Možnosti ovlivňování světelných podmínek při pěstování rostlin v současných interiérech
|
|
|
- Roman Pokorný
- před 5 lety
- Počet zobrazení:
Transkript
1 MENDELOVA UNIVERZITA V BRNĚ, ZAHRADNICKÁ FAKULTA V LEDNICI Možnosti ovlivňování světelných podmínek při pěstování rostlin v současných interiérech Diplomová práce Vedoucí diplomové práce: Ing. Tatiana Kuťková Csc. Vypracovala: Lucie Fikarová Lednice 2011
2 Prohlašuji, že jsem diplomovou práci na téma Možnosti ovlivňování světelných podmínek při pěstování rostlin v současných interiérech vypracovala samostatně, za odborného vedení vedoucí diplomové práce a použila jen pramenů, které jsou uvedeny v seznamu literatury. Souhlasím, aby práce byla uložena v knihovně Zahradnické fakulty Mendelovy univerzity v Brně a zpřístupněna ke studijním účelům. Lucie Fikarová
3 Děkuji mé vedoucí diplomové práce Ing. Tatianě Kuťkové Csc. za odborné vedení a rady při zpracování této diplomové práce. Dále bych chtěla poděkovat panu Ing. Stanislavu Hašovi za pomoc a cenné rady, hlavně při zpracování témat týkajících se světelné techniky.
4 OBSAH 1. ÚVOD CÍL PRÁCE LITERÁRNÍ ČÁST Světelné záření a jeho význam pro rostliny Fotosyntetické účinky světla Fotomorfogenetické účinky světelného záření Fotoperiodické účinky světla Světlo a ostatní pěstební faktory Světlo a teplota Světlo a vlhkost prostředí Vzájemné vztahy mezi vlhkostí, teplotou a intenzitou světla Světelně technické jednotky Svítivost Světelný tok Osvětlení Emisní spektrum Teplota chromatičnosti Index podání barev Barva světla Jednotky pro fotosynteticky aktivní záření Interiéry podle světelných podmínek Interiéry s nedostatkem světla Možnosti řešení nedostatečného osvětlení v interiérech Rostliny vhodné do interiérů s nedostatkem světla Interiéry s nadbytkem světla Možnosti řešení nadměrného osvětlení v interiéru Rostliny vhodné do interiérů s nadbytkem světla
5 4. MATERIÁL A METODIKA Hodnocení světelných podmínek pro potřeby pěstování rostlin Obecné charakteristiky hodnoceného interiéru Podmínky při měření Vyhodnocení vhodnosti umístění interiérových rostlin podle naměřených hodnot osvětlenosti Přisvětlování rostlin Modelové objekty Vstupní hala Zahradnické fakulty v Lednici Vstupní prostor kolejí Kancelář firmy Spektrum Holas v Havlíčkově Brodě VÝSLEDKY Hodnocení světelných podmínek ve vybraných interiérech z pohledu potřeb pro pěstování rostlin Přisvětlování rostlin Modelové objekty návrhy řešení prostor s nevhodnými světelnými podmínkami Vstupní hala budovy A Vstupní prostor kolejí Kancelář firmy Spektrum Holas v Havlíčkově Brodě DISKUZE ZÁVĚR RESUMÉ POUŽITÁ LITERATURA PŘÍLOHY
6 1. ÚVOD Lidé v současné době tráví většinu svého času v uzavřených prostorech různých budov, kde pracují, odpočívají, nakupují a podobně. Kontakt s přírodou jim tu často umožňují interiérové rostliny, které mají velmi pozitivní vliv na lidskou psychiku. Při pěstování rostlin v interiéru je limitujícím faktorem, který rozhoduje o tom, zda je možné rostliny v daném prostoru pěstovat, hlavně světlo. Teplota je většinou dána účelem, ke kterému interiér slouží, vzdušnou vlhkost můžeme ovlivňovat a zálivka, substrát i výživa jsou plně v našich rukou. Světelné podmínky můžeme také ovlivnit, ale je to náročnější a dražší než u ostatních pěstebních faktorů. Přesto se to v některých případech jistě vyplatí. Pozitivní působení a krásu živých rostlin je těžké něčím nahradit, právě proto, že jsou živé, rostou, vyvíjejí se a neustále se mění. V našich geografických podmínkách je nejčastějším problémem nedostatek světla v zimě, kdy jsou krátké dny a sluneční svit nemá takovou intenzitu, jakou by rostliny potřebovaly. U moderních prosklených staveb, jižních oken, zimních zahrad, apod. je naopak problémem nadbytek světla v létě, který může rostliny také poškodit. Diplomová práce je zaměřena na popis různých způsobů řešení obou těchto problémů, vytvoření metodiky hodnocení interiérů a na návrhy konkrétních řešení modelových objektů. Během jejího zpracování byla také zahájena spolupráce na soukromém projektu: Osvětlování interiérových rostlin. Cílem tohoto projektu je: 1. Na základě literárních podkladů provést analýzu této problematiky, objasnit metody hodnocení a měření světelného záření z hlediska fotobiologických procesů rostlin, 2. shromáždit údaje o působení rostlin v interiérech na jeho prostředí a zejména na člověka, jeho pracovní aktivitu a psychickou kondici, 3. vykonat pokusy a provést analýzu vlivu osvětlení zdroji s různými emisními spektry na barevné podání rostlin a provést příkladný výběr vhodných světelných zdrojů, provést analýzu jejich světelně technických vlastností (spektra, rozdělení svítivosti) 4. provést světelně technické výpočty několika variant osvětlovaných květinových ploch pro různé světelné náročnosti rostlin, 5. provést výběr vhodných sestav rostlin pro počítané květinové plochy, 3
7 6. navrhnout různé možnosti uspořádání osvětlovacích souprav pro umístění na podlahu, na podložky a pro závěsné stropní osvětlení, 7. zpracovat technickou dokumentaci a zajistit výrobu funkčních vzorků svítidel 8. publikovat všechny dosažené výsledky v časopise Světlo 9. uspořádat seminář na téma: Osvětlování interiérových rostlin 4
8 2. CÍL PRÁCE Cílem diplomové práce je poskytnout ucelený přehled možností řešení nevhodných světelných podmínek v interiérech z hlediska potřeb pro pěstování rostlin, vypracovat metodiku hodnocení těchto světelných podmínek, navrhnout řešení pro konkrétní interiéry s nevhodnými světelnými podmínkami, provést pokus s přisvětlováním rostlin a pracovat na dalších dílčích úkolech soukromého projektu. 5
9 3. LITERÁRNÍ ČÁST 3.1 Světelné záření a jeho význam pro rostliny Záření je pro rostliny v prvé řadě zdrojem energie. Dále rostliny informuje o vlastnostech prostředí, ve kterém se nacházejí. Na základě těchto informací mohou usměrňovat růst, vývoj i pohyby jednotlivých svých částí takovým způsobem, aby bylo zajištěno přežití a reprodukce v daném typu prostředí. Informační význam pro rostliny má intenzita ozáření, směr, ze kterého dopadá na rostlinu, spektrální složení a periodicita (Gloser, 1998). Světelné záření vyvolává u rostlin reakce fotosyntetické, fotomorfogenetické a fotoperiodické (viz Obrázek 1 v příloze I) Fotosyntetické účinky světla Pro rostlinu je důležité, aby fotosynteticky aktivní záření optimálně využívala, tj. aby vytvořila dostatek asimilátů pro udržení struktury pochodů stávajících i pro vývoj struktur nových, ale také aby nebyla poškozována nadbytkem dopadající energie. Proto se světelným podmínkám na stanovišti po celý život neustále aktivně přizpůsobuje. Světelné podmínky na stanovišti zjišťuje prostřednictvím fotosenzorů a informace přenáší specifickými signálními cestami. Podle těchto informací rostlina utváří svou vnější podobu (např. délku internodií a řapíků, velikost plochy listových čepelí), vnitřní struktury a ultrastruktury, např. uspořádání listového mezofylu, utváření fotosyntetického aparátu (Pavlová, 2005). Fotosyntéza je fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických syntézách, což je proces tvorby organických látek (primárně sacharidů) z látek anorganických, a to z oxidu uhličitého a vody. Fotosyntéza vyšších rostlin je vázána především na mezofylové buňky v listech a probíhá ve specializovaných organelách, které se nazývají chloroplasty. V nich probíhá fotosyntéza ve dvou fázích světelné a temnotní. Pro světelnou fázi, ve které probíhají fotochemické reakce, je charakteristická rovnice: 12 H 2 O + světelné záření 12 H O 2. Podstatou těchto primárních reakcí je tedy štěpení vody na světle (proces se nazývá fotolýza vody nebo také Hillova reakce), který poskytuje elektrony a vodíkové kationty (protony) pro tvorbu ATP a NADPH + H +. 6
10 Temnotní fázi fotosyntézy s charakteristickými enzymatickými reakcemi můžeme zjednodušeně popsat rovnicí: 6 CO H 2 C 6 H 12 O H 2 O. Sumárně pak můžeme celý proces fotosyntézy vyjádřit základní rovnicí: 6 CO H 2 O + světelné záření C 6 H 12 O O H 2 O (Hejnák, 2007). Množství záření kolísá v denních i ročních cyklech a navíc v porostech se mění i v závislosti na umístění listů v prostoru (Gloser, 1998). Pro fotosyntézu je důležitá pouze ta část spektra, která je absorbována asimilačními pigmenty, tedy přibližně v rozmezí vlnových délek 400 až 700 nm která se označuje jako fotosynteticky aktivní radiace - FAR (Hejnák, 2007). Také lidské oko se během vývoje přizpůsobovalo slunečnímu záření, a tak se hranice fotosynteticky účinné oblasti přibližně shodují s hranicemi viditelné části slunečního spektra (380 až 750 nm). Uvnitř tohoto intervalu je však závislost účinku záření na vlnové délce zcela jiná pro lidské oko než pro fotosyntézu: průměrné lidské oko je nejcitlivější na paprsky zelené vlnové délky asi 555 nm, kdežto zelené rostliny mohou nejlépe využívat paprsky červené ( nm) a modré ( nm), zelené paprsky z největší části propouštějí nebo odrážejí. Proto také byly zavedeny pro stejnou spektrální oblast dva různé pojmy, světlo a fotosynteticky účinné záření, jak ukazuje Obrázek 2 v Příloze I (Podešva, Véber, Kubín, 1968). Tok fotonů má maximální hustotu v oblasti kolem 620 nm. Největší množství energie, tj. maximum toku energie, je vyzařováno v oblasti kolem 500 nm (Pavlová, 2005). Záření dopadající na jednotku plochy měříme jako ozářenost v jednotkách intenzity ozáření (W*m -2 FAR), či jako hustotu toku fotosynteticky aktivních fotonů, která se udává v mol*m -2 *s -1 někdy též mol* s -1 *m -2 (Gloser, 1998). Světlo dopadající ve formě fotonů uvádí molekuly asimilačních pigmentů do vybuzeného excitovaného stavu (jde o tzv. nižší excitovaný stav, při kterém dochází k vypuzení elektronu v molekule chlorofylu na vyšší orbital). Takto elektronicky vybuzená molekula může proměnit excitační energii různými způsoby: a) Dochází k její vnitřní přeměně na kinetickou energii pohybu molekul, tj. na teplo b) Excitovaná molekula foton znovu vypudí fluorescence energie vypuzeného fotonu je nižší, fluorescencí se tak ztrácí 3 6 % světelné energie. c) Nejvýznamnější je způsob, při kterém dochází k přenosu excitační energie excitonu při tom vybuzená molekula bezprostředně přenáší svou excitační energii na blízkou neexcitovanou molekulu. Podobá se to vzájemnému působení mechanicky 7
11 spřažených kyvadel. Exciton je přenášen směrem k reakčnímu centru, protože čím se molekuly pigmentů nacházejí blíže reakčnímu centru, tím menší energii potřebují k excitaci (tedy i maximum jejich absorpce leží v delších vlnových délkách). Je to závislé na typu pigmentu a na způsobu jeho vazby k molekulám bílkovin. Karotenoidy a chlorofyl b potřebují vyšší excitační energii než chlorofyl a (Tůma, Tůmová, 1998). Možnosti přeměny excitační energie ukazuje Obrázek 3 v Příloze I. V rostlinách je obsaženo mnoho různých látek, které mají schopnost absorbovat sluneční záření. Látky, které odrážejí záření ve viditelné oblasti světla, se člověku jeví jako barevné a většinou se charakterizují jako barviva neboli pigmenty. Společným znakem těchto látek je přítomnost většího počtu konjugovaných dvojných vazeb v molekule. Chemicky, funkčně i co do lokalizace v rostlině i buňce se jedná o velmi heterogenní skupinu látek. Chlorofyly a karotenoidy barviva absorbující energii pro fotosyntézu jsou lipofilní povahy a jsou vázána v chloroplastech. Karotenoidy plní současně také funkci ochrannou, a to i v chromoplastech květů a plodů, kde jsou také součástí strategie sexuální reprodukce. Flavonoidy absorbují především záření UV a jejich funkce je ochranná. Jsou hydrofilní, lokalizované ve vakuolách. Nacházejí se nejčastěji v květech, ale také v povrchových vrstvách nadzemní části rostliny. Fytochromy, kryptochromy, fototropin barevné molekuly s funkcí fotosenzorickou se často jako pigmenty označují, ale na zbarvení rostlin se pro nízký počet svých molekul prakticky nepodílejí. Monitorují světelné podmínky na stanovišti a zprostředkují informaci pro děje fotomorfogenetické. Nacházejí se v cytoplazmě, jádře i plazmalemě. Na zbarvení rostlin se podílí i řada sekundárních metabolitů a dalších látek, které mají schopnost absorbovat záření v určitých oblastech světla. Tato energie se však mění na teplo a nevstupuje do životních procesů (Pavlová, 2005). Primární pochody fotosyntézy probíhají v tylakoidních membránách. Na většinu bílkovin integrovaných v tylakoidních membránách se váže velké množství asimilačních barviv, která jsou schopna absorbovat světelné záření. U vyšších rostlin se vyskytuje především chlorofyl a, ale i chlorofyl b a další pomocná barviva, která souhrnně označujeme jako akcesorická barviva. Jsou to hlavně karoteny a xantofyly. Pigmenty se v procesu fotosyntézy opotřebovávají, proto je nutná jejich obnova - až 8 % za 24 hodin (Tůma, Tůmová, 1998). Chlorofyly absorbují fotony v modré a červené oblasti spektra, v oblasti zelené je jejich absorpce minimální a jako zelené se člověku jeví (Pavlová, 2005). 8
12 Výjimečné postavení zde má chlorofyl a nejen proto, že je zastoupen v daleko největším množství, ale také pro schopnost některých molekul chlorofylu a přejít do excitovaného stavu a předat elektron na elektron- transportní systémy. Tímto donorem elektronů bývá chlorofyl a, který je umístěn v reakčním centru fotosystému, kde se zachytí excitační energie. V tylakoidních membránách jsou lokalizovány dva druhy reakčních center. Fotosystém I obsahuje především molekulu chlorofylu a, která se označuje P 700, neboť nejvíce absorbuje záření o vlnové délce 700 nm. Ve fotosystému II je vázána molekula P 680. Daleko méně je u vyšších rostlin zastoupen chlorofyl b (poměr asi 3:1) a je součástí pouze anténního systému, takže stejně jako další barviva má pouze akcesorickou funkci. Pro fungování fotosyntetického aparátu není nezbytný (Tůma, Tůmová, 1998). Karotenoidy absorbují fotony ve fialové a modré oblasti světla a jeví se jako žluté až oranžové. V chloroplastech vyšších rostlin se vyskytuje hlavně -karoten, z xantofylů pak lutein, violaxantin, zeaxantin a neoxantin. Karotenoidy jsou syntetizovány na světle i ve tmě, světlo jejich tvorbu výrazně stimuluje (Pavlová, 2005). Karotenoidy mají světlosběrnou funkci, ale i funkci ochrannou. Pokud molekuly chlororofylu absorbují více zářivé energie, než může být využito pro přenos elektronů, může dojít k aktivizování kyslíku, jehož některé formy mají destrukční účinky na tylakoidy. Pokud je ale nadbytečná energie převedena ke karotenoidům, zneškodní se přeměnou na teplo. Tato funkce karotenoidů je pro život rostlin nezbytná (Tůma, Tůmová, 1998). Při průchodu listem se záření kvalitativně i kvantitativně mění. Klesá podíl modré ( nm) a krátkovlnné červené složky ( nm), které jsou absorbovány fotosyntetickými pigmenty. Relativně stoupá záření v oblastech, v nichž je absorpce fotosyntetickými minimální nebo žádná, tedy v oblasti zelené ( nm) a dlouhovlnné červené ( nm). Absolutně se může zvyšovat množství záření v oblasti infračervené (nad 740 nm), neboť může vznikat při deexcitaci fotosyntetických pigmentů. Rychlost fotosyntézy závisí na množství využitelné energie, která dopadá na fotosynteticky aktivní pletiva (Pavlová, 2005). Vztah mezi rychlostí čisté fotosyntézy a ozářeností listu není lineární, nýbrž má tvar limitní křivky charakteristického tvaru. Tradičně bývá označována jako světelná křivka fotosyntézy. Její počátek leží u takové hodnoty ozářenosti, při které rychlost hrubé fotosyntézy je právě dostatečná na kompenzaci současně probíhající respirace (=kompenzační ozářenost, nebo-li světelný kompenzační bod). Hodnota kompenzační ozářenosti je závislá především na rychlosti respirace (a tudíž i na teplotě), a také na 9
13 strukturních charakteristikách listů. Obojí však může být závislé opět na druhu rostliny a podmínkách, za kterých roste. Nejčastěji se pohybuje v rozmezí 1 až 5 W*m -2 FAR, což odpovídá přibližně 5 až 20 mol*m -2 *s -1 (Gloser, 1998). Čím vyšší hodnotu má světelný kompenzační bod, tím je rostlina méně stínobytná (Faustus, Kincl, 1977). Při dosažení určité hodnoty ozářenosti již rychlost fotosyntézy nevzrůstá. Tato hodnota se nazývá saturační ozářenost. Obecně platí, že listy s nízkou hodnotou saturační ozářenosti mají i nízkou rychlost fotosyntézy při nasycení zářením (Gloser, 1998). Velikost plochy listové čepele určuje množství záření, které na ni dopadá. Na stanovištích zastíněných porostem jsou listové čepele větší a tenčí než na otevřených prostranstvích. Kutikula a epidermis dále rozhodují o množství záření, které do listu vstoupí. Trichomy, kutikulární a epikutikulární vosky a další struktury, např. různé typy žlázek, zvyšují množství odraženého záření a snižují množství fotosynteticky aktivního i tepelného záření, které do listu vstupuje. Epidermální buňky na vnější straně konvexní mohou naopak působit jako čočky a zvýšit množství energie vstupující do listu. Záření, které prošlo epidermis, vstupuje do mezofylu, jehož uspořádání také odpovídá světelným podmínkám na stanovišti. Listy na slunných stanovištích mají více vrstev palisádového parenchymu i vrstva houbového parenchymu je vyšší a listové čepele jsou tlustší než na stanovištích stinných. V palisádovém parenchymu je část fotosynteticky aktivního záření, které dopadá na chloroplasty, absorbována, zbytek záření prochází vakuolami nebo cytoplazmou mimo chloroplasty a vstupuje do buněk nižších vrstev listového mezofylu nebo do mezibuněčných prostor. Na rozhraních různých prostředí, např. na povrchu buněčných stěn, se záření rozptyluje (odráží se nebo lomí). Předpokládá se, že díky rozptylu, může být množství záření uvnitř listu vyšší než v okolním prostředí (existuje představa listu jako pasti na světlo). Odhaduje se, že foton může uvnitř listu urazit dráhu až několikanásobně delší, než je tloušťka listu. Světelným podmínkám odpovídá i vznik funkčního fotosyntetického aparátu a jeho uspořádání. Podle informací o světelných podmínkách je řízena exprese jaderných genů, které kódují proteiny fotosyntetického aparátu, např. proteiny LHC a ATPsyntázy, protein malé podjednotky Rubisco, katalytické proteiny Calvinova cyklu, prochlorofylidreduktázu nebo ferredoxin. U krytosemenných rostlin je světlo nezbytné k vlastní syntéze chlorofylu při fotochemické reakci prochlorofylidu na chlorofylid. 10
14 Ve srovnání s listy slunných stanovišť mají listy na stinných stanovištích více gran v thylakoidní membráně, vyšší poměr počtu fotosystémů II k počtu fotosystémů I a fotosystémy mají celkově vyšší obsah chlorofylu. Ve stromatu plastidů stinných listů je celkově méně proteinů než ve stromatu chloroplastů na stanovištích slunných. Listy slunných stanovišť mají naopak vyšší obsah karotenoidů, poměr fotosystémů II/I je blízký 2:1. Na aktuální světelné podmínky rostlina reaguje různými mechanismy. Změna postavení listu vzhledem ke směru dopadajícího záření je mechanismus, jímž rostlina může podle aktuálních podmínek na stanovišti aktivně regulovat množství energie dopadající na list. Tato pohybová reakce se nazývá heliotropismus. Největší množství záření dopadá na listovou čepel, stojí-li kolmo k dopadajícímu záření. Mnoho druhů rostlin celý den nastavuje čepele do této polohy. Za svítání jsou čepele postaveny téměř zcela vertikálně a natočeny vrchní stranou k východu, přes den sledují pohyb slunce po obloze a plynule přecházejí do polohy horizontální, kterou zaujímají v poledne. Na konci dne jsou opět v poloze téměř vertikální, vrchní strana je natočena k západu. V noci se čepele staví horizontálně a před svítáním se opět otáčejí k východu. Pokud je slunce kryto mraky, pohyb obvykle ustává. Tento typ reakce vede k tomu, že množství dopadajícího záření je maximální. Pokládá se za reakci kladnou a označuje se jako diaheliotropismus. Heliotropická reakce však může vést i ke snížení množství dopadajícího záření. Tento typ reakce je považován za negativní a označuje se jako paraheliotropismus. U některých druhů rostlin se charakter reakce mění podle dostupnosti vody. Aktivní přemisťování chloroplastů je další mechanismus, kterým rostlina může regulovat množství absorbovaného záření. Ve tmě a při nízkých ozářenostech jsou chloroplasty rozmístěny téměř homogenně v horní povrchové oblasti cytosolu, v rovině kolmé k dopadajícímu záření, což zvyšuje pravděpodobnost absorpce FAR. Při vysokých ozářenostech se chloroplasty přemisťují do bočních oblastí buňky, do roviny rovnoběžné s dopadajícím zářením, stíní se, množství záření, které na ně dopadá a může být absorbováno, se snižuje (Pavlová, 2005). Chloroplasty se staví na buněčné stěny rovnoběžně se směrem paprsků, a proto listy blednou (Faustus, Kincl, 1978) Fotomorfogenetické účinky světelného záření Viditelné záření ovlivňuje vývoj rostlin i při velmi nízkém toku, kdy ještě neprobíhá fotosyntéza. V tomto případě světlo nemá energetickou funkci, ale je signálem iniciujícím 11
15 vývojové změny v rostlinách. Morfogenetickou odpověď v rostlinách vyvolává červené a modré světlo (Masarovičová, 2008). Fotomorfogenezi lze definovat jako vývojovou strategii rostlin vystavených záření nebo jako integrovaný časový a prostorový sled růstových a vývojových změn indukovaných zářením (Procházka, 1998). Rostliny reagují pouze na záření určitých vlnových délek, o čemž svědčí tzv. akční spektra závislost odezvy na vlnové délce působícího záření. Nejaktivnějšími oblastmi spektra záření, které nejvíce ovlivňují růst a vývoj rostlin, je oblast červeného záření, s maximy vlnových délek 660 a 730 nm, a záření modré s absorpčními maximy 370, 450 a 480 nm. Růstově aktivní je i ultrafialové záření v oblasti UVA ( nm) a někdy i UVB-B záření v rozmezí vlnových délek nm. Záření dopadá na povrch rostlin, část se od povrchu odráží a část prochází rostlinnými pletivy, kde je absorbováno. V důsledku toho různými rostlinnými orgány prochází různě oslabené záření s odlišným zastoupením jednotlivých vlnových délek (Procházka, 1998). Vnímání světelného signálu rostlinnými buňkami vyžaduje přítomnost pigmentu fotoreceptoru, který se po absorpci světla stane fotochemicky aktivním (Masarovičová, 2008). Na informace o světelných poměrech na stanovišti, které rostliny pomocí fotoreceptorů získávají, dle svých možností aktivně a koordinovaně reagují utvářejí habitus i vnitřní struktury, modulují metabolismus a mění polohu orgánů i organel (Pavlová, 2005) Receptory záření Fotosenzory absorbují dopadající záření, energie však není využívána k fotosyntetické asimilaci anorganických látek a nevstupuje do energetického metabolismu rostliny, ale přináší informaci je zdrojem signálu. (Pavlová, 2005). Aby rostlina vůbec mohla využít informace přenášené zářením, je nutná jeho absorbce pomocí fotoreceptorů, dále pak převod a zesílení přijatého signálu (Gloser, 1998). Rostliny využívají oblasti spektra, které jsou nutné pro fotosyntézu, a oblasti k nim přilehlé, tj. oblast krátkovlnnou červenou, značenou R (z angl. Red; 550 až 700 nm), a dlouhovlnnou červenou, značenou FR (z angl. Far Red; 660 až 780 nm) a oblast modrou, značenou B (z angl. Blue; 425 až 490 nm), s přesahem do oblasti fialové (400 až 425 nm) a UV-A (320 až 400 nm) (Pavlová, 2005). 12
16 Červenou oblast (R i FR) monitorují fytochromy, modrou oblast s přesahem do oblasti UV-A kryptochromy a fototropin (Pavlová, 2005). Masarovičová (2008) uvádí i další receptory modrého záření např. cis- zeaxantin. Fytochromy a kryptochromy zprostředkují informaci o tom, zda je světlo nebo tma a jejich signál je předáván i mechanismu, který u rostlin měří čas. Fytochromy také přinášejí informaci o poměru R/FR, který rostlinu informuje o přítomnosti konkurujících rostlin na stanovišti nebo o přítomnosti vlastních fotosynteticky aktivních struktur, např. listů v rámci koruny jednoho stromu. Oblačnost může výrazně měnit celkové množství dopadající energie, nemění však poměr jednotlivých složek. Pohybové reakce jsou řízeny především kryptochromovým signálem. Fototropin zprostředkuje signál pro ty pohybové reakce, které reflektují pozici zdroje tropismy. Předpokládá se existence ještě dalších fotosenzorů, především pro oblast modrého světla a záření v oblasti UV-B (280 až 320 nm). Signální dráhy jednotlivých receptorů mají určité specifické složky, ale během přenosu signálu interferují nejen spolu ale i s dalšími signálními drahami. Výsledná fotomorfogenetická reakce (interpretace signálu) má komplexní charakter a odráží nejen přijaté signály, ale i schopnost rostliny na signály reagovat. Schopnost reagovat na světelné signály závisí na genetické výbavě i aktuálním stavu rostliny, např. na jejím stáří, množství dostupných asimilátů, minerálních látek i vody (Pavlová, 2005). Fytochrom Jedná se o fotoreverzibilní chromoprotein a relativní molekulovou hmotností a prostetickou skupinou tetrapyrolové struktury. Ten při absorbci záření o vlnových délkách s maximem 660 nm (red, R) mění svoji konfiguraci a po ozáření vlnovými délkami s maximem 730 nm (far-red, FR) se vrací do původní formy. Zároveň se mění i struktura celého bílkovinného komplexu i jeho biologická aktivita; biologicky aktivní je forma Pfr, i když v některých účincích fytochromu může být aktivní i forma Pr. (Procházka, 1998). Gloser (1998) a Tůma, Tůmová (1998) však uvádějí, že procesy v rostlinách jsou aktivovány jen formou Pfr, zatímco Pr je zjevně fyziologicky neúčinná. Ve svém funkčním stavu se fytochrom vyskytuje jako dimer s M r (Procházka, 1998). Tento dimer se skládá z 2 identických polypeptidů homodimerů a na každém z nich je přes sirný můstek připojena zvláštní prostetická skupina chromofor (Gloser, 1998). Neproteinová část je příbuzná modrým barvivům fykocyanům (Šebánek, 1983). Forma Pr modré barvy je cis izomerem. Absorpce červeného světla Pr formou vyvolá změnu konformace. To způsobí 13
17 změnu absorpčního spektra na formu Pfr absorbující tmavočervené světlo (Masarovičová 2008). Tato reakce se nazývá fototransformace (Pavlová, 2005). Forma Pfr je trans izomerem modrozelené barvy a je to fyziologicky aktivní forma. Formy fytochromů jsou vzájemně konvertibilní, tmavočervené světlo mění Pfr zpět na Pr (Masarovičová, 2008). Absorbční spektra obou forem pigmentu se překrývají, proto se nemůže konvertovat 100 % pigmentu na druhou formu. Osvětlením červeným světlem se dosáhne 85 % formy Pfr a 15 % Pr. Znamená to, že není možná přeměna všech molekul Pr na Pfr, neboť Pfr, i když s malou účinností, absorbuje i červené světlo. Část molekul Pfr se proto hned fototransformuje zpět na Pr. Podobně také Pr absorbuje malé množství tmavočerveného světla, které přemění část Pr na Pfr. Takto se dosáhne fotorovnováha neboli fotostacionární stav 97 % Pr a 3% Pfr. V podmínkách bílého světla obsahujícího červené i tmavočervené vlnové délky dochází k dynamickým přeměnám mezi Pr a Pfr (Masarovičová, 2008). K transformaci Pr na Pfr obvykle postačuje velmi nepatrné množství červeného záření asi 200 J * m -2, což je méně než 1 % přijaté energie z plného slunečního záření za dobu jedné minuty (Gloser, 1998). Forma Pr je poměrně stálá, zatímco forma Pfr se ve tmě rychle mění zpět na Pr (temnotní reverze) a zčásti se rozkládá. Celkový úbytek fytochromu je pak kompenzován syntézou de novo, při které je receptor syntetizován ve formě Pr. Účinky vyvolané ozářením R je možné zrušit ozářením FR jen během určité doby po ukončení působení R. Tato doba se nazývá únikový čas a může trvat od několika minut do několika hodin. Za tuto dobu tedy proběhne reakce vyvolaná ozářením R do takové fáze, ve které již není ovlivnitelná ozářením FR. To ukazuje, že k vyvolání některých jevů je nutná přítomnost aktivní formy Pfr jen po určitou, někdy velmi krátkou dobu (Procházka, 1998). Fytochrom je schopen reagovat i na velmi slabou ozářenost, od 5mW * m -2 (Kubín, 1983). Protože taková ozářenost je hluboko pod hodnotou požadovanou pro fotosyntézu, lze podle intenzity použitého záření odlišit vlivy zprostředkované fotosyntézou od vlivů ostatních (Procházka, 1998). Podle prahové hodnoty intenzity ozářenosti nutné k vyvolání dané odezvy lze fytochromem řízené reakce rozdělit na několik typů: a) Odezvy na velmi nízkou prahovou ozářenost (very low irradiance responses VLIR) v nich fytochrom reaguje již na ozářenost 50 mw *m -2 (Kubín, 1983). Příkladem je vnímání záření na počátku dne, při svítání. Tyto odezvy není možné ovlivnit ozářením FR, neboť při nich jsou pouze asi 0,02 % fytochromu ve formě Pfr a FR převádí na formu Pr jen 97 % přítomného fytochromu. (Procházka, 1998). Také klíčení semen 14
18 mnoha druhů plevelů, která jsou uložena dlouhodobě ve tmě v hloubce půdy, se může indukovat velmi malými dávkami světla (Masarovičová, 2008). b) Odezvy na nízkou prahovou ozářenost (low irrradiance responses LIR) prahová hodnota zářivého toku je asi 500 mw*m -2 (Kubín, 1983). Mezi tyto odezvy patří většina klasických fotoreverzibilních reakcí, např. stimulace klíčení, inhibice kvetení. Oba typy odpovědí (VLIR a LIR) se indukují malou dávkou záření. Celková dávka ozáření je funkcí toku záření a doby ozáření, přičemž při obou odpovědích platí zákon reciprocity. Potřebná dávka fotonů se může aplikovat buď větším tokem fotonů za krátký čas nebo malým tokem fotonů za dlouhý čas ozáření (Masarovičová, 2008). c) Odezvy na vysokou ozářenost (high irradiance responses HIR) tyto odezvy vyžadují obvykle dlouhodobé působení ozářenosti, které je vyšší než 5 W*m -2 (Kubín, 1983). Mají akční spektrum s maximy v oblasti FR a modrého záření a nejsou fotoreverzibilní. Regulace těchto procesů se zřejmě účastní fytochrom spolu s receptorem pro modré záření (Procházka, 1998). Odpovědi HIR nejsou fotoreverzibilní a neřídí se zákonem reciprocity. HIR odpovědi se týkají buď etiolovaných (např. inhibice prodlužování hypokotylu, syntéza antokyanů) nebo zelených rostlin (indukce kvetení, tvorba etylenu, zvětšení klíčních listů po zezelenání, atd.) (Masarovičová, 2008). Různé odpovědi zprostředkované fytochromy v odlišných světelných podmínkách, zařazené do 3 skupin (VLIR, LIR a HIR), jsou možné na základě existence více druhů fytochromů (A až E). V tabulce jsou shrnuté základní charakteristiky jednotlivých typů odpovědí. (Masarovičová, 2008) 15
19 V rostlinách se vyskytují 2 typy fytochromů. Rozdíl mezi nimi není v chromoforu, ale v bílkovinném nosiči tedy stejný chromofor je vázán na jiný proteinový dimer (Gloser, 1998). Fytochrom I se vyskytuje zejména v etiolovaných rostlinách a má absorpční maximum při 666 nm. Po ozáření se tento typ fytochromu rychle rozkládá. Fytochrom II je ve větším množství zastoupen v zelených rostlinách (poměr fytochromu I a II je přibližně 1:1), má absorpční maximum při 654 nm a je na světle poměrně stálý (Procházka, 1998). V semenech je uložen převážně fytochrom II. Pokud klíčení semene a další růst semenáčku probíhá na světle, je syntetizován opět fytochrom II. Při klíčení ve tmě však kromě syntézy tohoto typu fytochromu dochází k syntéze fytochromu I, kterého se může vytvářet až stokrát větší množství než typu II. Po osvětlení semenáčků dojde k inaktivaci příslušného genu a syntéza fytochromu I se zastavuje. Ve všech zelených rostlinách tedy naprosto převažuje typ II (Gloser, 1998). Fytochrom dále reguluje řadu růstových procesů: zakládání a expanzi listů, zakládání kořenů a dlouživý růst stonku. Roste-li stonek ve tmě, internodia jsou dlouhá, listy dvouděložných rostlin zakrňují, u jednoděložných se protahují a stonek nakonec ztrácí schopnost vzpřímeně růst. Těmto změnám, zvaným etiolizace, lze zabránit krátkodobým ozářením R, jehož účinek je možné zrušit následným ozářením FR (Procházka, 1998). Mimo zmíněné růstové vlivy se fytochrom účastní v jednom z nejdůležitějších životních pochodů odečítání času, tj. měření doby tmy a doby ozáření. Tento proces je nezbytný pro reakci rostlin na střídání dne a noci i pro reakci rostlin na střídající se roční období. Fytochromy mají v zelených rostlinách aktivní fyziologickou úlohu. Přítomnost R/FR reverzibilních pigmentů ve všech zelených rostlinách od řas po dvouděložné druhy naznačuje, že světlo těchto vlnových délek umožňuje rostlinám přizpůsobit se prostředí. Poměr toku fotonů ve spektrálním pásu okolo 660 nm (R) ku pásu okolo 730 nm (FR) v různých prostředích je odlišný. V porovnání se světlem v průběhu dne narůstá podíl tmavočerveného světla při západu slunce Jeho větší podíl je i v mělkých vrstvách půdy a zejména ve stínu jiných rostlin, kde se uplatňuje baldachýnový efekt. Ten vyplývá ze skutečnosti, že zelené listy absorbují pomocí chlorofylů značně červené světlo, ale pro tmavočervené světlo mají velkou propustnost. Se zvětšováním zastínění klesá poměr R/FR a též klesá podíl Pfr ku celkovému fytochromu, neboť se více Pfr konvertuje na Pr. Zjistilo se, že vysoký podíl tmavočerveného světla, tj. nízký poměr Pfr ku celkovému fytochromu se projeví větší rychlostí prodlužování stonků 16
20 heliofytů (rostlin výslunných stanovišť). Tato odpověď se označuje jako únik ze stínu. Podmínky zastínění způsobují omezení růstu listů a zvýšení růstu stonku do výšky. Sciofyty, rostliny dědičně adaptované na podmínky růstu pod dřevinami, rostou stejně na světle jako ve stínu. Fytochromy I a II mají vzájemně antagonistické úlohy. Kontinuální červené světlo absorbuje Pr I, zvýší se hladina Pfr II, čímž se stimuluje deetiolizace. Kontinuální tmavočervené světlo absorbuje Pfr II, konvertuje se na Pr II, čímž se inhibuje deetiolizace. Stimulace deetiolizace fytochromem I závísí na fotostacionárním stavu. Kontinuální tmavočervené světlo stimuluje a kontinuální červené světlo inhibuje deetiolizaci. Na přímém slunečním světle, které je bohaté na červené vlnové délky, je zezelenání etiolovaných rostlin zprostředkované fytochromem II. V klíčících rostlinách ve stínu jiných rostlin (v podrostu je světlo obohacené tmavočervenými vlnovými délkami) se zezelenání iniciuje primárně fytochromem I, který je nestálý, a proto se uplatňuje jen na začátku vývoje. Po zezelenání fytochrom I vystřídá fytochrom II, který při převaze tmavočerveného světla zruší inhibici růstu stonku a odpovědí je únik ze stínu. V dospělých rostlinách převažuje fytochrom II, který při převažujícím tmavočerveném světle částečně potlačí zezelenání a odpovědí je únik ze stínu, spočívající ve stimulaci prodlužovacího růstu. Tato odpověď je rychlá a je též reverzibilní, jestliže se rostliny vrátí do světla bohatého na červenou složku (Masarovičová, 2008). Receptory modrého záření K odpovědím na modré světlo patří fototropismus, orientovaná odpověď podle směru dopadajícího záření, pohyb chloroplastu v buňce, inhibice prodlužování hypokotylu, zavírání průduchů apod. Odpovědi na modré světlo nejsou reverzibilní jako odpovědi na fytochrom. Akční spektra pro odpovědi stimulované modrým světlem mají tři vrcholy mezi v oblasti od 400 do 500 nm. V rostlinách je vícero fotoreceptorů modrého světla, které mají různé chromofory - karotenoidy, flaviny a pterin (Masarovičová, 2008). Kryptochromy jsou fotoreceptory pro oblast modrého světla s přesahem do oblasti záření UV-A a zprostředkují informace pro metabolické, růstové, vývojové a pohybové reakce, nezprostředkují však informace o pozici zdroje. (Pavlová, 2005). Účastní se modrým světlem indukované inhibice prodlužování hypokotylu, expanze klíčních listů, zvýšené produkce antokyanů, regulace kvetení a synchronizace cirkadiánních rytmů (Masarovičová, 2008). Zatím byly nalezeny dva kryptochromy, značené CRY1 a CRY2 (z angl. Cryptochrome). Jsou přítomny v rostlinách po celou ontogenezi ve všech částech rostliny ve všech živých buňkách na světle i ve tmě (Pavlová, 2005). 17
21 Fototropin je fotoreceptor, který absorbuje záření v modré oblasti světla a zprostředkovává informaci o pozici zdroje záření. Rostlina tuto informaci interpretuje jako směrovanou pohybovou reakci tropismus. Fototropin je protein vázaný na plazmatickou membránu. Po absorbci světla se vytvoří stabilní produkt, který již není další absorbce schopen. Schopnost absorbovat světelné záření se spontánně obnoví ve tmě (Pavlová, 2005). Cis-zeaxantin se považuje za receptor modrého záření, které vede k pohybům svěracích buněk průduchů. Tento karotenoid je součástí fotoochranného xantofylového cyklu. V uzavřených buňkách je přítomný i brzy ráno, kdy fotoochrana ještě není potřebná, což potvrzuje i jeho nepřítomnost v mezofylových buňkách. Odpověď průduchů na světelné záření je integrovaným projevem dvou odlišných fotoreceptorových systémů, první závisí na fotosyntéze a druhý je odpovědí na modré světlo. Akční spektrum odpovědi průduchů na modré světlo je zřetelně odlišné od akčního spektra fotosyntézy. (Masarovičová, 2008). Ani záření ani fotoreceptory však nejsou zdrojem vlastních morfogenetických informací, ale pouze informují o stavu prostředí (Gloser, 1998). Růstové procesy probíhají na základě realizace genetické informace, ale tento buněčný aparát musí být schopen zpracovávat informace z prostředí a reagovat na ně (Tůma, Tůmová, 1998). To znamená, že po přijetí a zpracování signálu musí být přenesen na buněčnou úroveň, ze které může být signál dále přenesen na úroveň morfogenetickou (Procházka, 1998). Na morfologických změnách se pak podílí především intenzita světla, spektrální složení a fotoperiodismus (Tůma, Tůmová, 1998). Mechanismus působení fytochromu a kryptochromu na fyziologické procesy není dokonale znám, což je pochopitelné vzhledem k rozsahu zjištěných účinků. Zatímco kryptochrom je pevně zabudován v plazmatické membráně, fytochrom se vyskytuje také v cytosolu a dokonce i v jádře. Je proto velmi pravděpodobné, že regulační mechanismy mohou být v tomto případě nejméně trojího druhu: - Změny permeability a elektrického potenciálu plazmatické membrány - Změny v tvorbě proteinů. Je známo již více než 60 enzymů, jejichž tvorba je aktivována fytochromem. - Změny v koncentraci fytohormonů. Nové výzkumy ukazují, že neaktivní forma fytochromu Pr je v buňkách vcelku rovnoměrně rozptýlena, ovšem po přeměně na aktivní Pfr dochází k jejímu shlukování, a to zejména v blízkosti plazmatické membrány. Přitom bývá současně pozorována aktivace vodíkových pump a zvýšený příjem draslíku do buňky. Dochází též ke zvýšení koncentrace iontů vápníku 18
22 v cytosolu (vázaných na obvyklý nosič, bílkovinu calmodulin). Vápník je schopen aktivovat celou řadu enzymů v buňce, a má tedy zřejmě významnou funkci druhotného mediátora u všech světlem řízených reakcí (Gloser, 1998). Procesy fotomorfogeneze mají komplexní povahu. V jejich regulaci kooperují různé receptory a mohou zahrnovat i všechny reakce fytochromu (tj. VLIR, LIR i HIR). Nemají univerzální charakter a u různých druhů lze předpokládat rozdílnou regulaci širší srovnávací studie však zatím chybí (Procházka, 1998). Obecně zahrnuje fotomorfogeneze všechny projevy růstu a vývoje závislé na viditelném záření. Většinou sem řadíme pouze jevy zprostředkované specializovanými receptory záření a především fytochromem (vynecháváme tedy fenomén fotomorfogeneze spojený přímo s fotosyntézou), jež jsou nejšířeji prostudovány. Patří sem deetiolizace, morfogenetické reakce na zastínění či vysokou ozářenost a klíčení závislé na záření (Procházka, 1998). Deetiolizace Ve tmě rostlina investuje svůj rezervní materiál do dlouživého růstu internodií při potlačení funkce fotosyntetického aparátu netvoří se chlorofyl, listy jsou zakrnělé. Mluvíme též o skotomorfogenezi. Jde o adaptaci k dosažení světla. Na světle fotomorfogeneze jde o investici do aktivace fotosyntetického aparátu syntézy chlorofylu a růstu listů - převládá tendence stát se urychleně autotrofním organismem. Přechod mezi skotomorfogenezí a fotomorfogenezí se nazývá deetiolizace. V regulaci deetiolizace je nezbytná účast fytochromového systému, často i se spoluúčastí receptoru modrého záření. Plastogeneze - Chloroplasty se vyvíjejí z proplastidů, z nichž však ve tmě vznikají etioplasty. Prekurzor chlorofylu, prochlorofylid, je obsažen v etioplastu, nikoliv však bílkoviny světlosběrného komplexu ani jejich mrna. Účinkem záření, které je přímo absorbováno prochlorofylidem (působí jako primární receptor), dochází k jeho redukci na chlorofylid. Fytochrom reguluje především rychlost akumulace chlorofylu a syntézu bílkovin světlosběrného komplexu (kódovány v jádře) fotosystému II. Kinetika jejich syntézy a syntézy chlorofylu jsou si podobny, což ukazuje na vysokou komplexnost a koordinovanost procesů plastogeneze. Pod kontrolou fytochromu je dále proteosyntéza struktur, na nichž probíhá fotolýza vody (OEC, oxygen evolving complex), syntéza feredoxinu a malé podjednotky Rubisco (Procházka, 1998). Skoto a morfogeneze orgánů - U klíčních rostlinek dvouděložných druhů je jednou z prvních nápadných reakcí na světlo narovnání hypokotylu (i epikotylu). Tento ohyb je 19
23 pravděpodobně vyvoláván působením ethylenu, jehož syntéza je na světle zastavena. Působením světla se dále zrychluje růst plochy listů i délka řapíku. Velmi rychle, často během několika hodin, dochází k tvorbě chlorofylu a plně funkčních chloroplastů z proplastidů, tedy včetně řádně fungujících tylakoidních membrán a enzymů nutných pro zabezpečení asimilačních procesů (Gloser, 1998). Téměř všechny projevy deetiolizace jsou zprostředkovány fytochromem a receptorem modrého záření, ať již účinkují samostatně či spolupůsobí (Procházka, 1998). Světlem je zpomalována rychlost růstu stonku a jiných nadzemních částí. Na této inhibici se podílí oba systémy fotoreceptorů (fytochrom i kryptochrom). Modré světlo mívá často rychlejší a výraznější účinek (Gloser, 1998). Fotomorfogeneze na stanovišti Značná část fotomorfogenetických projevů rostliny je spojena s informací o poměru R/FR. Zvýšený podíl FR (nižší intenzita záření) charakterizuje svítání a soumrak. Rozsah tohoto podílu v podmínkách existence suchozemských rostlin kolísá v rozmezí od 0,05 (pod hustě zastíněným porostem) do 1,2 při plném slunečním záření nad porostem. Podíl modré složky se za svítání a za soumraku relativně zvyšuje. V proměnlivých podmínkách přirozených stanovišť je velmi obtížné odlišit fotomorfogenetické efekty závislé na rychlosti fotosyntézy od těch, jež jsou regulovány fytochromem a dalšími receptory. Příkladem morfogenetických efektů spojených s rychlostí fotosyntézy jsou např. změny morfologie a anatomické stavby listů v závislosti na intenzitě ozáření. Listy při vysoké ozářenosti mají obvykle menší plochu, jsou silnější, členitější, s delšími buňkami palisádového parenchymu či s větším počtem jeho vrstev. Jiným obecným rysem je prodlužování internodií při stoupajícím podílu FR, k němuž dochází jak při zastínění v porostu, tak jako efekt stmívání. V podstatě jde o snížení či anulování inhibice dlouživého růstu vyvolaného červenou složkou viditelného záření. Ta je spojena s vysokou hladinou Pfr. Zvýšený podíl FR vyvolává i inhibici větvení v paždí listů dolní části lodyhy a zvýšené ukládání asimilátů v nadzemních částech místo v kořenech. Efekt stmívání stimuluje i růst listů. Spolehlivost uváděného signalizačního a informačního systému tedy závisí především na kvalitě záření, a nikoli na ozářenosti, která více podléhá okamžitým změnám počasí. Komplexně se výše uváděné i další fotomorfogenetické projevy uplatňují ve dvou alternativních strategiích rostlin: v toleranci k zastínění a v úniku před zastíněním, mezi nimiž 20
24 existují i přechodné typy. V prvém případě při pomalém růstu a delší vegetační periodě je charakteristická vysoká účinnost fotosyntézy i při nižších intenzitách FAR, zvětšování listové plochy a poměrná necitlivost pro navození dlouživého růstu snižujícím se poměrem R/FR. Tato růstová reakce naopak odlišuje rostliny ze stínu unikající (Procházka, 1998). Podle požadavků na intenzitu záření rozdělujeme rostliny na: Světlomilné heliofyty, např. rostliny pouští, stepí, tunder a hor (Tůma, Tůmová, 1998). Tyto rostliny využívají lépe červené záření (Hejnák, 2007). Stínomilné sciofyty, které jsou přizpůsobené rozptýlenému světlu a využívají lépe paprsky modrofialové (Hejnák, 2007). Heliosciofyty rostliny tolerantní ke stoprocentnímu ozáření, ale snášejí i zastínění různého stupně (Tůma, Tůmová, 1998). Masarovičová (2008) a Faustus, Kincl (1978) uvádějí dělení rostlin na heliofyty a sciofyty, stejně jako Hejnák (2007). Procházka rostliny rozděluje rostliny ze stínu unikající a rostliny, které stín tolerují a uvádí, že je mezi nimi několik přechodných typů Fotoperiodické účinky světla Fotoperiodismus je metabolická, růstová či vývojová odezva rostlin na délku dne, tj. na dobu denního ozáření. Fotoperiodické reakce rostlin zahrnují schopnost rozlišovat mezi zářením a tmou, měřit čas a reagovat na změny doby ozáření morfogenetickými změnami. Doba, po kterou jsou rostliny vystaveny působení záření ve 24hodinovém cyklu, se nazývá fotoperioda a odtud i termín fotoperiodismus. Fotoperioda je nejspolehlivějším ekologickým signálem informujícím rostliny o ročním období a jako taková určuje u mnoha rostlinných druhů rozhodující trvání vegetativní fáze vývoje a řídí přechod do fáze reproduktivní. Kvetení není však jediným procesem ovlivněným fotoperiodou. Délka dne působí i na řadu dalších růstových i vývojových procesů. Internodia mnohých rostlin jsou velmi často kratší během krátkého dne (fotoperioda kratší než přibližně 10 hodin), některé rostliny s požadavkem jarovizace či rostliny dlouhodenní vytvářejí za krátkého dne přízemní listovou růžici. Také tvorba hlíz je většinou podporována krátkodenním režimem. Krátký den navozuje i procesy související se stárnutím, zráním a počátkem dormance. Fotoperiodická reakce rostlin je ovlivněna teplotou a stářím rostlin. Teplota nejen ovlivňuje délku fotoperiody nutnou k indukci kvetení, ale může plně změnit typ rostliny z krátkodenní na dlouhodenní a naopak. Rostliny mohou též měnit svou fotoperiodickou citlivost se stářím. 21
25 U většiny rostlin existuje kratší či delší období, kdy nejsou schopny reagovat na fotoperiodickou indukci kvetení. Toto období nazýváme juvenilita (Procházka, 1998). Pro fotoperiodickou reakci (odpověď) je rozhodující fotoperioda kritická. Kritická fotoperioda je fotoperioda mezní, hraniční, která navodí vývojovou změnu. Hodnota kritické fotoperiody se vztahuje vždy jen k určité vývojové změně, např. navození kvetení a k určitému druhu rostliny nebo nižší jednotce, např. populaci, kterou lze označit za ekotyp. Označení dlouhý nebo krátký den se vždy vztahuje jen ke kritické fotoperiodě. Krátkodenní reakce nastane, je-li délka aktuální fotoperiody kratší než kritická. Dlouhodenní reakce je navozena fotoperiodou delší, než je fotoperioda kritická. Fotoperiodismus byl objeven při studiu kvetení a charakteristika, tj. rostlina krátkodenní nebo dlouhodenní se vztahuje ke kvetení. Pokud se jedná o jinou vývojovou změnu, než je kvetení, je nutno ji definovat (Pavlová, 2005). Jen málo druhů zakládá květy zcela bez jakékoli vazby na délku fotoperiody (fotoperiodicky neutrální rostliny). Častěji se setkáváme s nároky na jistou délku dne, a podle toho rostliny rozdělujeme na dlouhodenní a krátkodenní (Gloser, 1998). Podle nároků na určitou fotoperiodu se rostliny dělí do několika skupin: 1) Rostliny fotoperiodicky citlivé a) Dlouhodenní vyžadují den delší než hodin a vykvétají i za nepřetržitého osvětlení (Tůma, Tůmová, 1998). Do této skupiny patří například Aechmea fasciata, Begonia tuberhybrida, Bryophyllum a další (Machovec, 1975). Dále také Achimenes, Calceolaria, Campanulla isophylla, Fuchsia, Stephanotis (Volf, Votruba, 1991) b) Krátkodenní osvětlení nesmí být zpravidla delší než hodin a na nepřetržitém světle nevykvetou, kvetou většinou za krátkého jarního nebo podzimního dne (Tůma, Tůmová, 1998). Příkladem této skupiny jsou Begonia elatior, Begonia x Gloire de Lorraine, Bougainvillea spectabilis, Camelia japonica, Coleus hybridus, Euphorbia fulgens, Euphorbia pulcherrima, Euphorbia milii (splendens), Kalanchoe blossfeldiana, Cattleya a Zygocactus (Machovec, 1975). c) Dlouhokrátkodenní vykvetou jen tehdy, když jsou po pěstování na dlouhém dni vystaveny určitému počtu krátkých dní d) Krátkodlouhodenní vyžadují zprvu krátké a potom dlouhé dny e) Intermediární vyžadují zvláštní periody, např. den 8 i 18-ti denní, tyto rostliny jsou poměrně vzácné. Tento typ rostlin uvádějí pouze autoři Podešva, Véber, Kubín (1968) 22
26 a Tůma, Tůmová (1998) jiní autoři, z jejichž děl byly použity informace, se o tomto typu rostlin nezmiňují. U všech fotoperiodicky citlivých rostlin může mít požadavek na fotoperiodu charakter: - Obligátní bez požadované induktivní délky dne vůbec netvoří květy - Fakultativní nakonec vykvetou v jakékoliv délce dne, nevhodná délka pouze jejich vývoj zpomaluje. Rovněž bylo dokázáno, že pro krátkodenní není důležitá délka světelné části dne, ale musí být přesně dodržena délka noci, která nesmí být přerušena ani krátkým osvětlením (obsahujícím červenou část spektra). U dlouhodenních rostlin naopak přerušení temné periody stimuluje kvetení rostlin 2) Rostliny fotoperiodicky neutrální kvetou za libovolné fotoperiody, ale začínají kvést až když mají vytvořen určitý počet listů (Tůma, Tůmová, 1998). Do této skupiny patří Aphelandra, Brovalia speciosa, Hydrangea, Pelargonium zonale, Pelargonium peltatum, Primula malacoides, Rosa, Sinningia hybrida, Zantendeschia, aj. (Machovec, 1975). Do řízení fotoperiodických reakcí je zapojen fytochromový systém (Tůma, Tůmová, 1998). Informaci o světelném signálu zprostředkují především fytochromy, u dlouhodenních rostlin se významněji podílí i kryptochrom. Tyto senzory předávají informaci mechanismu, který u rostlin měří čas vnitřnímu oscilátoru. Podstata měření času není zatím uspokojivě objasněna. Jisté je, že měření času je nezávislé na teplotě a že u rostlin senzory nejsou integrálními součástmi časoměrného mechanismu. Nevysvětlené je zatím také porovnání aktuální fotoperiody s fotoperiodou kritickou. Je však jisté, že významnou úlohu v tomto procesu hraje endogenní rytmicita rostlin. Délka fotoperiody je registrována v listech. List je fotoperiodicky citlivý jen v určité fázi svého vývoje. Fotoperiodický signál vyvolá v listu tvorbu fotoperiodického květního stimulu, který je z listů transportován do ostatních částí rostliny. V meristémech fotoperiodický signál vyvolá stav označovaný jako evokace, jehož výsledkem je tvorba květních částí. Listy velmi mladé nebo senescentní funkci fotoperiodického receptoru neplní. Není známo, zda nejsou schopny přijmout fotoperiodický signál nebo vytvořit fotoperiodický květní stimulus. Příjem fotoperiodického signálu, tvorba fotoperiodického stimulu a jeho transport do meristému jsou procesy, které se souborně nazývají fotoperiodická květní indukce. Fotoperiodický květní stimulus, jeho charakteristika a identifikace, byly po mnoho let 23
27 objektem intenzivní experimentální práce, nebyl však dosud identifikován. Jeho existence je doložena množstvím experimentů, v nichž byl přenesen roubováním listu z rostliny indukované ke kvetení na rostlinu neindukovanou, u níž kvetení vyvolal. Přenos fotoperiodického květního stimulu byl opakovaně ověřován a možnost jeho přenosu nejen mezi rostlinami stejného druhu, rostlinami různých druhů i mezi rostlinami různé fotoperiodické reakce byla doložena různými osobami na několika pracovištích (Pavlová, 2005). Příkladem je tzv. Čajlachjanova florigenová teorie - hypotetický květní hormon florigen, jehož množství závisí na počtu indukovaných period (Tůma, Tůmová, 1998). Později se předpokládalo, že florigen je směs dvou látek (anthezinu a giberilinu) nebo látek několika, nebyl vyloučen ani jeho fyzikální charakter (Pavlová, 2005). 3.2 Světlo a ostatní pěstební faktory Světlo a teplota V přírodě je hlavním zdrojem tepla, právě tak jako světla, sluneční záření. Životní pochody rostlin, tj. především asimilace a disimilace, narůstají a klesají vlivem teplot (Machovec, 1975). Rychlost růstu je určována množstvím energeticky bohatých látek, které se vytvoří při fotosyntéze. Pro fotosyntézu není potřebná vysoká teplota a může probíhat v širokém rozmezí teplot. U pokojových rostlin je v zimě omezována nedostatkem světla, nikoliv nedostatkem tepla. Fotosyntéza se zpomaluje, až zastavuje při určitých spodních a horních mezních hodnotách teploty, které jsou u jednotlivých druhů rostlin rozdílné. Teplomilné rostliny přestávají asimilovat při teplotě kolem 10 C. Při nedostatku světla v zimě vyšší teplota než 18 C nezvyšuje fotosyntetickou činnost a další zvyšování teploty v závislosti na světle nemá pro fotosyntézu význam (Volf, Votruba, 1991). U některých druhů rostlin probíhají v některých případech vlivem teplot určité změny ve fotoperiodické reakci. Například Zygocactus přestože je rostlinou krátkého dne, kvete při teplotách nižších než 17 C i za dlouhého dne. Podobně je to i u Begonia Gloire de Loraine, která je při teplotách vyšších než C rostlinou vysloveně krátkodenní, a která navzdory tomu kvete při nižších teplotách i za dlouhého dne (Machovec, 1975). Volf a Votruba uvádějí jako další příklad se stejnou reakcí krátkodenní Poinsetii (Euphorbia pulcherima). Teplota ovlivňuje také prodlužovací růst prodlužování buněk a internodií. Zvyšováním teploty při nedostatku světla se posiluje prodlužovací růst a to má za následek nepevnost 24
28 pletiv a stonků. Stavba rostlin není kompaktní a celkově se zhoršuje kvalita květin (Volf, Votruba, 1991). Proto podle množství světla, a to hlavně v zimě, upravujeme i tepelné poměry. Tam, kde jde o světlomilné rostliny, teploty raději snižujeme o 2-5 C v porovnání s normálními požadavky a rostliny udržujeme v sušším prostředí; v opačném případě se začnou vytahovat. U stínomilných rostlin nemusíme být tak úzkostliví (Machovec, 1975) Světlo a vlhkost prostředí Sluneční záření s narůstajícím množstvím energie zesiluje vypařování vody z rostlin a substrátu (Volf, Votruba, 1991). Všeobecně platí, že s ubýváním teploty a intenzity světla musí zákonitě ubývat i přísun vody a živin. Naopak při zvyšování teploty a intenzity světla je nutno dodávat větší dávky vody a výživy všem pěstovaným květinám. Tato zákonitost vyplývá ze všech životních pochodů a látkové výměny rostlin. Jakmile má rostlina všechny životní faktory v dostatečném množství, probíhají intenzivněji i fotosyntetické pochody. V opačném případě se všechny životní jevy omezují na minimum a rostlina se dostává do stavu vegetačního klidu. Z toho je jasné, že na podzim, v zimě a v předjaří je nutné zalévat mnohem opatrněji než na jaře, v létě a začátkem podzimu. Každé pravidlo má své výjimky a ani u zálivky to není jiné. Především je třeba se řídit teplotou a intenzitou výparu z rostlin. To znamená, že např. v létě, v období déletrvajících dešťů a za oblačného počasí, když velmi klesne teplota a sníží se i světelnost, budeme zalévat málo nebo méně; v tomto případě budeme dávat pozor, abychom rostliny v odpoledních hodinách již nenamáčeli. Naopak v zimě, v interiérech s ústředním topením a s teplotou nad 20 C budeme zalévat vydatněji tehdy, když mají alespoň minimální příkon světelné intenzity. Uvedené závislosti platí i pro kaktusy a sukulenty, u kterých v zimě, kdy je v našich podmínkách úplný nedostatek světla, je nutno dodržovat sníženou teplotu a méně zalévat. Nedostatek světla se u těchto rostlin nemůže nahradit ani teplotou, ani vodou, ani živinami (Machovec, 1975) Vzájemné vztahy mezi vlhkostí, teplotou a intenzitou světla Všechny výše uvedené faktory se jednak vzájemně podmiňují, jednak se v závislosti na sobě mění. Nelze si dost dobře představit sluneční záření, tedy určitý světelný požitek určité klimatické oblasti bez tomu odpovídající teploty, která zase ovlivňuje výpar a srážkové poměry, tedy i vlhkost prostředí. Uvedené faktory jsou vzájemně nezastupitelné, i když je možno na určitém stupni částečné nedostatky některých faktorů nahradit zčásti jiným faktorem. Příkladem je rozkvétání krátkodenních rostlin Zygocactus a dalších. Optimalizací jednoho faktoru lze vyvolat maximální účinek faktoru druhého, např. při optimální teplotě 25
29 probíhá asimilace v plné míře podle daného světelného požitku. Jakmile není teplota v optimu, nemůže se tento světelný požitek plně využít, ale zároveň může optimální teplota při nízké vzdušné vlhkosti rostliny poškozovat. Při volbě vhodných relací jednotlivých faktorů se řídíme analogií s podmínkami původního prostředí. Nesprávné korelační vztahy mezi jednotlivými faktory můžeme pozorovat přímo na rostlinách. Např. příliš dlouhý a slabý růst a malé listy prozrazují, že rostlina má nadbytek tepla, ale nedostatek světla. Naopak nadbytek světla, hlavně v oblasti ultrafialového záření a současný nedostatek tepla je příčinou zakrslého růžicovitého vzrůstu a velmi často i popálení mladých listů. Poměrně nejméně přímých korelačních vztahů je mezi světlem a vlhkostí prostředí. Světelné poměry jsou však úzce závislé na teplotě, což platí i o vlhkostních poměrech, vidíme, že nepřímých závislostí je tu mnoho. V přírodě vegetuje většina stínomilných rostlin zpravidla za zvýšené vlhkosti, jako např. kapradiny, plavuně, některé begónie, apod. Naproti tomu značné množství rostlin velmi náročných na světlo vegetuje v suchých podmínkách, např. kaktusy, sukulenty, většina cibulovin, apod. Aby se rostliny úspěšně vyvíjely, musí být všechny jmenované faktory ve vzájemné harmonii a vyrovnanosti. Pokud chceme, aby se dodržela rovnoměrnost vývoje, je nevyhnutelné, aby při poklesu jednoho z faktorů poklesly také oba dva další. Při zkracování délky dne a snižování světelnosti je třeba na podzim současně snižovat teplotu, relativní vzdušnou vlhkost a zálivku. Pokud tuto zásadu nedodržíme, budou se rostliny buď vytahovat, nebo hnít. Naopak se zvyšováním světelnosti brzo na jaře a v létě se současně zvyšuje teplota. U rostlin náročných na vlhkost prostředí je třeba úměrně zvyšovat intenzitu zalévání a postřikování. Zálivka a postřik jsou většinou ze všech uvedených faktorů nejlacinější, i když vyžadují největší péči. Teplota v interiéru je již předem dána (neudržuje se tu kvůli rostlinám, ale kvůli lidem, kteří zde žijí a k jakému účelu daný interiér slouží) a s ohledem na květiny ji lze měnit jen nepatrně, např. dodatečným vytápěním tropických skříní, apod. Získávání světla je v porovnání s ostatními faktory nejnákladnější složkou (Machovec, 1975). 3.3 Světelně technické jednotky Na základě mezinárodní dohody o sjednocení technických jednotek byla v roce 1974 vydána mezinárodní soustava jednotek SI. Tvoří ji: 26
30 - základní jednotka SI (termín, jednotka, její zkratka) - svítivost kandela (cd) - odvozená jednotka SI (její vyjádření pomocí základních jednotek) - jas kandela na čtverečný metr cd*m -2 - odvozené jednotky se zvláštním pojmenováním světelný tok lumen lm cd*sr -1 ; osvětlení lux lx lm*m -2 m -2 *cd*sr -1 (Chalupský, 1981) Svítivost Svítivost je definována jako fotometrická veličina, která vyjadřuje schopnost světelného zdroje vysílat v určitém směru do daného elementárního prostorového úhlu a v elementární časové jednotce energii, která vyvolá zrakový vjem (Haš, 2011). Svítivost světelného zdroje není ve všech směrech stejná. Světlo vycházející ze světelného zdroje se rozděluje do prostoru nerovnoměrně. Rozdělení svítivosti do různých směrů se zobrazuje křivkami svítivosti. (Büscher, 1964) Světelný tok Světelná energie je vyzařována v malých světelných kvantech fotonech, jejichž součet dává světelný tok. Každý foton představuje určité množství energie, které se mění podle délky světelné vlny, odpovídající určité barvě světla (Podešva, Véber, Kubín 1968). Jeden lumen je definován jako světelný tok vyzařovaný zdrojem o svítivosti 1 cd do jednoho steradiánu, tj. do jednotkového prostorového úhlu, který na povrchu koule o poloměru r vytíná plochu r 2 (Kubín, 1973). Světelný tok je důležitá veličina určující kvantitativně zdroj nebo svítidlo; je to tzv. účinnost - výkon (Chalupský 1981) Osvětlení Osvětlení je definováno jako světelný tok dopadající na jednotkovou plochu. Osvětlení rovné plochy je poměr světelného toku a její velikosti. Jednotkou osvětlení je jeden lux, tj. osvětlení plochy o velikosti 1 m 2 světelným tokem jednoho lumenu, čili kolmé osvětlení zdrojem o svítivosti 1 cd ze vzdálenosti 1 m (Kubín 1973). Když se vzdálenost bodového světelného zdroje od osvětlované plochy zvětší, intenzita osvětlení rychle klesá, a to se čtvercem vzdálenosti (Büscher 1961) Emisní spektrum Rozdělení zářivého toku, vysílaného světelným zdrojem, ukazuje emisní spektrum. Je to zobrazení zářivého toku tak, jak je rozdělen na jednotlivé vlnové délky v celé oblasti 27
31 viditelného záření, případně i v širším rozsahu optického záření (ultrafialové, infračervené záření). Emisní spektrum zásadně určuje kvalitativní vlastnosti světla, jeho teplotu chromatičnosti, index podání barev, vhodnost pro fotochemické nebo fotobiologické procesy (Haš, 2011). Emisní spektrum světelných zdrojů může být jednak spojité, jednak nespojité (diskontinuální, čárové). Spojité spektrum záření mají např. luminofory zářivek a vysokotlakých halogenidových výbojek (Podešva, Véber, Kubín, 1968). Vysílají jej také zdroje LED, jak uvádí Haš (2011). Diskontinuální spektrum se skládá jen z paprsků určitých vlnových délek, které se po rozkladu monochromátorem jeví jako samostatné čáry nebo pásy, oddělené většími nebo menšími mezerami. Mezery pak odpovídají vlnovým délkám, které ve spektru daného zdroje chybějí. Diskontinuální (čárové) spektrum je charakteristické pro elektrický výboj v plynech nebo parách kovů (Podešva, Véber, Kubín, 1968). Z hlediska účinku na rostliny je sice lhostejné, zda toto záření pochází z oboru spektra spojitého nebo nespojitého. To ovšem neznamená, že záření dané vlnové délky musí vyvolat stejný účinek, působí-li samostatně nebo působí-li současně ještě s paprsky jiných vlnových délek, a to i tehdy, je-li celkové množství energie dopadající na ozařovanou rostlinu v obou případech stejné (Kubín, 1973) Teplota chromatičnosti Některé umělé světelné zdroje, především žárovky a zářivky, bývají často charakterizovány tzv. teplotou chromatičnosti (teplotou barvy). Teplota chromatičnosti světelného zdroje je teplota, kterou by muselo mít absolutně černé těleso, aby jím vysílané záření způsobilo v normálním lidském oku při téže hustotě vyzařování stejný barevný dojem jako daný světelný zdroj. Údaj o teplotě chromatičnosti umělého světelného zdroje nestačí tedy k objektivnímu zhodnocení jeho spektrálních vlastností. Může být jen vodítkem pro přibližný odhad vzájemného poměru množství krátkovlnných a dlouhovlnných paprsků (Kubín, 1973). Světelné zdroje se z tohoto hlediska rozdělují do 3 skupin: Zdroje s teplotou chromatičnosti do 3300 K se označují jako zdroje teple bílé (warm white, ww). Rozsah K zahrnuje studené bílé zdroje (cool white, cw). Pro zdroje s teplotou chromatičnosti vyšší než 5000 K se používá název denní světlo (day light, d). Uvedené značení a jeho rozsahy nejsou normativní, někdy se mezi teplou bílou a studenou bílou vkládá ještě bílá teplota chromatičnosti K (Haš, 2011). 28
32 3.3.6 Index podání barev Teplota chromatičnosti může vyjádřit barvu světla jen u zdrojů se spojitým spektrem. Výbojové zdroje ale mají čárová nebo smíšená spektra, která neobsahují všechny barvy a barevné odstíny jako černé těleso. Proto se pro hodnocení barvy světla zavedl pojem index podání barev (R a ), který udává, jak věrně se jeví lidskému oku barvy osvětlovaného předmětu. Jeho nejvyšší hodnopta je 100. Podle indexu R a se zdroje rozdělují prakticky do 3 skupin: R a 90 až 100 jsou zdroje s vynikajícím podáním barev, R a 80 až 90 má dobré podání barev, R a menší než 80 má špatné podání barev (Haš, 2011) Barva světla Při označování světelných zdrojů se obvykle slučuje v jeden kód teplota chromatičnosti a index podání barev. Tento kód je označován jako barva světla. Je to trojmístné číslo. Jeho první číslicí je první číslice indexu podání barev. Druhé dvě číslice tvoří první dvě číslice teploty chromatičnosti. Např. barva 840 znamená, že světelný zdroj má index podání barev mezi hodnotami 80 a 90 a teplotu chromatičnosti 4000 K (Haš, 2011) Jednotky pro fotosynteticky aktivní záření Viditelné záření z hlediska jiných účinků než vidění člověka má pro zářivý tok jednotku watt (W), pro intenzitu ozáření W*m -2, pro dávku ozáření (to je působení záření po určitou dobu), jednotku W*s*m -2 (= J*m -2 ). Základní jednotky pro fotosyntetické ozařování (v rozsahu nm) jsou potom pro zářivý tok W FAR, pro intenzitu ozáření W*m -2 FAR. McCree, který studoval šíření a působení viditelného záření v rostlinách, zjistil u řady rostlin spektrální odraznost, spektrální propustnost a tím také absorbční spektra (poměr absorbovaného záření k celkovému dopadajícímu záření pro různé vlnové délky). Při zjišťování vztahu mezi intenzitou fotosyntézy a absorbovaným zářením dospěl k poznatku, že množství asimilátu je v lepší korelaci s tokem fotonů než se zářivým tokem. Energii v 1 molu kvant pro jednotlivé vlnové délky znázorňuje Obrázek 5 v příloze I. Energie fotonu je závislá na vlnové délce, podle vztahu: 19,878 e λ = (J). λ nm 29
33 Tato energie sdílená prostředím za 1 sekundu je zářivý tok 1 fotonu (W). Změříme-li vhodným přístrojem celkový zářivý tok světelného zdroje a naměřenou hodnotu podělíme zářivým tokem 1 fotonu, známe fotonový tok (množství fotonů za sekundu, rozměr je 1/s, rozměrová jednotka je souvztažná s energetickými jednotkami). Biologové při studiu vztahů mezi rychlostí fotosyntézy a dopadajícím zářením sledují energii aktivizující látkové množství 1μmolu. Nobel uvádí, že pro aktivaci jedné molekuly CO 2 je zapotřebí 8-10 fotonů. Jednotka pro hodnocení toku fotonů ale předpokládá, že počet fotonů, které aktivují jeden μmol látky, je rovný počtu molekul v 1μmolu, který určuje Avogadrova konstanta ( L = 6, /kmol). Celková energie aktivující množství látky 1 μmol se za tohoto předpokladu rovná hodnotě 700 E μm = 119,4 (1/λ) dλ (J/ μmol) 400 Podělíme-li nyní naměřený zářivý tok hodnotou energie fotonů pro 1 μmol (vyplývající z tohoto vzorce), dostaneme nikoliv počet fotonů, ale veličinu s rozměrem látkového množství (μmol*s -1, případně μmol*s -1 *m -2 ). Pak měříme energii hodnotami látkového množství, tedy nesouvztažnou jednotkou. Taková jednotka nemůže být sice schválena jako měrová jednotka žádnou mezinárodní institucí pro míry a váhy, ale s tichou dohodou je používána jako účelová jednotka. Tuto jednotku vzali v úvahu i výrobci měřicích přístrojů a používají ji i výrobci světelných zdrojů, které doporučují k asimilačnímu ozařování rostlin. Lze ji samozřejmě používat jen k účelu, pro který byla stanovena, tedy pro hodnocení fotosyntetických procesů ve spektrálním rozsahu nm. V literatuře se však často používá i pro procesy vyvolané zářením jiných vlnových délek, pro které není tato jednotka definována. Pro procesy, kde viditelné záření přivádí pouze impulzy pro rostlinné sensory, případně i ve spektrální oblasti mimo nm, a nezúčastní se přeměn hmoty a energie, se používají jen energetické jednotky - W, W*m -2 (Haš, 2011). 3.4 Interiéry podle světelných podmínek Na základě převládajících světelných podmínek v interiéru je možno je rozdělit na: - nadměrně osvětlené interiéry celoročně velmi dobře až nadměrně osvětlené, často orientované na jih, jihozápad či celoplošně prosklené. 30
34 - světlé interiéry s dobrými světelnými podmínkami, u kterých nehrozí silné přímé sluneční záření ani nedostatek světla. Svými podmínkami vyhovují většině pěstovaných rostlin. - tmavé nedostatečně osvětlené interiéry. Intenzita světla se v nich často pohybuje na hranici možností pěstování rostlin, někdy pouze kolem Lx (Kuťková, 2006) Interiéry s nedostatkem světla Světlo je jedním z limitujících faktorů úspěšného pěstování rostlin v interiérech. V našich zeměpisných šířkách se světelné podmínky výrazně zhoršují v zimním období. Je to spojeno s krátkým dnem v zimě, který je navíc mnohdy doprovázen zamračenou oblohou. Průměrné denní množství globálního záření je v prosinci asi desetkrát menší než v červnu a pětkrát menší než v březnu viz Obrázky 5,6,7 v Příloze I (Volf, Votruba, 1991). Nedostatek světla pro fotosyntézu rostlin v zimním období je většinou v současných interiérech spojen s vysokými teplotami. Tento nepoměr mezi světlem a teplotou, často doprovázený nízkou vlhkostí vzduchu, z důvodu vytápění interiérů ústředním topením, může způsobit řadu pěstitelských problémů. Nedostatkem světla mohou trpět interiéry staršího bytového fondu s malými okny, postavené na konci 19. nebo na začátku 20. století, pasáže, spojovací chodby a koridory ve školách, obchodních centrech, místnosti situované k severu a podobně (Kuťková, 2006). Brookes (1995) používá termín stinné stanoviště, které definuje jako stanoviště, které nedostává ani přímé ani nepřímé sluneční světlo, světelné poměry však nejsou nepříznivé. Stanoviště označené jako stinné dostává přibližně 25 % množství světla slunečného stanoviště. Podle autora tuto kvalitu světla najdete těsně u světlých oken, situovaných na sever i v stinné části slunečné místnosti, například u bočních stěn, kde sice rostliny nestojí v dosahu slunce, ale ani nejsou od slunečného okna vzdálené více než 1,5 až 2 m. Ale i stínomilné tropické rostliny je třeba na těchto stanovištích v zimních měsících přisunout blíže ke zdroji světla, protože dny v tropických oblastech jsou mnohem delší než na severní polokouli. I když je možné nepříznivý vliv nedostatku světla určitými vzájemnými korelacemi pěstitelských faktorů zmírnit, je nemožné ho úplně vyloučit. Pro asimilační reakci je třeba počítat přinejmenším s hodnotami, které se řádově pohybují ve stovkách luxů, kdežto pro fotoperiodickou reakci postačí hodnoty, pohybující se v rozmezí jednotek až několika málo desítek luxů. Astronomická délka dne tedy není identická s délkou fotoperiodicky účinného 31
35 dne, protože ještě i za svítání a za soumraku dosahuje intenzita osvětlení takových hodnot, které pro fotoperiodickou reakci úplně postačí (Machovec, 1975) Možnosti řešení nedostatečného osvětlení v interiérech Rostlina může růst a zdárně se vyvíjet jedině tehdy, má-li zachycené záření vhodné spektrální složení, dopadá-li toto záření na rostlinu v potřebném množství a je-li jeho příkon přiměřeně velký. Chceme-li vyhovět všem těmto požadavkům, musíme při výběru vhodného světelného zdroje uvážit spektrální rozložení vyzařované energie, přihlížet ke geometrickým vlastnostem zdroje a k úhrnnému množství fotosynteticky účinného záření, vysílaného za jednotku času. Bodové zdroje (žárovky, kompaktní zářivky, halogenidové výbojky, apod.) se vesměs vyznačují velkou hustotou vyzařování a můžeme s nimi tedy dosahovat velkých hustot ozáření, avšak v jejich těsné blízkosti, tj. na poměrně malé ploše. Záření bodových zdrojů se totiž řídí podle tzv. čtvercového zákona. Podle tohoto zákona hustoty zářivého toku ubývá se vzdáleností od bodového zdroje zpočátku velmi rychle a s rostoucím odstupem ozařované plochy stále pomaleji. K silnému ozáření velké plochy musíme tedy nutně použít větší počet bodových zdrojů, avšak lze jen obtížně dosáhnout toho, aby ozáření plochy bylo naprosto rovnoměrné. Plošné zdroje (lineární zářivky, lineární LED zdroje, atd.) mají sice poměrně malou hustotu vyzařování, zato však umožňují dosáhnout velmi homogenního ozáření značně velkých ploch, zejména použijeme-li jich více ve vhodném uspořádání. U plošných zdrojů neplatí v jejich nejbližším okolí čtvercový zákon a hustoty jejich zářivého toku ubývá v závislosti na vzdálenosti ozařovaného předmětu mnohem pomaleji než u zdrojů bodových (Podešva, Véber, Kubín, 1968). Pro použití umělého světla (záření) při pěstování rostlin má zvláštní význam komplexní povaha viditelného světla. Všechny zkoušené zdroje umělého světla, vhodné pro praktické použití, mají jiné spektrální složení a jinou intenzitu než přirozené světlo, které během dne a v jednotlivých fázích růstu a vývoje dopadá na rostliny. I takové zdroje umělého světla, které se svým spektrálním složením nejvíce přibližují přirozenému dennímu světlu, například zářivky s označením denní světlo jsou, přinejmenším aspoň v intenzitě záření, hluboko pod fyziologickým účinkem denního světla. Jiné zdroje umělého světla, např. xenonové vysokonapěťové lampy, ještě bližší barvou a intenzitou světla přirozenému dennímu světlu jsou zase provozně nákladné a k trvalejšímu osvětlování rostlin ekonomicky neúnosné. Pokud jsou umělé zdroje záření používány pouze jako doplňkové k přirozenému dennímu světlu používáme termín přisvětlování rostlin. Pokud jsou rostliny podrobeny výlučně vlivu 32
36 umělých zdrojů záření bez spolupůsobení světla venkovního, přirozeného, používá se termín ozařování rostlin (Podešva, Véber, Kubín, 1968) Umělé zdroje záření pro přisvětlování rostlin Zdroje luminiscenční Luminiscence je obecně definována jako přebytek záření daného tělesa vzhledem k jeho absolutní teplotě, má-li tento přebytek konečné trvání. Luminiscenční záření nijak nesouvisí s absolutní teplotou tělesa (jako např. u žárovek) vzniká přeměnou jiné formy energie než energie tepelné. Mechanismus luminiscence úzce souvisí a atomovou a molekulární stavbou zářící látky. V osvětlovací technice nabyla dosud praktického významu jen luminiscence plynů a par vyvolaná elektrickým výbojem a fotoluminiscence a elektroluminiscence některých pevných anorganických luminoforů, tzv. krystalofosforů (Kubín, 1973). Zdroje výbojové sodíkové výbojky Velká světelná účinnost je dána tím, že asi 85 % viditelného záření připadá na dvojitou spektrální čáru o vlnových délkách 589,0/589,6 nm, která leží v těsné blízkosti maxima spektrální citlivosti průměrného lidského oka. V okolí vlnové délky 590 nm je již poměrně vysoká i absorpce listových barviv a sodíkové výbojky jsou proto i velmi účinným zdrojem fotosynteticky účinného záření. Vzhledem k monochromatickému charakteru vyzařované energie jich však nelze samostatně používat při pěstování vyšších rostlin, neboť nedostatek krátkovlnných i dlouhovlnných paprsků brání jejich přirozenému vývoji (Kubín, 1973). Mají dlouhou životnost, která se pohybuje od 6000 do 8000 hodin (Musilová, 2009). Vysokotlaké sodíkové výbojky se používají hlavně k asimilačnímu přisvětlování ve sklenících. Mají vysoký měrný výkon a přeměňují s vysokou účinností elektrickou energii na viditelné záření (23 %). Výbojky s výkonem 400 W vydávají kolem 90 W FAR. Nejvíce energie vyzařují ve vlnových délkách 560 až 620 nm (Volf, Votruba, 1991). Index podání barev dosahuje hodnot pouze Zdroje využívající luminoforů zářivky Zářivky jsou nízkotlaké rtuťové výbojky s luminoforem, v nichž postupně dochází k dvojí energetické přeměně: 1. Výbojem ve rtuťových parách s velmi nízkým tlakem (řádově 0,01 toru) se nejdříve mění elektrická energie v ultrafialové záření, a to s neobyčejně velkou účinností okolo 60 %. Zároveň se ještě malá část příkonu (asi 2 %) přeměňuje ve viditelné záření s vlnovými délkami 404,7 407,8 435,8 546,1 577,0 579,1 nm. Tyto spektrální 33
37 čáry jsou charakteristické pro každou rtuťovou výbojku a je v nich soustředěna největší část energie, vyzařovaná těmito výbojkami v oblasti FAR. 2. Ultrafialové záření se přeměňuje v záření viditelné prostřednictvím vhodných látek, tzv. luminoforů, nanesených v tenké vrstvě na vnitřní stěně výbojové trubice. Účinnost této druhé přeměny závisí především na vlnové délce sekundárního (viditelného) záření a na fyzikálních i chemických vlastnostech luminoforu. Výsledná účinnost obou energetických přeměn v zářivce, tj. přeměny elektrické energie v ultrafialové záření a přeměny ultrafialového záření v záření fotosynteticky účinné může v optimálních případech dosahovat % (Kubín, 1973). Zářivky jsou v současné době nejúčinnější, a tedy i nejhospodárnější světelné zdroje. Pořizovací cena zářivky a jejího příslušenství (objímky, tlumivka, startér, kondenzátory) je sice značně vyšší než cena žárovky stejného příkonu, pracovní náklady jsou ovšem menší. K ekonomickému provozu přispívá i neobyčejně velká životnost zářivek (Podešva, Véber, Kubín, 1968). Zářivkových luminoforů je už dnes celá řada a značně se navzájem liší jak svými spektrálními vlastnostmi, tak svou energetickou účinností. Luminofory a jejich aktivátory se volí zpravidla tak, aby se světlo vydávané zářivkou blížilo svým charakterem buď světlu dennímu, nebo světlu žárovkovému (Podešva, Véber, Kubín, 1968) Snadná volba spektrálních vlastností zářivek vyústila v myšlenku, že by bylo možné vyrábět speciální zářivky pro pěstování rostlin, jejichž emisní spektrum by bylo přizpůsobeno absorpčnímu spektru asimilačních pigmentů (Kubín, 1973). Závislosti průměrné spektrální citlivosti rostlin na intenzitě fotosyntézy stále využívají někteří výrobci světelných zdrojů (Osram, Philips, GE, Megaman aj.) k vývoji a výrobě speciálních lineárních a kompaktních zářivek určených pro osvětlování rostlin. Ve spektru těchto zdrojů je silně potlačena zelenožlutá oblast, takže emitované světlo je purpurové (Haš, 2011). Tyto zářivky se doporučují pro pěstování rostlin v různých klimatizovaných zařízeních, tak pro pěstitelskou praxi. Přes velkou propagaci těchto zdrojů bylo ovšem prokázáno, že očekávaný efekt se nedostavil. Někteří autoři vysvětlují tento neúspěch takto: Dopadá-li na rostlinná pletiva záření, které je asimilačními orgány silně absorbováno, dochází k jejich pohlcení už v první vrstvě buněk pod epidermis. Značná část jejich energie nemůže být vůbec využita a ztrácí se v podobě tepla. Naproti tomu spodní vrstvy asimilačního pletiva nejsou dostatečně prozářeny 34



38 a trpí nedostatkem potřebné energie. Tento případ nemůže nastat, vyzařuje-li zdroj i záření jiných vlnových délek ve žlutozelené oblasti, které sice nejsou dokonale pohlcovány a větší měrou prostupují listovými pletivy. Záření propuštěné vrchními vrstvami pletiv se tak dostává i do spodních vrstev a dochází tak k rovnoměrnější distribuci záření v celém objemu listu. Asimilační pletivo tak nedosáhne nikde tzv. světelného nasycení a pracuje proto s větší účinností. Takové poměry mohou nastat nejen v pletivech vrchních listů, ale část záření může procházet i k nižším patrům listového zápoje rostlinného porostu (Kubín, 1973). Lineární zářivky v zahraniční odborné literatuře i ve firemních katalozích se průměr zářivkové trubice uvádí v osminách palce za písmenem T, takže zářivky o průměru 38 mm se označují T12, podobně průměru 26 mm odpovídá označení T8, průměru 16 mm T5 atd. Vhodné geometrické parametry, umožňující konstruovat materiálově úsporná svítidla s jednoduchou optikou s možností sestavovat je do estetických spojitých svíticích pásů nebo velkých ploch (Dvořáček, 2008). Kompaktní zářivky vyrábějí se 2 druhy - zářivky s vnějším předřadníkem (vyžadují odpovídající konstrukci svítidla, odlišnou od svítidel určených pro obyčejné žárovky) a zářivky s elektronickým integrovaným předřadníkem, které jsou určeny do žárovkových svítidel jako přímá a energeticky úspornější náhrada žárovky (Dvořáček, 2008). Příklady zářivek doporučovaných k osvětlování rostlin (modře jsou vyznačeny reflektorové zdroje) (Haš, Pavlíčková, ) 35


39 Vysokotlaké rtuťové výbojky s luminoforem Pro vysokotlaké rtuťové výbojky se používá vesměs luminoforů svítících červeně, čímž se zlepšuje spektrální složení vyzařovaného světla. Tyto luminofory jsou jiného složení než luminofory používané pro výrobu zářivek. Maximum ultrafialového záření připadá na dvojitou spektrální čáru s vlnovými délkami 365/366 nm. Luminofor musí být tedy buditelný především tímto zářením (Podešva, Véber, Kubín, 1968). Halogenidové výbojky lze z hlediska materiálu použitého na zhotovení vlastního hořáku rozdělit do dvou základních skupin: výbojky s křemenným hořákem a výbojky s keramickým hořákem. Vývojky s křemenným hořákem jsou vhodné jako přímá náhrada sodíkových výbojek v těch případech, kdy je žádoucí zlepšit podání barev bez větších investičních nákladů. Vyrábějí se většinou pro vyšší příkony (od 70 W) a jejich pořizovací cena je vyšší než u výbojek s keramickým hořákem, které se vyrábějí i pro menší příkony a jsou vhodné pro běžné použití v interiérech, kde umožňují dosáhnout velmi dobrého podání barev. U obou typů je potřeba použít předřadné zařízení (Dvořáček, 2009) U halogenidových výbojek je třeba počítat s poměrně velkým poklesem zářivého toku v průběhu jejich života. Maximální doba života výbojek je deset až patnáct tisíc hodin, ale po hodinách klesá světelný tok o 20 až 50 %. Při osvětlování rostlin s velkými světelnými požadavky je roční doba svícení až hodin, a je tedy třeba výbojky po dvou až dvou a půl letech vyměnit. Příklady halogenidových výbojek doporučovaných k osvětlování rostlin (modře jsou vyznačeny reflektorové zdroje) (Haš, Pavlíčková, ). 36
40 Elektroluminiscenční zdroje Vybudíme-li luminiscenci v pevném luminoforu, např. některém krystalofosforu, působením silného elektrického pole (stejnosměrného nebo střídavého), mluvíme o elektrolumincenci (Kubín, 1973). A právě tyto luminofory umožňují vyrábět bílé LED v celé škále náhradních teplot chromatičnosti, od 2500 K (teple bílá) až po 7000 K (denní světlo). Spektrum LED je spojité, takže obsahuje úplnou škálu barev a jejich odstínů. To umožňuje dosahovat vysokých hodnot indexů podání barev. Zdroje teple bílé soustřeďují převážnou část emitovaného záření do oblasti největší citlivosti lidského oka, ale preferují dlouhovlnnější část spektra. Zdroje neutrální nebo studené bílé opět největší část záření vysílají v oblasti největší citlivosti oka, ale preferují krátkovlnné záření. Všechny tyto zdroje mají propad emitované energie v oblasti asi nm. Hloubka tohoto propadu závisí na účelu použití zdrojů LED. Zdroje pro interiérové osvětlování mají spektrum vyrovnanější, maxima jsou nižší, propad v oblasti kolem 500 nm není tak hluboký, index podání barev dosahuje hodnoty R a = 80. Ve vývoji jsou i zdroje LED s luminofory, které při náhradní teplotě chromatičnosti 5500 K dosahují indexu podání barev až R a = 97. Spojité široké a energeticky vyrovnané spektrum je velice vhodné i pro všechny fotobiologické procesy v rostlinách. Je velice příznivé, že i při vyšších teplotách chromatičnosti dosahuje část spektra až k vlnové délce 800 nm, což má významný vliv na indukci květů a kvetení u všech rostlin, i u dlouhodenních, jejichž fytochrom far red vyžaduje ke své aktivaci právě toto dlouhovlnné červené záření. Dostatečný podíl záření v červené i modré oblasti u doporučovaných LED skýtá dostatek energie k fotosyntetickým procesům. Zdroje LED jsou vyráběny převážně jako reflektorové, s menším nebo větším úhlem poloviční svítivosti. To umožňuje volit jednoduchá svítidla a nezávislost rozdělení svítivosti na těchto svítidlech. Vzhledem k tomu, že tyto zdroje s jejich chladicím systémem mají zpravidla velmi působivý design, je příjemné je použít i bez svítidel, pouze s vhodně krytou objímkou. Zdroje LED se vyrábějí buď rotačně symetrické, nebo jako světelné lišty, zpravidla s délkami od 30 cm, velmi často jako stavebnicové, s možností spojit několik lišt za sebou. I světelné lišty už bývají konstruovány tak, že se použijí samostatně, bez svítidla, někdy jen s clonou proti oslnění. Zdroje LED mají ještě další příznivé vlastnosti. Jejich příkon a spotřeba energie při osvětlování rostlin mohou být zpravidla menší než u jiných zdrojů a jejich doba života je až hodin, což při osvětlování rostlin činí 10 až 20 let (Haš, Fikarová, 2011). 37
41 Navrhování osvětlení pro interiérové rostliny Nedostatečné denní osvětlení rostlin jak v intenzitě, tak i v délce osvětlení lze kompenzovat jen umělými zdroji světla. Výběr těchto zdrojů je závislý na účelu pěstování rostlin. Zcela jiné jsou požadavky na osvětlení v prostorách bez denního světla (fytotrony, pěstební komory), na přisvětlování rostlin ve sklenících a na osvětlování rostlin v interiérech s pobytem lidí. Jiné jsou požadavky na intenzity a doby osvětlení, ale i na kvalitu světla, vyjádřenou složením emitovaného záření a barvou světla (Haš, 2011). Vhodná svítidla Na rozdíl od přisvětlování produkčních rostlin a jiných produktů ve sklenících je zřejmé, že v interiérech nejen rostliny, ale i použitá svítidla musí plnit také funkci estetickou a dekorační (Hutla, Matouš, 2002). Měla by také správně usměrňovat světelný tok a jejich provedení je vhodnější jednoduší, méně nápadné, aby hlavní pozornost upoutávaly rostliny. Z hlediska usměrňování světelného toku je účelné používat reflektorové světelné zdroje, které mohou být osazeny v jakémkoliv esteticky vhodném svítidle. Jestliže reflektor usměrňuje světelný tok do kužele s menším vrcholovým úhlem, řeší reflektorový zdroj i problém oslnění. U některých světelných zdrojů, především u všech halogenidových výbojek, je zapotřebí počítat s tím, že ve svítidle je třeba umístit ještě předřadné přístroje. Je tedy možné použít jen ta svítidla, která jsou určena pro zvolenou výbojku (Haš, Pavlíčková, ). Proto je mj. vhodné, aby stavební architekti již při projektování konzultovali možnosti ozelenění moderního interiéru se zahradním architektem-specialistou na interiérovou zeleň a stanovili teplotní i světelné požadavky rostlin a v případě nedostatečného množství přirozeného záření připravili možnosti pro doplňkové osvětlení zdroji FAR. Bez této projektové přípravy je při instalaci rostlin velmi těžké optimální osvětlení vytvořit (Hutla, Matouš, 2002). Doba osvětlování Při provozování fotosyntetických svítidel je důležité znát, na jak dlouho je třeba je v průběhu dne zapnout. Lze vycházet i z jednoduchého pravidla, že průměrná minimální potřeba rostlin je asi lx po dobu dvanácti hodin denně. Výsledky pokusů orientovaných na praxi říkají, že: při vyšší intenzitě osvětlení je možné zkracovat jeho dobu, při střední nebo nízké intenzitě musí být doba osvětlení delší. 38
42 Zdá se, že pro většinu rostlin je nepodstatné, kdy se během 24 hodin svítí. Zpravidla se rostliny v interiéru osvětlují vegetačním osvětlením během dne nebo během pracovní doby či v době největšího návštěvnického provozu. Přirozené denní světlo musí v průměru podle intenzity záření a světelných nároků rostlin společně s doplňkovým vegetačním osvětlením pokrýt denní potřebu za dvanáct hodin. Obecně platí, že i v interiéru dobře osvětleném denním světlem je třeba rostliny s vysokými nároky přisvětlovat od poloviny září, rostliny se středními nároky od poloviny října a rostliny s nízkými nároky od poloviny listopadu. Následující doporučení přisvětlovacího rytmu a doby osvětlení vychází ze známých světelných nároků rostlin: dvě až tři hodiny v září a dubnu, tři až pět hodin v říjnu a v březnu, šest až deset hodin v listopadu a v únoru, jedenáct až šestnáct hodin v prosinci a lednu. Doplňkové vegetační osvětlení je vhodné ovládat časovými spínači. Podání barev Lidem i rostlinám vyhovuje, když spektrální složení světla zdroje zajišťuje co nejlepší podání barev. Pro intenzity osvětlení, které jsou uvažovány při osvětlování rostlin v interiéru, jsou nejvhodnější zdroje s neutrálním bílým světlem (3 300 až K), které příjemně zobrazuje bílou barvu i základní barevné odstíny. Zdroje k osvětlování interiérových rostlin musí mít barvu světla alespoň 842, popř V místech silně osvětlovaných denním světlem bývá vhodné použít zdroje s barvou 860, ale nejlépe 960. Z toho vyplývá, že zcela nevhodné jsou zdroje s purpurovou barvou (Flora, Fluora, Planta), které někteří výrobci dodávají s určením jako zdroje k osvětlování rostlin. Pro splnění požadavku na přírodní podání barev nejsou vhodné ani žádné teplotní zdroje (obyčejné i halogenové žárovky), ani rtuťové a sodíkové výbojky. Rovněž nejsou vhodné ani běžně prodávané zdroje k osvětlování interiérů, jejichž barva bývá obvykle 827 nebo 825. Nejvhodnější spektrum emitovaného záření mají některé luxusní lineární zářivky s barvou světla 965 (označované de Luxe, Biolux), jejichž podání barev je téměř takové jako na přirozeném denním světle. Lineární zářivky ale bývá jen málokde možné esteticky umístit nízko nad rostlinami. Lepšího estetického působení se dosáhne spíše bodovými zdroji - halogenidové výbojky, kompaktní zářivky (Haš, Pavlíčková, ). 39
43 Při vyšší teplotě chromatičnosti (nad 5000 K) může poklesnout podíl emitovaného záření v červené oblasti (Haš, Fikarová, 2011). Množství světelného záření Publikované doporučované hodnoty ozáření pro jednotlivé rostliny se uvádějí jak v jednotkách W*m -2 FAR, tak v jednotkách μmol*s -1 *m -2. Při výpočtech osvětlovacích soustav pro doplňkové přisvětlování je třeba uvedené fytotechnické jednotky převádět na fotometrické (lm, lx), protože v těchto jednotkách se uvádějí technické parametry světelných zdrojů. Vzájemné vztahy fytometrických a fotometrických jednotek jsou: F λ dλ [W FAR] 119,4 (F λ / λ ) dλ [μmol.s - V λ. F λ dλ [lm] kde F je spektrální zářivý tok, V je poměrná světelná účinnost monochromatického záření (ČSN ) = spektrální citlivost lidského oka (lm.w -1 ) Fotometrické a fotonové jednotky jsou závislé na vlnových délkách a proto jejich vzájemné poměry jsou pro různé světelné zdroje různé. Pro stanovení přepočítávacích faktorů je nutné mít jejich emisní spektra. Pro zjištění zářivých toků ve fytotechnických jednotkách použijeme údaj o světelném toku zdroje, který je udáván v technické dokumentaci. U reflektorových zdrojů nebývá uváděna hodnota světelného toku (lm), ale hodnota maximální svítivosti (v kandelách, cd). Vypočítané přepočítávací koeficienty pro různé světelné zdroje, doporučené pro osvětlování interiérových rostlin, jsou na Tabulce 1 v Příloze I a příklad výpočtu přepočítávacího koeficientu je v Příloze VII výpočty. V některých, zejména starších odborných pracích, se uvádějí doporučené hodnoty ozářenosti ve fotometrických jednotkách (luxech). Pokud byly při stanovování doporučovaných hodnot použity žárovky, je možné tyto hodnoty využít, převést je na fytotechnické jednotky a ty pak dále použít pro výpočty osvětlování jinými libovolnými zdroji. Pokud byly pro určení doporučovaných hodnot používány sodíkové nebo halogenidové výbojky, může být přepočet jen informativní. Přepočítávací koeficienty výbojek mají podle typů velké rozmezí: halogenidové výbojky 3,14 3,87 W*m -2 FAR, resp. 14,5 až 15,7 μmol.s -1 *m -2 na 1000 lx, sodíkové výbojky 2,30 2,74 W*m -2 FAR, resp. 11,30 až12,65 μmol.s -1 *m -2 na 1000 lx. 40
44 V takových případech je užitečné provést dříve uvedeným způsobem výpočet z emisního spektra nebo měření foto-a kvantum-metrem. Posoudí-li se v Tabulce 1 vzájemný poměr koeficientů k FAR a k mol, je zřejmé, že pro jednotlivé světelné zdroje je jen malý rozdíl, všechny hodnoty jsou v rozmezí 0,220 0,231. Z toho vyplývá, že pro doporučované zdroje k ozařování pokojových květin příliš nezáleží na tom, jaké měrové jednotky se k výpočtům osvětlení používají. V tabulce 1 jsou uvedeny ještě indexy fotoperiodické radiace, které zhodnocují vhodnost světelných zdrojů pro ovlivňování fotoperiody a kvetení. Index IFPR je podíl emitovaného záření v rozmezí vlnových délek nm (FPR), které aktivují fytochromy red a far red, k fotosynteticky aktivnímu záření v rozmezí vlnových délek nm (FAR): FPR IFPR = FAR Pro měření intenzity ozáření sluncem a oblohou se používají pyranometry (aktinometry, solarimetry), které zaznamenávají globální záření, to je celkové ozáření sluncem a oblohou na vodorovné rovině. Těchto přístrojů se používá i pro hodnocení fotosynteticky aktivního záření v denním světle. Pro měření denního záření i záření umělých zdrojů se vyrábějí speciální měřiče, Quantum metry, cejchované v uvedených jednotkách μmol*s -1 *m -2 (bývá uváděno též μmol*m -2 *s -1 ). Požadavkem pro tyto přístroje je, aby jejich měřené hodnoty v celém rozsahu FAR co nejlépe odpovídaly závislosti fotonového toku na vlnové délce a aby jejich meze citlivosti byly co nejpřesněji omezeny vlnovými délkami 400 a 700 nm. V současnosti tyto požadavky nejlépe splňuje přístroj LI-COR se senzory LI 190 (bodový), LI 191(lineární). Lze jím s vysokou přesností měřit všechny světelné zdroje, včetně slunečního záření. Různé způsoby měření intenzity záření znázorňuje Obrázek 8 v Příloze I. Pokud jsou k dispozici emisní spektra světelných zdrojů, pak je možné dříve uvedeným způsobem stanovit přepočítávací koeficienty k FAR, k mol, dopadající záření měřit luxmetrem a hodnoty fotosyntetické ozářenosti vypočítat. Výpočtem v oblasti nm lze stanovit i indexy IFPR. K měření intenzity osvětlení (lx) se ale musí použít pouze luxmetry, jejichž spektrální citlivost se perfektně shoduje s normativní křivkou citlivosti lidského oka V λ podle Mezinárodní komise pro osvětlování, CIE (ČSN ). Mnohé laciné luxmetry mívají značné odchylky své citlivosti od normativní křivky a chyby měření dosahují %. Při 41
45 používání všech přístrojů je nutné si uvědomovat, že se měří jen dopadající záření a nerespektuje se odraznost záření ani jeho propustnost listy a jinými orgány rostlin (Haš, 2011). Umístění umělých světelných zdrojů nad rostlinami U závěsných svítidel rozhoduje správná výška, resp. vzdálenost od rostlin, o účinku osvětlení. Příliš nízko zavěšené svítidlo nebo chybějící odstínění mohou u nejvyšších listů rostlin nebo na jejich špičkách vést k nevratným poškozením způsobeným vysokou teplotou. Při umístění na příliš velkou vzdálenost snížené množství světla může částečně nebo zcela ovlivnit ostatní růstové faktory, např. teplotu (Hutla, Matouš, 2002). Pro výpočet umístění světelných zdrojů nad skupinami rostlin je důležitý také způsob hodnocení ozářenosti (osvětlenosti). Při výpočtu pracovního osvětlení se hodnotí intenzita osvětlení na pracovní ploše, zpravidla vodorovné, ale i jinak orientované, pokud je to důležité z hlediska vytváření zrakového vjemu. Důležité je vždy to světlo, které se odráží od pozorovaného předmětu. Při osvětlování rostlin nás zajímá záření, které je rostlinou pohlceno. Charakteru působení světla na rostliny nejlépe odpovídá hodnotit světlo tak, jak dopadá na povrch polokoule, jejíž osa je vždy natočena ke světelnému zdroji. Protože ale vlivem indexu lomu rostlinné tkáně se při větších úhlech dopadu světla výrazně zvětšuje jeho odraz, uvažujeme, že světlo využitelné pro fotobiologické procesy dopadá na povrch kulového vrchlíku o výšce 3/5 poloměru polokoule. V tomto případě je poměr plochy základny vrchlíku a jeho kulové plochy 0,7 a průměrná intenzita využitelného dopadajícího záření je tedy rovna 7/10 světla dopadajícího na rovinu kolmou k přicházejícím paprskům (Haš, 2011). Vzorce pro výpočty a jejich postup je přehledně znázorněn v Příloze VII výpočty Rostliny vhodné do interiérů s nedostatkem světla Kvetoucí rostliny a rostliny s panašovanými listy se na stinných stanovištích necítí dobře (Brookes, 1992). Rostliny, které snáší stinná stanoviště, mají obvykle větší množství listů, které jsou široké a měkké (Hutla, Matouš, 2002). Dále bývají většinou nedělené a bez krycích chlupů. Anatomicky mají vždy silně vyvinuté asimilační pletivo (chlorenchym), v jehož buňkách jsou chloroplasty umístěny u buněčných stěn rovnoběžných s povrchem listu a průduchy jsou i na lícní (svrchní) straně listů dosti husté (Mareček, 2001). 42
46 U rostlin ozdobných listem rozlišujeme dvě hlavní skupiny: rostliny v první skupině zastavují koncem podzimu svůj růst a potřebují zimní klid, rostliny ve druhé skupině rostou dále i v zimě a zůstávají dekorativní. Rostliny pocházející z tropů, které v přírodě dostávají po celý rok denně asi 12 hodin slunečního svitu, rostou v mírném pásmu celý rok jen tehdy, když jim v zimním období vytvoříme umělým osvětlením optimální světelné podmínky v dostatečně teplé místnosti. V nevýhodě jsou často panašované rostliny, protože jejich žluté, krémové nebo bílé plochy listů neobsahují chlorofyl. Naproti tomu rostliny z mírného pásma zastaví svůj růst nebo ho alespoň podstatně zpomalí, když se dny zkracují a zima je přede dveřmi. Kvetoucí rostliny potřebují obvykle více světla k vytvoření a vývinu poupat (Brookes, 1992). K ozelenění těchto interiérů jsou vhodné rostliny nejnižších pater tropických lesů, např. Syngonium, Aglaonema, Philodendron, Cissus, Piper nigrum, mnohé kapradiny, či některé druhy temperátních lesů subtropického pásma, např. Aspidistra, Ophiopogon, Clivia, Fatsia, xfatshedera, Hedera a jiné (Kuťková, 2006). Vhodnými rostlinami jsou také Dieffenbachia, Dracaena, Chamaedorea, Cyperus, Spatiphyllum, Ficus elastica, Ficus lyrata, Howea forsteriana, Monstera deliciosa, Schefflera arboricola, Chlorophytum comosum, Peperomia, a další (Flehming, 2007) Interiéry s nadbytkem světla Problémem při pěstování rostlin nemusí být pouze nedostatek světla v zimě, ale i jeho nadbytek v letním období. Týká se to především budov s velkými prosklenými plochami, budov orientovaných na jih či jihozápad. Vysoká hodnota světla některým rostlinám vyhovuje, avšak za předpokladu, že se nejedná o přímé sluneční záření, ale o rozptýlené světlo. Tam, kde se jedná o přímé sluneční záření, musí být rostliny v letním období stíněny pomocí stínovek, rohoží, žaluzií či speciálních nátěrů skleněných ploch. Spolu s vysokou intenzitou slunečního záření dochází v těchto interiérech k výraznému přehřívání vzduchu v důsledku skleníkového efektu. Účinným opatřením je větrání (Kuťková, 2006). Brookes (1995) používá termín slunečné stanoviště, které definuje jako stanoviště, které celý den nebo část dne dostává přímé slunce. Autor dále uvádí, že okna situovaná k jihu dostávají sluneční světlo delší část dne, okna obrácená na východ několik hodin ráno a okna se západní expozicí několik hodin odpoledne. Intenzita slunečního záření také závisí na ročním období a geografické poloze místnosti. Okna na jih dostávají intenzivnější světlo, slunce však nezasahuje tak hluboko do místnosti jako ve východně nebo západně exponovaných místnostech. 43
47 3.4.4 Možnosti řešení nadměrného osvětlení v interiéru V období vysoké přírodní radiace vede zasklení průsvitnými skly na svislých a vodorovných plochách k silnému oteplení místnosti a také k nadměrné expozici rostlin. Hlavně při jižní orientaci oken je nezbytné sluneční záření odstínit, aby se zabránilo spálení či poškození listů a květů interiérových rostlin s výjimkou pouštních rostlin - kaktusů a sukulentů (Handbuch, 1998) Zatímco pojem stínění se vztahuje k nežádoucím ztrátám světla pro rostliny, stínováním se rozumí pěstitelské opatření k ochraně rostlin před nepříznivými účinky příliš silného slunečního záření. Snižují se nadměrné hodnoty ozářenosti a tak se předchází přehřívání rostlin, které způsobuje zpomalení růstu a v extrémních případech poškození rostlin popálením. Riziko popálení se zvyšuje při nízké vzdušné vlhkosti vzduchu (Volf, 2001) Mnoho druhů hrnkových květin nesnáší plné sluneční záření a nejcitlivější z nich se musí stínovat od března do září, méně citlivé od dubna do srpna. Zbytečně silné stínování však způsobuje zpomalení růstu, podporuje prodlužovací růst stonků, snižuje počet a kvalitu květů a zhoršuje vybarvení listů (Volf, Votruba, 1991). Stínování je však potřebné i u druhů tolerantních ke slunečnímu záření, a to jen po krátkou dobu v průběhu jejich množení, při pěstování choulostivějších mladých rostlin a dále po výsadbě nebo přesazení, než se obnoví schopnost kořenů přijímat dostatek vody. Citlivě také reagují neotužené rostliny, které se snadno popálí, jestliže jsou po nedostatku světla v zimě náhle vystaveny silnějšímu záření začátkem jara. V plném létě, kdy je záření nejsilnější, má být stínování silnější než na jaře a koncem léta (Volf, 2001). Stínovací zařízení má zabraňovat přehřívání rostlin a zároveň s větráním snižovat teplotu, nemá však bránit výměně vzduchu při větrání a nemá způsobovat zbytečné ztráty světla v případě slabého slunečního záření (Volf, Votruba, 1991). Díky stínicím zařízením je možné dosáhnout snížení účinků přímého slunečního záření v daném interiéru, případně zabránit jeho proniknutí úplně a snížit tím celkovou teplotu v místnosti. Na nadměrně exponovaných stanovištích se tak zabrání škodám na rostlinách (popálení). Vhodné jsou automaticky řízené systémy. Musí se přitom brát v úvahu nároky rostlin na světlo (fotobuňky ve výšce rostlin) a ne na potřeby staveb (Richtlinie, 2002). 44
48 Vnější stínění Má tu výhodu, že už vně oken je spolu s teplem odráženo. Zabraňuje většímu ohřevu místností a ulehčuje řízení teplot prostoru. Jejich pořizovací náklady jsou sice značně vyšší, ale také mnohem efektivnější než vnitřní stínidla. Prvky vnějšího stínění jsou vystaveny vnějším povětrnostním vlivům, a proto musí být vysoce kvalitní a trvanlivé. Ovládání je možné automaty (kombinace externí světelný senzor/fotobuňka) s nastavitelnou minimální a maximální hodnotou luxů. Jako externí stínování, i když ne tak efektivní, působí i markýzy a přístřešky (Handbuch 1998). Vnitřní stínění Vznik tepla je zde redukován jen nepatrně, takže musí být zřízeno odpovídající větrání, ventilace. Patří sem rolety a žaluziové systémy vyrobené z různých materiálů. Stínící účinek plastových systémů je 20 až 60 %. K dostání jsou v různých barvách a variantách, díky svému přirozenému působení jsou upřednostňovány světlé tóny. Doplňkové aluminiové pásky mohou docílit snížení vnikání tepla o 5 až 15 %, odrážejí totiž dlouhovlnné tepelné záření ve směru rostlin a povrchů (Richtlinie, 2002). Akrylové tkaniny mají životnost asi 10 let, s polyesterové 5 let. I tyto systémy je možné řídit automaticky. Stínicí účinek mají i klasické závěsy. Výběrem závěsů převážně z akrylového polyesteru nebo polyamidu tkanin, může být dosaženo zastínění 20-70%. Světlé a béžové tóny jsou vzhledem k lepší reprodukci barev vhodnější než tmavé odstíny. Systémy na odvádění a odrážení světla Prostřednictvím těchto zařízení je možné docílit kvalitní ochrany proti příliš intenzivnímu slunečnímu záření. Patří sem vysoce lesklé žaluziové rolety, které se instalují před, za nebo mezi dvojitá skla. Jsou poháněné motorem a jejich spouštění je řízeno dle světla na stanovišti. Proti vnikání světla do místnosti mohou být interiéry doplněny závěsy, světlými látkami, fóliemi, atd. Množství světla, které skrz tyto systémy do místnosti prochází, však většině rostlin v zimě k růstu nepostačuje a musí se tedy nahradit přídavným osvětlením (Richtlinie, 2002). Tento problém nastává i u speciálních skel - teplo-absorbující skla, protisluneční skla a speciální folie aplikované na izolační sklo (Handbuch, 1998) Rostliny vhodné do interiérů s nadbytkem světla Rostliny náročné na světlo často nemají žádné nebo málo malých listů, často mívají barevné listy (Hutla, Matouš, 2002). Největší světelnou intenzitu mají stanoviště vysokohorská, 45
49 pouštní a na mořském pobřeží, kde také rostliny jeví nejlépe vyvinutá přizpůsobení na ochranu před přílišným vlivem světla. Protože zdroj světelných a tepelných paprsků je společný, jsou také přizpůsobení k vyšší světelné intenzitě podobná přizpůsobení k vysoké teplotě. Z anatomicko-morfologických změn vyvolaných přílišným světlem je zmenšení plochy listů, silná vrstva kutikuly na pokožce a prodloužené buňky palisádového pletiva, v němž jsou chloroplasty uloženy jen na bočných stěnách buněk (protože přílišná světelná intenzita brzdí činnost chloroplastů). Jako ochranu před silným světlem vytvářejí rostliny slunných stanovišť i silné odění, mnohé listy stavějí hranou ke světlu, svrchní plochy listů se pokrývají silně lesklou, světlo odrážející vrstvou kutikuly nebo jsou silně ojíněné. Protože hlavně ultrafialové světlo brzdí prodlužovací růst stonků, jsou rostliny slunných stanovišť v horách většinou hustě trsnaté až polštářovité nebo kobercovité a keře se vyvinují jako nízké, hustě větvené keříky. Květy se však vyvinují do značné velikosti a jejich koruny bývají nápadně barevné (Dostál, 2001). K ozelenění nadměrně osvětlených interiérů jsou vhodné rostliny pocházející z otevřených stanovišť tropických savan, sucholesů, pouští a polopouští či rostliny subtropických oblastí z formací tvrdolistých lesů, stepí, pouští a polopouští. Konkrétně lze použít některé rody a druhy z čeledi Euphorbiaceae (Euphorbia tirucalli, E.trigona), Bromeliaceae (Aechmea fasciata, Puya, Streptocalyx) Crassulaceae, Cactaceae, Cycadaceae, Agavaceae. Dále lze použít takové rody a druhy jako je Dracaena draco, Ceropegia, Hoya, Tradescanthia sillamontana, Pachypodium, Beucarnea apod.. Nadměrně světlé interiéry vyhovují i celé řadě kvetoucích druhů (např. Anthurium andreanum hybridy, Spathiphyllum, Bougainvillea). Při konkrétním výběru druhů je nutno respektovat požadavky taxonu na teploty v zimě (Kuťková, 2006). Velmi světlé podmínky vyhovují také druhům Camellia japonica, Citrus, Musa acuminata, Myrtus communis, Sparmannia africana, Capsicum annuum, Pentas lanceolata, Senecio herreianus, Ceropegia woodi a dalším (Flehming, 2007). 46
50 4. MATERIÁL A METODIKA 4.1 Hodnocení světelných podmínek pro potřeby pěstování rostlin Cílem diplomové práce je vypracovat metodiku hodnocení světelných podmínek z pohledu potřeb pro pěstování rostlin. Po dohodě s vedoucí práce bude vybráno interiérů s interiérovými rostlinami, které budou takto zhodnoceny. Světelné podmínky budou hodnoceny na základě měření luxmetrem v blízkosti rostlin umístěných v interiéru v době, kdy se zhoršují přirozené světelné podmínky. Od konce října do začátku března v intervalu 14 dnů, vždy mezi 10. a 15. hodinou bude naměřena jedna hodnota a zaznamenána do tabulky. Jedná se většinou o interiéry veřejné, které jsou snáze přístupné pro měření Obecné charakteristiky hodnoceného interiéru Popis a lokalizace interiéru - u každého interiéru bude uveden název objektu, adresa a patro. Typ interiéru - každý veřejný interiér bude zařazen podle svého charakteru do kategorie, které vycházejí z hodnocení podle převládající funkce dle Kuťkové. Soukromé interiéry nebudou dále kategorizovány. Klasifikace veřejných interiérů podle převládající funkce dle Kuťkové (2006): 1. Interiéry s převládající společenskou funkcí 2. Interiéry s převládající shromažďovací funkcí 3. Pracovní interiéry 4. Historické interiéry 5. Interiéry určené k vykonávání různých sportovních, rekreačních a relaxačních činností 6. Multifunkční interiéry - obchody, obchodní centra, obchodní pasáže Expozice interiéru u každého interiéru bude uvedena orientace ke světovým stranám a použity tyto zkratky: J jih JV jihovýchod V východ SV severovýchod 47
51 S sever SZ severozápad Z západ JZ jihozápad H horní osvětlení Zeleň použitá v interiéru u každého interiéru bude uveden seznam druhů rostlin, které jsou zde použity. Pokud jsou interiérové rostliny na více stanovištích v místnosti a jsou zde různé světelné podmínky, budou hodnocena tato stanoviště zvlášť a pro každé z nich bude uvedena samostatná tabulka naměřených hodnot osvětlení Podmínky při měření Při každém měření bude zaznamenán údaj o meteorologické situaci. Mohou to být tyto situace: Jasno (zkratka J) plná intenzita záření Polojasno (zkratka P) intenzita záření je částečně snížena oblačností Zataženo (zkratka Z) pouze difuzní záření Přídavné osvětlení (zkratka PO) stanoviště je silně ovlivněno přídavným osvětlením Vyhodnocení vhodnosti umístění interiérových rostlin podle naměřených hodnot osvětlenosti Rozdělení stanovišť v interiérech podle světelných podmínek Naměřeno bude 10 hodnot pro každé stanoviště a podle průměru z naměřených hodnot s vynecháním 2 krajních, nenižší a nejvyšší (mohou být ovlivněny chybou či náhodnými extrémními světelnými podmínkami) budou hodnocená stanoviště v interiéru rozdělena do těchto kategorií osvětlenosti: 1.- Velmi světlá průměrná hodnota nad 2000 lx 2. - Světlá průměrná hodnota lx 3. - Tmavá průměrná hodnota lx 4. - Velmi tmavá průměrná hodnota do 300 lx 48
52 Rozdělení rostlin podle nároků na množství FAR Rostliny budou rozděleny do 3 skupin podle požadavků na množství FAR. Údaje o zařazení jednotlivých druhů do skupin budou zpracovány formou tabulky, která bude použita pro hodnocení vhodnosti umístění rostlin na daném stanovišti 1. - Vysoké požadavky na množství FAR pro udržení 1500 až 2000 lx - pro růst, popř. vykvetení nad 2000 lx 2. - Střední požadavky na množství FAR - pro udržení 500 až 750 lx - pro růst, popř. vykvetení 1500 až 2000 lx 3. - Nízké požadavky na množství FAR - pro udržení 300 až 500 lx Tabulka pro hodnocení vhodnosti umístění rostlin - pro růst, popř. vykvetení 500 až 750 lx (zpracováno na základě článku Světlo a rostlina v časopise Světlo, 2002, Machovec, 1975 a Martínek, ) Název rostliny Adiantum radianum 3 Aglaonema commutatum 3 Aloe vera 1 Anthurium andreanum 1 Anthurium sp. 2 Araucaria heterophylla 1 Asparagus densiflorus sprengeri 2 Aspidistra elatior 3 Begonia coralina Luzerna 2 Cissus rhombifolia 3 Clivia miniata 3 Codiaeum variegatum 2 Dasylirion longissimum 1 Dieffenbachia maculata 3 Dracena deremensis 3 Epipremnum pinnatum 3 Epipremnum pinnatum Neon 2 Euphorbia tirucalli 1 Euphorbia trigona 1 49 Zařazení do skupiny podle nároků na množství FAR
53 Ficus benjamina 2 Ficus elastica 3 Ficus lyrata 3 Hedera helix 2 Hibiscus rosa sinensis 1 Hipeastrum 1 Chamaedorea elegans 3 Chlorophytum comosum 3 Monstera deliciosa 3 Nephrolepis exaltata 2 Nerium oleander 1 Olea europaea 1 Pandanus veitchii 2 Peperomia obtusifolia 3 Philodendron erubescens 3 Phoenix canariensis 2 Radermachera sinica 2 Rhipsalis sp. 1 Sanseviera cylindrica 1 Sanseviera parva 3 Sanseviera trifasciata 3 Schefflera actinophylla 3 Schefflera arboricola 2 Spatiphyllum sp. 3 Stromante sp. 2 Synadenium grantii 1 Xantorea sp. 1 Zamioculcas zamifolia 3 Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Vhodné umístění číslo kategorie osvětlenosti i číslo skupiny podle požadavků na množství FAR se shoduje Částečně vhodné umístění - číslo kategorie osvětlenosti i číslo skupiny podle požadavků na množství FAR se liší pouze o 1 jednotku. Rostliny zde vegetují, ale bylo by možné sem umístit vhodnější druhy, které tu budou lépe kvést nebo se více a rychleji rozrůstat. Nevhodné umístění - číslo kategorie osvětlenosti i číslo skupiny podle požadavků na množství FAR se liší o 2 jednotky. Hrozí nebezpečí poškození rostlin. U náročných na světlo hlavně 50
54 v zimě, stínomilných hlavně v létě. Pokud zde tyto rostliny mají být umístěny je třeba jim podmínky uměle upravit přisvětlením nebo stíněním. 4.2 Přisvětlování rostlin Cílem pokusu je zjistit, jaký vliv bude mít přisvětlování na růst a vývoj různých rostlin v období s nepříznivými světelnými podmínkami. Ke sledování jsou použity druhy Cyclamen persicum (krátkodobá interiérová rostlina), Codiaeum variegatum (interiérová rostlina ozdobná listem) a Spatiphyllum sp. (dlouhodobá kvetoucí interiérová rostlina). Sledují se 4 skupiny rostlin: a) kontrolní skupina b) skupina ozařovaná kompaktní zářivkou barvy 827 c) skupina ozařovaná kompaktní zářivkou barvy 842 d) skupina ozařovaná halogenidovou výbojkou barvy 942 Osvětlení: Všechny rostliny se umístí tak, aby bylo přibližně stejné jejich denní osvětlení. Rostliny s doplňkovými světelnými zdroji se uspořádají tak, aby jednotlivé skupiny nebyly ovlivňovány zdroji jiných skupin. Intenzita doplňkového ozáření je ve všech případech 2000 lx. Činnost doplňkového osvětlení je řízena automaticky pomocí časového a soumrakového spínače. Osvětlení se zapíná každé ráno ve 4.00 hod., v průběhu dne se vypíná vždy, když intenzita venkovního osvětlení v místnosti překročí hodnotu 1500 lx. Večerní vypnutí osvětlení provede časový spínač vždy ve hod. Vývoj a růst rostlin: Z dostupných exemplářů rostlin stejného druhu se vybere po čtyřech rostlinách přibližně stejné velikosti, stejného počtu a velikosti listů, případně stejného počtu květů a poupat. Každých 14 dní se hodnotí: a) výška hlavního stvolu nebo nejvýše vyrostlého listu nebo květu b) počet listů c) celková listová plocha d) počet květů e) počet vyvinutých poupat 51
55 f) barva listů a květů (fotograficky, stále stejným přístrojem, digitální fotografie budou zpracovávány vždy jen automatickou úpravou) Na konci pokusu se dále bude hodnotit: g) celkový počet opadaných listů h) celkový počet nových listů i) celkový počet opadaných květů j) celkový počet nových poupat k) nárůst celkové listové plochy (včetně opadaných listů) Celková listová plocha se zjišťuje jen jednou za měsíc. Používá se metoda korelace plochy a šířky průměrného listu, korelační koeficient se zjistí na počátku pokusu a při dalším hodnocení se zjišťuje součet šířek všech listů na rostlině a násobením stanoveným korelačním koeficientem se zjistí plocha listů. Korelační koeficient se stanoví tak, že se tři listy různých velikostí od každého druhu obkreslí na milimetrový papír a spočítá se jejich plocha a změří se příslušná šířka. Jako korelační koeficient se používá průměrná hodnota podílů plochy a šířky jednotlivých listů. 4.3 Modelové objekty Po dohodě s vedoucí práce byly vybrány 3 modelové objekty: kancelář firmy Spektrum Holas v Havlíčkově Brodě, vstupní prostor kolejí a vstupní hala budovy A Zahradnické fakulty v Lednici. Ve dvou případech je řešeno nedostatečné, v jednom případě nadbytečné osvětlení Vstupní hala Zahradnické fakulty v Lednici Jedná se o prostor s velkým pohybem lidí. Je často využíván k prezentaci studentských prací a k dalším účelům, jako jsou např. ochutnávky a předváděcí akce na den otevřených dveří, ochutnávka salátů a podobně. V současné době jsou zde umístěny 3 nádoby s rostlinami, které jsou přisvětlována umělými zdroji. Ty sem byly instalovány pravděpodobně bez podrobnějšího návrhu a jejich intenzita ani podání barev není ideální Vstupní prostor kolejí Tento prostor se vyznačuje velkou intenzitou pohybu studentů. Slouží i ke krátkodobějšímu čekání (jsou tu 2 lavičky) a také je zde umístěna nástěnka, kde se vyvěšují letáky s různými informacemi. Zatím v prostoru nejsou žádné interiérové rostliny. Osvětlení dosahuje vysokých hodnot a teplota tu značně kolísá. 52
56 4.3.3 Kancelář firmy Spektrum Holas v Havlíčkově Brodě V tomto interiéru jsou přirozené světelné podmínky velmi nevhodné, co se týče potřeb pro pěstování interiérových rostlin. Místnost je orientována na západ, ale okno směřuje do dvora, takže ani přímo na okně není intenzita světla dostatečná pro náročnější druhy rostlin. Proto byl tento interiér zvolen pro vyzkoušení funkčních vzorků svítidel pro přisvětlování rostlin v praxi. 53
57 5. VÝSLEDKY 5.1 Hodnocení světelných podmínek ve vybraných interiérech z pohledu potřeb pro pěstování rostlin Světelné podmínky byly hodnoceny ve 14 veřejných interiérech a 1 soukromém, přičemž ve 4 z nich byla posuzována 2 stanoviště s různými světelnými podmínkami, takže celkem bylo posuzováno 19 stanovišť. Stanoviště byla hodnocena na základě měření luxmetrem každé 2 týdny v době od konce října do začátku března. Podle průměrné hodnoty bez dvou krajních naměřených hodnot byla stanoviště rozdělena do 4 kategorií: 1. - Velmi světlá průměrná hodnota nad 2000 lx 2. - Světlá průměrná hodnota lx 3. - Tmavá průměrná hodnota lx 4. - Velmi tmavá průměrná hodnota do 300 lx Pak byla podle tabulky, která je uvedena v kapitole Materiál a metodika, posouzena vhodnost umístění interiérových rostlin na daném stanovišti. Celkem bylo na stanovištích hodnoceno 69 rostlin, které byly rozděleny do kategorií: 1. vhodné (39 ks) 2. částečně vhodné (23 ks) 3. nevhodné (7 ks) Výsledky hodnocení shrnují následující tabulky: Tabulka rozdělení interiérů do kategorií Stanoviště Česká pošta Havlíčkův Brod Dům Pánů z Lipé Galerie Vaňkovka přízemí Galerie Vaňkovka východ v 1. Patře Metal Aliance Havlíčkův Brod Kategorie Tmavá Tmavá Tmavá Tmavá Tmavá 54
58 Nádraží v Podivíně u zdi Nemocnice Havlíčkův Brod u sloupu Okno v knihovně ZF v Lednici Poliklinika Havlíčkův Brod Rodinný dům Havlíčkův Brod Vstupní hala ZF v Lednici Nemocnice Havlíčkův Brod u oken Tmavá Tmavá Tmavá Tmavá Tmavá Tmavá Velmi světlá Plavecký bazén Havlíčkův Brod u oken Velmi světlá Plavecký bazén Havlíčkův Brod u zdi Vstupní hala budovy D ZF v Lednici Kancelář HB Auditing Havlíčkův Brod VZP Havlíčkův Brod Nádraží v Podivíně parapet Velmi světlá Velmi světlá Velmi tmavá Velmi tmavá Světlá Tabulka vyhodnocení vhodnosti použití rostlin na stanovištích Taxon Velmi světlá stanoviště Světlá stanoviště Tmavá stanoviště Velmi tmavá stanoviště Adiantum radianum Vhodné Aglaonema commutatum Vhodné Aloe vera Vhodné Anthurium andreanum Nevhodné Anthurium sp. Částečně vhodné Araucaria heterophylla Vhodné Asparagus densiflorus sprengeri Vhodné Aspidistra elatior Vhodné Begonia coralina Luzerna Vhodné Částečně vhodné Cissus rhombifolia Nevhodné Clivia miniata Vhodné Codiaeum variegatum Částečně vhodné 55
59 Dasylirion longissimum Dieffenbachia maculata Dracena deremensis Epipremnum pinnatum Epipremnum pinnatum Neon Euphorbia tirucalli Euphorbia trigona Ficus benjamina Vhodné Částečně vhodné Vhodné 2 x Částečně vhodné Částečně vhodné 2x vhodné 3 x vhodné 3 x vhodné Částečně vhodné Nevhodné 2 x částečně vhodné Ficus elastica Nevhodné Vhodné Ficus lyrata Hedera helix Hibiscus rosa sinensis Hipeastrum Chamaedorea elegans Vhodné Částečně vhodné Částečně vhodné 2 x vhodné umístění Chlorophytum comosum 3 x vhodné Monstera deliciosa Nevhodné Vhodné Nephrolepis exaltata Nerium oleander Olea europaea Pandanus veitchii Peperomia obtusifolia Philodendron erubescens Phoenix canariensis Radermachera sinica Sanseviera cylindrica Sanseviera parva Sanseviera trifasciata Schefflera actinophylla Vhodné Vhodné Vhodné Částečně vhodné Částečně vhodné Vhodné Částečně vhodné Vhodné Vhodné Vhodné Nevhodné Schefflera arboricola 3 x částečně vhodné Spatiphyllum sp. 2 x vhodné 2 x částečně vhodné 56
60 Stromante sp. Synadenium grantii Vhodné Částečně vhodné Částečně vhodné umístění Nevhodné Xantorea sp. Vhodné Zamioculcas zamifolia 2 xvhodné Částečně Vhodné Do kategorie tmavá spadá 11 hodnocených stanovišť. Sortiment rostlin je většinou vhodně zvolen, pouze ve 3 případech je zde umístěn nevhodný druh, ve 14 částečně vhodný, ve 26 případech je druh zvolen zcela vhodně. Nejčastějšími vhodně používanými druhy na těchto stanovištích jsou Spatiphyllum sp., Zamioculcas zamifolia, Chlorophytum comosum, Epipremnum pinnatum, Chamaedoreea elegans, Dieffenbachia maculata a Dracena deremensis 2 stanoviště spadají do kategorie velmi tmavá. 2 druhy jsou zde umístěny částečně vhodně, 1 nevhodně. Velmi světlých je 5 stanovišť. 10 druhů je zde umístěno vhodně, 4 částečně vhodně, 2 nevhodně. Světlé je pouze 1 stanoviště. 3 druhy jsou zde umístěny vhodně, 3 částečně vhodně. 5.2 Přisvětlování rostlin Cílem pokusu bylo posoudit vliv přisvětlování na různé druhy rostlin Cyclamen persicum, Codieum variegatum a Spatipihyllum sp. K přisvětlování byly použity tyto světelné zdroje: zářivka barvy 842, zářivka barvy 827, halogenidová výbojka barvy 942. Korelační koeficienty Korelační koeficienty pro výpočet listové plochy byly stanoveny následovně: Cyclamen 33,57; Spatiphyllum 92,66; Codieum 114,52. Tyto koeficienty byly použity pro všechny výpočty listových ploch. Osvětlení kontrolních rostlin Osvětlení kontrolních rostlin bylo měřeno při každém hodnocení pokusu vždy ve 12:00. Naměřené byly hodnoty uvedené v tabulce 1. 57
61 Tabulka 1 Datum Osvětlení v luxech ve 12: Tvorba květů Přisvětlování má rozhodně velký vliv na tvorbu květů obou pozorovaných kvetoucích druhů. Kontrolní rostliny během nepříznivého období stagnovaly a netvořily nové květy (kontrolní Cyclamen navíc po měsíci úplně uvadnul), zatímco osvětlované rostliny tvořily nové květy neustále. Jedinou výjimkou byl Cyclamen přisvětlovaný kompaktní zářivkou barvy 842, kde docházelo k problémům. Nejvíce nových poupat u Spatiphyllum se vytvořilo pod halogenidovou výbojkou. U kontrolní rostliny již založené květy rychle opadaly a nová poupata se začala tvořit až ke konci pokusu, když se zlepšily přirozené světelné podmínky. U Cyclamen se nejvíce poupat opět vytvořilo pod halogenidovou výbojkou. Původní rostlina pod zářivkou barvy 842 byla vyměněna a dále hodnocena pouze nová rostlina, protože již po prvních čtrnácti dnech původní rostlina přestala tvořit nové květy a opadala jí poupata, ale i po výměně bylo rozvíjení poupat a tvorba nových velmi pomalá. Původní rostlina začala opět kvést (byla ponechána pod zářivkou barvy 842) až 8.2. po měsíci a půl. I vyměněná rostlina po odkvětu již vytvořených květů (22.2.) začala stagnovat, vytvořila se pouze 3 nová poupata, která ani za měsíc (22.3. byl ukončen pokus) nerozkvetla. Kontrolní rostlina úplně zvadla 11.1., takže byla vyřazena z hodnocení byla jako kontrola použita rostlina původně osvětlovaná zářivkou barvy 842, která opět začala kvést. 58
62 Počet květů Spatiphyllum Charakteristika Halogenová Kompaktní Kompaktní Kontrola výbojka zářivka barvy zářivka barvy Celkový počet poupat Spatiphyllum nových Celkový počet opadaných květů Spatiphyllum Počet květů Cyclamen Charakteristika Halogenová Kompaktní zářivka Kompaktní zářivka Kontrola výbojka barvy 827 barvy 842 Celkový počet opadaných květů Cyclamen Celkový počet nových poupat Cyclamen Přírůstky Velikost přírůstků se lišila u různých rostlin pod různými zdroji. Největší přírůstky byly u Spatiphyllum pod zářivkou barvy 842, u Codieum pod halogenidovou výbojkou, ale rozdíl nebyl příliš velký. Cyclamen vytvořil nejvíce nových listů pod halogenidovou výbojkou a v tomto případě byl již rozdíl značný. U kontrolní rostliny Cyclamen i Spatiphyllum došlo k nárůstu počtu listů až od 8.2., kdy došlo ke zlepšení přirozených světelných podmínek. Listy Codieum začaly přirůstat ve stejnou dobu u všech skupin (pozorování 8.2.), ale výrazné rozdíly byly patrné v kvalitě kresby na listech. U kontrolní rostliny se kresba na prvních nových listech vůbec nevytvořila až do konce pokusu, až poslední list, který se vytvořil před ukončení pokusu měl na sobě patrnou kresbu (viz fotografická příloha). 59
63 Počet listů Codieum Charakteristika Halogenová výbojka Kompaktní zářivka barvy 827 Kompaktní zářivka barvy 842 Kontrola Celkový počet opadaných listů Codieum Celkový počet nových listů Codieum Listová plocha Codieum Charakteristika Halogenidová výbojka Kompaktní zářivka barvy 827 Kompaktní zářivka barvy 842 Kontrola Nárůst celkové listové plochy (včetně opadaných listů) Codieum 251,95 cm 2 183,23 cm 2 229,03 cm 2 160,69 cm 2 Počet listů Cyclamen Charakteristika Halogenová výbojka Kompaktní barvy 827 zářivka Kompaktní barvy 842 zářivka Kontrola Celkový opadaných listů Cyclamen počet Celkový počet nových listů Cyclamen Listová plocha Cyclamen Charakteristika Halogenová výbojka Kompaktní zářivka barvy 827 Kompaktní zářivka barvy 842 Kontrola Nárůst celkové listové plochy (včetně opadaných listů) Cyclamen 573,17 cm2 512,72 cm2 325,70 cm2 222,64 cm2 60
64 Počet listů Spatiphyllum Charakteristika Halogenová výbojka Kompaktní zářivka barvy 827 Kompaktní zářivka barvy 842 Kontrola Celkový počet opadaných listů Spatiphyllum Celkový počet nových listů Spatiphyllum Listová plocha Spatiphyllum Charakteristika Halogenová výbojka Kompaktní zářivka barvy 827 Kompaktní zářivka barvy 842 Kontrola Nárůst celkové listové plochy (včetně opadaných listů) Spatiphyllum 1292, , , ,70 Výška rostlin Výraznější změna výšky rostlin nastala pouze u Cyclamen bylo poprvé zaznamenáno výrazné prodloužení délky řapíků i stonků s květy o 3 až 4 cm. Nedošlo k němu u vyměněné rostliny pod zářivkou barvy 842, ale u původní rostliny, která byla použita jako kontrolní, k prodloužení došlo. Pravděpodobně byly rostliny ošetřeny retardanty a ty přestaly působit. 5.3 Modelové objekty návrhy řešení prostor s nevhodnými světelnými podmínkami Po dohodě s vedoucí práce byly vybrány 3 modelové objekty: specializovaná svítidla pro přisvětlování rostlin, vstupní prostor kolejí a vstupní hala budovy A Zahradnické fakulty v Lednici. Ve dvou případech je řešeno nedostatečné, v jednom případě nadbytečné osvětlení Vstupní hala budovy A V navrhovaném řešení se počítá se dvěma typy sesazovaných nádob, přičemž budou v prostoru umístěny tři kusy od každého typu. Všechny nádoby mají stříbrnou barvu. Větší nádoby Lechuza Cararo budou umístěny u stěny vedle dveří děkanátu, kde je minimální intenzita přirozeného světla, proto budou rostliny osvětlovány výkonnými svítidly HCI-PAR 30 35/942 NDL 30 D od firmy Osram (sestava Goliáš). K osázení těchto nádob byly zvoleny 61
65 druhy Codiaeum variegatum, Epipremnum pinnatum Neon a Ficus deltoidea. Všechny mají střední nároky na světlo, takže bez přisvětlování by zde být umístěny nemohly. Na druhou stranu nejsou vysoce náročné, takže by měly dobře prosperovat i pod umělým světlem. Tyto nádoby by měly být stabilně umístěny u stěny, nepočítá se s jejich přesouváním. Menší nádoby Lechuza Quadro 43 budou mobilní a počítá se s možností jejich přesouvání podle potřeb na využití haly. Po většinu času by měly být umístěny u stěny naproti stabilní výsadbě, kde je větší intenzita přirozeného světla, jež sem proniká dveřmi, které vedou přímo ven. Přesto tu budou umístěna svítidla k doplňkovému přisvětlování, protože intenzita přirozeného světla, zvláště v zimě je nedostatečná. Zvolena byla svítidla HCI-PAR 20 35/942 NDL 30 D (sestava David). K osázení mobilních nádob byly zvoleny druhy Schefflera arboricola Janine a Epipremnum pinnatum Marble queen. Tyto rostliny nemají příliš vysoké nároky na světlo, takže když budou občas přesunuty do horších světelných podmínek, nemělo by to nijak vadit. V současné době je místnost vymalovaná dosti tmavě, takže by bylo dobré stěny přemalovat světlejšími odstíny, které budou více odrážet světlo, a tím pádem bude místnost celkově působit světleji a rostliny více vyniknou. Pro tento návrh byly zvoleny barvy ve světle zeleném a světle oranžovém tónu, přičemž oranžové (RAL ) by měly být stěny s výsadbami a zelené (RAL ) 2 zbývající stěny. Momentálně jsou na lištách na stropě umístěna malá halogenová svítidla. V návrhu se počítá s jejich dalším využitím k osvětlování stolků se studentskými pracemi, které by se soustředily ke stěně bez výsadby vedle dveří ven Vstupní prostor kolejí Pro dlouhodobější ozelenění tohoto prostoru byly vybrány druhy Bougainvilea spectabilis a Trachycarpus fortunei, dále se v návrhu počítá s umístěním nádoby, ve které se bude výsadba obměňovat podle ročního období. Nádoby pro výsadbu byly zvoleny v barvě espresso, pro Trachycarpus a Bougainvilea je to nádoba Lechuza Classico 43, k sezónnímu osázení nádoba Lechuza Delta 20. Pro sezónní výsadbu byly vybrány cibuloviny (Narcisus, Hyacintus) - jaro, Impatiens walleriana - léto a Chrysanthemum - podzim. V zimě se počítá s tím, že se do nádoby umístí aranžmá z chvojí. Aby se zabránilo negativním vlivům přílišného přehřívání a nadbytku slunečního záření v létě, je nad nádobami s Bougainvilea spectabilis a se sezónní výsadbou navržena pohyblivá markýza. Trachycarpus bude umístěn blízko dveří ven, takže by zde mělo být větrání 62
66 dostatečně intenzivní a také se jedná o rostlinu, kterou je možné letnit venku, proto se zde se stínováním nepočítá. V zimě zde mohou být teploty velmi nízké. Trachycarpus by měl snést i teploty -5 C, ale před příchodem mrazů by měly být všechny rostliny přesunuty do skleníku a na jaře přeneseny zpět Kancelář firmy Spektrum Holas v Havlíčkově Brodě Cílem bylo vytvořit vzorové osázení nádob interiérovými rostlinami pro rozdílné typy svítidel a ukázat možnosti jejich využití v praxi. Svítidla se lišila jak svými vzhledovými vlastnostmi, tak intenzitou světla a typem světelného zdroje. Konkrétně bylo pracováno se svítidlem Klara (zdroj světla: halogenidová výbojka, intenzita světla: 10 mol*s -1 *m -2 ), Max (zdroj světla: 2 reflektorové světelné diody Master LED Spot Philips, intenzita světla 10 mol*s -1 *m -2 ) a Majel (zdroj světla: lineární zdroj Osram, intenzita světla 12 mol*s -1 *m -2 ) Pro každý světelný zdroj byly navrženy 2 varianty osázení, u nichž jedna byla zrealizována a nafocena, pro druhou byl vytvořen osazovací plán. Ve všech případech se u nerealizovaných variant počítá s použitím minerálního substrátu Seramis, u zrealizovaných a nafocených variant byl použit klasický zeminný substrát. Svítidlo Klára Toto svítidlo rovnoměrně osvětluje kruhovou plochu o průměru 0,5 m, proto byla pro první návrh zvolena nádoba Hydrocultur Gefässe Rund RAL 1013 (perlově bílá), která je také kruhová a má tento průměr. Její výška je 23 cm. Intenzita osvětlení je pod tímto svítidlem nejnižší (10 mol*s -1 *m -2 ), proto byly zvoleny rostliny s nižší náročností na světlo, kterým tato intenzita bude stačit. Jsou to Spatiphyllum sp., Dracena deremensis White stripe a Epipremnum pinnatum Marble Queen. Druhá, zrealizovaná, varianta využívá pouze část osvětlené plochy ve tvaru obdélníku 0,5 x 0,3 m. Na tuto plochu byly umístěny druhy Kalanchoe blossfeldiana, Codieum variegatum a Rhipsalis sp. Svítidlo Maks Toto svítidlo rovnoměrně osvětluje plochu 0,6 x 0,25 m. Těmto rozměrům odpovídá nádoba, zvolená pro první variantu návrhu, Vision RE weissaluminium (délka 60 cm, šířka 25 cm, výška 12 cm). Intenzita osvětlení je vyšší 15 mol*s -1 *m
67 K ozelenění této nádoby byly vybrány druhy Codieum variegatum, Guzmania hybr. a Aglaonema commutatum. Zrealizovaná varianta sestává z rostlin Dracena deremensis Yellow stripe, Cordyline stricta, Oxalis tuberosa, Fittonia albivensis a dvou Impatiens walleriana. Svítidlo Majel Intenzita osvětlení je o něco nižší než u předchozího svítidla, i když je použit také LED zdroj. Ozařovaná plocha je poměrně malá (pouze 0,3 x 0,2 m). Pod toto svítidlo byla v první variantě zvolena nádoba Lechuza Delta 20 stříbrná, která má sice délku 0,4 m, ale na okrajích budou umístěny na světelné záření méně náročné rostliny Hedera helix. Dále jsou v návrhu použity převislá Tradecantia albiflora Albovittata a kvetoucí Pentas lanceolata (2 ks). Zrealizovaná varianta je složena z rostlin Crossandra indibuliformis, Peperomia galuloides a Kalanchoe blossfeldiana. 64
68 6. DISKUZE Světelné záření je nejobtížněji ovlivnitelným faktorem při pěstování rostlin v interiéru a řešení nedostatku i nadbytku tohoto pěstebního faktoru je nejnákladnější. Nevhodné světelné podmínky se v současných interiérech vyskytují často, většinou během jednoho ročního období - zimy nebo léta. Existuje však široké spektrum možností, jak světelné podmínky vylepšit a tak umožnit rostlinám zdravý růst a vývoj během celého roku i na stanovištích s nevhodnými přirozenými světelnými podmínkami. Světelné záření je pro rostliny především zdrojem energie, ale kromě fotosyntetických má i další účinky, fotomorfogenetické a fotoperiodické. Působí na mnoho procesů v rostlinách, a proto je světelné záření jako pěstební faktor při pěstování interiérových rostlin nenahraditelné. Z výsledků hodnocení světelných podmínek ve vybraných interiérech vyplývá, že častějším problémem je nedostatek světla v zimě. Druhy pěstované na tmavých stanovištích jsou většinou vhodně zvoleny. Hodnocená velmi světlá stanoviště se nacházejí v nově postavených budovách s velkým podílem prosklených ploch. I na těchto stanovištích jsou druhy většinou umístěny vhodně. Optimální světelné podmínky byly zaznamenány pouze na 1 hodnoceném stanovišti. Zcela nevhodně zvolené druhy se na stanovištích nacházejí minimálně, ve většině případů umístění odpovídá úplně nebo částečně světelným podmínkám, které daný druh vyžaduje. Bohužel ve dvou případech jsou rostliny umístěny na zcela nevhodných velmi tmavých stanovištích. V kanceláři HB Auditing je pravděpodobné, že alespoň v létě budou podmínky o něco příznivější, přesto by bylo vhodné rostliny umístit blíže ke zdroji světla. V budově VZP je okno zakryto dřevěnou deskou, takže se zde světelné podmínky příliš nezlepší ani v létě. Jedná se o veřejnou instituci, kde by měla výsadba reprezentovat, proto by bylo nanejvýš vhodné sem umístit svítidla vhodná k přisvětlování rostlin s dostatečnou intenzitou, aby rostliny mohly plnit svoji funkci a působit příjemně, nikoli zuboženě. Pro řešení nedostatku přirozeného světelného záření existuje široký sortiment nejrůznějších umělých zdrojů světla. Ty se však značně liší svými vlastnostmi spektrálním složením, podáním barev, apod. Stává se, že i svítidla doporučovaná speciálně pro přisvětlování rostlin nejsou k tomuto účelu vhodná. Na internetu i v kamenných obchodech se můžeme setkat s žárovkami doporučovanými k přisvětlování rostlin, ale žárovky se pro přisvětlování rostlin příliš nehodí. Jsou málo účinné a hrozí riziko poškození rostlin teplem, které vydávají. Navíc jsou jejich prodej a výroba omezovány Evropskou unií a žárovky s vyššími příkony, které by se případně daly použít, už se nevyrábějí. Ani některé zářivky určené speciálně pro pěstování 65
69 rostlin nejsou vhodné pro své nepřirozené podání barev a neosvědčily se ani z hlediska pěstitelské praxe. Vhodné pro použití k osvětlování rostlin jsou bílé zářivky. Vhodnými zdroji jsou též halogenidové výbojky a také moderní zdroje LED. Halogenidová výbojka se velmi osvědčila i v pokusu s přisvětlováním rostlin. Halogenidové výbojky však potřebují předřadné zařízení, proto není možné je použít ve všech svítidlech. Zdroje LED mohou mít různé spektrum vyzařování podle použitého luminoforu. Jejich spektrum je spojité a mají vysoký index podání barev. Navíc mají nízkou spotřebu energie a dlouhou dobu života. Jsou to velmi perspektivní zdroje nejen pro přisvětlování rostlin a je pravděpodobné, že jejich sortiment se bude stále rozšiřovat. Zatím jsou poměrně nákladné, ale je pravděpodobné, že s rozšířením výroby cena klesne. Pokus s přisvětlováním rostlin v období s nepříznivými přirozenými světelnými podmínkami jednoznačně prokázal, že přisvětlování má velmi pozitivní vliv na růst a vývoj pozorovaných interiérových rostlin. Kontrolní neosvětlované rostliny prakticky zastavily svůj růst a vývoj, ale osvětlované rostliny dále přirůstaly a kvetly. Rozdíly mezi jednotlivými svítidly není možné tímto informativním pokusem prokázat, ale všechny světelné zdroje měly pozitivní vliv. Jen u zářivky barvy 842 docházelo k problémům s kvetením u Cyclamen, a i když byla původní pozorovaná rostlina vyměněna, problémy se opakovaly. Problematice nedostatku světelného záření při pěstování rostlin a možnostem řešení tohoto problému je věnováno poměrně velké množství literatury i v českém jazyce, naopak problematice nadbytku světelného záření a možnostem řešení je věnováno jen málo literárních zdrojů. V českém jazyce jsem ani žádný, který by se tomuto tématu věnoval, nenašla, čerpala jsem pouze ze zdrojů v němčině. Je však pravděpodobné, že i v České republice bude stanovišť s nadbytkem světelného záření s rozšiřující se výstavbou moderních prosklených staveb přibývat a proto by bylo dobré se podrobněji zabývat i možnostmi řešení tohoto problému. 66
70 7. ZÁVĚR Cílem práce bylo vytvořit přehled možností řešení nevhodných světelných podmínek z pohledu potřeb pro pěstování rostlin, podle navržené metodiky vyhodnotit stanoviště, kde jsou pěstovány interiérové rostliny a navrhnout konkrétní řešení pro modelové objekty. Podle navržené metodiky byla stanoviště v hodnocených interiérech rozdělena na velmi světlá, světlá, tmavá a velmi tmavá. Rostliny umístěné na těchto stanovištích byly rozděleny podle toho, jak odpovídá stanoviště jejich nárokům na světlo, do 3 kategorií vhodné, částečně vhodné a nevhodné. Z výsledků hodnocení vyplývá, že častějším problémem v naší geografické oblasti je nedostatek světla v zimě, ale v nových moderních prosklených stavbách je naopak pro rostliny limitující nadbytek světelného záření a přehřívání interiérů v létě. Nadbytek světla je méně častým problémem, je však pravděpodobné, že moderních prosklených budov bude přibývat, proto je důležité zabývat se i stíněním příliš exponovaných míst, aby i zde bylo možné bez problémů pěstovat interiérové rostliny. Rostlinné druhy na hodnocených stanovištích byly většinou vhodně zvoleny. Jako modelové objekty byly zvoleny: vstupní prostor ZF v Lednici, vstupní prostor kolejí ZF v Lednici a kancelář firmy Spektrum Holas v Havlíčkově Brodě. V rámci spolupráce na soukromém projektu Osvětlování interiérových rostlin byl založen informativní pokus k průběžnému sledování rostlin, osvětlovaných různými světelnými zdroji, s ohledem na jejich vývoj a kvalitativní parametry ve srovnání s rostlinami neosvětlovanými, který prokázal velmi pozitivní vliv přisvětlování v období s nepříznivými přirozenými světelnými podmínkami v zimě. Byly provedeny rozbory emisních spekter světelných zdrojů a vypočítány koeficienty pro převod fotometrických jednotek (lm) na fytotechnické jednotky (zářivý tok, W, fotonový tok, μmol*s -1 *m -2 ). Průběžně byly také zpracovávány podklady k článku pro časopis Světlo č.3/2011 a pro další článek, který bude uveřejněn v témže časopise. Na požádání garanta plánovaného semináře na téma světlo a rostliny v interiéru je připravována přednáška, která bude obsahovat výsledky prací na projektu. 67
71 . 8. RESUMÉ The main objective of this thesis is to provide a comprehensive outline of possible ways of dealing with inadequate lighting conditions of interiors in light of the needs of growing plants. Another goal was to evolve a methodology of the evaluation of lighting conditions, also to propose solutions for specific interiors with inadequate lighting conditions and finally to accomplish an experiment with lightening plants. The literature search describes the importance of light radiation for plants, its photosynthetic, fotomorfogenetic and fotoperiodic effects and also the relation of the light rays and other cultivative factors. Furthermore, there is presented a distinction of the interiors by the lighting conditions, the possibility of solving insufficiency or surplus of natural lighting in interiors and also plants suitable for interiors with these kinds of defects. For evaluating the light conditions in terms of growing indoor plants my own methodology have been developed. On grounds of this methodology 19 sites with interior plants have been interpreted. Possible solutions of these conditions were suggested for three model objects with inadequate lighting conditions including the suitable indoor plants and their containers. A lightening experiment was executed and proved very beneficial effect of lightening the indoors plants in winter period with worse natural light conditions. 68
72 9. POUŽITÁ LITERATURA BROOKES, J. Příjemný život s květinami. Pěstování a využití jejich barvy a tvaru při : úpravě bytového interiéru. 2. vyd. Bratislava: Príroda, s. ISBN BUESCHER, G. Svetlo a osvetlenie. 1. vyd. Bratislava: SVTL, s. Polytechnická knižnica. DOSTÁL. Světlomilné rostliny, In: MAREČEK, F. (editor) Zahradnický slovník naučný: R- Ž.. 1. vyd. Praha: Ústav zemědělských a potravinářských informací, s. ISBN DVOŘÁČEK, V. Světelné zdroje halogenidové výbojky (část 1), Světlo, č. 1, 2009, str [on-line] Dostupné z DVOŘÁČEK, V. Světelné zdroje halogenidové výbojky (část 2), Světlo, č. 2, 2009, str [on-line] Dostupné z DVOŘÁČEK, V. Světelné zdroje kompaktní zářivky, Světlo, č. 3, 2008, str. 43 [on-line] Dostupné z DVOŘÁČEK, V. Světelné zdroje lineární zářivky, Světlo, č. 2, 2008, str. 56 [on-line] Dostupné z DVOŘÁČEK, V. Světelné zdroje světelné diody, Světlo, č. 5, 2009, str. 69 [on-line] Dostupné z FLEHMIG, A. Pokojové rostliny: krok za krokem k zelenému ráji: jak na to. 1. české vyd. Praha: Svojtka & Co., s. ISBN GLOSER, J. Fyziologie rostlin. 2. rozš. vyd. Brno : Masarykova univerzita, 1998, 157 s. ISBN: Handbuch Innenraumbegrünung. 1. Vyd. Braunschweig: Thalacker Medien, 1998, 400 s. ISBN: HAŠ, S. FIKAROVÁ, L. Navrhování osvětlení pro interiérové rostliny. Světlo, č. 3, 2011 (v tisku) 69
73 HAŠ, S. HAJZLER, M. Fotosynteticky aktivní soustava ve skleníku Fata Morgana. Světlo, č. 4, str. 16 [on-line] dostupné z HAŠ, S. PAVLÍČKOVÁ, P. Osvětlování okrasných rostlin v interiérech, Světlo, č. 4,, str. 45 [on-line] dostupné z HAŠ, S. Světelné záření. Materiály k článku do časopisu Světlo, 2011 HAŠ, S. Zdroje světla pro osvětlování rostlin v interiérech. Materiály k článku do časopisu Světlo, 2011 HEJNÁK, V. Fyziologie rostlin. 1. vyd. Praha: Česká zemědělská univerzita v Praze, ISBN HLADKÝ L. Osvětlení z pohledu rostlin, Světlo, č. 4,, str. 42 [on-line] Dostupné z CHALUPSKÝ, L. Světlo a svítidla. 1. vyd. Praha: SNTL, s. Polytechnická knižnica. KERSTJENS, K. Richtlinie für die Planung, Ausführung und Pflege von Innenraumbegrünungen : Innenraumbegrünungsrichtlinie. Bonn: FLL, s. ISBN X KINCL, M. -- FAUSTUS, L. Základy fyziologie rostlin. 1. vyd. Praha: SPN, s. Učebnice pro vysoké školy. KUBÍN, Š. K definici fotosynteticky účinného záření. Informační zpravodaj Světelná technika, 1982/1983 KUBÍN, Š. Zdroje fotosynteticky účinného záření a metody jeho měření. 1.vyd. Praha: Academia, 1973, ISBN KUŤKOVÁ, T. Pěstování rostlin v současných interiérech, klasifikace interiéru pro potřeby zahradní a krajinářské tvorby. Materiály ke skriptům, 2006 MACHOVEC, J. a kol. Květiny v bytě. 1. vyd. Praha: Státní zemědělské nakladatelství, s. ISBN MAREČEK, F. Stínomilné rostliny, In: MAREČEK, F. (editor) Zahradnický slovník naučný: 70
74 R-Ž. 1. vyd. Praha: Ústav zemědělských a potravinářských informací, s. ISBN MARTÍNEK, J. Texty k probíraným rostlinám. Materiály k předmětu Interiérové květinářství, MASAROVIČOVÁ, E. Fyziológia rastlín. 2., dopl. Vyd. Bratislava: vydavateľstvo uk, s. Isbn: MATOUŠ M. HUTLA P. Světlo a rostlina. Světlo, č. 4, 2002, str. 14 [on-line] Dostupné z MUSILOVÁ, M. Světelné zdroje a ergonomie vidění. Brno, Bakalářská práce). Vysoká škola. Masarykova univerzita. Lékařská fakulta. Vedoucí práce: MUDr. Zdeňka Mašková [on-line] Dostupné z is.muni.cz/th/214921/lf_b/bakalarska_prace_musilova.doc PAVLOVÁ, L. Fyziologie rostlin. 1. vyd. Praha: Karolinum, s. Učební texty Univerzity Karlovy v Praze. ISBN PODEŠVA, J. -- KUBÍN, Š. -- VÉBER, K. Využití umělého světla při pěstování rostlin. 1. vyd. Praha: SZN, s. PROCHÁZKA, S. a kol. Fyziologie rostlin. 1. vyd. Praha: Academia, s. ISBN ŠEBÁNEK, J. Fyziologie rostlin. 1. vyd. Praha: SZN, s. TŮMA, J. Fyziologie rostlin. 1. Vyd. Hradec Králové : Gaudeamus, 1998, 266 s. ISBN: VOLF. Stínování, In: MAREČEK, F. (editor) Zahradnický slovník naučný: R-Ž vyd. Praha: Ústav zemědělských a potravinářských informací, s. ISBN VOLF, M. -- VOTRUBA, R. Základy skleníkového květinářství. 1. vyd. Praha: Brázda, s. ISBN ZABLOUDILOVÁ, L. Pokojové rostliny a jejich uplatnění ve veřejném interiéru. Diplomová práce. Lednice: MZLU v Brně, s. 71
75 10. PŘÍLOHY Příloha I Tabulky a obrázky k literární části Příloha II Ukázky spekter světelných zdrojů Příloha III - Tabulky hodnocení světelných podmínek ve vybraných interiérech pro potřeby pěstování rostlin Příloha IV Přisvětlování rostlin grafy s výsledky pokusu Příloha grafy 1 počet květů Příloha grafy 2 počet listů Příloha graf 3 přirozené osvětlení v průběhu pokusu Příloha V - Přisvětlování rostlin fotografická příloha k pokusu Příloha VI Modelové objekty Nákres č. 1 vstupní hala ZF v Lednici Nákres č. 2 Vstupní prostor kolejí ZF v Lednici Nákres č. 3 Kancelář firmy Spektrum Holas v Havlíčkově Brodě - svítidlo Klára Nákres č. 4 - Kancelář firmy Spektrum Holas v Havlíčkově Brodě svítidlo Maks Nákres č. 5 - Kancelář firmy Spektrum Holas v Havlíčkově Brodě - svítidlo Majel Příloha VII výpočty Příklad výpočtu převodního koeficientu Postup výpočtů při navrhování osvětlování pro interiérové rostlin 72
 Obrázek 2 (Hladký,")
76 Příloha I Tabulky a obrázky k literární části Obrázek 1 Fyziologické účinky záření (Kubín, 1973) Obrázek 2 (Hladký, 2009)
 Obrázek 5 Dlouhodobé průměry denního množství globálního záření (Volf, Votruba,")
77 Obrázek 3 Možnosti přeměny excitační energie (Pavlová, 2005) Obrázek 4 (Pavlová, 2005) Obrázek 5 Dlouhodobé průměry denního množství globálního záření (Volf, Votruba, 1991)
 Obrázek 8 Různé způsoby měření intenzity záření (Hladký,")
78 Obrázek 6 Počet dní bez slunečního svitu v zimních měsících (Podešva, Véber, Kubín, 1968) Obrázek 7 Délka dne v jednotlivých měsících (Volf, Votruba 1968) Obrázek 8 Různé způsoby měření intenzity záření (Hladký, )
79 Tabulka 1




")
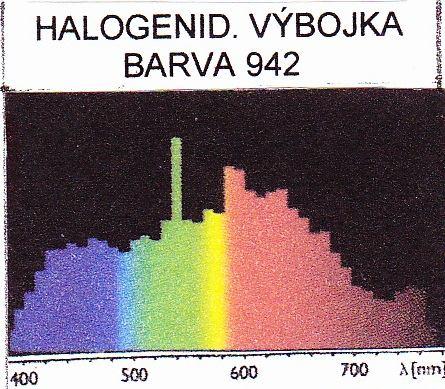
80 Příloha II Ukázky spekter světelných zdrojů Zářivky (Haš, 2011) Vysokotlaké sodíkové výbojky Halogenidové výbojky (Haš, Hajzler, 2008) (Haš, Hajzler, 2008) (Haš, 2011) Elektroluminiscenční zdroje Denní světlo (Dvořáček, 2009) (Haš, 2011)

81 Příloha III - Tabulky hodnocení světelných podmínek ve vybraných interiérech pro potřeby pěstování rostlin Popis a lokalizace interiérů Foto Nemocnice Havlíčkův Brod, 5. Patro Husova 2624 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí SV U oken: Hibiscus rosa sinensis, Cissus rhombifolia, Ficus benjamina, Synadenium grantii U sloupu: Pandanus weitchii, Dracena deremensis Průměrná hodnota: U oken: 2048,75 lx U sloupu: 456,875 lx Zařazení do skupiny: U oken: 1 U sloupu: 3 U oken: Hibiscus rosa sinensis vhodné umístění Cissus rhombifolia nevhodné umístění Ficus benjamina částečně vhodné umístění Synadenium grantii vhodné umístění U sloupu: Dracena deremensis vhodné umístění Pandanus weitchii částečně vhodné umístění Datum měření Podmínky při měření J J P P P Z P J J Z Naměřená hodnota v luxech u oken Naměřená hodnota v luxech u sloupu

82 Popis a lokalizace interiérů Foto Vstupní prostor budovy A Zahradnické fakulty v Lednici, přízemí Valtická 337 Lednice Typ interiéru Interiér s převládající shromažďovací funkcí Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek JZ, PO Aglaonema commutatum, Schefflera arboricola Průměrná hodnota: 1039,625 lx Zařazení do skupiny: 3 Aglaonema commutatum vhodné umístění Schefflera arboricola částečně vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J Z P P Z P P J P

83 Popis a lokalizace interiérů Foto Prosklená hala budovy D Zahradnické fakulty v Lednici, přízemí Valtická 337 Lednice Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí JZ, H Xantorea sp., Olea europaea, Dasylirion longissimum Průměrná hodnota: 7751,25 lx Zařazení do skupiny: 1 Xantorea sp. vhodné umístění Olea europaea vhodné umístění Dasylirion longissimum vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J Z P P Z P P J P






84 Popis a lokalizace interiérů Foto Nádraží Podivín, přízemí Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí JZ Na parapetu: Asparagus densiflorus Sprengeri, Synadenium grantii, Epipremnum pinnatum, Begonia coralina Luzerna, Hippeastrum sp. U zdi: Synadenium grantii, Begonia coralina Luzerna, Dieffenbachia maculata, Philodendron erubescens, Chlorophytum comosum Průměrná hodnota: Na parapetu: 1302, 75 lx U zdi: 420 lx Zařazení do skupiny: Na parapetu: 2 U zdi: 3 Na parapetu: Synadenium grantii částečně vhodné umístění Asparagus densiflorus Sprengeri vhodné umístění Epipremnum pinnatum částečně vhodné umístění Begonia coralina Luzerna - vhodné umístění Hippeastrum sp. částečně vhodné umístění U sloupu: Synadenium grantii nevhodné umístění Begonia coralina Luzerna částečně vhodné umístění Dieffenbachia maculata vhodné umístění Philodendron erubescens vhodné umístění Chlorophytum comosum vhodné umístění Datum měření Podmínky při měření J J P P P Z P P P J Naměřená hodnota v luxech u oken Naměřená hodnota v luxech u sloupu



85 Popis a lokalizace interiérů Foto Menza Zahradnické fakulty v Lednici, přízemí Valtická 337 Lednice Typ interiéru Expozice interiéru Zeleň použitá v interiéru Interiér s převládající společenskou funkcí JZ Epipremnum pinnatum Neon, Euphorbia trigona, Sanseviera cylindrica, Aloe vera Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Průměrná hodnota: lx Zařazení do skupiny: 1 Epipremnum pinnatum Neon částečně vhodné umístění Euphorbia trigona vhodné umístění Sanseviera cylindrica vhodné umístění Aloe vera vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J Z P P Z P P J P

86 Popis a lokalizace interiérů Foto Okno v knihovně Zahradnické fakulty v Lednici, přízemí Valtická 337 Lednice Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí SV Euphorbia tirucalli, Zamioculcas zamifolia, Chlorophytum commosum, Nephrolepis exaltata, Sanseviera parva Průměrná hodnota: 1055,125 lx Zařazení do skupiny: 3 Euphorbia tirucalli nevhodné umístění Zamioculcas zamifolia vhodné umístění Chlorophytum comosum vhodné umístění Nephrolepis exaltata částečně vhodné umístění Sanseviera parva vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J Z P P Z P P J P

87 Popis a lokalizace interiérů Foto Galerie Vaňkovka přízemí Ve Vaňkovce 1 Brno Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Multifunkční interiér - obchody, obchodní centra, obchodní pasáže H Schefflera arboricola, Stromante sp., Nephrolepis exaltata, Codieum variegatum, Chamaedorea elegans, Ficus benjamina, Anthurium sp., Dracena deremensis, Epipremnum pinnatum Průměrná hodnota: 517,125 Zařazení do skupiny: 3 Schefflera arboricola částečně vhodné umístění Stromante sp. částečně vhodné umístění Nephrolepis exaltata částečně vhodné umístění Codiaeum variegatum - částečně vhodné umístění Chamaedorea elegans vhodné umístění Ficus benjamina částečně vhodné umístění Anthurium sp. - částečně vhodné umístění Dracena deremensis vhodné umístění Epipremnum pinnatum vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P P Z J
88 Popis a lokalizace interiérů Foto Galerie Vaňkovka, východ v 1. patře Ve Vaňkovce 1 Brno Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Multifunkční interiér - obchody, obchodní centra, obchodní pasáže Schefflera actinophylla, Epipremnum pinnatum, Chamaedorea elegans Průměrná hodnota: 511,5 lx Zařazení do skupiny: 3 Schefflera actinophylla vhodné umístění Epipremnum pinnatum vhodné umístění Chamaedorea elegans vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P P Z J

89 Popis a lokalizace interiérů Foto Dům Pánů z Lipé 3. Patro Náměstí Svobody 17 Brno Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Multifunkční interiér - obchody, obchodní centra, obchodní pasáže H Spatiphyllum sp., Hedera helix Průměrná hodnota: 439,125 Zařazení do skupiny: 3 Spatiphyllum sp. vhodné umístění Hedera helix částečně vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P P Z J


90 Popis a lokalizace interiérů Foto Všeobecná zdravotní pojišťovna, přízemí Havlíčkovo nám. 170 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí Z Zamioculcas zamifolia, Epipremnum pinnatum, Spatiphylum sp. Průměrná hodnota: 116 lx Zařazení do skupiny: 4 Zamioculcas zamifolia částečně vhodné umístění Epipremnum pinnatum částečně vhodné umístění Spatiphylum sp. - částečně vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P J Z Z

91 Popis a lokalizace interiérů Foto Chodba před kanceláří Metal Aliance, 2. Patro Žižkova 280 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí J Sanseviera trifasciata, Aspidistra elatior Průměrná hodnota: 406,375 lx Zařazení do skupiny: 3 Sanseviera trifasciata vhodné umístění Aspidistra elatior vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P J Z Z

92 Popis a lokalizace interiérů Foto Česká pošta, 1. Patro Svatovojtěšská 58 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí J Phoenix canariensis Průměrná hodnota: 404,375 Zařazení do skupiny: 3 Phoenix canariensis částečně vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P J Z Z

93 Popis a lokalizace interiérů Foto Poliklinika, 4. Patro Nádražní 582 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiér s převládající shromažďovací funkcí JV Dracena deremensis, Ficus lyrata, Ficus benjamina, Dieffenbachia maculata, Chlorophytum comosum, Epipremnum pinnatum Průměrná hodnota: 473,125 lx Zařazení do skupiny: 3 Dracena deremensis vhodné umístění Ficus lyrata vhodné umístění Ficus benjamina částečně vhodné umístění Dieffenbachia maculata vhodné umístění Chlorophytum comosum vhodné umístění Epipremnum pinnatum vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P Z P Z P P Z Z

94 Popis a lokalizace interiérů Foto Kancelář HB Auditing, 1. patro Na Valech 154 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Pracovní interiér V Spatiphyllum sp., Radermachera sinica Průměrná hodnota: 222,5 lx Zařazení do skupiny: 4 Radermachera sinica nevhodné umístění Spatiphyllum sp. částečně vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P P Z Z P J J Z

95 Popis a lokalizace interiérů Foto Rodinný dům, 1. Patro U Stadionu 2009 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Soukromý S, H Spatiphyllum sp., Zamioculcas zamifolia, Monstera deliciosa, Schefflera arboricola, Dieffenbachia maculata, Anthurium andreanum, Adiantum radianum, Clivia miniata, Dracena deremensis Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Průměrná hodnota: 647,5 lx Zařazení do skupiny: 3 Spatiphyllum sp. vhodné umístění Zamioculcas zamifolia vhodné umístění Monstera deliciosa vhodné umístění Schefflera arboricola částečně vhodné umístění Dieffenbachia maculata vhodné umístění Anthurium andreanum nevhodné umístění Adiantum radianum vhodné umístění Clivia miniata vhodné umístění Dracena deremensis vhodné umístění Datum měření Podmínky při měření Naměřená hodnota v luxech J J P P Z P P Z J Z

96 Popis a lokalizace interiérů Foto Plavecký bazén, přízemí U Stadionu 3000 Havlíčkův Brod Typ interiéru Expozice interiéru Zeleň použitá v interiéru Vyhodnocení interiéru dle osvětlenosti Hodnocení vhodnosti použití rostlin z hlediska světelných podmínek Interiéry určené k vykonávání různých sportovních, rekreačních a relaxačních činností J U oken: Nerium oleander, Ficus benjamina U zdi: Ficus elastica, Monstera deliciosa, Araucaria heterophylla Průměrná hodnota: U oken: 5291,375 lx U zdi: 2902,25 lx Zařazení do skupiny: U oken: 1 U zdi: 1 U oken: Nerium oleander vhodné umístění Ficus benjamina částečně vhodné umístění U zdi: Ficus elastica nevhodné umístění Monstera deliciosa nevhodné umístění Araucaria heterophylla - vhodné umístění Datum měření Podmínky při měření J J P Z Z P P J J J Naměřená hodnota v luxech u oken Naměřená hodnota v luxech u zdi
97 Příloha IV Přisvětlování rostlin grafy s výsledky pokusu
98 Příloha grafy 1 Počet květů Spatiphyllum Halogenová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
99 Počet květů Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 ( provedena výměna rostliny.) Kontrola
100 Příloha grafy 2 Počet listů Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
101 Počet listů Spatiphyllum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
102 Počet listů Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
103 Příloha graf 3
104 Příloha V - Přisvětlování rostlin fotografická příloha k pokusu




105 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




106 Spatiphyllum Halogenidová výbojka Kontrola Zářivka barvy 842 Zářivka barvy 827




107 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




108 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola



109 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola




110 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola


111 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola



112 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola




113 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




114 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




115 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola




116 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
117 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola



118 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola
119 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




120 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




121 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola



122 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola



123 Pozorování Cyclamen Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




124 Spatiphyllum Halogenidová výbojka Zářivka barvy 842 Zářivka barvy 827 Kontrola



125 Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola




126 Kresba na listech - Codieum Halogenidová výbojka Zářivka barvy 827 Zářivka barvy 842 Kontrola
127 Příloha VI Modelové objekty Nákres č. 1 Vstupní prostor ZF v Lednici
128
129

130


 Nádoba: Hydrocultur")
131 Nákres č. 3 kancelář ve firmě Spektrum Holas svítidlo Klára Foto Návrh osázení k fotografii: Legenda Další varianta osázení: Legenda bílá) Nádoba: Hydrocultur Gefässe Rund RAL 1013 (perlově




132 Nákres č. 4 - Kancelář firmy Spektrum Holas v Havlíčkově Brodě svítidlo Maks Foto Návrh osázení k fotografii: Legenda Další varianta osázení: Legenda Nádoba: Vision RE weissaluminium




133 Nákres č. 5 - Kancelář firmy Spektrum Holas v Havlíčkově Brodě svítidlo Majel Foto Návrh osázení k fotografii: Legenda Další varianta osázení: Legenda Nádoba: Lechuza Delta 20 stříbrná
134 Příloha VII výpočty Příklad výpočtu převodního koeficientu K (F) K (F) mm F [mm* K (F)] F [mm*k (F)] 380 0,027 3,176 1,000 0,027 3, ,082 3,260 1,000 0,082 3, ,272 3,343 1,000 0,272 3, ,816 3,427 3,000 2,448 10, ,720 3,510 7,000 19,040 24, ,888 3,594 20, ,760 71, ,640 3,678 40, , , ,840 3,762 34, , , ,800 3,845 23, ,400 88, ,880 3,928 14, ,012 58, ,520 4,012 14, ,184 56, ,400 4,096 18, ,340 74, ,600 4,179 23, ,800 96, ,000 4,263 21, ,600 92, ,800 4,363 20, ,560 88, ,200 4,430 31, , , ,700 4,514 33, , , ,600 4,597 34, , , ,600 4,681 35, , , ,400 4,764 35, , , ,600 4,848 36, , , ,800 4,931 36, , , ,100 5,015 36, , , ,000 5,098 35, , , ,100 5,182 33, , , ,200 5,266 30, , , ,000 5,439 26, , , ,760 5,433 23, , , ,480 5,516 19, , , ,760 5,600 15, ,632 87, ,560 5,684 13, ,280 73, ,576 5,767 11,000 61,336 63, ,788 5,851 8,800 24,534 51, ,428 5,934 6,500 9,282 38, ,717 6,018 5,000 3,585 30, ,354 6,102 3,800 1,345 23, ,170 6,185 2,800 0,476 17, ,082 6,269 2,200 0,180 13, ,041 6,352 1,500 0,062 9, ,020 6,436 1,000 0,020 6, ,005 6,519 1,000 0,005 6,519 FAR , , ,467 FPR , , ,661 61,64% 1 klm = 3,229W 1 W = 0,310 klm 1 klm = 15,033 mol/s 1 mol/s = 0,067 klm 1 W = 4,65 mol/s 1 mol/s = 0,215 W



135 Spektrum k výpočtu Postup výpočtů při navrhování osvětlování pro interiérové rostliny
136

137
Vyjádření fotosyntézy základními rovnicemi
 FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
FOTOSYNTÉZA Fotochemický proces, při němž fotosynteticky aktivní pigmenty v zelených částech rostlin přijímají energii světelného záření a přeměňují ji na energii chemickou. Ta je dále využita při biologických
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
Fotosyntéza (2/34) = fotosyntetická asimilace
 = fotosyntetická asimilace") Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
Fotosyntéza (2/34) = fotosyntetická asimilace FOTO - protože k fotosyntéze je třeba fotonů Jedná se tedy o zachycování sluneční energie a přeměnu jednoduchých anorganických látek (CO 2 a H 2 O) na složitější
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
FOTOSYNTÉZA Správná odpověď:
 FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1
 FOTOSYNTÉZA Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1 Fotosyntéza (z řec. phos, photós = světlo) je anabolický děj probíhající u autotrofních organismů (řasy,
FOTOSYNTÉZA Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_07_BI1 Fotosyntéza (z řec. phos, photós = světlo) je anabolický děj probíhající u autotrofních organismů (řasy,
Fotosyntéza Ekofyziologie. Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni
 Fotosyntéza Ekofyziologie Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Fyziologické a ekologické aspekty fotosyntézy vliv stresů a proměnného prostředí na fotosyntézu; mechanismy
Fotosyntéza Ekofyziologie Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Fyziologické a ekologické aspekty fotosyntézy vliv stresů a proměnného prostředí na fotosyntézu; mechanismy
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
FOTOSYNTÉZA. Princip, jednotlivé fáze
 FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
FOTOSYNTÉZA Princip, jednotlivé fáze FOTOSYNTETICKÉ PIGMENTY - chlorofyl a modrozelený - chlorofyl b žlutozelený + karoteny, xantofyly žluté a oranžové zbarvení CHLOROFYL a, b CHLOROFYL a - nejdůležitější
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FOTOBIOLOGICKÉ POCHODY
 FOTOBIOLOGICKÉ POCHODY Základním zdrojem energie nutné pro život na Zemi je sluneční záření. Většina pochodů souvisí s přímým využitím zářivé energie pro metabolické pochody nebo pro orientaci organizmu
FOTOBIOLOGICKÉ POCHODY Základním zdrojem energie nutné pro život na Zemi je sluneční záření. Většina pochodů souvisí s přímým využitím zářivé energie pro metabolické pochody nebo pro orientaci organizmu
VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV
 VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV 1 Vladimír Špunda, 2 Otmar Urban, 1 Martin Navrátil 1 Přírodovědecká fakulta, Ostravská univerzita v Ostravě,
VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV 1 Vladimír Špunda, 2 Otmar Urban, 1 Martin Navrátil 1 Přírodovědecká fakulta, Ostravská univerzita v Ostravě,
ení k tvorbě energeticky bohatých organických sloučenin
 Fotosyntéza mimořádně významný proces, využívající energii slunečního zářenz ení k tvorbě energeticky bohatých organických sloučenin (sacharidů) z jednoduchých anorganických látek oxidu uhličitého a vody
Fotosyntéza mimořádně významný proces, využívající energii slunečního zářenz ení k tvorbě energeticky bohatých organických sloučenin (sacharidů) z jednoduchých anorganických látek oxidu uhličitého a vody
1- Úvod do fotosyntézy
 1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
1- Úvod do fotosyntézy Prof. RNDr. Petr Ilík, Ph.D. KBF a CRH, PřF UP FS energetická bilance na povrch Země dopadá 2/10 10 energie ze Slunce z toho 30% odraz do kosmu 47% teplo 23% odpar vody 0.02% pro
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Autor: Katka Téma: fyziologie (fotosyntéza) Ročník: 1.
 Ročník: 1.") Fyziologie Fotosyntéza Celým názvem: fotosyntetická asimilace - vznikla při ohrožení, že již nebudou anorg. l. rostliny začaly dělat fotosyntézu v atmosféře vzrostl počet O 2 = 1. energetická krize - nejdůležitější
Fyziologie Fotosyntéza Celým názvem: fotosyntetická asimilace - vznikla při ohrožení, že již nebudou anorg. l. rostliny začaly dělat fotosyntézu v atmosféře vzrostl počet O 2 = 1. energetická krize - nejdůležitější
FOTOSYNTÉZA. soubor chemických reakcí,, probíhaj v rostlinách a sinicích. z CO2 a vody jediný zdroj kyslíku ku pro život na Zemi
 Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
Fotosyntéza FOTOSYNTÉZA soubor chemických reakcí,, probíhaj hajících ch v rostlinách a sinicích ch zachycení a využit ití sluneční energie k tvorbě složitých chemických sloučenin z CO2 a vody jediný zdroj
aneb Fluorescence chlorofylu jako indikátor stresu
 Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
Měření fotosyntézy rostlin pomocí chlorofylové fluorescence aneb Fluorescence chlorofylu jako indikátor stresu Fotosyntéza: Fotosyntéza je proces, ve kterém je světelná energie zachycena světlosběrnými
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/ Anotace. Fotosyntéza světelná fáze. VY_32_INOVACE_Ch0214.
 Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
FOTOSYNTÉZA. CO 2 a vody. - soubor chemických reakcí. - probíhá v rostlinách a sinicích. - zachycení a využití světelné energie
 Fotosyntéza FOTOSYNTÉZA - soubor chemických reakcí - probíhá v rostlinách a sinicích - zachycení a využití světelné energie - tvorba složitějších chemických sloučenin z CO 2 a vody - jediný zdroj kyslíku
Fotosyntéza FOTOSYNTÉZA - soubor chemických reakcí - probíhá v rostlinách a sinicích - zachycení a využití světelné energie - tvorba složitějších chemických sloučenin z CO 2 a vody - jediný zdroj kyslíku
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
II. Rostlina a energie
 1 II. Rostlina a energie 2. Energie, přeměna látek, sluneční záření Živé organismy mají vysoký stupeň uspořádanosti a univerzálně spějí k rovnovážnému stavu. Životní procesy je nutné udržovat stálým dodáváním
1 II. Rostlina a energie 2. Energie, přeměna látek, sluneční záření Živé organismy mají vysoký stupeň uspořádanosti a univerzálně spějí k rovnovážnému stavu. Životní procesy je nutné udržovat stálým dodáváním
Fyziologie rostlin. 9. Fotosyntéza část 1. Primární fáze fotosyntézy. Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014
 Fyziologie rostlin 9. Fotosyntéza část 1. Primární fáze fotosyntézy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Fotosyntéza 1. část - úvod - chloroplasty - sluneční záření -
Fyziologie rostlin 9. Fotosyntéza část 1. Primární fáze fotosyntézy Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Fotosyntéza 1. část - úvod - chloroplasty - sluneční záření -
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
ZÁKLADNÍ FOTOMETRICKÉ VELIČINY
 ZÁKLADNÍ FOTOMETRICKÉ VELIČINY Ing. Petr Žák VÝVOJ ČLOVĚKA vývoj člověka přizpůsobení okolnímu prostředí (adaptace) příjem informací o okolním prostředí smyslové orgány rozhraní pro příjem informací SMYSLOVÉ
ZÁKLADNÍ FOTOMETRICKÉ VELIČINY Ing. Petr Žák VÝVOJ ČLOVĚKA vývoj člověka přizpůsobení okolnímu prostředí (adaptace) příjem informací o okolním prostředí smyslové orgány rozhraní pro příjem informací SMYSLOVÉ
2) Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.
 Povětrnostní činitelé studují se v ovzduší atmosféře (je to..) Meteorologie je to věda... Počasí. Meteorologické prvky. Zjišťují se měřením.") Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Pracovní list č. 2 téma: Povětrnostní a klimatičtí činitelé část. 1 Obsah tématu: Obsah tématu: 1) Vlivy působící na rostlinu 2) Povětrnostní činitelé a pojmy související s povětrnostními činiteli 3) Světlo
Energie fotonů je předávána molekulám chlorofylu A, který se zachyceným fotonem excituje (uvolní se energeticky bohatý elektron).
.") Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
Otázka: Fotosyntéza a biologické oxidace Předmět: Biologie Přidal(a): Ivana Černíková FOTOSYNTÉZA = fotosyntetická asimilace: Jediný proces, při němž vzniká v přírodě kyslík K přeměně jednoduchých látek
ROSTLINNÁ BUŇKA A JEJÍ ČÁSTI
 Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Gymnázium a Střední odborná škola pedagogická, Čáslav, Masarykova 248 M o d e r n í b i o l o g i e reg. č.: CZ.1.07/1.1.32/02.0048 TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Gymnázium, Ústí nad Orlicí, T. G. Masaryka 106 Seminář biologie Školní rok 2017/2018. Etiolované rostliny. Seminární práce. Lucie Fišarová (3.
 Gymnázium, Ústí nad Orlicí, T. G. Masaryka 106 Seminář biologie Školní rok 2017/2018 Etiolované rostliny Seminární práce Lucie Fišarová (3. A) Obsah 0. Úvod... 3 1. Etiolované rostliny... 4 1.1. Charakteristika...
Gymnázium, Ústí nad Orlicí, T. G. Masaryka 106 Seminář biologie Školní rok 2017/2018 Etiolované rostliny Seminární práce Lucie Fišarová (3. A) Obsah 0. Úvod... 3 1. Etiolované rostliny... 4 1.1. Charakteristika...
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
12-Fotosyntéza FRVŠ 1647/2012
 C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
C3181 Biochemie I 12-Fotosyntéza FRVŠ 1647/2012 Petr Zbořil 10/6/2014 1 Obsah Fotosyntéza, světelná fáze. Chlorofyly, struktura fotosyntetického centra. Komponenty přenosu elektronů (cytochromy, chinony,
Fotosyntéza. Ondřej Prášil
 Fotosyntéza 2 Ondřej Prášil prasil@alga.cz 384-340430 Obsah přednášky membrány a organely světlo termodynamika historie Fotosyntetické membrány Electron tomography Cells contain ~100 chlorosomes appressed
Fotosyntéza 2 Ondřej Prášil prasil@alga.cz 384-340430 Obsah přednášky membrány a organely světlo termodynamika historie Fotosyntetické membrány Electron tomography Cells contain ~100 chlorosomes appressed
LI-6400; Gazometrická stanovení fotosyntetických parametrů
 LI-6400; Gazometrická stanovení fotosyntetických parametrů 19. dubna 2011 Úvod List fixuje ve fotosyntéze CO 2, který proudí z vnější atmosféry. Současně ale uvolňuje CO 2 při respiračních pochodech. Na
LI-6400; Gazometrická stanovení fotosyntetických parametrů 19. dubna 2011 Úvod List fixuje ve fotosyntéze CO 2, který proudí z vnější atmosféry. Současně ale uvolňuje CO 2 při respiračních pochodech. Na
14. Fyziologie rostlin - fotosyntéza, respirace
 14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
14. Fyziologie rostlin - fotosyntéza, respirace Metabolismus -přeměna látek a energií (informací) -procesy: anabolický katabolický autotrofie Anabolismus heterotrofie Autotrofní organismy 1. Chemoautotrofy
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková
 Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Název: POZOROVÁNÍ PLASTIDŮ,VAKUOL, BUNĚČNÉ STĚNY Autor: Paed.Dr.Ludmila Pipková Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět: biologie Mezipředmětové vztahy: ekologie Ročník: 2.a 3.
Úvod do biologie rostlin Úvod PŘEHLED UČIVA
 Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Faktory počasí v ekologii - úvod
 Faktory počasí v ekologii - úvod Jakub Brom Laboratoř aplikované ekologie ZF JU Z ekologického hlediska nás zajímá, jak působí faktory počasí na organismy a zpětně, jak organismy působí na změnu těchto
Faktory počasí v ekologii - úvod Jakub Brom Laboratoř aplikované ekologie ZF JU Z ekologického hlediska nás zajímá, jak působí faktory počasí na organismy a zpětně, jak organismy působí na změnu těchto
Biosyntéza sacharidů 1
 Biosyntéza sacharidů 1 S a c h a r id y p o tr a v y (š k r o b, g ly k o g e n, sa c h a r o sa, a j.) R e z e r v n í p o ly sa c h a r id y J in é m o n o sa c h a r id y Trávení (amylásy - sliny, pankreas)
Biosyntéza sacharidů 1 S a c h a r id y p o tr a v y (š k r o b, g ly k o g e n, sa c h a r o sa, a j.) R e z e r v n í p o ly sa c h a r id y J in é m o n o sa c h a r id y Trávení (amylásy - sliny, pankreas)
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Fotosyntéza
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Molekulová spektroskopie 1. Chemická vazba, UV/VIS
 Molekulová spektroskopie 1 Chemická vazba, UV/VIS 1 Chemická vazba Silová interakce mezi dvěma atomy. Chemické vazby jsou soudržné síly působící mezi jednotlivými atomy nebo ionty v molekulách. Chemická
Molekulová spektroskopie 1 Chemická vazba, UV/VIS 1 Chemická vazba Silová interakce mezi dvěma atomy. Chemické vazby jsou soudržné síly působící mezi jednotlivými atomy nebo ionty v molekulách. Chemická
Ing. Pavel Hrzina, Ph.D. - Laboratoř diagnostiky fotovoltaických systémů Katedra elektrotechnologie K13113
 Sluneční energie, fotovoltaický jev Ing. Pavel Hrzina, Ph.D. - Laboratoř diagnostiky fotovoltaických systémů Katedra elektrotechnologie K13113 1 Osnova přednášky Slunce jako zdroj energie Vlastnosti slunečního
Sluneční energie, fotovoltaický jev Ing. Pavel Hrzina, Ph.D. - Laboratoř diagnostiky fotovoltaických systémů Katedra elektrotechnologie K13113 1 Osnova přednášky Slunce jako zdroj energie Vlastnosti slunečního
INTELIGENTNÍ KULTIVACE ROSTLIN
 Grow Light 300 INTELIGENTNÍ KULTIVACE ROSTLIN Lightdrop Grow Light je unikátní osvětlovací LED systém s možností regulace spektrálního složení a intenzity světla umožňující indukci intenzivního růstu a
Grow Light 300 INTELIGENTNÍ KULTIVACE ROSTLIN Lightdrop Grow Light je unikátní osvětlovací LED systém s možností regulace spektrálního složení a intenzity světla umožňující indukci intenzivního růstu a
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
INTELIGENTNÍ KULTIVACE ROSTLIN
 Grow Light 300 INTELIGENTNÍ KULTIVACE ROSTLIN Lightdrop Grow Light je unikátní osvětlovací LED systém s možností regulace spektrálního složení a intenzity světla umožňující indukci intenzivního růstu a
Grow Light 300 INTELIGENTNÍ KULTIVACE ROSTLIN Lightdrop Grow Light je unikátní osvětlovací LED systém s možností regulace spektrálního složení a intenzity světla umožňující indukci intenzivního růstu a
BIOLOGICKÁ MEMBRÁNA Prokaryontní Eukaryontní KOMPARTMENTŮ
 BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
BIOMEMRÁNA BIOLOGICKÁ MEMBRÁNA - všechny buňky na povrchu plazmatickou membránu - Prokaryontní buňky (viry, bakterie, sinice) - Eukaryontní buňky vnitřní členění do soustavy membrán KOMPARTMENTŮ - za
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO. Modelová rostlina suchozemská semenná neukončený růst specializované části
 Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Fotosyntéza Světelné reakce. Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni
 Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
Fotosyntéza Světelné reakce Ondřej Prášil Mikrobiologický ústav AVČR Laboratoř fotosyntézy v Třeboni Literatura Plant Physiology (L.Taiz, E.Zeiger), kapitola 7 pdf verze na požádání www.planthys.net Fotosyntéza
Škola: Gymnázium, Brno, Slovanské náměstí 7 III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN
 Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
Škola: Gymnázium, Brno, Slovanské náměstí 7 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Název projektu: Inovace výuky na GSN prostřednictvím ICT Číslo projektu: CZ.1.07/1.5.00/34.0940
Plazmové metody. Základní vlastnosti a parametry plazmatu
 Plazmové metody Základní vlastnosti a parametry plazmatu Atom je základní částice běžné hmoty. Částice, kterou již chemickými prostředky dále nelze dělit a která definuje vlastnosti daného chemického prvku.
Plazmové metody Základní vlastnosti a parametry plazmatu Atom je základní částice běžné hmoty. Částice, kterou již chemickými prostředky dále nelze dělit a která definuje vlastnosti daného chemického prvku.
11. ONTOGENEZE I: VEGETATIVNÍ FÁZE, FOTOMORFOGENEZE
 Ontogeneze je vývoj jedince. Ontogenetické změny, kterými rostlina prochází od svého zrodu přes dospívání, reprodukci až k stárnutí a odumírání, se uskutečňují s pomocí základních regulačních mechanismů
Ontogeneze je vývoj jedince. Ontogenetické změny, kterými rostlina prochází od svého zrodu přes dospívání, reprodukci až k stárnutí a odumírání, se uskutečňují s pomocí základních regulačních mechanismů
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Ekologie fotosyntézy
 Ekologie fotosyntézy Fotosyntéza Přeměna zářivé energie Slunce na energii chemických vazeb primární produkce organické hmoty fotochemický (Hillova reakce) a biochemický proces 1 mol přijatého CO 2 energetický
Ekologie fotosyntézy Fotosyntéza Přeměna zářivé energie Slunce na energii chemických vazeb primární produkce organické hmoty fotochemický (Hillova reakce) a biochemický proces 1 mol přijatého CO 2 energetický
Učební texty z fyziky 2. A OPTIKA. Obor zabývající se poznatky o a zákonitostmi světelných jevů. V posledních letech rozvoj optiky vynález a využití
 OPTIKA Obor zabývající se poznatky o a zákonitostmi světelných jevů Světlo je vlnění V posledních letech rozvoj optiky vynález a využití Podstata světla Světlo je elektromagnetické vlnění Zdrojem světla
OPTIKA Obor zabývající se poznatky o a zákonitostmi světelných jevů Světlo je vlnění V posledních letech rozvoj optiky vynález a využití Podstata světla Světlo je elektromagnetické vlnění Zdrojem světla
Otázky k předmětu Globální změna a lesní ekosystémy
 Otázky k předmětu Globální změna a lesní ekosystémy 1. Jaké jsou formy šíření energie v klimatickém systému Země? (minimálně 4 formy) 2. Na čem závisí množství vyzářené energie tělesem? (minimálně 3 faktory)
Otázky k předmětu Globální změna a lesní ekosystémy 1. Jaké jsou formy šíření energie v klimatickém systému Země? (minimálně 4 formy) 2. Na čem závisí množství vyzářené energie tělesem? (minimálně 3 faktory)
Růst a vývoj rostlin
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
Fluorescence (luminiscence)
") Fluorescence (luminiscence) Patří mezi luminiscenční metody fotoluminiscence. Luminiscence efekt, kdy excitované molekuly či atomy vyzařují světlo při přechodu z excitovaného do základního stavu. Podle
Fluorescence (luminiscence) Patří mezi luminiscenční metody fotoluminiscence. Luminiscence efekt, kdy excitované molekuly či atomy vyzařují světlo při přechodu z excitovaného do základního stavu. Podle
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Voda jako životní prostředí - světlo
 Hydrobiologie pro terrestrické biology Téma 6: Voda jako životní prostředí - světlo Sluneční světlo ve vodě Sluneční záření dopadající na hladinu vody je 1) cestou hlavního přísunu tepla do vody 2) zdrojem
Hydrobiologie pro terrestrické biology Téma 6: Voda jako životní prostředí - světlo Sluneční světlo ve vodě Sluneční záření dopadající na hladinu vody je 1) cestou hlavního přísunu tepla do vody 2) zdrojem
Vylepšování fotosyntézy
 Vylepšování fotosyntézy Využití fotosyntézy potraviny energie (paliva) Obojího bude podle predikcí potřebovat lidstvo čím dál tím víc. Energetické využití fotosyntézy potřeba nahrazení fosilních paliv
Vylepšování fotosyntézy Využití fotosyntézy potraviny energie (paliva) Obojího bude podle predikcí potřebovat lidstvo čím dál tím víc. Energetické využití fotosyntézy potřeba nahrazení fosilních paliv
Vitální barvení, rostlinná buňka, buněčné organely
 Vitální barvení, rostlinná buňka, buněčné organely Vitální barvení používá se u nativních preparátů a rozumíme tím zvýšení kontrastu určitých buněčných složek v živých buňkách, nebo tkáních pomocí barvení
Vitální barvení, rostlinná buňka, buněčné organely Vitální barvení používá se u nativních preparátů a rozumíme tím zvýšení kontrastu určitých buněčných složek v živých buňkách, nebo tkáních pomocí barvení
Metoda Live/Dead aneb využití fluorescenční mikroskopie v bioaugmentační praxi. Juraj Grígel Inovativní sanační technologie ve výzkumu a praxi
 Metoda Live/Dead aneb využití fluorescenční mikroskopie v bioaugmentační praxi Juraj Grígel Inovativní sanační technologie ve výzkumu a praxi Co je to vlastně ta fluorescence? Některé látky (fluorofory)
Metoda Live/Dead aneb využití fluorescenční mikroskopie v bioaugmentační praxi Juraj Grígel Inovativní sanační technologie ve výzkumu a praxi Co je to vlastně ta fluorescence? Některé látky (fluorofory)
LI-6400; Gazometrická stanovení fotosyntetických parametrů
 LI-6400; Gazometrická stanovení fotosyntetických parametrů 18. dubna 2008 Tato úloha by Vás měla seznámit s gazometrickými metodami stanovení fotosyntetické aktivity rostlin potažmo s přístrojem LI-6400,
LI-6400; Gazometrická stanovení fotosyntetických parametrů 18. dubna 2008 Tato úloha by Vás měla seznámit s gazometrickými metodami stanovení fotosyntetické aktivity rostlin potažmo s přístrojem LI-6400,
Světlo jako ekologický faktor
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 2 Světlo jako ekologický faktor
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 2 Světlo jako ekologický faktor
Buňka. Kristýna Obhlídalová 7.A
 Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
Buňka Kristýna Obhlídalová 7.A Buňka Buňky jsou nejmenší a nejjednodušší útvary schopné samostatného života. Buňka je základní stavební a funkční jednotkou živých organismů. Zatímco některé organismy jsou
CZ.1.07/1.5.00/
 [1] [3] [2] Číslo projektu Název školy Předmět CZ.1.07/1.5.00/34.0425 INTEGROVANÁ STŘEDNÍ ŠKOLA TECHNICKÁ BENEŠOV Černoleská 1997, 256 01 Benešov BIOLOGIE A EKOLOGIE Tematický okruh Téma Základy obecné
[1] [3] [2] Číslo projektu Název školy Předmět CZ.1.07/1.5.00/34.0425 INTEGROVANÁ STŘEDNÍ ŠKOLA TECHNICKÁ BENEŠOV Černoleská 1997, 256 01 Benešov BIOLOGIE A EKOLOGIE Tematický okruh Téma Základy obecné
Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké
 Fotosyntéza Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké rostliny. Zelené rostliny patří mezi autotrofy
Fotosyntéza Každá molekula kyslíku kterou právě dýcháme vznikla někdy v nějaké rostlině. Každý atom uhlíku našeho těla byl kdysi včleněn fotosyntézou do nějaké rostliny. Zelené rostliny patří mezi autotrofy
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
11. ONTOGENEZE I: VEGETATIVNÍ FÁZE, FOTOMORFOGENEZE
 Ontogeneze je vývoj jedince. Ontogenetické změny, kterými rostlina prochází od svého zrodu přes dospívání, reprodukci až k stárnutí a odumírání, se uskutečňují s pomocí základních regulačních mechanismů
Ontogeneze je vývoj jedince. Ontogenetické změny, kterými rostlina prochází od svého zrodu přes dospívání, reprodukci až k stárnutí a odumírání, se uskutečňují s pomocí základních regulačních mechanismů
Číslo a název projektu Číslo a název šablony
 Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Číslo a název projektu Číslo a název šablony DUM číslo a název CZ.1.07/1.5.00/34.0378 Zefektivnění výuky prostřednictvím ICT technologií III/2 - Inovace a zkvalitnění výuky prostřednictvím ICT SSOS_ZE_1.05
Luminiscence. emise světla látkou, která je způsobená: světlem (fotoluminiscence) fluorescence, fosforescence. chemicky (chemiluminiscence)
 fluorescence, fosforescence. chemicky (chemiluminiscence)") Luminiscence Luminiscence emise světla látkou, která je způsobená: světlem (fotoluminiscence) fluorescence, fosforescence chemicky (chemiluminiscence) teplem (termoluminiscence) zvukem (sonoluminiscence)
Luminiscence Luminiscence emise světla látkou, která je způsobená: světlem (fotoluminiscence) fluorescence, fosforescence chemicky (chemiluminiscence) teplem (termoluminiscence) zvukem (sonoluminiscence)
Co vás dnes čeká: Přednáška Fyziologie rostlin MB130P74. Katedra experimentální biologie rostlin, Z. Lhotáková
 Co vás dnes čeká: Přednáška 2: Specifika rostlinné buňky trocha opakování vakuola buněčná stěna plastidy Fotosyntetické struktury plastidy struktura, typy fotosyntetické pigmenty a jejich lokalizace Sluneční
Co vás dnes čeká: Přednáška 2: Specifika rostlinné buňky trocha opakování vakuola buněčná stěna plastidy Fotosyntetické struktury plastidy struktura, typy fotosyntetické pigmenty a jejich lokalizace Sluneční
Světlo je elektromagnetické vlnění, které má ve vakuu vlnové délky od 390 nm do 770 nm.
 1. Podstata světla Světlo je elektromagnetické vlnění, které má ve vakuu vlnové délky od 390 nm do 770 nm. Vznik elektromagnetických vln (záření): 1. při pohybu elektricky nabitých částic s nenulovým zrychlením
1. Podstata světla Světlo je elektromagnetické vlnění, které má ve vakuu vlnové délky od 390 nm do 770 nm. Vznik elektromagnetických vln (záření): 1. při pohybu elektricky nabitých částic s nenulovým zrychlením
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie
 synekologie b) autekologie c) demekologie") 1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
1. Ekologie zabývající se studiem jednotlivých druhů se nazývá: a) synekologie b) autekologie c) demekologie 2. Plocha lesa v ČR dle statistiky ročně: a) stoupá o cca 2 tis. ha b) klesá o cca 15 tis. ha
Název: Fotosyntéza. Autor: Mgr. Jiří Vozka, Ph.D. Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy
 Název: Fotosyntéza Autor: Mgr. Jiří Vozka, Ph.D. Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: chemie, biologie, matematika, fyzika Ročník: 5. Tématický celek:
Název: Fotosyntéza Autor: Mgr. Jiří Vozka, Ph.D. Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: chemie, biologie, matematika, fyzika Ročník: 5. Tématický celek:
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP. Šárka Poláková
 STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
Praktikum z experimentálních metod biofyziky a chemické fyziky I. Vypracoval: Jana Čurdová, Martin Kříž, Vít Marek. Dne: 2.3.
 Praktikum z experimentálních metod biofyziky a chemické fyziky I. Vypracoval: Jana Čurdová, Martin Kříž, Vít Marek. Dne:.3.3 Úloha: Radiometrie ultrafialového záření z umělých a přirozených světelných
Praktikum z experimentálních metod biofyziky a chemické fyziky I. Vypracoval: Jana Čurdová, Martin Kříž, Vít Marek. Dne:.3.3 Úloha: Radiometrie ultrafialového záření z umělých a přirozených světelných
BIORYTMY. Rytmicita procesů. Délka periody CZ.1.07/2.2.00/ Modifikace profilu absolventa biologických studijních oborů na PřF UP
 BIORYTMY EKO/EKŽO EKO/EKZSB Ivan H. Tuf Katedra ekologie a ŽP PřF UP v Olomouci Modifikace profilu absolventa : rozšíření praktické výuky a molekulárních, evolučních a cytogenetických oborů Rytmicita procesů
BIORYTMY EKO/EKŽO EKO/EKZSB Ivan H. Tuf Katedra ekologie a ŽP PřF UP v Olomouci Modifikace profilu absolventa : rozšíření praktické výuky a molekulárních, evolučních a cytogenetických oborů Rytmicita procesů
Buňka. Buňka (cellula) základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách
 základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách") Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Měření množství dopadající energie světla. Fotoinhibice, fotopoškození a fotoprotekční mechanismy
 Fotoinhibice, fotopoškození a fotoprotekční mechanismy Měření množství dopadající energie světla Ozářenost: W.m -2 (= J.s -1.m -2 ) (osvětlenost: ln.m -2 = lux)? Fotonová (kvantová) ozářenost: mol.s -1.m
Fotoinhibice, fotopoškození a fotoprotekční mechanismy Měření množství dopadající energie světla Ozářenost: W.m -2 (= J.s -1.m -2 ) (osvětlenost: ln.m -2 = lux)? Fotonová (kvantová) ozářenost: mol.s -1.m
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Závěrečná práce studentského projektu Fotosyntéza - Rostlinná barviva
 Gymnázium Jana Nerudy Závěrečná práce studentského projektu Fotosyntéza - Rostlinná barviva Evropský sociální fond Praha a EU Investujeme do vaší budoucnosti Pod vedením vedoucích práce Mgr. Jiřího Vozky,
Gymnázium Jana Nerudy Závěrečná práce studentského projektu Fotosyntéza - Rostlinná barviva Evropský sociální fond Praha a EU Investujeme do vaší budoucnosti Pod vedením vedoucích práce Mgr. Jiřího Vozky,
SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU
 SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU Nikola Burianová Experimentální biologie 2.ročník navazujícího studia Katedra Fyziky Ostravská univerzita v Ostravě OBSAH
SLEDOVÁNÍ VZTAHU MEZI OBSAHEM ENZYMU RUBISCO A KONCENTRACÍ CO 2 V CHLOROPLASTU Nikola Burianová Experimentální biologie 2.ročník navazujícího studia Katedra Fyziky Ostravská univerzita v Ostravě OBSAH
kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové
 Otázka: Růst a vývin rostlin Předmět: Biologie Přidal(a): Verunka kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové FÁZE RŮSTU lze
Otázka: Růst a vývin rostlin Předmět: Biologie Přidal(a): Verunka kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové FÁZE RŮSTU lze
Fluorescence chlorofylu
 Pro připomenutí Fluorescence chlorofylu Princip Fotochemické a nefotochemické zhášení fluorescence Excitace chlorofylu: plantphys.info Analýza zhášení (quenching analysis) Temnostní adaptace Kautského
Pro připomenutí Fluorescence chlorofylu Princip Fotochemické a nefotochemické zhášení fluorescence Excitace chlorofylu: plantphys.info Analýza zhášení (quenching analysis) Temnostní adaptace Kautského
Vybrané spektroskopické metody
 Vybrané spektroskopické metody a jejich porovnání s Ramanovou spektroskopií Předmět: Kapitoly o nanostrukturách (2012/2013) Autor: Bc. Michal Martinek Školitel: Ing. Ivan Gregora, CSc. Obsah přednášky
Vybrané spektroskopické metody a jejich porovnání s Ramanovou spektroskopií Předmět: Kapitoly o nanostrukturách (2012/2013) Autor: Bc. Michal Martinek Školitel: Ing. Ivan Gregora, CSc. Obsah přednášky
Biologie - Kvinta, 1. ročník
 - Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
- Kvinta, 1. ročník Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti Kompetence
SPEKTRÁLNÍ METODY. Ing. David MILDE, Ph.D. Katedra analytické chemie Tel.: ; (c) David MILDE,
 David MILDE,") SEKTRÁLNÍ METODY Ing. David MILDE, h.d. Katedra analytické chemie Tel.: 585634443; E-mail: david.milde@upol.cz (c) -2008 oužitá a doporučená literatura Němcová I., Čermáková L., Rychlovský.: Spektrometrické
SEKTRÁLNÍ METODY Ing. David MILDE, h.d. Katedra analytické chemie Tel.: 585634443; E-mail: david.milde@upol.cz (c) -2008 oužitá a doporučená literatura Němcová I., Čermáková L., Rychlovský.: Spektrometrické
Emise vyvolaná působením fotonů nebo částic
 Emise vyvolaná působením fotonů nebo částic PES (fotoelektronová spektroskopie) XPS (rentgenová fotoelektronová spektroskopie), ESCA (elektronová spektroskopie pro chemickou analýzu) UPS (ultrafialová
Emise vyvolaná působením fotonů nebo částic PES (fotoelektronová spektroskopie) XPS (rentgenová fotoelektronová spektroskopie), ESCA (elektronová spektroskopie pro chemickou analýzu) UPS (ultrafialová
Chemická kinetika. Chemické změny probíhající na úrovni atomárně molekulové nazýváme reakční mechanismus.
 Chemická kinetika Chemická reakce: děj mezi jednotlivými atomy a molekulami, při kterých zanikají některé vazby v molekulách výchozích látek a jsou nahrazovány vazbami v molekulách nově vznikajících látek.
Chemická kinetika Chemická reakce: děj mezi jednotlivými atomy a molekulami, při kterých zanikají některé vazby v molekulách výchozích látek a jsou nahrazovány vazbami v molekulách nově vznikajících látek.
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Luminiscence. Luminiscence. Fluorescence. emise světla látkou, která je způsobená: světlem (fotoluminiscence) chemicky (chemiluminiscence)
 chemicky (chemiluminiscence)") Luminiscence Luminiscence emise světla látkou, která je způsobená: světlem (fotoluminiscence) fluorescence, fosforescence chemicky (chemiluminiscence) teplem (termoluminiscence) zvukem (sonoluminiscence)
Luminiscence Luminiscence emise světla látkou, která je způsobená: světlem (fotoluminiscence) fluorescence, fosforescence chemicky (chemiluminiscence) teplem (termoluminiscence) zvukem (sonoluminiscence)
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
Dýchací řetězec. Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci)
") Dýchací řetězec Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 -->6 CO 2 + 6H 2 O + 38 ATP Dýchací
Dýchací řetězec Viz též přednášky prof. Kodíčka (snímky a blány v levém sloupci) Odbourávání glukosy (včetně substrátových fosforylací) C 6 H 12 O 6 + 6O 2 -->6 CO 2 + 6H 2 O + 38 ATP Dýchací
Buňka cytologie. Buňka. Autor: Katka www.nasprtej.cz Téma: buňka stavba Ročník: 1.
 Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,