Dostupnost živin jako faktor utvářející morfologii kořenů (trofomorfogeneze) Vliv dusíkatých látek, fosfátů, síranů a iontů železa
|
|
|
- Dalibor Brož
- před 5 lety
- Počet zobrazení:
Transkript
1 Dostupnost živin jako faktor utvářející morfologii kořenů (trofomorfogeneze) Vliv dusíkatých látek, fosfátů, síranů a iontů železa
2 Co je ovlivněno? Primární kořen Laterální kořen Kořenové vlásky (tvoří až 70% povrchu kořene)
3 Ionty železa dostatečná koncentrace Fe Podobný fenotyp jako u fosfátů neostatek železa vede k nárůstu délky kořenových vlásků a zvýšení jejich hustoty až 5x (zvýšená diferenciace epidermálních buněk) Jiná signální dráha mutant axr2 (auxin resistant) netvoří vlásky ani na plném ani Fe-deficientním mediu rozdíl proti fosfátu nízká koncentrace Fe
4 Sírany Nízká koncentrace síranů podporuje větvení kořenového systému Větší hustota zakládání laterálních kořenů Blíže ke kořenové špičce Aktivace enzymu nitrilázy, která se podílí na tvorbě IAA- zvýšení koncentrace auxinu.
5 Příjem nitrátů -Dobře rozpustné (v půdě 1-10 mm) -A.thaliana nemá mykorhizu veškerý transport se děje pomocí kořenů -Příjem nitrátů způsobí dpeolarizaci membrány kotransport H+, elektrofyziologická měření, 7 NRT genů u A.th -Dva mechanismy příjmu nitrátů vysokoafinitní a nízkoafinitní (nesaturovatelný) -Oba mechanismy regulované mnoha faktory, nitrát aktivující (hlavně VA) do určité míry pak inhibující, amonné ionty inhibují - Příjem nitrátů je centrálněřízen
6 Signalizace nitrátů -Asimilace nitrátů dusičnan dusitan amonný iont -Co je detekováno? -Ovlivnění exprese genů metabolismus dusíku, metabolismus sacharidů -Ovlivnění morfogeneze kořenového systému (poč. 70. léta n ječmeni, později generalizace) -Metabolická teorie rostoucí kořeny odčerpávají živiny ostatním, neplatí i mutanti v NR odpovídají na změny hladiny nitrátů

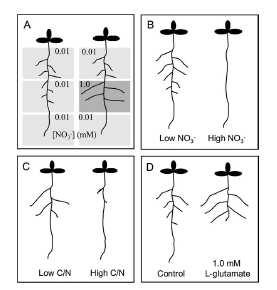
7 Vliv nerovnoměrné distribuce nitrátů v půdě (jako příklad navigace kořenů prostředím)
8

9 U arabidopsis se během indukovaného růstu LK neměníčetnost zakládaných kořenů ani rychlost jejich růstu během prvních stadií. Nedochází k zvětšování buněk, jen nárůst aktivity meristemu.
10 Dvě paralelní dráhy nitrátů jako signálních molekul Systémová řízená poměrem C:N v kořeni a prýtu, saturovatelnost (1mM nitrát vyvolá stejnou odpověď jako 50mM), nově založené LK, reverzibilní. Nezávislá na metabolizaci nitrátů, ale NR mutace zvyšuje citlivost na celkovou inhibici nitráty. - Změna obsahu auxinu v kořenech a prýtu během signalizace. -systémový efekt je oslaben v abi (ABA insensitive) mutantech (abi4-1,4-2, 5)

11 Lokální řízená transkripčními faktory, všechna stadia LK ANR1 Arabidopsis NITRATE REGULATED -MADS box transkripční faktor, akumulace mrna při nadbytku nitrátů, není v deficitních kořenech - NRT geny jako regulátory odpovědi AtNRT1.1 upstream k ANR1, podobná specifická odpověď na lokální nitráty, regulace dalších NRT genů -mutanti ANR1 neodpovídají na lokální zvýšení N, ale podrží si negativní odpověď na zvýšenou koncentraci nitrátu hypersenzitivita už 0,1 mm nitrát proti 10 mm ve WT.
12 -auxinový mutant axr4 také neodpovídá na lokální zvýšení nitrátu - U nodulujících rostlin je regulace nodulace a tvorby laterálních kořenů (hypernodulation aberrant root1- HAR1) - Amonné ionty blokují růst kořenů ve vysoké koncentraci a v závislosti na dosažitelnosti K+ - nejasná funkce - Glutamát jako zdroj dusíku vede k celkovému zkrácení kořenového systému a bohatšímu větvení. Druhově specifická reakce.
13
14 K čemu je to rostlině dobré? N jako indikátor obecné dostupnosti živin? N rychle difunduje a je dobrý indikátor kompetice?? poměr C/N... fotosyntéza/min. výživa... nadzemníčásti/kořen
15 Poměr C:N Uhlík z fotosyntézy nebo media, dusík z media Akumulace auxinu v hypokotylu LIN1 gen regulující rovnováhu, lin1 větví kořeny i při nízké fotosyntéze LIN1 je NRT2.1- vysokoafinitní transporter nitrátu


16 det3 mutant je na vysokém nitrátu deficientní v dlouživém růstu, s jiným zdrojem N na půdě se stejnými osmotickými vlastnostmi je srovnatelný s WT. Vliv na prýt
17
18
19 Asimilace a signalizace fosfátů Často nerozpustné anorganické sloučeniny, organicky vázaný Kyselé fosfatázy, organické kyseliny, vysokoafinitní transporter


20 Za normálních okolností stačí malá plocha kořene, při nedostatku jsou iniciovány kořenové vlásky, mutant v jejich tvorbě. rhd2 výrazně zaostává na deficientním mediu, ale ne na plném mediu. Signální dráha odlišná od železa mutant axr2 tvoří na fosfátově deficitním mediu normální vlásky.
21 Sytémová reakce na nedostatek fosfátů Na deficientním mediu vytváří většina rostlin klastry laterálních kořenů s determinovaným meristemem (několik dní) a následnou tvorbou množství kořenových vlásků. U A.thaliana roste množství laterálních kořenů až 50x (1uM x 1mM fosfát). Laterální kořeny na konci vývoje ztrácí citlivost na auxin a mitotickou aktivitu.

22 Odpověď na deficit fosfátů DR5 cycb PHOSPHATE TRANSPORTER2


23 Low phosphate insensitive (LPI) 11genů, neznámá funkce Ovlivňují expresi genů zodpovědných za správnou funkci meristemu (PLETHORA, SCARECROW, CRE, SHORT- ROOT)


24 Necitlivý k fosfátu - PHO3 Neznámá funkce, stálá nízká hladina interního fosfátu, hyperakumulace sacharidů, škrobu, anthokyanů. Ovlivnění sekrece?
25 Phosphate response regulator PHR1 transkripční faktor Myb, porucha regulace poměru kořen:prýt a exprese odpovídajících genů při fosfátovém hladovění. PHO2 je neobvyklá E2 ubiquitin transferáza a spolu s neg. regulátorem mikro RNA mir-399 tvoří signální fosfátovou dráhu. Hladovění vede k akumulaci P v listech. Phosphate defficiency response2 (PDR2). Monitoruje hladinu fosfátu a reguluje aktivitu meristemu. Mutant vykazuje hypersenzitivitu k fosfátovému hladovění.
26 Low phosphate root LPR oxidáza obsahující měď, zastaví růst prim. kořene v oblasti chudé na fosfáty. Phosphate starvation regulator - SUMOylovaný MYB traskripční faktor, aktivator genů fosfátového hladovění.
27 Další faktory ovlivňující odpověď na fosfátové hladovění Auxin se účastní tvorby laterálních kořenů inhibice TIBA,NPA, ale snížená citlivost. vyšší senzitivita primárních kořenů k inhibici auxinem a vyšší senzitivita laterálních kořenů k auxinu (10-100x). Zapojení TIR1 do signalizace deficience (zvýšení exprese, overexprese napodobuje deficientní fenotyp, mutace tir1-1 neindukuje laterální kořeny) ARF7 a 19 jsou esenciální pro tvorbu LK Cytokinin je negativním regulátorem laterálních kořenů při hladovění. Ethylen zodpovědnost za ukončení funkce meristemu Sacharidy

28
29 Jarovizace (vernalizace, vernalisation, яровизация) absolutní fakultativní
.")
 T. D.")
30 (Nešťastné) dějiny jarovizace Jev znám minimálně od zač. 20. století (J. Tournois , + na frontě) Gassner, G. (1918). Beiträge zur physiologiischen Charakteristik sommer- und winterannueller Gewächse, insbesondere der Getreidepflanzen. Z. Bot. 10, (1929?) T. D. Lysenko - jarovizace ozimů (nevymrznou, ale možná zplesniví) zásluhou N.I. Vavilova - bosonohý akademik Trofim Děnisovič Lysenko ( ) Nikolaj Ivanovič Vavilov ( )

 O situaci v")
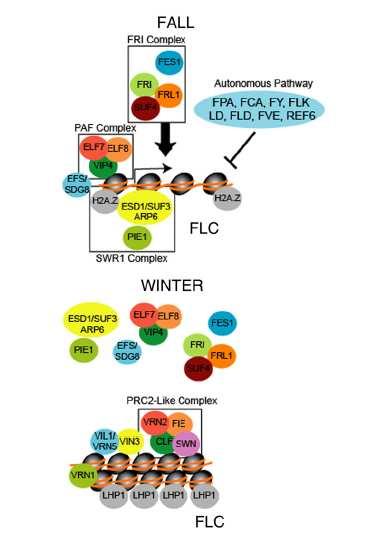
31 N. I. Vavilov + v gulagu 1943 (hlady?) O situaci v biologii
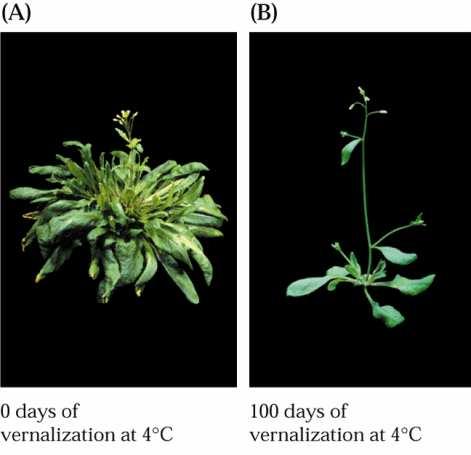
32 Chladový signál Aklimace rostliny se stávají odolné vůči mrazu, uvolnění vápníku a aktivace TCH genů, tvorba kryoprotektantů (sacharidy, polyaminy) Změna membránové fluidity Propojení vernalizace a aklimace HOS1- negativní regulátor chladové odpovědi (intenzita odpovědi, teplota) Mutanti vykazují chladovou odpověď při 19 C a kvetou výrazně dříve, omezená citlivost k vernalizaci E3 ligáza, cyklování podobné COP1
33
34
35 Mitotická stabilizace vernalizačního signálu DNA methylace C v GC páru Reset reprodukcí, dědí se v rámci buněčné linie Methylace je neg. regulátor vernalizace vede k poklesu methylace. Inhibice methylace vede k urychlení kvetení. Demethylace je katalyzovaná DNA- glykosylázami z rodiny DEMETER. Celkový vzorec methylací DNA je prakticky dán demethylací.

36 Remodelace chromatinu euchromatin x heterochromatin Heterochromatin tvoří spolu s inaktivovanými geny chromocentra
37 Pro tvorbu chromocenter je důležitá metylace DNA (DDM1- faktor chromatinové remodelace, MET1- methyltransferáza). Během života rostliny roste podíl HC ( u ddm1 méně) Rozvolnění center během dediferenciace (např. protoplastování) a tvorby kalusů. Porucha v tvorbě kalusů Kryptonite (KRP) H3K9 methylace Nutná pro fomování kalusů. ALE ddm1 může kalusy tvořit také. Negativní regulace tvorby kalusů Curly leaf (CLF) H3K27 methyltransferáza, mutanti spontánně tvoří struktury nediferencovaných buněk. Podobně LHP1 Like heterochromaitin protein 1) protein vážící methylovaný histon působí proti dediferenciaci.
38 Epigeneticky řízené geny FLC flowering locus C MADS TF, konstitutivní exprese, negativní regulátor, hladina snížená vernalizací přetrvává Modifikace hromatinu acetylace H3K9 a methylace H3K14 jsou znaky ativního genu - Methylace H3K9 a H3K27 jsou znaky neaktivního, reprimovaného genu FRI frigida mutant má redukovanou hladinu FLC
39 Odlišná citlivost ekotypů A.thaliana k vernalizaci je daná expresí funkčních FLC a FRI
40 Geny dále zapojené ve vernalizaci VIN3 -vernalization insensitive mrna jen po dobu chladu, TF podobná doména pro remodeling chromatinu, vin3 mají modifikace jako nevernalizované rostliny VRN2 polycomb protein, vrn2 nemají represivní modifikace, součást Polycomb repression complex (PRC2), kde je curly leaf, swinger and Fertilization independen endosperm komplex zajišťující represi FLC VRN1- DNA vážící, jednostranný nárůst metylace H3K27 VRN5- interaktor VIN3, stejný fenotyp,
41 FLC aktivátory 2 skupiny nezávislé na autonomní dráze a závislé na autonomní dráze Závislé na FRI genu frigida-like, frigida-essential, FLC v těchto mutantech není exprimován nepotřebují vernalizaci Nejasná funkce. Suprese mutace autonomních genů FLC je exprimván i na pozadí fri1. Mutace v těchto genech opět umlčí FLC. Geny pro remodelaci chromatinu (PAF1 komplex u kvasinek) snížení methylace H3K4, H3K36 EFS protein podobná funkce jako PAF komplex SWR1 komplex váže H2A, přítomnost tohoto histonu je markerem aktivního genu
42
43 Geny autonomní dráhy blokují expresi FLC RNA vážící proteiny TF a remodelace chromatinu Mutace vede k vysoké hladině FLC a hyperacetylaci H3a H4 (př. Flowering locus D je histon-deacetyláza)
44 Alternativní dráhy vernalizace MAF proteiny paralogy FLC, 5 členů -obecné represory kvetení -sdílí některé regulátory s FLC x některé jsou specifické -MAF1, 2, 3 jsou chladem reprimovány, MAF 5 je indukován a MAF4 neovlivněn -Mutace maf2 způsobuje zkrácení doby chladu nezbytné pro vyvolání odezvy -Overexprese MAF2 vede k necitlivosti k vernalizaci
45 Alternativní dráhy vernalizace AGL24 a SVP MADS-box TF -Fungují jako aktivátory kvetení v odpověď na vernalizaci -Vzájemná aktivace se SOC1 -Mutanti mají opožděný nástup kvetení -Obecná exprese ve vegetativních orgánech
46 Alternativní dráhy vernalizace AGL 19 agamous-like 19 (homolog SOC1) Ektopická exprese indukuje kvetení Nezávislé na SOC1, FLC Stejný vernalizační mechanismus jako FLC Represivní H3K27 methylace před vernalizací, méně po
47
48 Rozdílný mechanismus jednoděložné x dvojděložné Jednoděložné využívají proteinů světelné dráhy, vernalizační signál lze nahradit dlouhodobým SD.
49 VRN1 MADS TF, homolog APETALA1 VRN2 podobnost s genem CONSTANS, obsahuje tzv. CCT doménu, cíl vernalizace VRN3 homolog FT VRT2 - homolog SVT, represe VRN1 společně s VRN2 Není zatím jasné, v kteréčásti se vernalizace odehrává. Není jasný mechanismus paměti. Prozatím byly objeveny homology VIN3 s podobným akumulačním chováním.
50 Závěr Vernalizace se vyvinula později v evoluci nezávisle u jednoa dvojděložných. Využívá původního konzervovaného mechanismu regulace kvetení na základě délky fotoperiody. Vlastní mechanismus vernalizace využívá rozdílné skupiny proteinů v jedno- a dvojděložných rostlin.
51
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") 2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2) Reprodukce rostlin
 Reprodukce rostlin") 2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
MECHANIZMY EPIGENETICKÝCH PROCESŮ
 MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Životní cyklus rostliny. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení
 Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení") MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Látky jako uhlík, dusík, kyslík a. z vnějšku a opět z něj vystupuje.
 KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
KOLOBĚH LÁTEK A TOK ENERGIE Látky jako uhlík, dusík, kyslík a voda v ekosystémech kolují. Energii se do ekosystémů dostává z vnějšku a opět z něj vystupuje. Základní podmínky pro život na Zemi. Světlo
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Vápník. Deficience vápníku: - 0,4-1,5% DW. - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM)
") Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch
 Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Struktura chromatinu. Co je to chromatin?
 Struktura chromatinu Buněčné jádro a genová exprese Lenka Rossmeislová struktura-význam-modifikace Co je to chromatin? hmota, ze které jsou vytvořeny chromozomy DNA asociovaná s proteiny, které napomáhají
Struktura chromatinu Buněčné jádro a genová exprese Lenka Rossmeislová struktura-význam-modifikace Co je to chromatin? hmota, ze které jsou vytvořeny chromozomy DNA asociovaná s proteiny, které napomáhají
- Cesta GS GOGAT - Cesta GDH
 Buchanan 2000 Asimilace amonného iontu: - Cesta GS GOGAT - Cesta GDH Buchanan 2000 GS (glutaminsyntetáza, EC 6.3.1.2) - oktamerní protein o velikosti 350-400 kda, tvořený 8 téměř identickými podjednotkami
Buchanan 2000 Asimilace amonného iontu: - Cesta GS GOGAT - Cesta GDH Buchanan 2000 GS (glutaminsyntetáza, EC 6.3.1.2) - oktamerní protein o velikosti 350-400 kda, tvořený 8 téměř identickými podjednotkami
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Toxické látky v potravinách s nebezpečím onkologické aktivace
 Toxické látky v potravinách s nebezpečím onkologické aktivace Doc. MUDr. Pavel Dlouhý, Ph.D. Ústav hygieny 3. LF UK, Praha Rizikové faktory pro vznik nádorů Obezita Nadměrný příjem tuků? Nadměrná konzumace
Toxické látky v potravinách s nebezpečím onkologické aktivace Doc. MUDr. Pavel Dlouhý, Ph.D. Ústav hygieny 3. LF UK, Praha Rizikové faktory pro vznik nádorů Obezita Nadměrný příjem tuků? Nadměrná konzumace
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová
 autoi: E. Tylová, L. Moravcová") Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová Takto vypadají kontrolní, kultivované v roztoku obsahujícím všechny živiny. Pokud se vaše rostlinka vizuáln liší, kliknte
Klí k urování deficiencí kukuice seté (Zea mays) autoi: E. Tylová, L. Moravcová Takto vypadají kontrolní, kultivované v roztoku obsahujícím všechny živiny. Pokud se vaše rostlinka vizuáln liší, kliknte
Stav lesních půd drama s otevřeným koncem
 Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Fosfor: Projevy deficience P: - 0,2-0,5 % SH rostlin. - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1
 Fosfor: Buchanan 2000-0,2-0,5 % SH rostlin - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1 µm) - toxicita velmi zřídka nad 1% SH (např. Cajanus cajan při 0,3-0,5%, Vigna
Fosfor: Buchanan 2000-0,2-0,5 % SH rostlin - často limitující minerální živina (v substrátu většinou v koncentracích kolem 1 µm) - toxicita velmi zřídka nad 1% SH (např. Cajanus cajan při 0,3-0,5%, Vigna
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
Epigenetické mechanismy u rostlin
 Epigenetické mechanismy u rostlin Úvod srovnání rostlin a živočichů Klasické epigenetické systémy: transpozony, paramutace, nikleolární dominance Transgeny a viry zprostředkované umlčování genů Epigenetika
Epigenetické mechanismy u rostlin Úvod srovnání rostlin a živočichů Klasické epigenetické systémy: transpozony, paramutace, nikleolární dominance Transgeny a viry zprostředkované umlčování genů Epigenetika
2012/2013. Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol.
 2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
*Mléko a mléčné výrobky obsahují řadu bioaktivních
 www.bileplus.cz Mléko a mléčné výrobky obsahují řadu bioaktivních látek (vápník, mastné kyseliny, syrovátka, větvené aminokyseliny) ovlivňující metabolismus tuků spalování tuků Mléčné výrobky a mléčné
www.bileplus.cz Mléko a mléčné výrobky obsahují řadu bioaktivních látek (vápník, mastné kyseliny, syrovátka, větvené aminokyseliny) ovlivňující metabolismus tuků spalování tuků Mléčné výrobky a mléčné
Rekapitulace. Rostlina vládne buňkám, ne(jen) buňky rostlině.
 buňky rostlině.") Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
RŮST A VÝVOJ ROSTLIN. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1
 RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
Odborná škola výroby a služeb, Plzeň, Vejprnická 56, Plzeň. Číslo materiálu 19. Bc. Lenka Radová. Vytvořeno dne
 Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
Název školy Název projektu Číslo projektu Číslo šablony Odborná škola výroby a služeb, Plzeň, Vejprnická 56, 318 00 Plzeň Digitalizace výuky CZ.1.07/1.5.00/34.0977 VY_32_inovace_ZZV19 Číslo materiálu 19
Biochemie kosti. Anatomie kosti. Kostní buňky. Podpůrná funkce. Udržování homeostasy minerálů. Sídlo krvetvorného systému
 Biochemie kosti Podpůrná funkce Udržování homeostasy minerálů Sídlo krvetvorného systému Anatomie kosti Haversovy kanálky okostice lamely oddělené lakunami Kostní buňky Osteoblasty Osteocyty Osteoklasty
Biochemie kosti Podpůrná funkce Udržování homeostasy minerálů Sídlo krvetvorného systému Anatomie kosti Haversovy kanálky okostice lamely oddělené lakunami Kostní buňky Osteoblasty Osteocyty Osteoklasty
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP. Šárka Poláková
 STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
STANOVENÍ OBSAHŮ PŘÍSTUPNÝCH MIKROELEMENTŮ V PŮDÁCH BMP Šárka Poláková Přístupné mikroelementy Co jsou mikroelementy a jaká je jejich funkce v živých organismech Makrobiogenní prvky (H, C, O, N) Mikrobiogenní
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
05 Biogeochemické cykly
 05 Biogeochemické cykly Ekologie Ing. Lucie Kochánková, Ph.D. Prvky hlavními - biogenními prvky: C, H, O, N, S a P v menších množstvích prvky: Fe, Na, K, Ca, Cl atd. ve stopových množstvích I, Se atd.
05 Biogeochemické cykly Ekologie Ing. Lucie Kochánková, Ph.D. Prvky hlavními - biogenními prvky: C, H, O, N, S a P v menších množstvích prvky: Fe, Na, K, Ca, Cl atd. ve stopových množstvích I, Se atd.
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
kyslík ve vodě CO 2 (vápenato-)uhličitanová rovnováha alkalita
uhličitanová rovnováha alkalita") kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
kyslík ve vodě CO 2 ph (vápenato-)uhličitanová rovnováha alkalita elementární plyny s vodou nereagují, ale rozpouštějí se fyzikálně (N 2, O 2, ) plynné anorganické sloučeniny (CO 2, H 2 S, NH 3 ) s vodou
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA
 TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
jungle kompletní výživa rostlin Nahlédnutí pod pokličku indabox pro všechny typy pěstebních systémů /mírně odborné pojednání MEDICAL QUALITY GROWIN
 /mírně odborné pojednání kompletní výživa rostlin pro všechny typy pěstebních systémů JungleInDaBox je třísložkový komplex minerálního základu a synergicky působících biologických doplňků. Vysoká efektivita
/mírně odborné pojednání kompletní výživa rostlin pro všechny typy pěstebních systémů JungleInDaBox je třísložkový komplex minerálního základu a synergicky působících biologických doplňků. Vysoká efektivita
Autophagie a imunitní odpověd. Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha
 Autophagie a imunitní odpověd Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha Ostrava, 29. ledna 2019 Historie Nobel Prize 2016 Yoshinori Ōsumi https:p//nobeltpizrog/utplodss/2l018//06/ohsuiillchtul
Autophagie a imunitní odpověd Miroslav Průcha Klinická imunologie Nemocnice Na Homolce, Praha Ostrava, 29. ledna 2019 Historie Nobel Prize 2016 Yoshinori Ōsumi https:p//nobeltpizrog/utplodss/2l018//06/ohsuiillchtul
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
8. Polysacharidy, glykoproteiny a proteoglykany
 Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
FYZIOLOGIE ROSTLIN VÝŽIVA ROSTLIN 1) AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN
 AUTOTROFNÍ VÝŽIVA ROSTLIN 2) HETEROTROFNÍ VÝŽIVA ROSTLIN") FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
FYZIOLOGIE ROSTLIN Fyziologie rostlin, Biologie, 2.ročník 25 Podobor botaniky, který studuje životní funkce a individuální vývoj rostlin. Využívá poznatků z dalších odvětví biologie jako je morfologie,
Radiobiologický účinek záření. Helena Uhrová
 Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.)
") Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.) Botanická charakteristika: ČESNEK KUCHYŇSKÝ (ALLIUM SATIVUM L.) Pravlastí je Džungarsko (severní Čína) v Střední Asii,
Vliv selenu, zinku a kadmia na růstový vývoj česneku kuchyňského (Allium sativum L.) Botanická charakteristika: ČESNEK KUCHYŇSKÝ (ALLIUM SATIVUM L.) Pravlastí je Džungarsko (severní Čína) v Střední Asii,
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Nádherné akvarijní rostliny díky perfektně nastavené péči. Systémová péče o rostliny
 Nádherné akvarijní rostliny díky perfektně nastavené péči Systémová péče o rostliny sera péče o rostliny Vyvážený a kompletní systém pro individuální a účinnou péči o rostliny sera florena pro vodní rostliny,
Nádherné akvarijní rostliny díky perfektně nastavené péči Systémová péče o rostliny sera péče o rostliny Vyvážený a kompletní systém pro individuální a účinnou péči o rostliny sera florena pro vodní rostliny,
Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN,
 Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN, CHRÓM, Calcium, Magnesium Organické Minerály ORGANICKÉ MINERÁLY Zásadní zvláštností všech přípravků linie «Organické minerály»
Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN, CHRÓM, Calcium, Magnesium Organické Minerály ORGANICKÉ MINERÁLY Zásadní zvláštností všech přípravků linie «Organické minerály»
Chromatin. Struktura a modifikace chromatinu. Chromatinové domény
 Chromatin Struktura a modifikace chromatinu Chromatinové domény 2 DNA konsensus 5 3 3 DNA DNA 4 RNA 5 ss RNA tvoří sekundární strukturu s ds vlásenkami ds forms 6 of nucleic acids Forma točivost bp/turn
Chromatin Struktura a modifikace chromatinu Chromatinové domény 2 DNA konsensus 5 3 3 DNA DNA 4 RNA 5 ss RNA tvoří sekundární strukturu s ds vlásenkami ds forms 6 of nucleic acids Forma točivost bp/turn
N 2 + 8[H] + 16 ATP 2NH 3 + H 2 + 16ADP + 16P i
![N 2 + 8[H] + 16 ATP 2NH 3 + H 2 + 16ADP + 16P i](/thumbs/19/328137.jpg "N 2 + 8[H] + 16 ATP 2NH 3 + H 2 + 16ADP + 16P i") 1. Fixace N 2 v širším kontextu Biologická fixace vzdušného dusíku představuje z hlediska globální bilance N 2 důležitý proces jímž je plynný dusík asimilován do živé biomasy. Z povahy vazby mezi atomy
1. Fixace N 2 v širším kontextu Biologická fixace vzdušného dusíku představuje z hlediska globální bilance N 2 důležitý proces jímž je plynný dusík asimilován do živé biomasy. Z povahy vazby mezi atomy
J. Kubíček FSI Brno 2018
 J. Kubíček FSI Brno 2018 Fosfátování je povrchová úprava, kdy se na povrch povlakovaného kovu vylučují nerozpustné fosforečnany. Povlak vzniká reakcí iontů z pracovní lázně s ionty rozpuštěnými z povrchu
J. Kubíček FSI Brno 2018 Fosfátování je povrchová úprava, kdy se na povrch povlakovaného kovu vylučují nerozpustné fosforečnany. Povlak vzniká reakcí iontů z pracovní lázně s ionty rozpuštěnými z povrchu
Detlef Weigel ( )
") VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
Úvod do biochemie. Vypracoval: RNDr. Milan Zimpl, Ph.D.
 Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.
Úvod do biochemie Vypracoval: RNDr. Milan Zimpl, Ph.D. TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Co je to biochemie? Biochemie je chemií živých soustav.