Plazmidy podstatná složka genomu bakterií
|
|
|
- Michaela Staňková
- před 6 lety
- Počet zobrazení:
Transkript
1 Plazmidy podstatná složka genomu bakterií Definice: Plazmidy jsou extrachromozomální, autonomně se replikující genetické elementy, které se vyskytují v buňkách všech skupin mikroorganismů. Význam: 1. Podstatně ovlivňují základní biologické vlastnosti svých hostitelů, podílejí se na jejich diverzitě a evoluci 2. Jsou využívány k poznání základních molekulárněbiologických procesů v bakteriálních buňkách a horizontálnímu přenosu genů 3. Jsou prakticky a široce využívány v metodách MB a GI, zejména jako vektory
2 CHARAKTERISTIKA PLAZMIDŮ dsdna kružnicová nebo lineární, velikost: kb Základní typy plazmidů: kryptické - funkce neznámá epizomální- reverzibilní intergace do chromozomu hostitele konjugativní - schopné přenosu konjugací mobilizovatelné přenositelné za přítomnosti konjugativního plazmidu Příklady plazmidů: F-plazmidy (fertilitní faktor, konjugativní) zodpovědné za konjugaci, příp. mobilizaci jiných plazmidů R-plazmidy (R-faktory) zodpovědné za rezistenci k antibiotikům, řada z nich konjugativní kolicinogenní (Col-plazmidy) (bakteriocinogenní plazmidy) tvorba proteinů s antibiotikovým charakterem (Enterobacteriaceae) Virulenční plazmidy Ti-plazmidy (tumory indukující) tvorba nádorů u dvouděložných rostlin (Agrobacterium tumefaciens) Plazmidy odbourávající organické sloučeniny (Pseudomonas) Plazmidy podílející se na fixaci vzdušného dusíku (Rhizobium). Plazmidy používané jako vektory pro přenos DNA (pbr322, puc)

3 PLAZMIDY
4 Plazmidy archeí - málo prostudované, nejlépe u metanogenních archeí - většinou kryptické - některé mají charakter megaplazmidů (např. u Haloferax volcanii) velikost 690, 442 a 86 kb, - identifikované geny: ---- geny pro tvorbu plynových měchýřků ---- geny kódující restrikční endonukleázy a metylázy - konjugativní plazmidy (r. Sulfolobus) schopnost integrace do chromozomu, jednosměrný přenos

5
6 KLASIFIKACE PLAZMIDŮ označování: pxy123 Univerzální vlastností plazmidů je jejich inkompatibilita, kterou se rozumí neschopnost dvou plazmidů koexistovat společně v téže buňce (navzájem se vytěsňují). Kompatibilitou se rozumí schopnost dvou plazmidů koexistovat v jedné buňce bez selekčního tlaku a stabilně se dědit. Rozlišuje se mezi vektoriální a symetrickou inkompatibilitou. - vektoriální: je ztracen vždy jeden konkrétní plazmid ze dvou. - symetrická: každý z plazmidů je ztrácen při stejné pravděpodobnosti. Do stejné inkompatibilní skupiny náležejí navzájem inkompatibilní plazmidy. Inkompatibilní plazmidy jsou vzájemně příbuzné (využívají tentýž mechanismus kontroly replikace). V současné době je známo asi 30 inkompatibilních skupin u enterobaktérií, 9 u stafylokoků atd.
7 Důkaz přítomnosti plazmidu v bakteriálních buňkách 1. Elektroforéza po izolaci plazmidové DNA 2. Průkaz CCC DNA v elektronovém mikroskopu. 3. Centrifugační metody: - důkaz satelitního pruhu v sacharozovém gradientu podle S. - důkaz v CsCl-EB podle konformace CCC, LIN a OC. Charakterizace plazmidů - stanovení velikosti: elektronmikroskopicky nebo v agarozovém gelu, kde se nejdříve linearizuje - konstrukce restrikční mapy, sekvencování DNA - zařazení plazmidu do inkompatibilní skupiny. Za tímto účelem je plazmid přenesen do vhodných recipientních testovacích kmenů, které již obsahují známé plazmidy příslušných kompatibilních skupin. Předpokladem je to, aby oba plazmidy měly různé genetické markery, např. dva různé geny pro rezistence. Při Inc-testu se kmen pomnožuje nejdříve bez selekčního tlaku generací a pak se vzniklé kolonie testují na přítomnost obou plazmidů. Pro stanovení příbuznosti dvou plazmidů lze použít i srovnání jejich restrikčních map. Přesnější analýza určitých oblastí se pak může provést hybridizací. Poslední krok je sekvencování DNA.



8 Důkaz přítomnosti plazmidu v bakteriálních buňkách 1. Průkaz DNA v elektronovém mikroskopu 2. Centrifugační metody: důkaz satelitního pruhu po centrifugaci 3. Elektroforéza DNA v agarózovém gelu P P Ch Ch P
9 DŮKAZ PROTEINŮ KÓDOVANÝCH PLAZMIDEM (E. coli) A. Systém využívající minibuňky: není přítomen chromozom, ale plazmidy (inkubovány v minimálním mediu za přítomnosti značených aminokyselin (35S-metionin). Proteiny jsou detekovány na SDS-PAGE a prokázány autoradiograficky. B. Systém využívající maxibuňky. Po ozáření UV-světlem se chromozomové geny v důsledku poškození neexprimují, většina plazmidů za těchto podmínek zůstává díky své malé velikosti intaktní a může geny exprimovat. Důkaz tvorby proteinů probíhá analogicky jako u minibuněk. C. Systém translace in vitro (Zubayův systém), který se skládá z testované plazmidové DNA, ze supernatantu po centrifugaci lyzátu buněk E. coli, obsahujícího proteinové komponenty nutné pro transkripci a translaci (RNApolymeráza, ribozomy, translační faktory, 19 aminokyselin a jedné aminokyseliny značené, rntp, systému regenerujícího energii a další). Důkaz tvorby proteinů probíhá analogicky jako u minibuněk.
10 ODSTRAŇOVÁNÍ PLAZMIDŮ ( plasmid CURING, LÉČENÍ) změna fenotypu = důkaz, že určitá funkce je spjata s přítomností plazmidu A. Interkalační barviva: Akriflavin, akridinoranž, etidiumbromid a quinarcin patří mezi interkalační barviva, které se začleňují mezi sousední báze a zabraňují replikaci plazmidů. B. Coumermycin a novobiocin: Interference s účinkem DNA-gyrázy, který zavádí negativní superhelikální otáčky do kruhové dsdna. C. Rifampicin a mitomycin C: Rifampicin se váže na RNA polymerázu a zabraňuje tak transkripci. Mitomycin C je metabolicky aktivován na intermediát, který kroslinkuje DNA řetězce a blokuje tak transkripci. D. Natriumdodecylsulfát: Narušuje vazbu plazmidu k b. membráně E: Další metody: Zvýšená teplota, skladování kultur, regenerace protoplastů Komplementární přístup: přenos plazmidu do bezplazmidových buněk
11 LOKALIZACE ORI NA PLAZMIDOVÉ MOLE- KULE, STANOVENÍ ZPŮSOBU REPLIKACE Štěpení plazmidů v různých stadiích replikace restrikčním enzymem, sledování struktury v EM
12 VYHLEDÁVÁNÍ (IZOLACE) POČÁTKU REPLIKACE (ORI) NA PLAZMIDU Selektovatelný gen
 3.")
13 PODLE POČTU KOPIÍ SE PLAZMIDY DĚLÍ DO TŘÍ SKUPIN 1. S nízkým počtem kopií (1-2/chr) 2. Se středním počtem kopií (asi 15) 3. S vysokým počtem kopií (více jak 15) (dělení je umělé a hranice není pevná
14 STANOVENÍ POČTU PLAZMIDOVÝCH KOPIÍ POČET KOPIÍ = molární hmotnost chromozomu (kb) radioaktivita plazmidu (cpm) x molární hmotnost plazmidu (kb) radioaktivita chromozomu (cpm) -Ultracentrifugace v CsCl-EB s radioaktivně značenou DNA, stanovení radioaktivity v jednotlivých frakcích (pruzích) -Elektroforéza v gelu, stanovení množství DNA
15 STANOVENÍ ČETNOSTI REPLIKACE PLAZMIDŮ CENTRIFUGACÍ V GRADIENTU CsCl Plazmid, který se nereplikoval Plazmid, který se replikoval 1x 0,3 generace 0,5 generace 1,0 generace 15N 14N/15N 14N Plazmid, který se replikoval dvakrát nebo vícekrát
16 MECHANISMY REPLIKACE PLAZMIDŮ Začíná ve specifickém místě (oriv) vytvořením volné 3 OH skupiny (buď RNA-primer, nebo zlom DNA) malé plazmidy: mechanismus otáčivé kružnice (RC-plazmidy) velké plazmidy: (theta mechanismus) mechanismus podobný replikaci chromozomu je vyžadován Rep protein a další proteiny hostitele (DnaA, B, G aj); v místě ori je po vazbě iniciačních faktorů syntetizována primerová-rna (plazmidy typu ColE1 nevyžadují pro tvorbu primerové-rna žádný protein kódovaný plazmidem)
17 STRATEGIE KONTROLY POČTU PLAZMIDOVÝCH KOPIÍ Inhibitor cíl Plazmid kóduje difuzibilní inhibitor replikace, který se váže na cílovou sekvenci Přímá regulace Nepřímá regulace Vazba inhibitoru brání iniciaci replikace Vazba inhibitoru brání syntéze produktu nezbytného pro zahájení replikace Iteronová strategie Regulační protein (rep) nezbytný k zahájení replikace je vázán (titrován opakujícími se sekvencemi (iterony) poblíž ori. Replikace nastává po nasyntetizování dostatečného množství volného regulačního proteinu.
18 Regulace replikace ColE1 plazmidu a jeho derivátů Preprimer = cíl inhibitor rop (regulation of primer) rom (RNA One inhibition Moderator). Kissing komplex Párování RNAI a prnaprekurzoru (RNAII) zabraňuje maturaci primerového transkriptu
19 INTERAKCE RNAI S RNAII PŘI INCIACI REPLIKACE ColE1 ori V + Rop Mutace v genu rop vedou ke zvýšení počtu kopií plazmidu Po spárování RNAI a RNAII se změní sekundární struktura RNAII (preprimeru), který pak není RNázouH upraven na primer
20 Interakce RNAI, RNAII a proteinu Rom (~Rop) při iniciaci replikace ColE1 plazmidu Mutace v genu pro protein Rop vedou ke změně počtu plazmidových kopií

21 REGULACE REPLIKACE PLAZMIDU ColE1 RNAI (inhibitor) je syntetizován konstitutivně a ve vyšší koncentraci než než preprimerová RNAII. Rostoucí bakterie zvětšuje objem, čímž se snižuje pravděpodobnost interakce RNAI a RNAII a dochází k zahájení replikace. Další možnost regulace je dána působením proteinu Rop, jehož množství v buňkách může kolísat (regulace na úrovni transkripce nebo translace, případně jeho stabilita)
22 KONTROLA REPLIKACE ITERONOVÝCH PLAZMIDŮ Plazmid kóduje difuzibilní iniciační protein Rep, který se váže na řadu přímých opakování - iteronů (různý počet u různých plazmidů). Tento protein má následující vlastnosti: iniciuje nové cykly replikace může zabraňovat replikaci funguje jako autorepresor na úrovni transkripce Působení Rep je závislé na jeho koncentraci, která je ovlivňována počtem plazmidových kopií (resp. počtem iteronů) a rozdílnou afinitou Rep k iteronům (L a R) v oblasti ori.
23 PŮSOBENÍ PROTEINU RepA NA INICIACI REPLIKACE U ITERONOVÉHO PLAZMIDU psc101 Vazba DnaA a dalších chr. autoreprese aktivace replikace RepA R = CAAAGGTCTAGCAGCAGAATT-(TACAGA-R3) nízká koncentrace RepA replikace; vysoká koncentrace RepA autoreprese

24 KONTROLA REGULACE INICIACE REPLIKACE PLAZMIDU F čtyři L-iterony pět R-iteronů rep Rep Slabší vazba Rep iniciace replikace při vyšších koncentracích Rep represe Silnější vazba Rep zábrana replikace při nízkých koncentracích Rep Experimentální pozorování: delece R-iteronů vede ke zvýšení a adice dodatečných R-iteronů ke snížení počtu kopií
25 DVOJÍ PŮSOBENÍ RepA PROTEINU ITERONOVÝCH PLAZMIDŮ coupling model RepA se váže jen na jeden plazmid a iniciuje replikaci RepA se váže na dva plazmidy současně a spojuje je, čímž brání replikaci Důkaz EM
26 MECHANISMY ZAJIŠŤUJÍCÍ STABILNÍ UDRŽENÍ PLAZMIDŮ V BUŇKÁCH (SEGREGAČNÍ STABILITA PLAZMIDŮ) u vícekopiových plazmidů (>20) typu ColE1 je přítomen nekódující úsek (cer, 380 bp), který je cílem působení místně specifické rekombinázy (kódovaná geny xerc a xerd na chromozomu) Multimer resolution systems u větších plazmidů jsou rekombinázy kódovány plazmidovými geny u nízkokopiových plazmidů jsou přítomny aktivní mechanismy rozdělování: aktivní segregace: partitioning (lokud inc + proteiny Par(Sop)) postsegregační usmrcování buněk, které ztratily plazmid (interakce stabilního toxinu s nestabilním antitoxinem)
27 SYSTÉM PRO ROZDĚLOVÁNÍ MULTIMERNÍCH PLAZMIDOVÝCH FOREM Přímá opakování Resolváza
28 Xer FUNKCE E. Coli KATALYZUJE MÍSTNĚ-SPECIFICKOU REKOMBINACI V MÍSTECH CER PRO ROZKLAD PLAZMIDŮ ColE1 Proteiny XerC a XerD E.coli Působí na místo dif poblíž ter u E. coli
29 ZAJIŠTĚNÍ POČTU KOPIÍ SYSTÉMEM CER A XER Plazmid obsahuje cer Plazmid neobsahuje cer
30 MODEL PŮSOBENÍ FUNKCE SOP (PAR) U F PLAZMIDU Sop = stability of plasmid 3 kb Par Par = partitioning Proteiny hostitele Vnitřní membrána sopa kóduje protein, který zřejmě reguluje expresi sopb: protein SopB se váže spolu s proteiny hostitele na lokus sopc (místo pars) obsahující 12 tandemových 43 bprepetic a je ukotven na vnitřní membránu
31 Intracelulární lokalizace plazmidů Metoda: využití fluorescenčního značení proteinů vázajících se na sekvence plazmidů Závěry: 1. Plazmidy mají v buňkách specifickou nebo preferovanou lokalizaci: obvykle poblíž první a třetí čtvrtiny buňky 2. Lokalizace plazmidu v buňce závisí na jeho rozdělovacím systému 3. U rychle rostoucích buněk jsou plazmidy ve dvojicích nebo skupinách, které využívají společné připojovací místo na membráně (počet fluorescenčních signálů je nižší než počet plazmidových kopií) 4. Různé typy (druhy) plazmidů využívají různá místa pro přichycení při jejich dělení a v buňce se nacházejí na různých místech. Celkový závěr: platí obecný model, podle kterého je signál pro rozdělení plazmidu u dělících se buněk uprostřed a po rozdělení buňky se přesunuje do první a třetí čtvrtiny buňky.
32 Obecné schéma znázorňující rozdělování plazmidů Rozdělovací aparát (jeho složky nejsou dosud známy) Nově zreplikované plazmidy umístěné ve středu buňky se oddělí a přesunou do první a třetí čtvrtiny ještě před tvorbou septa Po rozdělení buňky se tyto polohy stávají v dceřiných buňkách opět středovými
33 MODELY ZNÁZORŇUJÍCÍ FUNKCI MÍST PAR U PLAZMIDŮ F A RP1 Fluorescenčně značené proteiny Par (ParA-GFP; ParB-GFP) A. Plazmid se váže na místo par na membráně. Při dělení buňky jsou kopie plazmidu přichyceny na rozdělená místa par a taženy do dceřinných buněk B. Dvě kopie plazmidu se nejdříve navážou na místo par a při jeho rozdělení jsou odděleny

34 Vizualizace polohy plazmidů F a P1 uvnitř buněk E. coli pomocí fluorescenčního barvení různé plazmidy se nacházejí v různých místech F - červená P1 - zelená Membrána - modrá
35 Obecný mechanismus genetického donucování (závislosti, addiction systems) Na plazmidu jsou přítomny dva geny, kódující produkty: 1. toxin (killer) - stabilnější 2. antitoxin (antidot, antikiller) - nestabilní Plazmid je přítomen Plazmid není přítomen
36
37 Ccd = couples cell division (geny týkající se buněčného dělení)
38 POSTSEGREGAČNÍ USMRCOVÁNÍ BEZPLAZMIDOVÝCH BUNĚK PŮSOBENÍM PROTEINOVÝCH SYSTÉMŮ Negativní regulace - v nadbytku operon Degradace antitoxinu Nefunkční antitoxin Smrt buňky
39 SLOŽKY PROTEINOVÝCH SYSTÉMŮ PRO UDRŽENÍ PLAZMIDU V BUŇCE Proteiny pro replikaci chromozomu buňky

40 RM-systémy typu II nesené na plazmidu (EcoRI)
41 C. Anti-sense-RNA-regulated systems
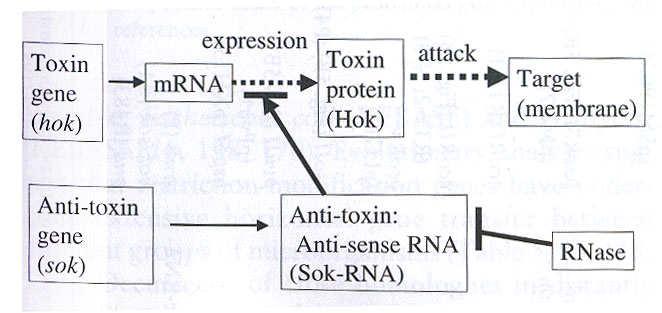
42 POSTSEGREGAČNÍ ZABÍJENÍ BEZPLAZMIDOVÝCH BUNĚK PŮSOBENÍM SYSTÉMU HOK/SOK U PLAZMIDU R1 hok = host killing (toxin); sok = suppression of killing (antitoxin) Antisense RNA je nestabilní a po ztrátě plazmidu z buněk rychle zmizí Hok protein je stabilní a po ztrátě plazmidu z buněk navodí kolaps membránového potenciálu
43 Plazmidy zodpovědné za rezistenci bakterií k antibiotikům (R-plazmidy) SLOŽKY R-PLAZMIDŮ 1. Resistance transfer factor (RTF) nese geny regulující DNA replikaci a počet kopií, geny pro přenos a někdy geny pro rezistenci k tetracyklinu. 2. R determinanta má různou velikost a nese další geny pro rezistenci k antibiotikům obvykle ampicilinu, chloramfenikolu, streptomycinu, kanamycinu, sulfonamidu v různých kombinacích. IS element + RTF (geny pro přenos a replikaci, + TetR) R determinanta (geny pro rezistence) R plazmid Vystavením buněk antibiotiku dochází k amplifikaci R-složky a zvětšuje se velikost celého plazmidu
44 ÚLOHA TRANSPOZONŮ PŘI EVOLUCI R-PLAZMIDŮ každý transpozon může být přenášen nezávisle Segment obsahující transpozony nesoucí determinanty rezistence Segment pro přenos determinant
45 Kolicinogenní plazmidy produkují látky s antibiotických charakterem PŘÍKLADY BAKTERIÁLNÍCH DRUHŮ PRODUKUJÍCÍCH BAKTERIOCINY Bakteriociny inhibují jednu nebo více základních funkcí (replikace, transkripce nebo translace, nebo energetický metabolismus). Na nárůstu citlivých buněk vytvářejí sterilní zóny lakuny. Pravé koliciny x defektní fágové částice (fágové bičíky, lytické enzymy)
46 STRUKTURA F PLAZMIDU (100 kb, 49% GC) 60 genů rozdělených podle funkce do 5 oblastí 1. Tranferová oblast 33 kb, 31 genů všechny funkce nezbytné pro přenos. Biosyntéza a sestavování F pilu (traa,b,c,e,f,g,h,l,k,q,u,v,w) Stabilizace párujících se buněk (trag,n) Konjugativní metabolismus DNA (trad,i,m,y,z) Regulace přenostu (traj,fino,finp) Povrchová exkluze (tras,t) 2. Vedoucí oblast Oblast, která je při přenosu DNA přenášena jako první. 13 kb. 3. Oblast replikace Tři oblasti s geny a sekvencemi podílejícími se na replikaci. 4. Oblast s inzerčními sekvencemi (IS3a,b, IS2, Tn1000) 5. Pif oblast Pif (phage inhibition by F). Tři geny pifc.
47 HLAVNÍ FUNKČNÍ OBLASTI F-PLAZMIDU
48
49 CHARAKTERISTIKA Ti-PLAZMIDŮ ( kb) Hostitel: Agrobacterium tumefaciens Agrobacterium rubi Agrobacterium rhizogenes (Ri-plazmidy) Ti způsobuje krčkové nádory (crown galls) Ri způsobuje vlasaté kořeny (hairy roots) Geny na plazmidu: T-DNA (15-45 kb) syntéza opinů a fytohormonů Geny pro virulenci Geny pro katabolismus opinů Ori Geny pro konjugaci Typy Ti-plazmidů (/podle typu opinu) nopalinový oktopinový agropinový sukcinamopionový Typy Ri-plazmidů manopinový agropinový
50 Struktura opinů 1. Rodina oktopinů Vznik kondenzací pyruvátu s některou aminokyselinou: - Pyruvát + arginin: oktopin - Pyruvát + lyzin: lyzopin - Pyruvát + histidin: histopin - Pyruvát + ornitin: kys. oktopinová 2. Rodina nopalinů Vznik kondenzací a-ketoglutarátu s aminokyselinami 3. Rodina agropinů Vznik kondenzací kys. glutamové s aminokyselinami
51 Chemická struktura opinů (zdroj C, příp. N)



52 Transformace rostlin navozená Ti-plazmidem Agr. tumefaciens 1. Poraněná rostlina Vytvoření krčkového nádoru 5. fytohormony opiny T-DNA 2. Infekce rostliny bakterií s Ti-plazmidem 4. Začlenění T-DNA do jádra rostlinné buňky 3. Přenos T-DNA do rostlinné buňky

53 Struktura Ti-plazmidu A. tumefaciens Geny pro přenos T-DNA do rostlinných buněk 25 bp LB Část plazmidu přenášená do rostliny 25 bp RB = sekvence nezbytná pro začlenění T-DNA do genomu rostliny Geny pro katabolismus opinů Geny zodpovědné za transformaci rostlinných pletiv Geny zodpovědné za tvorbu opinů v rostlinných buňkách
54 Mozaikovitá struktura Ti-PLAZMIDU Konjugativní přenos Ti-plazmidu do bakterií
55 PRŮBĚH PŘENOSU Ti-PLAZMIDU DO ROSTLINNÉ BUŇKY

56 PŘENOS T-DNA Z AGROBAKTERIA DO ROSTLINNÉ BUŇKY agrobakterium rostlinná buňka
57 PRŮBĚH PŘENOSU T-DNA Z AGROBAKTERIÍ DO ROSTLIN 1. Poranění rostliny, sekrece fenolických látek do prostředí (typu acetosyringonu: acetovanilon, hydroxyacetofenon aj.) 2. Reakce bakterií A. tumefaciens na fenolické látky (pozitivní chemotaxe) Aktivace genů vir na Ti-plazmidu Připojení bakterií k rostlinným buňkám (spolupůsobení chromozomových genů chva, chvb, psca) 3. Aktivace transkripce genů virb, C, D a E prostřednictvím proteinu kódovaného genem virg. 4. Působení produktů genů vir: Vznik jednovláknových zlomů na Ti-plazmidu (produkt genu vird) Tvorba jednovláknových kopií T-DNA 5. Vytvoření přenosového komplexu T-DNA a polypeptidů vird a vire 6. Přenos komplexu T-DNA do rostlinné buňky 7. Přenos T-DNA do jádra rostlinné buňky 8. Integrace T-DNA do chromozomu (různá místa, částečná homologie sekvencí RB a LB s místy začlenění)
58 FUNKCE PLAZMIDOVÝCH GENŮ VIRULENCE Geny vir = skupina genů v úseku asi 35 kb, velmi podobném u různých Ti-plazmidů Celkem asi 25 vir genů (7 operonů): vira,b,c,d,e a G, vytvářejících 20 polypeptidů a) tvorba jednovláknových zlomů na T-DNA: vird1, vird2 b) odkrucování jednovláknové T-DNA: vird1, vird2 a vire2 c) vytvoření přenosového komplexu T- DNA/proteiny: vird1, vird2 (virb) d) rozmezí hostitelů: virc
59 HLAVNÍ RYSY PŘENOSU T-DNA DO ROSTLINNÉ BUŇKY 1. Do rostlinné buňky je z Ti-plazmidu přenášena pouze T- DNA, ve formě ssdna 2. Při integraci T-DNA do rostlinného genomu dojde k definovanému začlenění 5 konce (1-2 bp BR), začlenění 3 konce je variabilní (vznik delecí bp) 3. Do rostlinného genomu se může začlenit více kopií T- DNA, i tandemově (vznik tandemů není jasný) 4. Pro přenos je nutná pouze 25 bp BR 5. Místo integrace je náhodné, nejsou vyžadovány úseky homologie preferenčně probíhá do transkripčně aktivních oblastí (často místa bohatá na AT páry) 6. Vlastní proces integrace T-DNA do genomu rostliny není jasný účast reparačních enzymů rostliny,? Produkty genů na T-DNA 7. V místě integrace dochází k přeskupením (delece, přeskupení apod.)
60 Symbiotické plazmidy Rhizobiaceae (psyn) Diazotrofie: klíčový proces v biosféře přeměna N2 na redukované formy (amonium) - volně žijící bakterie uskutečňují tento proces pro svou potřebu při omezením množství amoniaku nebo nitrátů - symbiotické mohou fixovat dusík jen po vytvoření mutualistických interakcí s leguminózami Předpoklady pro ustavení účinné symbiozy mezi bakteriemi a rostlinou: - chemotaktická invaze bakterií do kořenových buněk, kde se vytvoří specializovaný rostlinný orgán: nodula - bakterie mají 20 různých nodulačních (nod) genů ty se podílejí na syntéze nebo sekreci nodulačních faktorů (= silné mitogeny, přetvářející rostlinné buňky) - uvnitř rostlinných buněk vytvářejí bakterie bakteroidy, což jsou specializované buňky určené k fixaci dusíku. Bakteroidy dodávají rostlině fixovaný dusík a od rostliny odebírají fotosyntézou vytvářený uhlík.
61 Lokalizace symbiotických genů u rhizobií - Symbiotická fixace dusíku vyžaduje asi 60 genů - Geny pro symbiozu jsou na 150 kb až 1683 kb plazmidech (megaplazmidy), které představují 25-50% velikosti genomu - většina genů pro nodulaci a fixaci dusíku je na jediném symbiotickém plazmidu psyn, v některých případech na konjugativním transpozonu (502 kb) symbiotický ostrov - ztráta plazmidu vede k neschopnosti fixovat dusík Bakterie příbuzné rhizobiím byly zjištěny i u mravenců ve specializovaném orgánu pro recyklování dusíku. - Evoluce: přenos plazmidů nebo ostrovů vede k novým druhům
62 Degradativní plazmidy Nesou geny propůjčující bakteriím schopnost biologicky degradovat organické sloučeniny, které se běžně v přírodě nevyskytují. Ve většině případů kódují část degradační dráhy včetně regulačních elementů. Pseudomonas: salicylát, naftalen, kafr, octan, toluen, fenoly, xylen, dichlorfenoxyaceton, chlorbenzen TOL plazmidy (prototyp degradativních plazmidů) Plazmidy nesou geny pro degradaci toluenu, xylenu, benzylalkoholů, benzylaldehydů Příbuzné plazmidy: schopnost růst na salicylátu nebo naftalenu jako jediném zdroji C a E. 2,4-D-plazmid = model pro studium degradace chlorovaných aromatických látek (např. (2,4-D = dichlorfenoxyoctové kyseliny používané jako herbicid)
63 Virulenční plazmidy nesou geny zodpovědné za patogenitu a virulenci bakteriálních kmenů Obecné rysy virulenčních plazmidů enterobakterií - velikost kb, nízkokopiové (1-2 kopie) - podobné buď F plazmidu nebo R100 - v jedné buňce může být i více různých virulenčních plazmidů - např. Yersinia má tři různé plazmidy, z nichž každý přispívá výrazně k virulenci
64 Geny virulence na plazmidech Faktory virulence pro kolonizaci buněk a tkání - toxiny pro adhezi na epitel a invazi, - tvorba biofilmu - průnik bakterií buněčnou membránou hostitele, - systemické šíření do dalších tkání - intracelulární přežívání v mikrofágách. Mikrofág, též neutrofilní granulocyt, druh bílé krvinky, leukocyt fagocytující buňka schopná pohlcovat drobné cizorodé částice, např. bakterie. Virulenční geny obecně zvyšují přežívání v hostiteli, adherenci a invazivitu do buněk a někdy interferují s imunitními funkcemi hostitele
65 Virulenční plazmidy gramnegativních bakterií Výskyt: Čeleď Enterobacteriace A. normální mikroflóra gastronintestinálního traktu člověka Enterobacter, Klebsiella, Escherichia B. Patogeny: Salmonella, Shigella, Yersinia Patogenní kmeny Escherichia coli: Enterotoxigenní (ETEC) Enteroinvazivní (EIEC) Enterohemorhagické (EHEC) Enteropatogenní (EPEC) Enteroagregativní (EaggEC) Spektrum chorob odráží obsah genů virulence lokalizovaných na plazmidech, bakteriofágách, ostrovech patogenity (PAI), které nejsou u komensálů.
66 Virulenční plazmidy nesporulujících G+ patogenů G+ bakterie: nejzávažnější nosokomiální patogeny jsou stafylokoky a enterokoky Řada z nich je multirezistentní k antibiotikům. Virulenční plazmidy Staphylococcus aureus Extracelulární proteiny: toxiny vázané k určitým typům chorob Enterotoxiny - ETB: potravinové otravy TSST syndrom toxického šoku exfoliatin syndrom opařené kůže Virulenční plazmidy Enterococcus faecalis identifikováno 18 různých plazmidů (prototyp: pad1, 60 kb plasmid) Faktory virulence: adhesin, matrix binding proteiny, kapsulární polysacharidy, cytolyziny (lyze erytrocytů), želatinázy a proteázy schopné poškodit tkáně a buňky.
67
Klasifikace plazmid. Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: kb
 Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: 1-1000 kb Základní typyplazmid : kryptické - funkce neznámá epizomální - reverzibilní intergace do chromozomu hostitele konjugativní - schopné
Charakteristikaplazmid dsdna kružnicová nebo lineární, velikost: 1-1000 kb Základní typyplazmid : kryptické - funkce neznámá epizomální - reverzibilní intergace do chromozomu hostitele konjugativní - schopné
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy)
") BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy) Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza
BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy) Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Rezistence patogenů vůči antimikrobialním látkám. Martin Hruška Jan Dlouhý
 Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
ZÁKLADY BAKTERIÁLNÍ GENETIKY
 Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Konjugace. Přenos DNA zprostředkovaný konjugativními plazmidy. Donor recipient transkonjugant
 Konjugace Přenos DNA zprostředkovaný konjugativními plazmidy Donor recipient transkonjugant (Exkonjuganti - v rámci téhož druhu, transkonjuganti - v rámci různých druhů) Přenášené typy elementů (DNA):
Konjugace Přenos DNA zprostředkovaný konjugativními plazmidy Donor recipient transkonjugant (Exkonjuganti - v rámci téhož druhu, transkonjuganti - v rámci různých druhů) Přenášené typy elementů (DNA):
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE)
") MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Evoluce bakteriálních genomů
 Evoluce bakteriálních genomů Charakteristické rysy: Rychlé a rozsáhlé změny ve struktuře a informačním obsahu genomu - Vnitřní přestavby - Získávání a ztráty genů a genetických elementů Vývoj kmenů v rámci
Evoluce bakteriálních genomů Charakteristické rysy: Rychlé a rozsáhlé změny ve struktuře a informačním obsahu genomu - Vnitřní přestavby - Získávání a ztráty genů a genetických elementů Vývoj kmenů v rámci
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
IZOLACE, SEPARACE A DETEKCE PROTEINŮ I. Vlasta Němcová, Michael Jelínek, Jan Šrámek
 IZOLACE, SEPARACE A DETEKCE PROTEINŮ I Vlasta Němcová, Michael Jelínek, Jan Šrámek Studium aktinu, mikrofilamentární složky cytoskeletu pomocí dvou metod: detekce přímo v buňkách - fluorescenční barvení
IZOLACE, SEPARACE A DETEKCE PROTEINŮ I Vlasta Němcová, Michael Jelínek, Jan Šrámek Studium aktinu, mikrofilamentární složky cytoskeletu pomocí dvou metod: detekce přímo v buňkách - fluorescenční barvení
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Izolace nukleových kyselin
 Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Seminář izolačních technologií
 Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Seminář izolačních technologií Zpracoval: Karel Bílek a Kateřina Svobodová Podpořeno FRVŠ 2385/2007 a 1305/2009 Úpravy a aktualizace: Pavla Chalupová ÚMFGZ MZLU v Brně 1 Lokalizace jaderné DNA 2 http://www.paternityexperts.com/basicgenetics.html
Projekt realizovaný na SPŠ Nové Město nad Metují
 Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Projekt realizovaný na SPŠ Nové Město nad Metují s finanční podporou v Operačním programu Vzdělávání pro konkurenceschopnost Královéhradeckého kraje Modul 02 Přírodovědné předměty Hana Gajdušková 1 Viry
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
STAFYLOKOKOVÉ ENTEROTOXINY. Zdravotní nezávadnost potravin. Veronika Talianová, FPBT, kruh: 346 Angelina Anufrieva, FPBT, kruh: 336
 STAFYLOKOKOVÉ ENTEROTOXINY Zdravotní nezávadnost potravin Veronika Talianová, FPBT, kruh: 346 Angelina Anufrieva, FPBT, kruh: 336 OBSAH: Základní charakteristika Staphylococcus aureus Stafylokokové enterotoxiny
STAFYLOKOKOVÉ ENTEROTOXINY Zdravotní nezávadnost potravin Veronika Talianová, FPBT, kruh: 346 Angelina Anufrieva, FPBT, kruh: 336 OBSAH: Základní charakteristika Staphylococcus aureus Stafylokokové enterotoxiny
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA
 RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
VÝZNAM HORIZONTÁLNÍHO PŘENOSU GENETICKÉ INFORMACE PRO VZNIK ANTIBIOTICKÉ REZISTENCE. V. Bencko 1, P. Šíma 2
 V. Bencko 1, P. Šíma 2 1 Ústav hygieny a epidemiologie 1. LF UK a VFN, Praha 2 Laboratoř imunoterapie, Mikrobiologický ústav, v. v. i. AV ČR, Praha VÝZNAM HORIZONTÁLNÍHO PŘENOSU GENETICKÉ INFORMACE PRO
V. Bencko 1, P. Šíma 2 1 Ústav hygieny a epidemiologie 1. LF UK a VFN, Praha 2 Laboratoř imunoterapie, Mikrobiologický ústav, v. v. i. AV ČR, Praha VÝZNAM HORIZONTÁLNÍHO PŘENOSU GENETICKÉ INFORMACE PRO
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva
 Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
b) Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?
 Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?") 1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
 MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Multirezistentních gramnegativní tyčky: základy epidemiologie antibiotické rezistence
 B I O M E D I C AL Multirezistentních gramnegativní tyčky: základy epidemiologie antibiotické rezistence Jaroslav Hrabák CHARLES UNIVERSITY IN PRAGUE Obsah prezentace Jaké multirezistentní gramnegativní
B I O M E D I C AL Multirezistentních gramnegativní tyčky: základy epidemiologie antibiotické rezistence Jaroslav Hrabák CHARLES UNIVERSITY IN PRAGUE Obsah prezentace Jaké multirezistentní gramnegativní
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Mnohobuněčné kvasinky
 Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
Laboratoř buněčné biologie PROJEKT Mnohobuněčné kvasinky Libuše Váchová ve spolupráci s laboratoří Prof. Palkové (PřFUK) Do laboratoře přijímáme studenty se zájmem o vědeckou práci Kontakt: vachova@biomed.cas.cz
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Asociace půdních MO s cévnatými rostlinami
 Asociace půdních MO s cévnatými rostlinami Fylosféra Sphingomonas, Xantomonas Rizosféra+rizoplan -růst stimulují Pseudomonas, Achromobacter (auxiny, CK, gibereliny, glykolipidy) -dusík fixují volně žijící
Asociace půdních MO s cévnatými rostlinami Fylosféra Sphingomonas, Xantomonas Rizosféra+rizoplan -růst stimulují Pseudomonas, Achromobacter (auxiny, CK, gibereliny, glykolipidy) -dusík fixují volně žijící
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Nukleové kyseliny. Nukleové kyseliny. Genetická informace. Gen a genom. Složení nukleových kyselin. Centrální dogma molekulární biologie
 Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Centrální dogma molekulární biologie ukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Transkripce D R Translace rotein Mendel) Replikace 1869 objev nukleových kyselin (Miescher) 1944 nukleové kyseliny
Enterotoxiny Staphylococcus aureus. Jana Kotschwarová Andrea Koťová
 Enterotoxiny Staphylococcus aureus Jana Kotschwarová Andrea Koťová Obsah Charakteristika Staphylococcus aureus Vlastnosti Faktory virulence Enterotoxiny Patogeneze Výskyt Metody stanovení Prevence výskytu
Enterotoxiny Staphylococcus aureus Jana Kotschwarová Andrea Koťová Obsah Charakteristika Staphylococcus aureus Vlastnosti Faktory virulence Enterotoxiny Patogeneze Výskyt Metody stanovení Prevence výskytu
Zdrojem je mrna. mrna. zpětná transkriptáza. jednořetězcová DNA. DNA polymeráza. cdna
 Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
prokaryotní Znaky prokaryoty
 prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
prokaryotní buňka Znaky prokaryoty Základní stavební jednotka bakterií a sinic Mikroskopická velikost viditelné pouze v optickém mikroskopu Buňka neobsahuje organely Obsahuje pouze 1 biomembránu cytoplazmatickou
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Radiobiologický účinek záření. Helena Uhrová
 Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Biosyntéza a metabolismus bílkovin
 Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
Bílkoviny Biosyntéza a metabolismus bílkovin lavní stavební materiál buněk a tkání Prakticky jediný zdroj dusíku pro heterotrofní organismy eexistují zásobní bílkoviny nutný dostatečný přísun v potravě
Tématické okruhy pro státní závěrečné zkoušky
 Tématické okruhy pro státní závěrečné zkoušky Obor Povinný okruh Volitelný okruh (jeden ze dvou) Forenzní biologická Biochemie, pathobiochemie a Toxikologie a bioterorismus analýza genové inženýrství Kriminalistické
Tématické okruhy pro státní závěrečné zkoušky Obor Povinný okruh Volitelný okruh (jeden ze dvou) Forenzní biologická Biochemie, pathobiochemie a Toxikologie a bioterorismus analýza genové inženýrství Kriminalistické
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Infekce, patogenita a nástroje virulence bakterií. Karel Holada
 Infekce, patogenita a nástroje virulence bakterií Karel Holada khola@lf1.cuni.cz Klíčová slova Komenzalismus Mutualismus Parazitismus Normální flóra Patogenita Saprofyt Obligátní patogen Oportunní patogen
Infekce, patogenita a nástroje virulence bakterií Karel Holada khola@lf1.cuni.cz Klíčová slova Komenzalismus Mutualismus Parazitismus Normální flóra Patogenita Saprofyt Obligátní patogen Oportunní patogen
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
a) Primární struktura NK NUKLEOTIDY Monomerem NK jsou nukleotidy
 Primární struktura NK NUKLEOTIDY Monomerem NK jsou nukleotidy") 1 Nukleové kyseliny Nukleové kyseliny (NK) sice tvoří malé procento hmotnosti buňky ale významem v kódování genetické informace a její expresí zcela nezbytným typem biopolymeru všech živých soustav a)
1 Nukleové kyseliny Nukleové kyseliny (NK) sice tvoří malé procento hmotnosti buňky ale významem v kódování genetické informace a její expresí zcela nezbytným typem biopolymeru všech živých soustav a)
Nukleové kyseliny Replikace Transkripce, RNA processing Translace
 ukleové kyseliny Replikace Transkripce, RA processing Translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti
ukleové kyseliny Replikace Transkripce, RA processing Translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
Základy genetiky prokaryotické buňky
 Základy genetiky prokaryotické buňky Chromozomová (jaderná) DNA U prokaryot (bakterie, archea) dvouřetězcová většinou kružnicová U eukaryot dvouřetězcová lineární U DNA-virů dvouřetězcová lineární, jednořetězcová
Základy genetiky prokaryotické buňky Chromozomová (jaderná) DNA U prokaryot (bakterie, archea) dvouřetězcová většinou kružnicová U eukaryot dvouřetězcová lineární U DNA-virů dvouřetězcová lineární, jednořetězcová
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Polyfázová identifikace kmenů Aeromonas encheleia
 Polyfázová identifikace kmenů Aeromonas encheleia D. Nováková, A. Vávrová, P. Švec a I. Sedláček Česká sbírka mikroorganismů Charakterizace aeromonád G-, pohyblivé tyčky, kokotyčky, čeleď Aeromonadaceae
Polyfázová identifikace kmenů Aeromonas encheleia D. Nováková, A. Vávrová, P. Švec a I. Sedláček Česká sbírka mikroorganismů Charakterizace aeromonád G-, pohyblivé tyčky, kokotyčky, čeleď Aeromonadaceae
SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA
 SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA A. Transdukce E. coli, S. typhimurium, Bacillus, Klebsiella, Staphylococcus, Streptococcus Nespecifická (P22, P1, SPβ, φ11) abortivní Specifická (fág lambda) Jsou
SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA A. Transdukce E. coli, S. typhimurium, Bacillus, Klebsiella, Staphylococcus, Streptococcus Nespecifická (P22, P1, SPβ, φ11) abortivní Specifická (fág lambda) Jsou
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Exprese rekombinantních proteinů
 Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Investujeme do naší společné budoucnosti
 Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
Investujeme do naší společné budoucnosti TECHNICKÝ LIST TOLERANCE K ALS INHIBITORŮM U CUKROVÉ ŘEPY STRUČNĚ V roce 2014 firma SESVANDERHAVE veřejně oznámila nalezení rostlin cukrové řepy tolerantních k
ANTIBIOTIKA. Mgr. Marie Vilánková. Joalis s.r.o. Všechna práva vyhrazena
 ANTIBIOTIKA Mgr. Marie Vilánková 1 Antibiotika - látky působící na mikroorganismy Antibiotika = původně získávána biologickou cestou (produkována plísněmi nebo bakteriemi) Chemoterapeutika = chemický původ,
ANTIBIOTIKA Mgr. Marie Vilánková 1 Antibiotika - látky působící na mikroorganismy Antibiotika = původně získávána biologickou cestou (produkována plísněmi nebo bakteriemi) Chemoterapeutika = chemický původ,
Determinanty lokalizace nukleosomů
 METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti
METODY STUDIA CHROMATINU Topologie DNA a nukleosomů Struktura nukleosomu 1.65-1.8 otáčky Struktura nukleosomu 10.5 nt 1.8 otáčky 10n, 10n + 5 146 nt Determinanty lokalizace nukleosomů mechanické vlastnosti