Metabolismus sacharidů
|
|
|
- Jindřiška Dvořáková
- před 7 lety
- Počet zobrazení:
Transkript
1 Základy biochemie KB / B Metabolismus sacharidů Inovace studia biochemie prostřednictvím e-learningu Z / /0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky.
2 snova Přehled strukturních forem sacharidů. Glykoproteiny a glykosylace. Transportéry glukosy přes plasmatickou membránu. Glykolýza. Regulace glykolýzy. Glukoneogeneze. Regulace glykolýzy a glukoneogeneze. Substrátové cykly. oriho cyklus. Pentosafosfátová dráha.
3 Proč právě glukosa je univerzálním zdrojem energie?? 1. Pravděpodobně první sacharid tvořený z formaldehydových jednotek za prebiotických podmínek. 2. Glukosa má nízkou tendenci neenzymově glykosylovat proteiny díky preferenci cyklické formy. 3. K relativně vysoké stabilitě glukosy přispívá to, že všechny hydroxylové skupiny v β-d-glukose jsou v ekvatoriální poloze.
4 Převládajícími formami monosacharidů v roztoku jsou cyklické. Aldehydy tvoří poloacetalové, ketony poloketalové. Podle heterocyklů furanu furanosy, pyranu pyranosy. Furan Pyran
5 Formy D-glukosy. Uzavřením poloacetalového kruhu vzniká nové chirální centrum na uhlíku 1. Existují dva anomery D-glukosy α a β. 2 2 = 2 D-Glukosa α-d-glukopyranosa 2 (otevřená forma) β-d-glukopyranosa
6 yklická forma D-fruktosy 2 2 = 2 D-Fruktosa (otevřená forma) β-d-glukopyranosa (cyklická forma)
7 yklická forma sacharosy α β 2 Sacharosa (α-d-glukopyranosyl-(1 2)-β-D-fruktofuranosa)
8 yklická forma laktosy bsahuje galaktosu a glukosu spojené (1 4) glykosidovou vazbou. 2 2 β 1 4 α Laktosa (β-d-galaktopyranosyl-(1 4)-α-D-glukopyranosa)
9 yklická forma maltosy bsahuje dvě molekuly glukosy spojené (1 4) glykosidovou vazbou. 2 2 α 1 4 α Maltosa (α-d-glukopyranosyl-(1 4)-α-D-glukopyranosa)
10 Ketotriosa a aldotriosy Dihydroxyaceton (ketosa) D-Glyceraldehyd (aldosa) L-Glyceraldehyd (aldosa)
11 D-Aldosy se třemi až šesti uhlíky Konfigurace D se odvozuje od chirálního uhlíku, který je nejvzdálenější od aldehydové skupiny D-Glyceraldehyd D-Erythrosa 2 D-Threosa
12 D-Erythrosa D-Ribosa 2 D-Arabinosa
13 D-Threosa D-Xylosa 2 D-Lyxosa
14 D-Ribosa 2 D-Arabinosa D-Allosa D-Altrosa D-Glukosa D-Mannosa
15 D-Xylosa 2 D-Lyxosa D-Gulosa D-Idosa D-Galaktosa D-Talosa
16 D-Ketosy se třemi až šesti uhlíky Konfigurace D se odvozuje od chirálního centra nejdále od ketoskupiny Dihydroxyaceton D-Erythrulosa D-Ribulosa 2 D-Xylulosa D-Psikosa D-Fruktosa D-Sorbosa D-Tagatosa
17 Větvení glykogenu Řetězce glukos spojených glykosidovou vazbou α(1 4) jsou po deseti glukosových jednotkách větveny glykosidovou vazbou α(1 6). 2 2 α α 1 4 α(1 6)-Glykosidová vazba α
18 Struktury celulosy, škrobu a glykogenu Vazby β(1 4) vedou k rovným řetězcům, kdežto vazby α(1 4) k prohnutým strukturám elulosa (β[1 4]-glykosidová vazba) Škrob a glykogen (α[1 4]-glykosidová vazba)
19 Glykosaminoglykany aniontové polysacharidy pakující se disacharidové jednotky obsahující glukosamin nebo galaktosamin. Vazbou na proteiny tvoří proteoglykany S S S 3 N 3 hondroitin-6-sulfát Keratansulfát N 3 - S 3 eparin - NS 3 S Dermatansulfát N 3 yaluronát N 3
20 Glykoproteiny Glykosidové vazby mezi proteiny a sacharidy.vazby přes Asn (N-glykosidy), vazby přes Thr nebo Ser (-glykosidy). GlcNAc = N-Acetylglukosamin. Asn Ser 2 N N 2 N 2 2 N 3 N 3 N-vázaný GlcNAc -vázaný GalNAc
21 Glykosylace Glykosylace proteinů probíhá v lumen endoplasmatického retikula (ER) a v Golgiho komplexu. Příkladem je pankreatická elestasa [ ] katalyzující hydrolýzu proteinů (elastin) na místě Ala Xaa. Pankreatická elastasa je uvolňována z pankreatu ve formě zymogenu. Je syntetizována ribosomy na cytoplasmatické straně membrány ER. Signální sekvence 29 aminokyselin navede proelastasu do lumen a poté se odštěpí. Volný zymogen je glykosylován a poté, k dokončení glykosylace, putuje do Gogiho komplexu. becně: N-glykosylace začíná v ER a pokračuje v Golgiho komplexu. -Glykosylace probíhá pouze v Golgiho komplexu.

22 Elastasa vylučovaný glykoprotein s hydrolytickými vlastnostmi. Nachází se v krevním séru. ligosacharidové řetězce zaujímají podstatnou část molekuly.
23 Transportéry glukosy přes plasmatickou membránu. Např. GLUT1 a GLUT3 jsou téměř ve všech savčích buňkách. ladina glukosy v krvi je od 4 mm do 8 mm. Rodina transportérů glukosy: Název Tkáň K m (glukosa) Komentář GLUT1 Všechny savčí tkáně 1 mm Základní vstup glukosy. GLUT2 Játra a β-buňky pankreatu mm Regulace hladiny insulinu v pankreatu. V játrech odčerpává nadbytek glukosy z krve. GLUT3 Všechny savčí tkáně 1 mm Základní vstup glukosy. GLUT4 Svaly a tukové buňky 5 mm Množství se zvyšuje tréninkem. GLUT5 Tenké střevo ---- Primárně transportér fruktosy.
24 Glykolýza. ans Buchner a Eduard Buchner 1897 Příprava bezbuněčného extraktu kvasinek pro terapeutické účely. Konzervace sacharosou sacharosa byla rychle fermentována na ethanol. Nechtěně tak poprvé prokázali, že fermentace může probíhat mimo živou buňku. d roku 1860 (Luis Pasteur) fermentace může probíhat pouze v živých buňkách (vitalismus). Poté studována fermentace i ve svalech. Byly nalezeny naprosto shodné děje. Glykolýza byla plně objasněna v roce Gustav Embden, tto Mayerhof, arl Neuberg, Jacob Parnas, tto Warburg, Gerty ori a arl ori. Glykolýza bývá také označována jako Emden-Mayerhofova dráha.
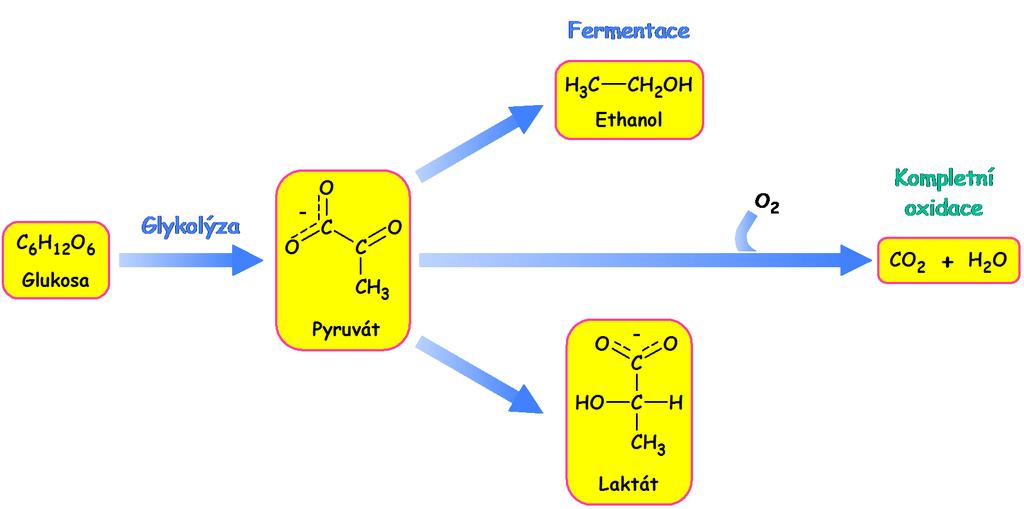
25 Glykolýza, také fermentace, probíhá anaerobně. Z Řečtiny Glyk- sladký, lysis, rozpad, ztráta sladkosti. Pochod společný pro prokaryotní i eukaryotní organismy. Glykolýzou se získává energie i bez přístupu kyslíku. U eukaryot probíhá glykolýza ve třech stupních v cytoplasmě: a) Převedení glukosy na fruktosa-1,6-bisfosfát (F-1,6-bisP) b) Štěpení F-1,6-bisP na dvě triosy c) Tvorba ATP při oxidaci tříuhlíkatých sloučenin na pyruvát. Pyruvát může být dále převeden na laktát (ve svalech) nebo na produkty fermentace jako je ethanol např. kvasinkami.
26 Katabolické dráhy glukosy


27 Schematický průběh glykolýzy V prvním stupni probíhají fosforylace, v druhém štěpení hexosy na dvě vzájemně převoditelné triosy a v třetím, při oxidaci tříuhlíkatých fragmentů na pyruvát, se tvoří ATP.
28 exokinasa - fosforylace glukosy Fosforylací se destabilizuje glukosa. Glukosa-6-fosfát neprochází buněčnou membránou. 2 2 P ATP exokinasa + ADP + + Glukosa Glukosa-6-fosfát (G-6P)
29 Fosfoglukosaisomerasa Izomerace: aldosa ketosa 2-2 P P P P 3 Glukosa-6-fosfát (G-6P) Glukosa-6-fosfát (otevřená forma) Fruktosa-6-fosfát (otevřená forma) Fruktosa-6-fosfát (F-6P)
30 Fosfofruktokinasa Allosterický enzym. Klíčová reakce nejen glykolýzy. 2-3 P ATP Fosfofruktokinasa 2-3 P 2 2 P ADP + + Fruktosa-6-fosfát (F-6P) Fruktosa-1,6-bisfosfát (F-1,6-BP)


31 Druhý stupeň glykolýzy Vytváří se dva vzájemně převoditelné triosafosfáty z hexosabisfosfátu.
32 Aldolasa Reakce je zvratem aldolové kondenzace. 2-2 P P 3 Dihydroxyacetonfosfát (DAP) 2-2 P 3 Aldolasa + Glyceraldehyd-3-fosfát (GAP) 2 P 3 2- Fruktosa-1,6-bisfosfát (otevřená forma)
33 Triosafosfátisomerasa Izomerace aldosa ketosa. Reakce je rychlá a reversibilní. V rovnováze se tvoří 96 % ketosy. V dalším průběhu se využívá aldosa. Triosafosfátisomerasa 2 P P 3 2- Dihydroxyacetonfosfát (DAP) Glyceraldehyd-3-fosfát (GAP)


34 Třetí stupeň glykolýzy xidací tříuhlíkatých fragmentů se tvoří ATP. xidují se obě triosy.
35 Glyceraldehyd-3-fosfátdehydrogenasa 2 P P + NAD + Glyceraldehyd-3-fosfátdehydrogenasa + P i + NAD P 3 2- Glyceraldehyd-3-fosfát (GAP) 1,3-Bisfosfoglycerát (1,3-BPG)
36 Glyceraldehyd-3-fosfátdehydrogenasa 1,3-bisfosfoglycerát je acylfosfát s vysokým potenciálem přenosu fosfátu na ADP za tvorby ATP. + NAD + xidace NAD + 2 P P P i Tvorba acyl-fosfátu (dehydratace) P P P 3 2-
37 Fosfoglycerátkinasa Fosforylace na úrovni substrátu. Vzhledem ke dvěma triosafosfátům se vytváří 2 x ATP. 2- P 3 + ADP Fosfoglycerátkinasa - + ATP 2 P 3 2-1,3-Bisfosfoglycerát 2 P Fosfoglycerát
38 Fosfoglycerátmutasa Reakce vyžaduje katalytické množství 2,3-bisfosfoglycerátu. - - Fosfoglycerátmutasa 2- P 3 2- P 3 3-Fosfoglycerát 2-Fosfoglycerát
39 Enolasa Dehydratace 2-fosfoglycerátu za tvorby 2-fosfoenolpyruvátu. Fosfoenolpyruvát má vysoký potenciál přenosu fosfátu na ADP za tvorby ATP. - P 3 2- Enolasa - 2- P Fosfoglycerát Fosfoenolpyruvát (PEP)
40 Pyruvátkinasa Dochází k přechodu enol-keto. Fosfát ve fosfoenolpyruvátu je vázán na nestabilní enol formu. Po přenosu fosfátu na ADP se stabilizuje keto forma P 3 Pyruvátkinasa - Fosfoenolpyruvát (PEP) ADP + + ATP 3 Pyruvát


41 Zachování redoxní rovnováhy Reoxidace NAD ve svalech vede ke tvorbě laktátu, u kvasinek (Sacharomycet) ke tvorbě ethanolu. Enzymy: laktátdehydrogenasa pyruvátdekarboxylasa alkoholdehydrogenasa
.")
42 Různé dráhy pyruvátu Laktát a ethanol se tvoří za účasti NAD (anaerobně). Acetyl oa se tvoří v mitochondrii v pyruvátdehydrogenasovém komplexu.
43 Tvorba ethanolu kvasinkami a dalšími mikroorganismy Pyruvátdekarboxylasa má jako koenzym thiaminpyrofosfát (z thiaminu, B 2 ). Druhým stupněm je redukce ethanalu NAD alkoholdehydrogenasou. - Pyruvátdekarboxylasa Alkoholdehydrogenasa 3 Pyruvát Ethanal (acetaldehyd) NAD + + NAD + 3 Ethanol
44 Laktátdehydrogenasa Enzym u řady mikroorganismů (laktátová fermentace). Ve svalech za nedostatku kyslíku anaerobní stav. - Laktátdehydrogenasa - 3 Pyruvát NAD + + NAD + 3 Laktát
45 Energetický výtěžek konverze glukosy na pyruvát Glukosa + 2 P i + 2 ADP + 2 NAD + = = 2 pyruvát + 2 ATP + 2 NAD Za anaerobních podmínek je energetický výtěžek: kj.mol -1 Výtěžek je jen zlomkem energie, kterou je možné získat z glukosy. Později srovnáme s aerobním výtěžkem.

46 Vstup galaktosy a fruktosy do glykolýzy
47 Převedení galaktosa-1-fosfátu na glukosa-1-fosfát Enzymy: 2 P Uridin P P - - Galaktosa-1-fosfát UDP-glukosa galaktosa-1-fosfáturidyltransferasa Galaktosa-1-fosfáturidyltransferasa UDP-galaktosa-4-epimerasa UDP-glukosa se regeneruje. 2 P - P - Uridin + 2 P 2- UDP-galaktosa Glukosa-1-fosfát 2 UDP-galaktosa-4-epimerasa P P - - Uridin UDP-glukosa
48 Laktasa štěpí laktosu na glukosu a galaktosu Laktasa + Laktosa Galaktosa Glukosa
49 Katarakt šedý zákal Porucha aktivity galaktosa-1-fosfáturidyltransferasy. Prezence aldosareduktasy v oční čočce vede ke tvorbě galacitolu, který je osmoticky aktivní a vtahuje do čočky vodu. Aldosareduktasa NAD + + NAD + 2 Galaktosa (cukerný aldehyd) 2 Galacitol (cukerný alkohol)
50 Laktosová intolerance Deficit laktasy, která štěpí laktosu na galaktosu a glukosu. Důsledky: U dětí klesá aktivita laktasy po odstavení na 10% hodnot po narození. Laktosa je zdroj energie pro střevní mikroorganismy, které ji fermentují na laktát a plyny (methan a vodík). Způsobuje to nadýmání. Vzniklý laktát je osmoticky aktivní, což způsobuje, že voda je vtahována do střev průjem. Způsobuje to také, že nejsou tráveny tuky a proteiny. Narušení metabolismu galaktosy galaktosemie. Kromě průjmů a možnosti narušení funkce jater, znamená zvýšená hladina galaktosy možnost tvorby očního zákalu.

51 V játrech vstupuje fruktosa do glykolýzy cestou fruktosa-1-fosfátu Enzymy: Fruktokinasa Fruktosa-1-fosfátaldolasa Triosakinasa.
52 Alternativní fosforylace fruktosy adiposní tkáni Alternativně může být fruktosa fosforylována na fruktosa-6- fosfát hexokinasou. Afinita hexokinasy ke glukose je však 20x vyšší než k fruktose. V játrech se proto může tvořit jen malé množství fruktosa-6-fosfátu. Analogické je to ve svalech. Fosforylace fruktosy na fruktosa-6-fosfát probíhá v adiposních buňkách, kde je převaha fruktosy.
53 Kontrolní mechanismy glykolýzy Kontrola glykolýzy je důležitá z těchto dvou důvodů: 1. Tvoří se ATP rozkladem glukosy na pyruvát 2. Tvoří se stavební jednotky k syntéze, např. mastných kyselin. V metabolických drahách jsou vhodným místem kontroly enzymy katalyzující prakticky ireversibilní reakce. Glykolýza: hexokinasa fosfofruktokinasa pyruvátkinasa

54 Fosfofruktokinasa klíčový enzym regulace savčí glykolýzy Jaterní enzym je 340 kda homotetramer obsahující katalytická a allosterická místa.
55 Inhibice a aktivace fosfofruktokinasy ATP je allosterický inhibitor; AMP ruší inhibiční účinek ATP. Aktivita enzymu roste, když poměr ATP / AMP klesá. Proč AMP a ne ADP? Při rychlém úbytku ATP se vznikající ADP rychle přeměňuje adenylátkinasou. ADP + ADP = ATP + AMP Malé změny v koncentraci ATP vedou k velkým změnám koncentrace AMP a tím ke zvýšení citlivosti regulace fosfofruktokinasy. Fosfofruktokinasa je také inhibována snížením p. Je to prevence tvorby nadbytku laktátu. Fosfofruktokinasa je inhibována citrátem. Nadbytek citrátu je znamením nadbytku biosyntetických prekurzorů. Není nutné odbourávat další glukosu. Aktivátorem fosfofruktokinasy je fruktosa-2,6-bisfosfát (F-2,6-bP). Aktivuje enzym tím, že zvyšuje jeho afinitu pro substrát. F-2,6-bP je allosterický aktivátor, který posouvá tetramerní enzym ze stavu T do stavu R.
56 2-3 P 2 P Fruktosa-2,6-bisfosfát (F-2,6-BP)
57 Allosterická inhibice fosfofruktokinasy ATP Nízká hladina ATP Reakční rychlost Vysoká hladina ATP [Fruktosa-6-fosfát]
58 A) Aktivace fosfofruktokinasy fruktosa-2,6-bisfosfátem B) ATP jako substrát, nadbytek inhibuje, přidání F-2,6-bP ruší inhibiční účinek ATP A) B) µm F-2,6-BP 100 Relativní rychlost µm 0 µm Relativní rychlost µm 0.1 µm 1 µm F-2,6-BP [Fruktosa-6-fosfát] (µm) [ATP] (µm)
59 Kontrola hladiny F-2,6-BP Fruktosa-2,6-bisfosfát se tvoří za katalýzy fosfofruktokinasou 2 (PFK2) a je hydrolyzována fruktosabisfosfatasou 2 (FBPasa2), což je bifukční enzym. Existuje v pěti isoenzymových formách. Forma L převažuje v játrech a forma M ve svalech. Forma L se podílí na udržování homeostéze krevní glukosy. Při vysoké hladině glukosy v krvi (insulin) se současně zvyšuje hladina fruktosa-6-fosfátu v játrech, což vede ke zvýšené tvorbě F-2,6-BP a tím ke zvýšení aktivity fosfofruktokinasy. Jaké kontrolní mechanismy fungují v játrech ve vztahu PFK2 a FBPasy2? Aktivity PFK2 a FBPasy2 jsou recipročně kontrolovány fosforylací Ser zbytku. Při nízké hladině glukosy (signalizuje glukagon přes proteinkinasovou kaskádu s camp) dojde k fosforylaci bifunkčního enzymu proteinkinasou A, což má za následek aktivaci FBPasy2 a inhibici PFK2. Snižuje se hladina F- 2,6-BP a zpomaluje se glykolýza. Při vysoké hladině glukosy, ztrácí bifunkční enzym fosfát, aktivuje se PFK2 a inhibuje FBPasa2, zvyšuje se hladina fruktosa-2,6-bisfosfátu a zrychluje glykolýza.
60 Úloha hexokinasy při regulaci glykolýzy exokinasa je inhibována produktem glukosa-6-fosfátem. Inhibice fosfofruktokinasy vede také k inhibici hexokinasy. Když je fosfofruktokinasa inaktivní, roste hladina fruktosa-6-fosfátu a tím i glukosa-6-fosfátu. V játrech je glukokinasa, která fosforyluje glukosu při vysokých koncentracích (glukokinasa je 60 x méně afinní ke glukose). Proto je rolí glukokinasy spíše fosforylovat glukosu pro tvorbu glykogenu a mastných kyselin. Dalším důvodem proč je klíčovým enzymem regulace glykolýzy fosfofruktokinasa a ne hexokinasa je, že glukosa-6-fosfát není pouze meziproduktem glykolýzy. Může přecházet na glykogen nebo se katabolizovat v pentosafosfátové dráze.


61 Úloha pyruvátkinasy při regulaci glykolýzy. Regulace pyruvátkinasy a regulace prostřednictvím F-2,6-BP brání při nízké hladině spotřebu glukosy játry ve prospěch mozku.

62 Reakční mechanismus glyceraldehyd-3-fosfátdehydrogenasy V aktivním místě jsou ys a is zbytky vázané nekovalentně na NAD +
63 Katalytický mechanismus glyceraldehyd-3-fosfátdehydrogenasy Glyceraldehyd-3-fosfát NAD + N N N 2 N N 2 N R 2 + N+ + R 1 S 1 R 2 + N+ + R 1 S emithioacetal
64 R 2 + N+ + N 2 R 1 S N N xidace 2 R 2 NAD N N 2 S N R 1 + N emithioacetal Thioesterový meziprodukt
65 R 2 NAD N N 2 N + N NAD + + R 2 N+ + N 2 N + N S R 1 NAD + NAD S R 1 Thioesterový meziprodukt Thioesterový meziprodukt
66 NAD + + R 2 N+ + N 2 S R 1 N + N Fosforylace P i 3 NAD + + R 2 N+ + N P S N R 1 N Thioesterový meziprodukt
67 GLUKNEGENEZE Syntéza glukosy z necukerných prekurzorů: Laktát, aminokyseliny (uhlíkatý řetězec glukogenních aminokyselin při hladovění) a glycerol. lavním místem glukoneogeneze jsou játra, malé množství v ledvinách, něco málo v mozku, kosterních svalech a srdečním svalu. Glukoneogeneze není zvratem glykolýzy. Denní spotřeba glukosy mozkem u dospělého člověka je 120 g, což je většina spotřeby těla (160 g). V tělních tekutinách je 20 g glukosy a zásoba ve formě glykogenu je 190 g. elkově je v těle zásoba glukosy asi na jeden den.
68 Ireversibilní kroky glykolýzy Aktuální G tvorby pyruvátu z glukosy je 84 kj/mol. Tři kroky jsou kritické (ireversibilní): a) exokinasa ( G = - 33 kj/mol) b) Fosfofruktokinasa ( G = -22 kj/mol) c) Pyruvátkinasa ( G = - 17 kj/mol)
69 Srovnání glykolýzy a glukoneogeneze I. Glukosa P i Glukosa-6-fosfatasa 2 Glukosa-6-fosfát ATP exokinasa ADP P i Fruktosabisfosfatasa Fosfoglukosaisomerasa Fruktosa-6-fosfát ATP ADP 2 Fruktosa-1,6-bisfosfát Aldolasa Fosfofruktokinasa Dihydroxyacetonfosfát Triosafosfátisomerasa Glyceraldehyd-3-fosfát
70 Srovnání glykolýzy a glukoneogeneze II. Dihydroxyacetonfosfát Triosafosfátisomerasa Glyceraldehyd-3-fosfát NAD + + P i NAD + + P i Glyceraldehyd-3-fosfátdehydrogenasa NAD + + NAD + + 1,3-Bisfosfoglycerát ADP ATP Fosfoglycerátkinasa ATP ADP 3-Fosfoglycerát Fosfoglycerátmutasa 2-Fosfoglycerát Enolasa 2 + GDP Fosfoenolpyruvátkarboxykinasa GTP Fosfoenolpyruvát ADP Pyruvátkinasa ATP xaloacetát Pyruvát P i + ADP ATP + 2 Pyruvátkarboxylasa
71 Karboxylace pyruvátu Překonání prvního ireversibilního kroku anaplerotická reakce pro cyklus trikarboxylových kyselin. Probíhá v matrix mitochondrie. Účastní se biotin jako prosthetická skupina pyruvátkarboxylasy (vázáno na koncovou aminoskupinu Lys). Pyruvátkarboxylasa je mitochondriální enzym, zatímco ostatní enzymy glukoneogeneze jsou cytoplasmatické. Reakce probíhá jen za přítomnosti acetyloa, který se na pyruvátkarboxylasu váže. AcetyloA allostericky aktivuje pyruvátkarboxylasu. Proč?
72 Pyruvátkarboxylasa a fosfoenolpyruvátkarboxykinasa (PEPK) Karboxylace probíhá v matrix. xaloacetát je redukován NAD malátdehydrogenasou na malát, který je transportován do cytosolu, kde je dekarboxylován a fosforylován. Dekarboxylace pohání jinak endergonní reakci. P Pyruvátkarboxylasa PEPK 2 - Pyruvát ATP ADP + P GTP i GDP + xaloacetát 2 Fosfoenolpyruvát (PEP)
73 Biotin N N S 4 Biotin Postranní řetězec (kyselina valerová)
74 Karboxybiotinylpyruvátkarboxylasa (biotin jako prosthetická skupina) - N N S ( 2 ) 4 Karboxybiotinyl-enzym N ( 2 ) 4 N Postranní řetězec lysinu
75 Mechanismus tvorby karboxybiotinu reakcí hydrogenuhličitanu s ATP. Adenosin P P P ATP ADP P - - Karboxyfosfát P i Biotinyl-enzym - N N + N N S ( 2 ) 4 N ( 2 ) 4 E S ( 2 ) 4 N ( 2 ) 4 E Karboxybiotinyl-enzym Biotinyl-enzym
76 Reakce karboxybiotinu s pyruvátem za tvorby oxaloacetátu přes enolformu pyruvátu jako meziproduktu (matrix) N Pyruvát N N - N Biotinyl-enzym N N Enolforma pyruvátu Karboxybiotinyl-enzym - xaloacetát
77 Reakce oxaloacetátu s GTP za katalýzy fosfoenolpyruvátkarboxykinasy je v cytosolu poháněna dekarboxylací oxaloacetátu. P P P - P - Guanosin - GDP + 2 PEPK 2 - xaloacetát GTP Fosfoenolpyruvát (PEP)
78 Další dva ireversibilní kroky: 2. Převedení fruktosa-1,6-bisfosfátu na fruktosa-6-fosfát: Fruktosa-1,6-bisfosfát + 2 Fruktosa-6-fosfát + P i Enzym: Fruktosa-1,6-bisfosfatasa. Allosterický enzym-aktivován citrátem, inhibován fruktosa-2,6-bisfosfátem a AMP. 3. Glukosa-6-fosfát tvorba volné glukosy. Ve většině tkání končí glukoneogeneze na tomto stupni (syntéza glykogenu atd.). Tvorba volné glukosy vyžaduje regulaci enzymu glukosa-6-fosfatasy. Enzym je přítomen jen ve tkáních, které udržují fyziologickou hladinu glukosy v krvi játra a částečně ledviny.
 působením glukosa-6-fosfatasy T1 transportuje G-6-P do")

79 Tvorba volné glukosy v dutinkách endoplasmatického retikula (ER) působením glukosa-6-fosfatasy T1 transportuje G-6-P do ER, T2 a T3 transportují P i a glukosu zpět do cytosolu. Glukosa-6-fosfatasa je stabilizována a 2+ -vazebným proteinem (SP)
80 Dráhy transportu oxaloacetátu Malát - aspartátový člunek (shutle). Srdeční sval a játra. Univerzální člunek - směr toku elektronů závisí na NAD / NAD + - ytosol Vnitřní mitochondrionální membrána Mitochondrion NAD + Malát Malát NAD Malátdehydrogenasa NAD Aminokyselina Aspartátaminotransferasa xaloacetát Dráha 1 Dráha 2 xaloacetát Malátdehydrogenasa NAD Aminokyselina Aspartátaminotransferasa α-ketokyselina - Aspartát Aspartát α-ketokyselina N + 3 N Glukoneogeneze PEP PEP
81 Glycerol-3-fosfátový člunek reoxidace NAD z glykolýzy za aerobních podmínek. Typické pro intenzivně pracující sval. Enzym je cytosolární glycerol-3-fosfátdehydrogenasa. Akceptorem elektronů FAD. Elektrony se transportují proti NAD koncentračnímu gradientu. NAD + + NAD + 2 ytosolická glycerol-3-fosfát dehydrogenasa 2 2 P P 3 2- Dihydroxyacetonfosfát ytosol Glycerol-3-fosfát Mitochondrionální glycerol-3-fosfát dehydrogenasa E-FAD 2 E-FAD Q 2 Q Matrix
82 esta glycerolu, který se tvoří hydrolýzou komplexních (zmýdelnitelných) tuků do glykolýzy / glukoneogeneze. 2 Glycerolkinasa 2 Glycerol-3-fosfátdehydrogenasa 2 2 ATP ADP 2 P 3 2- NAD + NAD P 3 2- Glycerol Glycerol-3-fosfát Dihydroxyacetonfosfát
83 Reciproká regulace glykolýzy a glukoneogeneze Glukoneogeneze a glykolýza jsou dva protichůdné pochody ideální regulace = jeden pochod aktivní a druhý neaktivní. Teoreticky mohou být oba pochody aktivní, protože jsou za podmínek v buňce exergonické. Množství a aktivity různých enzymů obou drah jsou pod kontrolou a proto nejsou obě dráhy současně vysoce aktivní. Rychlost glykolýzy je také dána koncentrací glukosy a rychlost glukoneogeneze koncentrací laktátu a dalších prekurzorů glukosy. Množství enzymů je kontrolováno hormonálně. ormony ovlivňují expresi genů a regulují degradaci mrna. Insulin, signál sytosti, stimuluje expresi fosfofruktokinasy, pyruvátkinasy a bifunkčního enzymu, který vede k tvorbě a degradaci fruktosa-2,6- bisfosfátu. Glukagon, signál hladovění, inhibuje expresi těchto enzymů a stimuluje tvorbu fosfoenolpyruvátkarboxykinasy a fruktosa-1,6-bisfosfatasy. Kontrola přes transkripci je pomalá.

84 Tři klíčové kroky glykolýzy a glukoneogeneze s vyznačením změn Gibbsovy energie v kj.mol -1

85 Reciproká regulace glykolýzy a glukoneogeneze v játrech
86 ormonální regulace glukoneogeneze při hladovění Nízká hladina glukosy v krvi (hladovění) Zvýšená sekrece glukagonu Zvýšená hladina [camp] Zvýšená rychlost fosforylace bifunkčního enzym Fosforylace bifunkčního enzymu proteinkinasou A, což má za následek aktivaci FBPasy2 a inhibici PFK2. Snižuje se hladina F-2,6- BP a zpomaluje se glykolýza. Inhibice fosfofruktokinasy a aktivace fruktosabisfosfatasy Zvýšená glukoneogeneze
87 Stechiometrie glukoneogeneze a zvratu glykolýzy Glukoneogeneze: Pyruvát + 4 ATP + 2 GTP + 2 NAD = = glukosa + 4 ADP + 2 GDP + 6 P i + 2 NAD G o = - 38 kj.mol -1 Na syntézu glukosy je spotřebováno 6 nukleosidtrifosfátů. Energeticky nevýhodnou (endergonní) reakci pohání hydrolýza ATP a GTP. Zvrat glykolýzy: 2 Pyruvát + 2 ATP + 2 NAD = glukosa + 2 ADP + 2 P i + 2 NAD + G o = + 84 kj.mol -1 Při odbourávání glukosy se získají jen 2 ATP.
88 Substrátové cykly Dvojice reakcí jako jsou fosforylace fruktosa-6-fosfátu na fruktosa-1,6- bisfosfát a jeho hydrolýza zpět na fruktosa-6-fosfát se nazývají SUBSTRÁTVÉ YKLY. bě nebývají současně plně aktivní. Přesto dochází často současně k oběma reakcím je to nedokonalost těchto reakcí cyklují tyto cykly se také nazývají JALVÉ (futile cycles). Jsou biologicky zajímavé. Jednou z jejich možných funkcí je zesílení metabolických signálů. Druhou možnou funkcí je produkce tepla hydrolýzou ATP. Příkladem je čmelák, který může za potravou již při 10 o. Je schopen dosáhnout potřebnou teplotu v hrudi současnou vysokou aktivitou fosfofruktokinasy a fruktosa-1,6-bisfosfatasy. ydrolýza ATP vytváří teplo. Tato bisfosfatasa není inhibována AMP!! To znamená, že enzym je určen k produkci tepla. Včela, která nemá v létacích svalech bisfosfatasu, nemůže při nízkých teplotách létat. U lidí existuje onemocnění maligní hypertermie, kdy dochází ke ztrátě kontroly, oba procesy probíhají současně plně a generují TEPL.

89 Substrátové cykly ykly poháněné ATP probíhají dvěma různými rychlostmi. Malá změna v rychlostech opačných reakcí vede k velké výsledné změně rychlosti.

90 Přehled možných drah glukosa-6-fosfátu
91 oriho cyklus Laktát a alanin tvořící se v kontraktilním svalstu jsou zdrojem energie pro jiné orgány. Tvořící se pyruvát ve svalech při intenzivním cvičení se nestačí odbourat aerobně a pokračování glykolýzy závisí na dostupnosti NAD +. Tvoří se laktát. Laktát je transportován krví do jater a zde je resyntetizována glukoneogenezí glukosa, která putuje do svalů. Alanin je druhým zdrojem uhlíku pro syntézu glukosy. Ve svalech je tvořen transaminací z pyruvátu, v játrech probíhá opačný proces. Alanin tak pomáhá udržovat rovnováhu dusíku v organismu. Erythrocyty postrádají mitochondrie a proto nemohou oxidovat kompletně glukosu.
92 Aminotransferasová reakce alanin pyruvát Koenzymem je pyridoxal-5-fosfát (PALP). 3 - Aminotransferasa 3 - N 3 + α-ketokyselina Aminokyselina Alanin Pyruvát


93 oriho cyklus
94 Isoenzymové formy laktátdehydrogenasy Laktátdehydrogensa katalyzuje vzájemný převod laktát pyruvát. LD je tetramer dvou typů 35 kd podjednotek. (heart) typ převažuje v srdečním svalu a M (muscle) v kontraktilním svalstvu a játrech. Podjednotky asociují tak, že vytvářejí pět typů tetramerů: 4, 3 M 1, 2 M 2, 1 M 3, a M 4 4 isoenzym má vyšší afinitu k substrátu než M 4, který je allostericky inhibován vysokou hladinou pyruvátu. 4 oxiduje laktát na pyruvát, který využívá srdeční sval za aerobních podmínek. Srdeční sval je vždy aerobní! M 4 funguje opačně převádí pyruvát na laktát což je v souladu s glykolýzou za anaerobnmích podmínek. statní isoenzymy mají vlastnosti mezi těmito dvěma krajními.


95 Vzájemná interakce glykolýzy a glukoneogeneze na úrovni tělesných orgánů
96 Pentosafosfátová dráha
97 V pentosafosfátové dráze se tvoří NADP a syntetizují se pentosy Pentosafosfátová dráha je pro všechny organismy zdrojem NADP pro reduktivní biosyntézy. Pentosafosfátová dráha má dvě fáze: A) xidační tvorbu NADP B) Neoxidační přeměnu sacharidů. Přeměna 3, 4, 5, 6 a 7 sacharidů na pentosy (nukleotidy) a přebytku 5 na meziprodukty glykolýzy. Tkáně s aktivní pentosafosfátovou dráhou (cytosol): nadledvinky, játra, varlata, adipozní tukové buňky, vaječníky, mléčná žláza a červené krvinky (erythrocyty). Základní schéma: Glukosa-6-fosfát + 2 NADP ribosa-5-fosfát + 2 NADP
98 Metabolické dráhy s potřebou NADP: Syntézy: Mastné kyseliny holesterol Neurotransmitery Nukleotidy Detoxifikace: Redukce oxidovaného glutathionu ytochrom P450 monooxygenasa
99 Biochemické reakce vzájemné přeměny sacharidů. 1. Izomerace aldosa / ketosa. Např. glukosa / fruktosa. 2. Epimerace změna konfigurace na chirálním uhlíku. Např. galaktosa / glukosa na Transketolasa přenos dvouhlíkatého štěou z ketosy na aldosu. 4. Transaldolasa přenos tříuhlíkatého štěpu z ketosy na aldosu. 5. Aldolasa štěpení resp. syntéza na principu aldolové kondenzace.


100 elkové schéma pentosafosfátové dráhy Ribulosa-5-fosfát je epimerován fosfopentosaepimerasou na xylulosa-5-fosfát a izomerován na ribosa-5-fosfát.
101 elkové schéma oxidační fáze pentosafosfátové dráhy: 2-2 P 3 Glukosa-6-fosfátdehydrogenasa 2-2 P 3 Laktonasa - 6-Fosfoglukonátdehydrogenasa NADP + NADP NADP + NADP P P 3 Glukosa-6-fosfát 6-Fosfoglukono-δ-lakton 6-Fosfoglukonát Ribulosa-5-fosfát
102 Reakce katalyzovaná glukosa-6-fosfátdehydrogenasou 2-2 P P 3 Glukosa-6-fosfátdehydrogenasa NADP + NADP + + Glukosa-6-fosfát 6-Fosfoglukono-δ-lakton
103 Reakce katalyzovaná laktonasou P 3 Laktonasa P Fosfoglukono-δ-lakton 6-Fosfoglukonát
104 Druhá oxidace za katalýzy 6-fosfoglukonátdehydrogenasou - 6-Fosfoglukonátdehydrogenasa NADP + NADP P Fosfoglukonát 2 P 3 2- Ribulosa-5-fosfát
105 Propojení pentosafosfátové dráhy s glykolýzou. Nadbytek pentos je přeměněn na meziprodukty glykolýzy. Schematicky: Výsledná suma reakcí: Xylulosa-5-fosfát + ribosa-5-fosfát 2 fruktosa-6-fosfát + glyceraldehyd-3-fosfát Analogicky: 3 Ribosa-5-fosfát 2 fruktosa-6-fosfát + glyceraldehyd-3-fosfát
106 Neoxidační fáze pentosafosfátové dráhy za účasti transketolasy 2 + Transketolasa P P 3 2 P P 3 2- Ribulosa-5-fosfát Ribosa-5-fosfát Glyceraldehyd-3-fosfát Sedoheptulosa-7-fosfát
107 Neoxidační fáze pentosafosfátové dráhy za účasti transaldolasy 2 2 Transaldolasa P 3 2 P P 3 2 P 3 2- Glyceraldehyd-3-fosfát Sedoheptulosa-7-fosfát Fruktosa-6-fosfát Erythrosa-4-fosfát
108 Neoxidační fáze pentosafosfátové dráhy transketolasa. + 2 Transketolasa P P P 3 2 P 3 2- Erythrosa-4-fosfát Xylulosa-5-fosfát Fruktosa-6-fosfát Glyceraldehyd-3-fosfát
109 Glukosa-6-fosfát je spotřebována jak glykolýzou, tak pentosafosfátovou drahou. Klíčovou roli v regulaci obou procesů hraje hladina NADP + v cytoplasmě. Dehydrogenace glukosa-6-fosfátu je ireversibilní proces. Nutná přítomnost NADP + jako příjemce elektronů. Z toho důvodu vyvolávají nízké hladiny NADP + inhibiční efekt dehydrogenace. Efekt je prohlubován skutečností, že NADP kompetuje s NADP + o aktivní místo enzymu. Poměr NADP + / NADP v jaterním cytosolu dobře živených krys je 0, 014, několika řády nižší než poměr NAD + / NAD za stejných podmínek je 700. NADP musí být bezprostředně spotřebován k syntézám. Neoxidační fáze pentosafosfátové dráhy je regulována dostupností substrátů.

110 Vstup glukosa-6-fosfátu do pentosafosfátové dráhy závisí na potřebě NADP, ribosa-5-fosfátu a ATP. Možnost 1-je třeba mnohem více ribosa-5-fosfátu než NADP.

111 Potřeba NADP a ribosa-5-fosfátu je vyvážená

112 Potřeba NADP je mnohem vyšší než potřeba ribosa-5-fosfátu.
113 Možnost 3 Glukosa-6-fosfát je kompletně oxidována na 2. Aktivní jsou tři skupiny reakcí: 1. xidativní fáze produkuje dvě molekuly NADP a jednu molekulu ribosa-5-fosfátu. 6 Glukosa-6-fosfátů + 12 NADP = 6-ribosa-5- fosfátů + 12 NADP Ribosa-5-fosfát je převedena na fruktosa-6-fosfát a glyceraldehyd-3-fosfát tranketolasou a transaldolasou. 6 Ribosa-5-fosfátů = 4 fruktosa-6-fosfáty + 2 GAP 3. Glukosa-6-fosfát je resyntetizována z fruktosa-6- fosfátu a glyceraldehyd-3-fosfátu glukoneogenezí. 4 Fruktosa-6-fosfáty + 2 GAP + 2 = 5 G-6-P + +

114 Potřeba NADP a ATP je vyrovnaná
115 Stechiometrie možností 1 a 2 Možnost 1 : 5 glukosa-6-fosfát + ATP 6 ribosa-5-fosfát + ADP + + Možnost 2 : Glukosa-6-fosfát + 2 NADP + 2 ribosa-5-fosfát + 2 NADP
116 Rovnice možnosti 3. Možnost 3 : 6 Glukosa-6-fosfát + 12 NADP ribosa-5-fosfát + 12 NADP ribosa-5-fosfát 4 fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát 4 fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát glukosa-6-fosfát + P i Suma tří reakcí: Glukosa-6-fosfát + 12 NADP NADP P i
117 Rovnice možnosti 4. 3 Glukosa-6-fosfát + 6 NADP NAD P i + 8 ADP 5 pyruvát NADP + 5 NAD + 8 ATP
118 K mechanismu transketolasy a transaldolasy Při transketolasové reakci je dvojuhlíkatý štěp vázán na koenzym, kterým je thiaminpyrofosfát (TPP). Transketolasa je homologní s enzymem E 1 pyruvátdehydrogenasového komplexu. Na aldosu se přenáší aktivovaný glykolaldehyd. Transaldolasa přenáší tříuhlíkatý dihydroxyaceton a na rozdíl od transketolasy, nemá prosthetickou skupinu. Zde se vytváří Schiffova báze mezi karbonylem ketosy (substrát) a ε- aminoskupinou Lys v aktivním místě enzymu. Tříuhlíkatý štěp vázaný na Lys je přenesen na aldosu. Je to analogie mechanismu aldolasy, enzymy jsou homologní.
119 Rovnováha mezi glykolýzou a glukoneogenezí v játrech vliv hladiny glukosy v krvi. lavním regulátorem glykolýzy a glukoneogeneze v játrech je fruktosa-2,6-bisfosfát. Fruktosa-2,6-bisfosfát aktivuje fosfofruktokinasu a inhibuje fruktosa 2,6-bisfosfatasu!!! Proč?? Při nízké hladině glukosy v krvi se z fruktosa-2,6- bisfosfátu uvolňuje fosfát za tvorby fruktosa-6-fosfátu. Fruktosa-6-fosfát se neváže jako aktivátor fosfofruktokinasu fosfofruktokinasu (váže se jen do akt. místa). Jakým způsobem je kontrolována koncentrace fruktosa- 2,6-bisfosfátu??? Na kontrole hladiny fruktosa-2,6-bisfosfátu se podílejí dva enzymy.
120 Fruktosa-2,6-bisfosfát.
121 Kontrolní mechanismus syntézy a odbourání fruktosa-2,6- bisfosfátu.
122 Rovnováha mezi glykolýzou a glukoneogenezí v játrech vliv hladiny glukosy v krvi. Fruktosa-2,6-bisfosfát je produktem reakce katalyzované fosfofruktokinasou 2 (PFK2). dštěpení fosfátu je katalyzované enzymem fruktosabisfosfatasa2 (FBPasa2) ba enzymy jsou součástí jednoho proteinového řetězce o délce 55 kd bifunkční enzym!!! o kontroluje zda bude aktivní PFK2 nebo FBPasa2? Za situace hladovění: Při nízké hladině glukosy je vylučován slinivkou hormon glukagon, který spouští uvnitř buněk camp kaskádu vedoucí k fosforylaci bifunkčního enzymu proteinkinasou A. Tato kovalentní modifikace aktivuje FBPasu2 a inhibuje PFK2. Převládá glukoneogeneze. Glukagon také aktivuje pyruvátkinasu v játrech. V opačném případě, při dostatku glukosy, je fosfát odštěpen, aktivuje se PFK2 a inhibuje FBAsa2. Urychluje se glykolýza!!!
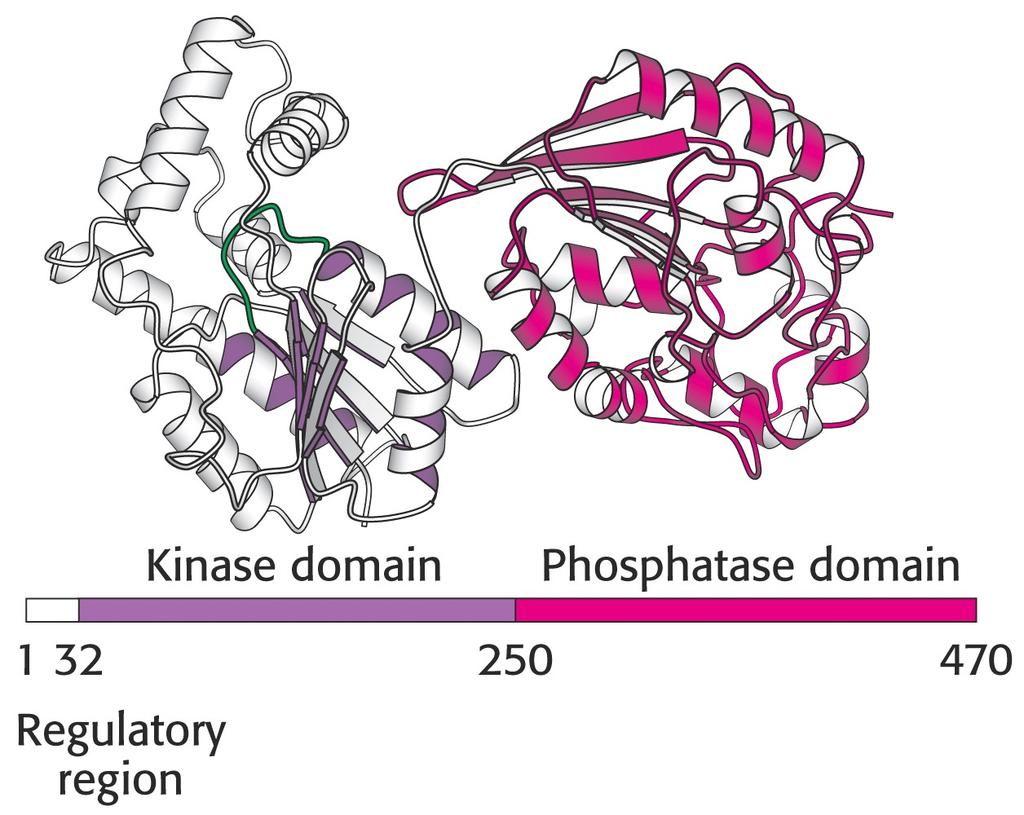
123 Bifunkční enzym fosfofruktokinasa-fosfofruktobisfosfatasa.
124 Pyruvátkinasa. Pyruvátkinasa je tetramer (57 kd podjednotka). Existuje řada izoenzymových forem kódovaných různými geny. L typ převažuje v játrech a M ve svalech a mozku. Jaterní enzym podléhá na rozdíl od svalového allosterické regulaci. Katalytická aktivita L formy je kontrolována reversibilní fosforylací. Aktivita M formy ne. Při nízké hladině glukosy funguje glukagon. Spouští se camp kaskáda dochází k fosforylaci pyruvátkinasy a tím ke snížení její aktivity. Tato hormonální regulace zabraňuje spotřebě glukosy játry!!! Glukosa je nutně potřebná k činnosti mozku a svalstva.
125 Kontrolní systém katalytické aktivity pyruvátkinasy. Regulace allosterickou kontrolou aktivátory, inhibitory a kovalentní modifikací.
METABOLISMUS SACHARIDŮ
 METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
METABLISMUS SAHARIDŮ GLUKNEGENEZE GLUKNEGENEZE entrální úloha glukosy Palivo Prekursor strukturních sacharidů a jiných molekul Syntéza glukosy z necukerných prekurzorů Laktát Aminokyseliny (uhlíkatý řetězec
Metabolismus sacharidů
 Základy biochemie KB / B Metabolismus sacharidů Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KB / B Metabolismus sacharidů Inovace studia biochemie prostřednictvím e-learningu Z.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Glykolýza a neoglukogenese
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Glykolýza a neoglukogenese z řečtiny glykos sladký, lysis uvolňování sled metabolických reakcí od glukosy přes fruktosa-1,6-bisfosfát
pátek, 24. července 15 GLYKOLÝZA
 pátek,. července 15 GLYKLÝZ sacharosa threalosa laktosa sacharasa threlasa laktasa D-glukosa D-fruktosa T T hexokinasa T hexokinasa glykogen - škrob fosforylasa D-galaktosa UD-galaktosa UD-glukosa fruktokinasa
pátek,. července 15 GLYKLÝZ sacharosa threalosa laktosa sacharasa threlasa laktasa D-glukosa D-fruktosa T T hexokinasa T hexokinasa glykogen - škrob fosforylasa D-galaktosa UD-galaktosa UD-glukosa fruktokinasa
CZ.1.07/2.2.00/ Obecný metabolismu. Metabolismus glukosy, glykolýza, glukoneogeneze (3).
.") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismu. Metabolismus glukosy, glykolýza, glukoneogeneze (3). Prof. RNDr. Pavel Peč, CSc.
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismu. Metabolismus glukosy, glykolýza, glukoneogeneze (3). Prof. RNDr. Pavel Peč, CSc.
1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu
 Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2018 1. Napište strukturní vzorce aminokyselin E a W a vzorce guanosinu a uracilu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Pentosový cyklus. osudy glykogenu. Eva Benešová
 Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Pentosový cyklus a osudy glykogenu Eva Benešová Pentosový cyklus pentosafosfátová cesta, fosfoglukonátová cesta nebo hexosamonofosfátový zkrat Funkce: 1) výroba NADPH 2) výroba ribosa 5-fosfátu 3) zpracování
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu
 Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Test pro přijímací řízení magisterské studium Biochemie 2019 1. Napište strukturní vzorce aminokyselin D a Y a vzorce adenosinu a thyminu U dalších otázek zakroužkujte správné tvrzení (pouze jedna správná
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Metabolismus sacharidů. VY_32_INOVACE_Ch0216.
 Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Odbourávání a syntéza glukózy
 Odbourávání a syntéza glukózy Josef Fontana EB - 54 Obsah přednášky Glukóza význam glukózy pro buňku, glykémie role glukózy v metabolismu transport glukózy přes buněčné membrány enzymy fosforylující a
Odbourávání a syntéza glukózy Josef Fontana EB - 54 Obsah přednášky Glukóza význam glukózy pro buňku, glykémie role glukózy v metabolismu transport glukózy přes buněčné membrány enzymy fosforylující a
Test pro přijímací řízení magisterské studium Biochemie Napište vzorce aminokyselin Q a K
 Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
Test pro přijímací řízení magisterské studium Biochemie 2017 1. Napište vzorce aminokyselin Q a K Dále zakroužkujte správné tvrzení (pouze jedna správná odpověď) 2. Enzym tyrozinkinasu řadíme do třídy
METABOLISMUS MONOSACHARIDŮ
 METABOLISMUS MONOSACHARIDŮ Metabolismus monosacharidů (zejména jejich katabolismus) je prakticky metabolismem glukosy. Ostatní monosacharidy z ní v případě potřeby vznikají, nebo jsou na ni několika reakcemi
METABOLISMUS MONOSACHARIDŮ Metabolismus monosacharidů (zejména jejich katabolismus) je prakticky metabolismem glukosy. Ostatní monosacharidy z ní v případě potřeby vznikají, nebo jsou na ni několika reakcemi
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
METABOLISMUS SACHARIDŮ PRINCIP Rozštěpené sacharidy vstřebávání střevní sliznicí do krevního oběhu dopraveny vrátnicovou žílou do jater. V játrech enzymaticky hexózy štěpeny na GLUKÓZU vyplavována do krve
OBECNÝ METABOLISMUS SACHARIDY I
 OBECNÝ METABOLISMUS SACHARIDY I Sacharidy Heterotrofní organismy (např. savci) sacharidy jsou hlavní živiny Autotrofní organizmy (např. rostliny) fixace C vedoucí k produkci sacharidů proces fotosyntézy
OBECNÝ METABOLISMUS SACHARIDY I Sacharidy Heterotrofní organismy (např. savci) sacharidy jsou hlavní živiny Autotrofní organizmy (např. rostliny) fixace C vedoucí k produkci sacharidů proces fotosyntézy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy
 9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
9. Citrátový cyklus, oxidační dekarboxylace pyruvátu a anaplerotické dráhy Obtížnost A Vyjmenujte kofaktory, které využívá multienzymový komplex pyruvátdehydrogenasy; které z nich řadíme mezi koenzymy
Cukry (Sacharidy) Sacharidy a jejich metabolismus. Co to je?
 Sacharidy a jejich metabolismus. Co to je?") Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
Sacharidy a jejich metabolismus Co to je? Cukry (Sacharidy) Organické látky, které obsahují karbonylovou skupinu (C=O) a hydroxylové skupiny (-O) vázané na uhlících Aldosy: karbonylová skupina na konci
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty
 sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
sloučeniny C, H, O Cukry = glycidy = sacharidy staré názvy: uhlohydráty, uhlovodany, karbohydráty triviální (glukóza, fruktóza ) vědecké (α-d-glukosa) organické látky nezbytné pro život hlavní zdroj energie
METABOLISMUS SACHARIDŮ
 METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
METABOLISMUS SAHARIDŮ A. Odbourávání sacharidů - nejdůležitější zdroj energie pro heterotrofy - oxidací sacharidů až na. získávají aerobní organismy energii ve formě. - úplná oxidace glukosy: složitý proces
Katabolismus. I. fáze aminokyseliny. II. fáze. III. fáze. proteiny. lipidy. polysacharidy. Glycerol + mastné kyseliny. glukosa ETS
 Katabolismus proteiny I. fáze aminokyseliny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS Acetyl-CoA (= AcSCoA nebo AcCoA) N 2 N N C 3 N N N C 2 C C C C 2 C 3 P P - 2 C
Katabolismus proteiny I. fáze aminokyseliny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS Acetyl-CoA (= AcSCoA nebo AcCoA) N 2 N N C 3 N N N C 2 C C C C 2 C 3 P P - 2 C
Energetický metabolizmus buňky
 Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Energetický metabolizmus buňky Buňky vyžadují neustálý přísun energie pro tvorbu a udržování biologického pořádku (život). Tato energie pochází z energie chemických vazeb v molekulách potravy (energie
Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech
 Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Citrátový cyklus Oxidace proteinů, tuků a cukrů jako zdroj energie v živých organismech 1. stupeň: OXIDACE cukrů, tuků a některých aminokyselin tvorba Acetyl-CoA a akumulace elektronů v NADH a FADH 2 2.
Propojení metabolických drah. Alice Skoumalová
 Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Propojení metabolických drah Alice Skoumalová Metabolické stavy 1. Resorpční fáze po dobu vstřebávání živin z GIT (~ 2 h) glukóza je hlavní energetický zdroj 2. Postresorpční fáze mezi jídly (~ 2 h po
Struktura sacharidů a nukleových kyselin
 Struktura sacharidů a nukleových kyselin Ústav lékařské chemie a klinické biochemie 2. LF UK a F Motol Matej Kohutiar 2017 snova přednášky I.Struktura sacharidů 1. Monosacharidy Reakce sacharidů 2. ligosacharidy
Struktura sacharidů a nukleových kyselin Ústav lékařské chemie a klinické biochemie 2. LF UK a F Motol Matej Kohutiar 2017 snova přednášky I.Struktura sacharidů 1. Monosacharidy Reakce sacharidů 2. ligosacharidy
Metabolismus sacharidů
 Metabolismus sacharidů Glukosa obsažená v celulose, škrobu a oligosacharidech nebo volná je nejrozšířenější organickou sloučeninou v přírodě. Pro chemotrofní organismy jsou sacharidy hlavní živinou, přičemž
Metabolismus sacharidů Glukosa obsažená v celulose, škrobu a oligosacharidech nebo volná je nejrozšířenější organickou sloučeninou v přírodě. Pro chemotrofní organismy jsou sacharidy hlavní živinou, přičemž
CZ.1.07/2.2.00/ Obecný metabolismu. Cyklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8).
 (8).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii Z.1.07/2.2.00/28.0171 becný metabolismu. yklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8). Prof. RNDr.
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii Z.1.07/2.2.00/28.0171 becný metabolismu. yklus trikarboxylových kyselin (citrátový cyklus, Krebsův cyklus) (8). Prof. RNDr.
Přehled energetického metabolismu
 Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
Přehled energetického metabolismu Josef Fontana EB 40 Obsah přednášky Důležité termíny energetického metabolismu Základní schéma energetického metabolismu Hlavní metabolické dráhy energetického metabolismu
SACHARIDY - Monosacharidy příručka pro učitele Obecné informace: Téma Monosacharidy se probírá v rozsahu jedné vyučovací hodiny.
 SACHARIDY - Monosacharidy příručka pro učitele Obecné informace: Téma Monosacharidy se probírá v rozsahu jedné vyučovací hodiny. Navazující učivo Před probráním tématu Monosacharidy probereme krátkou úvodní
SACHARIDY - Monosacharidy příručka pro učitele Obecné informace: Téma Monosacharidy se probírá v rozsahu jedné vyučovací hodiny. Navazující učivo Před probráním tématu Monosacharidy probereme krátkou úvodní
ANABOLISMUS SACHARIDŮ
 zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
zdroj sacharidů: autotrofní org. produkty fotosyntézy heterotrofní org. příjem v potravě důležitou roli hraje GLUKÓZA METABOLISMUS SACHARIDŮ ANABOLISMUS SACHARIDŮ 1. FOTOSYNTÉZA autotrofní org. 2. GLUKONEOGENEZE
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus. Metabolismus glykogenu (4). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus. Metabolismus glykogenu (4). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie, Přírodovědecká
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolismus sacharidů II
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus sacharidů II NUTNO ZNÁT VSTUP TĚCHTO ZÁKLADNÍCH MONOSACHARIDŮ DO GLYKOLÝZY Glykogen glukosa hlavní zdroj energie pro metabolismus
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolismus sacharidů II NUTNO ZNÁT VSTUP TĚCHTO ZÁKLADNÍCH MONOSACHARIDŮ DO GLYKOLÝZY Glykogen glukosa hlavní zdroj energie pro metabolismus
10. Metabolismus sacharidů
 10. Metabolismus sacharidů Obtížnost A Vysvětlete rozdíly v následujících dvojicích pojmů: aldosa/ketosa; redukující/neredukující sacharid; škrob/glykogen; homopolysacharid/heteropolysacharid; amylosa/amylopektin.
10. Metabolismus sacharidů Obtížnost A Vysvětlete rozdíly v následujících dvojicích pojmů: aldosa/ketosa; redukující/neredukující sacharid; škrob/glykogen; homopolysacharid/heteropolysacharid; amylosa/amylopektin.
SACHARIDY POJMY (odkaz na organickou chemii)
") Sacharidy SACARIDY PJMY (odkaz na organickou chemii) sacharidy, cukry, aldosy, ketosy monosacharidy, oligosacharidy, polysacharidy otevřené a cyklické formy monosacharidů, pyranosa, furanosa stereoisomerie
Sacharidy SACARIDY PJMY (odkaz na organickou chemii) sacharidy, cukry, aldosy, ketosy monosacharidy, oligosacharidy, polysacharidy otevřené a cyklické formy monosacharidů, pyranosa, furanosa stereoisomerie
Regulace metabolizmu lipidů
 Regulace metabolizmu lipidů Principy regulace A) krátkodobé (odpověď s - min): Dostupnost substrátu Alosterické interakce Kovalentní modifikace (fosforylace/defosforylace) B) Dlouhodobé (odpověď hod -
Regulace metabolizmu lipidů Principy regulace A) krátkodobé (odpověď s - min): Dostupnost substrátu Alosterické interakce Kovalentní modifikace (fosforylace/defosforylace) B) Dlouhodobé (odpověď hod -
Metabolické dráhy. František Škanta. Glykolýza. Repetitorium chemie X. 2011/2012. Glykolýza. Jaký je osud pyruátu bez přítomnosti kyslíku?
 Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
Repetitorium chemie X. 2011/2012 Metabolické dráhy František Škanta Metabolické dráhy xidativní fosforylace xidace mastných kyselin 1. fosforylace 2. štěpení hexosy na dvě vzájemně převoditelné triosy
POZNÁMKY K METABOLISMU SACHARIDŮ
 POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
POZNÁMKY K METABOLISMU SACHARIDŮ Prof.MUDr. Stanislav Štípek, DrSc. Ústav lékařské biochemie 1.LF UK v Praze Přehled hlavních metabolických cest KATABOLISMUS Glykolysa Glykogenolysa Pentosový cyklus Oxidace
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Metabolismus krok za krokem - volitelný předmět -
 Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Metabolismus krok za krokem - volitelný předmět - Vladimíra Kvasnicová pracovna: 411, tel. 267 102 411, vladimira.kvasnicova@lf3.cuni.cz informace, studijní materiály: http://vyuka.lf3.cuni.cz Sylabus
Integrace a regulace savčího energetického metabolismu
 Základy biochemie KBC / BCH Integrace a regulace savčího energetického metabolismu Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským
Základy biochemie KBC / BCH Integrace a regulace savčího energetického metabolismu Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským
Katabolismus - jak budeme postupovat
 Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Katabolismus - jak budeme postupovat I. fáze aminokyseliny proteiny polysacharidy glukosa lipidy Glycerol + mastné kyseliny II. fáze III. fáze ETS itrátový cyklus yklus trikarboxylových kyselin, Krebsův
Konsultační hodina. základy biochemie pro 1. ročník. Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa
 Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Konsultační hodina základy biochemie pro 1. ročník Přírodní látky Úvod do metabolismu Glykolysa Krebsův cyklus Dýchací řetězec Fotosynthesa Přírodní látky 1 Co to je? Cukry (Sacharidy) Organické látky,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Citrátový a glyoxylátový cyklus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Citrátový a glyoxylátový cyklus Buněčná respirace I. Fáze Energeticky bohaté látky jako glukosa, mastné kyseliny a některé aminokyseliny
Buněčné dýchání Ch_056_Přírodní látky_buněčné dýchání Autor: Ing. Mariana Mrázková
 Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
Registrační číslo projektu: CZ.1.07/1.1.38/02.0025 Název projektu: Modernizace výuky na ZŠ Slušovice, Fryšták, Kašava a Velehrad Tento projekt je spolufinancován z Evropského sociálního fondu a státního
CZ.1.07/2.2.00/ Obecný metabolismus. Energetický metabolismus (obecně) (1).
 (1).") mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 becný metabolismus Energetický metabolismus (obecně) (1). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Didaktické testy z biochemie 2
 Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
Didaktické testy z biochemie 2 Metabolismus Milada Roštejnská Helena Klímová br. 1. Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy Ukládány jako glykogen v játrech Ukládány Ukládány jako
AMPK AMP) Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze
 Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze") AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
8. Polysacharidy, glykoproteiny a proteoglykany
 Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Struktura a funkce biomakromolekul KBC/BPOL 8. Polysacharidy, glykoproteiny a proteoglykany Ivo Frébort Polysacharidy Funkce: uchovávání energie, struktura, rozpoznání a signalizace Homopolysacharidy a
Bp1252 Biochemie. #8 Metabolismus živin
 Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Bp1252 Biochemie #8 Metabolismus živin Chemické reakce probíhající v organismu Katabolické reakce přeměna složitějších látek na jednoduché, jsou většinou exergonické. Anabolické reakce syntéza složitějších
Sacharidy Klasifikace sacharidů
 Sacharidy Klasifikace sacharidů Sacharidy monosacharidy oligosacharidy polysacharidy aldosy ketosy disacharidy aldotetrosy ketotetrosy trisacharidy aldopentosy ketopentosy tertrasacharidy. aldohexosy ketohexosy.
Sacharidy Klasifikace sacharidů Sacharidy monosacharidy oligosacharidy polysacharidy aldosy ketosy disacharidy aldotetrosy ketotetrosy trisacharidy aldopentosy ketopentosy tertrasacharidy. aldohexosy ketohexosy.
CYKLUS TRIKARBOXYLOVÝCH KYSELIN A GLYOXYLÁTOVÝ CYKLUS
 YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
YKLUS TRIKARBXYLVÝ KYSELIN A GLYXYLÁTVÝ YKLUS BSA Základní charakteristika istorie Pyruvátdehydrogenasový komplex itátový cyklus dílčí reakce itátový cyklus výtěžek itátový cyklus regulace Anapleroticé
Otázka: Metabolismus. Předmět: Biologie. Přidal(a): Furrow. - přeměna látek a energie
: Furrow. - přeměna látek a energie") Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Otázka: Metabolismus Předmět: Biologie Přidal(a): Furrow - přeměna látek a energie Dělení podle typu reakcí: 1.) Katabolismus reakce, při nichž z látek složitějších vznikají látky jednodušší (uvolňuje
Sacharidy a polysacharidy (struktura a metabolismus)
") Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Sacharidy a polysacharidy (struktura a metabolismus) Sacharidy Živočišné tkáně kolem 2 %, rostlinné 85-90 % V buňkách rozličné fce: Zdroj a zásobárna energie (glukóza, škrob, glykogen) Výztuž a ochrana
Štěpení lipidů. - potravou přijaté lipidy štěpí lipázy gastrointestinálního traktu
 METABOLISMUS LIPIDŮ ODBOURÁVÁNÍ LIPIDŮ - z potravy nebo z tukových rezerv - hydrolytické štěpení esterových vazeb - vznik glycerolu a mastných kyselin - hydrolytické štěpení LIPÁZY (karboxylesterázy) -
METABOLISMUS LIPIDŮ ODBOURÁVÁNÍ LIPIDŮ - z potravy nebo z tukových rezerv - hydrolytické štěpení esterových vazeb - vznik glycerolu a mastných kyselin - hydrolytické štěpení LIPÁZY (karboxylesterázy) -
Biochemie jater. Eva Samcová
 Biochemie jater Eva Samcová Orgánová specializace Hlavní metabolické dráhy pro glukosu, mastné kyseliny a aminokyseliny jsou soustředěné okolo pyruvátu a acetyl-coa. Glukosa je primárním palivem pro mozek
Biochemie jater Eva Samcová Orgánová specializace Hlavní metabolické dráhy pro glukosu, mastné kyseliny a aminokyseliny jsou soustředěné okolo pyruvátu a acetyl-coa. Glukosa je primárním palivem pro mozek
Charakteristika složky 3) cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa
 cytochrom-c NADH-Q-reduktasa cytochrom-c- oxidasa ubichinon cytochromreduktasa") 8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
8. Dýchací řetězec a fotosyntéza Obtížnost A Pomocí následující tabulky charakterizujte jednotlivé složky mitochondriálního dýchacího řetězce. SLOŽKA Pořadí v dýchacím řetězci 1) Molekulový typ 2) Charakteristika
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/ Anotace. Metabolismus lipidů - odbourávání. VY_32_INOVACE_Ch0212
 Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu P VK Název školy: Gymnázium, Zábřeh, náměstí svobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
REGULACE ENZYMOVÉ AKTIVITY
 REGULACE ENZYMOVÉ AKTIVITY Proč je nutno regulovat enzymovou aktivitu? (homeostasa) Řada úrovní: regulace množství přítomného enzymu (exprese = proteosynthesa, odbourávání) synthesa vhodného enzymu (isoenzymy)
REGULACE ENZYMOVÉ AKTIVITY Proč je nutno regulovat enzymovou aktivitu? (homeostasa) Řada úrovní: regulace množství přítomného enzymu (exprese = proteosynthesa, odbourávání) synthesa vhodného enzymu (isoenzymy)
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Biosyntéza a metabolismus bílkovin
 Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
9. Dýchací řetězec a oxidativní fosforylace. mitochondriální syntéza ATP a fotosyntéza
 9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
9. Dýchací řetězec a oxidativní fosforylace mitochondriální syntéza ATP a fotosyntéza CHEMIOSMOTICKÁ TEORIE SYNTÉZY ATP Heterotrofní organismy získávají hlavní podíl energie (cca 90%) uložené ve struktuře
Metabolismus bílkovin. Václav Pelouch
 ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
ZÁKLADY OBECNÉ A KLINICKÉ BIOCHEMIE 2004 Metabolismus bílkovin Václav Pelouch kapitola ve skriptech - 3.2 Výživa Vyvážená strava člověka musí obsahovat: cukry (50 55 %) tuky (30 %) bílkoviny (15 20 %)
16a. Makroergické sloučeniny
 16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
16a. Makroergické sloučeniny Makroergickými sloučeninami v biochemii nazýváme skupinu látek umožňujících uvolnění značného množství energie v jednoduché reakci. Nelze je definovat prostě jako sloučeniny
Sacharidy Polyhydroxysloučeniny obsahující karbonylovou skupinu
 Sacharidy Polyhydroxysloučeniny obsahující karbonylovou skupinu o Tvoří strukturní molekuly (celulosa, chitin, pektiny, hemicelulosy atd.) o Jsou součástí nukleových kyselin o Pohotová energetická zásoba
Sacharidy Polyhydroxysloučeniny obsahující karbonylovou skupinu o Tvoří strukturní molekuly (celulosa, chitin, pektiny, hemicelulosy atd.) o Jsou součástí nukleových kyselin o Pohotová energetická zásoba
Metabolismus mikroorganismů
 Metabolismus mikroorganismů Metabolismus organismů Souvisí s metabolismem polysacharidů, bílkovin, nukleových kyselin a lipidů Cytoplazma, mitochondrie (matrix, membrána) H 3 PO 4 Polysacharidy Pentózový
Metabolismus mikroorganismů Metabolismus organismů Souvisí s metabolismem polysacharidů, bílkovin, nukleových kyselin a lipidů Cytoplazma, mitochondrie (matrix, membrána) H 3 PO 4 Polysacharidy Pentózový
Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996
 Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996 Šablona: III/2 č. materiálu: VY_32_INOVACE_CHE_414 Jméno autora: Třída/ročník: Mgr. Alena
Výukový materiál zpracován v rámci projektu EU peníze školám Registrační číslo projektu: CZ.1.07/1.5.00/34.0996 Šablona: III/2 č. materiálu: VY_32_INOVACE_CHE_414 Jméno autora: Třída/ročník: Mgr. Alena
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI
 METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
METABOLISMUS SLOUČENINY S MAKROERGNÍMI VAZBAMI Obsah Formy organismů Energetika reakcí Metabolické reakce Makroergické sloučeniny Formy organismů Autotrofní x heterotrofní organismy Práce a energie Energie
Citrátový cyklus. Tomáš Kučera.
 itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
itrátový cyklus Tomáš Kučera tomas.kucera@lfmotol.cuni.cz Ústav lékařské chemie a klinické biochemie 2. lékařská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole 2017 Schéma energetického
Ukázky z pracovních listů z biochemie pro SŠ A ÚVOD
 Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Ukázky z pracovních listů z biochemie pro SŠ A ÚVD 1) Doplň chybějící údaje. Jak se značí makroergní vazba? Kolik je v ATP makroergních vazeb? Co je to ADP Kolik je v ADP makroergních vazeb 1) Pojmenuj
Metabolismus lipidů. (pozn. o nerozpustnosti)
") Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
Metabolismus lipidů (pozn. o nerozpustnosti) Trávení lipidů Lipidy v potravě - většinou v hydrolyzovatelné podobě, především jako triacylglayceroly (TAG), fosfatidáty a sfingolipidy. V trávicím traktu
fce jater: (chem. továrna, jako 1. dostává všechny látky vstřebané GIT) METABOLICKÁ (jsou metabolicky nejaktivnější tkání v těle)
 METABOLICKÁ (jsou metabolicky nejaktivnější tkání v těle)") JÁTRA ústřední orgán intermed. metabolismu, vysoká schopnost regenerace krevní oběh játry: (protéká 20% veškeré krve, 10-30% okysl.tep.krve, která zajišťuje výživu buněk, zbytek-portální krev) 1. funkční
JÁTRA ústřední orgán intermed. metabolismu, vysoká schopnost regenerace krevní oběh játry: (protéká 20% veškeré krve, 10-30% okysl.tep.krve, která zajišťuje výživu buněk, zbytek-portální krev) 1. funkční
Sacharidy. Sacharidy. z jednoduchých monosacharidů kondenzací vznikají polysacharidy
 Sacharidy 1. Monosacharidy 2. Disacharidy 3. Polysacharidy Sacharidy nesprávně nazývány uhlovodany n ( 2 ) n - platí to pouze pro některé cukry přítomné ve všech rostlinných a živočišných buňkách vznik
Sacharidy 1. Monosacharidy 2. Disacharidy 3. Polysacharidy Sacharidy nesprávně nazývány uhlovodany n ( 2 ) n - platí to pouze pro některé cukry přítomné ve všech rostlinných a živočišných buňkách vznik
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Transport elektronů a oxidativní fosforylace
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Transport elektronů a oxidativní fosforylace Oxidativní fosforylace vs. fotofosforylace vyvrcholení katabolismu Všechny oxidační degradace
H 2 O, H + H 2 O, H + oligosacharidy. Příklad: hydrolýza škrobu (polysacharid) přes maltosu (disacharid) na glukosu (monosacharid).
 přes maltosu (disacharid) na glukosu (monosacharid).") Sacharidy Definice a klasifikace sacharidů Výraz karbohydráty (uhlovodany, atd.) vznikl na základě molekulového složení těchto sloučenin, neboť to může být vyjádřeno vzorcem C n (H 2 O) n, tedy jako hydráty
Sacharidy Definice a klasifikace sacharidů Výraz karbohydráty (uhlovodany, atd.) vznikl na základě molekulového složení těchto sloučenin, neboť to může být vyjádřeno vzorcem C n (H 2 O) n, tedy jako hydráty
Trávení a metabolismus
 Trávení a metabolismus Milada Roštejnská elena Klímová br. 1. Proces vylučování [1] 1 bsah (1. část) Zařazení člověka podle metabolismu Potrava sud potravy v lidském těle Trávení (obecně) Trávení sacharidů
Trávení a metabolismus Milada Roštejnská elena Klímová br. 1. Proces vylučování [1] 1 bsah (1. část) Zařazení člověka podle metabolismu Potrava sud potravy v lidském těle Trávení (obecně) Trávení sacharidů
Ukládání energie v buňkách
 Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
Ukládání energie v buňkách Josef Fontana EB - 58 Obsah přednášky Úvod do problematiky zásobních látek lidského organismu Přehled zásobních látek v těle Metabolismus glykogenu Struktura glykogenu Syntéza
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ Sacharidy v potravě: polysacharidy: škrob (brambory, pečivo, pudinky...) celulosa - ovoce, zelenina disacharidy: sacharosa (sladké jídlo) laktosa (mléko a výrobky z něj) maltosa
METABOLISMUS SACHARIDŮ Sacharidy v potravě: polysacharidy: škrob (brambory, pečivo, pudinky...) celulosa - ovoce, zelenina disacharidy: sacharosa (sladké jídlo) laktosa (mléko a výrobky z něj) maltosa
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
glukóza *Ivana FELLNEROVÁ, PřF UP Olomouc*
 Prezentace navazuje na základní znalosti Biochemie, stavby a transportu přes y Doplňující prezentace: Proteiny, Sacharidy, Stavba, Membránový transport, Symboly označující animaci resp. video (dynamická
Prezentace navazuje na základní znalosti Biochemie, stavby a transportu přes y Doplňující prezentace: Proteiny, Sacharidy, Stavba, Membránový transport, Symboly označující animaci resp. video (dynamická
Odbourávání lipidů, ketolátky
 dbourávání lipidů, ketolátky Josef Fontana EB - 56 bsah přednášky Energetický význam TAG Jednotlivé dráhy metabolismu lipidů lipidy jako zdroj energie degradace TAG v buňkách, β-oxidace MK tvorba a využití
dbourávání lipidů, ketolátky Josef Fontana EB - 56 bsah přednášky Energetický význam TAG Jednotlivé dráhy metabolismu lipidů lipidy jako zdroj energie degradace TAG v buňkách, β-oxidace MK tvorba a využití
33.Krebsův cyklus. AZ Smart Marie Poštová
 33.Krebsův cyklus AZ Smart Marie Poštová m.postova@gmail.com Metabolismus Metabolismus je souhrn chemických reakcí v organismu. Základní metabolické děje jsou: a) katabolické odbourávací (složité látky
33.Krebsův cyklus AZ Smart Marie Poštová m.postova@gmail.com Metabolismus Metabolismus je souhrn chemických reakcí v organismu. Základní metabolické děje jsou: a) katabolické odbourávací (složité látky
Sacharidy - polyhydroxyaldehydy nebo polyhydroxyketony (synonymen názvu je termín glycidy)
") /2 Sacharidy 1 Pojmy Sacharidy - polyhydroxyaldehydy nebo polyhydroxyketony (synonymen názvu je termín glycidy) Mnoho z nich, ne všechny, mají empirický vzorec ( 2 ) n ; některé také mohou obsahovat dusík,
/2 Sacharidy 1 Pojmy Sacharidy - polyhydroxyaldehydy nebo polyhydroxyketony (synonymen názvu je termín glycidy) Mnoho z nich, ne všechny, mají empirický vzorec ( 2 ) n ; některé také mohou obsahovat dusík,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Fotosyntéza
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Fotosyntéza Fotosyntéza pohlcení energie slunečního záření a její přeměna na chemickou energii rovnováha fotosyntetisujících a heterotrofních
Sacharidy: Přírodní organické látky v rostlinách i živočiších Ve struktuře: C, H, O (N, F, S)
") SACHARIDY (cukry) 1 Sacharidy: Přírodní organické látky v rostlinách i živočiších Ve struktuře: C, H, O (N, F, S) Dle počtu základních monosacharidových jednotek vázaných v jejich molekulách cukry 2 Biologický
SACHARIDY (cukry) 1 Sacharidy: Přírodní organické látky v rostlinách i živočiších Ve struktuře: C, H, O (N, F, S) Dle počtu základních monosacharidových jednotek vázaných v jejich molekulách cukry 2 Biologický
Biochemie I 2016/2017. Makromolekuly buňky. František Škanta
 Biochemie I 2016/2017 Makromolekuly buňky František Škanta Makromolekuly buňky ukry Tuky Bílkoviny ukry Jsou sladké Přehled strukturních forem sacharidů Monosacharidy Disacharidy Polysacharidy Ketotriosa
Biochemie I 2016/2017 Makromolekuly buňky František Škanta Makromolekuly buňky ukry Tuky Bílkoviny ukry Jsou sladké Přehled strukturních forem sacharidů Monosacharidy Disacharidy Polysacharidy Ketotriosa
Didaktické testy z biochemie 1
 Didaktické testy z biochemie 1 Trávení Milada Roštejnská elena Klímová Trávení br. 1. Trávicí soustava Rubrika A Z pěti možných odpovědí (alternativ) vyberte tu nejsprávnější. A B D E 1 Mezi monosacharidy
Didaktické testy z biochemie 1 Trávení Milada Roštejnská elena Klímová Trávení br. 1. Trávicí soustava Rubrika A Z pěti možných odpovědí (alternativ) vyberte tu nejsprávnější. A B D E 1 Mezi monosacharidy
Stanovení vybraných enzymů. Roman Kanďár
 Stanovení vybraných enzymů Roman Kanďár Takže prvně malé opakování ENZYM Protein (RNA) s katalytickou aktivitou Protein (RNA) kofaktor (prosthetická skupina, koenzym) Jaký je vlastně rozdíl mezi prosthetickou
Stanovení vybraných enzymů Roman Kanďár Takže prvně malé opakování ENZYM Protein (RNA) s katalytickou aktivitou Protein (RNA) kofaktor (prosthetická skupina, koenzym) Jaký je vlastně rozdíl mezi prosthetickou
Publikováno z 2. lékařská fakulta Univerzity Karlovy v Praze (http://www.lf2.cuni.cz)
") Publikováno z 2. lékařská fakulta Univerzity Karlovy v Praze (http://www.lf2.cuni.cz) Biochemie Napsal uživatel Marie Havlová dne 8. Únor 2012-0:00. Sylabus předmětu Biochemie, Všeobecné lékařství, 2.
Publikováno z 2. lékařská fakulta Univerzity Karlovy v Praze (http://www.lf2.cuni.cz) Biochemie Napsal uživatel Marie Havlová dne 8. Únor 2012-0:00. Sylabus předmětu Biochemie, Všeobecné lékařství, 2.
- metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy a jejich životním prostředím
 Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
Otázka: Obecné rysy metabolismu Předmět: Chemie Přidal(a): Bára V. ZÁKLADY LÁTKOVÉHO A ENERGETICKÉHO METABOLISMU - metabolismus soubor chemických reakcí probíhajících v živých organismech a mezi organismy
CHIRALITA William Thomson ( ) (Lord Kelvin, 1892)
 (Lord Kelvin, 1892)") CIRALITA William Thomson (1824-1907) (Lord Kelvin, 1892) I call any geometrical figure, or any group of points, chiral, and say it has chirality, if its image in a plane mirror, ideally realized, cannot
CIRALITA William Thomson (1824-1907) (Lord Kelvin, 1892) I call any geometrical figure, or any group of points, chiral, and say it has chirality, if its image in a plane mirror, ideally realized, cannot
METABOLISMUS SACHARIDŮ. Biochemický ústav LF MU (H.P., ET)
") METABOLISMUS SACHARIDŮ Biochemický ústav LF MU 2016 - (H.P., ET) 1 Sacharidy Hlavní ţivina pro lidský organismus Doporučený denní příjem sacharidů pro dospělé je přibližně 4 6 g/kg tělesné hmotnosti, sacharidy
METABOLISMUS SACHARIDŮ Biochemický ústav LF MU 2016 - (H.P., ET) 1 Sacharidy Hlavní ţivina pro lidský organismus Doporučený denní příjem sacharidů pro dospělé je přibližně 4 6 g/kg tělesné hmotnosti, sacharidy
Metabolismus pentóz, glykogenu, fruktózy a galaktózy. Alice Skoumalová
 Metabolismus pentóz, glykogenu, fruktózy a galaktózy Alice Skoumalová 1. Pentóza fosfátová dráha Přehledné schéma: Pentóza fosfátová dráha (PPP): Probíhá v cytozolu Všechny buňky Dvě části: 1) Oxidační
Metabolismus pentóz, glykogenu, fruktózy a galaktózy Alice Skoumalová 1. Pentóza fosfátová dráha Přehledné schéma: Pentóza fosfátová dráha (PPP): Probíhá v cytozolu Všechny buňky Dvě části: 1) Oxidační
Kofaktory enzymů. T. Kučera. (upraveno z J. Novotné)
") Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Kofaktory enzymů T. Kučera (upraveno z J. Novotné) Kofaktory enzymů neproteinová, nízkomolekulární složka enzymu ko-katalyzátor potřebný k aktivitě enzymu pomocné molekuly v enzymové reakci holoenzym (aktivní)
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová
 Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
Intermediární metabolismus - SOUHRN - Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba
METABOLISMUS SACHARIDŮ
 METABOLISMUS SACHARIDŮ GLUKOSA V KRVI Jedna z hlavních priorit metabolické regulace: Hladina glukosy v krvi nesmí poklesnout pod 3 mmol/l Hormonální regulace: insulin glukagon adrenalin kortisol ( snižuje
METABOLISMUS SACHARIDŮ GLUKOSA V KRVI Jedna z hlavních priorit metabolické regulace: Hladina glukosy v krvi nesmí poklesnout pod 3 mmol/l Hormonální regulace: insulin glukagon adrenalin kortisol ( snižuje
DYNAMICKÁ BIOCHEMIE. Daniel Nechvátal :: www.gymzn.cz/nechvatal
 DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
DYNAMICKÁ BIOCHEMIE Daniel Nechvátal :: www.gymzn.cz/nechvatal Energetický metabolismus děje potřebné pro zabezpečení života organismu ANABOLISMUS skladné reakce, spotřeba E KATABOLISMUS rozkladné reakce,
SACHARIDY. Vznik sacharidů v přírodě v buňkách autotrofů asimilací CO 2 v přítomnosti H 2 O
 SACHARIDY v těle člověka jen 2 % (v sušině) v rostlinách 85 90 % Funkce sacharidů v buňce: - zdroj energie (např. glukosa) - zásobní energetická surovina (škrob, glykogen) - zpevnění a ochrana buňky (celulosa,
SACHARIDY v těle člověka jen 2 % (v sušině) v rostlinách 85 90 % Funkce sacharidů v buňce: - zdroj energie (např. glukosa) - zásobní energetická surovina (škrob, glykogen) - zpevnění a ochrana buňky (celulosa,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Metabolusmus lipidů - katabolismus
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - katabolismus Trávení, aktivace a transport tuků Oxidace mastných kyselin Ketonové látky Úvod Oxidace MK je centrální
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Metabolusmus lipidů - katabolismus Trávení, aktivace a transport tuků Oxidace mastných kyselin Ketonové látky Úvod Oxidace MK je centrální