Biosyntéza vychází z terpenoidu geranylgeranyldifosfátu. Vznik ent-kaurenu je první krok vlastní dráhy biosyntézy giberelinů.
|
|
|
- Ladislava Novotná
- před 5 lety
- Počet zobrazení:
Transkript
1
2 Gibberelliny
3 Biosyntéza vychází z terpenoidu geranylgeranyldifosfátu. Vznik ent-kaurenu je první krok vlastní dráhy biosyntézy giberelinů.
4
5 Ze stejných počátků vychází také dráha biosyntézy brassinosteroidů a kyseliny abscisové.
6 Effects of Gibberellins General cell elongation. Breaking of dormancy in some plants which require stratification or light to induce germination. Promotion of flowering. Transport is non-polar, bidirectional producing general responses.
7 Další funkce GA Stimulates enzyme production (a-amylase) in germinating cereal grains for mobilization of seed reserves. (Beer!) Can cause parthenocarpic, seedless fruit development. Can delay senescence in leaves and citrus fruits. Induces maleness in dioecious flowers (sex expression).
 uninfected")
8 Discovered in association with Foolish disease of rice (Gibberella( fujikuroi) uninfected infected

9 The classic gibberellin response: elongation of stems.

10 Effects of Gibberellins The cabbage plants on the left were grown on their own The plants on the right were given gibberellic acid once a week for eight weeks Notice the long stems and flowers at the top of the plants on the right They are a result of this hormone
11 GA signals to aleurone layer to switch on genes that produce amylases -- enzymes which convert starch to sugar. Barley seedling GA seed Brewers germinate barley seeds, then, once the enzymes are made, they kill the seeds to make malted
12 CaM = kalmodulin Ca2+ vazebný protein či příbuzné kinázy jsou součástí každé vápníkové reakce buňky.
13 INTERMEZZO
14 [Ca2+]i transient je součástí téměř všech reakcí na podněty vnějšího i vnitřního prostředí rostliny. (př. sekrece a amylasy z aleuronu obilnin v reakci na GA) Bývá obvykle jednou z prvních/nejrychlejších reakcí na podnět.
15
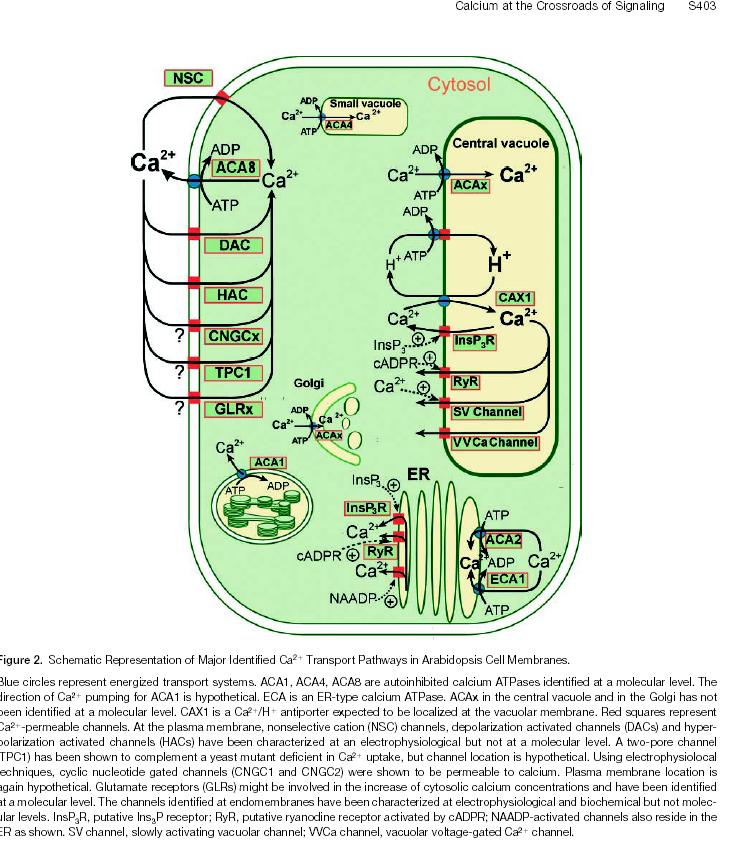
16

17 elativně nízká mobilita Ca2+ v cytoplasmě a rychlé odčerpávání možňuje kompartmentaci /prostorovou lokalizaci vápníkového signál
18 Reakce aleuronových buněk obilnin na GA je nejlépe dokumentovaný případ účasti α-podjednotky trimerní GTPázy na přenosu signálu u rostlin. Obecně trimerní GTPázy hrají u rostlin minoritní roli v recepci a přenosu signálu. Důležitými faktory jsou pravděpodobně receptorové kinázy a malé Rho GTPázy ROP.
19 INTERMEZZO pokračuje

20 Chybějící článek Jak receptorové kinázy aktivují Ropázy?
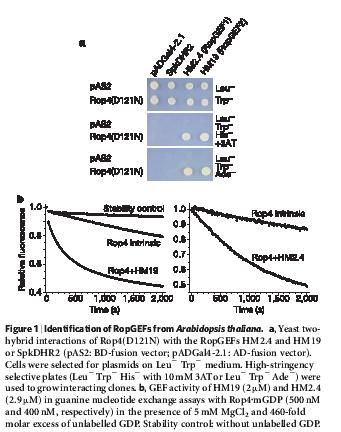
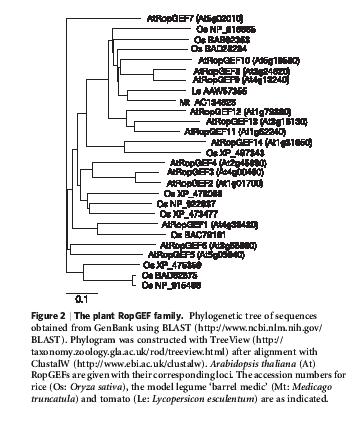
21 14 x u Arabidopsis.
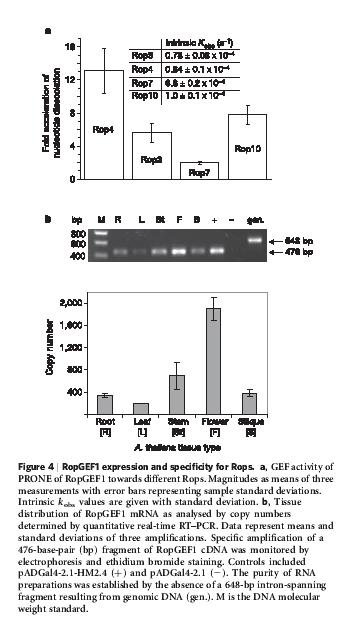
22
23 RLK RopGEF RopGTP pravděpodobně představuje široce využívaný signální modul v rostlinné buňce. Viz. dále např. podíl Rop na regulaci signálu kyseliny abscisové.
24 Zpět ke gibberelinům
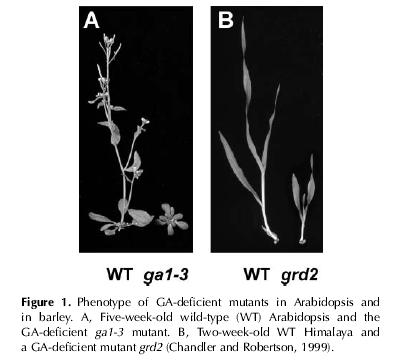
25


26 Green revolution
27

28
29
30 GA 20-oxidázy a GA 3-oxidázy katalyzují poslední kroky biosyntézy aktivních GAs. GA 2-oxidázy naopak inaktivují GAs. Aktivita signální dráhy GA je pevně negativně-zpětnovazebně spojena se syntézou GA. Např. u mutantů s konstitutivně aktivní GA dráhou (rga u Arabidopsis, slr1 u rýže) je snížena hladina GA 20-oxidázy a GA 3-oxidázy, a tedy také hladina GA.
31 Podobně je tomu u dalších fytohormonů!
32
33 GA obecně stimuluje ontogenetické fázové přechody a buněčné dloužení/diferenciaci. Tak působí proti identitě meristémů.
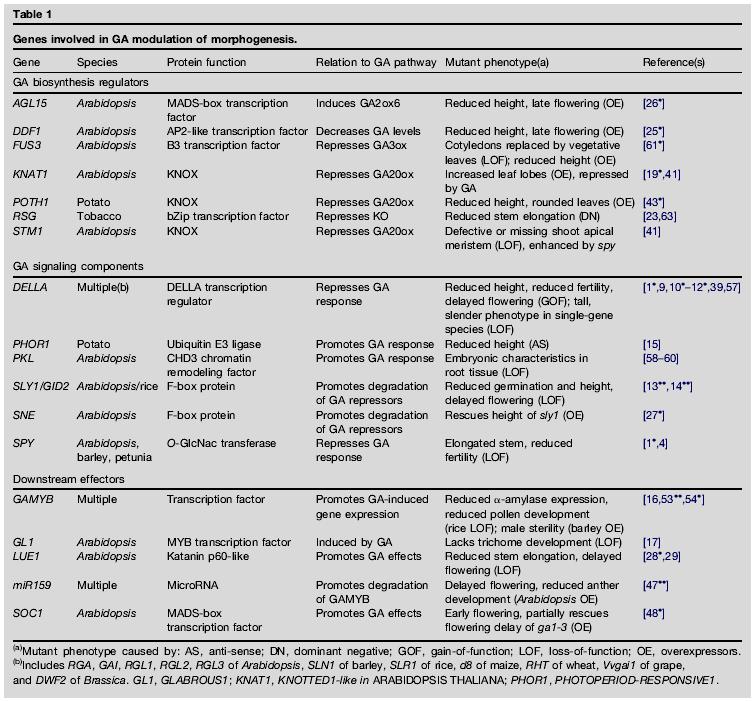
34
35 První známé bíolkoviny GA signální dráhy
36 SPY (SPINDLY,Arabidopsis)=Ser/Thr O- N-acetylglukosamin (GlcNAc) transferáza je negativním regulátorem GA signálu. Připojením GlcNAc na Ser či Thr interferuje s fosforylací, která předchází ubiquitinaci stabilizuje inhibitory GA signálu. Patří mezi geny, které šlechtitelé nevědomky modifikovali při šlechtění na zkrácení stébla = Zelená revoluce
37 Hlavními negativními regulátory GA jsou DELLA bílkoviny

38 GIBBERELINY nemají zřetelnou DNA vazebnou doménu a interagují tedy pravděpodobně s DN prostřednictvím jiných bílkovin. Arabidopsis = RGA, GAI a 3xRGL Rýže = SLR1
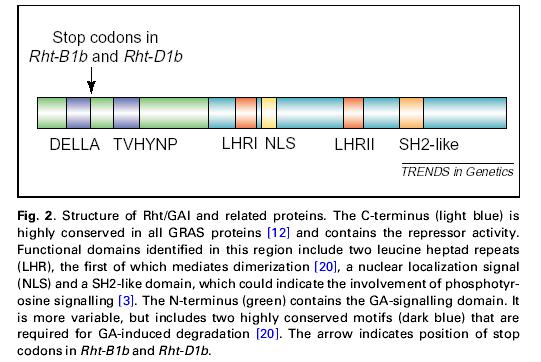
39
40 Pozitivními regulátory GA signálu jsou GID2(GA insensitive dwarf2) u rýže a SLY1 (SLEEPY1) u Arabidopsis
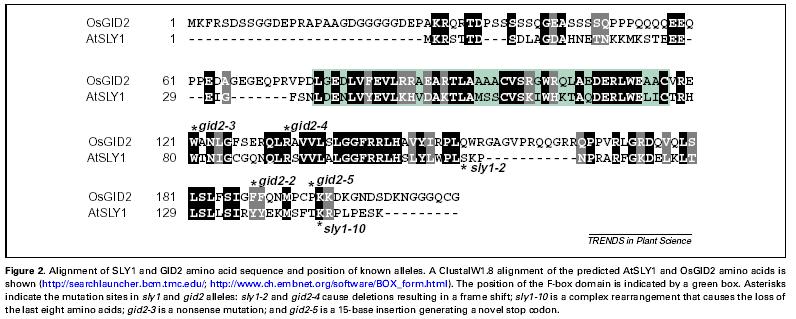
41
42 Oba kodují vysoce homologní F- box podjednotku SCF E3 ligázového komplexu.
43
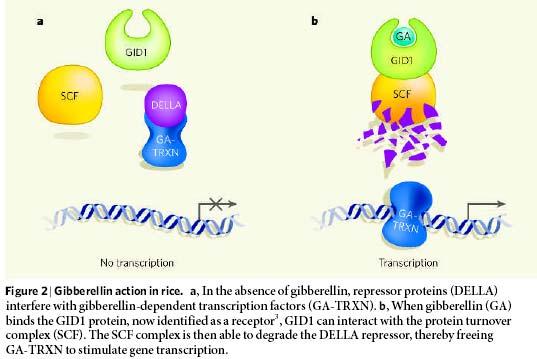
44

45

46 Receptorem GA v cytoplasmě je GID1 nový typ bílkovin příb. lipázám
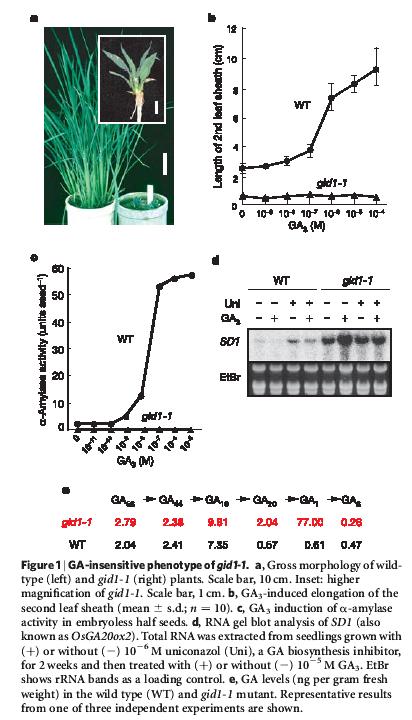
47
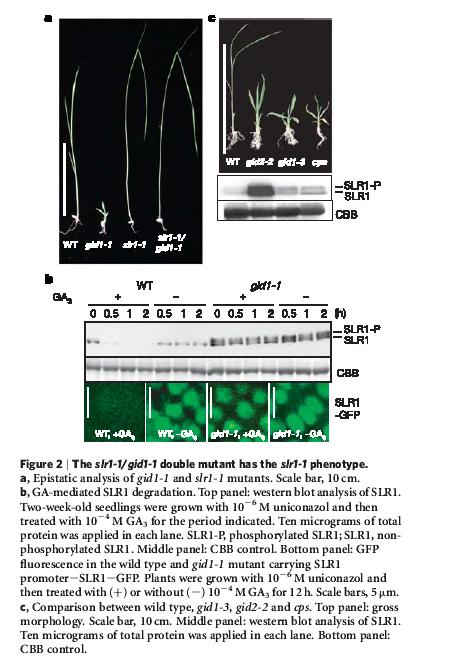
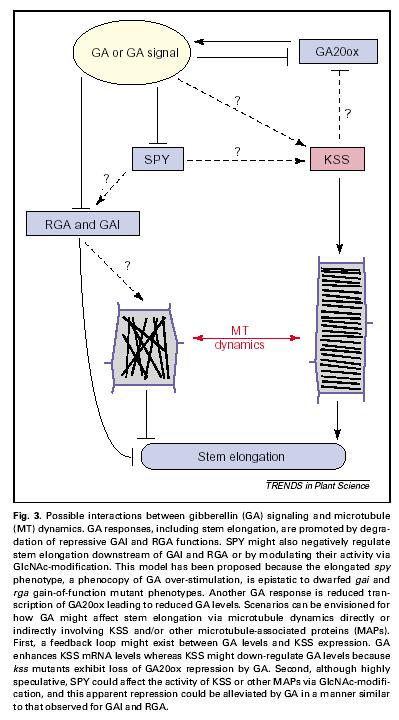
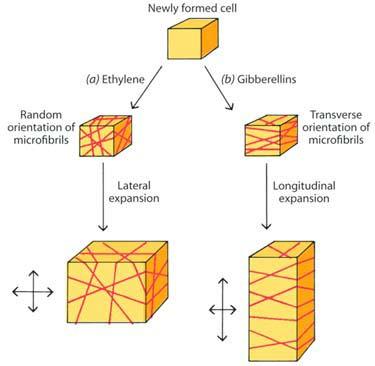
48

49 GA receptor je lokalizován v jádře.
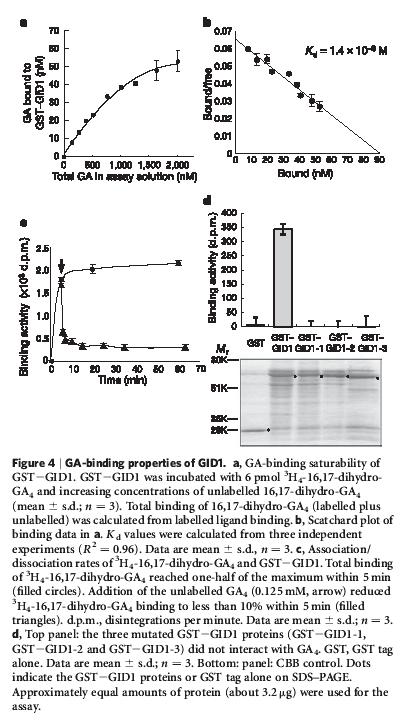
50
51

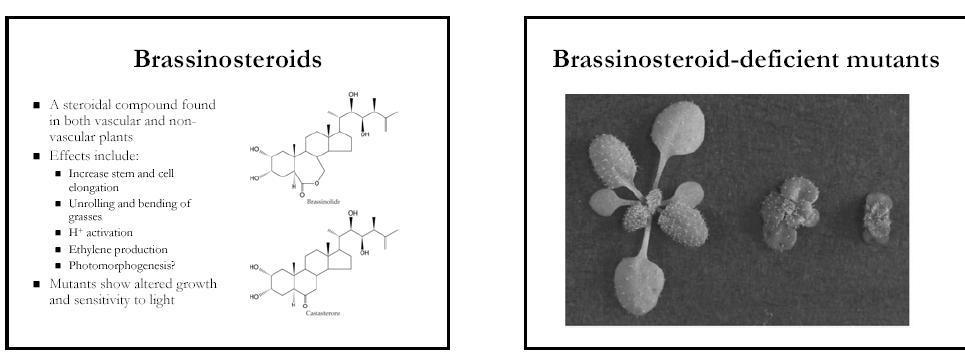
52 GID1 over-expr. GID1 a DELLA/SLR1 interagují
53
54 Jak přesně GID1 a SLR1 interagují s SCF GID2 není jasné. Ale podobnosti s dalšími dráhami jsou nápadné zvl. srovnej s TIR1 receptorem pro IAA z předchozí přednášky.
55 Degradace řízená E3- SCF ubiquitin ligázou je jádrem řady signálních drah u rostlin Podobně IAA (TIR1)

56 fra2 mutant ukazuje na souvislost GA a dynamiky MTs FRA2/AtKSS = katanin, stříhá MTs
57
58
59
60 Brassinosteroidy
61
62
63

64

65
66 BRI1 je RLK, která po aktivaci interaguje s dalšími kinázami. Pro další přenos signálu jsou klíčové fosfatázy.
67

68 BRI1 je příkladem důkladně "promutovaného" lokusu. Proto jsou


69 Arabidopsis má více než 600 RLKs a rýže přes Multiplikace RLKs je pravděpodobně spojena především s obranou proti fytopatogenům.
70
71

72
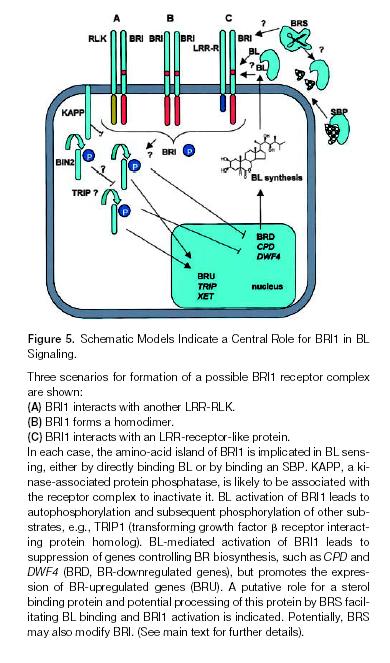
73

74 BIN2 je PK shaggy fam. a be BR fosforylací stimuluje degr TFs BES1 a BZR1 BSU1 je jaderná Fosfatáza. V tomto schematu schází KAPP která inaktivuje Aktivní BRI1/BAK1=SERK Komplex.

75 Signální endosom v BR dráze. Internalizovaný Aktivní Receptor V endosomu Je aktivní Dokud není Defosforylován. SERK3=BAK1
76
77 Eukaryotické dvoukomponentní signální moduly v recepci a přenosu signálů cytokininů a ethylénu.

78 Hlavně prokaryota AHK AHP ARR Eukaryota Klíčovou roli hraje "průtok" fosforylace. Histidin Aspartát a příp. ještě jednou
79 TFs "biol.hodiny"
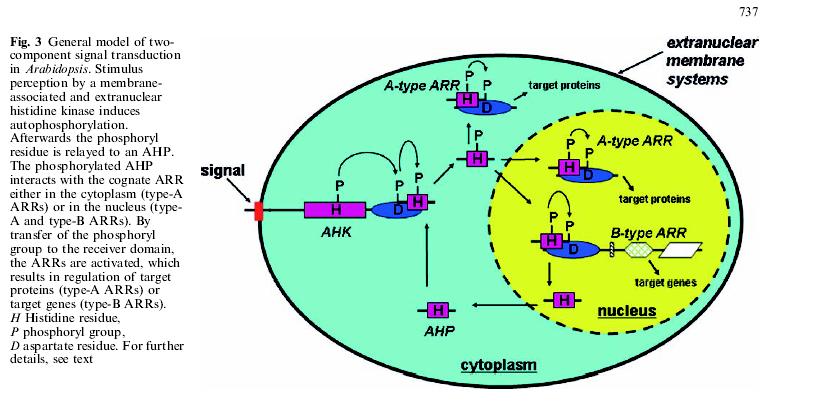
80
81
82 Cytokininy
83 Cytokinins
84 Cyt - (Gr.) hollow vessel; cell Kin - (Gr.) to move
85 isopentenyl transferáza Biosyntéza
86 Function of cytokinins Promotes cell division. Morphogenesis. Lateral bud development. Delay of senescence. Stomatal opening. Rapid transport in xylem stream.

87 Interaction of cytokinin and auxin in tobacco callus tissue

88 auxin cytokinin
89
 cre1/wol")

90 také WOL (wooden leg) cre1/wol chybí v kořeni floém a tvoří se jen xylém.
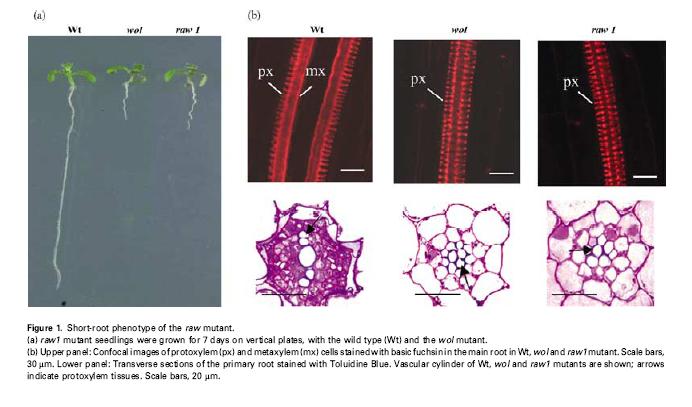
91

92 Studují to v Brně, výv. zárodečného vaku. TFs "biol.hodiny"
93
94
95
96 Ethylen
97 Ethylene H H \ / C = C / \ H H
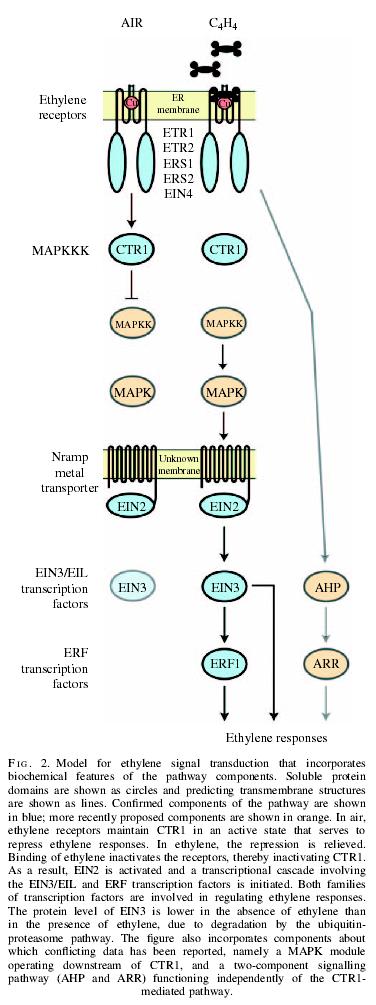
98 Functions of ethylene Gaseous in form. Rapid diffusion. Affects adjacent individuals. Fruit ripening. Senescence and abscission. Interference with auxin transport. Initiation of stem elongation and bud development.
99 Ethylene (C 2 H 4 ) Discovery of ethylene Illuminating gas - senescence and abscission -ethylene Bioassay (triple responses) Chemical nature of ethylene Gas (gas chromatography) Membrane permeable insoluble Biological significance of ethylene in plants Germination Seedling growth Organ senescence and abscission Fruit ripening (climacteric fruits - biosynthesis of ethylene)
100 Yang s cycle Transcriptional regulation inhibitor - stříbro
101 (ETO1) (e.g. CDPK)

102 Triple response of etiolated seedlings with ethylene short hypocotyl thick radial growth apical hook formation
103 Screening of ethylene response mutants Constitutive ethylene response in air (CER) ETO CTR Ethylene resistant/insensitive in ethylene (ETR/EIN)

104 etr1-1 with ethylene in dark Screening of mutagenized seed population with ethylene in dark
105 CTR1 je homolog RAF/MAPKKK kinázy a váže kys. fosfatidovou (PA)

106
107 Receptory etylénu jsou aktivní na ER. a interakcí se tam lokalizuje také CTR1
108

109 I v signalizaci etylénu hraje roli reg. degradace bílkovin.
110
111 Analýza signálních drah v protoplastech Jen Sheen lab
112 Arabidopsis Mesophyll Protoplasts Are Viable and Responsive


113 High Transfection Efficiency of Mesophyll Protoplasts Maize Arabidopsis Maize PEG-Calcium Electroporation


114 Functional Genomic Analyses of Plant Signal Transduction in Arabidopsis

115 Auxin Signaling in Maize Mesophyll Protoplasts

116 ytokinin Signaling in Arabidopsis Mesophyll Protoplasts
117 Stimuli Sensors/Receptors M A P K C itogenctivated rotein inase ascades Arabidopsis Genes MAPKKKs > 25 MAPKKs 9 MAPKs 23 /Groups Transcription Factors Primary Response Genes Secondary Response Genes Functional Genomic Approach
118 Příklad NPK1(MAPKKK) potlačuje auxinový signál.
119 Plant MAPK Signaling Cascades 2001 Plant Arabidopsis Tobacco Alfalfa Arabidopsis Tobacco Signal Bacterial Elicitor Oxidative Stress Fungal Elicitor Salt Stress Abiotic Sresses Mitosis Cytokinesis Sensor FLS2? AtHK1? MAPKKK AtMEKK1 AtANP1 (NtNPK1) AtMEKK1 NtNPK1 (AtANP1) MAPKK AtMKK4/5 (NtMEK2) (MsSIMKK) NtMEK2 (AtMKK4/5) (MsSIMKK) MsSIMKK (AtMKK4/5) (NtMEK2) AtMKK1/2 (NtSIPKK) NtMEK1 ** MAPK AtMPK3/6 AtMPK3/6 (NtWIPK/SIPK) (MsSAMK/SIMK *) NtSIPK/WIPK (AtMPK3/6) (MsSAMK/SIMK *) MsSIMK (AtMPK6) (NtSIPK) AtMPK4 (MsMMK2) NtNtf6 ** (MsMMK3) Target Genes PAL(+) GST6, HSP18.2 (+) GH3, ER7 (-) HMGR(+) PAL (+)??? Negative Regulator AtMKP1??? MsMP2C? AtPTP1? AtMKP1? AtPTP1??
120 Summary MAPK cascades play a central role in intracellular signaling MAPK cascades are involved in stress and hormonal signaling MAPK cascades mediate both positive and negative controls Manipulation of MAPK cascades confers multiple stress protection
121
122 ABA

123 Dormant buds of a hickory. ABA tends to promote dormancy while GA promotes growth.
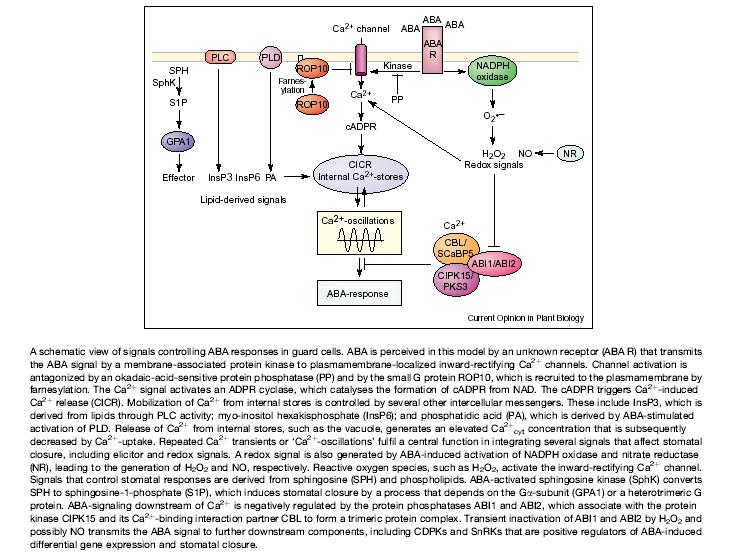
124
125 Functions of abscisic acid General growth inhibitor. Causes stomatal closure. Inhibits stomatal opening. Readily translocated. Produced in response to stress. Řada interakcí s cukry jako signály!

126 Viviparní kukuřice. např VP1
127 Hypotézy o působení ABA na vývoj semene. Důležité je rozlišení mateřské ABA, která přichází zvenčí (membr.rec.?) a endogenní embryonální (rozp. receptor?).
128 Existují pravděpodobně dva typy receptorů pro ABA - membránový a cytoplasmatický. Dosud nejsou známy.
129 KYSELINA KYSELINA ABSCISOVÁ ABSCISOVÁ

130 Abscisic acid je prekurzorem ABA
131
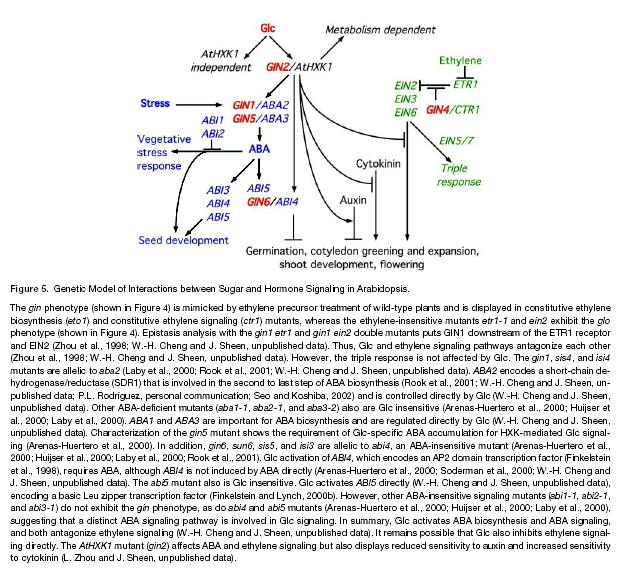
132 Nepřímá dráha štěpením violaxantinu!
133 ABA metabol ism
134 cadpr=cyklická ADP ribosa.
135 Rop10 je negativním regulátorem ABA drah.
136
137

138 ABA a cukry interagují
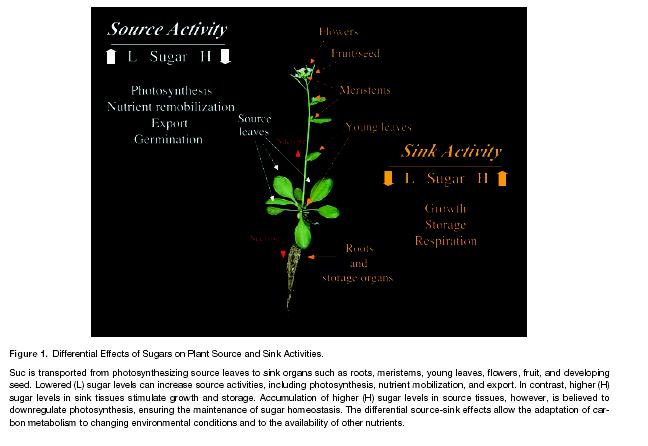
139 Sugar Signalling
140
141 Hexokináza (HXK volná či vázaná) katalyzuje aktivační fosforylaci glukosy, která tak vstupuje do metabolismu. HXK funguje jako SENZOR průtoku glukosy - sama vstupuje do jádra či prostřednictvím interagujících bílkovin (HIPs) ovlivňuje genovou expresi.

142 GIN2 je AtHXK1 Glukosa u WT brzdí deetiolaci.
143
144 Cukry interferují se všemi "fytohormony"
145
146 (met)ja kyselina (met)jasmonová
147 reakce na poranění a interakce s patogeny
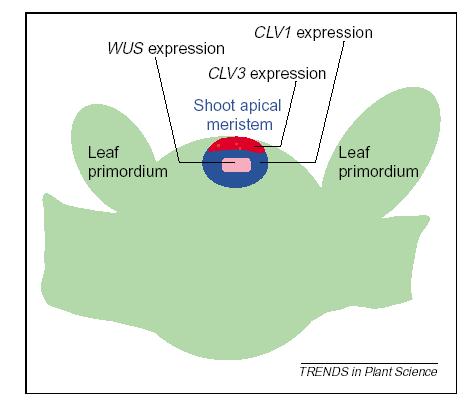
148 = PLA
149
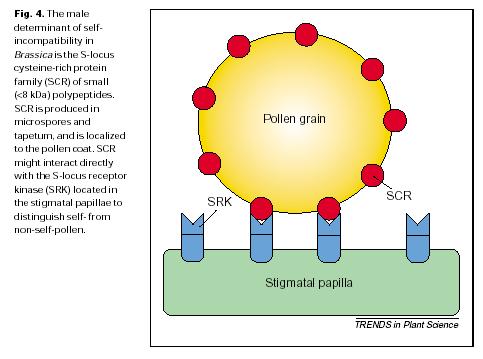
150 INTERAKCE S ETYLENEM A DALŠÍMI
151

152 a také E3 ligáza COI1 je F-box!
153 PEPTIDY JAKO SIGNÁLY
154
155

156
157

158
159
160
161
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př objevený Strigolacton
 Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding Protein1 byl objeven
Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding Protein1 byl objeven
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Cíl přednášky Popis základních principů hormonální regulace růstu a vývoje živočichů a rostlin,
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
REPETITORIUM REGULACÍ
 REPETITORIUM REGULACÍ Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding
REPETITORIUM REGULACÍ Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
3) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") 1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
1 2010 3) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Gray J (2003) Programmed Cell Death in Plants. University of Toledo,
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
PŘENOS SIGNÁLU V BUŇCE. Nela Pavlíková
 PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL
 STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Vápník. Deficience vápníku: - 0,4-1,5% DW. - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM)
") Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.)
") STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.) ZÁSOBNÍ PROTEINY A KYSELINA ABSCISOVÁ V ZYGOTICKÉ EMBRYOGENEZI HRACHU SETÉHO (PISUM SATIVUM L.) 1 Solnická P., 1 Klemš
STORAGE PROTEINS AND ABSCISIC ACID IN ZYGOTIC EMBRYOGENESIS OF PEA (PISUM SATIVUM L.) ZÁSOBNÍ PROTEINY A KYSELINA ABSCISOVÁ V ZYGOTICKÉ EMBRYOGENEZI HRACHU SETÉHO (PISUM SATIVUM L.) 1 Solnická P., 1 Klemš
Stomatální vodivost a transpirace
 Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Vodní režim rostlin Stomatální vodivost a transpirace Vliv faktorů prostředí - obecně Změny během dne Interakce různých faktorů Aklimace Adaxiální a abaxiální epidermis Ontogeneze Matematické modelování
Výzkumný ústav veterinárního lékařství v Brně
 LIPIDY: FUNKCE, IZOLACE, SEPARACE, DETEKCE FOSFOLIPIDY chemické složení a funkce v buněčných membránách; metody stanovení fosfolipidů fosfolipázy - produkty reakcí (ceramid, DAG = 2nd messengers) a stanovení
LIPIDY: FUNKCE, IZOLACE, SEPARACE, DETEKCE FOSFOLIPIDY chemické složení a funkce v buněčných membránách; metody stanovení fosfolipidů fosfolipázy - produkty reakcí (ceramid, DAG = 2nd messengers) a stanovení
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
Takahashi K & Yamanaka S. Cell 126, 2006,
 Obecné mechanismy buněčné signalizace Signalizace do buněčného jádra MUDr. Jan láteník, hd. Ústav lékařské biochemie 1.LF UK Somatické buňky lze přeprogramovat na pluripotentní kmenové buňky! řeprogramování
Obecné mechanismy buněčné signalizace Signalizace do buněčného jádra MUDr. Jan láteník, hd. Ústav lékařské biochemie 1.LF UK Somatické buňky lze přeprogramovat na pluripotentní kmenové buňky! řeprogramování
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Bringing Lights to Your Projects. by Visiocom.
 by Visiocom kryt čočka reflektor základna 12-ti čipové LED reflektor Síla slunce v jedinečném zlatém designu. VisioGrow. ovládací panel Studený vzduch Horký vzduch Hliníkové tělo Zadní panel Tichý ventilátor
by Visiocom kryt čočka reflektor základna 12-ti čipové LED reflektor Síla slunce v jedinečném zlatém designu. VisioGrow. ovládací panel Studený vzduch Horký vzduch Hliníkové tělo Zadní panel Tichý ventilátor
CZ.1.07/1.1.00/
 Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
Petr Tarkowski Rostlinné hormony malé molekuly s velkým významem Vzdělávání středoškolských pedagogů a studentů středních škol jako nástroj ke zvyšování kvality výuky přírodovědných předmětů CZ.1.07/1.1.00/14.0016
12. Fytohormony a regenerace rostlin mnohobuněčné organizmy Buňky pletiva orgány celek organizmus vzniká postupně genetického programu
 1 12. Fytohormony a regenerace rostlin Rostliny jsou mnohobuněčné organizmy. Buňky tvoří pletiva sestavená v orgány, orgány tvoří funkčně koordinovaný celek organizmus. Celek vzniká postupně z buňky jediné
1 12. Fytohormony a regenerace rostlin Rostliny jsou mnohobuněčné organizmy. Buňky tvoří pletiva sestavená v orgány, orgány tvoří funkčně koordinovaný celek organizmus. Celek vzniká postupně z buňky jediné
2012/2013. Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol.
 2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
2012/2013 Fyziologie rostlin: MB130P14, kolektiv přednášejících Albrechtová a kol. Místo konání: Viničná 7, 2. patro, B7, Zoologická posluchárna, 14:50-17:15 No. Téma: Přednášející CZ: Datum 1 Formování
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
(molekulární) biologie buňky
 biologie buňky") (molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
(molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
Intracelulární Ca 2+ signalizace
 Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI
 MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
Signalizace u rostlin II
 Fyziologie rostlin, LS 2013 Přednáška 7 Signalizace u rostlin II Další fytohormony, spolupráce fytohormonů; pohyby rostlin Jan Petrášek, PřFUK Gibereliny historie objevu a chemická struktura 30. léta 20.
Fyziologie rostlin, LS 2013 Přednáška 7 Signalizace u rostlin II Další fytohormony, spolupráce fytohormonů; pohyby rostlin Jan Petrášek, PřFUK Gibereliny historie objevu a chemická struktura 30. léta 20.
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách
 RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
RŮST = nevratné přibývání hmoty či velikosti rostliny spojené s fyziologickými pochody v buňkách Fáze růstu na buněčné úrovni: zárodečná (embryonální) dělení buněk meristematických pletiv prodlužovací
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Regulace enzymových aktivit
 Regulace enzymových aktivit Regulace enzymových aktivit: Změny množství enzymu v kompartmentu, buňce, orgánu: - změna exprese, degradace atd. - změna lokalizace Skutečné regulace: - aktivace/inhibice nízkomolekulárními
Regulace enzymových aktivit Regulace enzymových aktivit: Změny množství enzymu v kompartmentu, buňce, orgánu: - změna exprese, degradace atd. - změna lokalizace Skutečné regulace: - aktivace/inhibice nízkomolekulárními
Obecný metabolismus.
 mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
mezioborová integrace výuky zaměřená na rostlinnou biochemii a fytopatologii CZ.1.07/2.2.00/28.0171 Obecný metabolismus. Regulace glykolýzy a glukoneogeneze (5). Prof. RNDr. Pavel Peč, CSc. Katedra biochemie,
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
umožňují enzymatické systémy živé protoplazmy, nezbytný je kyslík,
 DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
DÝCHÁNÍ ROSTLIN systém postupných oxidoredukčních reakcí v živých buňkách, při kterých se z organických látek uvolňuje energie, která je zachycena jako krátkodobá energetická zásoba v ATP, umožňují enzymatické
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkn koordinovaný celek. Vývoj rostlin je
d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů
 Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů") MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
MBR2 2016 2) Membránový transport 1 d) Kanály e) Přenašeče a co-transportéry, mediátory difúze a sekundární aktivní transport f) Intracelulární transport proteinů d) Kanály Rostliny: iontové kanály a akvaporiny
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Jaderné receptory. ligand. cytoplazmatická membrána. jaderný receptor DNA. - ligandem aktivované transkripční faktory
 Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky
 fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky") FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
FM styrylové sloučeniny/barvičky (fy. Mol.Probes) fluoreskují po zapojení do membrány. Internalizují se endocyózou. Optimální je pro rostlinné buňky FM4-64 Cytokinese a endocytóza GFP či YFP-AtEB1 a CLIP170
Téma: FYTOHORMONY. Santner et al Praktikum fyziologie rostlin
 Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Téma: FYTOHORMONY Santner et al. 2009 Praktikum fyziologie rostlin Teoretický úvod: FYTOHORMONY Rostliny se vyvíjejí jako strukturovaný, organicky vyvážený a funkčně koordinovaný celek. Vývoj rostlin je
Signalizace je vlastně
 Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES
 BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES Vlašánková E.*, Kohout L.**, Klemš M.*, Hradilík J.* *Ústav botaniky a fyziologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická
BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES Vlašánková E.*, Kohout L.**, Klemš M.*, Hradilík J.* *Ústav botaniky a fyziologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická
Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy
 Fyziologie rostlin, LS 2013 Přednáška 5 Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy Jan Petrášek, PřFUK Signalizace - dráždivost Bohumil Němec: Vztahy rostlin k vnějšímu světu,
Fyziologie rostlin, LS 2013 Přednáška 5 Signalizace u rostlin I Signály, signální dráhy, auxiny a cytokininy Jan Petrášek, PřFUK Signalizace - dráždivost Bohumil Němec: Vztahy rostlin k vnějšímu světu,
RVR ) Vývoj květu a kontrola kvetení. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění
 Vývoj květu a kontrola kvetení. d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění") 2015 6) Vývoj květu a kontrola kvetení 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění 2 d) Vznik gamet Životní cyklus rostliny Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
2015 6) Vývoj květu a kontrola kvetení 1 d) Vznik gamet e) Mutace ve vývoji gametofytu f) Opylení, oplodnění 2 d) Vznik gamet Životní cyklus rostliny Mikrosporogeneze Megasporogeneze Vývoj samčího gametofytu
STANOVENÍ RYCHLOSTI KLÍČENÍ OBILEK JEČMENE
 Úloha č. Stanovení rychlosti klíčení obilek ječmene - - STANOVENÍ RYCHLOSTI KLÍČENÍ OBILEK JEČMENE DORMANCE A KLÍČENÍ SEMEN Termínem klíčení (obilek, semen) obilek rozumíme obnovení metabolické aktivity
Úloha č. Stanovení rychlosti klíčení obilek ječmene - - STANOVENÍ RYCHLOSTI KLÍČENÍ OBILEK JEČMENE DORMANCE A KLÍČENÍ SEMEN Termínem klíčení (obilek, semen) obilek rozumíme obnovení metabolické aktivity
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Fyziologie AUTOFAGIE. MUDr. JAN VARADY KARIM FNO
 Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Vodní režim rostlin. Mechanizmy pohybu průduchů. Obecné charakteristiky. Reakce průduchů na vlhkost vzduchu. Reakce průduchů na vodní stres
 Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Vodní režim rostlin Mechanizmy pohybu průduchů Obecné charakteristiky Reakce průduchů na světlo Reakce průduchů na vodní stres Reakce průduchů na vlhkost vzduchu Reakce průduchů na CO 2 Reakce průduchů
Tkáňové kultury rostlin. Mikropropagace
 Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Tkáňové kultury rostlin Mikropropagace IN VITRO KULTURY (EXPLANTÁTOVÉ KUTLURY, ROSTLINNÉ EXPLANTÁTY) Izolované rostliny, jejich orgány, pletiva či buňky pěstované in vitro ve sterilních podmínkách Na kultivačních
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 LRR/OBBC LRR/OBB Obecná biologie Orgány rostlin II. Mgr. Lukáš Spíchal, Ph.D. Cíl přednášky Popis anatomie, morfologie a funkce
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
Rostlinná cytologie. Přednášející: RNDr. Jindřiška Fišerová, Ph.D. Rostlinná cytologie, Katedra experimentální biologie rostlin PřF UK
 Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Rostlinná cytologie MB130P30 Přednášející: RNDr. Kateřina Schwarzerová,PhD. RNDr. Jindřiška Fišerová, Ph.D. Přijďte na katedru experimentální biologie rostlin vypracovat svou bakalářskou nebo diplomovou
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Buněčné základy růstu a morfogeneze, signalizace u rostlin fytohormony
 Buněčné základy růstu a morfogeneze, signalizace u rostlin fytohormony Růst a vývoj rostlin Zásadní rozdíly ve vývoji rostlin a živočichů Stavba rostlinného těla není předem určená Významný podíl nediferencovaných
Buněčné základy růstu a morfogeneze, signalizace u rostlin fytohormony Růst a vývoj rostlin Zásadní rozdíly ve vývoji rostlin a živočichů Stavba rostlinného těla není předem určená Významný podíl nediferencovaných
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
REGULACE TRANSLACE DEGRADACE BÍLKOVIN. 4. Degradace bílkovin. 4. Degradace bílkovin. 4. Degradace bílkovin
 4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
Bp1252 Biochemie. #11 Biochemie svalů
 Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
Prokaryotická X eukaryotická buňka. Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen)
") Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,
Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,