Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění řepky (Brassica napus L.)
|
|
|
- Tomáš Toman
- před 5 lety
- Počet zobrazení:
Transkript

 Metodika byla vypracovaná jako výstup")




1 Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění řepky (Brassica napus L.) Metodika byla vypracovaná jako výstup výzkumného projektu MZe NAZV QI 111A075 Využití biotechnologických metod, nových výchozích materiálů a efektivních postupů ve šlechtění ozimé řepky Autoři: prof. Ing. Vladislav Čurn, Ph.D., Ing. Lenka Havlíčková, Ph.D., Ing. Eva Jozová, Ing. Irena Jelínková, Mgr. Ing. Onřej Hejna, Ing. Vratislav Kučera, CSc., Ing. Miroslava Vyvadilová, CSc., Ing. Miroslav Klíma, Ph.D. České Budějovice, říjen 2014
2
3 Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění řepky (Brassica napus L.) Metodika byla vypracována jako výstup výzkumného projektu MZe NAZV QI 111A075 Využití biotechnologických metod, nových výchozích materiálů a efektivních postupů ve šlechtění ozimé řepky prof. Ing. Vladislav Čurn, Ph.D. Ing. Lenka Havlíčková, Ph.D. Ing. Eva Jozová Ing. Irena Jelínková Mgr. Ing. Ondřej Hejna Vratislav Kučera, CSc. Ing. Miroslava Vyvadilová, CSc. Ing. Miroslav Klíma, Ph.D. České Budějovice, 2014 i
4 Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění Vladislav Čurn a kol. vcurn@seznam.cz Biotechnologické centrum ZF JU v Českých Budějovicích, České Budějovice Vypracováno za podpory výzkumného projektu MZe NAZV QI 111A075 Využití biotechnologických metod, nových výchozích materiálů a efektivních postupů ve šlechtění ozimé řepky. Oponenty metodiky byli: prof. Ing. Jaroslava Ehrenbergerová, CSc., Mendelova univerzita Brno Ing. Petr Zehnálek, ÚKZÚZ ZS Hradec nad Svitavou Podíl autorů na vypracování metodiky: prof. Ing. Vladislav Čurn, Ph.D. (30%), Ing. Lenka Havlíčková, Ph.D. (30%), Ing. Eva Jozová, Ing. Irena Jelínková, Mgr. Ing. Ondřej Hejna (30%), Ing. Vratislav Kučera, CSc., Ing. Miroslava Vyvadilová, CSc., Ing. Miroslav Klíma, Ph.D. (10%) Text: 2014 Čurn V. et al. Foto: 2014 Čurn V. Vydáno bez jazykové úpravy ISBN: ii
5 Obsah: I. Cíl metodiky... 1 II. Vlastní popis metodiky... 3 II.1. Úvod... 3 II.2. Metodika izolace DNA... 7 Rostlinný materiál používaný pro izolaci DNA řepky... 7 Metody izolace DNA řepky... 7 Izolace DNA pomocí CTAB PVP (polyvinylpyrrolidon)... 7 Izolace DNA pomocí SDS... 9 II.3. Metodika analýzy DNA markerů Hybridní systémy CMS Ogu-INRA a Shaan 2A Detekce obnovitele fertility (Rfo) u systému Ogu-INRA Detekce rostlin se sterilní cytoplazmou u materiálu CMS typu Shaan 2A Detekce Rf u CMS typu Shaan 2A Detekce Rfo genu v mikrosporových embryích řepky u CMS systému Ogu- INRA Detekce autoinkompatibilních haplotypů detekce a selekce AI rostlin III. Srovnání novosti postupů IV. Popis uplatnění metodiky V. Ekonomické aspekty V. Seznam použité související literatury VI. Seznam publikací, které předcházely metodice iii
6 iv
7 I. Cíl metodiky Cílem metodiky je podat přehled o v praktickém šlechtění použitelných postupech pro selekci cílových rostlin postupech založených na technikách molekulární biologie, na technikách a přístupech molekulárního markerování. Řepka patří k našim nejvýznamnějším plodinám a tomu odpovídá i velký tlak na kvalitu a výkonost odrůd. Hybridní šlechtění a tvorba F 1 hybridů pak představuje velmi účinný nástroj pro zvýšení výnosového potenciálu, zlepšení celé řady agronomicky významných znaků a samozřejmě je i značnou výzvou pro šlechtitele, vzhledem k řadě problematických otázek, které s hybridním šlechtěním řepky souvisejí. Na IRC 2011 v Praze byla mimo jiné značná pozornost věnována i metodám molekulární biologie, molekulárním markerům a jejich aplikaci pro rozlišování genetických zdrojů o identifikaci samičích (udržovatelé pylové sterility) a samčích (obnovitelé pylové fertility) linií a stanovení stupně hybridity při výrobě hybridního osiva. Metodika je výsledkem řešení projektu NAZV QI 111A075 Využití biotechnologických metod, nových výchozích materiálů a efektivních postupů ve šlechtění ozimé řepky. Záměrem projektu bylo: zefektivnit šlechtitelské postupy pro tvorbu nových odrůd ozimé řepky s využitím nově vyvinutých, optimalizovaných a ověřených biotechnologických a analytických metod; vytvořit nové výchozí materiály pro šlechtění liniových a hybridních odrůd, vhodných pro potravinářské i technické využití; využít metody molekulární genetiky, explantátových kultur a nové expeditivní metody hodnocení pro výběr a tvorbu perspektivních genotypů; pomocí efektivních šlechtitelských postupů získat materiály pro tvorbu nových liniových a hybridních odrůd ozimé řepky, schopných konkurovat zahraničním odrůdám ve významných hospodářských znacích. Jedním z cílů projektu pak bylo vytvořit rodičovské komponenty a hybridy řepky na bázi systémů CMS s využitím biotechnologických metod a odvodit alelicky specifické markery pro selekci rostlin nesoucích geny obnovy fertility. Metodika navazuje na předchozí metodické výstupy projektu: Metodika analýzy molekulárních markerů pro účely popisu genetických zdrojů a hodnocení jejich diverzity a Metodika detekce a molekulární selekce autoinkompatibilních linií, kdy předmětem těchto výstupů bylo využití molekulárních metod při hodnocení genetické diverzity genových zdrojů řepky a pro účely výběru geneticky vzdálených rodičovských komponent pro křížení při tvorbě liniových a hybridních odrůd a dále selekce rostlin nesoucích znaky autoinkompatibility. Předkládaná metodika pak kompletuje tento soubor a je zaměřena na techniky selekce cílových rostlin z pohledu šlechtění hybridů na bázi CMS Ogu/INRA a Shaan 2A. 1
8 2
9 II. Vlastní popis metodiky II.1. Úvod Proč techniky molekulární biologie a proč snaha o jejich aplikaci v programech hybridního šlechtění řepky? Selekce s využitím markerů neboli MAS (Marker Assisted Selection) resp. modifikace tohoto postupu s větším důrazem na molekulární techniky MMAS (Molecular Marker Assisted Selection) je již řadu let aktuálním trendem ve šlechtění rostlin. Tento metodický přístup je založen na selekci cílových rostlin/potomstev na základě výsledků analýzy molekulárních markerů. Aplikací těchto postupů lze eliminovat vliv vnějšího prostředí, lze pak účinněji selektovat rostliny i na jiném základě, než-li pouhém fenotypovém výběru a přístupy MAS/MMAS lze i chápat jako cestu směrem ke genotypové selekci (GS), cílené selekci na základě důkladné znalosti genetické konstituce šlechtitelského ideotypu (Newell, Jannink 2013). Širší aplikace metodických postupů založených na MAS/MMAS může výrazně urychlit šlechtitelský proces a zefektivnit šlechtění (Varshney, Tuberosa 2007). Genetický marker (signální gen) se používá pro označení jasně se fenotypově projevujícího znaku s jednoduchou dědičností. Označení marker pak předpokládá spojení tohoto znaku genovou vazbou s jinými kvantitativními či kvalitativními znaky (Sozinov 1985). Genetické biochemické markery se musí vyznačovat dostatečnou genetickou a jí odpovídající fenotypovou variabilitou, vysokou expresivitou a penetrancí a rovněž vysokou heritabilitou, tj. nezávislostí na podmínkách prostředí (Černý, Šašek 1996). A ačkoli byly předchozí definice genetických markerů vysloveny ještě před érou PCR, OMICS a NextGen technologií, jejich platnost není a nemůže být zpochybňována. Výhodu markerů založených na polymorfismu DNA lze spatřovat v aplikovatelnosti u všech organismů, kde je zvládnutá technika izolace DNA, v jejich nezávislosti na podmínkách prostředí, značném počtu markerovacích technik a markerů, v možnosti použití nedestruktivní analýzy a možnosti jejich využití i pro analýzu rostlin ve velmi raných ontogenetických stádiích (Varshney et al. 2009). Techniky molekulární biologie technika molekulárních markerování pak nabízí šlechtiteli mocný nástroj pro jasné a přesné odhalení genetické konstituce analyzovaných rostlin z pohledů cílových znaků, nástroj pro velmi přesnou selekci cílových rostlin/genotypů. Přehled technik/markerů používaných u řepky je uveden v review: Čurn, Sáková, Graman (1995), Čurn, Sáková (1997) a Čurn, Žaludová (2007). Jednou z oblastí aplikace molekulárních markerů je pak selekce cílových rostlin/genotypů v programech hybridního šlechtění řepky. Hybridní odrůdy řepky ozimé jsou v našich podmínkách dlouhodobě pěstovány a od roku 1998, kdy byl uveden na český trh první komerční hybrid, se podíl hybridních odrůd v osetých 3
10 plochách řepky neustále zvyšuje. V roce 2008 došlo k výraznému nárůstu ploch hybridů v porovnání s plochami osetými liniovými odrůdami a podíl hybridních odrůd dosáhl 35 %. V roce 2011 podíl hybridních odrůd přesáhl hranici 50% a v roce 2014 pak dosáhl téměř 80% (Baranyk 2014). Zájem o hybridní odrůdy je dán nejen módní vlnou pěstování F 1 hybridů, ale u řepky došlo v důsledku hybridního šlechtění a zavedení hybridních odrůd k dosažení podstatného pokroku nejen ve výkonnosti, ale také ve stabilitě vysokých výnosů v rozdílných podmínkách a ročnících. Ve srovnání s liniovými odrůdami hybridní odrůdy disponují vyšším výnosovým potenciálem, poskytují stabilnější výnosy v různých pěstitelských podmínkách, jsou odolnější vůči stresu (zimovzdornost a suchovzdornost), lépe reagují na pozdější výsevy, účinněji využívají živiny a i přes vyšší cenu osiva hybridní odrůdy zajišťují stabilní vysokou produkci řepky a zajišťují ziskovost. Heterozní efektu u řepky dosahuje až 10% (Röbbelen 1985; Brandle, McVetty 1989; Kudla 1996; Zehnálek, Holubář 2004; Zehnálek et al. 2005; Zehnálek et al. 2006) a heterozní efekt se projevuje v podobě zvýšení výnosu semene, výnosu oleje, výšky rostlin, vitality a jistoty přezimování (Paulmann 1999; Koprna et al. 2009). V současnosti jsou v Evropě nejvíce rozšířené hybridní systémy MSL (Mänlicher Sterilität Lembke), CMS (Cytoplasmatic Male Sterility) Ogu-INRA a SAFECROSS (Syngenta). Ve světě, zejména v Číně je pak využíván i CMS systém Polima/Shaan. Z mnoha směrů šlechtění výrazně vyčnívá systém Ogura, který umožnil vytvářet restaurované hybridy z různých rodičovských kombinací. Původ tohoto typu cytoplazmatické samčí sterility je v literatuře často chybně udáván a jako zdroj sterility je mylně uváděna japonská odrůda ředkve 'Ogura' (Koprna et al. 2007). CMS byla poprvé v čeledi Brassicaceae popsána u japonských odrůd ředkve Raphanus sativus H. Ogurou a byla využívána pro účely šlechtění hybridních odrůd (Ogura 1968). "Ogura CMS type" byla záhy introdukována do významných zástupců rodu Brassica (B. oleracea a B. napus) na pracovišti INRA ve Francii a tímto krokem bylo umožněno širší využití CMS systému v hybridním šlechtění řepky (Pelletier et al. 1983, 1987; Budar et al. 2004). První CMS rostliny byly získány na základě mezirodové hybridizace a několikanásobným zpětným křížením (Bannerot et al. 1974) a bohužel kromě stabilního projevu sterility byla u získaných rostlin detekována celá řada nepříznivých vlastností (Bannerot et al. 1977). Tyto znaky byly způsobeny nekompatibilitou mezi jádrem a cytoplazmou a po fúzi protoplastů a získání cybridní kombinace, která nesla chloroplasty B. napus a mitochondrie R. sativus byly tyto negativní projevy cytoplazmy R. sativus eliminovány (Menczel et al. 1987; Pelletier et al. 1987). První hybridní odrůdy založené na systému Ogu-INRA registrované v České republice byly kompozitní, tj. osivo bylo tvořeno ze % pylově sterilním hybridním komponentem a z % opylovačem nejčastěji výkonnou odrůdou. Těmito hybridy byly Synergy (1998) a Betty (1999) (Zehnálek, Holubář, 2000). Další etapu hybridního šlechtění představovaly 3-liniové hybridy Embleme (2002), jehož finální generace štěpí v poměru 1:1 na plně fertilní a sterilní, dále topcross hybrid Spirit (2003), jehož finální generace štěpí na cca 70% pylově fertilních rostlin a 30% pylově sterilních. Aktuálně jsou pěstovány pylově fertilní restaurované hybridy (Zehnálek et al. 2005; Baranyk 2014). 4
11 Pro tvorbu restaurovaných hybridů je pak potřebné šlechtit sterilní mateřské komponenty (S cytoplazma a rfo geny obnovy fertility v jádře), udržovatele sterility (F cytoplazma a rfo geny obnovy fertility v jádře) a obnovitele fertility (úplný obnovitel s fertilní/sterilní cytoplazmou a dominantně homozygotní sestavou genů Rfo v jádře). V rámci sdružení Česká řepka je značná pozornost věnována i využití CMS systému Shaan 2A a šlechtění F 1 hybridních odrůd využívající tento systém sterility. Zcela originální a jedinečné jsou zejména nové genetické zdroje - sterilní linie a DH linie obnovitelů fertility s požadovanou 00 kvalitou pro systém CMS Shaan 2A. Tento systém se ukazuje jako velmi perspektivní pro tvorbu hybridů i v našich podmínkách a je rozpracován v evropských podmínkách k praktickému využití pouze v ČR. Jeho předností je zejména nepřítomnost vazby genu obnovy fertility s vysokým obsahem glukosinolátů či jeho stabilita v široké škále teplotních podmínek, na rozdíl od teplotně citlivého příbuzného CMS systému Polima (pol), jehož nestabilita limituje vlastní využití v hybridním šlechtění. Nedostatkem z pohledu selekce cílových rostlin je pak nedostupnost vhodných selekčních markerů. U řepky je možné pro produkci F 1 hybridů využít i systému autoinkompatibility (AI). U řepky se, obdobně jako u jiných brukvovitých rostlin, přirozeně vyskytuje sporofytický typ AI, kde pylový determinant je produktem buněk tapeta. K hybridnímu šlechtění jsou vhodné pouze AI linie s vysokou účinností AI reakce, aby byl podíl hybridního osiva maximální i v polních podmínkách. Podíl hybridního osiva je možno stanovit prakticky pouze na základě vhodného genetického markeru. Autoinkompatibilita je kontrolována jediným úsekem DNA tzv. S lokusem, který má velký počet alel (Ockendon 1974; de Nettancourt 1977). S haplotypy u rodu Brassica jsou podle svého fenotypového účinku na základě klasické genetické analýzy rozděleny do dvou tříd I a II (Nasrallah et al. 1991). S haplotypy třídy I jsou dominantní nad třídou II (Nasrallah, Nasrallah 1993). Většina pěstovaných genotypů řepky pak obsahuje v A genomu S haplotypy třídy I (homologní s S haplotypem S 47 B. rapa) a v C genomu S haplotypy třídy II (homologní s S haplotypem S 15 B. oleracea) (Okamoto et al. 2007; Zhang et al. 2008a). Autokompatibilita u řepky je pak vyvolána mutací v dominantním S haplotypu, která vede k supresi funkčního recesivního S haplotypu (Okamoto et al. 2007). Účinným nástrojem pro šlechtění a selekci žádoucích rostlin s funkčním projevem autoinkompatibility (obsahující funkční recesivní S haplotyp) je postup založený na využití molekulární analýzy. Selekční markery je možné detekovat v raných fázích ontogeneze a vhodnou kombinací molekulárního a klasického šlechtitelského postupu lze docílit účinné a přesné selekce. Předkládaná metodika je zacílena na využití molekulárních markerů na bázi analýzy DNA pro účely identifikace a selekce cílových genotypů používaných v hybridním šlechtění CMS systému Ogu/INRA a Shaan 2A. 5
12 6
13 II.2. Metodika izolace DNA Rostlinný materiál používaný pro izolaci DNA řepky děložní lístky (1-2 týdny po vyklíčení) plně vyvinuté pravé listy somatická embrya, kotyledonární pletiva DNA je izolována dle příslušného protokolu z čerstvého materiálu (odebraného ve skleníkových nebo v polních podmínkách a ihned zpracovávaného, popřípadě uchovávaného do doby izolace na ledu po dobu max 2 4 hod), nebo z materiálu zamraženého (po odebrání je materiál hluboce zmražen a uchováván při teplotě - 80 C, krátkodobě je možné uchování i při teplotě -20 C). Metody izolace DNA řepky DNA je izolována z materiálu čerstvého nebo zamraženého. O výtěžnosti, čistotě a možnostech použití takto získané DNA se zmiňujeme v závěru této kapitoly. DNA je možno izolovat mikroextrakčními metodami (komerčně dostupnými kity) nebo pomocí standardních metod izolace DNA. Izolace DNA pomocí CTAB PVP (polyvinylpyrrolidon) Metoda je založena na schopnosti CTAB (cetyltrimetylamoniumbromid) vytvářet komplex s nukleovými kyselinami, který je při vysoké koncentraci solí rozpustný (0,7 M NaCl), ale při snížené koncentraci solí (0,45 M NaCl) vytváří sraženinu (Murray a Thompson 1980). CTAB zároveň působí jako detergenční činidlo, které uvolňuje DNA z komplexu membrán a proteinů. Na základě rozdílné rozpustnosti CTAB v porovnání s DNA je lze oddělit a získat dostatečně čistou rostlinnou DNA. Přidání PVP (polyvinylpyrrolidon) odstraňuje kontaminanty a umožňuje získání čisté a kvalitnější DNA. 1. Připravíme si roztok 2x PVP CTAB a 1 % merkaptoethanolu, na jeden vzorek počítáme 500 µl roztoku. Připravený roztok dáme předehřát na 65 C. 2. Rostlinné pletivo můžeme rozdrtit v tekutém dusíku. Pokud nepoužíváme tekutý dusík do sterilních 1,5 ml mikrocentrifugačních zkumavek dáme rostlinné pletivo (cca 100 mg), ze kterého chceme izolovat DNA. Pro lepší drcení pletiva přidáme sterilní křemičitý písek. 3. Ke každému vzorku přidáme 500 µl předehřátého pufru, pletivo rozdrtíme a promícháme s pufrem. Necháme 45 min inkubovat při 65 C. Během inkubace každých cca 15 min lehce promícháme. 7
14 4. Po centrifugaci rpm 10 min se převede supernatant (500 µl) do nových 1,5 ml mikrocentrifugačních zkumavek a přidá se 500 µl chloroformu s IAA, směs se 10 min promíchává a následně centrifuguje 5 min při rpm. 5. Do nových mikrocentrifugačních zkumavek odpipetujeme vodnou fázi. Přidáme 1/5 objemu 5 % CTAB a směs se promíchá, opět se přidá 500 µl směsi chloroform IAA a 10 min protřepáváme. Centrifugujeme 5 min maximální rychlostí při pokojové teplotě. 6. Do nových mikrocentrifugačních zkumavek přepipetujeme vodnou fázi. Přidáme 2/3 objemu isopropanolu (přibližně 250 µl) a 2 3x lehce promícháme. Dáme na 30 min (až na noc) do mrazáku (-20 C). Centrifugujeme 5 min při 4 C maximální rychlostí. DNA by se měla zachytit na dně mikrocentrifugační zkumavky. Odstraníme supernatant. 7. Přidáme 300 µl 1x TE a necháme min inkubovat při 37 C. 8. Přidáme 2 objemy (600 µl) ledového (z mrazáku) 96 % ethanolu, 2 3x lehce promícháme. Vzorky dáme do mrazáku (-20 C) minimálně na 20 min, maximálně na 12 hodin (větší výtěžnost DNA). 9. Vzorky vyndáme z mrazáku a 10 min centrifugujeme maximální rychlostí při 4 C. DNA by měla vytvořit viditelný pelet na dně mikrocentrifugační zkumavky. Odstraníme supernatant. 10. Přidáme 1 ml ledového 70 % ethanolu, 2 3x lehce promícháme. Centrifugujeme 2 min maximální rychlostí při teplotě 4 C. Okamžitě odstraníme všechen supernatant, pro odstranění viditelných nečistot opakujeme tento krok 2x. Vzorky necháme dobře vysušit (max. 3 hodiny). 12. Podle množství peletu (DNA) přidáme µl 1x TE pufru nebo sterilní vody (rozpustit 40 min při 37 C). Pro dokonalé přečištění můžeme zopakovat postup od bodu 5. Přidáme 1 µl RNázy A a necháme 30 min inkubovat ve 37 C. Chemikálie: kapalný dusík ethanol (čistý, 96 % ethanol nedenaturovaný) šupinkový led 2x PVP CTAB extrakční pufr (2 % CTAB, 100 mm Tris HCl, ph=8,0, 20 mm EDTA, 1,4 M NaCl, 1 % 2-merkaptoetanol, 1 % PVP 40000) 2-merkaptoethanol 5 % CTAB chloroform IAA (24 : 1) 1x TE pufr sterilní isopropanol 70 % ethanol RNáza A 2 mg/ml Přístroje: chlazená centrifuga, sada automatických pipet, homogenizátory, vortex, třepací vodní lázeň nebo třepací dry-blok, termostat, analytické váhy, ph metr, mrazák, digestoř 8
15 Izolace DNA pomocí SDS Tento postup izolace je vhodný jako rychlá skríningová metoda, u řepky poskytuje dostatečné množství DNA pro provedení PCR analýz a vychází z postupu publikovaného v práci Edwards et al. (1991). 1. rostlinné pletivo rozdrtíme homogenizátorkem přímo v 1,5 ml mikrocentrifugační zkumavce ve 100 µl extrakčního pufru. 2. K homogenátu přidáme 300 µl extrakčního pufru a 5 s vortexujeme (tato extrakční směs může být ponechána při laboratorní teplotě i déle než 1 hodinu, dokud nejsou všechny vzorky homogenizovány). 3. Extrakt centrifugujeme 1 min při rpm a 300 µl supernatantu přeneseme do čisté mikrocentrifugační zkumavky. 4. K supernatantu přidáme 300 µl isopropanolu vychlazeného na -20 C, jemně promícháme (3 4x převrátit zkumavku) a směs necháme 2 min srážet. Centrifugujeme 5 min při rpm. 5. Osušený pelet rozpustíme v 50 µl TE pufru. DNA je stabilní ve 4 C déle než 1 rok. Chemikálie: SDS extrakční pufr (200 mm Tris HCl (ph=7,5), 250 mm NaCl, 25 mm EDTA, 0,5 % SDS) Isopropanol TE pufr (10 mm Tris HCl, ph=7,5; 0,1 mm EDTA) Přístroje: centrifuga, sada automatických pipet, homogenizátory, vortex, třepací vodní lázeň nebo třepací dry-blok, termostat, analytické váhy, ph metr, lednice 9
16 10
17 II.3. Metodika analýzy DNA markerů Hybridní systémy CMS Ogu-INRA a Shaan 2A Hybridní systémy Ogu-INRA i Shaan 2A jsou typem cytoplazmaticko-genové sterility, při které je výsledný genotyp mateřské pylově sterilní linie závislý nejen na "sterilní cytoplazmě" - plazmotypu S, ale také na přítomnosti genů obnovy fertility Rf (resp. Rfo) v jádře. Genetická konstituce mateřských pylově sterilních CMS linií je (S)rfrf, kde S značí sterilní cytoplazmu a rf jaderný gen pro obnovu fertility v homozygotně recesivní konstituci. Sterilita ("sterilní cytoplazma") je determinována mitochondriálními geny. Gen obnovy fertility Rf má dominantní charakter a interakce dominantního genu obnovy fertility v jádře a sterilní cytoplazmy vede k obnově fetrtility. Pylově fertilní udržovatelé sterility mají založení (F)rfrf, kde F značí nomální (fertilní) cytoplazmu. Úplný obnovitel fertility může mít cytoplazmu plazmotypu S nebo F, ale musí být dominantně homozygotní a v jádře nese genotyp RfRf. Pokud dojde k opylení sterilní linie heterozygotním obnovitelem fertility Rfrf, výsledkem je generace F 1, kde vyštěpuje 50 % fertilních a 50 % sterilní rostlin. Komponenta Cytoplazma Jádro Fenotyp (produkce pylu) CMS sterilní linie S rfrf Sterilní Udržovatel sterility F rfrf Fertilní Neúplný obnovitel fertility S/F Rfrf Fertilní Úplný obnovitel fertility S/F RfRf Fertilní Otcovské linie - obnovitelé fertility se vytvářejí z původních výchozích materiálů opakovaným zpětným křížením s perspektivními liniemi typu (F)rfrf, kterými jsou prakticky veškeré pylově fertilní odrůdy, šlechtitelské materiály a dihaploidní (DH) linie ozimé řepky. Pro urychlení a zefektivnění tvorby Rf linií se využívá metoda produkce dihaploidních (DH) linií technikou mikrosporových kultur (Vyvadilová a kol. 2008). Tato metoda umožňuje získat dominantní homozygoty pro Rf gen z kříženců s donory požadovaných znaků prakticky během jedné generace. Nevýhodou tohoto postupu je nicméně fakt, že selekce cílových rostlin homozygotních obnovitelů fertility je prováděna až v době květu (selekce rostlin fenotypově vykazujících pylovou fertilitu u genotypů nesoucích sterilní cytoplazmu) a ověření schopnosti obnovovat fertilitu je provedeno až na základě experimentálního křížení a hodnocení nasazení semen u F1 hybridů. Molekulární přístup umožňuje selekci rostlin nesoucích Rf gen (respektive dominantní alelu tohoto genu) v raných ontogenetických stadiích a pokud je vhodně kombinován s flow-cytometrickou analýzou lze takto rychle a spolehlivě selektovat diploidní dominanntě homozytotní rostliny (DH regeneranty). 11
18 Detekce obnovitele fertility (Rfo) u systému Ogu-INRA Detekce je založena na PCR detekci přítomnosti dominantní alely genu Rfo u analyzovaných rostlin. Pro PCR analýzu byly pro účely předkládané metodiky použity primery navržené v úseku genu PPR; PPR-B (GenBank AJ550021) RsPPRB-F a RsPPRB-R, které generovaly u rostlin nesoucích gen k obnově fertility amplikon o velikosti ~ 1,5kb. PCR reakci je možné doplnit o kontrolní primery amplifikující nekódující úsek o velikosti ~ 200bp ve všech vzorcích. Dále lze použít primery SG34U a SG34L (Primard-Brissed 2005), detekující rostliny nesoucí gen k obnově fertility přítomností amplikonu o velikosti ~ 1kb. Sekvence primerů používaných pro PCR analýzu. Primer Sekvence 5 3 Annealing RsPPRB-F GAAGCTCTTGCTACCCATCG 59 C RsPPRB-R TGACATGCTTCGATCTCGTC 59 C SG34U TATATTGTACCTTTGCCTCTTC 55 C SG34L CTTTTCTTTTAGTTTTTGGTTT 55 C PCR reakce pak probíhá v objemu 20 µl, v 1x reakčním pufru PPP (Top Bio), 1x BSA, 0,5 µm každého primeru (IDT) a 5-10 ng templátové DNA. Schéma pipetování systém PPP MM Top-Bio: 10 µl PPP master mixu 1 µl DNA 1 µl každého primeru (10 µm); či 0,5 µl v případě duplexové PCR 2 µl 10xBSA 5 µl dh 2 O (voda do objemu 20 µl) Amplifikace probíhá na MJ Research Thermocycler PTC 100, nebo Bioer XP Cycler při následujícím teplotním profilu pro primery RsPPRB-F/R: počáteční denaturace 5 min 95 C 35 cyklů: 30 s 95 C 45 s 59 C 2 min 72 C konečná elongace 10 min 72 C stop 4 C 12

, 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA")
19 Teplotní profil pro primery SG34U a SG34L zahrnoval: počáteční denaturace 2,5 min 94 C 30 cyklů: 30 s 94 C 1 min 55 C 2 min 72 C konečná elongace 5 min 72 C stop 4 C PCR produkty se rozdělují na 1,5% agarózovém gelu v 1x TBE pufru. Podmínky separace: 30 V 20min, 90 V 1hod a 30min. Jako marker se používá 100 bp DNA ladder (NEB) a DNA fragmenty se vizualizují barvením pomocí ethidium bromidu pod UV světlem. Chemikálie: PPP Master Mix (2x koncentrovaný: 150 mm Tris-HCl, ph 8,8 (při 25 C), 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA polymerázy, barvivo, stabilizátory a aditiva) templátová DNA primery dh 2 O (PCR dh 2 O, nebo Millipore dh 2 O) Přístroje: PCR thermocykler centrifuga, sada automatických pipet, mrazák Detekce genu Rf u rostlin řepky pomocí primeru RsPPRB-F/R (vlevo) a G34U a SG34L (vpravo). 102 A115 výchozí CMS 103 B115 udržovatel sterility /2 obnovitel fertility /3 obnovitel fertility 13
20 Detekce rostlin se sterilní cytoplazmou u materiálu CMS typu Shaan 2A Detekce je založena na PCR detekci přítomnosti příslušné sekvence u analyzovaných rostlin a na PCR-RFLP detekci rozdílných fragmentů u primerového páru S2A178F/R. Pro PCR analýzu byly pro účely předkládané metodiky použity primerové páry S2A178F/S2A178R a S2A1.3F/S2A1.3R. Sekvence primerů používaných pro PCR analýzu. Primer Sekvence 5 3 Annealing S2A178F TGGAGTACTTGGGATCAGCA 60 S2A178R ATCTCGCAGACGATCAAGGT 60 S2A1.3F AGTGGACCGGGTAGTGCTTA 57 S2A1.3R CCTCCAGACAGCTTCACTCC 57 PCR reakce pak probíhá v objemu 25 µl, v 1x reakčním pufru (PPP MM; Top-Bio), 12,5 pm primeru (IDT) a 10 ng templátové DNA. Schéma pipetování systém PPP MM Top-Bio: 12,5 µl PPP master mixu 1 µl DNA 1,25 µl každého primeru (10 µm) 9 µl dh 2 O (voda do objemu 25 µl) Amplifikace probíhá na MJ Research Thermocycler PTC 100, nebo Bioer XP Cycler při následujícím teplotním profilu: počáteční denaturace 3 min 94 C 35 cyklů: 30 s 94 C 30 60/57 C 1,5 min 72 C konečná elongace 10 min 72 C stop 4 C PCR produkty se rozdělují na 1,5% agarózovém gelu v 1x TBE pufru. Podmínky separace: 30 V 20min, 90 V 1hod a 30min. Jako marker se používá 100 bp DNA ladder (NEB) a DNA fragmenty se vizualizují barvením pomocí ethidium bromidu pod UV světlem. 14
21 Chemikálie: PPP Master Mix (2x koncentrovaný: 150 mm Tris-HCl, ph 8,8 (při 25 C), 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA polymerázy, barvivo, stabilizátory a aditiva) templátová DNA primery dh 2 O (PCR dh 2 O, nebo Millipore dh 2 O) Přístroje: PCR thermocykler centrifuga, sada automatických pipet, mrazák S použitím primerů S2A1.3F a S2A1.3R (amplifikujících nekódující úsek mtdna, lokalizovaný v horní části řetězce před vlastním orf224-1), byla amplifikací fragmentů o velikosti přibližně 1300bp detekována CMS mitochondriální sekvence přítomná v CMS rostlinách, obnovitelích fertility (odvozených z CMS) a u odvozených F 1 hybridů. U udržovatelů sterility nedocházelo k amplifikaci této sekvence, jelikož mají jako jediní fertilní cytoplazmu. Specifické primery S2A178F a S2A178F amplifikující část mitochondriálního genu orf224-1 (zodpovědného za CMS) vedly k detekci velikostně podobné sekvence i u udržovatelů sterility. Na základě výrazných odlišností v nefunkční sekvenci udržovatelů je však možné její nositele identifikovat použitím restrikčních enzymů v našem případě byla použita restrikční endonukleáza MseI. 15
22 Shaan 2A trnm Orf224-1 atp6 S2A1.3F/R Shaan 2A trnm Orf224-1 atp6 S2A178F/R Sekvenační analýza MseI PCR-RFLP CMS - CMS linie Shaan 2A mcms - udržovatel sterility Rf F 1 - obnovitel fertility - F 1 hybrid 16
23 Detekce Rf u CMS typu Shaan 2A Detekce je založena na PCR detekci přítomnosti fragmentů. Pro PCR analýzu byly pro účely předkládané metodiky použity primerové páry 3F a 4R, 14o1 a 14o4 (Formanová et al. 2010) a SSR primery KBrDP1F/R (Li et al. 2011), amplifikující oblast DNA, která je ve vazbě s genem pro obnovu fertility u CMS systému Polima a dále nově vyvinuté primery Rf84F/R. Sekvence primerů používaných pro PCR analýzu. Primer Sekvence 5 3 Annealing 3F GTTCTTACAGTGGCGTAAAGATGAG 57 C 4R AAGCTTCCACAGGTCCAGAGTAC 57 C Rf84F TGCCATTGTAAGAACTAAGAAGAGG 57 C Rf84R GCTTCTTAGTTCAACGATGGC 57 C 14o1 CTTCTTCGACAGAGAAACGTG 57 C 14o4 CAGAAGCTGATTCCTGATGATG 57 C KBrDP1F CACTCTTTTAGGAGAAGCACGAA 55 C KBrDP1R CCCGCATAGTTTGTCAGTTTCT 55 C PCR reakce pak probíhá v objemu 20 µl, v 1x reakčním pufru (PPP MM; Top-Bio), s optimální koncentrací primeru (IDT) a ~25-70 ng templátové DNA. Schéma pipetování systém PPP MM Top-Bio: 10 µl PPP master mixu 1 µl DNA (~25-70 ng) x µl každého primeru (0,5µM pro 3F/R, Rf84F/R a 14o4; 0,3 µm pro KBrDP1) x µl dh 2 O (voda do objemu 20 µl) Amplifikace probíhá na MJ Research Thermocycler PTC 100, nebo Bioer XP Cycler při následujícím teplotním profilu pro 3F/4R, Rf84F/R a 14o1/4: počáteční denaturace 5 min 94 C 30 s 57 C 1,5 min 72 C 35 cyklů: 30 s 94 C 30 s 57 C 1,5 min 72 C konečná elongace 10 min 72 C stop 4 C 17
24 Pro primery KBrDP1: počáteční denaturace 5 min 94 C 10 cyklů 40 s 94 C 45 s 60 C -0,5 C cyklus 45 s 72 C 30 cyklů: 40 s 94 C 45 s 55 C 45 s 72 C konečná elongace 10 min 72 C stop 4 C PCR produkty se rozdělují na 1,5% agarózovém gelu v 1x TBE pufru. Podmínky separace: 30 V 30min, 90 V 1hod a 30min. Jako marker se používá 100 bp DNA ladder (NEB) a DNA fragmenty se vizualizují barvením pomocí ethidium bromidu pod UV světlem. Chemikálie: PPP Master Mix (2x koncentrovaný: 150 mm Tris-HCl, ph 8,8 (při 25 C), 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA polymerázy, barvivo, stabilizátory a aditiva) templátová DNA primery dh 2 O (PCR dh 2 O, nebo Millipore dh 2 O) Přístroje: PCR thermocykler centrifuga, sada automatických pipet, mrazák Primerový pár 3F a 4R detekoval CMS rostliny společně s udržovateli sterility přítomností amplikonů o velikosti přibližně 500 bp a obnovitele fertility přítomností amplikonů přibližně 440 bp. V F1 generaci byly detekovány oba fragmenty. Sekvenační analýza těchto fragmentů prokázala deleci úseku o velikosti 62bp (oblast začínající od 140 nukleotidu forward primeru) u všech nositelů genu pro obnovu fertility. Na základě této analýzy byly navrženy nové primery Rf84F/R, amplifikující 164bp úsek u sterilních rostlin, udržovatelů a F 1 rostlin, zatímco 84bp fragment byl přítomen pouze u homozygotních nositelů Rf genu. 18
25 SCAR marker - primery4r/3f (Formanová et al. 2010) Na základě získané sekvence byl vyvinut nový molekulární marker k detekci Rf primeryrf84f/r Primerový pár 14o1 a 14o4 amplifikoval fragment o velikosti přibližně 700 bp u všech rostlin s výjimkou obnovitelů fertility. Detekce Rf 650 bp- SCAR marker primery 14oF/R (Formanová etal. 2010) Mikrosatelitový primerový pár KBrDP1 detekuje CMS rostliny a udržovatele sterility přítomností fragmentu o velikosti ~280 bp, zatímco u obnovitelů fertility a F 1 hybridů se amplifikuje druhý fragment o velikosti ~300 bp. PrimerKBrDP1 (Li et al., 2011) Rf udržovatel sterility pro S2 (DH) F1 F 1 hybrid CMS linie Shaan 2A mcms udržovatel sterility 19
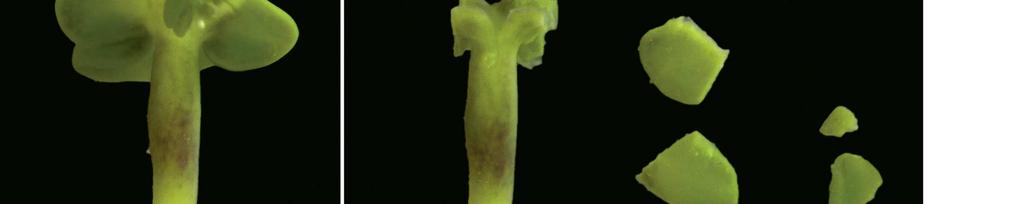
26 Detekce Rfo genu v mikrosporových embryích řepky u CMS systému Ogu-INRA Embrya byla získaná z pylových zrn F 1 rostlin jedné kombinace křížení, byla rozdělena do třech velikostních frakcí A 3,5mm; B 2,75-3,49 mm a C 2-2,74 mm. Detekce nositelů genu pro obnovu fertility byla provedena nedestruktivní metodou; odebráním kotyledonárního pletiva a následnou PCR detekcí se specifickými primery pro úsek genu PPR-B. Technika mikrosporových kultur a diploidizace pomocí trifluralinu v mikrosporové suspenzi byla provedena dle dříve publikovaného protokolu (Klíma et al. 2008). Za účelem genotypizace byly z každého embrya nedestruktivně (bez poškození apikálního meristému) v aseptických podmínkách odebrána kotyledonární pletiva k extrakci DNA. Embryo bylo přeneseno do regeneračního media (RM; Klíma et al. 2004). Nedostatečně veliká embrya nevhodná k nedestruktivnímu kotyledonárnímu odběru byla nejprve kultivována na DM médiu po dobu 2-3 týdnů. Následná kultivace embryí byla provedena dle protokolu Klíma et al. (2008) za účelem dosažení kořenících regenerantů schopných přenosu do půdy a získání osiva dihaploidních rostlin. DNA byla izolována dle protokolu Edwards et al. (1991) a detekce genu Rfo byla provedena pomocí specifických primerů RsPPRB-F/RsPPRB-R. Druhý použitý primerový pár byl použit jako pozitivní kontrola sloužící k amplifikaci vždy přítomného úseku B. napus odvozeného z AFLP markeru. Sekvence primerů používaných pro PCR analýzu. Primer Sekvence 5 3 Annealing RsPPRB-F GAAGCTCTTGCTACCCATCG 59 C RsPPRB-R TGACATGCTTCGATCTCGTC 59 C 208F TCGGGATGGAAACCATACTC 59 C 208R GGTCCCAGATAAGGGGAAAA 59 C Duplex PCR byla provedena v objemu 20 µl, tvořeného 1x reakčním pufrem PPP (Top Bio), 1x BSA, 0,25 µm každého primeru (IDT) a 5-10 ng templátové DNA. Schéma pipetování systém PPP MM Top-Bio: 10 µl PPP master mixu 1 µl DNA 0,5 µl každého primeru (10 µm) 2 µl 10xBSA 5 µl dh 2 O (voda do objemu 20 µl) 20

27 Amplifikace probíhá na MJ Research Thermocycler PTC 100, nebo Bioer XP Cycler při následujícím teplotním profilu: počáteční denaturace 5 min 95 C 35 cyklů: 30 s 95 C 45 s 59 C 2 min 72 C konečná elongace 10 min 72 C stop 4 C PCR produkty se rozdělují na 1,5% agarózovém gelu v 1x TBE pufru. Podmínky separace: 30 V 30min, 90 V 1hod a 30min. Jako marker se používá 100 bp DNA ladder (NEB) a DNA fragmenty se vizualizují barvením pomocí ethidium bromidu pod UV světlem. Chemikálie: PPP Master Mix (2x koncentrovaný: 150 mm Tris-HCl, ph 8,8 (při 25 C), 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA polymerázy, barvivo, stabilizátory a aditiva) templátová DNA primery dh 2 O (PCR dh 2 O, nebo Millipore dh 2 O) Přístroje: PCR thermocykler centrifuga, sada automatických pipet, mrazák 1-3 a 6 embrya nesoucí gen pro obnovu fertility 4-5 embrya bez genu pro obnovu fertility 21










28 22
29 Detekce autoinkompatibilních haplotypů detekce a selekce AI rostlin DNA byla izolována dle CTAB-PVP a detekce AI haplotypů byla provedena pomocí specifických primerů SLGa (Zhang et al. 2008b), které amplifikují cca 1 kb dlouhý fragment u AK rostlin. U AI rostlin, které mají funkční S haplotyp tento fragment není amplifikován. Jako vnitřní kontrola byl použit primerový pár SCO3 (Zeng et al. 2009), který amplifikuje fragment o délce 750 bp u všech rostlin (pozitivní kontrola správného průběhu PCR reakce). Sekvence primerů používaných pro PCR analýzu. Primer Sekvence 5 3 Annealing SLGa_F GATTCTACTTCCTCCTGCCT 58 C SLGa_R ATTACAGTCGCTAAGGCACC 58 C SCO3_F TGATGCAGGAACCTTGAAGC 58 C SCO3_R GAGCATCTTGCATGGAGGTC 58 C Duplex PCR byla provedena v objemu 20 µl, tvořeného 1x reakčním pufrem PPP (Top Bio), 1x BSA, 0,25 µm každého primeru (IDT) a 10 ng templátové DNA. Schéma pipetování systém PPP MM Top-Bio: 10 µl PPP master mixu 1 µl DNA 0,5 µl každého primeru (10 µm) 2 µl 10xBSA 5 µl dh 2 O (voda do objemu 20 µl) Amplifikace probíhá na MJ Research Thermocycler PTC 100, nebo Bioer XP Cycler při následujícím teplotním profilu: počáteční denaturace 3 min 95 C 35 cyklů: 30 s 94 C 45 s 58 C 1 min 72 C konečná elongace 10 min 72 C stop 4 C PCR produkty se rozdělují na 1,5% agarózovém gelu v 1x TBE pufru. Podmínky separace: 30 V 30min, 90 V 1hod a 30min. Jako marker se používá 100 bp DNA ladder (NEB) a DNA fragmenty se vizualizují barvením pomocí ethidium bromidu pod UV světlem. 23
30 Chemikálie: PPP Master Mix (2x koncentrovaný: 150 mm Tris-HCl, ph 8,8 (při 25 C), 40 mm (NH4)2SO4, 0,02% Tween 20, 5 mm MgCl2, 400 µm datp, 400 µm dctp, 400 µm dgtp, 400 µm dttp, 100 U/ml Taq DNA polymerázy, barvivo, stabilizátory a aditiva) templátová DNA primery dh 2 O (PCR dh 2 O, nebo Millipore dh 2 O) Přístroje: PCR thermocykler centrifuga, sada automatických pipet, mrazák 24
31 Schéma křížení donorů AI a donorů kvality (funkčních a nefunkčních S haplotypů) a vyštěpování haplotypů v populaci dihaploidních rostlin. Schéma selekce AI rostlin s funkčním S haplotypem. Duplex PCR s primery specifickými pro SLG-I, amplifikace ~1.1 kb fragmentu u AK rostlin, u AI rostlin nedochází k amplifikaci tohoto fragmentu. U všech rostlin pak dochází k amplifikaci 750 bp kontrolního fragmentu. Oba obrázky převzaty z Havlíčková et al. (2014). 25
32 26
33 III. Srovnání novosti postupů Předkládaná Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění je aktuální a nová, protože v současné době není v ČR k dispozici dostupná metodika zabývající se touto problematikou. Dosud dostupné informace jsou jen dílčí a rozptýlené ve vědeckých publikacích a monografiích, které se zabývají problematikou molekulárních markerů, komplexní zpracování selekčních markerů pak dostupné není. Molekulární markery představují ve srovnání s morfologickými či biochemickými markery kvalitativně nový přístup, který má na jedné straně obrovský potenciál využití, avšak na straně druhé i své limity. Reálná interpretace molekulárních dat obvykle vyžaduje kvalifikovanou volbu vhodných a optimalizovaných postupů. Využití analýzy molekulárních markerů pro popis a charakterizaci genotypů je předmětem předkládané metodiky. IV. Popis uplatnění metodiky Využití molekulárních technik při selekci rodičovských komponent v programech hybridního šlechtění v první části zahrnuje teoretický úvod do problematiky. V praktické části pak jsou uvedeny postupy izolace DNA a protokoly pro PCR detekci jednotlivých genů, resp. cílových genotypů. Metodika představuje soubor optimalizovaných návodů molekulární analýzy, na jejichž základě lze provádět rutinní analýzy DNA markerů u řepky s cílem selekce požadovaných genotypů. Výstupem analýzy je pak PCR produkt odpovídající detekované sekvenci. Molekulární markery nejsou dosud standardně pro účely selekce požadovaných genotypů používány, mohou být ale vhodným doplňkem klasických šlechtitelských selekčních postupů a oproti nim skýtají značné výhody. Uživatelé metodiky jsou pracoviště výzkumná a šlechtitelská, která mohou s výhodou využít předností analýzy molekulárních markerů rychlost analýzy, vysoká míra polymorfismu a nízké ovlivnění faktory vnitřního a vnějšího prostředí. Metodika bude uplatněna prostřednictvím šlechtitelské firmy OSEVA PRO s.r.o. S tímto subjektem byla uzavřena smlouva o uplatnění metodiky. 27
34 28
35 V. Ekonomické aspekty Předpokládané ekonomické přínosy metodiky jsou kalkulovány až do výše tis. Kč. Dalšími přínosy předkládané metodiky jsou rozšíření spektra technik a metodických postupů používaných v biotechnologické laboratoři, rozšíření portfolia technik a služeb prováděných v laboratoři, metodická a vzdělávací funkce. Přínosem pro šlechtění rostlin je úspora nákladů v procesu šlechtění zkrácení šlechtitelského procesu o 1 rok lze při celkových nákladech na jednu odrůdu a délce šlechtění (7-10 let) uspořit 800 tis. Kč, při vzniku nové odrůdy (s využitím molekulární selekce) by finanční přínos pro uživatele (šlechtitele) plynul z licenčního poplatku (nová odrůda by mohla být zaseta na 2500 ha po dobu 4 let s výnosem 2,8 t/ha, licenční poplatek je 60 Kč z 1 kg prodaného osiva, na 1 ha jsou zapotřebí asi 4 kg, takže 2500*60*4 = 600 tis Kč/rok x 4 roky= celkem finanční přínos je 2,4 mil. Kč. Do nákladů na zavedení postupů uvedených v metodice lze započítat pořízení nejnutnějšího investičního a neinvestičního vybavení pro provoz laboratoře a provedení úkonů a postupů uváděných v metodice v celkové minimální výši 1,100 tis. Kč. V případě automatizace izolace DNA a zpracování velkého objemu vzorků je pak nutné počítat s dalšími náklady na pořízení linky pro izolaci DNA pomocí paramagnetických partikulí v objemu cca 2,5 mil Kč a automatické čipové elektroforézy nebo fragmentačního analyzátoru ve výši 1,4 mil Kč. Další náklady pak představují náklady na chemikálie a na základě dlouhodobých kalkulací činí jednotková cena za analýzu jednoho vzorku 270 Kč (ceny za chemikálie). V uvedeném příkladu kalkulace nákladů nejsou uvedeny doplňkové náklady, odpisy, náklady na vzdělávání a vyškolení personálu laboratoře, náklady na vývoj metod, které jsou odvislé od konkrétní situace a vybavenosti (materiální i personální) pracoviště. Jako ekonomicky výhodnější se jeví analýza markerů ve specializovaných laboratořích, kde lze sdílet náklady na pořízení investic, výchovu a vzdělávání pracovníků, služby. 29
36 30
37 V. Seznam použité související literatury Bannerot H., Boulidard L., Cauderon Y., Tempe J. (1974). Transfer of cytoplasmic male sterility from Raphanus sativus to Brassica oleracea. Proc. Eucarpia Meeting Cruciferae, Dundee, Bannerot H., Boulidard L., Chupeau Y. (1977): Unexpected difficulties met with the radish cytoplasm in Brassica oleracea. Eucarpia Cruciferae Newsletter, 2: 16. Baranyk P. (2014): Stanovisko k odrůdové skladbě řepky pro rok 2014/15. Svaz pěstitelů a zpracovatelů olejnin, SPZO s.r.o. Praha. Brandle J.E., McVetty P.B.E. (1989): Heterosis and combining ability in hybrids derived from oilseed rape cultivars and inbred lines. Crop Science, 29: Budar F., Delourme R., Pelletier G. (2004): Male Sterility. In: Pua E.C., Douglas C.J. (eds.): Biotechnology in Agriculture and Forestry, Vol 54. Brassica. Springler-Verlag, pp Černý J., Šašek A. (1996): Bílkovinné signální geny pšenice obecné. ÚZPI, Praha. Čurn V. Žaludová (2007). Fingerprinting of Oilseed Rape Cultivars. In: Gupta S. (ed.): Rapeseed Breeding. (Advances in Botanical Research, Volume 45). Elsevier Publ., pp Čurn V., Sáková L. (1997). Využití biochemických markerů ve šlechtění řepky a dalších brukvovitých plodin. Genetika a šlechtění, 33: Čurn V., Sáková L., Graman J. (1995): Elektroforéza isoenzymů a neenzymatických bílkovin - metody a aplikace v genetice a šlechtění rostlin. Genetika a Šlechtění, 31: De Nettancourt D. (1977): Incompatibility in Angiosperms. Springer-Verlag, Berlin, Heidelberg, New York. Edwards K., Jihnstone C., Thompson C. (1991). A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res., 19: Formanová N., Stollar R., Gedd R., Mehé L., Laforest M., Landry B.S., and Brown G.G. (2010). High-resolution mapping of the Brassica napus Rfp restorer locus using Arabidopsis-derived molecular markers. Theor. Appl. Genet., 120: Havlíčková L., Jozová E., Klíma M., Kučera V., Čurn V. (2014): Detection of selfincompatible oilseed rape plants (Brassica napus L.) based on molecular markers for identification of the class I S haplotype. Genetics and Molecular Biology, 37: Klíma M., Vyvadilová M., Kučera V. (2008): Chromosome doubling effects of selected antimitotic agents in Brassica napus microspore culture. Czech J. Genet. Plant Breed., 42: Koprna R., Macháčková I., Horáček J., Ehrenbergerová J. 2007): Utilization of Ogu-INRA hybrid system in winter rapeseed breeding. MendelNet 07 Agro. Koprna, R., Kučera, V., Macháčková, I., Horáček, J., Ehrenbergerová, J. (2009): Development of fertility restorers of winter oilseed rape with low glucosinolate content for the CMS Ogu-INRA system. Czech Journal of Genetics and Plant Breeding, 45:
38 Kudla M. (1996): General combining ability of inbred lines and heterosis effect of winter oilseed rape F1 and F2 hybrids. In: Rośliny Oleiste, 1996, 17: Li Y., Liu Z., Cai Q., Yang G., He Q., and Lu P. (2011): Identification of a microsatellite marker linked to the fertility-restoring gene for a polima cytoplasmic male-sterile line in Brassica napus L. African Journal of biotechnology, 10: Menczel L., Morgan A., Brown S., Maliga P. (1987):Fusion-mediated combination of Ogura-type cytoplasmic male sterility with Brassica napus plastids using X-irradiated CMS protoplasts. Plant Cell Rep., 6: Murray H.G., Thompson W.F. (1980): Rapid isolation of high molecular weight DNA. Nucleilc Acids Res., 8: Nasrallah J.B., Nasrallah M.E. (1993): Pollen-stigma signaling in the sporophytic self-incompatibility response. Plant Cell, 5: Nasrallah J.B., Nishio T., Nasrallah M.E. (1991): The self-incompatibility genes of Brassica: Expression and use in genetic ablation of floral tissues. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: Newell M., Jannink J. (2013): Genomic selection in plant breeding. In: Fleury D., Whitford R. (eds.): Crop Breeding: Methods and Protocols. Humana Press, Springer New York, Heidelberg, Dordrecht, London. pp Ockendon D.J. (1974): Distribution of self-incompatibility allels and breeding structure of open-pollinated cultivars of Brussel sprounts. Heredity, 33: Ogura H. (1968): Studies on a new male-sterility in Japanese radish with special reference to utilisation of this sterility towards the practical raising of hybrid seeds. Mem. Fac. Agri Kagoshima Univ., 6: Okamoto S., Odashima M., Fujimoto R., Sato Y., Kitashiba H., Nishio T. (2007): Self-compatibility in Brassica napus is caused by independent mutations in S-locus genes. Plant J., 50: Paulmann W. (1999): Pokrok ve šlechtění hybridní řepky a pěstování MSL hybridních odrůd. Sborník, Seminář Svazu pěstitelů a zpracovatelů olejnin , Hluk, pp Pelletier G., Primard C., Vedel F., Chetrit P., Remy R., Rouselle P., Renard M. (1983): Intergeneric cytoplasmatic hybridization in Cruciferae by protoplast fusion. Molecular Genetics, 191: Pelletier G., Primard C., Vedel F., Chétrit P., Renard M., Delourme R. (1987): Molecular, phenotypic and genetic characterization of mitochondrial recombinants in rapeseed. Proceedings from 7th International Rapeseed Congress, Vol.I., Poznań, 1987, pp Primard-Brisset C., Poupard J.P., Horvais R., Eber F., Pelletier G., Renard M., Delourme R. (2005): A new recombined double low restorer line for the Ogu- INRA cms in rapeseed (Brassica napus L.). Theor Appl Genet, 111: Röbbelen G. (1985): Züchtung von Hybridraps. Bericht der Arbeitstagung Saatzuchtleiter, Gumpenstein, pp Sozinov A.A. (1985): Protein polymorphism and its importance in genetics, breeding and evolution. - In: Sozinov A.A. (ed.): Molekulyarnye Mekhanizmy Geneticheskikh Protsessov. Molekulyarnaya Genetika, Evolyutsiya i Molekulyarno-Geneticheskie Osnovy Selektsii. Nauka, Moscow, pp
39 Varshney R.K., Hoisington D.A., Nayak S.N., Graner A. (2009): Molecular plant breeding: methodology and achievements. Methods Mol. Biol., 513: Varshney R.K., Tuberosa R. (Eds.) (2007): Genomics-Assisted Crop Improvement. Vol 1 and 2. Springer, Dordrecht. Vyvadilová M., Klíma M., Kučera V. (2008): Metodika produkce dihaploidních linií pro šlechtění řepky ozimé. Výzkumný ústav rostlinné výroby, v.v.i., Praha. Zehnálek P., Holubář J. (2000): Přehled odrůd olejnin a kmínu ÚKZÚZ Brno, ISBN Zehnálek P., Holubář J. (2004): Přehled odrůd olejnin a kmínu ÚKZÚZ Brno, ISBN Zehnálek P., Holubář J., Mezlík T. (2005): Přehled odrůd olejnin a kmínu ÚKZÚZ Brno, ISBN Zehnálek P., Holubář J., Mezlík T. (2005): Přehled odrůd olejnin a kmínu ÚKZÚZ Brno, ISBN Zehnálek P., Holubář J., Mezlík T. (2006): Seznam doporučených odrůd 2006: Řepka olejka. ÚKZÚZ Brno, ISBN Zeng F., Yi B., Tu J., Fu T. (2009): Identification of AFLP and SCAR markers linked to the male fertility restorer gene of pol CMS (Brassica napus L.). Euphytica, 165: Zhang X., Ma C., Fu T., Li Y., Wang T., Chen O., Tu J., Shen J. (2008b): Development of SCAR markers linked to selfincompatibility in Brassica napus L. Mol. Breed., 21: Zhang X., Ma C., Tang J., Tang W., Tu J., Shen J., Fu T. (2008a): Distribution of S haplotypes and its relationship with restorer-maintainers of selfincompatibility in cultivated Brassica napus. Theor. Appl. Genet., 117:
40 34
41 VI. Seznam publikací, které předcházely metodice Havlíčková L., Jozová E., Klíma M. Kučera V., Čurn V. (2014): Detection of selfincompatible oilseed rape plants (Brassica napus L.) based on molecular markers for identification of the class I S haplotype. Genetics and molecular biology, 37: Žaludová J., Havlíčková L., Jozová E., Kučera V., Vyvadilová M., Klíma M., Čurn V. (2013): Marker assisted selection as a tool for detection of Brassica napus plants carrying self-incompatibility alleles, in hybrid breeding programs. Romanian Agricultural Research, 30: Havlíčková L., Čurn V., Jozová E., Kučera V., Vyvadilová M., Klíma M. (2012): Sequence analysis of the mtdna region correlated with Shaan 2A cytoplasmic male sterility in rapeseed (Brassica napus L.). Czech J. Genet. Plant Breed. 48: Čurn V., Nováková A., Šimáčková K., Ondřichová B., Bárta J. (2008): Molecular markers as a tool for plant breeding and variety identification. Prohnes J., Badenes M.L.(eds): Modern variety breeding for present and future needs. 18th Eucarpia General Congress, Valencia, 2008, p Kukolikova B., Curn V., Koprna R., Novakova A., Simackova K. (2008): The comparison of polymorphism and expressin of SLG gene in the varieties and SI/SC lines of oilseed rape. XX Int. Congress Genetics, Berlin, July 12-17, 2008, p. 76. Čurn V., Žaludová (2007): Fingerprinting of Oilseed Rape Cultivars. In: Gupta S. (ed.): Rapeseed Breeding. (Advances in Botanical Research, Volume 45). Elsevier Publ., pp Kukolikova B., Zaludova J., Curn V., Koprna R. (2007): Efficiency of marker assisted selection of self-incompatible oilseed rape varieties. - 6th Plant Genomics European Meetings, Tenerife, 3-6 October 2007, p Žaludová J., Kukolíková B., Čurn V. (2007): Marker-assisted selection of selfincompatible oilseed rape plants. Proc. 12th Int. Rapeseed Congress, Wuhan, China, Sciense Press USA, pp Žaludová J., Kukolíková B., Čurn V. (2006): Distribution of S-alleles in Brassica napus. - Proc. 28 th Sci. Conf. Oilseed Crops, Poznan, , p Čurn V. (2005): Molekulární markery - protokoly a návody pro cvičení. Biotechnologické centrum ZF JU, Č. Budějovice. Hájková P., Hrubý J., Pernová E., Čurn V., Žaludová J. (2005): Monitorig pěstebních ploch, přenos a detekce transgenů geneticky modifikované řepky olejky (Rassica napus L. var. napus), Sborník vědeckých prací VÚP Troubsko 15: Kubátová B., Čurn V. (2005): Soubor metodik a laboratorních protokolů. BC ZF JU a Laboratoř molekulární biologie rostlin KB BF JU, Č. Budějovice. 35
Metodika tvorby rodičovských komponent a hybridů ozimé řepky (Brassica napus L.) na bázi CMS
 na bázi CMS") Metodika tvorby rodičovských komponent a hybridů ozimé řepky (Brassica napus L.) na bázi CMS Metodika byla vypracována jako výstup výzkumného projektu MZe QI 111A075 Využití biotechnologických metod, nových
Metodika tvorby rodičovských komponent a hybridů ozimé řepky (Brassica napus L.) na bázi CMS Metodika byla vypracována jako výstup výzkumného projektu MZe QI 111A075 Využití biotechnologických metod, nových
UTILIZATION OF OGU-INRA HYBRID SYSTEM IN WINTER RAPESEED BREEDING VYUŽITÍ HYBRIDNÍHO SYSTÉMU OGU-INRA PŘI ŠLECHTĚNÍ ŘEPKY OZIMÉ
 UTILIZATION OF OGU-INRA HYBRID SYSTEM IN WINTER RAPESEED BREEDING VYUŽITÍ HYBRIDNÍHO SYSTÉMU OGU-INRA PŘI ŠLECHTĚNÍ ŘEPKY OZIMÉ Koprna R. 1, Macháčková I. 2, Horáček J. 3, Ehrenbergerová J. 4 1 OSEVA PRO
UTILIZATION OF OGU-INRA HYBRID SYSTEM IN WINTER RAPESEED BREEDING VYUŽITÍ HYBRIDNÍHO SYSTÉMU OGU-INRA PŘI ŠLECHTĚNÍ ŘEPKY OZIMÉ Koprna R. 1, Macháčková I. 2, Horáček J. 3, Ehrenbergerová J. 4 1 OSEVA PRO
Metodika detekce a molekulární selekce autoinkompatibilních linií řepky (Brassica napus L.)
") Metodika detekce a molekulární selekce autoinkompatibilních linií řepky (Brassica napus L.) Metodika byla vypracovaná jako výstup výzkumného záměru MSM 6007665806 Trvale udržitelné způsoby zemědělského
Metodika detekce a molekulární selekce autoinkompatibilních linií řepky (Brassica napus L.) Metodika byla vypracovaná jako výstup výzkumného záměru MSM 6007665806 Trvale udržitelné způsoby zemědělského
Metodika analýzy molekulárních markerů u jilmu, Ulmus L.
 Metodika analýzy molekulárních markerů u jilmu, Ulmus L. Metodika byla vypracovaná jako výstup projektu NAZV QI92A247 Charakterizace genetické struktury autochtonních populací jilmů pomocí DNA analýz,
Metodika analýzy molekulárních markerů u jilmu, Ulmus L. Metodika byla vypracovaná jako výstup projektu NAZV QI92A247 Charakterizace genetické struktury autochtonních populací jilmů pomocí DNA analýz,
Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genetických zdrojů řepky (Brassica napus L.) a hodnocení jejich diverzity
 a hodnocení jejich diverzity") Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genetických zdrojů řepky (Brassica napus L.) a hodnocení jejich diverzity Metodika byla vypracovaná jako výstup výzkumného projektu
Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genetických zdrojů řepky (Brassica napus L.) a hodnocení jejich diverzity Metodika byla vypracovaná jako výstup výzkumného projektu
Metodika časné detekce obnovitelů fertility pro CMS Ogu-INRA v mikrosporových embryích řepky olejky
 Metodika časné detekce obnovitelů fertility pro CMS Ogu-INRA v mikrosporových embryích řepky olejky Metodika byla vypracována jako výstup výzkumného projektu NAZV QJ 1510172 Využití nekonvenčních výchozích
Metodika časné detekce obnovitelů fertility pro CMS Ogu-INRA v mikrosporových embryích řepky olejky Metodika byla vypracována jako výstup výzkumného projektu NAZV QJ 1510172 Využití nekonvenčních výchozích
AKTUÁLNÍ POZNATKY V PĚSTOVÁNÍ, ŠLECHTĚNÍ, OCHRANĚ ROSTLIN A ZPRACOVÁNÍ PRODUKTŮ
 Výzkumný ústav pícninářský, spol. s r.o. Troubsko Zemědělský výzkum, spol. s r.o. Troubsko pod záštitou MZe ČR AKTUÁLNÍ POZNATKY V PĚSTOVÁNÍ, ŠLECHTĚNÍ, OCHRANĚ ROSTLIN A ZPRACOVÁNÍ PRODUKTŮ mediální partner
Výzkumný ústav pícninářský, spol. s r.o. Troubsko Zemědělský výzkum, spol. s r.o. Troubsko pod záštitou MZe ČR AKTUÁLNÍ POZNATKY V PĚSTOVÁNÍ, ŠLECHTĚNÍ, OCHRANĚ ROSTLIN A ZPRACOVÁNÍ PRODUKTŮ mediální partner
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Jednotné pracovní postupy zkoušení krmiv
 Národní referenční laboratoř Strana KVANTITATIVNÍ STANOVENÍ GENETICKÝCH MODIFIKACÍ METODOU qpcr POMOCÍ ROTOR-GENE PROBE PCR KITU Účel a rozsah Postup slouží ke kvantitativnímu stanovení genetických modifikací
Národní referenční laboratoř Strana KVANTITATIVNÍ STANOVENÍ GENETICKÝCH MODIFIKACÍ METODOU qpcr POMOCÍ ROTOR-GENE PROBE PCR KITU Účel a rozsah Postup slouží ke kvantitativnímu stanovení genetických modifikací
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KOLOREKTÁLNÍHO KARCINOMU Jednou z nejvhodnějších metod pro detekci minimální
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIDUÁLNÍ CHOROBY U KARCINOMU PANKREATU Jednou z nejvhodnějších metod pro detekci minimální reziduální
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Izolace genomové DNA ze savčích buněk, stanovení koncentrace DNA pomocí absorpční spektrofotometrie
 Izolace genomové DNA ze savčích buněk, stanovení koncentrace DNA pomocí absorpční spektrofotometrie IZOLACE GENOMOVÉ DNA Deoxyribonukleová kyselina (DNA) představuje základní genetický materiál většiny
Izolace genomové DNA ze savčích buněk, stanovení koncentrace DNA pomocí absorpční spektrofotometrie IZOLACE GENOMOVÉ DNA Deoxyribonukleová kyselina (DNA) představuje základní genetický materiál většiny
KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ
 KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ září 2014 KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ prof. Ing. Vladislav Čurn, Ph.D. Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta, Katedra speciální produkce
KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ září 2014 KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ prof. Ing. Vladislav Čurn, Ph.D. Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta, Katedra speciální produkce
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
ZPRÁVA ZA DÍLČÍ VÝSLEDKY ŘEŠENÍ VÝZKUMNÉHO PROGRAMU 3.d ZA ROK 2014
 ZPRÁVA ZA DÍLČÍ VÝSLEDKY ŘEŠENÍ VÝZKUMNÉHO PROGRAMU 3.d ZA ROK 2014 1. TITULNÍ LIST Podpora tvorby rostlinných genotypů s vysokou rezistencí k biotickým i abiotickým faktorům a diferencovanou kvalitou
ZPRÁVA ZA DÍLČÍ VÝSLEDKY ŘEŠENÍ VÝZKUMNÉHO PROGRAMU 3.d ZA ROK 2014 1. TITULNÍ LIST Podpora tvorby rostlinných genotypů s vysokou rezistencí k biotickým i abiotickým faktorům a diferencovanou kvalitou
ISSR (Inter simple sequence repeat) Jiří Košnar
 Jiří Košnar") ISSR (Inter simple sequence repeat) Jiří Košnar Přírodovědecká fakulta Jihočeské univerzity v Č. Budějovicích HISTORIE relativně nová metoda: Zietkiewicz E., Rafalski A., Labuda D. (1994): Genome fingerprinting
ISSR (Inter simple sequence repeat) Jiří Košnar Přírodovědecká fakulta Jihočeské univerzity v Č. Budějovicích HISTORIE relativně nová metoda: Zietkiewicz E., Rafalski A., Labuda D. (1994): Genome fingerprinting
Určeno pro obecné laboratorní užití. Není určeno pro použití v diagnostických postupech. POUZE PRO POUŽITÍ IN VITRO.
 Určeno pro obecné laboratorní užití. Není určeno pro použití v diagnostických postupech. POUZE PRO POUŽITÍ IN VITRO. PCR Nucleotide Mix Předem připravený roztok ultračistých PCR deoxynukleotidů (datp,
Určeno pro obecné laboratorní užití. Není určeno pro použití v diagnostických postupech. POUZE PRO POUŽITÍ IN VITRO. PCR Nucleotide Mix Předem připravený roztok ultračistých PCR deoxynukleotidů (datp,
Konečná zpráva hodnocení různých způsobů přípravy vzorků pro AMPLICOR HPV test firmy Roche
 Konečná zpráva hodnocení různých způsobů přípravy vzorků pro AMPLICOR HPV test firmy Roche Charakteristika testu: Set AMPLICOR HPV vyráběný firmou Roche je určený pro detekci vysoko-rizikových typů lidských
Konečná zpráva hodnocení různých způsobů přípravy vzorků pro AMPLICOR HPV test firmy Roche Charakteristika testu: Set AMPLICOR HPV vyráběný firmou Roche je určený pro detekci vysoko-rizikových typů lidských
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta
 Jihočeská univerzita v Českých Budějovicích OP Vzdělávání pro konkurenceschopnost CZ.1.072.4.00/12.0045 Koordinátor: Mgr. Martin Šlachta, Ph.D. Metodik: prof. Ing. Jan Frelich, CSc. Finanční manažerka:
Jihočeská univerzita v Českých Budějovicích OP Vzdělávání pro konkurenceschopnost CZ.1.072.4.00/12.0045 Koordinátor: Mgr. Martin Šlachta, Ph.D. Metodik: prof. Ing. Jan Frelich, CSc. Finanční manažerka:
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
P1 AA BB CC DD ee ff gg hh x P2 aa bb cc dd EE FF GG HH Aa Bb Cc Dd Ee Ff Gg Hh
 Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Heteroze jev, kdy v F1 po křížení geneticky rozdílných genotypů lze pozorovat zvětšení a mohutnost orgánů, zvýšení výnosu, životnosti, ranosti, odolnosti ve srovnání s lepším rodičem = heterózní efekt
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
MagPurix Blood DNA Extraction Kit 200
 MagPurix Blood DNA Extraction Kit 200 Kat. č. ZP02001-48 Doba zpracování: 50-60 minut pro MagPurix 12S 50-70 minut pro MagPurix 24 Použití Souprava MagPurix Blood DNA Extraction Kit 200 je určena pro izolátor
MagPurix Blood DNA Extraction Kit 200 Kat. č. ZP02001-48 Doba zpracování: 50-60 minut pro MagPurix 12S 50-70 minut pro MagPurix 24 Použití Souprava MagPurix Blood DNA Extraction Kit 200 je určena pro izolátor
Kdo jsme. Centrum strukturní a funkční genomiky rostlin Ústavu experimentální botaniky AV ČR, v.v.i.
 Kdo jsme Centrum strukturní a funkční genomiky rostlin Ústavu experimentální botaniky AV ČR, v.v.i. Partner Centra regionu Haná pro biotechnologický a zemědělský výzkum v Olomouci (projekt OP VaVpI) Centrum
Kdo jsme Centrum strukturní a funkční genomiky rostlin Ústavu experimentální botaniky AV ČR, v.v.i. Partner Centra regionu Haná pro biotechnologický a zemědělský výzkum v Olomouci (projekt OP VaVpI) Centrum
Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genových zdrojů a identifikace odrůd brambor (Solanum tuberosum L.
 Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genových zdrojů a identifikace odrůd brambor (Solanum tuberosum L.) Autoři: prof. Ing. Vladislav Čurn, Ph.D., Ing. Alena Nováková,
Metodika izolace DNA a analýzy molekulárních markerů pro účely popisu genových zdrojů a identifikace odrůd brambor (Solanum tuberosum L.) Autoři: prof. Ing. Vladislav Čurn, Ph.D., Ing. Alena Nováková,
DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR
 Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Úvod IntellMed, s.r.o., Václavské náměstí 820/41, 110 00 Praha 1 DIAGNOSTICKÝ KIT PRO DETEKCI MINIMÁLNÍ REZIUDÁLNÍ CHOROBY MRD EGFR Jednou z nejvhodnějších metod pro detekci minimální reziduální choroby
Izolace RNA. doc. RNDr. Jan Vondráček, PhD..
 Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Izolace RNA doc. RNDr. Jan Vondráček, PhD.. Metodiky izolace RNA celková buněčná RNA ( total RNA) zahrnuje řadu typů RNA, které se mohou lišit svými fyzikálněchemickými vlastnostmi a tedy i nároky na jejich
Seminář izolačních technologií
 Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace praktických cvičení molekulárně-biologických předmětů o sekvenční úlohy PRACOVNÍ PROTOKOL PRO PŘEDMĚT GENETCKÁ DIVERZITA Vypracováno
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace praktických cvičení molekulárně-biologických předmětů o sekvenční úlohy PRACOVNÍ PROTOKOL PRO PŘEDMĚT GENETCKÁ DIVERZITA Vypracováno
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin
 Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Inovace laboratorních úloh genetických předmětů metodikami pracujícími s ribonukleovými kyselinami pšenice Metodické návody pro laboratorní
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 ZÁKLADNÍ GENETICKÉ POJMY Genetika je nauka o dědičnosti a proměnlivosti znaků. Znakem se
Column DNA Lego Kit UNIVERZÁLNÍ SOUPRAVY PRO RYCHLOU IZOLACI ČISTÉ DNA (Katalogové číslo D201 + D202)
") Column DNA Lego Kit UNIVERZÁLNÍ SOUPRAVY PRO RYCHLOU IZOLACI ČISTÉ DNA (Katalogové číslo D201 + D202) Popis Column DNA Lego Kit je základ moderní stavebnicové (Lego) soupravy pro izolaci čisté DNA různého
Column DNA Lego Kit UNIVERZÁLNÍ SOUPRAVY PRO RYCHLOU IZOLACI ČISTÉ DNA (Katalogové číslo D201 + D202) Popis Column DNA Lego Kit je základ moderní stavebnicové (Lego) soupravy pro izolaci čisté DNA různého
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Transgenní řepka olejka (Brassica napus L.) její monitoring, molekulární detekce a vliv agrotechniky na eliminaci výdrolu
 její monitoring, molekulární detekce a vliv agrotechniky na eliminaci výdrolu") Transgenní řepka olejka (Brassica napus L.) její monitoring, molekulární detekce a vliv agrotechniky na eliminaci výdrolu Genové inženýrství umožňuje vnesení hospodářsky významných znaků do zájmových plodin
Transgenní řepka olejka (Brassica napus L.) její monitoring, molekulární detekce a vliv agrotechniky na eliminaci výdrolu Genové inženýrství umožňuje vnesení hospodářsky významných znaků do zájmových plodin
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION
 USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod
 Molekulární přístupy ve šlechtění rostlin Olomouc 14. února, 2017 Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod Jan Bartoš Ústav experimentální botaniky Olomouc, Czech Republic
Molekulární přístupy ve šlechtění rostlin Olomouc 14. února, 2017 Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod Jan Bartoš Ústav experimentální botaniky Olomouc, Czech Republic
PCR IN DETECTION OF FUNGAL CONTAMINATIONS IN POWDERED PEPPER
 PCR IN DETECTION OF FUNGAL CONTAMINATIONS IN POWDERED PEPPER Trojan V., Hanáček P., Havel L. Department of Plant Biology, Faculty of Agronomy, Mendel University of Agriculture and Forestry in Brno, Zemedelska
PCR IN DETECTION OF FUNGAL CONTAMINATIONS IN POWDERED PEPPER Trojan V., Hanáček P., Havel L. Department of Plant Biology, Faculty of Agronomy, Mendel University of Agriculture and Forestry in Brno, Zemedelska
cdna synthesis kit First-Strand cdna Synthesis System Verze 1.2
 cdna synthesis kit First-Strand cdna Synthesis System Verze 1.2 Obsah soupravy a její skladování Tato souprava pro reverzní transkripci obsahuje reagencie potřebné k provedení reverzní transkripce (RT)
cdna synthesis kit First-Strand cdna Synthesis System Verze 1.2 Obsah soupravy a její skladování Tato souprava pro reverzní transkripci obsahuje reagencie potřebné k provedení reverzní transkripce (RT)
1. Metodika. Protokol č. F1-4 Metodika: Srovnávací analýza efektivity přípravy rekombinantního proteinu ve fermentoru
 Protokol č.: F1-4 Datum: 20.12.2010 Metodika: analýza efektivity přípravy výběr z výsledků ze zkušebních provozů výroby antigenů. Vypracoval: Ing. Václav Filištein, Mgr. Tereza Chrudimská, Spolupracující
Protokol č.: F1-4 Datum: 20.12.2010 Metodika: analýza efektivity přípravy výběr z výsledků ze zkušebních provozů výroby antigenů. Vypracoval: Ing. Václav Filištein, Mgr. Tereza Chrudimská, Spolupracující
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316
 Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Tradice šlechtění šlechtění zlepšování pěstitelsky, technologicky a spotřebitelsky významných vlastností
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Tradice šlechtění šlechtění zlepšování pěstitelsky, technologicky a spotřebitelsky významných vlastností
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Genetické markery - princip a využití
 Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Genetické markery - princip a využití Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII. Martina Nováková, VŠCHT Praha
 MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Izolace nukleových kyselin
 Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
BÍLKOVINY HLÍZ BRAMBOR
 Jihočeská univerzita v Českých Budějovicích ZEMĚDĚLSKÁ FAKULTA BÍLKOVINY HLÍZ BRAMBOR jejich izolace a možnosti uplatnění Jan Bárta a kol. 19. května 2015, České Budějovice Kancelář transferu technologií
Jihočeská univerzita v Českých Budějovicích ZEMĚDĚLSKÁ FAKULTA BÍLKOVINY HLÍZ BRAMBOR jejich izolace a možnosti uplatnění Jan Bárta a kol. 19. května 2015, České Budějovice Kancelář transferu technologií
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
NOVÉ ODRŮDY OZIMÉ ŘEPKY OCEANIA A ORION A MÁKU SETÉHO ORBIS
 NOVÉ ODRŮDY OZIMÉ ŘEPKY OCEANIA A ORION A MÁKU SETÉHO ORBIS New Varieties of Winter Rapeseed and and Poppy Orbis Viktor VRBOVSKÝ OSEVA PRO s.r.o., Výzkumný ústav olejnin Opava Summary: The article presents
NOVÉ ODRŮDY OZIMÉ ŘEPKY OCEANIA A ORION A MÁKU SETÉHO ORBIS New Varieties of Winter Rapeseed and and Poppy Orbis Viktor VRBOVSKÝ OSEVA PRO s.r.o., Výzkumný ústav olejnin Opava Summary: The article presents
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ. I. Šubrt Společnost lékařské genetiky ČLS JEP
 EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
IZOLACE DNA (KIT DNeasy Plant Mini)
") 17.1 Izolace DNA (kit DNeasy Plant Mini) Strana 1 IZOLACE DNA (KIT DNeasy Plant Mini) 1 Účel a rozsah Postup slouží k získání deoxyribonukleové kyseliny (DNA) ze vzorku pomocí komerčního kitu DNeasy Plant
17.1 Izolace DNA (kit DNeasy Plant Mini) Strana 1 IZOLACE DNA (KIT DNeasy Plant Mini) 1 Účel a rozsah Postup slouží k získání deoxyribonukleové kyseliny (DNA) ze vzorku pomocí komerčního kitu DNeasy Plant
ODLIŠENÍ ODRŮD PŠENICE OBECNÉ TRITICUM AESTIVUM L. METODOU RAPD
 ODLIŠENÍ ODRŮD PŠENICE OBECNÉ TRITICUM AESTIVUM L. METODOU RAPD Distinguishing of Wheat Varieties (Tritium aestivum L.) by Method RAPD Zuzana Kohutová, Zuzana Kocourková, Hana Vlastníková, Petr Sedlák
ODLIŠENÍ ODRŮD PŠENICE OBECNÉ TRITICUM AESTIVUM L. METODOU RAPD Distinguishing of Wheat Varieties (Tritium aestivum L.) by Method RAPD Zuzana Kohutová, Zuzana Kocourková, Hana Vlastníková, Petr Sedlák
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Polymorfismus délky restrikčních fragmentů (RFLP)
") ÚSTAV LÉKAŘSKÉ BIOCHEMIE A LABORATORNÍ DIAGNOSTIKY 1. LF UK Polymorfismus délky restrikčních fragmentů (RFLP) Praktické cvičení z lékařské biochemie Všeobecné lékařství Martin Vejražka 2017/18 Obsah POLYMORFISMUS
ÚSTAV LÉKAŘSKÉ BIOCHEMIE A LABORATORNÍ DIAGNOSTIKY 1. LF UK Polymorfismus délky restrikčních fragmentů (RFLP) Praktické cvičení z lékařské biochemie Všeobecné lékařství Martin Vejražka 2017/18 Obsah POLYMORFISMUS
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Genetické markery Genetické markery = znaky, které informují o genotypu - vhodné jsou znaky,
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské. doc. RNDr. Ivan Mazura, CSc.
 Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 04 Inkompatibilita doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Inkompatibilní systémy vyšších rostlin Neschopnost rostlin tvořit semena Funkční gamety zachovány
Bi8240 GENETIKA ROSTLIN Prezentace 04 Inkompatibilita doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Inkompatibilní systémy vyšších rostlin Neschopnost rostlin tvořit semena Funkční gamety zachovány
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Genetické markery, markery DNA
 Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Obecná genetika Genetické markery, markery DNA Prof. Ing. Dušan GÖMÖRY, DrSc. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Šlechtění pšenice ozimé v Hrubčicích Ing. Eva Fučíková. Datum: 26. 11. 2014
 Šlechtění pšenice ozimé v Hrubčicích Ing. Eva Fučíková. Datum: 26. 11. 2014 Inovace studijních programů AF a ZF MENDELU směřující k vytvoření mezioborové integrace CZ.1.07/2.2.00/28.0302 Tato prezentace
Šlechtění pšenice ozimé v Hrubčicích Ing. Eva Fučíková. Datum: 26. 11. 2014 Inovace studijních programů AF a ZF MENDELU směřující k vytvoření mezioborové integrace CZ.1.07/2.2.00/28.0302 Tato prezentace
ALLELE FREQUENCY OF KIT GENE ASSOCIATED WITH TOBIANO SPOTTING PATTERN IN PAINT HORSE BREED
 ALLELE FREQUENCY OF KIT GENE ASSOCIATED WITH TOBIANO SPOTTING PATTERN IN PAINT HORSE BREED FREKVENCE ALEL GENU KIT ASOCIOVANÉHO SE STRAKATOSTÍ TOBIANO U KONÍ PLEMENE PAINT HORSE Chalupová P., Déduchová
ALLELE FREQUENCY OF KIT GENE ASSOCIATED WITH TOBIANO SPOTTING PATTERN IN PAINT HORSE BREED FREKVENCE ALEL GENU KIT ASOCIOVANÉHO SE STRAKATOSTÍ TOBIANO U KONÍ PLEMENE PAINT HORSE Chalupová P., Déduchová
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
RNA Blue REAGENS PRO RYCHLOU PŘÍPRAVU ČISTÉ A NEDEGRADOVANÉ RNA (katalogové číslo R011, R012, R013)
") RNA Blue REAGENS PRO RYCHLOU PŘÍPRAVU ČISTÉ A NEDEGRADOVANÉ RNA (katalogové číslo R011, R012, R013) Upozornění: RNA Blue obsahuje fenol a další toxické komponenty. Při kontaktu s kůží je nutné omytí velkým
RNA Blue REAGENS PRO RYCHLOU PŘÍPRAVU ČISTÉ A NEDEGRADOVANÉ RNA (katalogové číslo R011, R012, R013) Upozornění: RNA Blue obsahuje fenol a další toxické komponenty. Při kontaktu s kůží je nutné omytí velkým
Analýza DNA. Co zjišťujeme u DNA DNA. PCR polymerase chain reaction. Princip PCR PRINCIP METODY PCR
 o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Biotechnologické metody ve šlechtění rostlin
 KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ září 2014 prof. Ing. Vladislav Čurn, Ph.D. Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta, Katedra speciální produkce rostlinné Na Sádkách 1780, 370
KATEDRA SPECIÁLNÍ PRODUKCE ROSTLINNÉ září 2014 prof. Ing. Vladislav Čurn, Ph.D. Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta, Katedra speciální produkce rostlinné Na Sádkách 1780, 370
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém
 Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
OBSAH. PP Master Mixy... 11 PPP Master Mix 12 Plain PP Master Mix 13 Combi PPP Master Mix 14
 OBSAH DNA polymerázy pro PCR a pufry.................. 3 Taq DNA polymeráza 4 Taq DNA polymeráza Unis 5 TaqPurple DNA polymeráza 6 Taq DNA polymeráza 1.1 7 Combi Taq DNA polymeráza 8 LA DNA Polymerases
OBSAH DNA polymerázy pro PCR a pufry.................. 3 Taq DNA polymeráza 4 Taq DNA polymeráza Unis 5 TaqPurple DNA polymeráza 6 Taq DNA polymeráza 1.1 7 Combi Taq DNA polymeráza 8 LA DNA Polymerases
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
Nauka o dědičnosti a proměnlivosti
 Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základní pravidla dědičnosti
 Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Tkáňové kultury rostlin. Mikropropagace
 Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014. Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU
 V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
Národní program uchování a využití genetických zdrojů kulturních rostlin a agrobiodiverzity
 Národní program uchování a využití genetických zdrojů kulturních rostlin a agrobiodiverzity Jan Pelikán Výzkumný ústav pícninářský,spol. s r. o. Troubsko a Zemědělský výzkum, spol. s r. o. Troubsko Co
Národní program uchování a využití genetických zdrojů kulturních rostlin a agrobiodiverzity Jan Pelikán Výzkumný ústav pícninářský,spol. s r. o. Troubsko a Zemědělský výzkum, spol. s r. o. Troubsko Co
Tkáňový homogenizátor MagNA Lyser od společnosti Roche
 Izolace RNA Pracovní postup Homogenizace: Pozn. Postup homogenizace platí pouze pro izolaci RNA z nativní tkáně, v případě izolace z buněčné suspenze je tento krok vynechán a začíná se přídavkem homogenizačního
Izolace RNA Pracovní postup Homogenizace: Pozn. Postup homogenizace platí pouze pro izolaci RNA z nativní tkáně, v případě izolace z buněčné suspenze je tento krok vynechán a začíná se přídavkem homogenizačního
Ing. Prokop Šmirous, Ph.D. ŠLECHTĚNÍ KMÍNU V AGRITECU
 Ing. Prokop Šmirous, Ph.D. ŠLECHTĚNÍ KMÍNU V AGRITECU KRÁTCE Z HISTORIE Pesticidní pokusy od 70-80. let minulého stolení (herbicid Afalon, odrůda Rekord (1978)) Začátek šlechtitelské práce na kmínu v roce
Ing. Prokop Šmirous, Ph.D. ŠLECHTĚNÍ KMÍNU V AGRITECU KRÁTCE Z HISTORIE Pesticidní pokusy od 70-80. let minulého stolení (herbicid Afalon, odrůda Rekord (1978)) Začátek šlechtitelské práce na kmínu v roce
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Izolace bovinní genomové DNA z neinvazivně získaných biologických vzorků
 Izolace bovinní genomové DNA z neinvazivně získaných biologických vzorků Certifikovaná metodika č. 181 Jindřich Čítek Oto Hanuš Libor Večerek Eva Samková Lenka Hanusová Zuzana Křížová Irena Jelínková Robert
Izolace bovinní genomové DNA z neinvazivně získaných biologických vzorků Certifikovaná metodika č. 181 Jindřich Čítek Oto Hanuš Libor Večerek Eva Samková Lenka Hanusová Zuzana Křížová Irena Jelínková Robert
Metody analýzy DNA využívané ve Výzkumném a šlechtitelském ústavu Holovousy RNDr. Jana Čmejlová, Ph.D.
 Metody analýzy DNA využívané ve Výzkumném a šlechtitelském ústavu Holovousy RNDr. Jana Čmejlová, Ph.D. 1951 - Výzkumný ústav ovocnářský Holovousy 1997 - Výzkumný a šlechtitelský ústav ovocnářský Holovousy
Metody analýzy DNA využívané ve Výzkumném a šlechtitelském ústavu Holovousy RNDr. Jana Čmejlová, Ph.D. 1951 - Výzkumný ústav ovocnářský Holovousy 1997 - Výzkumný a šlechtitelský ústav ovocnářský Holovousy
IZOLACE DNA POMOCÍ CTAB PRO STANOVENÍ GMO METODOU PCR
 1253.1 Izolace DNA pomocí CTAB pro stanovení GMO Strana 1 IZOLACE DNA POMOCÍ CTAB PRO STANOVENÍ GMO METODOU PCR 1 Účel a rozsah Postup slouží k získání deoxyribonukleové kyseliny (DNA) ze vzorků krmiv
1253.1 Izolace DNA pomocí CTAB pro stanovení GMO Strana 1 IZOLACE DNA POMOCÍ CTAB PRO STANOVENÍ GMO METODOU PCR 1 Účel a rozsah Postup slouží k získání deoxyribonukleové kyseliny (DNA) ze vzorků krmiv
Důsledky selekce v populaci - cvičení
 Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Výzkumné centrum genomiky a proteomiky. Ústav experimentální medicíny AV ČR, v.v.i.
 Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Investujeme do naší společné budoucnosti
 Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Genetické markery. pro masnou produkci. Mgr. Jan Říha. Výzkumný ústav pro chov skotu, s.r.o.
 Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Genetické markery ve šlechtění skotu pro masnou produkci Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Genetické markery Polymorfní místa v DNA, které vykazují asociaci na sledované znaky Příčinné
Mendelova genetika v příkladech. Genetické markery
 Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
LABORATORNÍ PŘÍRUČKA. Laboratoř HLA systému a PCR diagnostiky
 LABORATORNÍ PŘÍRUČKA Transfuzní oddělení Laboratoř systému a PCR diagnostiky Účinnost od 15. 9. 2015 Verze č. 4 Tímto předpisem se ruší Verze č. 3, platnost od 1. 4. 2014 Odborný garant Jméno a příjmení,
LABORATORNÍ PŘÍRUČKA Transfuzní oddělení Laboratoř systému a PCR diagnostiky Účinnost od 15. 9. 2015 Verze č. 4 Tímto předpisem se ruší Verze č. 3, platnost od 1. 4. 2014 Odborný garant Jméno a příjmení,
Výuka genetiky na Přírodovědecké fakultě UK v Praze
 Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Cvičení č. 8. KBI/GENE Mgr. Zbyněk Houdek
 Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost
 Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost Registrační číslo: CZ.1.07/1. 5.00/34.0084 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Sada:
Výukový materiál zpracovaný v rámci operačního programu Vzdělávání pro konkurenceschopnost Registrační číslo: CZ.1.07/1. 5.00/34.0084 Šablona: III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Sada: