15. Glukoneogenéza, glykoneogenéza. Biosyntéza aminokyselin, lipidů a nukleotidů
|
|
|
- Luboš Matějka
- před 6 lety
- Počet zobrazení:
Transkript
1 15. Glukoneogenéza, glykoneogenéza Biosyntéza aminokyselin, lipidů a nukleotidů
2 15.1 BIOSYNTHESA SACHARIDŮ Je hlavní děj tvorby organické hmoty z CO 2 a H 2 O (autotrofy + sluneční zář.) Heterotrofy tvoří sacharidy z 3C(2C) až 4C molekul z katabolických drah = glukogenese též glukoneogenese (tvorba glukosy z necukerných zdrojů) Navazuje-li na glukoneogenesi tvorba oligo- a polysacharidů = glykoneogenese


3 VÝCHOZÍ 3C(2C) AŽ 4C MOLEKULY PRO GLUKONEOGENESI Pyruvát Laktát Glycerol meziprodukty citrátového cyklu (CC) u rostlin a některých mikroorganismů s glyoxylátovým cyklem i acetyl-coa
4 GLUKONEOGENESE - PŘEMĚNA PYRUVÁTU NA GLUKOSU Glukoneogenese probíhá hlavně v játrech, ale i v ledvinách (hl. při hladovění), méně ve střevních epiteliích Pyruvát tvoří zejména hepatocyty z laktátu (redukcí) a alaninu (transaminací) Coriův cyklus a glukoso-alaninový cyklus mezi játry a svaly K přeměně nelze využít celou cestu obrácené glykolysy, 3 její fosforylační reakce jsou nevratné : 1) fosfoenolpyruvát pyruvát 2) fruktosa-6-fosfát fruktosa-1,6-bisfosfát 3) glukosa glukosa-6-fosfát
5 1) PYRUVÁT A FOSFOENOLPYRUVÁT


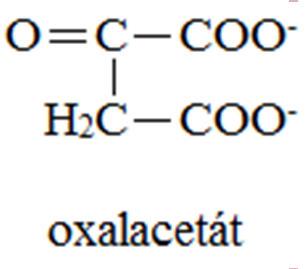
6
7 2) FRUKTOSA-1,6-BISFOSFÁT A FRUKTOSA-6- FOSFÁT
8 3) GLUKOSA-6-FOSFÁT A GLUKOSA
9 ENERGETICKÁ BILANCE A ŘÍZENÍ GLUKONEOGENESE Pro tvorbu 1 molekuly glukosy je třeba 2 molekul pyruvátu a 12 ATP 2x pyruvát + 12x ATP = 1 glc Glykolysa 1 molekuly glukosy poskytne 8 ATP, glukoneogenese je tedy energeticky dotována 1 glc = 8 ATP Glukoneogenesi zahajuje glukagon a adrenalin prostřednictvím camp


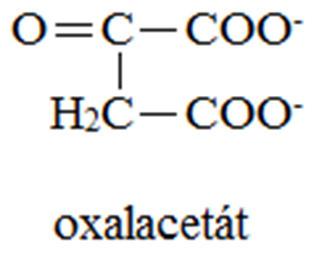
10 BIOSYNTHESA GLUKOSY Z DALŠÍCH PREKURZORŮ
11 GLUKOSA Z AMINOKYSELIN Glukogenních AMK je 18 (všechny kromě Lys a Leu) glukogenní i ketogenní jsou 4 AMK (Ile, Phe, Trp, Tyr) Lys a pravděpodobně i Leu jsou čistě ketogenní Obratlovci nemají glyoxylátový cyklus a nemohou tedy použít acetyl-coa (Lys, Leu) pro glukoneogenesi
12 VÝSLEDKY ODBOURÁVÁNÍ AMK Meziprodukty CC = substráty glukoneogenese 2-oxoglutarát: Arg, Glu, Gln, His, Pro sukcinyl-coa: Ile, Met, Val fumarát: Asp, Phe, Tyr oxalacetát: Asp, Asn pyruvát: Ala, Gly, Cys, Ser, Trp, Thr, acetyl-coa: Ile, Leu acetoacetát: Leu, Lys, Phe, Trp, Tyr
13 15.2 BIOSYNTHESA OLIGO- A POLYSACHARIDŮ Škrob zásobní polysacharid (PS) rostlin složen z α-amylosy (15-20 %) a α-amylopektinu (80-85 %) α-amylosa dlouhý lineární řetězec s vazbami α-d-glukos pouze 1 4 helikální struktura (+ jód = modrá b.) α-amylopektin velká rozvětvená molekula skládá se z jednotek α-d-glukosy s vazbami 1 4 a 1 6 po jednotkách glukos s vazbou 1 4 větvení vazbou 1 6 globulární struktura (+ jód = červená b.)
14 Glykogen: velmi rozvětvený polysacharid přítomen pouze u živočichů složen z jednotek α-d-glukosy s vazbami 1 4 a 1 6 po 3-7 jedn. glc s vazbou 1 4 větvení vazbou 1 6 zásobuje tělo glukosou, udržuje tak hladinu krevní glc v normě mezi jídly jaterní zásoby glykogenu vyčerpány už za 12-18h hladovění svalové zásoby glykogenu se úplně nevyčerpají nikdy
15 GLYKOGENESE - TVORBA GLYKOGENU 1) GLUKOSA-6-FOSFÁT

16 2) IZOMERISACE

17 3) AKTIVNÍ NUKLEOSID
18 4) RŮST ŘETĚZCE GLYKOGENU
19 GLYKOGENOLÝZA vztah glykoneogenese-glykogenolysa analogický vztahu glukoneogeneseglukolysa = nelze postupovat zpět pouhým otočením reakcí!!! probíhá vlastní reakční enzymatickou drahou štěpení glykogenu zahajuje fosforolysa (účast H 3 PO 4!)
20 1) FOSFOROLYSA
21 DVA ENZYMY JEDNÉ REAKCE svalová (dimer) a jaterní (monomer) glykogenfosforylasa jsou naprosto odlišné enzymy oba existují v dané tkáni v dalších dvou formách: glykogenfosforylasa a fosforylovaná v játrech aktivní forma ve svalech aktivní forma glykogenfosforylasa b defosforylovaná v játrech neaktivní forma ve svalech neaktivní forma, avšak aktivována přít. AMP po fosforolyse pokračuje jiný enzym linearisací v oblasti větvení
22 2) LINEARISACE VĚTVENÍ GLYKOGENU fosforolysa je zastavena 4 glc jednotky před větvením α1,4-transglykosylasa přenese trimer tří glukos na neredukující konec sousedního lineárního řetězce glykogenu na větvení nyní zbyla poslední glc, ta je odštěpena dle nejnovějších poznatků taktéž α1,4- transglykosylasou řetězec je zbaven větvícího bodu, linearisován, a tím připraven na opětovnou fosforolysu glykogenfosforylasou
23 3) GLUKOSA Z GLC-1-P výsledkem glykogenolysy je glc-1-p, který ještě není uvolněn do krve fosfoglukomutasa (PGM) opět izomerisuje glc-1-p přes glc-1,2-p 2 na využitelnější glc-6-p tkáně vybavené glc-6-fosfatasou (játra, ledviny) defosforylují glc-6-p a uvolní do krve čistou glukosu tkáně bez tohoto enzymu (svaly, mozek, tuková tkáň) jsou lakomci a krvi žádnou glc nedají glc-6-p si uschovají v cytosolu svých buněk pro svůj energetický metabolismus a glykolysou ho přemění na laktát glc-6-p mimo jiné odčerpává i pentosofosfátový cyklus
24 BIOSYNTHESA AMINOKYSELIN Schopnost biosynthesy aminokyselin je u různých organismů různá Rostliny dovedou vyrábět všechny aminokyseliny za použití NH 4+, NO 3 nebo NO 2- jako zdroje dusíku Bobovité rostliny v symbiose s určitými bakteriemi dokonce vycházejí z N 2 Mikroorganismy se ve schopnosti synthesy aminokyselin zřetelně liší Většina vyžaduje NH 4 + jako zdroj dusíku Některé bakterie a houby však jsou stejně jako rostliny schopné využívat i NO 3 a NO 2 - Existují i mikroorganismy, pro které jsou určité aminokyseliny esenciální
25 Živočichové musí používat k výrobě aminokyselin dusík organicky vázaný v aminokyselinách uvolněných hydrolysou bílkovin potravy nebo vlastních tkání Řadu kódovaných aminokyselin nedovedou vůbec synthetisovat a musí je získávat potravou nebo štěpením tkáňových bílkovin Říkáme jim esenciální (nepostradatelné) aminokyseliny, ostatním pak neesenciální nebo relativně postradatelné Pro člověka jsou esenciální větvené aminokyseliny valin, leucin a isoleucin, aromatické fenylalanin a tryptofan, dále threonin, methionin a lysin Histidin a arginin jsou postradatelné jen v dospělosti, v době vývoje v dětském věku jsou také esenciální Další dvě aminokyseliny jsou podmíněně postradatelné pouze při dostatku esenciálních aminokyselin, z nichž se tvoří; jsou to tyrosin, vznikající z fenylalaninu, a cystein synthetisovaný z methioninu
26 SYNTHESY NEESENCIÁLNÍCH AMINOKYSELIN Alanin a aspartát jsou vyráběny jednostupňovou přeměnou pyruvátu, resp. Oxalacetátu transaminační reakce s pyridoxalfosfátem jako kofaktorem a glutamátem jako zdrojem aminoskupiny nebo o obrácení procesu oxidační deaminace,tj. o redukční aminaci přes aminokyselinu Glutamát se tvoří redukční aminací z 2-oxoglutarátu, tedy obrácením reakce, která slouží k regeneraci 2-oxoglutarátu při transaminačních reakcích Glutamát je dále prekursorem další neesenciální aminokyseliny prolinu, který vzniká obrácením jeho odbourávání přes γ-semialdehyd Glutamát se dále spolu s aspartátem podílí na biosynthese argininu jako donor aminoskupiny Prekursorem argininu je nekódovaná aminokyselina ornithin a synthesa je součástí ureosynthetického cyklu Uhlíkové skelety aminokyselin pocházejí z meziproduktů glykolysy, pentosového nebo citrátového cyklu
27 O H 3 C C - COO Pyruvát O - O O C CH 2 C - COO Oxaloacetát O - O C O CH 2 CH 2 C - COO 2-Oxoglutarát Aminotransferasa 1 Aminokyselina 2-Oxokyselina Aminotransferasa 2 Aminokyselina 2-Oxokyselina Aminotransferasa 3 Aminokyselina 2-Oxokyselina H 3 C H C - COO + NH 3 Alanin O - O C H CH 2 C - COO + NH 3 Aspartát O - O C H CH 2 CH 2 C - COO + NH 3 Glutamát Asparaginsynthetasa O N H 2 C 4 H ATP Glutamin AMP + PP i - CH 2 C COO NH 3 + Asparagin Glutamát Glutaminsynthetasa O 2- O 3 PO C CH 2 5 ATP ADP H γ-glutamylfosfát (meziprodukt) - CH 2 C COO 5 NH 3 + NH 4 + P i O N H 2 C H CH 2 CH 2 C - COO + NH 3 Glutamát
28 GLU JE PREKURZOREM PRO, ORN A ARG O H C CH 2 H Glutamát CH 2 C COO NH Oxoglutarát H 2 N CH 2 CH 2 H CH 2 C COO NH Glutamát-5-semialdehyd Ornithin Samovolná cyklizace 3 H 2 C CH 2 Močovinový cyklus HC N C H 4 COO -Pyrrolin-5-karboxylát - NAD(P)H NAD(P) + + H 2 N N H 2 C N H CH 2 CH 2 Arginin H CH 2 C COO NH H H 2 C H C N H CH 2 C H Prolin COO -
29 Serin se synthetisuje z 3-fosfoglycerátu, meziproduktu glykolýzy Prvním stupněm je oxidace na 3-fosfohydroxypyruvát - tato α-oxokyselina je pak transaminována na 3- fosfoserin, jehož hydrolysou vzniká serin - COO NAD + NADH - COO Glutamát 2-Oxoglutarát - COO P i H H C CH 2 OH 2- OPO 3 1 C CH 2 O 2- OPO H 3 N C CH 2 H 2- OPO 3 3 HO - CH 2 C COO NH Fosfoglycerát 3-Fosfohydroxypyruvát 3-Fosfoserin Serin
30 Serin je prekursorem glycinu a cysteinu Při synthese glycinu se odstraní β-uhlíkový atom postranního řetězce serinu přesunem na tetrahydrofolát, nosič jednouhlíkových jednotek, Serin + tetrahydrofolát glycin + methylentetrahydrofolát + H 2 O V obráceném směru probíhá tato reakce při odbourávání glycinu Reakce je katalysována serinovou transhydroxymethylasou s PLP jako kofaktorem. Vazba mezi α- a β-atomy uhlíku serinu se stabilizuje tvorbou Schiffovy base serinu s PLP β-uhlíkový atom je pak přesunut na tetrahydrofolát Glycin může vznikat také z CO 2 a NH 4+ a methylentetrahydrofolátu za katalysy glycinsynthasou Obrácením výše uvedené reakce se pak může z takto vyrobeného glycinu tvořit serin
31 Serin přechází na cystein nahrazením kyslíkového atomu postranního řetězce za atom síry Jeho zdrojem je esenciální aminokyselina methionin, synthetisovaná z aspatrátu Meziprodukt této synthesy se kondensuje se serinem za vzniku cystathioninu - Ten je deaminován a štěpen na cystein a 2-oxoglutarát, zatímco při synthese methioninu se cystathion štěpí na druhé straně atomu síry a vzniká homocystein a pyruvát Enzymy obou reakčních stupňů jsou lyasy a používají PLP jako kofaktor.
32 Prekursory zbývajících esenciálních aminokyselin jsou pyruvát a aspartát Threonin a methionin se tvoří z homoserinu, vznikajícího z aspartátu přes aspartylfosfát a aspartátsemialdehyd Threonin vzniká fosforylací homoserinu, následnou eliminací fosfátu z γ-atomu uhlíku a substitucí β-uhlíku hydroxylovou skupinou Reakce je katalysováva PLP-enzymem threoninsynthetasou. Při biosynthese methioninu se tvoří z homoserinu nejprve o-sukcinyl-homoserin přenosem sukcinylové skupiny ze sukcinyl-coa V dalším stupni nahradí cystein sukcinát, čímž vznikne cystathion Ten se hydrolyticky štěpí na homocystein, pyruvát a NH 4 + Protože cystein může být methyován na methionin, je zřejmé, že cystathionin může sloužit jako meziprodukt při přeměně cysteinu na methionin u rostlina bakterií a při přeměně methiononu na cystein u savců
33 Histidin Pět atomů ze šesti histidinových má původ v 5- fosforibosyl-α-pyrofosfátu. Stejná látka je základem biosyntézy purinových a pyrimidinových bází nukleových kyselin. Šestý atom je z ATP Ostatní atomy z ATP se odštěpí jako 5-aminoimidazol-4-karboxamidribonukleotid, což je také meziprodukt syntézy purinů
34 BIOSYNTHESA LIPIDŮ Biosynthesa mastných kyselin Vyšší mastné kyseliny vznikají procesem, který se svým spirálovým charakterem velice podobá procesu odbourávání mastných kyselin Obrácená redukční spirála má prakticky identické meziprodukty, ovšem jednotlivé reakce provádějí jiné enzymy a do procesu je začleněná karboxylace Biosynthesa mastných kyselin je také v buňce lokalizovaná jinde Zatímco mastné kyseliny se odbourávají v mitochondriích, probíhá jejich biosynthesa v cytosolu Hlavními místy synthesy mastných kyselin jsou tukové tkáně
35 Výchozí látkou pro biosynthesu mastných kyselin je acetyl-coa, vznikající při odbourávání jiných mastných kyselin, při oxidační dekarboxylaci pyruvátu v rámci aerobního odbourávání sacharidů a z uhlíkových koster některých aminokyselin Podíváme-li se na princip výroby mastných kyselin vidíme, že jde o redukci acetylu na CH 2 - CH 2 - zbytky, tvořící obratle páteře mastné kyseliny Vodíky potřebné k redukci poskytuje NADPH získaný v pentosovém cyklu, u fototrofů též ve světlé fázi fotosynthesy Jako každý anabolický děj vyžaduje i výroba mastných kyselin energii
36 Uhlíkový řetězec mastné kyseliny se tedy buduje stupňovitě z dvouuhlíkových jednotek Ty však neposkytuje přímo acetyl-coa, ale energií bohatší produkt jeho karboxylace, malonyl-coa Meziprodukty biosynthesy jsou thioestery bílkoviny označené ACp ( Acyl Carrier Protein ) a ne CoA, jako při odbourávání mastných kyselin
37 Výchozí materiál, acetyl-coa, vzniká převážně v mitochondriích Membrány mitochondrií však pro něj nejsou propustné Přenos acetyl-coa do cytoslu, kde se odehrává biosynthesa mastných kyselin, se uskutečňuje buď karnitinem, nebo prostřednictvím přeměny na citrát Přenos karnitinem probíhá stejným mechanisnem jako přenos acylů vyšších mastných kyselin, ale v obráceném směru Druhou cestou, kterou se dostává acetyl-coa do cytosolu, je tvorba citrátu, který vzniká reakcí acetyl-coa s oxalacetátem za katalysy citrátsynthasou Citrát takto vytvořený přejde mitochondriální membránou do cytosolu a tam se účinkem citrátlyasy rozpadá za účasti ATP a CoA zpět na oxalacetát a acetyl-coa.
38 Proces biosynthesy mastných kyselin lze rozdělit do tří fází: A. Výroba malonyl-coa B. Synthesa palmitové kyseliny C. Další přeměny palmitátu A. Výroba malonyl-coa. Karboxylace acetyl-coa na energeticky bohatší malonyl-coa probíhá na enzymovém komplexu ligasy, acetyl- CoA-karboxylasy, který se skládá ze tří funkčních podjednotek: a) bílkovinného nosiče biotinu ( BCCP = Biotin Karboxyl Carrier Protein ), který má biotický kofaktor vazaný na jednom ze zbytků lysinu b) biotinkarboxylasy, která katalysuje za účasti ATP navázání CO 2 na BCCP, čímž je CO 2 převeden do aktivované formy c) karboxyltransferasy, která přenáší CO 2 z BCCPbiotinu na acetyl-coa za vzniku malonyl-coa

39 Reakci katalysovanou tímto enzymovým komplexem tedy vystihuje schéma
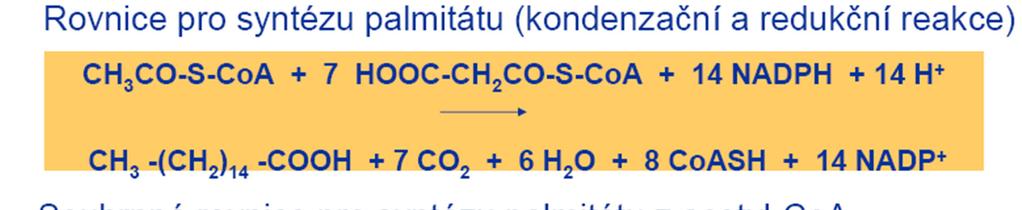
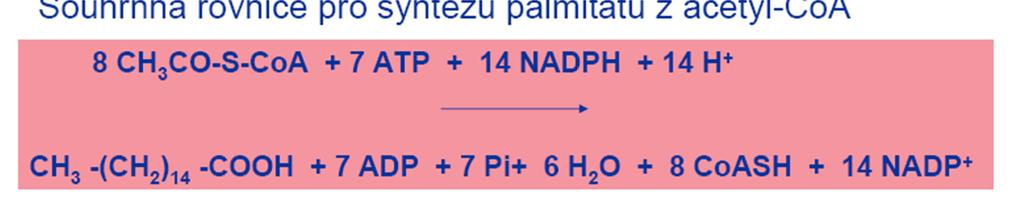
40 B. Synthesa palmitové kyseliny Sled reakcí, jimiž se z malonyl-coa syntetizuje základní šestnáctiuhlíkový řetězec palmitové kyseliny, zajišťuje multienzymový komplex synthetasy mastných kyselin Jeho součástí je ACP, jehož prosthetická skupina tvoří pohyblivé raménko, které slouží jako nosič substrátů a meziproduktů, vznikajících činností jednotlivých enzymů synthetasového komplexu váže je triolovou skupinou ( tzv. centrální SH, csh ) Malonyl-CoA, napojená na ACP, podstoupí sled reakcí kondensace, první redukce, dehydratace, druhá redukce a za odštěpení CO 2 prodlouží acyl ( v prvním cyklu acetyl ) o C 2 -jednotku Tento prodloužený acyl se přenese z csh raménka ACP na triolovou skupinu zbytku cysteinu jednoho z enzymů komplexu, β-oxoacylsynthetasy ( tzv. periférní SH, psh ) Na uvolněnou csh raménka ACP se připojí další malonylová skupina a proběhne nový sled reakcí Tyto cykly se opakují, až vznikne palmitový zbytek ( C 16 ), který se pak uvolní jako palmitoyl-coa
41 U většiny bakterií a v chloroplastech rostlin jsou ACP a enzymy synthetasy mastných kyselin diskrétní bílkoviny a jsou nekovalentně spojené v multienzymovou jednotku U živočichů jde o disociovatelný dimer, sestávající ze dvou polypeptidových podjednotek, z nichž každá má několik aktivních míst s různým typem enzymové aktivity
42 SLED REAKCÍ, KTERÝM SE MOLEKULA MASTNÉ KYSELINY VŽDY PRODLUŽUJE O DVA ATOMY UHLÍKU: 1. Transacylace. První cyklus zahrnuje přenos acetylu z acetyl-coa - nejprve na csh a pak transacylací na psh V dalších cyklech se přenáší z csh na psh acyl, který se v průběhu biosynthesy vytvořil Tím se uvolňuje csh pro vazbu malonylu ( viz 5. reakce )
43 2. Kondensace acylu s malonylem Při příblížení malonylové skupiny k csh pohyblivého raménka ACP se acyl ( v prvním cyklu acetyl ) uvolní z psh a napojí na malonyl účinkem tzv. kondensačního enzymu ( 3- oxoacylsynthetasy ) Ke spojení dochází adicí karbonylového uhlíku acylu na methylový uhlík malonylu za současné dekarboxylace; produktem je 3-oxoacyl
44 3. První redukce 3-oxoacylreduktasou za účasti NADPH se redukuje 3-oxoacyl na 3-hydoxyacyl 4. Dehydratace Účinkem lyasy (3-hydroxyacyldehydratasy, krotonasy) se odštěpí molekula vody a vznikne nenasycený acyl (enoyl)
45 5. Druhá redukce Enoyl se redukuje enoylreduktasou pomocí NADPH ( nebo NADH ) na nasycený acyl Reakce 3 až 5 probíhají na csh Vzniklý acyl se po ukončení cyklu acyltransferasou přenáší z csh na psh a na csh se váže nový malonyl a následuje nový cyklus reakcí 2 až 5 Jednotlivé cykly se opakují tak dlouho, až se dosáhne požadované délky řetězce Pak se proces zastaví allosterickou inhibicí Konečným produktem činnosti cytoplazmového multienzymového komplexu synthetasy mastných kyselin je zpravidla C 16 -kyselina
46 C. Další přeměny palmitátu Vyrobená mastná kyselina se uvolní z vazby na ACP a aktivuje se reakcí s CoA za účasti ATP a katalysy thiokinasou Vzniklý acyl-coa se může buď zapojit do synthesy lipidů, nebo se použije na výrobu kyselin s delším řetězcem, nenasycených kyselin nebo hydroxykyselin Tyto další úpravy však neprobíhají v cytosolu Řetězec se prodlužuje v mitochondriích přímo acetyl-coa bez použití malonyl-coa Nenasycené mastné kyseliny se pak tvoří specifickými NADP + -dyhydrogenasami;jsou stereospecifické a umožňují vznik cis-isomerů
47 Energetická stránka biosynthesy mastných kyselin Na prodloužení řetězce o C 2 -jednotku musí organismus vydat: 1 ATP na činnost biotinkarboxylasy při výrobě malonylu 2 NADPH ATP na dvojstupňovou redukci acetylu na CH 2 -CH 2 - Na výrobu mastné kyseliny o n uhlíkových atomech je tedy zapotřebí energie v jednotkách ATP = 7 ( n/2 1 ). Např. synthesa palmitové kyseliny stojí organismus 49 ATP. Pro srovnání při jejím zpětném odbourávání Lynenovou spirálou na acetyl-coa vzniká jen 5 (8 1) ATP 2ATP = 33 ATP, tedy něco přes 67% energie vynaložené na biosynthesu

48
49 BIOSYNTHESA PURINŮ A PYRIMIDINŮ Aminokyseliny jsou základ pro tvorbu všech ostatních biologických sloučenin dusíku, tzn. I dusíkatých složek nukleotidů, pyrimidinových i purinových bází Vzhledem k základnímu významu těchto látek pro funkci živé hmoty je jejich biosyntéza předpokladem pro růst a vývoj buněk, tkání i organismů a probíhá ve všech jejích formách
50 Biosyntesa pyrimidinů Pyrimidinové jádro vzniká ve formě volné báze z aspartátu Reakcí této aminokyseliny s karbamoylfosfátem vzniká N-karbamoylaspartát, který odštěpením vody tvoří cyklický amid, dihydroorotát Oxidací tohoto meziproduktu flavoproteinem dihydroorotátoxidasou (EC ) vzniká pyrimidinové jádro v orotátu, tj. karboxyuracilu Na tomto stupni reaguje pyrimidin a aktivovanou formou ribosafosfátu, 5-fosforibosyl-1-difosfátem za vzniku nukleotidu oriditin-5 -fosfátu Odštěpením karboxylové skupiny vzniká uridinmonofosfát (UMP), který dvojí reakcí s ATP přechází na UDP a UTP Tato látka je koenzymem glykosyltransferás a výchozí substrát pro syntézu RNA, ale také zdroj cytidintrifosfátu (CTP), který se z ní tvoří ligasovou reakcí s glutaminem

51 Zdroj:
52 Obě hlavní pyrimidinové složky nukleových kyselin vznikají společnou biosyntetickou cestou Biosyntéza probíhá v cytosolu a je tak oddělena od tvorby močoviny probíhající v mitochondriální matrix, ač oba pochody vycházejí z téhož substrátu karbamoylfosfátu Zdrojem této látky pro syntézu pyrimidinů je cytosolární karbamoylfosfátsynthasa (EC ), lišící se od mitochondriálního enzymu tím, že zdrojem dusíku není amoniak, ale glutamin Enzym není aktivován N-acetylglutamátem, ale je inhibován produktem celé biosyntetické cesty,utp Tento vztah je příkladem regulace biosyntézy mechanismem negativní zpětné vazby při dostatku pyrimidinových nukleotidů v buňce automaticky klesá syntéza výchozího substrátu jejich biosyntézy, karbamoylfosfátu
53 Biosynthesa purinů Na rozdíl od pyrimidinů se puriny syntetizují přímo ve formě nukleotidů Syntéza purinového jádra začíná tvorbou 5-fosforibosylaminu, tj. amonium-5 -nukleotidu reakcí glutaminu s 5-fosforibosyl-1- difosfátem Tato látka se acyluje glycinem za vzniku 5 fosforibosylglycinamidu Reakcí s 5,10-methylentetrahydrofolátem vzniká 5 -fosforibosyl-nformylglycinamid, který obsahuje všechny atomy imidazolového jádra purinu Přenosem amidového dusíku z glutaminu se mění amidová skupina na amidinovou v 5 -fosforibosyl-n-formylglycinamidinu nově zavedená aminoskupina je základ pro pyrimidinový kruh purinu V dalším kroku se ligasovou reakcí uzavírá imidazolový kruh na 5 fosforibosyl-5-aminoimidazol, který se dále karboxyluje CO 2 bez účasti známých karboxylačních koenzymů na 5 -fosforibosyl- 5.aminoimidazol-4-karboxylát V dalším reakčním stupni reakce se tvoří z karboxylu amid mechanismem analogickým tvorbě argininu z citrullinu zdrojem dusíku je aspartát, který se mění na fumarát
54 Formylací aminoskupiny 5 -fosforibosyl-5- aminoimidazol-4-karboxamidu 10- formyltetrahydrofolátem vzniklý 5 -fosforibosyl-5- formamidoimidazol-4-karboxamid obsahuje všechny atomy purinovéhojádra, které se pak uzavírá dehydratací za vzniku inosin-5 -fosfátu (IMP) Tato látka je výchozím substrátem pro syntézu obou významných purinových nukleotidů Náhradou kyslíku aminoskupinou z aspartátu (stejným mechanismem jako při tvorbě 5 -fosforibosyl- 5-aminoimidazol-4-karboxamidu) vzniká AMP Pro tvorbu GMP je nutná dehydrogenace IMP NAD + na xanthosin-5 -fosfát (XMP), v němž se pak nahrazuje karbonylový kyslík v poloze 6 aminoskupinou z glutaminu ligasovou reakcí

55 Zdroj:
56 Purinové jádro vzniká tedy jako mozaika částí procházejících z různých zdrojů Rozhodující úlohu při jeho tvorbě má tetrahydrofolátový systém přenosu C 1 -látek, který je proto nezbytnou složkou všech forem živé hmoty Protože efektivní funkce systému syntetizujícího puriny je esenciální pro rychle rostoucí a množící se buňky, jsou na ní mnohem více závislé buňky nádorové než normální somatické buňky Proto je jednou ze strategií v léčbě nádorových onemocnění blokáda tvorby purinů, např. antimetabolity kyseliny listové, které jsou mnohem toxičtější pro nádorové než pro somatické buňky
57 LITERATURA Mathews, Ch. K. Halde, K. E. Ahern, K. G., Biochemistry, 3. vydání, San Francisco, Benjamin Cummings, 1999, 1186 s., ISBN Voet, D. Voet, J. G. Pratt, Ch. W., Fundamentals of Biochemistry: life at the molecular level, 2. vydání, USA, Willey, 2006, 1264 s., ISBN
Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová
 Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová Vyberte esenciální aminokyseliny a) Asp, Glu b) Val, Leu, Ile c) Ala, Ser, Gly d) Phe, Trp Vyberte esenciální aminokyseliny a) Asp,
Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová Vyberte esenciální aminokyseliny a) Asp, Glu b) Val, Leu, Ile c) Ala, Ser, Gly d) Phe, Trp Vyberte esenciální aminokyseliny a) Asp,
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
Metabolizmus aminokyselin II
 Metabolizmus aminokyselin II Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol MUDr. Bc. Matej Kohutiar, Ph.D. matej.kohutiar@lfmotol.cuni.cz Praha 2018 Degradace uhlíkové kostry aminokyselin
Metabolizmus aminokyselin II Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol MUDr. Bc. Matej Kohutiar, Ph.D. matej.kohutiar@lfmotol.cuni.cz Praha 2018 Degradace uhlíkové kostry aminokyselin
Metabolismus mikroorganismů
 Metabolismus mikroorganismů Metabolismus organismů Souvisí s metabolismem polysacharidů, bílkovin, nukleových kyselin a lipidů Cytoplazma, mitochondrie (matrix, membrána) H 3 PO 4 Polysacharidy Pentózový
Metabolismus mikroorganismů Metabolismus organismů Souvisí s metabolismem polysacharidů, bílkovin, nukleových kyselin a lipidů Cytoplazma, mitochondrie (matrix, membrána) H 3 PO 4 Polysacharidy Pentózový
Metabolismus aminokyselin. Vladimíra Kvasnicová
 Metabolismus aminokyselin Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze dalších
Metabolismus aminokyselin Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze dalších
Metabolismus aminokyselin 2. Vladimíra Kvasnicová
 Metabolismus aminokyselin 2 Vladimíra Kvasnicová Odbourávání AMK 1) odstranění aminodusíku z molekuly AMK 2) detoxikace uvolněné aminoskupiny 3) metabolismus uhlíkaté kostry AMK 7 produktů 7 degradačních
Metabolismus aminokyselin 2 Vladimíra Kvasnicová Odbourávání AMK 1) odstranění aminodusíku z molekuly AMK 2) detoxikace uvolněné aminoskupiny 3) metabolismus uhlíkaté kostry AMK 7 produktů 7 degradačních
Metabolizmus aminokyselin II
 Metabolizmus aminokyselin II Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol dr. Matej Kohutiar, doc. Jana Novotná matej.kohutiar@lfmotol.cuni.cz Praha 2017 Degradace uhlíkové kostry aminokyselin
Metabolizmus aminokyselin II Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol dr. Matej Kohutiar, doc. Jana Novotná matej.kohutiar@lfmotol.cuni.cz Praha 2017 Degradace uhlíkové kostry aminokyselin
Propojení metabolických drah. Alice Skoumalová
 Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
Přehled energetického metabolismu
 Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Ukládání energie v buňkách
 Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Citrátový cyklus. Tomáš Kučera.
 itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Metabolizmus aminokyselin I
 Metabolizmus aminokyselin I Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol MUDr. Bc. Matej Kohutiar, Ph.D. matej.kohutiar@lfmotol.cuni.cz Praha 2018 snova I. přednáška: Metabolizmus a meziorgánové
Metabolizmus aminokyselin I Ústav lékařské chemie a klinické biochemie 2.LF UK a FN Motol MUDr. Bc. Matej Kohutiar, Ph.D. matej.kohutiar@lfmotol.cuni.cz Praha 2018 snova I. přednáška: Metabolizmus a meziorgánové
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
12. Metabolismus lipidů a glycerolu. funkce karnitinu a β-oxidace
 12. Metabolismus lipidů a glycerolu funkce karnitinu a β-oxidace LIPOPROTEINY Řadí se mezi složené lipidy Vznikají spojením (hydrofobními interakcemi nepolárních oblastí obou složek) lipidů se specifickými
12. Metabolismus lipidů a glycerolu funkce karnitinu a β-oxidace LIPOPROTEINY Řadí se mezi složené lipidy Vznikají spojením (hydrofobními interakcemi nepolárních oblastí obou složek) lipidů se specifickými
Štěpení lipidů. - potravou přijaté lipidy štěpí lipázy gastrointestinálního traktu
 METABOLISMUS LIPIDŮ ODBOURÁVÁNÍ LIPIDŮ - z potravy nebo z tukových rezerv - hydrolytické štěpení esterových vazeb - vznik glycerolu a mastných kyselin - hydrolytické štěpení LIPÁZY (karboxylesterázy) -
METABOLISMUS LIPIDŮ ODBOURÁVÁNÍ LIPIDŮ - z potravy nebo z tukových rezerv - hydrolytické štěpení esterových vazeb - vznik glycerolu a mastných kyselin - hydrolytické štěpení LIPÁZY (karboxylesterázy) -
Bp1252 Biochemie. #8 Metabolismus živin
 Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
14. Detoxikace amoniaku. ornithinový cyklus, odbourání nukleotidů
 14. Detoxikace amoniaku ornithinový cyklus, odbourání nukleotidů METABOLISMUS AMONIAKU Aminoskupiny se použijí: při synthese dusík obsahujících částí biomolekul sloučenin (aminokyseliny puriny a pirimidiny,
14. Detoxikace amoniaku ornithinový cyklus, odbourání nukleotidů METABOLISMUS AMONIAKU Aminoskupiny se použijí: při synthese dusík obsahujících částí biomolekul sloučenin (aminokyseliny puriny a pirimidiny,
Metabolismus bílkovin. Václav Pelouch
 ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus aminokyselin I. Jana Novotná 2. LF UK, Ústav lékařské chemie a klinické biochemie
 Metabolismus aminokyselin I Jana Novotná 2. LF UK, Ústav lékařské chemie a klinické biochemie Metabolismus aminokyselin PROTEINY Z POTRAVY GLYKOLÝZA KREBSŮV CYCLUS Proteosyntéza Trávení Transaminace TĚLESNÉ
Metabolismus aminokyselin I Jana Novotná 2. LF UK, Ústav lékařské chemie a klinické biochemie Metabolismus aminokyselin PROTEINY Z POTRAVY GLYKOLÝZA KREBSŮV CYCLUS Proteosyntéza Trávení Transaminace TĚLESNÉ
Struktura proteinů. - testík na procvičení. Vladimíra Kvasnicová
 Struktura proteinů - testík na procvičení Vladimíra Kvasnicová Mezi proteinogenní aminokyseliny patří a) kyselina asparagová b) kyselina glutarová c) kyselina acetoctová d) kyselina glutamová Mezi proteinogenní
Struktura proteinů - testík na procvičení Vladimíra Kvasnicová Mezi proteinogenní aminokyseliny patří a) kyselina asparagová b) kyselina glutarová c) kyselina acetoctová d) kyselina glutamová Mezi proteinogenní
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolismus dusíkatých látek
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus dusíkatých látek Oxidace aminokyselin Podíl AK na metabolické E se silně liší dle organismu a jeho momentálních potřeb, např.
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus dusíkatých látek Oxidace aminokyselin Podíl AK na metabolické E se silně liší dle organismu a jeho momentálních potřeb, např.
Aminokyseliny. Gymnázium a Jazyková škola s právem státní jazykové zkoušky Zlín. Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití
 Aminokyseliny Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek proteiny 18.7.2012 3. ročník čtyřletého G Určování postranních řetězců aminokyselin
Aminokyseliny Tematická oblast Datum vytvoření Ročník Stručný obsah Způsob využití Autor Kód Chemie přírodních látek proteiny 18.7.2012 3. ročník čtyřletého G Určování postranních řetězců aminokyselin
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Metabolismus sacharidů. VY_32_INOVACE_Ch0216.
 Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
PROTEINY. Biochemický ústav LF MU (H.P.)
") PROTEINY Biochemický ústav LF MU 2013 - (H.P.) 1 proteiny peptidy aminokyseliny 2 Aminokyseliny 3 Charakteristika základní stavební jednotky proteinů geneticky kódované 20 základních aminokyselin 4 a-aminokyselina
PROTEINY Biochemický ústav LF MU 2013 - (H.P.) 1 proteiny peptidy aminokyseliny 2 Aminokyseliny 3 Charakteristika základní stavební jednotky proteinů geneticky kódované 20 základních aminokyselin 4 a-aminokyselina
11. Metabolismus lipidů
 11. Metabolismus lipidů Obtížnost A Následující procesy a metabolické reakce, vedoucí ke zkrácení řetězce mastné kyseliny, vázané v triacylglycerolu, a vzniku acetyl-coa, seřaďte ve správném pořadí: a)
11. Metabolismus lipidů Obtížnost A Následující procesy a metabolické reakce, vedoucí ke zkrácení řetězce mastné kyseliny, vázané v triacylglycerolu, a vzniku acetyl-coa, seřaďte ve správném pořadí: a)
Biosyntéza a metabolismus bílkovin
 Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Biochemie jater. Eva Samcová
 Biochemie jater Eva Samcová Orgánová specializace Hlavní metabolické dráhy pro glukosu, mastné kyseliny a aminokyseliny jsou soustředěné okolo pyruvátu a acetyl-coa. Glukosa je primárním palivem pro mozek
Biochemie jater Eva Samcová Orgánová specializace Hlavní metabolické dráhy pro glukosu, mastné kyseliny a aminokyseliny jsou soustředěné okolo pyruvátu a acetyl-coa. Glukosa je primárním palivem pro mozek
Bílkoviny - proteiny
 Bílkoviny - proteiny Proteiny jsou složeny z 20 kódovaných aminokyselin L-enantiomery Chemická struktura aminokyselin R představuje jeden z 20 různých typů postranních řetězců R Hlavní řetězec je neměnný
Bílkoviny - proteiny Proteiny jsou složeny z 20 kódovaných aminokyselin L-enantiomery Chemická struktura aminokyselin R představuje jeden z 20 různých typů postranních řetězců R Hlavní řetězec je neměnný
Odbourávání a syntéza glukózy
 Odbourávání a syntéza glukózy Josef Fontana EB - 54 Obsah přednášky Glukóza význam glukózy pro buňku, glykémie role glukózy v metabolismu transport glukózy přes buněčné membrány enzymy fosforylující a
Odbourávání a syntéza glukózy Josef Fontana EB - 54 Obsah přednášky Glukóza význam glukózy pro buňku, glykémie role glukózy v metabolismu transport glukózy přes buněčné membrány enzymy fosforylující a
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová
 Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/ Anotace. Metabolismus lipidů - odbourávání. VY_32_INOVACE_Ch0212
 Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
DYNAMICKÁ BIOCHEMIE. Daniel Nechvátal :: www.gymzn.cz/nechvatal
 DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
Názvosloví cukrů, tuků, bílkovin
 Názvosloví cukrů, tuků, bílkovin SACARIDY CUKRY MNSACARIDY LIGSACARIDY PLYSACARIDY (z mnoha molekul monosacharidů) ALDSY KETSY -DISACARIDY - TRISACARIDY - TETRASACARIDY atd. -aldotriosy -aldotetrosy -aldopentosy
Názvosloví cukrů, tuků, bílkovin SACARIDY CUKRY MNSACARIDY LIGSACARIDY PLYSACARIDY (z mnoha molekul monosacharidů) ALDSY KETSY -DISACARIDY - TRISACARIDY - TETRASACARIDY atd. -aldotriosy -aldotetrosy -aldopentosy
Obecná struktura a-aminokyselin
 AMINOKYSELINY Obsah Obecná struktura Názvosloví, třídění a charakterizace Nestandardní aminokyseliny Reaktivita - peptidová vazba Biogenní aminy Funkce aminokyselin Acidobazické vlastnosti Optická aktivita
AMINOKYSELINY Obsah Obecná struktura Názvosloví, třídění a charakterizace Nestandardní aminokyseliny Reaktivita - peptidová vazba Biogenní aminy Funkce aminokyselin Acidobazické vlastnosti Optická aktivita
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Pentosový cyklus. osudy glykogenu. Eva Benešová
 Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Metabolismus lipidů. (pozn. o nerozpustnosti)
") Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
Gymnázium Vysoké Mýto nám. Vaňorného 163, 566 01 Vysoké Mýto
 Gymnázium Vysoké Mýto nám. Vaňorného 163, 566 01 Vysoké Mýto SUBSTITUČNÍ DERIVÁTY KARBOXYLOVÝCH O KYSELIN R C O X karboxylových kyselin - substituce na vedlejším uhlovodíkovém řetězci aminokyseliny - hydroxykyseliny
Gymnázium Vysoké Mýto nám. Vaňorného 163, 566 01 Vysoké Mýto SUBSTITUČNÍ DERIVÁTY KARBOXYLOVÝCH O KYSELIN R C O X karboxylových kyselin - substituce na vedlejším uhlovodíkovém řetězci aminokyseliny - hydroxykyseliny
Metabolismus dusíkatých látek
 Metabolismus dusíkatých látek Bílkoviny (aminokyseliny) Nukleové kyseliny (nukleotidy) Koloběh dusíku v biosféře Koloběh dusíku v biosféře: 1: působení hlízkovitých bakterií; 2: asimilace pomocí nitrátreduktasy
Metabolismus dusíkatých látek Bílkoviny (aminokyseliny) Nukleové kyseliny (nukleotidy) Koloběh dusíku v biosféře Koloběh dusíku v biosféře: 1: působení hlízkovitých bakterií; 2: asimilace pomocí nitrátreduktasy
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty
 sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Glykolýza a neoglukogenese
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Procvičování aminokyseliny, mastné kyseliny
 Procvičování aminokyseliny, mastné kyseliny Co je hlavním mechanismem pro odstranění aminoskupiny před odbouráváním většiny aminokyselin: a. oxidativní deaminace b. transaminace c. dehydratace d. působení
Procvičování aminokyseliny, mastné kyseliny Co je hlavním mechanismem pro odstranění aminoskupiny před odbouráváním většiny aminokyselin: a. oxidativní deaminace b. transaminace c. dehydratace d. působení
METABOLISMUS SACHARIDŮ
 METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolismus sacharidů II
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus sacharidů II NUTNO ZNÁT VSTUP TĚCHTO ZÁKLADNÍCH MONOSACHARIDŮ DO GLYKOLÝZY Glykogen glukosa hlavní zdroj energie pro metabolismus
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus sacharidů II NUTNO ZNÁT VSTUP TĚCHTO ZÁKLADNÍCH MONOSACHARIDŮ DO GLYKOLÝZY Glykogen glukosa hlavní zdroj energie pro metabolismus
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Syntéza a degradace mastných kyselin. Martina Srbová
 Syntéza a degradace mastných kyselin Martina Srbová Mastné kyseliny (fatty acids, FA) většinou sudý počet atomů uhlíku a lineární řetězec v esterifikované formě jako součást lipidů v neesterifikované formě
Syntéza a degradace mastných kyselin Martina Srbová Mastné kyseliny (fatty acids, FA) většinou sudý počet atomů uhlíku a lineární řetězec v esterifikované formě jako součást lipidů v neesterifikované formě
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Citrátový a glyoxylátový cyklus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolusmus lipidů - anabolismus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - anabolismus LIPIDY Zásobárna energie Hlavní složka buněčných membrán Pigmenty (retinal, karoten), kofaktory (vitamin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - anabolismus LIPIDY Zásobárna energie Hlavní složka buněčných membrán Pigmenty (retinal, karoten), kofaktory (vitamin
Metabolismus aminokyselin SOUHRN. Vladimíra Kvasnicová
 Metabolismus aminokyselin SOUHRN Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze
Metabolismus aminokyselin SOUHRN Vladimíra Kvasnicová Aminokyseliny aminokyseliny přijímáme v potravě ve formě proteinů: důležitá forma organicky vázaného dusíku, který tak může být v těle využit k syntéze
Integrace metabolických drah v organismu. Zdeňka Klusáčková
 Integrace metabolických drah v organismu Zdeňka Klusáčková Hydrolýza a resorpce základních složek potravy Přehled hlavních metabolických drah Biochemie výživy A) resorpční fáze (přísun živin) glukóza hlavní
Integrace metabolických drah v organismu Zdeňka Klusáčková Hydrolýza a resorpce základních složek potravy Přehled hlavních metabolických drah Biochemie výživy A) resorpční fáze (přísun živin) glukóza hlavní
Otázka: Metabolismus. Předmět: Biologie. Přidal(a): Furrow. - přeměna látek a energie
: Furrow. - přeměna látek a energie") Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Didaktické testy z biochemie 2
 Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
Biochemicky významné sloučeniny a reakce - testík na procvičení
 Biochemicky významné sloučeniny a reakce - testík na procvičení Vladimíra Kvasnicová Vyberte pravdivé(á) tvrzení o heterocyklech: a) pyrrol je součástí struktury hemu b) indol je součástí struktury histidinu
Biochemicky významné sloučeniny a reakce - testík na procvičení Vladimíra Kvasnicová Vyberte pravdivé(á) tvrzení o heterocyklech: a) pyrrol je součástí struktury hemu b) indol je součástí struktury histidinu
8. Polysacharidy, glykoproteiny a proteoglykany
 Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Metabolismus sacharidů
 Metabolismus sacharidů Glukosa obsažená v celulose, škrobu a oligosacharidech nebo volná je nejrozšířenější organickou sloučeninou v přírodě. Pro chemotrofní organismy jsou sacharidy hlavní živinou, přičemž
Metabolismus sacharidů Glukosa obsažená v celulose, škrobu a oligosacharidech nebo volná je nejrozšířenější organickou sloučeninou v přírodě. Pro chemotrofní organismy jsou sacharidy hlavní živinou, přičemž
Konsultační hodina. základy biochemie pro 1. ročník. Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa
 Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Inovace bakalářského studijního oboru Aplikovaná chemie http://aplchem.upol.cz
 Inovace bakalářského studijního oboru Aplikovaná chemie http://aplchem.upol.cz Z.1.07/2.2.00/15.0247 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Funkční
Inovace bakalářského studijního oboru Aplikovaná chemie http://aplchem.upol.cz Z.1.07/2.2.00/15.0247 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Funkční
Vzdělávací materiál. vytvořený v projektu OP VK. Anotace. Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20. Číslo projektu:
 Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Metabolismus lipidů. Biosyntéza mastných kyselin a triacylglycerolů. Lenka Fialová kařské biochemie 1. LF UK. Hlavní rysy biosyntézy mastných kyselin
 Metabolismus lipidů Biosyntéza mastných kyselin a triacylglycerolů Lenka Fialová Ústav lékal kařské biochemie 1. LF UK Hlavní rysy biosyntézy mastných kyselin syntéza MK může probíhat ve většině živočišných
Metabolismus lipidů Biosyntéza mastných kyselin a triacylglycerolů Lenka Fialová Ústav lékal kařské biochemie 1. LF UK Hlavní rysy biosyntézy mastných kyselin syntéza MK může probíhat ve většině živočišných
- metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy a jejich životním prostředím
 Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Bílkoviny. Charakteristika a význam Aminokyseliny Peptidy Struktura bílkovin Významné bílkoviny
 Bílkoviny harakteristika a význam Aminokyseliny Peptidy Struktura bílkovin Významné bílkoviny 1) harakteristika a význam Makromolekulární látky složené z velkého počtu aminokyselinových zbytků V tkáních
Bílkoviny harakteristika a význam Aminokyseliny Peptidy Struktura bílkovin Významné bílkoviny 1) harakteristika a význam Makromolekulární látky složené z velkého počtu aminokyselinových zbytků V tkáních
Metabolismus aminokyselin II. Močovinový cyklus
 Metabolismus aminokyselin II. Močovinový cyklus Osnova Zdroje dusíku jako odpadního produktu metabolismu aminokyselin. Meziorgánový tok aminokyselin. Zdroje aminodusíku pro močovinový cyklus Sled reakcí
Metabolismus aminokyselin II. Močovinový cyklus Osnova Zdroje dusíku jako odpadního produktu metabolismu aminokyselin. Meziorgánový tok aminokyselin. Zdroje aminodusíku pro močovinový cyklus Sled reakcí
fce jater: (chem. továrna, jako 1. dostává všechny látky vstřebané GIT) METABOLICKÁ (jsou metabolicky nejaktivnější tkání v těle)
 METABOLICKÁ (jsou metabolicky nejaktivnější tkání v těle)") JÁTRA ústřední orgán intermed. metabolismu, vysoká schopnost regenerace krevní oběh játry: (protéká 20% veškeré krve, 10-30% okysl.tep.krve, která zajišťuje výživu buněk, zbytek-portální krev) 1. funkční
JÁTRA ústřední orgán intermed. metabolismu, vysoká schopnost regenerace krevní oběh játry: (protéká 20% veškeré krve, 10-30% okysl.tep.krve, která zajišťuje výživu buněk, zbytek-portální krev) 1. funkční
Metabolismus proteinů a aminokyselin
 Metabolismus proteinů a aminokyselin Proteiny jsou nejdůležitější složkou potravy všech živočichů, nelze je nahradit ani cukry, ani lipidy. Je to proto, že organismus živočichů nedokáže ve svých metabolických
Metabolismus proteinů a aminokyselin Proteiny jsou nejdůležitější složkou potravy všech živočichů, nelze je nahradit ani cukry, ani lipidy. Je to proto, že organismus živočichů nedokáže ve svých metabolických
AMINOKYSELINY REAKCE
 CHEMIE POTRAVIN - cvičení AMINOKYSELINY REAKCE Milena Zachariášová (milena.zachariasova@vscht.cz) Ústav chemie a analýzy potravin, VŠCHT Praha REAKCE AMINOKYSELIN část 1 ELIMINAČNÍ REAKCE DEKARBOXYLACE
CHEMIE POTRAVIN - cvičení AMINOKYSELINY REAKCE Milena Zachariášová (milena.zachariasova@vscht.cz) Ústav chemie a analýzy potravin, VŠCHT Praha REAKCE AMINOKYSELIN část 1 ELIMINAČNÍ REAKCE DEKARBOXYLACE
 Aminokyseliny, proteiny, enzymy Základy lékařské chemie a biochemie 2014/2015 Ing. Jarmila Krotká Metabolismus základní projev života látková přeměna souhrn veškerých dějů, které probíhají uvnitř organismu
Aminokyseliny, proteiny, enzymy Základy lékařské chemie a biochemie 2014/2015 Ing. Jarmila Krotká Metabolismus základní projev života látková přeměna souhrn veškerých dějů, které probíhají uvnitř organismu
Aminokyseliny, peptidy a bílkoviny
 Aminokyseliny, peptidy a bílkoviny Dělení aminokyselin Z hlediska obsahu v živé hmotě Z hlediska významu ve výživě Z chemického hlediska Z hlediska rozpustnosti Dělení aminokyselin Z hlediska obsahu v
Aminokyseliny, peptidy a bílkoviny Dělení aminokyselin Z hlediska obsahu v živé hmotě Z hlediska významu ve výživě Z chemického hlediska Z hlediska rozpustnosti Dělení aminokyselin Z hlediska obsahu v
POZNÁMKY K METABOLISMU SACHARIDŮ
 POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
ANABOLISMUS SACHARIDŮ
 zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
NUTRACEUTIKA PROTEINY
 NUTRAEUTIKA PROTEINY VYUŽITÍ Proteiny, aminokyseliny, koncentráty většinou pro sportovní výživu Funkční potraviny hydrolyzáty Bílkovinné izoláty i v medicíně Fitness a wellness přípravky PROTEINY Sušená
NUTRAEUTIKA PROTEINY VYUŽITÍ Proteiny, aminokyseliny, koncentráty většinou pro sportovní výživu Funkční potraviny hydrolyzáty Bílkovinné izoláty i v medicíně Fitness a wellness přípravky PROTEINY Sušená
Aminokyseliny a dlouhodobá parenterální výživa. Luboš Sobotka
 Aminokyseliny a dlouhodobá parenterální výživa Luboš Sobotka Reakce na hladovění a stres jsou stejné asi 4000000 let Přežít hladovění a akutní stav Metody sledování kvality AK roztoků Vylučovací metoda
Aminokyseliny a dlouhodobá parenterální výživa Luboš Sobotka Reakce na hladovění a stres jsou stejné asi 4000000 let Přežít hladovění a akutní stav Metody sledování kvality AK roztoků Vylučovací metoda
Metabolismus. Source:
 Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno:
 Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Odbourávání lipidů, ketolátky
 dbourávání lipidů, ketolátky Josef Fontana EB - 56 bsah přednášky Energetický význam TAG Jednotlivé dráhy metabolismu lipidů lipidy jako zdroj energie degradace TAG v buňkách, β-oxidace MK tvorba a využití
dbourávání lipidů, ketolátky Josef Fontana EB - 56 bsah přednášky Energetický význam TAG Jednotlivé dráhy metabolismu lipidů lipidy jako zdroj energie degradace TAG v buňkách, β-oxidace MK tvorba a využití
7. Enzymy. klasifikace, názvosloví a funkce
 7. Enzymy klasifikace, názvosloví a funkce Jsou to přírodní katalyzátory, živočišné i rostlinné Umožňují průběh biochemických reakcí Nachází se ve veškerých živých systémech Enzymy vykazují druhovou specifitu
7. Enzymy klasifikace, názvosloví a funkce Jsou to přírodní katalyzátory, živočišné i rostlinné Umožňují průběh biochemických reakcí Nachází se ve veškerých živých systémech Enzymy vykazují druhovou specifitu
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Genetický kód. Jakmile vznikne funkční mrna, informace v ní obsažená může být ihned použita pro syntézu proteinu.
 Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Integrace a regulace savčího energetického metabolismu
 Základy biochemie KBC / BCH Integrace a regulace savčího energetického metabolismu Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským
Základy biochemie KBC / BCH Integrace a regulace savčího energetického metabolismu Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským
Kofaktory enzymů. T. Kučera. (upraveno z J. Novotné)
") Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
NaLékařskou.cz Přijímačky nanečisto
 alékařskou.cz Chemie 2016 1) Vyberte vzorec dichromanu sodného: a) a(cr 2 7) 2 b) a 2Cr 2 7 c) a(cr 2 9) 2 d) a 2Cr 2 9 2) Vypočítejte hmotnostní zlomek dusíku v indolu. a) 0,109 b) 0,112 c) 0,237 d) 0,120
alékařskou.cz Chemie 2016 1) Vyberte vzorec dichromanu sodného: a) a(cr 2 7) 2 b) a 2Cr 2 7 c) a(cr 2 9) 2 d) a 2Cr 2 9 2) Vypočítejte hmotnostní zlomek dusíku v indolu. a) 0,109 b) 0,112 c) 0,237 d) 0,120
Biochemie dusíkatých látek při výrobě vína
 Biochemie dusíkatých látek při výrobě vína Ing. Michal Kumšta www.zf.mendelu.cz Ústav vinohradnictví a vinařství kumsta@mendelu.cz Vzdělávací aktivita je součástí projektu CZ.1.07/2.4.00/31.0089 Projekt
Biochemie dusíkatých látek při výrobě vína Ing. Michal Kumšta www.zf.mendelu.cz Ústav vinohradnictví a vinařství kumsta@mendelu.cz Vzdělávací aktivita je součástí projektu CZ.1.07/2.4.00/31.0089 Projekt
CZ.1.07/2.2.00/ Obecný metabolismu. Cyklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8).
 (8).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii Z.1.07/2.2.00/28.0171 becný metabolismu. yklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8). Prof. RNDr.
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii Z.1.07/2.2.00/28.0171 becný metabolismu. yklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8). Prof. RNDr.
Nukleové kyseliny. obecný přehled
 Nukleové kyseliny obecný přehled Nukleové kyseliny objeveny r.1868, izolovány koncem 19.stol., 1953 objasněno jejich složení Watsonem a Crickem (1962 Nobelova cena) biopolymery nositelky genetické informace
Nukleové kyseliny obecný přehled Nukleové kyseliny objeveny r.1868, izolovány koncem 19.stol., 1953 objasněno jejich složení Watsonem a Crickem (1962 Nobelova cena) biopolymery nositelky genetické informace
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Lipidy. Nesourodá skupina látek Látky nerozpustné v polárních rozpouštědlech Složky: MK, alkoholy, N látky, sacharidy, kyselina fosforečná
 Lipidy Nesourodá skupina látek Látky nerozpustné v polárních rozpouštědlech Složky: MK, alkoholy, N látky, sacharidy, kyselina fosforečná Rozdělení: 1. neutrální lipidy (tuky, triacylglyceroly) 2. Vosky
Lipidy Nesourodá skupina látek Látky nerozpustné v polárních rozpouštědlech Složky: MK, alkoholy, N látky, sacharidy, kyselina fosforečná Rozdělení: 1. neutrální lipidy (tuky, triacylglyceroly) 2. Vosky
Tomáš Kuˇ. cera. Ústav lékaˇrské chemie a klinické biochemie 2. lékaˇrská fakulta, Univerzita Karlova v Praze.
 BIOCHEMIE SVALU Tomáš Kuˇ cera tomas.kucera@lfmotol.cuni.cz Ústav lékaˇrské chemie a klinické biochemie 2. lékaˇrská fakulta, Univerzita Karlova v Praze 2014 STRUKTURA KOSTERNÍHO SVALU svazky svalových
BIOCHEMIE SVALU Tomáš Kuˇ cera tomas.kucera@lfmotol.cuni.cz Ústav lékaˇrské chemie a klinické biochemie 2. lékaˇrská fakulta, Univerzita Karlova v Praze 2014 STRUKTURA KOSTERNÍHO SVALU svazky svalových