Membránové transportery - pokračování:
|
|
|
- Miluše Vávrová
- před 6 lety
- Počet zobrazení:
Transkript
1 Membránové transportery - Pumpy pohon ATP, PP pokračování: pro rostliny typické H+ pumpy plasmalemmy, tonoplastu Přenašeče Kanály

2 Plasmalemma: P H+-ATPáza Arabidopsis: genová rodina AHA (11 genů!)

3 Gradienty ph v rostoucích buňkách: příklad pyl. láček a koř. vlásků
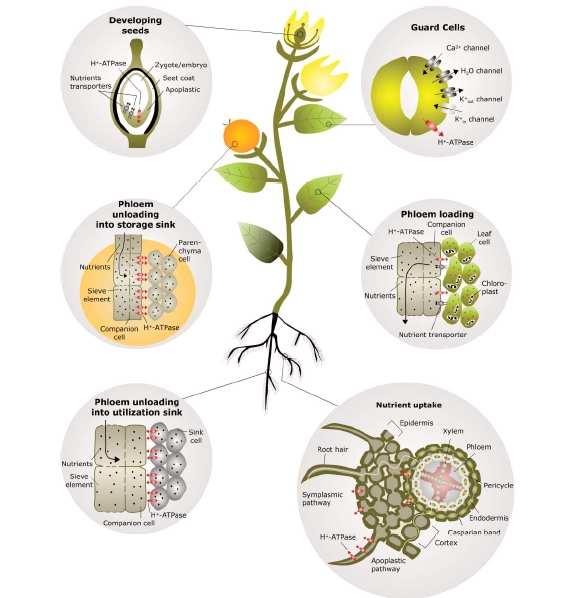
4 P-H+ATPázy a "loading"
5
6 Ca- ATPasa (další pumpa P- typu)
7 Dva typy rostlinných Ca2+ pump 1. IIA (ECA) - v ER, není aktivována CaM. 2. IIB (ACA) - v PM, ale i ER a tonoplast.mají autoinhibiční regulační doménu (fosforylace, vazba CaM). Obě jsou P-typu, tedy inhibovány ortovanadátem.
8

9 IIA - ECA IIB -ACA : CaM reg.

10 Fosforylace Ser45 CDPK blokuje aktivaci CAM! - tj. možnost jemného nastavení funkce.
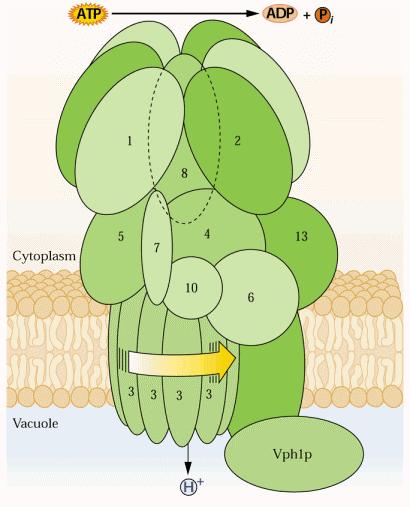
11 V-H+ATPasa
12
13 Připomeňme si: obrátíme-li elektromotor, dostaneme dynamo. F o F 1 ATP synthasa V - ATPasa
14 Specifický inhibitor V H+ATPázy

15 det/cop mutanti Dark Light Dark Light Wild Type det Mutant
16 det3 je mutant C podjednotky V- H+-ATPázy
17 V-H+ATPasa mutace letální pro pyl 13 podjednotek, 27 genů... kolik kombinací??

18 NaCl Rozdíly isoforem: např. regulace podmínkami E-subunit induk. stresem

19 V H+ATPáza se podílí na detoxikaci Cd

20 sůl teplo chlad... a samozřejmě i rozrůznění anatomické, vývojové...


21 Další možné role? kvasinky - homotypic vacuole fusion Drosophila - synaptic vesicle fusion savci - A4 jako ph sensor Arabidopsis - VHA-B1 v Glu signalling!! (Cho, Yoo, Sheen 2006)

22 glucose mannitol low light low nutrient high light

23 V-H+-PPasa (membránová pyrofosfatáza) rostlinná specialita v tonoplastu užívá jako zdroj energie pyrofosfát PPi ("odpad" při polymeraci DNA,RNA a syntetických proc. např. ADPG či UDPG) příbuzné pumpy ale i v plasmalemě bakterií a hmyzu
24 Evoluce rostlinné vakuoly je těsně spojena s evolucí V H+-PPasy, která je nutnou součástí funkčního tonoplastu (minoritně je ovšem lokalizována i do PM) za normálních podmínek i ve stresu.

25 AVP1 - vak. PPasa část proteinu v plasmalemě role v transportu auxinu? overexprese zvyšuje rezistenci k sol. stresu...
26
27 ABC transportery (ATP Binding Cassette) fungují nejen jako na ATP závislé pumpy, ale také jako iontové kanály, či jejich regulátory.

28 ATP vazebná místa.
29 Fungují jako pumpy či "flipázy"? jejich specifita kolísá a jsou klíčové pro řadu detoxifikačních transportů zvl. do vakuoly.
 ABC transporterem AtMRP2 Zatímco AtMRP1 transportuje jen konjugáty")
30 Transport dinitrofenolu konjugovaného glutathionem (A) a tetrapyrolového katabolitu chlorofylu (B) ABC transporterem AtMRP2 Zatímco AtMRP1 transportuje jen konjugáty glutathionu

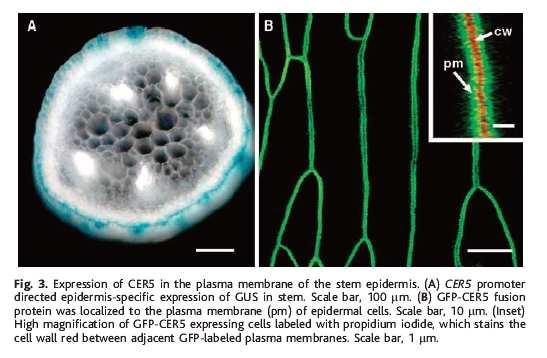
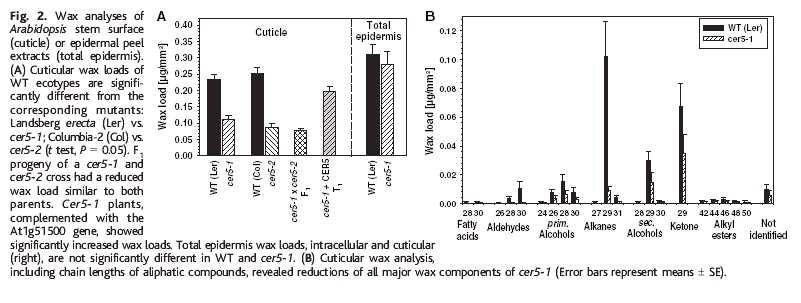
31 ABC a kutikula: CER5 kóduje ABC transp. wt cer5
32
33

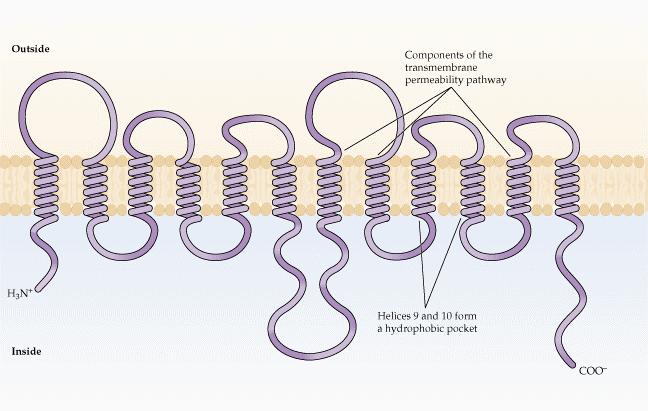
34 ABC a transport IAA


35 Modely transportu auxinu Klasicky: přenašeče PIN a AUX/LAX
36 Co dělají PGP (ABC) transportery?
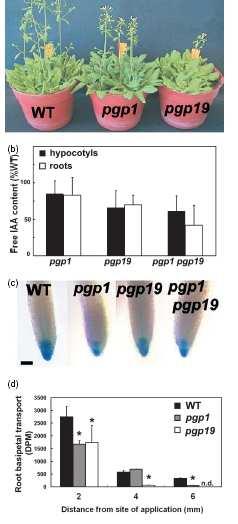

37 ABC a IAA "fenotypy" jen malé

38 Lokalizace klasických PIN IAA výtokových přenašečů není narušena
. Jeho exprese je aktivována IAA")
39 PGP1 je nepolárně lokalizován v mladých pletivech, ale polárně v diferencovaných b.kořene (f). Jeho exprese je aktivována IAA
40 Membránové transportery konkrétně : Pumpy Přenašeče saturovatelné, poháněny gradienty (pmf) Kanály

41 Saturační kinetika přenašeče

42 Model obecného přenašeče
43 Typy přenašečů uniport symport antiport
44

45 Jak studovat transportní procesy?

46 Heterologní exprese: oocyty, kvasinky
47
48 Symport H+/cukry při plnění floemu sucrose-h+ symporters

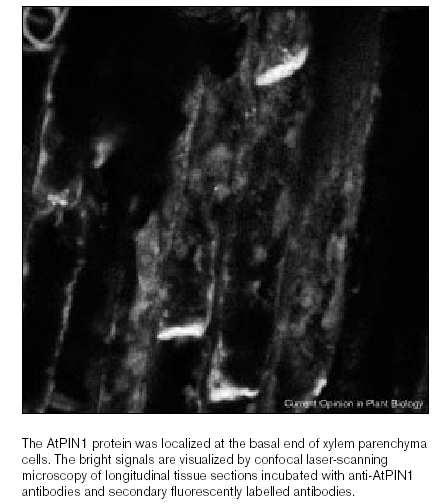
49 Arabidopsis SUC2 v průvodních buňkách

50 Bramborový SUT1 v sítkovicích


51 Vývojová regulace SUT1/2 mrkve sink source SUT1 SUT2

52 Proč různé přenašeče téhož iontu? Jemné ladění!

53 Jsou i výjimky z pohonu H+ Na + NO 3- coupling
54 Přenašeče auxinu (IAA) ABC transportery (MDR, PGP) PIN AUX/LAX
: AUX1/LAX1 PINs")
55 Kudy auxin teče? Přenašeče (efflux carriers): AUX1/LAX1 PINs inhibice fytotropiny (NPA)


56 IAA influx: AUX1 Swarup et al. 2001
 auxinu působením NPA!")
57 PIN přenašeče Fenotyp: jako inhibice transportu (efflux) auxinu působením NPA! pin1 mutant

58 PIN proteiny


59 Lokalizace členů PIN rodiny (I. Billou, K. Palme, J. Friml et al.)
60 Co pohání transport auxinu???? (Samozřejmě) žádá funkční membránu Interakce s řadou proteinů možná role ABC transportérů (PGP) K+ kanálů (TRH1) a dalších...
")
61 Přenašeče na tonoplastu (nejen CAM rostlinám se hodí odkládat malát do vakuoly a jde to nejen přes kanály)
62
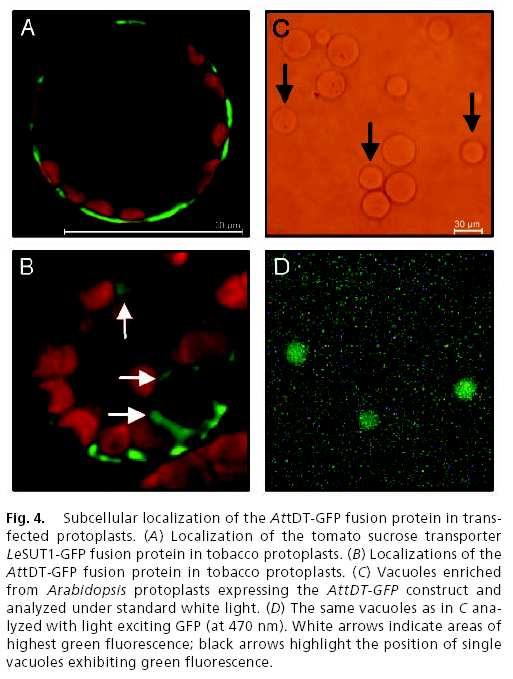
63
64
65 Membránové transportery konkrétně : Pumpy Přenašeče Kanály řádově vyšší propustnost! tedy je jich málo, biochemie nemusí být dost citlivá... elektrofyziologie!

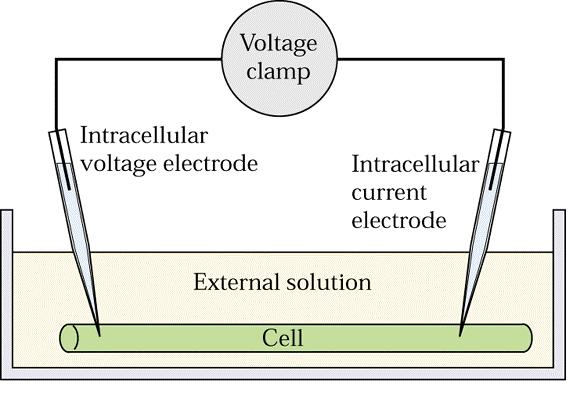
66 Měření membránových potenciálů

67

68 Micrograph of a patch pipette attached to the surface of a barley aleurone cell protoplast. (Photo courtesy of J. Schroeder and D. Bush.)
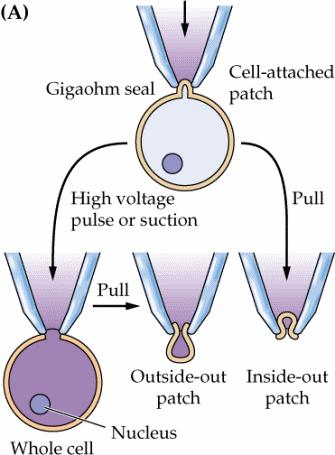
69 Patch-clamp
70
71 Patch-clamp techniky mohou Závislost membránového potenciálu na čase I-V křivky měřit i 1 kanál

72 I-V křivky (měřený) kationty ven anionty dovnitř anionty ven kationty dovnitř (vložený na membránu)
")
73 Měření selektivity kanálů - závislost I/V na složení média (outside-out patch, K+ kanály)
 100 mm K+ in 10 mm K+")
74 outside-out patch 100 mm KCl (in and out) 100 mm K+ in 10 mm K+ out
75 Co můžeme měřit - shrnutí Recordings of K+ currents in whole cells and in single K+-selective channels of guard cell protoplasts. (1) K+ currents recorded in the whole-cell configuration (see Web Figure 6.2.B) when the membrane potential is clamped at different values. Upward deflections show outward currents; downward deflections show inward currents. K+ concentrations were 105 mm in the pipette and cytoplasm, and 11 mm in the bathing solution outside the cell. (2) Data from the same experiment plotted as an I/V (current/voltage) curve, before and after the addition of Ba2+, an ion that blocks K+ channels. (3) Inward K+ current through a single channel in a membrane patch. (From Schroeder et al )
76 Podobně lze sledovat jakýkoli elektrogenní transport na membráně tedy třeba i činnost H+ pumpy! (Gehwolf et al. FEBS Lett 512:152, 2002)
77 Iontové kanály plasmalemmy: regulace napětím I křivky whole cell I-V

78 Závislost otvírání obec. kanálu na napětí

79 inward rectifying K+ kanál inhibitor - tetraethylammonium



80 K+ kanály typu shaker (inward rectifying) + regulační podjednotky! + fosforylace!! (ABA sig.?)


81 K+ kanály typu shaker (inward rectifying) KAT1 - isoformy! AKT1

82 akt1-1 mutant Rb+ uptake wt akt1-1 10, 100, 1000 µm Rb+ růst
 regulace Ca2 +!")

83 KCO1 (outward rectifying K+) regulace Ca2 +! inhlibice Ba+, vápník snižuje prahové napětí pro aktivaci
84 GORK (guard cell outward rectifying K+ channel) (exprese v oocytech)
 Na+ kanál příjem")
85 Na napětí nezávislý (a méně selektivní) Na+ kanál příjem sodíku??

86 Ca2+ kanály plasmalemy (wheat root plasma membrane, aktivace depolarizací)
 různě selektivní (Ca; Ca, K; Ca, Ba, Sr, Mg; maxication Rb až Mn) aktivované hyperpolarizací mechanosenzitivní nebo induk.")
87 Ca2+ kanály plasmalemy aktivované depolarizací (např. DACC, rca) různě selektivní (Ca; Ca, K; Ca, Ba, Sr, Mg; maxication Rb až Mn) aktivované hyperpolarizací mechanosenzitivní nebo induk. elicitory (ROS!) selektivnější (Ca; Ca, K; Ca, Ba) nezávislé na napětí... touch reaction in Arabidopsis
")
88 ... ale buňka má vápníkových kanálů mnohem víc, a k tomu ještě pumpy... (i na vakuole)

89 Iontové kanály tonoplastu a dalších endomembrán (pohon H+ gradientem)

90 Tonoplastový kanál pro jednomocné kationty (K+): regulace ph a Ca 2+ FV fast vacuolar channel: aktivace ph, inhibice Ca2+ VK vacuolar K+: inhibice ph, aktivace Ca2+

91 Vápníkové kanály tonoplastu a dalších endomembrán tonoplastový, aktivace IP3 tonoplastový, aktivace cadpr ( ryanodin. receptor ) voltage-activated, Ca2+ insensitive voltage-activated Ca2+ sensitive (SV)

92 CICR calcium induced calcium release Ca2+ stimuluje SV calcium channel
93 Aniontové kanály (Cl-) plasmalemmy (svěracích buněk) rychlý (R) pomalý (S) aktivace Ca2+!
 malát 10 mm 20 mm 50 mm 100 mm")
94 Tonoplastové kanály pro malát (VMAL) Rostlinná specialita! CAM rostliny! (Kalanchoe) malát 10 mm 20 mm 50 mm 100 mm
95 Kanály v regulaci vývojových dějů Příklad: vrcholový růst Zygota Fucus - relokalizace Ca2+ kanálů jako 1. krok



96 Gradienty Ca2+ a ph při vrcholovém růstu 100 nm Ca 450 nm Ca Fucus rhizoid... dtto kořenové vlásky... pylová láčka (Feijo et al.)
97 Obecný rys vrcholového růstu?... dále sekreční dráha a cytoskelet... (také TRH1 K+ transporter)

98 Transport vody přes membrány: aquaporiny

99 ... ale musí to jít rychle!

100 změny osmolarity Měření transportu vody



101 Jak měřit turgor?
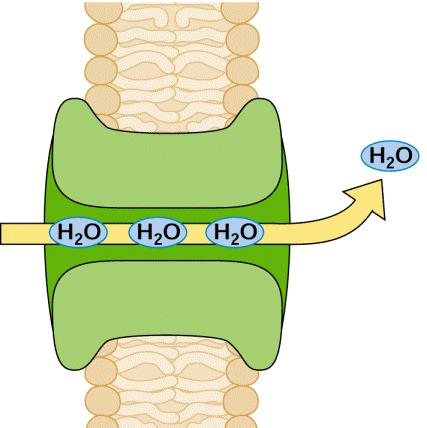
102

103 Tonoplast Intrinsic Proteins různé TIPs v různých typech vakuol! Arabidopsis: 35 genů, 10 prot. ve vakuole, kolik kombinací?
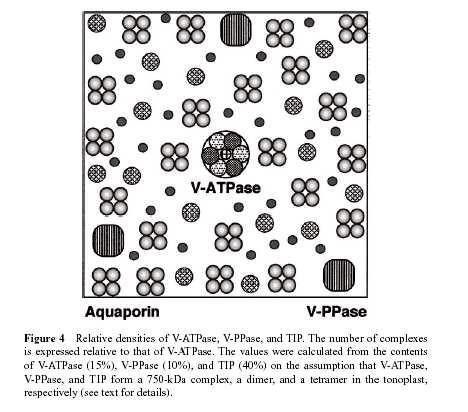
104
105 K čemu jsou akvaporiny? SPH simple permeability hypothesis regulace buněčného objemu (jsou nutné?) homeostáze objemu cytoplasmy (model??) zprostředkování pohybu vody... radiálně kořenem (ALE antisense PIP mutanti?) Vnímání a regulace osmotického (turgorového) tlaku TIPs přenášejí i NH3! (Hill et al. 2004, Martinoia et al. 2007)
106 Osmotické senzory rostlinné buňky

107 Model regulace akvaporinu turgorem

108 Stomatologie

109 Průduchy: integrace transportních systémů
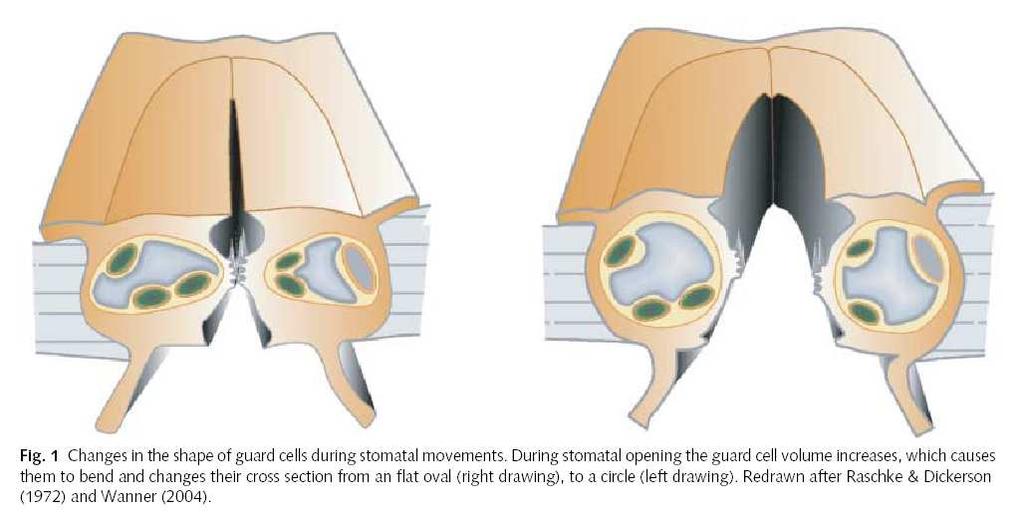
110
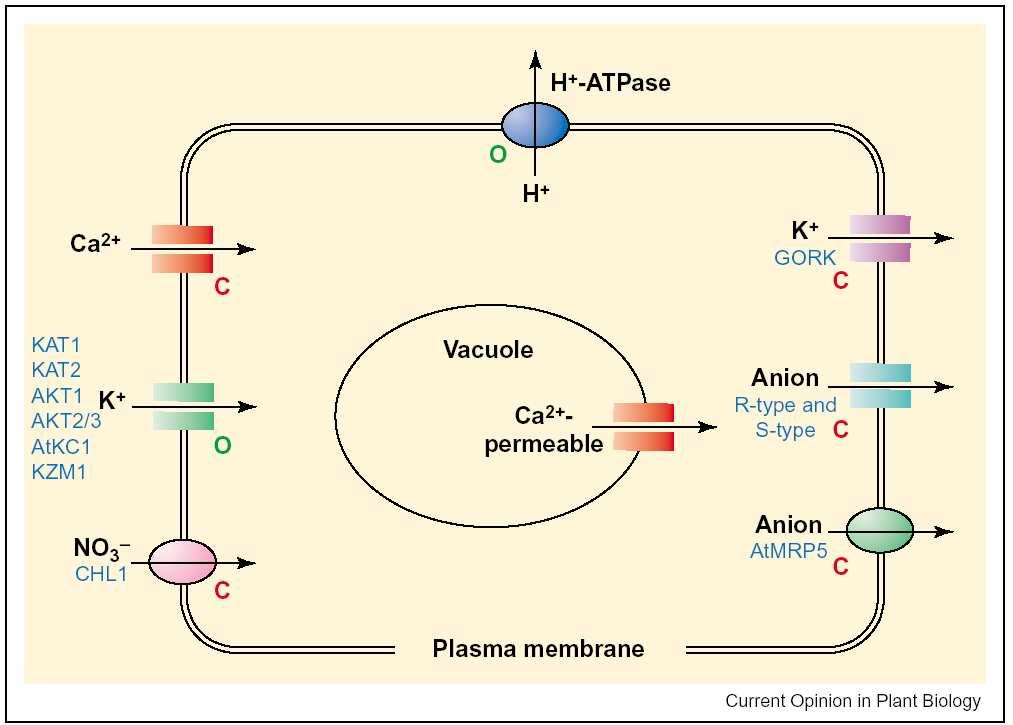
111


112 Kanály v plasmalemě svěracích buněk
113 Otvírání 1. H+ ATPasy pmf 2. hyperpolarizace K+ uptake přes inw. rect. 3. Cl- uptake symportem s H+ 4. produkce malátu 5. pmf na tonoplastu (H+ ATPase) 6. Cl- do vakuoly po spádu 7. malát do vakuoly přenašečem? 8. H+/K+ antiport 9. voda jde za solemi do vakuoly
114 Zavírání výtok K+ z vakuoly po spádu a depolarizace 2. výtok Cl- po spádu 3. otevření aniont. kanálů depolarizuje plasmalemu 4. aktivace outward rectifying K+ kanálu 5. voda následuje

115 Regulace ABA, světlo, CO 2 (... via ph...) zprostředkování Ca 2+
Membrány obsahují 1. sensory které umožňují reakci buňky na podněty z okolí 2. pumpy, přenašeče a kanály protransport látek skrz.
 Membrány obsahují 1. sensory které umožňují reakci buňky na podněty z okolí 2. pumpy, přenašeče a kanály protransport látek skrz. Mechanické vlastnosti membrán jsou obdivuhodné (např. Schopnost růstu a
Membrány obsahují 1. sensory které umožňují reakci buňky na podněty z okolí 2. pumpy, přenašeče a kanály protransport látek skrz. Mechanické vlastnosti membrán jsou obdivuhodné (např. Schopnost růstu a
Osnova. Základní vlastnosti biologických membrán Transport látek přes membránu
 Osnova Základní vlastnosti biologických membrán Transport látek přes membránu úvod pumpy přenašeče kanály transport vody Membránový transport v praxi regulace otevírání a zavírání svěracích buněk Membrány
Osnova Základní vlastnosti biologických membrán Transport látek přes membránu úvod pumpy přenašeče kanály transport vody Membránový transport v praxi regulace otevírání a zavírání svěracích buněk Membrány
Osnova. Základní vlastnosti biologických membrán Transport látek přes membránu
 Osnova Základní vlastnosti biologických membrán Transport látek přes membránu úvod pumpy přenašeče kanály transport vody Membránový transport v praxi regulace otevírání a zavírání svěracích buněk Membrány
Osnova Základní vlastnosti biologických membrán Transport látek přes membránu úvod pumpy přenašeče kanály transport vody Membránový transport v praxi regulace otevírání a zavírání svěracích buněk Membrány
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů
 Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů") MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA
 TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
Vodní režim rostlin. Transport kapalné vody
 Vodní režim rostlin Transport kapalné vody Transport vody přes membránu Příjem vody kořenem Radiální transport vody v kořenech Kořenový vztlak Příjem vody nadzemníčástí Základní charakteristiky transportu
Vodní režim rostlin Transport kapalné vody Transport vody přes membránu Příjem vody kořenem Radiální transport vody v kořenech Kořenový vztlak Příjem vody nadzemníčástí Základní charakteristiky transportu
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Průduchy regulace příjmu CO 2
 Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Úvod do biologie rostlin Transport látek TRANSPORT. Krátké, střední, dlouhé vzdálenosti
 Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Slide 1a TRANSPORT Krátké, střední, dlouhé vzdálenosti Slide 1b TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Slide 1c TRANSPORT Krátké, střední, dlouhé vzdálenosti Aktivní, pasivní Kapalin,
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
Rostlinná cytologie. Přednášející: RNDr. Jindřiška Fišerová, Ph.D. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Vápník. Deficience vápníku: - 0,4-1,5% DW. - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM)
") Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
10. Minerální výživa rostlin na extrémních půdách
 10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
10. Minerální výživa rostlin na extrémních půdách Extrémní půdy: Kyselé Alkalické Zasolené Kontaminované těžkými kovy Kyselé půdy Procesy vedoucí k acidifikaci (abnormálnímu okyselení): Zvětrávání hornin
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Rostlinná buňka jako osmotický systém
 Rostlinná buňka jako osmotický systém Voda se do rostlinné buňky i z ní pohybuje pouze pasivně, difusí. Hnací silou difuse vody jsou rozdíly tzv. vodního potenciálu ( ). Vodní potenciál je chemický potenciál
Rostlinná buňka jako osmotický systém Voda se do rostlinné buňky i z ní pohybuje pouze pasivně, difusí. Hnací silou difuse vody jsou rozdíly tzv. vodního potenciálu ( ). Vodní potenciál je chemický potenciál
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky Reakce průduchů na světlo
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Úvod do biologie rostlin Buňka ROSTLINNÁ BUŇKA
 Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
MBRO ) Membránový transport
 Membránový transport") MBRO1 2018 3) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů g) Sekreční dráha proteinů h) Rozpad
MBRO1 2018 3) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů g) Sekreční dráha proteinů h) Rozpad
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Transport živin do rostliny. Radiální a xylémový transport. Mimokořenová výživa rostlin.
 Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK
 Milan Chovanec Ústav fyziologie 2.LF UK") Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK Fyziologie srdce Akční potenciál v srdci (pracovní myokard) Automacie srdeční aktivity a převodní systém Mechanismus
Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK Fyziologie srdce Akční potenciál v srdci (pracovní myokard) Automacie srdeční aktivity a převodní systém Mechanismus
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
6. Mikroelementy a benefiční prvky. 7. Toxické prvky Al a těžké kovy, mechanismy účinku, obranné mechanismy rostlin
 1. Základní úvod do problematiky Historie studia minerální výživy rostlin, obecné mechanismy příjmu minerálních živin, transportní procesy na membránách. 2. Příjem minerálních živin kořeny rostlin a jejich
1. Základní úvod do problematiky Historie studia minerální výživy rostlin, obecné mechanismy příjmu minerálních živin, transportní procesy na membránách. 2. Příjem minerálních živin kořeny rostlin a jejich
4) Reakce rostlin k abiotickému stresu
 Reakce rostlin k abiotickému stresu") 2010 4) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení 1 Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing
2010 4) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení 1 Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing
RVR e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu
 Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu") 1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch
 Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
TRANSPORT PŘES BUNEČNÉ MEMBRÁNY
 TRANSPORT PŘES BUNEČNÉ MEMBRÁNY Plasmatická membrána - selektivně permeabilní bariera: esenciální molekuly (cukry, AA, lipidy.) vstupují do bunky; metabolické intermediáty zustávají v bunce; odpadní látky
TRANSPORT PŘES BUNEČNÉ MEMBRÁNY Plasmatická membrána - selektivně permeabilní bariera: esenciální molekuly (cukry, AA, lipidy.) vstupují do bunky; metabolické intermediáty zustávají v bunce; odpadní látky
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky
 fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky") FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
Vnitřní prostředí organismu. Procento vody v organismu
 Vnitřní prostředí organismu Procento vody v organismu 2 Vnitřní prostředí organismu Obsah vody v různých tkáních % VODY KREV 83% SVALY 76% KŮŽE 72% KOSTI 22% TUKY 10% ZUBNÍ SKLOVINA 2% 3 Vnitřní prostředí
Vnitřní prostředí organismu Procento vody v organismu 2 Vnitřní prostředí organismu Obsah vody v různých tkáních % VODY KREV 83% SVALY 76% KŮŽE 72% KOSTI 22% TUKY 10% ZUBNÍ SKLOVINA 2% 3 Vnitřní prostředí
Membránové potenciály
 Membránové potenciály Vznik a podstata membránového potenciálu vzniká v důsledku nerovnoměrného rozdělení fyziologických iontů po obou stranách membrány nestejná propustnost membrány pro různé ionty různá
Membránové potenciály Vznik a podstata membránového potenciálu vzniká v důsledku nerovnoměrného rozdělení fyziologických iontů po obou stranách membrány nestejná propustnost membrány pro různé ionty různá
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Tělesné kompartmenty tekutin. Tělesné kompartmenty tekutin. Obecná patofyziologie hospodaření s vodou a elektrolyty.
 Obecná patofyziologie hospodaření s vodou a elektrolyty. 2. 4. 2008 Tělesné kompartmenty tekutin Voda je v organismu kompartmentalizovaná do několika oddílů. Intracelulární tekutina (ICF) zahrnuje 2/3
Obecná patofyziologie hospodaření s vodou a elektrolyty. 2. 4. 2008 Tělesné kompartmenty tekutin Voda je v organismu kompartmentalizovaná do několika oddílů. Intracelulární tekutina (ICF) zahrnuje 2/3
10) Reakce rostlin k abiotickému stresu
 Reakce rostlin k abiotickému stresu") 2015 10) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
2015 10) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky. Reakce průduchů na vlhkost vzduchu. Reakce průduchů na vodní stres
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Anorganické látky v buňkách - seminář. Petr Tůma některé slidy převzaty od V. Kvasnicové
 Anorganické látky v buňkách - seminář Petr Tůma některé slidy převzaty od V. Kvasnicové Zastoupení prvků v přírodě anorganická hmota kyslík (O) 50% křemík (Si) 25% hliník (Al) 7% železo (Fe) 5% vápník
Anorganické látky v buňkách - seminář Petr Tůma některé slidy převzaty od V. Kvasnicové Zastoupení prvků v přírodě anorganická hmota kyslík (O) 50% křemík (Si) 25% hliník (Al) 7% železo (Fe) 5% vápník
Hypotonie děložní. MUDr.Michal Koucký, Ph.D. Gynekologicko-porodnická klinika VFN a 1.LF UK
 Hypotonie děložní MUDr.Michal Koucký, Ph.D. Gynekologicko-porodnická klinika VFN a 1.LF UK Fyziologie děložní kontraktility Interakce aktin vs. myosin v myocytech Myocyty propojeny pomocí gap a tight junctions
Hypotonie děložní MUDr.Michal Koucký, Ph.D. Gynekologicko-porodnická klinika VFN a 1.LF UK Fyziologie děložní kontraktility Interakce aktin vs. myosin v myocytech Myocyty propojeny pomocí gap a tight junctions
VAKUOLY - voda v rostlinné buňce
 VAKUOLY - voda v rostlinné buňce Úvod: O vakuole: Vakuola je membránová struktura, která je součástí většiny rostlinných buněk. Může zaujímat 30-90% objemu buňky. Vakuola plní v rostlinné buňce mnoho důležitých
VAKUOLY - voda v rostlinné buňce Úvod: O vakuole: Vakuola je membránová struktura, která je součástí většiny rostlinných buněk. Může zaujímat 30-90% objemu buňky. Vakuola plní v rostlinné buňce mnoho důležitých
5. Lipidy a biomembrány
 5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
5. Lipidy a biomembrány Obtížnost A Co je chybného na často slýchaném konstatování: Biologická membrána je tvořena dvojvrstvou fosfolipidů.? Jmenujte alespoň tři skupiny látek, které se podílejí na výstavbě
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
MEMBRÁNOVÝ TRANSPORT
 MEMBRÁNOVÝ TRANSPORT Membránový transport Soubor procesů umožňujících látkám různého typu překonat barieru biologické membrány. Buněčné membrány jsou polopropustné (semipermeabilní) Volný přístup přes
MEMBRÁNOVÝ TRANSPORT Membránový transport Soubor procesů umožňujících látkám různého typu překonat barieru biologické membrány. Buněčné membrány jsou polopropustné (semipermeabilní) Volný přístup přes
6) Reakce rostlin k abiotickým stresům
 Reakce rostlin k abiotickým stresům") MBRO2 2018 6) Reakce rostlin k abiotickým stresům 1 a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
MBRO2 2018 6) Reakce rostlin k abiotickým stresům 1 a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
Schéma epitelu a jeho základní složky
 Schéma epitelu a jeho základní složky Těsný spoj Bazální membrána Transcelulární tok Paracelulární tok LIS - Laterální intercelulární prostor Spojovací komplexy epiteliálních buněk Spojovací komplexy epiteliálních
Schéma epitelu a jeho základní složky Těsný spoj Bazální membrána Transcelulární tok Paracelulární tok LIS - Laterální intercelulární prostor Spojovací komplexy epiteliálních buněk Spojovací komplexy epiteliálních
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, Přírodovědecká fakulta UP
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, Přírodovědecká fakulta UP Funkce cytoplazmatické membrány Cytoplazmatická membrána odděluje vnitřní obsah buňky od vnějšího prostředí. Pro většinu látek
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, Přírodovědecká fakulta UP Funkce cytoplazmatické membrány Cytoplazmatická membrána odděluje vnitřní obsah buňky od vnějšího prostředí. Pro většinu látek
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Transport v rostlinách. Kateřina Schwarzerová Olga Votrubová
 Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
Transport v rostlinách Kateřina Schwarzerová Olga Votrubová Transport v rostlinách Rostlinou jsou transportovány především následující látky: Voda: přijímána většinou kořeny Minerální látky: obvykle přijímány
5) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") 2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
Stomatální vodivost a transpirace
 Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
FYTOREMEDIACE LÉČIV A JEJICH REZIDUÍ
 FYTOREMEDIACE LÉČIV A JEJICH REZIDUÍ Petr Soudek Ústav experimentální botaniky Akademie věd ČR Centralizovaný rozvojový projekt MŠMT č. C29: Integrovaný systém vzdělávání v oblasti výskytu a eliminace
FYTOREMEDIACE LÉČIV A JEJICH REZIDUÍ Petr Soudek Ústav experimentální botaniky Akademie věd ČR Centralizovaný rozvojový projekt MŠMT č. C29: Integrovaný systém vzdělávání v oblasti výskytu a eliminace
Eva Benešová. Dýchací řetězec
 Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
Eva Benešová Dýchací řetězec Dýchací řetězec Během oxidace látek vstupujících do různých metabolických cyklů (glykolýza, CC, beta-oxidace MK) vznikají NADH a FADH 2, které následně vstupují do DŘ. V DŘ
in Cl - Δµ s = RTln(C si /C so ) + zf(e i - E o ) MBR ) Membránový transport
 + zf(e i - E o ) MBR ) Membránový transport") MBR1 2016 3) Membránový transport d) Kanály e) Přenašeče a cotransportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů g) Sekreční dráha proteinů h) Rozpad proteinu
MBR1 2016 3) Membránový transport d) Kanály e) Přenašeče a cotransportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů g) Sekreční dráha proteinů h) Rozpad proteinu
Teoretický úvod: FOTOTROPISMUS. Praktikum fyziologie rostlin
 Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
BIOCHEMIE GIT. Tomáš Kuˇ. cera
 BIOCHEMIE GIT ˇ MALABSORP CNÍ SYNDROM Tomáš Kuˇ cera tomas.kucera@lfmotol.cuni.cz Ústav lékaˇrské chemie a klinické biochemie 2. lékaˇrská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole
BIOCHEMIE GIT ˇ MALABSORP CNÍ SYNDROM Tomáš Kuˇ cera tomas.kucera@lfmotol.cuni.cz Ústav lékaˇrské chemie a klinické biochemie 2. lékaˇrská fakulta, Univerzita Karlova v Praze a Fakultní nemocnice v Motole
Obecná fyziologie smyslů. Co se děje na membránách.
 Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Obecná fyziologie smyslů. Co se děje na membránách.
 Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Dynamický fluidní model membrány 2008/11
 Dynamický fluidní model membrány 2008/11") RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány Membránový
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány Membránový
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Lékařská chemie přednáška č. 3
 Lékařská chemie přednáška č. 3 vnitřní prostředí organismu transport látek v membráně Václav Babuška Vaclav.Babuska@lfp.cuni.cz Vnitřní prostředí organismu Procento vody v organismu 2 Vnitřní prostředí
Lékařská chemie přednáška č. 3 vnitřní prostředí organismu transport látek v membráně Václav Babuška Vaclav.Babuska@lfp.cuni.cz Vnitřní prostředí organismu Procento vody v organismu 2 Vnitřní prostředí
BUNĚČNÉ JÁDRO FYZIOLOGIE BUŇKY JADÉRKO ENDOPLASMATICKÉ RETIKULUM (ER)
") BUNĚČNÉ JÁDRO FYZIOLOGIE BUŇKY Buněčné jádro- v něm genetická informace Úkoly jádra-1) regulace dělení, zrání a funkce buňky; -2) přenos genetické informace do nové buňky; -3) syntéza informační RNA (messenger
BUNĚČNÉ JÁDRO FYZIOLOGIE BUŇKY Buněčné jádro- v něm genetická informace Úkoly jádra-1) regulace dělení, zrání a funkce buňky; -2) přenos genetické informace do nové buňky; -3) syntéza informační RNA (messenger
Fytoremediace II. Příjem látek. Petr Soudek
 Fytoremediace II. Příjem látek Petr Soudek PROČ ROSTLINY PŘIJÍMAJÍ TOXICKÉ KOVY? Stimulují vývoj a růst rostlin Účastní se biochemických reakcí (fotosyntéza, dýchání) Součást proteinů, jejich aktivace
Fytoremediace II. Příjem látek Petr Soudek PROČ ROSTLINY PŘIJÍMAJÍ TOXICKÉ KOVY? Stimulují vývoj a růst rostlin Účastní se biochemických reakcí (fotosyntéza, dýchání) Součást proteinů, jejich aktivace
I rostliny mají hormony!... co a jak s auxinem. Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha
 I rostliny mají hormony!... co a jak s auxinem Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha Rostliny a tvarová (a vývojová) různorodost Reakce na okolní prostředí Reakce na nepříznivé podmínky
I rostliny mají hormony!... co a jak s auxinem Eva Zažímalová Ústav experimentální botaniky AV ČR, Praha Rostliny a tvarová (a vývojová) různorodost Reakce na okolní prostředí Reakce na nepříznivé podmínky
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
*Ivana FELLNEROVÁ, PřF UP Olomouc*
 Faktory ovlivňující transport přes membrány Velikost molekul: Malé molekuly jako voda, kyslík, kysličník uhličitý mohou volně procházet přes membrány, na rozdíl od většiny větších molekul. Rozpustnost
Faktory ovlivňující transport přes membrány Velikost molekul: Malé molekuly jako voda, kyslík, kysličník uhličitý mohou volně procházet přes membrány, na rozdíl od většiny větších molekul. Rozpustnost
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 4. Membránové proteiny Ivo Frébort Lipidová dvojvrstva Biologické membrány Integrální membránové proteiny Transmembránové proteiny Kovalentně ukotvené membránové
Struktura a funkce biomakromolekul KBC/BPOL 4. Membránové proteiny Ivo Frébort Lipidová dvojvrstva Biologické membrány Integrální membránové proteiny Transmembránové proteiny Kovalentně ukotvené membránové
Magnetotaktické bakterie
 Magnetotaktické bakterie G- bakterie, objeveny v 60.l. 20.stol. koky, bacily, vibria, spirily; pohyb bičíky obligátně mikroaerofilní nebo anaerobní negativní aerotaxe výskyt: svrchní sedimenty ve vodě
Magnetotaktické bakterie G- bakterie, objeveny v 60.l. 20.stol. koky, bacily, vibria, spirily; pohyb bičíky obligátně mikroaerofilní nebo anaerobní negativní aerotaxe výskyt: svrchní sedimenty ve vodě
6) Reakce rostlin k abiotickým stresům
 Reakce rostlin k abiotickým stresům") MBRO2 2019 6) Reakce rostlin k abiotickým stresům 1 a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
MBRO2 2019 6) Reakce rostlin k abiotickým stresům 1 a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení c) Vliv vodního deficitu a zasolení na membránový transport
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Intracelulární Ca 2+ signalizace
 Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
glukóza *Ivana FELLNEROVÁ, PřF UP Olomouc*
 Prezentace navazuje na základní znalosti Biochemie, stavby a transportu přes y Doplňující prezentace: Proteiny, Sacharidy, Stavba, Membránový transport, Symboly označující animaci resp. video (dynamická
Prezentace navazuje na základní znalosti Biochemie, stavby a transportu přes y Doplňující prezentace: Proteiny, Sacharidy, Stavba, Membránový transport, Symboly označující animaci resp. video (dynamická
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
Obsah vody v rostlinách
 Transpirace 1/39 Obsah vody v rostlinách Obsah vody v protoplazmě (její hydratace) je nezbytný pro normální průběh životních funkcí buňky. Snížení obsahu vody má za následek i omezení životních dějů (pozorovatelné
Transpirace 1/39 Obsah vody v rostlinách Obsah vody v protoplazmě (její hydratace) je nezbytný pro normální průběh životních funkcí buňky. Snížení obsahu vody má za následek i omezení životních dějů (pozorovatelné
BIOMEMBRÁNY. Sára Jechová, leden 2014
 BIOMEMBRÁNY Sára Jechová, leden 2014 zajišťují ohraničení buněk- plasmatické membrány- okolo buněčné protoplazmy, bariéra v udržování rozdílů mezi prostředím uvnitř buňky a okolím a organel= intercelulární
BIOMEMBRÁNY Sára Jechová, leden 2014 zajišťují ohraničení buněk- plasmatické membrány- okolo buněčné protoplazmy, bariéra v udržování rozdílů mezi prostředím uvnitř buňky a okolím a organel= intercelulární
Přednášky z lékařské biofyziky Lékařská fakulta Masarykovy univerzity v Brně
 Přednášky z lékařské biofyziky Lékařská fakulta Masarykovy univerzity v Brně Biologické membrány a bioelektrické jevy Autoři děkují doc. RNDr. K. Kozlíkové, CSc., z LF UK v Bratislavě za poskytnutí některých
Přednášky z lékařské biofyziky Lékařská fakulta Masarykovy univerzity v Brně Biologické membrány a bioelektrické jevy Autoři děkují doc. RNDr. K. Kozlíkové, CSc., z LF UK v Bratislavě za poskytnutí některých
Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce)
") Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce) Úkol: Sepište krátký rukopis vědeckého původního článku na téma "Směrovaný transport auxinu přes plazmatickou membránu hraje úlohu
Úloha 5 k zápočtu z přednášky B130P16 (praktické základy vědecké práce) Úkol: Sepište krátký rukopis vědeckého původního článku na téma "Směrovaný transport auxinu přes plazmatickou membránu hraje úlohu
Program kursu Rostlinná buňka
 Program kursu Rostlinná buňka 1) Poznávání rostlinných buněk Buňka a vývoj jejího poznání Srovnání rostlinné a živočišné buňky Jak jsou buňky rozčleněny: membrány 2) Buněčné membrány a vakuoly rostlinných
Program kursu Rostlinná buňka 1) Poznávání rostlinných buněk Buňka a vývoj jejího poznání Srovnání rostlinné a živočišné buňky Jak jsou buňky rozčleněny: membrány 2) Buněčné membrány a vakuoly rostlinných
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11. *Ivana FELLNEROVÁ, PřF UP Olomouc*
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11 Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie PřF UP Olomouc 2008/11 Prezentace navazuje na základní znalosti z biochemie (lipidy, proteiny, sacharidy) Rozšiřuje přednášky: Stavba cytoplazmatické membrány
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Schéma rostlinné buňky
 Rostlinná buňka 1 2 3 5 vakuola 4 5 6 Rostlinná buňka je eukaryotní buňkou se základními charakteristikami tohoto typu buňky. Krom toho má některé charakteristiky typické pro rostlinné buňky, jako je předevšímř
Rostlinná buňka 1 2 3 5 vakuola 4 5 6 Rostlinná buňka je eukaryotní buňkou se základními charakteristikami tohoto typu buňky. Krom toho má některé charakteristiky typické pro rostlinné buňky, jako je předevšímř
Transport přes membránu
 Transport přes membránu Datum: 30. 12. 2012 Projekt: Využití ICT techniky především v uměleckém vzdělávání Registrační číslo: CZ.1.07/1.5.00/34.1013 Číslo DUM: VY_32_INOVACE_262 Škola: Akademie - VOŠ,
Transport přes membránu Datum: 30. 12. 2012 Projekt: Využití ICT techniky především v uměleckém vzdělávání Registrační číslo: CZ.1.07/1.5.00/34.1013 Číslo DUM: VY_32_INOVACE_262 Škola: Akademie - VOŠ,
Biologické membrány a bioelektrické jevy
 Přednášky z lékařské biofyziky Lékařská fakulta Masarykovy univerzity v Brně Biologické membrány a bioelektrické jevy Autoři děkují doc. RNDr. K. Kozlíkové, CSc., z LF UK v Bratislavě za poskytnutí některých
Přednášky z lékařské biofyziky Lékařská fakulta Masarykovy univerzity v Brně Biologické membrány a bioelektrické jevy Autoři děkují doc. RNDr. K. Kozlíkové, CSc., z LF UK v Bratislavě za poskytnutí některých
VORF Winslow Russel Briggs ( )
") VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
Bioimaging rostlinných buněk, CV.2
 Bioimaging rostlinných buněk, CV.2 Konstrukce mikroskopu (optika, fyzikální principy...) Rozlišení - kontrast Live cell microscopy Modulace kontrastu (Phase contrast, DIC) Videomikroskopia Nízký kontrast
Bioimaging rostlinných buněk, CV.2 Konstrukce mikroskopu (optika, fyzikální principy...) Rozlišení - kontrast Live cell microscopy Modulace kontrastu (Phase contrast, DIC) Videomikroskopia Nízký kontrast
Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim
 Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník
Číslo projektu CZ.1.07/1.5.00/34.0743 Název školy Autor Tematická oblast Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 31 Příjem a výdej, minerální výživa, způsob výživy, vodní režim Ročník