Buněčné základy růstu a morfogeneze, signalizace u rostlin fytohormony
|
|
|
- Dominik Bílek
- před 5 lety
- Počet zobrazení:
Transkript


1 Buněčné základy růstu a morfogeneze, signalizace u rostlin fytohormony


2 Růst a vývoj rostlin Zásadní rozdíly ve vývoji rostlin a živočichů Stavba rostlinného těla není předem určená Významný podíl nediferencovaných buněk (meristémy) Přítomnost buněčné stěny brání migraci buněk Poziční určení buněčného osudu Absence buněčných linií Neukončený vývoj
3 Růst a vývoj rostlin Růst nabývání velikosti, objemu, plochy či hmotnosti buněk či celých orgánů Diferenciace funkční rozrůznění buněk či orgánů podle jejich specializace Morfogeneze vývoj tvaru buňky či celé rostliny Vývoj je dán souhrnem procesů růstu a diferenciace, musí být pevně zasazen v čase a orientován prostorově Totipotence schopnost rostlinných buněk regenerovat za vhodných podmínek celou rostlinu. Dediferenciace opětovné nabytí schopnosti buněk realizovat více vývojových programů
, zralosti, stárnutí (senescence) Boyes et al, Plant Cell 13, 1499, 2001")
4 Růst a vývoj rostlin Fenologické fáze: - praktický a experimentální význam - etapy embryonální, juvenilní (vegetativní), zralosti, stárnutí (senescence) Boyes et al, Plant Cell 13, 1499, 2001



5 Růst a vývoj rostlin Růstové fáze: - buněčné dělení - objemový růst - diferenciace Buněčné dělení - v meristémech, neustálý průběh buněčného cyklu apikální meristém prýtu kořene Objemový růst - vrcholový a difúzní typ růstu, orgány s ukončeným a neukončeným typem růstu Diferenciace - specializace na úrovni buněk, pletiv a orgánů

6 Základní rozdíly v dělení živočišných a rostlinných buněk Převzato z přenášky D. Honyse, B130P14: Fyziologie rostlin.


7 Růst a vývoj rostlin Polarita buňky a rostliny dána směrovaným buněčným dělením a směrovaným buněčným růstem...důležitá role cytoskeletu! Před cytokinezí je polarita budoucí buněčné přepážky ustavena prostřednictvím preprofázového svazku mikrotubulů

8 Růst a vývoj rostlin Polarita buňky a rostliny dána směrovaným buněčným dělením a směrovaným buněčným růstem

9 Růst a vývoj rostlin Polarita buňky a rostliny dána směrovaným buněčným dělením a směrovaným buněčným růstem Během růstu buněk: komponenty buněčné stěny ukládány s určitou orientací pohyb sekrečních váčků - aktinový a mikrotubulární cytoskelet Ukládání celulózových mikrofibril: v interakci s cytoskeletem v kortikální cytoplazmě

10 dlouživý růst buněk zvětšuje se plocha buněčné stěny plocha plazmalemy objem vakuoly a plocha tonoplastu bez turgoru buňka neroste význam K + H 2 O syntéza celulózy hemicelulóz a pektinů lipidů proteinů


11 transport pektinů, hemicelulóz a proteinů do buněčné stěny Golgiho aparát syntéza celulózy
 Wolpert (1998)")



12 Orientace buněčného dělení, buněčné vrstvy a morfogeneze Bowman (2001) Wolpert (1998) Principles of Development Uvnitř orgánu kolmý na povrch rovnoběžný s povrchem Vně orgánu Základní aspekt morfogeneze Kontrola roviny buněčného dělení ve vztahu v povrchu rostlinného těla Povrch orgánu Převzato z přenášky D. Honyse, B130P14: Fyziologie rostlin.
 a druhá")
13 Primární apikální meristémy Meristémy Populace malých isodiametrických buněk Věčné trvání embryonálního charakteru meristému Část buněk nenastoupí cestu diferenciace, ale zachovává si dělivou schopnost po celou dobu existence vegetativního meristému iniciály / Kmenové buňky Nediferencované buňky: Po rozdělení si jedna dceřinná buňka zachová charakter iniciály (kmenové buňky) a druhá nastoupí diferenciační dráhu (může po několika dalších děleních) toitipotence iniciály x pluripotence kmnové buňky Převzato z přenášky D. Honyse, B130P14: Fyziologie rostlin, upraveno



14 Iniciace laterálních kořenů Přísná kontrola orientace buněčného dělení během morfogeneze Primordia laterálních kořenů vznikají periklinálním dělením pericyklu



15 Mezibuněčná komunikace Chemické signály Fytohormony (Auxin) du/botany/ s.com Fyzické propojení buněk Plasmodesmy Syncytium

")

16 totipotence rostlinných buněk podmínky úplná genetická výbava aparát schopný ji realizovat regenerace rostlin restituce (náhrada v ráně neobvyklé) 1902 Hypotéza Totipotence rostlinných buněk somatická embrya Gottlieb Haberlandt ( )
 Principles of")
17 Rostliny jsou přisedlé organizmy vysoká schopnost regenerace doplňování a nahrazování ztracených částí těla Většina rostlinných buněk není terminálně diferencovaných Regenerace rostlin z izolovaných somatických buněk Totipotence rostlinných buněk Wolpert (1998) Principles of Development


18 Buněčný cyklus u rostlin Buněčný cyklus sekvence pravidelně se opakujících událostí fáze G1, S, G2, M, G0 kontrolní body G1/S, G2/M 2C 2C dělení růst buňky zvýšení počtu organel a ribozomů replikace DNA syntéza strukturních proteinů jádra interfáze 2C vstup do G 0 nebo příprava na fázi S
19 Buněčný cyklus u rostlin regulace buněčného cyklu CKD cyklin dependentní kinázy zy: kinázy řízené regulačními proteiny- CYKLINY cykliny: vazba na základní katalytickou jednotku a vyskytují se jen v buňkách, které se aktivně dělí mitotické: A, B; G1 fáze: D aktivní komplex CDK-cyklin fosforylace dalších proteinů = reorganizace cytoskeletu a endomembránového systému v příslušné fázi cyklu např. při vstupu do M: fosforylace (inaktivace, destrukce) histonů H1 a laminů = kondenzace chromatinu a rozpad jaderné membrány


 Se vstupem do S-fáze je CDK")
20 Buněčný cyklus u rostlin regulace buněčného cyklu CKD cyklin dependentní kinázy kinázy řízené regulačními proteiny - cykliny fosforylace dalších proteinů = reorganizace cytoskeletu a endomembránového systému v příslušné fázi cyklu další regulace CDK: fosforylace / defosforylace specifickými kinázami / fosfatázami: v G1 fázi je CDK fosforylována v pozici Tyr 15 nebo Thr 14 (i obou) Se vstupem do S-fáze je CDK fosforylována v pozici Thr 160




.")

21 Buněčný cyklus u rostlin regulace buněčného cyklu Degradace cyklinů specifických pro jednotlivé fáze buněčného cyklu je pod kontrolou proteolytického aparátu (komplex ubikvitin ligázy SCF)..cykliny určené k degradaci označeny ubiquitinem Převzato z Buchanan et al 2000




22 Buněčný cyklus u rostlin Endoreduplikace -zmnožení jaderné DNA bez následné cytokineze -diferenciace v G2 fázi (místo G1) -v průběhu vývoje rostlin vyšší čím je orgán starší Převzato z Buchanan et al 2000

23 Co víme po více než sto letech? O typech signálů, které rostlina přijímá? O mechanizmech jejich zpracování? Přednáška B130P14: Fyziologie rostlin Katedra fyziologie rostlin, UK PřF, Jan Petrášek
24 rostliny příjem a přenos signálů přisedlé organizmy mnohobuněčné organizmy nutnost reagovat na okolí koordinace vnitřních procesů potřeba získávat informace z vnějšího i vnitřního prostředí signály: fyzikální (světlo, změny vektorového rozložení mechanických tlaků v pletivech, změny teploty ) chemické (fytohormony, cukry, gradienty iontů, patogeny... ) Většina signálů způsobuje změny genové exprese PŘÍJEM SIGNÁLU RECEPTORY SENSORY obvykle bílkovinného chrakteru - umístěny na membránách (plasmalemě, membránách organel, i v cytoplazmě) - bílkoviny (např. hexokinasa) či bílkovinné komplexy -vnímají jako signál změnu průtoku signální molekuly (např. cukru) -metabolické enzymy, membránové přenašeče

25 rostliny příjem a přenos signálů SLOŽKY PŘENOSU signálu obecné schéma příjmu a přenosu signálu fyziologický signál nebo s. z vnějšího prostředí vnější: světlo, teplota, dotek fyziologický: fytohormony, živiny receptorová kináza, receptor spjatý s G-proteinem signální dráha kinázy, vápník, lipidy, ph odpověď exprese genů, uspořádání cytoskeletu, enzym. aktivita vyhodnocení informace = reakce na signál změny genové exprese změny v organizaci a aktivitách cytoplasmy
 zesílení signálu 2) jeho přenosu 3) změnách růstu a metabolismu Fig. 18.1, 2.: Buchanan et al.")
26 Přenos signálu v rostlinách Typy signálu lze členit dle různých hledisek na externí, interní, biotické a abiotické. Reakce rostliny spočívá v: 1) zesílení signálu 2) jeho přenosu 3) změnách růstu a metabolismu Fig. 18.1, 2.: Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000

27 Přenos signálu v rostlinách Přenos signálu: zachycení receptorem, aktivace druhých poslů vyvolání odpovědi buňky převzato od J. Petráška
28 Přenos signálu v rostlinách Signál míra informace v čase či prostoru Reakce rostliny na signál je závislá na: vývojovém stavu předchozích vlivech prostředí vnitřních biologických hodinách, které specifikujíčas v rámci roku a dne!! DůleD ležitost časování odpovědi di na signál příkladu klíčení semene - pokud je špatně načasováno, nelze zvrátit Zea mays, mutace v prekurzoru ABA klíčící semena na mateřské rostlině
29 Přenos signálu v rostlinách Konečná reakce rostliny (resp. buňky) bývá projevem integrace různých signálních informací kooperace mezi dvěma či více druhy signálů u rostlin např. známa v případě signalizace červeným a modrým světlem vnímání gravitace a světla světla a dostupnosti nitrátů kooperace růstových regulátorů (fytohormonů) různých minerálních látek Existence společných mechanismů pro více signálních drah

30 Příjem signálu v rostlinách - receptory 1) pomocí receptorů v plazmatické membráně / cytosolu / jádře / ER Taiz and Zeiger 2010

31 Příjem signálu v rostlinách - receptory 2) vlastní změnou membránového potenciálu, která otevírá iontové kanály Fig : Buchanan et al., Biochemistry and Molecualr Biology of Plants, 2000 převzato od J. Petráška

32 Úrovně signalizace v rostlinném těle Komplexita regulace signálních drah: složité vztahy na úrovních: druhých poslů metabolických reakcí přestavby podpůrných struktur cytoskeletu membránového systému Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000



33 Příjem signálu v rostlinách - receptory Receptory v plazmatické membráně jsou u rostlin pravděpodobně analogické živočišným receptorům (ale důkazy pro všechny typy nejsou prozatím úplné) A receptor vázanv zaný na trimerický G protein B - receptor vázanv zaný na enzym C receptor vázanv zaný na iontový kanál SIGNÁLN LNÍ DRÁHA: Sled reakcí, které vedou od receptoru aktivovaného signálem k buněčné reakci Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000
34 Přehled důležitých signálních drah u rostlin Fosfatidyl inositolový signáln lní systém Fosfolipáza C enzym vázaný na plasmalemu často aktivována G-proteinem štěpení fosfatidylinositol 4,5 bisfosfátu (PIP2) inositol 1,4,5 trisfosfát (IP3) diacyl glycerol (DAG) IP3 a DAG fungují jako druzí poslové Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000
 mění její citlivost k Ca 2+ (PKC působí v mnoha signálních drahách spojených s dělením a")
35 Přehled důležitých signálních drah u rostlin Fosfatidyl inositolový signáln lní systém IP 3 aktivuje Ca 2+ kanály v membránách endoplazmatického retikula výsledný signál je zprostředkován přes kalmodulin-dependentní protein kinázy DAG zůstává vázaný na membránu může aktivovat proteinkinázu C (PKC) mění její citlivost k Ca 2+ (PKC působí v mnoha signálních drahách spojených s dělením a růstem buněk) Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000
36 Přehled důležitých signálních drah u rostlin Protein kinázové kaskády nejrozšířenějším způsobem regulace genové exprese Výsledkem kinázové kaskády je aktivace transkripčních faktorů (TF) v cytoplazmě či přímo v jádře. MAPK - protein kinázy aktivující mitogeny = transkripční faktory Ca2+/ kalmodulinové kinázy po reakci s Ca2+/ kalmodulinem aktivují TF návazné fosforylace slouží k zesílení signálu a případné další regulaci Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000
37 Přehled důležitých signálních drah u rostlin Řízená proteolýza signálních molekul, např. transkripčních faktorů. Protein určený k degradaci je označen ubiquitinem ve třech následných krocích: 1) Aktivační enzym E1 za spotřeby ATP aktivuje ubiquitin 2) konjugační enzym E2 ho dále napojí na komplex ubiquitin ligázy E3 3) na komplexu E3 je již vázán protein určený k degradaci a dochází k vazbě ubiquitinu Degradace proteinů následně probíhá v proteasomu ubiquitin 76 AMK protein, vysoce konzervovaný
38 Přehled důležitých signálních drah u rostlin Některé receptory fytohormonů jsou součástí ubiquitinové proteolytické dráhy (př. auxin, giberelin, k. jasmonová) v jádře: část vazebného místa IAA!přenos signálu v rostlinách často představuje INAKTIVACI REPRESORŮ část vazebného místa IAA komplex E3 ligázy negativní regulace se ukázala podle mat. modelů jako rychlejší komplex E3 ligázy a AUX/IAA represoru - ubiquitinace x živočichové spíš pozitivní (aktivační) signální dráhy AUX/IAA represor degradace 26s proteasomem
39 Rostlinná neurobiologie Nově formovaná oblast rostlinné fyziologie, která si klade za cíl studovat podstatu adaptivního chování rostlin. Alternativní název Sensorická biologie rostlin či biologie vnímání rostlin. Sdružuje více pohledů: Oblasti studia: - buněčná a molekulární biologie - elektrofyziologie - ekologie - anatomie, adaptace, signalizace během symbiózy a reakce na patogeny, rostlinná imunita, paměť a učení, komunikace mezi rostlinami, rostlinná inteligence Prezentace na příště... Jméno rostlinná neurobiologie podle Brenner et al Rostliny odpovídají na okolní svět změnami v morfologii, pohyby, reagují na elektrické podněty, přenášejí akční potenciál, tvoří signální molekuly a ovlivňující chování jiných organismů Brenner E, et al (2006) Plant neurobiology: an integrated view of plant signaling. Trends Plant Sci 11:

40 FYTOHORMONY Santner A. et al, Nature Chemical Biology 5, (2009)
41 Fytohormony Julius von Sachs ( ) hypotéza chemických signálů (morfogenů) Signální molekuly s růstově regulační funkcí Základních 6 skupin - auxiny - cytokininy - gibereliny - kyselina abscisová - etylén - brasinosteroidy Další látky s růstovr stově-regulační funkcí: - polyaminy - kyselina jasmonová - kyselina salicylová - fenolické látky - peptidové hormony - deriváty karotenoidů (strigolaktony) Santner et al. 2009, Nature Chemical Biology 5 (5),
42 Fytohormony Signální molekuly v podobě organických nízkomolekulárních látek Přítomné v nízkých koncentracích (10-6 až 10-9 M) Při vyvolání růstové či vývojové odpovědi se samy chemicky nemění Vývojové procesy jsou zajištěny změnou koncentrace určitého typu fytohormonu a současně modulací citlivosti pletiv k němu Působí v místě svého vzniku nebo i v jiných částech rostliny -transport! Spolupůsobení faktorů okolního prostředí (světelné a tepelné záření, gravitace, elektrická pole) Podrobněji o fytohormonech pojednává semestrální přednáška KEBR PřFUK Fytohormony B130P15
43 Analogie živočišných a rostlinných hormonů sporná Velmi obecný mechanismus účinku je společný: Biosyntéza HORMON Metabolismus Receptor Aktivace výkonných struktur (systém druhých poslů) a případné následné ovlivnění genové exprese Živočišné hormony: - syntéza ve specializovaných tkáních - transportovány na přednostní místo spotřeby - vyvolávají relativně specifickou fyziologickou odpověď - působí v úzké koncentrační škále mezi 10-8 až M Rostlinné hormony: - není jasně patrný rozdíl mezi místem biosyntézy a spotřeby - mají značně pleiotropní (různorodé) účinky (mohou vyvolávat kvalitativně rozdílné účinky v různých pletivech, ale v závislosti na koncentraci i ve stejném pletivu) - koncentrační rozmezí širší mezi 10-6 až 10-9 M převzato od J. Petráška

44 Syntetické látky s růstově-regulační funkcí chemicky připravené látky se strukturou podobnou fytohormonům -váží se na receptory pro fytohormony -vyvolávají růstovou odpověď rostliny -mohou mít společný mechanismus -mohou narušovat transport konkrétního hormonu - často stabilnější, lépe aplikovatelné použití v zahradnictví, agronomii PREZENTACE Commercial Uses of Gibberellins prezentace
45 Nároky na fytohormony...aneb čím izolovanější buňka je, tím závislejší se stává na tom co jí dodáme zvenčí Rostlina Zvýšení nároků na exogenní fytohormony Buňka Kalus, buněčná kultura Protoplast Protoplastová kultura Snížení nároků na exogenní fytohormony Buňka Organogeneze, embryogeneze Rostlina převzato od J. Petráška
 Kögl (Holandsko) 1933 izoloval tuto látku z lidské moči a následně identifikoval jako kyselinu indolyl-3-octovou (IAA) teprve 1946 potvrzena ve vyšších rostlinách kyselina")
46 Auxiny historie objevu Nejdéle známé fytohormony Darwin 1880, Paal 1919, Went 1928 v pokusech s koleoptilí ovsa prokázali existenci účinné látky (izolace z fermentační směsi) podporující prodlužování buněk (auxein=růst) Kögl (Holandsko) 1933 izoloval tuto látku z lidské moči a následně identifikoval jako kyselinu indolyl-3-octovou (IAA) teprve 1946 potvrzena ve vyšších rostlinách kyselina indolyl-3-octová

47 Auxiny historie objevu F.W. Went, , koleoptilový test na auxiny
48 Auxiny - chemické struktura endogenní auxiny IAA IBA 4-Cl-IAA Kyselina indolyl-3-octová Kyselina indolyl-3-máselná Kyselina 4-chlor-indolyl-3-octová syntetické auxiny Kyselina fenyloctová Kyselina 2,4-dichlorfenoxyoctová Kyselina 1-naftyloctová PAA 2,4-D NAA převzato od J. Petráška


 Biosyntéza")

49 Auxiny - biosyntéza Biosyntéza hlavně v mladých apikálních pletivech a rychle se dělících buňkách tj. meristémech, embryu, vyvíjejících se plodech 1) Biosyntéza vycházející z L- tryptofanu: 2) Biosyntéza nezávislá na L- tryptofanu: - Prekurzorem indol či indolylglycerol-fosfát Taiz and Zeiger 2002
50 Auxiny - metabolismus Konjugace IAA: - kovalentní vazba nízkomolekulárních látek - je reverzibilní - konjugované formy IAA jsou neaktivní (mohou se hromadit) - funguje jako rychlý regulační mechanismus - glukosylestery, myoinositol estery, konjugáty s aminokyselinami (aspartát) Funkce konjugátů: - zásobní pool auxinu (klíčící semene), až 50-90% celkového obsahu IAA - možné rychlé uvolnění aktivní formy (rychlejší než syntéza) - detoxifikační - transportní - ochrana před degradací





51 Polární transport auxinů regulační funkce Chemiosmotická teorie transportu auxinu z buňky do buňky: Odlišná prostupnost IAA a IAA - přes plazmatickou membránu IAA může procházet membránou IAA- potřeba přenašeče - Disociace molekul IAA vyšší v cytosolu (vyšší ph oproti apoplastu) -IAA - je zachytávána v cytoplasmě v tzv. aniontové pasti, Polarita toku je dána polárním umístěním přenašečů
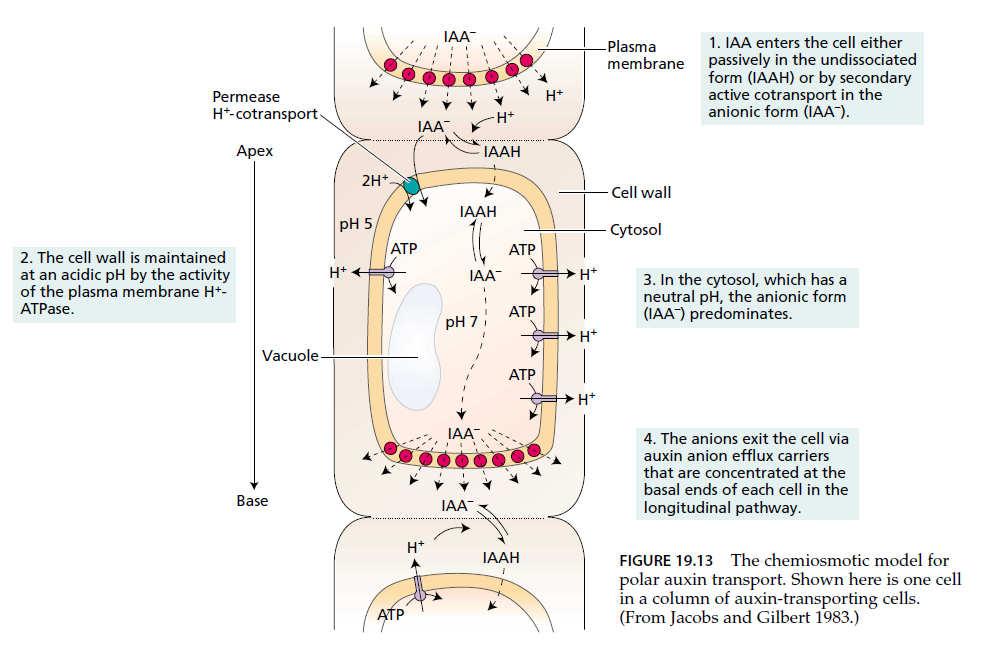
52



53 Polární transport auxinů regulační funkce Rostlina Směr toku auxinu Kořen Přenašeč auxinu ven z buňky Přenašeč auxinu do buňky Parenchymatické buňky protofloému (Podle Grebe, BioEssays 26, 719, 2004) převzato od J. Petráška

 lokalizace PIN1 proteinu na bazální straně buněk Petrášek and Friml, 2009, Development 136,")
54 Polární transport auxinů přenašeče auxinu přenašeče auxinu do buňky AUX1/LAX rodina transmembránových proteinů, podobné aminokyselinovým permeázám pin1 mutant Arabidopsis převzato z Taiz and Zeiger 2002 přenašeče auxinu z buňky PIN ABC přenašeče - rodina proteinů PIN1 - PIN8 (Arabidopsis) (ATP binding casette) lokalizace PIN1 proteinu na bazální straně buněk Petrášek and Friml, 2009, Development 136,
 - regulace samotnými")
55 Přenašečové molekuly podléhají konstitutivnímu cyklování Přenašečové molekuly cyklují mezi plasmatickou membránou a endomembránovým systémem: - regulace inhibitory transportu auxinu (PBA, TIBA) - regulace samotnými auxiny

56 Auxiny mechanismus účinku Auxinem stimulovaná aktivace H + -ATPázy: - umožňuje snižovat ph buněčné stěny a rozvolňovat tak její strukturu, důsledkem je tzv. kyselý růst převzato od J. Petráška

57 Auxiny fyziologické účinky Stimulace prodlužovacího růstu segmenty koleoptilí ovsa H 2 O IAA
58 Auxiny fyziologické účinky Regulace tropismů - pozitivní gravitropismus kořene - negativní gravitropismus stonku Gravitropismus kořene ohyb dolů: - hromadění auxinu na spodní straně, kde je taková koncentrace inhibiční pro buněčnou elongaci Gravitropismus stonku ohyb nahoru: - hromadění auxinu na spodní straně, kde je taková koncentrace aktivační pro buněčnou elongaci Friml 2003, Current Opinion in Plant Biology 6, 1-6 Paciorek et al. 2005, Nature 435 (7046), převzato od J. Petráška
 účinky auxinu Paciorek et al.")
59 Auxiny fyziologické účinky odlišná citlivost a odpověď k auxinu Regulace tropismů - pozitivní gravitropismus kořene - negativní gravitropismus stonku stonek pupen kořen pleiotropní (různorodé) účinky auxinu Paciorek et al. 2005, Nature 435 (7046), Friml 2003, Current Opinion in Plant Biology 6, 1-6


60 Auxiny fyziologické účinky Regulace tropismů fototropismus stonku: - hromadění auxinu na od světla odvrácené straně, kde je taková koncentrace aktivační pro buněčnou elongaci - zřejmě souvisí se signalizací prostřednictvím fosforylace receptoru modrého světla fototropinu DR5::GUS světlo převzato od J. Petráška
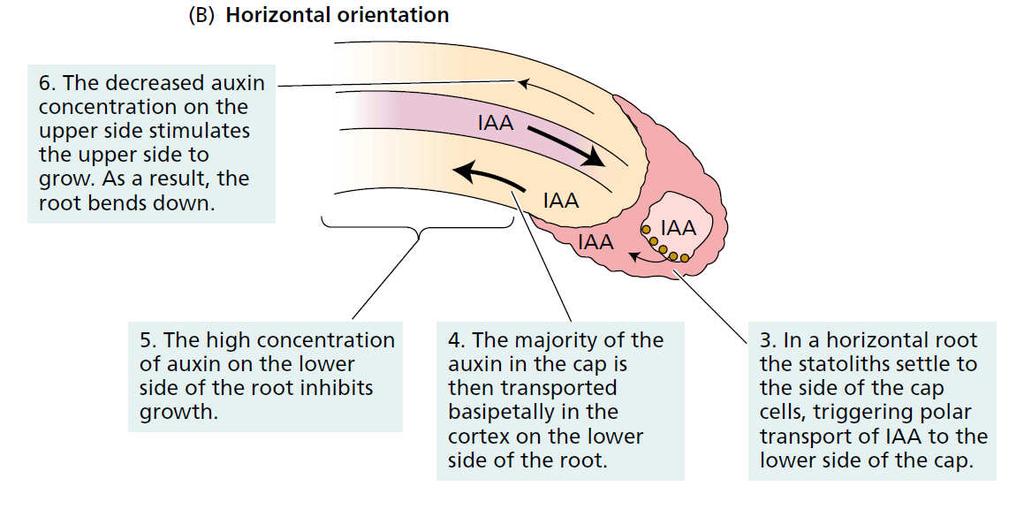

61



62 Auxiny fyziologické účinky Udržování apikální dominance vzrostný vrchol produkcí auxinu a jeho transportem bazipetálně udržuje svou dominanci odstranění apikálního meristému uvolňuje inhibici růstu bočních pupenů boční (axilární) pupeny
63 Auxiny fyziologické účinky Regulace embryogeneze, zakládání laterálních orgánů Tanaka et al.,.cell. Mol. Life Sci. 63, , 2006 převzato od J. Petráška






64 Auxiny fyziologické účinky Podpora tvorby hlíz či plodů, stimulace zakořeňování normální plod odstranění nažek po odstranění nažek, stříkáno auxinem zbytnělé receptákulum nažka růst receptákula je stimulován auxinem produkovaným v nažkách Callis 2005, Nature 435, převzato od J. Petráška
65 Cytokininy historie objevu Haberlandt 1913 látka v cévních svazcích stimulující buněčné dělení Overbeek 1940 látka v mléčném endospermu z nezralých ořechů kokosu Skoog 1950 pokusy s in vitro kulturami tabáku, stimulace dělení po aplikaci částí vaskulárního pletiva na explantát, úspěch i s kvasničným extraktem a autoklávovanou DNA Miller 1954 izolace účinné látky z klávované DNA slanečka, pojmenována kinetin první syntetický cytokinin Letham 1963 první přirozeně se vyskytující se cytokinin (zeatin) izolován z nezralého endospermu kukuřice převzato od J. Petráška


66 Cytokininy chemická struktura N 6 -substituované deriváty adeninu podle postranního řetězce se dělí na isoprenoidní a aromatické Sakakibara, Annu Rev Plant Biol , 431 převzato od J. Petráška
67 Cytokininy biosyntéza Isopentenyl transferáza (IPT) agrobakterium Vychází z isopentenyl pyrofosfátu (= AMAPP dimetylalyl difosfát) rostliny + adenosin fosfátu ATP / ADP (AMP) probíhá hlavně v kořenech, embryích, mladých listech a plodech, všude tam, kde se buňky dělí -signalizace na krátkou i dlouho vzdálenost -transport xylémem převzato od J. Petráška

68 PREZENTACE Vývoj tumoru a role etylénu při jeho morfogenezi
 a cytokininu")

69 Cytokininy fyziologické účinky Stimulace buněčného dělení v explantátových kulturách Poměr hladin auxinu (A) a cytokininu (CK) určující pro organogenezi: -vyrovnaný poměr kalus Převaha auxinu Převaha cytokininu model organogeneze in vitro Skoog a Miller 1957


70 Cytokininy fyziologické účinky Stimulace buněčného dělení v meristémech, narušení apikální dominance. Rostlina tabáku exprimující ipt gen z agrobakteria zvýšení syntézy cytokininů - účinek podobný exogenní aplikaci: narušena apikální dominance aktivita laterálních meristémů Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 intaktní apex apikální meristém Odstranění apexu prorůstání úžlab. pupenu úžlabní pupen IPT izopentenyl transferáza (CK biosyntéza) CKX cytokinin oxidáza (CK degradace) auxin inhibice IPT


71 Cytokininy fyziologické účinky Zpomalení stárnutí rostliny tabáku exprimující gen ipt pod kontrolou promotoru indukovaného senescencí jsou nesmrtelné. Cytokininy stimulují tvorbu chlorofylu. ovlivnění vývoje chloroplastů Za tmy: etioplasty (prolamelární těleso) ošetřeno cytokininy vývoj thylakoidů Chory et al. 1994, courtesy of J. Chory, American Society of Plant Biologists Taiz and Zeiger 2002
72 Cytokininy fyziologické účinky cytokininy podporují redistribuci živin a asimilátů v rostlině -mění vztahy zdroj / sink -podílejí se (spolu s auxinem) na poměru vývoje kořenů / prýtu -též v závislosti na obsahu živin v daném orgánu radioaktivně 14 C značená aminokyselina kontrola (ošetřeno vodou) neošetřeno 50mM kinetinc neošetřeno neošetřeno 50mM kinetin

 produkce vlastních cytokininů")


73 Cytokininy fyziologické účinky regulační funkce fytohormonů může být zneužita i pateogenními organismy (bakterie, hmyz, nematoda) produkce vlastních cytokininů nebo stimulace jejich produkce samotnou rostlinou kořenové tumory - Agrobacterium prolomení dormace pupenů čarověníky Agrobacterium tumefaciens. Corynebacterium fascians - aktinomyceta
74 Gibereliny historie objevu a chemická struktura 30. léta 20. století Kurosawa Japonsko, choroba rýže vyvolaná houbou Gibberella fujikuroi (Fusarium moniliforme v imperfektním stadiu), vyvolává dlouživý růst a následné poléhání rostlin 50. léta 20. století kyselina giberelová (GA 3 ) identifikována v extraktu z houby a následně nalezena též ve vyšších rostlinách Dnes známo okolo 130 různých forem giberelinů Jsou to tetracyklické diterpeny, slabé organické kyseliny, základem tetracyklický ent-giberelanový skelet Dělení podle počtu atomů uhlíku na gibereliny C 19 a C 20, řazení dle čísel podle posloupnosti objevů. převzato od J. Petráška


75 Gibereliny historie objevu a chemická struktura První objevené a současně nejaktivnější jsou gibereliny G 19 - GA 1, GA 3, GA 4 a GA 7. Aktivita ovlivněna přítomností karboxylových skupin v poloze 7 a 19 a přítomností hydroxylových skupin. převzato od J. Petráška
 Inaktivace nejčastěji hydroxylací v poloze 2. či reverzibilní konjugací s cukry. Taiz, Zeiger, Plant Physiology online převzato od J.")
76 Gibereliny biosyntéza a metabolismus Fáze 1 v plastidech: Až po ent-kauren. Fáze 2 v ER: Od ent-kaurenu až po aldehyd GA 12. Fáze 3 v cytoplazmě: Vzájemné přeměny a inaktivace giberelinů. Jsou pod vývojovou kontrolou (světlo, teplota) Inaktivace nejčastěji hydroxylací v poloze 2. či reverzibilní konjugací s cukry. Taiz, Zeiger, Plant Physiology online převzato od J. Petráška

77


78 Gibereliny fyziologické účinky Stimulace prodlužovacího růstu stonku spojena s okyselením buněčné stěny a aktivitou hydroláz - též podpořeno buněčné dělení v meristémech. Elongace listové pochvy u zakrslých semenáčků rýže po aplikaci GA 3 Elongace stonku u trpasličího semenáčku hrachu po aplikaci GA 3 Kontrola 100 pg GA 3 1 ng GA 3 Kontrola Figs , 17.7.: Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 GA 3 převzato od J. Petráška

 Kontrola Paclobutrazol")
79 Gibereliny fyziologické účinky Stimulace prodlužovacího růstu stonku Aplikace inhibitorů cytochrom P450 monooxygenáz sníží množství giberelinů v rostlině a způsobí cílenou zakrslost, praktické využití v zemědělství Poinssetia netransformovaná rostlina transrormace cdna pro giberelin-2-oxidázu (degradace GA 1 ) Kontrola Paclobutrazol inhibitor syntézy GA

80 Gibereliny fyziologické účinky Stimulace kvetení navození kvetení v jinak neinduktivních podmínkách: exogenní dodání giberelinu umožní obejít jarovizaci Gibereliny jsou nezbytné při stimulaci kvetení u mrkve požadující tzv. jarovizaci. Chlad indukuje tvorbu giberelinů. Pro indukci kvetení existují 4 vzájemně propojené dráhy indukované těmito 4 stimuly: Fotoperioda přes FT protein Počet listů a nízká teplota Sacharóza Gibereliny -GA, bez chladu +GA, bez chladu -GA, v chladu Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 převzato od J. Petráška
 14 týdenní sekvoj aplikace GA4/GA7 aplikace")
81 Gibereliny fyziologické účinky gibereliny regulují přechod z juvenilní do dospělé fáze jehličnany dlouhá juvenilní fáze (20let) 14 týdenní sekvoj aplikace GA4/GA7 aplikace GA4/GA7

82 Gibereliny fyziologické účinky Stimulace klíčení semene gibereliny nahromaděny v embryu vyvíjejícího se semene na počátku klíčení se transportují do aleuronové vrstvy buněk stimulace exprese enzymu α-amylázy hydrolyzující škrob vzniklé cukry poté stimulují prodlužování buněk hypokotylu klíčícího embrya Taiz, Zeiger, Plant Physiology online
83 Kyselina abscisová historie objevu a chemická struktura 60. léta 20. století látka identifikována nezávisle při studiu dormance u javoru jako dormin (navozující dormanci) současně při studiu opadu mladých plodů bavlny jako abscisin (navozující opad) (dnes je úloha při opadu přisuzována etylénu, ne ABA) Kyselina abscisová je seskviterpen, přirozeně se vyskytuje v podobě několika isomerů, aktivní je pouze (S)-ABA. převzato od J. Petráška

84 Kyselina abscisová - biosyntéza a metabolismus Biosyntéza z karotenoidu violaxantinu (C40) Tvorba: v dormantních orgánech (pupeny, semena hlízy) v rychle rostoucích pletivech Transport floémem i xylémem. Obsah stoupá při nedostatku vody, zasolení, chladu či poranění. ABA představuje hormon odpovědi na stres Konjugace ABA s glukózou inaktivace ve vakuole

85 Kyselina abscisová fyziologické účinky Regulace vodního hospodářství snížením transpirace při nedostatku vody. Při nedostatku vody stoupá biosyntéza ABA v kořenech transport xylémem do nadzemníčásti svěracích buňkách průduchů dochází ke zvýšení výtoku iontů K + a uzavření průduchů Kontrola ABA Commelina communis, pokožkové buňky s průduchy Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 převzato od J. Petráška

86 Kyselina abscisová fyziologické účinky ABA signalizace při odpovědi na vodní stres koncentrace ABA ve xylému se při vodním stresu mění až ze 10nM to 3000nM
 transportujících anionty do apoplastu výsledná")
87 Kyselina abscisová fyziologické účinky Regulace vodního hospodářství snížením transpirace při nedostatku vody. ABA aktivuje Ca 2+ kanály v plazmatické membráně vzestup Ca 2+ otevře další Ca 2+ kanály na tonoplastu a indukuje aktivitu aniontových kanálů (Cl - ) transportujících anionty do apoplastu výsledná depolarizace membrány podpoří aktivitu kanálů transportujících K + ionty ven z buňky. Zmenšující se obsah iontů v buňce vede ke ztrátě vody a uzavírání průduchu Julian Schroeder et al.
88 Kyselina abscisová fyziologické účinky Regulace embryogeneze a zrání semen Hromadění ABA předchází vlastní dehydrataci semena Indukce proteinů LEA (late embryogenesis abundant) - mají roli v odolnosti proti desikaci - současně ABA stimuluje produkci zásobních proteinů v semeni Indukce dormance semen a pupenů, inhibice klíčení - hromadění ABA v dormantních orgánech V semenech vysoká hladina ABA brání klíčení, které nastane až po snížení její hladiny. Mutantní rostliny s poruchou biosyntézy ABA mají narušenou dormanci a projevují viviparii semen semena klíčí ještě na mateřské rostlině. Zea mays, mutace v prekurzoru ABA Taiz, Zeiger, Plant Physiology online převzato od J. Petráška
89 Kyselina abscisová fyziologické účinky Zvýšení odolnosti proti suchu, zasolení, chladu či poškození ABA působí na expresi hydrofilních, termostabilních proteinů ochraňujících strukturu biomembrán. Stimulace tvorby laterálních kořenů reakce na nedostatek vody. Inhibice prodlužovacího růstu a urychlení stárnutí. převzato od J. Petráška
90 Etylén historie objevu a chemická struktura 1901 Dimitrij Neljubov, popsal aktivní složku svítiplynu: etylén, která indukovala v blízkosti plynových lamp opad listů 30. léta 20. století důkaz tvorby ethylenu rostlinami, v té době již využíván na defoliaci a dozrávání plodů Jediný plynný fytohormon Nejjednodušší uhlovodík H 2 C=CH 2 převzato od J. Petráška

91 Etylén - biosyntéza a metabolismus Biosyntéza z L-methioninu přes S-adenosylmethionin (SAM) a kyselinu 1-aminocyklopropan-1- karboxylovou (ACC) Oxidací ACC vzniká etylén. Biosyntéza ACC je regulována dalšími fytohormony auxiny, cytokininy, brasinosteroidy též stresovým působením Produkce etylénu téměř všemi částmi rostlin, nejvíce v rostoucích částech Největší produkce během zrání, stárnutí a opadu plodů a květů a při stresu.

 - uzavírání háčku hypokotylu - ztráta")

92 Etylén fyziologické účinky Trojná odezva v reakci na aplikaci etylénu: - inhibice dloužení - stimulace radiálního růstu (reorganizace mikrotubulů potažmo celulóz. mikrofibril) - uzavírání háčku hypokotylu - ztráta gravitropismu semenáčky salátu stimulace tvorby kořenového vlášení kontrola + etylén vzduch etylén Semenáčky hrachu převzato od J. Petráška

93 Etylén fyziologické účinky Stimulace dozrávání plodů aktivace hydrolytických enzymů Rajče nesoucí antisense- ACC-oxidázu wt ihnibice syntézy etylenu Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000



94 Etylén fyziologické účinky Stimulace dozrávání plodů aktivace hydrolytických enzymů u některých druhů: jablka, rajčata, hrušky, banány, avokádo, fíky, mango, broskve - zvýšená intenzita tzv. klimakterického dýchání Taiz and Zeiger, 2002 zpětná vazba autokatalyticky stimulovaná syntéza etylénu










95 Etylén fyziologické účinky Stimulace opadu listů u stárnoucích listů s poklesem hladiny auxinu dochází ke zvýšené tvorbě etylénu v opadové zóně bříza WT transformant necitlivý k etyléu...ale supraoptimální dávky auxinu stimulují produkci etylénu Agent Orange defoliant na bázi auxinu použitý ve vietnamské válce
96 Brasinosteroidy historie objevu a chemická struktura 1979 první rostlinný steroidní regulátor brassinolid (BR) izolován z pylu řepky (z 227 kg pylu 4mg brassinolidu) 1982 kastasteron izolován z kaštanu Dnes popsáno více než 40 příbuzných látek Jsou to rostlinné steroidní látky, přítomné ve velmi nízkých koncentracích M převzato od J. Petráška
97 Brasinosteroidy biosyntéza a metabolismus Vychází z izopentenyldifosfátu, První biologicky aktivní je kampesterol Nejsou transportovány na dlouhé vzdálenosti, biosyntéza hlavně v nadzemní části
 dané zvětšováním buněk na adaxiální straně Kontrola před testem Aplikace BR Fig. 17.")
98 Brasinosteroidy fyziologické účinky Stimulace dlouživého růstu např. odklonění čepele listu rýže (lamina inclination test) dané zvětšováním buněk na adaxiální straně Kontrola před testem Aplikace BR Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000

99 Brasinosteroidy fyziologické účinky Stimulace dlouživého růstu mutanty s postiženou biosyntézou brassinolidu jsou zakrslé zakrslý vzrůst je dán nejen menší velikostí ale i menším počtem buněk listu stimulace buněčného růstu: indukce enzymů rozvolňujících b.s. stimulace příjmu vody akvaporiny wt det2 dim většinou účinkují nedaleko místa syntézy externě dodané transportovány xylémem transpiračním proudem z listů transport ojedinělý stimulace růstu pylové láčky interakce s etylénem podporují jeho produkci zvyšují odolnost proti suchu a nízkým teplotám Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 převzato od J. Petráška
100 Brasinosteroidy fyziologické účinky kontrola dlouživého růstu záleží na uspořádání mikrotubulů (a celulózových mikrofibril) huseníček: wild type mikrotubuly zeleně -BR +BR exogenně BR deficientní mutant
101 Polyaminy Studovány u eukaryotních organismů již přes 50 let, v rostlinách nejčastěji putrescin, spermin a spermidin. Biosyntéza 1.aminokyselin argininu a ornitinu 2.S-adenozylmethioninu (SAM) jeho dekarboxylací SAM dekarboxylázou. Nejvíce se vyskytují v meristémech. Stimulace klíčení, růstu kalusu, somatické embryogeneze atd. Díky polykationtovému charakteru se váží na DNA, ovlivňují její konformaci a regulují genovou expresi Kusano et al. 2008, Planta
 pleiotropní účinky u rostlin bramboru: malý vzrůst, malé listy, předčasná senescence. wt Antisense SAM Fig. 17.71.: Buchanan et al.")
102 Polyaminy v rostlinách se nacházejí ve vyšších koncentracích než základní fytohormony je třeba i vyšších koncentrací (mmol) pro vyvolání odezvy Zablokování exprese SAM dekarboxylázy (narušení biosyntézy) pleiotropní účinky u rostlin bramboru: malý vzrůst, malé listy, předčasná senescence. wt Antisense SAM Fig : Buchanan et al., Biochemistry and Molecular Biology of Plants, 2000 převzato od J. Petráška
 Methylester Ja (MeJa) převzato od J.")
103 Kyselina jasmonová (jasmínová) Poprvé u rostlin izolována z esenciálních olejů Jasminum grandiflorum Biosyntéza z kyseliny linolenové hladina Ja stoupá při stresových reakcích po poranění či napadení patogenem Stimulace genů pro inhibitory proteáz produkovaných patogeny. Kyselina jasmonová (Ja) Methylester Ja (MeJa) převzato od J. Petráška
 zvyšujících rezistenci k virovému, houbovému či bakteriálnímu napadení SA se")
104 Kyselina salicylová Kyselina salicylová (SA) byla jako aktivní látka izolována již v 19 století z vrbové kůry, její hlavní využití je v medicíně jako analgetikum (aspirin). U rostlin je účinek SA především spojen s ochranou před patogeny, SA indukuje tvorbu proteinů (pathogenesis-related) zvyšujících rezistenci k virovému, houbovému či bakteriálnímu napadení SA se účastní na hypersensitivní reakci (nekrozy napadených listů) a systemické rezistenci vůči patogenům indukce termogenního procesu v květech Sauromatum guttatum
105 Fenolické látky Velmi široká skupina sekundárních metabolitů růstově regulační funkci mají jen některé. Účastní se např. při inhibici dlouživého růstu (kyselina skořicová) či při zprostředkování infekce půdními bakteriemi (Agrobacterium tumefaciens) u dvouděložných (syringon, acetosyringon). převzato od J. Petráška

106 Peptidové hormony Příkladem peptidového hormonu (faktoru) je systemin (18 AMK) role při odpovědi na poranění patogenem transportován floémem do míst, kde ještě poranění není spouští syntézu JA, která vyvolá tvorbu ochranných inhibitorů proteáz převzato od J. Petráška
")


107 Strigolaktony Strigolaktony terpenoidní látky odvozené od karoteonidů Stimulují klíčení parazitických rostlin (Striga tropická fotosyntetizující, zárazy - nezelené) Stimulují prorůstání mykorhizních hub signál pro jejich větvení Hrách setý Orobanche vulgaris Striga Inhibují větvení stonku rostlin Transportovány z kořenů do nadzemních částí Umehara et al., Gomez-Roldan et al., Nature 455, 2008 wt Biosyntetický mutant převzato od J. Petráška
 a cytokininu (CK) určující pro organogenezi: -převaha A kořeny -vyrovnaný poměr kalus, -převaha CK diferenciace")
108 Interakce hormonálních signálů - příklady Auxiny-cytokininy Stimulace buněčného dělení v explantátových kulturách Poměr hladin auxinu (A) a cytokininu (CK) určující pro organogenezi: -převaha A kořeny -vyrovnaný poměr kalus, -převaha CK diferenciace prýtu
 je pod stálou kontrolou více hormonálních faktorů -")

109 Interakce hormonálních signálů -příklady Auxiny-cytokininy vývoj laterálních orgánů (postranních kořenů, listů, květů) je pod stálou kontrolou více hormonálních faktorů - zřejmě se účastní i strigolaktony - jsou transportovány vzhůru a jejich účinek je nadřazen auxinovému a cytokininovému účinku Leyser, Plant Cell Environment, 2009 převzato od J. Petráška

110 Interakce hormonálních signálů - příklady ABA-Gibereliny alfa amyláza blokována ABA, - stimulována GA Dodání ABA působí inhibičně na celý proces indukce klíčení giberelinem: na transkripci a syntézu α-amylázy na její sekreci z aleuronových buněk do endospermu Působení GA je v přímém protikladu stimuluje α-amylázy a následné rozvolnění endospermu pomocí hydroláz převzato od J. Petráška

111 Děkuju za pozornost Taiz, Zeiger, Plant Physiology online
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy
 Fyziologie rostlin, LS 2013 Přednáška 5 Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy Jan Petrášek, PřFUK Signalizace - dráždivost Bohumil Němec: Vztahy rostlin k vnějšímu světu,
Fyziologie rostlin, LS 2013 Přednáška 5 Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy Jan Petrášek, PřFUK Signalizace - dráždivost Bohumil Němec: Vztahy rostlin k vnějšímu světu,
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
CZ.1.07/1.1.00/
 Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
2012/2013. Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol.
 2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Vznik dřeva přednáška
 Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
Vznik dřeva přednáška strana 2 2 Rostlinné tělo a růst strana 3 3 Růst - nejcharakterističtější projev živých organizmů - nevratné zvětšování hmoty či velikosti spojené s činností živé protoplazmy - u
Průduchy regulace příjmu CO 2
 Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
Průduchy regulace příjmu CO 2 Průduchy: regulace transpiračního proudu / výměny plynů transpiration photosynthesis eartamerica.com Průduchy svěrací buňky - zavírání při ztrátě vody (poklesu turgoru) -
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy
 3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy Co to jsou hormony? termín hormon poprvé použitý v medicíně před 100 lety, pochází z řečtiny ηορμονε = stimulovat (ale nezahrnuje
3. Fytohormony a růstové regulátory I. auxiny, gibereliny a cytokininy Co to jsou hormony? termín hormon poprvé použitý v medicíně před 100 lety, pochází z řečtiny ηορμονε = stimulovat (ale nezahrnuje
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Stavba dřeva. Základy cytologie. přednáška
 Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Základy cytologie přednáška Buňka definice, charakteristika strana 2 2 Buňky základní strukturální a funkční jednotky živých organismů Základní charakteristiky buněk rozmanitost (diverzita) - např. rostlinná
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN
 Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Otázka: Výživa rostlin, vodní režim rostlin, růst a pohyb rostlin Předmět: Biologie Přidal(a): Cougee AUTOTROFNÍ A HETEROTROFNÍ VÝŽIVA ROSTLIN, VODNÍ REŽIM ROSTLIN, RŮST A POHYB ROSTLIN 1. autotrofní způsob
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO. Modelová rostlina suchozemská semenná neukončený růst specializované části
 Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
Úvod do biologie rostlin Pletiva Slide 1 ROSTLINNÉ TĚLO Modelová rostlina suchozemská semenná neukončený růst specializované části příjem vody a živin + ukotvení fotosyntéza rozmnožovací potřeba struktur
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Rostlinná pletiva II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis struktury a funkce rostlinných
12. Fytohormony a regenerace rostlin mnohobuněčné organizmy Buňky pletiva orgány celek organizmus vzniká postupně genetického programu
 1 12. Fytohormony a regenerace rostlin Rostliny jsou mnohobuněčné organizmy. Buňky tvoří pletiva sestavená v orgány, orgány tvoří funkčně koordinovaný celek organizmus. Celek vzniká postupně z buňky jediné
1 12. Fytohormony a regenerace rostlin Rostliny jsou mnohobuněčné organizmy. Buňky tvoří pletiva sestavená v orgány, orgány tvoří funkčně koordinovaný celek organizmus. Celek vzniká postupně z buňky jediné
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Šetlík, Seidlová, Šantrůček 2. REGULACE RŮSTU
 2. REGULACE RŮSTU 2.1. FYTOHORMONY Koordinace metabolismu, růstu a vývoje u složitých vícebuněčných organismů závisí na předávání informace mezi buňkami a orgány pomocí signálů. U rostlin jsou růstové
2. REGULACE RŮSTU 2.1. FYTOHORMONY Koordinace metabolismu, růstu a vývoje u složitých vícebuněčných organismů závisí na předávání informace mezi buňkami a orgány pomocí signálů. U rostlin jsou růstové
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové
 Otázka: Růst a vývin rostlin Předmět: Biologie Přidal(a): Verunka kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové FÁZE RŮSTU lze
Otázka: Růst a vývin rostlin Předmět: Biologie Přidal(a): Verunka kvantitativní změna přirůstá hmota, zvětšuje se hmotnost a rozměry rostliny rostou celý život a rychleji než živočichové FÁZE RŮSTU lze
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
3. ČÁST - RŮST A VÝVOJ
 Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. Obecné problémy růstu a vývoje 3. ČÁST - RŮST A VÝVOJ I velmi dokonalé znalosti
Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. Obecné problémy růstu a vývoje 3. ČÁST - RŮST A VÝVOJ I velmi dokonalé znalosti
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Růst a vývoj rostlin
 I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
I N V E S T I C E D O R O Z V O J E V Z D Ě L Á V Á N Í TENTO PROJEKT JE SPOLUFINANCOVÁN EVROPSKÝM SOCIÁLNÍM FONDEM A STÁTNÍM ROZPOČTEM ČESKÉ REPUBLIKY Pracovní list č. 11 Růst a vývoj rostlin Pro potřeby
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky. Reakce průduchů na vlhkost vzduchu. Reakce průduchů na vodní stres
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
3) Membránový transport
 Membránový transport") MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
MBR1 2016 3) Membránový transport a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy 1 Prokaryotická buňka Eukaryotická buňka 2 Pohyb vody první reakce klidných
OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.)
") Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
Úloha č. 18 Obnova apikální dominance na klíčních rostlinách hrachu - 1 - OBNOVA APIKÁLNÍ DOMINANCE NA KLÍČNÍCH ROSTLINÁCH HRACHU (Pisum sativum L.) OBECNÁ CHARAKTERISTIKA RŮSTOVÝCH KORELACÍ Jednotlivé
pátek, 24. července 15 BUŇKA
 BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
BUŇKA ŽIVOČIŠNÁ BUŇKA mitochondrie ribozom hrubé endoplazmatické retikulum cytoplazma plazmatická membrána mikrotubule lyzozom hladké endoplazmatické retikulum Golgiho aparát jádro jadérko chromatin volné
3 a) Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek
 Fyzikální principy. 5 Chemický potenciál (µ s ) (volná energie na jeden mol: J/mol) * = chemický potenciál roztoku s za standartních podmínek") MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
MBRO1 1 2 2017 3) Membránový transport Prokaryotická buňka Eukaryotická buňka a) Fyzikální principy b) Regulace pohybu roztoků membránami a jejich transportéry c) Pumpy Pohyb vody první reakce klidných
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky Reakce průduchů na světlo
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
Mendělejevova tabulka prvků
 Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Mendělejevova tabulka prvků V sušině rostlin je obsaženo přibližně 45% uhlíku, 42% kyslíku, 6,5% vodíku, 1,5% dusíku a 5% minerálních prvků. Tzv. organogenní prvky (C, O, H, N) představují tedy 95% veškerých
Transport živin do rostliny. Radiální a xylémový transport. Mimokořenová výživa rostlin.
 Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Transport živin do rostliny Radiální a xylémový transport. Mimokořenová výživa rostlin. Zóny podél kořene, jejich vztah s anatomií a příjmem živin Transport iontů na střední vzdálenosti Radiální transport
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Rostlinná cytologie. Přednášející: RNDr. Jindřiška Fišerová, Ph.D. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Minerální výživa na extrémních půdách. Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů
 Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
Minerální výživa na extrémních půdách Půdy silně kyselé, alkalické, zasolené a s vysokou koncentrací těžkých kovů Procesy vedoucí k acidifikaci půd Zvětrávání hornin s následným vymýváním kationtů (draslík,
3) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
3. ČÁST - RŮST A VÝVOJ
 Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. Obecné problémy růstu a vývoje 3. ČÁST - RŮST A VÝVOJ Růst rostlin je velmi
Učební text k přednášce Bi4060 na přírodovědecké fakultě MU v Brně. Určeno pouze ke studijním účelům. Autor textu Jan Gloser. Obecné problémy růstu a vývoje 3. ČÁST - RŮST A VÝVOJ Růst rostlin je velmi
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Vladimír Vinter
 Embryo (zárodek) Vývoj embrya (embryogeneze) trvá různě dlouhou dobu (např. u pšenice 20-25 dnů). U některých rostlin jsou embrya zcela nediferencovaná, např. u orchidejí. Zygota je výrazně polární buňka
Embryo (zárodek) Vývoj embrya (embryogeneze) trvá různě dlouhou dobu (např. u pšenice 20-25 dnů). U některých rostlin jsou embrya zcela nediferencovaná, např. u orchidejí. Zygota je výrazně polární buňka
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Buňka. Buňka (cellula) základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách
 základní stavební a funkční jednotka organismů, schopná samostatné existence. Cytologie nauka o buňkách") Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Buňka Historie 1655 - Robert Hooke (1635 1703) - použil jednoduchý mikroskop k popisu pórů v řezu korku. Nazval je, podle podoby k buňkám včelích plástů, buňky. 18. - 19. St. - vznik buněčné biologie jako
Buňka cytologie. Buňka. Autor: Katka www.nasprtej.cz Téma: buňka stavba Ročník: 1.
 Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Buňka cytologie Buňka - Základní, stavební a funkční jednotka organismu - Je univerzální - Všechny organismy jsou tvořeny z buněk - Nejmenší životaschopná existence - Objev v 17. stol. R. Hooke Tvar: rozmanitý,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Fyziologie rostlin. 3. Ontogeneze rostlin. Celistvost rostlin. část 2. Rostlinné regulátory. Alena Dostálová, Ph.D.
 Fyziologie rostlin 3. Ontogeneze rostlin. Celistvost rostlin. část 2. Rostlinné regulátory Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Rostlinné regulátory - regulátory růstu
Fyziologie rostlin 3. Ontogeneze rostlin. Celistvost rostlin. část 2. Rostlinné regulátory Alena Dostálová, Ph.D. Pedagogická fakulta ZČU, letní semestr 2013/2014 Rostlinné regulátory - regulátory růstu
Úvod do biologie rostlin Buňka ROSTLINNÁ BUŇKA
 Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
Slide 1a ROSTLINNÁ BUŇKA Slide 1b Specifické součásti ROSTLINNÁ BUŇKA Slide 1c Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna Slide 1d Specifické součásti ROSTLINNÁ BUŇKA buněčná stěna plasmodesmy Slide
Vakuola. Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich
 Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
Vakuola Dutina uvnitř protoplastu, která u dospělých buněk zaujímá 30 až 90 % jejich objemu. Je ohraničená na svém povrchu membránou zvanou tonoplast. Tonoplast je součástí endomembránového systému buňky
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Vodní režim rostlin. Obsah vody, RWC, vodní potenciál a jeho komponenty: Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy,
 Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Vodní režim rostlin Úvod Klima, mikroklima Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch
 Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
Půda - 4 složky: minerálníčástice organickéčástice voda vzduch kameny a štěrk písek (částice o velikosti 2-0,05mm) prachovéčástice (0,05-0,002mm) jílovéčástice (méně než 0,002mm) F t = F m + F d F d =
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
Vodní režim rostlin. Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické.
 a homoiohydrické.") Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Vodní režim rostlin Úvod Adaptace, aklimace: rostliny vodní, poikilohydrické (řasy, mechy, lišejníky, kapradiny, vyšší rostliny) a homoiohydrické. Obsah vody, RWC, vodní potenciál a jeho komponenty: charakteristika,
Abiotický stres - sucho
 FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
FYZIOLOGIE STRESU Typy stresů Abiotický (vliv vnějších podmínek) sucho, zamokření, zasolení půd, kontaminace prostředí toxickými látkami, chlad, mráz, vysoké teploty... Biotický (způsobený jiným druhem
Úvod do biologie rostlin Úvod PŘEHLED UČIVA
 Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Slide 1a Slide 1b Systém Slide 1c Systém Anatomie Slide 1d Systém Anatomie rostlinná buňka stavba a funkce Slide 1e Systém Anatomie rostlinná buňka stavba a funkce buněčná stěna, buněčné membrány, membránové
Fyziologie rostlin - maturitní otázka z biologie (3)
") Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Otázka: Fyziologie rostlin Předmět: Biologie Přidal(a): Isabelllka FOTOSYNTÉZA A DÝCHANÍ, VODNÍ REŽIM ROSTLINY, POHYBY ROSTLIN, VÝŽIVA ROSTLIN (BIOGENNÍ PRVKY, AUTOTROFIE, HETEROTROFIE) A)VODNÍ REŽIM VODA
Prokaryota x Eukaryota. Vibrio cholerae
 Živočišná buňka Prokaryota x Eukaryota Vibrio cholerae Dělení živočišných buněk: buňky jednobuněčných organismů (volně žijící samostatné jednotky) buňky mnohobuněčných větší morfologické i funkční celky
Živočišná buňka Prokaryota x Eukaryota Vibrio cholerae Dělení živočišných buněk: buňky jednobuněčných organismů (volně žijící samostatné jednotky) buňky mnohobuněčných větší morfologické i funkční celky
Schéma rostlinné buňky
 Rostlinná buňka 1 2 3 5 vakuola 4 5 6 Rostlinná buňka je eukaryotní buňkou se základními charakteristikami tohoto typu buňky. Krom toho má některé charakteristiky typické pro rostlinné buňky, jako je předevšímř
Rostlinná buňka 1 2 3 5 vakuola 4 5 6 Rostlinná buňka je eukaryotní buňkou se základními charakteristikami tohoto typu buňky. Krom toho má některé charakteristiky typické pro rostlinné buňky, jako je předevšímř
FOTOSYNTÉZA Správná odpověď:
 FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
FOTOSYNTÉZA Správná odpověď: 1. Mezi asimilační barviva patří 1. chlorofyly, a) 1, 2, 4 2. antokyany b) 1, 3, 4 3. karoteny c) pouze 1 4. xantofyly d) 1, 2, 3, 4 2. V temnostní fázi fotosyntézy dochází
ROSTLINNÁ PLETIVA I. Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list.
 je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list.") ROSTLINNÁ PLETIVA I Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list. Orgány jsou složeny lž z buněk, které tvoří uvnitř orgánů ů odlišná uskupení pletiva.
ROSTLINNÁ PLETIVA I Tělo cévnatých rostlin (kormus) je rozdělené strukturně ifunkčně na orgány: kořen, stonek a list. Orgány jsou složeny lž z buněk, které tvoří uvnitř orgánů ů odlišná uskupení pletiva.
Aplikované vědy. Hraniční obory o ţivotě
 BIOLOGICKÉ VĚDY Podle zkoumaného organismu Mikrobiologie (viry, bakterie) Mykologie (houby) Botanika (rostliny) Zoologie (zvířata) Antropologie (člověk) Hydrobiologie (vodní organismy) Pedologie (půda)
BIOLOGICKÉ VĚDY Podle zkoumaného organismu Mikrobiologie (viry, bakterie) Mykologie (houby) Botanika (rostliny) Zoologie (zvířata) Antropologie (člověk) Hydrobiologie (vodní organismy) Pedologie (půda)
Základy buněčné biologie
 Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Stomatální vodivost a transpirace
 Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
- pro učitele - na procvičení a upevnění probírané látky - prezentace
 Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
Číslo projektu Název školy Autor Tematická oblast CZ.1.07/1.5.00/34.0743 Moravské gymnázium Brno s.r.o. RNDr. Monika Jörková Biologie 10 obecná biologie Organely eukaryotní buňky Ročník 1. Datum tvorby
ontogeneze listu zpočátku všechny buňky mají meristematický charakter, růst všemi směry (bazální, marginální a apikální meristémy listu)
") Anatomie listu ontogeneze listu epidermis mezofyl vaskularizace vliv ekologických podmínek na stavbu listů listy jehličnanů listy suchomilných rostlin listy vlhkomilných rostlin listy vodních rostlin opadávání
Anatomie listu ontogeneze listu epidermis mezofyl vaskularizace vliv ekologických podmínek na stavbu listů listy jehličnanů listy suchomilných rostlin listy vlhkomilných rostlin listy vodních rostlin opadávání
15. DÝCHÁNÍ ROSTLIN A ŽIVOČICHŮ, RŮST A POHYBY ROSTLIN
 15. DÝCHÁNÍ ROSTLIN A ŽIVOČICHŮ, RŮST A POHYBY ROSTLIN A. Biologický význam dýchání, fáze dýchání, funkce mitochondrií B. Růst a vývoj rostlin, růstové zóny, regulátory růstu, pohyby a dráždivost rostlin
15. DÝCHÁNÍ ROSTLIN A ŽIVOČICHŮ, RŮST A POHYBY ROSTLIN A. Biologický význam dýchání, fáze dýchání, funkce mitochondrií B. Růst a vývoj rostlin, růstové zóny, regulátory růstu, pohyby a dráždivost rostlin
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Bi8240 GENETIKA ROSTLIN Prezentace 07 Rezistence rostlin k abiotickým faktorům doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz Abiotické faktory 1. Nízké teploty ( chladuvzdornost, mrazuvzdornost, zimovzdornost)
Cvičení z fyziologie rostlin. Organogeneze in vitro
 Cvičení z fyziologie rostlin Organogeneze in vitro Teoretický úvod: Jedním z nedůležitějších faktorů ovlivňujících růst a morfogenezi v tkáňových kulturách rostlin je složení kultivačního média. Média
Cvičení z fyziologie rostlin Organogeneze in vitro Teoretický úvod: Jedním z nedůležitějších faktorů ovlivňujících růst a morfogenezi v tkáňových kulturách rostlin je složení kultivačního média. Média
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
DÝCHÁNÍ. uložená v nich fotosyntézou, je z nich uvolňována) Rostliny tedy mohou po určitou dobu žít bez fotosyntézy
 Rostliny tedy mohou po určitou dobu žít bez fotosyntézy") Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Dýchání 2/38 DÝCHÁNÍ Asimiláty vzniklé v rostlinných buňkách fotosyntézou mají různé funkce: stavební, zásobní, enzymatické aj. Zásobní látky jsou v případě potřeby využívány (energie, uložená v nich fotosyntézou,
Stav lesních půd drama s otevřeným koncem
 Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
Stav lesních půd drama s otevřeným koncem Pavel Rotter Ca Mg Lesní půda = chléb lesa = Prvek K význam pro výživu rostlin příznaky nedostatku podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů ve
VAKUOLA. membránou ohraničený váček membrána se nazývá tonoplast. běžná u rostlin, zvířata specializované funkce či její nepřítomnost
 VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
Z Buchanan et al. 2000
 Průběh buněčného cyklu Z Buchanan et al. 2000 Změny v uspořádání mikrotubulů v průběhu buněčného cyklu A interfáze, kortikální mikrotubuly uspořádané v cytoplasmě pod plasmalemou B konec G2 fáze, mikrotubuly
Průběh buněčného cyklu Z Buchanan et al. 2000 Změny v uspořádání mikrotubulů v průběhu buněčného cyklu A interfáze, kortikální mikrotubuly uspořádané v cytoplasmě pod plasmalemou B konec G2 fáze, mikrotubuly
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Buňka. Autor: Mgr. Jitka Mašková Datum: Gymnázium, Třeboň, Na Sadech 308
 Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
Buňka Autor: Mgr. Jitka Mašková Datum: 27. 10. 2012 Gymnázium, Třeboň, Na Sadech 308 Číslo projektu Číslo materiálu CZ.1.07/1.5.00/34.0702 VY_32_INOVACE_BIO.prima.02_buňka Škola Gymnázium, Třeboň, Na Sadech
Morfogeneze rostlinné buňky. Rostlinná cytologie morfogeneze rostlinné buňky, Katedra experimentální biologie rostlin PřF UK
 Morfogeneze rostlinné buňky Morfogeneze = vývoj tvaru, změna tvarových vlastností http://www.biologyreference.com/co-dn/differentiation-in-plants.html Embryogenní (meristematická) buňka Dospělá buňka,
Morfogeneze rostlinné buňky Morfogeneze = vývoj tvaru, změna tvarových vlastností http://www.biologyreference.com/co-dn/differentiation-in-plants.html Embryogenní (meristematická) buňka Dospělá buňka,