Obecný metabolismus.
|
|
|
- Kristina Marková
- před 7 lety
- Počet zobrazení:
Transkript
1 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/ Obecný metabolismus. Oxidativní fosforylace (9). Prof.RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká fakulta UP Olomouc
2 Oxidativní fosforylace. Stručná charakteristika. Denní energetická potřeba 70 kg muže je asi kj. Tomuto množství odpovídá 83 kg ATP. Běžná zásoba ATP je 250 g. Pro dosažení potřebného množství energie je nutné recyklovat ADP na ATP. Každá molekula ATP je recyklována 300x za den. Potřebná energie se získává oxidací redukovaných koenzymů NADH a FADH 2. Elektrony z redukovaných koenzymů se přenáší přes čtyři velké komplexy proteinů na kyslík. Přenos je lokalizován ve vnitřní mitochondriální membráně a nazývá se dýchací řetězec nebo elektronový transportní řetězec. NADH + ½ O 2 + H + H 2 O + NAD + ; DG o = kj/mol Tři komplexy elektronového transportního řetězce využívají energii přenosu elektronů k transportu protonů z matrix do mezimembránového prostoru. Generuje se tak ph gradient a transmembránový elektrický potenciál, který vytváří protonmotivní sílu.
3 Oxidativní fosforylace. Stručná charakteristika ATP je syntetizován, když protony putují zpět do matrix přes proteinový komplex ATPasy. ADP + P i + H + H 2 O + ATP ; DG o = + 30,5 kj/mol Oxidace energeticky bohatých sloučenin a fosforylace ADP jsou spojeny s protonovým gradientem přes vnitřní mitochondriální membránu. Proces se nazývá respirace nebo buněčná respirace. Respirace je proces tvorby ATP při kterém anorganická látka (jako je molekulární kyslík) slouží jako terminální akceptor elektronů. Donorem elektronů může být organická nebo anorganická látka. Proces se uskutečňuje v mitochondrii.
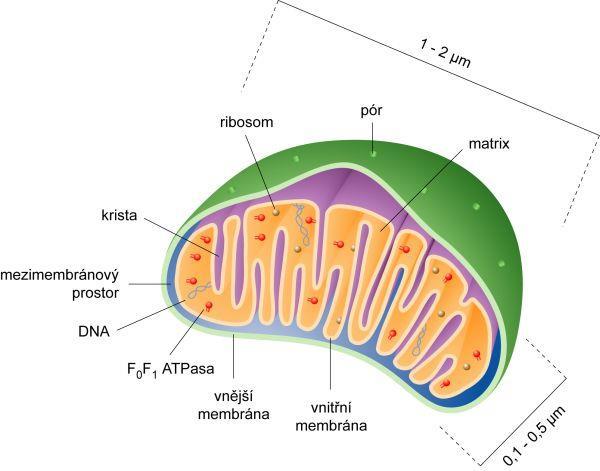
4 Obrázek mitochondrie.
5 Komplexy přenosu elektronů. Elektrony jsou přenášeny z NADH na O 2 přes řetězec tří velkých proteinových komplexů: a) NADH-Q oxidoreduktasa (komplex I) b) Q-cytochrom c oxidoreduktasa (komplex III) c) cytochrom c oxidasa (komplex IV) Čtvrtý velký proteinový komplex sukcinát-q reduktasa obsahuje sukcinádehydrogenasu generující FADH 2 v citrátovém cyklu. Elektrony z FADH 2 vstupují do dýchacího řetězce v Q- cytochromoxidoreduktase. Sukcinát-Qreduktasa nepumpuje protony!! Komplexy I, III a IV jsou svým způsobem na sebe navazující a spojené (supramolekulární komplex) nazývaný respirasom. Elektrony mezi komplexy přenáší speciální přenašeč. Koenzym Q také nazývaný ubichinon, v redukované formě ubihydrochinon.
6 Komplexy přenosu elektronů. Ubichinon je hydrofobní chinon (má řetězec složený z isoprenových jednotek), který rychle difunduje přes vnitřní mitochondriální membránu. Nejznámější koenzym Q (savčí) má 10 isoprenových jednotek (Q 10 ). Ubichinon přenáší elektrony z NADH-Q oxidoreduktasy na Q- cytochrom c oxidoreduktasu ve formě ubihydrochinonu. Elektrony z FADH 2 generované citrátovým cyklem jsou přeneseny na ubichinon a poté na Q-cytochrom c oxidoreduktasový komplex. Chinony obecně mohou existovat ve třech oxidačních stavech: plně oxidovaná forma Q; s jedním elektronem semichinon QH -, po ztrátě protonu vznikne semichinonový radikál Q.- ; plně redukovaná ubichinn je ubichinol QH 2. Druhým přenašečem elektronů je malý rozpustný protein cytochrom c. Přenáší elektrony z komplexu III na komplex IV.
7 Složky dýchacího řetězce. 0 NADH + H + I I Fe 2+ Komplex NADH - Q reduktasa FADH 2-50 Volná energie ve vztahu k O 2 (kj.mol -1 ) e - Ubichinon (Q) e - II e - Fe 2+ III Fe 2+ e - Komplex cytochrom c Sukcinát - dehydrogenasa reduktasa Cytochrom c e - Fe 2+ Fe 2+ IV e - Komplex cytochrom c oxidasa /2 O 2 H 2 O
8 Oxidační stavy ubichinonu (chinonů) O - H 3 C H 3 C O O O. H + CH 3 R Semichinon, iont radikál Q.- H 3 C H 3 C O O CH 3 e - H + + H 3 C H 3 C O OH CH 3 e H + O H O R O R O CH 3 10 O - OH Oxidov aná forma Q Semichinon Redukov aná forma QH 2 (ubichinon) (ubichinol) H 3 C H 3 C O OH CH 3
9 Oxidační stavy flavinů. H 3 C N O NH 2 e H + H 3 C H N O NH H 3 C N CH 2 N O H 3 C N R N H O HO OH Flavinmononukleotid (redukovaný) (FMNH 2 ) OH CH 2 OH Flavinmononukleotid (oxidovaný) (FMN)
10 Komplexy elektronového řetězce. NADH-Qoxiodoreduktasa (komplex I). Obrovský enzym (> 900 kd), asi 46 polypeptidových řetězců. Protonová pumpa. Integrální protein ve vnitřní mitochondriální membráně. Prosthetickou skupinou je FMN redukovaný FMNH 2. Elektrony jsou poté přeneseny na Fe-S klastry. Atom Fe je buď v oxidovaném stavu Fe 3+ nebo v redukovaném Fe 2+. Klastry nepumpují protony. Konečným akceptorem elektronů v komplexi I je ubichinon (Q) přecházející na ubichinol (QH 2 ). Katalyzovaná reakce: NADH + Q + 5 H + matrix NAD + + QH H + cytoplasma
11 Komplex I přenos elektronů spojený s pumpováním protonů do matrix 4 H + Mezimembránov ý prostor Mezimembránov ý prostor Hotov ost Q QH 2 Q QH 2 Q Matrix NADH FMN Matrix NAD + (Fe-S) (Fe-S) (Fe-S) 2 H + e -
12 Komplexy elektronového řetězce. Sukcinát-Qreduktasa (komplex II) je součást vnitřní mitochondriální membrány. Elektrony z FADH 2 jsou přenášeny přes klastry Fe-S na ubichinon. Komplex II nepumpuje protony. Další dva enzymy glycerol-3-fosfátdehydrogenasa a acyl CoAdehydrogenasa (mastné kyseliny) přenáší elektrony z FADH 2 na Q za tvorby QH 2. Oba enzymy také nepumpují protony. Příklad 2Fe-2S klastru. Všechny Fe-S klastry podléhají oxidačněredukční reakci na atomu Fe. Cys S Fe 2- S S Fe 2- Cys Cys S S S Cys
13 Komplexy elektronového řetězce. Q-cytochrom c oxidoreduktasa (komplex III nebo cytochromreduktasa). Funkcí komplexu III je přenos elektronů z QH 2 na oxidovaný cytochrom c (Cyt c). Komplex III je ve vodě rozpustný a pumpuje protony ven z matrix. Pumpuje 2 H +. Je to polovina protonů než pumpuje komplex I (menší termodynamická síla). Rovnice: QH Cyt c ox + 2 H + matrix Q + 2 Cyt c red + 4 H + cytoplasma Komplex III obsahuje dva typy cytochromů b a c 1. Cytochromy jsou proteiny obsahující hem jako prosthetickou skupinu. Přenášejí elektrony. Stejný hem jako obsahují myoglobin a hemoglobin. Enzym také obsahuje protein železo síra s 2Fe-2S centrem Rieskeho centrum.
14 Mechanismus komplexu III Q cyklus. Prv ní část Q cyklu Cyt c 2 H + Druhá část Q cyklu Cyt c 2 H + Cyt c 1 Cyt c 1 QH 2 Q QH 2 Q Hotov ost Q Q Q.- Q o Q i Hotov ost Q Q.- Q o QH 2 Hotov ost Q Q i
15 Q cyklus. QH 2 transportuje dva elektrony na komplex III cytochrom c, který akceptuje pouze jeden elektron. Mechanismus spojení přenosu jednoho elektronu s transportem protonů z matrix do mezimembránového prostoru se nazývá Q cyklus. Dvě molekuly QH 2 se váží postupně na komplex III na který předávají dva elektrony a dva H +. Tyto protony se uvolňují do cytoplasmy. První molekula QH 2 se váže na první vazebné místo Q o a dva elektrony putují v komplexu na dvě různá místa. Jeden elektron putuje nejdříve na Rieskeho 2Fe-2S klastr a poté na cytochrom c 1 a konečně na oxidovaný cytochrom c, který redukuje. Redukovaná molekula cytochromu c difunduje dále na cytochrom c oxidasu. Druhý elektron putuje přes dva hemy cytochromu b na druhé vazebné místo Q i.
16 Q cyklus. V tomto místě je Q redukuván na semichinon radikál aniont (Q.- ) elektronem z první molekuly QH 2. Nově plně oxidovaný Q opouští první Q místo a obnovuje pool Q. Druhá molekula QH 2 se váže na Q o vazebné místo a reaguje analogicky jako první molekula. Jeden elektron je přenesen na cytochrom c. Druhý elektron putuje přes dvě hemové skupiny na cytochrom b k částečně redukovanému ubichinonu vázanému na Q i místě. Po vstupu elektronu z druhé QH 2 molekuly převezme tento chinonový radikál dva protony z matrix za tvorby QH 2. Sumárně: 2 QH 2 + Q + 2 Cyt c + 2 H + matrix 2 Q + QH Cyt c reduk H + cytoplasma V jednom Q cyklu jsou oxidovány dvě molekuly QH 2 na Q a jedna je redukována na QH 2
17 Cytochrom c oxidasa. Cytochrom c oxidasa (komplex IV) katalyzuje transfer elektronů z redukovaného cytochromu c na na molekulární kyslík (terminální akceptor elektronů). 4 Cyt c reduk + 8 H + matrix + O 2 4 Cyt c ox + 2 H 2 O + 4 H + cytoplasma Čtyři elektrony jsou přeneseny na kyslík a současně jsou pumpovány protony z matrix do cytoplasmy. Reakce je termodynamicky výhodná. Nejvíce je prostudována hovězí cytochrom c oxidasa, která se skládá z 13 podjednotek z nichž 3 jsou kódovány mitochondriální DNA. Enzym obsahuje 2 skupiny hemu A a tři měďnaté ionty seskupené do dvou center. Jedno centrum označené Cu A /Cu A obshuje dva Cu ionty vázané cysteinovými vedlejšími skupinami. Centrum přijímá první elektrony z Cyt c.
18 Cytochrom c oxidasa. Zbývající Cu (Cu B ) je koordinačně vázán na tři His z nichž jeden je kovalentně vázán na Tyr. Měďnatá centra oscilují mezi oxidačními stavy Cu 2+ a Cu +. Další součástí enzymu jsou dvě molekuly hemu A označované jako hem a a hem a 3. Hemy se liší redukčním potenciálem, protože jsou lokalizovány v různých místech molekuly enzymu s různým okolím. Elektron putuje z Cyt c na Cu A /Cu A, na hem a, na hem a 3, na Cu B a na kyslík!! Hem a 3 a Cu B jsou přímo propojeny s aktivním místem kde dochází k redukci kyslíku.
19 Cytochrom c oxidasa. Elektrony ze dvou molekul redukovaného Cyt c putují cytochrom c oxidasou jeden se zachytí na Cu B a druhý na a 3. Obě redukovaná místa předávají elektrony na kyslík. Když se molekulární kyslík naváže, přebírá elektrony z obou míst za tvorby peroxidu (O 2 2- ), který tvoří můstek mezi oběma místy. Váží se další dvě molekuly Cyt c a elektrony putují do aktivního místa. Přidáním elektronu a H + na každý atom kyslíku dochází k redukci dvou iont kyslíkových skupin na Cu B OH a Fe 3+ - OH. Reakcí se dvěma dalšími H + se uvolní dvě molekuly H 2 O a enzym se vrací do původního stavu. 4 Cyt c reduk + 4 H + matrix + O 2 4 Cyt c ox + 2 H 2 O Všechny čtyři protony v této reakci mají původ v matrix!!! Celková rovnice reakce katalyzované cytochrom c oxidasou: 4 Cyt c reduk + 8 H + matrix + O 2 4 Cyt c ox + 2 H 2 O + 4 Cyt c cytoplasma
20 Toxické deriváty molekulárního kyslíku superoxidový radikál, peroxidy. Molekulární kyslík je ideálním akceptorem elektronů. Pokud probíhá čtyřelektronová redukce vzniká bezpečná voda. Částečná redukce vede ke tvorbě nebezpečných produktů. Transfer jednoho elektronu na O 2 vzniká superoxidový aniont!! Transfer dvou elektronů vede ke tvorbě peroxidu. O 2 + e - O 2.- ; O e - O 2 2- Obě sloučeniny jsou nebezpečné! Strategií bezpečné katalýzy cytochrom c oxidasy je neuvolňovat meziprodukty. Vždy se však nějaké množství superoxidových aniontů a peroxidů uvolní. Z peroxidů a superoxidů se můžou formovat OH., což jsou reaktivní deriváty kyslíku (ROS = reactive oxygen species). Jejich oxidační destruktivní účinky se projevují při stárnutí a vyvolání řady onemocnění.
21 Toxické deriváty molekulárního kyslíku superoxidový radikál, peroxidy. Buněčnou obranou před působením superoxidových radikálů je enzym superoxiddismutasa: O H + O 2 + H 2 O 2 Dismutace je reakce při které je jeden reaktant převeden na dva různé produkty. Peroxid vodíku vytvořený v superoxiddismutasové reakci je rozkládám všudypřítomným hemovým proteinem katalasou (také se jedná o dismutaci): 2 H 2 O 2 O H 2 O Oba enzymy mají vysokou k cat blížící se difůzi. Při likvidaci peroxidu vodíku hraje také významnou roli glutathionperoxidasa. Významně se uplatňují antioxidanty jako např. vitaminy E a C.
22 Mechanismus cytochrom c oxidasy. Označeny jsou chemické protony (tvorba vody), další 4 H + jsou pumpovány z matrix do cytoplasmy. 2 Cyt c Cu A / Cu A O 2 Hem a Fe 2+ Cu + Hem a 3 Cu B Fe 2+ Cu + Fe 2+ Cu + O - O 2 H 2 O 2 Cyt c Cu A / Cu A Fe 2+ Cu + Hem a Fe + HO OH Cu Cu B Hem a 3 2 H + 2 H +
23 Elektronový transportní řetězec. MEZIMEMBRÁNOVÝ PROSTOR H + H + H + I I III IV Q QH 2 HOTOVOST Q M A T R I X I I O 2 H 2 O FADH 2 NADH CITRÁTOVÝ CYKLUS ACETYL CoA
24 Protonový gradient pohání syntézu ATP. Transfer elektronů z NADH na O 2 je silně exergonní proces: NADH + 1/2O 2 + H + H 2 O + NAD + ; DG o = - 220, 1 kj.mol -1 Proces je spojen s endergonickou syntézou ATP: ADP + P i + H + ATP + H 2 O ; DG o = + 30, 5 kj/mol -1 Ve vnitřní mitochondriální membráně je lokalizován integrální protein původně nazývaný mitochondriální ATPasa nebo F 1 F o ATPasa (F o čti ef ó, na oligomycin citlivá podjednotka). Původně enzym katalyzoval hydrolýzu ATP. Současný název ATPsynthasa. Další název je komplex V. Dlouho badatelé hledali spojení mezi transportem elektronů a syntézou ATP, analogii substrátové fosforylace. V roce 1961 přišel P. Mitchell s chemiosmotickou hypotézou tvorby ATP.
25 Chemiosmotická hypotéza tvorby ATP. Podstata hypotézy: Transport protonů a syntéza ATP jsou spojeny protonovým gradientem vytvořeným přes mitochondriální membránu. Koncentrace H + je nízká v matrix, elektrické pole v matrix nese negativní náboj. Protony putují zpět do matrix, aby došlo k vyrovnání náboje. Vnitřní mitochondriální membrána protony nepropouští. Protony vstupují do integrálního proteinu ATPsynthasa. Energeticky nerovnovážné rozdělení protonů se nazývá protonmotivní síla. Protonmotivní síla má dvě složky: chemický gradient a gradient náboje. Chemický gradient reprezentuje gradient ph. Gradient náboje tvořen nerovnovážnou distribucí protonů tvořící chemický gradient. Obě komponenty pohání syntézu ATP.
26 Chemiosmotická hypotéza tvorby ATP. V mezimembránovém prostoru je ph o 1,4 jednotky nižší než v matrix. Membránový potenciál je 0, 14 V, vně je kladný. Membránový potenciál odpovídá volné energii 21, 8 kj.mol -1 na mol protonů. K důkazu chemiosmotické hypotézy byl použit uměle vytvořený systém. Jako respirační řetězec byl použit bakteriorhodopsin, což je membránový protein z halobakterie, který po osvětlení pumpuje protony. Halobakterie, správně Haloarchaea, je třída archeí z kmene Euryarchaeota nejedná se tedy o bakterie (existují v solných roztocích Mrtvé moře). Byly vytvořeny syntetické váčky obsahující bakteriorhdopsin a mitochondriální ATPasu získanou z hovězího srdce. Po osvětlení váčky syntetizovaly ATP. ATPasa je v liposomu zařazena opačně než v mitochondrii.
27 Důkaz platnosti chemiosmotické hypotézy. Bakteriorhodopsin H + H + ATPsynthasa ADP + Pi ATP
28 ATPsynthasa. Tvarem ATPsynthasa připomíná kuličku a tyčinku. Část připomínající tyčinku se nazývá F o podjednotka a je integrální součástí vnitřní mitochondriální membrány. Kulička o rozměrech 85 Å nazvaná F 1 podjednotka je umístěna v matrix mitochondrie. Podjednotka F 1 má katalytickou aktivitu synthasy, izolovaná vykazuje ATPasovou aktivitu. Podjednotka F 1 je složena z pěti typů polypeptidových řetězců (a 3, b 3, g, d, e) s vyznačenou stechiometrií. Podjednotky a a b, které tvoří kuličku F 1 jsou uspořádány střídavě do hexamerního kruhu. Obě váží nukleotidy, ale jen b podjednotka působí přímo katalyticky. Pod kuličkou jsou proteiny středové tyčinky složené z g a e podjednotek. Podjednotka g tvoří helikální vlákno zasahující do středu a 3 b 3 hexameru a narušuje symetrii a 3 b 3 hexameru: každá z podjednotek b se nachází v interakci s jinou stranou (částí) g.

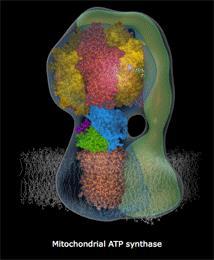
29 Model ATPsynthasy.
30 ATPsynthasa. Podjednotka F o je hydrofobní segment lokalizovaný ve vnitřní mitochondriální membráně. F o obsahuje protonový kanál, který je sestavený z 10 až 14 podjednotek c, které jsou umístěny v membráně. Jedna podjednotka a je na povrchu kanálku. Obě podjednotky F 1 a F o jsou spojeny dvěma způsoby: středovou ag tyčkou (vláknem) a vnější spojkou tvořenou jednou a podjednotkou, dvěma b podjednotkami a d podjednotkou.
31 Tok protonů přes ATPsynthasu vede ke tvorbě a uvolnění ATP. ATPsynthasa katalyzuje tvorbu ATP z ADP a orthofosfátu: ADP 3- + HPO H + ATP 4- + H 2 O Oba substráty mají na fosfáty navázán Mg 2+. Atom kyslíku ADP atakuje atom P orthofosfátu za tvorby pentakovalentního meziproduktu, který uvolňuje vodu a přechází na ATP. 3 - O O - O - O O O O - O O - O - H H O R P P + O - P OH + H + R P P P O - - O O O O O - O O O - ADP P i Pentakov alentní meziprodukt O O - O O - R P P O O O ATP O - P O + H 2 O O -
32 Tok protonů přes ATPsynthasu vede ke tvorbě a uvolnění ATP. Ukázalo se ATPsynthasa katalyzuje spojení ADP a orthofosfátu i bez toku protonů. ATP však se z katalytického místa neuvolňuje!! Pro uvolnění ATP je nutný tok protonů přes podjednotku F o. Rolí protonového gradientu není tvorba ATP, ale jeho uvolnění z katalytického místa. Protonmotivní síla vytváří tři aktivní místa, která se sekvenčně proměňují a mění svoji funkci. ATPsynthasa je složena z pohyblivé části a stacionární části. Pohyblivá část reprezentuje rotor složený z c prstence a ge stonku. Stacionární část neboli stator je složen ze zbytku molekuly. Tvorbu a uvolnění ATP charakterizuje potvrzená hypotéza: binding-change mechanism mechanismus vazebné změny (konformace).
33 Tok protonů přes ATPsynthasu vede ke tvorbě a uvolnění ATP. Průtokem protonů se otáčí rotor měnící konformaci statorové části ve třech stupních: 1) ATP a P i se váží, 2) Syntéza ATP, 3) Uvolnění ATP. Pohybem g rotoru nejsou tři b podjednotky ekvivalentní. V každém momentu je jedna b podjednotka ve stavu L (loose = otevřená). Tato konformace váže ADP a P i. Druhá podjednotka je ve stavu T (tight = pevně sevřený). Dochází k tvorbě ATP. Obě tyto konformace neuvolňují ATP ani ADP nebo P i. Poslední podjednotka je ve stavu O (open = otevřený), uvolňuje ATP. Rotor g rotuje ve směru hodinových ručiček po 120 o.
, b podjednotky jsou")
34 Model F 1 ATPsynthasy. Trojúhelník naznačuje g podjednotku (rotor), b podjednotky jsou naznačeny ve stavech L (zelená), T (lila) a O (žlutá).
 a orthofosfát (menší).")
35 Pohled shora na F 1 ATPsynthasu. Nahoře výpočet atomové hustoty. Dole modré kroužky naznačují ADP (větší) a orthofosfát (menší).

36 F 1 ATPsynthasa struktura z polypeptidových řetězců. Vpravo dole model s označením podjednotek.
37 Z historie bádání o ATPsynthase Herman KALCKAR objevil souvislost mezi syntézou ATP a respirací Ephraim RACKER izoloval části ATP synthasy Peter MITCHELL publikoval chemiosmotickou hypotézu tvorby ATP Paul D. BOYER postuloval hypotézu syntézy ATP na základě konformačních změn enzymu Paul D. BOYER Binding Change Mechanisms. Klíčovým krokem je mechanické uvolnění syntetizovaného ATP John E. WALKER sekvenoval gen pro syntézu ATPsynthasy John E. WALKER a spol. publikoval strukturu ATPsynthasy publikovány tři různé postupy modelování rotace spojené se syntézou ATP. Mikroskopicky Masasuke YOSHIDA.

38 Nobelova cena za chemii 1978 Peter MITCHELL. The Nobel Prize in Chemistry 1978 was awarded to Peter Mitchell "for his contribution to the understanding of biological energy transfer through the formulation of the chemiosmotic theory".



39 Nobelova cena za objev v chemii The Nobel Prize in Chemistry 1997 was divided, one half jointly to Paul D. Boyer and John E. Walker "for their elucidation of the enzymatic mechanism underlying the synthesis of adenosine triphosphate (ATP)" and the other half to Jens C. Skou "for the first discovery of an ion-transporting enzyme, Na+, K+ -ATPase"

40 3 D zobrazení prstence c podjednotek jako hlavní součásti F o ATPsynthasy
41
42 Schematický nákres F o F 1 ATPsynthasy. H + F o F o Vnitřní mitochondriální membrána stopka H + Kotv a (b + d) F 1 - ATPsynthasa M A T R I X a 3 b 3 a 3 b 3 d M A T R I X
43 Pohyblivý model podjednotky F o tři segmenty. Tvorba ATP z ADP a P i.
44 Tok protonů přes c kanálky roztáčí rotor (g). Mechanismus toku protonů je spojen se strukturou podjednotek a a c F o podjednotky ATPsynthasy. Stacionární podjednotka a prochází membránou a je složena z 10 až 14 podjednotek c. Struktura a obsahuje dva hydrofilní polokanálky, které neprochází celou membránou. Protony vstupující do kanálku neprojdou přes membránu. Podjednotka a je konstruována tak, že každý polokanálek přímo souvisí s jednou podjednotkou c. Kanálek je tvořen helikální polypeptidovou strukturou obsahující uprostřed řetězce Asp (61). Proton putující kanálkem z cytoplasmatické strany se naváže na Asp. Podjednotka c s navázaným protonem se pootočí na místo s nízkou koncentrací protonů a tam se proton uvolní.
45 Tok protonů přes c kanálky roztáčí rotor (g). Pohyb protonů půlkanálkem z míst s vysokou koncentrací protonů (cytoplasma) do míst s nízkou koncentrací protonů (matrix) pohání rotaci podjednotky c. Podjednotka c s protonizovaným Asp se uvolní z kontaktu s cytoplasmatickým půlkanálkem a další c podjednotka ji nahradí. Když se c podjednotka pohybuje, podjednotka a je nepohyblivá (stacionární). Každý proton, který vstoupí do půlkanálku a podjednotky na straně cytoplasmy se pomocí c podjednotky dostane do matrix.
46 Tok protonů přes c kanálky roztáčí rotor (g). Počet c podjednotek v c kruhu se pohybuje mezi 10 až 14. Každá otáčka g (rotoru) o 360 o vede k syntéze tří molekul ATP. Uvažujme 10 c podjednotek, potom každá vytvořená molekula ATP spotřebuje 10 / 3 = 3,33 protonů. Pro zjednodušení uvažujme 3 protony z matrix pro každý ATP!! Uvažujme, že odpočívající člověk spotřebuje denně 85 kg ATP, potom musí projít ATPsynthasou 3,3 x protonů.

47 Podjednotka c ATPsynthasy (F o ).

48 Model F 1 F o ATPsynthasy.

49

50 Složky protonvodivé jednotky ATP synthasy. Cytoplasmov ý půlkanálek Matrixov ý půlkanálek Podj ednotka a
51 Vstup elektronů z cytoplasmy do dýchacího řetězce. Jednou z funkcí dýchacího řetězce je regenerace NAD + pro využití v glykolýze. NADH neprochází vnitřní mitochondriální membránou, protože není propustná pro NAD + a NADH. Jednou z cest elektronů z cytoplasmy do dýchacího řetězce je glycerol-3-fosfátový člunek. Elektrony jsou přeneseny na QH 2 (ubichinol). Ubichinol předává elektrony na komplex III transportního řetězce a proto se vytvoří jen 2 ATP!! Glycerol-3-fosfátový člunek je typické pro svalové buňky. Srdeční sval a játra přenáší elektrony (NADPH) z cytoplasmy do mitochondrie malát-aspartátovým člunkem.
52 Glycerol-3-fosfátový člunek (kosterní svaly). NAD H + + H + NAD + O CH 2 OH CH 2 OPO 3 2- Cytoplasmová glycerol fosfát dehydrogenasa HO CH 2 OH H 2- CH 2 OPO 3 Glyceronfosfát Glycerol fosfát CYTOPLASMA E - FAD H 2 E - FAD Mitochondriální glycerol fosfát dehydrogenasa Q QH 2 MATRIX
53 Malát-aspartátový člunek. Malát-aspartátový člunek je zprostředkován dvěma membránovými transportéry a čtyřmi enzymy. Elektrony z NADH jsou přeneseny v cytoplasmě na oxaloacetát za tvorby L-malátu, který je přenesen přes vnitřní mitochondriální membránu jako výměna za 2-oxoglutarát. Malát je reoxidován NAD + v matrix za tvorby NADH. Reakci katalyzuje enzym citrátového cyklu malátdehydrogenasa. Vzniklý oxaloacetát neprochází membránou, je podroben transaminaci za tvorby aspartátu, který je transportován do cytoplasmy jako výměna s glutamátem. Glutamát předává aminoskupinu na oxaloacetát za tvorby aspartátu a 2-oxoglutarátu. Aspartát je v cytoplasmě deaminován za tvorby oxaloacetátu a cyklus je restartován.
54 Malát aspartátový člunek (srdce, játra) NAD + NADH oxaloacetát glutamát Malát 2 - oxoglutarát CYTOPLASMA aspartát Malát MATRIX 2 - oxoglutarát aspartát NAD + NADH oxaloacetát glutamát
55 Vstup ADP do mitochodrie ATP-ADP translokasa. ATP ani ADP neprochází volně vnitřní mitochondriální membránou. Funkci plní specifický transportní protein ATP-ADP translokasa. (antiporter, 30kD). ADP 3- cytoplasma + ATP 4- matrix ADP 3- matrix + ATP 4- cytoplasma ATP-ADP translokasa tvoří zhruba 15% membránových proteinů. Dospělý člověk transportuje denně ekvivalent své váhy ATP!! Translokasa transportuje ADP do matrix a ATP z matrix do cytoplasmy. ATP i ADP se váží na translokasu bez Mg 2+. Transport je energeticky náročný. Spotřebuje se při něm až čtvrtina energie vytvořené respirací (regenerace membrány). Inhibice tohoto procesu zastavuje buněčnou respiraci!!.
56 Mechanismus mitochondriální ATP-ADP translokasy. ADP (cyt) ATP (matrix) ATP (cytoplasma) ADP (matrix)
57 Další mitochondriální transportéry. Fosfátový transportér, který spolupůsobí s ATP-ADP translokasou zprostředkovává elektroneutrální výměnu H 2 PO 4 - za OH -. Kombinace působení těchto dvou transportérů vede k výměně cytoplasmového ADP a P i za matrixový ATP poháněn vstupem H + do matrix a výstupem OH - do cytoplsmy. Oba tyto transportéry, které doprovází ATP synthasu vytváří velký komplex nazvaný ATP synthasom. Další homologní transportéry : dikarboxylátový transportér přenáší malát, sukcinát a fumarát z matrix jako výměnu za P i. Pyruvát z cytoplasmy vstupuje pyruvátovým transportérem do mitochondriální membrány výměnou za OH -.
58 Regulace buněčné respirace. Kompletní oxidace glukosy poskytuje 30 ATP. Dle publikace: Hinkle, P.C., Kumar, M.A., Resetar, A. a Harris, D.L., Biochemistry 30: 3576, Syntéza molekuly ATP je poháněna tokem tří protonů přes ATP synthasu. Další jeden proton je spotřebován na transport ATP z matrix do cytoplasmy!! Proto NADH = 2, 5 ATP a FADH 2 = 1, 5 ATP. P/O, for ATP delivered to the outside of mitochondria, would be 10/(3 + 1) = 2.5 (NADH) and 6/(3 + 1) = 1.5 (succinate), each considerably lower than then textbook values of 3 and 2. Subsequently, careful reevaluation of the P/O ratios for NADH and succinate (FADH 2 ) indicated that the experimental values are close to 2.5 and 1.5
59 Výtěžek ATP z celkové oxidace glukosy. Glykolýza: 2 ATP Pyruvátdehydrogenasa - 2 NADH = 5 ATP. Citrátový cyklus (přímo): 2 ATP Oxidativní fosforylace: 2 molekuly NADH (reoxidace z glykolýzy) = 3 ATP V případě transportu glycerol-3-fosfátovým člunkem! 2 molekuly FADH 2 = 3 ATP 6 molekul NADH = 15 ATP Součet: 30 ATP. Tradičně v učebnicích 36 ATP.
60 Regulace buněčné respirace. Rychlost oxidativní fosforylace je dána potřebou ATP. Elektronový transport je pevně vázán na fosforylaci. Když není k dispozici ADP, elektronový transport se zastaví. S rostoucí koncentrací ADP, roste rychlost fosforylce. Regulace rychlosti oxidativní fosforylace ADP se nazývá respirační kontrola nebo akceptorová kontrola. Hladina ADP ovlivňuje citrátový cyklus. Při nízké hladině ADP (odpočívající sval) se nespotřebovávají NADH a FADH 2. CC se zpomaluje je málo NAD + a FAD. V opačném případě se CC aktivuje. Elektrony netečou z bohatých molekul (glukosa) na kyslík pokud není potřeba syntetizovat ATP.
61 Regulované odpojení. Některé organismy mají schopnost odpojovat oxidativní fosforylaci od syntézy ATP jako způsob tvorby tepla. Hibernující živočichové mají hnědou adiposní tkáň bohatou na mitochondrie. Barva je tvořena kombinací zelených cytochromů a mnoha mitochondrií. Vnitřní mitochondriální membrána těchto živočichů obsahuje velké množství odpojovacího proteinu (UCP-1, uncoupling protein) jinak thermogeninu (dimer 33 kd podobá se ATP-ADP translokase). UCP-1 generuje teplo zkratem protonového gradientu přes UCP-1 tečou protony zpět do matrix mimo ATP synthasu. UCP-1 je aktivován chladem vyvolanou hydrolýzou triacylglycerolů vzniklé mastné kyseliny aktivují thermogenin. Ženy mají hnědou adiposní tkáň na prsou (hrudník) a krku aktivovanou chladem.
62 Inhibice oxidativní fosforylace. Inhibice elektronového transportního řetězce. Rotenon využívaný jako insekticid, amytal barbiturátové sedativum inhibují elektronový transport v NADH-Q oxidoreduktase tím, že zabraňují vstupu substrátu NADH. Při této inhibici není zasažen tokelektronů z FADH 2 (sukcinátdehydrogenasa) Antimycin A antibiotikum, interferuje s tokem elektronů z cytochromu b H Q-cytochrom c oxidoreduktasy. Kyanid, azid (N 3- ) a oxid uhelnatý blokují elektronový tok v cytochrom c oxidase. Kyanid a azid reagují z Fe 3+ hemu a 3. Inhibice elektronového toku inhibuje také ATP syntézu. Inhibice ATP synthasy. Oligomycin, antibiotikum a dicyklohexylkarbodiamid (DCC) zabraňují toku protonů přes ATP synthasu.
63 Inhibice oxidativní fosforylace. Odpojení (rozpojení) elektronového transportu od syntézy ATP. Na příklad 2,4-dinitrofenol (DNP) a řada dalších aromatických kyselých sloučenin. Tyto látky přenáší protony zpět z cytoplasmy do matrix mimo ATP synthasu. Nedochází k tvorbě ATP. Zvyšuje se spotřeba kyslíku a oxidace NADH. NO 2 Často se energie uvolňuje jako teplo. DNP je součástí herbicidů a fungicidů. Inhibice exportu ATP. ATP-ADP translokasa je inhibována již malými koncentracemi atraktylosidu (rostlinný glykosid) nebo bongkrekovou kyselinou (antibiotikum). Atraktylosid se váže na translokasu na straně cytoplasmy, bongkreková kyselina v matrix mitochondrie. OH NO 2 2,4 - dinitrofenol (DNP)
64 Inhibice a inhibitory oxidativní fosforylace. NADH NADH - Qoxidoreduktasa Rotenon, amytal QH 2 Q - cytochrom c oxidoreduktasa Antimycin A Cytochrom c Cytochrom c oxidasa CN -, N 3 - a CO O 2
65 Ústřední motiv bioenergetiky. Oxidativní fosforylace je realizována na bázi protonového gradientu. Protonové gradienty pohání řadu energii vyžadujících procesů jako je aktivní transport Ca 2+ iontů v mitochondrii, vstup některých aminokyselin a sacharidů do bakterií, pohyb bakteriálních bičíků a přenos elektronů z NADP + na NADPH. Protonový gradient se také uplatňuje při uvolnění tepla (hibernace). Obecně: Protonové gradienty jsou ústředním převoditelným platidlem biologických systémů.
66 Protonový gradient je převoditelná forma volné energie. Aktivní transport Protonový potenciál Produkce tepla Protonový gradient Rotace bičíku ATP Syntéza NADPH
OXIDATIVNÍ FOSFORYLACE
 OXIDATIVNÍ FOSFORYLACE OBSAH Mitochondrie Elektronový transport Oxidativní fosforylace Kontrolní systém oxidativního metabolismu. Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů Dýchací
OXIDATIVNÍ FOSFORYLACE OBSAH Mitochondrie Elektronový transport Oxidativní fosforylace Kontrolní systém oxidativního metabolismu. Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů Dýchací
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy
 Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
Dýchací řetězec, oxidativní fosforylace, mitochondriální transportní systémy JAN ILLNER Dýchací řetězec & oxidativní fosforylace Tvorba energie v živých systémech ATP zdroj E pro biochemické procesy Tvorba
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
MitoSeminář II: Trochu výpočtů v bioenergetice. Souhrn. MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK
 MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
MitoSeminář II: Trochu výpočtů v bioenergetice MUDr. Jan Pláteník, PhD. Ústav lékařské biochemie 1.LF UK (se zahrnutím cenných připomínek, kterými přispěl prof. MUDr. Jiří Kraml, DrSc.) 1 Dýchacířet etězec
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Transport elektronů a oxidativní fosforylace
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Elektronový transport a oxidativní fosforylace
 Elektronový transport a oxidativní fosforylace Mitochondrie Elektronový transport xidativní fosforylace Kontrolní systém oxidativního metabolismu. Moto: dpočívající lidské tělo spotřebuje asi 420 kj energie
Elektronový transport a oxidativní fosforylace Mitochondrie Elektronový transport xidativní fosforylace Kontrolní systém oxidativního metabolismu. Moto: dpočívající lidské tělo spotřebuje asi 420 kj energie
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Bp1252 Biochemie. #8 Metabolismus živin
 Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
CZ.1.07/2.2.00/ Obecný metabolismus. Energetický metabolismus (obecně) (1).
 (1).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
Úvod do buněčného metabolismu Citrátový cyklus. Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK
 Úvod do buněčného metabolismu Citrátový cyklus Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK METABOLISMUS = přeměna látek v organismu - má stránku chemickou (látkovou) - reakce anabolické
Úvod do buněčného metabolismu Citrátový cyklus Prof. MUDr. Jiří Kraml, DrSc. Ústav lékařské biochemie 1. LF UK METABOLISMUS = přeměna látek v organismu - má stránku chemickou (látkovou) - reakce anabolické
Citrátový cyklus a Dýchací řetězec. Milada Roštejnská Helena Klímová
 Citrátový cyklus a Dýchací řetězec Milada oštejnská elena Klímová 1 bsah 1 Citrátový cyklus Citrátový cyklus (reakce) Citrátový cyklus (schéma) espirace (dýchání) Vnější a vnitřní respirace Dýchací řetězec
Citrátový cyklus a Dýchací řetězec Milada oštejnská elena Klímová 1 bsah 1 Citrátový cyklus Citrátový cyklus (reakce) Citrátový cyklus (schéma) espirace (dýchání) Vnější a vnitřní respirace Dýchací řetězec
>>> E A1 + E A2. . aktivační energie potřebná k reakci bez přítomnosti katalyzátoru E A E A1. energie potřebná ke vzniku enzym-substrátového komplexu
 Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Enzymy Charakteristika enzymů- fermentů katalyzátory biochem. reakcí biokatalyzátory umožňují a urychlují průběh rcí v organismu nachází se ve všech živých systémech z chemického hlediska jednoduché nebo
Aerobní odbourávání cukrů+elektronový transportní řetězec
 Aerobní odbourávání cukrů+elektronový transportní řetězec Dochází k němu v procesu jménem aerobní respirace. Skládá se z kroků: K1) Glykolýza K2) oxidativní dekarboxylace pyruvátu K3) Krebsův cyklus K4)
Aerobní odbourávání cukrů+elektronový transportní řetězec Dochází k němu v procesu jménem aerobní respirace. Skládá se z kroků: K1) Glykolýza K2) oxidativní dekarboxylace pyruvátu K3) Krebsův cyklus K4)
E ŘEŠENÍ KONTROLNÍHO TESTU ŠKOLNÍHO KOLA
 Ústřední komise Chemické olympiády 48. ročník 2011/2012 ŠKOLNÍ KOLO kategorie A a E ŘEŠENÍ KONTROLNÍ TESTU ŠKOLNÍ KOLA KONTROLNÍ TEST ŠKOLNÍ KOLA (60 BODŮ) ANORGANICKÁ CEMIE 16 BODŮ Úloha 1 8 bodů Napište
Ústřední komise Chemické olympiády 48. ročník 2011/2012 ŠKOLNÍ KOLO kategorie A a E ŘEŠENÍ KONTROLNÍ TESTU ŠKOLNÍ KOLA KONTROLNÍ TEST ŠKOLNÍ KOLA (60 BODŮ) ANORGANICKÁ CEMIE 16 BODŮ Úloha 1 8 bodů Napište
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Dýchací řetězec (DŘ)
") Dýchací řetězec (DŘ) Vladimíra Kvasnicová animace na internetu: http://vcell.ndsu.nodak.edu/animations/etc/index.htm http://vcell.ndsu.nodak.edu/animations/atpgradient/index.htm http://www.wiley.com/college/pratt/0471393878/student/animations/oxidative_phosphorylation/index.html
Dýchací řetězec (DŘ) Vladimíra Kvasnicová animace na internetu: http://vcell.ndsu.nodak.edu/animations/etc/index.htm http://vcell.ndsu.nodak.edu/animations/atpgradient/index.htm http://www.wiley.com/college/pratt/0471393878/student/animations/oxidative_phosphorylation/index.html
Předmět: KBB/BB1P; KBB/BUBIO
 Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
Předmět: KBB/BB1P; KBB/BUBIO Energie z mitochondrií a chloroplastů Cíl přednášky: seznámit posluchače se základními principy získávání energie v mitochondriích a chloroplastech Klíčová slova: mitochondrie,
9. Dýchací řetězec a oxidativní fosforylace. mitochondriální syntéza ATP a fotosyntéza
 9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
B4, 2007/2008, I. Literák
 B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
B4, 2007/2008, I. Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují pořádek ve světě, který spěje k čím dál většímu chaosu Druhá věta termodynamiky: Ve vesmíru nebo jakékoliv izolované
Citrátový cyklus. Tomáš Kučera.
 itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Otázka: Metabolismus. Předmět: Biologie. Přidal(a): Furrow. - přeměna látek a energie
: Furrow. - přeměna látek a energie") Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Ukázky z pracovních listů z biochemie pro SŠ A ÚVOD
 Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno:
 Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Bruno Sopko Rychlost chemické reakce je dána změnou Gibbsovy energie a aktivační energií: Tudíž zrychlení reakce pomocí katalýzy může být vyjádřeno: Z předchozí rovnice vyplývá: Pokud katalýza při 25
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
DYNAMICKÁ BIOCHEMIE. Daniel Nechvátal :: www.gymzn.cz/nechvatal
 DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
12-Fotosyntéza FRVŠ 1647/2012
 C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Sacharidy a polysacharidy (struktura a metabolismus)
") Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Enzymologie. Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů.
 ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
ENZYMOLOGIE 1 Enzymologie Věda ležící na pomezí fyz. ch. a bioch. Zabývá se problematikou biokatalyzátorů. Jak je možné, že buňka dokáže utřídit hrozivou změť chemických procesů, které v ní v každém okamžiku
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Buněčný metabolismus. J. Vondráček
 Buněčný metabolismus J. Vondráček Téma přednášky BUNĚČNÝ METABOLISMUS základní dráhy energetického metabolismu buňky a dynamická podstata jejich regulací glykolýza, citrátový cyklus a oxidativní fosforylace,
Buněčný metabolismus J. Vondráček Téma přednášky BUNĚČNÝ METABOLISMUS základní dráhy energetického metabolismu buňky a dynamická podstata jejich regulací glykolýza, citrátový cyklus a oxidativní fosforylace,
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
Dýchací řetězec (Respirace)
") Dýchací řetězec (Respirace) Buněčná respirace (analogie se spalovacím motorem) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 ---------> 6 CO 2 + 6H 2 O + 38 ATP Oxidativní
Dýchací řetězec (Respirace) Buněčná respirace (analogie se spalovacím motorem) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 ---------> 6 CO 2 + 6H 2 O + 38 ATP Oxidativní
Lékařská chemie a biochemie modelový vstupní test ke zkoušce
 Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
Lékařská chemie a biochemie modelový vstupní test ke zkoušce 1. Máte pufr připravený smísením 150 ml CH3COOH o c = 0,2 mol/l a 100 ml CH3COONa o c = 0,25 mol/l. Jaké bude ph pufru, pokud přidáme 10 ml
ANABOLISMUS SACHARIDŮ
 zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
Metabolické dráhy. František Škanta. Glykolýza. Repetitorium chemie X. 2011/2012. Glykolýza. Jaký je osud pyruátu bez přítomnosti kyslíku?
 Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
METABOLISMUS SACHARIDŮ
 METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
5. Lipidy a biomembrány
 5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Přednáška 6: Respirace u rostlin
 Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
Přednáška 6: Respirace u rostlin co vás v s dnes čeká: Co rostliny získávají respirací Procesy respirace: glykolýza Krebsův cyklus dýchací řetězec oxidativní fosforylace faktory ovlivňující rychlost respirace
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Citrátový a glyoxylátový cyklus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Didaktické testy z biochemie 2
 Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolusmus lipidů - katabolismus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - katabolismus Trávení, aktivace a transport tuků Oxidace mastných kyselin Ketonové látky Úvod Oxidace MK je centrální
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - katabolismus Trávení, aktivace a transport tuků Oxidace mastných kyselin Ketonové látky Úvod Oxidace MK je centrální
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74. Katedra experimentální biologie rostlin, Z. Lhotáková
 FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
FOTOSYNTÉZA I. Přednáška Fyziologie rostlin MB130P74 Katedra experimentální biologie rostlin, Z. Lhotáková proteinové komplexy thylakoidní membrány - jsou kódovány jak plastidovými tak jadernými geny 1905
- metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy a jejich životním prostředím
 Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Kofaktory enzymů. T. Kučera. (upraveno z J. Novotné)
") Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Energetika a metabolismus buňky
 Předmět: KBB/BB1P Energetika a metabolismus buňky Cíl přednášky: seznámit posluchače s tím, jak buňky získávají energii k životu a jak s ní hospodaří Klíčová slova: energetika buňky, volná energie, enzymy,
Předmět: KBB/BB1P Energetika a metabolismus buňky Cíl přednášky: seznámit posluchače s tím, jak buňky získávají energii k životu a jak s ní hospodaří Klíčová slova: energetika buňky, volná energie, enzymy,
Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová
 Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová Vyberte esenciální aminokyseliny a) Asp, Glu b) Val, Leu, Ile c) Ala, Ser, Gly d) Phe, Trp Vyberte esenciální aminokyseliny a) Asp,
Metabolismus aminokyselin - testík na procvičení - Vladimíra Kvasnicová Vyberte esenciální aminokyseliny a) Asp, Glu b) Val, Leu, Ile c) Ala, Ser, Gly d) Phe, Trp Vyberte esenciální aminokyseliny a) Asp,
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
Efektivní adaptace začínajících učitelů na požadavky školské praxe
 Mezipředmětová integrace tělesná výchova biologie chemie Biochemie pro učitele tělesné výchovy III.: aerobní metabolismus (průvodce studiem) Filip Neuls, Ph.D. Průvodce studiem Z pohledu tělesného zatížení
Mezipředmětová integrace tělesná výchova biologie chemie Biochemie pro učitele tělesné výchovy III.: aerobní metabolismus (průvodce studiem) Filip Neuls, Ph.D. Průvodce studiem Z pohledu tělesného zatížení
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Biologické redoxní děje Biological redox processes. Tisková verze Print version Prezentace Presentation
 Biologické redoxní děje Biological redox processes Tomáš Kučera 2011 Tisková verze Print version Prezentace Presentation Biologické redoxní děje Slide 1a Biologické redoxní děje Biologické redoxní děje
Biologické redoxní děje Biological redox processes Tomáš Kučera 2011 Tisková verze Print version Prezentace Presentation Biologické redoxní děje Slide 1a Biologické redoxní děje Biologické redoxní děje
BÍLKOVINY. V organismu se nedají nahradit jinými sloučeninami, jen jako zdroj energie je mohou nahradit sacharidy a lipidy.
 BÍLKOVINY o makromolekulární látky, z velkého počtu AMK zbytků o základ všech organismů o rostliny je vytvářejí z anorganických sloučenin (dusičnanů) o živočichové je musejí přijímat v potravě, v trávicím
BÍLKOVINY o makromolekulární látky, z velkého počtu AMK zbytků o základ všech organismů o rostliny je vytvářejí z anorganických sloučenin (dusičnanů) o živočichové je musejí přijímat v potravě, v trávicím
BUŇKA A ENERGIE. kajman brýlový Caiman crocodilus Kostarika, 2004. Biologie 6, 2015/2016, Ivan Literák
 BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 6, 2015/2016, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
Přehled energetického metabolismu
 Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
Redoxní děj v neživých a živých soustavách
 Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Enzymy Enzymy Katalyzují chemické reakce, kdy se mění substrát na produkt Katalytickým působením se snižuje aktivační energie reagujících molekul substrátu, tím se reakce urychlí Za přítomnosti enzymu
Propojení metabolických drah. Alice Skoumalová
 Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
1- Úvod do fotosyntézy
 1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Opakování
 Slabé vazebné interakce Opakování Co je to atom? Opakování Opakování Co je to atom? Atom je nejmenší částice hmoty, chemicky dále nedělitelná. Skládá se z atomového jádra obsahujícího protony a neutrony
Slabé vazebné interakce Opakování Co je to atom? Opakování Opakování Co je to atom? Atom je nejmenší částice hmoty, chemicky dále nedělitelná. Skládá se z atomového jádra obsahujícího protony a neutrony
Metabolismus. Source:
 Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
Source: http://www.roche.com/ http://www.expasy.org/ Metabolismus Source: http://www.roche.com/sustainability/for_communities_and_environment/philanthropy/science_education/pathways.htm Metabolismus -
V organismu se bílkoviny nedají nahradit žádnými jinými sloučeninami, jen jako zdroj energie je mohou nahradit sacharidy a lipidy.
 BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
BÍLKOVINY Bílkoviny jsou biomakromolekulární látky, které se skládají z velkého počtu aminokyselinových zbytků. Vytvářejí látkový základ života všech organismů. V tkáních vyšších organismů a člověka je
Metabolismus. - soubor všech chemických reakcí a příslušných fyzikálních procesů, které souvisejí s aktivními projevy života daného organismu
 Metabolismus Obecné znaky metabolismu Získání a využití energie - bioenergetika Buněčné dýchání (glykolysa + CKC + oxidativní fosforylace) Biosynthesa sacharidů + fotosynthesa Metabolismus lipidů Metabolismus
Metabolismus Obecné znaky metabolismu Získání a využití energie - bioenergetika Buněčné dýchání (glykolysa + CKC + oxidativní fosforylace) Biosynthesa sacharidů + fotosynthesa Metabolismus lipidů Metabolismus
Metabolismus lipidů. (pozn. o nerozpustnosti)
") Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
BUŇKA A ENERGIE. kajman brýlový Caiman crocodilus Kostarika, Biologie 8, 2017/2018, Ivan Literák
 BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 8, 2017/2018, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
BUŇKA A ENERGIE kajman brýlový Caiman crocodilus Kostarika, 2004 Biologie 8, 2017/2018, Ivan Literák ENERGIE, KATALÝZA, BIOSYNTÉZA Živé organismy vytvářejí a udržují POŘÁDEK VE SVĚTĚ, KTERÝ SPĚJE K ČÍM
VAKUOLA. membránou ohraničený váček membrána se nazývá tonoplast. běžná u rostlin, zvířata specializované funkce či její nepřítomnost
 VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY
 Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
Inovace profesní přípravy budoucích učitelů chemie CZ.1.07/2.2.00/15.0324 Text zpracovala Mgr. Taťána Štosová, Ph.D PŘÍRODNÍ LÁTKY Obsah 1 Úvod do problematiky přírodních látek... 2 2 Vitamíny... 2 2.
Repetitorium chemie 2016/2017. Metabolické dráhy František Škanta
 Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
Repetitorium chemie 2016/2017 Metabolické dráhy František Škanta Metabolické dráhy Primární metabolismus-trávení Metabolismus sacharidů Glykolýza Krebsův cyklus Oxidativní fosforylace Metabolismus lipidů
Metabolismus aminokyselin 2. Vladimíra Kvasnicová
 Metabolismus aminokyselin 2 Vladimíra Kvasnicová Odbourávání AMK 1) odstranění aminodusíku z molekuly AMK 2) detoxikace uvolněné aminoskupiny 3) metabolismus uhlíkaté kostry AMK 7 produktů 7 degradačních
Metabolismus aminokyselin 2 Vladimíra Kvasnicová Odbourávání AMK 1) odstranění aminodusíku z molekuly AMK 2) detoxikace uvolněné aminoskupiny 3) metabolismus uhlíkaté kostry AMK 7 produktů 7 degradačních
16a. Makroergické sloučeniny
 16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Konsultační hodina. základy biochemie pro 1. ročník. Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa
 Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. ENZYMY I úvod, názvosloví, rozdělení do tříd
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ENZYMY I úvod, názvosloví, rozdělení do tříd Úvod z řeckého EN ZYME (v kvasinkách) biologický katalyzátor, protein (RNA) liší se od chemických
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti ENZYMY I úvod, názvosloví, rozdělení do tříd Úvod z řeckého EN ZYME (v kvasinkách) biologický katalyzátor, protein (RNA) liší se od chemických
Vzdělávací materiál. vytvořený v projektu OP VK. Anotace. Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20. Číslo projektu:
 Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
CYKLUS TRIKARBOXYLOVÝCH KYSELIN A GLYOXYLÁTOVÝ CYKLUS
 YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
Název: Fotosyntéza, buněčné dýchání
 Název: Fotosyntéza, buněčné dýchání Výukové materiály Autor: Mgr. Blanka Machová Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: Biologie, chemie Ročník: 2. Tematický
Název: Fotosyntéza, buněčné dýchání Výukové materiály Autor: Mgr. Blanka Machová Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: Biologie, chemie Ročník: 2. Tematický
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
FOTOSYNTÉZA. Princip, jednotlivé fáze
 FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/ Anotace. Fotosyntéza světelná fáze. VY_32_INOVACE_Ch0214.
 Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Metabolismus bílkovin. Václav Pelouch
 ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 4. Membránové proteiny Ivo Frébort Lipidová dvojvrstva Biologické membrány Integrální membránové proteiny Transmembránové proteiny Kovalentně ukotvené membránové
Struktura a funkce biomakromolekul KBC/BPOL 4. Membránové proteiny Ivo Frébort Lipidová dvojvrstva Biologické membrány Integrální membránové proteiny Transmembránové proteiny Kovalentně ukotvené membránové