Signalizace je vlastně
|
|
|
- Pavlína Konečná
- před 5 lety
- Počet zobrazení:
Transkript
1 Signalizace je vlastně komunikace a komunikace je základem života.
2 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Signalizace světlem
3 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Signalizace světlem
4 Signalizace, regulace, komunikace a integrace v buňce, pletivu a Několik poznámek. organizmu Kvantitativně převažujícím prvkem regulace genové exprese je represe (a její odblokování). Většina bílkovin je polyfunkčních a jejich exprese a funkce je regulována na mnoha úrovních najednou. Signální dráhy se integrují na úrovni společného regulačního bílkovinného intermediátu (příklad fosf.), druhého posla, promotoru, procesu či struktury. Signál je zesilován, či zeslabován - při tom šum okolí může být pozitivně využit k zesílení signálu = stochastická resonance.
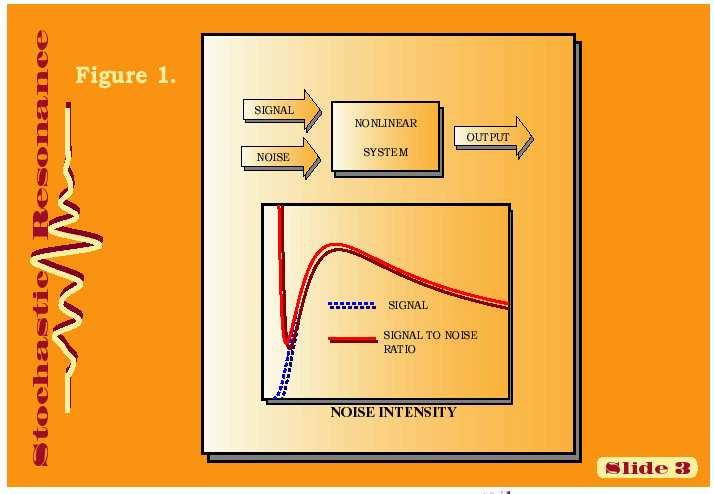
5
6 Z toho, že organismus vládne buňkám (nejen buňky organizmu), také plyne, že také buňka a organizmus vládne signálním drahám a sítím. Buňka není jen výsledkem propletence procesů/struktur, které v ní probíhají/strukturují, ale také jejich tvůrcem.
 Keyboard Signal")



7 Analogy view of cell signaling transduction pathway Input (command) Keyboard Signal perception Endogenous: phytohormone Exogenous: environmental cue CPU (Central Processing Unit) Chip Signal Transduction Network Compiling, Integration, processing Output (performance) Printer Signal Response Morphogenesis change, Growth development differentiation
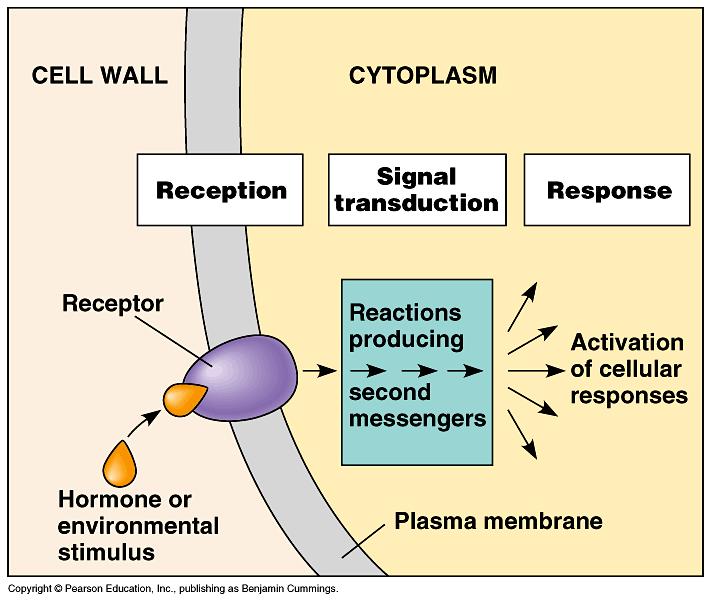
8
")
9 A wide range of disparate external and internal signals is monitored by plants and used to compute appropriate developmental responses Molecular Cell Biology 2: (2001)
10 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Signalizace světlem

11
12 Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA
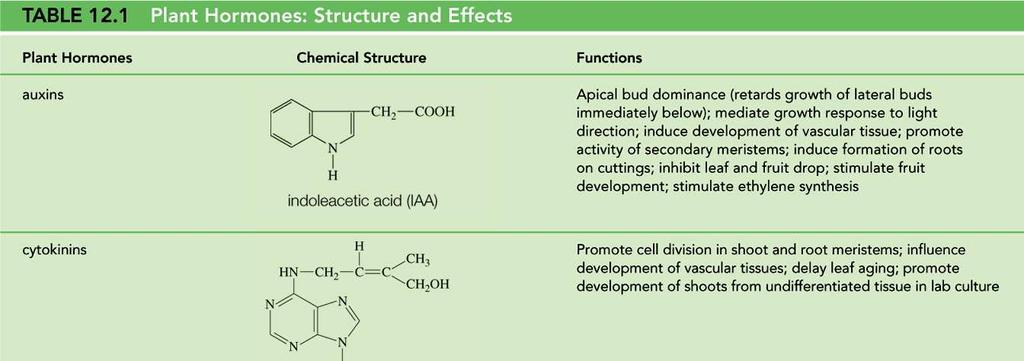
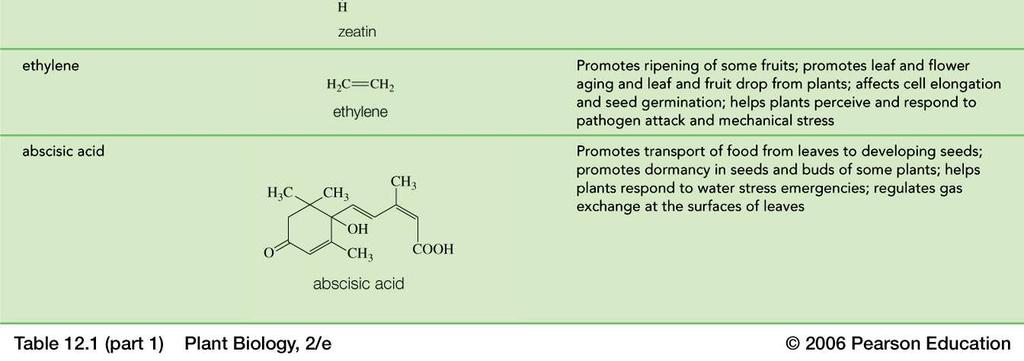
13
14
15
16 Main approaches used to study plant hormone perception, signal transduction biosynthesis and responses 1. Molecular genetics approach hormone biosynthetic mutants vs. hormone insensitive mutants 2. Biochemistry approach hormone binding protein (photo-affinity probe, affinity chromatography, immunopurification) 3. Inhibitors approach kinase/phosphatase, G-protein, PKC etc. inhibitors
17 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz Peptidové hormony Signalizace světlem
18 Degradace bílkovin je stejně důležitý regulační krok jako jejich syntéza. Signální dráhy často obsahují vysoce specifickou/regulovanou degradaci bílkoviny jako důležitý regulační krok. (Vzpomeňme na cykliny )
19 Proteolýza je ovšem také konstitutivní proces. Až 30% translatovaných bílkovin je nefunkčních.
20 Degradace buněč ěčných bílkovinb 17-1 Proteolytické dráhy u eukaryot - 1. vakuolární/lysozomální - 2. Na ubiquitinu-proteasomu závislá degradace - 3. post-proteasomálni degradace : Tricorn, TPII? - 4. Degradace membranových proteinů

21 17-2 Hlavní proteolytické dráhy eukaryot Mitochondria Autophagosome Mitochondrial /plastid proteolytic systems Lysosome/ endosome endosomelysosome system cooperation nuclear proteins cytoplasmic proteins Nucleus Ubiquitinproteasome system ER proteins endosome-lysosome pathway degrades extracellular and cellsurface proteins ubiquitin-proteasome pathway degrades proteins from the cytoplasm, nucleus and ER mitochondria (and chloroplasts) have their own proteolytic system that are of bacterial origin
22 Degradace membránových proteinů. ATP dep. membr.. komplexy AAA proteases mediate the degradation of membrane proteins in bacteria, mitochondria and chloroplasts (i.e., compartments of eubacterial origin) combine proteolytic and chaperone activities in one system, acting as qualitycontrol machineries model substrate polypeptides containing hydrophilic domains at either side of the membrane can be completely degraded by either of two AAA proteases found in mitochondria, if solvent-exposed domains are in an unfolded state - a short protein tail protruding from the membrane surface is sufficient to allow the proteolytic attack of an AAA protease that facilitates domain unfolding at the opposite side Leonhard et al. (2000) Mol. Cell 5,

23 ClpAP je proteázový komplex aktivní v plastidech (homol. E.coli). Brání hromadění nefunkčních bílkovin v plastidech. Podobně je tomu v mitochondriích.
24 Vakuolárn rní/lysozomální degradace 17-5 macroautophagy is the equivalent of forming intracellular endosomes (phagosomes) that fuse to the lysosome and result in the breakdown of its contents vakuola Cuervo and Dice (1998) J. Mol. Med. 76, 6-12.
25 Bílkoviny určené k degradaci proteasomem jsou modifikovány ubiquitinem. Prvním známým proteinem ubq. in vivo v aktivní formě byl u rostlin fytochrom. Většina bílkovin je před ubiquitinací specificky fosforylována.
26 Ubiquitinová dráha 17-6 E1 - ubiquitin activating enzyme E2 - ubiquitin conjugating enzyme E3 - ubiquitin ligase ~ denotes high-energy thioester bond DUB, deubiquinating enzyme

27 Syntet. z fusních tandemových prekursorů 3 až 6. Jsou štepeny deubiqutinačními enzymy/proteasami = DUB 76 AA rostl. od kvasinek/živočichů se liší 2/3 AA. Ubq. se kovalentně váže na Lys cílové bílkoviny C -Gly.
28 E1 ubq. Aktivace E2 ubq. Konjugace E3 ubq. Ligace ubq. na Lys cílové bílkoviny. E3 zajišťuje specifitu rozpoznání. Na bílkovině je ubq. více Lys a to opakovaně = polyubiquitin.
29 ~1200 E3 Genes! [Vierstra, 2003 TIPS]
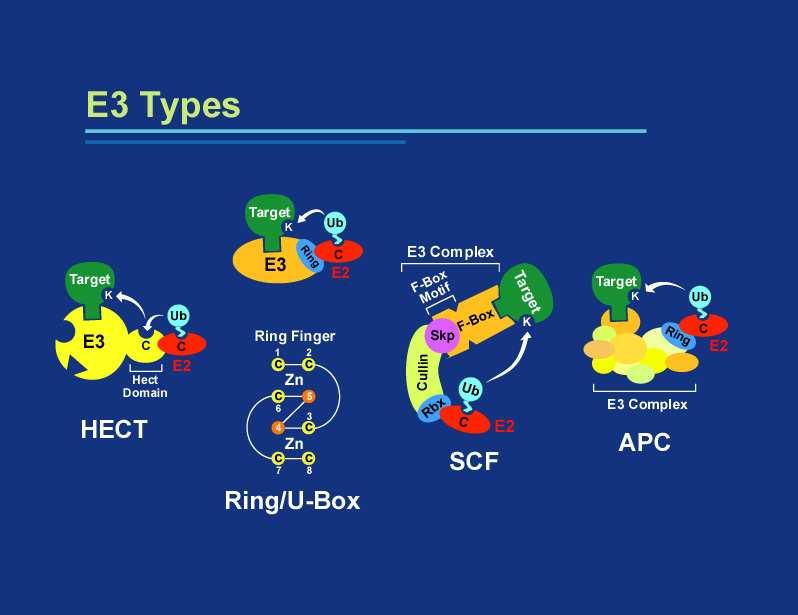
30
31

32 F-box odpovídá za specifitu interakce se substrátem.
33 Monoubiquitinace může sloužit jako regulační modifikace. Např. pro třídění do endocytotické dráhy a vakuoly/lysozomuči modifikuje např. transkripci TF-ubq.
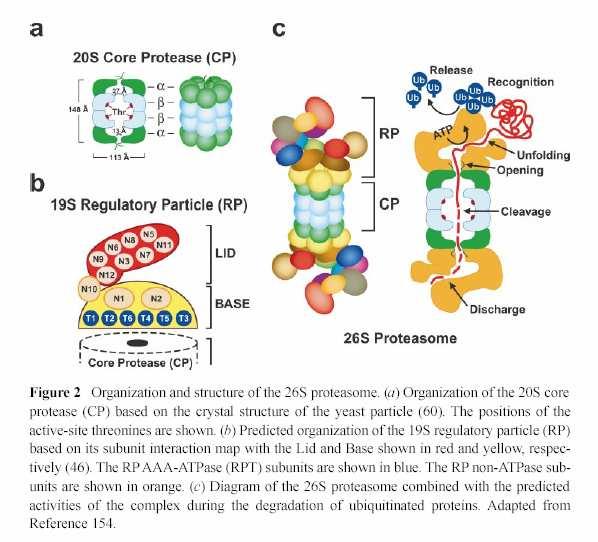
34 Proteasom
35 Topologie proteasomu zajišťuje, že proteázovou aktivitou nebudou nespecificky zasaženy cytoplasmatické bílkoviny. U Arabidopsis každá podjednotka = dva geny - to zn. různé subtypy proteasomu.
36 CSN (COP9 / signalosom) komplex byl poprvé objeven u Arabidopsis (viz. dále = světlo jako signál). Jeho podjednotky a celková organizace jsou homologní "víku" RP proteasomu, ale funguje nezávisle, jako regulátor ubiquitinace a aktivity proteazómu mu.
37
38 CSN kontroluje aktivitu E3 SCF ligázy prostřednictvím neddylace či deneddylace (NEDD8 či RUB1 jsou peptidy podobné ubiquitinu) a degradaci bílkovin interakcemi s proteasomem (alternativní víko ).
39 Bílkoviny se mnohonásobně liší poločasem životnosti a ten se prudce mění s měnícím se diferenciačním/regulačním stavem buňky. Klíčové proteiny signálních drah (včetně transkripčních faktorů) bývají velmi labilní.
40 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz Peptidové hormony Rotlinné hormony a jejich signální dráhy Signalizace světlem
41 Auxin
42

43 Hladina aktivního auxinu je regulována modifikacemi/konjugacemi - reverzibilními i ireverzibilními.

44 Polar transport of Auxin Chemoosmotická teorie transportu auxinu potvrzena objevem AUX1 vtokového a PIN výtokových přenašečů a jejich polární lokalizace. Transport jde ovšem také pře ABC transportery (PGP, závislé na ATP) a je možná také sekrece z IAA naplněných váčků jako v případě neurotransmiterů.
45 Auxin - přenos signálu
46
47 Fungují PINy jako senzory? Co je receptorem auxinu???
48 F-BOX SCF-E3 LIG. KOMPLEXU TIR1 JE RECEPTOREM AUXINU.

49 Auxin signal transduction E3 ubiquitin ligase complex 49
50 Mutanti monopteros(mp) a bodenlos(bdl) nemají kořen. MP kóduje člena ARF (auxin response transcript. factors) rodiny transkripčních aktivátorů ARF5. (Prosím neplést s malými GTPázami!). BDL kóduje člena rodiny inhibitorů ARFů AUX/IAA, IAA12. Tyto bílkoviny se vyznačují minutovým poločasem životnosti. S ARFy tvoří heterodimery a tak blokují jejich aktivitu. Transkripce AUX/IAA genů je stimulována IAA (okamžitě) - tak byly objeveny.

51 ABP1 Auxin Binding Protein1 byl objeven biochemicky je lokalizován převážně do membrány ER, ale malá frakce "uniká" a je aktivní na povrchu buňky. Pokus s blokováním reakce protoplastů na IAA protilát. prokázal jeho podíl na reakci na IAA.
52 Řada mutantů s postiženým proteasomem (a také CSN) má auxinový fenotyp - bývají auxin rezistentní.
53 Model regulace ARFů AUX/IAA a jejich auxinem stimulovanou proteolýzou. BDL a další AUX/IAA geny jsou také cílem transkripční stimulace.
54 De novo syntéza AUX/IAA represorů umožňuje potlačení signálu = atenuaci. Podobně kinázy fungují jako přenašeče signálů jen díky protipůsobícím fosfatázám. a tak je to i s Ca2+ a dalšími.
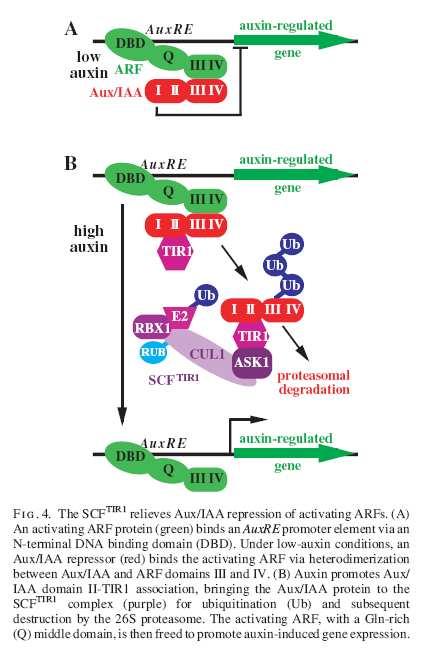
55
56 TIR1 a spol. jsou regulovány neddylací/rub1 modifikací a tedy také CSN.
57 NEDD8=RUB1
58 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Rotlinné hormony a jejich signální dráhy Signalizace světlem

59

60


61 Effect of Gibberellins on growth and development d1 d1 WT WT +GA 1 +GA 1 Cabbage, a long-day plant, remains GA 1 stimulates stem elongation in corn dwarf mutant, but has no effect on tall WT a rosette under short day, but it can be induced to bolt and flower by applications of GAs
62 Signální dráha giberelinů

63

64

65
66
67 Oba kodují vysoce homologní Fbox podjednotku SCF E3 ligázového komplexu.

68

69 Signální dráha GA - analogická k auxinu
70
71
72
73

74 Signální dráha JA?


75 SHRNUTÍ - Ubiquitin-ligase based receptors ( auxin, jasmonates and gibberellin) GENES & DEVELOPMENT 22: (2008)
76 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Rotlinné hormony a jejich signální dráhy Signalizace světlem
77 Two-component or phospho-relay circuitry Prokaryota Eukaryota
78

79
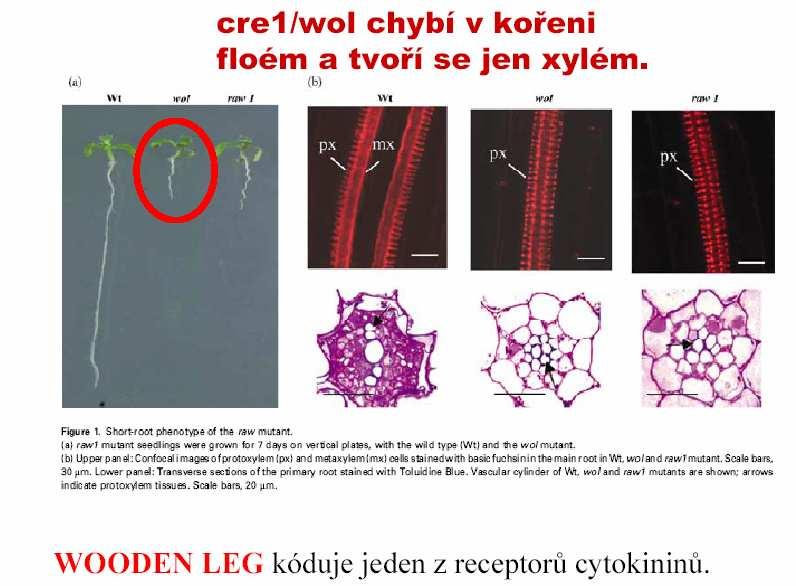
80
81

82 ARR A-typu se podílejí na negativně zpětnovazebném potlačení odpovědi na cytokininy.
83
84
85
86
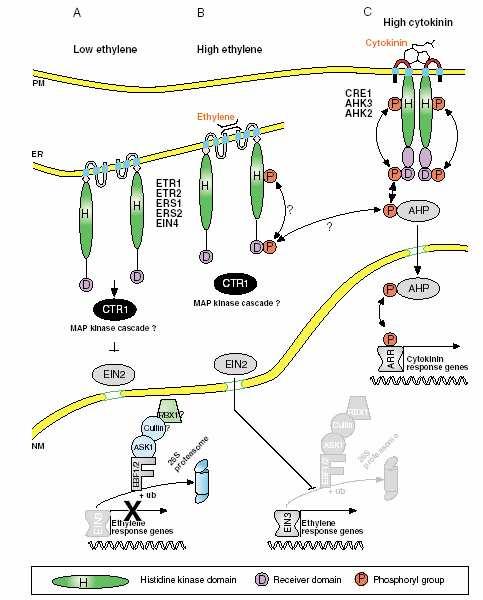
87 Signální dráha etylenu
88
89
90 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Rotlinné hormony a jejich signální dráhy Signalizace světlem
91
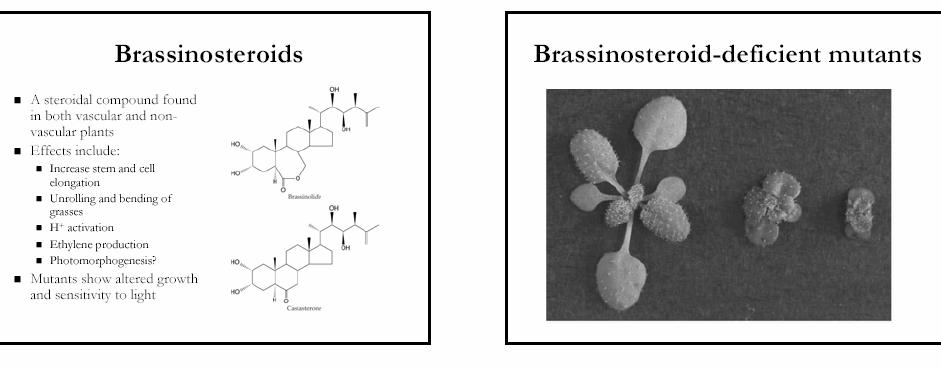
92

93
94 Brassinosteroid signaling pathway Development 133, (2006)

95

96 Příklady signalizací pomocí RLK
97 ABA

98
99 ABA různé signální dráhy Různé receptory?
100
101 Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních drah Auxin Giberellin Jasmonát (?) Signální dráhy založené na dvoukomponentní dráze Cytokininy Etylen Brassinosteroidy a signalizace pomocí proteinkináz ABA = mnoho funkcí a receptorů Peptidové hormony Signalizace světlem
102 PEPTIDY jako hormony
103
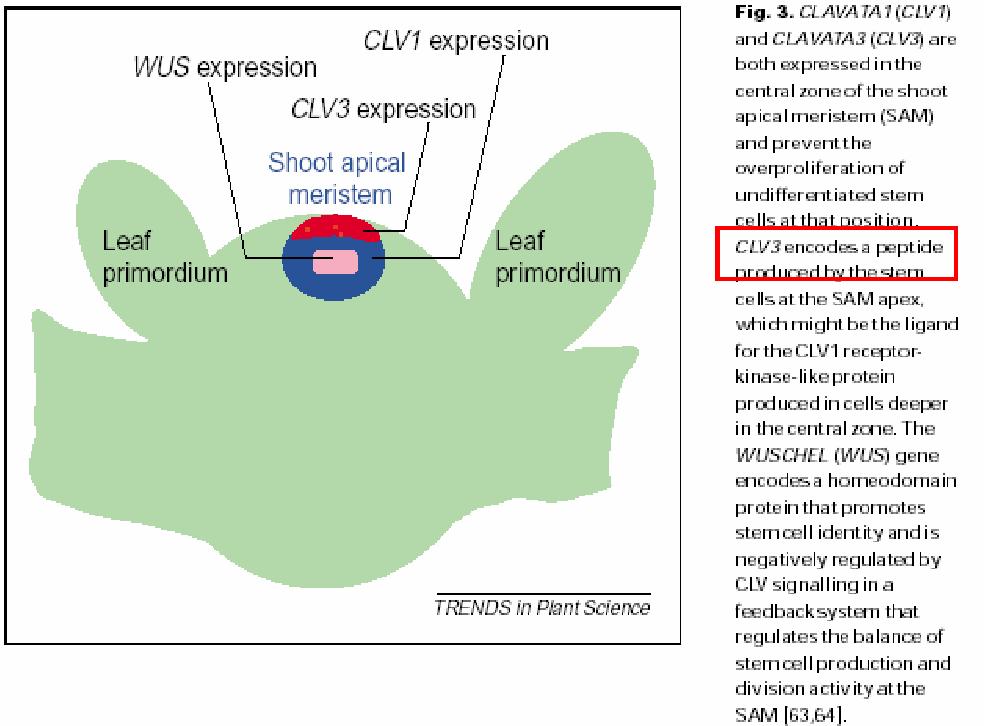
104
105 KONVERGENCE SIGNÁLNÍCH DRAH
106
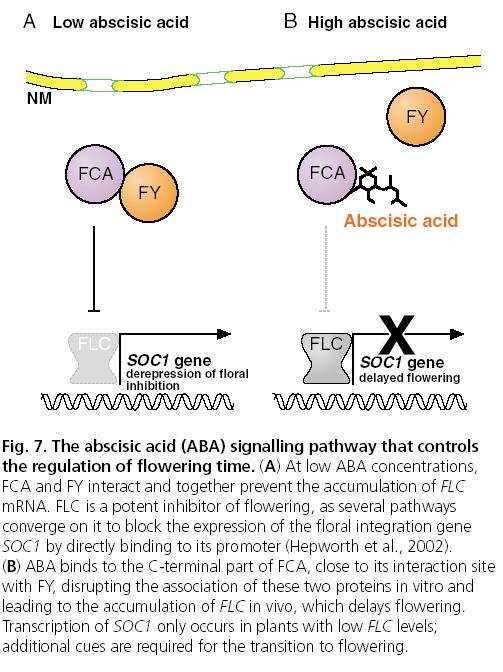
107 ABA GA

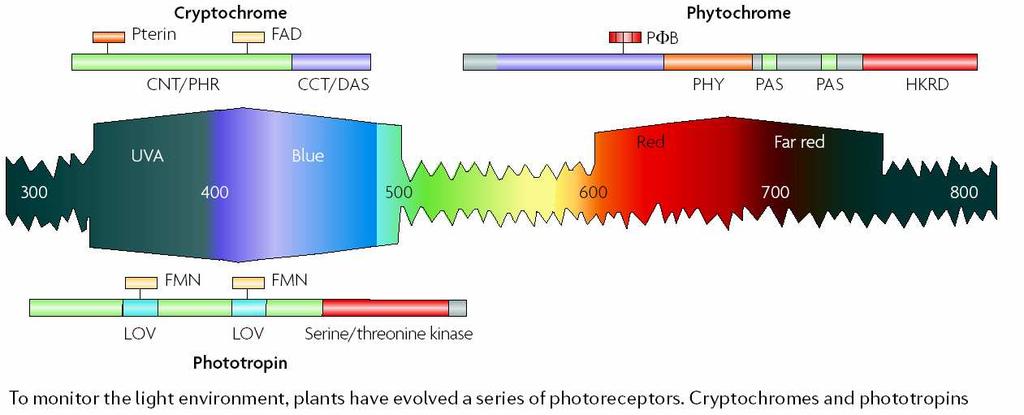
108
109 SVĚTLO jako SIGNÁL
110 Světlo Kvalita Kvantita Směr Trvání/Perioda UV B nm UV-A nm Modré s nm Červené s nm Dl. Červ. S nm
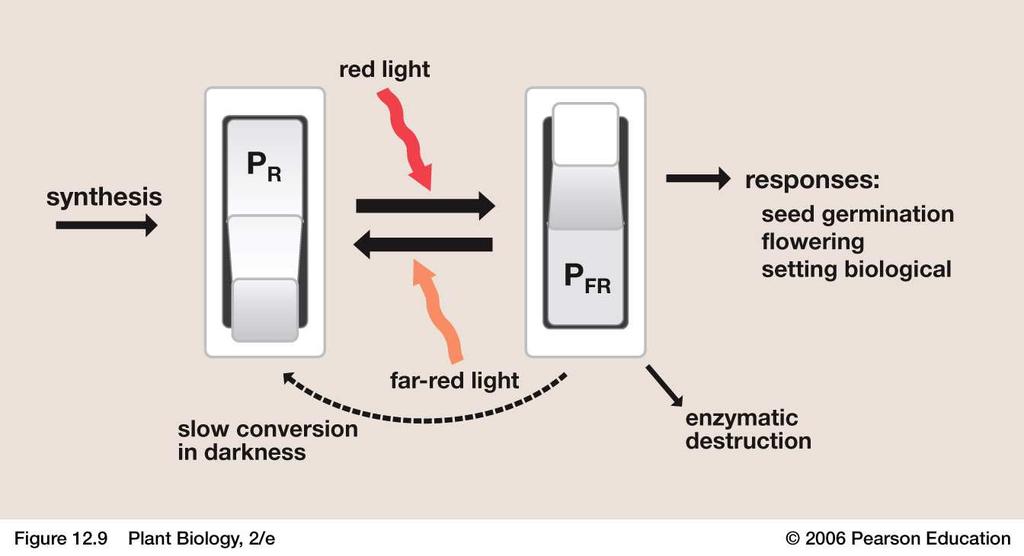
111 Light environment: sunlight UV B & A Visible light: nm or µm
112 Light quality variation during the day
113 Light quality variation due to location The light quality alters drastically depending on the situation. Direct sunlight Skylight (blue sky!) Under shade
114 Light quantity variation in nature The light intensity in nature varies at least ten orders of magnitude
115 A Light darkness directional light shade direct light reflected light cotyledon hypocotyl shade avoiding characteristic photomorphogenic phototropic etiolated


116 Borthwick s Experiment in Grand Rapids Lettuce (1954)
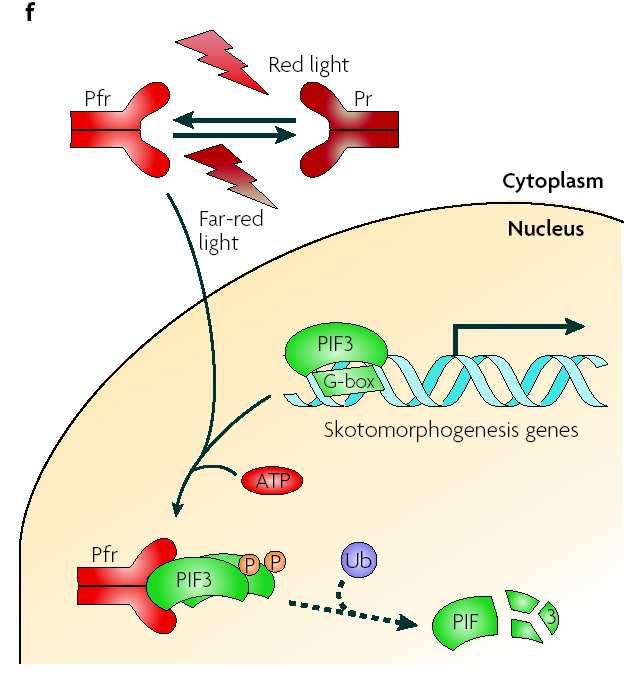
117 Fotoreceptory Fytochromy Kryptochromy Fototropiny (NPH1) Neznámý rec. UV-B

118
119

120 Absorption spectra of the two forms of Phytochrome
121
122 Fytochromy 5 genů v Arabidopsis (PhyA - PhyE) Všechny exprimovány +/- všude > posttranslační regulace PhyA a PhyB dominantní a nejvíce studovány PhyA je foto-labilní Phy B,C,D,E jsou foto-stabilní PhyA je nejen nestabilní na světle (poločas rozpadu PfrA ~ 1h), ale dochází také k poklesu jeho transkripce zprostředkovaného fytochromovým systémem
123 Phytochromes consist of a polypeptide (~125 kda, 1200 aa) and a covalently linked tetrapyrrole chromophore in the N-terminal part. Phytochromes are homodimers through the C-terminal dimerization (heterodimer?) All phytochromes seem to have identical chromophore The protein part of phytochrome is encoded by multigene family (phytochrome gene family) in all higher plants examined

124 Far-red darkness
125
126 Phy v Pfr formě jsou kinázy Autofosforylace, fosf. regulatory (Aux/IAA, TFs), kryptochromy Jen PfrB jde do jádra (FR to inhibuje) kin. hodiny. PhyA obě formy v jádře po osvětleníkin. 15min.
127
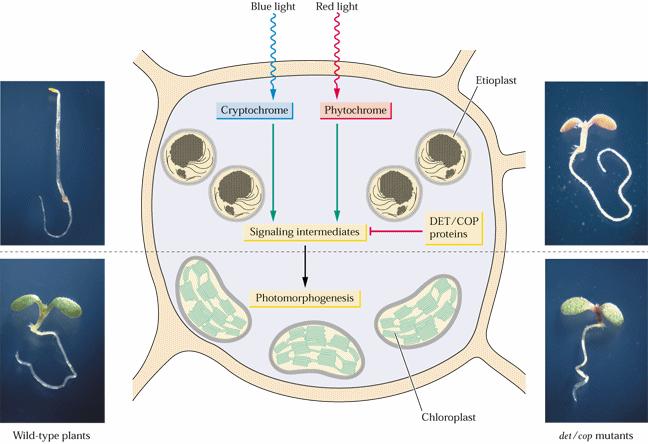
128 Fytochromoví mutanti Jak je hledat?

129 Jsou i na světle částečně slepí - a tedy vypadají jako ve tmě. cotyledon Photomorphogenic development Hypocotyl Skotomorphogenic development or Etiolation root
130
131 Phytochrome function specificity during early Arabidopsis development phya: phyb: phyc: phyd: phye: Far-red red Red Red Red Red De-etiolation etiolation (major) De-etiolation etiolation (major) Primary leaf expansion De-etiolation etiolation (minor) Internode suppression
132 PhyB se např. účastní úniku ze stínu
133
134 Kryptochromy Vyvinuly se z fotolyáz nezávisle u rostlin a živočichů
135 Cryptochromes Many plant responses were not R-FR reversible and had action spectra with peaks in the blue and near-uv. There must be a BL receptor(s). Due to their elusive nature, Gressel (1977) described BL receptors as cryptochromes. hy4 mutants showed a lack of hypocotyl growth inhibition under blue light,but were normal under red and far-red. Dark BL BL Wild-type cry1 adapted from Neff and Chory, 1998 The sequence of Hy4 was reported in 1993, and it was similar to DNA photolyase (Ahmad and Cashmore 1993, Sancar 1994) yet has no photolyase activity. Re-designated Cryptochrome 1 (Lin et al., 1995) Involvement in circadian rhythms; led to discovery of animal crys (Cashmore, 2003) cry s are phosphorylated when illuminated (Shilatin et al., 2003, Bouvy et al., 2003). The timing of phosphorylation agrees well with the time course of early physiology (Folta and Spalding, 2001)

136 The C-Terminus CCT of cry1 Regulates Photomorphogenesis Ectopic overexpression of the CRY1 C-terminal extension results in a constitutive-photomorphogenic phenotype. Cashmore, 2003
137 Pozitivní a negativní regulace fotomorfogeneze
138



139 Xing Wang Deng, Yale COP Mutants First isolated by Deng et al., Constitutive Photomorphogenic phenotype: expanded cotyledons, short hypocotyls, light-regulated gene expression patterns in darkness 1996 Mayer et al. show that COP1 mutation affects expression of many genes not just specific to photomorphogenesis.
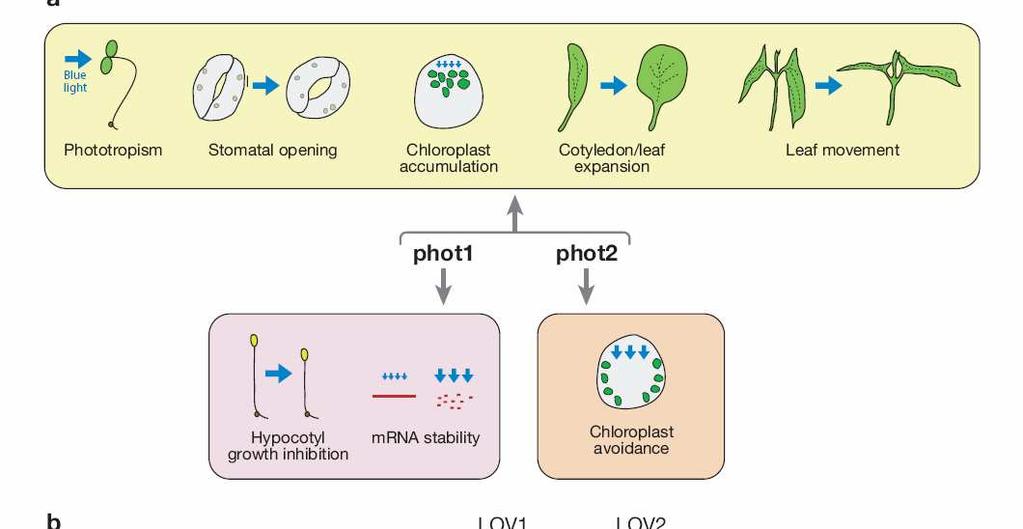
 COP9 byla")

140 CSN komplex (viz. nahoře) COP9 byla první známá podjednotka signalosomu.
141 HY5 is a positive regulator of photomorphogenesis and acts downstream of multiple photoreceptors Blue Red Far-red red hy1, hy2: Normal Long Long hy3/phyb: Normal Long Normal phya: Normal Normal Long Hy4/cry1: Long Normal Normal hy5: Long Long Long
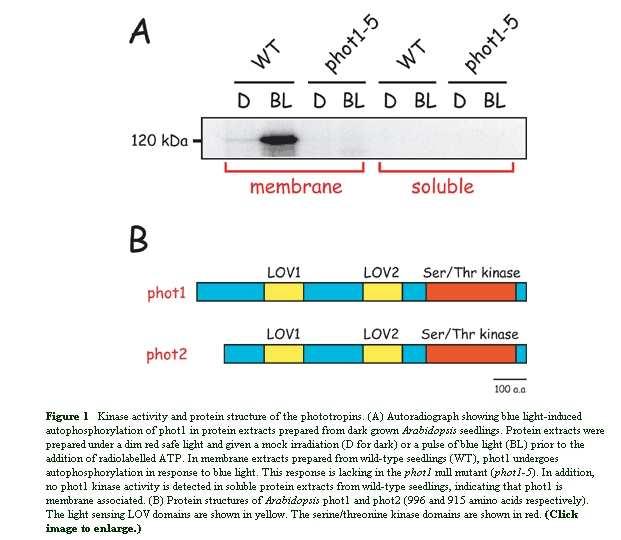
142 Far-red light Red light Blue/UV-A light phya phyb CRY1 CRY2 COPs HY5 etc Photomorphogenesis
143 HY5 and COP1 HY5 encode a bzip type DNA-binding transcription factor HY5 binds to promoters of light-regulated genes and mediates their light activation HY5 is a nuclear protein and directly interact with nuclear localized COP1

144 Komplexní regulace Hy5 Transkripční regulace světlem Degradace pomocí COP1 za tmy Fosforylace

145 Hy5 integruje signály z fytochromů a kryptochromů
146
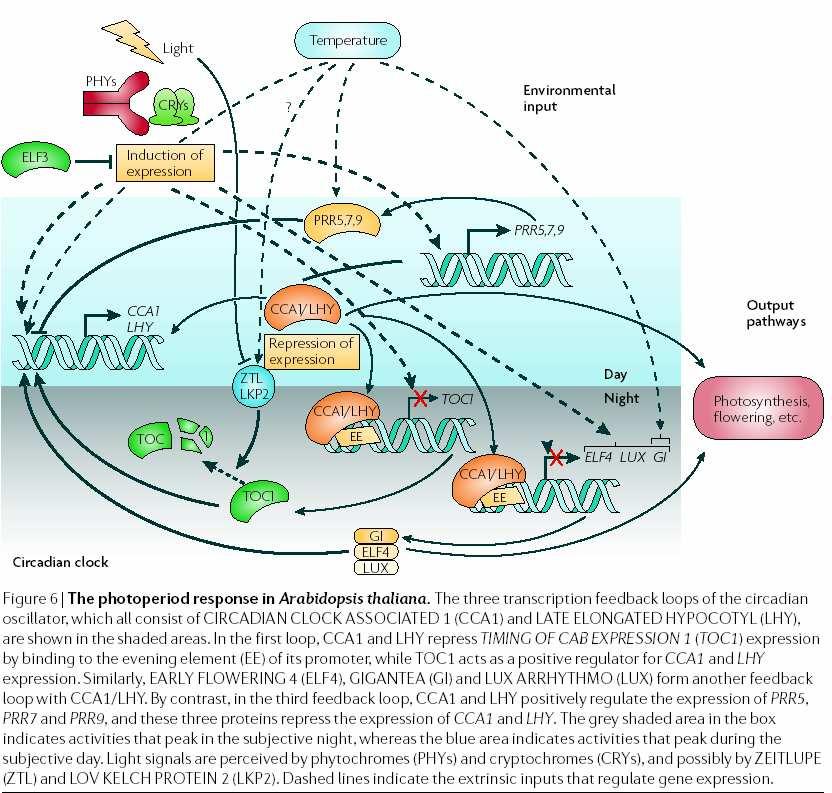
147 Fototropismus a FOTOTROPINY
148 opět auxin a jeho transport

149 CHLOROPLAST DARK WEAK BLUE LIGHT STRONG BLUE LIGHT MOVEMENTS -LEMNA
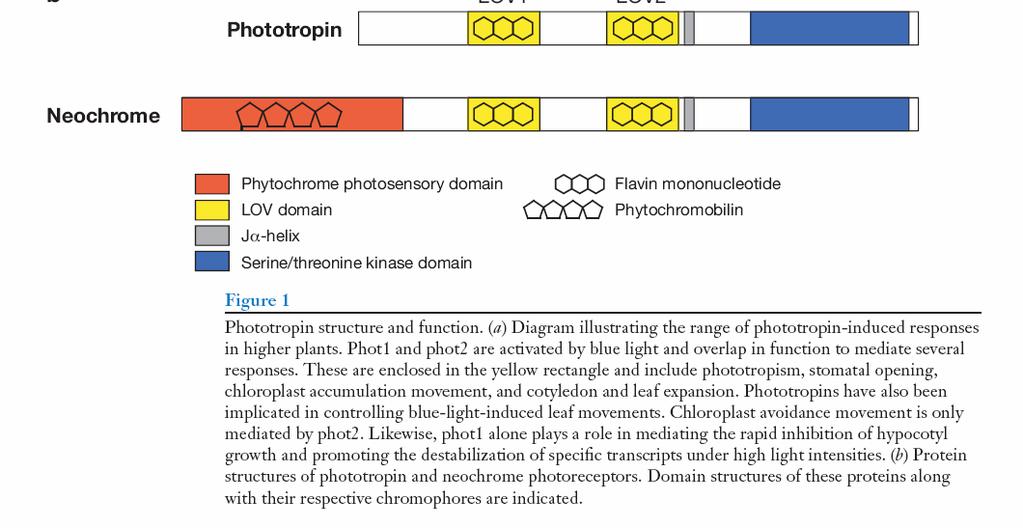
150 FOTOTROPINY Adiantum kapradina
151
152 Mechanismus asociace s membránou je nejasný.
153 Po osvětlení je část Phot1 bílkoviny uvolněna do cytoplazmy. V kortexu etiolovaného hypokotylu je Phot1 bílkovina přednostně polarizovaně kolokalizována s Pin1 na příčných stěnách.
154 Fotoperiodická signalizace a biologické hodiny
155

156
157 "Biologické hodiny" tikají v každé buňce rostliny a s vnějším časovým cyklem komunikují přes fytochormový/kryptochromový systém.
158 Transkriptomická analýza Arabidopsis ukázala, že asi 6000 genů je exprimováno v diurnálním rytmu, z toho asi 500 přímo v závislosti na "centrálním oscilátoru/hodinách"
159
160
161
Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu
 Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace genové exprese je represe (a její odblokování). Většina bílkovin
Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace genové exprese je represe (a její odblokování). Většina bílkovin
Signalizace je vlastně komunikace. a komunikace je základem života.
 Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace. a komunikace je základem života.
 Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů
 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
MFPSB 1. b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze
 Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze") 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner Martin Fellner 2010 2 a) Fotomorfogeneze
1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner Martin Fellner 2010 2 a) Fotomorfogeneze
9) Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer
 Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer") 2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
Příběh pátý: Auxinová signalisace
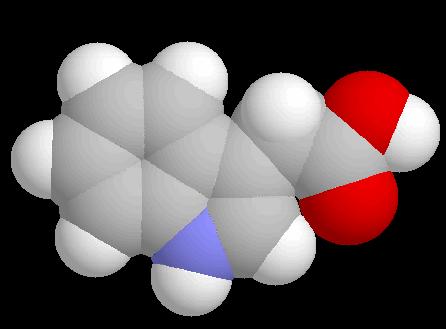
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
5) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") 2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
RVR e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu
 Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu") 1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
19. století. b) Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR
 Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR") SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
1) Úloha světla a fytochromů ve vývoji a růstu rostlin
 Úloha světla a fytochromů ve vývoji a růstu rostlin") S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
2) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
4) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
(molekulární) biologie buňky
 biologie buňky") (molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
(molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
VORF Winslow Russel Briggs ( )
") VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
Biosyntéza a degradace proteinů. Bruno Sopko
 Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př objevený Strigolacton
 Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding Protein1 byl objeven
Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding Protein1 byl objeven
2) Reprodukce rostlin
 Reprodukce rostlin") 2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora
 Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora Klinika dětské anesteziologie a resuscitace, Lékařská fakulta MU, Fakultní nemocnice Brno DNA transkripce
Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora Klinika dětské anesteziologie a resuscitace, Lékařská fakulta MU, Fakultní nemocnice Brno DNA transkripce
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
PŘENOS SIGNÁLU V BUŇCE. Nela Pavlíková
 PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL
 STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Joanne Chory (1955) VORF-4.
 VORF-4.") VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") 2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
Prokaryotická X eukaryotická buňka. Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen)
") Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,
Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
1 2015 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen MBR 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka
Detlef Weigel ( )
") VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
Teoretický úvod: FOTOTROPISMUS. Praktikum fyziologie rostlin
 Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS
 ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS IZOLACE FOSFOPROTEOMU A JEHO VYUŽITÍ PŘI STUDIU ÚČINKU CYTOKININŮ NA ROSTLINU Černý M., Brzobohatý B. Department
ISOLATION OF PHOSPHOPROTEOM AND ITS APPLICATION IN STUDY OF THE EFFECT OF CYTOKININ ON PLANTS IZOLACE FOSFOPROTEOMU A JEHO VYUŽITÍ PŘI STUDIU ÚČINKU CYTOKININŮ NA ROSTLINU Černý M., Brzobohatý B. Department
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI
 MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
MOŽNOSTI VYUŽITÍ BIOLOGICKY AKTIVNÍCH LÁTEK PŘI MOŘENÍ OSIVA SÓJI POSSIBILITIES OF USE BIOLOGICALLY ACTIVE AGENT FOR SOY SEED TREATMENT PAVEL PROCHÁZKA, PŘEMYSL ŠTRANC, KATEŘINA PAZDERŮ, JAROSLAV ŠTRANC
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
2) Reprodukce rostlin
 Reprodukce rostlin") 1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
1 2010 2) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Klíčení semen Speciální číslo Plant Cell, vol. 216 (June 2004) Supplement, pp. S1 S245, zaměřené na Plant Reproduction (Reprodukce rostlin)
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky
 Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
REPETITORIUM REGULACÍ
 REPETITORIUM REGULACÍ Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding
REPETITORIUM REGULACÍ Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA. nové...př. 2008 objevený Strigolacton Auxin DOMINANTNÍ MORFOGEN ABP1 Auxin Binding
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Rekapitulace. Rostlina vládne buňkám, ne(jen) buňky rostlině.
 buňky rostlině.") Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Rekapitulace Rostliny jsou od živočichů tak daleko, jak jen lze ( univerzální outgroup ). Mnohobuněčnost vznikla vícekrát tj. u rostlin a u živočichů nezávisle. Charakteristické rysy rostlinného organismu:
Regulace translace REGULACE TRANSLACE LOKALIZACE BÍLKOVIN V BUŇCE. 4. Lokalizace bílkovin v buňce. 1. Translační aparát. 2.
 Regulace translace 1. Translační aparát 2. Translace 3. Bílkoviny a jejich posttranslační modifikace a jejich degradace 5. Translace v mitochondriích a chloroplastech REGULACE TRANSLACE LOKALIZACE BÍLKOVIN
Regulace translace 1. Translační aparát 2. Translace 3. Bílkoviny a jejich posttranslační modifikace a jejich degradace 5. Translace v mitochondriích a chloroplastech REGULACE TRANSLACE LOKALIZACE BÍLKOVIN
Fyziologie AUTOFAGIE. MUDr. JAN VARADY KARIM FNO
 Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
Jaderné receptory. ligand. cytoplazmatická membrána. jaderný receptor DNA. - ligandem aktivované transkripční faktory
 Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
4) Reprodukce rostlin
 Reprodukce rostlin") MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
MBR 1 2015 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1 buňka 2 buňky Meristém
3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
REGULACE TRANSLACE DEGRADACE BÍLKOVIN. 4. Degradace bílkovin. 4. Degradace bílkovin. 4. Degradace bílkovin
 4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
4) Reprodukce rostlin
 Reprodukce rostlin") MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
MBR1 2016 4) Reprodukce rostlin g) Tvorba semen h) Dozrávání embrya i) Vývoj endospermu 1 2 g) Tvorba semen Semeno krytosemenných: - embryo - endosperm - testa (obal) Vývoj embrya Globulární stádium 1
Biosyntéza vychází z terpenoidu geranylgeranyldifosfátu. Vznik ent-kaurenu je první krok vlastní dráhy biosyntézy giberelinů.
 Gibberelliny Biosyntéza vychází z terpenoidu geranylgeranyldifosfátu. Vznik ent-kaurenu je první krok vlastní dráhy biosyntézy giberelinů. Ze stejných počátků vychází také dráha biosyntézy brassinosteroidů
Gibberelliny Biosyntéza vychází z terpenoidu geranylgeranyldifosfátu. Vznik ent-kaurenu je první krok vlastní dráhy biosyntézy giberelinů. Ze stejných počátků vychází také dráha biosyntézy brassinosteroidů
7) Senescence a programová smrt buňky (PCD)
 Senescence a programová smrt buňky (PCD)") MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
MBRO2 1 2018 7) Senescence a programová smrt buňky (PCD) c) Senescence a rostlinné hormony d) Vývojová PCD e) PCD a reakce rostlin ke stresu Nejnovější review: Obecné: Kast DJ, Dominguez R (2017) Current
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Joanne Chory ( ) narozena v Massachusetts. má 4 sourozence. manžel Stephen Worland, biochemik. děti Katherine and Joe
 narozena v Massachusetts. má 4 sourozence. manžel Stephen Worland, biochemik. děti Katherine and Joe") 23.10.2017 VORF-4 2017 Joanne Chory (19.3. 1955) 1 narozena v Massachusetts 2 Salk Institute for Biological Studies lant Molecular and Cellular Biology Laboratory 10010 North Torrey ines Road La Jolla,
23.10.2017 VORF-4 2017 Joanne Chory (19.3. 1955) 1 narozena v Massachusetts 2 Salk Institute for Biological Studies lant Molecular and Cellular Biology Laboratory 10010 North Torrey ines Road La Jolla,
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES
 BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES Vlašánková E.*, Kohout L.**, Klemš M.*, Hradilík J.* *Ústav botaniky a fyziologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická
BRASSINOSTEROIDS AND WATER STRESS BRASSINOSTEROIDY A VODNÍ STRES Vlašánková E.*, Kohout L.**, Klemš M.*, Hradilík J.* *Ústav botaniky a fyziologie rostlin, Agronomická fakulta, Mendelova zemědělská a lesnická