5. Sekvenování, přečtení genetické informace, éra genomiky.
|
|
|
- Silvie Vávrová
- před 9 lety
- Počet zobrazení:
Transkript
1 5. Sekvenování, přečtení genetické informace, éra genomiky. Minulá přednáška nastínila zrod molekulární biologie a představila některé možnosti, jak pracovat s DNA - jak ji analyzovat na základě velikosti pomocí elektroforézy, jak najít nějaký známý úsek DNA pomocí hybridizace se značenou sondou, jak zaklonovat kus DNA a vytvořit tak rekombinantní DNA (základ genového inženýrství) a také jak rychle namnožit vybraný úsek DNA do miliónu kopií pomocí PCR. Zde si představíme metody, které nám umožňují přímo DNA číst, písmenko po písmenku, nukleotid po nukleotidu. Jak minulá přednáška ukázala, DNA řetězec je příliš malinký na to, abychom i nejcitlivějšími mikroskopy byli schopni přímo vidět jednotlivé nukleotidy. Například snímek vpravo ukazuje asi maximum toho, co jsme schopni pomocí elektronového mikroskopu z řetězce DNA vidět - jednotlivá písmenka (nukleotidy) v něm rozhodně nerozeznáme. Proto je nutné použít různé molekulárně biologické triky a nepřímé metody, které nám jsou schopné sekvenci DNA určit. S první takovou metodou, která byla a je dodnes rozumně aplikovatelná, přišel v roce 1977 Frederick Sanger, který za ní v roce 1980 získal Nobelovu cenu, a podle kterého je tato metoda pojmenována jako Sangerova metoda sekvenování DNA: F. SANGER, S. NICKLEN, AND A. R. COULSON: DNA sequencing with chain-terminating inhibitors. PNAS 74(12), , Tato metoda využívá stejného principu jako PCR, kdy je templátová DNA množena pomocí primerů a polymerázy. V případě sekvenační reakce se ovšem použije pouze jeden primer a tudiž dochází k syntéze jen jednoho řetězce v jednom směru. Dalším specifikem sekvenační reakce je (v její originální podobě, dnes je to jinak), že se dělá ve čtyřech zkumavkách, které všechny mají totožné složky reakce (dntp jako stavební bloky, templát, primer a polymerázu), ale v každé je pak navíc jeden tzv. dideoxynukleotid (ddatp v první zkumavce, ddttp v druhé, ddgtp ve třetí a ddctp ve čtvrté). Tento nukleotid je malinko pozměněný, chybí mu OH skupina na 3 uhlíku:

2 To způsobí, že po začlenění takovéhoto nukleotidu do nově syntetizovaného řetězce již nemůže být připojen další nukleotid (který vyžaduje pro vazbu právě OH skupinu v tomto místě) a tudíž se za tímto nukleotidem syntéza nového řetězce zastaví. Protože v každé reakci jsou namíchány vždy všechny čtyři normální nukleotidy plus je tam přimíchán navíc tento dideoxynukleotid, je vždy věcí náhody, kdy se tento dideoxynukleotid vmezeří do syntézy a kde se tedy nový řetězec ukončí, ale vždy je to na pozici, kde by v normální sekvenci byla normální forma nukleotidu (tedy pokud je v reakci přítomen ddgtp, tak nový řetězec vždy bude končit v místě, kde se normálně nachází báze G, resp. kde se v templátu nachází komplementární C): Jak jsme viděli v minulé přednášce, pomocí gelové elektroforézy můžeme rozdělit fragmenty DNA podle velikosti, tj. pokud pustíme na gel vedle sebe tyto čtyři reakce, jsme podle délky jednotlivých fragmentů schopni přečíst pořadí nukleotidů v sekvenci (je třeba mít velmi citlivou elektroforézu, která dokáže rozdělit od sebe dva fragmenty, které se liší pouze o jediný nukleotid):

3 Dnes již existují dokonalejší metody, které ovšem využívají stejného principu, ale jednotlivé reakce jsou značeny čtyřmi různými fluorescenčními značkami. Tím je možné původně čtyři reakce sloučit do jedné zkumavky a též je analyzovat v jednom proužku gelu, dnes na tzv. kapilární elektroforéze, kdy fragmenty se opět na základě velikosti rozdělí tím, jak putují kapilárou, a na konci kapiláry je detektor schopný zaznamenat barvu právě projíždějícího fragmentu: Výsledkem sekvenační reakce, analyzované na moderním sekvenátoru pomocí kapilární elektroforézy a laserové detekce fluorescenčně značených fragmentů je chromatogram s "peaky" jednotlivých fragmentů tak, jak projížděly detektorem. Pořadí těchto "peaků" pak koresponduje se sekvencí DNA:

4 Tímto způsobem se dnes sekvenuje, tj. čte sekvence DNA, v naprosté většině případů, kdy potřebujeme přečíst sekvenci jednoho konkrétního fragmentu DNA. Touto metodou byl také kompletně přečten celý lidský genom (viz. dále), akorát bylo zapotřebí provést milióny takových sekvenačních reakcí! Dnes již existují dokonalejší metody, které umožňují přečíst sekvenci miliónů fragmentů DNA najednou (tzv. Next Generation Sequencing - viz dále). Jak najít gen, resp. mutaci v něm, způsobující lidské onemocnění? Pamatujete na Mendela? Je třeba vystopovat, jak se určitý znak (v tomto případě onemocnění) dědí z generace na generaci. Proto je třeba mít k dispozici poměrně rozsáhlý rodokmen, resp. dostatečný počet pacientů pokud možno s co největším počtem příbuzných, kteří se mohou zúčastnit analýzy. To dělá hledání lidských genů, spojených s určitou chorobou, takovým problémem bohužel není možné lidské pacienty křížit dle potřeby, jako to můžeme udělat s modelovým organismem (myš, octomilka). Pak je potřeba velmi rozsáhlé a pečlivé práce za použití DNA fingerprintingu zkoušet všechny možné variabilní markery (sekvence DNA, jejichž pozici v genomu známe a které vykazují rozdíly, když se porovnávají mezi jedinci), které v DNA máme k dispozici a pokusit se vystopovat, se kterým markerem se náš neznámý hledaný gen spolu dědí (pamatujete na míchání genů pomocí rekombinací?). Když se tohle podaří, tak budeme vědět, že někde v blízkosti (bavíme se ca. o stotisících až milionech bazí) této značky se ten gen nachází. Pak je potřeba pomocí postupného sekvenování projít (chromosome walking) takovýto úsek chromozomu pomocí hybridizace najdeme bakteriální klon s přilehlým úsekem, čímž se dostaneme zase o kousek dále, a takto můžeme prosekvenovat celý úsek a hledat kýženou mutaci:
 dědí z generace na generaci.")
5 Pokud srovnáme takto získanou sekvenci mezi mnoha jedinci s chorobou a mnoha jedinci bez ní, máme šanci najít záměnu, kterou budou mít všichni nemocní, ale ne zdraví jedinci. Po mnoha dalších ověření je pak možné zjistit, jestli tato záměna je skutečně příčinou choroby. Takhle nějak se postupovalo při hledání mutací způsobujících dědičná onemocnění před nástupem moderních metod, zejména Next Generation Sequencing, které toto výrazně zjednodušily (dnes už se neprocházíme po neznámých úsecích chromozomů, ale rovnou porovnáváme celé genomy postižených a zdravých jedinců). Ale nepředbíhejme a podívejme se na historii přečtení kompletní genetické informace člověka. Po té, co se vědci několikrát prošli po neznámých úsecích chromozomů, si uvědomili, že by takto mohli osekvenovat celý lidský genom, řekli si něco ve smyslu: Co takhle projít se po celém genomu? A tak se v roce 1990 zrodil Human Genome Project (HUGO), který si dal za cíl přečíst kompletní genetickou informaci člověka do 15 let. Mnoho laboratoří po světě si rozdělilo lidský genom na menší části na základě hrubé mapy lidského genomu. Kousky sekvencí z nejrůznějších částí genomu byly namapovány (metodou hybridizace - FISH) na jednotlivé chromozomy a od těchto známých kousků se vždy začínalo. Lidský genom byl rozsekán na menší úseky (několik desítek až stovek tisíců bazí) a z těchto úseků byla vytvořena genomová knihovna v bakteriích, které nesly tzv. BAC klony (Bacterial Artificial Chromosome). Opět metodou hybridizace byly identifikovány klony, které v sobě nesly ony známé namapované kousky chromozomů. Po identifikaci takového klonu měli vědci v rukou již relativně malý kus lidského genomu, o kterém věděli, na jakém chromozomu a kde se nachází. Tento klon pak mohli znovu rozkouskovat ještě na menší kousky a metodou Chromosome Walking mohli celý tento kus postupně prosekvenovat. Později přišli s ještě rychlejším a efektivnějším řešením, jak tyto kusy lidského genomu v BAC klonech prosekvenovat - metodou Random Shotgun. Při něm BAC klon rozkouskovali na relativně krátké fragmenty a všechny naslepo začali
.")
6 sekvenovat. Doufali, že když jich prosekvenují hodně, tak z nich na základě překryvů zpětně poskládají celý úsek klonu - a to se většinou dařilo. Protože celé HUGO konzorcium postupovalo takto hierarchicky, tj. rozdělili si velké úseky lidského genomu a tyto velké úseky si pak rozdělili na menší a ty sekvenovali, ale vždy věděli, s kterou oblastí genomu pracují, mluví se o tzv. HIERARCHICAL SHOTGUN přístupu. V roce 1998 přišel velký vizionář, úspěšný badatel a byznysman v jedné osobě - Craig Venter - s nápadem využít metodu Random Shotgun na celý lidský genom najednou, tj. netvořit BAC klony, které budou namapovány na jednotlivé chromozomy, ale prostě vzít celou lidskou DNA, rozbít ji na kousky a naslepo sekvenovat, osekvenovat milióny sekvencí a doufat, že z nich půjde díky překryvům zpětně poskládat celý

7 lidský genom. Za tímto účelem nakoupila firma Celera Genomics stovky sekvenátorů, které ve dne v noci skoro rok chrlily jednu sekvenci za druhou. Nakonec to bylo 27 milionů sekvencí o průměrné délce 540 bazí (tj sekvenačních reakcí denně). Tímto množstvím osekvenovali 5x více nukleotidů, než obsahuje lidský genom, což jim umožnilo poskládat 90 % lidského genomu (přesněji řečeno jeho euchromatinové části, tj. aktivní DNA ve které je naprostá většina genů) - přečtení 5x více sekvencí zajistilo, že zbylo jen málo míst, do kterých se při náhodném čtení fragmentů netrefili ani jednou a že většina fragmentů zároveň tvořila překryvy: Projekt firmy Celera Genomics donutil i konsorcium HUGO urychlit svou práci a tak nakonec v roce 2001 byl oficiálně osekvenován lidský genom (v praxi to znamenalo, že bylo přečteno a poskládáno ca. 90% euchromatinové DNA), přičemž HUGO konsorcium publikovalo svou verzi v Nature a Celera Genomics v Science. Od té doby byly postupně dokončeny i problematická místa a dnes je více než 99% lidské euchromatinové DNA poskládáno - v ENSEMBL Genome Browser se můžete prouzdat kompletním lidským genomem. Člověk má mnohem méně genů, než jsme předpokládali Velkým překvapením byl počet genů v lidském genomu, který byl po prvním poskládání v roce 2001 odhadnut na ca , namísto odhadovaných ! Ve srovnání s mnohem jednodušším organismem, jakým je moucha octomilka, která má genů, máme ani ne dvakrát tolik. Odpověď na to, kde je v DNA schována výrazně větší složitost organismů, jako je člověk, naznačuje tabulka níže. Počet protein-kódujících lokusů není výrazně větší u mnohem složitějšího organismu, ale je třeba si uvědomit, že velmi často z jednoho lokusu (genu) vzniká více variant transkriptů (alternativním sestřihem) a tím i více různých proteinů. Tj. neplatí vždy 1 gen = 1 protein, a u složitějších organismů je zpravidla více různých variant transkriptů a proteinů z jednoho lokusu, než u jednodušších (viz. tabulka).
 - přečtení 5x více sekvencí zajistilo, že zbylo jen málo míst, do kterých se při náhodném čtení fragmentů netrefili ani jednou a že většina fragmentů")
8 člověk octomilka známé protein-kódující geny pseudogeny RNA geny exony transkripty Toto zjištění ještě více ztěžuje definování genu, jednoduchá definice není - je genem lokus na chromozomu, který kóduje nějaký protein? Co když takový lokus kóduje více variant proteinů (a ty varianty mohou mít někdy i docela různé funkce). Měl by být genem nazýván každý jednotlivý transkript? Ale co když všechny transkripty z jednoho lokusu sdílejí většinu své sekvence a liší se jen mírně a je i těžké rozlišit funkce jednotlivých variant proteinů? Zůstaňme tedy u toho, že genem budeme nazývat místo na chromozomu (lokus), který kóduje nějaký protein nebo více variant podobných proteinů, které vznikají alternativním sestřihem transkriptů daného lokusu (genem mohou být nazývány i lokusy kódující pouze RNA, například ribozomální RNA). Na obrázku dole je například lokus CG42249 (modře), chcete-li gen, v genomu octomilky, který kóduje 2 alternativní transkripty RC a RB (exony naznačeny žlutými obdélníčky), podle nichž vznikají 2 různé varianty proteinu, které sdílejí svou první polovinu, ale liší se tou druhou polovinou (varianta RC vzniká vyštěpením posledních 4 exonů varianty RB a napojením dalších exonů za nimi): NEXT GENERATION SEQUENCING - sekvenování nové generace V letech následujících po dokončení lidského genomu, který byl přečten za pomocí klasické metody sekvenování dle Sangera (viz. výše), se objevily zcela nové technologie, které posunuly sekvenování do úplně jiné dimenze - často jsou nazývány Next Generation Sequencing (zkráceně též NextGen). Pro ilustraci síly těchto nových metod můžeme použít příklad: na osekvenování celého lidského genomu potřebovaly desítky laboratoří po celém světě více než 11 let, Celera Genomics se stovkami přístrojů a plně automatizovaným provozem potřebovala 2 roky, dnes je to samé možné udělat na jediném přístroji (který se vejde na obyčejný stůl) při obsluze jedním člověkem ani ne za 3 týdny! Principem, který umožňuje během jediné sekvenační reakce přečíst až stovky miliónů bazí, je ukotvení miliónů sekvenačních reakcí na čip (ne větší než několik cm) a simultánní detekce každé jednotlivé reakce pomocí velmi citlivých metod s obrovským rozlišením (zachycení miliónů signálů na ploše několikacentrimetrového čipu). Na obrázku níže je příklad jednoho z přístupů, který zvolila firma Illumina - nejprve uchytí na skleněnou destičku - čip - milióny krátkých fragmentů DNA, kterou chce osekvenovat. Uchycení a tudíž rozmístění jednotlivých fragmentů je zcela náhodné. Každý fragment je na svém místě namnožen, což

9 zvýší signál pro detekci, a poté je čip promýván fluorescenčně značenými nukleotidy, které jsou chemicky modifikovány tak, že se na komplementární DNA fragmentu naváže vždy jen jeden nukleotid (syntéza nepokračuje, dokud v dalším kroku nedojde k modifikaci, umožňující navázání dalšího nukleotidu). Po navázání nukleotidů citlivý detektor sejme obrázek jako je ten dole (malinký výřez, ve skutečnosti jsou tam milióny takových bodů), takže počítač ví, jaký konkrétní nukleotid ze 4 možných byl v kterém konkrétním místě v 1. kroku navázán. Pak pokračuje syntéza a po každém kroku je sejmut obraz. Počítač pak z těchto obrazů poskládá sekvenci každého jednotlivého fragmentu v daném místě, tudíž získá najednou sekvence miliónů fragmentu z jednoho malého čipu. Na základě překryvů (DNA je nafragmentována náhodně na malé kousky) ji pak poskládá do víceméně souvislé DNA (pokud původně šlo o souvislou DNA). Popis různých přístupů Next Generation Sequencing v Nature Methods. Éra genomiky Možnosti přečíst a analyzovat celé genomy nás přenesly do éry genomiky. Genomický přístup v bádání znamená, že se badatel nezabývá jen jedním nebo několika málo geny najednou, ale analyzuje nějaký biologický problém na celogenomové úrovni. Můžete se například ptát, jak se liší určitý gen, zodpovědný za nějakou vlastnost, mezi člověkem a jeho nejbližším příbuzným šimpanzem - to by byl přístup klasické genetiky. Dnes už ovšem známe celý genom jak člověka, tak šimpanze a proto můžeme porovnat, jak se liší kompletní genetická informace těchto dvou příbuzných druhů, ptát se například, kde v DNA je zakódováno to, co nás dělá lidmi? Nebo při hledání mutace, která nás předurčuje k určitému typu rakoviny, již nemusíte postupovat kousek po kousku, gen po genu, ale můžete srovnat rovnou celé genomy pacientů trpících touto formou rakoviny a hledat v nich, jakou záměnu oproti většině lidí mají všichni tito pacienti společnou. Možná to nebude jedna záměna, ale více, roztroušené po celém genomu a právě jejich kombinace nás bude činit náchylnějšími - bez možnosti srovnávat celé genomy najednou je nepředstavitelné tyto komplexní záležitosti odhalit. Při nalezení nějakého organismu s výjimečnými vlastnostmi můžeme prozkoumat celý jeho genom a snáze zjistit, co ho dělá tak výjimečným. A nemusíme skončit jen na úrovni DNA a číst celé genomy. Podobně můžeme analyzovat i RNA a ptát se, které všechny geny v daném organismu či jen určité tkáni jsou za daných podmínek aktivní, tj. se přepisují do mrna? Pak mluvíme o tzv. transkriptomice, protože namísto jednoho konkrétního transkriptu analyzujeme kompletní sadu všech RNA, které se za daných podmínek přepisují, tj. analyzujeme kompletní transkriptom. Toto bylo možné již dříve pomocí microarrays - čipů, které detekovaly celé sady RNA, ale pouze u vybraných organismů, u kterých byl transkriptom znám a někdo si dal práci připravit čipy pro vybrané transkripty. Dnes je ovšem možné díky Next Generation Sequencing, resp. jeho konkrétní aplikaci zvané RNAseq, přečíst transkriptom i organismů, o nichž nic nevíme, prostě osekvenujeme veškeré RNA přítomné v daném vzorku.

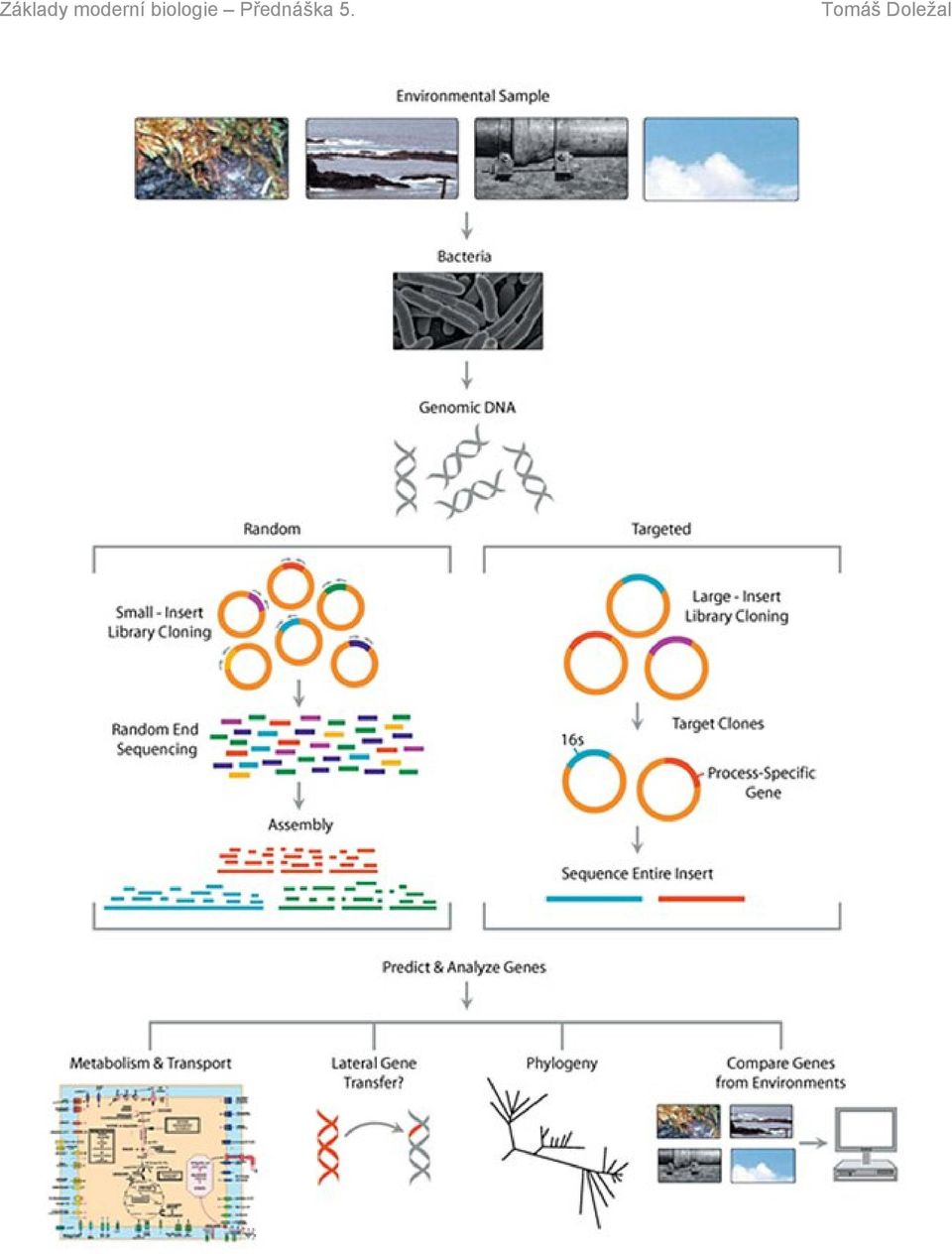
10 Je třeba zdůraznit, že se nesekvenuje přímo RNA, ale RNA převedená reverzní transkripcí na DNA. Reverzní transkripce je jedna z komplikací původního centrálního dogmatu, které popisovalo tok genetické informace pouze ve směru DNA RNA protein. Ovšem některé viry jsou schopné přepsat svou RNA zpět do DNA pomocí enzymu reverzní transkriptáza, který je využíván právě při čtení transkriptů a transkriptomů. A nakonec nemusíme skončit pouhou genomikou, tj. analýzou celého genomu jednoho organismu. Můžeme číst genomy více organismů najednou, nebo dokonce genomy všech organismů vyskytujících se v daném vzorku, například všech mikroorganismů v určité půdě za daných podmínek. Pak hovoříme o tzv. METAGENOMICE. METAGENOMIKA = genomika společenstev z prostředí - analýza DNA mikroorganismů izolovaných z prostředí, které se nedají snadno kultivovat, pomocí nové generace sekvenování METATRANSKRIPTOMIKA = analýza aktivity všech genů v prostředí i bez jejich předchozí znalosti, můžeme porovnávat aktivity neznámých genů v různých prostředích a izolovat tak nové geny a jejich funkce V tomto duchu se rozhodl i Craig Venter, po té co byl nucen opustit Celera Genomics a založil si Institut Craiga Ventera, prozkoumat mikroorganismy této planety. Vybavil se luxusní jachtou Sorcerer II a začal brázdit světové oceány a pomocí filtrů sbírat veškeré mikroorganismy, které tam zachytí. Jejich DNA pak zasílal do svého institutu, kde je sekvenují (metagenomika v praxi). Například expedice do Sargasového moře odhalila více než 1800 nových organismů a popsala více než 1,2 miliónu nových genů. Též detekovali, že v tomto mikrobiálním světě dochází k intenzivnímu horizontálnímu přenosu genů. Craig Venter se tak po 170 letech vrátil k Darwinově cestě na H.M.S. Beagle, tentokrát ovšem na molekulární úrovni.

11
12

6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
Výzkumné centrum genomiky a proteomiky. Ústav experimentální medicíny AV ČR, v.v.i.
 Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Výzkumné centrum genomiky a proteomiky Ústav experimentální medicíny AV ČR, v.v.i. Systém pro sekvenování Systém pro čipovou analýzu Systém pro proteinovou analýzu Automatický sběrač buněk Systém pro sekvenování
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Poziční klonování Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s metodou pozičního klonování genů
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Sekvenování genomů Ing. Hana Šimková, CSc. Cíl přednášky - seznámení se strategiemi celogenomového sekvenování,
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů
 Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?
Využití metagenomiky při hodnocení sanace chlorovaných ethylenů in situ Výsledky pilotních testů Stavělová M.,* Macháčková J.*, Rídl J.,** Pačes J.** * Earth Tech CZ, s.r.o ** ÚMG AV ČR PROČ METAGENOMIKA?
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Sekvenování nové generace. Radka Reifová
 Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Sekvenování nové generace Radka Reifová Prezentace ke stažení www.natur.cuni.cz/zoologie/biodiversity v záložce Přednášky 1. Přehled sekvenačních metod nové generace 2. Využití sekvenačních metod nové
Genetika zvířat - MENDELU
 Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Genetika zvířat DNA - primární struktura Několik experimentů ve 40. a 50. letech 20. století poskytla důkaz, že genetický materiál je tvořen jedním ze dvou typů nukleových kyselin: DNA nebo RNA. DNA je
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Sekvenování příští generace (Next Generation Sequencing, NGS)
") Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
Sekvenování příští generace (Next Generation Sequencing, NGS) Přednáška 6, 2013/14 Ivo Papoušek Next generation sequencing poptávka po nízkonákladovém sekvenování vyvolala tlak na vývoj high-throughput
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Analýza transkriptomu Ing. Hana Šimková, CSc. Cíl přednášky - seznámení s moderními metodami komplexní
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Věda v prostoru. Voda v pohybu. Buněční detektivové. Svědkové dávné minulosti Země
 6+ Věda v prostoru Jak vědci pracují v laboratoři? Proč je zelená víc než jen obyčejná barva? Jak můžeme použít prášek do pečiva ke sfouknutí svíčky? Získejte odpovědi na všechny otázky v tomto vzrušujícím
6+ Věda v prostoru Jak vědci pracují v laboratoři? Proč je zelená víc než jen obyčejná barva? Jak můžeme použít prášek do pečiva ke sfouknutí svíčky? Získejte odpovědi na všechny otázky v tomto vzrušujícím
Molekulární biotechnologie č.9. Cílená mutageneze a proteinové inženýrství
 Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
4. Centrální dogma, rozluštění genetického kódu a zrod molekulární biologie.
 4. Centrální dogma, rozluštění genetického kódu a zrod molekulární biologie. Od genu k proteinu - centrální dogma biologie Geny jsou zakódovány v DNA - Jakým způsobem? - Jak se projevují? Již v roce 1902
4. Centrální dogma, rozluštění genetického kódu a zrod molekulární biologie. Od genu k proteinu - centrální dogma biologie Geny jsou zakódovány v DNA - Jakým způsobem? - Jak se projevují? Již v roce 1902
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno
 Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Retinoblastom Mgr. Veronika Peňásová vpenasova@fnbrno.cz Laboratoř molekulární diagnostiky, OLG FN Brno Klinika dětské onkologie, FN Brno Retinoblastom (RBL) zhoubný nádor oka, pocházející z primitivních
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod
 Molekulární přístupy ve šlechtění rostlin Olomouc 14. února, 2017 Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod Jan Bartoš Ústav experimentální botaniky Olomouc, Czech Republic
Molekulární přístupy ve šlechtění rostlin Olomouc 14. února, 2017 Charakterizace hybridních trav pomocí cytogenetických a molekulárních metod Jan Bartoš Ústav experimentální botaniky Olomouc, Czech Republic
Molekulární genetika
 Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Molekulární genetika Genetické inženýrství Technologie rekombinantní DNA Vektor Genomová DNA Štěpení RE Rozštěpení stejnou RE, lepivé konce Ligace Transformace Bakteriální chromozóm Rekombinantní vektor
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ. I. Šubrt Společnost lékařské genetiky ČLS JEP
 EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
EKONOMICKÉ ASPEKTY GENETICKÝCH VYŠETŘENÍ I. Šubrt Společnost lékařské genetiky ČLS JEP Lékařská genetika Lékařský obor zabývající se diagnostikou a managementem dědičných onemocnění Genetická prevence
Biologie - Oktáva, 4. ročník (přírodovědná větev)
") - Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
- Oktáva, 4. ročník (přírodovědná větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k
Biologie - Oktáva, 4. ročník (humanitní větev)
") - Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
- Oktáva, 4. ročník (humanitní větev) Biologie Výchovné a vzdělávací strategie Kompetence k řešení problémů Kompetence komunikativní Kompetence sociální a personální Kompetence občanská Kompetence k podnikavosti
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK
 ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
ové technologie v analýze D A, R A a proteinů Stanislav Kmoch Centrum aplikované genomiky, Ústav dědičných metabolických poruch, 1.LFUK Motto : "The optimal health results from ensuring that the right
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Co se o sobě dovídáme z naší genetické informace
 Genomika a bioinformatika Co se o sobě dovídáme z naší genetické informace Jan Pačes, Mgr, Ph.D Ústav molekulární genetiky AVČR, CZECH FOBIA (Free and Open Bioinformatics Association) hpaces@img.cas.cz
Genomika a bioinformatika Co se o sobě dovídáme z naší genetické informace Jan Pačes, Mgr, Ph.D Ústav molekulární genetiky AVČR, CZECH FOBIA (Free and Open Bioinformatics Association) hpaces@img.cas.cz
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Mikročipy v mikrobiologii
 Mikročipy v mikrobiologii doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 Obsah přednášky 1) Charakteristika biočipů, DNA microarrays a DNA chip 2) Výroba čipů, charakteristika
Mikročipy v mikrobiologii doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 Obsah přednášky 1) Charakteristika biočipů, DNA microarrays a DNA chip 2) Výroba čipů, charakteristika
Využití rekombinantní DNA při studiu mikroorganismů
 Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Využití rekombinantní DNA při studiu mikroorganismů doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 1 2 Obsah přednášky 1) Celogenomové metody sekvenování 2) Sekvenování H.
Ondřej Scheinost Nemocnice České Budějovice, a.s.
 Ondřej Scheinost Nemocnice České Budějovice, a.s. Nové technologie přelomové období principy technologií klinická použitelnost chips (arrays) sekvenační technologie Důležitost genetických informací i další
Ondřej Scheinost Nemocnice České Budějovice, a.s. Nové technologie přelomové období principy technologií klinická použitelnost chips (arrays) sekvenační technologie Důležitost genetických informací i další
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Sekvence. Genom. Základní informace. Výstupy z výukové jednotky
 Sekvence Základní informace Následující text je součástí učebních textů předmětu Analýza sekvencí DNA a je určen hlavně pro studenty Matematické biologie. Může být ovšem přínosný i pro další studenty biologických
Sekvence Základní informace Následující text je součástí učebních textů předmětu Analýza sekvencí DNA a je určen hlavně pro studenty Matematické biologie. Může být ovšem přínosný i pro další studenty biologických
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Molekulárně biologické metody princip, popis, výstupy
 & Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
& Molekulárně biologické metody princip, popis, výstupy Klára Labská Evropský program pro mikrobiologii ve veřejném zdravotnictví (EUPHEM), ECDC, Stockholm NRL pro herpetické viry,centrum epidemiologie
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
PŘEHLED SEKVENAČNÍCH METOD
 PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
PŘEHLED SEKVENAČNÍCH METOD Letní škola bioinformatiky 2014, Brno Ing.Matej Lexa, Phd (FI MU Brno) CO JE TO SEKVENACE A CO SE BUDE SEKVENOVAT? POŘADÍ NUKLEOTIDU V DNA SEKVENOVÁNÍ DNA od manuálních metod
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 17. - 21. 6. 2013 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Struktura a analýza rostlinných genomů Jan Šafář
 Struktura a analýza rostlinných genomů Jan Šafář Ústav experimentální botaniky AV ČR, v.v.i Centrum regionu Haná pro biotechnologický a zemědělský výzkum Proč rostliny? Proč genom? Norman E. Borlaug Zelená
Struktura a analýza rostlinných genomů Jan Šafář Ústav experimentální botaniky AV ČR, v.v.i Centrum regionu Haná pro biotechnologický a zemědělský výzkum Proč rostliny? Proč genom? Norman E. Borlaug Zelená
Ústav experimentální medicíny AV ČR úspěšně rozšířil přístrojové vybavení pro vědce z peněz evropských fondů
 Ústav experimentální medicíny AV ČR úspěšně rozšířil přístrojové vybavení pro vědce z peněz evropských fondů Ústav úspěšně dokončil realizaci dvou investičních projektů s využitím prostředků z Operačního
Ústav experimentální medicíny AV ČR úspěšně rozšířil přístrojové vybavení pro vědce z peněz evropských fondů Ústav úspěšně dokončil realizaci dvou investičních projektů s využitím prostředků z Operačního
Analýza DNA. Co zjišťujeme u DNA
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů, záměny), chromosomové aberace (numerické, strukturní) Polymorfismy konkrétní
Osekvenované genomy. Pan troglodydes, 2005. Neandrtálec, 2010
 GENOMOVÉ PROJEKTY Osekvenované genomy Haemophilus influenze, 1995 první osekvenovaná bakterie Saccharomyces cerevisiae, 1996 první osekvenovaný eukaryotický organimus Caenorhabditis elegans, 1998 první
GENOMOVÉ PROJEKTY Osekvenované genomy Haemophilus influenze, 1995 první osekvenovaná bakterie Saccharomyces cerevisiae, 1996 první osekvenovaný eukaryotický organimus Caenorhabditis elegans, 1998 první
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION
 USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
USING OF AUTOMATED DNA SEQUENCING FOR PORCINE CANDIDATE GENES POLYMORFISMS DETECTION VYUŽITÍ AUTOMATICKÉHO SEKVENOVÁNÍ DNA PRO DETEKCI POLYMORFISMŮ KANDIDÁTNÍCH GENŮ U PRASAT Vykoukalová Z., Knoll A.,
APLIKACE METAGENOMIKY PRO HODNOCENÍ PRŮBĚHU SANAČNÍHO ZÁSAHU NA LOKALITÁCH KONTAMINOVANÝCH CHLOROVANÝMI ETHYLÉNY
 APLIKACE METAGENOMIKY PRO HODNOCENÍ PRŮBĚHU SANAČNÍHO ZÁSAHU NA LOKALITÁCH KONTAMINOVANÝCH CHLOROVANÝMI ETHYLÉNY Monika Stavělová 1, Jakub Rídl 2, Maria Brennerová 3, Hana Kosinová 1, Jan Pačes 2 1 AECOM
APLIKACE METAGENOMIKY PRO HODNOCENÍ PRŮBĚHU SANAČNÍHO ZÁSAHU NA LOKALITÁCH KONTAMINOVANÝCH CHLOROVANÝMI ETHYLÉNY Monika Stavělová 1, Jakub Rídl 2, Maria Brennerová 3, Hana Kosinová 1, Jan Pačes 2 1 AECOM
1. Definice a historie oboru molekulární medicína. 3. Základní laboratorní techniky v molekulární medicíně
 Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
Obsah Předmluvy 1. Definice a historie oboru molekulární medicína 1.1. Historie molekulární medicíny 2. Základní principy molekulární biologie 2.1. Historie molekulární biologie 2.2. DNA a chromozomy 2.3.
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII. Martina Nováková, VŠCHT Praha
 MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNĚ BIOLOGICKÉ METODY V ENVIRONMENTÁLNÍ MIKROBIOLOGII Martina Nováková, VŠCHT Praha MOLEKULÁRNÍ BIOLOGIE V BIOREMEDIACÍCH enumerace FISH průtoková cytometrie klonování produktů PCR sekvenování
MOLEKULÁRNÍ BIOLOGIE. 2. Polymerázová řetězová reakce (PCR)
") MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
MOLEKULÁRNÍ BIOLOGIE 2. Polymerázová řetězová reakce (PCR) Náplň praktik 1. Izolace DNA z buněk bukální sliznice - izolační kit MACHEREY-NAGEL 2. PCR polymerázová řetězová reakce (templát gdna) 3. Restrikční
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014. Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU
 V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
V. letní škola metod molekulární biologie nukleových kyselin a genomiky 16. - 20. 6. 2014 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU Zemědělská 1, Budova A, 4. patro (učebny dle programu)
Možné účinky XENOBIOTIK
 Možné účinky XENOBIOTIK přímý toxický účinek -látka působí pouhou svou přítomností na kritickém místě v organismu biochemický účinek - látka interaguje s cílovou molekulou (receptorem), ovlivní nějaký
Možné účinky XENOBIOTIK přímý toxický účinek -látka působí pouhou svou přítomností na kritickém místě v organismu biochemický účinek - látka interaguje s cílovou molekulou (receptorem), ovlivní nějaký
EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva
 EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva Iva Buriánková Katedra ekologie PřF UP KLONOVÁNÍ GENŮ KLONOVÁNÍ GENŮ Klonování
EKO/MEM - Molekulární ekologie mikroorganizmů Klonování a sekvenování přírodní DNA základ pro fylogenetickou analýzu společenstva Iva Buriánková Katedra ekologie PřF UP KLONOVÁNÍ GENŮ KLONOVÁNÍ GENŮ Klonování
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Biotechnologický kurz. III. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
Biotechnologický kurz Biotechnologický kurz III. letní škola metod molekulární biologie nukleových kyselin a genomiky 18. - 22. 6. 2012 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně
b) Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?
 Jak se změní sekvence aminokyselin v polypeptidu, pokud dojde v pozici 23 k záměně bázového páru GC za TA (bodová mutace) a s jakými následky?") 1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
1.1: Gén pro polypeptid, který je součástí peroxidázy buku lesního, má sekvenci 3'...TTTACAGTCCATTCGACTTAGGGGCTAAGGTACCTGGAGCCCACGTTTGGGTCATCCAG...5' 5'...AAATGTCAGGTAAGCTGAATCCCCGATTCCATGGACCTCGGGTGCAAACCCAGTAGGTC...3'
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE
 TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
TEST: GENETIKA, MOLEKULÁRNÍ BIOLOGIE 1) Důležitým biogenním prvkem, obsaženým v nukleových kyselinách nebo ATP a nezbytným při tvorbě plodů je a) draslík b) dusík c) vápník d) fosfor 2) Sousedící nukleotidy
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Analýza DNA. Co zjišťujeme u DNA DNA. PCR polymerase chain reaction. Princip PCR PRINCIP METODY PCR
 o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
o zjišťujeme u DN nalýza DN enetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní mutace,
GENETIKA dědičností heredita proměnlivostí variabilitu Dědičnost - heredita podobnými znaky genetickou informací Proměnlivost - variabilita
 GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
GENETIKA - věda zabývající se dědičností (heredita) a proměnlivostí (variabilitu ) živých soustav - sleduje rozdílnost a přenos dědičných znaků mezi rodiči a potomky Dědičnost - heredita - schopnost organismu
Biotechnologický kurz. II. letní škola metod molekulární biologie nukleových kyselin a genomiky
 Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Biotechnologický kurz Biotechnologický kurz II. letní škola metod molekulární biologie nukleových kyselin a genomiky 20. - 24. 6. 2011 Ústav morfologie, fyziologie a genetiky zvířat AF MENDELU v Brně Zemědělská
Co zjišťujeme u DNA ACGGTCGACTGCGATGAACTCCC ACGGTCGACTGCGATCAACTCCC ACGGTCGACTGCGATTTGAACTCCC
 Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Analýza DNA Co zjišťujeme u DNA Genetickou podstatu konkrétních proteinů Mutace bodové (sekvenční delece nebo inzerce nukleotidů), chromosomové aberace (numerické, strukturální) Polymorfismy konkrétní
Molekulární biotechnologie. Nový obor, který vznikl koncem 70. let 20. století (č.1)
") Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace
 Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Nukleosidy, nukleotidy, nukleové kyseliny, genetická informace Centrální dogma Nukleové kyseliny Fosfátem spojené nukleotidy (cukr s navázanou bází a fosfátem) Nukleotidy Nukleotidy stavební kameny nukleových
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Elektroforéza Sekvenování
 Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
Elektroforéza Sekvenování Výsledek PCR Elektroforéza V molekulární biologii se používá k separaci nukleových kyselin a bílkovin Principem je pohyb nabitých molekul v elektrickém poli Gelová, polyakrylamidová
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti NUKLEOVÉ KYSELINY 3 složky Nukleotidy dusík obsahující báze (purin či pyrimidin) pentosa fosfát Fosfodiesterová vazba. Vyskytuje se mezi
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Referenční lidský genom. Rozdíly v genomové DNA v lidské populaci. Odchylky od referenčního genomu. Referenční lidský genom.
 Referenční lidský genom Rozdíly v genomové DNA v lidské populaci Zdroj DNA: 60% sekvencí pochází ze sekvenování DNA od jednoho dárce (sekvenování a sestavování BAC klonů) některá místa genomu se nepodařilo
Referenční lidský genom Rozdíly v genomové DNA v lidské populaci Zdroj DNA: 60% sekvencí pochází ze sekvenování DNA od jednoho dárce (sekvenování a sestavování BAC klonů) některá místa genomu se nepodařilo
Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií
 poznatků genetiky a evolucionistických teorií") Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
Obecná genetika Zesouladení ( sjednocení ) poznatků genetiky a evolucionistických teorií Ing. Roman Longauer, CSc. Ústav zakládání a pěstění lesů, LDF MENDELU Brno Tento projekt je spolufinancován Evropským
Klonování. Co to vlastně je?
 Kristýna Boháčová Klonování Co to vlastně je? Klonování je podle běžné definice vytváření nového jedince geneticky identického (shodného) s předlohou Tyto dva jedinci se poté označují jako klony Tento
Kristýna Boháčová Klonování Co to vlastně je? Klonování je podle běžné definice vytváření nového jedince geneticky identického (shodného) s předlohou Tyto dva jedinci se poté označují jako klony Tento
Microfluidic systems, advantages and applications Monika Kremplová, Mgr.
 Název: Školitel: Microfluidic systems, advantages and applications Monika Kremplová, Mgr. Datum: 21. 6. 2013 Reg.č.projektu: CZ.1.07/2.3.00/20.0148 Název projektu: Mezinárodní spolupráce v oblasti "in
Název: Školitel: Microfluidic systems, advantages and applications Monika Kremplová, Mgr. Datum: 21. 6. 2013 Reg.č.projektu: CZ.1.07/2.3.00/20.0148 Název projektu: Mezinárodní spolupráce v oblasti "in
2 Inkompatibilita v systému Rhesus. Upraveno z A.D.A.M.'s health encyclopedia
 2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
2 Inkompatibilita v systému Rhesus Upraveno z A.D.A.M.'s health encyclopedia 3 Inkompatibilita v systému Rhesus Úkol 7, str.119 Které z uvedených genotypových kombinací Rh systému u manželů s sebou nesou
v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH
 RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání
RÁMCOVÝ VZDĚLÁVACÍ PROGRAM PRO ZÍSKÁNÍ SPECIALIZOVANÉ ZPŮSOBILOSTI v oboru KLINICKÁ GENETIKA PRO ODBORNÉ PRACOVNÍKY V LABORATORNÍCH METODÁCH 1. Cíl specializačního vzdělávání Cílem specializačního vzdělávání
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Mendelova genetika v příkladech. Genetické markery
 Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Mendelova genetika v příkladech Genetické markery Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Hodnocení genetické proměnlivosti Fenotypový
Izolace, klonování a analýza DNA
 Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Molekulární genetika II zimní semestr 4. výukový týden ( )
") Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Ústav biologie a lékařské genetiky 1.LF UK a VFN, Praha Molekulární genetika II zimní semestr 4. výukový týden (27.10. 31.10.2008) prenatální DNA diagnostika presymptomatická Potvrzení diagnózy Diagnostika
Laboratoř molekulární patologie
 Laboratoř molekulární patologie Ústav patologie FN Brno Prof. RNDr. Jana Šmardová, CSc. 19.11.2014 Složení laboratoře stálí členové Prof. RNDr. Jana Šmardová, CSc. Mgr. Květa Lišková Mgr. Lenka Pitrová
Laboratoř molekulární patologie Ústav patologie FN Brno Prof. RNDr. Jana Šmardová, CSc. 19.11.2014 Složení laboratoře stálí členové Prof. RNDr. Jana Šmardová, CSc. Mgr. Květa Lišková Mgr. Lenka Pitrová
Replikace, transkripce a translace
 Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Replikace, transkripce a translace Pravděpodobnost zařazení chybné báze cca 1:10 4, reálně 1:10 10 ; Proč? Výběr komplementární base je zásadní pro správnost mezigeneračního předávání genetické informace
Na rozdíl od genomiky se funkční genomika zaměřuje na dynamické procesy, jako je transkripce, translace, interakce protein - protein.
 FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
FUNKČNÍ GENOMIKA Co to je: Oblast molekulární biologie která se snaží o zpřístupnění a využití ohromného množství dat z genomových projektů. Snaží se popsat geny, a proteiny, jejich funkce a interakce.
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací