Co je vidět v buňkách? Inoue, 50. léta: vláknité struktury (meióza mikrospor Lilium longiflorum)
|
|
|
- Šimon Šimek
- před 7 lety
- Počet zobrazení:
Transkript
1 Cytoskelet
2 Co je vidět v buňkách? Inoue, 50. léta: vláknité struktury (meióza mikrospor Lilium longiflorum)

3 Aktin a tubulin tubulin (a DNA) v pylové láčce tabáku aktin v pylové láčce tabáku tubulin v epidermis listů tabáku aktin v rhizodermis Arabidopsis


4 Co je vidět v buňkách? kořen Arabidopsis Ledbetter MC, Porter KR (1964): Morphology of Microtubules of Plant Cell.
5 Endosperm, Haemanthus sp.

6 Obecné rysy cytoskeletu vlákna jsou polymery z několika málo typů podjednotek diverzitu a nejrůznější funkce jim udílejí asociované proteiny

7 Obecné funkce cytoskeletu vazba organel a multimolekulárních komplexů

8 Obecné funkce cytoskeletu řízený vnitrobuněčný pohyb proudění cytoplasmy cytoplasmic streaming


9 ER and clathrin-coated vesicles associated with AF and MT řízený vnitrobuněčný pohyb

10 Obecné funkce cytoskeletu buněčná polarita

11 Vznik polárního vlákna z monomerů obecný princip u aktinu i tubulinu

12 Vznik polárního vlákna z monomerů
13 Základní cytoskeletální systémy eukaryot Aktinový (mikrofilamenta) všude (prokaryotní homolog MreB) talin-rfp Tubulinový (mikrotubuly) všude (prokaryotní homolog FtsZ) Intermediální filamenta klasická zatím jen u živočichů keratin, vimentin, neurofilamenta, laminy... MAP4-GFP
 farmakologické")

14 Metody studia cytoskeletu mikroinjekce nepřímá fluorescence infiltrace agrobakterií do listů tabáku exprese fúzních fluorescenčních proteinů přímá fluorescence (faloidin) farmakologické působení (stabilizující x destabilizující látky) tubulindestabilizing tubulin-stabilizing actin-destabilizing actin-stabilizing oryzalin taxol latrunculin B phalloidin nocodazole propyzamid cytochalasin D
15 Aktinový cytoskelet

16 Vznik polárního vlákna aktinu aktin
-decorated actin filament displays a helical rope-like appearance; the thicker part of a turn is directed to the pointed end of a filament (direction of the arrowhead).")
17 Vznik polárního vlákna z monomerů (a). S1 (myosin fragment)-decorated actin filament displays a helical rope-like appearance; the thicker part of a turn is directed to the pointed end of a filament (direction of the arrowhead). (b), part of an actin filament bundle from a REF-52 fibroblast after S1 decoration. Bars, 0.1 um T. M. Svitkina and G.G. Borisy, "Methods in Enzymology", volume "Molecular Motors and the Cytoskeleton: Part B", 298: , 1998.

18 Dynamika aktinových vláken: polymerace a treadmilling pointed end barbed end ATP-bound ADP-bound
19 Proteiny ovlivňující dynamiku aktinu



20 Proteiny vázající G-aktin: regulace dostupnosti podjednotek sequestered G-actin here: thymosine β4 may be vertebrate-specific? fosfolipáza C Profilin Tβ4 a profilin vážou přednostně ATP-G-aktin udržují vysokou pohotovostní hladinu aktinu pro rychlou polymeraci

21 Profilin Tradescantia stamen hairs
22 Profilin
 10 µm, (B) 16 µm, (C) 22 µm, and (D) 62 µm. Bar, 10 µm.")
 Control cell.")
23 Injekce profilinu inhibuje růst pylových láček a narušuje strukturu aktinu Pollen tubes injected with increasing doses of native pollen profilin. Leftmost cell - before injection, rightmost 20 min after injection. (A) 10 µm, (B) 16 µm, (C) 22 µm, and (D) 62 µm. Bar, 10 µm. F-actin distribution of profilin-injected cells after chemical fixation and Alexa-phalloidin staining. Twenty minutes after injection cells were chemically fixed and stained with 0.3 µm Alexa-phalloidin. (A) Control cell. (B) Cell injected with a low concentration of profilin (~30 µm). (C) Cell injected with a higher concentration of profilin (>60 µm). Vidali et al. 2001
24 Stříhání mikrofilament: ADF/cofilin cofilin = Actin Depolymerizing Factor (ADF) Condeelis 2001
25 Stříhání mikrofilament: ADF/cofilin Condeelis 2001

26 Změny hladiny ADF1 u Arabidopsis overexprese: vymizení svazků mikrofilament (cables) menší buňky inhibice: více svazků mikrofilament Dong et al sense cdna anti-sense cdna ADF1
27 Změny hladiny ADF1 u Arabidopsis WT AtADF1-O AtADF1-U Dong et al. 2001
: WT: 16,57 ± 0,95 ADF1-O: 16,46 ± 1,10 AtADF1-U: 25,61 ± 1,90 Dong et al.")
28 Změny hladiny ADF1 u Arabidopsis overexprese: omezení růstu buněk/ogánů inhibice: podpora růstu buněk/ogánů Počet listů v růžici před vykvetením (vliv na indukci kvetení): WT: 16,57 ± 0,95 ADF1-O: 16,46 ± 1,10 AtADF1-U: 25,61 ± 1,90 Dong et al ADF je jedním z klíčových regulátorů aktinového cytoskeletu!

29 Profilin versus Cofilin Deeks et al. 2002
30 Nukleace aktinových filament de novo


31 Nukleace aktinových filament de novo Arp2/3 komplex regulátory: WASP SCAR/WAWE Condeelis, 2001

32 Nukleace aktinových filament de novo

33

34 SCAR/WAWE: regulátory Arp2/3 Zhang, X., et al. Plant Cell 2005;17: The evolutionarily conserved Rac-WAVE-Arp2/3 pathway links actin filament nucleation to cell morphogenesis. WAVE translates Rac-GTP signals into Arp2/3 activation by regulating the stability and/or localization of the activator subunit Scar/WAVE. The WAVE complex includes: 1) Sra1/PIR121/CYFIP1, 2) Nap1/NAP125, 3) Abi-1/Abi-2, 4) Brick1(Brk1)/HSPC300, 5) Scar/WAVE Arabidopsis BRICK1/HSPC300 Is an Essential WAVE-Complex Subunit that Selectively Stabilizes the Arp2/3 Activator SCAR2. Current Biology, Volume 16, Issue 9, Pages J. Le, E. Mallery, C. Zhang, S. Brankle, D. Szymanski WT itb1/scar2
 arp2 brk1 arpc5")
35 WAVE-complex stabilizes the Arp2/3 activator SCAR2. WT arp2 arpc5 sra1 brk1 (BRICK) arp2 brk1 arpc5 brk1
36 Podjednotky Arp2/3 komplexu jsou evolučně staré Arabidopsis: ARPC1/Arc40p/p40 (2) ARPC2/Arc35p/p35 (2) ARPC3/Arc18p/p21 (1) ARPC4/Arc19p/p20 (1) ARPC5/Arc15p/p15 (1) McKinney et al. 2002; Mathur 2005
37 Arabidopsis: mutanti třídy distorted1 Podjednotky a regulátory Arp2/3 komplexu! (Mathur, 2005)

38 Arabidopsis: mutanti třídy distorted WT crooked spirrig wurm

39 autofluorescence GFP-mTalin WT wurm

40 hypokotyl GFP-mTalin trichomy Golgi agregáty! WT crooked

41 Další způsoby nukleace: forminy (FH2 proteiny) overexpression of a partial yeast formin causes formation of ectopic actin structures overexpression of formins stimulates filament formation in vitro
 FH12-FH21 bez tm. domény (Class II) Zigmond, Curr. Opin. Cell Biol. 2003; Cvrčková et al.")
42 Doménová struktura forminů Pro-rich, profilin-binding, nukleace Pro A rich FH1 FH2 B FH1 repeat FH2 aktivace Rho GTPázou C FH2 D PTEN FH1 FH2 E FH1 FH2 coiled F FH1 coil FH2 Arabidopsis: 21 genů! FH1-FH11 transmembránová doména (Class I) FH12-FH21 bez tm. domény (Class II) Zigmond, Curr. Opin. Cell Biol. 2003; Cvrčková et al., BMC Genomics 2004


43 Kráčející nukleační komplex dimerizační rozhraní FH1 FH2 leaky cap model - živočišný Otomo et al. Nature 433: , 2005 u rostlin? vazba na MT?
 AtFH1:GFP")

44 Overexprese AtFH1 v pylových láčkách kontrola talin AtFH1 (FH1/2) AtFH1:GFP Membránová lokalizace forminů třídy I typická pro rostliny (Cheung a Wu, 2004)
45 Proč mají rostliny právě tyto geny v tolika kopiích? Kromě funkčního rozrůznění i jemné ladění pro proměnlivé podmínky v čase (rostlina je přisedlá) v (mikro)prostoru buňky obklopené stěnou? Možnosti se nevylučují (viz též Nasmyth, Dirick, Surana, Amon, Cvrčková, 1991)

 (Ingouff et al.")
46 Specifické funkce isoforem forminů AtFH1: bundling AtFH5: cytokineze endospermu AtFH6: zvýšená exprese v hálkách indukovaných hlísticemi, suprimuje mutaci bni1 u kvasinky AtFH4/AtFH8: kořenově specifické, slabý defekt na kořenových vláscích mutant fh5 má zpožděnou celularizaci endospermu (jinde redundantní) (Ingouff et al. 2005) mutace ve forminech u různých organismů často vedou k defektům v cytokinezi
47 Intermediální filamenta

48 Intermediální filamenta: konvergence? regulace např. fosforylací (CDK... jaderná lamina) Výjimka z pravidla: rovnocenné konce!
49 Epitopy příbuzné IF v rostlinách? interfáze kolokalizace s MT? profáze mitóza a cytokineze wheat root tip, anti-keratin

50 Biochem J July 15; 261(2):

51 Mikroinjekce značených protilátek anti-if DIOC6 - membrány

52 Jaderná IF - laminy Asi jen u živočichů?
53 Jak u rostlin? nacházíme laminové epitopy, ale v genomu A. th. není ortolog rekonstituce jader Xenopus v bezbuněčných extraktech z tabáku (Lu and Zhai 2001) savčí LBR se lokalizuje do jaderné membrány v tabáku (Irons et al. 2003)
 Portion of the peripheral lamina with a complex structure (lam), connected with the internal matrix (inm). Individual filaments are observed (arrow).")
54 Cell Biology International Volume 27, Issue 3, 2003, Pages C. de la Torre Fig. 1. Ultrastructural organisation of the plant NM in resinless sections. (A) Portion of the peripheral lamina with a complex structure (lam), connected with the internal matrix (inm). Individual filaments are observed (arrow). (B) A delicate anastomosed network of filaments forms the internal matrix: nm knobbed filaments (arrows) and thicker ones, covered by globular structures (big arrow;dense aggregates (da). (C) Higher magnification of a branched filament of the internal matrix, showing the junction of two filaments and a typical 25 nm knob. Bar in (A), 0.1 µm; in (B), 0.5 µm; and in (C), 0.1 µm. Identification of the IF proteins by Western blot in the onion NM. (D) The anti-numa serum, S2, reveals three onion isoforms, the major one at 220 kd (arrow). (E) Antibodies against chicken B2and A lamins, and a serum against the chicken lamina (Ls) recognise in all cases protein spots at 65 kd with pi varying from 5.65 to 6.8. (F) 2-D blot with serum 288 against LeMFP1. The 90 kd AcMFP1 shows up to 12 basic spots. In situ localisation of the IF proteins in the NM. (G) NuMA is associated with the internal matrix network and accumulates in small foci with a punctate pattern. (H) Lamin-like proteins accumulate not only at the peripheral lamina, but also in internal foci. (I) AcMFP1 accumulation in large replication foci from a late-s phase NM.

55 Rostliny sice nemají laminy, ale...
56 Mikrotubulární cytoskelet

57 Mikrotubulární systémy rostlinné buňky

58 Tradescantia, injected bovine tubulin

59 existuje preference směrů Helikální uspořádání MT


60 Helikální uspořádání MT

61 světlo SPR1, SPR2 (spiral) etiolované Arabidopsis spiral1 (spr1) mutants show a right-handed helical growth in roots and etiolated hypocotyls due to impaired directional growth of rapidly expanding cells.


62 SPR1: malý protein na +koncích MT SPR1 encodes a small protein. Its localization to cortical microtubules (MTs) suggests that SPR1 maintains directional cell expansion by regulating cortical MT functions.


63
 mor1 lefty1, lefty2: mutace tubulinu Točí se rostlina bez mikrotubulů doleva?")
64 Točivost orgánů mutantů: směr nemusí souhlasit se směrem mt spirály Pravotočivé: spr1 Levotočivé: inhibitory (taxol) mor1 lefty1, lefty2: mutace tubulinu Točí se rostlina bez mikrotubulů doleva? A jak to dělá??
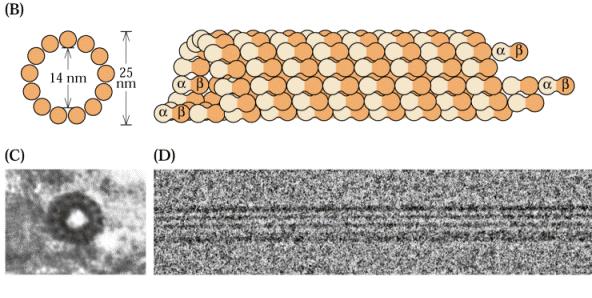
65 Struktura mikrotubulů
66 Dynamika přestaveb mikrotubulů animace
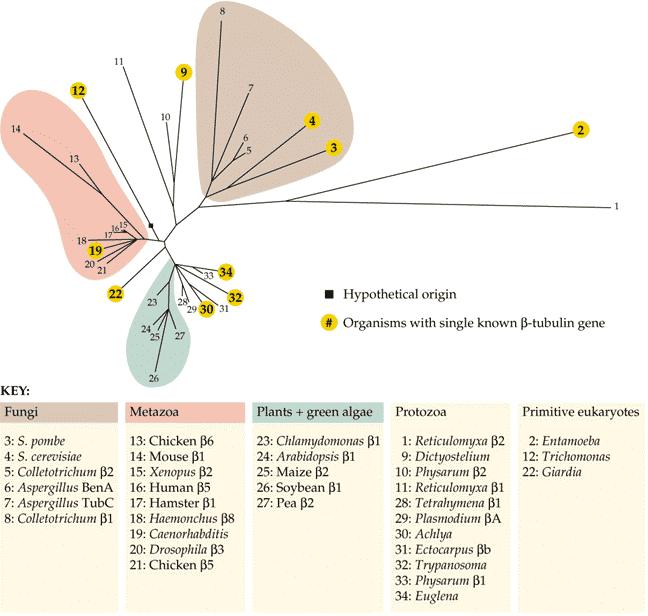
67 Rodiny tubulinových genů

68

69 tubulinové háčky v isol. fragmoplastech v nadbytku tubulinu


70 Nukleace mikrotubulů γ-tubulin mikrotubulární organizační centra (MTOC) rostliny nemají žádná definovaná MTOC

71 MT recovery after drug-induced disassembly



72 Tobacco cell ghosts; Murata et al., Nat.Cell.Biol. Oct. 2005

73 Nukleace γ-tubulinem - model

74 Modulace dynamiky mikrotubulů - MAPs
75
76 End binding protein (EB1) EB1 váže +konce MT


77 Jak se proteiny mohou dostat na +konec MT?


78 Katanin - p60 a p80 ATPase subunit, which functions to sever microtubules regulates the activity of the ATPase and localizes the protein to the centrosomes, not present in plants Mutace botero, fragile fibre2: brittle swollen semi-dwarf, disordered cortical mt

79 botero1 mutant BOT1 is required for organizing cortical microtubules into transverse arrays in interphase cells, and that this organization is required for consolidating, rather than initiating, changes in the direction of cell expansion.
 - defekt migrace neuronů LIS1 váže +konce MT, interakce s dyneinem ale rostliny nemají")
80 TON1 (tonneau) defekt PPB a organizace MT homolog LISSENCEPHALY1 (H.s. LIS1) - defekt migrace neuronů LIS1 váže +konce MT, interakce s dyneinem ale rostliny nemají dynein wt ton1 (Traas et al. 1995)

")
81 MAP65: crosslinking ( rostlinná specialita ) Arabidopsis má 9 homologů MAP65


82 pleiade mutace v MAP65 (Muller et al. 2002) mnohojaderné buňky (kořenově specifické!)
83
")
84 MOR1 (microtubule organization) (MAP215)

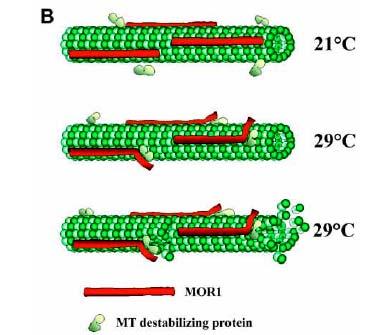

85 Modely funkce MOR1



86 MOR1 Wasteneys 2003/2004: mikrofibrily se uspořádají i samy 2-6-dichlorobenzonitril

87 Asociace mezi kortikálními mt a stěnou onion root

88 Asociace mezi kortikálními MT a stěnou
89 Interakce MT a buněčné stěny Hechtovy provazce cibule

90

")
91 Chara: jak se to měřilo (aktin)


92 ZWICHEL kóduje kinesin! (jeden z mnoha!)

93

")
94 Mutant chup1 (chloroplast unusual positioning) CHUP1: actin binding!

95


96 Regulace struktury a funkce Jako všude: proteinkinázy a fosfatázy cytoskeletu kořen A.th. po staurosporinu (mikrotubuly)

97 Aktin: malé GTPázy (Rop, Arf)

98 Příklad: diferenciace epidermálních buněk (Panteris a Galatis 2005)
 BRICK (Z. mays) SPIKE1 (multidomén. adaptor.")
99 Regulační aspekty: ví se hlavně o aktinu distorted mutace (ARP2/3) BRICK (Z. mays) SPIKE1 (multidomén. adaptor. protein) RICs/ROPs

100 mikrotubuly celulóz. mikrofibrily aktin (Panteris a Galatis 2005)
101 Regulace tvaru epidermálních buněk: RIC/ROP systém Rop Interacting CRIBs: efektory ROP, kontrolující aktin i tubulin! RIC1: MT bundling RIC4: actin polymerization ROP2/4: inhibice RIC1 stimulace RIC4 ROP GTPázy = Rho of plants (Rho GTPázy) CRIB doména = Cdc42/Rac interactive binding domain


102 TON2 kóduje protein fosfatázu A wt ton2 wt ton2-14 ton2-13
103 Cytoskelet kvasinek


104 Aktinový cytoskelet během buněčného cyklu actin patches actin ring actin DNA Saccharomyces cerevisiae Schizosaccharomyces pombe actin

105 Mitotické vřeténko během buněčného cyklu




106 Tubulinový cytoskelet během buněčného cyklu S. cerevisiae TUB3-GFP

107 Spindle pole body (SPB) strukturní proteiny Spc72 Spc72 Cnm67 Spc42 jaderná membrána Spc110 Spc97 Spc98 Tub4 γ-tubulinový komplex cmt vnější plak 1. intermediární vrstva 2. intermediární vrstva centrální plak HB Cmd1 Kar1 cytoplasma regulační proteiny Mps1 satelit vnitřní plak Cdc31 pmt jádro Mps2 Ndc1
108 SPB během buněčného cyklu M G1 G2 S START

109 SPB během buněčného cyklu
110 Molekulové motory
111 Molekulové motory Po aktin. filamentech: až 100 µm/s (Chara), typicky jednotky µm/s proudění cytoplasmy, pohyb organel typický motor - myosin Po mikrotubulech: cca 150 nm/s (řádově pomalejší) pohyby chromosomů a mitotického vřeténka, membr. váčků, organizace mikrotubulů... kinesiny (typicky k +konci) a dyneiny (k -konci)

112 Pohyby organel: plastidy jdou za světlem Mougeotia
113 MOLEKULOVÉ MOTORY DEFINICE Proteiny, které jsou schopné posouvat se na úkor energie ATP po cytoskeletálních vláknech vždy jedním určitým směrem. Hydrolýza ATP vede k cyklickým konformačním změnám jejich molekul, což umožňuje jejich pohyb axonemální dynein (Gibbons) kinezin (Brady, Vale, Scholey) cytoplazmatický dynein (Lye, Scholey, McIntosh)
114 FUNKCE MOLEKULOVÝCH MOTORŮ transport váčků (endocytóza, exocytóza, přenos signálů, atd.) transport molekul nebo komplexů (mrna) pohyb organel proudění cytoplazmy segregace chromosomů v mitóze a meióze splývání jader (karyogamie) buněčná polarita, morfogeneze vliv na dynamiku mikrotubulů komunikace svalový stah



115 Nadrodiny molekulových motorů C-konec myoziny dyneiny kineziny ocas tělo N-konec krček motorová doména mikrofilamenta IF mikrotubuly
116 Nadrodiny molekulových motorů S. cer. A. th. počet tříd myoziny kineziny nezař. dyneiny 1 0 funkční redundance
117 Kineziny Arabidopsis má 61 kinezinů pohyb po mikrotubulech oběma směry (zajišťován různými proteiny), směr určuje krček většina (živočišné a 40 u A.th.) + end-directed často role v cytokinezi (PAKPs phragmoplast-associated kinesin proteins) zbytek (21) end-directed ATK1 funkce vřeténka ZWI regulace calmodulinem KCH calponin homology domain
118 náklad Kineziny a dyneiny kinezin + náklad dynein - β α (+) konec (-) konec

119 mechanismus pohybu kinezinu

120 mechanismus pohybu kinezinu

121 mechanismus pohybu kinezinu

122 mechanismus pohybu kinezinu

123 mechanismus pohybu kinezinu ANIMACE 1
124 Myoziny myozin I náklad + aktinové filamentum (+) konec (-) konec 70 nm

125 mechanismus pohybu myozinu

126 mechanismus pohybu myozinu
127 mechanismus pohybu myozinu

128 mechanismus pohybu myozinu

129 mechanismus pohybu myozinu ANIMACE 2

130 funkce myozinů

131 M y o z i n y některé třídy specifické jen pro určité skupiny organismů Arabidopsis má 17 myosinů (class VIII, XI, XIII)
132 Regulace molekulových motorů regulace přes Ca 2+ a calmodulin IQ motivy myozinů KCBP kinezin injekce Ca 2+ urychluje pohyb chromosomů fosforylace
133 Interakce motorů operujících na mikrotubulech a mikrofinamentech interakce obou skupin motorů koordinace buněčného transportu (MT long-distance, MF short-distance) SMY1 suprimuje mutaci v MYO2 (fyzická interakce jejich ocasových domén) S. cer. KCBP je z části kinezin, z části myozin - A. th. přímá interakce MyoVa s KhcU - savci
134 Cytoplasmic streaming Pohyb organel, váčků a různého materiálu podél aktinového cytoskeletu (~ 70 µm/s), které provádějí motorové proteiny operující na aktinových filamentech. kolokalizace mikrofilament a proudů cytoplasmy depolymerizace mikrofilament inhibuje streaming depolymerizace mikrotubulů neinhibuje streaming kosedimentace myozinu s váčky imunolokalizace myozinů: - myozin I kolem jádra, mitochondrií, chloroplastů - myozin II kolem mitochondrií, chloroplastů - myozin V kolem malých organel (váčky)

135 Buněčné dělení kortikální mikrotubuly preprofázový prstenec vytváření dělicího vřeténka segregace chromosomů tvorba plazmatické destičky ANIMACE 3
136 segregace chromosomů cytoplazma S. cerevisiae jádro + dynein _ MTOC + Kar3 Kar3 + + Cin8 Kar3 kmt pmt _ + MTOC JM cmt (Kip1) aktinové vlákno Kar3 Cin8 Kip1 dynein
137 segregace chromosomů cytoplazma S. cerevisiae jádro JM + dynein _ + _ MTOC Kar3 Cin8 pmt + MTOC cmt (Kip1) aktinové vlákno Kar3 Cin8 Kip1 dynein
138 segregace chromosomů S. cerevisiae cytoplazma jádro JM dynein _ + _ MTOC Kar3 Cin8 pmt + MTOC cmt (Kip1) aktinové vlákno Kar3 Cin8 Kip1 dynein
139 segregace chromosomů rostliny motorové proteiny operující na mikrotubulech zvýšená hladina exprese kinezinů během mitózy (KatB/C, KCBP, TKRP125) lokalizace některých kinezinů podél mikrotubulů vřeténka (N- i C-koncové) injekce KCBP-protilátky zablokuje mitózu v metafázi
140 tvorba plazmatické destičky motorové proteiny operující na mikrotubulech asociace váčků s mikrotubuly fragmoplastu depolymerizace mikrotubulů inhibuje tvorbu destičky depolymerizace mikrofilament neinhibuje tvorbu destičky (vede pouze k abnormální pozici destičky) lokalizace některých motorů u fragmoplastu; N-koncové kineziny (TKRP125) i C-koncové (KatA a KCBP) injekce KCBP-protilátky po anafázi blokuje tvorbu destičky


141 buněčná komunikace MyoVIII funguje v plasmodesmech mikrotubuly a mikrofilamenta jsou nezbytné pro regulovaný transport makromolekul přes plazmodesmy pravděpodobná účast molekulových motorů mikrovilus naruby? Baluška et al. 2001


142 náklad tělo motorová doména
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Cytoskelet a molekulární motory: Biologie a patologie. Prof. MUDr. Augustin Svoboda, CSc.
 Cytoskelet a molekulární motory: Biologie a patologie Prof. MUDr. Augustin Svoboda, CSc. Cytosol: tekutá hmota, vyplňující prostor uvnitř buňky mezi organelami. Ve světelném mikroskopu se jeví jako amorfní
Cytoskelet a molekulární motory: Biologie a patologie Prof. MUDr. Augustin Svoboda, CSc. Cytosol: tekutá hmota, vyplňující prostor uvnitř buňky mezi organelami. Ve světelném mikroskopu se jeví jako amorfní
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
VAKUOLA. membránou ohraničený váček membrána se nazývá tonoplast. běžná u rostlin, zvířata specializované funkce či její nepřítomnost
 VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
CYTOPLAZMA. -Prostor mezi organelami, inkluzemi, ribozomy a jádrem
 Cytoskelet CYTOPLAZMA -Prostor mezi organelami, inkluzemi, ribozomy a jádrem -Viskózní až gelový charakter -Prostředí pro mnohé metabolické děje: např. glykolýza translace syntéza sacharosy pentózová cesta
Cytoskelet CYTOPLAZMA -Prostor mezi organelami, inkluzemi, ribozomy a jádrem -Viskózní až gelový charakter -Prostředí pro mnohé metabolické děje: např. glykolýza translace syntéza sacharosy pentózová cesta
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
B9, 2015/2016, I. Literák, V. Oravcová CYTOSKELETÁLNÍ PRINCIP BUŇKY
 B9, 2015/2016, I. Literák, V. Oravcová CYTOSKELETÁLNÍ PRINCIP BUŇKY CYTOSKELETÁLNÍ PRINCIP BUŇKY mikrotubuly střední filamenta aktinová vlákna CYTOSKELETÁLNÍ PRINCIP BUŇKY funkce cytoskeletu - udržovat
B9, 2015/2016, I. Literák, V. Oravcová CYTOSKELETÁLNÍ PRINCIP BUŇKY CYTOSKELETÁLNÍ PRINCIP BUŇKY mikrotubuly střední filamenta aktinová vlákna CYTOSKELETÁLNÍ PRINCIP BUŇKY funkce cytoskeletu - udržovat
1. AKTINOVY CYTOSKELET (mikrofilamenta)
") CYTOSKELET - pohyb bunek, zmeny tvaru bunek - pohyb organel, bunecné procesy (napr. separace chromosomu) - vyzaduje energii (ATP) - CYTOSKELETON = cytoplasmaticky systém vláken - nutný pro bunecný pohyb,
CYTOSKELET - pohyb bunek, zmeny tvaru bunek - pohyb organel, bunecné procesy (napr. separace chromosomu) - vyzaduje energii (ATP) - CYTOSKELETON = cytoplasmaticky systém vláken - nutný pro bunecný pohyb,
BUNĚČNÁ MOTILITA A MOLEKULÁRNÍ MOTORY
 BUNĚČNÁ MOTILITA A MOLEKULÁRNÍ MOTORY 1 VÝZNAM BUNĚČNÉ MOTILITY A MOLEKULÁRNÍCH MOTORŮ V MEDICÍNĚ Příklad: Molekulární motor: dynein Onemocnění: Kartagenerův syndrom 2 BUNĚČNÁ MOTILITA A MOLEKULÁRNÍ MOTORY
BUNĚČNÁ MOTILITA A MOLEKULÁRNÍ MOTORY 1 VÝZNAM BUNĚČNÉ MOTILITY A MOLEKULÁRNÍCH MOTORŮ V MEDICÍNĚ Příklad: Molekulární motor: dynein Onemocnění: Kartagenerův syndrom 2 BUNĚČNÁ MOTILITA A MOLEKULÁRNÍ MOTORY
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
3. Nukleocytoplasmatický kompartment rostlinných buněk
 3. Nukleocytoplasmatický kompartment rostlinných buněk Co je nukleocytoplasmatický kompartment a jak vypadá u typické rostlinné buňky Jádro buněčné Nositel naprosté většiny genetické informace buňky Jak
3. Nukleocytoplasmatický kompartment rostlinných buněk Co je nukleocytoplasmatický kompartment a jak vypadá u typické rostlinné buňky Jádro buněčné Nositel naprosté většiny genetické informace buňky Jak
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Úvod do biologie rostlin Buňka ROSTLINNÁ BUŇKA
 Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
NEMEMBRÁNOVÉ ORGANELY. Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly
 Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly") NEMEMBRÁNOVÉ ORGANELY Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly RIBOSOMY Částice složené z rrna a proteinů, skládají se z velké kulovité
NEMEMBRÁNOVÉ ORGANELY Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly RIBOSOMY Částice složené z rrna a proteinů, skládají se z velké kulovité
MITÓZA V BUŇKÁCH KOŘÍNKU CIBULE
 Cvičení 6: BUNĚČNÝ CYKLUS, MITÓZA Jméno: Skupina: MITÓZA V BUŇKÁCH KOŘÍNKU CIBULE Trvalý preparát: kořínek cibule obarvený v acetorceinu V buňkách kořínku cibule jsou viditelné různé mitotické figury.
Cvičení 6: BUNĚČNÝ CYKLUS, MITÓZA Jméno: Skupina: MITÓZA V BUŇKÁCH KOŘÍNKU CIBULE Trvalý preparát: kořínek cibule obarvený v acetorceinu V buňkách kořínku cibule jsou viditelné různé mitotické figury.
Prohlášení: V Praze, 29. 4. 2011. Podpis
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Martina Benáková MAPs proteiny asociované s mikrotubuly u rostlin MAPs plant microtubule associated proteins Bakalářská práce
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Martina Benáková MAPs proteiny asociované s mikrotubuly u rostlin MAPs plant microtubule associated proteins Bakalářská práce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
Stavba dřeva. Základy cytologie. přednáška
 Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Současná formulace: Buňka je minimální jednotka, která vykazuje všechny znaky živých soustav
 Buněčná teorie: Počátky formování: 1840 a dále, Jan E. Purkyně myšlenka o analogie rostlinného a živočišného těla (buňky zrníčka) Schwann T. Virchow R. nové buňky vznikají pouze dělením buněk již existujících
Buněčná teorie: Počátky formování: 1840 a dále, Jan E. Purkyně myšlenka o analogie rostlinného a živočišného těla (buňky zrníčka) Schwann T. Virchow R. nové buňky vznikají pouze dělením buněk již existujících
MBR ) Architektura buňky. e) Plastidy f) Mitochondrie a peroxizómy g) Cytoskelet
 Architektura buňky. e) Plastidy f) Mitochondrie a peroxizómy g) Cytoskelet") MBR 2015 1) Architektura buňky 1 e) Plastidy f) Mitochondrie a peroxizómy g) Cytoskelet e) Plastidy 2 Vyskytují se v autotrofních eukaryotech. U rostlin se vyskytují téměř ve všech buňkách. Plastidy produkují:
MBR 2015 1) Architektura buňky 1 e) Plastidy f) Mitochondrie a peroxizómy g) Cytoskelet e) Plastidy 2 Vyskytují se v autotrofních eukaryotech. U rostlin se vyskytují téměř ve všech buňkách. Plastidy produkují:
DUM č. 1 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 1 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Charakteristika buněčného cyklu eukaryot
projekt GML Brno Docens DUM č. 1 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Charakteristika buněčného cyklu eukaryot
Endocytóza o regulovaný transport látek v buňce
 . Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
. Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Interakce buněk s mezibuněčnou hmotou. B. Dvořánková
 Interakce buněk s mezibuněčnou hmotou B. Dvořánková Obsah přednášky Buňka a její organely Extracelulární matrix Interakce buněk s ECM i navzájem Kultivace buněk in vitro Buněčné jádro Alberts: Molecular
Interakce buněk s mezibuněčnou hmotou B. Dvořánková Obsah přednášky Buňka a její organely Extracelulární matrix Interakce buněk s ECM i navzájem Kultivace buněk in vitro Buněčné jádro Alberts: Molecular
základem veškerého aktivního pohybu v živočišné říši je interakce proteinových vláken CYTOSKELETU
 Lukáš Hlaváček, Katedra zoologie Přf UP Olomouc, 2010 POHYB je jeden ze základních životních projevů pro život je nezbytný POHYB na všech úrovních: subcelulární (pohyb v rámci buňky) celulární (pohyb buňky)
Lukáš Hlaváček, Katedra zoologie Přf UP Olomouc, 2010 POHYB je jeden ze základních životních projevů pro život je nezbytný POHYB na všech úrovních: subcelulární (pohyb v rámci buňky) celulární (pohyb buňky)
základem veškerého aktivního pohybu v živočišnéříši je interakce proteinových vláken CYTOSKELETU
 POHYB je jeden ze základních životních projevů pro život je nezbytný POHYB na všech úrovních: subcelulární (pohyb v rámci buňky) celulární (pohyb buňky) orgánový pohyb (pohyb orgánu) organizmální pohyb
POHYB je jeden ze základních životních projevů pro život je nezbytný POHYB na všech úrovních: subcelulární (pohyb v rámci buňky) celulární (pohyb buňky) orgánový pohyb (pohyb orgánu) organizmální pohyb
Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Úlohy: Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Úlohy: Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
Nejmenší jednotka živého organismu schopná samostatné existence. Výměnu látek Růst Pohyb Rozmnožování Dědičnost
 BUŇKA Nejmenší jednotka živého organismu schopná samostatné existence Buňka je schopna uskutečňovat základní funkce organismu: obrázky použity z Nečas: BIOLOGIE LIDSKÉ TĚLO Alberts: ZÁKLADY BUNĚČNÉ BIOLOGIE
BUŇKA Nejmenší jednotka živého organismu schopná samostatné existence Buňka je schopna uskutečňovat základní funkce organismu: obrázky použity z Nečas: BIOLOGIE LIDSKÉ TĚLO Alberts: ZÁKLADY BUNĚČNÉ BIOLOGIE
- pro učitele - na procvičení a upevnění probírané látky - prezentace
 Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
Rozmnožování buněk Vertikální přenos GI. KBI / GENE Mgr. Zbyněk Houdek
 Rozmnožování buněk Vertikální přenos GI KBI / GENE Mgr. Zbyněk Houdek Buněčný cyklus Buňky vznikají z bb. a jedinou možnou cestou, jak vytvořit více bb. je jejich dělením. Vertikální přenos GI: B. (mateřská)
Rozmnožování buněk Vertikální přenos GI KBI / GENE Mgr. Zbyněk Houdek Buněčný cyklus Buňky vznikají z bb. a jedinou možnou cestou, jak vytvořit více bb. je jejich dělením. Vertikální přenos GI: B. (mateřská)
Mitóza a buněčný cyklus
 Mitóza a buněčný cyklus Něco o chromosomech - Chromosom = 1 molekula DNA + navázané proteiny -V diploidní buňce jsou od každého chromosomu 2 kopie (= homologní chromosomy) - Homologní chromosomy nesou
Mitóza a buněčný cyklus Něco o chromosomech - Chromosom = 1 molekula DNA + navázané proteiny -V diploidní buňce jsou od každého chromosomu 2 kopie (= homologní chromosomy) - Homologní chromosomy nesou
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
- význam: ochranná funkce, dodává buňce tvar. jádro = karyon, je vyplněné karyoplazmou ( polotekutá tekutina )
") Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
Otázka: Buňka a dělení buněk Předmět: Biologie Přidal(a): Štěpán Buňka - cytologie = nauka o buňce - rostlinná a živočišná buňka jsou eukaryotické buňky Stavba rostlinné (eukaryotické) buňky: buněčná stěna
Kosterní svalstvo tlustých a tenkých filament
 Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
Cytologie. Přednáška 2010
 Cytologie Přednáška 2010 Buňka 1.Velikost 6 200 µm, průměrná velikost 20um 2. JÁDRO a CYTOPLAZMA 3. ORGANELY (membránové) 4. CYTOPLAZMATICKÉ INKLUZE 5. CYTOSKELET 6. Funkční systémy eukaryotické buňky:
Cytologie Přednáška 2010 Buňka 1.Velikost 6 200 µm, průměrná velikost 20um 2. JÁDRO a CYTOPLAZMA 3. ORGANELY (membránové) 4. CYTOPLAZMATICKÉ INKLUZE 5. CYTOSKELET 6. Funkční systémy eukaryotické buňky:
Univerzita Karlova v Praze - 1. lékařská fakulta. Buňka. Ústav pro histologii a embryologii
 Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
Univerzita Karlova v Praze - 1. lékařská fakulta Buňka. Stavba a funkce buněčné membrány. Transmembránový transport. Membránové organely, buněčné kompartmenty. Ústav pro histologii a embryologii Doc. MUDr.
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
Základy buněčné biologie
 Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Univerzita Palackého v Olomouci. Bakalářská práce
 Univerzita Palackého v Olomouci Bakalářská práce Olomouc 2015 Veronika Kapustová Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra buněčné biologie a genetiky Fenotypová charakterizace nadexpresní
Univerzita Palackého v Olomouci Bakalářská práce Olomouc 2015 Veronika Kapustová Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra buněčné biologie a genetiky Fenotypová charakterizace nadexpresní
Pohyb přípravný text kategorie A, B
 ÚSTŘEDNÍ KOMISE BIOLOGICKÉ OLYMPIÁDY BIOLOGICKÁ OLYMPIÁDA 2005/2006 40. ROČNÍK Pohyb přípravný text kategorie A, B Ivan ČEPIČKA Petr L. JEDELSKÝ Magdalena KUBEŠOVÁ Jana LIŠKOVÁ Jan MATĚJŮ Vendula STRÁDALOVÁ
ÚSTŘEDNÍ KOMISE BIOLOGICKÉ OLYMPIÁDY BIOLOGICKÁ OLYMPIÁDA 2005/2006 40. ROČNÍK Pohyb přípravný text kategorie A, B Ivan ČEPIČKA Petr L. JEDELSKÝ Magdalena KUBEŠOVÁ Jana LIŠKOVÁ Jan MATĚJŮ Vendula STRÁDALOVÁ
Morfogeneze rostlinné buňky. Rostlinná cytologie morfogeneze rostlinné buňky, Katedra experimentální biologie rostlin PřF UK
 Morfogeneze rostlinné buňky Morfogeneze = vývoj tvaru, změna tvarových vlastností http://www.biologyreference.com/co-dn/differentiation-in-plants.html Embryogenní (meristematická) buňka Dospělá buňka,
Morfogeneze rostlinné buňky Morfogeneze = vývoj tvaru, změna tvarových vlastností http://www.biologyreference.com/co-dn/differentiation-in-plants.html Embryogenní (meristematická) buňka Dospělá buňka,
BUNĚČNÝ CYKLUS. OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky. Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí
 (1 BUNĚČNÝ CYKLUS BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace kontrolní body molekulární brzdy Jednobuněčné
(1 BUNĚČNÝ CYKLUS BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace kontrolní body molekulární brzdy Jednobuněčné
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Přeměna chemické energie v mechanickou
 Přeměna chemické energie v mechanickou Molekulám schopným této energetické přeměny se říká molekulární motory. Nejklasičtějším příkladem je svalový myosin (posouvá se po aktinu), ale patří sem i ATP-syntáza
Přeměna chemické energie v mechanickou Molekulám schopným této energetické přeměny se říká molekulární motory. Nejklasičtějším příkladem je svalový myosin (posouvá se po aktinu), ale patří sem i ATP-syntáza
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Buňky, tkáně, orgány, soustavy
 Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Eukaryotická buňka. Stavba. - hlavní rozdíly:
 Eukaryotická buňka - hlavní rozdíly: rostlinná buňka živočišná buňka buňka hub buněčná stěna ano (celulóza) ne ano (chitin) vakuoly ano ne (prvoci ano) ano lysozomy ne ano ne zásobní látka škrob glykogen
Eukaryotická buňka - hlavní rozdíly: rostlinná buňka živočišná buňka buňka hub buněčná stěna ano (celulóza) ne ano (chitin) vakuoly ano ne (prvoci ano) ano lysozomy ne ano ne zásobní látka škrob glykogen
Univerzita Karlova v Praze, 1. lékařská fakulta
 Univerzita Karlova v Praze, 1. lékařská fakulta Tkáň svalová. Obecná charakteristika hladké a příčně pruhované svaloviny (kosterní a srdeční). Funkční morfologie myofibrily. Mechanismus kontrakce. Stavba
Univerzita Karlova v Praze, 1. lékařská fakulta Tkáň svalová. Obecná charakteristika hladké a příčně pruhované svaloviny (kosterní a srdeční). Funkční morfologie myofibrily. Mechanismus kontrakce. Stavba
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
25.2.2014. Genomika. Obor genetiky, který se snaží. stanovit úplnou genetickou informaci. organismu a interpretovat ji v. termínech životních pochodů.
 Genomika Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat ji v termínech životních pochodů. 1 Strukturní genomika stanovení sledu nukleotidů genomu organismu,
Genomika Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat ji v termínech životních pochodů. 1 Strukturní genomika stanovení sledu nukleotidů genomu organismu,
Laboratoř růstových regulátorů Miroslav Strnad
 Laboratoř růstových regulátorů Miroslav Strnad Fyziologie rostlinné buňky, cytoskelet, buněč ěčné dělení a onkogeneze [kap 1] Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR ER
Laboratoř růstových regulátorů Miroslav Strnad Fyziologie rostlinné buňky, cytoskelet, buněč ěčné dělení a onkogeneze [kap 1] Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR ER
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Univerzita Karlova v Praze Přírodovědecká fakulta. Molekulární biologie a biochemie organismů
 Univerzita Karlova v Praze Přírodovědecká fakulta Molekulární biologie a biochemie organismů Petra Schiebertová Nukleace aktinu v rostlinných buňkách Bakalářská práce Vedoucí bakalářské práce: RNDr. Kateřina
Univerzita Karlova v Praze Přírodovědecká fakulta Molekulární biologie a biochemie organismů Petra Schiebertová Nukleace aktinu v rostlinných buňkách Bakalářská práce Vedoucí bakalářské práce: RNDr. Kateřina
Cytoskelet, buněčné interakce a extracelulární matrix
 Bi8120 Aplikovaná buněčná biologie 7.3.2012 Cytoskelet, buněčné interakce a extracelulární matrix doc. RNDr. Renata Veselská, Ph.D., M.Sc. Ústav experimentální biologie Přírodovědecká fakulta MU Program
Bi8120 Aplikovaná buněčná biologie 7.3.2012 Cytoskelet, buněčné interakce a extracelulární matrix doc. RNDr. Renata Veselská, Ph.D., M.Sc. Ústav experimentální biologie Přírodovědecká fakulta MU Program
DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje.
 Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
Inovace studia molekulární. a buněčné biologie
 Inovace studia molekulární I n v e s t i c e d o r o z v o j e v z d ě l á v á n í a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární I n v e s t i c e d o r o z v o j e v z d ě l á v á n í a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
Buněčné dělení ŘÍZENÍ BUNĚČNÉHO CYKLU
 BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 KBB/ZGEN Základy genetiky Dana Šafářová KBB/ZGEN Základy genetiky Rozsah: 2+1
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 KBB/ZGEN Základy genetiky Dana Šafářová KBB/ZGEN Základy genetiky Rozsah: 2+1
Buněčné jádro a viry
 Buněčné jádro a viry Struktura virionu Obal kapsida strukturni proteiny povrchove glykoproteiny interakce s receptorem na povrchu buňky uvnitř nukleocore (ribo )nukleova kyselina, virove proteiny Lokalizace
Buněčné jádro a viry Struktura virionu Obal kapsida strukturni proteiny povrchove glykoproteiny interakce s receptorem na povrchu buňky uvnitř nukleocore (ribo )nukleova kyselina, virove proteiny Lokalizace
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
ENERGIE BUNĚČNÁ RESPIRACE FOTOSYNTÉZA. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 ENERGIE BUNĚČNÁ RESPIRACE FOTOSYNTÉZA 2013 Doc. MVDr. Eva Bártová, Ph.D. ZÍSKÁVÁNÍ a PŘENOS ENERGIE BUŇKOU 1. termodynamická věta - různé formy energie se mohou navzájem přeměňovat 2. termodynamická věta
ENERGIE BUNĚČNÁ RESPIRACE FOTOSYNTÉZA 2013 Doc. MVDr. Eva Bártová, Ph.D. ZÍSKÁVÁNÍ a PŘENOS ENERGIE BUŇKOU 1. termodynamická věta - různé formy energie se mohou navzájem přeměňovat 2. termodynamická věta
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Cytologie I, stavba buňky
 Cytologie I, stavba buňky Ústav pro histologii a embryologii Předmět: Histologie a embryologie 1, B01131, obor Zubní lékařství Datum přednášky: 1.10.2013 Buňka je základní strukturální a funkční jednotka
Cytologie I, stavba buňky Ústav pro histologii a embryologii Předmět: Histologie a embryologie 1, B01131, obor Zubní lékařství Datum přednášky: 1.10.2013 Buňka je základní strukturální a funkční jednotka
Mitochondrie Buněčné transporty Cytoskelet
 Přípravný kurz z biologie Mitochondrie Buněčné transporty Cytoskelet 5. 11. 2011 Mgr. Kateřina Caltová Mitochondrie Mitochondrie semiautonomní organely vlastní mtdna, vlastní proteosyntetický aparát a
Přípravný kurz z biologie Mitochondrie Buněčné transporty Cytoskelet 5. 11. 2011 Mgr. Kateřina Caltová Mitochondrie Mitochondrie semiautonomní organely vlastní mtdna, vlastní proteosyntetický aparát a
A. chromozómy jsou rozděleny na 2 chromatidy spojené jen v místě centromery. B. vlákna dělícího vřeténka jsou připojena k chromozómům
 Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin
 Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Lokalizace proteinu ARPC2 v rostlinné buňce Bc. Kateřina Šlajcherová Praha, 2013 školitelka: RNDr. Kateřina Schwarzerová,
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Lokalizace proteinu ARPC2 v rostlinné buňce Bc. Kateřina Šlajcherová Praha, 2013 školitelka: RNDr. Kateřina Schwarzerová,
Z Buchanan et al. 2000
 Průběh buněčného cyklu Z Buchanan et al. 2000 Změny v uspořádání mikrotubulů v průběhu buněčného cyklu A interfáze, kortikální mikrotubuly uspořádané v cytoplasmě pod plasmalemou B konec G2 fáze, mikrotubuly
Průběh buněčného cyklu Z Buchanan et al. 2000 Změny v uspořádání mikrotubulů v průběhu buněčného cyklu A interfáze, kortikální mikrotubuly uspořádané v cytoplasmě pod plasmalemou B konec G2 fáze, mikrotubuly
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
Co vás dnes čeká: Přednáška 2: Specifika rostlinné buňky trocha opakování vakuola buněčná stěna plastidy Fotosyntetické struktury
 Co vás dnes čeká: Přednáška 2: Specifika rostlinné buňky trocha opakování vakuola buněčná stěna plastidy Fotosyntetické struktury Sluneční záření - energie Eukaryontní buňky: Rozdíly mezi rostlinnou a
Co vás dnes čeká: Přednáška 2: Specifika rostlinné buňky trocha opakování vakuola buněčná stěna plastidy Fotosyntetické struktury Sluneční záření - energie Eukaryontní buňky: Rozdíly mezi rostlinnou a
Pohyb buněk a organismů
 Pohyb buněk a organismů Pohybové buněčné procesy: Vnitrobuněčný transpost organel, membránových váčků Pohyb chromozómů při dělení buněk Cytokineze Lokomoce buněk (améboidní a řasinkový pohyb) Svalový pohyb
Pohyb buněk a organismů Pohybové buněčné procesy: Vnitrobuněčný transpost organel, membránových váčků Pohyb chromozómů při dělení buněk Cytokineze Lokomoce buněk (améboidní a řasinkový pohyb) Svalový pohyb
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
8 cyklinů (A, B, C, D, E, F, G a H) - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů
 - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů") Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
BIOLOGIE BUŇKY II Struktura buňky Buněčný cyklus
 BIOLOGIE BUŇKY II Struktura buňky Buněčný cyklus 10.10.2016 Nejjednodušší forma života (viry neschopnost samostatné reprodukce) Základní stavební a funkční jednotka organismů schopná se dělit Spojeno s
BIOLOGIE BUŇKY II Struktura buňky Buněčný cyklus 10.10.2016 Nejjednodušší forma života (viry neschopnost samostatné reprodukce) Základní stavební a funkční jednotka organismů schopná se dělit Spojeno s
Bp1252 Biochemie. #11 Biochemie svalů
 Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
BIOLOGIE BUŇKY. Aplikace nanotechnologií v medicíně zimní semestr 2016/2017. Mgr. Jana Rotková, Ph.D.
 BIOLOGIE BUŇKY Aplikace nanotechnologií v medicíně zimní semestr 2016/2017 Mgr. Jana Rotková, Ph.D. OBSAH zařazení v systému organismů charakterizace buňky buněčné organely specializace buněk užitečné
BIOLOGIE BUŇKY Aplikace nanotechnologií v medicíně zimní semestr 2016/2017 Mgr. Jana Rotková, Ph.D. OBSAH zařazení v systému organismů charakterizace buňky buněčné organely specializace buněk užitečné
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin
 Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Role komplexu ARP2/3 v rostlinné buňce Bc. Petra Schiebertová Praha, 2013 Školitelka: RNDr. Kateřina Schwarzerová,
Univerzita Karlova v Praze Přírodovědecká fakulta Katedra experimentální biologie rostlin Role komplexu ARP2/3 v rostlinné buňce Bc. Petra Schiebertová Praha, 2013 Školitelka: RNDr. Kateřina Schwarzerová,
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Souhrn 4. přednášky. Genetické metody
 Souhrn 4. přednášky Genetické metody Plasmidy (kvasinkové elementy) Integrace (plasmidy, PCR, kazety) Teplotně-sensitivní mutanty (esenciálních genů) Tetrádová analýza Syntetická letalita, epistase, suprese
Souhrn 4. přednášky Genetické metody Plasmidy (kvasinkové elementy) Integrace (plasmidy, PCR, kazety) Teplotně-sensitivní mutanty (esenciálních genů) Tetrádová analýza Syntetická letalita, epistase, suprese
FYZIOLOGIE BUŇKY BUŇKA 5.3.2015. Základní funkce buněk: PROKARYOTICKÁ BUŇKA. Funkce zajišťují základní životní projevy buněk: EUKARYOTICKÁ BUŇKA
 FYZIOLOGIE BUŇKY BUŇKA - nejmenší samostatná morfologická a funkční jednotka živého organismu, schopná nezávislé existence buňky tkáně orgány organismus - fyziologie orgánů a systémů založena na komplexní
FYZIOLOGIE BUŇKY BUŇKA - nejmenší samostatná morfologická a funkční jednotka živého organismu, schopná nezávislé existence buňky tkáně orgány organismus - fyziologie orgánů a systémů založena na komplexní
Biologie 12, 2017/2018, Ivo Papoušek, Ivan Literák BUNĚČNÝ CYKLUS A JEHO REGULACE
 Biologie 12, 2017/2018, Ivo Papoušek, Ivan Literák BUNĚČNÝ CYKLUS A JEHO REGULACE BUNĚČNÝ CYKLUS PROGRAMOVANÁ BUNĚČNÁ SMRT KONTINUITA ŽIVOTA: R. R. Virchow: Virchow: buňka buňka z buňky, z buňky, živočich
Biologie 12, 2017/2018, Ivo Papoušek, Ivan Literák BUNĚČNÝ CYKLUS A JEHO REGULACE BUNĚČNÝ CYKLUS PROGRAMOVANÁ BUNĚČNÁ SMRT KONTINUITA ŽIVOTA: R. R. Virchow: Virchow: buňka buňka z buňky, z buňky, živočich
Mitochondrie. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Mitochondrie Krátká historie objevu mitochondrií Jako granulární struktury pozorovány v buňkách od poloviny 19. století 1886, Richard Altmann: popsal pozorování bioblastů a navrhl hypotézu, že se jedná
Mitochondrie Krátká historie objevu mitochondrií Jako granulární struktury pozorovány v buňkách od poloviny 19. století 1886, Richard Altmann: popsal pozorování bioblastů a navrhl hypotézu, že se jedná