ONTOGENETIC TRANSITIONS (Leyser and Day Chap.9B)
|
|
|
- Bohuslav Kučera
- před 5 lety
- Počet zobrazení:
Transkript
1
2 ONTOGENETIC TRANSITIONS (Leyser and Day Chap.9B)
3 Often supported by the increase in GAs.
4 EMF EMF embryonic embryonicflower
5 Repression of many developmental programmes (often on the level of chromatin conformation) is the major regulatory mechanism in development.
6 Transition from embryonic to postembryonic ABA supports dormant state (some mutants are viviparous) GAs help to move into germination gibberellic acid 1 mutants (unable to synth. GAs) germinate only upon GAs addition (remember effects of light on germination ) GAs inactivate/supress embryogenic develop. programme allowing progression into juvenile phase
7 leafy cotyledon 1 LEC1
8
9
10 LEC1 leafy cotyledon 1 low level of storage proteins and dessication sensitive. excised embryos in vitro develop trichoms on the cotyledons CCAAT-box TF
11 LEC1 is necessary to promote cotyledon identity and storage subst. Works in embryos = mature plants of lec1 look like WT!!
12 OX - LEC1
13 Transition to post-embryon. dev. requires inactivation! of LEC1 action. brought about by
14
15 PKL pickle mutant accum. higher amount of storage products, in small plantlets accum. of storage lipids/oil bodies in roots able to produce calluses and somatic embryos
16 LEC1 je TF příbuzný rodině CCAAT-box bílkovin. PKL je členem CHD3 rodiny regulátorů chromatinu. by pkl mutant LEC1 expression stops, but starts again PKL keeps LEC1 suppressed
17 PKL is expressed throughout the plant and ontogenesis
18 U pkl mutanta je LEC1 exprimován i po klíčení. Pkl vykazuje giberelinový fenotyp tmavě zelené listy, menší a pozdně kvetoucí (1d zpoždění)s akumulací giberelinů. pkl looks like GAs mutants dark green late flowering PKL je expr. konstitutivně tedy regulace gibereliny probíhá na úrovni bílkoviny. pkl, gai dvojitý mutant kvete o 33 dní později
19 pkl and GAs double mutants have additive phenotypes pkl flowers 1 day later gai (GAs insensitive) flowers 3 days later double pkl x gai1 flowers 33d later!
20
21
22 Phase transitions in postgermination development
23 Morphology of phytomer changes from juv. to adult to flowering.
24 bolting stem
25 Some cauline leaves are initiated before flowering.
26 Vývojové přechody během vývoje po vyklíčení Juvenilní (trichomy jen adaxiálně) a dospělé fytomery Arabidopsis se vyznačují absencí dlouživého růstu stonku. Podpůrné listeny květenství byly pravděpodobně založeny (anlage) již před přechodem do kvetení. Změny ve stavbě fytomer při vývojových přechodech.
27 CHANGES IN AXILLARY SAMs
28 1.Úžlabní pupeny zakládány dodatečně Akropetálně. 2.Simultánní zakládání listu a pupene basipetálně. 3. Úžlabní květní pupen je podepřen přechodně residuem listové anlage (ex- Prese AS1).
29 1.Úžlabní pupeny zakládány dodatečně Akropetálně. 2.Simultánní zakládání listu a pupene basipetálně. 3. Úžlabní květní pupen je podepřen přechodně residuem listové anlage (ex- Prese AS1).
30 1.Úžlabní pupeny zakládány dodatečně Akropetálně. 2.Simultánní zakládání listu a pupene basipetálně. 3. Úžlabní květní pupen je podepřen přechodně residuem listové anlage (ex- Prese AS1).
31
32 Identita květenství a květů Floral meristem identity = FMI př. geny
33 LEAFY --lfy lfy je je dominantní regulátor/udržovatel identity květního meristému A --mutant mutant vytváří vytváří místo místo květů květů listovité listovité sekundární sekundární výhony výhony B --35S::LFY 35S::LFY exprimuje exprimujelfy Lfy konstitutivně konstitutivně a vytváří vytváří květy květy místo místo sekundárních sekundárních výhonů výhonů z z růžice. růžice.
34 LEAFY --lfy lfy je je dominantní regulátor/udržovatel identity květního meristému A --mutant mutant vytváří vytváří místo místo květů květů listovité listovité sekundární sekundární výhony výhony B --35S::LFY 35S::LFY exprimuje exprimujelfy Lfy konstitutivně konstitutivně a vytváří vytváří květy květy místo místo sekundárních sekundárních výhonů výhonů z z růžice. růžice.
35 Mutant phenotype becomes less severe (also in ap1) in older nodes. however lfy x ap1 double = almost complete conversion of flowers into shoots (i.e. they can partially compensate each other)
36 lfy plant spec. TF místo květů výhony ap1 - (MADS-box homl.)- podobně, ale místo kalicha listeny, v jejichž úžlabí vznikají sekundární květy cal (MADS-box homl.)- = WT phen. ap1 cal místo květů opakovaně se zakládající mristémy květenství. tfl1 - květenství předčasně ukončeno koncovým květem. ap1x lfy - úplná konverze květů ve vegetativní výhony
37 cal homozygot je bez genotypu ap1 cal dvojitý mutat zakládá na místě květů opakovaně sekundární meristémy



38 BoCAL Locus in Broccoli, Cauliflower, and Purple Cauliflower Fig 1a. De Cicco, broccoli B. oleracea v. italica Fig 1b. Yeh Erh Fu, cauliflower B. oleracea v. botrytis Fig 1c. Violet Queen, purple cauliflower B. oleracea v. italica
39 LFY je TF specifický pro rostliny CAL a AP1 jsou MADS-box homology TFL1 je podobný inhibitoru RAF kináz (RKIP).
40 Jen LFY je exprimován ve veget. fázi slabě v listových primordiích. 24h po květní indukcí se LFY exprese přesouvá do SAM a sílí. Až po 72h se objevuje exprese AP1 a CAL.
41 Exprese AP1 je silně zpožděna a oslabena u lfy. LFY se váže do AP1 promotoru. Podobně reguluje CAL. LFY expression is very low in ap1 x cal double mutants. AP1 a CAL zase pozitivní zpětnou vazbou zesilují syntézu LFY.
42 POSITIVE FEEDBACK LOOP

43 lfy plant spec. TF místo květů výhony ap1 - (MADS-box homl.)- podobně, ale místo kalicha listeny, v jejichž úžlabí vznikají sekundární květy cal (MADS-box homl.)- = WT phen. ap1 cal místo květů opakovaně se zakládající mristémy květenství. tfl1 - květenství předčasně ukončeno koncovým květem. ap1x lfy - úplná konverze květů ve vegetativní výhony
44 TFL1 blokuje expresi FMI genů v meristému květenství TFL1 expr. slabě pod veget. SAM TFL1 expr.silně pod meristémem květenství. U mutantů lfy a ap1 cal je expr. i v pův. květních meristémech. WT TFL1 je inhibován jejich WT alelami. Jak vypadá AP1 či LFY 35S::overexpresor?

45 LEAFY --lfy lfy je je dominantní regulátor/udržovatel identity květního meristému A --mutant mutant vytváří vytváří místo místo květů květů listovité listovité sekundární sekundární výhony výhony B --35S::LFY 35S::LFY exprimuje exprimujelfy Lfy konstitutivně konstitutivně a vytváří vytváří květy květy místo místo sekundárních sekundárních výhonů výhonů z z růžice. růžice.
46 FMI genes initiate transcription of ABC genes of flower organ identity

47
48 TFL1 as an inhibitor of phytomere progression Architektura květenství je řízena přetlačováním mezi TFL1 a geny květní identity (FMI geny). TFL1 celkově zpomaluje přechod a průchod generativní fází. Evolučně původní se zdá být květenství ukončené terminálním květem jako u tfl1.
49 Neukončená květenství se zdají být evolučně odvozená Homolog u hledíku CEN (CENTRORADIALIS) byl rekrutován evolučně nezávisle. Není expr. ve vegetativní fázi, ale v generativní funguje jako TFL1.
50 The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1 Lilac Pnueli1, Lea Carmel-Goren2, Dana Hareven1, Tamar Gutfinger1, John Alvarez3, Martin Ganal4, Daniel Zamir2 and Eliezer Lifschitz1,* 1Department of Biology, Technion-Israel Institute of Technology, Haifa 32000, Israel 2Department of Genetics and Field Crops, Faculty of Agriculture, Hebrew University, Rehovot 70700, Israel 3Department of Biological Sciences, Monash University, Clayton, Victoria 3168, Australia 4Institute for Plant Genetics, Corrensstrasse 3, D Gatersleben, Germany Development 125, (1998) 1979 SUMMARY Vegetative and reproductive phases alternate regularly during sympodial growth in tomato. In wild-type indeterminate plants, inflorescences are separated by three vegetative nodes. In determinate plants homozygous for the recessive allele of the SELF-PRUNING (SP) gene, sympodial segments develop progressively fewer nodes until the shoot is terminated by two consecutive inflorescences. We show here that the SP gene is the tomato ortholog of CENTRORADIALIS and TERMINAL FLOWER1, genes which maintain the indeterminate state of inflorescence meristems in Antirrhinum and Arabidopsis respectively. The sp mutation results in a single amino acid change (P76L), and the mutant phenotype is mimicked by overexpressing the SP antisense RNA. Ectopic and overexpression of the SP and CEN transgenes in tomato rescues the indeterminate phenotype, conditions the replacement of flowers by leaves in the inflorescence and suppresses the transition of the vegetative apex to a reproductive shoot. The SELF-PRUNING gene is expressed in shoot apices and leaves from very early stages, and later in inflorescence and floral primordia as well. This expression pattern is similar to that displayed by the tomato ortholog LEAFY and FLORICAULA. Comparison of the sympodial, day-neutral shoot system of tomato and the monopodial, photoperiod-sensitive systems of Arabidopsis and Antirrhinum suggests that flowering genes that are required for the processing of floral induction signals in Arabidopsis and Antirrhinum are required in tomato to regulate the alternation between vegetative and reproductive cycles in sympodial meristems. Key words: Growth habit, Reproductive switching, Sympodial shoot, Determinate meristem, SELF-PRUNING (SP), CENTRORADIALIS (CEN)/TERMINAL FLOWER1 (TFL1), Tomato


51
52 Indeterminate and determinate shoots of tomato. (A) Indeterminate (SP) shoot: One full-size sympodial segment is shown. It consists of three leaves and a terminal inflorescence. The third leaf of such a unit (No. 3) appears above the inflorescence because it is united with the new, fast growing sympodial unit. Arrows indicate three consecutive inflorescences. The insert features a scorpioid (zigzag) tomato inflorescence. (B) Determinate (sp/sp) shoot. Only one nodal leaf separates the first two inflorescences. Arrows mark the terminal inflorescence (TI) and an axillary shoot (AS) developing below the older inflorescence. (C) Shoot of an sp double mutant. Note the distance between inflorescences and the termination of the shoot just as in the determinate, (sp/sp) plants, in B.


53
54 SIGNAL INTEGRATION IN FLORAL TRANSITION
55
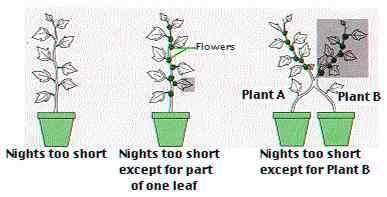
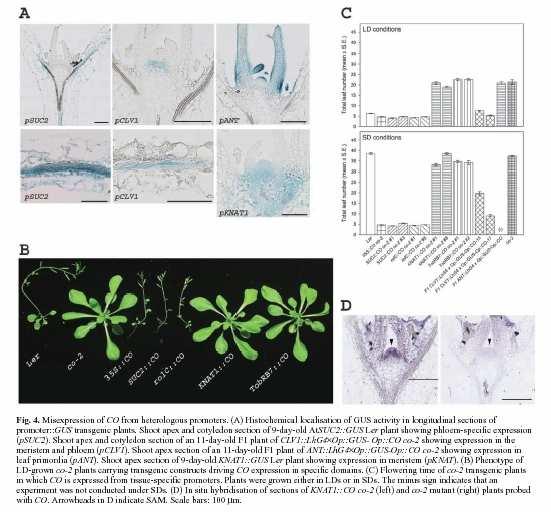
56
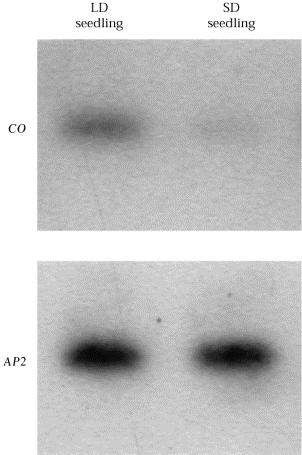
57

58
59
60

61 GRAFTING EXPERIMENTS IN ARABIDOPSIS
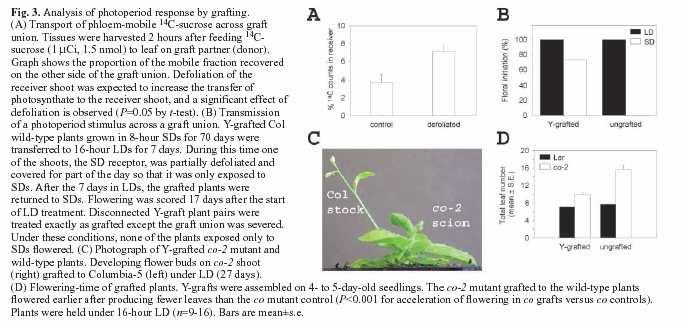
62
63
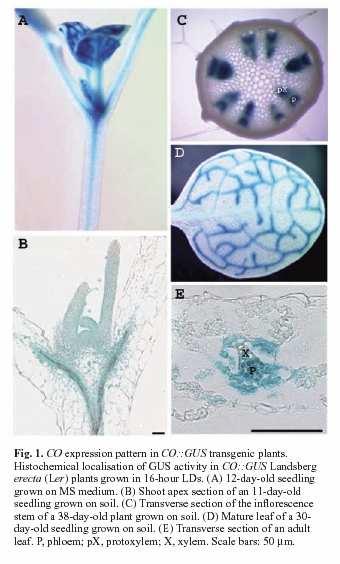
64 CO sám o sobě není florigen. Je exprimován a působí výhradně v doprovodných buňkách floemu aktivací FT. FT je exprimován v různých pletivech a aktivován různými drahami.je pravděpodobné, že působí buněčně neautonomě (23kDa). Že by součást florigenu? CO působí ještě přes další FT nezávislou dráhu.
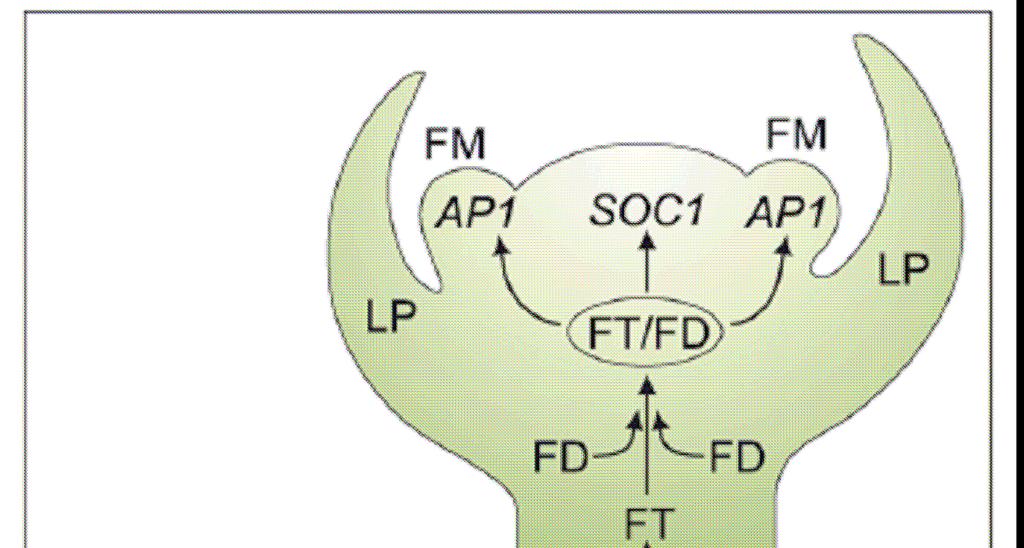


65
66 Síť integrované signalizace

67 Fytochrom FytochromB zpožďuje zpožďuje nástup nástup kvetení kvetení proto proto mutant mutant kvete kvete časněji časněji než než WT. WT. Fytochrom FytochromA naopak naopak urychluje urychluje nástup nástup kvetení. kvetení.
68 4 x signalling pathways integrated
69
70 AGL20=SOC1
71 Autonomní dráha kvetení Geny zjišťovány jako mutanti, kteří kvetou normálně za induktivních podmínek, ale později v neindukt. podmínkách. Př. FCA RNA vaz. bílkovina; LD TF. Působí přes inhibici inhibitoru FLC tj. mutace nepůsobí zpoždění kvetení u flc.
72 AGL20=SOC1
73 K integraci drah fotoperiodické, vernalizační a autonomní dochází na úrovni genů FT a AGL20=SOC1 Integrace signalizační Integrace na úrovni promotoru INTEGRATION OF PATHWAYS ON THE PROMOTOR LEVEL.
74 Vliv giberelinů ga1 (silně postižená syntéza giberelinů) kvete i na LD pozdě. spindly (spy WT je tetratrikopept. rep. bílkovina) s konstitutivně aktivní giberelinovou dráhou kvete dříve. Gibereliny zřejmě stimulují všechny ontogenetické vývojové přechody.
75 Je izolována řada recesivních mutantů s velmi časným kvetením - dráha aktivně inhibující kvetení embryonic flower 1 a 2 (emf) - TF a VRN2 homolog early flowering 1 a 2 (elf). terminal flower2 (tfl2, homol. heterochromatin 1) často regulátory chromatinu OFTEN CHROMATIN REGULATORS

76 EMF EMF embryonic embryonicflower
77 O regulaci přechodu z juvenilní do dospělé (schopnost přejít do generativní f.) fáze vegetativního vývoje se ví, že je částečně! regulována faktory přechodu do generativní fáze (světlo, gibereliny,co,fca). hasty (hst) mutant komplementárně k např. 35S::LFY (méně dospělých listů, stejně juven. jako WT) tvoří méně juvenilních listů než WT, ale stejný počet dospělých. Tedy je inhibitorem přechodu do dospělé fáze.
78 JUVENILE AND ADULT PHASES ARE DISTINGUISHED BASED ON COMPETENCE TO FLOWER
79 SHOOT BRANCHING
80 Větvení vegetativního výhonu úžlabní pupeny - většinou iniciují několik listů a zastavují vývoj.
81 Apikální dominance a IAA Transgenní (mutantní) rostliny s vyšší konc.iaa mají snížené větvení a naopak. axr1 se sníženou citlivostí k IAA je více větvený - má nižší apikální dominanci.
82
83 AND CYTOKININS
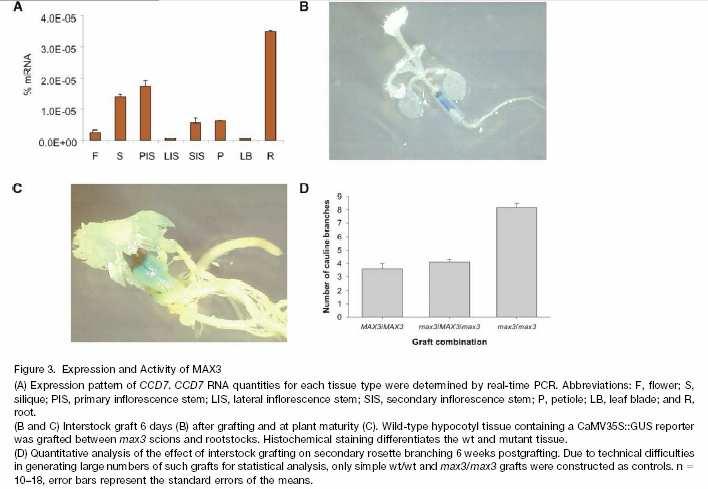
84 úloha cytokininů zvýšená hladina stimuluje růst postranních pupenů
85 biosyntéza cytokininů probíhá silně v kořenech. je stimulována dostatkem dusíku tak růst postraních výhonů je spoluregulován dostupností živin v půdě IAA z nadzemníčásti naopak podporuje růst postranních kořenů
86 Dekapitace u hrachu vede k náhlému vzrůstu transportu cytokininů z kořenů do nadzemního výhonu. Inhibice transportu cytokininů není ovšem hlavním účinkem IAA - dekapitace funguje i u rostlin oddělených od kořenů. IAA ALSO INHIBITS SYNTHESIS OF CYTOKINS
87 Mutants Arabidopsis max (more axillary growth) Pea rms (ramosus) Petunia dad (decreased apical dominance)
88 Mutace ramosus (rms) u hrachu je jich známo 5. Původně nejstudovanější je rms1. Dekapitovaný rms1 nereaguje na bloček s IAA - tato funkce je níže v regulaci apikální dominance (pod IAA).

89 RMS1 homol. to MAX4 and DAD1
90
91 RMS1 se podílí na vzniku akropetálně transportovaného signálů (z kořenů do nadzemí) WT podnož působí WT větvení na rms1 roubu. WT roub je neovlivněn rms1 podnoží. Kousek WT stonku uprostřed upraví větvení jen nad ne pod. u rms1 je 15x nižší transport cytokininů - tj. RMS1 nepůsobí přes cytokininy.
92 Koordinace růstu rostliny vnitřním a vnějším prostředím.
 je homolog")
93 MAX4 (more axillary branching) je homolog MAX3(RMS1)







94 Fig. 1. Branching phenotype of 35-d-old WT Arabidopsis and max4 mutant.

95

96

97
98
99 max3 nemá podstatně změněny hladiny IAA a ABA. max3 má chloroplastový lokalizační signál - transitní peptid.

100 a je importován do plastidů a hromadí se ve stromatu
101 HPLC GC induk. induk. Bakterie exprimující MAX3 štěpí karotenoidy
102
103





104


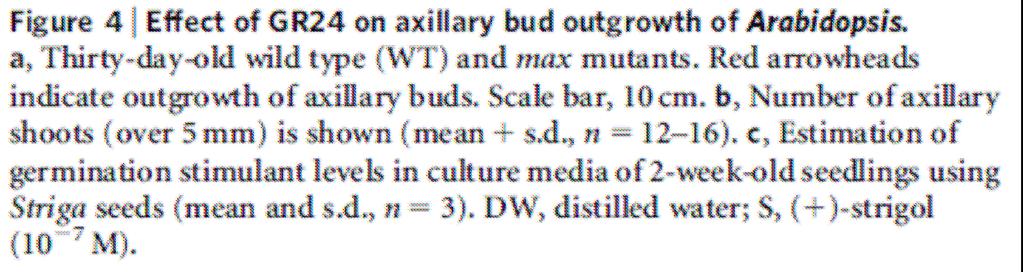
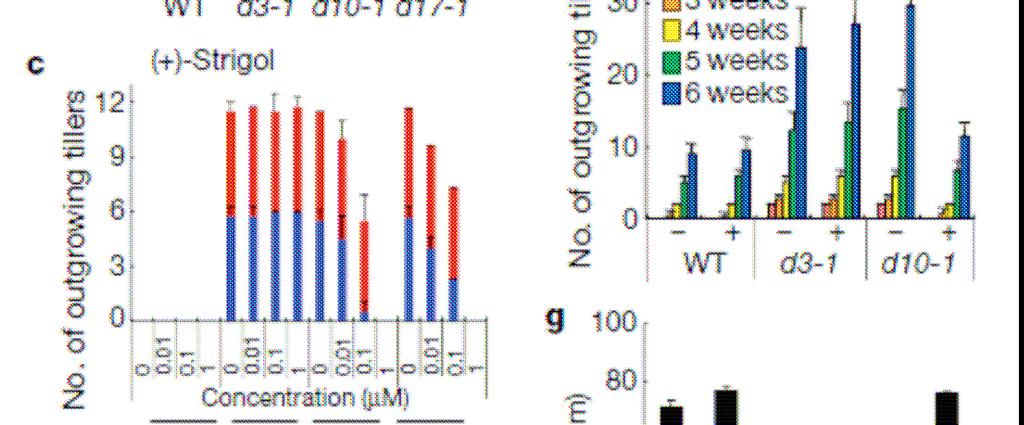
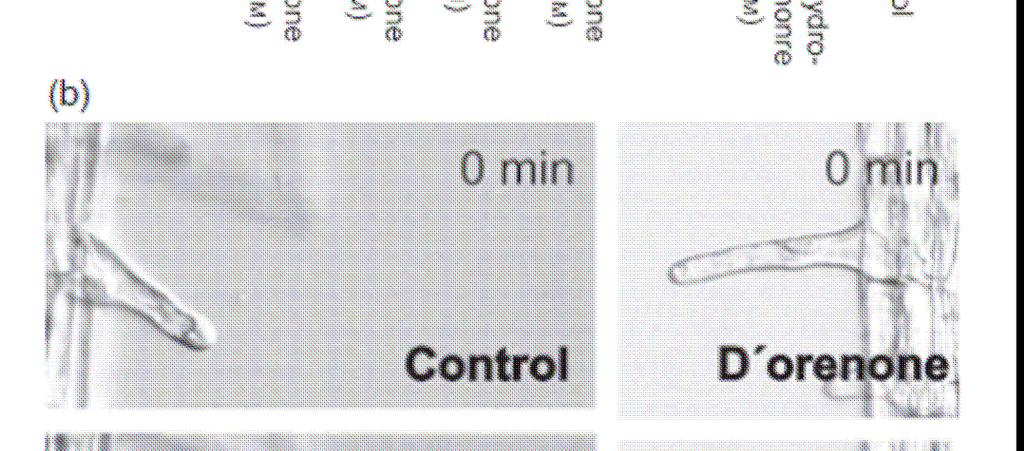
105
106

107




108


109
110
111 also rice got MAX homologues
112 Model Ottoline Leyser 2007
113 Fig. 2. Model for MAX pathway function through the regulation of auxin transport capacity. In WT stems the MAX pathway down-regulates auxin transporters; thus auxin transport capacity in the stem is limited and saturated by apically-derived auxin. Buds are unable to export auxin; therefore they cannot outgrow. In max mutants there is more auxin transport capacity in the stem, and this means that the buds are able to export auxin and outgrowth (indicated by the yellow arrow). Green squares represent auxin transporters and the arrows represent auxin that is being carried down the stem. 70 Ongaro and Leyser April
114 Fig. 3. Model for the hormonal control of shoot branching in Arabidopsis. Auxin is transported down the stem in the polar transport stream. The capacity of the stream is regulated by the MAX pathway, in which MAX3, MAX4, and MAX1 act in the synthesis of a novel upwardly mobile hormone that regulates PIN1 levels through MAX2. For bud activation, buds must be able to export auxin and thus MAX-limited auxin transport capacity in the stem prevents bud outgrowth. In addition, the concentration of auxin in the polar transport stream is monitored by the AXR1/AFB pathway to regulate cytokinin synthesis. High auxin down-regulates cytokinin synthesis, inhibiting bud activation. The model predicts, rather paradoxically, that because of the increased capacity for auxin transport in the stem of max mutants, there will be high auxin and thus low cytokinin in the stem, but none the less increased branching, because of the high transport capacity (see Fig. 2). Orthologues of the MAX genes in pea (RMS) and petunia (DAD) are written in brackets.
115 Závěry kap. 9 Setrvalá produkce fytomer tvořících rostlinný organismus je umožněna neukončeností vývoje meristémů; to je předpokladem vývojové plasticity rostlin. Tato neukončenost souvisí s udržováním dynamické rovnováhy mezi tvorbou a diferenciací buněk v meristému. Typ fytomer je ovlivněn vývojovým stádiem rostliny. Vývojové, fyziologické a vnější procesy rozhodují o tom kolik fytomer vznikne a kdy dojde k přechodu do nové vývojové fáze (z vegetativní do generativní).
116 Srovnání vývoje rostlin a živočichů
117 vývoj květu odpovídá mechanismu založenému na kaskádách transkripčních faktorů. vývoj rostlinného embrya již podstatně méně. není podobnost mezi embryogenezí živočichů a vývojem květu důkazem pokračující embryogeneze u rostlin?
118 U živočichů i rostlin hraje klíčovou roli represe genové exprese na úrovni chromatinu (polycomb - např. CLF).
119 Diferenciace založená na asymetrickém dělení buňky
120 Diferenciace založená na mezibuněčné signalizaci Mechanismy tvorby uspořádání jsou vždy založeny na zpětnovazebných regulačních obvodech, které zesilují počáteční malé rozdíly.
121 U živočichů hraje důležitou roli tkáňová kompartmentalizace (závislá na mezibuněčné adhezi), která zabraňuje nepatřičným migracím. BOUNDARIES AGAINST MIGRTIONS
122 ektopická lokalizace anteriorních buněk v posteriorní zóně s morfogenem WG značí nové místo překryvu s DPP a novou špičku křídla DPP=decapentaplegic anterior - poster. grad. WG=wingless dorso-vetral grad.
123 Pro přežívání živočišných buněk jsou důležité tzv. "přežívací signály" od okolních buněk. Jinak nastupuje PCD. Apoptóza působená nedostakem "přežívacích signálů" je hlavním rysem morfogeneze/vývoje živočichů.
124 rostliny mají různé cesty získávání živin a energie.

125 Totipotence Totipotencea neukončená neukončenáembryogenese embryogenese rostlin rostlin
126 Závěr Přes nezávislý vznik mnohobuněčnosti a nezávislou evoluci mají mechanismy regulující vývoj mnohobuněčných rostlin a živočichů důležité společné rysy na molekulární úrovni v určování buněčného vývojového osudu a na teoretické/biokybernetické úrovni při vzniku pravidelných uspořádání. Jsou mezi nimi ovšem také fundamentální rozdíly vyplývající z odlišnosti vývojových constrains vyplývajících z přisedlosti a autotrofie rostlin. Tím nejdůležitějším rysem odlišujícím vývoj rostlin a živočichů je kontinuální (celoživotní) organogeneze rostlin na rozdíl od omezené terminální organogeneze živočichů. Tím je umožněna nesrovnatelně větší morfologická plasticita rostlin.
127 Haseloff/teaching/MCBPart1B/ Lecture1.html
128
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") 2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2) Reprodukce rostlin
 Reprodukce rostlin") 2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Rekapitulace. Rostlina vládne buňkám, ne(jen) buňky rostlině.
 buňky rostlině.") Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Životní cyklus rostliny. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení
 Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení") MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.)
") Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Detlef Weigel ( )
") VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Obecná a srovnávací odontologie. Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód
 Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky
 Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.)
") CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
CONTRIBUTION TO UNDERSTANDING OF CORRELATIVE ROLE OF COTYLEDON IN PEA (Pisum sativum L.) PŘÍSPĚVEK K POZNÁNÍ KORLAČNÍ FUNKCE DĚLOHY U HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav Biologie rostlin,
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Znak Stupnice hodnocení Hodnoty / Values Descriptor Evaluation scale Poznámka / Note. EVIGEZ č. deskr. /Desc. no. Poř.
 Vybrané popisné deskriptory/ Selected Characterisation and Evaluation Descriptors - Vicia sativa, V. pannonica, V.villosa (Ing. Miroslav Hýbl, PhD, AGRITEC Šumperk s.r.o. 2001) 1. MORFOLOGICKÉ ZNAKY /
Vybrané popisné deskriptory/ Selected Characterisation and Evaluation Descriptors - Vicia sativa, V. pannonica, V.villosa (Ing. Miroslav Hýbl, PhD, AGRITEC Šumperk s.r.o. 2001) 1. MORFOLOGICKÉ ZNAKY /
Laboratoř na čipu. Lab-on-a-chip. Pavel Matějka
 Laboratoř na čipu Lab-on-a-chip Pavel Matějka Typy analytických čipů 1. Chemické čipy 1. Princip chemického čipu 2. Příklady chemických čipů 3. Příklady analytického použití 2. Biočipy 1. Princip biočipu
Laboratoř na čipu Lab-on-a-chip Pavel Matějka Typy analytických čipů 1. Chemické čipy 1. Princip chemického čipu 2. Příklady chemických čipů 3. Příklady analytického použití 2. Biočipy 1. Princip biočipu
Database systems. Normal forms
 Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Aktivita CLIL Chemie I.
 Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
2012/2013. Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol.
 2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA
 GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost.
 Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE
 MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE BRNO 2012 Bc. NELA DAŇKOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Vliv strigolaktonu na větvení stonku rostlin
MENDELOVA UNIVERZITA V BRNĚ AGRONOMICKÁ FAKULTA DIPLOMOVÁ PRÁCE BRNO 2012 Bc. NELA DAŇKOVÁ Mendelova univerzita v Brně Agronomická fakulta Ústav biologie rostlin Vliv strigolaktonu na větvení stonku rostlin
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Just write down your most recent and important education. Remember that sometimes less is more some people may be considered overqualified.
 CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Gymnázium, Brno, Slovanské nám. 7 WORKBOOK. Mathematics. Teacher: Student:
 WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
12. ONTOGENEZE II : KVETENÍ, FOTOPERIODISMUS
 12. ONTOGENEZE II : KVETENÍ, 12.1 REGULACE PŘECHODU Z VEGETATIVNÍ DO REPRODUKTIVNÍ FÁZE Přechod od vegetativního růstu ke kvetení je zásadní změnou růstu a organogeze. Představuje ukončení opakované tvorby
12. ONTOGENEZE II : KVETENÍ, 12.1 REGULACE PŘECHODU Z VEGETATIVNÍ DO REPRODUKTIVNÍ FÁZE Přechod od vegetativního růstu ke kvetení je zásadní změnou růstu a organogeze. Představuje ukončení opakované tvorby
Litosil - application
 Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
Co je populační biologie. Specifika populační biologie. rostlin
 Co je populační biologie Specifika populační biologie rostlin Co je populační biologie? Disciplína studující změny velikosti populace v čase. N t+1 = N t + N t+1 = N t + B Birth N t+1 = N t + B - D Birth
Co je populační biologie Specifika populační biologie rostlin Co je populační biologie? Disciplína studující změny velikosti populace v čase. N t+1 = N t + N t+1 = N t + B Birth N t+1 = N t + B - D Birth
Ustavování polarity a os listu
 Shrnutí 2. Podélná osa je determinována již na úrovni zygoty, u krytosemenných vajíčka (žádná buňka není sama). Klíčovou roli hraje polární transport auxinu a fixace tělního rozvrhu kombinatorikou transkripčních
Shrnutí 2. Podélná osa je determinována již na úrovni zygoty, u krytosemenných vajíčka (žádná buňka není sama). Klíčovou roli hraje polární transport auxinu a fixace tělního rozvrhu kombinatorikou transkripčních
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
Zelené potraviny v nových obalech Green foods in a new packaging
 Energy News1 1 Zelené potraviny v nových obalech Green foods in a new packaging Již v minulém roce jsme Vás informovali, že dojde k přebalení všech tří zelených potravin do nových papírových obalů, které
Energy News1 1 Zelené potraviny v nových obalech Green foods in a new packaging Již v minulém roce jsme Vás informovali, že dojde k přebalení všech tří zelených potravin do nových papírových obalů, které
Jan F. Humplík Laboratoř růstových regulátorů & Oddělení chemické biologie a genetiky, CRH, ÚEB AV ČR
 Jan F. Humplík Laboratoř růstových regulátorů & Oddělení chemické biologie a genetiky, CRH, ÚEB AV ČR Fenotyp = vnější projevy genotypu + vliv prostředí sensors computer analysis plants database storage
Jan F. Humplík Laboratoř růstových regulátorů & Oddělení chemické biologie a genetiky, CRH, ÚEB AV ČR Fenotyp = vnější projevy genotypu + vliv prostředí sensors computer analysis plants database storage
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Introduction to MS Dynamics NAV
 Introduction to MS Dynamics NAV (Item Charges) Ing.J.Skorkovský,CSc. MASARYK UNIVERSITY BRNO, Czech Republic Faculty of economics and business administration Department of corporate economy Item Charges
Introduction to MS Dynamics NAV (Item Charges) Ing.J.Skorkovský,CSc. MASARYK UNIVERSITY BRNO, Czech Republic Faculty of economics and business administration Department of corporate economy Item Charges
WORKSHEET 1: LINEAR EQUATION 1
 WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable
WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
PART 2 - SPECIAL WHOLESALE OFFER OF PLANTS SPRING 2016 NEWS MAY 2016 SUCCULENT SPECIAL WHOLESALE ASSORTMENT
 PART 2 - SPECIAL WHOLESALE OFFER OF PLANTS SPRING 2016 NEWS MAY 2016 SUCCULENT SPECIAL WHOLESALE ASSORTMENT Dear Friends We will now be able to buy from us succulent plants at very good wholesale price.
PART 2 - SPECIAL WHOLESALE OFFER OF PLANTS SPRING 2016 NEWS MAY 2016 SUCCULENT SPECIAL WHOLESALE ASSORTMENT Dear Friends We will now be able to buy from us succulent plants at very good wholesale price.
Automatika na dávkování chemie automatic dosing
 Automatika na dávkování chemie automatic dosing Swimmingpool Technology Autodos 700 Automatické dávkování Autodos Autodos automatic dosing Autodos 700 je jedno-kanálové zaøízení, pro mìøení a dávkování.
Automatika na dávkování chemie automatic dosing Swimmingpool Technology Autodos 700 Automatické dávkování Autodos Autodos automatic dosing Autodos 700 je jedno-kanálové zaøízení, pro mìøení a dávkování.
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
PC/104, PC/104-Plus. 196 ept GmbH I Tel. +49 (0) / I Fax +49 (0) / I I
 / I Fax +49 (0) / I I") E L E C T R O N I C C O N N E C T O R S 196 ept GmbH I Tel. +49 (0) 88 61 / 25 01 0 I Fax +49 (0) 88 61 / 55 07 I E-Mail sales@ept.de I www.ept.de Contents Introduction 198 Overview 199 The Standard 200
E L E C T R O N I C C O N N E C T O R S 196 ept GmbH I Tel. +49 (0) 88 61 / 25 01 0 I Fax +49 (0) 88 61 / 55 07 I E-Mail sales@ept.de I www.ept.de Contents Introduction 198 Overview 199 The Standard 200
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
3) Růst a vývoj. a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů KFZR 1
 Růst a vývoj. a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů KFZR 1") 1 2010 3) Růst a vývoj a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů Raghavan V (2006) Double Fertilization. Embryo and Endosperm Development In Flowering Plants. Springer.
1 2010 3) Růst a vývoj a) Embryogeneze a cytokineze b) Meristém a vývoj rostliny c) Vývoj listů a kořenů Raghavan V (2006) Double Fertilization. Embryo and Endosperm Development In Flowering Plants. Springer.
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
Popisné deskriptory u zahradního mečíku - Characterisation and evaluation descriptors Gladiolus L. (Petr Novák, VÚKOZ Průhonice 2004)
") Popisné deskriptory u zahradního mečíku - Characterisation and evaluation descriptors Gladiolus L. (Petr Novák, VÚKOZ Průhonice 2004) č. znaku poř. číslo znak stupnice hodnoty/ values character scale Poznámka
Popisné deskriptory u zahradního mečíku - Characterisation and evaluation descriptors Gladiolus L. (Petr Novák, VÚKOZ Průhonice 2004) č. znaku poř. číslo znak stupnice hodnoty/ values character scale Poznámka
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných
RŮST A VÝVOJ ROSTLIN. Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1
 RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
RŮST A VÝVOJ ROSTLIN Mgr. Alena Výborná Gymnázium, SOŠ a VOŠ Ledeč nad Sázavou VY_32_INOVACE_01_1_11_BI1 Růst = nezvratné zvětšování rozměrů a hmotnosti rostliny. Dochází ke změnám tvaru a vnitřního uspořádání
Dostupnost živin jako faktor utvářející morfologii kořenů (trofomorfogeneze) Vliv dusíkatých látek, fosfátů, síranů a iontů železa
 Vliv dusíkatých látek, fosfátů, síranů a iontů železa") Dostupnost živin jako faktor utvářející morfologii kořenů (trofomorfogeneze) Vliv dusíkatých látek, fosfátů, síranů a iontů železa Co je ovlivněno? Primární kořen Laterální kořen Kořenové vlásky (tvoří
Dostupnost živin jako faktor utvářející morfologii kořenů (trofomorfogeneze) Vliv dusíkatých látek, fosfátů, síranů a iontů železa Co je ovlivněno? Primární kořen Laterální kořen Kořenové vlásky (tvoří
Základní pravidla dědičnosti - Mendelovy a Morganovy zákony
 Obecná genetika Základní pravidla dědičnosti - Mendelovy a Morganovy zákony Ing. Roman LONGAUER, CSc. Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je
Obecná genetika Základní pravidla dědičnosti - Mendelovy a Morganovy zákony Ing. Roman LONGAUER, CSc. Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49
 Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
Střední průmyslová škola strojnická Olomouc, tř.17. listopadu 49 Výukový materiál zpracovaný v rámci projektu Výuka moderně Registrační číslo projektu: CZ.1.07/1.5.00/34.0205 Šablona: III/2 Anglický jazyk
II_ _Listening Pracovní list č. 2.doc II_ _Listening Pracovní list č. 3.doc II_ _Listening Řešení 1,2.doc
 Název školy: ZŠ Brno, Měšťanská 21, Brno -Tuřany Název práce: Listening Pořadové číslo: II_2-01-06 Předmět: Anglický jazyk Třída: 9. AC Téma hodiny: Problémy Vyučující: Mgr. Milena Polášková Cíl hodiny:
Název školy: ZŠ Brno, Měšťanská 21, Brno -Tuřany Název práce: Listening Pořadové číslo: II_2-01-06 Předmět: Anglický jazyk Třída: 9. AC Téma hodiny: Problémy Vyučující: Mgr. Milena Polášková Cíl hodiny:
DATA SHEET. BC516 PNP Darlington transistor. technický list DISCRETE SEMICONDUCTORS Apr 23. Product specification Supersedes data of 1997 Apr 16
 zákaznická linka: 840 50 60 70 DISCRETE SEMICONDUCTORS DATA SHEET book, halfpage M3D186 Supersedes data of 1997 Apr 16 1999 Apr 23 str 1 Dodavatel: GM electronic, spol. s r.o., Křižíkova 77, 186 00 Praha
zákaznická linka: 840 50 60 70 DISCRETE SEMICONDUCTORS DATA SHEET book, halfpage M3D186 Supersedes data of 1997 Apr 16 1999 Apr 23 str 1 Dodavatel: GM electronic, spol. s r.o., Křižíkova 77, 186 00 Praha
Zubní pasty v pozměněném složení a novém designu
 Energy news4 Energy News 04/2010 Inovace 1 Zubní pasty v pozměněném složení a novém designu Od října tohoto roku se začnete setkávat s našimi zubními pastami v pozměněném složení a ve zcela novém designu.
Energy news4 Energy News 04/2010 Inovace 1 Zubní pasty v pozměněném složení a novém designu Od října tohoto roku se začnete setkávat s našimi zubními pastami v pozměněném složení a ve zcela novém designu.
PITSTOP VY_22_INOVACE_26
 PITSTOP VY_22_INOVACE_26 Vzdělávací oblast: Jazyk a jazyková komunikace Vzdělávací obor: Anglický jazyk Ročník: 9. PITSTOP 1/ Try to complete the missing words. Then listen and check your ideas. Eight
PITSTOP VY_22_INOVACE_26 Vzdělávací oblast: Jazyk a jazyková komunikace Vzdělávací obor: Anglický jazyk Ročník: 9. PITSTOP 1/ Try to complete the missing words. Then listen and check your ideas. Eight
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY. Číslo Servisní Informace Mechanika: 5-2013
 Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY Servisní Informace Datum vydání: 20.2.2013 Určeno pro : AMS, registrované subj.pro montáž st.měř. Na základě SI VDO č./datum: Není Mechanika
Mechanika Teplice, výrobní družstvo, závod Děčín TACHOGRAFY Servisní Informace Datum vydání: 20.2.2013 Určeno pro : AMS, registrované subj.pro montáž st.měř. Na základě SI VDO č./datum: Není Mechanika
STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.)
") STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.) ZÁSOBNÍ PROTEINY A KYSELINA ABSCISOVÁ V ZYGOTICKÉ EMBRYOGENEZI HRACHU SETÉHO (PISUM SATIVUM L.) 1 Solnická P., 1 Klemš
STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.) ZÁSOBNÍ PROTEINY A KYSELINA ABSCISOVÁ V ZYGOTICKÉ EMBRYOGENEZI HRACHU SETÉHO (PISUM SATIVUM L.) 1 Solnická P., 1 Klemš
 11.12. 100 ΕΙΣΟΔΟΣ = E / ENTRANCE = E = = 1174 550 ΤΥΠΟΠΟΙΗΜΕΝΟ ΚΥ = 2000 (ΕΠΙΛΟΓΗ: 2100) / CH STANDARD = 2000 (OPTIONAL: 2100) 243 50 ΚΥ/CH + 293 ΚΥ/CH +103 100 ΚΥ /CH 6 11 6 20 100 0,25 ΚΑ (CO) + 45
11.12. 100 ΕΙΣΟΔΟΣ = E / ENTRANCE = E = = 1174 550 ΤΥΠΟΠΟΙΗΜΕΝΟ ΚΥ = 2000 (ΕΠΙΛΟΓΗ: 2100) / CH STANDARD = 2000 (OPTIONAL: 2100) 243 50 ΚΥ/CH + 293 ΚΥ/CH +103 100 ΚΥ /CH 6 11 6 20 100 0,25 ΚΑ (CO) + 45
Nová éra diskových polí IBM Enterprise diskové pole s nízkým TCO! Simon Podepřel, Storage Sales 2. 2. 2011
 Nová éra diskových polí IBM Enterprise diskové pole s nízkým TCO! Simon Podepřel, Storage Sales 2. 2. 2011 Klíčovéatributy Enterprise Information Infrastructure Spolehlivost Obchodní data jsou stále kritičtější,
Nová éra diskových polí IBM Enterprise diskové pole s nízkým TCO! Simon Podepřel, Storage Sales 2. 2. 2011 Klíčovéatributy Enterprise Information Infrastructure Spolehlivost Obchodní data jsou stále kritičtější,
Růst a vývoj rostlin
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
Prostředí je vždy důležité při formování fenotypu
 Evoluce fenotypu IV Prostředí je vždy důležité při formování fenotypu Fenotypová plasticita schopnost organismu vytvářet různý fenotyp v závislosti na vnějších podmínkách (jeden genotyp odpovídá mnoha
Evoluce fenotypu IV Prostředí je vždy důležité při formování fenotypu Fenotypová plasticita schopnost organismu vytvářet různý fenotyp v závislosti na vnějších podmínkách (jeden genotyp odpovídá mnoha
Configuration vs. Conformation. Configuration: Covalent bonds must be broken. Two kinds of isomers to consider
 Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
TechoLED H A N D B O O K
 TechoLED HANDBOOK Světelné panely TechoLED Úvod TechoLED LED světelné zdroje jsou moderním a perspektivním zdrojem světla se širokými možnostmi použití. Umožňují plnohodnotnou náhradu žárovek, zářivkových
TechoLED HANDBOOK Světelné panely TechoLED Úvod TechoLED LED světelné zdroje jsou moderním a perspektivním zdrojem světla se širokými možnostmi použití. Umožňují plnohodnotnou náhradu žárovek, zářivkových
 If there is any inconsistency of weather forecast between Local Weather Station and this unit, the Local Weather Station's forecast should prevail. The trend pointer displayed on the LCD indicates the
If there is any inconsistency of weather forecast between Local Weather Station and this unit, the Local Weather Station's forecast should prevail. The trend pointer displayed on the LCD indicates the
18.VY_32_INOVACE_AJ_UMB18, Frázová slovesa.notebook. September 09, 2013
 1 (Click on the text to move to the section) Worksheet Methodology Sources 2 Decide which words are considered prepositions (předložky) and which are particles (částice) Source: SWAN, Michael a Catharine
1 (Click on the text to move to the section) Worksheet Methodology Sources 2 Decide which words are considered prepositions (předložky) and which are particles (částice) Source: SWAN, Michael a Catharine
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Název společnosti: VPK, s.r.o. Vypracováno kým: Ing. Michal Troščak Telefon: Datum:
 Pozice Počet Popis 1 SCALA2 3-45 A Datum: 2.7.217 Výrobní č.: 98562862 Grundfos SCALA2 is a fully integrated, self-priming, compact waterworks for pressure boosting in domestic applications. SCALA2 incorporates
Pozice Počet Popis 1 SCALA2 3-45 A Datum: 2.7.217 Výrobní č.: 98562862 Grundfos SCALA2 is a fully integrated, self-priming, compact waterworks for pressure boosting in domestic applications. SCALA2 incorporates
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI
 MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
PERSPEKTIVES OF WEGETABLE WASTE COMPOSTING PERSPEKTIVY KOMPOSTOVÁNÍ ZELENINOVÉHO ODPADU
 PERSPEKTIVES OF WEGETABLE WASTE COMPOSTING PERSPEKTIVY KOMPOSTOVÁNÍ ZELENINOVÉHO ODPADU Mach P., Tesařová M., Mareček J. Department of Agriculture, Food and Environmental Engineering, Faculty of Agronomy,
PERSPEKTIVES OF WEGETABLE WASTE COMPOSTING PERSPEKTIVY KOMPOSTOVÁNÍ ZELENINOVÉHO ODPADU Mach P., Tesařová M., Mareček J. Department of Agriculture, Food and Environmental Engineering, Faculty of Agronomy,
SOIL ECOLOGY the general patterns, and the particular
 Soil Biology topic No. 5: SOIL ECOLOGY the general patterns, and the particular patterns SOIL ECOLOGY is an applied scientific discipline dealing with living components of soil, their activities and THEIR
Soil Biology topic No. 5: SOIL ECOLOGY the general patterns, and the particular patterns SOIL ECOLOGY is an applied scientific discipline dealing with living components of soil, their activities and THEIR
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
THE GROWTH OF COTYLEDONARY BUDS OF PEA (Pisum sativum L.)
") THE GROWTH OF COTYLEDONARY BUDS OF PEA (Pisum sativum L.) RŮST DĚLOŽNÍCH PUPENŮ HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav biologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická
THE GROWTH OF COTYLEDONARY BUDS OF PEA (Pisum sativum L.) RŮST DĚLOŽNÍCH PUPENŮ HRACHU (Pisum sativum L.) Mikušová Z., Hradilík J. Ústav biologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická
Tkáňové kultury rostlin. Mikropropagace
 Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
M-LOCK Magnetický zámek pro křídlové sklněné dveře. M-LOCK Magnetic lock for glass swing doors
 M-LOCK Magnetický zámek pro křídlové sklněné dveře M-LOCK Magnetic lock for glass swing doors M-L ock Serratura Magnetica Magnetický zámek M-Lock Přednosti (výhody) 31 mm Extrémně malé rozměry Extrémně
M-LOCK Magnetický zámek pro křídlové sklněné dveře M-LOCK Magnetic lock for glass swing doors M-L ock Serratura Magnetica Magnetický zámek M-Lock Přednosti (výhody) 31 mm Extrémně malé rozměry Extrémně
Czech Republic. EDUCAnet. Střední odborná škola Pardubice, s.r.o.
 Czech Republic EDUCAnet Střední odborná škola Pardubice, s.r.o. ACCESS TO MODERN TECHNOLOGIES Do modern technologies influence our behavior? Of course in positive and negative way as well Modern technologies
Czech Republic EDUCAnet Střední odborná škola Pardubice, s.r.o. ACCESS TO MODERN TECHNOLOGIES Do modern technologies influence our behavior? Of course in positive and negative way as well Modern technologies
Transportation Problem
 Transportation Problem ١ C H A P T E R 7 Transportation Problem The transportation problem seeks to minimize the total shipping costs of transporting goods from m origins (each with a supply s i ) to n
Transportation Problem ١ C H A P T E R 7 Transportation Problem The transportation problem seeks to minimize the total shipping costs of transporting goods from m origins (each with a supply s i ) to n
Číslo projektu: CZ.1.07/1.5.00/34.0036 Název projektu: Inovace a individualizace výuky
 Číslo projektu: CZ.1.07/1.5.00/34.0036 Název projektu: Inovace a individualizace výuky Autor: Mgr. Libuše Matulová Název materiálu: Education Označení materiálu: VY_32_INOVACE_MAT27 Datum vytvoření: 10.10.2013
Číslo projektu: CZ.1.07/1.5.00/34.0036 Název projektu: Inovace a individualizace výuky Autor: Mgr. Libuše Matulová Název materiálu: Education Označení materiálu: VY_32_INOVACE_MAT27 Datum vytvoření: 10.10.2013