Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu
|
|
|
- Anna Ševčíková
- před 5 lety
- Počet zobrazení:
Transkript
1 Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace genové exprese je represe (a její odblokování). Většina bílkovin je polyfunkčních a jejich exprese a funkce je regulována na mnoha úrovních najednou. Signální dráhy se integrují na úrovni společného regulačního bílkovinného intermediátu (příklad fosf.), druhého posla, promotoru, procesu či struktury. Signál je zesilován, či zeslabován - při tom šum okolí může být pozitivně využit k zesílení signálu = stochastická resonance.
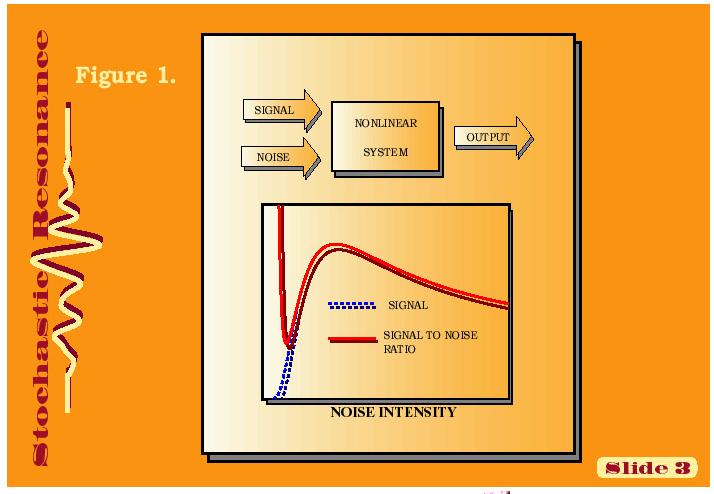
2
3 Z toho, že organismus vládne buňkám (nejen buňky organizmu), také plyne, že také buňka a organizmus vládne signálním drahám a sítím. Buňka není jen výsledkem propletence procesů/struktur, které v ní probíhají/strukturují, ale také jejich tvůrcem.
4 Degradace bílkovin je stejně důležitý regulační krok jako jejich syntéza. Signální dráhy často obsahují vysoce specifickou/regulovanou degradaci bílkoviny jako důležitý regulační krok. (Vzpomeňme na cykliny )
5 Proteolýza je ovšem také konstitutivní proces. Až 30% translatovaných bílkovin je nefunkčních.
6 Degradace buněč ěčných bílkovinb 17-1 Proteolytické dráhy u eukaryot - 1. vakuolární/lysozomální - 2. Na ubiquitinu-proteasomu závislá degradace - 3. post-proteasomálni degradace : Tricorn, TPPII? - 4. Degradace membranových proteinů
7 Hlavní proteolytické dráhy eukaryot 17-2 Mitochondria Mitochondrial /plastid proteolytic systems Lysosome/ endosome endosomelysosome system cooperation cytoplasmic proteins Ubiquitinproteasome system endosome-lysosome pathway degrades extracellular and cellsurface proteins ubiquitin-proteasome pathway degrades proteins from the cytoplasm, nucleus and ER mitochondria (and chloroplasts) have their own proteolytic system that are of bacterial origin Autophagosome nuclear proteins Nucleus ER proteins
8 Degradace membránových proteinů. ATP dep. membr. komplexy AAA proteases mediate the degradation of membrane proteins in bacteria, mitochondria and chloroplasts (i.e., compartments of eubacterial origin) combine proteolytic and chaperone activities in one system, acting as qualitycontrol machineries - model substrate polypeptides containing hydrophilic domains at either side of the membrane can be completely degraded by either of two AAA proteases found in mitochondria, if solventexposed domains are in an unfolded state - a short protein tail protruding from the membrane surface is sufficient to allow the proteolytic attack of an AAA protease that facilitates domain unfolding at the opposite side Leonhard et al. (2000) Mol. Cell 5,

9 ClpAP je proteázový komplex aktivní v plastidech (homol. E.coli). Brání hromadění nefunkčních bílkovin v plastidech. Podobně je tomu v mitochondriích.
10 Vakuolárn rní/lysozomální degradace 17-5 macroautophagy is the equivalent of forming intracellular endosomes (phagosomes) that fuse to the lysosome and result in the breakdown of its contents vakuola Cuervo and Dice (1998) J. Mol. Med. 76, 6-12.
11 Bílkoviny určené k degradaci proteasomem jsou modifikovány ubiquitinem. Prvním známým proteinem ubq. in vivo v aktivní formě byl u rostlin fytochrom. Většina bílkovin je před ubiquitinací specificky fosforylována.
12 Ubiquitinová dráha 17-6 E1 - ubiquitin activating enzyme E2 - ubiquitin conjugating enzyme E3 - ubiquitin ligase ~ denotes high-energy thioester bond DUB, deubiquinating enzyme

13 Syntet. z fusních tandemových prekursorů 3 až 6. Jsou štepeny deubiqutinačními enzymy/proteasami = DUB 76 AA rostl. od kvasinek/živočichů se liší 2/3 AA. Ubq. se kovalentně váže na Lys cílové bílkoviny C -Gly.
14 E1 ubq. Aktivace E2 ubq. Konjugace E3 ubq. Ligace ubq. na Lys cílové bílkoviny. E3 zajišťuje specifitu rozpoznání. Na bílkovině je ubq. více Lys a to opakovaně = polyubiquitin.
15 E3 ubiquitin ligázy základní typy E3 ubiquitin ligáz u rostlin: kol.1300 genů Arabidopsis kóduje podjednotky E3 ligáz. s HECT domains (E6AP-related proteins) - monomerní 17x u A.t. s Ring/U-Boxem (VHL, SCF, APC, MDM2, c-cbl, etc.) 480x RING a 64x U-Box u A.t SCF komplex 4 podjednotky - podjednotka řídící specifitu F-box 700x u A.t. APC ale také 37x AtE2 = potenciálě obrovské množství spec. ubq. komplexů.

16 vedle 700 F-boxů má Arabidopsis 21 SKP bílkovin. Opět velká kombinatorika komplexů existují ovšem preferované kombinace.
17 F-box odpovídá za specifitu interakce se substrátem.
18 SCF- dependentní ubiquitinace u kvasinek F-box proteins mediate substrate selectivity in degrading various yeast proteins many (all?) of the substrates need to be phosphorylated to be recognized by the F-box protein WD40 and leucinerich repeats (LRRs) present in F-box proteins mediate protein-protein interactions u rostlin je to podobné
19 Monoubiquitinace může sloužit jako regulační modifikace. Např. pro třídění do endocytotické dráhy a vakuoly/lysozomu či modifikuje např. transkripci TF-ubq.
 To initiate sister chromatid separation, the APC has to ubiquitinate the anaphase inhibitor")
20 Anaphase promoting complex (APC) The anaphase-promoting complex (also termed cyclosome ) is a ubiquitin-protein ligase that controls important transitions in mitosis by ubiquitinating regulatory proteins consists of many different proteins, including some related to SCF (e.g., ring protein) To initiate sister chromatid separation, the APC has to ubiquitinate the anaphase inhibitor securin, whereas exit from mitosis requires the ubiquitination of B-type cyclins Gieffers et al. (2001) Mol. Cell 7,
21 Proteasom

22
23 Topologie proteasomu zajišťuje, že proteázovou aktivitou nebudou nespecificky zasaženy cytoplasmatické bílkoviny. U Arabidopsis každá podjednotka = dva geny - to zn. různé subtypy proteasomu.
24 CSN (COP9 / signalosom) komplex byl poprvé objeven u Arabidopsis (viz. dále = světlo jako signál). Jeho podjednotky a celková organizace jsou homologní "víku" RP proteasomu, ale funguje nezávisle, jako regulátor ubiquitinace a aktivity proteazómu mu.
25
26 CSN kontroluje aktivitu E3 SCF ligázy prostřednictvím neddylace či deneddylace (NEDD8 či RUB1 jsou peptidy podobné ubiquitinu) a degradaci bílkovin interakcemi s proteasomem (alternativní víko ).
27 N-koncové pravidlo Met, Thr, Ser, Gly a Val na N stabilizují bílkovinu, zatímco Lys, Arg, His ji destabilizují.

28 N-koncová acetylace stabilizuje bílkovinu.
29 U některých prot. je N Met odštěpen aminopeptidázou a novou počáteční AA bývá Glu nebo Asp;takové bílkoviny se stávají substrátem ubiquitinace teprve po přidání N - Arg.
30 Bílkoviny se mnohonásobně liší poločasem životnosti a ten se prudce mění s měnícím se diferenciačním/regulačním stavem buňky. Klíčové proteiny signálních drah (včetně transkripčních faktorů) bývají velmi labilní.

31 Příklady proteolytických bílk. a ovlivněných procesů u rostlin.
32 Inhibitory proteasomu The peptide aldehydes, MG 132, MG 115, and PSI, inhibit the complex's chymotrypsinlike activity in a potent but reversible manner. Lactacystin is a natural, irreversible, nonpeptide, cell permeable inhibitor that is more selective than peptide aldehydes but less selective than peptide boronates, another class of proteasome inhibitors.
33 SVĚTLO jako SIGNÁL

34
35 Světlo UV B nm UV-A nm Modré s nm Červené s nm Dl. Červ. S nm


36 Borthwick s Experiment in Grand Rapids Lettuce (1952)
37 Fytochromy Kryptochromy Fototropiny Neznámý rec. UV-B Fotoreceptory

38
39 Phytochrome Exists in Two Photointraconvertible Forms -synthesized as Pr in darkness -converted by red light (max= 666 nm) to Pfr -Pfr is biologically active form -Far-red light (730 nm) converts to Pr

40
41



42

43 Early Developmental Effects of Phytochromes Phytochromes regulate (100 s of processes described) -- stem elongation -- cotyledon expansion -- chloroplast development (greening) -- apical hook opening -- gene expression Microarray analyses show that the early effects of phytochromes are to induce the expression of transcription factors that then alter the expression of genes involved in photomorphogenic development (Tepperman et al., 2001).
44 Fytochromy jsou syntetizovány v Pr formě a vedle fotokonverze existuje temnostní reverze Pfr na Pr u mnoha dvouděložných (není u jednoděložných, trav)
45
46 PhyA je foto-labilní Phy B,C,D,E jsou foto-stabilní
47 PhyA je nejen nestabilní na světle (PfrA-1h p.r.), ale dochází také k poklesu jeho transkripce zprostředkovaného fytochromovým systémem
48 Phy v Pfr formě jsou kinázy Autofosforylace, fosf. regulatory (Aux/IAA, TFs), kryptochromy Jen PfrB jde do jádra (FR to inhibuje) kin. hodiny. PhyA obě formy v jádře po osvětlení- kin. 15min. PfrB váže TF PIF3 transkr. světlem regul. genů

49

50 PKS, NDPK2 PIF3 and PHYB only interact if phyb is in Pfr state.

51 Interaction between PIF3 and PHYB is dependent upon state of PHYB PIF3 bound to bead, PHYB radioactively labeled. Extracts were treated with R or FR, spun down, and electrophoresed. PIF3 and PHYB only interact if phyb is in Pfr state. Ni et al., 1999
52 Fytochromoví mutanti Jak je hledat?
53 Jsou i na světle částečně slepí - a tedy vypadají jako ve tmě. THE PLANT CELL, Vol 3, Issue , Copyright 1991 by American Society of Plant Biologists RESEARCH ARTICLES The hy3 Long Hypocotyl Mutant of Arabidopsis Is Deficient in Phytochrome B D. E. Somers, R. A. Sharrock, J. M. Tepperman and P. H. Quail University of California, Berkeley/U.S. Department of Agriculture, Plant Gene Expression Center, Albany, California 94710
54
55 PhyB se např. účastní úniku ze stínu
56
57 Kryptochromy Vyvinuly se z fotolyáz nezávisle u rostlin a živočichů
 described BL receptors as cryptochromes. hy4 mutants showed a lack of hypocotyl growth inhibition under blue light,but were normal under red and far-red.")
58 Cryptochromes Many plant responses were not R-FR reversible and had action spectra with peaks in the blue and near-uv. There must be a BL receptor(s). Due to their elusive nature, Gressel (1977) described BL receptors as cryptochromes. hy4 mutants showed a lack of hypocotyl growth inhibition under blue light,but were normal under red and far-red. Dark BL Wild-type adapted from Neff and Chory, 1998 The sequence of Hy4 was reported in 1993, and it was similar to DNA photolyase (Ahmad and Cashmore 1993, Sancar 1994) yet has no photolyase activity. Re-designated Cryptochrome 1 (Lin et al., 1995) Involvement in circadian rhythms; led to discovery of animal crys (Cashmore, 2003) cry s are phosphorylated when illuminated (Shilatin et al., 2003, Bouvy et al., 2003). The timing of phosphorylation agrees well with the time course of early physiology (Folta and Spalding, 2001) BL cry1


59 The C-Terminus CCT of cry1 Regulates Photomorphogenesis Ectopic overexpression of the CRY1 C-terminal extension results in a constitutive-photomorphogenic phenotype. Is cry functioning through a mechanism involving COP1? Cashmore, 2003
60 Pozitivní a negativní regulace fotomorfogeneze

61



62 Xing Wang Deng, Yale COP Mutants First isolated by Deng et al., Constitutive Photomorphogenic phenotype: expanded cotyledons, short hypocotyls, light-regulated gene expression patterns in darkness 1996 Mayer et al. show that COP1 mutation affects expression of many genes not just specific to photomorphogenesis.
63 CSN komplex (viz. nahoře) COP9 byla první známá podjednotka signalosomu.


64 The hy5 mutant long hypocotyl under light conditions, particularly blue (Koorrneef et al., 1980). Encodes a B-zip transcription factor that is presumably a positive regulator of photomorphogenesis. Plants grown for days in light show different levels of HY5 HY5 level correlates with advanced photomorphogenic development. HY5 accumulates rapidly in light and is not as detectable upon transfer of plants to darkness fluence rate How does HY5 regulate photomorphogenesis? Is it simply present only in light and acting as a positive regulator? Is it more complex? It seems to be acting in a manner that is antagonistic to COP1. Osterlund et al., 2000
 Conclusions: The WD repeat domain is necessary and sufficient for interaction, yet binding is strongly enhanced by the")
65 Does COP1 Interact with CRY? Yeast 2-hybrid assay between COP1 truncations and CCT2 (CRY2 c-terminus) Conclusions: The WD repeat domain is necessary and sufficient for interaction, yet binding is strongly enhanced by the coiled-coil and Zn binding domains. CCT2-GUS interacts similarly. Coimmunoprecipitation Using anti-cry2, COP1 can be coimmunoprecipitated, demonstrating likely interaction in vivo. Wang et al., 2001

66 COP and CCT1 Co-Localize to the Nucleus GFP::COP1 GFP::CCT1

67 Proposed Model for Cryptochrome Function COP1, CRY, HY5 Interaction to Regulate Degradation of HY5 Hellmann and Estelle, 2002
68 v jádře ve tmě, na světle v cytoplasmě v jádře v mitochondriích i plastidech novější hypotéza

69
70 Fototropismus a FOTOTROPINY
71 opět auxin a jeho transport

72 DARK CHLOROPLAST MOVEMENTS -LEMNA WEAK BLUE LIGHT STRONG BLUE LIGHT

73 FOTOTROPINY Adiantum kapradina

74
75 Mechanismus asociace s membránou je nejasný.
76 Po osvětlení je část Phot1 bílkoviny uvolněna do cytoplazmy. V kortexu etiolovaného hypokotylu je Phot1 bílkovina přednostně polarizovaně kolokalizována s Pin1 na příčných stěnách.
77
78 Fotoperiodická indukce kvetení a biologické hodiny
79
80
81

82 Biological clocks jsou odolné vůči změnám teploty Two concepts associated with biological clocks Free running period Entrainment by light
83 "Biologické hodiny" tikají v každé buňce rostliny a s vnějším časovým cyklem komunikují přes fytochormový/kryptochromový systém.
84 Transkriptomická analýza Arabidopsis ukázala, že asi 6000 genů je exprimováno v diurnálním rytmu, z toho asi 500 přímo v závislosti na "centrálním oscilátoru/hodinách"
85 cyklus prostředí "hodiny" GATING

86 na krátkém dnu
87
88 Rostlinné "hormony" Auxins Gibberellins Cytokinins Abscisic acid Ethylene Brassinosteroids JA, SA

89

90

91

92 Signals are perceived at the cell membrane and transduced to the nucleus, resulting in a change in gene expression Receptor proteins, often with carbohydrate sidechains One to many 2 signals: proteins, ions, or the morphogens themselves. The last transducer is a gene regulator protein.
93 Auxiny
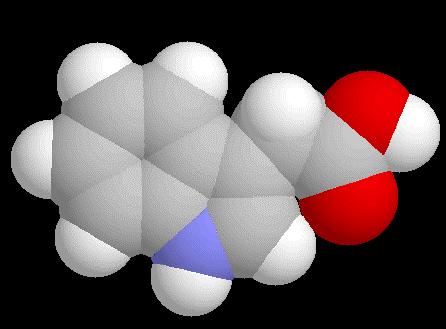
94 Auxin
95

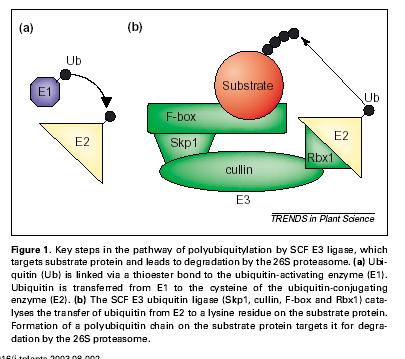
96 Hladina aktivního auxinu je regulována modifikacemi/konjugacemi - reverzibilními i ireverzibilními.

97 Polar transport of Auxin Chemoosmotická teorie transportu auxinu potvrzena objevem AUX1 vtokového a PIN výtokových přenašečů a jejich polární lokalizace. Transport jde ovšem také pře ABC transportery (PGP, závislé na ATP) a je možná také sekrece z IAA naplněných váčků jako v případě neurotransmiterů.
98 Auxin - přenos signálu

99
100 Fungují PINy jako senzory? Co je receptorem auxinu???


101 ABP1 Auxin Binding Protein1 byl objeven biochemicky je lokalizován převážně do membrány ER, ale malá frakce "uniká" a je aktivní na povrchu buňky. Pokus s blokováním reakce protoplastů na IAA protilát. prokázal jeho podíl na reakci na IAA.
102 Mutanti monopteros(mp) a bodenlos(bdl) nemají kořen. MP kóduje člena ARF (auxin response transcript. factors) rodiny transkripčních aktivátorů ARF5. (Prosím neplést! s malými GTPázami). BDL kóduje člena rodiny inhibitorů ARFů AUX/IAA, IAA12. Tyto bílkoviny se vyznačují minutovým poločasem životnosti. S ARFy tvoří heterodimery a tak blokují jejich aktivitu. Transkripce AUX/IAA genů je stimulována IAA (okamžitě) - tak byly objeveny.
103 Řada mutantů s postiženým proteasomem (a také CSN) má auxinový fenotyp - bývají auxin rezistentní.
104 Model regulace ARFů AUX/IAA a jejich auxinem stimulovanou proteolýzou. BDL a další AUX/IAA geny jsou také cílem transkripční stimulace.
105 De novo syntéza AUX/IAA represorů umožňuje potlačení signálu = atenuaci. Podobně kinázy fungují jako přenašeče signálů jen díky protipůsobícím fosfatázám. a tak je to i s Ca2+ a dalšími.
106 F-BOX SCF-E3 LIG. KOMPLEXU TIR1 JE RECEPTOREM AUXINU.
107 TIR1
108 Receptorem auxinu je TIR1 a jeho homology: F-box podjednotka SCF E3 ligázy.




109
110 TIR1 a spol. jsou regulovány neddylací/rub1 modifikací a tedy také CSN.


111 NEDD8=RUB1
112
Signalizace je vlastně komunikace. a komunikace je základem života.
 Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace. a komunikace je základem života.
 Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně komunikace a komunikace je základem života. Signalizace, regulace, komunikace a integrace v buňce, pletivu a organizmu Několik poznámek. Kvantitativně převažujícím prvkem regulace
Signalizace je vlastně
 Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů
 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
Buněčný cyklus - principy regulace buněčného Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Průběh mitózy v buněčné kultuře fibroblastů Buněčný cyklus Kinázy závislé na cyklinech
9) Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer
 Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer") 2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
MFPSB 1. b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze
 Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze") 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner Martin Fellner 2010 2 a) Fotomorfogeneze
1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner Martin Fellner 2010 2 a) Fotomorfogeneze
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění
 Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Buněčný cyklus - principy regulace buněčného růstu a buněčného dělění Mitóza Dr. B. Duronio, The University of North Carolina at Chapel Hill Buněčný cyklus Kinázy závislé na cyklinech kontrolují buněčný
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
1) Úloha světla a fytochromů ve vývoji a růstu rostlin
 Úloha světla a fytochromů ve vývoji a růstu rostlin") S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
RVR e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu
 Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu") 1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
5) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") 2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
4) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
19. století. b) Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR
 Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR") SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
2) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
Růst a vývoj rostlin - praktikum MB130C78 Blok I Hormonální regulace vývoje rostlin Cvičení MB130C78: Růst a vývoj rostlin. Katedra experimentální biologie rostlin, PřF UK, Jan Petrášek Úlohy: 1. Pozitivní
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
VORF Winslow Russel Briggs ( )
") VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Database systems. Normal forms
 Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Database systems Normal forms An example of a bad model SSN Surnam OfficeNo City Street No ZIP Region President_of_ Region 1001 Novák 238 Liteň Hlavní 10 26727 Středočeský Rath 1001 Novák 238 Bystřice
Gymnázium, Brno, Slovanské nám. 7 WORKBOOK. Mathematics. Teacher: Student:
 WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
WORKBOOK Subject: Teacher: Student: Mathematics.... School year:../ Conic section The conic sections are the nondegenerate curves generated by the intersections of a plane with one or two nappes of a cone.
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Buněčný cyklus. When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants.
 Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Buněčný cyklus cyklus buněčných procesů začínajících
Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Buněčný cyklus cyklus buněčných procesů začínajících
(molekulární) biologie buňky
 biologie buňky") (molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
(molekulární) biologie buňky Buňka základní principy Molecules of life Centrální dogma membrány Metody GI a MB Interakce Struktura a funkce buňky - principy proteiny, nukleové kyseliny struktura, funkce
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Biosyntéza a degradace proteinů. Bruno Sopko
 Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
PŘENOS SIGNÁLU V BUŇCE. Nela Pavlíková
 PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
Just write down your most recent and important education. Remember that sometimes less is more some people may be considered overqualified.
 CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
CURRICULUM VITAE - EDUCATION Jindřich Bláha Výukový materiál zpracován v rámci projektu EU peníze školám Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Bc. Jindřich Bláha. Dostupné z Metodického
3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFR 1 2017 3) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky
 Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
Evoluce fenotypu V Existence trade-offs záleží na proximátních mechanismech ovlivňujících znaky Parameters of body size and developmental time: the growth rate the initial weight the ICG Celkový vztah
6. Buňky a rostlina. Mají rostliny kmenové buňky?
 6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
6. Buňky a rostlina Mají rostliny kmenové buňky? Biotechnologické využití pluripotence rostlinných buněk: buněčné a tkáňové kultury rostlin in vitro, vegetativní množení rostlin Komunikace mezi buňkami
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora
 Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora Klinika dětské anesteziologie a resuscitace, Lékařská fakulta MU, Fakultní nemocnice Brno DNA transkripce
Má tajemný clusterin u dětí v septickém stavu aktivitu chaperonu? J. Žurek, P.Košut, M. Fedora Klinika dětské anesteziologie a resuscitace, Lékařská fakulta MU, Fakultní nemocnice Brno DNA transkripce
Proteiny Genová exprese. 2013 Doc. MVDr. Eva Bártová, Ph.D.
 Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Proteiny Genová exprese 2013 Doc. MVDr. Eva Bártová, Ph.D. Bílkoviny (proteiny), 15% 1g = 17 kj Monomer = aminokyseliny aminová skupina karboxylová skupina α -uhlík postranní řetězec Znát obecný vzorec
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
Struktura a funkce biomakromolekul KBC/BPOL 3. Enzymy a proteinové motory Ivo Frébort Enzymová katalýza Mechanismy enzymové katalýzy o Ztráta entropie při tvorbě komplexu ES odestabilizace komplexu ES
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
Biosensors and Medical Devices Development at VSB Technical University of Ostrava
 VŠB TECHNICAL UNIVERSITY OF OSTRAVA FACULTY OF ELECTRICAL ENGINEERING AND COMPUTER SCIENCE Biosensors and Medical Devices Development at VSB Technical University of Ostrava Ing. Martin Černý Ph.D. and
VŠB TECHNICAL UNIVERSITY OF OSTRAVA FACULTY OF ELECTRICAL ENGINEERING AND COMPUTER SCIENCE Biosensors and Medical Devices Development at VSB Technical University of Ostrava Ing. Martin Černý Ph.D. and
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA
 GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
GUIDELINES FOR CONNECTION TO FTP SERVER TO TRANSFER PRINTING DATA What is an FTP client and how to use it? FTP (File transport protocol) - A protocol used to transfer your printing data files to the MAFRAPRINT
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Joanne Chory (1955) VORF-4.
 VORF-4.") VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
Struktura a funkce biomakromolekul KBC/BPOL
 Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
Struktura a funkce biomakromolekul KBC/BPOL 2. Posttranslační modifikace a skládání proteinů Ivo Frébort Biosyntéza proteinů Kovalentní modifikace proteinů Modifikace proteinu může nastat předtím než je
DUM č. 11 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
projekt GML Brno Docens DUM č. 11 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 30.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Princip genové exprese, intenzita překladu
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost.
 Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Tento materiál byl vytvořen v rámci projektu Operačního programu Vzdělávání pro konkurenceschopnost. Projekt MŠMT ČR Číslo projektu Název projektu školy Klíčová aktivita III/2 EU PENÍZE ŠKOLÁM CZ.1.07/1.4.00/21.2146
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Fyziologie AUTOFAGIE. MUDr. JAN VARADY KARIM FNO
 Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Prokaryotická X eukaryotická buňka. Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen)
") Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,
Prokaryotická X eukaryotická buňka Hlavní rozdíl organizace genetického materiálu (u prokaryot není ohraničen) Cytoplazmatická membrána osemipermeabilní ofosfolipidy, bílkoviny otransport látek, receptory,
2N Voice Alarm Station
 2N Voice Alarm Station 2N Lift1 Installation Manual Version 1.0.0 www.2n.cz EN Voice Alarm Station Description The 2N Voice Alarm Station extends the 2N Lift1/ 2N SingleTalk with an audio unit installed
2N Voice Alarm Station 2N Lift1 Installation Manual Version 1.0.0 www.2n.cz EN Voice Alarm Station Description The 2N Voice Alarm Station extends the 2N Lift1/ 2N SingleTalk with an audio unit installed
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL
 STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E.
 Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E. Pokorný Mendel University of Agriculture and Forestry, Department of
Characterization of soil organic carbon and its fraction labile carbon in ecosystems Ľ. Pospíšilová, V. Petrášová, J. Foukalová, E. Pokorný Mendel University of Agriculture and Forestry, Department of
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Regulace translace REGULACE TRANSLACE LOKALIZACE BÍLKOVIN V BUŇCE. 4. Lokalizace bílkovin v buňce. 1. Translační aparát. 2.
 Regulace translace 1. Translační aparát 2. Translace 3. Bílkoviny a jejich posttranslační modifikace a jejich degradace 5. Translace v mitochondriích a chloroplastech REGULACE TRANSLACE LOKALIZACE BÍLKOVIN
Regulace translace 1. Translační aparát 2. Translace 3. Bílkoviny a jejich posttranslační modifikace a jejich degradace 5. Translace v mitochondriích a chloroplastech REGULACE TRANSLACE LOKALIZACE BÍLKOVIN
VY_32_INOVACE_06_Předpřítomný čas_03. Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace
 VY_32_INOVACE_06_Předpřítomný čas_03 Autor: Růžena Krupičková Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace Název projektu: Zkvalitnění ICT ve slušovské škole Číslo projektu: CZ.1.07/1.4.00/21.2400
VY_32_INOVACE_06_Předpřítomný čas_03 Autor: Růžena Krupičková Škola: Základní škola Slušovice, okres Zlín, příspěvková organizace Název projektu: Zkvalitnění ICT ve slušovské škole Číslo projektu: CZ.1.07/1.4.00/21.2400
2. Entity, Architecture, Process
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Praktika návrhu číslicových obvodů Dr.-Ing. Martin Novotný Katedra číslicového návrhu Fakulta informačních technologií ČVUT v Praze Miloš
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Praktika návrhu číslicových obvodů Dr.-Ing. Martin Novotný Katedra číslicového návrhu Fakulta informačních technologií ČVUT v Praze Miloš
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
REGULACE TRANSLACE DEGRADACE BÍLKOVIN. 4. Degradace bílkovin. 4. Degradace bílkovin. 4. Degradace bílkovin
 4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
4. Degradace bílkovin Degradace - několik proteolytických cest, specifických pro určitý buněčný kompartment REGULACE TRANSLACE DEGRADACE BÍLKOVIN 4. Degradace bílkovin 4. Degradace bílkovin Degradace bílkovin
Autofagie a výživa u kriticky nemocného pacienta
 Autofagie a výživa u kriticky nemocného pacienta Igor Satinský Nemocnice Havířov Mezioborová JIP Colours of Sepsis, Ostrava, 28.1.2015 Autofagie a výživa u kriticky nemocného pacienta Igor Satinský Nemocnice
Autofagie a výživa u kriticky nemocného pacienta Igor Satinský Nemocnice Havířov Mezioborová JIP Colours of Sepsis, Ostrava, 28.1.2015 Autofagie a výživa u kriticky nemocného pacienta Igor Satinský Nemocnice
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Teoretický úvod: FOTOTROPISMUS. Praktikum fyziologie rostlin
 Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
Teoretický úvod: FOTOTROPISMUS Praktikum fyziologie rostlin 1 Teoretický úvod: FOTOTROPISMUS Fototropismus náleží mezi vitální ohybové pohyby rostlin. Řadí se mezi pohyby paratonické povahy, tj. je vyvolán
Endosomy, vakuola a ti druzí
 Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
Endocytóza Endosomy, vakuola a ti druzí Endocytické váčky mají clathrinový obal Various receptors and clathrin protein coats recognize vesicle types, and these are delivered (along cytoskeleton pathways?)
TRANSLACE - SYNTÉZA BÍLKOVIN
 TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
TRANSLACE - SYNTÉZA BÍLKOVIN Translace - překlad genetické informace z jazyka nukleotidů do jazyka aminokyselin podle pravidel genetického kódu. Genetický kód - způsob zápisu genetické informace Kód Morseovy
Aktivita CLIL Chemie I.
 Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
Škola: Gymnázium Bystřice nad Pernštejnem Jméno vyučujícího: Mgr. Marie Dřínovská Aktivita CLIL Chemie I. Název aktivity: Uhlíkový cyklus v přírodě Carbon cycle Předmět: Chemie Ročník, třída: kvinta Jazyk
Dusík. - nejdůležitější minerální živina (2-5% SH)
") Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Dusík - nejdůležitější minerální živina (2-5% SH) - dostupnost dusíku ovlivňuje: - produkci biomasy a její distribuci - ontogenetický vývoj - hormonální rovnováhu (cytokininy, ABA) - rychlost fotosyntézy
Detlef Weigel ( )
") VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Litosil - application
 Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
Litosil - application The series of Litosil is primarily determined for cut polished floors. The cut polished floors are supplied by some specialized firms which are fitted with the appropriate technical
Configuration vs. Conformation. Configuration: Covalent bonds must be broken. Two kinds of isomers to consider
 Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
Stereochemistry onfiguration vs. onformation onfiguration: ovalent bonds must be broken onformation: hanges do NT require breaking of covalent bonds onfiguration Two kinds of isomers to consider is/trans:
WORKSHEET 1: LINEAR EQUATION 1
 WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable
WORKSHEET 1: LINEAR EQUATION 1 1. Write down the arithmetical problem according the dictation: 2. Translate the English words, you can use a dictionary: equations to solve solve inverse operation variable