MFPSB 1. b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze
|
|
|
- Dagmar Kubíčková
- před 5 lety
- Počet zobrazení:
Transkript
1 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner Martin Fellner 2010
2 2 a) Fotomorfogeneze Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer Rockwell NC et al. (2006) Phytochrome Structure and Signaling Mechanisms. Annu Rev Plant Biol 57: p




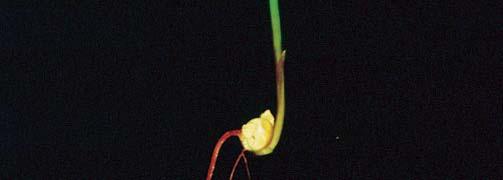



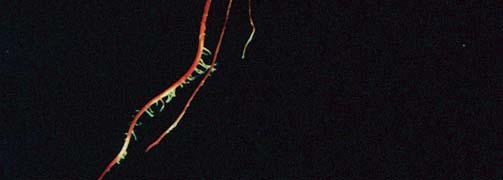






3 3 Růst ve tmě (etiolizovaný růst, skotomorfogeneze) Růst na světle (fotomorfogeneze)
4 4 Fotomorfogeneze = světlem indukované změny ve vývoji a růstu rostliny. Proces, při kterém světlo jako signál změní vývoj rostliny tak, aby mohla pro svůj další růst využít světlo jako energii. Světlo Základní fotomorfogenické reakce: Tma - inhibice prodlužování - stimulace syntézy chlorofylu - stimulace růstu listu - regulace klíčení
5 5 Při fotomorfogenezi je světlo zachycováno pigmenty, které jsou součástí fotoreceptorů: červeného světla: fytochromy A až E (phytochromes) modrého světla a UV-A: kryptochromy a fototropiny (cryptochromes, phototropins) Absorpce světla aktivace signálních drah Aktivace světlem indukovaných genů



6 6 Fytochrom funguje v jádře aktivuje transkripční faktory. Je však lokalizovaný původně v cytoplazmě => musí být přemísťěn do jádra. Sharma R (2001) Current Science 80: Fytochrom se přemísťuje ř ť do jádra vlivem světla Vizualizace pomocí GFP (green fluorescent protein; GFP aktivovaný světlem emituje fluorescenční záření) - Pohyb phyb indukován R, inhibován FR;! do jádra putuje pouze ve formě Pfr, pohyb je pomalý - Pohyb phya indukován FR; putuje v obou formách; pohyb je rychlý. Konstrukt Promotor PHYB GFP Transformace rostlin phya-gfp phyb-gfp Sledování exprese PHYB v buňkách a pletivech
 Jaderný protein PIF3 (phytochrome y interacting")
 - reaguje s C- terminálním koncem fytochromu B Pfr tvoří komplex s PIF3")

7 7 Regulace genové exprese fytochromem B PhyB je syntetizován v cytolazmě ve formě PrB PIC iniciuje transkripci MYB genů (CCA1, LHY) Jaderný protein PIF3 (phytochrome y interacting factor3): ) -přesná funkce není známa transkripční faktor reagující s G-boxem (= část promotoru genu MYB) - reaguje s C- terminálním koncem fytochromu B Pfr tvoří komplex s PIF3 Nové informace Kim J et al. (2003) Plant Cell 15: Ve formě PfrB je fytochrom transportován do jádra PfrB se váže k dimeru PIF3 Transkripční faktor MYB aktivuje transkripci LHCB
8 8 b) Úloha COP1 ve fotomorfogenezi rostlin Genetický skríning Selekce mutantů Identifikace genů cop1 (constitutive photomorphogenesis) - etiolizované rostliny ukazují fenotyp rostlin rostoucích na světle
9 9 Mutovaný (poškozený) gen COP1 vede ke konstitutivní fotomorfogenezi Zdravý gen COP1 negativní regulátor fotomorfogeneze COP1 centrální tál gen fotomorfogeneze f rostlin
10 10 Struktura Arabidopsis COP1 (AtCOP1) NES RING Coiled-coil NIS WD40 N-terminální doména: RING finger a coiled-coil doména RING finger a coiled-coil doména vnesení do cop1 mutanta vede k osvobození letálního fenotypu N-terminální doména má základní funkci ve vývoji rostlin NES = leucine-rich Nuclear Export Signal NIS = Nuclear Import Signal C-terminální doména: WD40
11 11 Interakce Arabidopsis COP1 s dalšími proteiny NES RING Coiled-coil NIS WD40 COP10 CIP8 SPA1-4, COP1, CIP1, CIP4, CIP7 HY5, HYH, STO, HFR1, CRY1, CRY2, phya, phyb Nezmapovaný faktor: LAF1
12 12 COP1 funguje jako E3 ubiquitin ligáza Negativní funkce COP1 byla odhalena pomocí HY5 HY5 bzip transkripční faktor: - akumuluje se v reakci na světlo - v reakci na světlo aktivuje fotomorfogenické geny - ve tmě dochází k degradaci HY5 pomocí proteolýzy zprostředkované proteazomem Proteolýza zprostředkovaná proteazomem vyžaduje protein ubiquitin ubiquitin protein ubiquitin protein RING finger E3 ubiquitin ligáza Proteazom
13 13 COP1 podezřelý z toho, že jde od HY5 E3 ubiquitin ligázu důkazy: -COP1 interaguje přímo s HY5 - degradace HY5 je narušena v mutantu cop1 - mutant cop1 s bodovou mutací v WD40 špatně interaguje s HY5 - HY5 se stabilizuje na světle, když COP1 je z jádra odstraněn COP1 = E3 ubiquitin ligáza COP1 rovněž ubiquitinuje HYH a transkripční faktory jiných typů: LAF1 (Myb) nebo HFR1 (bhlh) COP1 funguje jako hlavní vypínač: ve tmě vypíná fotomorfogenezi degradací transkripčních faktorů, které aktivují specifické geny fotomorfogeneze
14 14 phya fotoreceptor dlouhovlného červeného světla (FR); na světle extrémně labilní CRY2 fotoreceptor modrého světla; na světle extrémně labilní COP1 přímo interaguje s phya a CRY2 a směřuje je k ubiquitinaci proteazomem Ochrana před nadměrnou aktivací signálních drah phyb fotoreceptor červeného světla; na světle stabilní CRY1 fotoreceptor modrého světla; na světle stabilní COP1 přímo interaguje s phyb a CRY1, ale nemá vliv na jejich stabilitu COP1 reguluje interakci i phyb a CRY1 s terčovými č proteiny Schéma úlohy COP1 ve fotomofogenezi
15 15 Schéma úlohy COP1 ve fotomofogenezi

 Plant J (April 2008) Nový transkripční faktor BIT1 (MYB transcription factor Blue Insensitive Trait 1)")
 a tak aktivuje jeho expresi.")
16 Funkce COP1 je modulována nukleoplazmatickou translokací 16 Mutace Leu zbytků v doméně COP1 odpovědné za export proteinu z jádra Zvýšená akumulace mutovaného COP1 v jádře Transgenní rostliny overexprimující COP1 s mutovanou doménou zodpovědnou za import proteinu do jádra Update 2008 Hong SH et al. (2008) Plant J (April 2008) Nový transkripční faktor BIT1 (MYB transcription factor Blue Insensitive Trait 1) Tma: degradace BIT1 působením COP1. BL: BIT1 stabilizován CRY1, není degradován. BIT1 se váže k promotoru Pdbs (člen light-harvesting complex genů) a tak aktivuje jeho expresi. Snížena akumulace mutovaného COP1 v jádře = konstitutivní fotomorfogeneze
17 17 Faktory regulující COP1 1) Proteiny SPA - kvasinkový dvouhybridní skríning - genetické interakce - purifikace proteinových komplexů - regulují funkci COP1 v signální dráze phya - obsahují N-terminální kinase-like doménu, coiled-coil doménu, WD40 na C-konci - WD40 vysoce homologní k WD40 doméně COP1 - fyzicky interagují s COP1 prostřednictvím coiled-coil domény NES RING Coiled-coil NIS WD40 COP10 CIP8 SPA1-4, COP1, CIP1, CIP4, CIP7 HY5, HYH, STO, HFR1, cry1, cry2, phya, phyb Mechanizmus regulace COP1 proteiny SPA není znám. - zvyšují aktivitu (fosforylací COP1) - usnadňují interakci COP1 se substráty - stabilizují COP1 komplexy


18 18 2) Kryptochromy - na světle CRY1 a CRY2 jsou fosforylovány => stávají se aktivními - na světle CRY1 a CRY2 negativně regulují aktivitu COP1 asociací C-terminální domény CRY (CCT) s doménou WD40 proteinu COP1 - transgenní rostliny exprimující CCT1, CCT2 => cop1 fenotyp - transgenní rostliny s mutací v CCT narušena interakce s COP1 => bez cop1 fenotypu

n CRY Neaktivní")

19 19 Navržený mechanismus regulace COP1 kryptochromy Modré světlo => přenos elektronů uvnitř PHR => změna konformace CCT => autofosforylace CRY => aktivace kryptochromu Modré světlo Aktivní (Pi)n CRY Neaktivní PHR CCT PHR CCT Kináza Fosfatáza PHR CCT PHR = Photolyase-related doména; N- terminální doména Aktivace kryptochromu Deaktivace COP1
20 20 3) COP/DET/FUS Další geny: DET (DE ETIOLATED) FUS (FUSCA) Mutanti s fenotypem konstitutivní fotomorfogeneze Součástí dvou velkých komplexů: COP9 signalozom CDD komplex COP9 signalozom konzervovaný u rostlin a živočichů; spojen s multikomplexy E3 ubiquitin ligázy; reguluje aktivitu E3 CDD komplex nalezen pouze u rostlin Vztah mezi COP, DET a FUS není zatím znám: - mutace cop/det/fus potlačuje jadernou akumulaci COP1 -stabilita CDD komplexu je postižena mutací v COP9 signalozomu - mutace cop10 vede ke změně velikosti komplexu COP1
21 21 c) COP1 a tumorogeneze COP1 velice konzervovaný u ryb, obojživelníků, ptáků a savců; nebyl identifikován u Drosophily, ale byl nalezen u komára MmCOP1 role v tumorogenezi a reakcích ke stresům Struktura savčího COP1 (MmCOP1) NE umožňuje interakci COP1 s jadernou obálkou COP1 migruje mezi jádrem a cytoplazmou. Má ale odlišné import a export signály (NIS, NES) Coiled-coil doména self dimerizace i COP1; interaguje s DET1 COP1 interaguje s c-jun, p53 a MVP prostřednictvím WD40
22 22 Funkce COP1 v savčích buňkách Stejně jako u rostlin, živočišný COP1 je zapojen v ubiquitinaci proteinů. Jeho úloha u savců je však mnohem méně známa než u rostlin Tři známé substráty COP1: p53 (tumor suppressor protein) c-jun (proto-oncogene) MVP (major vault protein) p53 (tumor suppressor protein) COP1 degraduje p53: Overexprese COP1 Inhibice transkripce a apoptózy závislé na p53 COP1 byl overexprimován ve vysokém procentu v endokarcinomech prsu a vaječníků Eliminace COP1 Zastavení buněčného cyklu v G1 fázi Update 2008 He L et al. (2007) Nature 447: Transkripční factor p53 reguluje expresi genů kódujících microrna (mir-34), které potlačují buněčné dělení. Hladiny microrna jsou radikálně sníženy v nádorových buňkách.
23 23 U rostlin je aktivita COP1 regulována světlem. Otázka: Co reguluje aktivitu COP1 u živočichů? Co způsobuje zvýšení či redukci exprese COP1? p53 Poškození DNA Protein ATM fosforyluje COP1 na Ser 387 Transport COP1 z jádra do cytoplazmy a autodegradace COP1 Rozpad komplexu COP1-p53 Zastavení ubiquitinace a degradace p53 Apoptóza Update 2008 Poškození DNA důsledek UV a ionizačního záření P53 je nejčastěji mutovaným supresorem e Benjamin CL et al. (2008) Photochem Photobiol 84: UVB indukuje mutaci (C=>T a CC=>TT) v genu p53. Mutace jsou spojené s vývojem rakoviny kůže. Mutace p53 - detekovány v myší kůži dopředu před vypuknutím rakoviny. Využití: Testování opalovacích krémů. Bensaad K, Vousden KH (2007) Trends in Cell Biology 17: Rakoviny ukazují metabolické změny vedoucí k upregulaci glykolýzy a spotřeby glukózy. p53 reguluje metabolismus glukózy a oxidativní stres - potlačování vzniku nádorů
24 24 c-jun MVP UV záření UV záření Akumulace c-jun Fosforylace MVP Iniciace transkripce AP-1 (activator protein) Rozpad komplexu COP1-MVP Redukce transkripce p53 Redukce transkripce AP-1 (activator protein) Tumorogeneze Apoptóza
25 Ionizační 25 záření UV záření Stres Poškození DNA COP1 Transport do cytoplazmy P P COP1 p53 p53 Apoptóza ATM P c-jun P c-jun P c-jun p53 AP-1 COP1 Tumorogenesis Transport do cytoplazmy P MVP P MVP COP1 MVP = major vault protein Transport do cytoplazmy COP1 COP1 = constitutive photomorphogenesis 1 ATM = ataxia telangiectasia mutated AP-1 = activator protein p53 = tumor suppressor protein c-jun = proto-oncogene
26 26 Použitá literatura Hong SH et al. (2008) Plant J (April 2008) Benjamin CL et al. (2008) Photochem Photobiol 84: Bensaad K, Vousden KH (2007) Trends in Cell Biology 17: He L et al. (2007) Nature 447: Jiao Y et al. (2007) Nature Reviews in Genetics 8: Taiz L, Zeiger E (2006) Plant Physiology. 4th Edition, Sinauer Asociates, Inc. Publishers, Sunderland, Massachusetts Hoeller D et al. (2006) Nature Reviews Cancer 6: Dornan D et al. (2006) Science 313: Burger A et al. (2006) Neoplasia 8: Yi C et al. (2005) Cancer Res 65: Yi C, Deng XW (2005) Trends in Cell Biology 15: Dornan D et al. (2004) Cancer Research 64: Wertz IE et al. (2004) Science 303: Dornan D et al. (2004) Nature 429: Bianchi E et al. (2003) J Biol Chem 278: Yi C et al. (2002) BMC Cell Biology 3: 30-46
27 27 Kdo jsou? Xing-Wang Deng Yale University, New Haven Vishva M. Dixit Genentech, Inc., San Francisco Joanne Chory The Salk Institute, La Jolla? Chunling Yi Yale University, New Haven David Dornan Genentech, Inc., San Francisco
28 28 Klíčová slova pro příští přednášku Boron Boron transporter Boron channel Boron transport NIP5;1 Borate transport NaBC1 Na + -coupled borate transporter Boric acid BOR1 Xylem loading Google: NCBI:
Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů
 1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
1 Úloha genu COP1 v rostlinné fotomorfogenezi a tumorogenezi u živočichů a) Fotomorfogeneze b) Úloha COP1 ve fotomorfogenezi rostlin c) COP1 a tumorogeneze Martin Fellner 2015 2 a) Fotomorfogeneze Briggs
9) Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer
 Fotomorfogeneze RVR. Schäfer E, Nagy F (eds) (2006) Photomorphogenesis in Plants and Bacteria, 3rd ed., Springer") 2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
2015 9) Fotomorfogeneze a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů 1 Briggs
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
2014 3) Role světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) Reakce rostlin zprostředkované fytochromy 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
1) Úloha světla a fytochromů ve vývoji a růstu rostlin
 Úloha světla a fytochromů ve vývoji a růstu rostlin") S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
S 2018 1) Úloha světla a fytochromů ve vývoji a růstu rostlin a) Vlastnosti a lokalizace fytochromů b) eakce rostlin zprostředkované fytochromy c) Ekologické funkce fytochromů d) Buněčný a molekulární
3) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
1 2014 3) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Whitelam GC, Halliday KJ (eds) (2007) Light and Plant
4) Role světla a fytochromů ve vývoji a růstu rostlin
 Role světla a fytochromů ve vývoji a růstu rostlin") 1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
1 2010 4) Role světla a fytochromů ve vývoji a růstu rostlin c) Ekologické funkce fytochromů d) Buněčný a molekulární mechanismus funkce fytochromů Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
4) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFR 1 2018 4) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
5) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") 2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
2010 5) Reakce rostlin k modrému světlu a) Fototropismus b) Rychlá inhibice prodlužovacího růstu c) Stimulace otevírání průduchů (stomat) 1 Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors,
6) Interakce auxinů a světla ve vývoji a růstu rostlin
 Interakce auxinů a světla ve vývoji a růstu rostlin") SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
SFZR 1 2015 6) Interakce auxinů a světla ve vývoji a růstu rostlin Martin Fellner Laboratoř růstových regulátorů PřF UP v Olomouci a ÚEB AVČR SFZR 2 Vývoj organismu regulují signály (faktory) Vnitřní Vnější
2) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
SFR 1 2017 2) Reakce rostlin k modrému světlu b) Fotoreceptory c) Přenos signálu Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR, Spudich JL (eds) (2005)
19. století. b) Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR
 Fotoreceptory c) Přenos signálu. b) Fotoreceptory modrého světla: Kryptochromy (cryptochromes) PHR") SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
SFR 2018 2) Reakce rostlin k modrému světlu 1 19. století Charles a Francis Darwin Studium fototropismu koleoptile 2 b) Fotoreceptory c) Přenos signálu Počátek 90. let Identifikace fotoreceptorů Identifikace
4) Reakce rostlin k modrému světlu
 Reakce rostlin k modrému světlu") SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
SFZR 1 2015 4) Reakce rostlin k modrému světlu a) Fotobiologie reakcí zprostředkovaných modrým světlem Whitelam GC, Halliday KJ (eds) (2007) Light and Plant Development Blackwell Publishing Briggs WR,
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RVR e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu
 Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu") 1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
1 2015 9) Fotomorfogeneze e) Fotobiologie reakcí zprostředkovaných modrým světlem f) Fotoreceptory g) Přenos signálu Briggs WR, Spudich JL (eds) (2005) Handbook of Photosensory Receptors, Wiley-VCH Schäfer
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
2) Reprodukce rostlin
 Reprodukce rostlin") 2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
2010 1 2) Reprodukce rostlin a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu 2 Životní cyklus rostliny 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") 2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
2015 2) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis
 u Arabidopsis") TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
TUBULIN-FOLDING COFACTOR A (TFC A) u Arabidopsis Mikrotubuly Formace heterodimerů α/βtubulinu Translace α a β -tubulin monomerů chaperonin c-cpn správný folding α-tubulin se váže na TFC B a β na TFC
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
SFZR 1 2016 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
7) Dormance a klíčení semen
 Dormance a klíčení semen") 2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
2015 7) Dormance a klíčení semen 1 a) Dozrávání embrya a dormance b) Klíčení semen 2 a) Dozrávání embrya a dormance Geny kontrolující pozdní fázi vývoje embrya - dozrávání ABI3 (abscisic acid insensitive
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2017-2018 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu. Školitel Mgr.
Životní cyklus rostliny. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení
 Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu. a) Indukce kvetení") MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
MBRO 208 4) Reprodukce rostlin Životní cyklus rostliny 2 a) Indukce b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Martin Fellner Laboratoř růstových regulátorů PřF
Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin
 1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
1 2014 Rostlinné hormony brasinosteroidy a jejich úloha ve vývoji a růstu rostlin Hayat S, Ahmad A (2011) Brassinosteroids: a class of plant hormone. Springer, Berlin 2 Vývoj organismu regulují signály
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Příběh pátý: Auxinová signalisace
 Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Příběh pátý: Auxinová signalisace Co je auxin? Derivát tryptofanu Příbuzný serotoninu a melatoninu Všechny deriváty přítomny jak u živočichů, tak u rostlin IAA Serotonin Serotonin: antagonista auxinu Přítomen
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
MBR ) Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu
 Reprodukce rostlin. a) Indukce kvetení. b) Vývoj květu - stručná morfologie. c) Genetická a molekulární analýza vývoje květu") MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
MBR1 2016 4) Reprodukce rostlin 1 a) Indukce kvetení b) Vývoj květu - stručná morfologie c) Genetická a molekulární analýza vývoje květu Životní cyklus rostliny 2 3 a) Indukce kvetení Indukce kvetení přeprogramování
Joanne Chory (1955) VORF-4.
 VORF-4.") VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
VORF-4 2015 Joanne Chory (1955) 1 Salk Institute for Biological Studies Plant Molecular and Cellular Biology Laboratory 10010 North Torrey Pines Road La Jolla, California 92037 USA http://pbio.salk.edu/pbioc/
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2015-2016 1 Název Funkční analýza jaderných proteinů fosforylovaných pomocí mitogenaktivovaných proteinkináz. Školitel
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Detlef Weigel ( )
") VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
VORF-8 2015 Detlef Weigel (15. 12. 1961) 1 Max Planck Institute for Developmental Biology Department of Molecular Biology Spemannstrasse 37-39 D-72076 Tübingen Germany http://www.weigelworld.org/ Max Planck
5. Příjem, asimilace a fyziologické dopady anorganického dusíku. 5. Příjem, asimilace a fyziologické dopady anorganického dusíku
 5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
5. Příjem, asimilace a fyziologické dopady anorganického dusíku Zdroje dusíku dostupné v půdě: Amonné ionty + Dusičnany = největší zdroj dusíku v půdě Organický dusík (aminokyseliny, aminy, ureidy) zpracování
a) Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) Bor v živočišných tkáních
 Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) Bor v živočišných tkáních") 1 Transport boru v rostlinách a živočiších. Jeho úloha v růstu a vývoji. a) Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) Bor v živočišných tkáních Martin Fellner 2010
1 Transport boru v rostlinách a živočiších. Jeho úloha v růstu a vývoji. a) Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) Bor v živočišných tkáních Martin Fellner 2010
Transport boru v rostlinách a živočiších. Jeho úloha v růstu a vývoji.
 1 Transport boru v rostlinách a živočiších. Jeho úloha v růstu a vývoji. a) Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) v živočišných tkáních Martin Fellner 2017 Xu F
1 Transport boru v rostlinách a živočiších. Jeho úloha v růstu a vývoji. a) Úvod b) Funkce boru v rostlinách c) Mechanizmy transportu boru v rostlinách d) v živočišných tkáních Martin Fellner 2017 Xu F
Růst a vývoj rostlin - praktikum MB130C78
 Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Růst a vývoj rostlin - praktikum MB130C78 Blok 3 Role aktinového cytoskeletu v morfogenezi rostlinných buněk - analýza fenotypu Úlohy: 1. Kvantifikace počtu zkroucených a správně tvarovaných trichomů u
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Bakalářské práce. Magisterské práce. PhD práce
 Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Bakalářské práce Magisterské práce PhD práce Témata bakalářských prací na školní rok 2018-2019 1 Název Fenotypová analýza vybraných dvojitých mutantů MAPK v podmínkách abiotického stresu Školitel Mgr.
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV
 VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV 1 Vladimír Špunda, 2 Otmar Urban, 1 Martin Navrátil 1 Přírodovědecká fakulta, Ostravská univerzita v Ostravě,
VLIV SPEKTRÁLNÍHO SLOŽENÍ FOTOSYNTETICKY AKTIVNÍ RADIACE NA INDUKCI FOTOSYNTÉZY TERMOOPTICKÝ JEV 1 Vladimír Špunda, 2 Otmar Urban, 1 Martin Navrátil 1 Přírodovědecká fakulta, Ostravská univerzita v Ostravě,
VORF Winslow Russel Briggs ( )
") VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
VORF-2 2015 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA Prof.
Buněčné jádro a viry
 Buněčné jádro a viry Struktura virionu Obal kapsida strukturni proteiny povrchove glykoproteiny interakce s receptorem na povrchu buňky uvnitř nukleocore (ribo )nukleova kyselina, virove proteiny Lokalizace
Buněčné jádro a viry Struktura virionu Obal kapsida strukturni proteiny povrchove glykoproteiny interakce s receptorem na povrchu buňky uvnitř nukleocore (ribo )nukleova kyselina, virove proteiny Lokalizace
Bi8240 GENETIKA ROSTLIN
 Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
Bi8240 GENETIKA ROSTLIN Prezentace 02 Reprodukční vývoj Indukce kvetení doc. RNDr. Jana Řepková, CSc. repkova@sci.muni.cz 1. Indukce kvetení a tvorba květů 2. Tvorba reprodukčních orgánů a gamet 3. Opylení,
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
Joanne Chory ( ) narozena v Massachusetts. má 4 sourozence. manžel Stephen Worland, biochemik. děti Katherine and Joe
 narozena v Massachusetts. má 4 sourozence. manžel Stephen Worland, biochemik. děti Katherine and Joe") 23.10.2017 VORF-4 2017 Joanne Chory (19.3. 1955) 1 narozena v Massachusetts 2 Salk Institute for Biological Studies lant Molecular and Cellular Biology Laboratory 10010 North Torrey ines Road La Jolla,
23.10.2017 VORF-4 2017 Joanne Chory (19.3. 1955) 1 narozena v Massachusetts 2 Salk Institute for Biological Studies lant Molecular and Cellular Biology Laboratory 10010 North Torrey ines Road La Jolla,
Auxin - nejdéle a nejlépe známý fytohormon
 Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
Auxin - nejdéle a nejlépe známý fytohormon Auxin je nejdéle známým fytohormonem s mnoha popsanými fyziologickými účinky Darwin 1880, Went 1928 pokusy s koleoptilemi trav a obilovin prokázali existenci
ONKOGENETIKA. Spojuje: - lékařskou genetiku. - buněčnou biologii. - molekulární biologii. - cytogenetiku. - virologii
 ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
Fotomorfogeneze: vliv světla na procesy vývoje rostlin ve výuce biologie na školách
 UNIVERZITA KARLOVA V PRAZE Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Učitelství biologie pro SŠ (dvouoborové) Bc. Libor Sedlecký Fotomorfogeneze: vliv světla na procesy vývoje rostlin
UNIVERZITA KARLOVA V PRAZE Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Učitelství biologie pro SŠ (dvouoborové) Bc. Libor Sedlecký Fotomorfogeneze: vliv světla na procesy vývoje rostlin
8 cyklinů (A, B, C, D, E, F, G a H) - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů
 - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů") Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
RŮST A VÝVOJ. Diferenciace rozlišování meristematických buněk na buňky specializované
 RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
RŮST A VÝVOJ Růst nevratný nárůst hmoty způsobený činností živé protoplasmy hmota a objem buněk, počet buněk, množství protoplasmy kvantitativní změny Diferenciace rozlišování meristematických buněk na
ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv
 u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv") Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Urbanová Anna ve srovnání s eukaryoty (životnost v řádu hodin) u prokaryot kratší (životnost v řádu minut) na životnost / stabilitu molekuly mají vliv strukturní rysy mrna proces degradace každá mrna v
Buněčné dělení ŘÍZENÍ BUNĚČNÉHO CYKLU
 BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
IV117: Úvod do systémové biologie David Šafránek 29.10.2008 Obsah Spojitý deterministický model transkripční regulace Obsah Spojitý deterministický model transkripční regulace Schema transkripční regulace
Senescence v rozvoji a léčbě nádorů. Řezáčová Martina
 Senescence v rozvoji a léčbě nádorů Řezáčová Martina Replikační senescence Alexis Carrel vs. Leonard Hayflick and Paul Moorhead Diferencované bb mohou prodělat pouze omezený počet dělení - Hayflickův limit
Senescence v rozvoji a léčbě nádorů Řezáčová Martina Replikační senescence Alexis Carrel vs. Leonard Hayflick and Paul Moorhead Diferencované bb mohou prodělat pouze omezený počet dělení - Hayflickův limit
Buněčný cyklus. When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants.
 Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Buněčný cyklus cyklus buněčných procesů začínajících
Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Buněčný cyklus cyklus buněčných procesů začínajících
Síra. Deficience síry: řepka. - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH
 Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
Síra řepka - 0,2-0,5% SH, nedostatek při poklesu obsahu síranů pod 0,01% SH - toxicita není příliščastá (nad 4000 mg SO 4 2- l -1 ), poškození může vyvolat SO 2 (nad 1-1,5 mg m 3 1 ) fazol Deficience síry:
AMPK AMP) Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze
 Tomáš Kuc era. Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze") AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
AMPK (KINASA AKTIVOVANÁ AMP) Tomáš Kuc era Ústav lékar ské chemie a klinické biochemie 2. lékar ská fakulta, Univerzita Karlova v Praze 2013 AMPK PROTEINKINASA AKTIVOVANÁ AMP přítomna ve všech eukaryotních
Regulace růstu a vývoje
 Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Regulace růstu a vývoje REGULACE RŮSTU A VÝVOJE ROSTLINNÉHO ORGANISMU a) Regulace na vnitrobuněčné úrovni závislost na rychlosti a kvalitě metabolických drah, resp. enzymů a genů = regulace aktivity enzymů
Protinádorová imunita. Jiří Jelínek
 Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Signalizace a komunikace. Rostlinná cytologie - signalizace, Katedra experimentální biologie rostlin PřF UK
 Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
Signalizace a komunikace Annu. Rev. Plant Biol. 2008.59:67-88 Development, 117 (1993), pp. 149 162 Meristémy trvale dělivá pletiva Periklinální dělení Antiklinální dělení http://www.mun.ca/biology/desmid/brian/biol3530/devo_07/ch07f05.jpg
VORF Winslow Russel Briggs ( )
") VORF-2 2018 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA http://dpb.carnegiescience.edu/labs/briggs-lab
VORF-2 2018 Winslow Russel Briggs (29.4. 1928) 1 Department of Plant Biology Carnegie Institution for Science (dříve Carnegie Institution of Washington) 260 Panama Street Stanford, CA 94 305 USA http://dpb.carnegiescience.edu/labs/briggs-lab
Inhibitory ATR kinasy v terapii nádorů
 Inhibitory ATR kinasy v terapii nádorů J.Vávrová, M Řezáčová Katedra radiobiologie FVZ Hradec Králové UO Brno Ústav lékařské chemie LF Hradec Králové UK Praha Cíl léčby: zničení nádorových buněk zachování
Inhibitory ATR kinasy v terapii nádorů J.Vávrová, M Řezáčová Katedra radiobiologie FVZ Hradec Králové UO Brno Ústav lékařské chemie LF Hradec Králové UK Praha Cíl léčby: zničení nádorových buněk zachování
Takahashi K & Yamanaka S. Cell 126, 2006,
 Obecné mechanismy buněčné signalizace Signalizace do buněčného jádra MUDr. Jan láteník, hd. Ústav lékařské biochemie 1.LF UK Somatické buňky lze přeprogramovat na pluripotentní kmenové buňky! řeprogramování
Obecné mechanismy buněčné signalizace Signalizace do buněčného jádra MUDr. Jan láteník, hd. Ústav lékařské biochemie 1.LF UK Somatické buňky lze přeprogramovat na pluripotentní kmenové buňky! řeprogramování
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU. Jana Novotná
 RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU Jana Novotná Co jsou to cytokiny? Skupina proteinů a peptidů (glykopeptidů( glykopeptidů), vylučovaných živočišnými buňkami a ovlivňujících buněčný růst (též růstové
RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU Jana Novotná Co jsou to cytokiny? Skupina proteinů a peptidů (glykopeptidů( glykopeptidů), vylučovaných živočišnými buňkami a ovlivňujících buněčný růst (též růstové
Intermediární metabolismus. Vladimíra Kvasnicová
 Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Intermediární metabolismus Vladimíra Kvasnicová Vztahy v intermediárním metabolismu (sacharidy, lipidy, proteiny) 1. po jídle (přísun energie z vnějšku) oxidace CO 2, H 2 O, urea + ATP tvorba zásob glykogen,
Respirace. (buněčné dýchání) O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3
 O 2. Fotosyntéza Dýchání. Energie záření teplo BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3") Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
Respirace (buněčné dýchání) Fotosyntéza Dýchání Energie záření teplo chem. energie CO 2 (ATP, NAD(P)H) O 2 Redukce za spotřeby NADPH BIOMASA CO 2 (-COO - ) = -COOH -CHO -CH 2 OH -CH 3 oxidace produkující
5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy
 Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy") SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
SFZR 1 2014 5) Fyziologie rostlinných hormonů auxinů: receptory a signální dráhy a) Auxinový receptor TIR1 b) Auxinový receptor ABP1 c) Kooperace receptorů TIR1 a ABP1 Estelle M et al. (2011) Auxin Signaling:
Jaderné receptory. ligand. cytoplazmatická membrána. jaderný receptor DNA. - ligandem aktivované transkripční faktory
 Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
Jaderné receptory Jaderné receptory - ligandem aktivované transkripční faktory - pokud není znám ligand ORPHAN receptors - ligand nalezen adopted orphan ligand DNA cytoplazmatická membrána jaderný receptor
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
Fyziologie AUTOFAGIE. MUDr. JAN VARADY KARIM FNO
 Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Fyziologie AUTOFAGIE MUDr. JAN VARADY KARIM FNO 29.1.2019 Autofagie?? Autofagie Self-eating Regulovaný katabolický jev Degradace a recyklace buněčných cytoplasmatických komponent: malfunkční a staré proteiny,
Intracelulární Ca 2+ signalizace
 Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
Studium genetické predispozice ke vzniku karcinomu prsu
 Univerzita Karlova v Praze 1. lékařská fakulta Studium genetické predispozice ke vzniku karcinomu prsu Petra Kleiblová Ústav biochemie a experimentální onkologie, 1. LF UK - skupina molekulární biologie
Univerzita Karlova v Praze 1. lékařská fakulta Studium genetické predispozice ke vzniku karcinomu prsu Petra Kleiblová Ústav biochemie a experimentální onkologie, 1. LF UK - skupina molekulární biologie
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
FYZIOLOGIE ROSTLIN. Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz
 FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
FYZIOLOGIE ROSTLIN Přednášející: Doc. Ing. Václav Hejnák, Ph.D. Tel.: 224382514 E-mail: hejnak @af.czu.cz Studijní literatura: Hejnák,V., Zámečníková,B., Zámečník, J., Hnilička, F.: Fyziologie rostlin.
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Výzkumný ústav veterinárního lékařství v Brně
 LIPIDY: FUNKCE, IZOLACE, SEPARACE, DETEKCE FOSFOLIPIDY chemické složení a funkce v buněčných membránách; metody stanovení fosfolipidů fosfolipázy - produkty reakcí (ceramid, DAG = 2nd messengers) a stanovení
LIPIDY: FUNKCE, IZOLACE, SEPARACE, DETEKCE FOSFOLIPIDY chemické složení a funkce v buněčných membránách; metody stanovení fosfolipidů fosfolipázy - produkty reakcí (ceramid, DAG = 2nd messengers) a stanovení
Toxické látky v potravinách s nebezpečím onkologické aktivace
 Toxické látky v potravinách s nebezpečím onkologické aktivace Doc. MUDr. Pavel Dlouhý, Ph.D. Ústav hygieny 3. LF UK, Praha Rizikové faktory pro vznik nádorů Obezita Nadměrný příjem tuků? Nadměrná konzumace
Toxické látky v potravinách s nebezpečím onkologické aktivace Doc. MUDr. Pavel Dlouhý, Ph.D. Ústav hygieny 3. LF UK, Praha Rizikové faktory pro vznik nádorů Obezita Nadměrný příjem tuků? Nadměrná konzumace
Přehled pedagogické činnosti - doc. RNDr. Tomáš Obšil, Ph.D.
 Přehled pedagogické činnosti - doc. RNDr. Tomáš Obšil, Ph.D. Pedagogická činnost Akademický rok 2003/2004 Přednáška: Biofyzikální chemie (C260P43) volitelná pro všechny obory rozsah: 2/1, Zk, letní semestr
Přehled pedagogické činnosti - doc. RNDr. Tomáš Obšil, Ph.D. Pedagogická činnost Akademický rok 2003/2004 Přednáška: Biofyzikální chemie (C260P43) volitelná pro všechny obory rozsah: 2/1, Zk, letní semestr
Biosyntéza a degradace proteinů. Bruno Sopko
 Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Biosyntéza a degradace proteinů Bruno Sopko Obsah Proteosyntéza Post-translační modifikace Degradace proteinů Proteosyntéza Tvorba aminoacyl-trna Iniciace Elongace Terminace Tvorba aminoacyl-trna Aminokyselina
Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR. Mgr. Silvie Dudová
 Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR Mgr. Silvie Dudová Centrum základního výzkumu pro monoklonální gamapatie a mnohočetný myelom, ILBIT LF MU Brno Laboratoř experimentální
Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR Mgr. Silvie Dudová Centrum základního výzkumu pro monoklonální gamapatie a mnohočetný myelom, ILBIT LF MU Brno Laboratoř experimentální
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
arise from animals and plant from
 Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Jediným způsobem jak může vzniknou nová buňka
Buněčný cyklus When a cell arises, there must be a previous cell, just as animals can only arise from animals and plant from plants. (Rudolf Wirchow, 1858) Jediným způsobem jak může vzniknou nová buňka
Laboratoř růstových regulátorů Miroslav Strnad. ové kultury. Olomouc. Univerzita Palackého & Ústav experimentální botaniky AV CR
 Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
Laboratoř růstových regulátorů Miroslav Strnad Tkáňov ové kultury Olomouc Univerzita Palackého & Ústav experimentální botaniky AV CR DEFINICE - růst a vývoj rostlinných buněk, pletiv a orgánů lze účinně
4) Reakce rostlin k abiotickému stresu
 Reakce rostlin k abiotickému stresu") 2010 4) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení 1 Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing
2010 4) Reakce rostlin k abiotickému stresu a) Vodní deficit b) Zasolení a osmotické přizpůsobení a jeho role v toleranci k suchu a zasolení 1 Jenks M et al. (2005) Plant Abiotic Stress. Blackwell Publishing
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
Elementy signálních drah. cíle protinádorové terapie
 Elementy signálních drah cíle protinádorové terapie Martin Pešta, Ondřej Topolčan Department of Internal Medicine II, Faculty of Medicine in Pilsen, Charles University in Prague, Czech Republic Cílená
Elementy signálních drah cíle protinádorové terapie Martin Pešta, Ondřej Topolčan Department of Internal Medicine II, Faculty of Medicine in Pilsen, Charles University in Prague, Czech Republic Cílená
Signalizace je vlastně
 Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Signalizace je vlastně komunikace a komunikace je základem života. Přenos signálu Rotlinné hormony a jejich signální dráhy Signální dráhy založené na regulované proteolýze Komponenty proteolytických degradačních
Příběh šestý: Co mají společného signální dráhy?
 Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Příběh šestý: Co mají společného signální dráhy? Paralelismus v recepci auxinů a jasmonátů SCF-komplex Receptor = F-box protein Struktura SCF komplexu SCF komplex = RING-BOX1 + Skp1 + Cullin + F-box protein
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
1 Biochemické animace na internetu
 1 Biochemické animace na internetu V dnešní době patří internet mezi nejužívanější zdroje informací. Velmi často lze pomocí internetu legálně stáhnout řadu již vytvořených výukových materiálů sloužících
1 Biochemické animace na internetu V dnešní době patří internet mezi nejužívanější zdroje informací. Velmi často lze pomocí internetu legálně stáhnout řadu již vytvořených výukových materiálů sloužících
Světlo: vliv na časový systém, pozornost a náladu. Helena Illnerová Fyziologický ústav AV ČR, v. v. i.
 Světlo: vliv na časový systém, pozornost a náladu Helena Illnerová Fyziologický ústav AV ČR, v. v. i. CIRKADIANNÍ RYTMY ve spánku bdění v tělesné teplotě v chování v příjmu pití a potravy v tvorbě hormonů
Světlo: vliv na časový systém, pozornost a náladu Helena Illnerová Fyziologický ústav AV ČR, v. v. i. CIRKADIANNÍ RYTMY ve spánku bdění v tělesné teplotě v chování v příjmu pití a potravy v tvorbě hormonů
Vápník. Deficience vápníku: - 0,4-1,5% DW. - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM)
") Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
Vápník - 0,4-1,5% DW - cytoplasmatická koncentrace vápníku velmi nízká (0,1-0,2µM) - stavební, signální funkce, stabilizace membrán - vápnomilné x vápnostřežné druhy Deficience vápníku: - poškození meristemů,
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění