and they are mobile as a secretory unit (Neumann et al. 2003, Hanton et al. 2005, Sparkes et al. 2009). The motility of the ERES/Golgi is ensured probably by acto-myosin complex (Hawes et al. 2008). 11
22 1.3.2 Endosomal space and recycling of the plasma membrane proteins One of the well-described pathways of endosomal sorting pathway in mammalian cells is the sorting of lysosomal proteins. Here, the transport from TGN to early endosome (EE) (anterograde) of lysosome-destined, receptor-bound acid hydrolases and plasma membrane receptors mediated by clathrin-coated vesicles (CCVs) (Braulke and Bonifacino 2009). The transport of acid hydrolases to the lysosomes proceeds via maturating of the late endosomes/multivesicular bodies (MVB) without the assistance of the vesicle transport (Woodman 2007). The retrograde recycling (from EE to TGN) of sorting receptors is mediated by so-called retromer-coated vesicles (Bonifacino and Rojas 2006) with the cooperation of clathrin and adaptor proteins (Popoff et al. 2007). Clathrin coats are involved also in transport steps on the cytoplasmic face of the plasma membrane, namely in the receptor-mediated and fluid-phase endocytosis. During endocytosis, the clathrin coat is assembled on the cytoplasmic face of the plasma membrane forming pits that invaginate to pinch off and become free CCVs. The proteins are destined to the lysosome throught late endosome/mvb or recycled to the cell surface after series of sorting events that occur in several organelles (Maxfield and McGraw 2004). In higher plants, two compartments in the post-golgi space are identified, TGN with the function of EE (TGN/EE) and prevacuolar compartment (PVC) that later forms the vacuole (Lam et al. 2007, Robison et al. 2008). The participation of clathrin in the endocytotic internalization of plasma membrane proteins was demonstrated for PIN auxin efflux carriers (Dhonukshe et al. 2007). From recent studies of sorting nexin/retromer localization it seems that CCVs formed at the TGN/EE interface are destined for plasma membrane recycling events rather than for an anterograde trafficking to PVC. Thus PVC probably functions as final destinaton for vacuolar targeting proteins without receptor recycling events (Niemes et al. 2010). Moreover, clathrin has been proposed to take a place in defense response, salt-stress response and cytokinesis (Konig et al. 2008, Leborgne-Castel et al. 2008, Karahara et al. 2009). The secretory pathway of the post-golgi (or recycling) vesicles, designated for exocytosis, ends up by docking and vesicle fusion at the plasma membrane with the assistance of SNARE proteins (Hay and Sheller 1997, Sanderfoot and Raikhel 1999). Prior to the process of the fusion with the plasma membrane vesicles are reversibly tethered to the plasma membrane by octameric protein complex, exocyst (He and Guo 2009). The exocyst protein complex has been described in angiosperm (Hála et al. 2008) as well as in land plants (Eliáš et al. 2003, Synek et al. 2006). 12
23 1.3.3 Dynamics of the auxin transporters in the endosomal space As mentioned above, proteins that are secreted to the PM might undergo repeated endocytosis and recycling via recycling endosomes (REs) or mature into PVC/MVBs before being targeted to the vacuole for the degradation. This protein trafficking plays an important role in rapid non-genomic regulation of the auxin signalling. The polarity of auxin transport and the formation of auxin gradients are directed by the coordinated localization of auxin efflux proteins (PINs) to the plasma membrane. The regulation of amount and positioning of PINs within the plasma membrane is achived by their constitutive cycling between plasma membrane an endosomal compartment (Geldner et al. 2001). This process is inhibited with brefeldin A (BFA) that interferes with ARF-GEF GNOM. After BFA treatment PINs are accumulated in the endosomal compartments and thus auxin transport is inhibited (Steinmann 1999). In addition, it has been proposed that auxin itself inhibits endocytosis of PIN1 transporters and as the consequence it promotes its own transport (Paciorek 2005). When compared with PIN1 auxin efflux carrier trafficking, auxin influx carrier AUX1 dynamics display different sensitivities to trafficking inhibitors and is independent of GNOM. Interestingly, the localization of newly synthesized PIN1 seems to be apolar (random within PM) until the polarization via recycling through endosomal compartment (Dhonukshe et al. 2008). ARF-GEF-dependent endosomal sorting is also involved in the trafficking of PINs to the lytic vacuolar pathway through the prevacuolar compartment (PVC), from which PIN proteins might be retrieved again into the trans-golgi network through the assistance of the retromer complex subunits SORTING NEXIN 1 (SNX1) and VACUOLAR PROTEIN SORTING 29 (VPS29) (Jaillais et al., 2006; Jaillais et al. 2007` Kleine-Vehn et al., 2008b). Alltogether,factors of directed protein targeting in plants are (at least) the following: 1) the lipid (sterol)/protein composition and/or possible domains on the targeted membrane 2) the vesicle coat protein composition determinates its localization and fate 3) the spatiotemporal dimension of the endomembrane internalization, i.e. the activation of the protein cargo within the vesicle/endosome by effector proteins (for example by (de)phosphorylation), protein-protein interaction and/or assembly of functional protein complexes. This machinery of vesicle formation, directed endomembrane transport, vesicle docking, tethering and fusion is underlined by the well co-ordinated network of signalling molecules (small GTPases) and cytoskeleletal proteins. 13
24 1.4 Cytoskeleton and endomembrane traffic interactions in the formation auxin gradients Cytoskeleton, a dynamic network of proteins Plant cytoskeleton is dynamic protein structure, which ensures number of cellular processes that are essential for maintaining cell shape, intracellular transport and cell division. In plants, cytoskeleton consists of two major protein polymers: microtubules (MTs) and actin filaments (AFs), which form dynamics tracks and scaffolds within the cells (reviewed by Kueh et al. 2009). Cytoskeleton as the dynamic network of continuously changing polymers quickly reacts by its reorganization to various internal and external stimuli. Its features, resulting from the structure of the actin and tubulin polymers, designate its function as a vector for polar transport of molecules and organels within the cells. Proteins interacting with the cytoskeletal polymers modulate its structure and function. The question is how is this system coordinated? Where is the polarity landmark, which directs cytoskeletal network in plants? What are the signalling cascades? Could be auxin the signal for the remodelling? There is probably a strong feedback between auxin and cytoskeleton polarity. Cytoskeleton together with endomembrane trafficking machinery seemingly directs auxin transport through targeting its membrane transporters and vice-versa the establishment of auxin gradients could re-establish via effector proteins the cytoskeletal polarity Microtubules MTs are hollow cylindres about 25 nm in diameter, being composed of protofilaments, which are polymers of alpha ( ) and beta ( ) tubulin dimers. Typically, protofilaments arrange themselves in an imperfect helix with one turn of the helix containing 13 tubulin dimers, each from a different protofilament. MTs are highly dynamic structures capable of very rapid response to the endogenous and exogenous signals. They originate from dispersed nucleation sites containing γ-tubulin and polymerize by GTP-dependent manner into protofilaments (Lambert et al. 1993). The γ-tubulin is a present in a form of a ring, which might serve as a template for holow MTs (Pastuglia et al. 2006). The speed of tubulin turnover (in the range of tens of seconds) depends on the levels of free GTP and the bulk of free and tubulin monomers. The elongation occurs towards the (+) end. In vitro, heterodimers can add or dissociate at either end of a microtubule, but there is greater tendency for subunits to add at the plus end, where b-tubulin is exposed. Microtubules can undergo treadmilling, with addition of tubulin heterodimers at the plus end and dissociation of tubulin heterodimers at the minus end. Tubulins are encoded by numebr of genes that code for proteins with differences in charge and possible function. This microheterogeneity is 14
25 supplemented by various post-translational modifications. Tubulin has two functional domains, polymerization and regulatory. Polymerization, autoregulatory domain contains a region that binds GTP and mediates the link between dimers and protofilaments (Fosket et al. 1992). Regulatory domain is localized at the carboxyl-terminal region of both tubulin subunits and contains binding sites for calcium (Ca 2+ ) and microtubule-associated proteins (MAPs) (Sackett et al. 1985). MAPs can be roughly divided into two groups. The first group includes structural MAPs, mediating interactions among MTs and other cell structures, stabilizing MTs and governing their polymerization (Mandelkow 1995). Structural MAPs reduce the critical threshold concentration of tubulin required for MTs polymerization and thereby increase its speed (Nick et al. 1998). There are many linker proteins between cortical MTs and plasma membrane, phospoliphase D being identified as a one of them (Gardiner et al. 2001). Severing of the MTs at the cortical cytoplasm of the isotropic growing root hairs is ensured by plant homologes of animal proteins katanin (Bartolini and Gundersen 2006). Also homologues of animal +TIPs proteins ((+) end tracking proteins) were identified in plants, namely SPR1 (SPIRAL), CLASP (CLIP-associated protein) and EB1 (END BINDING) (Hamada 2007). These MTs (+) end residing proteins were described as regulatory proteins of MTs instability and also function as sensors of environmental determinants in the cell (reviewed by Sedbrook and Kaloriti 2008). The second group of MAPs includes motor proteins. Kinesins as a microtubular motors generate movement along the MT by conversion of chemical energy (ATP hydrolysis) to kinetic energy (Miki et al. 2005). In plants, MTs are organized in the specific structures during the cell cycle. During preprophase they form the preprophase band (PPB) of MTs marking the site of the future localization of newly formed cell plate. The importance of MTs is obvious during mitosis and cytokinesis, where they form mitotic spindle and phragmoplast, respectively. In the interphase cells, MTs are localized in the cortical cytoplasm forming the network of cortical microtubules (cmt) underlining cell wall. cmts are generally parallel to each other and in most cases placed perpendicular to the preferred cell growth (Nick et al. 1998). The orientation of cmt plays an important role in the maintenance of cell polarity by positioning of newly synthesized cellulose microfibrils on the inner side of the cell wall (Giddings and Staehelin 1991). The interaction between cell wall and MTs has been described in several reviews (for example Wasteneys and Ambrose 2009). The importance of the possible feedback control and complicated bi-directional regulation between the CSCs machinery and cortical microtubule array and the contribution of MTs and AFs domains within the plasma 15
26 membrane for the polarization of cell expansion in various plant cell types has been recently summarized by Szymanski (2009) Actin filaments AFs are composed of two linear polymers of actin globular monomers (G-actin) covered with the spectrum of associated proteins. In the vegetative cells of Arabidopsis thaliana, there are prevalently three actin isoforms present, coded by ACTIN 2, ACTIN7 and ACTIN8 genes (Kandasamy et al. 2009). Two single filaments (F-actin) are interlaced (head-to-tail) forming microfilament and are polarized with its fast-growing barbed end and slow-growing pointed end. The elongation is about 10-times faster at the barbed end as a result of ATP hydrolysis. When the rate of polymerization at the barbed is equal to the rate of depolymerization on the pointed end, the filaments elongates on one end and shorten on the other and in total it moves by so called treadmilling. The speed of polymerization relies on the concentration rate of G- actin, free ATP and presence of Ca 2+, Mg 2+ a K + and associated proteins. Dynamic behavior of AFs highly depends on the presence actin binding proteis (ABPs), which were identified in more than 70 classes in eukaryotes. ABPs could be roughly divided into following groups: nucleating factors, capping proteins, severing proteins and side-binding proteins (reviewed by Staiger et al. 2010). These proteins control the ratio of G-actin and F- actin, participate in the nucleation of AFs, control the speed of polymerization and are involved in organizing of filaments into higher structures. The monomer binding proteins are profilins, ACTIN-DEPOLYMERIZING FACTOR (ADF) proteins and CYCLASE ASSOCIATED proteins (CAP). These proteins control the turnover of actin filaments (reviewed by Staiger et al. 2010). Profilin makes complex with G- actin and this binding prevents spontaneous nucleation at the filament pointed end. In the case of uncapped, barbed end of AFs, profilin contributes to the filament elongation by delivering monomer actin (Staiger et al. 1997). Formins, plasma membrane associated actin nucleation proteins interact with profilin and are major actin filament nucleators in plants (Cvrčková et al. 2004). ADF proteins regulate actin dynamics by keeping the pool of monomeric actin. CAP proteins bind ATP-G-actin as well as ADP-G-actin facilitating nucleotide exchange on actin. The monomer binding proteins work probably in synergy. CAP proteins with the ability of monomer binding and nucleotide exchange might function in between the ADF-mediated depolymerization and profilin-based shuttling of ATP-G-actin onto actin filament barbed ends (Staiger et al. 2010). Proposed model of actin dynamics in plant cells assumes that the major pool of the monomer actin is profilin-bound, preventing its spontaneous nucleation and pointed end growth. New filaments with available non-capped barbed ends are created by 16
27 nucleating factors (formins or the Arp 2/3 complex) either de novo or along the side of mother filaments or grow from the end of fragments or from recently severed filaments with free barbed ends by cooperation of actin with ABPs (Staiger et al. 2010). Capping proteins are class of ABPs that bind and cap actin filament ends. The best characterized is CP protein from Arabidopsis. It prevents subunit loss and addition of actin monomers by capping the barbed ends of the AFs (Huang et al. 2006). Fimbrin, villin and LIM domain proteins are ABPs, which allow the formation of actin network and higher structures and thus function as a side-binding and severing proteins (Thomas et al. 2009). Another group of proteins associated with AFs are molecular motors as myosins, which use energy from ATP to move along the AFs (Asada and Collings 1997, Shimmen and Yokota 2004, Shimmen, 2007). Actin bundles as continuously remodelled arrays serve as a tracks for the motor based transport of various organelles and cargos throughout the cells, GA stacks and ER (Boevink et al. 1998), mitochondria (Sparkes et al. 2008), peroxisomes (Mathur et al. 2002), chloroplasts (Takagi et al. 2009), membrane vesicles and endomembrane compartments (Geldner et al. 2001, Grebe et al. 2003, Dhonukshe et al. 2008). In addition, the network of dynamic AFs build framework around the organelles and positions them within cytoplasmic domains (Oikawa et al. 2003). In the interphase cell AFs are localized in the cortical cytoplasm and although they do not co-align with cmts they seem to stabilize them at the plasma membrane (Shibaoka et al. 1994). AFs were found along the MTs in PPB and in phragmoplast (Kakimoto and Shibaoka 1987). Although some of the actin bundles are often aligned with the long axes of the cells, actin is present around the interphase nucleus and in cytoplasmic strands, where together with associated proteins ensure cytoplasmic streaming, organelle movements and vesicle trafficking (Kamiya et al. 1981). Depolymerisation of AFs induces changes in the localization of cortical cellulose microfibrils and affects polarized growth (Baskin and Bivens 1995). After the depolymerization of AFs with latrunculin B, there were found aggregates of CSCcontaining Golgi stacks and patchy distribution of membrane located CSCs mimicking Golgi mislocalization (Crowell et al. 2009). AFs play the very important role during diffuse cell growth as well as in the polarized apical growth of root hairs, trichomes and pollen tubes (reviewed by Hussey et al. 2006, Szymanski 2009). In the tip growth actin serves as a scaffold for transport and docking of secretory vesicles at the plasma membrane (reviewed by Staiger et al. 2010). It was reported, that actin polymerisation is necessary for the membrane vesicle accumulation at the tip of pollen tubes, while actin depolymerization accompany vesicular docking and fusion at the plasma membrane (Lee et al. 2008). Next to ABPs, small GTPases play crucial role in these 17
28 processes. In a growing pollen tubes small GTPases Rop (Rho of plants) were found having similarity with Rho GTPases of animals and yeast (Lin et al. 1996). These signalling molecules are involved in the organization of AFs by the modelling of Ca 2+ flow (Li et al. 1999). Tip-localized Rop-GTPases function through antagonistically working ROPinteractive CRIB motif containing proteins RIC3 and RIC4 (Hwang et al. 2005). Actinrelated protein (Arp 2/3) complex is one of the possible nucleators of actin in plants (Staiger and Blanchoin 2006). This complex is activated by the Wiskott-Aldrich Syndrome Protein (WAVE) protein complex. Some of the subunits of WAVE complex appear to be ROP effectors that translate ROP-GTPase signals into nucleation response of AFs. The WAVE-Arp 2/3 activation might generate new actin nucleation domains at the cortical cytoplasm and thus contribute into co-ordinated cell expansion (Szymansky 2009) Examples of MTs and AFs interactions Both microtubular and actin network are involved in the regulation of cell directional elongation and expansion and in the establishment of cell polarity. Both networks display distinct ways to organize cytoplasm and define the cortical boundaries with increased cell wall elasticity. Nice example of cooperation of the MTs with AFs is the model of their participation in transport and targeting CSCs within the plasma membrane. Actin cables and their myosin motors ensure the transport of CSC-loaded GA through cytoplasm to the cortical region where GA docks by kinesin motors to cmts. Then, CSCs are transported to the plasma membrane by CSC-containing vesicles. The function of MTs thus lies in the local GA unloading, trafficking of CSC-vesicles and their fusion with the plasma membrane (Szymansky 2009). The physical interaction between these two polymers could be facilitated by its molecular motors kinesin and myosin, the interaction can be mediated by +TIPs proteins, formins, MAPs etc. (reviewed by Collings 2008, Petrášek and Schwarzerová 2009). The affinity between phospholipase D and the ß-tubulin was shown and as well as between PLD and actin7, suggesting the interaction between PLD and the cytoskeletal proteins (Ho et al. 2009). However, the signalling mechanism underlining the interconnection between these two cytoskeletal networks still reminds unclear Cytoskeleton in vesicle trafficking As mentioned above, small GTPases appear to be very important in trafficking events as well as in controlling cytoskeletal changes and signalling events in the cells. In yeast and animal cells the connection between clathrin-mediated endocytosis and Arp2/3-mediated actin assembly was reported (Galleta and Cooper 2009). Although there were indentified many 18
29 actin binding proteins in Arabidopsis, there is still missing the direct proof of a link between actin and clathrin-mediated endocytic machinery. Some evidence about cytoskeletal requirement in the vesicle trafficking events comes from tip-growing cells such as root hairs and pollen tubes. F-actin was found to generate strong meshwork at the tips of the root hair cells, where it coordinates secretory vesicle docking and fusion at the apex. It was reported that actin polymerization is necessary for the vesicle accumulation and actin depolymerization found to be important for vesicle docking and fusion at the plasma membrane (Lee et al. 2008). Both oscillating cytosolic calcium gradients and tip-localized Rho-GTPases in growing root tips target actin cytoskeleton, which in the feedback relation regulates their positioning (Cole and Fowler 2006) Cytoskeleton in the targeting of auxin transporters As mentioned above the intracellular polarization of auxin transporters is linked to the vesicle trafficking processes (Steinmann et al. 1999, Geldner et al. 2003, Kleine-Vehn et al. 2006). Pharmacological experiments suggested that their constitutive cycling in the endosomal space seems to be actin- and myosin dependent (Geldner et al. 2001; Holweg et al 2004; Holweg 2007). Moreover, it seems that with respect to the cytoskeleton, apical-basal localization of PIN proteins follows distinct pathways as suggested by experiments in Arabidopsis thaliana roots. Here, the process of cycling and translocation of endosomes with PIN proteins from one membrane domain to the other, so-called transcytosis (Kleine-Vehn et al. 2008a), has been shown to involve preferential involvement of AFs in the apical-basal targeting of PINs and MTs preferentially in basal targeting (Kleine-Vehn et al. 2008b). The participation of MTs in PIN protein positioning was proposed earlier using anti-tubulin drugs (Boutté et al. 2006). However the information about mechanism of the participation of cytoskeleton in auxin efflux carrier trafficking and targeting is limited. Cytochalasin D (Cyt D), drug known to disrupt actin filaments influenced both PIN1 polar localization and auxin transport (Butler et al. 1998; Geldner et al. 2001). Brefeldin A (BFA), which is known to interfere with anterograde protein transport, induced accumulation of PIN1 proteins in endosomal compartments (Geldner et al. 2001), reduced auxin efflux and also caused remodelling of actin filaments (chapter 3.1 of this thesis; Petrášek et al. 2003). The accumulation of PIN1 in intracellular compartments upon BFA treatment is reversible and this reversibility is uncoupled in the presence of Cyt D, indicating that this process is actindependent (Geldner et al. 2001). Moreover, endocytosis of membrane vesicles is inhibited by auxin itself, which leads to the promotion of its own efflux (Paciorek et al. 2005). This inhibitory effect of auxin can thus be used as powerful tool for uncovering its own action. 19

30 Fig.3: The model of a self-referring regulatory circuit between polar auxin transport and actin organization where auxin promotes its own transport by shaping actin filaments based on results of Maisch and Nick 2007 and Nick et al The level of actin filament bundling is critical for the efficient auxin efflux carrier deposition. Modified from Nick et al Stabilizing effect of auxin transport inhibitors TIBA (2,3,5-triiodobenzoic acid) and PBA (2-(1-pyrenoyl) benzoic acid) on actin filaments and its role in impairing vesicle traffic was demonstrated next to plant cells also in other eukaryotes (Dhonukshe et al. 2008). Actin bundling induced by overexpressing mouse actin-associated protein talin impairs cell division synchrony and increases its sensitivity to another inhibitor of auxin transport 1-Nnaphthylphthalamic acid (NPA). Normal actin organization and synchronized cell division was re-established by addition of auxins that are subject of auxin efflux in the tissue, IAA and NAA (Maisch and Nick 2007). All these findings point to the idea that auxin could control its own transport by changing actin filament dynamics and thus vesicle trafficking of own transporters (Nick et al. 2009; Fig. 3). Epidermal pavement cells of Arabidopsis thaliana and their jigsaw morphology are favourite model to study the establishment of cell polarity during cell shaping. Fine actin meshwork and microtubules (Fu et al. 2005) and its ROP (Rho of plants) GTPase-dependent reorganization have been shown to play important role in pavement cell development (Settleman 2005). Interestingly, it is speculated that PIN proteins, auxin and their interactions with ROP GTPases might play role in the development of lobes (Yang 2008). ROP proteins appear to interact directly with upstream regulators of the Arp 2/3 complex. Arp2/3 complex plays role in the modulation of the spatial distribution of cortical F-actin and activates the polymerization of some types of actin filaments in cells with complex shapes (Li et al. 2003). ROP function is dependent on the class 1 ADP-ribosylation factors (ARFs), which are core 20
31 components of the vesicle transport machinery involved in the polar localization of PIN auxin efflux carriers (Xu and Scheres 2005, Boutté et al.2007, Geldner 2009). 21
32 22
33 2 Objectives and outlines This thesis is focused on some components of cell polarity establishment and their cooperation, mainly during auxin-triggered morphogenesis. Auxin regulates patterning of plant body by its asymmetrical distribution and polar flow along the spatio-temporal axes. Direction of the auxin flow is mediated by polar distribution of auxin transport proteins within single cell. The main objective of the work presented in this thesis has been to describe the role of the cytoskeleton as a dynamic tracks guiding and positioning the membrane vesicles containing among others auxin efflux proteins. In the chapter 3.1 of this thesis the connection between the protein trafficking processes and the actin remodelling is implied. Brefeldin A (BFA), the drug known to disrupt anterograde induced substantial and reversible remodelling of actin filaments around BFAinduced endomembrane compartments, while cortical actin filaments remained intact. Thus, actin remodelling upon BFA treatment suggested interesting connection between endomembrane dynamics and actin filaments. The data from this paper were already part of my diploma thesis and other aspects connected with action of auxin transport inhibitors were discussed there, but results with actin remodelling form an important background for other chapters of this thesis. The most important question that had remained from this work was whether the process of actin remodelling upon BFA treatment is active and what is the role of membrane trafficking in these processes. Based on the above mentioned results with BFA interfering with auxin transport and actin filaments remodelling, we have complemented our analysis with the spectrum of inhibitors of the retrograde transport from the plasma membrane and cytoskeletal drugs and monitored in vivo their effect on the endocytosis marker FM 4-64 uptake and PIN1 auxin efflux carrier localization in tobacco BY-2 cells. It was possible to dissect both the requirement of cytoskeleton and endocytosis for the trafficking of auxin efflux carriers and also suggest the general requirement of cytoskeleton for plant endocytosis. The results from these studies are summarised in the chapter 3.2. During the experiments, where endocytosis in tobacco BY-2 suspension-cultured cells was tracked with endocytosis marker FM 4-64, the unexpected effect of this marker and other marker from FM group on the plasma membrane protein localization was observed. Detailed analysis of FM styryl dyes that transiently affected the localization and activity of plasma membrane-localized auxin transporters suggests that they should be used with appropriate circumspection. Results of this analysis are described in the chapter 3.3 of this thesis. 23
34 Since our studies using BY-2 cells suggested the importance of intact and dynamic cytoskeleton in the processes of vesicle trafficking and PIN protein targeting, we have started the studies of this topic in planta. To obtain complete data on the role of cytoskeleton in the processes of protein targeting and auxin transport, a microscopical phenotypic screen of all available Arabidopsis thaliana plants with mutated genes for cytoskeletal proteins and their important interactors was initiated. The rate of endocytosis, scored by FM 4-64 uptake in the root epidermal and cortex cells, and parallel immunofluorescence localization of PINs has been determined. The goal of this work is to identify new determinants of PIN polar targeting. The data from the first part of the screen including mutants in all available actin proteins, Arp 2/3 complex subunits and its effectors from WAVE activating complex and actin monomer binding proteins are summarized in the chapter
 Do phytotropins inhibit auxin efflux by impairing vesicle traffic?")
35 3 Results 3.1 Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Petrášek, J., Černá, A., Schwarzerová, K., Elčkner, M., Morris, D. and Zaţímalová, E. (2003) Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiol, 131, This paper is concentrated on the mechanism of action of phytotropin 1-Nnaphthylphthalamic acid (NPA) and vesicle trafficking inhibitor brefeldin A (BFA) on auxin transport in BY-2 (Nicotiana tabacum L. cv Bright-Yellow 2) suspension cells. Both of these compounds decreased auxin efflux. Moreover, BFA in contrast to NPA had substantial effect on actin cytoskeleton. These data suggest that BFA action on auxin transport and disruption of actin cytoskeleton are connected. It could be predicted that actin cytoskeleton and endomembrane trafficking might participate on localization of auxin efflux carriers within the plasma membrane. My participation in this work was in preparation and cultivation of the plant material, visualization of cytoskeletal proteins, drug treatments, in vivo studies, microscopy and image analysis, interpretaion of the obtained data. These data were already part of my diploma thesis, they are presented here because they form the background for other three chapters that form this thesis devoted to the co-operation between cytoskeleton and vesicle trafficking events. BFA-induced remodelling of perinuclear actin filaments in the interphase BY-2 cells 25
36 26
37 Do Phytotropins Inhibit Auxin Efflux by Impairing Vesicle Traffic? 1 Jan Petrášek, Adriana Černá, Kateřina Schwarzerová, Miroslav Elčkner, David A. Morris 2, and Eva Zažímalová* Institute of Experimental Botany, The Academy of Sciences of the Czech Republic, Rozvojová 135, CZ Prague 6, Czech Republic (J.P., M.E., D.A.M., E.Z.); and Department of Plant Physiology, Faculty of Science, Charles University, Viničná, CZ Prague 2, Czech Republic (J.P., A.Č., K.S.) Phytotropins such as 1-N-naphthylphthalamic acid (NPA) strongly inhibit auxin efflux, but the mechanism of this inhibition remains unknown. Auxin efflux is also strongly decreased by the vesicle trafficking inhibitor brefeldin A (BFA). Using suspension-cultured interphase cells of the BY-2 tobacco (Nicotiana tabacum L. cv Bright-Yellow 2) cell line, we compared the effects of NPA and BFA on auxin accumulation and on the arrangement of the cytoskeleton and endoplasmic reticulum (ER). The inhibition of auxin efflux (stimulation of net accumulation) by both NPA and BFA occurred rapidly with no measurable lag. NPA had no observable effect on the arrangement of microtubules, actin filaments, or ER. Thus, its inhibitory effect on auxin efflux was not mediated by perturbation of the cytoskeletal system and ER. BFA, however, caused substantial alterations to the arrangement of actin filaments and ER, including a characteristic accumulation of actin in the perinuclear cytoplasm. Even at saturating concentrations, NPA inhibited net auxin efflux far more effectively than did BFA. Therefore, a proportion of the NPA-sensitive auxin efflux carriers may be protected from the action of BFA. Maximum inhibition of auxin efflux occurred at concentrations of NPA substantially below those previously reported to be necessary to perturb vesicle trafficking. We found no evidence to support recent suggestions that the action of auxin transport inhibitors is mediated by a general inhibition of vesicle-mediated protein traffic to the plasma membrane. The polar transport of auxins (such as indole-3- acetic acid [IAA]) plays a crucial role in the regulation of growth and development in plants (Davies, 1995). Much experimental evidence supports the proposal by Rubery and Sheldrake (1974) and Raven (1975) that auxin transport polarity results from the differential permeabilities of each end of transporting cells to auxin anions (IAA ) and undissociated auxin molecules (IAA; for review, see Goldsmith, 1977). IAA (a weak organic acid) is relatively lipophilic and can readily enter cells by diffusion from the more acidic extracellular space; the IAA anion, on the other hand, is hydrophilic and does not cross membranes easily. As a consequence, auxins tend to accumulate in plant cells by a process of anion trapping and exit the symplast with the intervention of transmembrane auxin anion efflux carriers (Goldsmith, 1977). There is now overwhelming evidence 1 This work was supported by the European Union, International Cooperation Copernicus (grant no. ERBIC15 CT to E.Z.); by the Ministry of Education, Youth, and Sports of the Czech Republic (project no. LN00A081); and by the UK Royal Society and the Academy of Sciences of the Czech Republic under the European Science Exchange Scheme (grant to D.A.M.). 2 Present address: Division of Cell Sciences, School of Biological Sciences, University of Southampton, Bassett Crescent East, Southampton SO16 7PX, UK. * Corresponding author; eva.zazim@ueb.cas.cz; fax Article, publication date, and citation information can be found at that the differential efflux of IAA anions from the two ends of auxin-transporting cells results from an asymmetric (polar) distribution of such carriers (Goldsmith, 1977; Lomax et al., 1995). Genes encoding putative auxin influx and efflux carriers have been identified from Arabidopsis and other species (for review, see Morris, 2000; Muday and DeLong, 2001; Friml and Palme, 2002). It has been shown that efflux carrier proteins, encoded by members of the PIN (PIN-FORMED) gene family, and possibly influx carriers (encoded by AUX1), are targeted to specific regions of the plasma membrane (PM) in auxintransporting cells (Bennett et al., 1996; Gälweiler et al., 1998; Müller et al., 1998; Swarup et al., 2001; for review, see Friml and Palme, 2002). Studies employing specific inhibitors of components of the polar auxin transport process have played a major role in shaping our understanding of the polar auxin transport machinery. The most widely used inhibitor of auxin efflux is 1-N-naphthylphthalamic acid (NPA), a well-characterized member of a group of inhibitors known as phytotropins (Katekar and Geissler, 1980; Rubery, 1990). The application of NPA to various plant tissues results in the inhibition of auxin efflux carrier activity and, as a consequence, increases auxin accumulation in cells (for review, see Morris, 2000). Although the mechanism of its inhibitory action on polar auxin transport remains obscure, it seems to be mediated by a specific, high affinity, NPA-binding protein (NBP; Sussman and Gardner, 1980; Rubery, 1990). Observations on zuc- 254 Plant Physiology, January 2003, Vol. 131, pp , American Society of Plant Biologists
38 Inhibition of Auxin Efflux in BY-2 Tobacco Cells chini (Cucurbita pepo) hypocotyl cells have shown that the NBP is probably a peripheral membrane protein located on the cytoplasmic face of the PM and associated with the cytoskeleton (Cox and Muday, 1994; Dixon et al., 1996; Butler et al., 1998; but compare with Bernasconi et al., 1996). Protein synthesis inhibitors such as cycloheximide (CH) rapidly uncouple carrier-mediated auxin efflux and the inhibition of efflux by NPA (Morris et al., 1991). In the short term, however, CH has no effect on either the specific and saturable binding of NPA or on auxin efflux itself, suggesting that the NBP and the efflux catalyst may interact through a third, rapidly turned over protein (Morris et al., 1991; for discussion, see Morris, 2000; Luschnig, 2001). Although the identity of the NBP and its mechanism of action on auxin efflux carriers remain unknown, the Arabidopsis tir3 (transport inhibitor response 3) mutant exhibits a reduced number of NPA-binding sites and a reduction in polar auxin transport (Ruegger et al., 1997). Thus, TIR3 (renamed BIG by Gil et al., 2001, to reflect the unusually large size 566 kd of the protein it encodes) may code for an NBP or may be required for NBP expression, localization, or stabilization (for discussion see Gil et al., 2001; Luschnig, 2001). In addition to polar auxin transport inhibitors, drugs that inhibit Golgi-mediated vesicle traffic, such as brefeldin A (BFA) and monensin, also very rapidly inhibit auxin efflux carrier activity in zucchini hypocotyl tissue (Wilkinson and Morris, 1994; Morris and Robinson, 1998), and in suspension-cultured tobacco (Nicotiana tabacum L. cv Xanthi XHFD8) cells (Delbarre et al., 1998). They also inhibit polar auxin transport through tissue (Robinson et al., 1999). However, the time lag for inhibition of efflux carrier activity by BFA (minutes) is considerably shorter than the lag for inhibition of efflux activity by protein synthesis inhibitors (up to 2 h; Morris et al., 1991). This implies that efflux catalysts turn over very rapidly in the PM without a requirement for concurrent protein synthesis, a situation that contrasts sharply with the inhibitory action of NPA on auxin efflux, which does require concurrent protein synthesis (see above). Results of a detailed comparison of the effects of CH and BFA on efflux carrier activity revealed that efflux carrier proteins probably cycle between the PM and an unidentified intracellular compartment (Robinson et al., 1999; compare with Delbarre et al., 1998). This possibility has been strongly supported by the observation that AtPIN1, a member of a family of putative Arabidopsis auxin efflux carrier proteins (see Friml and Palme, 2002), is rapidly and reversibly internalized after BFA treatment of Arabidopsis roots (Geldner et al., 2001). A link has been suggested recently between the inhibitory action of polar auxin transport inhibitors on auxin efflux and their inhibitory effects on the actin-dependent vesicle trafficking and cycling of efflux carrier proteins (Geldner et al., 2001). Treatment of Arabidopsis roots with the auxin transport inhibitor 2,3,5-triiodobenzoic acid (TIBA) prevented the BFA-induced internalization of the putative auxin efflux carrier AtPIN1 and prevented the traffic of internalized PIN1 to the PM after BFA washout. This would have the effect of reducing the density of carriers in the PM available for auxin efflux. The authors found similar effects of TIBA on a rapidly turned-over PM-ATPase and the KNOLLE gene product (a syntaxin involved in vesicle docking; see Muday and Murphy, 2002). As a consequence, a rather general action of auxin transport inhibitors on membrane-trafficking processes was suggested, rather than a specific effect on auxin efflux carriers (Geldner et al., 2001). The site-directed traffic of auxin efflux carrier proteins involves not only the Golgi-mediated secretory system itself, but also the participation of components of the cytoskeletal system. The application of cytochalasin (an actin-depolymerizing agent) reduced polar auxin transport in maize (Zea mays) coleoptiles (Cande et al., 1973) and in zucchini hypocotyls (Butler et al., 1998). Moreover, cytochalasin D has been shown recently to block the cycling of PIN1 between endosomal compartments and the PM in Arabidopsis roots (Geldner et al., 2001). These observations are consistent with an important role for actin filaments (AFs) in the proper localization and function of components of the auxin efflux carrier complex (for review, see Muday, 2000; Muday and Murphy, 2002). Evidence from a careful in vitro biochemical analysis of the association between the NBP and the cytoskeleton in membrane preparations from zucchini hypocotyls indicates a strong link between NPA action and the actin cytoskeleton (Butler et al., 1998). Only treatments that stabilized F-actin (phalloidin), but not those that stabilized microtubules (MTs; taxol), increased NPA-binding activity. Furthermore, direct interaction between the high-affinity NBP and F-actin was proven by F-actin affinity chromatography in the same system (Hu et al., 2000). The processes that regulate the cycling of the efflux carrier proteins and that direct their traffic to specific areas of the PM remain unknown, although recent observations are beginning to provide insights into possible mechanisms. Gil et al. (2001) have reported that tir3 (see above) and doc1 (dark overexpression of CAB; Li et al., 1994) are allelic mutants of a gene (BIG) that has significant identity with the CAL/O (CALOS- SIN/PUSHOVER) gene. The product of CAL/O is involved in the regulation of synaptic vesicle cycling in Drosophila melanogaster (Richards et al., 1996). BIG (TIR3) is required for normal auxin transport in plants and is probably associated with the actin cytoskeleton (Cox and Muday, 1994; Butler et al., 1998). Because the cycling of putative auxin efflux carrier proteins involves BFA-sensitive and actin-dependent vesicle traffic to the PM (Geldner et al., 2001), BIG Plant Physiol. Vol. 131,
39 Petrášek et al. may play a similar role in plants to that of CAL/O in D. melanogaster, and regulate this directed vesicle traffic. The observations discussed above associate the inhibition of auxin efflux carrier activity by NPA with effects of the compound on actin-dependent and Golgi vesicle-mediated targeting of efflux carrier protein to the PM. Nevertheless, almost nothing is known about the effects (if any) of NPA or other phytotropins on the organization of components of the cytoskeleton or on the vesicle secretory system. Given the possibility that phytotropins might have a general effect on vesicle traffic to the PM and on the cycling of proteins between the PM and endosomal compartments (as suggested by Geldner et al., 2001; but see Discussion ), some physical disruption of the secretory pathway and/or cytoskeleton might be expected to occur after the application of these compounds. However, to the best of our knowledge, no such disruption has been reported so far. Here, we report an investigation to compare the action of BFA and NPA on both auxin accumulation and on the arrangement and structure of components of the secretory pathway and the cytoskeleton (AFs, MTs, and endoplasmic reticulum [ER]) in suspension-cultured BY-2 tobacco cells. Using a new quantitative method to study the rearrangement of AFs and the formation of actin clusters in the perinuclear region of cells, we show that although both of these compounds increase auxin accumulation by inhibiting auxin efflux, only BFA has an effect on the structure of AFs and the ER. Our observations lead us to suggest that although radial and perinuclear (but possibly not cortical) AFs and ER are required for normal auxin efflux, the inhibitory action of NPA on efflux does not involve any changes in the cytoskeleton and ER. RESULTS Effects of NPA and BFA on the Accumulation of Auxin Figure 1. The effect of NPA on the net accumulation of [ 3 H]NAA (2 nm) in 2-d-old BY-2 cells. A, Time course of net [ 3 H]NAA accumulation in the absence (F, control) and in the presence of 10 M (E), 50 M (f), and 200 M ( ) NPA. Error bars SEs of the mean (n 3). B, The effect of concentration of NPA on net [ 3 H]NAA accumulation, measured over 20 min, by 2-d-old BY-2 cells. Results expressed as mean radioactivity per 0.5 ml of cell suspension (cell density cells ml 1 ). Error bars SEs of the mean (n 4). The rate of [ 3 H]-labeled naphthalene-1-acetic acid ([ 3 H]NAA) accumulation by BY-2 cells is shown in Figures 1A and 2A. After an initial period of rapid uptake lasting 3 to 10 min depending on experiment, uptake settled to a slower, steady rate that was maintained for up to 40 min. Accumulation was extremely sensitive to NPA and was stimulated approximately 3-fold in the presence of 10 or 50 m NPA (Fig. 1A). An NPA concentration dependence study indicated that [ 3 H]NAA accumulation was maximally stimulated by as little as 1.0 m NPA and that the stimulatory effect of NPA began to decline rapidly at concentrations around or greater than 100 m (Fig. 1B). Because of the greatly reduced stimulation of [ 3 H]NAA accumulation at high concentrations of NPA, possibly caused by toxic side effects not directly related to auxin efflux, the maximum concentration of NPA employed in subsequent cytological observations was restricted to 50 m. A similar picture emerged in the case of BFA (Fig. 2), although the maximum stimulation of [ 3 H]NAA accumulation (at between 10 and 40 m BFA) was lower than that caused by NPA (compare Figs. 1A with 2A, and 1B with 2B). As with NPA, high concentrations of BFA (100 m) reduced the stimulation of [ 3 H]NAA accumulation (Fig. 2B). Over the uptake period used here, no significant metabolism of [ 3 H]NAA by BY-2 cells was detected. Apart from a small amount of label that remained at the origin in all chromatography solvents used (less than 10% of the total label recovered), the recovered ethanol-soluble radioactivity migrated as a single spot that had the same mobility on cellulose thinlayer plates as authentic [ 3 H]NAA (data not shown). 256 Plant Physiol. Vol. 131, 2003

.")
40 Inhibition of Auxin Efflux in BY-2 Tobacco Cells BFA significantly increased auxin accumulation (Figs. 1 and 2), their effects on cytoskeleton arrangement differed considerably. Although the fine cortical AFs and MTs retained their transverse orientation after a 30-min treatment with 20 m BFA (Fig. 3, B and H), BFA had a dramatic effect on the arrangement of the radial and perinuclear AFs (Fig. 3E). Fine AFs in the transvacuolar strands collapsed and actin became concentrated in clusters around the nucleus (Fig. 3E). We have developed a simple procedure for the evaluation of quantitative changes in actin aggregation in the perinuclear region, utilizing image analysis software. This method is based on the fact that relative fluorescence intensities of regions with aggregated actin are very different from the background fluorescence intensities. Thus, aggregation of actin increases the degree of variation in fluorescence intensities of individual pixels in the area of interest. Full details of the procedure are described in Materials and Methods (see also Fig. 4). This procedure was used to evaluate the effects of BFA and it was shown that the degree of actin aggregation noticeably increased with duration of treatment (Fig. 4E). The highest BFA concentration tested (100 m) was shown to be inhibitory for actin aggregation in the same way that it was inhibitory for auxin accumula- Figure 2. The effect of BFA on the net accumulation of [ 3 H]NAA (2 nm) in 2-d-old BY-2 cells. A, Time course of net [ 3 H]NAA accumulation in the absence (F, control) and in the presence of 20 M (E), 40 M (f), and 100 M ( ) BFA. Error bars SEs of the mean (n 3). B, The effect of concentration of BFA on net [ 3 H]NAA accumulation, measured over 20 min, by 2-d-old BY-2 cells. Results expressed as mean radioactivity per 0.5 ml of cell suspension (cell density cells ml 1 ). Error bars SEs of the mean (n 4). Effects of BFA and NPA on AFs and MTs To test the reaction of the cytoskeleton to the application of agents that modify polar auxin transport, the arrangement of both AFs and MTs in BFA- and NPA-treated cells was studied during a 30-min incubation period in parallel with the auxin accumulation measurements described above. Because the cell populations used to measure auxin accumulation were predominantly in interphase, we investigated the arrangement of the interphase cytoskeleton (MTs and AFs in the cortical cytoplasm and AFs in the transvacuolar strands and perinuclear region). Typical interphase BY-2 tobacco cells contained fine and transversally oriented AFs (Fig. 3A) and MTs (Fig. 3G) in the cortical cytoplasm, together with radially oriented AFs in transvacuolar strands and in the perinuclear region (Fig. 3D). There were no MTs in transvacuolar strands and around the nucleus in interphase cells (Fig. 3G). Although both NPA and Figure 3. The effect of BFA and NPA on the arrangement of AFs and MTs in 2-d-old BY-2 cells. Control cells with fine AFs in the cortical region (A), radially oriented AFs in transvacuolar strands (D), and transversely oriented cortical MTs (G). B, E, and H, AFs and cortical MTs after 30-min incubation in 20 M BFA. Modification of AFs staining pattern in cortical (B) and perinuclear region (E), where AFs in transvacuolar strands are pulled down, forming clusters around the nucleus. H, Unaffected arrangement of cortical MTs. C, F, and I, AFs and cortical MTs after 30-min incubation in 50 M NPA. Unaltered AFs staining pattern in the cortical (C) and perinuclear region (F). I, Transversally oriented cortical MTs with no obvious changes. Each image is representative of cells in the treatment specified. Approximately 1,000 cells per treatment were examined. Scale bars 10 m. Plant Physiol. Vol. 131,
.")
41 Petrášek et al. tion (compare Fig. 4E with 2B). The effect of BFA on AFs was reversible and 30 min after washout of BFA with fresh medium, the actin clusters disappeared and the ODV parameter decreased again to control values (Fig. 4F). In contrast to BFA, 30 min of incubation in 50 m NPA did not cause any changes in the arrangement of AFs in cortical region (Fig. 3C; compare with Fig. 3, A and B) as well as around the nucleus and in the transvacuolar strands (Fig. 3F; compare with Fig. 3, D and E). Correspondingly, cortical MTs were also unaffected after 30 min in 50 m NPA (Fig. 3I; compare with Fig. 3, G and H). The Effect of BFA and NPA on the ER Figure 4. The quantification of BFA effect on AFs. Grabbed images of tetramethylrhodamine B isothiocyanate (TRITC)-phalloidin-stained control (A) and BFA-treated cells (C) after transformation to complementary colors. B and D, Interactively applied circular measuring frame over the perinuclear region for the measurement of the relative optical density variation (ODV) parameter. See Materials and Methods for details. E, Relative ODV in control (white columns) and in the presence of 20 M (shaded columns), 40 M (checked columns), and 100 M (black columns) BFA. F, Relative ODV after 30-min incubation in 20 M BFA (time 0 min, shaded column) and after washout and subsequent 30-min incubation in fresh medium without BFA (time 30 min, white column) and in the presence of 20 M BFA in the medium (time 30 min, shaded column). Error bars SEs of the mean (n 10 optical fields, 30 cells assessed in each). In addition to the Golgi apparatus, the plant ER has also been shown to be sensitive to BFA treatment (Henderson et al., 1994). Therefore, we investigated if the ER was also affected in cells in which BFA or NPA stimulated the accumulation of auxin. The behavior of ER in interphase cells of BY-2 after NPA or BFA treatment was followed in vivo using cells transformed with the pbin m-gfp5-er plant binary vector coding for the ER-localized fusion protein (mgfp5-er). In exponentially growing control interphase cells, ER was present in the form of a tubular network penetrating not only the cortical layer of cytoplasm (Fig. 5A), but also the transvacuolar strands and perinuclear region (Fig. 5D). Within this network, small motile bodies were observed (video sequence can be seen at cz/laboratory_of_hormonal_regulations/bfamovies. htm). The movement of these bodies was observed over the surface of the network of ER tubules that constantly changed its orientation and pattern. Treatment of cells with 20 m BFA for 30 min resulted in disintegration of the fine tubular network of ER, the formation of large sheets of ER, and the aggregation of the signal into a large number of bright fluorescent spots (Fig. 5B; video sequence can be seen at regulations/bfamovies.htm). However, the first observable effects of BFA were clear after only 5 min (data not shown), when disintegration of the tubular network and formation of fluorescent spots started. On the other hand, even after 30 min of 20 m BFA treatment, there were still cells with no obvious damage of ER. The accumulation of GFP fluorescence was also observed in the perinuclear region (Fig. 5E). Moreover, the movement of small motile bodies inside ER tubules decreased during a 30-min incubation in 20 m BFA and had almost stopped by the end of that time period. Longer treatment with 20 m BFA (7 h) resulted in the formation of large sheets of ER and intensively fluorescing bodies of irregular shape and size (data not shown). In contrast to BFA, a 30-min incubation in 50 m NPA had no observable effects on either ER structure 258 Plant Physiol. Vol. 131, 2003
 fluorescence in control cells. Optical cuts through cortical region (A) and perinuclear region (D).")
42 Inhibition of Auxin Efflux in BY-2 Tobacco Cells Figure 5. The effect of BFA and NPA on the arrangement of ER in 2-d-old BY-2 cells expressing mgfp5-er. A and D, Green fluorescent protein (GFP) fluorescence in control cells. Optical cuts through cortical region (A) and perinuclear region (D). B and E, GFP fluorescence in cells after 30-min incubation in 20 M BFA. The formation of bright fluorescent spots (B) and large sheets (B, asterisks) in the cortical layer are shown. Bright fluorescent spots in transvacuolar strands and in the perinuclear region (E). Video files showing control cells and the effect of 30-min incubation in 20 M BFA can be seen at BFAmovies.htm. C and F, GFP fluorescence in cells after 30-min incubation in 50 M NPA. Unaltered ER in the cortical (C) and perinuclear region (F). Bars 10 m. or arrangement (Fig. 5, C and F); furthermore, no changes in the movement of small bodies were found. The effects of both NPA and BFA on the cell structures examined and on auxin accumulation are summarized in Table I. DISCUSSION The Actions of NPA and BFA on Auxin Accumulation Differ In suspension-cultured tobacco cells, NAA accumulation (inhibition of efflux) is controlled predominantly by the activity of NPA-sensitive auxin efflux carriers (Delbarre et al., 1996). Our results reveal a major discrepancy between the concentration of NPA required to saturate the inhibition of auxin efflux (1 m) and those reported to be necessary to inhibit either the BFA-sensitive cycling of PIN1 between the PM and an internal compartment (200 m; Geldner et al., 2001), or PIN1 internalization in the tir3 mutant of BIG (150 m; Gil et al., 2001). These observations suggest that the stimulation of NAA accumulation in tobacco cells by NPA (Delbarre et al., 1996; Petrášek et al., 2002; this report) is unlikely to have resulted from perturbation of efflux carrier cycling. We show here that in suspension-cultured cells of BY-2 tobacco, the stimulation of NAA accumulation by NPA was markedly reversed at concentrations of NPA around 10 to 30 m. In another tobacco cell line (VBI-0), although high concentrations of NPA (up to at least 100 m) did not cause such a reversal, abnormalities in cell division and loss of cell polarity occurred at concentrations of NPA even as low as 10 m (Petrášek et al., 2002). Therefore, at concentrations of NPA not much greater than those necessary to saturate auxin accumulation, both cell behavior and auxin transport may be perturbed by mechanisms that have nothing to do with the specific effects of NPA on the auxin efflux machinery. The inhibitory effect of NPA on auxin efflux is extremely rapid. In the suspension-cultured tobacco cells used in our experiments, the stimulation of NAA accumulation by NPA occurred without measurable time lag. This indicates that in cell suspensions, in which NPA would be expected to reach binding sites very rapidly, the inhibitory effect of the compound on auxin efflux carrier activity is very fast. This rapid response contrasts with the rather long treatment periods used in the experiments of Geldner et al. (2001; 2 h) and of Gil et al. (2001; 3 h) to study the effects of NPA and BFA on PIN1 cycling and localization. Unfortunately, no information on the kinetics of the inhibitory effect of NPA on efflux carrier cycling was provided. Consistent with its reported inhibitory effect on efflux carrier traffic to the PM (Delbarre et al., 1998; Morris and Robinson, 1998; Robinson et al., 1999; Geldner et al., 2001; see introduction), BFA also strongly promoted NAA accumulation by BY-2 cells in a concentration-dependent manner. However, as with NPA, high concentrations of BFA (above 30 m) reversed the stimulation of auxin accumulation. This biphasic response is correlated with the action of BFA on the actin cytoskeleton (discussed below). In the cell suspensions used in our experiments, the maximum stimulation of NAA accumulation observed (22.7% after 20 min at 10 m BFA) was substantially less than the maximum stimulation caused by NPA treatment (130.2% after 20 min at 1 m). These results imply that although BFA may be effective in inhibiting membrane vesicle cycling, even under conditions where the response to BFA is maximal, a small proportion of carrier proteins continues to catalyze auxin efflux across the PM. Evidence for this possibility comes from the results of Geldner et al. (2001). These Table I. Summary of the effects of NPA ( M) and BFA ( M) on cell structures and auxin accumulation in suspension-cultured 2-d-old BY-2 tobacco cells Compound Microtubules Actin Filaments Endoplasmic Reticulum Auxin Accumulation NPA a b BFA b, Structure af- a, No differences from control observed. fected and/or auxin accumulation increased. Plant Physiol. Vol. 131,
43 Petrášek et al. authors demonstrated that treatment of Arabidopsis seedling roots with 50 m BFA for 2 h resulted in internalization of PIN1. However, close examination of their Figure 1B illustrating this result (Geldner et al., 2001) suggests appreciable residual fluorescence in the PM from PIN1 markers. BFA But Not NPA Affects the Cytoskeleton Although a role for the cytoskeleton in polar auxin transport has been established (for review, see Muday, 2000), only few data are available on the state of AFs and MTs after disruption of polar auxin transport with inhibitors. Thus, we compared the effects of NPA and BFA on the arrangement and structure of the cytoskeleton under the same conditions as those used to study the effects of these compounds on NAA accumulation. In agreement with Saint-Jore et al. (2002), who reported that treatment of BY-2 cells with BFA did not affect cortical MTs and AFs, we also found no effect of BFA on cortical cytoskeletal components. However, in the perinuclear region we found that BFA caused actin to form prominent clusters. To our knowledge, this is the first report of this phenomenon in plants. The lack of information about perinuclear actin possibly stems from the fact that most previous studies have concentrated on AFs in the cortical layer of cytoplasm, and the perinuclear region has largely been overlooked (compare with Satiat-Jeunemaitre et al., 1996; Saint-Jore et al., 2002). Recently, Waller et al. (2002) reported an increased membrane association of cortical actin and bundling of cortical AFs in maize epidermal cells after BFA treatment. This suggests that under some circumstances, even cortical actin can be modified by treatment with BFA. Because phalloidin (used here for actin cytoskeleton visualization) binds preferentially to F-actin, it is likely that the newly formed actin clusters produced in the perinuclear region consist of the filamentous form of actin. A possible explanation for the formation of the perinuclear actin clusters is that actin may play a role in the process of ER-Golgi apparatus fusion after BFA treatment (compare with Ritzenthaler et al., 2002). The quantitative image analysis showed that perinuclear actin cluster formation increased with increasing concentrations of BFA up to 40 m but declined substantially with further increase in BFA concentrations to 100 m. A possible interpretation is that at high concentrations of BFA, the ER-Golgi hybrid compartment (Ritzenthaler et al., 2002) is not formed. As a consequence, AFs are unable to reorganize into clusters. The reduction in actin cluster formation at high BFA concentrations correlates with the reduced stimulation of NAA accumulation under the same conditions. One possibility is that the two processes are functionally connected. In contrast to BFA, the phytotropin NPA did not cause any changes in the arrangement of either MTs or AFs at concentrations that clearly inhibit NAA transport. The lack of effect on the arrangement of MTs is consistent with similar observations by Hasenstein et al. (1999) on maize root cells, who found that the inhibition of auxin transport by NPA was not accompanied by changes in the orientation of cortical MTs. Several reports strongly point to an association of the NBP and F-AFs (Cox and Muday, 1994; Butler et al., 1998; Hu et al., 2000). Although this association seems essential for the inhibitory action of NPA on auxin efflux, the binding of NPA to the NBP appears not to disrupt the association of the NBP with the actin cytoskeleton (Hu et al., 2000; this report). Thus, we conclude that the inhibitory action of NPA on auxin efflux from plant cells is not associated with disruption of the cytoskeletal system. BFA But Not NPA Affects the ER To follow possible mechanisms underlying the changes in perinuclear actin organization, we also investigated the structure of ER in interphase BY-2 cells. Using cells expressing a fusion protein containing GFP and ER-signaling and ER-retention amino acid (His-Asp-Glu-Leu, HDEL) sequences, both mobile particles and a static polygonal network of tubules were observed as reported for other fusion proteins containing an HDEL retention signal (Boevink et al., 1996; Haseloff et al., 1997). Because proteins containing an HDEL retention sequence might occasionally escape from the ER to the cis-golgi apparatus, where HDEL binds to a specific receptor (Boevink et al., 1998), the possibility cannot be excluded that the fluorescence signal could also be observed in the structure of the cis-golgi. However, it is unlikely that the mobile particles seen in control cells in this study are Golgi stacks because they did not move in the stop and go fashion characteristic of Golgi stacks (Nebenführ et al., 1999). One possibility is that they are the small, dilated cisternae of ER described previously in Brassicaceae and tobacco guard cells by Hawes et al. (2001). Treatment of cells with BFA resulted in the appearance of brightly fluorescing static spots at the surface of the ER sheets. Similar results were reported by Boevink et al. (1999) and Batoko et al. (2000) for the transient expression of a GFP-HDEL-containing protein. These fluorescing spots may be accumulations of GFP in the ER, but a positive identification has not yet been made (C. Hawes, personal communication). The disintegration of the ER that was observed in our experiments is in agreement with results of Henderson et al. (1994), who showed disruption of ER after 3 h of treatment with BFA in maize root cells by immunofluorescence microscopy with anti-hdel antibody. Ritzenthaler et al. (2002) reported that up to 20 min, treatment with BFA caused no visible alteration in ER morphology in BY-2 cells. Our results indicated that the first observable changes in ER-targeted GFP distribution 260 Plant Physiol. Vol. 131, 2003
44 Inhibition of Auxin Efflux in BY-2 Tobacco Cells in BY-2 cells can be seen in as little as 5 min after BFA application, when disintegration of the tubular network and formation of fluorescent spots started. In contrast to BFA treatments, NPA had no effect on ER-targeted GFP distribution in BY-2 cells. To our knowledge, no other data about phytotropin effects on ER are yet available. The NPA Enigma Geldner et al. (2001) reported that TIBA (and possibly also other auxin transport inhibitors) prevented the BFA-induced internalization of PIN1 and the traffic of internalized PIN1 to the PM after the BFA washout. Because similar effects of TIBA were observed on the cycling of a PM-ATPase and of the syntaxin KNOLLE, these authors suggested that auxin transport inhibitors affect auxin efflux by generally interfering with membrane trafficking processes. To generalize from these findings, however, may be premature, not least because TIBA is not a good representative of auxin transport inhibitors. First, TIBA does not fulfill the structural requirements of typical phytotropins and acts as a weak auxin antagonist (Katekar and Geissler, 1980). Although it inhibits auxin transport (Katekar and Geissler, 1980; Rubery, 1990), its high-affinity binding to maize microsomal preparations is only partially displaced by NPA (Depta et al., 1983), suggesting different loci of action of TIBA and phytotropins. Second, it has long been established that TIBA is itself a weak auxin, which unlike NPA, undergoes carriermediated polar transport on carriers that can be competed by IAA, 2,4-dichlorophenoxyacetic acid, and NAA (Depta and Rubery, 1984). Third, work in our laboratory has shown that in BY-2 tobacco cells, the stimulation of NAA accumulation (2 nm) is saturated by as little as 1 m TIBA (H. Slizowska, M. Elčkner, and E. Zažímalová, unpublished data). Last, and more importantly, unlike NPA and IAA, at the concentration of 25 m used by Geldner et al. (2001), TIBA causes substantial cytoplasmic acidification (Depta and Rubery, 1984). This, in turn, is likely to have nonspecific side effects on many cellular processes, possibly including secretory mechanisms. We believe that there are other good reasons to be cautious in accepting the suggestion that auxin transport inhibitors act by generally interfering with vesicle traffic and turnover. The concentration of NPA stated to be necessary to bring about a similar reduction in PIN1 cycling to that caused by 25 m TIBA (200 m NPA; Geldner et al., 2001) is about two orders of magnitude greater than the concentration of NPA required to saturate inhibition of auxin efflux (1 3 m; Petrášek et al., 2002; this report). Also, in suspension-cultured tobacco cells, concentrations of NPA exceeding about 50 m reduce the stimulation of NAA accumulation substantially, possibly as a result of nonspecific side effects on the cells unrelated to the specific regulation of auxin efflux (Petrášek et al., 2002; this report). Finally, as discussed above, NPA has no effect on the arrangement of the cytoskeleton or ER in BY-2 cells, even at concentrations well above those that saturate the inhibition of auxin efflux. Therefore, NPA cannot inhibit actin-dependent vesicle traffic in general by an indirect action on the structure of the cytoskeleton and ER. In conclusion, although the vesicle trafficking inhibitor BFA mimics some of the physiological effects of the phytotropin NPA, particularly insofar as they both inhibit carrier-mediated auxin efflux across the PM, our results clearly suggest that they do so by affecting different cellular mechanisms. MATERIALS AND METHODS Plant Material Cells of tobacco (Nicotiana tabacum L. cv Bright-Yellow 2) line BY-2 (Nagata et al., 1992) were cultivated in darkness at 26 C on an orbital incubator (IKA KS501, IKA Labortechnik, Staufen, Germany; 120 rpm, orbital diameter 30 mm) in liquid medium (3% [w/v] Suc, 4.3 g L 1 Murashige and Skoog salts, 100 mg L 1 inositol, 1 mg L 1 thiamin, 0.2 mg L 1 2,4-dichlorophenoxyacetic acid, and 200 mg L 1 KH 2 PO 4 [ph 5.8]) and subcultured weekly. Stock BY-2 calli were maintained on media solidified with 0.6% (w/v) agar and subcultured monthly. Transgenic cells and calli were maintained on the same media supplemented with 100 g ml 1 kanamycin and 100 g ml 1 cefotaxim. All chemicals were obtained from Sigma (St. Louis) unless otherwise stated. Transformation of BY-2 Cells The basic transformation protocol of An (1985) was used. A 2-mL aliquot of 3-d-old BY-2 cells was co-incubated for 3 d with 100 L of an overnight culture of Agrobacterium tumefaciens strain C58C1 carrying pbin m-gfp5-er plant binary vector (gift of Dr. Jim Haseloff, University of Cambridge, UK). It codes for ER-localized GFP variant mgfp5-er, a thermotolerant derivative of mgfp4-er (Haseloff et al., 1997), and contains a C-terminal ER retention signal sequence (HDEL). Incubated cells were then washed three times in 50 ml of liquid medium containing 100 g ml 1 cefotaxim and plated onto solid medium containing 100 g ml 1 kanamycin and 100 g ml 1 cefotaxim. Kanamycin-resistant colonies appeared after 3 to 4 weeks of incubation in darkness at 27 C. Cell suspension cultures established from these were maintained as described above, with the addition of 100 g ml 1 kanamycin and 100 g ml 1 cefotaxim to the cultivation medium. Effects of NPA and BFA on Cytoskeleton Arrangement Appropriate volumes of a 25 mm stock solution of BFA in 96% (w/v) ethanol were added to cell cultures to give final concentrations of 20, 40, and 100 m. NPA (synthesized in the Institute of Experimental Botany, Prague; compare with Petrášek et al., 2002) was added to cell cultures from 5 mm stock solution in 96% (w/v) ethanol to a final concentration of 50 m (determined by reference to NPA concentration studies; see above). Equivalent volumes of 96% (w/v) ethanol were added to all control cultures. Cell cultures were treated with BFA or NPA for 30 min with continuous shaking at room temperature (approximately 25 C) before microscopic examination (see below). When required, washout of BFA was performed with fresh cultivation medium after filtration of treated suspensions on 50-mmdiameter cellulose filter paper discs on a Nalgene filter holder (Nalge Company, Rochester, NY). Aliquots of 10 ml of cell suspension were washed three times (10 min each time) in 50 ml of fresh cultivation medium, and cells were examined immediately. Plant Physiol. Vol. 131,
45 Petrášek et al. Visualization of AFs AFs were visualized by the method of Kakimoto and Shibaoka (1987) modified according to Olyslaegers and Verbelen (1998). Filtered cells were fixed for 10 min in 1.8% (w/v) paraformaldehyde (PFA) in standard buffer (50 mm PIPES [ph 7.0], supplemented with 5 mm MgCl 2, and 10 mm EGTA). After a subsequent 10-min fixation in standard buffer containing 1% (v/v) glycerol, cells were rinsed twice for 10 min with standard buffer. Then, 0.5 ml of the resuspended cells were incubated for 35 min with the same volume of 0.66 m TRITC-phalloidin prepared freshly from 6.6 m stock solution in 96% (w/v) ethanol by dilution (1:10 [v/v]) in phosphatebuffered saline (PBS; 0.15 m NaCl, 2.7 mm KCl, 1.2 mm KH 2 PO 4, and 6.5 mm Na 2 HPO 4 [ph 7.2]). Cells were than washed three times for 10 min in PBS and observed immediately. Visualization of MTs MTs were visualized as described by Wick et al. (1981) with the modifications described by Mizuno (1992). After 30 min of prefixation in 3.7% (w/v) PFA in MT-stabilizing buffer consisting of 50 mm PIPES, 2 mm EGTA, and 2 mm MgSO 4 (ph 6.9) at 25 C, the cells were subsequently fixed in 3.7% (w/v) PFA and 1% (w/v) Triton X-100 in MT-stabilizing buffer for 20 min. After treatment with an enzyme solution (1% [w/v] macerozyme and 0.2% [w/v] pectinase) for 7 min at 25 C, the cells were attached to poly-l-lyscoated coverslips and treated with 1% (w/v) Triton X-100 in MT-stabilizing buffer for 20 min. Subsequently, the cells were treated with 0.5% (w/v) bovine serum albumin in PBS and incubated with a monoclonal mouse antibody against -tubulin (DM 1A, Sigma) for 45 min at 25 C (dilution 1:500 [v/v] in PBS). After washing with PBS, a secondary fluorescein isothiocyanate (FITC)-conjugated anti-mouse antibody (Sigma), diluted 1:80 (v/v) in PBS, was applied for 1 h at 25 C. Coverslips with cells were carefully washed in PBS, rinsed with water, and embedded in Mowiol solution (Polysciences, Warrington, PA). Microscopy and Image Analysis Both fixed and live preparations were observed with an epifluorescence microscope (Eclipse E600, Nikon, Tokyo) equipped with appropriate filter sets for FITC and TRITC fluorescence detection. mgfp5-er fluorescence was observed using the FITC filter set. Images and time lapse scans were grabbed with a monochrome integrating CCD camera (COHU 4910, COHU Inc., San Diego) and digitally stored. The organization of both the cytoskeleton and ER were studied microscopically in at least 10 independent experiments. Representative images in Figures 3 and 5 show an arrangement typical for around 1,000 cells evaluated in each treatment. Subtle differences reflect the variability of staining pattern and appearance of cells. After this careful microscopic examination, LUCIA image analysis software (Laboratory Imaging, Prague) was used for the evaluation of the effect of BFA on perinuclear actin aggregation. Images of TRITC-phalloidinstained AFs were transformed to complementary colors (Fig. 4, A and C) and a circular measuring frame was applied interactively over the perinuclear region (Fig. 4, B and D). The ODV parameter was measured. The ODV parameter is the sd of optical density values under the circular measuring frame, where the bigger the ODV, the higher the aggregation of actin. These optical density values reflect the relative fluorescence intensities of individual pixels. The method measures the coherency of the fluorescent signal in the perinuclear region; the less coherent the signal, the greater the extent of actin aggregation. Approximately 300 cells in 10 optical fields were assessed for each sample. Auxin Accumulation Measurement Auxin accumulation by cells was measured according to the method of Delbarre et al. (1996), modified by Petrášek et al. (2002). The accumulation by the cells of [ 3 H]NAA (specific radioactivity 935 GBq mmol 1, synthesized at the Isotope Laboratory, Institute of Experimental Botany, Prague), was measured in 0.5-mL aliquots of cell suspension (cell density about cells ml 1, as determined by counting cells in Fuchs-Rosenthal hemocytometer). Each cell suspension was filtered, resuspended in uptake buffer (20 mm MES, 40 mm Suc, and 0.5 mm CaSO 4, ph adjusted to 5.7 with KOH), and equilibrated for 45 min with continuous orbital shaking. Equilibrated cells were collected by filtration, resuspended in fresh uptake buffer, and incubated on the orbital shaker for 1.5 h in darkness at 25 C. [ 3 H]NAA was added to the cell suspension to give a final concentration of 2 nm. After a timed uptake period (depending on experiment, see above), 0.5-mL aliquots of suspension were withdrawn and accumulation of label was terminated by rapid filtration under reduced pressure on 22-mm-diameter cellulose filters. The cell cakes and filters were transferred to scintillation vials, extracted in ethanol for 30 min, and radioactivity was determined by liquid scintillation counting (Packard Tri-Carb 2900TR scintillation counter, Packard Instrument Co., Meriden, CT). Counts were corrected for surface radioactivity by subtracting counts obtained for aliquots of cells collected immediately after the addition of [ 3 H]NAA. Counting efficiency was determined by automatic external standardization, and counts were corrected automatically. NPA or BFA were added as required from ethanolic stock solutions to give the appropriate final concentration (see above). In time course experiments, aliquots of cell suspension were removed at timed intervals varying from 0 to 40 min from the start of experiments; the concentration dependence of auxin accumulation in response to NPA or BFA was determined after a 20-min uptake period. Metabolism of Labeled Compounds Possible distortion of the results of auxin accumulation studies by metabolism of the [ 3 H]NAA taken up by the cells was checked. Cells of BY-2 were incubated for 30 min as described in the presence of 2 nm [ 3 H]NAA. At the end of the incubation period, 10-mL aliquots of the incubated suspensions were quickly filtered on paper with gentle suction, washed rapidly with 5 ml of uptake buffer, and the cell cake was transferred to 2 ml of prechilled ethanol and stored at 80 C until required. Cell debris was removed by filtration under gentle pressure through cellulose filters. Radioactive compounds in the extracts were separated by chromatography on cellulose thin-layer plates (POLYGRAM CEL 300 UV 254, Macherey-Nagel, Düren, Germany), together with samples of the labeled auxins. The plates were developed in three independent solvent systems: (a) isopropanol:26% (v/v) ammonia:water (10:1:1 [v/v]), (b) chloroform:ethanol:glacial acetic acid (95:1:5 [v/v]), and (c) chloroform:ethanol:glacial acetic acid (75:20:5 [v/v]). Each chromatogram strip was cut into 20 sequential segments, eluted in ethanol, and counted by liquid scintillation counting. ACKNOWLEDGMENTS The authors thank Dr. Jim Haseloff (University of Cambridge, UK) for the pbin m-gfp5-er binary vector, and Miss Andrea Hourová (Institute of Experimental Botany, Prague) for her excellent technical assistance. Received August 9, 2002; returned for revision September 10, 2002; accepted October 12, LITERATURE CITED An G (1985) High efficiency transformation of cultured tobacco cells. Plant Physiol 79: Batoko H, Zheng HQ, Hawes C, Moore I (2000) A Rab1 GTPase is required for transport between the endoplasmic reticulum and Golgi apparatus and for normal Golgi movement in plants. Plant Cell 12: Bennett MJ, Marchant A, Green HG, May ST, Ward SP, Millner PA, Walker AR, Schulz B, Feldmann KA (1996) Arabidopsis AUX1 gene: a permease-like regulator of root gravitropism. Science 273: Bernasconi P, Patel BC, Reagan JD, Subramanian MV (1996) The N-1- naphthylphthalamic acid-binding protein is an integral membrane protein. Plant Physiol 111: Boevink P, Martin B, Oparka K, Santa Cruz S, Hawes C (1999) Transport of virally expressed green fluorescent protein through the secretory pathway in tobacco leaves is inhibited by cold shock and brefeldin A. Planta 208: Boevink P, Oparka K, Santa Cruz S, Martin B, Betteridge A, Hawes C (1998) Stacks on tracks: The plant Golgi apparatus traffics on an actin/er network. Plant J 15: Boevink P, Santa Cruz S, Hawes C, Harris N, Oparka KJ (1996) Virusmediated delivery of the green fluorescent protein to the endoplasmic reticulum of plant cells. Plant J 10: Plant Physiol. Vol. 131, 2003
46 Inhibition of Auxin Efflux in BY-2 Tobacco Cells Butler JH, Hu SQ, Brady SR, Dixon MW, Muday GK (1998) In vitro and in vivo evidence for actin association of the naphthylphthalamic acidbinding protein from zucchini hypocotyls. Plant J 13: Cande WZ, Goldsmith MHM, Ray PM (1973) Polar auxin transport and auxin-induced elongation in the absence of cytoplasmic streaming. Planta 111: Cox DN, Muday GK (1994) NPA binding activity is peripheral to the plasma membrane and is associated with the cytoskeleton. Plant Cell 6: Davies PJ (1995) The plant hormone concept: concentration, sensitivity and transport. In PJ Davies, ed, Plant Hormones: Physiology, Biochemistry and Molecular Biology, Ed 2. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp Delbarre A, Muller P, Guern J (1998) Short-lived and phosphorylated proteins contribute to carrier-mediated efflux, but not to influx, of auxin in suspension-cultured tobacco cells. Plant Physiol 116: Delbarre A, Muller P, Imhoff V, Guern J (1996) Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1-acetic acid, and indole-3-acetic acid in suspensioncultured tobacco cells. Planta 198: Depta H, Eisele K-H, Hertel R (1983) Specific inhibitors of auxin transport: action on tissue segments and in vitro binding to membranes from maize coleoptiles. Plant Sci Lett 31: Depta H, Rubery PH (1984) A comparative study of carrier participation in the transport of 2,3,5-triiodobenzoic acid, indole-3-acetic acid, and 2,4- dichlorophenoxyacetic acid by Cucurbita pepo L. hypocotyl segments. J Plant Physiol 115: Dixon MW, Jacobson JA, Cady CT, Muday GK (1996) Cytoplasmic orientation of the naphthylphthalamic acid-binding protein in zucchini plasma membrane vesicles. Plant Physiol 112: Friml J, Palme K (2002) Polar auxin transport old questions and new concepts? Plant Mol Biol 49: Gälweiler L, Guan C, Müller A, Wisman E, Mendgen K, Yephremov A, Palme K (1998) Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282: Geldner N, Friml J, Stierhof YD, Jürgens G, Palme K (2001) Auxin transport inhibitors block PIN 1 cycling and vesicle trafficking. Nature 413: Gil P, Dewey E, Friml J, Zhao Y, Snowden KC, Putterill J, Palme K, Estelle M, Chory J (2001) BIG: a calossin-like protein required for polar auxin transport in Arabidopsis. Genes Dev 15: Goldsmith MHM (1977) The polar transport of auxin. Annu Rev Plant Physiol 28: Haseloff J, Siemering KR, Prasher DC, Hodge S (1997) Removal of a cryptic intron and subcellular localisation of green fluorescent protein are required to mark transgenic Arabidopsis plants brightly. Proc Natl Acad Sci USA 94: Hasenstein KH, Blancaflor EB, Lee JS (1999) The microtubule cytoskeleton does not integrate auxin transport and gravitropism in maize roots. Physiol Plant 105: Hawes C, Saint-Jore C, Martin B, Zheng HQ (2001) ER confirmed as the location of mystery organelles in Arabidopsis plants expressing GFP! Trends Plant Sci 6: Henderson J, Satiat-Jeunemaitre B, Napier R, Hawes C (1994) Brefeldin A-induced disassembly of the Golgi apparatus is followed by disruption of the endoplasmic reticulum in plant cells. J Exp Bot 45: Hu SQ, Brady SR, Kovar DR, Staiger CJ, Clark GB, Roux SJ, Muday GK (2000) Identification of plant actin binding proteins by F-actin affinity chromatography. Plant J 24: Kakimoto T, Shibaoka H (1987) Actin filaments and microtubules in the preprophase band and phragmoplast of tobacco cells. Protoplasma 140: Katekar GF, Geissler AE (1980) Auxin transport inhibitors: IV. Evidence of a common mode of action for a proposed class of auxin transport inhibitors: the phytotropins. Plant Physiol 66: Li HM, Altschmied L, Chory J (1994) Arabidopsis mutants define downstream branches in the phototransduction pathway. Genes Dev 8: Lomax TL, Muday GK, Rubery PH (1995) Auxin transport. In PJ Davies, ed, Plant Hormones: Physiology, Biochemistry and Molecular Biology, Ed 2. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp Luschnig C (2001) Auxin transport: why plants like to think BIG. Curr Biol 11: Mizuno K (1992) Induction of cold stability of microtubules in cultured tobacco cells. Plant Physiol 100: Morris DA (2000) Transmembrane auxin carrier systems dynamic regulators of polar auxin transport. Plant Growth Regul 32: Morris DA, Robinson JS (1998) Targeting of auxin carriers to the plasma membrane: differential effects of brefeldin A on the traffic of auxin uptake and efflux carriers. Planta 205: Morris DA, Rubery PH, Jarman J, Sabater M (1991) Effects of inhibitors of protein synthesis on transmembrane auxin transport in Cucurbita pepo L. hypocotyl segments. J Exp Bot 42: Müller A, Guan CH, Gälweiler L, Tänzler P, Huijser P, Marchant A, Parry G, Bennett M, Wisman E, Palme K (1998) AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J 17: Muday GK (2000) Maintenance of asymmetric cellular localization of an auxin transport protein through interaction with the actin cytoskeleton. J Plant Growth Regul 19: Muday GK, DeLong A (2001) Polar auxin transport: controlling where and how much. Trends Plant Sci 6: Muday GK, Murphy AS (2002) An emerging model of auxin transport regulation. Plant Cell 14: Nagata T, Nemoto Y, Hasezava S (1992) Tobacco BY-2 cell line as the Hela cell in the cell biology of higher plants. Int Rev Cytol 132: 1 30 Nebenführ A, Gallagher LA, Dunahay TG, Frohlick JA, Mazurkiewicz AM, Meehl JB, Staehelin LA (1999) Stop-and-go movements of plant Golgi stacks are mediated by the acto-myosin system. Plant Physiol 121: Olyslaegers G, Verbelen JP (1998) Improved staining of F-actin and colocalization of mitochondria in plant cells. J Microsc Oxford 192: Petrášek J, Elčkner M, Morris DA, Zažímalová E (2002) Auxin efflux carrier activity and auxin accumulation regulate cell division and polarity in tobacco cells. Planta 216: Raven JA (1975) Transport of indoleacetic acid in plant cells in relation to ph and electrical potential gradients, and its significance for polar IAA transport. New Phytol 74: Richards S, Hillman T, Stern M (1996) Mutations in the Drosophila pushover gene confer increased neuronal excitability and spontaneous synaptic vesicle fusion. Genetics 142: Ritzenthaler C, Nebenführ A, Movafeghi A, Stussi-Garaud C, Behnia L, Pimpl P, Staehelin LA, Robinson DG (2002) Reevaluation of the effects of brefeldin A on plant cells using tobacco bright yellow 2 cells expressing Golgi-targeted green fluorescent protein and COPI antisera. Plant Cell 14: Robinson JS, Albert AC, Morris DA (1999) Differential effects of brefeldin A and cycloheximide on the activity of auxin efflux carriers in Cucurbita pepo L. J Plant Physiol 155: Rubery PH (1990) Phytotropins: receptors and endogenous ligands. Symp Soc Exp Biol 44: Rubery PH, Sheldrake AR (1974) Carrier-mediated auxin transport. Planta 188: Ruegger M, Dewey E, Hobbie L, Brown D, Bernasconi P, Turner J, Muday G, Estelle M (1997) Reduced naphthylphthalamic acid binding in the tir3 mutant of Arabidopsis is associated with a reduction in polar auxin transport and diverse morphological defects. Plant Cell 9: Satiat-Jeunemaitre B, Steele C, Hawes C (1996) Golgi-membrane dynamics are cytoskeleton dependent: a study on Golgi stack movement induced by brefeldin A. Protoplasma 191: Saint-Jore CM, Evins J, Batoko H, Brandizzi F, Moore I, Hawes C (2002) Redistribution of membrane proteins between the Golgi apparatus and endoplasmatic reticulum in plants is reversible and not dependent on cytoskeletal networks. Plant J 29: Sussman MR, Gardner G (1980) Solubilization of the receptor for N-1- naphthylphthalamic acid. Plant Physiol 66: Swarup R, Friml J, Marchant A, Ljung K, Sandberg G, Palme K, Bennett M (2001) Localization of the auxin permease AUX1 suggests two functionally distinct hormone transport pathways operate in the Arabidopsis root apex. Genes Dev 15: Waller F, Riemann M, Nick P (2002) A role of actin-driven secretion in auxin-induced growth. Protoplasma 219: Wick SM, Seagull RW, Osborn M, Weber K, Gunning BES (1981) Immunofluorescence microscopy of organized microtubule arrays in structurally stabilized meristematic plant cells. J Cell Biol 89: Wilkinson S, Morris DA (1994) Targeting of auxin carriers to the plasma membrane: effects of monensin on transmembrane auxin transport in Cucurbita pepo L. tissue. Planta 193: Plant Physiol. Vol. 131,

47 3.2 Addresing the role of cytoskeleton in the endocytosis of PIN1 auxin efflux carrier in BY-2 cells This chapter describes the participation of cytoskeleton in the vesicle trafficking processes. Inhibitors of vesicle trafficking and cytoskeletal dynamic were used and their in vivo effect monitored by FM 4-64 uptake and auxin efflux carrier PIN1-GFP localization in tobacco BY-2 cells. This approach allowed to determine the requirement of cytoskeleton for the endocytosis in suspension-cultured tobacco cells. Our description of the action of these inhibitors in BY-2 cells can be further utilized as a tool to study protein localization in the plant secretory and endocytic pathways. The chapter is presented here in the form of the extended text that is in the preparation for the publication. My participation was in the experimental part, in vivo studies, drug treatments, confocal microscopy, analyses and interpretation of the data, designing and coordination of the experiments and writing of the manuscript. Endocytosis of FM 4-64 styryl dye by tobacco BY-2 cells 27
48 28
49 Abstract The process of endocytosis involving the formation and internalization of the plasma membrane (PM) vesicles has been shown to play an important role in the positioning of PM auxin transporters, namely PIN auxin efflux carriers. However, there is still limited amount of data on the role of cytoskeleton in this process in general and also in the positioning of PINs in particular. The goal of this in vivo study was to characterize the impact of endocytic inhibitors and auxins on the endocytosis itself and PIN1 targeting in particular and to determine the cytoskeletal requirements for these processes in BY-2 tobacco cells. The inhibitors of various stages of endocytosis as well as auxins, previously reported to inhibit endocytosis, were used and followed microscopically their impact on the uptake of FM 4-64 dye and PIN1-GFP distribution after brefeldin A treatment. It is shown here that the uptake of FM 4-64 was effectively blocked by tyrosine kinase inhibitor Tyrphostin A23 suggesting that this inhibitor does not interfere only with the loading of clathrin-coated vesicles, but it affects also other trafficking processes dependent on tyrosine kinase activity. Dynasore, the specific inhibitor of clathrin-mediated endocytosis in mammalian cells was used in plants for the first time here and its inhibitory effect on both FM 4-64 uptake and formation of BFA-induced PIN1 aggregates confirmed clathrin-driven endocytosis of PIN1 in BY-2 cells. By using cytoskeletal drugs it is shown that clathrin-mediated endocytosis of FM 4-64 is dependent on both actin filaments and microtubules. Moreover, previously reported remodelling of actin filaments in the perinuclear region after BFA treatment is shown here to include active cooperation between actin cytoskeleton and membrane vesicles, suggesting the role of actin filaments in the endocytosis. Introduction Endocytosis involves internalization of the PM along with membrane proteins and lipids. It is implicated in various processes initiated at the cell surface. These include uptake of ions, nutrients and hormones, signalling, maintenance of PM proteins and lipids and many other processes essential for the membrane homeostasis, cell-to-cell communication and the contact external environment. In animal cells, PM proteins are continuously internalized by variety of mechanisms. After internalization into the vesicles, the proteins are transported to endosomes where they undergo sorting. From the endosomes they are either recycled to the PM or kept in the endocytic pathway through late endosome to the lysosomes for degradation (Robinson et al. 2008). It has been long-term debate whether plant cells possess endocytosis or not, mainly 29
50 form the fact that the turgor pressure in plants would make endocytosis difficult) (Cram 1980). Up to now the importance of endocytosis for plant cells has been shown in the number of studies. There are two major types of endocytosis in plant cells: fluid phase endocytosis (FPE) and receptor-mediated endocytosis (RME). The major difference among them is in the kinetics of the uptake. FPE demonstrates linear uptake kinetics without an evidence of saturation and the binding to the cell surface depends on the temperature. RME saturates with ligand concentration and is not temperature dependent (Aniento and Robinson, 2005). The receptor serves like a sensor for transduction of a signal to the downstream signalling cascades not only at the cell surface but subcellular trafficking of the PM receptors has to be considered as integral part of the signal transduction cascades (von Zastrow and Sorkin, 2007). Despite the fact that there has been published several papers about candidates of PM proteins that are internalized into the cells in plants (Holstein, 2002; Geldner, 2004) the demonstration of internalization of a protein endocytosis through clathrin-coated pits and their recycling to the PM was published quite recently (Geldner and Jürgens 2006, Dhonukshe et al. 2007, Perez-Gomez 2007). To test whether plant cells posses molecular machinery necessary for RME, human transferin receptor (htfr) was expressed in Arabidopsis protoplasts and results indicated that htfr has probably similar trafficking properties as in animal cells and those cycles between PM and endosomal compartment (Ortiz-Zapater et al. 2006). The update on RME and trafficking in plants considering ligand-regulated receptor endocytosis, putting endosomal signalling in plants into more general evolutionary context has been published recently (Geldner and Robatzek 2008). The polarized intracellular localization of auxin efflux proteins (PINs) is ensured by the co-ordinated trafficking of PIN-containing vesicles to various PM domains. Interestingly, the localization of newly synthesized PIN1 seems to be apolar and random within PM until the polarization starts via recycling through endosomal compartment (Dhonukshe et al. 2008). The regulation of the amount and positioning of PINs within the PM is achieved by constitutive cycling between PM a endosomal compartment (Geldner et al. 2001). This process is inhibited with brefeldin A (BFA) that interferes with the activity of GNOM, the adenosine rybosilation factor (ARF)-guanine exchange factor (GEF). As a results, the PIN proteins are found in the endosomal compartments (Steinmann 1999). Indeed, the application of BFA decrease the auxin efflux in tobacco cells (Petrášek et al., 2003; chapter 3.1. of this thesis) Moreover, endocytosis of membrane vesicles is inhibited by auxin itself, which leads to the promotion of its own efflux (Paciorek et al. 2005). The polarization of auxin transporters is linked to the vesicle trafficking (Steinmann et al. 1999, Geldner et al. 2003, Kleine-Vehn et al. 2006) and their constitutive cycling in the 30
51 endosomal space seems to be actomyosin-dependent (Geldner et al. 2001; Holweg et al 2004; Holweg 2007). Moreover, it seems that with respect to the cytoskeleton the apical-basal localization of PIN proteins follows distinct pathways as suggested by experiments in Arabidopsis thaliana roots. Here, the process of cycling and translocation of endosomes with PIN proteins from one membrane domain to the other, so-called transcytosis (Kleine-Vehn et al. 2008a), has been shown to involve actin filaments in the apical-basal targeting of PINs and microtubules preferentially in basal targeting (Kleine-Vehn et al. 2008b). The participation of microtubules in PIN positioning was proposed earlier using anti-tubulin drugs (Boutté et al. 2006). However the information about mechanism of cytoskeleton participation in auxin efflux carrier trafficking and targeting is limited. Mutual interaction between cytoskeleton and auxin transport and between cytoskeleton and its involvement in the localization of auxin transporters is generally expected to play a role on two levels. These are level of fast responses (nongenomic) and level of differential gene expression and downstream events playing a role in long-term interactions. In principle, there are several approaches that can be used to investigate this topic. One of the most fruitful approach includes the treatment of cells with cytoskeletal drugs and inhibitors of vesicle trafficking and subsequent tracking of their effect on the endocytosis in general and/or on particular protein transport and distribution within the cell. Tyrphostins, the structural analogues of tyrosine are inhibitors of tyrosine kinases and function by binding to the active sites of the enzymes. Some of them, for example tyrphostin A23, have been reported as inhibitors of endocytosis via inhibiting the interaction of YXXΦ motif (tyrosine-containing motif) of the receptor protein with μ2 subunit of adaptor complexes that link the cytosolic domains of integral membrane proteins to the clathrin coat involved in vesicle formation. So cold control tyrphostins (as tyrphostin A51) are active as inhibitors of tyrosine kinase activity, but they are incapable of inhibiting the YXXΦ motif/μ2 interaction and they did not inhibit endocytosis (reviewed by Robinson et al. 2008). Moreover Tyrphostin A23 (Tyr A23) has been reported as potent broad range protein tyrosine kinase inhibitor (Levitzki and Gilon 1991) next to Tyrphostin A51 (Tyr A51), which has been reported as potent inhibitor of EGFR (epidermal growth factor receptor) in animal cells (Levitzki 1990). These data are consistent with Tyr A23 inhibition of the YXXΦ motif/μ2 interaction in intact cells and with the possibility that different tyrphostins may be used to inhibit specific membrane trafficking events in eukaryotic cells (Banbury et al. 2003). In plants, there have been reported use of tyrphostin as an inhibitors of recruitment of endocytic cargo into clathrin mediated pathway in Arabidopsis protoplasts (Ortiz-Zapater et al. 2006) 31
52 and inhibitors of PIN2 protein internalization in Arabidopsis roots without affecting subcellular trafficking (Dhonukshe et al. 2007). Wortmannin is known inhibitor of phosphoinositol-3-kinase (PI3-kinase) activity. PI3- kinase is essential for the formation of internal vesicles and for the attachment of the small subunit of retromer and the sortin nexins in yeast and mammals (Robinson et al and refs. therein). Morphologically, wortmannin by prevention of retrograde transport (Poupon et al. 2003) is known to cause a dilation or swelling of multivesicular bodies (MVBs) in mammalian cells (Reaves et al.1996, Shpetner et al. 1996, Bright et al. 2001) and PVCs (prevacuolar compartment)/mvbs in plant cells (Tse et al. 2004, Jaillais et al. 2006). There has been long-term debate what is the source of membrane for rapid enlargement of these organelles. Wortmannin-induced homotypic fusion of MVBs/PVCs in BY-2 suspension cells has been demonstrated recently (Wang et al. 2009). Wortmannin is also supposed to be the inhibitor of protein trafficking downstream of the internalization event at the PM (Aniento and Robinson 2005) and it has been shown to inhibit protein sorting to the vacuole (Matsuoka et al., 1995). It also inhibits the uptake of FM 1-43 dye in BY-2 suspension cells (Emans et al., 2002). Filipin specifically binds sterols (Miller 1984) and in higher doses it has been reported to shift or deplete sterols in membranes of animal cells, leading to the inhibition of endocytosis (Marella et al. 2002). In plant cells, filipin also binds to sterols (Grebe et al. 2003) and inhibits FM4-64 uptake, as well as the endocytosis of some PM proteins (AUX1, PIN1) in Arabidopsis root cells (Kleine-Vehn et al. 2006). Dynasore is a small molecule that has been reported recently as highly specific, cellpermeable inhibitor of dynamin GTPase function. It blocks completely compensatory synaptic vesicle (clathrin-mediated) endocytosis (Newton et al., 2006). Two types of coated pit intermediates accumulate during dynasore treatment, U-shaped, half formed pits and O- shaped, fully formed pits that are captured while pinching off. Thus, dynamin acts at two steps during clathrin coat formation; GTP hydrolysis is probably needed at both steps (Marcia et al. 2006). Despite the fact, that in plants the clathrin-mediated endocytosis has been proposed (Ortiz-Zapater et al. 2006, Geldner and Jürgens 2006, Dhonukshe et al. 2007, Perez- Gomez 2007) the use of dynasore in plants has not been reported so far. Brefeldin A inhibits the function of ARF-GEF GTPases by interacting with their GEFs (Jackson and Casanova 2000). In mammalian and yeast cells are ARF GTPases involved in the formation of transport vesicles by recruiting coat proteins to intracellular membranes. BFA affects COP I coat formation, participating in retrograde transport and inhibiting transport of proteins from ER to Golgi and inducing protein relocation from the GA to ER. 32
53 Moreover, there have been described so-called BFA-compartments, structures formed from trans golgi network (TGN) and endosomal elements which remained after the reabsorbtion of GA into ER. BFA may also act on endosomes and induce endosomes to become tubulated in mammalian cells (Lippincott-Schwartz et al.1991). In plant cells BFA induced the formation of GA-ER hybrid compartments (Ritzenthaler et al. 2002), BFA-compartments (Satiat- Jeunemaitre et al., 1996; Tse et al. 2004) or the loss of Golgi cis-cisternae (Hess et al., 2000). Moreover, BFA induces endosomal compartments aggregation in Arabidopsis root cells (Geldner et al. 2003, Grebe et al. 2003). BFA-compartments were found to surround Golgi stacks, indicating that early steps of vesicle trafficking stay probably intact during the BFA action and that the BFA-induced aggregates are considered to be endosomes. Different reaction upon BFA treatment is caused by the sensitivity of individual classes of ARF-GEFs residing in ER, GA or TGN/EE. It seems that GNOM demonstrates a new class of ARF- GEFs, specific for the endosomal recycling in plants and that its homologue GNOM-LIKE play the role in the ER-GA transport (Richter et al. 2007). Recently published study in BY-2 cells demonstrates that BFA induce TGN-derived as well as Golgi-derived aggregates and these are physically different from each other, suggesting that Golgi and TGN are probably functionally distinct organelles in plants (Lam et al. 2009). Interestingl, the presence of PMderived sterols within the compartments confirms running endocytosis during the BFA inhibition (Grebe et al. 2003). Indeed, the presence of cell wall polysaccharides in the BFA compartment (Satiat-Jeunemaitre and Hawes, 1992) may reflect the accumulation of secretory vesicles, rather than being interpreted as endocytic in origin (Baluška et al. 2002). Nevertheless, the huge number of vesicles constituting the BFA compartment in Arabidopsis roots can only be the result of a continual vesicle input from both secretory and endocytic pathways. Also plant hormone auxin was described as an inhibitor of endocytosis in plants. Naturally occurring indole-3-acetic acid (IAA) as well as its synthetic analogues naphthalene- 1-acetic acid (1-NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D) prevented internalization of PINs, PM ATPse and cell wall pectins in response to BFA and decreased FM 4-64 endocytic tracer uptake in Arabidopsis thaliana roots. This mode of auxin action leads to the promotion of its own efflux by the feedback control mechanism of abundance its transporters within PM (Paciorek et al. 2005). Such an inhibitory effect of auxin can be used as powerful tool for uncovering the nongenomic fast effects of auxin itself. There have been published only quite a few studies searching for the mechanism of endocytosis and protein trafficking in plants that use cytoskeletal drugs. Cytochalasin D (Cyt 33
54 D), drug known to disrupt actin filaments, was used to suggest the role of intact cytoskeleton for the function of auxin efflux carrier as well as for its trafficking (Butler et al. 1998; Geldner et al. 2001). The accumulation of PIN1 in the intracellular compartments upon BFA treatment is reversible and this reversibility is uncoupled in the presence of Cyt D, indicating that this process is actin-dependent (Geldner et al. 2001). In pollen tubes, depolymerisation of actin filaments with cytochalasin D completely abolished plasma membrane retrieval of fluorescent dye, whereas depolymerization of microtubules with oryzalin had no effect (Lisboa et al. 2008). In this study inhibitors of endocytosis and auxins were used in BY-2 cells and their effect tested on the dynamics of FM 4-64 uptake and specifically on the formation of BFA-induced aggregations of PIN1-GFP. By this approach it is suggested that clathrin-mediated endocytosis of FM4-64 is dependent on both actin and microtubular cytoskeleton and that remodelling of actin filaments and not microtubules is needed for vesicle recycling in the endosomal space. Materials and methods Plant material and gene constructs Tobacco BY-2 cell line (N. tabacum L. cv. Bright Yellow 2; Nagata et al. 1992) and BY- 2 cells transformed with Arabidopsis thaliana PIN1::PIN1:GFP (Benková et al., 2003; Petrášek and Zaţímalová, 2006) were cultured in darkness at 27 C on an orbital incubator (IKA KS501, IKA Labortechnik, at 120 rpm (orbital diameter 30 mm) in liquid medium (3% sucrose, 4.3 g l -1 Murashige and Skoog salts, 100 mg l -1 inositol, 1 mg l -1 thiamin, 0.2 mg l -1 2,4-dichlorophenoxyacetic acid and 200 mg l -1 KH 2 PO 4, ph 5.8) supplemented for transformed cells with 100 mg l -1 kanamycin and 100 mg l -1 cefotaxim, and sub-cultured weekly. Chemicals Styryl dye FM 4-64 (Molecular Probes, catalogue number T13320), BFA, tyrphostin A23, Tyrphostin A51, wortmannin, filipin, dynasore, cytochalasin D, oryzalin and auxins NAA, IAA, 2,4-D (all Sigma-Aldrich) were kept as 20, 50, 50, 50, 10, 20, 80, 10, 57 mm and 10, 20, 10 mm stock solutions, respectively, in DMSO, stored in -20ºC. 34
55 Application of FM dyes, inhibitors and auxins Unless otherwise indicated, the following conditions were used. FM 4-64 (final concentration 2 μm) was added to 1 ml of 2 3-day-old BY-2 cell suspension under continuous shaking in multi-well plates, and samples were observed by confocal microscopy at the times indicated. The inhibitors (tyrphostin A23, Tyrphostin A51, wortmannin, filipin, dynasore, cytochalasin D, oryzalin) and auxins (1-NAA, IAA, 2,4-D) were added directly to the cultivation medium to final concentrations of 50, 50, 33, 15, 80, 20, 15 μm and 10, 5, 5 μm respectively for 30 min pretreatment. For the PIN1 localization experiments transgenic BY-2 cell line expressing under native promoter intragenic translational fusion of PIN1 from Arabidopsis thaliana and GFP was used (Petrášek and Zaţímalová, 2006). 30 min pretreatment with above-mentioned inhibitors was followed by 30 min concomitant treatment with with BFA (50 μm) and analyzed by confocal microscopy. The appropriate amount of the solvent (DMSO) was added to controls. Microscopy For the in vivo microscopy, Zeiss LSM 5 DUO confocal microscope with a 40x C- Apochromat water immersion objective (NA = 1.2) was used. Fluorescence signals for GFP (excitation 488 nm, emission nm) and FM 4-64 (excitation 561 nm, emission >575 nm) were detected. All treatments were performed at least triplicate with the same results, with a minimum of 50 cells evaluated in total each treatment and representative images are shown. 35
56 Results The characterization of the effect of vesicle trafficking inhibitors on the FM 4-64 uptake in BY-2 cells Tyrphostin A23 caused delay in the internalization of the FM 4-64 in comparison with control cells (Figure 1A), endocytic vesicles in the cytoplasm appeared after 20 minutes of the FM 4-64 treatment (Figure 1B). In contrast, tyrphostin A51, previously reported as a control tyrphostin in the respect of blocking endocytosis, accelerated the FM 4-64 uptake, as soon as after few minutes there were observed vesicles containing the dye. Moreover, the PM membrane was poorly stained suggesting quite rapid FM-64 uptake (Figure 1C). Wortmannin blocked the FM 4-64 uptake at the PM, there were no endocytic vesicles observed (Figure 1D), even after 1 h after the addition of the dye into the media (data not shown). Filipin decreased the amount of the FM 4-64 vesicles cells compared with the control cells (compare Figure 1A and 1E), however it did not block the endocytosis completely. Dynasore delayed the uptake of FM 4-64 and the dye could not be internalized into BY-2 cells staining just the PM and adjacent cortical cytoplasm (Figure 1F). Alltogether, effects of used inhibitors on the endocytosis reflect their target and mode of action within the cells. However, the clear difference between tyrphostins A23 and A51 observed here was unexpected and possibly reflects the fact that endocytic machinery of BY-2 has numerous targets for these tyrosine kinase inhibitors and that this inhibition of endocytosis is connected with this mode of action and not with the specific block of tyrosinebased internalization motif. The characterization of the effect of auxins on the FM 4-64 uptake in BY-2 cells Synthetic auxins 1-NAA (Figure 2B), 2,4-D (Figure 2D) as well as endogenous auxin IAA (Figure 2C) slowed down the uptake of FM 4-64 into BY-2 cells in comparison with control (Figure 2A). For all auxins, 5 µm concentrations were tested and no remarkable differences observed.. These results indicate that the sensitivity FM 4-64 uptake in BY-2 cultured cells seems to be comparable for IAA and synthetic auxins 1-NAA and 2,4-D. Cytoskeleton is involved in the FM 4-64 uptake from the plasma membrane in BY-2 cells 36
57 To uncover possible role of cytoskeleton in the processes of endocytosis cytoskeletal drugs were used and followed their effect on the uptake of FM 4-64 dye in vivo. The inhibitor of actin polymerization Cyt D inhibited the endocytosis of FM 4-64, there were no FM containing vesicles observed after 20 min treatment (Figure 3B). The inhibitor of tubulin polymerization oryzalin markedly slowed down the FM 4-64 uptake (Figure 3C) when compared with control cells (Figure 3A), but its effect was not so dramatic as in case of Cyt D. These data indicate the need of both actin and microtubular cytoskeleton during the endocytosis of FM 4-64 dye. They also indirectly suggest that actin filaments are slightly more important for the endocytosis than microtubules. BFA-induced intracellular accumulation of PIN1 protein is differentially triggered by vesicle trafficking inhibitors, auxins and cytoskeletal drugs To follow the requirement of endocytosis and cytoskeleton for the internalization step of the constitutive cycling of PIN1 protein BY-2 cells were incubated 30 min in the medium containing the inhibitor and then BFA was added for other 30 min. This approach was chosen, because in BY-2 cells, in contrast to root tip cells of Arabidopsis (Steinmann et al. 1999; Geldner et al. 2001) the application of inhibitors of endocytosis, auxins or cytoskeletal drugs alone did not induce observable relocalizations of PIN1-GFP that is always at the plasma membrane (data not shown). Thus, blocking anterograde trafficking with BFA helped to uncover retrograde pathways for direct observations in vivo. Tyrphostin A23 prevented the formation of BFA-induced PIN1 aggregates (Figure 4C), while Tyrphostin A51 did not have such an effect (Figure 4D). After the wortmannin treatment accumulations of the PIN1-GFP were observed (Figure 4E), but they were morphologically distinct from aggregations induced by BFA alone (Figure 4A). Wortmannin caused more massive accumulations of PIN1-GFP in the perinuclear region as well as in the cortical cytoplasm (Figure 4E). Filipin did not prevent the formation of BFA-induced PIN1- GFP aggregates (Figure 4F). Both 1-NAA and IAA did not prevent BFA-induced PIN1-GFP aggregations (Figure 4H and 4I). Interestingly 2,4-D, which is not good substrate for auxin efflux carrier in contrast to IAA and 1-NAA, prevented BFA-induced internalization of PIN1-GFP. These data suggest that the inhibitory action of particular auxin on the endocytosis (Paciorek et al. 2005) is seemingly connected with its ability to be transported out of the cells. 37
58 20 min 15 min 10 min 5 min Cyt D enhanced the formation of BFA-induced PIN1-GFP aggregations, which were regularly forming one big aggregation of the PIN1-GFP in the cytoplasm (Figure 4K). In contrast, oryzalin did not prevent the formation of BFA-induced PIN1-GFP aggregations and had no additional visible effect (Figure 4L). These data suggest the actin filaments involvement in the trafficking of PIN1-containing endosomes and that these vesicle trafficking events are not dependent on microtubules. Untreated Tyrphostin A23 Tyrphostin A51 Wortmannin Filipin A B C D E F Dynasore Figure 1: Differential effects of tyrphostin A23, tyrphostin A51, wortmannin, filipin and dynasore on the uptake of FM 4-64 dye. In vivo confocal microscopy of FM 4-64 (2 μm), the uptake of the dye by 3-day-old tobacco BY-2 cells after 30 min pre-treatment with inhibitors. (A) Control cells with equal amount of solvent. (B) Tyrphostin A23 (50 μm) inhibition of the endocytosis of FM (C) Tyrphostin A51 (50 μm) acceleration of the endocytosis (note the absence of the PM staining). (D) Wortmannin (33 μm) inhibition of endocytosis of the dye at the PM. (E) Filipin (15 μm) reduction the amount of FM-labelled endosomes. (F) Dynasore (80 μm) delay of the uptake of FM 4-64 and the accumulation of the vesicles at the cortical cytoplasm after 20 min incubation in FM Scale bars 10 μm. 38




, the")




 1- NAA (5 μm).")
 2,4-D (5 μm).")


59 20 min 15 min 10 min 5 min Untreated NAA IAA 2,4-D A B C D Figure 2: The effect of auxins on the internalization of the FM 4-64 dye. In vivo confocal microscopy of FM 4-64 (2 μm), the uptake of the dye by 3-day-old tobacco BY-2 cells after 30 min pre-treatment with the auxin. (A) Control cells with equal amount of solvent. (B) 1- NAA (5 μm). (C) IAA (5 μm). (D) 2,4-D (5 μm). For all auxins tested significant delay in the process of FM 4-64 uptake in contrast to control cells is observed. Scale bars 10 μm. 39







60 20 min 15 min 10 min 5 min Untreated Cytochalasin D Oryzalin A B C Figure 3: Inhibitors of cytoskeletal polymerization inhibited internalization of FM In vivo confocal microscopy of FM 4-64 (2 μm), the uptake of the dye by 3-day-old tobacco BY-2 cells after 30 min pre-treatment with cytoskeletal drugs. (A) Control cells with equal amount of solvent. (B) Cytochalasin D (40 μm) inhibition of the endocytosis of FM 4-64, there were no vesicles present even after 20 min. (C) Oryzalin (15 μm) delay of the FM 4-64 uptake. Scale bars 10 μm. 40



.")
 Intracellular aggregation of PIN1-GFP after 30 min of BFA (50")
 pre-treatment prevented the BFA-induced")
 Tyrphostin A51 (50 μm) pre-treatment did not prevent the")

 Filipin (15 μm) did not prevent the BFA-induced (G) Dynasore")
 prevented BFA-induced internalization of PIN1-GFP (J),")
 Cytochalasin D (40 μm) pre-treatment enhanced the BFA-induced")

61 Untreated Wm + BFA IAA + BFA A E I BFA Fil + BFA 2,4-D + BFA B F J Tyr A23 + BFA Dns + BFA CytD + BFA C G K Tyr A51 + BFA NAA + BFA Ory + BFA D H L Figure 4: The interference of trafficking inhibitors, auxins and cytoskeletal drugs after BFA-induced inhibition of anterograde trafficking of PIN1-GFP.. In vivo confocal microscopy of 3-day-old tobacco BY-2 cells transformed with PIN1:: PIN1:GFP after 30 min pre-treatment with inhibitors or auxins followed by 30 minutes BFA treatment (50 μm). (A) Control cells with equal amount of solvent. (B) Intracellular aggregation of PIN1-GFP after 30 min of BFA (50 μm) alone, without pre-treatment with the inhibitor (C) Tyrphostin A23 (50 μm) pre-treatment prevented the BFA-induced PIN1-GFP intracellular aggregation. (D) Tyrphostin A51 (50 μm) pre-treatment did not prevent the BFA-induced PIN1-GFP intracellular aggregation. (E) Wortmannin (33 μm) pre-treatment did not prevent the BFAinduced PIN1-GFP intracellular aggregation, but aggregates differ from the BFA effect alone (B). (F) Filipin (15 μm) did not prevent the BFA-induced PIN1-GFP intracellular aggregation. (G) Dynasore (80 μm) pre-treatment prevented the BFA-induced PIN1-GFP intracellular aggregation. (H, I, J) 2,4-D (5 μm) prevented BFA-induced internalization of PIN1-GFP (J), while IAA (5 μm) (I) and NAA (5 μm) (H) did not have this effect. (K) Cytochalasin D (40 μm) pre-treatment enhanced the BFA-induced PIN1-GFP intracellular aggregation. (L) Oryzalin (15 μm) pre-treatment did not prevent the BFA-induced PIN1-GFP intracellular aggregation. Scale bars 20 μm. 41
62 Discussion The use of endocytic inhibitors is a powerful tool for identification of plant endocytic and secretory pathways. The effect of the most inhibitors and substances applied in the present study has been previously described, however on distinct plant materials and sometimes with different results. In this study, we used BY-2 cells to compare their action in one system to obtain the information that would help us to describe the endocytic pathways within these cells. FM 4-64 was used as an endocytic marker of fluid phase and clathrin-mediated endocytosis. The action of both used tyrphostins A23 as well as A51 on FM 4-64 endocytosis was quite surprising. There was previously published the specific inhibitory effect of Tyr A23 on the protein recruitment without affecting the FM 4-64 internalization in Arabidopsis roots (Dhonukshe et al. 2007), as well as in Arabidopsis protoplasts (Ortiz-Zapater et al. 2006). However, in our experiments with BY-2 suspension cells, tyrphostin A23 strongly inhibited FM 4-64 uptake. These data are in agreement with recently published paper by Lam et al. 2009, who showed that the effect of tyrphostine A23 is dose-dependent. Tyrphostin A51 is widely used as inactive, control tyrphostin for the endocytosis studies (Banbury et al. 2003). In our experiments tyrphostine A51 did not inhibit the endocytosis, but its effect was oposite, such an effect of tyrphostine A51 was not observed so far and therefore care must be taken in future studies using this tyrphostin. The clear difference between tyrphostins A23 and A51 observed in our work was unexpected and possibly reflects the fact that endocytic machinery of BY-2 has numerous targets for these tyrosine kinase inhibitors and that this inhibition of endocytosis is connected with this mode of action and not with the specific block of tyrosinebased internalization motif. The reaction of cells to wortmannin treatment is in agreement with already published results (Emans et al., 2002, Wang et al. 2009). Wortmannin can be used as a potent inhibitor of endocytosis in BY-2 suspension cell lines. Filipin did not have as dramatic inhibitory effect on FM 4-64 uptake as in Arabidopsis root cells (Kleine-Vehn et al. 2006), however there was observed delay in the dye uptake, caused probably by the PM sterol binding. The inhibition of endocytosis with dynasore, reported here for the first time in plants, confirmed the clathrinmediated endocytosis of FM 4-64 in plants. Auxins were used in our work as previously described inhibitors of endocytosis (Paciorek et al. 2005), though the mechanism of this inhibition is unclear. It was shown that FM 4-64 endocytosis was inhibited by all auxins tested IAA, 1-NAA and 2,4-D. Therefore, although these three auxin use to various extent both passive (slower) and active (faster) transport 42
63 inside tobacco cells (Delbarre et al. 1996) this is not reflected on their ability to generally block internalization of FM dye, when they are used in the same concentrations. Our experiments with inhibitors of actin and tubulin polymerization indicated the need of both actin and microtubular cytoskeleton for the FM 4-64 endocytosis and so it could be suggested that the fluid phase and clathrin-mediated endocytosis of FM 4-64 is dependent on both actin filaments and microtubules. BFA as an inhibitor of anterograde vesicle transport was used to selectively visualize trafficking pathway running in the retrograde direction. PIN1-GFP protein as the marker of clathrin-dependent trafficking (recycling) pathways was used. The regulation of the amount and positioning of PIN1 protein within the PM is achieved by BFA-sensitive constitutive cycling between PM a endosomal compartment (Geldner et al. 2001). After incubating the cells in BFA-enriched media, agreggates of PIN1-GFP were observed within the cytoplasm of BY-2 cells. As has been recently reported, BFA induces TGN-derived as well as Golgiderived aggregates in BY-2 cells (Lam et al. 2009). Indeed, the presence of cell wall polysaccharides in the BFA compartment (Satiat-Jeunemaitre and Hawes, 1992) may reflect the accumulation of secretory vesicles, rather than being interpreted as endocytic in origin (Baluška et al. 2002). Moreover, during the BFA inhibition, it was found that endocytosis may proceed (Grebe et al. 2003). The observation of BFA-induced PIN1-GFP aggregations after tyrphostin A23 suggested that besides blocking endocytosis from the PM (as shown by FM 4-64) it specifically block formation of BFA-induced PIN1-GFP aggregations. In this respect, the action of tyrphostin 23 is different from tyrphostin 51, which is inactive in the agreement with results of Dhonukshe et al. (2007) and Ortiz-Zapater et al. (2006). Also dynasore blocked the formation of PIN1-GFP-containing BFA-induced vesicles giving the community another tool for addresing the recycling of PIN proteins, specifically with respect to the role of plasma membrane dynamins. Wortmanin probably does not interfere with the vesicle budding and transport in the endosomal space as there are formed BFA-induced PIN1-containing aggregations. However, they seem to be slightly distinct than without wortmannin pretreatment. All of the used auxins inhibited endocytosis but only 2,4-D, which not the substrate for auxin efflux prevented the formation of BFA-induced aggregations of PIN1-GFP.. These data suggest that the inhibitory action of particular auxin on the endocytosis (Paciorek et al. 2005) is seemingly connected with its ability to be transported out of the cells. Those auxins, which are more accumulated inside cells inhibit more intensively the endocytosis, including the endocytosis of their efflux carriers, to enhance their own efflux. 43
64 Disruption of both actin filaments or microtubules inhibited endocytosis of FM-64. However, the formation of BFA-induced aggregations of PIN1-GFP was not prevented by both actin and tubulin drugs. But only in case of depolymerization of actin cytoskeleton, the formation of much bigger aggreagates of PIN1 was observed. These data suggest the actin filaments involvement in the trafficking of PIN1-containing endosomes and that these vesicle trafficking events are not dependent on microtubules. Previously reported remodelling of actin filaments in the perinuclear region of tobacco cells after BFA treatment (Petrášek et al. 2003) could include active co-operation between actin cytoskeleton and membrane vesicles, suggesting the role of actin filaments in the endocytosis. Moreover, from the above in vivo observed data it could be speculated that in BY-2 cells PIN1-containing BFA-induced aggregations might be formed from the actively cycling PIN1 vesicles in the endosomal space (between PM an GA) rather than from fusion of the endosomes or de novo endocytic vesicles originating either at the PM or at the membranes of RE/EE/TGN. This active BFA trapping of recycling PIN1 vesicles supports the dependency of this trafficking on intact actin. Mictrotubules seem to be important for the endocytotic proceses running from the PM, as suggested by our FM 4-64 experiments, rather than in vesicle endosomal trafficking. Obtained results together with the information on the particular inhibitor are summarized in Table 1. 44











65 FM 4-64 endocytosis FM 4-64 endocytosis Remarks PIN1 localization after BFA Remarks on the mechanism of action References Control Tyrphostin A Complete inhibition Analogue of tyrosine, blocking tyrosine kinases, binding in Tyr pocket of proteins disable the AP-2 protein binding and to internalization of the protein by CCV Aniento and Robinson 2005, Robinson et al Tyrphostin A Weak PM signal, enhanced endocytosis Analogue of tyrosine, blocking tyrosine kinases, not reported as a inhibitor of endocytosis, potent inhibitor of EGFR (epidermal growth factor receptor) in animal cells Aniento and Robinson 2005, Robinson et al Levitzki 1990 Wortmannin Only PM signal Inhibitor of phosphoinositol-3 kinase activity, it blocks plant endocytosis downstream of the internalization event at the PM, protein sorting to the vacuole, and FM 1-43 uptake in BY-2 cells Aniento and Robinson 2005, Robinson et al Matsuoka et al Emans et al.,2002 Filipin Less endocytic vesicles Labels plant 3-β-hydroxysterols, Inhibits FM4-64 uptake, Inhibitor of endocytosis of PM resident proteins Grebe et al Kleine-Vehn et al., 2006 Dynasore Endocytosis slowed down Permeable inhibitor of dynamin GTPase function, that blocks completely compensatory synaptic vesicle (clathrinmediated) endocytosis,not used in plants so far Newton et al NAA Endocytosis slowed down Reported as a inhibitor of endocytosis and protein trafficking in plants Paciorek et al IAA Endocytosis slowed down Reported as a inhibitor of endocytosis and protein trafficking in plants Paciorek et al ,4-D Endocytosis slowed down s Reported as a inhibitor of endocytosis and protein trafficking in plants Paciorek et al Cytochalasin D Endocytosis slowed down Inhibitor of actin polymerization, disrupts AFs Oryzalin Endocytosis slowed down s MTs targeting, Inhibitor of MTs polymerization Table 1: The effect of inhibitors of vesicle trafficking, auxins and cytoskeletal drugs on the processes of endocytosis and auxin efflux carrier PIN1 internalization after BFA in BY-2 cells. + running endocytosis, - inhibited endocytosis 45
66 References Aniento F, Robinson D (2005) Testing for endocytosis in plants. Protoplasma 226: 3-11 Baluška F, Hlavačka A, Šamaj J, Palme K, Robinson D, Matoh T, McCurdy D, Menzel D, Volkmann D (2002) F- actin-dependent endocytosis of cell wall pectins in meristematic root cells. Insights from brefeldin A- induced compartments. Plant Physiol 130: Banbury D, Oakley J, Sessions R, Banting G (2003) Tyrphostin A23 inhibits internalization of the transferrin receptor by perturbing the interaction between tyrosine motifs and the medium chain subunit of the AP-2 adaptor complex. J Biol Chem 278: Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J (2003) Local, effluxdependent auxin gradients as a common module for plant organ formation. Cell 115: Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J (2003) Local, effluxdependent auxin gradients as a common module for plant organ formation. Cell 115: Boutté Y, Crosnier M, Carraro N, Traas J, Satiat-Jeunemaitre B (2006) The plasma membrane recycling pathway and cell polarity in plants: studies on PIN proteins. J Cell Sci 119: Bright N, Lindsay M, Stewart A, Luzio J (2001) The relationship between lumenal and limiting membranes in swollen late endocytic compartments formed after wortmannin treatment or sucrose accumulation. Traffic 2: Butler J, Hu S, Brady S, Dixon M, Muday G (1998) In vitro and in vivo evidence for actin association of the naphthylphthalamic acid-binding protein from zucchini hypocotyls. Plant J 13: Dhonukshe P, Aniento F, Hwang I, Robinson D, Mravec J, Stierhof Y, Friml J (2007) Clathrin-mediated constitutive endocytosis of PIN auxin efflux carriers in Arabidopsis. Curr Biol 17: Dhonukshe P, Grigoriev I, Fischer R, Tominaga M, Robinson D, Hašek J, Paciorek T, Petrášek J, Seifertová D, Tejos R, Meisel L, Zazímalová E, Gadella TJ, Stierhof Y, Ueda T, Oiwa K, Akhmanova A, Brock R, Spang A, Friml J (2008) Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc Natl Acad Sci U S A 105: Dhonukshe P, Tanaka H, Goh T, Ebine K, Mähönen A, Prasad K, Blilou I, Geldner N, Xu J, Uemura T, Chory J, Ueda T, Nakano A, Scheres B, Friml J (2008) Generation of cell polarity in plants links endocytosis, auxin distribution and cell fate decisions. Nature 456: Emans N, Zimmermann S, Fischer R (2002) Uptake of a fluorescent marker in plant cells is sensitive to brefeldin A and wortmannin. Plant Cell 14: Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P, Delbarre A, Ueda T, Nakano A, Jürgens G (2003) The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxindependent plant growth. Cell 112: Geldner N, Friml J, Stierhof Y, Jürgens G, Palme K (2001) Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413: Geldner N, Hyman D, Wang X, Schumacher K, Chory J (2007) Endosomal signaling of plant steroid receptor kinase BRI1. Genes Dev 21: Geldner N, Jürgens G (2006) Endocytosis in signalling and development. Curr Opin Plant Biol 9:
67 Geldner N, Richter S, Vieten A, Marquardt S, Torres-Ruiz R, Mayer U, Jürgens G (2004) Partial loss-of-function alleles reveal a role for GNOM in auxin transport-related, postembryonic development of Arabidopsis. Development 131: Geldner N, Robatzek S (2008) Plant receptors go endosomal: a moving view on signal transduction. Plant Physiol 147: Grebe M, Xu J, Möbius W, Ueda T, Nakano A, Geuze H, Rook M, Scheres B (2003) Arabidopsis sterol endocytosis involves actin-mediated trafficking via ARA6-positive early endosomes. Curr Biol 13: Hess M, Müller M, Debbage P, Vetterlein M, Pavelka M (2000) Cryopreparation provides new insight into the effects of brefeldin A on the structure of the HepG2 Golgi apparatus. J Struct Biol 130: Holstein S (2002) Clathrin and plant endocytosis. Traffic 3: Holweg C (2007) Acto-Myosin motorises the flow of auxin. Plant Signal Behav 2: Holweg C, Nick P (2004) Arabidopsis myosin XI mutant is defective in organelle movement and polar auxin transport. Proc Natl Acad Sci U S A 101: Jackson C, Casanova J (2000) Turning on ARF: the Sec7 family of guaninenucleotide-exchange factors. Trends Cell Biol 10: Jaillais Y, Fobis-Loisy I, Miège C, Rollin C, Gaude T (2006) AtSNX1 defines an endosome for auxin-carrier trafficking in Arabidopsis. Nature 443: Kleine-Vehn J, Dhonukshe P, Sauer M, Brewer P, Wiśniewska J, Paciorek T, Benková E, Friml J (2008b) ARF GEF-dependent transcytosis and polar delivery of PIN auxin carriers in Arabidopsis. Curr Biol 18: Kleine-Vehn J, Dhonukshe P, Swarup R, Bennett M, Friml J (2006) Subcellular trafficking of the Arabidopsis auxin influx carrier AUX1 uses a novel pathway distinct from PIN1. Plant Cell 18: Kleine-Vehn J, Langowski L, Wisniewska J, Dhonukshe P, Brewer P, Friml J (2008) Cellular and molecular requirements for polar PIN targeting and transcytosis in plants. Mol Plant 1: Lam S, Cai Y, Tse Y, Wang J, Law A, Pimpl P, Chan H, Xia J, Jiang L (2009) BFA-induced compartments from the Golgi apparatus and trans- Golgi network/early endosome are distinct in plant cells. Plant J 60: Levitzki A, Gilon C (1991) Tyrphostins as molecular tools and potential antiproliferative drugs. Trends Pharmacol Sci 12: Lippincott-Schwartz J, Yuan L, Tipper C, Amherdt M, Orci L, Klausner R (1991) Brefeldin A's effects on endosomes, lysosomes, and the TGN suggest a general mechanism for regulating organelle structure and membrane traffic. Cell 67: Lisboa S, Scherer G, Quader H (2008) Localized endocytosis in tobacco pollen tubes: visualisation and dynamics of membrane retrieval by a fluorescent phospholipid. Plant Cell Rep 27: Marella M, Lehmann S, Grassi J, Chabry J (2002) Filipin prevents pathological prion protein accumulation by reducing endocytosis and inducing cellular PrP release. J Biol Chem 277: Matsuoka K, Bassham D, Raikhel N, Nakamura K (1995) Different sensitivity to wortmannin of two vacuolar sorting signals indicates the presence of distinct sorting machineries in tobacco cells. J Cell Biol 130: Nagata T, Nemoto Y, Hasezawa S (1992) Tobacco BY-2 cell line as the "HeLa" cell in the cell biology of higher plants. International Review of Cytology 130:
68 Newton A, Kirchhausen T, Murthy V (2006) Inhibition of dynamin completely blocks compensatory synaptic vesicle endocytosis. Proc Natl Acad Sci U S A 103: Ortiz-Zapater E, Soriano-Ortega E, Marcote M, Ortiz-Masiá D, Aniento F (2006) Trafficking of the human transferrin receptor in plant cells: effects of tyrphostin A23 and brefeldin A. Plant J 48: Paciorek T, Zazímalová E, Ruthardt N, Petrášek J, Stierhof Y, Kleine-Vehn J, Morris D, Emans N, Jürgens G, Geldner N, Friml J (2005) Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 435: Petrášek J, Černá A, Schwartzerová K, Elčkner M, Morris D, Zazímalová E (2003) Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiol 131: Petrášek J, Mravec J, Bouchard R, Blakeslee J, Abas M, Seifertová D, Wisniewska J, Tadele Z, Kubeš M, Čovanová M, Dhonukshe P, Skůpa P, Benková E, Perry L, Kreček P, Lee O, Fink G, Geisler M, Murphy A, Luschnig C, Zaţímalová E, Friml J (2006) PIN proteins perform a ratelimiting function in cellular auxin efflux. Science 312: Poupon V, Stewart A, Gray S, Piper R, Luzio J (2003) The role of mvps18p in clustering, fusion, and intracellular localization of late endocytic organelles. Mol Biol Cell 14: Pérez-Gómez J, Moore I (2007) Plant endocytosis: it is clathrin after all. Curr Biol 17: R Richter S, Geldner N, Schrader J, Wolters H, Stierhof Y, Rios G, Koncz C, Robinson D, Jürgens G (2007) Functional diversification of closely related ARF-GEFs in protein secretion and recycling. Nature 448: Ritzenthaler C, Nebenführ A, Movafeghi A, Stussi-Garaud C, Behnia L, Pimpl P, Staehelin L, Robinson D (2002) Reevaluation of the effects of brefeldin A on plant cells using tobacco Bright Yellow 2 cells expressing Golgi-targeted green fluorescent protein and COPI antisera. Plant Cell 14: Robinson D, Jiang L, Schumacher K (2008) The endosomal system of plants: charting new and familiar territories. Plant Physiol 147: Satiat-Jeunemaitre B (1992) Spatial and temporal regulations in helicoidal extracellular matrices: comparison between plant and animal systems. Tissue Cell 24: Satiat-Jeunemaitre B, Cole L, Bourett T, Howard R, Hawes C (1996) Brefeldin A effects in plant and fungal cells: something new about vesicle trafficking? J Microsc 181: Shpetner H, Joly M, Hartley D, Corvera S (1996) Potential sites of PI-3 kinase function in the endocytic pathway revealed by the PI-3 kinase inhibitor, wortmannin. J Cell Biol 132: Steinmann T, Geldner N, Grebe M, Mangold S, Jackson C, Paris S, Gälweiler L, Palme K, Jürgens G (1999) Coordinated polar localization of auxin efflux carrier PIN1 by GNOM ARF GEF. Science 286: Tse Y, Mo B, Hillmer S, Zhao M, Lo S, Robinson D, Jiang L (2004) Identification of multivesicular bodies as prevacuolar compartments in Nicotiana tabacum BY-2 cells. Plant Cell 16: von Zastrow M, Sorkin A (2007) Signaling on the endocytic pathway. Curr Opin Cell Biol 19: Wang H, Wang G, Yu Y, Wang Y (2009) The role of phosphoinositide-3- kinase/akt pathway in propofolinduced postconditioning against focal cerebral ischemia-reperfusion injury in rats. Brain Res 1297:
 Probing plant membranes with FM dyes: tracking, dragging or blocking? Plant J, 61, 883-892.")
69 3.3 Probing plant membranes with FM dyes: tracking, dragging or blocking? Jelínková, A., Malínská, K., Simon, S., Kleine-Vehn, J., Pařezová, M., Pejchar, P., Kubeš, M., Martinec, J., Friml, J., Zaţímalová, E. and Petrášek, J. (2010) Probing plant membranes with FM dyes: tracking, dragging or blocking? Plant J, 61, This chapter has been published in the form of technical advance and points out the necessity of critical use of some tools used in biologyand vice versa the possibility of use their side effect in further research. During the experiments that used endocytosis FM styryl markers I have noticed unexpected effect of these markers on the localization of PIN1 plasma membrane localized auxin efflux carrier. This paper characterizes this effect in detail and emphasize the need for circumspection during in vivo studies of membrane proteins performed using simultaneous labelling with FM dyes. My contribution to this paper was in the original observation of the effect of FM-dyes on plasma membrane protein localization in suspension cells, analyses of the data, participation on designing and co-ordination of the experiments and finalizing the manuscript. Transient relocalization of PIN1-GFP after FM 4-64 treatment 49
70 50


71 The Plant Journal (2010) doi: /j X x TECHNICAL ADVANCE Probing plant membranes with FM dyes: tracking, dragging or blocking? Adriana Jelínková 1,2,, Kateřina Malínská 1,,*, Sibu Simon 1, Jürgen Kleine-Vehn 3, Markéta Pařezová 1, Přemysl Pejchar 1, Martin Kubeš 1, Jan Martinec 1, Jiří Friml 3, Eva Zažímalová 1 and Jan Petrášek 1,2 1 Institute of Experimental Botany, Academy of Sciences of the Czech Republic, Rozvojová 263, Prague 6, Czech Republic, 2 Department of Plant Physiology, Faculty of Science, Charles University, Viničná 5, Prague 2, Czech Republic, and 3 Department of Plant Systems Biology, Flanders Institute for Biotechnology, and Department of Plant Biotechnology and Genetics, Ghent University, 9052 Ghent, Belgium Received 12 August 2009; revised 23 November 2009; accepted 2 December * For correspondence (fax ; malinska@ueb.cas.cz). These authors contributed to this paper equally. SUMMARY Remarkable progress in various techniques of in vivo fluorescence microscopy has brought an urgent need for reliable markers for tracking cellular structures and processes. The goal of this manuscript is to describe unexplored effects of the FM (Fei Mao) styryl dyes, which are widely used probes that label processes of endocytosis and vesicle trafficking in eukaryotic cells. Although there are few reports on the effect of styryl dyes on membrane fluidity and the activity of mammalian receptors, FM dyes have been considered as reliable tools for tracking of plant endocytosis. Using plasma membrane-localized transporters for the plant hormone auxin in tobacco BY-2 and Arabidopsis thaliana cell suspensions, we show that routinely used concentrations of FM 4-64 and FM 5-95 trigger transient re-localization of these proteins, and FM 1-43 affects their activity. The active process of re-localization is blocked neither by inhibitors of endocytosis nor by cytoskeletal drugs. It does not occur in A. thaliana roots and depends on the degree of hydrophobicity (lipophilicity) of a particular FM dye. Our results emphasize the need for circumspection during in vivo studies of membrane proteins performed using simultaneous labelling with FM dyes. Keywords: FM 4-64, FM 1-43, plasma membrane, tobacco BY-2 cells, endocytosis. INTRODUCTION FM dyes are fluorescent markers that were originally developed as membrane potential sensors (Grinvald et al., 1988) and later optimized to stain synaptic vesicles in vivo (Betz and Bewick, 1992; Betz et al., 1992, 1996; Gaffield and Betz, 2006). They are widely used in tracking processes of membrane trafficking in eukaryotic organisms (Gaffield and Betz, 2006), including plants (Bolte et al., 2004; Aniento and Robinson, 2005). The amphiphilic character of FM molecules (Figure 1) allows their penetration through the cell wall followed by insertion into the plasma membrane (PM) and subsequent tracking of intracellular membrane dynamics. The tail region of the FM molecule is responsible for its lipophilicity. A higher number of carbons in the tail increases the lipophilicity of the molecule, but PM staining becomes irreversible (Betz et al., 1996). The composition of the bridge region determines the fluorescence wavelength of the molecule. More double bonds in this region shift emission to the red part of the spectrum. In contrast to the hydrophobic environments of the PM, fluorescence of FM dyes is strongly reduced in a polar solvent (water). This makes FM dyes optimal for selective tracking of membrane dynamics. Moreover, positively charged head regions prevent the movement of the dye across the membrane via phospholipid flip-flopping. These chemical properties make FM dyes useful tools in tracking endocytosis and vesicle trafficking in living cells. The most commonly used FM dyes in plant biology are FM 4-64 (N-(3-triethylammoniumpropyl)-4-(6-(4-(diethylamino)phenyl)hexatrienyl)pyridinium dibromide) and FM 1-43 (N-(3-triethylammoniumpropyl)-4-(4-(dibutylamino)styryl) pyridinium dibromide). In addition to tracking endocytosis (Emans et al., 2002), these dyes are often used in tracking ª 2010 Institute of Experimental Botany, ASCR 1 Journal compilation ª 2010 Blackwell Publishing Ltd
72 2 Adriana Jelínková et al. Figure 1. Structure of the FM dyes (Betz et al., 1996) used in this study. secretory pathways (Dettmer et al., 2006), as after some time they end up in the vacuolar membrane, and thus can also be exploited for studies of vacuolar dynamics (Kutsuna and Hasezawa, 2002). There are also several other organelles stained by FM dyes (for review see Bolte et al., 2004). Because of differences in the speed of incorporation into endomembranes, FM 1-43 and FM 4-64 can be used as selective dyes for exo- and endocytosis in pollen tubes (Zonia and Munnik, 2008). Moreover, a slightly less lipophilic variant of the FM 4-64 molecule, red-fluorescing FM 5-95 (N-(3-trimethylammoniumpropyl)-4-(6-(4-(diethylamino)phenyl)hexatrienyl) pyridinium dibromide), can be used as an alternative. The hydrophobicity of the above mentioned FM dyes gradually decreases from FM 4-64 through FM 5-95 to FM 1-43 (information provided in Molecular Probes The Handbook). Recent progress in plant molecular biology has provided evidence for the existence of plant clathrin-dependent endocytosis of PINFORMED (PIN) auxin efflux carriers (Dhonukshe et al., 2007). It appears that, after fluid phase endocytosis (Oparka, 1990), clathrin-dependent endocytosis is the major endocytic route in plants (Perez-Gomez and Moore, 2007). Thus, FM dyes are now considered as preferential markers for this type of endocytosis, being blocked largely at the PM of the cells where the clathrin molecule is not functional (Dhonukshe et al., 2007). The suitability of FM dyes as markers for endocytic pathways has recently been supported by experiments involving micro-injection of FM 4-64 directly into the cytosol of Nicotiana tabacum BY-2 cells and Tradescantia virginiana stamen hair cells (van Gisbergen et al., 2008). Moreover, a set of Förster Resonance Energy Transfer (FRET) data measured between FM 4-64 and cytoplasmic GFP (Griffing, 2008) also showed that this dye is taken up into cells preferentially by endocytosis. However, even though FM dyes are undoubtedly valuable tools for several applications, few reports on their side effects have been published. Styryl-based dyes have been shown to influence membrane fluidity (Rodes et al., 1995). In addition, they block mechanotransduction channels (Gale et al., 2001) as well as the activity of muscarinic and nicotinic acetylcholine receptors (for review, see Gaffield and Betz, 2006). Therefore, careful investigation of their effects on cellular structures and their activities would help in understanding the mechanism of FM dye action and validate their exploitation as endocytotic markers. Here we show that some concentrations of FM dyes, even much lower than those widely used, have the potential to disturb the localization and activity of integral PM proteins, such as auxin carriers, in cell suspension cultures. To characterize this phenomenon, we used tobacco BY-2 or Arabidopsis cell cultures expressing translational fusions with GFP using either the auxin efflux carrier PIN1 (PIN1:: PIN1:GFP; Benkova et al., 2003; Petrášek et al., 2006) or the multi-drug resistance (MDR) type conditional auxin efflux carrier PGP4/ABCB4 (PGP4::PGP4:GFP; Cho et al., 2007; Terasaka et al., 2005; Yang and Murphy, 2009) from A. thaliana. In both cases, transient internalization of GFP-tagged proteins from PM was observed upon treatment with FM 4-64 and FM 5-95, but not with FM The internalization was not blocked by inhibitors of endocytosis or cytoskeletal drugs, but was sensitive to the metabolic drug sodium azide. Moreover, all three dyes interfered to various degrees with the activity of endogenous tobacco auxin carrier proteins and phospholipid metabolism. RESULTS FM 4-64 induces transient re-localization of plasma membrane proteins in BY-2 cells We used FM styryl dyes for characterization of endosomal trafficking of proteins involved in the auxin efflux machinery in a tobacco BY-2 cell line. It was very easy to reproduce already published data describing the use of FM dyes (Bolte et al., 2004). However, in addition to this, other previously uncharacterized effects of FM 4-64 treatment were repeatedly observed. Direct application of 2 lm FM 4-64 to BY-2 cells stably expressing either PIN1 GFP or PGP4 GFP resulted in transient internalization of these proteins from the PM. Control cells showed localization of PIN1 GFP and PGP4 GFP in the PM and the cortical cytoplasm (Figure 2a,b, 0 min, and Figure S1a). After 2 min of treatment with FM 4-64, a remarkable increase in the internal pool of both fusion proteins was observed as punctate GFP signal (Figure S1). This phenomenon clearly preceded the endocytosis of FM 4-64, because the dye was still localized mainly on the PM 2 min after its application (Figure S1). As FM 4-64 internalization proceeded (Figure 2a,b, 10 min), FM-labelled endosomes did not co-localize with FM-induced PIN1 GFP patches (Figure 2c,d). Similarly, no co-localization between PGP4 GFP patches and FM-decorated endosomes was observed (data not shown). Prolonged incubation with FM 4-64 (up to 30 min) resulted in remarkable endocytosis of FM 4-64, but PIN1 GFP as well as PGP4 GFP vesicles disappeared from the cortical cytoplasm, restoring a ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
 In vivo confocal microscopy of PIN1 GFP (a) and PGP4 GFP (b) in 3-day-old tobacco BY-2 cells; treatment with 2 lm FM 4-64 for the indicated times.")

 Longitudinal sections through the cortical layer of the cytoplasm showing internalized PIN1 GFP and FM 4-64 after 10 min incubation with 2 lm FM 4-64.")
73 Side-effects of FM dyes in plants 3 Figure 2. FM 4-64-stimulated transient internalization of plasma membrane-located auxin carriers. (a, b) In vivo confocal microscopy of PIN1 GFP (a) and PGP4 GFP (b) in 3-day-old tobacco BY-2 cells; treatment with 2 lm FM 4-64 for the indicated times. PIN1 GFP and PGP4 GFP localized at transversal plasma membranes of control cells (upper row) and were internalized after 10 min of incubation with FM 4-64 (middle row). Notice the patchy pattern in the green channel. Restored plasma membrane localization of PIN1 GFP and PGP4 GFP after 30 min of treatment with FM 4-64 was accompanied by endocytosis of FM 4-64 (bottom row). (c) Longitudinal sections through the cortical layer of the cytoplasm showing internalized PIN1 GFP and FM 4-64 after 10 min incubation with 2 lm FM (d) Scattergram created from the oval region indicated in (c), demonstrating no co-localization of PIN1 GFP and FM 4-64 fluorescence. Scale bars = 20 lm. (a) (b) (c) (d) 250 Intensity of FM 4-64 (red channel) Intensity of PIN1::PIN1:GFP (green channel) Absolute frequency control-like situation (i.e. localization on the PM; Figure 2a,b, 30 min). Movie S1 shows PIN1 GFP-containing vesicles 3 min after FM 4-64 application in comparison with the control (Movie S2). Altogether, the results show that application of FM 4-64 dye, at a concentration even lower than that routinely used for tracking of endocytosis, results in transient re-localization of at least two integral PM proteins in exponentially growing BY-2 cells. FM dye-induced transient internalization of plasma membrane proteins is cell suspension culture-specific, and may be tissue- and/or species-specific To address the question of whether the above described effect of FM 4-64 on PM proteins is common in plant cell suspensions, we studied this effect in a cell suspension derived from A. thaliana PIN1::PIN1:GFP plants (Benkova et al., 2003). PIN1 GFP localization in the Arabidopsis PM differs from that in BY-2 cells; the fluorescence intensity is less homogeneous and some intracellular accumulation is seen (Figure 3a, 0 min). Nevertheless, 10 min after addition of FM 4-64, similar transient re-localization of PIN1 GFP to that reported for BY-2 cells was observed (Figure 3a, 10 min). FM-labelled endosomes did not co-localize with FM-induced PIN1 GFP patches. In comparison with BY-2, the recovery of PIN1 GFP PM localization appeared to be slower (Figure 3a, 40 min). The FM 4-64 effect was also examined in A. thaliana seedlings expressing another auxin efflux carrier, PIN2 GFP (Xu and Scheres, 2005) or aquaporin water channel PIP2 GFP (Cutler et al., 2000). PIN2 GFP (Figure 3b) or PIP2 GFP (Figure 3c) seedlings were treated with FM The dye was washed out and images of root epidermis and cortex at various time points up to 1 h after addition of FM dye were collected. Neither of these two proteins showed enhanced FM-induced internalization compared to mock treatment. Although a tissue and even species specificity cannot be excluded, FM dye-induced transient internalization of PM proteins appears to be cell suspension culture-specific. FM dye-induced transient internalization of plasma membrane proteins correlates with dye hydrophobicity To establish whether the above-mentioned effects of FM 4-64 are specific for this particular dye or whether they are ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
 In vivo confocal microscopy of PIN1 GFP in 3-day-old Arabidopsis cell suspension culture; treatment with 2 lm FM 4-64 for the indicated times.")

 was observed within 30 min. Scale bars = 20 lm. more general, we compared them with similar FM dyes: the less hydrophobic FM 5-95 and FM 1-43.")
74 4 Adriana Jelínková et al. (a) 0 min 10 min 40 min (b) 0 min 30 min (c) 0 min 30 min Figure 3. FM dye-induced transient internalization of plasma membrane proteins is cell suspension culture-specific. (a) In vivo confocal microscopy of PIN1 GFP in 3-day-old Arabidopsis cell suspension culture; treatment with 2 lm FM 4-64 for the indicated times. PIN1 GFP localized at plasma membranes of control cells (0 min) and was internalized after 10 min of incubation with FM Notice the patchy pattern in the green channel. Plasma membrane localization of PIN1 GFP was restored after 40 min of treatment with FM (b, c) Five-day-old Arabidopsis seedlings expressing PIN2 GFP (b) or PIP2 GFP (c) were incubated for 30 min in the presence or absence of FM4-64 dye. No apparent PIN2 or PIP2 internalization (in green) was observed within 30 min. Scale bars = 20 lm. more general, we compared them with similar FM dyes: the less hydrophobic FM 5-95 and FM To secure full saturation of the PM with FM dyes (2 lm), providing a comparable situation at the beginning of the experiment, a protocol of FM staining on ice and subsequent observation at room temperature was used (Emans et al., 2002). This approach is often used because, at the beginning of the experiment, when the PM is saturated with FM dye on ice, endocytosis does not occur; later on, it can be easily restored upon transfer of cells to room temperature. For all these experiments, a PIN1 GFP-transformed BY-2 cell line was used. After loading the cells with FM 4-64 on ice and subsequent transfer to room temperature, the PIN1 GFP re-localization resembled that observed using a protocol without preincubation on ice (Figure 4a). It was also transient. Although with slower kinetics, FM 5-95 also induced internalization of PIN1 GFP (Figure 4b) and PGP4 GFP (data not shown). In contrast, the least hydrophobic dye among those tested (FM 1-43) never induced any observable re-localization of PIN1 GFP (Figure 4c) or PGP4 GFP (data not shown), even after prolonged incubation. The effect of all three FM dyes was compared as the relative decrease of the PIN1 GFP fluorescence intensity at the plasma membrane after 15 min incubation (Figure S1e). These results suggest a positive correlation between the FM dye hydrophobicity and the ability to re-localize integral PM proteins. Active metabolism, but neither endocytosis nor cytoskeleton dynamics, are necessary for FM-induced internalization of plasma membrane proteins To assess possible mechanisms triggered by the addition of FM dyes, the metabolic inhibitor sodium azide (NaN 3 ) was used to block ATP-dependent processes in BY-2 cells. Both FM 4-64 endocytosis and PIN1 GFP internalization were totally prevented by 15 mm NaN 3 (Figure 5a). This indicates an active, energy-dependent process involved in PIN1 GFP re-localization. On the other hand, treatments that effectively block FM dye endocytosis, including prolonged incubation on ice (Figure 5b) as well as application of the phosphatidylinositol 3-kinase inhibitor wortmannin (Figure 5c), were not effective in blocking the transient internalization of PIN1 GFP caused by FM Interestingly, FM dye-induced formation of PIN1 GFP patches was even more pronounced on ice, was not transient (over short time periods) and persisted for at least 1 h. Figure S2 shows that de-polymerization of neither actin filaments (treated with 500 nm latrunculin B) nor microtubules (treated with 20 lm oryzalin) is able to abolish the FM dye effect on PIN1 GFP localization completely. Interestingly, de-polymerization of actin filaments (in contrast to the de-polymerization of microtubules) slightly decreased the amount of re-localized PIN1 GFP, suggesting partial dependence of this process on the actin cytoskeleton. ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
, FM 5-95 (b) or FM 1-43 (c) at 26 C.")
75 Side-effects of FM dyes in plants 5 Figure 4. FM-induced transient internalization of plasma membrane proteins correlates with the dye hydrophobicity. In vivo confocal microscopy of PIN1 GFP in 3-day-old tobacco BY-2 cells after 15 min preincubation on ice followed by 15 min treatment with FM 4-64 (a), FM 5-95 (b) or FM 1-43 (c) at 26 C. Notice no internalization of PIN1 GFP after FM 1-43 treatment, in contrast to FM 4-64 and FM Scale bars = 20 lm. (a) (b) (c) To address the question of whether FM-induced PIN1 GFP patches are formed from the PM or from the endosomal pool of PIN1 GFP, brefeldin A (BFA), an inhibitor of anterograde vesicle trafficking in plant cells, was used. BFA compartments were formed close to the nucleus after 1 h incubation in 20 lm BFA for FM-treated as well as control cells (data not shown). Treatment with BFA did not prevent the extensive formation of PIN1 GFP-containing patches after FM 4-64 application (Figure 5d). In cells treated with BFA but without FM 4-64, no similar PIN1 GFP patches close to the PM were observed even after 30 min (data not shown). BFA compartments containing PIN proteins usually started to form after 40 min of BFA treatment, mostly in the vicinity of the nucleus. Their formation in all cells was completed after 1 h of incubation with BFA. Therefore, the BFA compartment is not yet developed in Figure 5(d). Altogether, FM 4-64-induced patches are not formed from vesicles that possibly have not fused with the PM, but are rather derivatives of the PM itself. FM-induced dragging of PM proteins into cells is an active process that does not require phosphatidylinositol 3-kinase-dependent pathways. FM dyes interfere with the activity of integral plasma membrane auxin carriers To test how FM dyes interfere with the activity of integral PM auxin carriers, the radioactively labelled synthetic auxins 2,4-dichlorophenoxyacetic acid ([ 3 H]2,4-D) and naphthalene- 1-acetic acid ([ 3 H]NAA) were used as markers for active auxin influx and efflux, respectively (Petrášek et al., 2006). The active auxin influx decreased immediately after the addition of 2 lm FM 1-43 and to lesser extent after addition of 2 lm FM 4-64 or FM 5-95, as indicated by significantly decreased auxin accumulation in 2-day-old wild-type BY-2 cells (Figure 6a). The active auxin efflux was also partly influenced by FM 1-43 (i.e. the accumulation of [ 3 H]NAA increased); however, FM 4-64 and FM 5-95 had no significant effect (Figure 6b). It appears that the transient re-localization of auxin carriers caused by FM 4-64 and FM 5-95 is not sufficient to change the overall balance of auxin transport across membranes, and so changes in accumulation of labelled auxin are hardly detectable in living cells. Nevertheless, the most hydrophilic FM dye FM 1-43, which did not cause internalization of auxin carriers from the PM, had a significant effect on the auxin accumulation in cells, suggesting another mode of direct interference with auxin carrier activity. FM dyes interfere with phospholipid metabolism To test the influence of FM dyes on the PM, we investigated the dynamics of phosphatidylcholine (PC) hydrolysis. PC is a major component of the PM and a substrate of phospholipases; it plays a crucial role in many signal transduction processes in plants (Meijer and Munnik, 2003; Wang, 2004). Level of fluorescently labelled diacylglycerol (BOPIPY-DAG), a direct product of BODIPY-PC hydrolysis by the action of non-specific phospholipase C (NPC) or a non-direct product of BODIPY-PC hydrolysis by the action of phospholipase D (PLD), was used as a marker of these enzyme activities. The level of BOPIPY-DAG significantly decreased after treatment of the BY-2 cells by virtually all tested FM dyes. In 2 lm FM 1-43-treated cells, the level of BODIPY-DAG decreased to approximately 60% within 15 min compared to non-treated cells. The effect of FM 5-95 was less obvious, and the decrease in BOPIPY-DAG caused by FM 4-64 was almost non-significant (Figure 6c). When cells were treated for 30 min, there was no further decrease in BODIPY-DAG (data not shown). Thus, FM 4-64 does not alter the levels of DAG significantly. However, as in the case of auxin carrier activity, the most hydrophilic FM dye FM 1-43 strongly inhibits enzyme(s) of phospholipid metabolism in cells. ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
 Accumulation of radioactively labelled synthetic auxin [ 3 H]2,4-D reflecting the activity of auxin influx carriers.")
76 6 Adriana Jelínková et al. (a) (a) (b) (b) (c) (c) (d) Figure 6. The activity of endogenous auxin influx and efflux carriers after treatment with 2 lm FM 4-64, FM 5-95 or FM (a) Accumulation of radioactively labelled synthetic auxin [ 3 H]2,4-D reflecting the activity of auxin influx carriers. (b) Accumulation of radioactively labelled synthetic auxin [ 3 H]NAA reflecting the activity of auxin efflux carriers. (c) Production of fluorescently labelled BODIPY-DAG reflecting enzyme activities of phospholipases. Error bars represent SEs of the means (n = 4). DISCUSSION Figure 5. Testing the mechanism of FM-induced internalization of plasma membrane proteins. In vivo confocal microscopy of PIN1 GFP in 3-day-old tobacco BY-2 cells; treatment with 2 lm FM 4-64 for the indicated times. Transversal confocal sections. (a) Total inhibition of both FM 4-64 endocytosis and internalization of PIN1 GFP after 5 min pre-incubation with 15 mm sodium azide (NaN 3 ) followed by 15 min incubation with 2 lm FM (b) Pre-incubation on ice for 15 min followed by addition of 2 lm FM 4-64 and subsequent 20 min incubation on ice. Notice patches of PIN1 GFP and blocked endocytosis of FM (c, d) Pre-incubation with 50 lm wortmannin (c) or 20 lm brefeldin A (d) for 30 min followed by 10 min incubation with 2 lm FM Notice patches of PIN1 GFP after both treatments. Scale bars = 20 lm. The results presented here show a phenomenon that has not been described before, i.e. that treatment with styryl dyes of the FM family can change the localization as well as the activity of at least some integral PM proteins in plant cells. FM4-64 and FM5-95 influenced the localization of integral PM proteins. This effect was transient and related to the early phases of the interaction between FM dyes and the PM. In addition, application of FM 1-43 did not interfere with the trafficking of PM proteins, but it significantly decreased the activity of auxin transporters and phospholipid metabolism. These findings are important, especially given the fact that FM dyes are widely used for tracking the dynamics of the PM. The properties of FM molecules (Betz et al., 1996) imply ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
77 Side-effects of FM dyes in plants 7 that they cannot penetrate the PM, and so enter the cell interior only by PM internalization. This has been verified recently by testing the FRET between FM 4-64 and cytoplasmic GFP (Griffing, 2008) and by micro-injection of FM 4-64 into the cell interior (van Gisbergen et al., 2008). In addition to clathrin-mediated endocytosis (Dhonukshe et al., 2007) and sterol endocytosis (Kleine-Vehn et al., 2006; Klima and Foissner, 2008), FM 4-64 also appears to be a good marker for all other PM internalization processes in plants (Bolte et al., 2004). No toxicity or similar unwanted side effects were reported after application of optimized concentrations of FM dyes (usually between 1 and 30 lm; for review, see Bolte et al., 2004). In BY-2 tobacco cells, saturation of the PM was achieved using 17 lm FM 4-64 (Bolte et al., 2004), and this concentration did not inhibit cell division or cell-cycle progression (Kutsuna and Hasezawa, 2002). In our study, 2 lm of FM dye was enough to induce transient re-localization of PM proteins. Thus, the effective concentrations of FM dyes used in our experiments are almost 10 times lower than that reported as optimal (Bolte et al., 2004). Interestingly, FM-induced changes in the localization of integral PM proteins were not observed in A. thaliana root epidermis and cortex cells. However, tissue and even species specificity cannot be excluded. Among other possible explanations, the composition of PM in various cell types and extracellular space conditions may play critical roles here. The dye must be hydrophilic enough to pass through the cell wall and hydrophobic enough to be incorporated preferentially into the PM. In agreement with this, Meckel et al. (2004) showed that, in the highly turgescent guard cells of Vicia faba, only the most hydrophobic dye FM 4-64 can be used for tracking endocytosis, while FM 1-43 non-specifically labelled various cytoplasmic structures and mitochondria. With the current knowledge of the mechanism by which FM dyes incorporate into the PM, it is not easy to explain their effect on re-localization of integral PM proteins unambiguously. While it is not clear whether there is a preferential target for FM dyes among the spectrum of plant PM phospholipids, it is obvious that all FM dyes are positively charged and prefer specific lipids in prokaryotes. In the PM of Bacillus subtilis or Escherichia coli, FM 4-64, FM 1-43 and FM 5-95 were incorporated into the same lipid spiral-like structure enriched with anionic phospholipid phosphatidyl glycerol (Barak et al., 2008). Thus binding to specific lipids could result in lipid phase segregation, preferentially destabilizing proteins with specific requirements for lipid environment. As the degree of mis-localization of the PM auxin carriers correlated with the hydrophobicity of FM dyes, we can assume that FM 4-64, as the most hydrophobic (lipophilic) form of FM dye tested in our experiments, is incorporated into the lipid bilayer very effectively. The rapid change in the composition of the PM could trigger transient processes until a new steady state of the PM with FM dye incorporated is reached. These processes could include steric competition between FM dyes and some PM proteins, temporary changes in PM fluidity, lipid phase segregation, or a combination of any of these processes. Consequently, changes in the physical properties of the phospholipid bilayer could result in the release of some material from the PM as in the case described here, where the formation of particles that lack FM dye but contain some PM proteins was observed. At present, we are not able to conclude whether these particles are in the form of vesicles, lipid droplets or other structures. A further aspect worth mentioning is that FM dyes differ significantly in their ability to associate with a lipid bilayer. It was shown that neuronal membranes were half-saturated with FM 4-64 at a concentration approximately 10 times lower than required for half saturation with FM 1-43 (Zhu and Stevens, 2008). Therefore, a membrane can hold 10 times more molecules of FM 1-43 than of FM Thus, it can be speculated that membrane fluidity, which is known to be altered by styryl-based dyes (Rodes et al., 1995), is influenced more by FM 1-43 than by FM It is also known that alternations of membrane fluidity affect phospholipase activities (Villaverde and Verstraeten, 2003); this would explain the significant reduction of DAG levels in FM 1-43-treated cells. Various treatments of BY-2 cells were performed to examine the character of FM dye-induced PIN1 GFP or PGP4 GFP patches and the mechanism of their internalization. Treatment with BFA, which in plants act as an inhibitor of anterograde vesicle trafficking, showed that FM induced patches were derivatives of PM and are not formed from secretory vesicles that had not been able to fuse with the PM. To check the involvement of endocytosis, low temperature and the phosphatidylinositol-3-kinase inhibitor wortmannin, which have all been successfully shown to block endocytosis in plants (Emans et al., 2002; Dettmer et al., 2006), were tested. As none of the treatments prevented the formation of FM-induced patches, we concluded that it was not this type of endocytosis, but some other process, that was stimulated by the addition of FM dyes. This was further supported by distinct localizations of FM-induced PIN1 GFP or PGP4 GFP patches and FM-stained vesicles, and also by the overall different timing of endocytosis labeled with FM and appearance of patches. Prolonged incubation on ice prevented formation of FM-labelled endosomes but increased and prolonged the FM-induced re-localization effect. This suggests the mechanism requiring lower amounts of energy than the clathrin-dependent one. The formation of FM-induced PIN1 or PGP4 patches was blocked only after sodium azide treatment. This suggests that the process of FM-induced PM protein re-localization is energy-dependent, possibly via an ATP-driven step. Alternatively, FM-induced internalization could also be indirectly dependent on the cell energy ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
78 8 Adriana Jelínková et al. pool, as reported in yeast for the transmembrane potentialinduced re-distribution of specific transporter proteins (Grossmann et al., 2007). A well-established auxin accumulation assay (Delbarre et al., 1996; Petrášek et al., 2006) was used to test the effect(s) of FM dyes on the function of proteins of interest, i.e. to monitor the activity of tobacco BY-2 endogenous auxin carriers in vivo. In contrast to the formation of FM-induced patches, FM 1-43 was the most active in inhibiting cellular auxin influx and efflux, followed by FM 5-95 and FM The timing of the FM 1-43 inhibitory action on the auxin influx and efflux activities was fully comparable to the effects of widely used specific inhibitors of active auxin influx (1-naphthoxyacetic acid) and efflux (1-naphthylphthalamic acid) (Petrášek et al., 2003; Hoyerová et al., 2008). This means that both the specific inhibitors of auxin influx and efflux, as well as FM 1-43, modify auxin accumulation immediately after their application, without any lag phase. This suggests direct action on carrier proteins residing in the PM. As FM 1-43 did not induce any re-localization of PM auxin carriers, its action on auxin transporters might be related to an inhibitory effect on phospholipase(s) reflected in changed levels of DAG. DAG is a PM-located second messenger directly involved in the phosphoinositidedependent signalling system, which plays an important role in plant signalling processes. Alternatively, FM 1-43 could also interfere with the function of auxin carrier molecules by steric blockage, etc. Similar effects were observed with FM 1-43 for mechanotransduction channels of murine hair cells, neuronal cells (Gale et al., 2001; Drew and Wood, 2007) and muscarinic acetylcholine receptors (Mazzone et al., 2006). Interestingly, the inhibitory action of FM dyes on both active auxin influx and efflux was inversely proportional to their hydrophobicity. The action of FM 4-64 and 5-95 might have been attributed to the fact that after their use there are fewer auxin carriers at the PM. However, this does not apply to FM 1-43, which did not induce any internalization of these proteins. Generally, after application of these dyes to living cells, all carriers and other proteins are affected in parallel. In the case of inhibition of influx, intracellular auxin accumulation is decreased, and, in contrast, in the case of inhibition of efflux, auxin accumulation is increased. Thus the sum of these auxin transport processes across the PM, resulting in the net capacity for auxin accumulation in living cells, does not directly reflect the individual activity of auxin carriers. Only the use of auxins that are preferential substrates for either influx or efflux carriers gives an indication of the effect(s) of FM dyes on auxin transporter activity. Such treatments showed that FM dyes, particularly FM 1-43, do influence the activity of auxin transport processes across the PM, and that they do so immediately after application. Altogether, even though the FM dyes are very useful tools for tracking endocytosis in vivo, they also can induce transient internalization of at least some PM proteins and alter their activity, mainly in suspension-cultured cell lines. Therefore, a very careful characterization of action of these dyes should be performed under specific experimental conditions before they are used as endocytotic markers. The data also indicate that FM dyes do not label all endocytic pathways in plants. EXPERIMENTAL PROCEDURES Plant material and gene constructs The tobacco BY-2 cell line (N. tabacum L. cv. Bright Yellow 2; Nagata et al., 1992) and BY-2 cells transformed with A. thaliana PIN1:: PIN1:GFP (Benkova et al., 2003; Zažímalová et al., 2007) were cultivated in darkness at 25 C on an orbital incubator (IKA KS501, IKA Labortechnik, at 120 rpm (orbital diameter 30 mm) in liquid medium (3% sucrose, 4.3 g L )1 Murashige and Skoog salts, 100 mg L )1 inositol, 1 mg L )1 thiamin, 0.2 mg L )1 2,4- dichlorophenoxyacetic acid and 200 mg L )1 KH 2 PO 4, ph 5.8) supplemented (transformed cells only) with 100 mg L )1 kanamycin and 100 mg L )1 cefotaxim, and sub-cultured weekly. Calli were maintained on the same medium solidified with 0.6% w/v agar. A gene construct containing A. thaliana PGP4::PGP4:GFP in pgptv GFP (Cho et al., 2007) was introduced into Agrobacterium tumefaciens strain GV2260 and then into tobacco cells by co-cultivation as described previously (An, 1985; Petrášek et al., 2003). The PGP4- transformed lines were cultivated in liquid medium supplemented with 100 mg L )1 kanamycin, 300 mg L )1 claforan and 50 mg L )1 hygromycin B. Plants were grown on Murashige and Skoog (MS) plates (with sucrose) under a 16 h light/8 h dark photoperiod at 21/18 C. Experiments were performed during the light period. Uptake experiments using 2 lm FM4-64 were performed on 5-day-old seedlings expressing PIN2::PIN2 GFP (Xu and Scheres, 2005) or 35S::PIP2:GFP (Cutler et al., 2000) in liquid MS medium. The A. thaliana cell suspension culture was derived from PIN1::PIN1:GFP-expressing plants (Benkova et al., 2003) as described by Petrášek et al. (2006). Chemicals Boron-dipyrromethane-phosphatidylcholine [2-decanoyl 1-(O-(11- (4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-proprionyl)amino)undecyl)sn-glycero-3-phosphocholine] (BODIPY-PC) was obtained from Invitrogen ( Phospholipase A 2 (PLA 2 ) from bee venom, PC-PLC from Bacillus cereus and PLD from cabbage were obtained from Sigma-Aldrich Co. ( HPLC-grade chloroform, ethanol, methanol and high-performance thin layer chromatography (HP-TLC) plates were obtained from Merck ( All other chemicals were of analytical grade. FM dyes 4-64, 5-95 and 1-43 (Molecular Probes, catalogue numbers T13320, T23360, T3163), brefeldin A, wortmannin, latrunculin B and oryzalin (all Sigma-Aldrich, were kept as 20, 50, 10, 2.5 and 57 mm stock solutions, respectively, in DMSO at )20 C. Sodium azide was added from a 1.5 M stock solution in water to a final concentration of 15 mm. Application of FM dyes and inhibitors FM dyes were added to 1 ml of 2 3-day-old BY-2 cell suspension under continuous shaking in multi-well plates, and samples were observed at the time indicated. Alternatively, the protocol described by Emans et al. (2002) was used. 1 ml of a 2 3-day-old BY-2 cell ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
79 Side-effects of FM dyes in plants 9 suspension was placed on ice for 15 min and then supplemented with 2 lm FM dye. After another 15 min of incubation on ice, cells were transferred to an orbital shaker at 26 C and observed at the time indicated. Brefeldin A, sodium azide and wortmannin were added directly to the cultivation medium to final concentrations of 20 lm, 15 mm and 50 lm, respectively. The same amount of solvent was added to controls. Seedlings were primed for 5 min on ice with MS medium containing 2 lm FM4-64, subsequently rinsed three times in liquid MS medium, and analysed by confocal microscopy. Microscopy For in vivo microscopy, a Zeiss LSM 5 DUO confocal microscope with a 40 C-Apochromat objective (NA = 1.2 W) was used ( Fluorescence signals for GFP (excitation 488 nm, emission nm), FM 4-64 and FM 5-59 (both excitation 561 nm, emission >575 nm) were detected. Sequential scanning was used to avoid any cross-talk between fluorescence channels. To separate FM1-43 and GFP fluorescence, spectral fluorescence detection and subsequent linear unmixing were used. Lambda series (excitation 488 and 561 nm) were collected using the Zeiss META system. Nineteen channels (10.7 nm wide) of the META detector were used to span an emission range of nm. Linear unmixing was performed using Zeiss LSM IMAGE EXAMINER software. Reference spectra of GFP and FM1-43 were acquired from single labelled specimens. The scattergram of fluorescence intensities in the red and green channels was constructed using Zeiss IMAGE EXAMINER software. Independent experiments were performed in triplicate with the same results, and representative images are shown. Auxin accumulation assay Auxin accumulation by BY-2 cells was measured according to the method described by Delbarre et al. (1996), as modified by Petrášek et al. (2003). Accumulation by the cells of [ 3 H]NAA (specific radioactivity 935 GBq mmol )1, synthesized at the Isotope Laboratory, Institute of Experimental Botany, Prague, Czech Republic) or [ 3 H]2,4-D (specific radioactivity 20 Ci mmol )1, American Radiolabeled Chemicals Inc., was measured in 0.5 ml aliquots of cell suspension (cell density approximately cells ml )1, as determined by counting cells in a Fuchs Rosenthal haemocytometer). FM dyes were added at the beginning of the accumulation assay. The values for the net accumulation of auxin (with the value at time 0 subtracted) after 20 min of incubation with radioactive auxin and FM dye were plotted relative to the control accumulation without FM dyes. Determination of non-specific phospholipase C activity Fluorescent substrate (0.66 lg ml )1 BODIPY-PC) was added to a 3-day-old BY-2 cell suspension. The suspension was diluted to g FW ml )1. After 10 min of incubation, FM dye was added to the final concentration of 10 lm. After incubation on an orbital shaker at 26 C in darkness for 15 min, the cells were harvested for the extraction of lipids and HP-TLC analysis of reaction products. Lipids were extracted by a modification of the method described by Bligh and Dyer (1959). Briefly, 4 ml of methanol/chloroform (2/1 v/v) was added to harvested cells. After 30 min of extraction, 2 ml 0.1 M KCl was added, and the mixture was left at 4 C for at least 30 min. Then samples were centrifuged for 15 min at 420 g. The lower phase was evaporated to dryness using a vacuum evaporator and re-dissolved in ethanol. Samples were applied to HP-TLC silica gel-60 plates using a Linomat IV sampler (Camag, camag.com). After 10 min of saturation, plates were developed in a horizontal developing chamber (Camag) in a mobile phase of chloroform/methanol/water (65/25/4 v/v/v) (Scherer et al., 2002). Plates were dried and scanned using a video camera (Kodak DC 120, under UV light for computer-assisted quantification (Kodak ds 1D). Identification of individual spots was based on comparison with fluorescently labelled lipid standards. ACKNOWLEDGEMENTS We thank Jan Malínský (Institute of Experimental Medicine, Prague, the Czech Republic) for helpful discussion, Hyung-Taeg Cho (Chungnam National University, Daejeon, Korea) and Angus Murphy (Purdue University, West Lafayette, IN, USA) for PGP4:: PGP4:GFP gene constructs, and Ben Scheres (Utrecht University, Utrecht, the Netherlands) and Chris Somerville (University of California, Berkeley, CA, USA) for A. thaliana seeds expressing PIN2 GFP and PIP2 GFP proteins, respectively. This work was supported by the Grant Agency of the Academy of Sciences of the Czech Republic (grant number KJB to J.P.), by the Grant Agency of Charles University (grant number to A.J. and S.S.) and by a grant from the Ministry of Education, Youth and Sports of the Czech Republic to E.Z. (LC06034). SUPPORTING INFORMATION Additional Supporting Information may be found in the online version of this article: Figure S1. FM 4-64-stimulated transient internalization of plasma membrane-located auxin carriers. Figure S2. De-polymerization of the actin or microtubule cytoskeleton does not abolish FM 4-64-induced internalization of plasma membrane-located auxin carriers. Movie S1. Re-localization of PIN1 GFP into patches in the cytoplasm after 3 min treatment with 2 lm FM Movie S2. PIN1 GFP localization after 3 min mock treatment with the appropriate amount of solvent (DMSO). Please note: As a service to our authors and readers, this journal provides supporting information supplied by the authors. Such materials are peer-reviewed and may be re-organized for online delivery, but are not copy-edited or typeset. Technical support issues arising from supporting information (other than missing files) should be addressed to the authors. REFERENCES An, G. (1985) High-efficiency transformation of cultured tobacco cells. Plant Physiol. 79, Aniento, F. and Robinson, D.G. (2005) Testing for endocytosis in plants. Protoplasma, 226, Barak, I., Muchova, K., Wilkinson, A.J., O Toole, P.J. and Pavlendova, N. (2008) Lipid spirals in Bacillus subtilis and their role in cell division. Mol. Microbiol. 68, Benkova, E., Michniewicz, M., Sauer, M., Teichmann, T., Seifertova, D., Jurgens, G. and Friml, J. (2003) Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell, 115, Betz, W.J. and Bewick, G.S. (1992) Optical analysis of synaptic vesicle recycling at the frog neuromuscular-junction. Science, 255, Betz, W.J., Mao, F. and Bewick, G.S. (1992) Activity-dependent fluorescent staining and destaining of living vertebrate motor-nerve terminals. J. Neurosci. 12, Betz, W.J., Mao, F. and Smith, C.B. (1996) Imaging exocytosis and endocytosis. Curr. Opin. Neurobiol. 6, Bligh, E.G. and Dyer, W.J. (1959) A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, Bolte, S., Talbot, C., Boutté, Y., Catrice, O., Read, N.D. and Satiat-Jeunemaitre, B. (2004) FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. J. Microsc. 214, ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
80 10 Adriana Jelínková et al. Cho, M., Lee, S. and Cho, H. (2007) P-glycoprotein 4 displays auxin efflux transporter-like action in Arabidopsis root hair cells and tobacco cells. Plant Cell, 19, Cutler, S.R., Ehrhardt, D.W., Griffitts, J.S. and Somerville, C.R. (2000) Random GFP :: cdna fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. Proc. Natl Acad. Sci. USA, 97, Delbarre, A., Muller, P., Imhoff, V. and Guern, J. (1996) Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1-acetic acid, and indole-3-acetic acid in suspension-cultured tobacco cells. Planta, 198, Dettmer, J., Hong-Hermesdorf, A., Stierhof, Y.D. and Schumacher, K. (2006) Vacuolar H + -ATPase activity is required for endocytic and secretory trafficking in Arabidopsis. Plant Cell, 18, Dhonukshe, P., Aniento, F., Hwang, I., Robinson, D., Mravec, J., Stierhof, Y. and Friml, J. (2007) Clathrin-mediated constitutive endocytosis of PIN auxin efflux carriers in Arabidopsis. Curr. Biol. 17, Drew, L.J. and Wood, J.N. (2007) FM1-43 is a permeant blocker of mechanosensitive ion channels in sensory neurons and inhibits behavioural responses to mechanical stimuli. Mol. Pain, 3, 1. Emans, N., Zimmermann, S. and Fischer, R. (2002) Uptake of a fluorescent marker in plant cells is sensitive to brefeldin A and wortmannin. Plant Cell, 14, Gaffield, M.A. and Betz, W.J. (2006) Imaging synaptic vesicle exocytosis and endocytosis with FM dyes. Nat. Protoc. 1, Gale, J.E., Marcotti, W., Kennedy, H.J., Kros, C.J. and Richardson, G.P. (2001) FM1-43 dye behaves as a permeant blocker of the hair-cell mechanotransducer channel. J. Neurosci. 21, van Gisbergen, P., Esseling-Ozdoba, A. and Vos, J. (2008) Microinjecting FM4-64 validates it as a marker of the endocytic pathway in plants. J. Microsc. 231, Griffing, L. (2008) FRET analysis of transmembrane flipping of FM4-64 in plant cells: is FM4-64 a robust marker for endocytosis? J. Microsc. 231, Grinvald, A., Frostig, R.D., Lieke, E. and Hildesheim, R. (1988) Optical imaging of neuronal activity. Physiol. Rev. 68, Grossmann, G., Opekarová, M., Malinsky, J., Weig-Meckl, I. and Tanner, W. (2007) Membrane potential governs lateral segregation of plasma membrane proteins and lipids in yeast. EMBO J. 26, 1 8. Hoyerová, K., Perry, L., Hand, P., Laňková, M., Kocábek, T., May, S., Kottová, J., Pačes, J., Napier, R. and Zažímalová, E. (2008) Functional characterization of PaLAX1, a putative auxin permease, in heterologous plant systems. Plant Physiol. 146, Kleine-Vehn, J., Dhonukshe, P., Swarup, R., Bennett, M. and Friml, J. (2006) Subcellular trafficking of the Arabidopsis auxin influx carrier AUX1 uses a novel pathway distinct from PIN1. Plant Cell, 18, Klima, A. and Foissner, I. (2008) FM dyes label sterol-rich plasma membrane domains and are internalized independently of the cytoskeleton in characean internodal cells. Plant Cell Physiol. 49, Kutsuna, N. and Hasezawa, S. (2002) Dynamic organization of vacuolar and microtubule structures during cell cycle progression in synchronized tobacco BY-2 cells. Plant Cell Physiol. 43, Mazzone, S.B., Mori, N., Burman, M., Palovich, M., Belmonte, K.E. and Canning, B.J. (2006) Fluorescent styryl dyes FM1-43 and FM2-10 are muscarinic receptor antagonists: intravital visualization of receptor occupancy. J. Physiol. 575, Meckel, T., Hurst, A.C., Thiel, G. and Homann, U. (2004) Endocytosis against high turgor: intact guard cells of Vicia faba constitutively endocytose fluorescently labelled plasma membrane and GFP-tagged K + -channel KAT1. Plant J. 39, Meijer, H.J.G. and Munnik, T. (2003) Phospholipid-based signaling inplants. Annu. Rev. Plant Biol. 54, Nagata, T., Nemoto, Y. and Hasezawa, S. (1992) Tobacco BY-2 cell-line as the HeLa cell in the cell biology of higher plants. Int. Rev. Cytol. 132, Oparka, K.J. (1990) What is phloem unloading? Plant Physiol. 94, Perez-Gomez, J. and Moore, I. (2007) Plant endocytosis: it is clathrin after all. Curr. Biol. 17, R217 R219. Petrášek, J., Černá, A., Schwarzerová, K., Elčkner, M., Morris, D.A. and Zažímalová, E. (2003) Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiol. 131, Petrášek, J., Mravec, J., Bouchard, R. et al. (2006) PIN proteins perform a ratelimiting function in cellular auxin efflux. Science, 312, Rodes, J.F., Berreurbonnenfant, J., Tremolieres, A. and Brown, S.C. (1995) Modulation of membrane fluidity and lipidic metabolism in transformed rat fibroblasts induced by the sesquiterpenic hormone farnesylacetone. Cytometry, 19, Scherer, G.F.E., Paul, R.U., Holk, A. and Martinec, J. (2002) Down-regulation by elicitors of phosphatidylcholine-hydrolyzing phospholipase C and upregulation of phospholipase A in plant cells. Biochem. Biophys. Res. Commun. 293, Terasaka, K., Blakeslee, J.J., Titapiwatanakun, B. et al. (2005) PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots. Plant Cell, 17, Villaverde, M.S. and Verstraeten, S.V. (2003) Effects of thallium(i) and thallium(iii) on liposome membrane physical properties. Arch. Biochem. Biophys. 417, Wang, X. (2004) Lipid signaling. Curr. Opin. Plant Biol. 7, Xu, J. and Scheres, B. (2005) Dissection of Arabidopsis ADP-RIBOSY- LATION FACTOR 1 function in epidermal cell polarity. Plant Cell, 17, Yang, H.B. and Murphy, A.S. (2009) Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 59, Zažímalová, E., Křeček, P., Skůpa, P., Hoyerová, K. and Petrášek, J. (2007) Polar transport of the plant hormone auxin the role of PIN-FORMED (PIN) proteins. Cell. Mol. Life Sci. 64, Zhu, Y. and Stevens, C.F. (2008) Probing synaptic vesicle fusion by altering mechanical properties of the neuronal surface membrane. Proc. Natl Acad. Sci. USA, 105, Zonia, L. and Munnik, T. (2008) Vesicle trafficking dynamics and visualization of zones of exocytosis and endocytosis in tobacco pollen tubes. J. Exp. Bot. 59, ª 2010 Institute of Experimental Botany, ASCR Journal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), doi: /j X x
81 Supporting Information Figure S1. FM 4-64-stimulated transient internalization of plasma membranelocated auxin carriers, fast effects, mock treatments and the quantification. In vivo confocal microscopy of PIN1-GFP (a) and PGP4-GFP (b) in 3-day-old tobacco BY-2 cells; treatment with 2 µm FM 4-64 for indicated times. (c) Mock treatment for figure 2a. DMSO 1: (v/v) was added to PIN1-GFP expressing cells and images were taken at 2 minutes, 10 minutes and 30 minutes after the addition of DMSO. No changes in the localization of PIN1-GFP were observed compared to the non-treated cells and no increase in the formation of PIN1-GFP patches was observed. (d) Mock treatment for figure 4b. Cells were pre-incubated on ice for 15 minutes, than DMSO 1:10000 (v/v) was added and cells further incubated on ice. Images were captured at 15 and 30 minutes after the addition of DMSO. DMSO did not induce the formation of PIN1-GFP vesicles as observed after FM4-64 treatment. (e) Mean plasma membrane grey levels after 15 min treatment with FM dyes reflecting the relocalization of the PIN1-GFP into the endosomal space. Note that FM 4-64 has the most remarkable effect. After the interactive thresholding of the plasma membranes (indicated by yellow bars in the image on the right) the mean value of fluorescence intensity was recorded for both time points: just after the application of the respective dye and after 15 min incubation. Cells treated for 15 minutes with DMSO (1:10000 v/v) or 2µM FM dyes were compared to the non-treated cells, mean grey levels (8 bit resolution) reflect the intensity of the fluorescence in the plasma membrane. All cell chains containing three and more cells from 9 optical fields were analyzed. Error bars represent SD. 1
82 The test of equality of variances indicated that the variances of the grey levels of the plasma membrane after 15 min of treatment were significantly different from control cells both for FM 4-64-treated cells (F=3.2, p=0.004) and FM 1-43-treated cells (F=2.98, p=0.002) and significantly equal for FM 5-59-treated cells (F=1.01, p=0.486). Therefore, a two sample t-tests that do not assume equal variances were performed for the comparison of mean grey levels reflecting the plasma membrane fluorescence of control and FM 4-64 and FM 1-43-treated cells. For FM 5-59, a two sample t-test assuming equal variances was performed. The mean grey levels for FM 4-64-treated cells (M=62.77; SD=24.85; n=22) were significantly smaller than in control cells (M=122.78; SD=44.48; n=28) as tested by the two-sample t-test for unequal variances, t(44)=6.04, p<= Differences in grey values for FM1-43 and FM 5-59 were statistically insignificant using the t test for unequal and equal variances, respectively (p>=0.005). Figure S2. Depolymerization of actin or microtubule cytoskeleton does not abolish FM 4-64-induced internalization of plasma membrane-located auxin carriers. In vivo confocal microscopy of PIN1-GFP in 3-day-old tobacco BY-2 cells treated with latrunculin B (a) or oryzalin (b). Cells were stained with 2 µm FM 4-64 for 15 minutes. In cells with depolymerized actin is PIN1GFP localization less affected by FM4-64 dye, whereas depolarization of microtubules results in increased relocalization of PIN1GFP. To depolymerize actin cytoskeleton, BY2 cells were incubated with 500nM Latrunculin B for 10 minutes. Subsequently 2µM FM 4-64 was added and images were taken at times indicated. For depolymerization of microtubules 20µM oryzalin was used. 2
83 Video S1. Relocalization of PIN1-GFP into patches in the cytoplasm after 3 min of treatment with 2 µm FM Frame rate 7 frames/s. Video S2. PIN1-GFP after 3 min mock treatment with the appropriate amount of the solvent (DMSO). Frame rate 7 frames/s. 3


 2 min 10")
 30 min")





 FM")

84 Figure S1 2 min 0 min (a) PIN1-GFP PIN1-GFP PGP4-GFP PGP4-GFP (c) 2 min 10 min FM 4-64 (d) 30 min PIN1-GFP/ice (e) Mean plasma membrane grey levels after 15 min Control FM 4-64 FM 5-95 Merged Merged PIN1-GFP/DMSO (b) FM 4-64 FM min 30 min
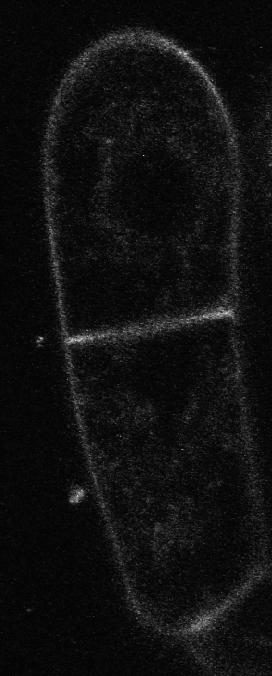

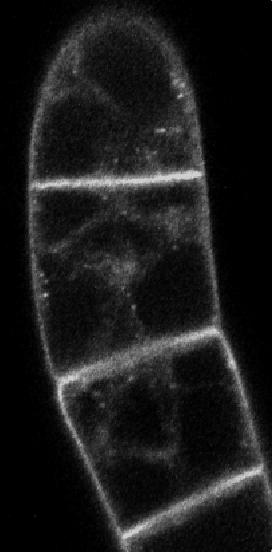
85

86 3.4 Cytoskeletal proteins mediating endocytosis and targeted deposition of auxin transporters in Arabidopsis thaliana The primary objective of this study is to find all possible cytoskeleton and cytoskeletonassociated proteins important for the processes of endocytosis and polarized targeting of PIN auxin efflux proteins. Available Arabidopsis thaliana mutants are tested in the microscopical screen for the rate of endocytosis in the root tip endodermis and cortex cells, scored by an in vivo FM 4-64 staining, and for the localization of PIN1 and PIN2 the root apex. The chapter is presented here in the form of the text that is in the preparation for the publication. It contains the first set of data form the analysis of actin and some actin associated proteins. The screening of other mutants is on progress. I am the only one who performed all experiments described here (collection of the available mutants, cultivation of plants, confocal microscopy, immunofluorescence stainings inw hole mounts). I have been also involved in the interpretation of the data, designing and coordination of the experiments and writing of the manuscript. Whole mount immunolocalization of PIN1 auxin efflux carrier in the root of wild type Arabidopsis thaliana seedlings 51
87 52
88 Abstract Thanks to its sessile life strategy, the polarity of plant body reflects the polarity of single cells. The polarity is maintained by asymmetric distribution of various molecules downstream from extra and intracellular signals. Directional transport of auxin plays an important role in the pattern formation, morphogenesis, and directional growth responses. The positioning of PIN auxin efflux transporters has been shown to be crucial in the setting of auxin gradients. It is dependent on the plasma membrane deposition of membrane vesicles and their constitutive cycling between plasma membrane and endosomal space. Although some evidences support the idea of differential actin and microtubular cytoskeleton dependence of PIN protein trafficking, there is a significant lack of the information on the role of cytoskeleton in this process. In this paper we use combination of live cell imaging and immunofluorescence techniques to search for the molecular players of actin filaments (AFs) and proteins proteins associated with AFs in the mechanisms of endocytosis and directed PIN1 protein targeting. Seedlings of Arabidopsis thaliana carrying mutations in actin genes (ACT1, ACT2, ACT7, ACT11), Arp 2/3 complex genes (ARP2, ARP3, ARPC2, ARPC5), WAVE complex components genes (BRK1, NAP1, SRA1) and actin monomer binding proteins genes (ADF1, ADF4, ADF5, PRF1, PRF2) were tested for the the rate of the endocytosis using FM-4-64 uptake by root epidermal and cortex cells. Simultaneously, PIN1 protein was immunolocalized in whole mounts by indirect immunofluorescence. We show here that with only few exceptions all mutations presented here either reduced the rate of FM 4-64 endocytosis and its progression or changed the appearance of endocytic vesicles. The localization of PIN1 in the root tip stele cells did not show any significant changes in all mutants screened here with one exception. Plant carrying the mutation in the ARPC5 gene coding for the subunit of Arp2/3 complex had prominent PIN1 plasma membrane staining and some granulated signal through the cytoplasm. Our results suggest that actin and actinassociated proteins analyzed here seem to be not primarily important for PIN1 localization within the basal plasma membrane of root provasculature. However, they are essential for proper membrane trafficking and endocytic vesicle formation and dynamics which makes in turn part of the PIN1 protein targeting and cycling. The analysis of other mutations is in progress. 53
89 Introduction The polarization of auxin transporters is linked to the vesicle trafficking (Steinmann et al. 1999, Geldner et al. 2003, Kleine-Vehn et al. 2006) and their constitutive cycling in the endosomal space seems to be actin- and myosin dependent (Geldner et al. 2001; Holweg et al 2004; Holweg 2007). Moreover, it seems that with respect to the cytoskeleton, apical-basal localization of PIN proteins follows distinct pathways as suggested by experiments in Arabidopsis thaliana roots. Here, the process of cycling and translocation of endosomes with PIN proteins from one membrane domain to the other, so-called transcytosis (Kleine-Vehn et al. 2008a), has been shown to involve preferential involvement of actin filaments in the apical-basal targeting of PINs and microtubules preferentially in basal targeting (Kleine-Vehn et al. 2008b). The participation of microtubules in PIN positioning was proposed earlier using anti-tubulin drugs (Boutté et al. 2006). However the information about mechanism of the participation of cytoskeleton in auxin efflux carrier trafficking and targeting is limited. Mutual interaction between cytoskeleton and auxin transport and between cytoskeleton and its involvement in the localization of auxin transporters is generally expected to play a role on two levels. These are level of fast responses (nongenomic) and level of differential gene expression and downstream events playing a role in long-term interactions. The treatment of cells with cytoskeletal drugs, inhibitors of vesicle trafficking and auxin transport inhibitors. Cytochalasin D (Cyt D), drug known to disrupt actin filaments influenced both PIN1 polar localization and auxin transport (Butler et al. 1998; Geldner et al. 2001). Brefeldin A (BFA), which is known to interfere with anterograde protein transport, induced accumulation of PIN1 proteins in endosomal compartments (Geldner et al. 2001), reduced auxin efflux and also caused remodelling of actin filaments (Petrášek et al. 2003). The accumulation of PIN1 in intracellular compartments upon BFA treatment is reversible and this reversibility is uncoupled in the presence of Cyt D, indicating that this process is actindependent (Geldner et al. 2001). Moreover, endocytosis of membrane vesicles is inhibited by auxin itself, which leads to the promotion of its own efflux (Paciorek et al. 2005). This inhibitory effect of auxin can thus be used as powerful tool for uncovering its own action. Stabilizing effect of auxin transport inhibitors TIBA (2,3,5-triiodobenzoic acid) and PBA (2-(1-pyrenoyl) benzoic acid) on actin filaments and its role in impairing vesicle traffic was demonstrated next to plant cells also in other eukaryotes (Dhonukshe et al. 2008). Actin bundling induced by overexpressing mouse actin-associated protein talin impairs cell division synchrony and increases its sensitivity to another inhibitor of auxin transport 1-Nnaphthylphthalamic acid (NPA). Normal actin organization and synchronized cell division was re-established by addition of auxins that are subject of auxin efflux in the tissue, IAA and 54
90 NAA (Maish and Nick 2007). All these findings point to the idea that auxin could control its own transport by changing actin filament dynamics and thus vesicle trafficking of own transporters (Nick et al. 2009). As summarized above, there is quite solid evidence about the role of cytoskeleton in the transport of auxin, but all possible mechanisms are still point of debate. There is significant lack of systematic knowledge on the components linking the dynamics of cytoskeleton and processes of endocytosis in plants and about components necessary for the trafficking of auxin transporters, in particular. Therefore, detailed classification of all cytoskeleton and cytoskeleton-associated proteins in terms of their involvement in endocytosis and auxin efflux transporter localization is highly needed. In our laboratory, we have started with the collection of mutants in cytoskeletal and associated proteins and at the moment we have around 30 homozygous mutant lines and other 30 lines from which homozygous lines should be isolated. The analysis was done in all of the available actin protein mutants and some actin-associated proteins. The seeds were ordered at the Nottingham Arabidopsis Stock centre (NASC).For the selection of T-DNA insertion and other mutant lines, the information from exhaustive reviews on the actin-binding (Higaki et al. 2007, Staiger et al. 2010) and microtubule-binding proteins (Buschmann and Lloyd 2008) as well as vegetative actin genes (Kandasamy et al. 2007) and tubulin genes (Ludwig 1987, Oppenheimer 1988, Kopczak et al. 1992, Snustad 1992) is used. In vegetative cells of Arabidopsis thaliana are highly expressed three actin isoforms: ACTIN 2, ACTIN7 and ACTIN8 (Kandasamy et al. 2009). Dynamic behaviour of actin highly depends on the presence actin binding proteins (ABPs, there were identified more than 70 classes in eucaryotes). The ABPs could be roughly divided into following groups: nucleating factors, capping proteins, severing proteins and sidebinding proteins (reviewed by Staiger et al. 2010). In this manuscript data from the first part of the screen of the so far available mutants in actin proteins, actin nucleating proteins and their subunits and some of actin monomer binding proteins are presented. In general, they show that vegetative actins as well as reproductive actins play its role in endocytosis and trafficking of the membrane vesicles in Arabidopsis root cells. Data from screening of Arp 2/3 mutants and its activators clearly suggest that these actin nucleation factors are involved in the endocytosis (see Deeks and Hussey, 2005) of diffusely-growing plant cells. 55
91 Protein ACT1 ACT2 ACT7 ACT11 ARP2 ARP3 ARPC2B ARPC5 BRK1 NAP1 SRA1 ADF1 ADF4 ADF5 PRF1 PRF2 Locus AT2G37620 AT3G18780 AT5G09810 AT3G12110 AT3G27000 AT1G13180 AT2G33385 AT4G01710 AT2G22640 AT2G35110 AT5G18410 AT3G46010 AT5G59890 AT2G16700 AT2G19760 AT4G29350 Mutant analyzed (NASC line) N656765, T-DNA, hm N25256, EMS, hm N670149, T-DNA, hm N665143, T-DNA, hm N577920, T-DNA, hm N675737, T-DNA, hm N T-DNA, hm N T-DNA, hm N86554, EMS, TILLING, ns N538799, T-DNA, ht N T-DNA, ht N T-DNA, hm N621551, T-DNA, ht N662447, T-DNA, hm N656528, T-DNA, hm N T-DNA, hm Original reference Nairn et al McKinney et al.1995 McDowell et al.1996 Huang et al.1997 Mathur et al Markus et al Mathur et al Mathur et al Zimmermann et al Hulskamp et al Hulskamp et al Carlier M et al.1997 Sweetlove et al Alonso et al Huang et al.1996 Christensen et al Annotation Preferential expression during reproductive development, root moderate expression Essential for root hair tip growth, single mutation mild phenotypes Essential for root growth, epidermal cell specification, cell division and root architecture Preferential expression during reproductive development, root moderate expression Arp 2/3 complex protein, protein binding, expressed during seedling growth Arp 2/3 complex protein, ATP, actin and protein binding, expressed during seedling growth Subunit of Arp 2/3 complex, F-actin organization in trichomes, structural molecule activity Subunit of Arp 2/3 complex, cell expansion of trichomes WAVE complex component, lobe formation of pavement cells, morphogenesis of stomata and root hairs WAVE complex component, trichomes morphogenesis, WAVE complex component, actin based cell morphogenesis Key organization of actin, turnover of actin, monomer and filament binding Key organizator of actin, turnover of actin, monomer and filament binding Key organizator of actin, turnover of actin, monomer and filament binding Expressed in vegetative organs, regulates actin polymerization at PM sites References Kandasamy et al Kandasamy et al Kandasamy et al Huang et al.1997 Mathur et al. 2003a Mathur et al. 2003a Mathur et al Mathur et al. 2003b Djakovic et al Deeks et al Li et al Dong et al Dong et al Dong et al Deeks et al Expressed in vegetative organs Christensen et al Table1: Actin and actin-associated proteins of Arabidopsis thaliana and corresponding mutant lines used in this work. hm - ordered from NASC as a homozygous line, ht - ordered from NASC as heterozygote line, ns - not specified. 56
92 Materials and methods Plant material Plants were grown on vertically-oriented agar plates containing Arabidopsis medium (AM, MS/2, 1.5% agar, 1% sucrose, ph 5.8) under a 16/8 h light/dark photoperiod at 21/18 C. Experiments were performed during the light period in 4-day-old seedlings of individual mutants of Arabidopsis thaliana cv. Col-0 obtained from NASC (Nottingham Arabidopsis Stock Centre). The list of analyzed mutants is described in Table 1. For all individual experiments wild type Col-0 plants were always included, analyses were done in triplicates. Application of FM 4-64 dye Seedlings were primed for 5 min on ice with MS medium containing 2 μm FM 4-64 (Molecular Probes, catalogue n. T13320) and subsequently rinsed three times in liquid MS medium and analysed by confocal microscopy at the times indicated. Whole mount immunolocalization of PIN1 protein PIN1 in the root tip was immunolocalized in whole-mounts using automated system for immunocytochemistry (InsituPro VSi, Intavis, Germany). The protocol (InsituPro VS protocol Arabidopsis whole mount 1, Robert and Offringa, Leiden, Netherlands) was optimized. The 4-day-old Arabidopsis seedlings were fixed in 4% paraformaldehyde in MTSB (50 mm PIPES, 5 mm EGTA, 5 mm MgSO 4, ph 7 adjusted with KOH) for 1 h under the vacuum. Samples were washed with MTSB with 0.1% Triton (5 10 min) and with deionised water (5 10 min). Cell wall was then digested with 0.05% pectolyase in MTSB with mannitol (0.45M) for min and samples washed with MTSB with 0.1% Triton (5 10 min). Incubation with 10% DMSO and 3% NP-40 in MTSB for 1 h followed. After another washing in MTSB with 0.1% Triton (5 10 min) seedlings were pre-incubated in 2% BSA in MTSB (1h, 37 C) and incubated with the epitope-specific primary monoclonal antibody against PIN1 (PIN1 (ak-13): sc-27161, Santa Cruz Biotechnology) in 3% BSA in MTSB (5 h, 37 C). After extensive washing with MTSB with 0.1% Triton (8 10 min), the seedlings were incubated with the secondary antibody (Alexa 488, Molecular Probes, catalogue n. A-11059) in 3% BSA in MTSB for another 3h (37 C). Finally, the samples were washed with MTSB with 0.1% Triton (5 10 min) and with deionised water (5 10 min) and and mounted into 50% glycerol with 0.02% sodium azide. 57
93 Microscopy Zeiss LSM 5 DUO confocal microscope with a 40x C-Apochromat objective (NA = 1.2 W) was used. Fluorescence signals of Alexa 488 (excitation 488 nm, emission nm) and FM 4-64 (excitation 561 nm, emission >575 nm) were detected. Results and discussion The screen data presented within this manuscript involve mutations in reproductive actins ACT1 and ACT11, vegetative actins ACT2 and ACT7, in proteins of actin-related protein complex Arp 2/3 ARP2, ARP3 and some of its subunits ARPC2B and ARPC5, in proteins of WAVE Arp 2/3 activating complex: BRK1, NAP1, and SRA1 and in actin monomer binding proteins: ADF1, ADF4, ADF5, PRF1 and PRF2. Most of the mutant seeds were ordered from NASC as homozygous lines (see Table 1). The genotyping of all lines is in progress. The screening of mutants in actin genes involved the members of both vegetative actins (ACT2 and ACT7) and generative actins (ACT1 and ACT11). According to Genevestigator ( data vegetative actins are much more expressed in root tissues than generative, but the generative have also some background expression in these tissues. T- DNA insertion homozygous mutants in ACT1, ACT7 and ACT11 genes and EMS mutant for ACT2 were analyzed, respective mutants were named act2, act7 and act1, act11 for the purpose of this screen (the identity of the particular mutant is given in Table 1). 3-day-old seedlings were either incubated in FM 4-64 (2 μm) and traced its endocytotic uptake or the roots were fixed and stained with PIN1 antibody to uncover aberrations in the auxin efflux localization within PM by the particular actin mutation (Figure 1). All mutants, except the reproductive actin mutation act1 (Figure 1B) inhibited endocytosis of FM4-64 dye. However, even there after 20 minutes of incubation with FM 4-64 unusual clusters of endocytic vesicles were observed (Figure 1B). In the act2 mutant the inhibition of the uptake of FM 4-64 dye was observed, at all particular times of the uptake there was remarkably less endocytic vesicles through the cytoplasm of the endodermis and the cortex cells (Figure 1C). Endocytosis was altered also in act7 mutant, where the inhibition of FM 4-64 uptake by the epidermal and cortex cells was accompanied by the accumulation of membrane vesicles at the plasma membrane of epidermal cells after 20 min (Figure 1D). Unexpectedly, mutation in reproductive act11 caused inhibition of endocytosis as well and endocytic vesicles accumulated at the outer side of epidermal cell layer (Figure 1E). Moreover, the epidermal, as well as the cortex cells of act7 and act11 mutant roots (Figure 1I, J) appeared thicker and swollen comparing with the wild type (Figure 1F) or the other two mutation act1 and act2 58
94 roots (Figure 2G, H). This phenotype of root swelling and bulge roots has been reported in act7-4 (Gilliand et al., 2003). Interestingly, this effect was observed here also in reproductive act11 mutant. These results indicate that ACT11 isoform might be important for the vegetative tissues as well, although its expression here is mild (Genevestigator). The bulging effect could be partly caused by inhibition of endocytosis caused by the mutation in ACT7 or ACT11 gene. The endocytosis is not inhibited fully by the mutation of the single actin isoform, which is in agreement with the fact that only double mutants of vegetative actins had drastical dwarfism (Kandasamy et al. 2009). Therefore, it is planned to include also all available double mutants to the screen. All of the actin mutations studied in this screen were found not to interfere with PIN1 protein localization at the PM (Figure 1L-1O) comparing with the wild type PIN1 localization (Figure 1K). Interestingly, ACT7 has been reported earlier to be induced by auxin (Kandasamy et al. 2001) and therefore it could be speculated that auxin maxima generated in the root tip or during lateral root formation could be connected with the selective upregulation of actin genes that would afterwards enhance processes of constitutive cycling and transcytosis of PIN carriers (Kleine-Vehn et al. 2008). It is planned to test this hypotheses with the addition of PIN2 immunolocalization in the epidermal and cortex cells. In general, presented data show that vegetative actins as well as reproductive actins play its role in endocytosis and trafficking of the membrane vesicles in Arabidopsis root cells. These results further support the idea that the actin remodelling observed after BFA around membrane vesicles in BY-2 cells (Petrášek et al. 2003) represents the active process of mutual co-operation between actin filaments and membrane vesicles. The screening of mutants in the genes coding Arp 2/3 complex subunits and its activators involved ARP2, ARP3, ARPC2b, ARPC5 and WAVE complex components BRK1, NAP1 and SRA1proteins. The Arp 2/3 complex is one of the possible nucleators of actin in plants (Staiger and Blanchoin 2006), although more important role play the formin-assisted nucleation (Cvrčková et al. 2004). This complex is activated by WAVE (BRK1, NAP1, SRA1) protein complex. Some of the subunits of WAVE complex appear to be ROP (Rho of Plants) effectors that translate ROP-GTPase signals into nucleation response of actin filaments. The WAVE-Arp 2/3 activation might generate new actin nucleation domains at the cortical cytoplasm and thus to contribute into coordinated cell expansion (Szymansky 2009). T-DNA insertion homozygous mutants in ARP2, ARP3, ARPC2b, ARPC5 genes, EMS TILLING heterozygous mutant in BRK1 and T-DNA insertion heterozygous mutants in NAP1 and SRA1 genes were analyzed. Respective mutants were named arp2, arp3, arpc2b, arpc5, brk1, nap1 and sra1 for the purpose of this screen (the identity of the particular mutant is given in Table 1). Endocytosis of both arp2 and arp3 mutants was inhibited, there were 59
95 almost no endocytic vesicles noticed within the cytoplasm even after 20 min of the FM 4-64 incubation (Figure 2B and 2C). Moreover, arp3 mutant showed some aberrations in the shape of the epidermal cells (Figure 2C), which is in accordance with the phenotype of dis1-1 mutant (Mathur et al. 2003). The uptake of FM 4-64 in arpc2b mutant was slightly slowed down and these root tips had altered shapes of cortex cells (Figure 2D). The most remarkable impact was observed in arpc5 (Figure 2E). These mutants had aberrant cell shapes in all layers of the root tip. However, the endocytosis was not inhibited but altered, after 20 minutes of the incubation in FM 4-64 large endocytic vesicles accumulated at the cortical cytoplasm. Similar phenotype was reported for the ARPC5 mutation crooked, which had aberrant cell shape and disbalanced regulation of dense and fine F-actin (Mathur et al. 2003b). The endocytosis in brk1 mutant was slowed down (Figure 2F) when compared with the wild type and after 20 minutes there were not endocytic vesicles in the cytoplasm of the cortex cells. NAP1 gene mutation caused accumulation of the endocytic vesicles at the cortical cytoplasm, more likely at the PM, after min of the FM 4-64 incubation (Figure 2G). In the sra1 the uptake of FM 4-64 was slowed down as well. Any of the mutations in Arp 2/3 complex proteins neither in WAVE activator proteins did not seem to interfere with PIN1 protein basal localization in stele cells of the root tip (Figure 2J-L and 2N-P) comparing with wild type PIN1 staining (Figure 2I), except the arpc5 mutant. In the stele cells of this mutant very strong signal of the PIN1 protein was present at the PM and moreover granulated staining of the PIN1 throughout the cytoplasm of stele cells waas observed (Figure 2M). Data from screening of Arp 2/3 mutants and its activators clearly suggest that these actin nucleation factors are involved in the endocytosis (see Deeks and Hussey, 2005) of diffusely-growing plant cells. The dynamic network of actin filaments is regulated by several ABPs (actin binding proteins). Profilins (PRF) and actin depolymerising factors (ADFs) belong into the group of actin monomer-binding proteins. These proteins regulate the size and activity of the actin subunit pool. ADFs regulate actin dynamics and bind to G-actin as well as to F-actin. Other group of actin monomer-binding proteins are profiling. Profilin makes complex with G-actin preventing spontaneous nucleation. In the case of uncapped, barbed end of actin filament, profilin contributes to filament elongation by delivering monomer actin (Staiger et al. 1997). The screening of mutants in the genes coding for actin monomer-binding proteins involved ADF1, ADF4, ADF5, PRF1 and PRF2 proteins. T-DNA insertion homozygous mutants in ADF1, ADF5, PRF1 and PRF2 genes and T-DNA insertion heterozygous mutant in ADF4 gene were analyzed. Respective mutants were named adf1, adf4, adf5, prf1 and prf2 for the purpose of this screen (the identity of the particular mutant is given in Table 1). In the adf1 60
96 mutants the endocytosis of FM 4-64 was inhibited (Figure 3B) in comparison with ADF4 gene mutation (Figure 3C) and ADF5 gene mutation (Figure 3D), where the process of endocytosis at the times indicated was comparable with the wild type root of the seedling (Figure 4A). This difference could be caused partly due to the fact that adf4 line was heterozygous, but the difference between adf1 and adf5 rather reflect tissue specific expression in the PRF family (Ruzicka et al., 2007). prf1 (Figure 3E) and prf2 (Figure 3F) mutations affected the rate of endocytosis. The PIN1 protein location and distribution within PM of the stele cells of the Arabidopsis seedlings of the adf mutants (Figure 3H-J) and profilin mutants (Figure 3K and 3L) did not show any significant changes in basal PIN1 localization comparing with the wild type PIN1 staining (Figure 3G). 61
 FM 4-64 uptake by wild type cells. (B) Reproductive actin ACT1 gene mutation with no influence on the dye uptake, but vesicles are clustered.")
97 Figure 1: In vivo confocal microscopy of FM 4-64 (2 μm) uptake into the epidermal and cortex cells of the root tip in 3-day-old seedlings of Arabidopsis thaliana mutants in actin genes and immunolocalization of PIN1 protein in these mutants. (A) FM 4-64 uptake by wild type cells. (B) Reproductive actin ACT1 gene mutation with no influence on the dye uptake, but vesicles are clustered. (C) Vegetative actin ACT2 gene mutation with slower uptake of FM 4-64 related to time of incubation. (D) Vegetative actin ACT7 gene mutation with the inhibition of the endocytosis, the most remarkable in the more inner tissues of cortex cells. Note that the epidermal as well as cortex cells of act7 mutant are swollen in contrast to wild type cells (A). (E) Reproductive actin ACT11 gene mutation with the inhibition of the FM 4-64 uptake by the root cells, after 20 min treatment FM 4-64 vesicles are accumulated at the outer side of epidermal cells. Cells are swollen. (F-J) The thickness of roots of act7 (I) and of act11 mutant (J) is increased in comparison with wild type (F) or act1 (G) and act2 (H) mutants, staining 5 min with FM (L-O) All actin mutations with no changes in the localization of PIN1 protein at basal plasma membranes of root provasculature in comparison with wild type PIN1 staining (G). RC root cap, EP epidermis, CX cortex, ST stele. Scale bars 10 μm. Scale bars (F-J) 100 μm. 62
 FM 4-64 uptake by wild type cells. (B) ARP2 gene mutation with inhibited endocytosis of FM 4-64 dye.")
98 Figure 2 In vivo confocal microscopy of FM 4-64 (2 μm) uptake into the epidermal and cortex cells of the root tip in 3-day-old seedlings of Arabidopsis thaliana mutants in Arp 2/3 complex and WAVE complex genes and immunolocalization of PIN1 protein in these mutants. (A) FM 4-64 uptake by wild type cells. (B) ARP2 gene mutation with inhibited endocytosis of FM 4-64 dye. (C) Disturbed FM 4-64 endocytosis in arp3 mutant, distorted shape of epidermal cells. (D) arpc2b mutant with slowed-down uptake of FM 4-64 and altered cortex cell layer. (E) arpc5 mutation with aberrant cell shaping along whole root tip and the accumulations of the endocytic vesicles at the cortical cytoplasm after 20 min treatment. (F) Delayed FM4-64 endocytosis in brk1 mutant. (F) NAP1 gene mutation with the accumulation of the endocytic vesicles in the cortical cytoplasm-pm interface after min of the FM 4-64 incubation. (G) sra1 mutant with slowed-down uptake of FM 4-64 in time dependent manner comparing with wild type. All mutations exceptarpc5 did not seem to interfere with PIN1 protein basal localization in stele cells (I-P and M) when compared with wild type PIN1 staining (I). arpc5 stele provasculature cells had very strong signal of the PIN1 protein at the PM and granulated staining of the PIN1 throughout the cytoplasm (M). RC root cap, EP epidermis, CX cortex, ST stele. Scale bars10 μm. 63
 FM 4-64 uptake by wild type cells. (B) adf1 mutant with inhibited endocytosis of FM4-64.")
99 Figure 3 In vivo confocal microscopy of FM 4-64 (2 μm) uptake into the epidermal and cortex cells of the root tip in 3-day-old seedlings of Arabidopsis thaliana mutants in ADF and PRF genes and immunolocalization of PIN1 protein in these mutants. (A) FM 4-64 uptake by wild type cells. (B) adf1 mutant with inhibited endocytosis of FM4-64. (C) adf4 and (D) adf5 with no inhibition of FM 4-64 uptake. (E) prf1 mutant and (F) prf2 mutant with inhibited endocytosis. The adf1, adf4 and adf5 mutations (H-J) as well as prf1 and prf2 mutants (K, L) with PIN1 localized on the basal side of the PM with no changes in comparison with control cells (H). RC root cap, EP epidermis, CX cortex, ST stele. Scale bars 10 μm. 64
.")
100 From our data can be concluded, that all tested homozygous lines of actin and its associated proteins had reduced rate of FM 4-64 endocytosis. Simultaneously, there were observed changes in vesicle dynamics and appearance (Figure 4). The phenomena observed in the screen for FM 4-64 uptake by root epidermal and cortex cells could be classified in several categories that could serve for the comparisons during the continuation of this screen. As shown in Figure 4 these categories were observed: 1) Reduced quantity of vesicles in the endodermis and the cortex cells (Figure 4B) 2) Inhibition of vesicle endoctosis from the outer side of epidermis and accumulation of vesicles in the cortical cytoplasm (Figure 4C, D) 3) Inhibition of vesicle endocytosis from the PM and accumulation of the vesicles at the PM of the epidermal and cortex cells (Figure 4E, G) 4) different types of vesicles and vesicle aggregations (Figure 4F). Wild type random vesicles Inhibited formation of vesicles Cytoplasmic clusters of vesicles One-side aggregation Cortical cytoplasm vesicles Tubular vesicles PM-attached vesicles Figure 4 The phenotypes of the vesicles and vesicle formations of FM 4-64 endocytic dye (after 20 min incubation) in the endodermis cell layer of the roots of Arabidopsis thaliana mutants (A) Wild type random vesicles. (B) Inhibited formation of vesicles as observed in act2, act7, arp2, arp3, sra1, prf2 mutants (C) act1 cytoplasmic clusters. (D) One-side aggregates of act11 mutant (E) Cortical cytoplasm vesicles of arpc5 mutant (F) Tubular vesicles observed in brk1 mutant (G) PM-attached vesicles observed in nap1 and prf1 mutant. Scale bars 5 μm. The localization of PIN1 in the root tip stele cells scored by indirect immunofluorescence did not show any significant changes within mutants screened here. The only visible effect was seen in the arpc5 mutant, where it seems that there was more PIN protein present at the basal side of the PM as well as throughout the cytoplasm (Figure 3E). However, in this mutant there was affected cell shaping of all layers of the root (Figure 3M) so the inhibition seems to be more complex. Since arpc5 mutation led to the preferential accumulation of FM 4-64 vesicles in the cortical cytoplasm, it could be speculated that this effct is connected with the massive abundance of PIN1 in this region. 65
101 Results presented here point to the fact that actin might not play a direct role in the polar localization and docking of the PIN1 protein within the PM of stele cells of the Arabidopsis cell root tip. However, actin and specially its dynamics (the most impact had mutations of Arp 2/3 complex and its subunits) seem to be essential for vector trafficking of the endocytic vesicles downstream from the PM internalization. The defects in cell trafficking lead to aberrations in cell polarity and shaping as seen in some of the mutants. To further understand the participation of cytoskeletal proteins in polar PIN1 targeting the screen should be finished with other actin-associated proteins, tubulins and its associated proteins. References Aniento F, Robinson D (2005) Testing for endocytosis in plants. Protoplasma 226: 3-11 Boutté Y, Crosnier M, Carraro N, Traas J, Satiat-Jeunemaitre B (2006) The plasma membrane recycling pathway and cell polarity in plants: studies on PIN proteins. J Cell Sci 119: Boutté Y, Ikeda Y, Grebe M (2007) Mechanisms of auxin-dependent cell and tissue polarity. Curr Opin Plant Biol 10: Buschmann H, Lloyd C (2008) Arabidopsis mutants and the network of microtubule-associated functions. Mol Plant 1: Butler J, Hu S, Brady S, Dixon M, Muday G (1998) In vitro and in vivo evidence for actin association of the naphthylphthalamic acid-binding protein from zucchini hypocotyls. Plant J 13: Carlier M, Laurent V, Santolini J, Melki R, Didry D, Xia G, Hong Y, Chua N, Pantaloni D (1997) Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: implication in actin-based motility. J Cell Biol 136: Christensen H, Ramachandran S, Tan C, Surana U, Dong C, Chua N (1996) Arabidopsis profilins are functionally similar to yeast profilins: identification of a vascular bundle-specific profilin and a pollen-specific profilin. Plant J 10: Deeks M, Cvrčková F, Macheský L, Mikitová V, Ketelaar T, Ţárský V, Davies B, Hussey P (2005) Arabidopsis group Ie formins localize to specific cell membrane domains, interact with actinbinding proteins and cause defects in cell expansion upon aberrant expression. New Phytol 168: Deeks M, Kaloriti D, Davies B, Malhó R, Hussey P (2004) Arabidopsis NAP1 isessential for Arp2/3-dependent trichome morphogenesis. Curr Biol 14: Deeks M, Hussey P (2005) Arp2/3 and scar: Plants move to the fore. NATURE REVIEWS MOLECULAR CELL BIOLOGY: Dhonukshe P, Grigoriev I, Fischer R, Tominaga M, Robinson D, Hašek J, Paciorek T, Petrášek J, Seifertová D, Tejos R, Meisel L, Zaţímalová E, Gadella TJ, Stierhof Y, Ueda T, Oiwa K, Akhmanova A, Brock R, Spang A, Friml J (2008) Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc Natl Acad Sci U S A 105: Djakovic S, Dyachok J, Burke M, Frank M, Smith L (2006) BRICK1/HSPC300 functions with SCAR and the ARP2/3 complex to regulate epidermal cell shape in Arabidopsis. Development 133: Dong C, Kost B, Xia G, Chua N (2001) Molecular identification and characterization of the Arabidopsis 66
102 AtADF1, AtADFS and AtADF6 genes. Plant Mol Biol 45: Friml J, Palme K (2002) Polar auxin transport--old questions and new concepts? Plant Mol Biol 49: Fu Y, Gu Y, Zheng Z, Wasteneys G, Yang Z (2005) Arabidopsis interdigitating cell growth requires two antagonistic pathways with opposing action on cell morphogenesis. Cell 120: Geldner N (2009) Cell polarity in plants: a PARspective on PINs. Curr Opin Plant Biol 12: Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P, Delbarre A, Ueda T, Nakano A, Jürgens G (2003) The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 112: Gälweiler L, Guan C, Müller A, Wisman E, Mendgen K, Yephremov A, Palme K (1998) Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282: Gilliland LU, Pawloski LC, Kandasamy MK, Meagher RB (2003) Arabidopsis actin gene ACT7 plays an essential role in germination and root growth. Plant J 33: Higaki T, Sano T, Hasezawa S (2007) Actin microfilament dynamics and actin side-binding proteins in plants. Curr Opin Plant Biol 10: Holweg C (2007) Acto-Myosin motorises the flow of auxin. Plant Signal Behav 2: Holweg C, Nick P (2004) Arabidopsis myosin XI mutant is defective in organelle movement and polar auxin transport. Proc Natl Acad Sci U S A 101: Huang S, An Y, McDowell J, McKinney E, Meagher R (1997) The Arabidopsis ACT11 actin gene is strongly expressed in tissues of the emerging inflorescence, pollen, and developing ovules. Plant Mol Biol 33: Huang S, McDowell J, Weise M, Meagher R (1996) The Arabidopsis profilin gene family. Evidence for an ancient split between constitutive and pollen-specific profilin genes. Plant Physiol 111: Hülskamp M, Misŕa S, Jürgens G (1994) Genetic dissection of trichome cell development in Arabidopsis. Cell 76: Jelínková A, Malínska K, Simon S, Kleine- Vehn J, Pařezová M, Pejchar P, Kubeš M, Martinec J, Friml J, Zaţímalová E, Petrášek J (2010) Probing plant membranes with FM dyes: tracking, dragging or blocking? Plant Journal 61: Kandasamy MK, Gilliland LU, McKinney EC, Meagher RB (2001) One plant actin isovariant, ACT7, is induced by auxin and required for normal callus formation. Plant Cell 13: Kleine-Vehn J, Dhonukshe P, Sauer M, Brewer P, Wiśniewská J, Paciorek T, Benková E, Friml J (2008b) ARF GEFdependent transcytosis and polar delivery of PIN auxin carriers in Arabidopsis. Curr Biol 18: Kleine-Vehn J, Dhonukshe P, Swarup R, Bennett M, Friml J (2006) Subcellular trafficking of the Arabidopsis auxin influx carrier AUX1 uses a novel pathway distinct from PIN1. Plant Cell 18: Kleine-Vehn J, Langowski L, Wisniewská J, Dhonukshe P, Brewer P, Friml J (2008a) Cellular and molecular requirements for polar PIN targeting and transcytosis in plants. Mol Plant 1: Kopczak S, Haas N, Hussey P, Silflow C, Snustad D (1992) The small genome of Arabidopsis contains at least six expressed alpha-tubulin genes. Plant Cell 4: Li S, Blanchoin L, Yang Z, Lord E (2003) The putative Arabidopsis arp2/3 complex controls leaf cell morphogenesis. Plant Physiol 132: Ludwig S, Oppenheimer D, Silflow C, Snustad D (1987) Characterization of 67
103 the alpha-tubulin gene family of Arabidopsis thaliana. Proc Natl Acad Sci U S A 84: Ludwig S, Oppenheimer D, Silflow C, Snustad D (1987) Characterization of the alpha-tubulin gene family of Arabidopsis thaliana. Proc Natl Acad Sci U S A 84: Maisch J, Nick P (2007) Actin is involved in auxin-dependent patterning. Plant Physiol 143: Mathur J, Mathur N, Kernebeck B, Hülskamp M (2003a) Mutations in actin-related proteins 2 and 3 affect cell shape development in Arabidopsis. Plant Cell 15: Mathur J, Mathur N, Kirik V, Kernebeck B, Srinivas B, Hülskamp M (2003b) Arabidopsis CROOKED encodes for the smallest subunit of the ARP2/3 complex and controls cell shape by region specific fine F-actin formation. Development 130: McDowell J, An Y, Huang S, McKinney E, Meagher R (1996) The arabidopsis ACT7 actin gene is expressed in rapidly developing tissues and responds to several external stimuli. Plant Physiol 111: McKinney E, Ali N, Traut A, Feldmann K, Belostotsky D, McDowell J, Meagher R (1995) Sequence-based identification of T-DNA insertion mutations in Arabidopsis: actin mutants act2-1 and act4-1. Plant J 8: Nairn C, Winesett L, Ferl R (1988) Nucleotide sequence of an actin gene from Arabidopsis thaliana. Gene 65: Nick P, Han M, An G (2009) Auxin stimulates its own transport by shaping actin filaments. Plant Physiol 151: Oppenheimer D, Haas N, Silflow C, Snustad D (1988) The beta-tubulin gene family of Arabidopsis thaliana: preferential accumulation of the beta 1 transcript in roots. Gene 63: Paciorek T, Zaţímalová E, Ruthardt N, Petrášek J, Stierhof Y, Kleine-Vehn J, Morris D, Emans N, Jürgens G, Geldner N, Friml J (2005) Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 435: Petrášek J, Černá A, Schwartzerová K, Elčkner M, Morris DA, Zaţímalová E (2003) Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiology 131: Petrášek J, Friml J (2009) Auxin transport routes in plant development. Development 136: Petrášek J, Mravec J, Bouchard R, Blakeslee J, Abas M, Seifertová D, Wisniewská J, Tadele Z, Kubeš M, Čovanová M, Dhonukshe P, Skůpa P, Benková E, Perry L, Kreček P, Lee O, Fink G, Geisler M, Murphy A, Luschnig C, Zaţímalová E, Friml J (2006) PIN proteins perform a ratelimiting function in cellular auxin efflux. Science 312: Ruzicka D, Kandasamy M, McKinney E, Burgos-Rivera B, Meagher R (2007) The ancient subclasses of Arabidopsis Actin Depolymerizing Factor genes exhibit novel and differential expression. Plant J 52: Settleman J (2005) Intercalating Arabidopsis leaf cells: a jigsaw puzzle of lobes, necks, ROPs, and RICs. Cell 120: Snustad D, Haas N, Kopczak S, Silflow C (1992) The small genome of Arabidopsis contains at least nine expressed beta-tubulin genes. Plant Cell 4: Steinmann T, Geldner N, Grebe M, Mangold S, Jackson C, Paris S, Gälweiler L, Palme K, Jürgens G (1999) Coordinated polar localization of auxin efflux carrier PIN1 by GNOM ARF GEF. Science 286: Xu J, Scheres B (2005) Cell polarity: ROPing the ends together. Curr Opin Plant Biol 8: Yang Z (2008) Cell polarity signalling in Arabidopsis. Annu Rev Cell Dev Biol 24: Zimmermann I, Saedler R, Mutondo M, 68
104 Hulskamp M (2004) The Arabidopsis GNARLED gene encodes the NAP125 homolog and controls several actinbased cell shape changes. Mol Genet Genomics 272:
105 70
106 4 Materials and Methods Plant material and gene constructs Tobacco BY-2 cell line (N. tabacum L. cv. Bright Yellow 2; Nagata et al. 1992) and BY- 2 cells transformed with Arabidopsis thaliana PIN1::PIN1:GFP (Benková et al., 2003; Petrášek and Zaţímalová, 2006) were cultured in darkness at 27 C on an orbital incubator (IKA KS501, IKA Labortechnik, at 120 rpm (orbital diameter 30 mm) in liquid medium (3% sucrose, 4.3 g l -1 Murashige and Skoog salts, 100 mg l -1 inositol, 1 mg l -1 thiamin, 0.2 mg l -1 2,4-dichlorophenoxyacetic acid and 200 mg l -1 KH 2 PO 4, ph 5.8) supplemented for transformed cells with 100 mg l -1 kanamycin and 100 mg l -1 cefotaxim, and sub-cultured weekly. Plants were grown on vertically-oriented agar plates containing Arabidopsis medium (AM, MS/2, 1.5% agar, 1% sucrose, ph 5.8) under a 16/8 h light/dark photoperiod at 21/18 C. Experiments were performed during the light period in 4-day-old seedlings of individual mutants of Arabidopsis thaliana cv. Col-0 obtained from NASC (Nottingham Arabidopsis Stock Centre). Chemicals Styryl dye FM 4-64 (Molecular Probes, catalogue number T13320), BFA, tyrphostin A23, Tyrphostin A51, wortmannin, filipin, dynasore, cytochalasin D, oryzalin and auxins NAA, IAA, 2,4-D (all Sigma-Aldrich) were kept as 20, 50, 50, 50, 10, 20, 80, 10, 57 mm and 10, 20, 10 mm stock solutions, respectively, in DMSO, stored in -20ºC. Application of FM dyes and inhibitors Unless otherwise indicated, the following conditions were used. FM 4-64 (final concentration 2 μm) was added to 1 ml of 2 3-day-old BY-2 cell suspension under continuous shaking in multi-well plates, and samples were observed by confocal microscopy at the times indicated. The inhibitors (tyrphostin A23, Tyrphostin A51, wortmannin, filipin, dynasore, cytochalasin D, oryzalin) and auxins (1-NAA, IAA, 2,4-D) were added directly to the cultivation medium to final concentrations of 50, 50, 33, 15, 80, 20, 15 μm and 10, 5, 5 μm respectively for 30 min pretreatment. For the PIN1 localization experiments transgenic 71
107 BY-2 cell line expressing under native promoter intragenic translational fusion of PIN1 from Arabidopsis thaliana and GFP was used (Petrášek and Zaţímalová, 2006). 30 min pretreatment with above-mentioned inhibitors was followed by 30 min concomitant treatment with with BFA (50 μm) and analyzed by confocal microscopy. The appropriate amount of the solvent (DMSO) was added to controls. Seedlings were primed for 5 min on ice with MS medium containing 2 μm FM 4-64 (Molecular Probes, catalogue n. T13320) and subsequently rinsed three times in liquid MS medium and analysed by confocal microscopy at the times indicated. Immunolocalization of PIN1 protein PIN1 in the root tip was immunolocalized in whole-mounts using automated system for immunocytochemistry (InsituPro VSi, Intavis, Germany). The protocol (InsituPro VS protocol Arabidopsis whole mount 1, Robert and Offringa, Leiden, Netherlands) was optimized. The 4-day-old Arabidopsis seedlings were fixed in 4% paraformaldehyde in MTSB (50 mm PIPES, 5 mm EGTA, 5 mm MgSO 4, ph 7 adjusted with KOH) for 1 h under the vacuum. Samples were washed with MTSB with 0.1% Triton (5 10 min) and with deionised water (5 10 min). Cell wall was then digested with 0.05% pectolyase in MTSB with mannitol (0.45M) for min and samples washed with MTSB with 0.1% Triton (5 10 min). Incubation with 10% DMSO and 3% NP-40 in MTSB for 1 h followed. After another washing in MTSB with 0.1% Triton (5 10 min) seedlings were pre-incubated in 2% BSA in MTSB (1h, 37 C) and incubated with the epitope-specific primary monoclonal antibody against PIN1 (PIN1 (ak-13): sc-27161, Santa Cruz Biotechnology) in 3% BSA in MTSB (5 h, 37 C). After extensive washing with MTSB with 0.1% Triton (8 10 min), the seedlings were incubated with the secondary antibody (Alexa 488, Molecular Probes, catalogue n. A-11059) in 3% BSA in MTSB for another 3h (37 C). Finally, the samples were washed with MTSB with 0.1% Triton (5 10 min) and with deionised water (5 10 min) and and mounted into 50% glycerol with 0.02% sodium azide. Microscopy For in vivo microscopy, a Zeiss LSM 5 DUO confocal microscope with a 40 C- Apochromat objective (NA = 1.2 W) was used ( Fluorescence signals for GFP (excitation 488 nm, emission nm), FM 4-64 and FM 5-59 (both excitation 561 nm, emission >575 nm) were detected. Sequential scanning was used to avoid any 72
108 crosstalk between fluorescence channels. To separate FM1-43 and GFP fluorescence, spectral fluorescence detection and subsequent linear unmixing were used. Lambda series (excitation 488 and 561 nm) were collected using the Zeiss META system. Nineteen channels (10.7 nm wide) of the META detector were used to span an emission range of nm. Linear unmixing was performed using Zeiss LSM IMAGE EXAMINER software. Reference spectra of GFP and FM1-43 were acquired from single labelled specimens. The scattergram of fluorescence intensities in the red and green channels was constructed using Zeiss IMAGE EXAMINER software. 73
109 74
110 5A Conclusions and prospects The main objective of the work presented in this thesis was to describe the role of the cytoskeleton and vesicle trafficking in the process of auxin efflux carrier targeting to the site of its action, i.e. to the plasma membrane. The connection between auxin transport, vesicular (protein) trafficking and actin cytoskeleton was uncovered. In our previous work, the inhibitor of anterograde transport pathways brefeldin A (BFA) inhibited auxin efflux and caused actin filaments remodelling in BY-2 tobacco suspension cells. Therefore, this remodelling raised various questions how the state of actin and auxin transport is connected and whether actin and endomembrane network co-operation is essential for auxin efflux carrier trafficking and thus thier function. Results presented in this thesis were obtained from both model cell line of tobacco BY-2 cells and roots of Arabidopsis thaliana seedlings. Using the pharmacological approach and in vivo observation of endocytosis and PIN1 localization in tobacco BY-2 cells it was suggested for the first time that clathrin-mediated endocytosis of FM 4-64 is actin- as well as tubulin-dependent. The BFA-induced remodelling of actin filaments, observed in our previous work, was suggested to reflect some kind of active process, because pharmacological actin disruption led to the drastic changes in the appearance of BFA-induced compartments. This was not observed with tubulin drugs. Moreover, detailed knowledge on the action of inhibitors of endocytosis, namely tyrphostins, filipin, wortmannin and dynasore was obtained. In the course of experiments with the commonly used in vivo trackers of the endocytosis, FM styryl dyes, unexpected phenomena was detected. Using plasma membrane-localized transporters for the plant hormone auxin in tobacco BY-2 and Arabidopsis thaliana cell suspensions it was shown that routinely used concentrations of FM 4-64 and FM 5-95 trigger transient re-localization of these proteins, and FM 1-43 affects their activity. The active process of re-localization is blocked neither by inhibitors of endocytosis nor by cytoskeletal drugs. It does not occur in Arabidopsis thaliana roots and depends on the degree of hydrophobicity (lipophilicity) of a particular FM dye. These results emphasize the need for circumspection during in vivo studies of membrane proteins performed using simultaneous labelling with FM dyes. To further characterize the cytoskeletal requirements for vesicle trafficking and protein targeting in planta, a phenotypic microscopical screen of all available Arabidopsis thaliana mutants in genes coding for cytoskeletal and their associated proteins was initiated. The obtained mutants were tested for their rate of endocytosis using FM 4-64 and the localization 75
111 of PIN1 and PIN2 in Arabidopsis thaliana root apex. The purpose of this work, which is still in progress, is the identification of new determinants of PIN polar targeting. The data presented here represent the first part of the screen, covering all available actin proteins, Arp 2/3 complex proteins and its subunits, proteins of WAVE activating complex and actin monomer binding proteins. The results presented here point to the idea, that actin might not play a direct role in anchoring of the PIN1 protein within the basal plasma membrane of the stele provascular parenchyma cells. Since the most remarkable impact on the endocytosis and partly also on the localization of PIN1 had mutations of Arp 2/3 complex and its subunits, it seem that actin dynamics is essential for vector trafficking of the endocytic vesicles downstream from the plasma membrane internalization. Various defects in the endocytosis were observed in mutants in actin and its polymerization regulators. To further understand the participation of cytoskeletal proteins in polar PIN1 targeting it is necessary to complete our screen with other actin-associated proteins, tubulin and tubulin-associated proteins. Other mutants and double mutants will be requested from NASC and other research teams. Altogether, results from both BY-2 cells and Arabidopsis roots suggest that actin seems to be not so important for PIN1 polar localization/targeting within the basal side of the plasma membrane. However, intact actin is essential for proper endomembrane trafficking, endocytic vesicle formation and dynamics that make in turn part of the PIN1 protein targeting and cycling. It could be that intact actin might be essential for vesicle trafficking events, while endosomes as sorting organelles might move along microtubules travelling to their respective destinations (plasma membrane, trans-golgi network, and vacuole), similar as it is mammalian cells. A future investigation, including finishing the screen and subsequent usage of endosomal organelle markers, genetic and pharmacological manipulations of actin and tubulin proteins and its associates will solve this puzzle. 76
112 5B Výsledky a závěry Hlavním cílem této disertační práce bylo popsat roli cytoskeletu a vesikulárního transportu v procesech směrovaného ukládání přenašečů auxin ven z buněk do místa jejich působení, tj. do plazmatické membrány. Propojení mezi transportem auxinu, transportem vesikulů (proteinů) a aktinovým cytoskeletem bylo jiţ v minulosti naznačeno. V naší předchozí práci bylo zjištěno, ţe v buňkách tabákové kultury BY-2 inhibitor anterográdního transportu brefeldin A (BFA) zpomaluje tok auxinu z buněk a způsobuje přestavbu aktinových filament. Tento účinek na stav aktinového cytoskeletu vyvolal řadu otázek: Jak souvisí stav aktinu s auxinovým transportem? Zdal-li funkční propojení aktinu s endomembránovým systémem je nezbytné pro cyklování auxinových přenašečů a tím pro jejich funkci. Výsledky prezentované v této disertační práci byly získány s pouţitím modelu buněčné linie tabáku BY-2 a semenáčků rostlin Arabidopsis thaliana. S vyuţitím farmakologického přístupu a in vivo pozorování endocytózy a lokalizace proteinů PIN1 v tabákových buňkách BY-2 bylo poprvé navrţeno, ţe endocytóza barvy FM 4-64 zprostředkovaná klatrinem je závislá jak na aktinové, tak i na mikrotubulární síti. Dále bylo ukázáno, ţe shlukování aktinu stimulované působením BFA pozorované v naší předchozí práci je zřejmě aktivní proces, neboť farmakologické narušení aktinu vedlo k drastickým změnám ve vzhledu shluků stimulovaných BFA. Toto nebylo pozorováno po pouţití inhibitorů mikrotubulární dynamiky. Navíc byly získány podrobné znalosti o působení inhibitorů endocytosy u rostlinných buněk: tyrfostinů, filipinu, wortmanninu a dynasoru. Během experimentů s rutinně uţívanými in vivo FM styrylovými endocytickými barvami byl zjištěn jejich nečekaný účinek. S vyuţitím fluorescenčně značených transportérů auxinu lokalizovaných na plazmatické membráně v tabákových buňkách BY-2 a v buněčné suspenzi Arabidopsis thaliana bylo pozorováno, ţe běţně pouţívané koncentrace FM 4-64 a FM 5-95 způsobují přesun těchto proteinů a ţe FM 1-43 ovlivňuje jejich funkci. Tento aktivní proces re-lokalizace není blokován ani inhibitory endocytózy, ani inhibitory cytoskeletu. Zmíněný efekt nebyl pozorován u buněk kořenů Arabidopsis thaliana a závisí na stupni hydrofobicity příslušného FM barviva. Získané výsledky zdůrazňují potřebu obezřetnosti při studiu značených membránových proteinů in vivo souběţně s pouţitím FM barev. K prohloubení znalosti o účasti cytoskeletu v procesech transportu membránových váčků a směrovaném ukládání proteinů v celistvé rostlině byla zahájena fenotypová studie všech dostupných mutantů v genech kódujících cytoskeletální a asociované proteiny v huseníčku Arabidopsis thaliana. U doposud získaných mutantních rostlin byla testována rychlost 77
113 endocytózy pomocí FM 4-64 a současně byla sledována změna lokalizace proteinu PIN1 v kořenové špičce Arabidopsis thaliana. Cílem této doposud probíhající studie, je identifikace nových faktorů podílejících se na polárním umísťování PIN proteinů. Zde prezentovaná data, která představují její první část, zahrnují všechny nám dostupné mutantní linie v genech kódujících aktiny, v proteinech Arp 2/3 komplexu, jeho podjednotkách a aktivátorech WAVE a v proteinech vázajících se na na monomery aktinu. Získané výsledky naznačují, ţe aktin zřejmě nehraje přímou roli v ukotvení proteinů PIN1 na bazální část plazmatické membrány buněk středního válce kořene. Vzhledem k tomu, ţe největší dopad na endocytózu a částečně také na lokalizaci proteinu PIN1 měly mutace v komplexu Arp 2/3 a jeho podjednotkách, dá se předpokládat ţe je to právě dynamika aktinu, která je nezbytná pro směrovaný transport endomembránových váčků ve směru od plasmatické membrány. Různé poruchy v endocytóze byly pozorovány i u mutantů v aktinu a regulačních faktorech jeho polymerace. Pro ucelenější představu o účasti cytoskeletálních proteinů v polárním směrování PIN proteinů je nutné doplnit tuto studii o další mutanty v aktin-asociovaných proteinech, tubulinech a jejich asociovaných proteinech. Potřebné mutantní rostliny (případně i dvojití mutanti) budou objednány z NASC a získány od jiných výzkumných týmů. Shrneme-li výsledky této disertační práce, lze jak u modelu buněk BY-2 tak i u kořenů Arabidopsis konstatovat, ţe aktinová vlákna pravděpodobně nejsou tak důleţitý pro polární umísťování a lokalizaci PIN1 proteinů na bazální stranu plasmatické membrány. Nicméně, intaktní aktinová dynamika jsou nepostradatelná pro endomembránový transport, tvorbu endocytických váčků a jejich dynamiku, tedy procesy hrající klíčovou úlohu ve směrování a cyklování PIN1 proteinů. Je moţné, ţe neporušená aktinová dynamika je důleţitá pro transport váčků, zatímco endosomy jako organely ve kterých probíhá třídění proteinů se pohybují do míst svého budoucího působení (plazmatická membrána, trans-golgiho aparát a vakuola) podél mikrotubulů, podobně jako je tomu u savčích buněk. Budoucí výzkum zaměřený na dokončení popsané fenotypové studie a následné vyuţití markerů endosomálního systému spolu s genetickými a farmakologickými manipulacemi aktinových a tubulinových proteinů a jejich interaktorů napomůţe k objasnění této problematiky. 78
114 6 References Aloni R, Aloni E, Langhans M, Ullrich C (2006) Role of cytokinin and auxin in shaping root architecture: regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann Bot 97: Aniento F, Robinson D (2005) Testing for endocytosis in plants. Protoplasma 226: 3-11 Ashe H, Briscoe J (2006) The interpretation of morphogen gradients. Development 133: Bartolini F, Gundersen G (2006) Generation of noncentrosomal microtubule arrays. J Cell Sci 119: Baskin T, Bivens N (1995) Stimulation of radial expansion in arabidopsis roots by inhibitors of actomyosin and vesicle secretion but not by various inhibitors of metabolism. Planta 197: Benková E, Ivanchenko M, Friml J, Shishkova S, Dubrovsky J (2009) A morphogenetic trigger: is there an emerging concept in plant developmental biology? Trends Plant Sci 14: Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J (2003) Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115: Bennett M, Marchant A, Green H, May S, Ward S, Millner P, Walker A, Schulz B, Feldmann K (1996) Arabidopsis AUX1 gene: a permease-like regulator of root gravitropism. Science 273: Bennett S, Alvarez J, Bossinger G, Smyth D (1995) Morphogenesis in PINOID mutants of Arabidopsis Thaliana. Plant Journal: Bhalerao R, Bennett M (2003) The case for morphogens in plants. Nat Cell Biol 5: Boevink P, Oparka K, Santa Cruz S, Martin B, Betteridge A, Hawes C (1998) Stacks on tracks: the plant Golgi apparatus traffics on an actin/er network. Plant J 15: Bonifacino J, Rojas R (2006) Retrograde transport from endosomes to the trans- Golgi network. Nat Rev Mol Cell Biol 7: Boutté Y, Crosnier M, Carraro N, Traas J, Satiat-Jeunemaitre B (2006) The plasma membrane recycling pathway and cell polarity in plants: studies on PIN proteins. J Cell Sci 119: Boutté Y, Ikeda Y, Grebe M (2007) Mechanisms of auxin-dependent cell and tissue polarity. Curr Opin Plant Biol 10: Braulke T, Bonifacino J (2009) Sorting of lysosomal proteins. Biochem Biophys Acta 1793: Buschmann H, Lloyd C (2008) Arabidopsis mutants and the network of microtubule-associated functions. Mol Plant 1: Butler J, Hu S, Brady S, Dixon M, Muday G (1998) In vitro and in vivo evidence for actin association of the naphthylphthalamic acid-binding protein from zucchini hypocotyls. Plant J 13: Chen J, Shimomura S, Sitbon F, Sandberg G, Jones A (2001) The role of auxinbinding protein 1 in the expansion of tobacco leaf cells. Plant J 28:
115 Cole R, Fowler J (2006) Polarized growth: maintaining focus on the tip. Curr Opin Plant Biol 9: Crowell E, Bischoff V, Desprez T, Rolland A, Stierhof Y, Schumacher K, Gonneau M, Höfte H, Vernhettes S (2009) Pausing of Golgi bodies on microtubules regulates secretion of cellulose synthase complexes in Arabidopsis. Plant Cell 21: Cvrčková F, Rivero F, Bavlnka B (2004) Evolutionarily conserved modules in actin nucleation: lessons from Dictyostelium discoideum and plants. Review article. Protoplasma 224: Delbarre A, Muller P, Imhoff V, Guern J (1996) Comparison of mechanisms controlling uptake and accumulation of 2,4-dichlorophenoxy acetic acid, naphthalene-1-acetic acid, and indole-3- acetic acid in suspension-cultured tobacco cells. Planta 198: Dhonukshe P, Aniento F, Hwang I, Robinson D, Mravec J, Stierhof Y, Friml J (2007) Clathrin-mediated constitutive endocytosis of PIN auxin efflux carriers in Arabidopsis. Curr Biol 17: Dhonukshe P, Grigoriev I, Fischer R, Tominaga M, Robinson D, Hašek J, Paciorek T, Petrášek J, Seifertová D, Tejos R, Meisel L, Zaţímalová E, Gadella TJ, Stierhof Y, Ueda T, Oiwa K, Akhmanova A, Brock R, Spang A, Friml J (2008) Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc Natl Acad Sci U S A 105: Dhonukshe P, Tanaka H, Goh T, Ebine K, Mähönen A, Prasad K, Blilou I, Geldner N, Xu J, Uemura T, Chory J, Ueda T, Nakano A, Scheres B, Friml J (2008) Generation of cell polarity in plants links endocytosis, auxin distribution and cell fate decisions. Nature 456: Driskell O, Mironov A, Allan V, Woodman P (2007) Dynein is required for receptor sorting and the morphogenesis of early endosomes. Nat Cell Biol 9: Dubrovsky J, Sauer M, Napsucialy- Mendivil S, Ivanchenko M, Friml J, Shishkova S, Celenza J, Benková E (2008) Auxin acts as a local morphogenetic trigger to specify lateral root founder cells. Proc Natl Acad Sci U S A 105: E FD, C ML (1992) Structural and Functional Organization of Tubulin. 43: Eliáš M, Drdová E, Ziak D, Bavlnka B, Hála M, Cvrčková F, Soukupová H, Ţarský V (2003) The exocyst complex in plants. Cell Biol Int 27: Friml J (2003) Auxin transport - shaping the plant. Curr Opin Plant Biol 6: 7-12 Friml J (2010) Subcellular trafficking of PIN auxin efflux carriers in auxin transport. Eur J Cell Biol 89: Friml J, Benková E, Blilou I, Wisniewská J, Hamann T, Ljung K, Woody S, Sandberg G, Scheres B, Jürgens G, Palme K (2002a) AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 108: Friml J, Palme K (2002) Polar auxin transport--old questions and new concepts? Plant Mol Biol 49: Friml J, Vieten A, Sauer M, Weijers D, Schwarz H, Hamann T, Offringa R, Jürgens G (2003b) Efflux-dependent auxin gradients establish the apicalbasal axis of Arabidopsis. Nature 426: Friml J, Yang X, Michniewicz M, Weijers D, Quint A, Tietz O, Benjamins R, 80
116 Ouwerkerk P, Ljung K, Sandberg G, Hooykaas P, Palme K, Offringa R (2004) A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux. Science 306: Fu Y, Gu Y, Zheng Z, Wasteneys G, Yang Z (2005) Arabidopsis interdigitating cell growth requires two antagonistic pathways with opposing action on cell morphogenesis. Cell 120: Galletta B, Cooper J (2009) Actin and endocytosis: mechanisms and phylogeny. Curr Opin Cell Biol 21: Gardiner J, Harper J, Weerakoon N, Collings D, Ritchie S, Gilroy S, Cyr R, Marc J (2001) A 90-kD phospholipase D from tobacco binds to microtubules and the plasma membrane. Plant Cell 13: Geldner N (2009) Cell polarity in plants: a PARspective on PINs. Curr Opin Plant Biol 12: Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P, Delbarre A, Ueda T, Nakano A, Jürgens G (2003) The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 112: Geldner N, Friml J, Stierhof Y, Jürgens G, Palme K (2001) Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413: Giddings T, Staehelin L (1991) Microtubule-mediated control of microfibril deposition: A reexamination of the hypothesis. In C Lloyd, ed, The Cytoskeletal Basis of Plant Growth and Form. Academic Press, London, pp Goldberg R, de Paiva G, Yadegari R (1994) Plant embryogenesis: zygote to seed. Science 266: Grebe M, Xu J, Möbius W, Ueda T, Nakano A, Geuze H, Rook M, Scheres B (2003) Arabidopsis sterol endocytosis involves actin-mediated trafficking via ARA6-positive early endosomes. Curr Biol 13: Grebe M, Xu J, Scheres B (2001) Cell axiality and polarity in plants--adding pieces to the puzzle. Curr Opin Plant Biol 4: Gälweiler L, Guan C, Müller A, Wisman E, Mendgen K, Yephremov A, Palme K (1998) Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282: Hála M, Cole R, Synek L, Drdová E, Pečenková T, Nordheim A, Lamkemeyer T, Madlung J, Hochholdinger F, Fowler J, Ţárský V (2008) An exocyst complex functions in plant cell growth in Arabidopsis and tobacco. Plant Cell 20: Hamada T (2007) Microtubule-associated proteins in higher plants. J Plant Res 120: Hanton S, Renna L, Bortolotti L, Chatre L, Stefano G, Brandizzi F (2005) Diacidic motifs influence the export of transmembrane proteins from the endoplasmic reticulum in plant cells. Plant Cell 17: Hawes C, Osterrieder A, Hummel E, Sparkes I (2008) The plant ER-Golgi interface. Traffic 9: Hay J, Scheller R (1997) SNAREs and NSF in targeted membrane fusion. Curr Opin Cell Biol 9: He B, Guo W (2009) The exocyst complex in polarized exocytosis. Curr Opin Cell Biol 21:
117 Henderson J, Bauly J, Ashford D, Oliver S, Hawes C, Lazarus C, Venis M, Napier R (1997) Retention of maize auxinbinding protein in the endoplasmic reticulum: quantifying escape and the role of auxin. Planta 202: Higaki T, Sano T, Hasezawa S (2007) Actin microfilament dynamics and actin side-binding proteins in plants. Curr Opin Plant Biol 10: Ho A. Y., Day D A., Brown M., Marc Jan (2009) Arabidopsis phospholipase Dδ as an initiator of cytoskeleton-mediated signalling to fundamental cellular processes. Functional Plant Biology 36: Holweg C (2007) Acto-Myosin motorises the flow of auxin. Plant Signal Behav 2: Holweg C, Nick P (2004) Arabidopsis myosin XI mutant is defective in organelle movement and polar auxin transport. Proc Natl Acad Sci U S A 101: Huang S, Gao L, Blanchoin L, Staiger C (2006) Heterodimeric capping protein from Arabidopsis is regulated by phosphatidic acid. Mol Biol Cell 17: Hussey P, Ketelaar T, Deeks M (2006) Control of the actin cytoskeleton in plant cell growth. Annu Rev Plant Biol 57: Hwang I, Robinson D (2009) Transport vesicle formation in plant cells. Curr Opin Plant Biol 12: Hwang J, Gu Y, Lee Y, Yang Z (2005) Oscillatory ROP GTPase activation leads the oscillatory polarized growth of pollen tubes. Mol Biol Cell 16: Islam M, Niwa Y, Takagi S (2009) Lightdependent intracellular positioning of mitochondria in Arabidopsis thaliana mesophyll cells. Plant Cell Physiol 50: Jaillais Y, Fobis-Loisy I, Miège C, Rollin C, Gaude T (2006) AtSNX1 defines an endosome for auxin-carrier trafficking in Arabidopsis. Nature 443: Jelínková A, Malínska K, Simon S, Kleine- Vehn J, Pařezová M, Pejchar P, Kubeš M, Martinec J, Friml J, Zaţímalová E, Petrášek J (2010) Probing plant membranes with FM dyes: tracking, dragging or blocking? Plant Journal 61: Jönsson H, Krupinski P (2010) Modeling plant growth and pattern formation. Curr Opin Plant Biol 13: 5-11 Kakimoto T., Shibaoka H. (1987) Actin filaments and microtubules in the preprophase band and phragmoplast of tobacco cells. Protoplasma 140: Karahara I, Suda J, Tahara H, Yokota E, Shimmen T, Misaki K, Yonemura S, Staehelin L, Mineyuki Y (2009) The preprophase band is a localized center of clathrin-mediated endocytosis in late prophase cells of the onion cotyledon epidermis. Plant J 57: Kieffer M, Neve J, Kepinski S (2010) Defining auxin response contexts in plant development. Curr Opin Plant Biol 13: Kleine-Vehn J, Dhonukshe P, Sauer M, Brewer P, Wiśniewská J, Paciorek T, Benková E, Friml J (2008b) ARF GEFdependent transcytosis and polar delivery of PIN auxin carriers in Arabidopsis. Curr Biol 18: Kleine-Vehn J, Dhonukshe P, Swarup R, Bennett M, Friml J (2006) Subcellular trafficking of the Arabidopsis auxin influx carrier AUX1 uses a novel pathway distinct from PIN1. Plant Cell 18:
118 Kleine-Vehn J, Langowski L, Wisniewská J, Dhonukshe P, Brewer P, Friml J (2008a) Cellular and molecular requirements for polar PIN targeting and transcytosis in plants. Mol Plant 1: Kopczak S, Haas N, Hussey P, Silflow C, Snustad D (1992) The small genome of Arabidopsis contains at least six expressed alpha-tubulin genes. Plant Cell 4: Kramer E (2002) A mathematical model of pattern formation in the vascular cambium of trees. J Theor Biol 216: Kramer E (2006) How far can a molecule of weak acid travel in the apoplast or xylem? Plant Physiol 141: Krecek P, Skupa P, Libus J, Naramoto S, Tejos R, Friml J, Zazímalová E (2009) The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol 10: 249 Kueh H, Mitchison T (2009) Structural plasticity in actin and tubulin polymer dynamics. Science 325: König S, Ischebeck T, Lerche J, Stenzel I, Heilmann I (2008) Salt-stress-induced association of phosphatidylinositol 4,5- bisphosphate with clathrin-coated vesicles in plants. Biochem J 415: Lam S, Siu C, Hillmer S, Jang S, An G, Robinson D, Jiang L (2007) Rice SCAMP1 defines clathrin-coated, transgolgi-located tubular-vesicular structures as an early endosome in tobacco BY-2 cells. Plant Cell 19: Lambert A (1993) Microtubule-organizing centers in higher plants. Curr Opin Cell Biol 5: Leyser O (2009) The control of shoot branching: an example of plant information processing. Plant Cell Environ 32: Li H, Lin Y, Heath RM, Zhu MX, Yang Z (1999) Control of Pollen Tube Tip Growth by a Rop GTPase Dependent Pathway That Leads to Tip-Localized Calcium Influx. Plant Cell 11: Li S, Blanchoin L, Yang Z, Lord E (2003) The putative Arabidopsis arp2/3 complex controls leaf cell morphogenesis. Plant Physiol 132: Lin Y, Wang Y, Zhu J, Yang Z (1996) Localization of a Rho GTPase Implies a Role in Tip Growth and Movement of the Generative Cell in Pollen Tubes. Plant Cell 8: Lokerse A, Weijers D (2009) Auxin enters the matrix--assembly of response machineries for specific outputs. Curr Opin Plant Biol 12: Ludwig S, Oppenheimer D, Silflow C, Snustad D (1987) Characterization of the alpha-tubulin gene family of Arabidopsis thaliana. Proc Natl Acad Sci U S A 84: Maisch J, Nick P (2007) Actin is involved in auxin-dependent patterning. Plant Physiol 143: Mandelkow E, Song Y, Mandelkow E (1995) The microtubule lattice-- dynamic instability of concepts. Trends Cell Biol 5: Marchant A, Kargul J, May S, Muller P, Delbarre A, Perrot-Rechenmann C, Bennett M (1999) AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake within root apical tissues. EMBO J 18: Mathur J, Mathur N, Hülskamp M (2002) Simultaneous visualization of peroxisomes and cytoskeletal elements reveals actin and not microtubule-based 83
119 peroxisome motility in plants. Plant Physiol 128: Maxfield F, McGraw T (2004) Endocytic recycling. Nat Rev Mol Cell Biol 5: Michniewicz M, Zago M, Abas L, Weijers D, Schweighofer A, Meskiene I, Heisler M, Ohno C, Zhang J, Huang F, Schwab R, Weigel D, Meyerowitz E, Luschnig C, Offringa R, Friml J (2007) Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 130: Miki H, Okada Y, Hirokawa N (2005) Analysis of the kinesin superfamily: insights into structure and function. Trends Cell Biol 15: Mravec J, Skůpa P, Bailly A, Hoyerová K, Kreček P, Bielach A, Petrášek J, Zhang J, Gayková V, Stierhof Y, Dobrev P, Schwartzerová K, Rollick J, Seifertová D, Luschnig C, Benková E, Zaţímalová E, Geisler M, Friml J (2009) Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 459: Muday G, DeLong A (2001) Polar auxin transport: controlling where and how much. Trends Plant Sci 6: Müller A, Guan C, Gälweiler L, Tänzler P, Huijser P, Marchant A, Parry G, Bennett M, Wisman E, Palme K (1998) AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J 17: Signaling to The Microtubular Cytoskeleton in Plants. (1998) International Review of Cytology 184: Napier R, David K, Perrot-Rechenmann C (2002) A short history of auxin-binding proteins. Plant Mol Biol 49: Neumann U, Brandizzi F, Hawes C (2003) Protein transport in plant cells: in and out of the Golgi. Ann Bot 92: Nick P, Bergfeld R, Schafer E, Schopfer P (1990) Unilateral reorientation of microtubules at the outer epidermal wall during photo- and gravitropic curvature of maize coleoptiles and sunflower hypocotyls. Planta 181: Nick P, Han M, An G (2009) Auxin stimulates its own transport by shaping actin filaments. Plant Physiol 151: Niemes S, Langhans M, Viotti C, Scheuring D, San Wan Yan M, Jiang L, Hillmer S, Robinson D, Pimpl P (2010) Retromer recycles vacuolar sorting receptors from the trans-golgi network. Plant J 61: Noh B, Murphy A, Spalding E (2001) Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development. Plant Cell 13: Oikawa K, Kasahara M, Kiyosue T, Kagawa T, Suetsugu N, Takahashi F, Kanegae T, Niwa Y, Kadota A, Wada M (2003) Chloroplast unusual positioning1 is essential for proper chloroplast positioning. Plant Cell 15: Okada K, Ueda J, Komaki M, Bell C, Shimura Y (1991) Requirement of the Auxin Polar Transport System in Early Stages of Arabidopsis Floral Bud Formation. Plant Cell 3: Oppenheimer D, Haas N, Silflow C, Snustad D (1988) The beta-tubulin gene family of Arabidopsis thaliana: preferential accumulation of the beta 1 transcript in roots. Gene 63: Paciorek T, Zaţímalová E, Ruthardt N, Petrášek J, Stierhof Y, Kleine-Vehn J, Morris D, Emans N, Jürgens G, Geldner 84
120 N, Friml J (2005) Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 435: Palme K, Gälweiler L (1999) PIN-pointing the molecular basis of auxin transport. Curr Opin Plant Biol 2: Pastuglia M, Azimzadeh J, Goussot M, Camilleri C, Belcram K, Evrard J, Schmit A, Guerche P, Bouchez D (2006) Gamma-tubulin is essential for microtubule organization and development in Arabidopsis. Plant Cell 18: Petrášek J, Černá A, Schwartzerová K, Elčkner M, Morris DA, Zaţímalová E (2003) Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiology 131: Petrášek J, Friml J (2009) Auxin transport routes in plant development. Development 136: Petrášek J, Mravec J, Bouchard R, Blakeslee J, Abas M, Seifertová D, Wisniewská J, Tadele Z, Kubeš M, Čovanová M, Dhonukshe P, Skůpa P, Benková E, Perry L, Kreček P, Lee O, Fink G, Geisler M, Murphy A, Luschnig C, Zaţímalová E, Friml J (2006) PIN proteins perform a ratelimiting function in cellular auxin efflux. Science 312: Petrášek J, Schwartzerová K (2009) Actin and microtubule cytoskeleton interactions. Curr Opin Plant Biol 12: Popoff V, Mardones G, Tenza D, Rojas R, Lamaze C, Bonifacino J, Raposo G, Johannes L (2007) The retromer complex and clathrin define an early endosomal retrograde exit site. J Cell Sci 120: Prokhnevsky A, Peremyslov V, Dolja V (2008) Overlapping functions of the four class XI myosins in Arabidopsis growth, root hair elongation, and organelle motility. Proc Natl Acad Sci U S A 105: Raven J (1975) Transport of indoleacetic acid in plant cells in relation to ph and electrical potential gradients, and its significance for polar IAA transport. New Phytologist 74: Reinhardt D, Pesce E, Stieger P, Mandel T, Baltensperger K, Bennett M, Traas J, Friml J, Kuhlemeier C (2003) Regulation of phyllotaxis by polar auxin transport. Nature 426: Robinson D, Jiang L, Schumacher K (2008) The endosomal system of plants: charting new and familiar territories. Plant Physiol 147: Rubery P, Sheldrake A (1974) Carriermediated auxin transport. Planta 118: S. Sabatini D (1999) An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. In HW Beis, J. Murfett, T. Guilfoyle, J. Malamy, P. Benfey, O. Leyser, N. Bechtold, P. Weisbeek and B. Scheres, ed, Vol 99, Cell, pp Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoyle T, Malamy J, Benfey P, Leyser O, Bechtold N, Weisbeek P, Scheres B (1999) An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell: Sackett D, Bhattacharyya B, Wolff J (1985) Tubulin subunit carboxyl termini determine polymerization efficiency. J Biol Chem 260: Sainsbury F, Collings D, Mackun K, Gardiner J, Harper J, Marc J (2008) Developmental reorientation of transverse cortical microtubules to longitudinal directions: a role for actomyosin-based streaming and partial 85
121 microtubule-membrane detachment. Plant J 56: Sanderfoot A, Raikhel N (1999) The specificity of vesicle trafficking: coat proteins and SNAREs. Plant Cell 11: Sedbrook J, Kaloriti D (2008) Microtubules, MAPs and plant directional cell expansion. Trends Plant Sci 13: Settleman J (2005) Intercalating Arabidopsis leaf cells: a jigsaw puzzle of lobes, necks, ROPs, and RICs. Cell 120: Shimmen T (2007) The sliding theory of cytoplasmic streaming: fifty years of progress. J Plant Res 120: Shimmen T, Yokota E (2004) Cytoplasmic streaming in plants. Curr Opin Cell Biol 16: Snustad D, Haas N, Kopczak S, Silflow C (1992) The small genome of Arabidopsis contains at least nine expressed beta-tubulin genes. Plant Cell 4: Sparkes I, Frigerio L, Tolley N, Hawes C (2009) The plant endoplasmic reticulum: a cell-wide web. Biochem J 423: Sparkes I, Teanby N, Hawes C (2008) Truncated myosin XI tail fusions inhibit peroxisome, Golgi, and mitochondrial movement in tobacco leaf epidermal cells: a genetic tool for the next generation. J Exp Bot 59: Staiger C, Blanchoin L (2006) Actin dynamics: old friends with new stories. Curr Opin Plant Biol 9: Staiger C, Poulter N, Henty J, Franklin- Tong V, Blanchoin L (2010) Regulation of actin dynamics by actin-binding proteins in pollen. J Exp Bot 61: Steinmann T, Geldner N, Grebe M, Mangold S, Jackson C, Paris S, Gälweiler L, Palme K, Jürgens G (1999) Coordinated polar localization of auxin efflux carrier PIN1 by GNOM ARF GEF. Science 286: Synek L, Schlager N, Eliáš M, Quentin M, Hauser M, Ţárský V (2006) AtEXO70A1, a member of a family of putative exocyst subunits specifically expanded in land plants, is important for polar growth and plant development. Plant J 48: Szymanski D (2009) Plant cells taking shape: new insights into cytoplasmic control. Curr Opin Plant Biol 12: Tanaka H, Dhonukshe P, Brewer P, Friml J (2006) Spatiotemporal asymmetric auxin distribution: a means to coordinate plant development. Cell Mol Life Sci 63: Teale W, Paponov I, Palme K (2006) Auxin in action: signalling, transport and the control of plant growth and development. Nat Rev Mol Cell Biol 7: Thomas C, Tholl S, Moes D, Dieterle M, Papuga J, Moreau F, Steinmetz A (2009) Actin bundling in plants. Cell Motil Cytoskeleton 66: Titapiwatanakun B, Blakeslee J, Bandyopadhyay A, Yang H, Mravec J, Sauer M, Cheng Y, Adamec J, Nagashima A, Geisler M, Sakai T, Friml J, Peer W, Murphy A (2009) ABCB19/PGP19 stabilises PIN1 in membrane microdomains in Arabidopsis. Plant J 57: Uggla C, Moritz T, Sandberg G, Sundberg B (1996) Auxin as a positional signal in pattern formation in plants. Proc Natl Acad Sci U S A 93:
122 Venis M, Napier R (1992) Plant hormone receptors: past, present and future. Biochem Soc Trans 20: Wasteneys G, Ambrose J (2009) Spatial organization of plant cortical microtubules: close encounters of the 2D kind. Trends Cell Biol 19: Wolpert L (1969) Positional information and the spatial pattern of cellular differentiation. J Theor Biol 25: 1-47 Wolpert L (1998) Pattern formation in epithelial development: the vertebrate limb and feather bud spacing. Philos Trans R Soc Lond B Biol Sci 353: Xu J, Scheres B (2005) Cell polarity: ROPing the ends together. Curr Opin Plant Biol 8: Yang Z (2008) Cell polarity signalling in Arabidopsis. Annu Rev Cell Dev Biol 24: Zhang J, Nodzynski T, Penčík A, Rolčík J, Friml J (2010) PIN phosphorylation is sufficient to mediate PIN polarity and direct auxin transport. Proc Natl Acad Sci U S A 107: Zaţímalová E, Murphy A, Yang H, Hoyerová K, Hošek P (2010) Auxin transporters--why so many? Cold Spring Harb Perspect Biol 2: a Zaţmalová E, Kreček P, Skůpa P, Hoyerová K, Petrášek J (2007) Polar transport of the plant hormone auxin - the role of PIN- FORMED (PIN) proteins. Cell Mol Life Sci 64:
123 88
124 Addenda Following two publications were done with my participation during my stay in CIB CSIC, Department of Biology, Madrid Spain.
125 Z-DNA, a new in situ marker for transcription ˇ ORIGINAL PAPER A. Cerná, 1 A. Cuadrado, 2 N. Jouve, S.M. Díaz de la Espina, C. De la Torre Centro de Investigaciones Biológicas, CSIC, Madrid, Spain; 1 Institute of Experimental Botany, ASCR, Prague, Czech Republic and 2 Departamento de Biología Celular y Genética, Universidad de Alcalá de Henares, Madrid, Spain 2004, European Journal of Histochemistry Z-DNA forms transiently behind the active RNA polymerases, because of the mechanical torsional stress produced during transcription. In this paper, we explore the possibility that the distribution of Z-DNA stretches signals the sites related to nuclear transcription. To localize transcription, the in situ assay for active RNA polymerases, that allows the elongation of the already initiated transcripts but no initiation of new ones (runon experiments), was carried out in isolated nuclei of Allium cepa L. root meristems. Both nucleolar and non-nucleolar sites appeared labelled. Nucleoli were most active in transcription than the multiple non-nucleolar foci altogether. In situ immunodetection of Z-DNA provided images that were comparable to those obtained after the run-on assay, with one exception: while Z-DNA and transcription sites were scattered throughout the whole nucleus, Z-DNA also accumulated in the nuclear periphery, where no transcription foci were detected in the run-on assays. The peripheral Z-conformation signals might correspond to dsrna segments present in the premrna in the process of their export to cytoplasm. The Z-containing structures nearly disappeared when non-nucleolar RNA polymerase II-dependent transcription had been previously abolished by the adenosine analogue DRB (5,6- dichloro-1-β-d-ribofuranosylbenzimidazole). This inhibition selectively decreased the amount of all nucleoplasmic Z structures. On the other hand, the inhibition of the nucleolar RNA polymerase I by cordycepin (3 -deoxyadenosine) prevented the presence of Z-DNA in nucleoli. We propose to use the in situ immunodetection of Z-DNA as a marker of the transcription level in both nucleolus and non-peripheral nucleoplasmic regions of nuclei. Co-detection of Z-DNA and of intermediate filament (IF) proteins, the major components of the nuclear matrix, was also carried out. The IFA antibody recognises a conserved epitope essential for dimerization of the multiple IF proteins. They co-localized with most nucleolar Z-DNA, but not with the nucleoplasmic ones. In the nuclear periphery, the Z- positive signals were adjacent to the IF proteins constituting the lamina, though both signals did not often co-localize. Key words: Allium cepa, Z-DNA immunodetection, in situ run-on transcription, FISH of rdna, nucleoli, intermediate filaments. Correspondence: Consuelo de la Torre, CIB-CSIC Ramiro de Maeztu 9. E Madrid, Spain. delatorrec@cib.csic.es Paper accepted January 8, 2004 European Journal of Histochemistry 2004; vol. 48:49-56 Z-DNA is a configuration alternative to the normal B-one. It forms behind an active RNA polymerase because of the topological stress produced during transcription (Wittig et al. 1989). It can also be produced in the local regions of short complementary base-pairing of the otherwise single-stranded DNA (Herbert and Rich, 2001). In the Z-DNA, the helix axis zig-zags back and forth, hence its name, while the base pairs are located nearly perpendicular to it. Z-DNA represents a more elongated left-handed helix than the right handed B-configuration, with the phosphate groups closer than in the B-form. Z-DNA is a moving and transient target. Alternating purine-pyrimidine sequences favours and stabilises such a conformation, GpC repeats being more efficient than the CpG ones (Rich and Zhang, 2003).The left-handed DNA structure is stabilizedd by DNA negative supercoiling (Peck et al., 1982), and by cations such as Ca 2+, spermine and spermidine, and cytosine methylation at C5, but not at C8 (Kawai et al., 1996), as well as by a high salt environment (Behe and Felsenfeld, 1981). More than 10 years ago it was shown that Z-DNA was preferentially formed near the 5 ends of genes, i.e. near the site of transcription initiation (Schroth et al., 1992). Moreover, the potential Z-DNA sequences are distributed throughout the whole genome in a specific pattern that can be followed both in interphase nuclei and in metaphase chromosomes. In the insect polytene chromosomes, they are more abundant in the heterochromatic regions and in telomeres, i.e. in the untranscribed DNA regulatory sequences (Arndt-Jovin et al., 1985), even when these sequences are mostly devoid from any active role in promoting transcription. Two main proteins bind to Z-DNA. The first of them, WP900 is an enantiomer of daunorubicin that actually favours the conversion of B- into Z- DNA. The second protein is the adenosine deaminase (ADAR) enzyme that edits RNA. ADAR 49
126 A. Cernà ˇ et al. anchors to Z-DNA to slide along newly synthesized RNA. ADAR produces small changes in the spontaneously formed double-stranded regions of premrna, deaminating adenosine to give rise to inosine, while uracil is replaced by cytosine. When inosine is read as guanine in the ribosomes, modified proteins can appear after the editing process (Herbert and Rich, 1991). RNA editing can also create or destroy protein splice sites (Herbert, 1996). Z-DNA is involved in the cellular defence against pox viruses attach (Rich and Zhang, 2003). In this paper we explore the possibility that the distribution of Z-DNA stretches in the nucleus may correspond to transcription-related sites. For this, we have carried out immunodetection of Z-DNA in the nuclei of the plant proliferating cells of Allium cepa root meristems. The assays were performed both in control conditions as well as after preferential inhibition of non-nucleolar transcription by DRB, a treatment that secondarily unravels the rdna in the nucleolar organizer region (Le Panse et al. 1999). Immunodetection was also carried out under preferential inhibition of the RNA polymerase I-dependent nucleolar transcription, by using cordycepin (3 -deoxyadenosine). This drug also collapses without blocking the (non-nucleolar) transcription going on in the Balbiani rings of insect polytene chromosomes (Díez et al., 1973). The set of assays here used proves that the in situ evaluation of Z-DNA is a reliable marker for ongoing transcription sites, where multiple active RNA polymerases are clustered (Martin and Pombo, 2003). It discerns the transcription sites both in nucleolus and nucleoplasm. The data suggest that the antibody also detect Z-RNA in local regions of double-stranded RNA in pre-mrna that becomes adjacent to the internal face of nuclear envelope while waiting for export to cytoplasm. Materials and Methods Isolation of nuclei Root meristem cells from Allium cepa L. bulbs were grown in filtered tap water at room temperature. Isolation of nuclei was performed as previously described (Mínguez and Moreno, 1993). Drug treatment Two day-old roots growing from bulbs were treated either with DRB (50 mm, 6h) or with cordycepin (10 4 M, 4h) and then extensively washed in running tap water before being used for the experiments. Primary antibodies As primary antibodies we have used the monoclonal anti-brdurd antibody (from Sigma) and the sheep polyclonal antibody to Z-DNA (AbCam, Cambridge, UK). Secondary antibodies The following secondary antibodies were used: goat anti-rabbit polyclonal to sheep IgG FITC-conjugated antibody (Abcam, dilution 1:200) (wavelength: 488/518 nm) was used. In the case of the pta71 probe from whole 45S pre-ribosomal RNA, it was directly labelled with TRITC falloidin (wavelength: 514/610 nm) or goat anti-mouse secondary antibodies from Molecular Probes: Alexa 488 (wavelength: 495/519 nm) and Alexa 546 (wavelength: 556/573 nm). Run-on in situ assay for active RNA polymerases Experiments of Br-UTP incorporation were carried out in isolated onion root meristematic cell nuclei. We followed the protocol published by Wansink et al. (1993), modificated by De Cárcer and Medina (1999). Freshly isolated nuclei were washed in glycerol buffer (25% glycerol, 20 mm Tris/HCl, ph 7.5, 5 mm MgCl 2, 0.5 mm EGTA, 25 units RNasin (Plus RNase inhibitor, Promega) for 5 min at 4 C. Then they were permeabilized in the same buffer, which was supplemented with 1% Nonidet-P40, for 5 min at 4 C and washed again in washing buffer for 3 min at 4 C. In these steps, centrifugation was at 1000g for 10 min at 4 C. Incubation was performed in the transcription solution (25% glycerol, 100 mm KCl, 50 mm Tris/HCl, ph 7.5, 5 mm MgCl2, 0.5 mm EGTA), supplemented with 0.5 mm ATP, 0.5 mm CTP, 0.5 mm GTP, 0.5 mm Br-UTP, 25 units/ml RNasin, 100 µg/ml trna and 0.1 mm PMSF, for 30 min at 37 C. The reaction was stopped by plunging the tube into ice and adding cold (0 C) paraformaldehyde up to a 4% final concentration. Nuclei were fixed under these conditions for 30 min. Finally, they were repeatedly washed in washing buffer. Sterile materials were used throughout the procedure. For light microscope observation, nuclei were dropped onto round, polylysine-coated coverslips. Br-UTP was revealed by immunofluorescence with the undiluted monoclonal antibody anti-brudr (Sigma). 50
127 Original Paper Fluorescence in situ hybridization The isolated nuclei were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS: 1.4 mm H2KPO4, 8 mm Na 2 HPO 4, 2.7 mm KCl, 3.7 mm NaCl, ph 7.4) for 30 min, then extensively washed in 0.05% Tween in PBS, followed by 10 min permeabilization with 1% Triton-X in PBS (all at 4 C). After 10 min washing with 0.05% Tween in PBS, the nuclei were incubated with the hybridization solution (15 µl of 100% formamide, 6 µl of 50% dextran sulphate, 3 µl 20 SSC,1 µl 10% SDS in water, 1 µl salmon DNA (10 ng/µl), the wheat pta71 probe (a 9kb fragment from Triticum aestivum containing the 18S-5.8S.25S rdna and intergenic spacers (Gerlach and Bedbrook, 1979), that was directly labelled with TRITC falloidin (wavelength: 514/610 nm), in distilled H 2 O up to a final volume of 30 µl/eppendorf. It has been previously denaturated for 10 min and then incubated on ice for 5 min.then the nuclei were placed in the programmable thermal controller with a selected programme (75 C 5 min, 55 C 2 min; 50 C 0.30 min; 45v 1 min; 42 C 2 min; 40 C 5 min; 38 C 5 min). Once the programme was finished, the eppendorfs were placed at 37 C in a humid chamber and let them hybridize for 2h. After hybridization the nonspecifically bound and the weakly hybridized probe was removed by extensively washing in 0.05% Tween in PBS.The nuclei were then incubated with a 1:10 dilution of the primary sheep polyclonal antibody to Z-DNA (Abcam), left overnight at room temperature. This incubation was followed by 1h incubation at the room temperature with the rabbit polyclonal to sheep IgG FITC-conjugated secondary antibody (Abcam, dilution 1:200). In all these steps centrifugation at 1000g was provided. After washing in 0.05% Tween in PBS, the nuclei were spread on coverslips and directly stained with 1 mg/ml 4,6-diamino-2-phenylindole (DAPI) and mounted, using Vectashield/Vector as the anti-fading mounting medium. Immunofluorescence Immunofluorescence was performed either on coverslips containing a layer of isolated nuclei (for runon and Z-DNA immunolocalization experiments) or in eppendorfs containing the nuclear fractions, that after incubation with the secondary antibody were dropped on coverslips (for FISH and Z-DNA immunolocalizaton experiments). For the Z-DNA and IFA detection the nuclei were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 30 min. Then extensively washed in 0.05% Tween in PBS, followed by 10 min permeabilization with 1% Triton-X in PBS (all at 4 C). After 10 min washing with 0.05% Tween in PBS the nuclei were incubated 10 min with 20 mm glycin in PBS solution, then washed and blocked for 30 min in 2% BSA in PBS. After blocking the nuclei were incubated with the primary sheep polyclonal antibody to Z-DNA (Abcam, dilution 1:10) overnight, at room temperature. The incubation with the primary antibody was followed by 1h incubation at the room temperature with the anti-rabbit polyclonal to sheep IgG FITC- conjugated antibody (Abcam, 1:200 dilution). After each step nuclei were recovered by centrifugation at 1000g. After washing in 0.05% Tween in PBS the nuclei were dropped on coverslips and directly stained with 1mg/mL DAPI and mounted using Vectashield. Slides containing nuclei after the run-on reaction were blocked directly with 2% bovine serum albumin (BSA) in PBS for 30 min at room temperature. Samples were then incubated with the first antibody diluted in blocking solution, for 1 h at 37 C. In the co-localization experiments, they were washed and incubated again with the other first antibody, under the same conditions. After extensive washing in PBS, slides were incubated with the secondary antibody or antibodies, also for 1 h, at 37 C. After a final washing, they were mounted on slides, using Vectashield as anti-fading mounting medium. In controls, the first antibody was omitted. After labelling, preparations were stained with 1 mg/ml DAPI. Confocal microscopy was performed with a Laser Scanning Spectral Confocal Microscope (Leica TCS-SP2-AOBS-UV) supplied with an argon-krypton laser and also with UV (for DAPI). A 63X/ mm immersion objective was used. Digital images were processed with Adobe Photoshop Results In situ assay for active RNA polymerases (run-on experiments) BrUTP-incorporation after the in situ assay for active RNA polymerases detects transcription at discrete sites in the nucleolus and nucleoplasm of plant cells (Morcillo et al., 1976; González- Melendi et al., 2000). Isolated nuclei were used to perform the in situ assay for active RNA polymerases that allows the completion of the elongat- 51
128 A. Cernà ˇ et al. ing RNA chains without allowing the initiation of new transcripts. The assay requires the addition of the four nucleotides. Br-UTP instead of UTP was one of the nucleotides added to the assay, so that its incorporation into the growing RNA chains could be directly detected by immunofluorescence. The use of this run-on assay provided an image of the topological distribution of the transcription foci in the nuclei (Figure 1A). While the nucleoli are dark roundish regions in the blue DAPI-stained nucleus, the green foci in the nucleus represent the place where the elongating chains of RNA are being completed.the largest positive Br-UTP signals correspond to nucleoli, while the non-nucleolar transcription foci are smaller and appear dispersed throughout the whole nucleus. The total amount of nucleoplasmic transcription was usually smaller than that of nucleolar transcription. Most of the transcription foci in nucleoplasm appeared as dots, differing in size. There was no preferential location of transcription foci apposed to nuclear envelope. Distribution of Z-DNA to discrete transcription sites Z-DNA is a highly immunogenic structure and there are poly- and monoclonal antibodies to detect it. Moreover, these antibodies are not species-specific, so that they can be used in different materials. As far as we know this is the first time one of these antibodies is used to detect Z-DNA in plant cells. Under control conditions, immunodetection of Z- DNA showed a distribution pattern (Figure 1B) slightly different from the topological transcription pattern obtained after the run-on experiments (Figure 1A). On one side, Z-DNA occupied a larger portion of both nucleolar masses than the signals produced by the residual transcription in the already initiated nascent RNAs. Secondly, there was an important peripheral layer of signals positive to the Z-configuration with no clear correlate in the in situ assay. The accumulation of signals in the nuclear periphery did not took place in conditions where the non-nucleolar transcription has been prevented by the use of a preferential inhibitor of the non-nucleolar transcription dependent on RNA polymerase II (Figure 1C). These signals might correspond to prem-rna sequences in the process of editing during its export from nucleus, as double-stranded RNA regions pack in the Z-configuration and editing of mrna apparently continues in the cytoplasm (Herbert and Rich, 2001). The Z-DNA spots dispersed throughout these nuclei display a pattern fully compatible with the nucleoplasmic transcription sites. In order to detect whether changes in the pattern of transcription may modify the topology of Z-DNA, we impaired the transcription of polymerase I and II with drugs.the transcription of the RNA polymerase II was preferentially inhibited by the use of the adenosine analogue DRB. Its use diminished the nucleoplasmic foci that contained Z-DNA (Figure 1 C). Under this treatment, the continuous layer of Z- DNA apposed to the internal part of the nuclear envelope in the control cells (Figure 1D) had also disappeared, as commented above. Few foci of Z- DNA were still observed in the nucleoplasm of these cells. On the other hand, in the DRB-treated cells, Z- DNA dispersed throughout the whole nucleoli with multiple small positive dots (Figure 1 E). This dispersion is best seen in the left nucleolus of Figure 1C.There appears to be a small Z-DNA spot for each of the ribosomal genes transcribing in a nucleolus, as their spacing occur every 3 nm, the size of each of the whole transcriptional unit known as Christmas tree. Sometimes, their coalescence provide strong continuous signals instead, as also seen in portions of the right nucleolus in Figure 1C. The DRB secondary effect on unravelling and dispersion of nucleolar rdna sequences was described in mammalian cells (Le Panse et al., 1999). Converserly, cordycepin, another adenosine analogue that preferentially inhibits the transcription by RNA polymerase I, prevented indeed the detection of Z-DNA in the nucleoli (Figure 1D). Simultaneously, the amount of Z-DNA increased and displayed a dispersed pattern in these nuclei, with accumulation of Z-signals in the nuclear periphery, as again occurred under control unimpeded transcription (Figure 1B). Finally, the co-detection by FISH of the rdna codifying for the 45S ribosomal RNA precursor (pta71 probe directly labelled with rhodamine.dutp by nick translation to reduced any background signal) and of Z-DNA immunodetection requires additional efforts to try to find a compatible protocol. Thus, when FISH was carried out overnight on the slides, the detection of rdna was perfect but the detection of Z-DNA was very poor. Converserly, when FISH was reduced to only 2 hours in the nuclear suspension, the rdna signals were greatly reduced in comparison to control, while the immunodetection of Z-DNA reached a level similar to controls (data not shown). 52

129 Original Paper Figure 1. Nuclei of Allium cepa root meristematic cells, under different RNA transcription conditions. A and B: Control conditions. Detection of active RNA polymerases by the in situ assay using Br-UTP. Blue, DAPI staining of nuclear DNA; light green, immunodetection of Br-UTP. The two nucleoli appear relatively poor in DNA in comparison with other nuclear regions, because the large amount of proteins they have. Both have active transcription sites. B: Immunodetection of Z-DNA. Green, Z-DNA; Blue, DAPI. Z-DNA is mostly concentrated in nucleoli, but also distributed throughout nucleoplasm. There is an enrichment in the Z-signals in the peripheral part of nuclei in comparison to the distribution of the transcription foci in A. C: Cell treated with DRB, the preferential inhibitor of non-nucleolar transcription. Z-DNA has diminished in comparison to D, while unravelling of one of the two nucleoli is clearly seen, while the multiple dot signals present in the rightmost nucleolus seems to have a fixed distribution pattern. (Colours as in B). D: Cell treated with cordycepin, the preferential inhibitor of nucleolar transcription. Z-DNA is missing from nucleoli (darker region in this nucleus), while it is still dispersed in the nucleoplasm. (Colours as in B). E. Control nucleus. Immunodetection of intermediate filament (IF) proteins. Red, IFA; blue, DAPI. The IF-proteins localize to a nuclear network pervaiding the nucleus, with a denser distribution in nucleoli and in the nuclear periphery, corresponding to the lamina and nucleolar matrices, respectively. F: Control nucleus. Co-detection of Z-DNA and IF proteins. Both nucleolar and peripheral distribution of Z-DNA are evident. Z-DNA co-localizes with the IF proteins in the nucleoli. However, co-localization is not frequent in the nucleoplasmic signal. The inset at the right of this particular figure shows the topological distribution of IF-proteins and Z-signals distribution in an enlarged region of the nuclear envelope. The Z-signals concentrate underneath the lamina that contains the IF antigens, though they do not co-localize. G: Prominent Z-DNA in the nucleoli of the cells treated with DRB to selectively depress the RNA polymerase II transcription taking place in the nucleoplasm. H: Lack of any Z-DNA signals in the nucleoli of cells treated with cordycepin, the drug that preferentially inhibits the nucleolar RNA polymerase I-dependent transcription. There is again distribution of Z-signals close but not co-localizing with the IF proteins of the nuclear lamina. 53
130 author Z-DNA sites and the nuclear network of intermediate filaments The use of the IFA antibody, against a conserved region essential for dimerization and present in most nuclear IF proteins, showed that the IF protein made up by a loose intranuclear network in control nuclei (Figure 1E). These proteins (contrasted in red) were enriched in the nucleoli, and also in the nuclear periphery. In this particular nucleus, there is retraction of the nuclear DNA content (blue, DAPI signal). This is a common observation after the processing steps it has been submitted. In order to analyze whether the distribution of Z-DNA may be conditioned by the IF presence, co-detection of IF and Z-DNA was carried out both in control conditions (Figure 1F) and under inhibition of nucleoplasmic (Figure 1G) and nucleolar transcription (Figure 1H). The data show adjacent positioning but not co-localization of IF and Z-signals, both in nucleoli and also underneath the internal membrane of the nuclear envelope. This is better seen in the inset located at the right of Figure 1F. In it, it is possible to visualize how the IF proteins are closer to nuclear envelope than their adjacent Z-signals. In the nucleoli, however, there appears to be co-localization of most IF proteins with Z-DNA (Figure 1H), though this may result from the fine and overlapping dispersion of both nucleolar components. Discussion Though it is known for a long time that plants posses Z-DNA binding proteins (Lafer et al., 1985), to our knowledge, this work is the first that shows cross-reactivity of a polyclonal antibody against Z-DNA with plant nuclei. The present results show that indirect immunodetection of the moving and transient Z-DNA is a reliable marker for active transcription. This assay presents some advantages over alternative ones. Thus, Z-DNA immunodetection is faster and cheaper than the in situ assay for active RNA polymerases (run-on), which has being used for comparison in this work. Some patients with systemic lupus erithematosus produce antibodies against Z-DNA (Lafer et al., 1983). Z-DNA is highly antigenic so that there are available commercial monoclonal and polyclonal anti-z-dna antibodies (Lafer et al. 1981) that cross-react with the left hand conformation of DNA irrespective from the species. It is known that the domain of the RNA editing enzyme that anchors to Z-DNA to edit nascent RNA recognizes the B-DNA to Z-DNA transition (Kim et al., 2000). Z-DNA forms as a consequence of the operation of the RNA polymerase I in the nucleolus and also of the RNA polymerase II during the synthesis of pre-messenger RNAs in the non-nucleolar region of the plant nuclei, as a consequence of the DNA negative supercoiling produced behind the progressing polymerases (Liu and Wang, 1987). The already formed Z-DNA becomes the target of DNA topoisomerases that relax such a structure in around 15 min after transcription is turned off (Wolft et al., 1995, 1996).This is supported by data showing the recognition of Z-DNA by topoisomerase II in Drosophila (Arndt-Jovin et al., 1993). It would be possible to increase the amount of Z- DNA by stabilizing it through the addition of cations as spermine, spermidine and Ca 2+ (Behe and Felsenfeld, 1981) and high salt. Thus, stabilized Z- DNA could be detected in the nuclear matrices isolated after high salt extraction where proteins of the intermediate filament family have also being detected (Moreno Díaz de la Espina, 1995). Their co-detection with specific sequences as those constituting the MAR sequences (Matrix Attachment Regions) in Arabidopsis (Van Drunen et al., 1997) may be possible. The topological transcription pattern for both RNA polymerase I and II in Allium is similar in plant and animal cells (Wansick et al., 1993). By changing the in vivo conditions of the cell it would be possible to favour the discernment of transcription that depends on a single of the different RNA polymerases. Thus, making the ionic requirements more strict for one single polymerase will ensure the recording of one of the three transcription systems that coexist in the eukaryotic cell. An intriguing fact deserved our attention.there is an abundant Z-DNA signal inside the nuclear envelope in control conditions. It does not correlate with the presence of Br-UTP incorporation, and it fades away after inhibition of the non-nucleolar transcription. It could be thought that this signal result from the flow of pre-mrnas towards cytoplasm in the nuclei that are actively transcribing in the proliferating cells. The Z-conformation may also be formed in the double-stranded regions of RNA (Davis et al., 1986). This hypothesis deserves to be carefully explored. Finally, the easy detection of Z-DNA will favour 54
131 Original Paper the still missing study to define some putative functional role of this DNA, apart from being the mechanical consequence of transcription. We are aware of the many questions on Z-DNA that still remain open, but the potential of the in situ immunodetection of Z-DNA as an easy marker of transcription is already clear. Acknowledgements We thank Ms. M. Carrascosa, Mr. J.L. Marcilla and Mrs. M Carnota for their excellent technical contribution. Funding The work has been partially supported by the Dirección General de Investigación del Ministerio de Ciencia y Tecnología of Spain (Project BMC ). Adriana Černà was supported by a fellowship from the Spanish Agency of Internacional Cooperation (AECI), from the Ministerio de Asuntos Exteriores, Spain. References Arndt-Jovin DJ, Robert-Nicoud M, Baurschmidt P, Jovin TM. Immunofluorescence localization of Z-DNA in chromosomes: Quantitation by scanning microphotometry and computer-assisted image analysis. J Cell Biol 1985;101: Arndt-Jovin DJ, Udvardy A, Garner MM, Ritter S, Jovin T. Z-DNA binding and inhibition by GTP of Drosophila topoisomerase II. Biochem 1993;32: Behe M, Felsenfeld G. Effect of methylation on a synthetic polynucleotide: the B-Z transition in poly(dg-m5dc). Poly(dG-m5dC). Proc Natl Acad Sci USA 1981;78: Davies P.W, Hall K., Cruz P., Tinoco I and Nelson T.: The tetraribonucleotide rcpg pcpg forms a left-handed Z-RNA double helix. Nucleic Acids Res 1986; 14: De Cárcer G, Medina J. Simultaneous localization of transcription and early processing markers allows dissection of functional domains in the plant cells nucleolus. J Struct Biol 1999; 128: Díez JL. Effect of cordycepin (3 -desoxyadenosine) on polytene chromosomes of Chironomus pallidivittatus salivary glands. Chromosoma 1973, 42: Fernández-Gómez ME, Medina FJ, Díaz de la Espina SM. Effects of DRB on the ultrastructural organization and RNA metabolism of plant nucleus. In: Harris JR, Zbarsky IB, eds. Nuclear Structure and Function. Plenum Press, New York, pp , Gerlach W.L. and Bedbrook J.R.: Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res 1979; 7: González-Melendi P, Beven A, Boudonck K, Abranches R, Wells B, Dolan L. and Shaw P.:The nucleus: a highly organized but dynamic structure. J Microsc 2000; 198: Herbert A. RNA editing, introns and evolution. Trends Genet 1996; 12:6-9. Herbert A, Rich A.The role of binding domains for dsrna and Z-DNA in the in vivo editing of minimal substrates by ADAR1. Proc Natl Acad Sci USA 2001; 98: Jackson DA, Hassan AB, Errington RJ, Cook PR.Visualization of focal sites of transcription within human nuclei. EMBO J 1993; 12: Kim YG, Lowenhaupt K, Maas S, Herbert A, Schwartz T, Rich A. The zab domain of the human RNA editing enzyme ADAR1 recognizes Z-DNA when surrounded by B-DNA. J Biol Chem 2000; 275, Lafer EM, Moller A, Nordheim A, Stollar BD, Rich A. Antibodies specific for left-handed DNA. Proc Natl Acad Sci USA 1981; 78: Lafer EM, Valle RP, Moller A, Norheim A, Schur PH, Rich A, Stollar B-D. Z-DNA specific antibodies in human systemic lupus erithematosus. J Clin Invest 1983; 71: Lafer EM, Sousa R, Rosen B, Hsu A, Rich A. Isolation and characterization of Z-DNA binding proteins from wheat germ. Biochem 1985; 24: Le Panse S, Masson C, Héliot L, Chassery J-M, Junéra HR, Hernández- Verdún D. 3-D organization of ribosomal transcription units after DRB inhibition of RNA polymerase II transcription. J Cell Sci 1999; 112: Liu LF, Wang JC. Supercoiling of the DNA template during transcription. Proc Natl Acad Sci USA 1987; 84: Martin S, Pombo A. Transcription factories: quantitative studies of nanostructures in the mammalian nucleus. Chrom Res 2003; 11: Moreno Díaz de la Espina S. Nuclear matriz isolated from plant cells. Intern Rev Cytol 1995; 162B: Moreno Díaz de la Espina S. and De la Torre C.: Coiled-coil and intermediate filament-proteins in the plant nucleoskeleton. In: C. Lloyd and Y. Blume, eds. The Plant Cytoskeleton. NATO Monographies Series Morcillo G, De la Torre C, Giménez-Martín G. Nucleolar transcription during plant mitosis. In situ assay for RNA polymerase activity. Exp Cell Res 1976; 102: Peck LJ, Nordheim A, Rich A, Wang JC. Flipping of cloned d(pgpg)n DNA sequences from right to left-handed helical structure by salt, Co(III), or negative supercoiling. Proc Natl Acad Sci USA 1982; 79: Rich A, Zhang S. Z-DNA: the long road to biological function. Nat Rev Genet 2003; 4: Wansink DG, Schul W, van der Kraan I, van Steensel B, van Driel R, de Jong L. Fluorescent labeling of nascent RNA reveals transcription by RNA polymerase II in domains scattered throughout the nucleus. J Cell Biol 1993; 122: Wolfl S,Wittig B, Rich A. Identification of transcriptionally induced Z- DNA segments in the human C-MYC gene. Biochim. Biophys. Acta 1995; 1264: Wolfl S, Martínez C, Rich A, Majzoub JA. Transcription of the human corticotrophin-releasing hormone gene in NPLC cells is correlated with Z-DNA formation. Proc Natl Acad Sci USA 1996; 93:
132 90
133 Chromosoma DOI /s RESEARCH ARTICLE Triplex configuration in the nick-free DNAs that constitute the chromosomal scaffolds in grasshopper spermatids Adriana Černá & Carmen López-Fernández & José Luis Fernández & Susana Moreno Díaz de la Espina & Consuelo de la Torre & Jaime Gosálvez Received: 7 June 2007 /Revised: 7 August 2007 / Accepted: 7 August 2007 # Springer-Verlag 2007 Abstract After applying proper deoxyribonucleic acid (DNA) probes, fluorescence in situ hybridization (FISH) showed that the 8/9 centromeres one per chromatid of the male haploid complement (X0) of Pyrgomorpha conica grasshopper colocalized at the spermatid blunt end, where the spermatozoa flagellum inserts. A bundle of aligned 4,6- diamidino-2-phenylindole-positive chromatid scaffolds, which formed the central spermatid core, was observed after DNA breakage detection followed by FISH. Modular nature of scaffold DNA was occasionally evident. The technique also showed that in the early spermatid, the chromatid scaffolds lacked any DNA nick, whereas abundant breaks accumulated in the surrounding loops. Moreover, immunodetection showed that scaffold DNA participated in the formation of triplex DNA, while this configuration was absent from the loops. During spermatid maturation, triplex DNA disappeared from the scaffold in parallel with loop retraction, while protamines replace Communicated by S. Gerbi A. Černá : S. Moreno Díaz de la Espina : C. de la Torre (*) Centro de Investigaciones Biológicas, CSIC, Ramiro de Maeztu, 9, Madrid 8040, Spain delatorrec@cib.csic.es C. López-Fernández : J. Gosálvez Departamento de Biología. Facultad de Ciencias, Universidad Autónoma de Madrid, Madrid 8049, Spain J. L. Fernández Laboratorio de Genética, Centro Oncológico de Galicia, A Coruña, Spain A. Černá Institute of Experimental Botany, CSAV, Prague, Czech Republic histones. Thus, the presence of triplex DNA in the chromatid scaffold correlates with the anchoring of expanded DNA loops to it. After loop retraction, the scaffolds of all chromatids coiled as a single unit in the spermatid head. This cooperative coiling produced enlargement and tilting of the distal telomeric signals, which were distributed along the spermatid head according to the length of each chromosome. We propose that specific DNA sequences dispersed throughout the whole chromatid fold forward and backward coaxially to chromatid length, forming individual scaffold modules whose linear assembly accounts for the minimum length of each individual chromatid. Finally, the core of the grasshopper male spermatid should be considered as a single chromosome in which the DNA scaffolds of the whole set of the nonhomologous chromosomes of the haploid complement are interconnected. This pattern of chromatin organization applies probably to other elongated spermatids. Keywords Chromatid scaffold. DNA loops. Triplex DNA. DNA breaks. Sperm chromatin structure. Spermiogenesis Introduction The morphology of the spermatid nuclei changes as basic proteins substitute for histones in Drosophila (Hennig 2003) and extensive chromatin condensation is being achieved (Jayaramaiah-Raja and Renkawitz-Pohl 2005). During Drosophila spermiogenesis, histones are modified by sitespecific methylation, acetylation and monoubiquitination. Then, histones are transiently replaced by transition proteins and these, in turn, by protamines (Rathke et al. 2007). The process is essentially analogous to that taking place in mammalian spermiogenesis (Govin et al. 2004).
134 Chromosoma However, there are not protamines in the spermatozoa of the house cricket. Instead, they contain five histones resembling the F1, F3, F2a1, F2a2, and F2b of calf thymus, plus a new basic protein that appears in late spermatids or in spermatozoa (Tessier and Pallota 1973). This additional protein is an arginine-rich histone with unique features (Pallota and Tessier 1976). During spermiogenesis, abundant deoxyribonucleic acid (DNA) breaks appear, as reported both in mammalian (Sakkas et al. 1995) and in insect species (Rathke et al. 2007). The production of DNA nicks during spermiogenesis will obviously ameliorate the torsional strain produced in unconstrained supercoils not only during the replacement of DNA-binding proteins but also during the extreme chromatin condensation that characterizes this process. The induction of DNA nicks seems to depend on topoisomerase II because, in short-term cultures of mouse testicular cells, inhibitors of this enzyme prevents their appearance (Labergé and Boissonneault 2005). It is largely unknown how sperm chromatin structure avoids any illegitimate DNA recombination to take place during repair of the physiological double-strand breaks taking place in spermiogenesis. Triplex DNA (also known as H-DNA) is a relatively rare conformation of native DNA in which two pyrimidine nucleotide strands share a common purine strand (Felsenfeld and Rich 1957). Triplex DNA motifs leave single-stranded DNA apt to hybridize with complementary single-stranded DNAs or ribonucleic acids. Triplex DNA is credited to be operative in chromatin organization. Thus, short triplex DNA motifs self-assemble to give rise to highly condensed and ordered structures (Goobes et al. 2002). In insects, triplex DNA accumulates in the polytene chromosome bands inactive in transcription, where it is involved in the ectopic pairing of nonhomologous genome regions (Burkholder et al. 1991). Thus, some triplex DNA sequences are probably responsible for the nonhomologous recombinational DNA repair of double-strand breaks. This mechanism is the only one possible in the spermatid uninemic chromosomes, as it is the case in the uninemic G1 chromosomes of somatic cells (Takata et al. 1998). In the human genome, triplex DNA accumulates also on gene promoters, suggesting a possible inhibitory role on any transcription but the tissue-specific one (Goñi et al. 2004). Little is known about the actual state of chromatin organization in the spermatid, and this applies to all species. The chromatid is the basic unit of the eukaryotic chromosome. There is a single chromatid per chromosome after the postmeiotic telophase II and also after the somatic mitosis. Visualization of unexpected features of the scaffolds supporting the chromatids constituting the spermatid head of a grasshopper was possible because of their exquisite ordering and extreme packing. The present research was aimed to look for distribution of triplex DNA, if any, and of DNA nicks during the chromatin remodeling taking place throughout spermiogenesis in the grasshopper Pyrgomorpha conica. Spermatogenesis of this insect was selected because we had previously developed a series of specific DNA probes for telomeric and centromeric sequences (López- Fernández et al. 2004, 2006), which were perfect tools to follow their distribution in the spermatid head. Materials and methods Testes of P. conica (Orthoptera: Pyrgomorphidae) were removed from adult males. Some of them were fixed in 3:1 ethanol/acetic acid (phase contrast microscopy and conventional fluorescence in situ hybridization [FISH]). Other animals were kept alive for fresh utilization. Fluorescence in situ hybridization For conventional in situ hybridization, either a specific DNA clone (PyrAlu-2p) derived from a P. conica DNA library specific for pericentromeric heterocromatin, a DNA probe with a poly-ttagg sequence to label telomeres, or a whole genomic DNA was used. DNA probes were biotinylated by nick translation, denatured, and hybridized overnight at room temperature on dried slides. Slides were washed twice in 50% formamide in 2 salt sodium citrate (SSC), ph 7.0, for 5 min, and twice in 2 SSC at ph 7.0, for 3 min, at room temperature. The hybridized probe was detected with streptavidin-cy3 (1:200; Sigma-Aldrich Química SA, Madrid, Spain). Cells were counterstained with 4,6-diamidino-2-phenylindole (DAPI) or propidium iodide in Vectashield (Vector, Burlingame, CA). Preparation of nucleoids To prepare nucleoids in agarose microgels, meiocytes were released from testes by mechanical separation to obtain single cell suspensions. Cell concentration was adjusted to five to ten cells per field, under 40 objective (approx. 10 million cells/ml). Cells were collected in a 1.5-ml Eppendorf tube. After 5 min sedimentation, the supernatant was transferred to another tube and centrifuged at 1,000 g for 5 min. The cell pellet was resuspended in fresh saline solution. Cells were mixed with low-melting-point agarose at 37 C at a final concentration of 0.7%. Fifteen microliters of the mixture were layered onto a glass slide precoated with 0.65% standard agarose dried at 80 C, covered with a mm coverslip, and allowed to solidify at 4 C. To remove nuclear proteins to get the nucleoids, coverslips were gently removed. Then, the slides were placed in the lysing solution (0.4 M Tris HCl, 2 M NaCl, 1% sodium
135 Chromosoma dodecyl sulfate [SDS], ph 7.5), at room temperature, for 20 min. To find out whether the possible presence of disulfide bonds may affect the appearance of nucleoids, a different lysing solution containing 2 M 1, 4-dithio-DL-threitol (DTT, from Fluka Chemie AG, Germany), 2 M NaCl, and 1% SDS, ph 7.5, was used. Its DTT component will release any S S links present between the nuclear proteins. FISH detection of DNA breaks (DBD-FISH) To combine DNA breakage detection (DBD) with FISH (Fernández et al. 1998), half of each slide was covered with the insect cells, while human blood leukocytes (control) occupied the other half. After the lysing solution was used to remove proteins, the resultant nucleoids were washed in 0.9% NaCl for 10 min before being incubated in an alkaline unwinding solution (0.03 M NaOH, 1 M NaCl) for 2.5 min at 7 C, in the dark. This process transforms DNA breaks and alkali-labile sites into short single-stranded DNA regions. After neutralizing for 5 min with 0.4 M Tris-HCl, nucleoids were washed in TBE buffer (0.445 M Tris/borate, M disodium ethylenediamine tetraacetic acid) for 2 min, dehydrated by their passage throughout 70, 90, and 100% ethanol baths (2 min in each), and air-dried. For in situ hybridization, a P. conica whole genome probe (4.3 ng/μl in 50% formamide/2 SSC, 10% dextran sulfate, 100 mm calcium phosphate, ph 7.0; 1 SSC is M Na citrate, 0.15 M NaCl, ph 7.0), which had been biotin-labeled by nick-translation, was denatured and hybridized overnight at room temperature on dried slides. In parallel, a whole DNA probe using human DNA was prepared under similar conditions and used as an internal hybridization control. After hybridization, slides were washed twice in 50% formamide/2 SSC ph 7.0, for 5 min, and twice in 2 SSC ph 7.0 for 3 min, at room temperature. The hybridized probe was detected with streptavidin-cy3 (1:200; Sigma-Aldrich Química SA). Cells were counterstained with DAPI or propidium iodide in Vectashield (Vector). Triplex DNA detection To detect triplex DNA, immunofluorecence was performed on agarose microgels containing a layer of nuclei of spermatozoa from fresh testis tissues. The microgels were permeabilized for 10 min with 1% Triton-X in phosphatebuffered saline (PBS) and washed for 10 min with 0.05% Tween in PBS. After incubation with 20 mm glycine in PBS solution, the slides were incubated with primary antitriplex antibodies overnight, in a humid chamber. Extensive washing in 0.05% Tween in PBS (3 10 min) and 1 h incubation with goat anti-mouse secondary antibody Alexa 488 (Molecular Probes, wavelength=495/519 nm) followed. After washing with 0.05% Tween in PBS, the nuclei were counterstained with DAPI in Vectashield (Vector). The primary monoclonal antibody Jel 466, prepared from mice immunized with poly[d(tm5c)].poly[d(ga)], or Jel 318, the triplex-specific antibody that binds to poly[d(t) l-poly[d(a)]poly[d(t)], were used at 1:20 dilutions (Ohno et al. 2002). FISH detection of DNA breaks (DBD-FISH) signals and anti-triplex DNA was sequentially performed. The protocol included first the detection of the antibody against triplex DNA, followed by the DBD-FISH procedure to discern DNA breaks. The alkali solution used for DNA denaturation removes most of the detected antibody. After performing DBD-FISH, digital images were captured with a motorized Leica DML microscope to fix targeted regions on the slide. After DBD-FISH, the slides were reanalyzed and photographed, under a DMRB epifluorescence microscope (Leica, Wetzlar, Germany), equipped with a Leica CCD black and white 12-bit camera. A PL Fluotar 100 objective and appropriate fluorescence filters for DAPI and Cy3 were used. Direct-color photographs were acquired with a Photometrics CoolSnap CCD mounted on a motorized Leica DML microscope. Results Markers for sperm chromatin distribution Different stages could be recognized in the process during which nuclei of young roundish spermatids transform into elongated sperm heads (stages 1 to 6 in Fig. 1a). The spermatid blunt end connects to a long flagellum (stages 3 to 6 in Fig. 1a) and produces an oriented long sperm with a distal thin end and a proximal end containing bulk chromatin (Fig. 1a). The elongated sperm nuclei (stage 7 in Fig. 1b) are the final stages found within the gonad, as the different mature spermatids here shown will be subsequently released to the deferent tubuli. The P. conica grasshopper displays XX/XO sex determinism. Thus, 8/9 chromatids, corresponding to all the uninemic (unreplicated) chromosomes of its male haploid complement, are contained in each spermatid. A very distinctive feature of this species is that all its chromosomes are acrocentric. Conventional FISH with DNA probes (PyrAlu-2p) that hybridize to DNA sequences in pericentromeric constitutive heterochromatin (López-Fernández et al. 2006) showed the location of all the centromeres in the diplotene chromosomes (pink, in Fig. 2a). All of the chromosomes here displayed are bivalents that, in this species, tend to have a chiasma close to centromeres. Then,

, where a maximum number of nine centromeres were detected.")
136 Chromosoma Fig. 1 Morphology of P. conica spermatid nuclei. Numbers 1 to 6 in a and number 7 in b show progressive stages of spermatid maturation taking place during male spermiogenesis in this species. Bars=400 μm some bivalents look like a metacentric chromosome, and the chiasma might be confused with a centromere, although this is not the case. FISH signals for detecting centromeres show how they concentrate in the blunt end of the young roundish spermatids (yellow in Fig. 2b,d), where a maximum number of nine centromeres were detected. Frequently, the proximity among the different centromeres barred their resolution as separate units. The centromeres concentrated at the spermatid blunt end (Fig. 2c,d) early in the spermiogenesis and remained in this position throughout the whole spermiogenesis, as shown in the fully elongated spermatids (Fig 2e). Distribution of telomeres in the spermatid was also revealed by using this time the poly-ttagg DNA probe for telomeric sequences. The size of telomeres in this species visualized with this DNA probe is very small when compared with those in other Orthopteran species, even in Fig. 2 Conventional FISH using pericentromeric and telomeric DNA probes. a Diplotene showing the pericentromeric localization of PyrAlu-p2 DNA probe (pink) in all of the bivalent chromosomes, in the meiosis of this species. Their frequent chiasma tends to be formed close to centromeres, so that these bivalents might be confused with metacentric chromosomes. b Localization of the pericentromeric DNA probe at the proximal end of roundish early spermatids. c, d Roundish spermatids showing polarized distribution of this DNA probe under phase contrast and FISH, respectively. e Elongating sperm head showing the same polarized distribution of the pericentromeric DNA probe. f Telomeric DNA sequences detected by FISH after applying a poly-ttagg DNA probe. These sequences are distributed in around eight sites along the whole length of the elongating spermatid. These signals correspond to the telomeres distal to centromeres, while those corresponding to telomeric sequences in the pericentromeric heterochromatin remain undetected. g Telomeric signals in a final stage spermatid. Note that the final coiling of all chromatid scaffolds forming the spermatid core tiltes the telomeres of the different chromatids. h Elongated spermatid nuclei showing the coiling achieved by the spermatid nuclei (SybGreen II staining) at late spermiogenesis. Bars=300 μm

.")
137 Chromosoma Fig. 3 DNA loops release in the DTT-treated elongating spermatids. a c In response to the DTT-induced disulfide bond disruption, spermatids display a central core surrounded by a peripheral halo of chromatin. b Enlarged detail of a. c Early spermatid nuclei showing the chromatin halo formed by DNA loops after SybGreen II staining. d, e No dispersed chromatin halo is observed in the absence of DTT, i.e., when the disulfide bridges remain undisturbed. Bars: a, c d=40 μm; b=25 μm the highly undercondensed meiotic chromosomes (López- Fernández et al. 2004). In the spermatid cells, the FISH hybridization signals for these telomeric DNA probes corresponded only to the distal telomeres, i.e., to telomeres localized at the noncentromeric end of each chromosome arm (Fig. 2f,g). On the other hand, the interstitial telomere sequences localized close to or embedded into the constitutive pericentromeric heterocromatin were not visualized. However, because all chromosomes are acrocentric in this species, the apparently missing set of proximal telomeres should be located at the spermatid proximal end and hence close to or embedded into the constitutive heterochromatin localized by the PyrAlu-2p DNA probe (at the bottom of Fig. 2f,g). Coiling and looping of the spermatid core After achieving maximum chromatin condensation, the bundle of chromatid scaffolds forming the spermatid core coiled (Fig. 2g). This modified the morphology of the FISH signals corresponding to the distal telomeres distributed throughout the whole spermatid length in the mid-spermatid. Fig. 4 Early spermatid nuclei sequentially processed for visualization of DNA nicks and triplex DNA, after previous release of DNA loops induced by DTT. a Presence of DNA breaks after DBD-FISH by using a whole genome DNA probe (red). Individual chromatid scaffolds (blue, DAPI + ) align coaxially to the spermatid length, forming the spermatid core. The set of chromatid scaffolds seem to be formed by nick-free DNA. b Immunolocalization of triplex DNA shows that it concentrates in the spermatid core, i.e., where the chromatid scaffolds are located. In spermatids at this developmental stage, their accumulation is especially evident in the scaffolds regions associated to centromeres or in centromeres themselves. Gaps in the triplex DNAcontaining scaffolds proximal to centromeres are clearly observed. c Merge of images a and b showing that nick-free DNA and triplex DNA colocalize in the central spermatid core, where the chromatid scaffolds are located. Bars=40 μm
.")
138 Chromosoma The telomeric signals clearly enlarged and became tilted in relation to the spermatid long axis (Fig. 2g). The cooperative coiling of the bundle of DNA chromatid scaffolds was also visualized in the most mature elongated spermatid heads, by using the SybGreen DNA fluorescent dye (Fig. 2h). No traces of FISH hybridization were detected under the maximum spermatid coiling achieved by late spermiogenesis. When the intact unfixed spermatids are embedded in an agarose microgel and then subjected to a lysis buffer, chromosomal proteins are lost (Fernández et al. 2002). Addition of DTT to the buffer used to break the protein disulfide bonds resulted in the observation of DNA loops forming a halo along the spermatid axis or core (Fig. 3b,c), independently from the spermatid maturation stage (compare Fig. 3c,d). DNA loop release was homogeneously distributed along the spermatid whole length, and the emerging loops were similar in size (Fig. 3a,b), although their density was lower at the distal than at the proximal (centromeric) spermatid ends (Fig. 3c). In open contrast, no conspicuous DNA loops were observed when the lysing buffer did not contain any DTT, although SDS was still present (Fig. 3d,e). However, a few scaffold widenings were observed before maximum compaction of the spermatid core was achieved (Fig. 3d). They may correspond to spontaneous and probably physiological relaxation of some specific DNA loops. It should be worthy to analyze whether these slightly expanded regions could be related to the low level of spermatid-specific transcription. Looped and scaffold DNAs. The uneven distribution of DNA breaks and triplex DNA To localize the possible presence of DNA breaks altogether with constitutive alkaline-labile sites, the intact unfixed spermatids embedded in an agarose microgel were subjected to the DBD-FISH protocol (Fernández et al. 1998), after using a lysing buffer containing DTT to break any possible disulfide bonds between proteins. Under these conditions, a bundle of DAPI + caffolds (blue), as those seen in the early spermatid of Fig. 4a, were neatly observed. Various DAPI + scaffolds ran in a parallel position, coaxially to the spermatid length. They finished in a large granule at the spermatid blunt end where all centromeres, a scaffoldspecific structure, were clustered (blue, bottom part of Fig. 4a c). Using a whole genome DNA probe, it was observed that DNA breaks (red) were homogenously distributed in the early spermatids (Fig. 4a,c). They did not quench the signal produced by DAPI intercalation in the different scaffold Fig. 5 DBD-FISH and triplex DNA localization during spermatid elongation and, also, in fully elongated spermatids. a e A series of elongating spermatids showing the regional distribution of DNA loops containing DNA breaks (red, FISH with a whole genome DNA probe). Note that the final chromatin region showing DNA loops with DNA breaks corresponds to the pericentromeric or centromeric regions. In blue, DAPI counterstaining to show the main chromatin body in the sperm head. f When the spermatid achieves a maximum condensation and its central core coils, FISH signals were no longer visible. g i A series of selected spermatids showing triplex DNA (green) distributed along the spermatid head and their DAPI counterstain (blue). Observe colocalization of triplex DNA with the most positive regions to DAPI, which should correspond to those enriched in AT. j When the spermatid head coils, again, triplex DNA is not detected any longer. Bars= 100 μm
 that were distributed along the spermatid chromatid scaffolds (Fig. 4a,c, red).")
139 Chromosoma DNAs. Hence, scaffold DNA was apparently nick-free. On the other hand, abundant DNA nicks were present in the DNA loops (expanded by a previous treatment with DTT) that were distributed along the spermatid chromatid scaffolds (Fig. 4a,c, red). Triplex DNA was also immunolocalized by specific antibodies (Burkholder et al. 1991), in the protein-depleted spermatids suspended in microgels. Triplex DNA (green) was absent from the dispersed peripheral loops. It accumulated instead on the bundle of chromatid scaffolds forming the longitudinal spermatid core (Fig. 4b). Centromeric scaffold DNA regions were specially enriched in triplex DNA and formed a large granule. Interstitial segments of the DNA scaffolds corresponding to centromeres and pericentromeric regions, were apparently discontinuous or displayed different compaction levels (green signals at bottom of Fig. 4b,c). Other intercalar triplex DNA signals, distributed all along the whole length of the spermatid core, accumulated in structures resembling dots (green in color). The elongated spermatids of Fig. 5, which are more mature than those in Fig. 4, after their lysis in the absence of DTT, showed their DAPI + chromatid scaffolds (blue) aggregated in a tight solid spermatid core (Fig. 5a f,j). Discrete clusters of hybridization signals corresponding to DNA nicks (red) were identified by FISH (Fig. 5a e). They seem to correspond to previous loops that had not yet fully retracted toward the bundle of DAPI + chromatid scaffolds (Fig. 5a e), as they disappeared earlier in the telomeric (upper part, Fig. 5c e) than in the centromeric spermatid ends (bottom of Fig. 5c e). In the nearly mature spermatids, chromatid scaffolds remained nick-free. In relation to triplex DNA detection in the mature long spermatids, the most intense signals (green) colocalized with the brighter DAPI + (blue) regions, i.e., scaffold DNA sequences relatively enriched in AT (Fig. 5g i). Triplex DNA (green) disappeared gradually from the DAPI + scaffolds (blue), from distal to proximal regions of the spermatid. Thus, disappearance of triplex DNA from the spermatid core roughly followed the compaction and retraction of DNA loops observed by DBD-FISH (Fig. 5b d), in spite that this last process depends on the formation of disulfide bridges linking chromosomal proteins. When the bundle of chromatid scaffolds forming the spermatid core was completely coiled, no FISH signals for DNA breaks (red) or signals for triplex DNA (green) remained in these spermatids (Fig. 5f,j). Fig. 6 Model for the association of chromatid scaffolds in the spermatid and for looped DNA distribution in each individual chromatid scaffold. Only three out of the 8/9 chromatid scaffolds contained in the spermatid head are depicted. a In the spermatid head, all the FISH-detected centromeres occupy the blunt spermatid tip (as observed in Fig. 2b,d), where the proximal telomeres remain undetected, because their embedding in the pericentromeric heterochromatin. On the other hand, the distal telomeres are detected at different distances from centromeres (as observed in Fig. 2f,g). b Cooperative coiling of the different chromatid scaffolds in the spermatid. The telomeric signals grow and tilt in relation to the spermatid core, making evident their coiling (as observed in Fig. 2g). c Model for the distribution of DNA in each of the individual scaffolds forming the spermatid cores. Four contiguous modules of scaffold-specific DNA sequences are depicted between the centromeric spermatid end and the end where the telomere of the longest chromosome of this grasshopper complement is located. Scaffold DNA folds forward and backward, coaxially to the chromosomal length, while DNA loops emerge from such scaffold in some of the regions where triplex DNA structures are located. Retraction of DNA loops toward the scaffold by late spermiogenesis will make that both looped DNA and its binding proteins appose to the chromosomal scaffold. DNA loop retraction occurs earlier in the distal telomeric than in the centromeric regions of the chromatid scaffold (as observed in Fig. 5c,d)
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
SOIL ECOLOGY the general patterns, and the particular
 Soil Biology topic No. 5: SOIL ECOLOGY the general patterns, and the particular patterns SOIL ECOLOGY is an applied scientific discipline dealing with living components of soil, their activities and THEIR
Soil Biology topic No. 5: SOIL ECOLOGY the general patterns, and the particular patterns SOIL ECOLOGY is an applied scientific discipline dealing with living components of soil, their activities and THEIR
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Gymnázium, Brno, Slovanské nám. 7 WORKBOOK. Mathematics. Teacher: Student:
 WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky
 fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky") FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost.
 Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Litosil - application
 Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Aktivita CLIL Chemie I.
 Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
PC/104, PC/104-Plus. 196 ept GmbH I Tel. +49 (0) / I Fax +49 (0) / I I
 / I Fax +49 (0) / I I") E L E C T R O N I C C O N N E C T O R S 196 ept GmbH I Tel. +49 (0) 88 61 / 25 01 0 I Fax +49 (0) 88 61 / 55 07 I E-Mail sales@ept.de I www.ept.de Contents Introduction 198 Overview 199 The Standard 200
E L E C T R O N I C C O N N E C T O R S 196 ept GmbH I Tel. +49 (0) 88 61 / 25 01 0 I Fax +49 (0) 88 61 / 55 07 I E-Mail sales@ept.de I www.ept.de Contents Introduction 198 Overview 199 The Standard 200
VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL. S R. O.
 VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL. S R. O. Návrh konceptu konkurenceschopného hotelu v době ekonomické krize Diplomová práce 2013 Návrh konceptu konkurenceschopného hotelu v době ekonomické krize Diplomová
VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL. S R. O. Návrh konceptu konkurenceschopného hotelu v době ekonomické krize Diplomová práce 2013 Návrh konceptu konkurenceschopného hotelu v době ekonomické krize Diplomová
Czech Republic. EDUCAnet. Střední odborná škola Pardubice, s.r.o.
 Czech Republic EDUCAnet Střední odborná škola Pardubice, s.r.o. ACCESS TO MODERN TECHNOLOGIES Do modern technologies influence our behavior? Of course in positive and negative way as well Modern technologies
Czech Republic EDUCAnet Střední odborná škola Pardubice, s.r.o. ACCESS TO MODERN TECHNOLOGIES Do modern technologies influence our behavior? Of course in positive and negative way as well Modern technologies
USING VIDEO IN PRE-SET AND IN-SET TEACHER TRAINING
 USING VIDEO IN PRE-SET AND IN-SET TEACHER TRAINING Eva Minaříková Institute for Research in School Education, Faculty of Education, Masaryk University Structure of the presentation What can we as teachers
USING VIDEO IN PRE-SET AND IN-SET TEACHER TRAINING Eva Minaříková Institute for Research in School Education, Faculty of Education, Masaryk University Structure of the presentation What can we as teachers
2. Entity, Architecture, Process
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Praktika návrhu číslicových obvodů Dr.-Ing. Martin Novotný Katedra číslicového návrhu Fakulta informačních technologií ČVUT v Praze Miloš
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Praktika návrhu číslicových obvodů Dr.-Ing. Martin Novotný Katedra číslicového návrhu Fakulta informačních technologií ČVUT v Praze Miloš
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA
 GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
Configuration vs. Conformation. Configuration: Covalent bonds must be broken. Two kinds of isomers to consider
 Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
Biosensors and Medical Devices Development at VSB Technical University of Ostrava
 VŠB TECHNICAL UNIVERSITY OF OSTRAVA FACULTY OF ELECTRICAL ENGINEERING AND COMPUTER SCIENCE Biosensors and Medical Devices Development at VSB Technical University of Ostrava Ing. Martin Černý Ph.D. and
VŠB TECHNICAL UNIVERSITY OF OSTRAVA FACULTY OF ELECTRICAL ENGINEERING AND COMPUTER SCIENCE Biosensors and Medical Devices Development at VSB Technical University of Ostrava Ing. Martin Černý Ph.D. and
Introduction to MS Dynamics NAV
 Introduction to MS Dynamics NAV (Item Charges) Ing.J.Skorkovský,CSc. MASARYK UNIVERSITY BRNO, Czech Republic Faculty of economics and business administration Department of corporate economy Item Charges
Introduction to MS Dynamics NAV (Item Charges) Ing.J.Skorkovský,CSc. MASARYK UNIVERSITY BRNO, Czech Republic Faculty of economics and business administration Department of corporate economy Item Charges
CHAPTER 5 MODIFIED MINKOWSKI FRACTAL ANTENNA
 CHAPTER 5 MODIFIED MINKOWSKI FRACTAL ANTENNA &KDSWHUSUHVHQWVWKHGHVLJQDQGIDEULFDW LRQRIPRGLILHG0LQNRZVNLIUDFWDODQWHQQD IRUZLUHOHVVFRPPXQLFDWLRQ7KHVLPXODWHG DQGPHDVXUHGUHVXOWVRIWKLVDQWHQQDDUH DOVRSUHVHQWHG
CHAPTER 5 MODIFIED MINKOWSKI FRACTAL ANTENNA &KDSWHUSUHVHQWVWKHGHVLJQDQGIDEULFDW LRQRIPRGLILHG0LQNRZVNLIUDFWDODQWHQQD IRUZLUHOHVVFRPPXQLFDWLRQ7KHVLPXODWHG DQGPHDVXUHGUHVXOWVRIWKLVDQWHQQDDUH DOVRSUHVHQWHG
The Over-Head Cam (OHC) Valve Train Computer Model
 Valve Train Computer Model") The Over-Head Cam (OHC) Valve Train Computer Model Radek Tichanek, David Fremut Robert Cihak Josef Bozek Research Center of Engine and Content Introduction Work Objectives Model Description Cam Design
The Over-Head Cam (OHC) Valve Train Computer Model Radek Tichanek, David Fremut Robert Cihak Josef Bozek Research Center of Engine and Content Introduction Work Objectives Model Description Cam Design
WORKSHEET 1: LINEAR EQUATION 1
 WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable
WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable
Compression of a Dictionary
 Compression of a Dictionary Jan Lánský, Michal Žemlička zizelevak@matfyz.cz michal.zemlicka@mff.cuni.cz Dept. of Software Engineering Faculty of Mathematics and Physics Charles University Synopsis Introduction
Compression of a Dictionary Jan Lánský, Michal Žemlička zizelevak@matfyz.cz michal.zemlicka@mff.cuni.cz Dept. of Software Engineering Faculty of Mathematics and Physics Charles University Synopsis Introduction
Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E.
 Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E. Pokorný Mendel University of Agriculture and Forestry, Department of
Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E. Pokorný Mendel University of Agriculture and Forestry, Department of
CHAIN TRANSMISSIONS AND WHEELS
 Second School Year CHAIN TRANSMISSIONS AND WHEELS A. Chain transmissions We can use chain transmissions for the transfer and change of rotation motion and the torsional moment. They transfer forces from
Second School Year CHAIN TRANSMISSIONS AND WHEELS A. Chain transmissions We can use chain transmissions for the transfer and change of rotation motion and the torsional moment. They transfer forces from
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Caroline Glendinning Jenni Brooks Kate Gridley. Social Policy Research Unit University of York
 Caroline Glendinning Jenni Brooks Kate Gridley Social Policy Research Unit University of York Growing numbers of people with complex and severe disabilities Henwood and Hudson (2009) for CSCI: are the
Caroline Glendinning Jenni Brooks Kate Gridley Social Policy Research Unit University of York Growing numbers of people with complex and severe disabilities Henwood and Hudson (2009) for CSCI: are the
CARBONACEOUS PARTICLES IN THE AIR MORAVIAN-SILESIAN REGION
 UHLÍKATÉ ČÁSTICE V OVZDUŠÍ MORAVSKO- SLEZSKÉHO KRAJE CARBONACEOUS PARTICLES IN THE AIR MORAVIAN-SILESIAN REGION Ing. MAREK KUCBEL Ing. Barbora SÝKOROVÁ, prof. Ing. Helena RACLAVSKÁ, CSc. Aim of this work
UHLÍKATÉ ČÁSTICE V OVZDUŠÍ MORAVSKO- SLEZSKÉHO KRAJE CARBONACEOUS PARTICLES IN THE AIR MORAVIAN-SILESIAN REGION Ing. MAREK KUCBEL Ing. Barbora SÝKOROVÁ, prof. Ing. Helena RACLAVSKÁ, CSc. Aim of this work
Just write down your most recent and important education. Remember that sometimes less is more some people may be considered overqualified.
 CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
AUXIN BINDING PROTEIN 1 (ABP1) and its role in the auxin management in plant cells
 and its role in the auxin management in plant cells") AUXIN BINDING PROTEIN 1 (ABP1) and its role in the auxin management in plant cells Department of Experimental Plant Biology, Faculty of Science Milada Čovanová Summary of Ph.D. dissertation thesis List
AUXIN BINDING PROTEIN 1 (ABP1) and its role in the auxin management in plant cells Department of Experimental Plant Biology, Faculty of Science Milada Čovanová Summary of Ph.D. dissertation thesis List
Effect of ph on the denitrifying enzyme activity in pasture soils in relation to the intrinsic differences in denitrifier communities
 University of South Bohemia in České Budějovice Faculty of Science Effect of ph on the denitrifying enzyme activity in pasture soils in relation to the intrinsic differences in denitrifier communities
University of South Bohemia in České Budějovice Faculty of Science Effect of ph on the denitrifying enzyme activity in pasture soils in relation to the intrinsic differences in denitrifier communities
THE ASSOCIATION OF SERUM BILIRUBIN AND PROMOTER VARIATIONS IN UGT1A1 WITH ATHEROSCLEROSIS
 THE ASSOCIATION OF SERUM BILIRUBIN AND PROMOTER VARIATIONS IN UGT1A1 WITH ATHEROSCLEROSIS Vítek L 1,2, Jiráskov sková A 1, Šmídová H 1, Dostálov lová G 3, Bělohlávek J 3, Kadlčkov ková L, Linhart,, A 3
THE ASSOCIATION OF SERUM BILIRUBIN AND PROMOTER VARIATIONS IN UGT1A1 WITH ATHEROSCLEROSIS Vítek L 1,2, Jiráskov sková A 1, Šmídová H 1, Dostálov lová G 3, Bělohlávek J 3, Kadlčkov ková L, Linhart,, A 3
Effect of temperature. transport properties J. FOŘT, Z. PAVLÍK, J. ŽUMÁR,, M. PAVLÍKOVA & R. ČERNÝ Č CTU PRAGUE, CZECH REPUBLIC
 Effect of temperature on water vapour transport properties J. FOŘT, Z. PAVLÍK, J. ŽUMÁR,, M. PAVLÍKOVA & R. ČERNÝ Č CTU PRAGUE, CZECH REPUBLIC Outline Introduction motivation, water vapour transport Experimental
Effect of temperature on water vapour transport properties J. FOŘT, Z. PAVLÍK, J. ŽUMÁR,, M. PAVLÍKOVA & R. ČERNÝ Č CTU PRAGUE, CZECH REPUBLIC Outline Introduction motivation, water vapour transport Experimental
Transportation Problem
 Transportation Problem ١ C H A P T E R 7 Transportation Problem The transportation problem seeks to minimize the total shipping costs of transporting goods from m origins (each with a supply s i ) to n
Transportation Problem ١ C H A P T E R 7 Transportation Problem The transportation problem seeks to minimize the total shipping costs of transporting goods from m origins (each with a supply s i ) to n
Chapter 7: Process Synchronization
 Chapter 7: Process Synchronization Background The Critical-Section Problem Synchronization Hardware Semaphores Classical Problems of Synchronization Critical Regions Monitors Synchronization in Solaris
Chapter 7: Process Synchronization Background The Critical-Section Problem Synchronization Hardware Semaphores Classical Problems of Synchronization Critical Regions Monitors Synchronization in Solaris
SEZNAM PŘÍLOH. Příloha 1 Dotazník Tartu, Estonsko (anglická verze) Příloha 2 Dotazník Praha, ČR (česká verze)... 91
 Příloha 2 Dotazník Praha, ČR (česká verze)... 91") SEZNAM PŘÍLOH Příloha 1 Dotazník Tartu, Estonsko (anglická verze)... 90 Příloha 2 Dotazník Praha, ČR (česká verze)... 91 Příloha 3 Emailové dotazy, vedení fakult TÜ... 92 Příloha 4 Emailové dotazy na vedení
SEZNAM PŘÍLOH Příloha 1 Dotazník Tartu, Estonsko (anglická verze)... 90 Příloha 2 Dotazník Praha, ČR (česká verze)... 91 Příloha 3 Emailové dotazy, vedení fakult TÜ... 92 Příloha 4 Emailové dotazy na vedení
CZ.1.07/1.5.00/
 Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
Silicified stems of upper Paleozoic plants from the Intra Sudetic and Krkonoše Piedmont basins
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Geologie Studijní obor: Paleobotanika Mgr. Václav Mencl Zkřemenělé stonky svrchnopaleozoických rostlin z vnitrosudetské a podkrkonošské
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Geologie Studijní obor: Paleobotanika Mgr. Václav Mencl Zkřemenělé stonky svrchnopaleozoických rostlin z vnitrosudetské a podkrkonošské
Air Quality Improvement Plans 2019 update Analytical part. Ondřej Vlček, Jana Ďoubalová, Zdeňka Chromcová, Hana Škáchová
 Air Quality Improvement Plans 2019 update Analytical part Ondřej Vlček, Jana Ďoubalová, Zdeňka Chromcová, Hana Škáchová vlcek@chmi.cz Task specification by MoE: What were the reasons of limit exceedances
Air Quality Improvement Plans 2019 update Analytical part Ondřej Vlček, Jana Ďoubalová, Zdeňka Chromcová, Hana Škáchová vlcek@chmi.cz Task specification by MoE: What were the reasons of limit exceedances
Exocyst subunit AtSEC15b: its role in plant cell morphogenesis and characterization of its Rab interacting partner
 CHARLES UNIVERSITY IN PRAGUE Faculty of Science Department of Experimental Plant Biology ACADEMY OF SCIENCES OF THE CZECH REPUBLIC Institute of Experimental Botany Laboratory of Cell Biology PhD Thesis
CHARLES UNIVERSITY IN PRAGUE Faculty of Science Department of Experimental Plant Biology ACADEMY OF SCIENCES OF THE CZECH REPUBLIC Institute of Experimental Botany Laboratory of Cell Biology PhD Thesis
DC circuits with a single source
 Název projektu: utomatizace výrobních procesů ve strojírenství a řemeslech egistrační číslo: Z..07/..0/0.008 Příjemce: SPŠ strojnická a SOŠ profesora Švejcara Plzeň, Klatovská 09 Tento projekt je spolufinancován
Název projektu: utomatizace výrobních procesů ve strojírenství a řemeslech egistrační číslo: Z..07/..0/0.008 Příjemce: SPŠ strojnická a SOŠ profesora Švejcara Plzeň, Klatovská 09 Tento projekt je spolufinancován
FIRE INVESTIGATION. Střední průmyslová škola Hranice. Mgr. Radka Vorlová. 19_Fire investigation CZ.1.07/1.5.00/
 FIRE INVESTIGATION Střední průmyslová škola Hranice Mgr. Radka Vorlová 19_Fire investigation CZ.1.07/1.5.00/34.0608 Výukový materiál Číslo projektu: CZ.1.07/1.5.00/21.34.0608 Šablona: III/2 Inovace a zkvalitnění
FIRE INVESTIGATION Střední průmyslová škola Hranice Mgr. Radka Vorlová 19_Fire investigation CZ.1.07/1.5.00/34.0608 Výukový materiál Číslo projektu: CZ.1.07/1.5.00/21.34.0608 Šablona: III/2 Inovace a zkvalitnění
GENERAL INFORMATION RUČNÍ POHON MANUAL DRIVE MECHANISM
 KATALOG CATALOGUE RUČNÍ POHONY PRO VENKOVNÍ PŘÍSTROJE, MONTÁŽ NA BETONOVÉ SLOUPY MANUAL DRIVE MECHANISM FOR THE ACTUATION OF OUTDOOR TYPE SWITCHING DEVICES MOUNTED ON THE CONCRETE POLES TYP RPV ISO 9001:2009
KATALOG CATALOGUE RUČNÍ POHONY PRO VENKOVNÍ PŘÍSTROJE, MONTÁŽ NA BETONOVÉ SLOUPY MANUAL DRIVE MECHANISM FOR THE ACTUATION OF OUTDOOR TYPE SWITCHING DEVICES MOUNTED ON THE CONCRETE POLES TYP RPV ISO 9001:2009
DOPLNĚK K FACEBOOK RETRO EDICI STRÁNEK MAVO JAZYKOVÉ ŠKOLY MONCHHICHI
 MONCHHICHI The Monchhichi franchise is Japanese and held by the Sekiguchi Corporation, a famous doll company, located in Tokyo, Japan. Monchhichi was created by Koichi Sekiguchi on January 25, 1974. Sekiguchi
MONCHHICHI The Monchhichi franchise is Japanese and held by the Sekiguchi Corporation, a famous doll company, located in Tokyo, Japan. Monchhichi was created by Koichi Sekiguchi on January 25, 1974. Sekiguchi
Melting the ash from biomass
 Ing. Karla Kryštofová Rožnov pod Radhoštěm 2015 Introduction The research was conducted on the ashes of bark mulch, as representatives of biomass. Determining the influence of changes in the chemical composition
Ing. Karla Kryštofová Rožnov pod Radhoštěm 2015 Introduction The research was conducted on the ashes of bark mulch, as representatives of biomass. Determining the influence of changes in the chemical composition
LOGOMANUÁL / LOGOMANUAL
 LOGOMANUÁL / LOGOMANUAL OBSAH / CONTENTS 1 LOGOTYP 1.1 základní provedení logotypu s claimem 1.2 základní provedení logotypu bez claimu 1.3 zjednodušené provedení logotypu 1.4 jednobarevné a inverzní provedení
LOGOMANUÁL / LOGOMANUAL OBSAH / CONTENTS 1 LOGOTYP 1.1 základní provedení logotypu s claimem 1.2 základní provedení logotypu bez claimu 1.3 zjednodušené provedení logotypu 1.4 jednobarevné a inverzní provedení
VY_32_INOVACE_06_Předpřítomný čas_03. Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace
 VY_32_INOVACE_06_Předpřítomný čas_03 Autor: Růžena Krupičková Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace Název projektu: Zkvalitnění ICT ve slušovské škole Číslo projektu: CZ.1.07/1.4.00/21.2400
VY_32_INOVACE_06_Předpřítomný čas_03 Autor: Růžena Krupičková Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace Název projektu: Zkvalitnění ICT ve slušovské škole Číslo projektu: CZ.1.07/1.4.00/21.2400
EU peníze středním školám digitální učební materiál
 EU peníze středním školám digitální učební materiál Číslo projektu: Číslo a název šablony klíčové aktivity: Tematická oblast, název DUMu: Autor: CZ.1.07/1.5.00/34.0515 III/2 Inovace a zkvalitnění výuky
EU peníze středním školám digitální učební materiál Číslo projektu: Číslo a název šablony klíčové aktivity: Tematická oblast, název DUMu: Autor: CZ.1.07/1.5.00/34.0515 III/2 Inovace a zkvalitnění výuky
Škola: Střední škola obchodní, České Budějovice, Husova 9. Inovace a zkvalitnění výuky prostřednictvím ICT
 Škola: Střední škola obchodní, České Budějovice, Husova 9 Projekt MŠMT ČR: EU PENÍZE ŠKOLÁM Číslo projektu: CZ.1.07/1.5.00/34.0536 Název projektu školy: Výuka s ICT na SŠ obchodní České Budějovice Šablona
Škola: Střední škola obchodní, České Budějovice, Husova 9 Projekt MŠMT ČR: EU PENÍZE ŠKOLÁM Číslo projektu: CZ.1.07/1.5.00/34.0536 Název projektu školy: Výuka s ICT na SŠ obchodní České Budějovice Šablona
Využití hybridní metody vícekriteriálního rozhodování za nejistoty. Michal Koláček, Markéta Matulová
 Využití hybridní metody vícekriteriálního rozhodování za nejistoty Michal Koláček, Markéta Matulová Outline Multiple criteria decision making Classification of MCDM methods TOPSIS method Fuzzy extension
Využití hybridní metody vícekriteriálního rozhodování za nejistoty Michal Koláček, Markéta Matulová Outline Multiple criteria decision making Classification of MCDM methods TOPSIS method Fuzzy extension
Návrh a implementace algoritmů pro adaptivní řízení průmyslových robotů
 Návrh a implementace algoritmů pro adaptivní řízení průmyslových robotů Design and implementation of algorithms for adaptive control of stationary robots Marcel Vytečka 1, Karel Zídek 2 Abstrakt Článek
Návrh a implementace algoritmů pro adaptivní řízení průmyslových robotů Design and implementation of algorithms for adaptive control of stationary robots Marcel Vytečka 1, Karel Zídek 2 Abstrakt Článek
Mikrokvadrotor: Návrh,
 KONTAKT 2011 Mikrokvadrotor: Návrh, Modelování,, Identifikace a Řízení Autor: Jaromír r Dvořák k (md( md@unicode.cz) Vedoucí: : Zdeněk Hurák (hurak@fel.cvut.cz) Katedra řídicí techniky FEL ČVUT Praha 26.5.2011
KONTAKT 2011 Mikrokvadrotor: Návrh, Modelování,, Identifikace a Řízení Autor: Jaromír r Dvořák k (md( md@unicode.cz) Vedoucí: : Zdeněk Hurák (hurak@fel.cvut.cz) Katedra řídicí techniky FEL ČVUT Praha 26.5.2011
Why PRIME? 20 years of Erasmus Programme Over 2 million students in total Annually
 What is PRIME? Problems of Recognition In Making Erasmus European-wide research project Conducted by ESN with the support of the European Commission Two editions: 1 st in 2009 Follow-up in 2010 Why PRIME?
What is PRIME? Problems of Recognition In Making Erasmus European-wide research project Conducted by ESN with the support of the European Commission Two editions: 1 st in 2009 Follow-up in 2010 Why PRIME?
Britské společenství národů. Historie Spojeného království Velké Británie a Severního Irska ročník gymnázia (vyšší stupeň)
") Název vzdělávacího materiálu: Číslo vzdělávacího materiálu: Autor vzdělávací materiálu: Období, ve kterém byl vzdělávací materiál vytvořen: Vzdělávací oblast: Vzdělávací obor: Vzdělávací předmět: Tematická
Název vzdělávacího materiálu: Číslo vzdělávacího materiálu: Autor vzdělávací materiálu: Období, ve kterém byl vzdělávací materiál vytvořen: Vzdělávací oblast: Vzdělávací obor: Vzdělávací předmět: Tematická
Klepnutím lze upravit styl předlohy. nadpisů. nadpisů.
 1/ 13 Klepnutím lze upravit styl předlohy Klepnutím lze upravit styl předlohy www.splab.cz Soft biometric traits in de identification process Hair Jiri Prinosil Jiri Mekyska Zdenek Smekal 2/ 13 Klepnutím
1/ 13 Klepnutím lze upravit styl předlohy Klepnutím lze upravit styl předlohy www.splab.cz Soft biometric traits in de identification process Hair Jiri Prinosil Jiri Mekyska Zdenek Smekal 2/ 13 Klepnutím
Tabulka 1 Stav členské základny SK Praga Vysočany k roku 2015 Tabulka 2 Výše členských příspěvků v SK Praga Vysočany Tabulka 3 Přehled finanční
 Příloha I Seznam tabulek Tabulka 1 Stav členské základny SK Praga Vysočany k roku 2015 Tabulka 2 Výše členských příspěvků v SK Praga Vysočany Tabulka 3 Přehled finanční odměny pro rozhodčí platný od roku
Příloha I Seznam tabulek Tabulka 1 Stav členské základny SK Praga Vysočany k roku 2015 Tabulka 2 Výše členských příspěvků v SK Praga Vysočany Tabulka 3 Přehled finanční odměny pro rozhodčí platný od roku
SPECIAL THEORY OF RELATIVITY
 SPECIAL THEORY OF RELATIVITY 1. Basi information author Albert Einstein phenomena obsered when TWO frames of referene moe relatie to eah other with speed lose to the speed of light 1905 - speial theory
SPECIAL THEORY OF RELATIVITY 1. Basi information author Albert Einstein phenomena obsered when TWO frames of referene moe relatie to eah other with speed lose to the speed of light 1905 - speial theory
místo, kde se rodí nápady
 místo, kde se rodí nápady a private european network of information centres on materials and innovative products. Created in 2001 in Paris, it provides members with a large selection of specific, reproducible
místo, kde se rodí nápady a private european network of information centres on materials and innovative products. Created in 2001 in Paris, it provides members with a large selection of specific, reproducible
Dynamic Development of Vocabulary Richness of Text. Miroslav Kubát & Radek Čech University of Ostrava Czech Republic
 Dynamic Development of Vocabulary Richness of Text Miroslav Kubát & Radek Čech University of Ostrava Czech Republic Aim To analyze a dynamic development of vocabulary richness from a methodological point
Dynamic Development of Vocabulary Richness of Text Miroslav Kubát & Radek Čech University of Ostrava Czech Republic Aim To analyze a dynamic development of vocabulary richness from a methodological point
DATA SHEET. BC516 PNP Darlington transistor. technický list DISCRETE SEMICONDUCTORS Apr 23. Product specification Supersedes data of 1997 Apr 16
 zákaznická linka: 840 50 60 70 DISCRETE SEMICONDUCTORS DATA SHEET book, halfpage M3D186 Supersedes data of 1997 Apr 16 1999 Apr 23 str 1 Dodavatel: GM electronic, spol. s r.o., Křižíkova 77, 186 00 Praha
zákaznická linka: 840 50 60 70 DISCRETE SEMICONDUCTORS DATA SHEET book, halfpage M3D186 Supersedes data of 1997 Apr 16 1999 Apr 23 str 1 Dodavatel: GM electronic, spol. s r.o., Křižíkova 77, 186 00 Praha
STUDY EDITS FOR BETTER TRANSPORT IN THE CENTRE OF NÁCHOD
 CZECH TECHNICAL UNIVERSITY IN PRAGUE Faculty of transportation sciences Title of project STUDY EDITS FOR BETTER TRANSPORT IN THE CENTRE OF NÁCHOD 2006 Petr Kumpošt Basic information about town Náchod Náchod
CZECH TECHNICAL UNIVERSITY IN PRAGUE Faculty of transportation sciences Title of project STUDY EDITS FOR BETTER TRANSPORT IN THE CENTRE OF NÁCHOD 2006 Petr Kumpošt Basic information about town Náchod Náchod
Fourth School Year PISTON MACHINES AND PISTON COMPRESSORS
 Fourth School Year PISTON MACHINES AND PISTON COMPRESSORS 1. Piston machines Piston machines are classified as machines working with volume changes. It means that the working medium is closed by a in a
Fourth School Year PISTON MACHINES AND PISTON COMPRESSORS 1. Piston machines Piston machines are classified as machines working with volume changes. It means that the working medium is closed by a in a
Stojan pro vrtačku plošných spojů
 Střední škola průmyslová a hotelová Uherské Hradiště Kollárova 617, Uherské Hradiště Stojan pro vrtačku plošných spojů Závěrečný projekt Autor práce: Koutný Radim Lukáš Martin Janoštík Václav Vedoucí projektu:
Střední škola průmyslová a hotelová Uherské Hradiště Kollárova 617, Uherské Hradiště Stojan pro vrtačku plošných spojů Závěrečný projekt Autor práce: Koutný Radim Lukáš Martin Janoštík Václav Vedoucí projektu:
UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ DISERTAČNÍ PRÁCE
 UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ DISERTAČNÍ PRÁCE 2009 Mgr. Petra Horáková UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ Katedra analytické chemie Elektrochemická analýza nukleotidových
UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ DISERTAČNÍ PRÁCE 2009 Mgr. Petra Horáková UNIVERZITA PARDUBICE FAKULTA CHEMICKO-TECHNOLOGICKÁ Katedra analytické chemie Elektrochemická analýza nukleotidových
Energy vstupuje na trh veterinárních produktů Energy enters the market of veterinary products
 Energy news2 1 Energy vstupuje na trh veterinárních produktů Energy enters the market of veterinary products Doposud jste Energy znali jako výrobce a dodavatele humánních přírodních doplňků stravy a kosmetiky.
Energy news2 1 Energy vstupuje na trh veterinárních produktů Energy enters the market of veterinary products Doposud jste Energy znali jako výrobce a dodavatele humánních přírodních doplňků stravy a kosmetiky.
On large rigid sets of monounary algebras. D. Jakubíková-Studenovská P. J. Šafárik University, Košice, Slovakia
 On large rigid sets of monounary algebras D. Jakubíková-Studenovská P. J. Šafárik University, Košice, Slovakia coauthor G. Czédli, University of Szeged, Hungary The 54st Summer School on General Algebra
On large rigid sets of monounary algebras D. Jakubíková-Studenovská P. J. Šafárik University, Košice, Slovakia coauthor G. Czédli, University of Szeged, Hungary The 54st Summer School on General Algebra
The tension belt serves as a tension unit. After emptying the belt is cleaned with a scraper.
 Second School Year BELT AND WORM CONVEYORS They are machines for transporting piece or loose materials even for great distances. In loaders and unloaders it is not necessary to stop the conveyor. The transport
Second School Year BELT AND WORM CONVEYORS They are machines for transporting piece or loose materials even for great distances. In loaders and unloaders it is not necessary to stop the conveyor. The transport
Database systems. Normal forms
 Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49
 Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
Enabling Intelligent Buildings via Smart Sensor Network & Smart Lighting
 Enabling Intelligent Buildings via Smart Sensor Network & Smart Lighting Petr Macháček PETALIT s.r.o. 1 What is Redwood. Sensor Network Motion Detection Space Utilization Real Estate Management 2 Building
Enabling Intelligent Buildings via Smart Sensor Network & Smart Lighting Petr Macháček PETALIT s.r.o. 1 What is Redwood. Sensor Network Motion Detection Space Utilization Real Estate Management 2 Building
Anatomie, histologie a embryologie
 Anatomie, histologie a embryologie Témata: - Kořen, kořenový vrchol, kořenový meristém - Základní radiální a longitudinální zonace kořene - Primární růst kořene a sekundární tloustnutí - Zakládání a vývin
Anatomie, histologie a embryologie Témata: - Kořen, kořenový vrchol, kořenový meristém - Základní radiální a longitudinální zonace kořene - Primární růst kořene a sekundární tloustnutí - Zakládání a vývin
Úloha cytoskeletu v morfogenezi rostlinných buněk
 Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Shrnutí dizertační práce Úloha cytoskeletu v morfogenezi rostlinných buněk Lenka Havelková Vedoucí dizertační práce:
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Shrnutí dizertační práce Úloha cytoskeletu v morfogenezi rostlinných buněk Lenka Havelková Vedoucí dizertační práce:
CZ.1.07/1.5.00/
 Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
1, Žáci dostanou 5 klíčových slov a snaží se na jejich základě odhadnout, o čem bude následující cvičení.
 Moje hlavní město Londýn řešení: 1, Žáci dostanou 5 klíčových slov a snaží se na jejich základě odhadnout, o čem bude následující cvičení. Klíčová slova: capital, double decker bus, the River Thames, driving
Moje hlavní město Londýn řešení: 1, Žáci dostanou 5 klíčových slov a snaží se na jejich základě odhadnout, o čem bude následující cvičení. Klíčová slova: capital, double decker bus, the River Thames, driving
Eurogranites 2015 Variscan Plutons of the Bohemian Massif
 Accommodation & meeting point 1: Hotel Luník on Friday, 3 July, 17.30 (1) Prague public transport (MHD) by red metro line C get off at I. P. Pavlova station further on foot according to map below, ~ 5
Accommodation & meeting point 1: Hotel Luník on Friday, 3 July, 17.30 (1) Prague public transport (MHD) by red metro line C get off at I. P. Pavlova station further on foot according to map below, ~ 5
Vliv metody vyšetřování tvaru brusného kotouče na výslednou přesnost obrobku
 Vliv metody vyšetřování tvaru brusného kotouče na výslednou přesnost obrobku Aneta Milsimerová Fakulta strojní, Západočeská univerzita Plzeň, 306 14 Plzeň. Česká republika. E-mail: anetam@kto.zcu.cz Hlavním
Vliv metody vyšetřování tvaru brusného kotouče na výslednou přesnost obrobku Aneta Milsimerová Fakulta strojní, Západočeská univerzita Plzeň, 306 14 Plzeň. Česká republika. E-mail: anetam@kto.zcu.cz Hlavním
Unit 3 Stereochemistry
 Unit 3 Stereochemistry Stereoisomers hirality (R) and (S) Nomenclature Depicting Asymmetric arbons Diastereomers Fischer Projections Stereochemical Relationships Optical Activity Resolution of Enantiomers
Unit 3 Stereochemistry Stereoisomers hirality (R) and (S) Nomenclature Depicting Asymmetric arbons Diastereomers Fischer Projections Stereochemical Relationships Optical Activity Resolution of Enantiomers
[ 1 ] PRAC. Perspective from a Member State. MUDr. Jana Mladá 2015 Státní ústav pro kontrolu léčiv
![[ 1 ] PRAC. Perspective from a Member State. MUDr. Jana Mladá 2015 Státní ústav pro kontrolu léčiv](/thumbs/83/87566435.jpg "[ 1 ] PRAC. Perspective from a Member State. MUDr. Jana Mladá 2015 Státní ústav pro kontrolu léčiv") [ 1 ] PRAC Perspective from a Member State [ 2 ] Today EU regulatory drug safety network PRAC conclusions and recommendations at national level Modern history of pharmacovigilance in Czech Republic [ 3
[ 1 ] PRAC Perspective from a Member State [ 2 ] Today EU regulatory drug safety network PRAC conclusions and recommendations at national level Modern history of pharmacovigilance in Czech Republic [ 3
TELEGYNEKOLOGIE TELEGYNECOLOGY
 TELEGYNEKOLOGIE TELEGYNECOLOGY Račanská E. 1, Huser M. 1, Schwarz D. 2, Šnábl I. 2, Ventruba P. 1 1) Gynekologicko porodnická klinika LF MU a FN Brno 2) Institut biostatistiky a analýz LF a PřF MU Abstrakt
TELEGYNEKOLOGIE TELEGYNECOLOGY Račanská E. 1, Huser M. 1, Schwarz D. 2, Šnábl I. 2, Ventruba P. 1 1) Gynekologicko porodnická klinika LF MU a FN Brno 2) Institut biostatistiky a analýz LF a PřF MU Abstrakt
MEMOSPRAY INTELIGENTNÍ TRYSKOVÝ KOMPLET PRO TECHNOLOGIE POVRCHOVÝCH ÚPRAV MATERIÁLŮ. Martin Pavliska
 MEMOSPRAY INTELIGENTNÍ TRYSKOVÝ KOMPLET PRO TECHNOLOGIE POVRCHOVÝCH ÚPRAV MATERIÁLŮ Martin Pavliska Hennlich Industrietechnik, spol. s r.o.,českolipská 9, 412 01 Litoměřice, ČR, pavliska@hennlich.cz Abstrakt
MEMOSPRAY INTELIGENTNÍ TRYSKOVÝ KOMPLET PRO TECHNOLOGIE POVRCHOVÝCH ÚPRAV MATERIÁLŮ Martin Pavliska Hennlich Industrietechnik, spol. s r.o.,českolipská 9, 412 01 Litoměřice, ČR, pavliska@hennlich.cz Abstrakt
Národní informační den společných technologických iniciativ ARTEMIS a ENIAC
 Národní informační den společných technologických iniciativ ARTEMIS a ENIAC 21. března 2011, Praha Pravidla a podmínky účasti v projektech ARTEMIS a ENIAC v ČR Úvod k finančním pravidlům JTIs (ARTEMIS
Národní informační den společných technologických iniciativ ARTEMIS a ENIAC 21. března 2011, Praha Pravidla a podmínky účasti v projektech ARTEMIS a ENIAC v ČR Úvod k finančním pravidlům JTIs (ARTEMIS
Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY. Číslo Servisní Informace Mechanika: 5-2013
 Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY Servisní Informace Datum vydání: 20.2.2013 Určeno pro : AMS, registrované subj.pro montáž st.měř. Na základě SI VDO č./datum: Není Mechanika
Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY Servisní Informace Datum vydání: 20.2.2013 Určeno pro : AMS, registrované subj.pro montáž st.měř. Na základě SI VDO č./datum: Není Mechanika
Škola: Střední škola obchodní, České Budějovice, Husova 9. Inovace a zkvalitnění výuky prostřednictvím ICT
 Škola: Střední škola obchodní, České Budějovice, Husova 9 Projekt MŠMT ČR: EU PENÍZE ŠKOLÁM Číslo projektu: CZ.1.07/1.5.00/34.0536 Název projektu školy: Výuka s ICT na SŠ obchodní České Budějovice Šablona
Škola: Střední škola obchodní, České Budějovice, Husova 9 Projekt MŠMT ČR: EU PENÍZE ŠKOLÁM Číslo projektu: CZ.1.07/1.5.00/34.0536 Název projektu školy: Výuka s ICT na SŠ obchodní České Budějovice Šablona
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49
 Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky
 Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
WYSIWYG EDITOR PRO XML FORM
 WYSIWYG EDITOR PRO XML FORM Ing. Tran Thanh Huan, Ing. Nguyen Ba Nghien, Doc. Ing. Josef Kokeš, CSc Abstract: In this paper, we introduce the WYSIWYG editor pro XML Form. We also show how to create a form
WYSIWYG EDITOR PRO XML FORM Ing. Tran Thanh Huan, Ing. Nguyen Ba Nghien, Doc. Ing. Josef Kokeš, CSc Abstract: In this paper, we introduce the WYSIWYG editor pro XML Form. We also show how to create a form
The Military Technical Institute
 The Military Technical Institute The security is essential, the security is a challenge Jitka Čapková Petr Kozák Vojenský technický ústav, s.p. The Czech Republic security legislation applicable to CIS:
The Military Technical Institute The security is essential, the security is a challenge Jitka Čapková Petr Kozák Vojenský technický ústav, s.p. The Czech Republic security legislation applicable to CIS:
Fytomineral. Inovace Innovations. Energy News 04/2008
 Energy News 4 Inovace Innovations 1 Fytomineral Tímto Vám sdělujeme, že již byly vybrány a objednány nové lahve a uzávěry na produkt Fytomineral, které by měly předejít únikům tekutiny při přepravě. První
Energy News 4 Inovace Innovations 1 Fytomineral Tímto Vám sdělujeme, že již byly vybrány a objednány nové lahve a uzávěry na produkt Fytomineral, které by měly předejít únikům tekutiny při přepravě. První
CZ.1.07/1.5.00/34.0527
 Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
Projekt: Příjemce: Digitální učební materiály ve škole, registrační číslo projektu CZ.1.07/1.5.00/34.0527 Střední zdravotnická škola a Vyšší odborná škola zdravotnická, Husova 3, 371 60 České Budějovice
Kvalita cigaretového tabáku v ČR- marketingové šetření se zaměřením na konečného spotřebitele
 Kvalita cigaretového tabáku v ČR- marketingové šetření se zaměřením na konečného spotřebitele Diplomová práce Bc. Eva Švejdová Vysoká škola hotelová v Praze 8, spol. s r. o. Katedra Hotelnictví Studijní
Kvalita cigaretového tabáku v ČR- marketingové šetření se zaměřením na konečného spotřebitele Diplomová práce Bc. Eva Švejdová Vysoká škola hotelová v Praze 8, spol. s r. o. Katedra Hotelnictví Studijní
2N Voice Alarm Station
 2N Voice Alarm Station 2N Lift1 Installation Manual Version 1.0.0 www.2n.cz EN Voice Alarm Station Description The 2N Voice Alarm Station extends the 2N Lift1/ 2N SingleTalk with an audio unit installed
2N Voice Alarm Station 2N Lift1 Installation Manual Version 1.0.0 www.2n.cz EN Voice Alarm Station Description The 2N Voice Alarm Station extends the 2N Lift1/ 2N SingleTalk with an audio unit installed
Výukový materiál v rámci projektu OPVK 1.5 Peníze středním školám
 VY_22_INOVACE_AJOP40764ČER Výukový materiál v rámci projektu OPVK 1.5 Peníze středním školám Číslo projektu: Název projektu: Číslo šablony: CZ.1.07/1.5.00/34.0883 Rozvoj vzdělanosti II/2 Datum vytvoření:
VY_22_INOVACE_AJOP40764ČER Výukový materiál v rámci projektu OPVK 1.5 Peníze středním školám Číslo projektu: Název projektu: Číslo šablony: CZ.1.07/1.5.00/34.0883 Rozvoj vzdělanosti II/2 Datum vytvoření:
ROLZ-2. Portable AV/Conference Center. Assembly Instructions
 1 ROLZ-2 Portable AV/Conference Center Assembly Instructions Rolz-2 Portable AV/Conference Center Part Drawing Description Qty Part Drawing Description Qty Hardware List A 1 ½ Flat Head Screw 2 EA P-1
1 ROLZ-2 Portable AV/Conference Center Assembly Instructions Rolz-2 Portable AV/Conference Center Part Drawing Description Qty Part Drawing Description Qty Hardware List A 1 ½ Flat Head Screw 2 EA P-1
EXACT DS OFFICE. The best lens for office work
 EXACT DS The best lens for office work EXACT DS When Your Glasses Are Not Enough Lenses with only a reading area provide clear vision of objects located close up, while progressive lenses only provide
EXACT DS The best lens for office work EXACT DS When Your Glasses Are Not Enough Lenses with only a reading area provide clear vision of objects located close up, while progressive lenses only provide
Dynamic programming. Optimal binary search tree
 The complexity of different algorithms varies: O(n), Ω(n ), Θ(n log (n)), Dynamic programming Optimal binary search tree Různé algoritmy mají různou složitost: O(n), Ω(n ), Θ(n log (n)), The complexity
The complexity of different algorithms varies: O(n), Ω(n ), Θ(n log (n)), Dynamic programming Optimal binary search tree Různé algoritmy mají různou složitost: O(n), Ω(n ), Θ(n log (n)), The complexity
VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL.S R.O.
 VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL.S R.O. Bc. Nina Baťková Ovlivňuje reklama způsob ţivota společnosti? Diplomová práce 2014 Ovlivňuje reklama způsob ţivota společnosti? Diplomová práce Bc. Nina Baťková
VYSOKÁ ŠKOLA HOTELOVÁ V PRAZE 8, SPOL.S R.O. Bc. Nina Baťková Ovlivňuje reklama způsob ţivota společnosti? Diplomová práce 2014 Ovlivňuje reklama způsob ţivota společnosti? Diplomová práce Bc. Nina Baťková
Friction drives have constant or variable drives (it means variators). Friction drives are used for the transfer of smaller outputs.
. Friction drives are used for the transfer of smaller outputs.") Third School Year FRICTION DRIVES 1. Introduction In friction drives the peripheral force between pressed wheels is transferred by friction. To reach peripheral forces we need both a pressed force and
Third School Year FRICTION DRIVES 1. Introduction In friction drives the peripheral force between pressed wheels is transferred by friction. To reach peripheral forces we need both a pressed force and
Aplikovaná chemie a biochemie. Přednáška č. 3. Proteinové techniky (2)
") Aplikovaná chemie a biochemie Přednáška č. 3 Proteinové techniky (2) Proteiny (2): funkce jednotlivých domén; posttranslační modifikace (fosforylace, acetylace, glykosylace); regulace odborávání proteinu
Aplikovaná chemie a biochemie Přednáška č. 3 Proteinové techniky (2) Proteiny (2): funkce jednotlivých domén; posttranslační modifikace (fosforylace, acetylace, glykosylace); regulace odborávání proteinu