Využití vektorů při klonování DNA
|
|
|
- Sabina Doležalová
- před 6 lety
- Počet zobrazení:
Transkript

1 školní rok 2015/2016, kurz Bi6400 Využití vektorů při klonování DNA Jan Šmarda Ústav experimentální biologie Přírodovědecká fakulta MU 1



2 Klonování = proces tvorby klonů Klon: soubor geneticky identických buněk (resp. organismů), odvozených ze společného předka Klonování DNA: tvorba klonů DNA 2
3 Klonování = proces tvorby klonů Klon DNA: soubor identických molekul DNA, připravených např. množením rekombinantních molekul DNA v hostitelské buňce (in vivo) nebo PCR (in vitro) Rekombinantní molekula DNA: molekula DNA vytvořená spojením dvou cizorodých fragmentů DNA, např. klonované DNA s klonovacím vektorem 3

4 Paul Berg otec klonování DNA nar v USA emeritní profesor Stanfordovy univerzity Nobelova cena v oboru Chemie v roce 1980 (spolu s F. Sangerem a W. Gilbertem) za příspěvek k výzkumu nukleových kyselin 4
5 Klonování DNA umožňuje izolovat z komplexního genomu jeho dílčí úseky (např. geny), ty ve formě rekombinantních molekul mnohonásobně namnožit a zpřístupnit dalšímu studiu Využití klonování DNA: izolace genů studium regulačních oblastí, které řídí genovou expresi fyzikální a genetická analýza genomů exprese cizích genů v nepříbuzných hostitelích (heterologní exprese), např. za účelem produkce potřebných látek základ genového inženýrství (příprava transgenních organismů) 5

6 Klonování DNA-příklady využití 6
7 Postup klonování DNA výběr hostitelského organismu a klonovacího vektoru příprava vektorové DNA příprava klonované DNA vytvoření a přenos rekombinantní DNA do organismu hostitele selekce hostitelských buněk obsahujících rekombinantní DNA a jejich pomnožení identifikace ( screening ) klonů buněk nesoucích rekombinantní DNA Klonovací vektory se používají pro přenos klonované DNA do buněk a její následnou propagaci 7
8 Klonovací vektory - funkce zajištění přenosu studované DNA do hostitelské buňky zajištění propagace vlastní struktury vektoru, včetně klonované sekvence v hostitelské buňce zajištění segregace do dceřiných hostitelských buněk mnohonásobným množením transformované buňky vzniká klon identických buněk, z nichž každá obsahuje jednu nebo více kopií rekombinantní DNA: nesený gen je klonovaný 8
9 Klonovací vektory-základní informace nejčastěji kružnicové molekuly DNA schopné autonomní replikace obvykle odvozené z plazmidů nebo virů navíc obsahují pomocné sekvence různého původu usnadňující vložení cizorodé DNA, její expresi, selekci transformantů, apod. výhodná je malá velikost (do 10 kb) usnadnění purifikace a manipulace 9
10 Typy vektorů Plazmidové: např. pbr322, puc18 (=derivát pbr322), BAC Fágové: - odvozené od bakteriofága Lambda - odvozené od fága M13 Kosmidy: hybridy mezi plazmidy a fágy Vektory pro eukaryontní buňky: - kvasinkové - rostlinné - živočišné plasmid-bolivar-rodriguez (1977, UCSF, USA) 10
")

11 pbr322: nejúspěšnější vektor vytvořen Bolivarem a Rodriquezem - dvěma mexickými postdoky v laboratoři Herberta Boyera UCSF v roce 1977 (objevitel enzymu EcoRI) nikdy nebyl patentován, rychle šířen vědeckou komunitou zdarma výhody: kombinace rezistence na ampicilin a tetracyklin vysoká rychlost replikace nízké riziko konjugačních přenosů
 většina odvozena od plazmidu ColE1 bakterie E.")
12 Plazmidové vektory jednoduchá manipulace přirozený výskyt v mnoha druzích bakterií schopny nezávislé replikace, včetně jakéhokoliv fragmentu DNA, který je do nich uměle včleněn variabilita ve velikosti: jednotky stovky kb (pro klonování obvykle 2-15 kb) většina odvozena od plazmidu ColE1 bakterie E. coli 12
13 Neintegrační plazmidy extrachromozomové molekuly DNA, obvykle kruhové, dvouřetězcové, tvořící nadšroubovici Přirozené plazmidy obsahují vlastní počátky replikace replikují se nezávisle na chromozomu menší plazmidy využívají ke své replikaci enzymy hostitelské buňky, větší si kódují vlastní Integrační plazmidy - epizomy molekuly DNA, které se replikují jako součásti bakteriálního chromozomu v určité fázi se vyčlení a existují jako samostatné elementy 13
14 Klasifikace plazmidů dle znaků kódovaných plazmidovými geny fertilitní F plazmidy: nesou pouze geny tra (řídící konjugaci), stimulují konjugativní přenos plazmidů (např. F-plazmid E. coli) rezistenční R plazmidy) nesou geny, které hostitelům poskytují rezistenci k jednomu nebo více antibiotikům (např. RP4 plazmid Pseudomonas) Col plazmidy kódují koliciny, které zabíjejí jiné bakterie (např. ColE1 plazmid E. coli) degradativní plazmidy umožňují hostiteli metabolizovat neobvyklé molekuly jako např. toluen nebo kyselinu salicylovou (TOL plazmid Pseudomonas putida) virulentní plazmidy propůjčují hostiteli virulenci (patogenní vlastnosti) - např. Ti plazmid Agrobacterium tumefaciens, způsobující tvorbu nádorů u dvouděložných rostlin 14
15 Přirozené plazmidy podbuněčné elementy DNA disponující schopností nezávislé replikace a občasného přechodu do jiné buňky forma symbiotického vztahu: plazmid poskytuje hostiteli určitou výhodu (např. rezistenci na škodlivou látku), hostitel poskytuje plazmidu možnost propagace zodpovídají za šíření genů pro rezistenci k antibiotikům je zakázáno provádět takové experimenty, které by mohly způsobit rozšíření nových genů pro rezistenci k antibiotikům v patogenních bakteriích 15
16 Selekce plazmidů Geny zajišťující rezistenci na antibiotika, které jsou přítomné v plazmidech velmi zjednodušují sledování přítomnosti plazmidů v buňkách fungují jako selekční markery 16
17 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 17
 podmínkou je přítomnost počátku replikace (místa ori) místo ori je definováno specifickou nukleotidovou")
18 Replikace plazmidů základní předpoklad pro využití plazmidů jako klonovacích systémů (namnožení fragmentu DNA v buňce) podmínkou je přítomnost počátku replikace (místa ori) místo ori je definováno specifickou nukleotidovou sekvencí 18
19 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 19
20 Tvorba více kopií plazmidu v buňce deriváty ColE1 jsou multi-copy plazmidy ColE1 divokého typu cca 15 kopií v 1 buňce uměle vylepšené deriváty několik set kopií faktory řídící počet kopií plazmidů v buňce nejsou jasné Výhody - snadná purifikace s vysokými výtěžky - vysoká exprese klonovaného genu Nevýhody: - pomalejší růst hostitelské buňky - nadměrná exprese klonovaného genu může být pro buňku zátěž 20
21 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 21
22 Plazmidy a rozmezí hostitelů některé plazmidy se replikují v rozmanitých bakteriálních druzích (široké rozmezí hostitelů): RP4, RSF1010, pc194 většina vektorů užívaných pro klonování se replikuje jen v úzkém rozmezí hostitelů Výhoda: - snížení rizika rozšíření pozměněné genetické informace Nevýhoda: - pokud potřebujeme experimentovat s jinými bakteriemi než E. coli musíme si připravit vlastní vektor se specifickým ori 22
23 Pendlující (bifunkční, shuttle ) vektory pjk3-1, pkt240, PGC3311 možný přenos mezi dvěma bakteriálními druhy 2 místa ori v jednom plazmidu 23
24 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 24
 poloha klonovacího místa nesmí narušit funkci oblasti ori nebo jiné důležité funkce plazmidu")
25 Klonovací místo místo, ve kterém lze plazmid snadno otevřít unikátní (zastoupené pouze jednou v molekule plazmidu) restrikční místo do kterého se začleňuje cizorodá DNA větší počet různých restrikčních míst se cíleně seskupuje do krátkého úseku DNA (mnohočetné klonovací místo, polylinker) poloha klonovacího místa nesmí narušit funkci oblasti ori nebo jiné důležité funkce plazmidu přerušení kódující sekvence začleněním klonovaného fragmentu lze využít pro screening transformantů 25
26 Klonovací místo insert se může do vektoru začlenit v obou orientacích (pokud jsou konce molekul DNA kompatibilní) 26
27 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 27
28 Přenos plazmidových vektorů do hostitelských buněk podmínka klonování buňka může být transformována jen jednou molekulou DNA z každé buňky vyroste na agarové plotně jedna kolonie každá kolonie je klonem identických buněk nesoucích stejnou molekulu DNA 28
29 Přenos plazmidových vektorů do hostitelských buněk hostitelské bakterie umožňují snadné namnožení (propagaci) klonované DNA 29

30 Transformace: přenos molekuly DNA do bakteriální buňky bakterie se liší v míře schopnosti přijímat cizorodou DNA většina přijímá jen omezené množství Stanley N. Cohen (nar. 1935) Stanford University bakterie musí být chemicky nebo fyzikálně ošetřeny, aby se účinnost transformace zvýšila: musí být přivedeny do stavu kompetence 30
31 Příprava kompetentních buněk E. coli 70. léta 20. století: suspendování buněk E. coli v ledovém solném roztoku vede k navození stavu kompetence buňky se vyhladoví v chlazeném roztoku CaCl 2 (v tomto stavu se mohou zamrazit) sůl usnadňuje přichycení DNA na vnější buněčný povrch (nikoliv průnik stěnou) pohyb DNA dovnitř buňky je nastává po krátkém zvýšení teploty na 42 o C ( teplotní šok ), kterým se zvýší fluidita membrány další možnost - elektroporace 31

 možnost oboustranného pohybu různých molekul do buňky a z")
32 Transformace bakterií elektroporací promytí buněk vodou (odmytí elektrolytů z růstového média) krátký elektrický puls o vysokém napětí vznik dočasných otvorů v buněčném obalu vstup DNA do buňky Výhody vysoká účinnost přenosu použitelnost u různých typů bakterií Nevýhody nutnost optimalizace několika parametrů (růstové podmínky, teplota, délka pulzu, napětí) možnost oboustranného pohybu různých molekul do buňky a z buňky 32
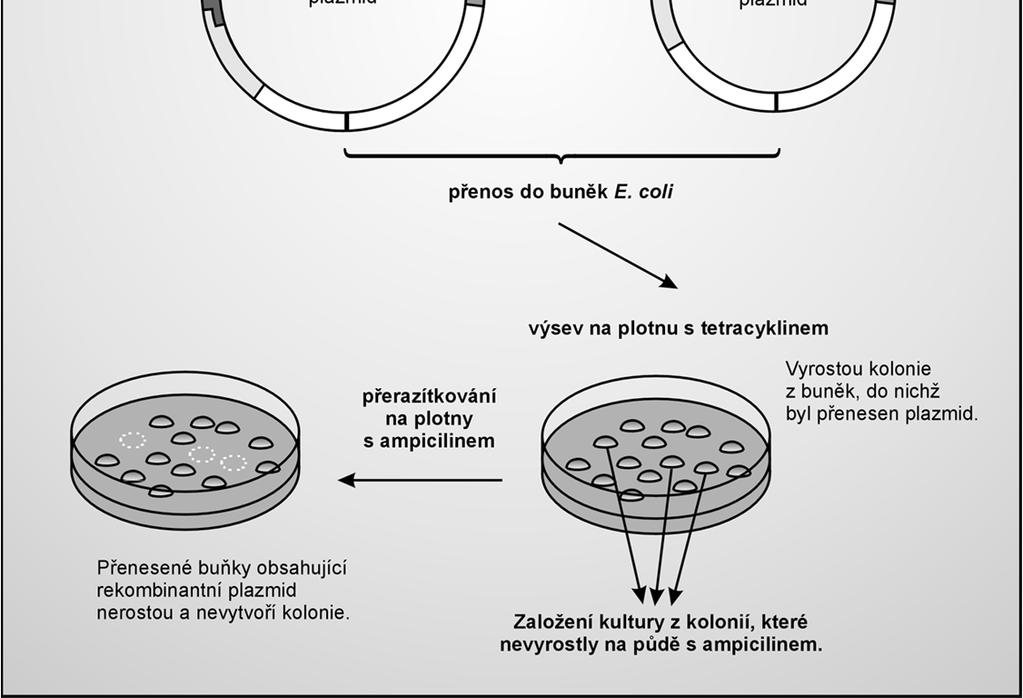
33 Po transformaci bakterií následuje selekce transformantů po transformaci se bakteriální buňky krátce inkubují v růstovém médiu bez selekce (zotavení bakterií, exprese selekčního markeru) výsev na agarové plotny obsahující selekční činidlo selekce transformantů výtěžek cca transformovaných buněk na 1 ng plazmidu 33
34 Požadavky na plazmidové vektory autonomní replikace v bakteriální buňce tvorba více kopií v buňce schopnost stabilního udržení cizorodé DNA při replikaci, nepřenosnost do dalších buněk konjugací (bezpečnost práce s novými konstrukty) přítomnost restrikčních (klonovacích) míst, využitelných pro klonování snadný a účinný přenos do hostitelských buněk přítomnost jednoho nebo více markerů pro selekci hostitelské buňky (např. rezistence na antibiotikum) malá velikost (zvýšení účinnosti transformace, jednoduchá izolace) 34
 procesy ligace i transformace jsou málo účinné: DNA přijme max. 1% bakteriálních buněk (E.")
35 Selekční markery nezbytné pro funkci vektoru pomáhají odlišit transformované buňky (menšinu) od většiny netransformovaných buněk (tj. zabránit v růstu netransformovaným buňkám) procesy ligace i transformace jsou málo účinné: DNA přijme max. 1% bakteriálních buněk (E. coli), v praxi obvykle méně selekce transformantů je obvykle zajištěna plazmidovým genem, který hostitelským buňkám poskytne rezistenci na antibiotikum 35
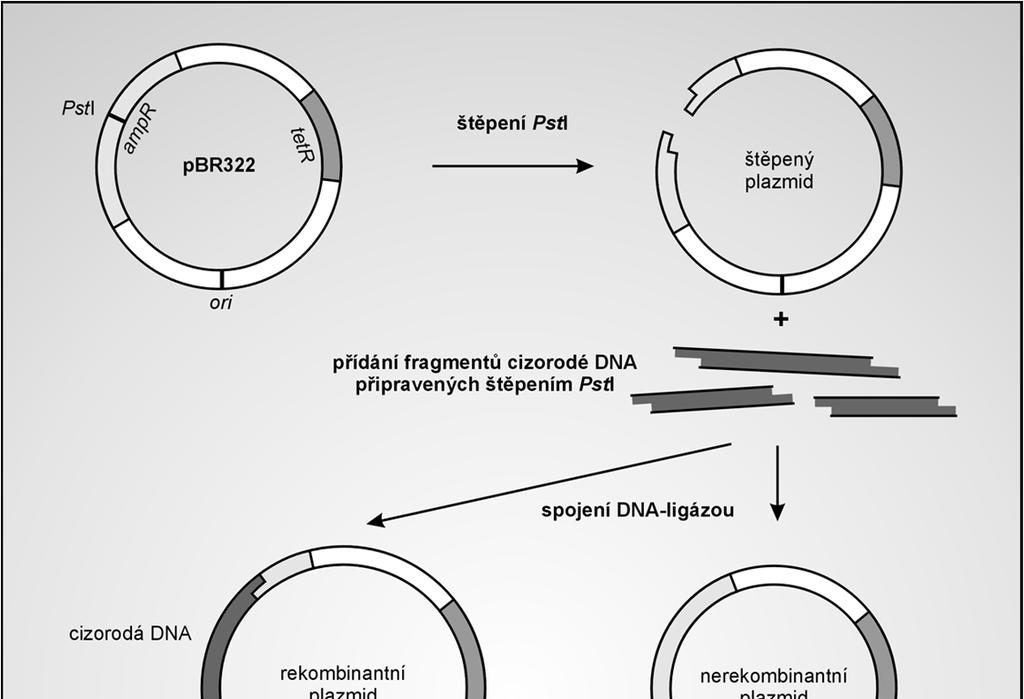
36 Princip identifikace transformantů bakterie E. coli jsou citlivé na ampicilin a tetracyklin pokud nesou plazmid pbr322 získávají rezistenci na tato antibiotika Amp rezistenci zajišťuje gen bla další 2 geny kódují enzymy, které detoxifikují Tet 36
, např.")

37 Gen bla kóduje enzym β-laktamázu Alexander Fleming ( ) β-laktamáza hydrolyzuje β-laktamová antibiotika (příbuzná penicilinu), např. ampicilin, která účinně zabíjejí bakterie tím, že zabraňují tvorbě jejich buněčné stěny β-laktamázy mění tato antibiotika do formy, která není pro bakterii toxická 37
38 Ne přítomnost plazmidu, ale exprese nesených genů rozhoduje o rezistenci není vhodné transformanty okamžitě vysévat na selekční médium po teplotním šoku je třeba buňky krátce kultivovat v médiu bez antibiotik čas je nutný pro expresi genu, který hostitelské buňce zajistí přežití v selekčních podmínkách 38
39 Identifikace bakteriálních kolonií obsahujících rekombinantní plazmidy ( screening ) selekce transformantů neřeší problém varianty neseného plazmidu: selekce nerozlišuje prázdný a rekombinantní vektor Nutný screening tranformantů, např: restrikční analýzou plazmidové DNA inzerční inaktivací alfa-komplementací 39

40 Restrikční analýza začlenění klonované DNA změní molekulovou hmotnost a restrikční vzorec (přítomnost nových restrikčních míst) je třeba izolovat plazmidy z jednotlivých klonů transformantů, štěpit je vhodnou restrikční endonukleázou a provést gelovou elektroforézu zjištění orientace klonovaného fragmentu 40
41 Inzerční inaktivace slouží přímé identifikaci kolonií nesoucích rekombinantní plazmid úspěšné začlenění fragmentu DNA do klonovacího místa se projeví na fenotypu kolonií transformovaných bakterií klonovací místo je ve vektoru (např. pbr322) umístěno v jednom z genů zodpovědných za rezistenci hostitelské buňky k antibiotiku inzerce klonované DNA způsobí ztrátu funkce tohoto genu buňky nesoucí rekombinantní plazmid jsou k danému antibiotiku citlivé, buňky nesoucí prázdný vektor jsou rezistentní 41
42 Inzerční inaktivace 42


43 Razítkování kolonií přenos kolonií v identické vzájemné poloze z jedné misky na druhou 43
 je umístěno v blízkosti 5 -konce genu lacz kódujícího β-galaktozidázu buňky obsahující plazmid puc8 jsou amp r a mohou tvořit β-galaktozidázu přítomnost klonovacího místa nenarušuje")
44 Inzerční inaktivace: modro-bílý test klonovací místo (např. plazmidu puc18) je umístěno v blízkosti 5 -konce genu lacz kódujícího β-galaktozidázu buňky obsahující plazmid puc8 jsou amp r a mohou tvořit β-galaktozidázu přítomnost klonovacího místa nenarušuje čtecí rámec lacz, pouze k tomuto genu přidává několik kodónů, funkce produktu (hydrolýza laktózy) tím není narušena (modré kolonie na plotnách s chromogenním substrátem) 44


45 Inzerční inaktivace: modro-bílý test včleněním klonovaného fragmentu DNA do klonovacího místa vektoru se přeruší sekvence lacz, kolonie jsou amp r, ale β-galaktozidázu tvořit nemohou (bílé kolonie na plotnách s chromogenním substrátem) 45
46 Kultivační podmínky pro detekci aktivity β-galaktozidázy v bakteriích místo štěpení laktózy na galaktózu testujeme odlišnou reakci katalyzovanou tímto enzymem: Chromogenním substrátem je látka analogická laktóze X- gal (5-bromo-4-chloro-3-indolyl-β-D-galaktozid), která se β-galaktozidázou štěpí na zabarvený produkt induktorem enzymu je IPTG (izopropylthiogalaktozid) X-gal a IPTG se přidávají spolu s Amp do agarového média 46

47 X-gal je bezbarvý substrát, který je β-galaktozidázou konvertován na tmavě modrý produkt. Modrá kolonie: buňky produkují β-galaktozidázu - plazmid je intaktní, žádný klonovaný fragment Bílá kolonie: buňky netvoří β-galaktozidázu - plazmid má přerušen gen lacz, klonování bylo (patrně) úspěšné 47
48 Komplementace puc18 nese jen část genu lacz, zbytek genu poskytuje hostitelská bakterie (princip komplementacedoplnění funkce) umožňuje udržet nízkou velikost plazmidu Ullman, Jacob, Monod,
49 Výhody a navýhody modro-bílého testu Výhody: jednoduše poskytuje informace o úspěšnosti ligace zajišťuje selekci transformantů (je Amp R ) bílá barva kolonie signalizuje, že přijatý plazmid nevznikl recirkularizací prázdného vektoru, ale došlo k ztrátě funkce LacZ, pravděpodobně v důsledku inzerční inaktivace Nevýhody: pokud je inzert malý a nepřerušuje čtecí rámec, β-galaktozidáza může mít dostatek aktivity pro zmodrání kolonií bílá kolonie není vždy zárukou úspěšného klonování (delece v MCS, včlenění nežádoucího úseku DNA, který poruší čtecí rámec) 49
 kyvadlové vektory: obsahují dva počátky replikace možnost propagace ve dvou různých organismech (např. E.coli a B.")
50 Vektory pro speciální účely expresní vektory: obsahují promotor, kterým lze zajistit produkci cizího proteinu v hostitelských buňkách (vhodné jsou inducibilní systémy) kyvadlové vektory: obsahují dva počátky replikace možnost propagace ve dvou různých organismech (např. E.coli a B. subtilis) 50
51 Expresní vektory upraveny nejen pro klonování DNA, ale také pro její expresi obsahují signály pro: - iniciaci transkripce (promotor) u transkripčních fúzních vektorů - iniciaci transkripce a translace (promotor, RBS a iniciační kodón) u translačních fúzních vektorů cizorodá DNA může být v expresních vektorech podrobena transkripci in vitro 51
52 2 typy expresních vektorů 1. Transkripční fúzní vektor obsahuje promotor translační signály poskytuje klonovaná DNA 2. Translační fúzní vektor obsahuje signály pro iniciaci transkripce i translace klonovaný fragment se začleňuje do kódující oblasti genu ve vektoru inzert musí respektovat čtecí rámec 52
53 Transkripční fúzní vektor 53

54 Translační fúzní vektor 54
 možno snadno získat smyslové i protismyslné transkripty klonovaného")
55 Vektory pgem expresní vektory MCS je obklopeno dvěma opačně orientovanými fágovými promotory (T7 a SP6) možno snadno získat smyslové i protismyslné transkripty klonovaného genu 55
56 Vektory pro klonování a expresi genů v eukaryotických buňkách primární klonování se provádí v bakteriích E. coli, následuje přenos do eukaryotických buněk v eukaryotických buňkách se často sleduje funkce genu, interakce jeho produktu s jinými proteiny, atd. studium možností regulace a zdokonalení syntézy důležitých metabolických produktů (např. hormonů inzulin) změny vlastností organizmu (např. zvýšení odolnosti užitkových rostlin k herbicidům) větší důraz na expresi daného genu než na tvorbu genových knihoven 56
57 Kvasinkové epizomové vektory založené na přirozených kvasinkových plazmidech 2 µm, které se vyskytují u většiny kmenů S. cerevisiae kopií na buňku samostatná replikace v kvasinkách obsahují místo ori pro replikaci v E. coli (kyvadlové vektory) využití: např. při výrobě léčiv z klonovaných genů nevýhoda: absence selekčního markeru REP zapojeny do replikace plazmidu FLP zapojen do intramolekulární rekombinace 57
58 Kvasinkové epizomové vektory (YEps) selekce založena na komplementaci auxotrofních mutací hostitelského kmene např. v genech trp1, ura3, leu2, his3 (podíl na biosyntéze aminokyselin) hostitelský kmen musí nést příslušnou mutaci (kvasinka neporoste v médiu postrádajícím příslušnou aminokyselinu) nedostatek antibiotik, ke kterým jsou kvasinky citlivé 58

59 Kvasinkový epizomový vektor nese marker pro rezistenci k antibiotiku pro selekci v E. coli nízká stabilita v kvasinkových buňkách plynoucí z občasné chybné segregace při mitóze občasná integrace do kvasinkového chromozomu homologní rekombinací (gen fungující jako selekční marker je velmi podobný mutované formě genu, která se nachází v chromozomové DNA) 59
60 Klonování v kvasinkovém epizomovém vektoru YEp13 60
61 Kvasinkový integrační plazmid (Yip) bakteriální plazmid nesoucí kvasinkové geny neschopen samostatné replikace v kvasinkách integruje se do chromozomu homologní rekombinací nízká účinnost transformace obtížné získání rekombinantního vektoru po transformaci výrazně vyšší stabilita než u autonomně se replikujících plazmidů 61
62 Umělý kvasinkový chromozom (YAC) Obsahuje: dvě kvasinkové telomery (správná replikace konců, ochrana před exonukleázami, udržování lineární podoby chromozomu) centromeru (rozdělení do sesterských buněk) sekvence pro autonomní replikaci (počátky replikace po celé délce chromozomu) selektovatelné signální markery replikuje se v buňkách E. coli i kvasinkách (kyvadlový vektor) vhodný pro klonování velkých fragmentů DNA (0,2 2 Mb) 62
63 Umělý kvasinkový chromozom (YAC) Výhody: vysoká klonovací kapacita lze klonovat celé eukaryotické geny, včetně regulačních sekvencí Nevýhody: nižší stabilita inzertu (možnost vnitřní rekombinace) obtížná purifikace 63
 restrikčním štěpením před klonováním se provádí další restrikční štěpení - vznikají dvě lineární ramena")
64 Klonování v YAC propagace v E. coli v kruhové plazmidové formě odstranění vyplňovacího fragmentu mezi telomerami ( stuffer ) restrikčním štěpením před klonováním se provádí další restrikční štěpení - vznikají dvě lineární ramena nesoucí selekční markery ligace insertu mezi ramena (podobně jako u fága Lambda) transformace kvasinkových buněk s částečně narušenou buněčnou stěnou (sféroplastů) selekce založená na komplementaci auxotrofních mutací (rekombinanti obsahují obě ramena) 64


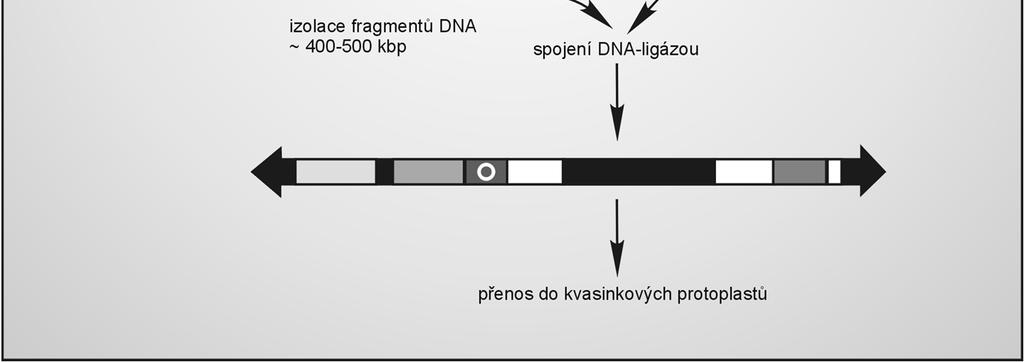
65 Klonování v YAC 65



 vhodné pro")
66 Vektory odvozené od fága Lambda Výhody: rekombinantní DNA lze sbalit do kapsidů a přenést do hostitelských buněk infekcí (o několik řádů vyšší účinnost přenosu než při transformaci plazmidovou DNA) v jedné zkumavce lze uchovávat ve formě fágových virionů celou genovou knihovnu (např. několik miliónů rekombinantních klonů) vhodné pro klonování větších fragmentů DNA (výhodné pro tvorbu genových knihoven) 66

67
 rozhodují vlivy okolí, genetická charakteristika hostitele i fága existují mutanti fága Lambda")
68 Lysogenní a lytický cyklus Lambda je temperovaný bakteriofág (po infekci E. coli může podstoupit lytický nebo lysogenní cyklus) rozhodují vlivy okolí, genetická charakteristika hostitele i fága existují mutanti fága Lambda podstupující pouze lytický cyklus (jasné plaky) divoký typ poskytuje zakalené plaky 68
")

 ničí represor")

69 Lysogenie exprese téměř všech fágových genů je vypnuta fágovým represorem (produkt genu ci) spontánní represe není absolutní jednou za každých 10 5 buněčných dělení se profág spontánně vyčlení a vstupuje do lytické dráhy reparační mechanismy aktivované poškozením DNA (UV záření) ničí represor CI (přechod na lytický cyklus) genom Lambda se integruje do genomu hostitele místně specifickou rekombinací a spolu s ním se replikuje 69
70 Genom fága Lambda dvouřetězcová lineární DNA ( bp), 50 genů 12 nespárovaných, ale komplementárních nukleotidů na obou koncích (lepivé konce) párování koncových sekvencí je vzhledem k jejich délce stabilní v infikované buňce E. coli se DNA fága Lambda převádí do kružnicové formy: důsledek kovalentní vazby katalyzované bakteriální DNA-ligázou 70
71 Lepivé konce DNA fága Lambda 71
72 Lytický cyklus kružnicová DNA se nejdříve replikuje obdobně jako plazmidy: vznik dceřiných kruhových molekul později se replikace přepíná na systém otáčivé kružnice: vznik dlouhých lineárních molekul DNA, obsahujících mnoho spojených kopií genomu Lambda (konkatemery) 72
73 Sestavování fágových částic exprese fágových genů sestavení prázdné fágové hlavy enzymy rozeznávají specifická místa na konkatemerech a provedou v nich asymetrická štěpení: vznik lepivých konců ( cohesive end sites - cos) oblast vymezená 2 místy cos je sbalena a přenesena do fágové hlavy připojení bičíku 73
74 Životní cyklus fága Lambda 74
75 Vektory fága Lambda mají 2 omezení molekulu DNA Lambda lze zvětšit jen asi o 5%, což odpovídá přidání pouze cca 3 kb nové DNA (větší molekuly se nazabalí do fágové hlavy) genom Lambda obsahuje cílová místa pro všechny běžné restriktázy (komplikace pro klonování, riziko rozpadu vektoru) 75
 cca 15 kb zmenšený genom není lyzogenní a prochází pouze lytickým cyklem 76")
76 Zvýšení klonovací kapacity vektoru lambda odstranění postradatelných genů, které zajišťují lysogenní cyklus (integraci a vyštěpení profága) cca 15 kb zmenšený genom není lyzogenní a prochází pouze lytickým cyklem 76
77 Odstranění cílových míst pro RE mutageneze in vitro (štěpení RE, modifikace konců DNA, zpětná ligace) přirozená selekce - hostitelem je např. kmen E. coli produkující EcoRI - EcoRI zničí většinu molekul DNA fága Lambda - pokud se objeví plaky - reprezentují mutované fágy, v nichž došlo k pozměnění cílových míst pro EcoRI - opakovanými cykly infekce lze získat molekuly Lambda, ve kterých budou chybět všechna místa EcoRI 77
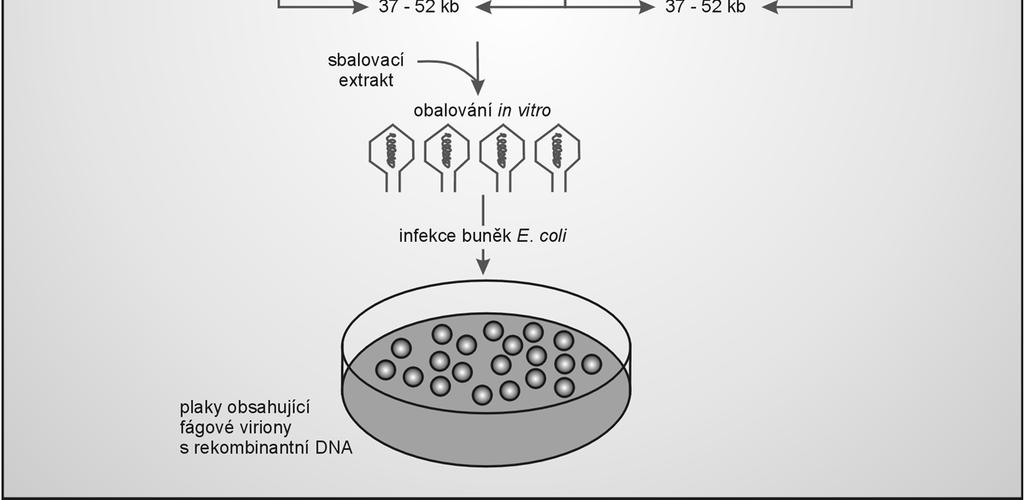
78 Sbalování DNA in vitro usnadňuje tvorbu virionů při klonování DNA přenos fágové DNA do buněk transfekcí je méně účinný než infekcí (větší velikost než obvyklé plazmidy) fágovou DNA lze účinněji přenést do buněk prostřednictvím fágových částic, které se sestaví pomocí sbalovacích extraktů, obsahujících prekurzory fágových hlav a bičíků rekombinantní lineární molekula DNA zakončená místy cos se sbalí do fágových kapsidů vytvořenými viriony se infikují hostitelské buňky E. coli, kde se rekombinantní DNA pomnoží 78
79 Dva typy vektorů odvozených od fága Lambda: Inzerční a substituční Inzerční vektory menší delece postradatelných fágových genů jsou sbalovatelné i bez insertu cizorodá DNA se vkládá do 1 restrikčního místa maximální klonovací kapacita - cca 13 kb 79
80 Příklad inzerčního vektoru: λgt10 obsahuje 1 unikátní klonovací místo (EcoRI) v genu ci schopnost tvorby životaschopných částic je zachována klonovací kapacita: 8 kb EcoRI vektor štěpí na 2 fragmenty (pravé a levé rameno) ligací se klonovaný fragment vkládá do místa EcoRI lepivé konce různých molekul se párují: vznik konkatemerů štěpení v místech cos a včlenění rekombinantní DNA do fágových hlav probíhá při sbalování in vitro : životaschopné fágové částice infekce bakterií na misce: tvorba plak 80
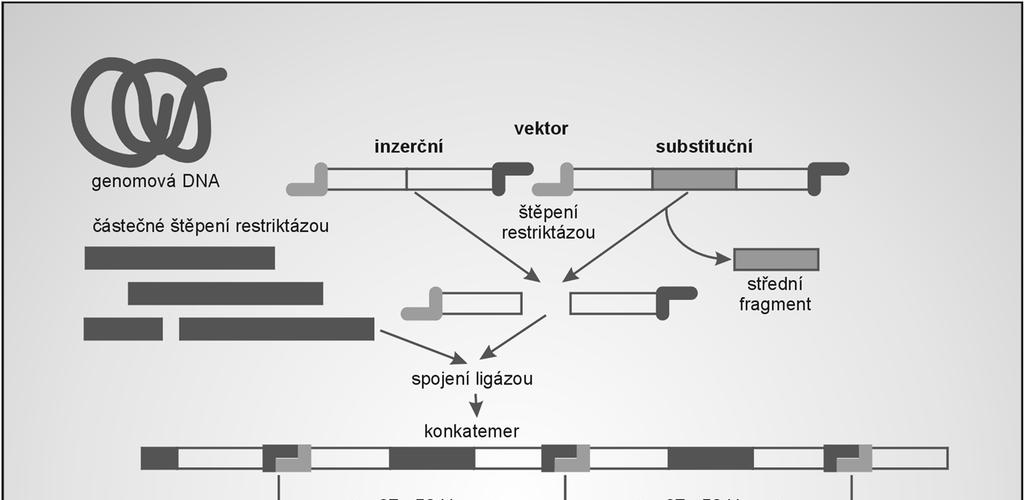
81 81
82 Dva typy vektorů odvozených od fága Lambda: Inzerční a substituční Substituční vektory mají deletovány téměř všechny sekvence, které nejsou nezbytné pro lytický cyklus - cca 40% genomu jsou k dispozici buď jako připravená ramena (nedají se množit bez insertu), nebo mají místo insertu vycpávku DNA ( stuffer fragment ) z libovolné sekvence, aby vektor zůstal sbalovatelný při klonování se vycpávka vyštěpí a nahradí klonovaným fragmentem, který může být velký až 24 kb. 82
83 Substituční (nahrazovací) vektory deriváty fága Lambda vyšší klonovací kapacita (až 24 kb) využívají přítomnosti sekvencí, které nejsou nezbytné pro lytický cyklus, ale jejich absence by znemožnila tvorbu životaschopných fágových částic (fágová hlava požaduje určitou minimální velikost DNA, ale nezáleží na nukleotidové sekvenci) mají 2 rozpoznávací místa pro restriktázu určenou pro klonování, která lemují nepotřebnou DNA (tzv. stuffer, vycpávka) používají se pro tvorbu genomových knihoven 83
84 Příklad substitučního vektoru: EMBL4 2 klonovací BamHI místa vymezují postradatelný fragment stuffer štěpením se uvolňuje stuffer + pravé a levé rameno ramena mají tendenci se spojovat (lepivé konce) 84
 infekce trávníkových bakterií, tvorba")
85 Klonování do substitučního vektoru štěpení vektoru enzymem BamHI oddělení vzniklých fragmentů elektroforézou eluce ramen z gelu ligace ramen s klonovaným fragmentem, který má kompatibilní konce rekombinantní fágová DNA v podobě konkatemeru je štěpena in vitro před začleněním do fágových hlaviček ( DNA packaging ) infekce trávníkových bakterií, tvorba plak 85
86 Výhody substitučních vektorů preferenčně jsou klonovány větší fragmenty (pokud je fragment menší než 8 kb nejsou částice životaschopné) možnost defosforylace inzertu: zamezení tvorby většího počtu tandemově uspořádaných insertů rozlišení rekombinantních a prázdných vektorů: střední fragment může kódovat β-galaktozidázu: plaky původního (nerekombinovaného) fága jsou modré na médiu obsahujícím X-gal 86

87 87
88 Kosmidy hybridy mezi fágovou DNA a bakteriálním plasmidem plazmidy, které obsahují místo cos spojení výhod plazmidových vektorů a vektorů odvozených od fága Lambda využívají toho, že sbalování probíhá in vitro nejen s genomy Lambda, ale také s jakoukoliv molekulou DNA nesoucí místa cos ve správné vzdálenosti 88
89 Využití kosmidů pro klonování štěpení, ligace insertů a purifikace rekombinantních klonů se provádí stejně jako při práci s plazmidy ligační podmínky se nastaví tak, aby byla podporována tvorba konkatemerů místo transformace bakterií se ligační směs podrobí sbalování in vitro ( DNA packaging ) vzniknou fágové částice, které zajistí přenos rekombinantní DNA do bakterií, ale nejsou životaschopné (nedochází k tvorbě plak, v kosmidu nejsou přítomny fágové geny) kosmid se bude v bakteriích replikovat jako plazmid (selekce bakteriálních kolonií) 89
90 Příklad kosmidu: pjb8 90
91 Výhody kosmidů klonovací kapacita kb možnost propagace a purifikace konvenčními metodami pro práci s plasmidy přirozená selekce pro velké inzerty (všechny kolonie jsou rekombinantní; nerekombinantní kosmidy jsou příliš malé, aby se začlenily do hlavičky fága) 91
 pro konjugaci")
, nesoucí 10 genů pouze jedna mezigenová sekvence o")
92 Bakteriofág M13 vláknitý bakteriofág infikující E.coli připojuje se na špičky povrchových výrůstků bakterií (pilusy) pro konjugaci neinfikuje bakterie F - dlouhý vláknitý tvar fágové částice obsahuje jednořetězcovou kružnicovou molekulu DNA (6,4 kb), nesoucí 10 genů pouze jedna mezigenová sekvence o velikosti 507 nukleotidů, kam je možné klonovat 92
93 Životní cyklus fága M13 obsahuje jednořetězcovou DNA jednovláknová molekula DNA M13 uvnitř buňky funguje jako templát pro syntézu komplementárního vlákna a replikuje se DNA M13 se nevčleňuje do genomu hostitele, ale přechází do obou dceřiných buněk při dělení nové fágové částice se tvoří průběžně, hostitelské bakterie pokračují v růstu 93
94 Replikace DNA fága M13 jednořetězcová DNA se uvnitř buňky mění na dvouřetězcovou DNA (replikační forma - RF) RF se replikuje mechanismem otáčivé kružnice, jehož produktem je lineární jednořetězcová DNA vzniklé vlákno se opět mění na RF nebo se začleňuje do dceřiných fágových virionů 94
95 Tvorba částic fága M13 při replikaci fágové DNA se v buňce hromadí mnoho kopií molekul podobných plazmidům zároveň se exprimují fágové geny fágový protein 5 se váže k ssdna a zahajuje její přesun k membráně, kde dojde k sestavení dceřiných fágových částic viriony nezpůsobují lýzi buňky (matné plaky) 95
96 Výhody vektorů odvozených od M13 replikace zahrnuje tvorbu ds DNA, kterou lze z buněk izolovat jako plazmidovou DNA a dále s ní jako s plazmidem manipulovat (použít ke klonování) významně usnadněno získání ssdna: do fágových virionů jsou sbalovány jednořetězcové kružnicové molekuly DNA, které lze snadno izolovat velikost virionu vláknitého fága je určena délkou molekuly DNA: délka klonované DNA není omezena (lze klonovat fragmenty DNA, jejichž velikost dosahuje několikanásobku délky fágového genomu) infikované buňky jsou životaschopné: nevznikají plaky, ale zóny pomaleji rostoucích buněk jsou viditelné - lze získat fágy o vysokém titru 96


97 Klonování ve vektorech M13 97
98 Klonovací kapacita vektorů Klonovací vektor Běžné plazmidové vektory λ Inzerční vektory λ Substituční vektory Kosmidové vektory PAC vektory BAC vektory YAC vektory Velikost inzertu 0-10 kb 0-10 kb 9-23 kb kb kb 300 kb Mb 98
99 Klonování ve vyšších rostlinách vektory založené na přirozeně se vyskytujících plazmidech bakterie Agrobacterium tumefaciens vektory založené na rostlinných virech možnost přímého přenosu genů do rostlin (bez vektorů) 99

100 Vektory odvozené od plazmidu Ti Agrobacterium tumefaciens není známý žádný plazmid specifický pro rostliny bakteriální plazmid Ti ( tumorinducing ) má pro rostliny velký význam bakterie A. tumefaciens pronikají do dvouděložné rostliny v místě poranění, přenášejí do rostlinných buněk Ti plazmid a vyvolají zde onemocnění crown gall zával plynoucí z nadměrné proliferace buněk 100
101 Ti plazmidy Agrobacterium tumefaciens A. tumefaciens: bakterie, která vyvolává tvorbu nádorů na tělech rostlin přenáší Ti ( tumor-inducing ) plazmid do rostlinných buněk část Ti plazmidu se začleňuje do chromozomové DNA Funkční elementy Ti plazmidu: oblast T, která se integruje do chromozomu rostlinné buňky oblast vir, která nese geny, jejichž produkty napomáhají přenosu T oblasti značná velikost plazmidu (cca 200 kb) Pokud se do T-DNA začlení žádaná nukleotidová sekvence, dojde k jejímu přenosu do rostlinných buněk 101

102 Ti plazmid Agrobacterium tumefaciens T-DNA obsahuje cca 8 genů, které se v hostitelské rostlinné buňce exprimují a jsou zodpovědné za: její nadměrnou proliferaci syntézu opinů, které bakterie potřebují pro svou výživu 102

103 Využití Ti plazmidu k přenosu genů do rostlin Problém: Ti plazmid o velikosti 200 kb nemá vhodná unikátní restrikční místa Řešení: 1. strategie binárního vektoru 2. strategie kointegrace 103
104 Využití Ti plazmidu k přenosu genů do rostlin 1. strategie binárního vektoru T-DNA nemusí být fyzicky spojena se zbytkem plazmidu buňky lze transformovat dvěma plazmidy: - malým plazmidem nesoucím T-DNA (obsahuje unikátní klonovací místo) - velkým plazmidem obsahujícím běžné sekvence Ti plazmidu s výjimkou T-DNA Pokud jsou oba plazmidy v jedné buňce A. tumefaciens, tak se doplňují: - T-DNA nesená malým plazmidem se přenese do rostlinné chromozomové DNA pomocí proteinů, které kóduje velký plazmid 104
105 Využití Ti plazmidu k přenosu genů do rostlin 2. strategie kointegrace klonovaný gen se vloží do běžného plazmidu s klonovanou krátkou sekvencí T-DNA tento plazmid se přenese do A.tumefaciens nesoucí Ti plazmid klonovaný gen se do Ti plazmidu přenese homologní rekombinací při infekci se přirozeně integruje do rostlinného chromozomu 105
106 Ne všechny rostlinné buňky přijmou exogen přirozeným průnikem A. tumefaciens s Ti plazmidem do rostliny (např. infekce poraněného stonku) se klonovaný gen objeví pouze v buňkách závalu malý praktický význam z hlediska biotechnologií jsou potřeba metody, kterými se gen dostane do všech buněk organismu 106

107 Regenerace rostlin z transformovaných buněk infekce rostlinných buněk A. tumefaciens v kultuře selekce transformantů regenerace rostliny z transformovaných buněk všechny buňky rostliny ponesou klonovaný gen 107
108 Podmínka regenerace rostliny: úprava Ti plazmidu musí ztratit svůj nádorotvorný potenciál odstraněním nádorových genů z T-DNA se nenaruší infekčnost infekčnost určuje oblast vir a 2 opakující se sekvence (25 bp) vymezující T-DNA oblast mezi opakujícími se sekvencemi může být změněna, aniž by došlo k ohrožení jejího přenosu do rostliny Plazmid pbin19 mezi repeticemi obsahuje: klonovací místa v genu lacz gen pro rezistenci ke kanamycinu Je to binární vektor: počáteční manipulace v E. coli, následuje přenos do A. tumefaciens a pak do rostliny selekce transformovaných rostlinných buněk na médiu s kanamycinem 108


109 Přenos genů do rostlinných buněk in vitro a regenerace rostliny 109
110 110
 problém: u drozofily nejsou známy žádné plazmidy vektory proto založeny na transpozonu")
111 Vektory pro hmyz vývoj hmyzích vektorů souvisí s atraktivností modelového systému octomilky (Drosophila melanogaster) problém: u drozofily nejsou známy žádné plazmidy vektory proto založeny na transpozonu (P-elementu) 111
112 Transpozony krátké úseky DNA (obvykle kratší než 10 kb), které se v buněčných chromozomech přemísťují z jednoho místa na druhé P-elementy jsou transpozony octomilky (2,9 kb): tvořeny 3 geny a koncovými převrácenými repeticemi 112
113 P-elementy 1 gen kóduje transpozázu invertované repetice představují rozpoznávací sekvence, které transpozáze umožňují identifikovat konce transpozonu mohou se pohybovat i mezi chromozomy a plazmidem 113
114 Vektory založené na P-elementech plazmidy nesoucí 2 modifikované P-elementy, z nichž jeden obsahuje inzerční místo pro klonovanou DNA v genu kódujícím transpozázu a druhý postrádá koncové repetice vložením klonované DNA se transpozáza inaktivuje, takže tento P- element je inaktivní druhý P-element je rovněž defektní, protože jeho konce nemohou být transpozázou rozeznány transpozáza druhého z P-elementů řídí přenos P-elementu s integrovaným genem do chromozomu hostitele 114
115 Klonování ve vektorech založených na P-elementech klonovaný gen se integruje do příslušného restrikčního místa v jednom z P-elementů mikroinjekce plazmidu do embryí octomilky transpozáza druhého z P-elementů řídí přenos P-elementu s integrovaným genem do některého z chromozomů octomilky pokud k přenosu dojde v zárodečné linii, vyvine se dospělá moucha obsahující klonovaný gen ve všech svých buňkách 115
 polyhedrin vytváří ochranný krystal kolem viru a zůstává nerozpustný v běžných podmínkách s výjimkou")
116 Vektory založené na bakulovirech používané k produkci proteinů v hmyzích buňkách bakuloviry jsou viry, které se běžně vyskytují u hmyzu genom bakuloviru obsahuje gen pro polyhedrin, jehož produkt se na konci infekčního cyklu v hmyzích buňkách hromadí ve formě inkluzních částic (silný promotor) polyhedrin vytváří ochranný krystal kolem viru a zůstává nerozpustný v běžných podmínkách s výjimkou zásaditých podmínek blízkých těm, kterou jsou ve střevech (zvyšuje životaschopnost viru mimo tělo hostitele) tento gen lze nahradit genem jiným a dosáhnout tak jeho výrazné exprese v hmyzích buňkách 116
117 Vektory pro klonování u savců využití : technologie gene knock-out nebo knock-in - studium genových funkcí produkce rekombinantních proteinů v buněčných kulturách genové terapie 117
118 Vektory pro savčí buňky replikace extrachromozomových elementů typu plazmidů je v savčích buňkách obtížná stabilní klony vznikají po přenosu DNA do buněk a jejím začlenění do chromozomu 118
 (vysoká konstitutivní exprese) polyadenylační signál (zvýšení stability mrna) počátek replikace příslušného viru (např. SV40) počátek replikace E.")
119 Vektory pro savčí buňky odvozené od plazmidů Obsahují signály, které savčí buňka vyžaduje pro zajištění genové exprese: obvykle virového původu (SV40, CMV) promotor (např. CMV) (vysoká konstitutivní exprese) polyadenylační signál (zvýšení stability mrna) počátek replikace příslušného viru (např. SV40) počátek replikace E. coli (ori) bakteriální selekční marker (bla) selekční marker pro savčí buňky neo (rezistence na toxické látky, např. G418 geneticin) 119
 obsahuje rané a pozdní geny Polyomavirus SV40")
120 Klonovací vektory pro savčí buňky odvozené z virů první klonování u savců provedeno s vektorem odvozeným z opičího viru SV40 (1979) SV40 infikuje několik savčích druhů u některých hostitelů probíhá lytický cyklus, u jiných lyzogenní genom (5,2 kb) obsahuje rané a pozdní geny Polyomavirus SV40 120
121 Vektory odvozené od viru SV40 rané geny: exprimují se dříve než pozdní kódují proteiny účastnící se replikace virové DNA pozdní geny: kódují proteiny virového obalu Stejný problém jako u fága lambda: systém sbalování omezuje velikost klonované DNA cizí DNA musí nahradit jeden nebo více postradatelných virových genů 121
122 Další vektory virového původu Odvozené od adenovirovů papilomavirů retrovirů 122

123 Retrovirové vektory velmi účinné, protože životní cyklus retrovirů zahrnuje zpětnou transkripci a integraci virové DNA do genomu infikované buňky mají charakter plazmidů, do kterých byly klonovány určité retrovirové sekvence studovaná DNA se začlení do virové sekvence daného plazmidu a rekombinantní plazmidová DNA se izoluje z bakterií DNA se přenáší do buněk transfekcí virové částice se z buněk uvolňují pučením a infikují cílové buňky integrace do genomové DNA je náhodná (stádium proviru) transgen je velmi stabilní a účinně se exprimuje 123

124 Klonování retrovirovými vektory živočišné buňky pěstované v kultuře jsou transfekovány retrovirovým vektorem obsahujícím žádanou DNA např. metodou precipitace fosforečnanem vápenatým v buňkách, které DNA přijaly, se tvoří rekombinantní retrovirové částice, které nesou cizorodou DNA a pučením buňky opouštějí tyto rekombinantní virové částice se použijí pro infekci žádaných buněk v infikovaných buňkách retroviry zajistí stabilní integraci dané DNA do chromozomu tvorbou provirů využití možnosti trans komplementace retrovirových funkcí defektním pomocným ( helper ) virem nebo hostitelem funkce, které nelze trans komplementovat, musí kódovat samotný vektor (LTR a místo psi) 124

125 Retroviry RNA genom v infikované buňce se RNA kopíruje do dvouřetězcové DNA zpětnou transkriptázou zpětná transkriptáza je obsažena ve virionu a vstupuje do buňky zároveň s RNA DNA se cirkularizuje a integruje do DNA hostitelské buňky působením integrázy integrovaná DNA je ohraničena sekvencemi LTR LTR obsahuje promotor pro transkripci genů gag, pol, env transkripty se sbalují do virových částic pomocí domény psi sestavené částice pučí ven z buňky, aniž ji lyzují a získávají tak glykoproteiny z hostitelské buněčné membrány 125
126 Klonování do retrovirových vektorů využití možnosti propagace v E. coli vytvoření rekombinantní verze plazmidu a její namnožení v bakteriích transfekce speciální buněčné linie ( helper cells ), která ve svém genomu obsahuje geny gag, pol a env transfekované buňky budou tvořit viriony obsahující RNA kopii vytvořeného konstruktu virové částice mohou infikovat jiné buňky, které neobsahují integrované esenciální geny přítomnost zpětné trankriptázy a integrázy ve virionech zajistí konverzi RNA do DNA a její stabilní integraci do genomu další viriony se nevytvoří 126
127 127
 mikroinjekce embryonálních kmenových buněk (mikroinjekce, vložení zpět do")
128 Klonování DNA bez vektoru nejefektivnějším přenosem do savčích buněk je mikroinjekce plazmidová nebo lineární DNA mikroinjikovaná do jádra savčích buněk se začleňuje do chromozomu mikroinjekce do oplodněné vaječné buňky se uplatňuje při tvorbě knock-out myši (krátká kultivace in vitro, implantace do náhradní matky) mikroinjekce embryonálních kmenových buněk (mikroinjekce, vložení zpět do embrya, implantace do náhradní matky) vede k tvorbě chimérických myší (směs upravených a neupravených buněk) 128

129 Přenosy genů do zárodečných myších buněk mikroinjekcí Význam: umožňují studium funkce genů v kontextu intaktního organismu myši, které tímto způsobem získaly cizí geny se nazývají transgenní Princip: DNA se mikroinjekcí vpraví do prvojádra oplozeného myšího vajíčka po kultivaci in vitro se vajíčka přenesou do pseudobřezí myši, kde se dále vyvíjejí u přibližně 10% potomstva bude cizorodá DNA integrována do genomu všech buněk pokud se cizorodá DNA vyskytuje v zárodečných buňkách, bude se přenášet křížením do dalšího potomstva stejně, jako kterýkoliv jiný gen 129

130 Přenosy genů do myší prostřednictvím embryonálních kmenových (ES) buněk ES buňky jsou uměle kultivované buňky odvozené z raných myších embryí, tzv. blastocyst, které mohou být kultivovány in vitro a později raným embryím opět předány jsou totipotentní mohou se podílet na vývoji všech tkání Princip: přenos DNA do ES buněk v kultuře selekce stabilních transfektantů přenos transfektantů do blastocysty mikroinjekcí přenos blastocysty do pseudobřezí myši 130

131 Přenosy genů do myší prostřednictvím embryonálních kmenových (ES) buněk Výsledek: některá z mláďat budou obsahovat jednak buňky odvozené z transfekovaných ES buněk a jednak normálních buněk. Myši představující směs dvou různých buněčných typů chimérické myši. U některých mláďat mohou být buňky odvozené z transfekovaných ES buněk v zárodečné linii: - jejich křížením se transfekovaný gen bude přenášet na potomstvo jako stabilní znak (transgenní myši). - pokud byl normální gen pozměněn ve smyslu úplné ztráty funkce, označují se tyto myši jako knockout myši. 131
132 Souhrn technik používaných při genetických manipulacích s myšími ES buňkami 132

133 133
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Genetika bakterií. KBI/MIKP Mgr. Zbyněk Houdek
 Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Genetika bakterií KBI/MIKP Mgr. Zbyněk Houdek Bakteriofágy jako extrachromozomální genomy Genom bakteriofága uvnitř bakterie profág. Byly objeveny v bakteriích už v r. 1915 Twortem. Parazitické org. nemají
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.12. Využití poznatků molekulární biotechnologie. Transgenní rostliny.
 Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
Molekulární biotechnologie č.12 Využití poznatků molekulární biotechnologie. Transgenní rostliny. Transgenní organismy Transgenní organismus: Organismus, jehož genom byl geneticky modifikován cizorodou
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE)
") MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
MUTAGENEZE INDUKOVANÁ TRANSPOZONY (TRANSPOZONOVÁ MUTAGENEZE) Nejrozšířenější použití transpozonů je mutageneza za účelem lokalizace genů a jejich charakterizace. Výhody: 1. vyšší frekvence mutace než při
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Genomika (KBB/GENOM) Fyzické mapování Fyzické cytogenetické a fyzické molekulární mapy Ing. Hana Šimková, CSc. Cíl přednášky
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Genové knihovny a analýza genomu
 Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Genové knihovny a analýza genomu Klonování genů Problém: genom organismů je komplexní a je proto obtížné v něm najít a klonovat specifický gen Klonování genů Po restrikčním štěpení genomové DNA pocházející
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Využití živých organismů pro uskutečňování definovaných chemických procesů pro průmyslové nebo komerční aplikace Organismus je geneticky upraven metodami genetického
Zdrojem je mrna. mrna. zpětná transkriptáza. jednořetězcová DNA. DNA polymeráza. cdna
 Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
Obsah přednášky 1) Klonování složených eukaryotických genů 2) Úprava rekombinantních genů 3) Produkce rekombinantních proteinů v expresních systémech 4) Promotory 5) Vektory 6) Reportérové geny Zdrojem
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
Mendelova genetika v příkladech. Transgenoze rostlin. Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno
 Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
Mendelova genetika v příkladech Transgenoze rostlin Ing. Petra VESELÁ, Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem
MIKROBIOLOGIE V BIOTECHNOLOGII
 Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
Biotechnologie MIKROBIOLOGIE V BIOTECHNOLOGII Termín biotechnologie byl poprvé použit v roce 1917 Procesy, při kterých se na tvorbě výsledného produktu podílejí živé organismy Širší definice: biotechnologie
ZÁKLADY BAKTERIÁLNÍ GENETIKY
 Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
Zdroj rozmanitosti mikrorganismů ZÁKLADY BAKTERIÁLNÍ GENETIKY Různé sekvence nukleotidů v DNA kódují různé proteiny Různé proteiny vedou k různým organismům s různými vlastnostmi Exprese genetické informace
BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy)
") BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy) Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza
BAKTERIÁLNÍ TRANSPOZONY (mobilní elementy) Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
analýza dat a interpretace výsledků
 Genetická transformace bakterií III analýza dat a interpretace výsledků Předmět: Biologie ŠVP: Prokaryotní organismy, genetika Doporučený věk žáků: 16-18 let Doba trvání: 45 minut Specifické cíle: analyzovat
Genetická transformace bakterií III analýza dat a interpretace výsledků Předmět: Biologie ŠVP: Prokaryotní organismy, genetika Doporučený věk žáků: 16-18 let Doba trvání: 45 minut Specifické cíle: analyzovat
Molekulární biotechnologie č.9. Cílená mutageneze a proteinové inženýrství
 Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Molekulární biotechnologie č.9 Cílená mutageneze a proteinové inženýrství Gen kódující jakýkoliv protein lze izolovat z přírody, klonovat, exprimovat v hostitelském organismu. rekombinantní protein purifikovat
Metody molekulární biologie
 Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Metody molekulární biologie 1. Základní metody molekulární biologie A. Izolace nukleových kyselin Metody využívající různé rozpustnosti Metody adsorpční Izolace RNA B. Centrifugační techniky o Princip
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 2.2.2018 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Klonování gen a genové inženýrství
 Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Klonování gen a genové inženýrství Genové inženýrství užite né termíny Rekombinantní DNA = DNA, ve které se nachází geny nejmén ze dvou zdroj, asto ze dvou zných druh organism Biotechnologie = manipulace
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Exprese rekombinantních proteinů
 Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Exprese rekombinantních proteinů Exprese rekombinantních proteinů je proces, při kterém můžeme pomocí různých expresních systémů vytvořit protein odvozený od konkrétního genu, nebo části genu. Tento protein
Školení GMO Ústav biochemie a mikrobiologie
 Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
Školení GMO Ústav biochemie a mikrobiologie 8.2.2019 Agrobacterium tumefaciens OZNÁMENÍ o uzavřeném nakládání první a druhé kategorie rizika na Ústavu biochemie a mikrobiologie VŠCHT a Ústavu biotechnologie
BAKTERIÁLNÍ REZISTENCE
 BAKTERIÁLNÍ REZISTENCE Petr Zouhar, Fyziologický ústav AV ČR, v. v. i.; UK v Praze, PřF, Katedra fyziologie V této úloze se v hrubých rysech seznámíte s některými metodami používanými v běžné molekulárně
BAKTERIÁLNÍ REZISTENCE Petr Zouhar, Fyziologický ústav AV ČR, v. v. i.; UK v Praze, PřF, Katedra fyziologie V této úloze se v hrubých rysech seznámíte s některými metodami používanými v běžné molekulárně
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Molekulární základy dědičnosti. Ústřední dogma molekulární biologie Struktura DNA a RNA
 Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Molekulární základy dědičnosti Ústřední dogma molekulární biologie Struktura DNA a RNA Ústřední dogma molekulární genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace DNA RNA
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316
 Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Využití houbových organismů v genovém inženýrství MIKROORGANISMY - bakterie, kvasinky a houby využíval
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 Využití houbových organismů v genovém inženýrství MIKROORGANISMY - bakterie, kvasinky a houby využíval
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í I ti d j dělá á í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním
ÚLOHA C Klonování PCR produktu do plasmidu
 Jméno a učo: Datum: ÚLOHA C Klonování PCR produktu do plasmidu TEORETICKÝ ÚVOD Při klonování PCR produktů do plasmidů se využívá vlastnosti Taq polymerasy, a jiných non-proofreading polymeras, přidávat
Jméno a učo: Datum: ÚLOHA C Klonování PCR produktu do plasmidu TEORETICKÝ ÚVOD Při klonování PCR produktů do plasmidů se využívá vlastnosti Taq polymerasy, a jiných non-proofreading polymeras, přidávat
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Rezistence patogenů vůči antimikrobialním látkám. Martin Hruška Jan Dlouhý
 Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
Rezistence patogenů vůči antimikrobialním látkám Martin Hruška Jan Dlouhý Pojmy Patogen (patogenní agens, choroboplodný zárodek nebo původce nemoci) je biologický faktor (organismus), který může zapřičinit
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 2.4 GENETICKÉ MANIPULACE in vitro - nekonvenční techniky, kterými lze modifikovat rostlinný
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/ B.Mieslerová (KB PřF UP v Olomouci)
") Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 2011 B.Mieslerová (KB PřF UP v Olomouci) VYUŽITÍ HOUBOVÝCH ORGANISMŮ V GENOVÉM INŽENÝRSTVÍ MIKROORGANISMY
Zvyšování konkurenceschopnosti studentů oboru botanika a učitelství biologie CZ.1.07/2.2.00/15.0316 2011 B.Mieslerová (KB PřF UP v Olomouci) VYUŽITÍ HOUBOVÝCH ORGANISMŮ V GENOVÉM INŽENÝRSTVÍ MIKROORGANISMY
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Molekulární biotechnologie. Nový obor, který vznikl koncem 70. let 20. století (č.1)
") Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Molekulární biotechnologie Nový obor, který vznikl koncem 70. let 20. století (č.1) Molekulární biotechnologie je založena Na přenosu genů z jednoho organismu do druhého Jeden organismus má gen, který
Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií
 Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Téma bakalářské práce: Úloha protein-nekódujících transkriptů ve virulenci patogenních bakterií Nové odvětví molekulární biologie se zabývá RNA molekulami, které se nepřekládají do proteinů, ale slouží
Speciace neboli vznik druhů. KBI/GENE Mgr. Zbyněk Houdek
 Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
Speciace neboli vznik druhů KBI/GENE Mgr. Zbyněk Houdek Co je to druh? Druh skupina org., které mají společné určité znaky. V klasické taxonomii se jedná pouze o fenotypové znaky. V evoluční g. je druh
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ
 BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
BUŇKA ZÁKLADNÍ JEDNOTKA ORGANISMŮ SPOLEČNÉ ZNAKY ŽIVÉHO - schopnost získávat energii z živin pro své životní potřeby - síla aktivně odpovídat na změny prostředí - možnost růstu, diferenciace a reprodukce
Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
 MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
MASARYKOVA UNIVERZITA V BRNĚ Přírodovědecká fakulta Ústav experimentální biologie Oddělení genetiky a molekulární biologie Interakce proteinu p53 s genomovou DNA v kontextu chromatinu glioblastoma buněk
Příprava rekombinantních molekul pro diagnostické účely
 1 Příprava rekombinantních molekul pro diagnostické účely doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 2 Obsah přednášky 1) Pojem rekombinantní DNA 2) Historické milníky
1 Příprava rekombinantních molekul pro diagnostické účely doc. RNDr. Milan Bartoš, Ph.D. bartosm@vfu.cz Přírodovědecká fakulta MU, 2014 2 Obsah přednášky 1) Pojem rekombinantní DNA 2) Historické milníky
Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor)
") RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
RNAi Rich Jorgensen a kolegové vložili gen produkující pigment do petunií (použili silný promotor) Místo silné pigmentace se objevily rostliny variegované a dokonce bílé Jorgensen pojmenoval tento fenomén
Zajištění exprese klonovaných genů a její optimalizace
 Zajištění exprese klonovaných genů a její optimalizace Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripční úroveň Síla promotoru a jeho charakter Terminátor
Zajištění exprese klonovaných genů a její optimalizace Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripční úroveň Síla promotoru a jeho charakter Terminátor
Zajištění exprese klonovaných genů a její optimalizace
 Zajištění exprese klonovaných genů a její optimalizace 1 Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripční úroveň Síla promotoru a jeho charakter Terminátor
Zajištění exprese klonovaných genů a její optimalizace 1 Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripční úroveň Síla promotoru a jeho charakter Terminátor
Ivo Papoušek. Biologie 8, 2015/16
 Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
Ivo Papoušek Biologie 8, 2015/16 Doporučená literatura: Metody molekulární biologie (2005) Autoři: Jan Šmarda, Jiří Doškař, Roman Pantůček, Vladislava Růžičková, Jana Koptíková Izolace nukleových kyselin
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Kyselina hyaluronová. Kyselina hyaluronová. Streptococcus equi subsp. produkovaná kyselina hyaluronová a. Autor prezentace: Mgr.
 Kyselina hyaluronová Streptococcus equi subsp. zooepidemicus a jím produkovaná kyselina hyaluronová a glukuronidáza Marcela Tlustá Biotechnologická laborato Meyer a Palmer, 1934 Extracelulární matrix,
Kyselina hyaluronová Streptococcus equi subsp. zooepidemicus a jím produkovaná kyselina hyaluronová a glukuronidáza Marcela Tlustá Biotechnologická laborato Meyer a Palmer, 1934 Extracelulární matrix,
Zajištění exprese klonovaných genů a její optimalizace
 Zajištění exprese klonovaných genů a její optimalizace Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripce Síla promotoru Terminátor transkripce Stabilita
Zajištění exprese klonovaných genů a její optimalizace Faktory ovlivňující expresi klonovaných genů A. Regulační sekvence pro genovou expresi 1. Transkripce Síla promotoru Terminátor transkripce Stabilita
Struktura a organizace genomů
 CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Hybridizace nukleových kyselin
 Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Hybridizace nukleových kyselin Tvorba dvouřetězcových hybridů za dvou jednořetězcových a komplementárních molekul Založena na schopnosti denaturace a renaturace DNA. Denaturace DNA oddělení komplementárních
Obsah přednášky. 1) Exprese v Escherichia coli 2) Exprese v Saccharomyces cerevisiae 3) Exprese v Pichia pastoris 4) Exprese v hmyzích buňkách
 Exprese v Escherichia coli 2) Exprese v Saccharomyces cerevisiae 3) Exprese v Pichia pastoris 4) Exprese v hmyzích buňkách") Obsah přednášky 1) Exprese v Escherichia coli 2) Exprese v Saccharomyces cerevisiae 3) Exprese v Pichia pastoris 4) Exprese v hmyzích buňkách Exprese v Escherichia coli proteiny větší než malé proteiny
Obsah přednášky 1) Exprese v Escherichia coli 2) Exprese v Saccharomyces cerevisiae 3) Exprese v Pichia pastoris 4) Exprese v hmyzích buňkách Exprese v Escherichia coli proteiny větší než malé proteiny
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
Izolace nukleových kyselin
 Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
Izolace nukleových kyselin Požadavky na izolaci nukleových kyselin V nativním stavu z přirozeného materiálu v dostatečném množství požadované čistotě. Nukleové kyseliny je třeba zbavit všech látek, které
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA
 RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
RESTRIKCE A MODIFIKACE FÁGOVÉ DNA po jednom cyklu Kmeny E. coli K a K(P1) + mají vzájemně odlišnou hostitelskou specifitu (K a P1) = obsahují odlišné RM-systémy Experimentální důkaz přítomnosti a působení
SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA
 SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA A. Transdukce E. coli, S. typhimurium, Bacillus, Klebsiella, Staphylococcus, Streptococcus Nespecifická (P22, P1, SPβ, φ11) abortivní Specifická (fág lambda) Jsou
SYSTÉMY ZPROSTŘEDKOVANÉHO PŘENOSU DNA A. Transdukce E. coli, S. typhimurium, Bacillus, Klebsiella, Staphylococcus, Streptococcus Nespecifická (P22, P1, SPβ, φ11) abortivní Specifická (fág lambda) Jsou
REPLIKACE A REPARACE DNA
 REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
REPLIKACE A REPARACE DNA 1 VÝZNAM REPARACE DNA V MEDICÍNĚ Příklad: Reparace DNA: enzymy reparace nukleotidovou excizí Onemocnění: xeroderma pigmentosum 2 3 REPLIKACE A REPARACE DNA: Replikace DNA: 1. Podstata
Elektronoptický snímek viru mozaikové choroby tabáku. Mozaiková choroba tabáku. Schéma viru mozaikové choroby tabáku
 Obecná virologie Viry lat. virus šťáva, jed, v lékařské terminologii infekční činitel 1879 1882: první pokusný přenos virového onemocnění (mozaiková choroba tabáku) 1898: první pokusný přenos živočišného
Obecná virologie Viry lat. virus šťáva, jed, v lékařské terminologii infekční činitel 1879 1882: první pokusný přenos virového onemocnění (mozaiková choroba tabáku) 1898: první pokusný přenos živočišného
Zkušební okruhy k přijímací zkoušce do magisterského studijního oboru:
 Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
Biotechnologie interakce, polarita molekul. Hydrofilní, hydrofobní a amfifilní molekuly. Stavba a struktura prokaryotní a eukaryotní buňky. Viry a reprodukce virů. Biologické membrány. Mikrobiologie -
19.b - Metabolismus nukleových kyselin a proteosyntéza
 19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
19.b - Metabolismus nukleových kyselin a proteosyntéza Proteosyntéza vyžaduje především zajištění primární struktury. Informace je uložena v DNA (ev. RNA u některých virů) trvalá forma. Forma uskladnění
ZÁKLADY VIROLOGIE. Obecná charakteristika virů. Chemické složení virů. Stavba viru. Bílkoviny
 Obecná charakteristika virů ZÁKLADY VIROLOGIE Nebuněčné mikroorganismy Genetické elementy, který se množí pouze uvnitř živé buňky Vnitrobuněčná a mimobuněčná forma Extracelulární forma virové částice se
Obecná charakteristika virů ZÁKLADY VIROLOGIE Nebuněčné mikroorganismy Genetické elementy, který se množí pouze uvnitř živé buňky Vnitrobuněčná a mimobuněčná forma Extracelulární forma virové částice se
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Základy molekulární biologie KBC/MBIOZ Ivo Frébort 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci. reg. č.: CZ.1.07/2.2.00/
 Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Implementace laboratorní medicíny do systému vzdělávání na Univerzitě Palackého v Olomouci reg. č.: CZ.1.07/2.2.00/28.0088 Hybridizační metody v diagnostice Mgr. Gabriela Kořínková, Ph.D. Laboratoř molekulární
Konjugace. Přenos DNA zprostředkovaný konjugativními plazmidy. Donor recipient transkonjugant
 Konjugace Přenos DNA zprostředkovaný konjugativními plazmidy Donor recipient transkonjugant (Exkonjuganti - v rámci téhož druhu, transkonjuganti - v rámci různých druhů) Přenášené typy elementů (DNA):
Konjugace Přenos DNA zprostředkovaný konjugativními plazmidy Donor recipient transkonjugant (Exkonjuganti - v rámci téhož druhu, transkonjuganti - v rámci různých druhů) Přenášené typy elementů (DNA):
Nebuněčný život (život?)
") Nebuněčný život (život?) Nebuněčný život (život?) 1. viry 2. viroidy (infekční RNA) 3. satelity (subvirální infekční jednotky, jejichž replikace buňkou je zajištěna koinfekcí pomocným virem ) (a) satelitní
Nebuněčný život (život?) Nebuněčný život (život?) 1. viry 2. viroidy (infekční RNA) 3. satelity (subvirální infekční jednotky, jejichž replikace buňkou je zajištěna koinfekcí pomocným virem ) (a) satelitní
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém
 Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Nové přístupy v modifikaci funkce genů: CRISPR/Cas9 systém Lesk a bída GM plodin Lesk a bída GM plodin Problémy konstrukce GM plodin: 1) nízká efektivita 2) náhodnost integrace transgenu 3) legislativa
Exprese genetické informace
 Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
Exprese genetické informace Tok genetické informace DNA RNA Protein (výjimečně RNA DNA) DNA RNA : transkripce RNA protein : translace Gen jednotka dědičnosti sekvence DNA nutná k produkci funkčního produktu
MOBILNÍ GENETICKÉ ELEMENTY. Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 MOBILNÍ GENETICKÉ ELEMENTY Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. Demerec (1937) popsal nestabilní mutace u D. melanogaster B. McClintocková (1902-1992, Nobelova cena 1983) ukázala ve
MOBILNÍ GENETICKÉ ELEMENTY Lekce 13 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. Demerec (1937) popsal nestabilní mutace u D. melanogaster B. McClintocková (1902-1992, Nobelova cena 1983) ukázala ve
Detekce Leidenské mutace
 Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
Detekce Leidenské mutace MOLEKULÁRNÍ BIOLOGIE 3. Restrikční štěpení, elektroforéza + interpretace výsledků Restrikční endonukleasy(restriktasy) bakteriální enzymy štěpící cizorodou dsdna na kratší úseky
25.2.2014. Genomika. Obor genetiky, který se snaží. stanovit úplnou genetickou informaci. organismu a interpretovat ji v. termínech životních pochodů.
 Genomika Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat ji v termínech životních pochodů. 1 Strukturní genomika stanovení sledu nukleotidů genomu organismu,
Genomika Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat ji v termínech životních pochodů. 1 Strukturní genomika stanovení sledu nukleotidů genomu organismu,
Molekulárn. rní. biologie Struktura DNA a RNA
 Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Molekulárn rní základy dědičnosti Ústřední dogma molekulárn rní biologie Struktura DNA a RNA Ústřední dogma molekulárn rní genetiky - vztah mezi nukleovými kyselinami a proteiny proteosyntéza replikace
Bílkoviny a rostlinná buňka
 Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Bílkoviny a rostlinná buňka Bílkoviny Rostliny --- kontinuální diferenciace vytváření orgánů: - mitotická dělení -zvětšování buněk a tvorba buněčné stěny syntéza bílkovin --- fotosyntéza syntéza bílkovin
Struktura a funkce nukleových kyselin
 Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Struktura a funkce nukleových kyselin ukleové kyseliny Deoxyribonukleová kyselina - DA - uchovává genetickou informaci Ribonukleová kyselina RA - genová exprese a biosyntéza proteinů Složení A stavební
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Objevy, které umožnily cíleně manipulovat s DNA
 Genové inženýrství Genové inženýrství se zabývá vytvářením pozměněných či nových genů a jejich zaváděním do organizmů s cílem rekonstruovat jejich genetickou výbavu. Metodickým základem genového inženýrství
Genové inženýrství Genové inženýrství se zabývá vytvářením pozměněných či nových genů a jejich zaváděním do organizmů s cílem rekonstruovat jejich genetickou výbavu. Metodickým základem genového inženýrství
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Obsah přednášky: RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Studijní materiály na: http://www.zoologie.upol.cz/osoby/fellnerova.htm Obsah přednášky: Obecná charakteristika virů velikost a morfologie
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Studijní materiály na: http://www.zoologie.upol.cz/osoby/fellnerova.htm Obsah přednášky: Obecná charakteristika virů velikost a morfologie
Okruhy otázek ke zkoušce
 Okruhy otázek ke zkoušce 1. Úvod do biologie. Vznik života na Zemi. Evoluční vývoj organizmů. Taxonomie organizmů. Původ a vývoj člověka, průběh hominizace a sapientace u předků člověka vyšších primátů.
Okruhy otázek ke zkoušce 1. Úvod do biologie. Vznik života na Zemi. Evoluční vývoj organizmů. Taxonomie organizmů. Původ a vývoj člověka, průběh hominizace a sapientace u předků člověka vyšších primátů.
Petr Müller Masarykův onkologický ústav. Genová terapie
 Genová terapie Petr Müller Masarykův onkologický ústav Genová terapie =terapie využívající vpravení exogenní DNA do buněk či tkání organismu za účelem opravy fenotypu (deficience či mutace genu, vrozené
Genová terapie Petr Müller Masarykův onkologický ústav Genová terapie =terapie využívající vpravení exogenní DNA do buněk či tkání organismu za účelem opravy fenotypu (deficience či mutace genu, vrozené
DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje.
 Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
Název: Viry. Autor: PaedDr. Pavel Svoboda. Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy. Předmět, mezipředmětové vztahy: biologie
 Název: Viry Výukové materiály Autor: PaedDr. Pavel Svoboda Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: biologie Ročník: 2. (1. vyššího gymnázia) Tematický
Název: Viry Výukové materiály Autor: PaedDr. Pavel Svoboda Název školy: Gymnázium Jana Nerudy, škola hl. města Prahy Předmět, mezipředmětové vztahy: biologie Ročník: 2. (1. vyššího gymnázia) Tematický
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Základy molekulární biologie KBC/MBIOZ Klonování a genetické modifikace Sci-fi Skutečnost 6. Molekulární biotechnologie a transgenní organismy Dolly the Sheep Nadexprese proteinů Genetické modifikace a
Základy genetiky prokaryotické buňky
 Základy genetiky prokaryotické buňky Chromozomová (jaderná) DNA U prokaryot (bakterie, archea) dvouřetězcová většinou kružnicová U eukaryot dvouřetězcová lineární U DNA-virů dvouřetězcová lineární, jednořetězcová
Základy genetiky prokaryotické buňky Chromozomová (jaderná) DNA U prokaryot (bakterie, archea) dvouřetězcová většinou kružnicová U eukaryot dvouřetězcová lineární U DNA-virů dvouřetězcová lineární, jednořetězcová
Transgeneze u ptáků: očekávání vs. realita
 Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva
 Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
Citlivost a rezistence mikroorganismů na antimikrobiální léčiva Sylva Janovská Univerzita Pardubice Fakulta chemicko-technologická Katedra biologických a biochemických věd Centralizovaný rozvojový projekt
Obsah přednášky: RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Studijní materiály na: http://www.zoologie.upol.cz/osoby/fellnerova.htm Obsah přednášky: Obecná charakteristika virů velikost a morfologie
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Studijní materiály na: http://www.zoologie.upol.cz/osoby/fellnerova.htm Obsah přednášky: Obecná charakteristika virů velikost a morfologie
Exprese genetického kódu Centrální dogma molekulární biologie DNA RNA proteinu transkripce DNA mrna translace proteosyntéza
 Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Exprese genetického kódu Centrální dogma molekulární biologie - genetická informace v DNA -> RNA -> primárního řetězce proteinu 1) transkripce - přepis z DNA do mrna 2) translace - přeložení z kódu nukleových
Izolace, klonování a analýza DNA
 Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Izolace, klonování a analýza DNA Ing. Pavel Kotrba, Ph.D., Ing. Zdeněk Knejzlík, Ph.D., Ing. Zdeněk Chodora Ústav biochemie a mikrobiologie, VŠCHT Praha HTpavel.kotrba@vscht.czTH, HTzdenek.knejzlik@vscht.czTH,
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
Základy molekulární biologie KBC/MBIOZ 4. Metody molekulární biologie I Izolace DNA a RNA Specifické postupy pro baktérie, kvasinky, rostlinné a živočišné tkáně U RNA nutno zabránit kontaminaci RNasami
studium množství určitého transkriptu v daném vzorku a v množství dané molekuly mrna v dané buňce a v daném
 Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Analýza genové exprese Analýza genomu genomika Analýza transkriptomu transkriptomika Analýza proteinu - proteomika Analýza transkripce studium množství určitého transkriptu v daném vzorku a v daném čase
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.