IV117: Úvod do systémové biologie
|
|
|
- Kamil Růžička
- před 6 lety
- Počet zobrazení:
Transkript
1 IV117: Úvod do systémové biologie David Šafránek
2 Obsah
3 Obsah
4 Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru demetylace uvazovana bez omezeni represor v aktivnim i pasivnim stavu aktivita = konc. aktivniho represoru demetylace jen pro aktivni receptory fine tuned model robustni model
5 Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru demetylace uvazovana bez omezeni represor v aktivnim i pasivnim stavu aktivita = konc. aktivniho represoru demetylace jen pro aktivni receptory fine tuned model robustni model


6 Robustnost citlivost na parametry X X K1 K3 K2 K1 K3 K2 Y1 Y2 Y3 Y1 Y2 Y3
7 Robustnost diferenciace buněk morfogeneze v eukaryotických organismech buňky při vývoji rozčleněny do tkání prostorové formování tkání řízeno signální dráhou citlivou na specifické proteiny morfogeny nejprve všechny buňky stejné (získány dělením vajíčka) postupně diferenciovány a tvarovány prostorovými gradienty morfogenů
8 Diferenciace buněk morfogeneze
9 Diferenciace buněk morfogeneze
10 Diferenciace buněk morfogeneze
11 Diferenciace buněk morfogeneze
12 Diferenciace buněk morfogeneze

13 Diferenciace buněk morfogeneze
14 Robustnost diferenciace buněk morfogeneze morfogeneze je řízena specifickými signálními dráhami receptory citlivé na morfogeny cílovým bodem jsou promotory genů figurujících v developmentálních transkripčních sítích nastavení hranic regionů je robustní vůči fluktuacím a změnám v genetické regulaci způsobeno markantním zvyšováním rychlosti degradace morfogenu s jeho rostoucí koncentrací morfogen degradace receptor
 vyvíjí se z vajíčka (cca 7 dní) experimentálně byla zjištěna")

15 Morfogeneze octomilky obecné Drosophila melanogaster (také vinná muška ) vyvíjí se z vajíčka (cca 7 dní) experimentálně byla zjištěna robustní morfogeneze A. Eldar et.al. Robustness of the BMP morphogen gradient in Drosophila embryonic patterning. Nature. 419, (2002).
16 Obsah
17 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů
18 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů - problém rozlišitelnosti
19 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů - problém rozlišitelnosti vazba proteinu na patřičnou část DNA elongace aminokyselin volba správného antikodonu (trna) pro příslušný kodon v mrna další vazby terciálních struktur (např. interakce proteinů, interakce ligandů s receptorem,...)...
20 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů - problém rozlišitelnosti vazba proteinu na patřičnou část DNA elongace aminokyselin volba správného antikodonu (trna) pro příslušný kodon v mrna další vazby terciálních struktur (např. interakce proteinů, interakce ligandů s receptorem,...)... výběr špatné komponenty (chyba) může mít fatální následky
21 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů - problém rozlišitelnosti vazba proteinu na patřičnou část DNA elongace aminokyselin volba správného antikodonu (trna) pro příslušný kodon v mrna další vazby terciálních struktur (např. interakce proteinů, interakce ligandů s receptorem,...)... výběr špatné komponenty (chyba) může mít fatální následky jak minimalizovat možnost chyby?
22 Stupně volnosti (nepřesnosti) v organismu některé interakce uvnitř organismu závisí na selekci správných komponent při působení rušivých faktorů - problém rozlišitelnosti vazba proteinu na patřičnou část DNA elongace aminokyselin volba správného antikodonu (trna) pro příslušný kodon v mrna další vazby terciálních struktur (např. interakce proteinů, interakce ligandů s receptorem,...)... výběr špatné komponenty (chyba) může mít fatální následky jak minimalizovat možnost chyby? mechanismus kinetické korektury kinetic proofreading J.J. Hopfield, Kinetic proofreading: a new mechanism for reducing errors in biosynthetic processes requiring high specificity. Proc. Natl. Acad. Sci. U.S.A. 71: , (1974).
23 Translace
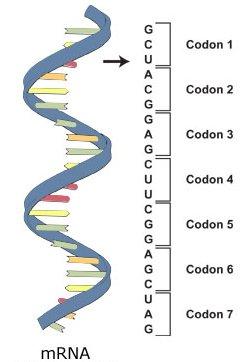
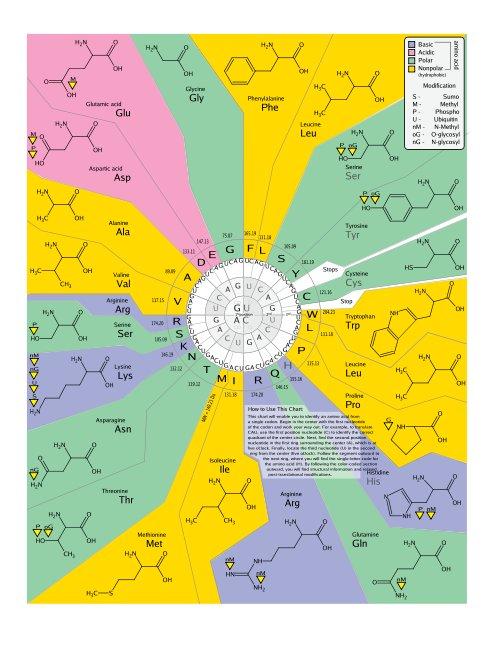
24 Mapování antikodonů dle genetického kódu

25 Mapování antikodonů v ribozómu
26 Mapování antikodonů v ribozómu?
27 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu
28 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu porušení genetického kódu
29 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu porušení genetického kódu chyby v translaci nastávají řádově s frekvencí 10 4
30 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu porušení genetického kódu chyby v translaci nastávají řádově s frekvencí 10 4 proteiny mají řádově 100 aminokyselin
31 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu porušení genetického kódu chyby v translaci nastávají řádově s frekvencí 10 4 proteiny mají řádově 100 aminokyselin tedy pravděpodobnost vytvoření proteinu s jednou chybnou aminokyselinou je = 0.01 = 1%
32 Mapování antikodonů vlivem působení rušivých faktorů (např. fluktuace teploty) může dojít k navázání nesprávného antikodonu porušení genetického kódu chyby v translaci nastávají řádově s frekvencí 10 4 proteiny mají řádově 100 aminokyselin tedy pravděpodobnost vytvoření proteinu s jednou chybnou aminokyselinou je = 0.01 = 1% větší frekvence chyb by vedla k dysfunkčnímu proteinu!
33 Modelování procesu translace nejprve vytvoříme jednoduchý model předpoklad rovnovážného stavu vazby trna kodon chybovost určena přímo pravděpodobností výběru nesprávného antikodonu
34 Modelování procesu translace nejprve vytvoříme jednoduchý model předpoklad rovnovážného stavu vazby trna kodon chybovost určena přímo pravděpodobností výběru nesprávného antikodonu provedeme rozšíření modelu v němž bude možnost chyby redukována mechanismem proofreadingu
35 Model translace c c... správný antikodon (trna)
36 Model translace c + C c... správný antikodon (trna) C... kodon (mrna)
37 Model translace c + C cc c... správný antikodon (trna) C... kodon (mrna) cc... vazba kodon antikodon
38 Model translace c + C cc v A c... správný antikodon (trna) C... kodon (mrna) cc... vazba kodon antikodon A... správná aminokyselina
39 Model translace c + C cc v A... rychlost formace vazby kodon antikodon k c... rychlost rozpadu vazby kodon antikodon v... rychlost vytvoření vazby A s předchozí aminokyselinou
40 Model translace c + C cc v A formování proteinu (vazeb mezi aminokyselinami) je řádově pomalejší než interakce na kodonu v << v << k c
41 Model translace c + C cc v A formování proteinu (vazeb mezi aminokyselinami) je řádově pomalejší než interakce na kodonu v << v << k c v nemá z krátkodobého hlediska vliv na rovnovážný stav reakcí a k c
42 Model translace c + C cc
43 Model translace c + C cc [cc] dt = [c][c] k c[cc]
44 Model translace c + C cc [cc] dt [cc] dt = 0 = [c][c] k c[cc]
45 Model translace c + C cc [cc] dt [cc] dt = [c][c] k c[cc] = 0 [c][c] = k c[cc]
46 Model translace c + C cc [cc] dt [cc] dt = [c][c] k c[cc] = 0 [c][c] = k c[cc] [cc] = [c][c]
47 Model translace c + C cc [cc] dt [cc] dt zavedeme K c = k c = [c][c] k c[cc] = 0 [c][c] = k c[cc] [cc] = [c][c]
48 Model translace c + C cc [cc] dt [cc] dt zavedeme K c = k c = [c][c] k c[cc] = 0 [c][c] = k c[cc] [cc] = [c][c] a dostáváme: [cc] = [c][c] K c
49 Model translace c + C cc v A pomocí kvazi-stabilní aproximace lze nyní vyjádřit rychlost r A vazebného přiřazení aminokyseliny A k formovanému proteinu: r A = v[cc]
50 Model translace c + C cc v A pomocí kvazi-stabilní aproximace lze nyní vyjádřit rychlost r A vazebného přiřazení aminokyseliny A k formovanému proteinu: r A = v[cc] = v[c][c] K c
51 Model translace správný vs. nesprávný antikodon Kc? Kd
52 Model translace nesprávný antikodon d + C k d k d dc v B C... kodon (mrna) d... nesprávný antikodon (trna) dc... vazba kodon antikodon B... nesprávná aminokyselina
53 Model translace nesprávný antikodon d + C k d k d dc v B koncentrace různých trna je za normálních okolností přibližně shodná ([d] [c]) rychlost začlenění různých aminokyselin do proteinu je také přibližně shodná (stále uvažujeme v)
54 Model translace nesprávný antikodon d + C k d k d dc v B stejnou procedurou jako pro c dostáváme rychlost r B přiřazení aminokyseliny B k proteinu: r B = v[dc]
55 Model translace nesprávný antikodon d + C k d k d dc v B stejnou procedurou jako pro c dostáváme rychlost r B přiřazení aminokyseliny B k proteinu: kde K d = k d k d r B = v[dc] = v[d][c] K d
56 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A
57 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c
58 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d
59 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d,
60 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d, a tedy: E st = K c K d
61 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d, a tedy: E st = K c K d = k c k d k d
62 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d, a tedy: E st = K c K d = k c k d k d k c k d
63 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d, a tedy: E st = K c K d = k c k d k d k d rozhodující úlohu hraje tedy rychlost rozpadu dočasné vazby kodon antikodon
64 Model translace analýza chybovosti srovnáme-li v rovnovážném stavu obě situace, dostáváme míru chybovosti E st : E st = r B r A = v[d][c] K d v[c][c] K c K c K d jelikož maximální možná rychlost syntézy je limitována difůzí, jsou rychlosti obou syntéz srovnatelné, k d, a tedy: E st = K c K d = k c k d k d k d rozhodující úlohu hraje tedy rychlost rozpadu dočasné vazby kodon antikodon chybný antikodon vázán nestabilně: k d >> k c
65 Model translace analýza chybovosti experimentálně byla zjištěna míra chybovosti translace u živých organismů E = 10 4
66 Model translace analýza chybovosti experimentálně byla zjištěna míra chybovosti translace u živých organismů E = 10 4 rychlost rozpadu vazby kodon-antikodon byla zjištěna 100 krát rychlejší pro chybný antikodon
67 Model translace analýza chybovosti experimentálně byla zjištěna míra chybovosti translace u živých organismů E = 10 4 rychlost rozpadu vazby kodon-antikodon byla zjištěna 100 krát rychlejší pro chybný antikodon dle uvedeného modelu bychom dostali E st = 10 2
68 Model translace analýza chybovosti experimentálně byla zjištěna míra chybovosti translace u živých organismů E = 10 4 rychlost rozpadu vazby kodon-antikodon byla zjištěna 100 krát rychlejší pro chybný antikodon dle uvedeného modelu bychom dostali E st = 10 2 uvedený model je v rozporu se skutečností
69 Model translace analýza chybovosti experimentálně byla zjištěna míra chybovosti translace u živých organismů E = 10 4 rychlost rozpadu vazby kodon-antikodon byla zjištěna 100 krát rychlejší pro chybný antikodon dle uvedeného modelu bychom dostali E st = 10 2 uvedený model je v rozporu se skutečností jaký faktor tedy snižuje chybovost translace?
70 Model translace II c c... antikodon
71 Model translace II c + C c... antikodon C... kodon (mrna)
72 Model translace II c + C cc c... antikodon C... kodon (mrna) cc... vazba kodon antikodon
73 Model translace II c + C cc m c C c, c... antikodon, značí chemickou modifikaci C... kodon (mrna) cc, c C... vazba kodon antikodon
74 Model translace II c + C cc m c C v A c, c... antikodon, značí chemickou modifikaci C... kodon (mrna) cc, c C... vazba kodon antikodon A... aminokyselina
75 Model translace II c + C cc m c C c C v A l c c + C c, c... antikodon, značí chemickou modifikaci C... kodon (mrna) cc, c C... vazba kodon antikodon A... aminokyselina
76 Model translace II c + C cc m c C c C v A l c c + C... rychlost formace vazby kodon antikodon k c... rychlost rozpadu vazby kodon antikodon v... rychlost přiřazení A do formovaného proteinu
77 Model translace II c + C cc m c C c C v A l c c + C... rychlost formace vazby kodon antikodon k c... rychlost rozpadu vazby kodon antikodon v... rychlost přiřazení A do formovaného proteinu m... rychlost chemické modifikace navázané trna
78 Model translace II c + C cc m c C c C v A l c c + C... rychlost formace vazby kodon antikodon k c... rychlost rozpadu vazby kodon antikodon v... rychlost přiřazení A do formovaného proteinu m... rychlost chemické modifikace navázané trna l c... rychlost vyklouznutí trna z kodonu
79 Model translace II c + C cc m c C c C v A l c c + C formování proteinu (vazeb mezi aminokyselinami) je v řádově pomalejší časové škále než interakce na kodonu v <<, k c, l c, m
80 Model translace II c + C cc m c C c C v A l c c + C formování proteinu (vazeb mezi aminokyselinami) je v řádově pomalejší časové škále než interakce na kodonu v <<, k c, l c, m v nemá z krátkodobého hlediska vliv na rovnovážný stav ostatních reakcí (, k c, m a l c)
81 Model translace II c + C cc m c C l c c + C
82 Model translace II c + C cc [c C] dt m c C = m[cc] l c[c C] l c c + C
83 Model translace II c + C cc [c C] dt [c C] dt = 0 m c C = m[cc] l c[c C] l c c + C
84 Model translace II c + C cc [c C] dt [c C] dt m c C = m[cc] l c[c C] = 0 m[cc] = l c[c C] l c c + C
85 Model translace II c + C cc [c C] dt [c C] dt [c C] = m l c [cc] m c C = m[cc] l c[c C] = 0 m[cc] = l c[c C] l c c + C
86 Model translace II c + C cc [c C] dt [c C] dt [c C] = m l c [cc] = m l c m c C = m[cc] l c[c C] = 0 m[cc] = l c[c C] l c c + C [c][c] K c, K c = k c + m
87 Model translace II c + C cc [c C] dt [c C] dt [c C] = m l c [cc] = m l c m c C = m[cc] l c[c C] = 0 m[cc] = l c[c C] l c c + C [c][c] K c, K c = k c + m pro rychlost začlenění A do proteinu tedy dostáváme: r A = v[c C]
88 Model translace II c + C cc m c C l c c + C [c C] dt = m[cc] l c[c C] [c C] dt = 0 m[cc] = l c[c C] [c C] = m l c [cc] = m l c [c][c] K c, K c = k c + m pro rychlost začlenění A do proteinu tedy dostáváme: r A = v[c C] = v m l c [c][c] K c
89 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C
90 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C v v obou případech přibližně shodné k d m přibližně shodné pro všechny trna, m << k c, k d modifikace trna nemá vliv na rozpad vazby s kodonem
91 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C v v obou případech přibližně shodné k d m přibližně shodné pro všechny trna, m << k c, k d modifikace trna nemá vliv na rozpad vazby s kodonem K d K c = k d +m k d k c +m
92 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C v v obou případech přibližně shodné k d m přibližně shodné pro všechny trna, m << k c, k d modifikace trna nemá vliv na rozpad vazby s kodonem K d K c = k d +m k d k c +m k d + m + m
93 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C v v obou případech přibližně shodné k d m přibližně shodné pro všechny trna, m << k c, k d modifikace trna nemá vliv na rozpad vazby s kodonem K d K c = k d +m k d k c +m k d + m + m k d k c
94 Model translace II (špatný vs. správný antikodon) c + C cc d + C k d k d dc m c C c C m d C d C v A l c c + C v B l d d + C v v obou případech přibližně shodné k d m přibližně shodné pro všechny trna, m << k c, k d modifikace trna nemá vliv na rozpad vazby s kodonem K d K c = k d +m k d k c +m k d + m + m k d k c = l d l c
95 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d
96 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d pro míru chybovosti ve stabilním stavu nyní dostáváme: E st = r B r A
97 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d pro míru chybovosti ve stabilním stavu nyní dostáváme: E st = r B = l c K c r A l d K d
98 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d pro míru chybovosti ve stabilním stavu nyní dostáváme: E st = r B = l c K c r A l d K d jelikož K d K c l d l c
99 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d pro míru chybovosti ve stabilním stavu nyní dostáváme: E st = r B = l c K c r A l d K d jelikož K d K c l d l c lze psát: E st ( K c K d ) 2
100 Model translace II (nesprávný vs. správný antikodon) r A = v[c C] = v m l c r B = v[d C] = v m l d [c][c] K c [d][c] K d pro míru chybovosti ve stabilním stavu nyní dostáváme: E st = r B = l c K c r A l d K d jelikož K d K c l d l c lze psát: E st ( K c K d ) 2 nyní dle experimentálně zjištěných dat E st (10 2 ) 2 = 10 4
101 Analýza chybovosti translace shrnutí translace je příklad mechanismu při němž dochází k problému rozlišitelnosti vlivem rušivých elementů může dojít hybě chyba při rozlišení dvou komponent může vést k dysfunkci organismus omezuje možnost chyby pomocí mechanismu proofreadingu proofreading spočívá v chemické modifikaci interagujících látek
102 Škálovatelnost mechanismu proofreadingu účinek proofreadingu lze umocnit zvýšením počtu postupných chemických modifikací interagujících látek c + C kc cc m 1 c 1 C m 2 c 2 C m n c n C i {1,..., n}. c i C l m i c + C v A
103 Škálovatelnost mechanismu proofreadingu účinek proofreadingu lze umocnit zvýšením počtu postupných chemických modifikací interagujících látek c + C kc cc m 1 c 1 C m 2 c 2 C m n c n C i {1,..., n}. c i C l m i c + C v A d + C k d k d dc m 1 d 1 C m 2 d 2 C m n d n C i {1,..., n}. d i C l m i d + C v B
104 Škálovatelnost mechanismu proofreadingu účinek proofreadingu lze umocnit zvýšením počtu postupných chemických modifikací interagujících látek c + C kc cc m 1 c 1 C m 2 c 2 C m n c n C i {1,..., n}. c i C l m i c + C v A d + C k d k d dc m 1 d 1 C m 2 d 2 C m n d n C i {1,..., n}. d i C l m i d + C v B E st ( K c K d ) n+1
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
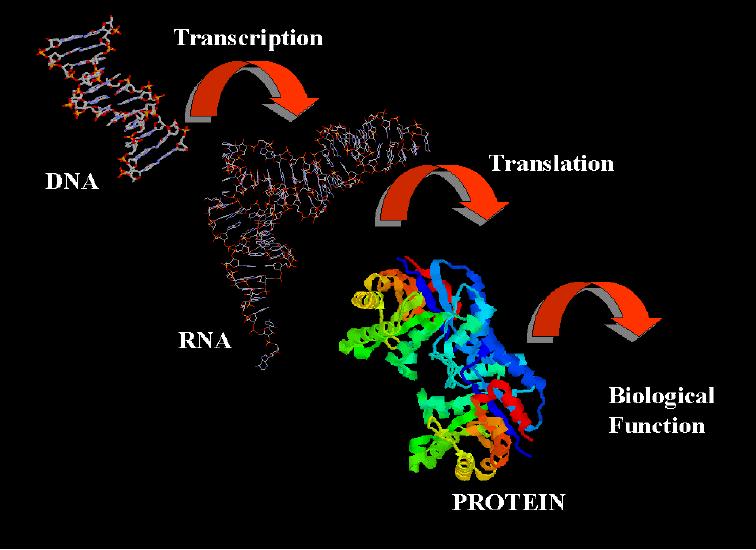
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
Nukleové kyseliny Replikace Transkripce translace Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Život závisí na schopnosti buněk skladovat,
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Translace (druhý krok genové exprese)
") Translace (druhý krok genové exprese) Od RN k proteinu Milada Roštejnská Helena Klímová 1 enetický kód trn minoacyl-trn-synthetasa Translace probíhá na ribosomech Iniciace translace Elongace translace
Translace (druhý krok genové exprese) Od RN k proteinu Milada Roštejnská Helena Klímová 1 enetický kód trn minoacyl-trn-synthetasa Translace probíhá na ribosomech Iniciace translace Elongace translace
TRANSLACE - SYNTÉZA BÍLKOVIN
 TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
Centrální dogma molekulární biologie
 řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
řípravný kurz LF MU 2011/12 Centrální dogma molekulární biologie Nukleové kyseliny 1865 zákony dědičnosti (Johann Gregor Mendel) 1869 objev nukleových kyselin (Miescher) 1944 genetická informace v nukleových
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Nukleové kyseliny. DeoxyriboNucleic li Acid
 Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Molekulární lární genetika Nukleové kyseliny DeoxyriboNucleic li Acid RiboNucleic N li Acid cukr (deoxyrobosa, ribosa) fosforečný zbytek dusíkatá báze Dusíkaté báze Dvouvláknová DNA Uchovává genetickou
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Schéma průběhu transkripce
 Molekulární základy genetiky PROTEOSYNTÉZA A GENETICKÝ KÓD Proteosyntéza je složitý proces tvorby bílkovin, který zahrnuje proces přepisu genetické informace z DNA do kratšího zápisu v informační mrna
Molekulární základy genetiky PROTEOSYNTÉZA A GENETICKÝ KÓD Proteosyntéza je složitý proces tvorby bílkovin, který zahrnuje proces přepisu genetické informace z DNA do kratšího zápisu v informační mrna
(Vývojová biologie) Embryologie. Jiří Pacherník
 Embryologie. Jiří Pacherník") (Vývojová biologie) Embryologie Jiří Pacherník jipa@sci.muni.cz Podpořeno projektem FRVŠ 524/2011 buňka -> tkáně -> orgány -> organismus / jedinec Základní procesy na buněčné úrovni dělení buněk proliferace
(Vývojová biologie) Embryologie Jiří Pacherník jipa@sci.muni.cz Podpořeno projektem FRVŠ 524/2011 buňka -> tkáně -> orgány -> organismus / jedinec Základní procesy na buněčné úrovni dělení buněk proliferace
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 8.10.2008 Obsah Metody dynamické analýzy Obsah Metody dynamické analýzy Shrnutí biologický systém definován interakcemi mezi jeho komponentami interakce
IV117: Úvod do systémové biologie David Šafránek 8.10.2008 Obsah Metody dynamické analýzy Obsah Metody dynamické analýzy Shrnutí biologický systém definován interakcemi mezi jeho komponentami interakce
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Studijní materiály pro bioinformatickou část ViBuChu. úloha II. Jan Komárek, Gabriel Demo
 Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Modelování biochemických procesů: Deterministický model transkripční regulace
 Modelování biochemických procesů: Deterministický model transkripční regulace David Šafránek Seminář ParaDiSe 1.10.2007 Obsah Základní pojmy Spojitý deterministický model chemických reakcí Spojitý deterministický
Modelování biochemických procesů: Deterministický model transkripční regulace David Šafránek Seminář ParaDiSe 1.10.2007 Obsah Základní pojmy Spojitý deterministický model chemických reakcí Spojitý deterministický
Genetický kód. Jakmile vznikne funkční mrna, informace v ní obsažená může být ihned použita pro syntézu proteinu.
 Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Genetický kód Jakmile vznikne funkční, informace v ní obsažená může být ihned použita pro syntézu proteinu. Pravidla, kterými se řídí prostřednictvím přenos z nukleotidové sekvence DNA do aminokyselinové
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Genetika. Genetika. Nauka o dědid. dičnosti a proměnlivosti. molekulárn. rní buněk organismů populací
 Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
Genetika Nauka o dědid dičnosti a proměnlivosti Genetika molekulárn rní buněk organismů populací Dědičnost na úrovni nukleových kyselin Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci
Obecná a srovnávací odontologie. Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód
 Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
Základní morfogenetické procesy
 Základní morfogenetické procesy 502 Základní morfogenetické procesy Mechanismy, které se uplatňují v ontogenesi, tedy při vývoji jedince od zygoty k mnohobuněčnému organismu Buněčná úroveň diferenciace
Základní morfogenetické procesy 502 Základní morfogenetické procesy Mechanismy, které se uplatňují v ontogenesi, tedy při vývoji jedince od zygoty k mnohobuněčnému organismu Buněčná úroveň diferenciace
Kontrola genové exprese
 Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Majeská Čudejková 3. Proteosyntéza Centrální dogma molekulární biologie Rozluštění genetického kódu in vitro Marshall Nirenberg a Heinrich Matthaei zjistili,
Základy molekulární biologie KBC/MBIOZ Mária Majeská Čudejková 3. Proteosyntéza Centrální dogma molekulární biologie Rozluštění genetického kódu in vitro Marshall Nirenberg a Heinrich Matthaei zjistili,
Modelov an ı biologick ych syst em u Radek Pel anek
 Modelování biologických systémů Radek Pelánek Modelování v biologických vědách typický cíl: pomocí modelů se snažíme pochopit, jak biologické systémy fungují model zahrnuje naše chápání simulace ukazuje,
Modelování biologických systémů Radek Pelánek Modelování v biologických vědách typický cíl: pomocí modelů se snažíme pochopit, jak biologické systémy fungují model zahrnuje naše chápání simulace ukazuje,
Biologická léčiva. Co jsou to biosimilars a jak se vyrábějí. Michal Hojný
 Biologická léčiva Co jsou to biosimilars a jak se vyrábějí Michal Hojný Zadání Jsou to opravdu generické kopie originálů? Jsou tam nějaká nebezpečí při výrobě? Jsou ty léky úplně stejné? Jak těžké je vyrobit
Biologická léčiva Co jsou to biosimilars a jak se vyrábějí Michal Hojný Zadání Jsou to opravdu generické kopie originálů? Jsou tam nějaká nebezpečí při výrobě? Jsou ty léky úplně stejné? Jak těžké je vyrobit
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Obecná biologie a genetika B53 volitelný předmět pro 4. ročník
 Obecná biologie a genetika B53 volitelný předmět pro 4. ročník Charakteristika vyučovacího předmětu Vyučovací předmět vychází ze vzdělávací oblasti Člověk a příroda, vzdělávacího oboru Biologie. Mezipředmětové
Obecná biologie a genetika B53 volitelný předmět pro 4. ročník Charakteristika vyučovacího předmětu Vyučovací předmět vychází ze vzdělávací oblasti Člověk a příroda, vzdělávacího oboru Biologie. Mezipředmětové
Využití metod strojového učení v bioinformatice David Hoksza
 Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Nejmenší jednotka živého organismu schopná samostatné existence. Výměnu látek Růst Pohyb Rozmnožování Dědičnost
 BUŇKA Nejmenší jednotka živého organismu schopná samostatné existence Buňka je schopna uskutečňovat základní funkce organismu: obrázky použity z Nečas: BIOLOGIE LIDSKÉ TĚLO Alberts: ZÁKLADY BUNĚČNÉ BIOLOGIE
BUŇKA Nejmenší jednotka živého organismu schopná samostatné existence Buňka je schopna uskutečňovat základní funkce organismu: obrázky použity z Nečas: BIOLOGIE LIDSKÉ TĚLO Alberts: ZÁKLADY BUNĚČNÉ BIOLOGIE
Radiobiologický účinek záření. Helena Uhrová
 Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA
 TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
TRANSPORT PŘES MEMBRÁNY, MEMBRÁNOVÝ POTENCIÁL, OSMÓZA 1 VÝZNAM TRANSPORTU PŘES MEMBRÁNY V MEDICÍNĚ Příklad: Membránový transportér: CFTR (cystic fibrosis transmembrane regulator) Onemocnění: cystická fibróza
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Molekulární diagnostika pletencové svalové dystrofie typu 2A
 Molekulární diagnostika pletencové svalové dystrofie typu 2A Lenka Fajkusová Centrum molekulární biologie a genové terapie Fakultní nemocnice Brno Pletencové svalové dystrofie (Limb Girdle Muscular Dystrophy
Molekulární diagnostika pletencové svalové dystrofie typu 2A Lenka Fajkusová Centrum molekulární biologie a genové terapie Fakultní nemocnice Brno Pletencové svalové dystrofie (Limb Girdle Muscular Dystrophy
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Biologie buňky. systém schopný udržovat se a rozmnožovat
 Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Biologie buňky 1665 - Robert Hook (korek, cellulae = buňka) Cytologie - věda zabývající se studiem buňek Buňka ozákladní funkční a stavební jednotka živých organismů onejmenší známý uspořádaný dynamický
Buňky, tkáně, orgány, soustavy
 Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
Lidská buňka buněčné organely a struktury: Jádro Endoplazmatické retikulum Goldiho aparát Mitochondrie Lysozomy Centrioly Cytoskelet Cytoplazma Cytoplazmatická membrána Buněčné jádro Jadérko Karyoplazma
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 17.9.2008 Obsah Informace o předmětu Úvod Historie Základní pojmy a principy Obsah Informace o předmětu Úvod Historie Základní pojmy a principy Náplň předmětu
IV117: Úvod do systémové biologie David Šafránek 17.9.2008 Obsah Informace o předmětu Úvod Historie Základní pojmy a principy Obsah Informace o předmětu Úvod Historie Základní pojmy a principy Náplň předmětu
Základy molekulární a buněčné biologie. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Exprese rekombinantních proteinů
 Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Nukleové kyseliny Replikace Transkripce translace
 Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
Nukleové kyseliny Replikace Transkripce translace Figure 4-3 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-4 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-5 Molecular
jedné aminokyseliny v molekule jednoho z polypeptidů hemoglobinu
 Translace a genetický kód Srpkovitý tvar červených krvinek u srpkovité anémie: důsledek záměny Srpkovitý tvar červených krvinek u srpkovité anémie: důsledek záměny jedné aminokyseliny v molekule jednoho
Translace a genetický kód Srpkovitý tvar červených krvinek u srpkovité anémie: důsledek záměny Srpkovitý tvar červených krvinek u srpkovité anémie: důsledek záměny jedné aminokyseliny v molekule jednoho
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Tématické okruhy pro státní závěrečné zkoušky
 Tématické okruhy pro státní závěrečné zkoušky Program / Obor Povinný okruh Volitelný okruh (jeden ze tří) Mikrobiologie a buněčná biologie Mikrobiologie životního prostředí Obor: Mikrobiologie Bioinženýrství
Tématické okruhy pro státní závěrečné zkoušky Program / Obor Povinný okruh Volitelný okruh (jeden ze tří) Mikrobiologie a buněčná biologie Mikrobiologie životního prostředí Obor: Mikrobiologie Bioinženýrství
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Genová etiologie nemocí
 Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Nukleové kyseliny Replikace Transkripce, RNA processing Translace
 Nukleové kyseliny Replikace Transkripce, RNA processing Translace Figure 6-2 Molecular Biology of the Cell ( Garland Science 2008) replikace Figure 4-8 Molecular Biology of the Cell ( Garland Science
Nukleové kyseliny Replikace Transkripce, RNA processing Translace Figure 6-2 Molecular Biology of the Cell ( Garland Science 2008) replikace Figure 4-8 Molecular Biology of the Cell ( Garland Science
A. chromozómy jsou rozděleny na 2 chromatidy spojené jen v místě centromery. B. vlákna dělícího vřeténka jsou připojena k chromozómům
 Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Karlova univerzita, Lékařská fakulta Hradec Králové Obor: všeobecné lékařství - test z biologie Vyberte tu z nabídnutých odpovědí (1-5), která je nejúplnější. Otázka Odpověď 1. Mezi organely membránového
Digitální učební materiál
 Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Odborná biologie, část biologie Společná pro
Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Odborná biologie, část biologie Společná pro
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 24.9.2008 Obsah Modelové organismy Získávání biologických dat Modely a simulace in silico Obsah Modelové organismy Získávání biologických dat Modely a simulace
IV117: Úvod do systémové biologie David Šafránek 24.9.2008 Obsah Modelové organismy Získávání biologických dat Modely a simulace in silico Obsah Modelové organismy Získávání biologických dat Modely a simulace
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Syntéza a postranskripční úpravy RNA
 Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Syntéza a postranskripční úpravy RNA 2016 1 Transkripce Proces tvorby RNA na podkladu struktury DNA Je přepisován pouze jeden řetězec dvoušroubovice DNA templátový řetězec Druhý řetězec se nazývá kódující
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc.
 Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Garant předmětu GEN: prof. Ing. Jindřich Čítek, CSc. Garant předmětu GEN1: prof. Ing. Václav Řehout, CSc. Další vyučující: Ing. l. Večerek, PhD., Ing. L. Hanusová, Ph.D., Ing. L. Tothová Předpoklady: znalosti
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Replikace, transkripce a translace
 Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Molekulární diagnostika
 Molekulární diagnostika Odry 11. 11. 2010 Michal Pohludka, Ph.D. Buňka základní jednotka živé hmoty Všechny v současnosti známé buňky se vyvinuly ze společného předka, tedy buňky, která žila asi před 3,5-3,8
Molekulární diagnostika Odry 11. 11. 2010 Michal Pohludka, Ph.D. Buňka základní jednotka živé hmoty Všechny v současnosti známé buňky se vyvinuly ze společného předka, tedy buňky, která žila asi před 3,5-3,8
BUNĚČ ORGANISMŮ KLÍČOVÁ SLOVA:
 BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
BUNĚČ ĚČNÁ STAVBA ŽIVÝCH ORGANISMŮ KLÍČOVÁ SLOVA: Prokaryota, eukaryota, viry, bakterie, živočišná buňka, rostlinná buňka, organely buněčné jádro, cytoplazma, plazmatická membrána, buněčná stěna, ribozom,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
GENETIKA dědičností heredita proměnlivostí variabilitu Dědičnost - heredita podobnými znaky genetickou informací Proměnlivost - variabilita
 GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
7. Měření fluorescence při excitaci kontinuálním světlem ( steady-state )
") 7. Měření fluorescence při excitaci kontinuálním světlem ( steady-state ) Steady-state měření Excitujeme kontinuálním světlem, měříme intenzitu emise (počet emitovaných fotonů) Obvykle nedetekujeme všechny
7. Měření fluorescence při excitaci kontinuálním světlem ( steady-state ) Steady-state měření Excitujeme kontinuálním světlem, měříme intenzitu emise (počet emitovaných fotonů) Obvykle nedetekujeme všechny
Projdou či neprojdou III: Pohyb částic v kapalině - difúze
 Projdou či neprojdou III: Pohyb částic v kapalině - difúze Shrnutí Žáci pozorují difúzi- rozptyl částic v kapalině. Na základě Brownova pohybu se molekuly v kapalném prostředí vlivem tepelného pohybu zcela
Projdou či neprojdou III: Pohyb částic v kapalině - difúze Shrnutí Žáci pozorují difúzi- rozptyl částic v kapalině. Na základě Brownova pohybu se molekuly v kapalném prostředí vlivem tepelného pohybu zcela
ÚVOD DO STUDIA BUŇKY příručka pro učitele
 Obecné informace ÚVOD DO STUDIA BUŇKY příručka pro učitele Téma úvod do studia buňky je rozvržen na jednu vyučovací hodinu. V tomto tématu jsou probrány a zopakovány základní charakteristiky živých soustav
Obecné informace ÚVOD DO STUDIA BUŇKY příručka pro učitele Téma úvod do studia buňky je rozvržen na jednu vyučovací hodinu. V tomto tématu jsou probrány a zopakovány základní charakteristiky živých soustav
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Termodynamika a živé systémy. Helena Uhrová
 Termodynamika a živé systémy Helena Uhrová Základní pojmy termodynamiky soustava izolovaná otevřená okolí vlastnosti soustavy znaky popisující soustavu stav rovnováhy tok m či E =0 funkce stavu - soubor
Termodynamika a živé systémy Helena Uhrová Základní pojmy termodynamiky soustava izolovaná otevřená okolí vlastnosti soustavy znaky popisující soustavu stav rovnováhy tok m či E =0 funkce stavu - soubor
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
NEMEMBRÁNOVÉ ORGANELY. Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly
 Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly") NEMEMBRÁNOVÉ ORGANELY Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly RIBOSOMY Částice složené z rrna a proteinů, skládají se z velké kulovité
NEMEMBRÁNOVÉ ORGANELY Ribosomy Centrioly (jadérko) Cytoskelet: aktinová filamenta (mikrofilamenta) intermediární filamenta mikrotubuly RIBOSOMY Částice složené z rrna a proteinů, skládají se z velké kulovité
MOLEKULÁRNÍ ZÁKLADY DĚDIČNOSTI
 Maturitní téma č. 33 MOLEKULÁRNÍ ZÁKLADY DĚDIČNOSTI NUKLEOVÉ KYSELINY - jsou to makromolekuly tvořené řetězci vzájemně spojených nukleotidů. Molekula nukleotidu sestává z : - pětiuhlíkatého monosacharidu
Maturitní téma č. 33 MOLEKULÁRNÍ ZÁKLADY DĚDIČNOSTI NUKLEOVÉ KYSELINY - jsou to makromolekuly tvořené řetězci vzájemně spojených nukleotidů. Molekula nukleotidu sestává z : - pětiuhlíkatého monosacharidu
Autokláv reaktor pro promíchávané vícefázové reakce
 Vysoká škola chemicko technologická v Praze Ústav organické technologie (111) Autokláv reaktor pro promíchávané vícefázové reakce Vypracoval : Bc. Tomáš Sommer Předmět: Vícefázové reaktory (prof. Ing.
Vysoká škola chemicko technologická v Praze Ústav organické technologie (111) Autokláv reaktor pro promíchávané vícefázové reakce Vypracoval : Bc. Tomáš Sommer Předmět: Vícefázové reaktory (prof. Ing.
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Základy buněčné biologie
 Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Maturitní otázka č. 8 Základy buněčné biologie vypracovalo přírodozpytné sympózium LP, AM & DK na konferenci v Praze, 1. Máje 2014 Buňka (cellula) je nejmenší známý útvar, který je schopný všech životních
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele)
") Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Genetika - maturitní otázka z biologie (2)
") Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):
Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):