Základy biochemie KBC/BCH. Fotosyntéza. Inovace studia biochemie prostřednictvím e-learningu CZ / /0407
|
|
|
- Františka Horáčková
- před 8 lety
- Počet zobrazení:
Transkript
1 Základy biochemie KB/B Fotosyntéza Inovace studia biochemie prostřednictvím e-learningu Z / /0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky.
2 snova Úvod hloroplasty Světlo absorbující pigmenty Světelná reakce alvinův - Bensonův cyklus (RUBIS) Fotorespirace Rostliny 4 a AM
3 Fotosyntéza - úvod Fotosyntéza fixuje ročně asi tun uhlíku, což reprezentuje kj energie. Fotosyntéza u rostlin je dvoustupňový proces, při kterém se světelná energie využívá k oxidaci 2 : [ ] Současně získané elektrony se využívají k redukci 2 : 4 [ ] + 2 ( 2 ) + 2 Dva stupně fostosyntézy se tradičně uvádí jako světelná fáze a temnostní fáze. Místem fotosyntézy u eukaryot (vyšší rostliny a řasy) jsou chloroplasty. Buňky obsahují 1 až 1000 chloroplastů tvaru elipsoidu o rozměrech asi 5 µm délky.

4 hloroplast kukuřice Vnější membrána Lamela Vnitřní membrána Mezimembránový prostor Grana Stroma Prostor thylakoidu

5 Fotosyntetizující cyanobakterie Anabaena (zvětšení 450 x). Předkové, kteří se evolučně přetvořili na současné chloroplasty.
6 Světlo absorbující pigmenty hlorofyl - cyklický tetrapyrrol s centrálním atomem Mg 2+. Prekurzorem je stejně jako u hemu protoporfyrin IX. lavní formy chlorofylu u rostlin a cyanobakterií jsou: chlorofyl a (hl a) a chlorofyl b (hl b), u fotosyntetizujících bakterií jsou to bakteriochlorofyl a (Bhl a) a bakteriochlorofyl b (Bhl b). hlorofyly jako vysoce konjugované sloučeniny silně absorbují viditelné světlo. Malé rozdíly v chemické struktuře silně ovlivňují jejich spektra. Fotosystémy I a II, obsahující mimo jiné chlorofyly, jsou lokalizovány v thylakoidní membráně chloroplastů. Fotosystém I (PS 700) je transmembránový multipodjednotkový komplex (10), absorbuje světlo kratší vlnové délky než 700 nm. Vytváří NADP. Fotosystém II (PS 680), transmembránový komplex 20 podjednotek, přenos elektronů z vody na plastochinon (plastochinol). Absorbuje světlo vlnových délek kratších než 680 nm. ytochrom bf (obdoba komplexu III mitochondrií!!!). ytochrom bf generuje protonový gradient přes thylakoidní membránu, což vede ke tvorbě ATP.
7 Struktury chlorofylů 2 R 1 R I N N + 3 II 4 R 3 3 N N Mg 2+ + Fe 2+ 3 IV N N III 3 3 N N 3 2 V R hlorofyl (Fe-protoporfyrin IX) R 1 R 2 R 3 R 4 hlorofyl a hlorofyl b Bakteriochlorofyl a Bakteriochlorofyl b a a a a 3 P P P nebo G P P = G = 2 2 Fytylový postranní řetězec Geranylový postranní řetězec a značí, že mezi 3 a 4 se nevyskytuje dvojná vazba.

8 Absorpční spektrum chlorofylu a
9 Absorpční spektra různých fotosyntetických pigmentů hlorofyl b Sluneční spektrum Absorbance hlorofyl a Karotenoidy Fykoerythrin Fykocyanin Vlnová délka (nm)
10 Světlosběrné komplexy Světlosběrné komplexy obsahují fotosyntetická reakční centra místa, kde se odehrává primární reakce světelné fáze fotosyntézy a anténní komplex obsahující další chlorofyly a přídavné pigmenty jako jsou karotenoidy, např. β-karoten β-karoten 3 3
11 Fotosyntetický anténní komplex. Zachycené fotony náhodně migrují anténním komplexem až vstoupí do reakčního centra (tmavě zelené). Foton Foton Foton Foton
12 Světelné reakce Elektrony excitovaná molekula může uvolnit excitační energii několika způsoby: a)vnitřní konverze na kinetickou energii, pohyb molekul, teplo. U molekul chlorofylu k těmto stavům nedochází. b)fluorescence, je emitován foton o vyšší vlnové délce (nižší energie). U rostlin jen asi 3 až 6 % absorbované světelné energie. c)excitační transfer, excitovaná molekula přenáší excitační energii na sousední molekuly chlorofylu s nižší excitační energií než anténní chlorofyly. d)fotooxidace, světlem excitovaná donorová molekula je oxidována přenosem elektronu na akceptorovou molekulu. Takovým donorem je excitovaný chlorofyl. Fotooxidovaný chlorofyl, kationtový radikál, se vrací do původního stavu oxidací jiné molekuly.
13 Reakční centrum Fotoindukované oddělení nábojů probíhá v reakčním centru. Elektrony extrahované z počáteční sloučeniny jsou excitované a představují redukční sílu mohou redukovat další sloučeniny a skladovat tak světelnou energii ve formě chemických sloučenin.

14 Struktura fostosystému II. D1 a D2 podjednotky (32kd) navázány dva chlorofyly (speciální, také P680)-přenos elektronů na feofytin, poté na plastochinony. Feofytin chlorofyl se dvěma + namísto centrálního Mg 2+. elková reakce: 2 Q Q 2 Stroma D2 D1 Dutina thylakoidu Specifický pár chlorofylů Manganové centrum

15 Struktura fotosystému I. Dvě podjednotky psaa a psab jsou žlutě a části obdobné PS II jsou červeně a modře, chlorofyly včetně speciálního páru zeleně, vyznačeny klastry Fe-S. 4Fe-4S klastry Stroma Dutina thylakoidu Specifický pár chlorofylů Pohled shora
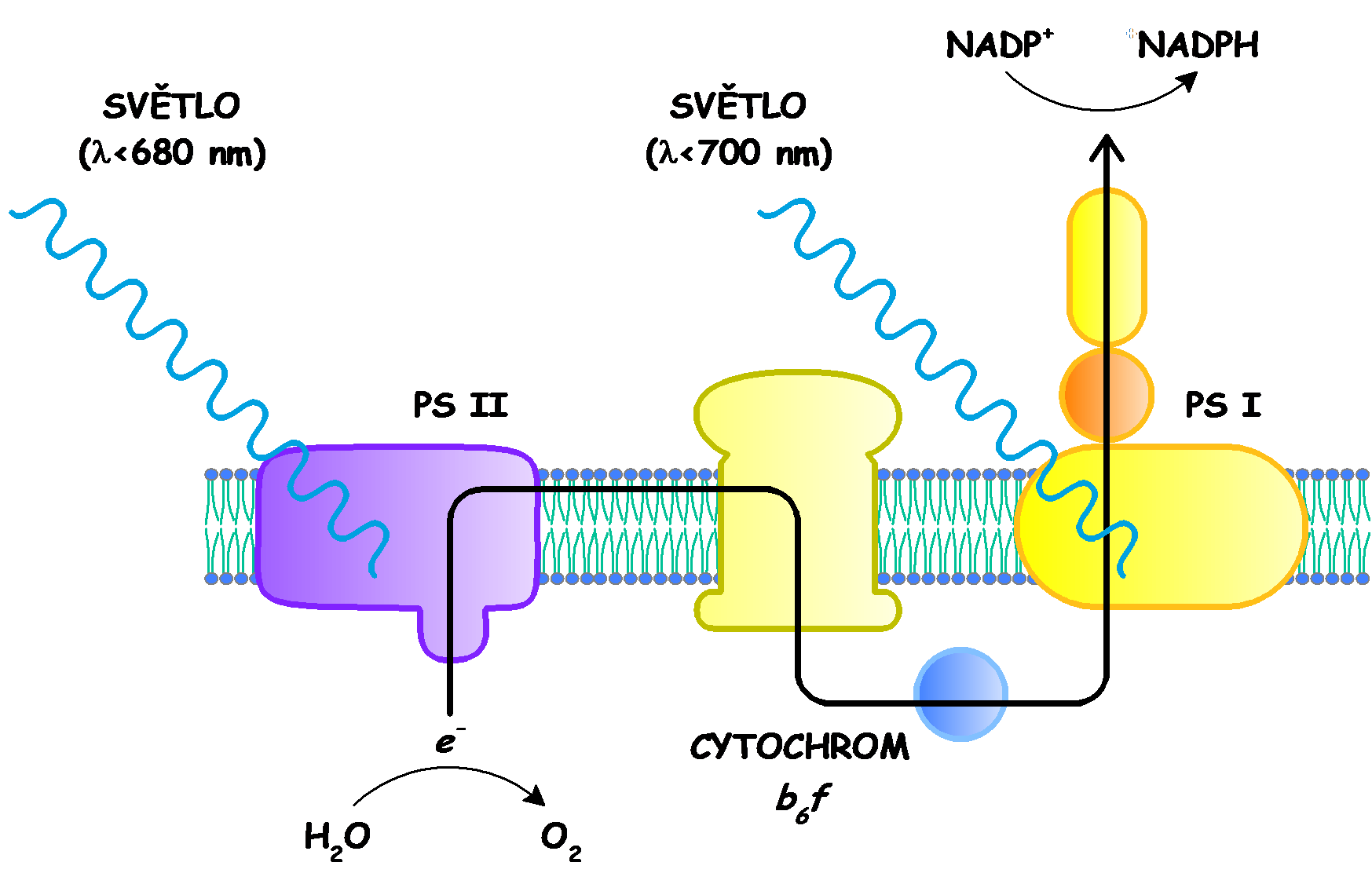
16 Dva fotosystémy
17 Fotolýza vody tvorba kyslíku Součástí PS II (P680, fotosystém II) je centrum uvolňující kyslík (oxygen evolving center - E). V tomto komplexním útvaru dochází působením světelných kvant (fotonů) k rozkladu molekul vody na kyslík, čtyři protony a čtyři elektrony e - Elektrony uvolněné z vody vstupují do PSII, kde jsou světelnými kvanty uvedeny excitací do vyšších energetických hladin označený P680 * a posléze putují řetězcem přes Q 2.
18 Pravděpodobné schéma uvolnění kyslíku ze dvou molekul vody v Mn centru lavní roli hrají 4 ionty Mn v různém oxidačním stavu, vážící silně kyslíkaté sloučeniny, jeden a 2+ a l -. S 0 až S 4 vždy jednoelektronová oxidace elektrony vstupují do PSII.. + a l Mn + Mn 3+ S 0 Mn 4+ e -, + + a l Mn Mn 3+ S 1 Mn 4+ Mn 4+ Mn 4+
19 Pravděpodobné schéma uvolnění kyslíku v Mn centru a + + S 1 e - a + + S 2 l + 3+ Mn Mn 3+ Mn 4+ l + 3+ Mn Mn 4+ Mn 4+ Mn 4+ Mn 4+
20 Pravděpodobné schéma uvolnění kyslíku v Mn centru a + + S 2 e -, + a + S 3 l + 3+ Mn Mn 4+ Mn 4+ l + 4+ Mn Mn 4+ Mn 4+ Mn 4+ Mn 4+
21 Pravděpodobné schéma uvolnění kyslíku v Mn centru a l + 4+ Mn Mn 4+ S 3 Mn 4+ e -, + + a l + 5+ Mn + Mn 4+ S 4 Mn 4+ Mn 4+ Mn 4+
22 Pravděpodobné schéma uvolnění kyslíku v Mn centru + S 4 + S 4 a l + 5+ Mn + Mn 4+ Mn 4+ a l + 5+ Mn + Mn 4+ Mn 4+ Mn 4+ Mn 4+
23 Pravděpodobné schéma uvolnění kyslíku v Mn centru K uvolnění 2 je nutná oxidace a l + 5+ Mn + Mn 4+ S 4 Mn a l Mn + Mn 3+ S 0 Mn 4+ Mn 4+ Mn 4+
24 Plastochinon (Q) plastochinol (Q 2 ) (n = 6-10) n Plastochinon 2[ ] n Plastochinol
25 Z-schéma světelné fáze fotosyntézy Elektrony mohou dosáhnout na ferredoxin-nadp + reduktasu (FNR), která katalyzuje redukci NADP +. xidace vody a tok elektronů přes Q cyklus generuje transmembránový protonový gradient s vyšší koncentrací protonů na straně thylakoidní dutiny (lumen - uvnitř). Energie gradientu se uplatňuje při tvorbě ATP. Různé prosthetické skupiny fotosyntetického aparátu rostlin mohou být seřazeny podle redukčního potenciálu v diagramu zvaném Z-schéma. Elektrony uvolněné z P680 po absorpci fotonů jsou nahrazovány elektrony z fotolýzy vody. Každý elektron prochází řetězcem přenašečů do hotovosti plastochinonových molekul. Vytvořený redukovaný plastochinol redukuje cytochrom b 6 f komplex za současného transportu protonů do thylakoidů. ytochrom b 6 f přenáší elektrony na plastocyanin (P) a ten regeneruje fotooxidovaný P700 v PSI. Elektrony emitované z PSI redukují přes řetězec přenašečů NADP + na NADP. Necyklický proces. V cyklickém procesu se elektrony vrací zpět na cytochrom b 6 f a přitom se přenášejí další protony do thylakoidu.
26 Diagram Z-schéma PSI -1.4 P700* A 0 A1 F X F A Necyklická cesta PSII P680* hl a Pheo a yklická cesta F B Fd e - 2 NADP FNR (stroma) -0.4 hν 2 NADP -0.2 Q A Q B 8 + (stroma) cyt b 6 ξ (V) 0 Q PL Manganový komplex hν 8 + (thylakoid) Q [4Fe-4S] cyt f ytochrom b 6 f P P e - S (thylakoid) S 4 Z +1.2 P680
27 ytochrom b 6 f komplex a plastocyanin (P) ytochrom b 6 f komplex je integrální membránový proteinový komplex obsahující řadu složek analogicky jako např. mitochondriální komplex III a cytochrom bc 1. Elektrony z cytochrom b 6 f komplexu putují Q cyklem (viz mitochondrie!!). Při transportu jednoho elektronu se translokují dva protony do thylakoidu. Tedy čtyři elektrony z 2 2 vedou k translokaci 8 protonů. Plastocyanin (P) periferní membránový protein (99 aminokyselin) - člen skupiny modrých měďnatých proteinů lokalizovaný na vnitřní straně thylakoidů. xidační stavy redoxních center cyklují mezi u 2+ a u + stavy.
28 Lokalizace systémů PSII, PSI, cytochromu b 6 f a ATP-synthasy na thylakoidech Fotosystém PSI ATP-synthasa Volná membrána (stromatální lamela) Fotosystém PSII ytochrom b 6 f Převrstvená membrána (granum)
29 Lokalizace systémů PSII, PSI, cytochromu b 6 f a ATP-synthasy PSI se nachází na neuzavřených stromatálních lamelách v dotyku se stromatem, kde je k dosažení NADP +. PSII je lokalizován většinou mezi uzavřenými složenými grany, mimo přímý kontakt se stromatem. ytochrom b 6 f je rovnoměrně distribuován v celé membráně. ATPasa je lokalizována na vnější straně membrány.
30 Směr protonového gradientu fotosystému II Stroma (vysoké p) Q 2 2 Q Lumen thylakoidu (nízké p)


31 ytochrom bf V každém Q cyklu jsou přes cytochrom bf transportovány 4 protony. 1. část: Q Pc(u 2+ ) = Q + 2 Pc (u + ) (v thylakoidu) 2. část: yt.bf (redukuje) + Q (z poolu) = Q 2 (protony ze stromatu) Po reoxidaci Q 2 se protony uvolní do thylakoidu.
32 Jagedorfův pokus. Syntéza ATP jako důsledek p gradientu. Membrána thylakoidu ATP ADP + P i p 7 p 4 p 4 + p 8 Inkubace po dobu několika hodin Náhlá změna externího p a přídavek ADP a P i p 4 p 4


33 Srovnání fotosyntézy a oxidativní fosforylace
.")
34 Fotosystém I využívá světelnou energii k tvorbě redukovaného ferredoxinu silného redukčního činidla (obsahuje 2Fe-2S). Elektrony jsou přenášeny na ferredoxin-nadp reduktasu.
35 Tok elektronů fotosystémem I na ferredoxin. Pozitivní náboj P700 se doplňuje elektrony z plastocyaninu Ferredoxin 4Fe-4S hinon (A 1 ) P700 hlorofyl (A 0 ) Plastocyanin
36 Dráha elektronů z vody na NADP + při fotosyntéze

37 Struktura ferredoxin-nadp + reduktasy Flavin Vazebné místo NADP + Ferredoxin
38 Ferredoxin NADP + reduktasa - mechanismus + + Fd red + + Fd red Fd ox Fd ox FAD FAD FAD 2 Semichinonová forma NADP + + NADP +
39 Energetický výtěžek světelné (primární) fáze fotosyntézy Absorpce 4 fotonů PSII generuje 1 molekulu 2 a uvolní se 4 protony do dutiny thylakoidu. Dvě molekuly plastochinolu jsou oxidovány komplexem bf za uvolnění 8 protonů do dutiny thylakoidu (lumen). Elektrony ze 4 molekul redukovaného plastocyaninu jsou za účasti dalších 4 absorbovaných fotonů předány na ferredoxin. Čtyři molekuly redukovaného ferredoxinu generují 2 molekuly NADP. elková reakce tvorby NADP: NADP stroma = NADP lumen Dvanáct protonů v lumen vstupuje do catpsynthasy (chloroplastová). Tvoří se celkem 3 molekuly ATP. elková reakce fotofosforylace: NADP stroma = NADP lumen 3 ADP P i lumen = 3 ATP stroma 2 NADP ADP + 3 P i + + = NADP + 3 ATP + 2
40 yklická fotofosforylace yklická fotofosforylace je alternativní drahou fotosyntézy. Elektrony redukovaného ferredoxinu mohou být převedeny na cytochrom bf komplex namísto na NADP +. Elektrony prochází zpět bf komplexem, redukují plastocyanin, který je posléze reoxidován P yklus se uzavírá. Výsledkem je pumpování protonů komplexem bf. Vzniklý protonový gradient pohání syntézu ATP. Tvoří se jen ATP a ne NADP! Proces je aktivní za situace, kdy je nadbytek NADP, prakticky nedostatek NADP +. yklická fotofosforylace je ve smyslu zisku ATP produktivnější než necyklická. Dva absorbované fotony poskytují 1 molekulu ATP, kdežto při necyklické je třeba na 3 ATP 8 fotonů. Na procesu se nepodílí fotosystém II, netvoří se kyslík fotolýzou vody.
41 Temnostní (sekundární) fáze fotosyntézy alvin Bensonův cyklus Metabolická dráha, ve které rostliny inkorporují 2 a tvoří sacharidy, byla objasněna mezi léty 1946 až 1953 M. alvinem a A. Bensonem. Pracovali s buňkami řas a zkoumali produkty vstupu značeného Analýzou buněk řas po 5 s po expozici značeným 14 2 zjistili, že jediným produktem je 3-fosfoglycerát (3PG) s označeným karboxylem. Dále zjistili, že akceptorem 2 je pentosa ribulosa-1,5-bisfosfát a enzymem podílejícím se na reakci je ribulosa-1,5-bisfosfátkarboxylasa (RUBIS). alvin-bensonův cyklus probíhá ve stromatu chloroplastů a lze ho rozdělit na dvě části: Produkční ve které tři molekuly ribulosa-1,5-bisfosfát reagují se třemi molekulami 2 za tvorby šesti molekul glyceraldehyd-3-fosfátu (GAP) za spotřeby 9 molekul ATP a 6 molekul NADP. yklický charakter procesu činí dráhu ekvivalentní tvorbě jedné molekuly GAP ze tří molekul 2. GAP se může využít k biosyntéze. Regenerační ve které uhlíkové atomy zbylých pěti GAP jsou přeměněny v procesu podobném pentosafosfátové dráze na tři ribulosa- 5-fosfáty, které proces zahajovaly.

42 Dráha elektronů z vody na NADP + při fotosyntéze
43 Fosforylce tří ribulosa-5-fosfátů, vstup tří 2 -karboxylace (RUBIS), tvorba šesti 3-fosfoglycerátů a šesti 1,3 - bisfosfoglycerátů ATP ADP Fosforibulosakinasa 2 P 3 2 P 3 Ribulosa-1,5-bisfosfát (Ru-1,5-BP) 2 Ribulosabisfosfát karboxylasa RUBIS - 2 P P 3 3-Fosfoglycerát (3-PG) ATP ADP Fosfoglycerátkinasa P 3 2 P 3 1,3-Bisfosfoglycerát (1,3-BPG) P 3 Ribulosa-5-fosfát (Ru-5-P) 2 P 3 Ribulosa-5-fosfát (Ru-5-P)
44 Redukce šesti 1,3-bisfosfoglycerátů na glyceraldehyd-3-fosfát a start NADP NADP regenerační fáze P 3 2 P 3 1,3-Bisfosfoglycerát (1,3-BPG) Glyceraldehyd-3-fosfát dehydrogenasa 2 P 3 Glyceraldehyd-3-fosfát (GAP) Triosafosfátisomerasa 2 P 3 PRDUKTY 2 Dihydroxyacetonfosfát (DAP) Aldolasa 2 P 3 2 P 3 Erythrosa-4-fosfát (E-4-P) Aldolasa 2 P 3 2 P 3 Fruktosa-1,6-bisfosfát (F-1,6-BP) 2 P 3 Sedoheptulosa-1,7-bisfosfát (S-1,7-BP)
45 Pokračování regenerační fáze 2 P 3 2 P 3 2 P 3 2 Fruktosabisfosfatasa Fruktosa-1,6-bisfosfát (F-1,6-BP) P i 2 2 P 3 Sedoheptulosa-1,7-bisfosfát (S-1,7-BP) 2 P 3 Ribosa-5-fosfát (R-5-P) 2 P 3 Xylulosa-5-fosfát (Xu-5-P) Sedoheptulosabisfosfatasa 2 P i 2 P 3 Fruktosa-6-fosfát (F-6-P) Transketolasa 2 P 3 Transketolasa Sedoheptulosa-7-fosfát (S-7-P)
46 Závěr regenerační fáze - tvorba ribulosa-1,5-bisfosfátu 2 2 P 3 Ribosa-5-fosfát (R-5-P) 2 P 3 Xylulosa-5-fosfát (Xu-5-P) Ribosafosfátisomerasa Fosfopentosaepimerasa P 3 Ribulosa-5-fosfát (Ru-5-P) 2 P 3 Ribulosa-5-fosfát (Ru-5-P) ATP ADP 2 P 3 Fosforibulokinasa 2 P 3 Ribulosa-1,5-bisfosfát (Ru-1,5-BP)
47 Jednotlivé reakce alvin Bensonova cyklu sm, z celkového počtu jedenácti reakcí alvin-bensonova cyklu, je zvratem glykolýzy nebo reakcí glukoneogeneze známé z biochemie živočichů. Jediné tři reakce z jedenácti jsou katalyzovány enzymy, které se v živočišné říši nevyskytují: Fosforibulosakinasa katalyzující tvorbu ribulosa-1,5-bisfosfátu z ribulosa-5-fosfátu. Ribulosabisfosfátkarboxylasa katalyzující vstup 2 do sacharidu. Sedoheptulosabisfosfatasa (SBPasa) katalyzující odštěpení fosfátu ze sedoheptulosa-1,7-bisfosfátu na sedoheptulosa-7-fosfát.
48 Ribulosabisfosfátkarboxylasa (RuBP karboxylasa) RuBP karboxylasa je nejdůležitější enzym na Zemi. Vyznačuje se nízkou aktivitou (k cat = 3 s -1 ), ale jeho obsah v zeleném listu přesahuje 50 % všech proteinů. Je tedy i nejrozšířenějším proteinem biosféry. RuBPkarboxylasa z vyšších rostlin a většiny fotosyntetických mikroorganismů je složena z osmi velkých podjednotek kódovaných chloroplastovou DNA (L je složena ze 477 aminokyselin) a osmi malých podjednotek (S složených z 123 aminokyselin) kódovaných jadernou DNA. K aktivitě RuBP karboxylasa je nutný Mg 2+ vázaný na katalyticky významnou karbamátovou skupinu (-N- - ), která se vytváří reakcí nesubstrátového 2 s ε-aminoskupinou Lys 201. Reakci in vivo katalyzuje RuBP karboxylasaaktivasa za spotřeby ATP.
49 Mechanismus katalýzy ribulosabisfosfátkarboxylasy Reakce je silně exergonní ( G o = - 35, 1 kj.mol -1 ), dáno štěpením oxokyseliny na dva stabilní 3-fosfoglyceráty. ENZYM B 2 P 3-2 P 3 2 P 3-2 P 3 Ribulosa-1,5-bisfosfát 2 P 3 Endiolátová forma 2 P 3 xokyselina + 2 P P 3 2 P Fosfoglycerát P 3-2 P 3 ydratovaný meziprodukt 3-Fosfoglycerát
50 Biosyntéza sacharidů elková stechiometrie alvin-bensonova cyklu (GAP = glyceraldehyd-3-fosfát) ATP + 6 NADP GAP + 9 ADP + 8 P i + 6 NADP + Vytvořený GAP může být využit k řadě biosyntéz, buď vně nebo uvnitř chloroplastu. Může být převeden na fruktosa-6-fosfát dalším postupem cyklu a poté na glukosa-1-fosfát (G1P) (fosfoglukosaisomerasa a fosfoglukosamutasa). G1P je prekurzor všech dalších sacharidů rostlin. lavní složka škrobu, α-amylosa, je syntetizována ve stromatu chloroplastu jako dočasná skladovatelná forma glukosy. α-amylosa je také syntetizována jako dlouhodobá skladovatelná forma v jiných částech rostlin jako jsou listy, semena a kořeny. elulosa tvořena dlouhými řetězci β(1 4)-vázanými glukosovými jednotkami je hlavním polysacharidem rostlin. Je syntetizována z UDP-glukos v rostlinné plasmové membráně a vylučována do extracelulárního prostoru.
51 Syntéza škrobu. Škrob je méně větvený než glykogen. elkově je reakce poháněna hydrolýzou PP i. Škrob je syntetizován a skladován v chloroplastech. Aktivní glukosa = ADP-glukosa. - γ P β P α P ADENSIN ATP - P - Glukosa-1-fosfát ADP-glukosafosforylasa - PP i Anorganická fosfatasa 2 P i 2 P P ADENSIN ADP-Glukosa Synthasa škrobu ADP α-amylosa (n jednotek) α-amylosa (n + 1 jednotek)
52 První část syntézy α-amylosy Aktivace (fosforylace) glukosa-1-fosfátu. - γ P β P α P ADENSIN ATP - P - Glukosa-1-fosfát ADP-glukosafosforylasa - PP i Anorganická fosfatasa 2 P i 2 P P ADENSIN - - ADP-Glukosa
53 Druhá část syntézy α-amylosy. Prodlužování řetězce na neredukujícím konci molekuly α-amylasy. Enzym: Synthasa škrobu 2 P P ADENSIN ADP-Glukosa Synthasa škrobu ADP α-amylosa (n jednotek) α-amylosa (n + 1 jednotek)
54 Sacharosa hlavní transportní sacharid pro nefotosyntetizující buňky - nejdůležitější produkt fotosyntézy Sacharosa je syntetizována v cytosolu a proto musí být její dvě komponenty (G3P a DA) transportovány z chloroplastu systémem antiporter s fosfátem. Dvě triosy se kombinují za tvorby fruktosa-6-fosfátu, která se následně převede na glukosa-1-fosfát. G1P je aktivován UTP (uridintrifosfát) na UDP-glukosu. Sacharosa-6-fosfát se tvoří za katalýzy enzymem sacharosafosfátsynthasa. dštěpení fosfátu se uskutečňuje sacharosafosfátfosfatasou. UDP-Glukosa + Fruktosa-6-fosfát Sacharosafosfátsynthasa UDP Sacharosa-6-fosfát 2 Sacharosafosfátfosfatasa P i Sacharosa
55 Regulace alvin Bensonova cyklu Kontrolní body každé metabolické dráhy jsou na enzymových stupních vzdálených od rovnováhy (velká negativní hodnota G). Enzymy katalyzující reakce s velkou změnou volné energie: Ribulosabisfosfátkarboxylasa; G (kj.mol -1 ) = - 41, 0 Fruktosabisfosfatasa; G (kj.mol -1 ) = - 27, 2 Sedoheptulosabisfosfatasa; G (kj.mol -1 ) = - 29, 7 Katalytická účinnost těchto enzymů je silně ovlivňována osvětlením!! Aktivita RUBISA je ovlivňována těmito dvěma na světle závislými faktory: p. Po osvětlení se zvyšuje p ve stromatu z hodnoty 7, 0 na hodnotu 8, 0. Protony jsou pumpovány ze stromatu do dutin thylakoidů. RUBIS má ostré p optimum 8, 0!! [Mg 2+ ] Vstup protonů do thylakoidů je kompenzován transportem Mg 2+ do stromatu.
56 Standardní a fyziologické změny volné energie reakcí alvin-bensonova cyklu Stupeň Enzym G o (kj.mol -1 ) G (kj.mol -1 ) 1. Fosforibulokinasa - 21, 8-15, 8 2. Ribulosabisfosfátkarboxylasa - 35, 1-41, Fosfoglycerátkinasa + glyceraldehyd-3-fosfátdehydrogenasa + 18, 0-6, 7 5. Triosafosfátisomerasa - 7, 5-0, 8 6. Aldolasa - 21, 8-1, 7 7. Fruktosabisfosfatasa - 14,2-27,2 8. Transketolasa + 6, 3-3, 8 9. Aldolasa - 23, 4-0, Sedoheptulosabisfosfatasa - 14,2-29,7 11. Transketolasa + 0, 4-5, Fosfopentosaepimerasa + 0, 8-0, Ribosafosfátisomerasa + 2, 1-0, 4
57 Regulace alvin Bensonova cyklu Analog přechodového stavu (katalytického mechanismu RUBISa) karboxyarabinitol-1-fosfát (A1P). Tuto látku, která inhibuje RUBISo, syntetizuje řada rostlin pouze za tmy. Blokuje se tak aktivita RUBISA ve tmě. ba další enzymy jsou také aktivovány zvýšením p a [Mg 2+ ]. Navíc jsou ovlivňovány NADP. To je druhý regulační systém závislý na redoxním potenciálu stromatu. Thioredoxin, protein obsahující 105 aminokyselin, má redukovatelnou disulfidovou skupinu. Redukovaný thioredoxin aktivuje jak fruktosabisfosfatasu, tak sedoheptulosabisfosfatasu. Redoxní hladina thioredoxinu závisí na systému ferredoxinthioredoxinreduktasy. Jeho redoxní stav závisí na osvětlení. Thioredoxin deaktivuje fosfofruktokinasu klíčový enzym glykolýzy. Závěr: Světlo stimuluje alvin-bensonův cyklus a deaktivuje glykolýzu, kdežto tma má opačný efekt.
58 A-1-P inhibitor ribulosabisfosfátkarboxylasy (tma) 2 P 3-2 Karboxyarabinitol-1-fosfát (A-1-P)
 Je přítomen téměř ve všech organismech pro život savců je")
59 Thioredoxin je 1kD oxidoreduktasa - enzym obsahující dithioldisulfid v aktivním místě, antioxidant) Je přítomen téměř ve všech organismech pro život savců je esenciální.
60 Světlem indukovaná funkce ferredoxin-thioredoxinreduktasy. Redukovaný thioredoxin aktivuje řadu enzymů alvinova cyklu. PSI red hν PSI * red PSI ox Ferredoxin ox Ferredoxin red Ferredoxin-thioredoxin reduktasa red Ferredoxin-thioredoxin reduktasa ox S S S S Thioredoxin ox Thioredoxin red S S S S Bisfosfatasa aktivní Bisfosfatasa inaktivní S S S S
61 Fotorespirace d roku 1960 je známo, že osvětlené rostliny spotřebovávají kyslík a uvolňují 2 drahou odlišnou od oxidativní fosforylace. Při nízkých hladinách 2 a vysokých 2 může proces fotorespirace převládnout nad fixací 2. Překvapení: kyslík kompetuje s 2 jako substrát RUBISa. Přesnější název: ribulosabisfosfátkarboxylasa-oxygenasa (RUBIS). Při oxygenasové reakci se vytváří z RuBP 3-fosfogylcerát a fosfoglykolát. Fosfoglykolát je hydrolyzován na glykolát a ten je oxidován za tvorby 2 v sérii enzymových reakcí v peroxisomech a mitochondrii. Fotorespirace je devastující proces jehož výsledkem je neužitečná spotřeba světlem vytvořeného NADP a ATP!!
62 Pravděpodobný mechanismus oxygenasové reakce RUBISA 2 P 3 2 P 3 2 P 3 - ENZYM B 2 P 3 Ribulosa-1,5-bisfosfát 2 P 3 Endiolátová forma 2 P 3 xokyselina + 2 P P P 3 - Fosfoglykolát 3-Fosfoglycerát 2 P 3 ydratovaný meziprodukt
63 xygenasová reakce RUBISA. hloroplast peroxisom (glyoxysom) - 2 P P 3 i + - Fosfoglykolátfosfatasa 2 P 3-2 Glykolátoxidasa - 3-Fosfoglycerát Fosfoglykolát Glykolát / Katalasa Glyoxylát 2 2 P 3 2 P 3 Ribulosa-1,5-bisfosfát karboxylasa/oxygenasa 2-2 P 3 3-Fosfoglycerát Glycerátkinasa ADP ATP - 2 Glycerát NAD + NAD ydroxypyruvátreduktasa - 2 ydroxypyruvát Ribulosa-1,5-bisfosfát LRPLAST PERXISM
64 xygenasová reakce RUBISA. Peroxisom mitochondrie + 2 N Glyoxylát Glycin - N 3 TRANSAMINAE N Serin 2 ydroxypyruvát PERXISM MITNDRIE
65 Rostliny 4 koncentrují na povrchu listu 2. Řada rostlin se evolučně přizpůsobila kolísání koncentrace 2 a 2 tak, že koncentrují ve fotosyntetických buňkách 2 a zabraňují fotorespiraci. Např. cukrovka, cukrová třtina, kukuřice a mnoho plevelných rostlin. Typické pro tropické oblasti. List má charakteristickou anatomii pod vrstvou buněk tzv. mesofylu jsou soustředěny žilky cévních svazků. V mesofylových buňkách není ribulosabisfosfátkarboxylasa!! Kutikula Svrchní epidermis Palisádový mezofyl oubový mezofyl Xylem Floem évní svazek 2 2 Spodní epidermis Průduch (stoma) Svěrací buňky
66 yklus 4 M. atch a R. Slack (1960) V mesofylu reaguje vstupující 2 ve formě 3 - s fosfoenolpyruvátem za tvorby oxaloacetátu (čtyři uhlíky, proto 4 ). xaloacetát je redukován NADP na malát, který je transportován do buněk cévních svazků, kde je oxidativně dekarboxylován na 2, pyruvát a NADP. Takto koncentrovaný 2 vstupuje do alvinova cyklu. Pyruvát je transportován zpět do mesofylu, kde je fosforylován za katalýzy pyruvát-fosfátdikinasy za tvorby fosfoenolpyruvátu. Při aktivaci přechází ATP na AMP + PP i. PP i je posléze hydrolyzován všudepřítomnou pyrofosfatasou na 2 x P i. xid uhličitý je koncentrován v buňkách cevních svazků na úkor spotřeby 2 ATP/ 2. Fotosyntéza 4 rostlin spotřebovává celkem 5 ATP na rozdíl od 3 rostlin, které spotřebují 3 ATP.
67 Metabolická dráha 4 rostlin. Mezofyl buňky pochvy cévních svazků 2 VZDU 2 P i Karbonátanhydrasa ATP + P i AMP + PP i 3 - P i - NADP NADP Pyruvátfosfát dikinasa P 3 - Fosfoenolpyruvát (PEP) PEP karboxylasa Malátdehydrogenasa - xaloacetát MEZFYLVÁ BUŇKA - Malát BUŇKA PVY ÉVNÍ SVAZKU 3 Jablečný enzym - Pyruvát 2 + NADP NADP + ALVINŮV YKLUS 3-Fosfoglycerát
68 AM rostliny jako varianta rostlin 4 Dráha byl poprvé popsána u tučnolistých rostlin a odtud název AM crassulacean acid metabolism. U AM rostlin je časově oddělen záchyt 2 a alvin-bensonův cyklus. AM rostliny otevírají stomata (póry v listech) v noci a zachycují 2 drahou 4 ve formě malátu. Pro zachycení 2 je nutné velké množství fosfoenolpyruvátu. To se získává štěpením škrobu a glykolýzou. Během dne je malát štěpen za tvorby 2 vstupujícího do alvin Bensonova cyklu a pyruvátu použitého na resyntézu škrobu. elý tento proces je realizován také z důvodu zabránění ztrátě vody!!! Stejnou drahou probíhá fotosyntéza u sukulentů.
FOTOSYNTÉZA ZÁKLAD ŽIVOTA NA ZEMI
 FOTOSYNTÉZA ZÁKLAD ŽIVOTA NA ZEMI Pavel Peč Katedra biochemie Přírodovědecké fakulty Univerzita Palackého v Olomouci Fotosyntéza fixuje na Zemi ročně asi 1011 tun uhlíku, což reprezentuje 1018 kj energie.
FOTOSYNTÉZA ZÁKLAD ŽIVOTA NA ZEMI Pavel Peč Katedra biochemie Přírodovědecké fakulty Univerzita Palackého v Olomouci Fotosyntéza fixuje na Zemi ročně asi 1011 tun uhlíku, což reprezentuje 1018 kj energie.
Základy biochemie KBC / BCH. Fotosyntéza. Inovace studia biochemie prostřednictvím e-learningu CZ / /0407
 Základy biochemie KB / B Fotosyntéza Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky.
Základy biochemie KB / B Fotosyntéza Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky.
Obsah. Chloroplasty. Světlo absorbující pigmenty. Světelná reakce. Calvinův - Bensonův cyklus (RUBISCO) Fotorespirace. Rostliny C 4 a CAM
 Fotorespirace. Rostliny C 4 a CAM") FOTOSYNTÉZA Obsah hloroplasty Světlo absorbující pigmenty Světelná reakce alvinův - Bensonův cyklus (RUBISO) Fotorespirace Rostliny 4 a AM Typ výživy a metabolismus Zdroj energie Zdroj uhlíku Zdroj vodíku
FOTOSYNTÉZA Obsah hloroplasty Světlo absorbující pigmenty Světelná reakce alvinův - Bensonův cyklus (RUBISO) Fotorespirace Rostliny 4 a AM Typ výživy a metabolismus Zdroj energie Zdroj uhlíku Zdroj vodíku
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Fotosyntéza
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Metabolické dráhy. František Škanta. Glykolýza. Repetitorium chemie X. 2011/2012. Glykolýza. Jaký je osud pyruátu bez přítomnosti kyslíku?
 Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
35.Fotosyntéza. AZ Smart Marie Poštová
 35.Fotosyntéza AZ Smart Marie Poštová m.postova@gmail.com Fotosyntéza - úvod Syntéza glukosy redukcí CO 2 : chlorofyl + slun.zareni 6 CO 2 + 12H 2 O C 6 H 12 O 6 + 6O 2 + 6H 2 O (Kyslík vzniká fotolýzou
35.Fotosyntéza AZ Smart Marie Poštová m.postova@gmail.com Fotosyntéza - úvod Syntéza glukosy redukcí CO 2 : chlorofyl + slun.zareni 6 CO 2 + 12H 2 O C 6 H 12 O 6 + 6O 2 + 6H 2 O (Kyslík vzniká fotolýzou
12-Fotosyntéza FRVŠ 1647/2012
 C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
FOTOSYNTÉZA. Princip, jednotlivé fáze
 FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
Biosyntéza sacharidů 1
 Biosyntéza sacharidů 1 S a c h a r id y p o tr a v y (š k r o b, g ly k o g e n, sa c h a r o sa, a j.) R e z e r v n í p o ly sa c h a r id y J in é m o n o sa c h a r id y Trávení (amylásy - sliny, pankreas)
Biosyntéza sacharidů 1 S a c h a r id y p o tr a v y (š k r o b, g ly k o g e n, sa c h a r o sa, a j.) R e z e r v n í p o ly sa c h a r id y J in é m o n o sa c h a r id y Trávení (amylásy - sliny, pankreas)
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
Fotosyntéza a Calvinův cyklus. Eva Benešová
 Fotosyntéza a Calvinův cyklus Eva Benešová Fotosyntéza světlo CO 2 + H 2 O O 2 + (CH 2 O) světlo 6CO 2 + 6H 2 O 6O 2 + C 6 H 12 O 6 Opět propojení toku elektronů se syntézou ATP. Zachycení světelné energie
Fotosyntéza a Calvinův cyklus Eva Benešová Fotosyntéza světlo CO 2 + H 2 O O 2 + (CH 2 O) světlo 6CO 2 + 6H 2 O 6O 2 + C 6 H 12 O 6 Opět propojení toku elektronů se syntézou ATP. Zachycení světelné energie
FOTOSYNTÉZA. soubor chemických reakcí,, probíhaj v rostlinách a sinicích. z CO2 a vody jediný zdroj kyslíku ku pro život na Zemi
 Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/ Anotace. Fotosyntéza světelná fáze. VY_32_INOVACE_Ch0214.
 Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vyjádření fotosyntézy základními rovnicemi
 FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
Repetitorium chemie 2015/2016. Metabolické dráhy František Škanta
 Repetitorium chemie 2015/2016 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů Oxidace
Repetitorium chemie 2015/2016 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů Oxidace
Biochemie II 2009/10. Metabolismus. František Škanta
 Biochemie II 2009/10 Metabolismus František Škanta Metabolické dráhy Primární metabolismus Metabolismus sacharidů Glykolýza Krebsův cyklus xidativní fosforylace Metabolismus lipidů xidace mastných kyselin
Biochemie II 2009/10 Metabolismus František Škanta Metabolické dráhy Primární metabolismus Metabolismus sacharidů Glykolýza Krebsův cyklus xidativní fosforylace Metabolismus lipidů xidace mastných kyselin
Dýchací řetězec. Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci)
") Dýchací řetězec Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 -->6 CO 2 + 6H 2 O + 38 ATP Dýchací
Dýchací řetězec Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 -->6 CO 2 + 6H 2 O + 38 ATP Dýchací
9. Dýchací řetězec a oxidativní fosforylace. mitochondriální syntéza ATP a fotosyntéza
 9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
Fotosyntéza (2/34) = fotosyntetická asimilace
 = fotosyntetická asimilace") Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Repetitorium chemie 2016/2017. Metabolické dráhy František Škanta
 Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
FOTOSYNTÉZA. CO 2 a vody. - soubor chemických reakcí. - probíhá v rostlinách a sinicích. - zachycení a využití světelné energie
 Fotosyntéza FOTOSYNTÉZA - soubor chemických reakcí - probíhá v rostlinách a sinicích - zachycení a využití světelné energie - tvorba složitějších chemických sloučenin z CO 2 a vody - jediný zdroj kyslíku
Fotosyntéza FOTOSYNTÉZA - soubor chemických reakcí - probíhá v rostlinách a sinicích - zachycení a využití světelné energie - tvorba složitějších chemických sloučenin z CO 2 a vody - jediný zdroj kyslíku
ANABOLISMUS SACHARIDŮ
 zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Dýchací řetězec (Respirace)
") Dýchací řetězec (Respirace) Buněčná respirace (analogie se spalovacím motorem) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 ---------> 6 CO 2 + 6H 2 O + 38 ATP Oxidativní
Dýchací řetězec (Respirace) Buněčná respirace (analogie se spalovacím motorem) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 ---------> 6 CO 2 + 6H 2 O + 38 ATP Oxidativní
14. Fyziologie rostlin - fotosyntéza, respirace
 14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74. Katedra experimentální biologie rostlin, Z. Lhotáková
 FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
Fyziologie rostlin. 9. Fotosyntéza část 1. Primární fáze fotosyntézy. Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014
 Fyziologie rostlin 9. Fotosyntéza část 1. Primární fáze fotosyntézy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Fotosyntéza 1. část - úvod - chloroplasty - sluneční záření -
Fyziologie rostlin 9. Fotosyntéza část 1. Primární fáze fotosyntézy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Fotosyntéza 1. část - úvod - chloroplasty - sluneční záření -
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Fotosyntéza Světelné reakce. Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni
 Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
FOTOSYNTÉZA. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1
 FOTOSYNTÉZA Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1 Fotosyntéza (z řec. phos, photós = světlo) je anabolický děj probíhající u autotrofních organismů (řasy,
FOTOSYNTÉZA Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1 Fotosyntéza (z řec. phos, photós = světlo) je anabolický děj probíhající u autotrofních organismů (řasy,
Energie fotonů je předávána molekulám chlorofylu A, který se zachyceným fotonem excituje (uvolní se energeticky bohatý elektron).
.") Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké
 Fotosyntéza Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké rostliny. Zelené rostliny patří mezi autotrofy
Fotosyntéza Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké rostliny. Zelené rostliny patří mezi autotrofy
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
ení k tvorbě energeticky bohatých organických sloučenin
 Fotosyntéza mimořádně významný proces, využívající energii slunečního zářenz ení k tvorbě energeticky bohatých organických sloučenin (sacharidů) z jednoduchých anorganických látek oxidu uhličitého a vody
Fotosyntéza mimořádně významný proces, využívající energii slunečního zářenz ení k tvorbě energeticky bohatých organických sloučenin (sacharidů) z jednoduchých anorganických látek oxidu uhličitého a vody
METABOLISMUS SACHARIDŮ
 METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Předmět: KBB/BB1P; KBB/BUBIO
 Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
1- Úvod do fotosyntézy
 1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
- metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy a jejich životním prostředím
 Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
FOTOBIOLOGICKÉ POCHODY
 FOTOBIOLOGICKÉ POCHODY Základním zdrojem energie nutné pro život na Zemi je sluneční záření. Většina pochodů souvisí s přímým využitím zářivé energie pro metabolické pochody nebo pro orientaci organizmu
FOTOBIOLOGICKÉ POCHODY Základním zdrojem energie nutné pro život na Zemi je sluneční záření. Většina pochodů souvisí s přímým využitím zářivé energie pro metabolické pochody nebo pro orientaci organizmu
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
B4, 2007/2008, I. Literák
 B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
FOTOSYNTÉZA Správná odpověď:
 FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
LÁTKOVÝ A ENERGETICKÝ METABOLISMUS
 LÁTKOVÝ A ENERGETICKÝ METABOLISMUS Metabolismus = neustálý příjem, přeměna a výdej látek = probíhá po celou dobu života rostliny Dva typy procesů : ANABOLICKÉ KATABOLICKÉ ANABOLISMUS - energie se spotřebovává
LÁTKOVÝ A ENERGETICKÝ METABOLISMUS Metabolismus = neustálý příjem, přeměna a výdej látek = probíhá po celou dobu života rostliny Dva typy procesů : ANABOLICKÉ KATABOLICKÉ ANABOLISMUS - energie se spotřebovává
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Sacharidy a polysacharidy (struktura a metabolismus)
") Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Cukry (Sacharidy) Sacharidy a jejich metabolismus. Co to je?
 Sacharidy a jejich metabolismus. Co to je?") Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Praktické cvičení č. 11 a 12 - doplněno
 Praktické cvičení č. 11 a 12 - doplněno Téma: Metabolismus eukaryotické buňky Pomůcky: pracovní list, učebnice botaniky Otázky k opakování: Co je anabolismus a co je katabolisimus? Co jsou enzymy a jak
Praktické cvičení č. 11 a 12 - doplněno Téma: Metabolismus eukaryotické buňky Pomůcky: pracovní list, učebnice botaniky Otázky k opakování: Co je anabolismus a co je katabolisimus? Co jsou enzymy a jak
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Transport elektronů a oxidativní fosforylace
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Pentosový cyklus. osudy glykogenu. Eva Benešová
 Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Konsultační hodina. základy biochemie pro 1. ročník. Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa
 Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
CZ.1.07/2.2.00/ Obecný metabolismus. Energetický metabolismus (obecně) (1).
 (1).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Škola: Gymnázium, Brno, Slovanské náměstí 7 III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN
 Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
pátek, 24. července 15 GLYKOLÝZA
 pátek,. července 15 GLYKLÝZ sacharosa threalosa laktosa sacharasa threlasa laktasa D-glukosa D-fruktosa T T hexokinasa T hexokinasa glykogen - škrob fosforylasa D-galaktosa UD-galaktosa UD-glukosa fruktokinasa
pátek,. července 15 GLYKLÝZ sacharosa threalosa laktosa sacharasa threlasa laktasa D-glukosa D-fruktosa T T hexokinasa T hexokinasa glykogen - škrob fosforylasa D-galaktosa UD-galaktosa UD-glukosa fruktokinasa
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy
 Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
2. ČÁST - METABOLICKÉ PROCESY
 Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. 2. ČÁST - METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických
Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. 2. ČÁST - METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Metabolismus sacharidů. VY_32_INOVACE_Ch0216.
 Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
B METABOLICKÉ PROCESY
 B METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických sloučenin a reakcí při přeměnách látek v živých buňkách je hlavní náplní vědního oboru biochemie. Pro rostlinného fyziologa jsou
B METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických sloučenin a reakcí při přeměnách látek v živých buňkách je hlavní náplní vědního oboru biochemie. Pro rostlinného fyziologa jsou
2. ČÁST - METABOLICKÉ PROCESY
 Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. 2. ČÁST - METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických
Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. 2. ČÁST - METABOLICKÉ PROCESY Poznávání neuvěřitelně velkého množství chemických
Autor: Katka Téma: fyziologie (fotosyntéza) Ročník: 1.
 Ročník: 1.") Fyziologie Fotosyntéza Celým názvem: fotosyntetická asimilace - vznikla při ohrožení, že již nebudou anorg. l. rostliny začaly dělat fotosyntézu v atmosféře vzrostl počet O 2 = 1. energetická krize - nejdůležitější
Fyziologie Fotosyntéza Celým názvem: fotosyntetická asimilace - vznikla při ohrožení, že již nebudou anorg. l. rostliny začaly dělat fotosyntézu v atmosféře vzrostl počet O 2 = 1. energetická krize - nejdůležitější
SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU
 SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU Nikola Burianová Experimentální biologie 2.ročník navazujícího studia Katedra Fyziky Ostravská univerzita v Ostravě OBSAH
SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU Nikola Burianová Experimentální biologie 2.ročník navazujícího studia Katedra Fyziky Ostravská univerzita v Ostravě OBSAH
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
Fyziologie buňky. RNDr. Zdeňka Chocholoušková, Ph.D.
 Fyziologie buňky RNDr. Zdeňka Chocholoušková, Ph.D. Přeměna látek v buňce = metabolismus Výměna látek mezi buňkou a prostředím Buňka = otevřený systém probíhá výměna látek i energií s prostředím Některé
Fyziologie buňky RNDr. Zdeňka Chocholoušková, Ph.D. Přeměna látek v buňce = metabolismus Výměna látek mezi buňkou a prostředím Buňka = otevřený systém probíhá výměna látek i energií s prostředím Některé
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
MitoSeminář II: Trochu výpočtů v bioenergetice. Souhrn. MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK
 MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů.
 Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů. Šárka Gregorová, 2013 Poznámka: protože se tyhle dvě státnicové otázky z velké
Fotosyntéza. Dýchání a fotosyntéza, struktura a funkce antén a reakčních center, energetika transportu elektronů a protonů. Šárka Gregorová, 2013 Poznámka: protože se tyhle dvě státnicové otázky z velké
Energetický metabolismus rostlin. respirace
 Energetický metabolismus rostlin Zdroje E: fotosyntéza respirace Variabilní využívání: - orgánové a pletivové rozdíly (kořen, prýt, pokožka, ) - změny při vývoji a diferenciaci - vliv dostupnosti vody,
Energetický metabolismus rostlin Zdroje E: fotosyntéza respirace Variabilní využívání: - orgánové a pletivové rozdíly (kořen, prýt, pokožka, ) - změny při vývoji a diferenciaci - vliv dostupnosti vody,
BUŇKA A ENERGIE. kajman brýlový Caiman crocodilus Kostarika, 2004. Biologie 6, 2015/2016, Ivan Literák
 BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
Ukázky z pracovních listů z biochemie pro SŠ A ÚVOD
 Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Energetický metabolismus rostlin
 Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
Energetický metabolismus rostlin Sylabus - témata (Fischer, Duchoslav) 1. Energie v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta, spřažení
DYNAMICKÁ BIOCHEMIE. Daniel Nechvátal :: www.gymzn.cz/nechvatal
 DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Metabolismus. Source:
 Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
OXIDATIVNÍ FOSFORYLACE
 OXIDATIVNÍ FOSFORYLACE OBSAH Mitochondrie Elektronový transport Oxidativní fosforylace Kontrolní systém oxidativního metabolismu. Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů Dýchací
OXIDATIVNÍ FOSFORYLACE OBSAH Mitochondrie Elektronový transport Oxidativní fosforylace Kontrolní systém oxidativního metabolismu. Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů Dýchací
Ekologie fotosyntézy
 Ekologie fotosyntézy Fotosyntéza Přeměna zářivé energie Slunce na energii chemických vazeb primární produkce organické hmoty fotochemický (Hillova reakce) a biochemický proces 1 mol přijatého CO 2 energetický
Ekologie fotosyntézy Fotosyntéza Přeměna zářivé energie Slunce na energii chemických vazeb primární produkce organické hmoty fotochemický (Hillova reakce) a biochemický proces 1 mol přijatého CO 2 energetický
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
BUŇKA A ENERGIE. kajman brýlový Caiman crocodilus Kostarika, Biologie 8, 2017/2018, Ivan Literák
 BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 8, 2017/2018, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 8, 2017/2018, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
FOTOSYNTÉZA. Fotosyntéza je fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených
 B + + B + FOTOSYNTÉZA Fotosyntéza je fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta
B + + B + FOTOSYNTÉZA Fotosyntéza je fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
FOTOSYNTÉZA V DYNAMICKÝCH
 FOTOSYNTÉZA V DYNAMICKÝCH ANIMACÍCH Výukový program vytvořený v programu Macromedia Flash Milada Roštejnská Helena Klímová Praha 2008 Univerzita Karlova v Praze, Přírodovědecká fakulta FOTOSYNTÉZA V DYNAMICKÝCH
FOTOSYNTÉZA V DYNAMICKÝCH ANIMACÍCH Výukový program vytvořený v programu Macromedia Flash Milada Roštejnská Helena Klímová Praha 2008 Univerzita Karlova v Praze, Přírodovědecká fakulta FOTOSYNTÉZA V DYNAMICKÝCH
5. ENZYMOVÉ REAKCE FOTOSYNTÉZY
 5.1. ALVINŮV YKLUS Oxid uhličitý vstupuje do organické vazby a redukuje se v reduktivním pentosovém cyklu (PR, pentose reducing cycle), který se také nazývá fotosyntetický cyklus redukce oxidu uhličitého
5.1. ALVINŮV YKLUS Oxid uhličitý vstupuje do organické vazby a redukuje se v reduktivním pentosovém cyklu (PR, pentose reducing cycle), který se také nazývá fotosyntetický cyklus redukce oxidu uhličitého
VLIV ZVÝŠENÉ KONCENTRACE OXIDU UHLIČITÉHO NA DENNÍ CHOD OBSAHU A AKTIVITY ENZYMU RUBISCO
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA CHEMICKÁ ÚSTAV CHEMIE POTRAVIN A BIOTECHNOLOGIÍ FACULTY OF CHEMISTRY INSTITUTE OF FOOD SCIENCE AND BIOTECHNOLOGY VLIV ZVÝŠENÉ KONCENTRACE
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA CHEMICKÁ ÚSTAV CHEMIE POTRAVIN A BIOTECHNOLOGIÍ FACULTY OF CHEMISTRY INSTITUTE OF FOOD SCIENCE AND BIOTECHNOLOGY VLIV ZVÝŠENÉ KONCENTRACE
aneb Fluorescence chlorofylu jako indikátor stresu
 Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Glykolýza a neoglukogenese
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Otázka: Metabolismus. Předmět: Biologie. Přidal(a): Furrow. - přeměna látek a energie
: Furrow. - přeměna látek a energie") Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Bp1252 Biochemie. #8 Metabolismus živin
 Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty
 sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení
 Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení Ročník 1.
Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 30 Metabolismus, fotosyntéza, dýchání, glykolýza, kvašení Ročník 1.
Metabolismus bílkovin. Václav Pelouch
 ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
Energetický metabolismus rostlin
 Energetický metabolismus rostlin Sylabus - témata (Fischer, Šantrůček) 1. Základy energetiky v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta,
Energetický metabolismus rostlin Sylabus - témata (Fischer, Šantrůček) 1. Základy energetiky v živých systémech Formy energie a základní principy přeměny energií; změny volné energie, rovnovážná konstanta,
Metabolismus sacharidů
 Základy biochemie KB / B Metabolismus sacharidů Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KB / B Metabolismus sacharidů Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem