Paměťové stopy - engramy
|
|
|
- Vlasta Havlová
- před 9 lety
- Počet zobrazení:
Transkript
1 Paměťové stopy - engramy
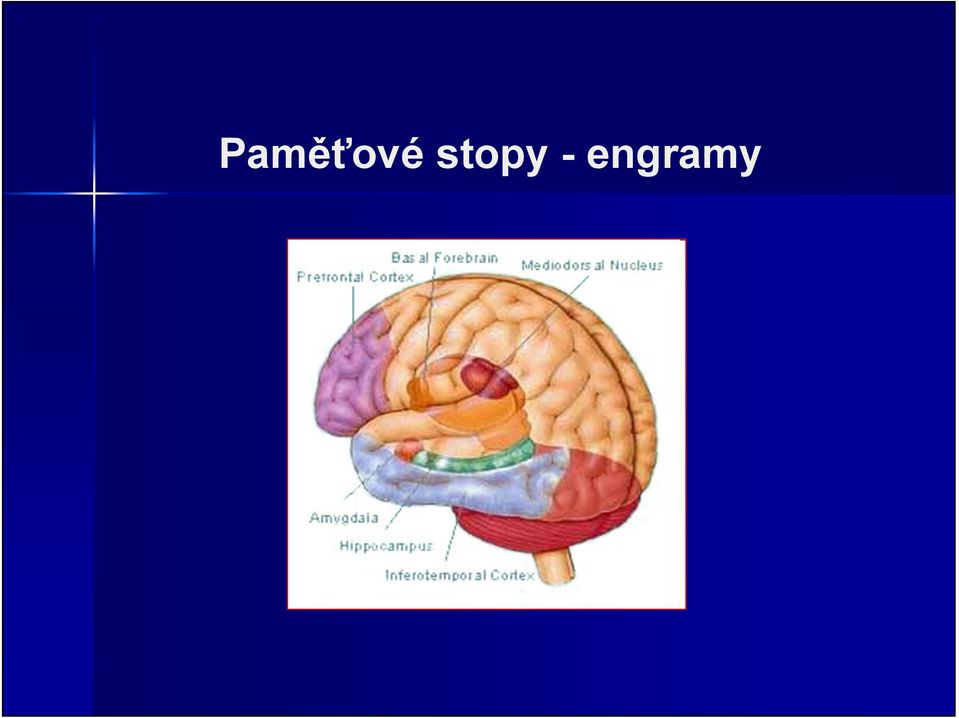
2 Obsah přednášky V této přednášce se budeme zabývat paměťovými stopami - ENGRAMY, jejich povahou, a některými fenomény s nimi spojenými Částečně si zopakujeme látku s přednášky č. 2, která pojednávala o mechanismech paměti

3 Engram paměťová stopa Tento pojem (navržený Richardem Semonem v r. 1921) značí hypotetický mechanismus jímž je uchována v paměti informace. - Semon, R. (1921), The Mneme. London: George Allen & Unwin Alternetivně může také značit vzpomínku, kousek informace v paměti Richard Semon německý zoolog a evoluční biolog, nepříliš známý, ale jeho myšlenky byly velmi vlivné Semon prosazoval koncept, že mentální stavy jsou zprostředkovány změnami v nervové činnosti Paměťová stopa není zpravidla lokalizována na jednom místě, ale je často distribuována v mozkových okruzích v podobě změn v synaptické plasticitě, které vedou ke změněné neuronální funkci, potažmo s pozorovatelné změně v chování

4 Engram a synapse

5 Lashleyho hledání engramu Karl Spencer Lashley ( ), byl americký psycholog a behaviorista, který byl ovlivněn behavioristou Johnem B. Watsonem Pokoušel se hledat, kde by engram mohl v mozku být lokalizován, a to za pomoci metody, kdy zvířata trénoval v různých úlohách (brightness discrimination a různá bludiště) a následně jim chirurgicky odnímal části mozkové kůry, a sledoval jak se jejich chování změnilo (povšimněte si, že tento koncept je v podstatě modifikován dodnes. Lashley ve hledání engramu neuspěl, nepodařilo se mu najít konkrétní strukturu, která by odpovídala za chování v těchto bludištích. Jeho zjištění postavilo teoretický rámec pro koncept distribuované paměťové stopy Lashleyho nálezy naznačovaly, že čím větší byl objem odňaté mozkové kůry, tím bylo poškození chování větší, ale že příliš nezáleželo na tom, kterou část kůry odejmul

6 Lashleyho principy V roce 1950 shrnul Lashley 30 let svého bádání do dvou principů (K. Lashley. In search of the engram. Symposia of the Society for Experimental Biology, 4: , 1950) Princip ekvipotenciality (Equipotentiality Principle) po odstranění jedné korové oblasti může jiná oblast kortexu nahradit funkci té odstraněné, tzn. že by ve finále nezáleželo na tom, kde je léze provedena Princip vlivu objemu (The Mass Action Principle) postižení je tím větší, čím větší je objem hmoty, která je chirurgicky odstraněna Jinými slovy Lashley zastával názor, že paměťová stopa není v mozku lokalizována na jednom místě, ale že je distribuována ve více strukturách, a že příliš nezáleží na tom, kde je léze provedena, ale spíš, jak je toto poškození rozsáhlé
 postižení je tím větší, čím větší je objem hmoty, která je chirurgicky odstraněna")
7 Význam Lashleyho myšlenek Oba zmíněné Lashleyho principy vyvolávaly kontroverze již po svém uvedení. Měly však svůj nezastupitelný význam pro další myšlenkový rozvoj koncepce distribuovaného kódování paměti, mnohočetných paměťových systémů (multiple memories theory). Dnes víme, že v mnoha případech paměťových funkcí distribuce paměťových stop skutečně existuje, avšak na povaze konkrétních mozkových struktur přece jenom záleží. Zajímavost: Lashley např. provedl lézi primární vizuální kůry (oblast V1) a následná neschopnost potkanů řešit úlohu brightness discrimination (rozlišování jasu) jej vedla k chybnému závěru, že tato korová oblast zodpovídá za tento typ učení

8 Richard F. Thompson a studium mrkacího reflexu u králíka Později badatel Richard F. Thompson studoval klasické podmiňování mrkacího reflexu u králíka (eye-blink coniditoning) a prováděl léze cerebella a jiných oblastí, a dospěl k závěru, že mozeček může být sídlem tohoto engramu. Thompson také studovat jádro nc. interpositus lateralis (LIP; hluboké jádro mozečku) a zjistil, že pokud je chemicky inaktivováno, dochází k deficitu v tomto typu podmiňování, ukázal tedy, že LIP je také zapojeno v paměťové stopě tohoto typu chování Zde je důležité zdůraznit, že ačkoliv Thompsonovy závěry byly v podstatě správné, je podmiňování mrkacího reflexu je velmi neodvozený typ, asociativního, avšak nedeklarativního typu paměti, který se vyskytuje i u velmi jednoduchých živočichů (vzpomeňte na podmiňování zatahovacího reflexu u Aplysie).
 a zjistil, že pokud je chemicky inaktivováno, dochází k deficitu v tomto typu podmiňování, ukázal tedy, že LIP je také zapojeno v paměťové stopě")
9 Paměťové stopy v případě deklarativní paměti Tvorba deklarativní paměti zahrnuje komplexní interakci mezi limbickými strukturami (především hipokampem, amygdalou, entorhinální kůrou, subikulem, septálními jádry) a neokortikálními oblastmi. Paměťové stopy jsou zde rovněž distribuovány, ačkoliv mohou být např. během učení v určitých případech distribuovány převážně v hipokampu, viz experiment Mosera a Moserové, kdy částečná hipokampální léze poškodila vybavení v úloze MWM (pokud byla úloha naučena s intaktním celým hipokampem, po částečné lézi nebylo funkční vybavení), avšak zvířete s malou částí hipokampu byla stále schopná se učit.
, avšak zvířete s malou částí hipokampu byla stále")
10 Mnohočetnost paměťových fenoménů Existuje řada paměťových systémů (typů paměti) a paměťových procesů (mechanismů tvorby paměti) Základní molekulární mechnismy změn synaptické plasticity, které jsou podkladem učení a paměti, jsou víceméně univerzální

11 Marrův model formace archikortikálních stop (1971) David Courtnay Marr ( ) britský neurobiolog a psycholog, zabýval se především viděním V r navrhnul autoasociativní model archikortikální (hipokampální) reprezentace paměti, kdy aktivované neurony vytvářely vzájemná spojení na základě zvýšení synaptické účinnosti a následně po vytvoření sítě stačila aktivace části buněk k obnovení funkce celé sítě Model založen na modifikaci synaptických spojení pomocí zkušenosti, umožňoval vybavení paměťové stopy i na základě částečné informace, model se stal východiskem pro mnoho hypotéz o funkci hipokampu, z nichž některé byly potvrzeny. "Simple memory: a theory for archicortex." Phil. Trans. Royal Soc. London, 262:23-81 (1971)

12 Změnu v účinnosti v synaptickém přenosu jako základ paměťových stop Hebbův zákon: When an axon of cell A is near enough to excite a cell B and repeatedly and persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A's efficacy, as one of the cells firing B, is increased. Cells that fire together wire together Donald Hebb Typu synaptické plasticity: LTP, LTD, ale existují i krátkodobé změny v účinnosti, jako např. synaptická facilitace, augmentace, post-tetanická potenciace (pozorovány i na nervosvalovém spojaní) trvají od desítek až stovek ms do řádu desítek minut, z často je za jejich indukci zodpovědné přetrvávající presynaptické zvýšení koncentrace vápníku
 trvají od desítek až stovek ms do řádu desítek minut, z často je za jejich")
13 Evokované potenciály v hipokampu EPSP V hipokampu vzniká populační excitační postsynaptický potenciál v odezvě na dráždění aferentních vláken Population spike Populační excitační postsynaptický potenciál (fepsp) v gyrus dentatus drážděním perforující dráhy (svazku přicházejícího z entorhinální kůry) EPSP (intracelulárně u větších vláken, popř. pomocí techniky terčíkového zámku fepsp (field-epsp) extracelulárně, připadá v úvahu u struktur s pravidelným uspořádáním neuronů do vrstev a lamin viz hipokampus
 extracelulárně, připadá v úvahu u struktur s pravidelným uspořádáním")
14 Dlouhodobá potenciace v hipokampu jako kandidát na neuronální substrát paměťové stopy LTP popsána poprvé v r v hipokampu anestetikovaného králíka dlouhodobé zvýšení EPSP po tetanickém dráždění Později popsána i u volně pohyblivých zvířat, včetně potkanů a myší Obecně se LTP dělí na časnou (E-LTP) a pozdní fázi (L-LTP). E-LTP nezávisí na proteosyntéze, pozdní vyžaduje tvorbu nových bílkovin. Lákavé přirovnání E-LTP ~ krátkodobá paměť L-LTP ~ dlouhodobá paměť Není to tak jednoznačné Baseline Tetanus LTP Bliss and Lømo (1973) J. Physiol. 232:

15 LTP vykazuje specificitu Během dráždění určitého vstupu dochází k potenciaci pouze u tohoto stimulovaného vstupu, a ne u sousedních vstupů (pakliže nejsou tyto vstupy funkčně spřaženy, popř. stimulovány současně viz asociativita a kooperativita)

16 LTP vykazuje asociativitu Associativita představuje fenomén, kdy relativně slabá stimulace na jednom vstupu není dostatečná aby vyvolala LTP, současné silná stimulace na jiném, funkčně asociovaném vstupu vyvolá LTP na obou vstupech. Často je pozorována např.na synapsích Schafferových kolaterál z hipokampální oblasti CA3 do CA1

17 LTP vykazuje kooperativitu LTP může být indukována buď silným tetanickým drážděním jednoho vstupu, nebo kooperativně slabším drážděním více vstupů současně. Pokud je stimulován slabě jen jeden vstup, nedosáhne depolarizace hodnoty potřebné k indukci LTP. Pokud je však takovouto slabší stimulací drážděno více konvergujících vstupů, může dojít k sumaci těchto efektů a následné tvorbě dlouhodobé potenciace. Někteří autoři zastávají názor, že rozdíl mezi kooperativitou a asociativitou je pouze sémantický

18 Mechnismy podmiňující indukci časné LTP Aktivace AMPA receptorů deplarizace odblokuje hořčík z ústí NMDA receptorů Po aktivaci NMDA receptorů vtok Ca 2+ do buňky Ca 2+ se váže na kalmodulin komplex vápník/kalmodulin aktivuje CaMKII (kalcium/kalmodulin-závislá kináza II) Paralelní aktivace adenylylcyklázy tvorba camp aktivace PKA CaMKII - fosforylace AMPA receptorů a inzerce nových AMPA receptorů do postsynaptické membrány CaMKII posléze podstupuje autofosforylaci a stává se konstitutivně aktivní (i po návratu Ca 2+ na normální hladinu) - přechod k udržovací fázi časné LTP
 - přechod k udržovací fázi")
19 Exprese pozdní LTP udržovací fáze Mnoho signálních kaskád (CaMKII, PKC) může konvergovat aktivací extracellular signal-regulated kinase (ERK). Ta je patrně spojovacím článkem mezi časnou a pozdní LTP. ERK fosforyluje řadu proteinů a signálních molekul (jak cytoplazmatických, tak jaderných), včetně transkripčních faktorů. Klíčovou molekulu pro udržovací fázi pozdní LTP je proteinkináza Mzeta (PKMζ) Je to atypická izoforma PKC, která postrádá regulační podjednotku a je trvale aktivní. Tato molekula je syntetizována a je nezbytnou a dostačující podmínkou pro L-LTP Zároveň je tato molekula substrátem dlouhodobé paměti viz později
 Je to atypická izoforma PKC, která postrádá regulační podjednotku a je trvale")
20 V rámci LTP existuje více možností zasílení synaptické účinnosti Inzerce nových receptorů do postsynaptické membrány větší odpovědí na stejné množství neuropřenašeče Zvýšení výlevu prostřednictvím retrográdního signálu nejasné, uvažuje se o NO, popř. extracelulárních adhezních molekulách Rašení nových synaptických knoflíků Ztenčení krčku dendritického trnu změna elektrických vlastností vyšší excitabilita

21 Dlouhodobá deprese - LTD Opak dlouhodobé potenciace, jde o dlouhodobé snížení účinnosti synaptického přenosu následkem repetitivní stimulace presynaptických vstupů Je to vlastně protějšek LTP, z fyziologického hlediska se zdá být intuitivně nezbytný ( what goes up, must go down ) Vyskytuje se např. v hipokampu (nízkofrekvenční stimulace) nebo v Purkyňových buňkách mozečku (silná stimulace)
22 Dlouhodobá deprese - LTD Na synapsích Schafferových kolaterál (spojení hipokampálních oblastí CA3 a CA1) lze vyvolat LTP i LTD tetanickým dráždění, klíčovým faktorem pro to, který fenomén bude indukován, je frakvence dráždění (viz obrázek). Nízkofrekvenční stimulace vede ke vzniku LTD. LTD je odlišná od tzv. synaptické depotenciace, což je de facto návrat LTP na klidovou hladinu. V mozečku se LTD účastní motorického učení.
23 Dlouhodobá deprese - LTD LTD je podmíněna zpravidla aktivací NMDA receptorů (ne vždy), ale o nižší hodnotě, a zvýšením intracelulárního vápníku, avšak na nižší hladinu, než jaká je pozorována u LTP. Někdy se LTD mohou účastnit také metabotropní glutamátové receptory (mglur), popř snad i kanabinoidní receptory (také spřažené s G-proteinem) Indukce LTD zahrnuje aktivaci fosfatáz (defosforylujcíí enzymy), které následně sníží citlivost glutamátových receptorů a tím redukují postsynaptickou odpověď na neuropřenašeč. Může zahrnovat také internalizaci AMPA receptorů z postsynaptické membrány do intracelulárních kompartmentů.
24 Homosynaptická a heterosynaptická LTD Homosynaptická LTD vzniká předchozím drážděním stejné dráhy, kde je posléze sledována amplituda EPSP LTP Homosynaptická LTD Heterosynaptická LTD vzniká tetanickým drážděním odlišného vstupu do neuronu než kde je posléze měřena synaptická odpověď - pozorována poprvé jako korelát LTP v hipokampu, indukcí LTP na jednom vstupu do neuronu došlo k LTD na jiných sousedících synapsích do tohoto neuronu
25 Studium vztahu LTP a učení/paměti Je LTP (popř. LTD méně studovaná) skutečně podkladem učení a paměti?
26 Teoretická koncepce studia vztahu LTP-paměť Zkušenost Změna synaptické účinnnosti (přirozená plasticita) Paměť Tetanizace Změna účinnosti synaptického přenosu (LTP) Zvýšení evokované odpovědi (EPSP) Typy studií zabývajících se vztahem LTP - paměť: Vliv učení na synaptickou sílu a LTP Vliv tetanizace (saturace LTP) na učení Vliv manipulace (např.blokády) synaptické plasticity na učení paměť
27 Manipulace (blokáda) LTP ovlivňuje učení Studie R. Morrise ukázala, že po aplikaci antagonisty NMDA receptorů, látky D,L- AP5, došlo jak k zablokování indukce LTP tak neschopnosti učení ve vodním bludišti Vidíte, že potkani, jimž byl aplikován D,L- AP5 hledali během probe trialu ostrůvek víceméně náhodně, na rozdíl od situace po aplikaci L-AP5 neaktivní izomer. Blokáda indukce LTP, přestože blokuje učení, však automaticky neimplikuje, že LTP je substrátem učení, je možné, že pouze dochází k zablokování nějakých společných mechanismů.
28 Saturace LTP poškozuje učení Původní studie Mosera et al. ukázala, že pokud byli potkani tetanizováni na většině synapsí v hipokampu, tzn. došlo k saturaci LTP, poškodilo to učení a paměť. To naznačuje vztah mezi LTP a učením. Je tu však velké ALE. Tito potkani nebyli před saturací LTP obeznámeni s pravidly úlohy. Moser et al., 1998
29 Intaktní předtrénování pravidlům úlohy odstraní negativní vliv saturace LTP na učení Pokud zvířata podstoupí před saturací LTP tzv. NSP (non-spatial pretraining; neprostorové předtrénování), deficit vzniklý saturací LTP již není pozorován. Indukce LTP tedy v tomto případě spíše zodpovídá za procedurální aspekt úlohy to je právě ten, který si zvířata osvojí během NSP NSP zpravidla realizováno tréninkem potkanů v bazénu bez orientačních bodů (záclona), přičemž poloha ostrůvku se náhodně mění. Zvířata se tedy naučí pravidla úlohy, tzn. že je třeba hledat ostrůvek, že thigmotaxe nikam nevede, pouze se nenaučí, KDE ostrůvek je.
30 Dochází při učení skutečně k indukci LTP? Whitlock JR, Heynen AJ, Shuler MG, Bear MF. Learning induces long-term potentiation in the hippocampus. Science. 2006; 313(5790): Whitlock a jeho kolegové trénovali potkany v úloze pasivního vyhýbání (inhibitory avoidance) a zjistili, že u některých synapsí dochází vlivem učení k indukci LTP U mnoha z měřených neuronů však LTP indukována není. Je to rovněž podpůrným důkazem pro distributed memory encoding
31 Inhibice PKMζ vede k vymizení LTP a k vymazání již ustavené dlouhodobé paměťové stopy Pastalkova E, Serrano P, Pinkhasova D, Wallace E, Fenton AA, Sacktor TC. Storage of spatial information by the maintenance mechanism of LTP. Science. 2006;313(5790): Specifický blokátor PKMζ byl aplikován do hipokampu a zablokoval jak pozdní LTP, tak vybavení naučené informace avšak neovlivní nové učení, tzn. že PKM ζ nehraje patrně roli v indukci časné LTP Prokázáno bylo, že syntéza PKMζ je nezbytnou a dostačující podmínkou pro udržovací fázi pozdní LTP
32 Inhibice proteinkinázy Mzeta
33 Otázky do budoucna Je LTP indukována také jinými typy učení než pasivním vyhýbáním? Maže inhibice PKMζ pomocí peptidu ZIP také jiné druhy paměti než place avoidance, což je kognitivní úloha vysoce závislá na hipokampu? Je toto vymazání paměti patrné i v jiných strukturách v mozku než je hipokampus? Některé určitě, např. chuťovou averzi v inzulárním kortexu, viz: Shema R, Sacktor TC, Dudai Y. Rapid erasure of long-term memory associations in the cortex by an inhibitor of PKM zeta. Science; 317(5840):951-3 Velmi nedávno bylo prokázáno, že u řady typů paměti tento mechanismus funguje, konkrétně pro prostorovou, operantní paměť a klasické podmiňování u savců Serrano P, Friedman EL, Kenney J, Taubenfeld SM, Zimmerman JM, Hanna J, Alberini C, Kelley AE, Maren S, Rudy JW, Yin JC, Sacktor TC, Fenton AA. PKMzeta maintains spatial, instrumental, and classically conditioned long-term memories. PLoS Biol Dec 23;6(12):
34 Některé nedávné nálezy ohledně PKM zeta Inhibice PKMzeta peptidem ZIP byla schopna zablokovat také rozvoj neuroplastické bolesti ZIPping to pain relief: the role (or not) of PKMζ in chronic pain. Price TJ, Ghosh S. Mol Pain Feb 22;9:6. doi: / Review. Inhibice PKMzeta peptidem ZIP byla schopna zablokovat také rozvoj neuroplastické bolesti Inhibice PKMzeta peptidem ZIP byla schopna zablokovat také rozvoj neuroplastické bolesti Inhibition of PKMzeta in nucleus accumbens core abolishes long-term drug reward memory. Li YQ, Xue YX, He YY, Li FQ, Xue LF, Xu CM, Sacktor TC, Shaham Y, Lu L. J Neurosci Apr 6;31(14): doi: /JNEUROSCI Zdá se, že některé senisitiační v neuropatické popleti a sensitiazce na drogy szměny jsou schopny reagovat na inhibici proteinkinázy Mzeta
35 Nejnovější negativní důkaz o PKMzeta a LTP Letos vyšla práce popisující transgenní potkany s konstitutivní a podmíněnou delecí PKMζ a u nich nebylo pozorování žádné zhoršení paměti ani LTP. Naopak aplikace ZIP těmto potkanům vedla k vymazání LTP, zdá se že ZIP tedy blokuje doposud neznámé procesy, kromě aktivity na PKMζ Volk LJ, Bachman JL, Johnson R, Yu Y, Huganir RL. PKM-ζ is not required for hippocampal synaptic plasticity, learning and memory. Nature. 2013;493(7432):420-3
36 Lateralizace paměťové stopy Lateralizace má obecně více významů jednak rozdíly mezi jednotlivými polovinami jednotlivých bilaterálních mozkových struktur (např. mírně rozdílný objem pravého a levého hipokampu, měřený volumetricky (např. u lidí i u potkanů zajímavostí je, že albinotičtí potkani Wistar jsou v hipokampu lateralizováni podobně jako člověk, zatímco pigmentovaná zvířata z kmene Long-Evans jsou hipokampálně lateralizována opačně). Další význam tohoto pojmu lateralizace určité funkce (často u lidí) např pomocí fmri bylo zjištěno, že při určitých mentálních úlohách se äktivuje jedna strana mozku více než druhá (PFC, hipokampus) implikuje, že funkce sídlí ve větší míře v jedné z polovin - lateralita V experimentální rovině např. potkani trénováni naučit se MWM, kterým byl každý den před tréninkem vypnut jeden z hipokampů pomocí TTX (+ zakrytí opačného oka), naučí se také úlohu říká se, že paměťová stopa byla lateralizována do jednoho hipokampu tento typ lateralizace je poměrně málo studován Bureš a Fenton, 90.léta v naší laboratoři Tento typ lateralizace lze snadno detekovat pomocí testu vybavení, kdy vypneme natrénovanou hemisféru a zvířeti pak zbývá k navigaci pouze hemisféra neivní vybavení bylo skutečně poškozeno
37 Interhemisferální transfer paměťové stopy Bureš a Fenton ukázali, že kromě lateralizace může u této stopy docházet také ke spontánnímu či nucenému přesunu paměťové stopy z jedné natrénované hemisféry do druhé. Spontánní transfer je vyvolán zpravidla krátkou expozicí prostředí s oběma hemisférami intaktními Nucený transfer je vyvolán ponecháním obou hemisfér intaktních a odkrytím obou očí
38 Interhipokampální syntéza lateralizovaných engramů Fenton a spol. ukázali, že je možné každou hemisféru natrénovat na 2 orientační body (použili celkem pouze 4 orientační body, ABCD). Levá AB, pravá CD, posléze testovali úspěšnost s oběma hemisférami intaktními Výkonnost AB (stejně jako při osvojení) > pouze AC (syntéza stále vysoko nad hranicí náhody) > AE (s novým, neznámým orientačním bodem; na hranici náhody) Fenton AA, Arolfo MP, Nerad L, Bures J. Interhippocampal synthesis of lateralized place navigation engrams. Hippocampus, 1995 Tento typ experimentů vyžaduje precizní kontrolu orientačních bodů, Fenton a spol tento pokus prováděli v MWM ve tmě, body ABCD byly pouze svítící obrazce.
39 Patofyziologie paměťových stop Amnézie, hypermnézie (Solomon Šereševskijneuropsycholog A. Lurija) Amnézie retrográdní, anterográdní nemusí být abslutní, mohou být gradované Například pacient H.M. (s resekcí MTL včetně hipokampu) trpí anterográdní amnézií a časově gradovanou retrográdní amnézií nepamatuje si události těsně před operací, ale velmi staré vzpomínky zachovány.
Koncepce paměti jako schopnosti modifikovat své chování na základě zkušenosti
 Mechanismy paměti Co se dozvíme dnes? Behaviorální koncept paměti Fyziologie synaptického spojení Mechanismy jednoduchého učení u modelového organismu (Aplysie) Synaptická plasticita Dlouhodobá potenciace
Mechanismy paměti Co se dozvíme dnes? Behaviorální koncept paměti Fyziologie synaptického spojení Mechanismy jednoduchého učení u modelového organismu (Aplysie) Synaptická plasticita Dlouhodobá potenciace
Výběr literatury. Kandel E.R. et al.: Principles of Neural Science, Third edition, Prentice-Hall International Inc., USA, 1991.
 Výběr literatury Kandel E.R. et al.: Principles of Neural Science, Third edition, Prentice-Hall International Inc., USA, 1991. O Keefe J. a Nadel L., Hippocampus as a Cognitive Map. London, Clarendon Press,
Výběr literatury Kandel E.R. et al.: Principles of Neural Science, Third edition, Prentice-Hall International Inc., USA, 1991. O Keefe J. a Nadel L., Hippocampus as a Cognitive Map. London, Clarendon Press,
LTP A LTD MECHANIZMY UČENÍ A PAMĚTI, FARMAKOLOGICKÉ OVLIVNĚNÍ
 LTP A LTD MECHANIZMY UČENÍ A PAMĚTI, FARMAKOLOGICKÉ OVLIVNĚNÍ SOUHRN LTP AND LTD THE MECHANISMS OF LEARNING AND MEMORY AND POSSIBLE PHARMACOLOGICAL INFLUENCING Richard Rokyta Univerzita Karlova, 3. lékařská
LTP A LTD MECHANIZMY UČENÍ A PAMĚTI, FARMAKOLOGICKÉ OVLIVNĚNÍ SOUHRN LTP AND LTD THE MECHANISMS OF LEARNING AND MEMORY AND POSSIBLE PHARMACOLOGICAL INFLUENCING Richard Rokyta Univerzita Karlova, 3. lékařská
do paměťových stop Úvod do učení a paměti Aleš Stuchlík
 I mozek a mysl: zpracování informací a konstrukce světa 2 Svět otištěný do paměťových stop Aleš Stuchlík Úvod do učení a paměti Přežití v každodenním světě často vyžaduje, aby jedinec pružně reagoval na
I mozek a mysl: zpracování informací a konstrukce světa 2 Svět otištěný do paměťových stop Aleš Stuchlík Úvod do učení a paměti Přežití v každodenním světě často vyžaduje, aby jedinec pružně reagoval na
Obecná citlivost neuronů i na chemickou modulaci. Neurony nekomunikují pouze synapticky, ale i mimosynapticky. Informační polévka.
 Šířen ení signálů a synapse Synapse, místa přerušení elektrického vedení. Zpomalení, převod na chemickou řeč. Neurony tedy nekomunikují pouze AP, ale i chemicky. Prostor pro zpracování informací. Plasticita
Šířen ení signálů a synapse Synapse, místa přerušení elektrického vedení. Zpomalení, převod na chemickou řeč. Neurony tedy nekomunikují pouze AP, ale i chemicky. Prostor pro zpracování informací. Plasticita
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti EEG - elektroencefalogram Skalpové EEG Intrakraniální EEG > 1 cm < 1 cm Lokální potenciály Extracelulární akční potenciály ~ 1 mm ~ 1 um EEG - elektroencefalogram
Elektrofyziologické metody a studium chování a paměti EEG - elektroencefalogram Skalpové EEG Intrakraniální EEG > 1 cm < 1 cm Lokální potenciály Extracelulární akční potenciály ~ 1 mm ~ 1 um EEG - elektroencefalogram
Neuroplasticita Celoživotní schopnost nervových buněk mozku stavět, přestavovat, rušit a opravovat svoji tkáň. Celoživotní potenciál mozku
 Neuroplasticita Celoživotní schopnost nervových buněk mozku stavět, přestavovat, rušit a opravovat svoji tkáň. Celoživotní potenciál mozku přizpůsobit se strukturálně i funkčně novým podnětům či změněným
Neuroplasticita Celoživotní schopnost nervových buněk mozku stavět, přestavovat, rušit a opravovat svoji tkáň. Celoživotní potenciál mozku přizpůsobit se strukturálně i funkčně novým podnětům či změněným
Spánek. Neurobiologie chování a paměti. Eduard Kelemen. Národní ústav duševního zdraví, Klecany
 Spánek Neurobiologie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Spánek Spánková stadia a architektura spánku Role spánku při konsolidaci paměti behaviorální studie Role spánku
Spánek Neurobiologie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Spánek Spánková stadia a architektura spánku Role spánku při konsolidaci paměti behaviorální studie Role spánku
Změny v mozku na základě zkušeností
 Změny v mozku na základě zkušeností - vliv prostředí na strukturu a funkci mozku - stárnutí mozku často klesá schopnost těchto změn - Kritická perioda doba, do kdy je ještě vliv prostředí účinný - zlom
Změny v mozku na základě zkušeností - vliv prostředí na strukturu a funkci mozku - stárnutí mozku často klesá schopnost těchto změn - Kritická perioda doba, do kdy je ještě vliv prostředí účinný - zlom
Experimentální studium učení a paměti
 Experimentální studium učení a paměti Kurz z cyklu Pokroky v Neurovědách Aleš Stuchlík & Štěpán Kubík Učení a paměť obecné koncepty, dělení paměti, mechanismy, terminologie (A. Stuchlík) Paměť, učení a
Experimentální studium učení a paměti Kurz z cyklu Pokroky v Neurovědách Aleš Stuchlík & Štěpán Kubík Učení a paměť obecné koncepty, dělení paměti, mechanismy, terminologie (A. Stuchlík) Paměť, učení a
Fyziologie synapsí. Rostislav Tureček. Ústav experimentální medicíny, AVČR Oddělení neurofyziologie sluchu.
 Fyziologie synapsí Rostislav Tureček Ústav experimentální medicíny, AVČR Oddělení neurofyziologie sluchu turecek@biomed.cas.cz Signály v nervovém systému 1) Elektrické 2) Chemické Thomas C. Südhof Nobel
Fyziologie synapsí Rostislav Tureček Ústav experimentální medicíny, AVČR Oddělení neurofyziologie sluchu turecek@biomed.cas.cz Signály v nervovém systému 1) Elektrické 2) Chemické Thomas C. Südhof Nobel
Spánek. kurz Neurobiologie chování a paměti. RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR
 Spánek kurz Neurobiologie chování a paměti RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR Spánek Spánková stadia a spánková architektura Role spánku při konsolidaci
Spánek kurz Neurobiologie chování a paměti RNDr. Eduard Kelemen, Ph.D. Národní ústav duševního zdraví Fyziologický ústav AVČR Spánek Spánková stadia a spánková architektura Role spánku při konsolidaci
Základní buněčné a fyziologické mechanismy paměti. MUDr. Jakub Hort, PhD. Neurologická klinika UK, 2.LF a FN Motol
 Základní buněčné a fyziologické mechanismy paměti MUDr. Jakub Hort, PhD. Neurologická klinika UK, 2.LF a FN Motol Poradna pro poruchy paměti FN Motol SYNDROM DEMENCE poškození paměti + jeden další příznak:
Základní buněčné a fyziologické mechanismy paměti MUDr. Jakub Hort, PhD. Neurologická klinika UK, 2.LF a FN Motol Poradna pro poruchy paměti FN Motol SYNDROM DEMENCE poškození paměti + jeden další příznak:
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti RNDr. Eduard Kelemen, PhD Národní ústav duševního zdraví, Klecany Elektrofyziologické metody studia chování a paměti EEG a LFP théta, gama, pomalé
Elektrofyziologické metody a studium chování a paměti RNDr. Eduard Kelemen, PhD Národní ústav duševního zdraví, Klecany Elektrofyziologické metody studia chování a paměti EEG a LFP théta, gama, pomalé
Neuroanatomie chování a paměti
 Neuroanatomie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Neuroanatomie chování a paměti Historické poznámky Neuronová doktrína Gnostická pole Metody studia lokalizace funkcí
Neuroanatomie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Neuroanatomie chování a paměti Historické poznámky Neuronová doktrína Gnostická pole Metody studia lokalizace funkcí
Fyziologický vývoj mozku v dětském věku
 Fyziologický vývoj mozku v dětském věku MUDr. Zuzana Ludvíková Konference Mensa ČR 19.11.2014 Lidský mozek Obsahuje přes 1000 miliard nervových buněk Pokud pracuje naplno odčerpávají neurony 20% z celkové
Fyziologický vývoj mozku v dětském věku MUDr. Zuzana Ludvíková Konference Mensa ČR 19.11.2014 Lidský mozek Obsahuje přes 1000 miliard nervových buněk Pokud pracuje naplno odčerpávají neurony 20% z celkové
Digitální učební materiál
 Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Nervová soustava Společná pro celou sadu oblast
Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Nervová soustava Společná pro celou sadu oblast
Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK
 Milan Chovanec Ústav fyziologie 2.LF UK") Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK Fyziologie srdce Akční potenciál v srdci (pracovní myokard) Automacie srdeční aktivity a převodní systém Mechanismus
Fyziologie srdce I. (excitace, vedení, kontrakce ) Milan Chovanec Ústav fyziologie 2.LF UK Fyziologie srdce Akční potenciál v srdci (pracovní myokard) Automacie srdeční aktivity a převodní systém Mechanismus
1/50 ROLE HIPOKAMPU V UČENÍ, CHOVÁNÍ A PAMĚTI
 1/50 ROLE HIPOKAMPU V UČENÍ, CHOVÁNÍ A PAMĚTI 2/50 HIPOKAMPUS U HLODAVCŮ U laboratorních potkanů zabírá hipokampus poměrně velkou část mozku Dělení dorzální ventrální Oba oddíly zapojeny v odlišných mozkových
1/50 ROLE HIPOKAMPU V UČENÍ, CHOVÁNÍ A PAMĚTI 2/50 HIPOKAMPUS U HLODAVCŮ U laboratorních potkanů zabírá hipokampus poměrně velkou část mozku Dělení dorzální ventrální Oba oddíly zapojeny v odlišných mozkových
Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy)
 mozek mícha Periferní nervový systém (nervy)") Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Vybrané typy chování, učení a typy paměti závislé na hipokampu
 Vybrané typy chování, učení a typy paměti závislé na hipokampu Hipokampus u hlodavců U laboratorních potkanů zabírá hipokampus poměrně velkou část mozku Dělení dorzální ventrální Oba oddíly zapojeny v
Vybrané typy chování, učení a typy paměti závislé na hipokampu Hipokampus u hlodavců U laboratorních potkanů zabírá hipokampus poměrně velkou část mozku Dělení dorzální ventrální Oba oddíly zapojeny v
Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie
 Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie Záznam elektrické aktivity mozku Buněčné mechanizmy vzniku Principy registrace Základní vlastnosti Experimentální využití
Elektrofyziologie - využití při studiu neuronálních mechanizmů paměti a epilepsie Záznam elektrické aktivity mozku Buněčné mechanizmy vzniku Principy registrace Základní vlastnosti Experimentální využití
Neuroanatomie chování a paměti
 Neuroanatomie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Fyziologický ústav AVČR, Praha https://www.youtube.com/watch?v=ami3hbgrj6o Neuroanatomie chování a paměti Neuronová
Neuroanatomie chování a paměti Eduard Kelemen Národní ústav duševního zdraví, Klecany Fyziologický ústav AVČR, Praha https://www.youtube.com/watch?v=ami3hbgrj6o Neuroanatomie chování a paměti Neuronová
Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy)
 mozek mícha Periferní nervový systém (nervy)") Buňka Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Buňka Neuron Nervová soustava Centrální nervový systém (CNS) mozek mícha Periferní nervový systém (nervy) Základní stavební jednotky Neuron přenos a zpracování informací Gliové buňky péče o neurony, metabolická,
Intracelulární Ca 2+ signalizace
 Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
Intracelulární Ca 2+ signalizace Vytášek 2009 Ca 2+ je universální intracelulární signalizační molekula (secondary messenger), která kontroluje řadu buměčných metabolických a vývojových cest intracelulární
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Mechanismy hormonální regulace metabolismu. Vladimíra Kvasnicová
 Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Mechanismy hormonální regulace metabolismu Vladimíra Kvasnicová Osnova semináře 1. Obecný mechanismus působení hormonů (opakování) 2. Příklady mechanismů účinku vybraných hormonů na energetický metabolismus
Šíření signálů a synapse
 Šíření signálů a synapse Šíření signálů a synapse Synapse, místa přerušení elektrického vedení. AP a místní potenciály. Zpomalení, převod na chemickou řeč. Neurony tedy nekomunikují pouze AP, ale i chemicky.
Šíření signálů a synapse Šíření signálů a synapse Synapse, místa přerušení elektrického vedení. AP a místní potenciály. Zpomalení, převod na chemickou řeč. Neurony tedy nekomunikují pouze AP, ale i chemicky.
Nervová soustává č love ká, neuron r es ení
 Nervová soustává č love ká, neuron r es ení Pracovní list Olga Gardašová VY_32_INOVACE_Bi3r0110 Nervová soustava člověka je pravděpodobně nejsložitěji organizovaná hmota na Zemi. 1 cm 2 obsahuje 50 miliónů
Nervová soustává č love ká, neuron r es ení Pracovní list Olga Gardašová VY_32_INOVACE_Bi3r0110 Nervová soustava člověka je pravděpodobně nejsložitěji organizovaná hmota na Zemi. 1 cm 2 obsahuje 50 miliónů
NEUROPLASTICITA PLASTICITA MOZKU A MOTORICKÉ UČENÍ NEUROPLASTICITA 10.10.2013 NEUROPLASTICITA NEUROPLASTICITA - HISTORIE
 NEUROPLASTICITA PLASTICITA MOZKU A MOTORICKÉ UČENÍ Je schopnost adaptace, přestavby a reorganizace CNS v jeho molekulární formě a funkci. IMPLIKACE PRO NEUROREHABILITACI Bc. Hana Kafková RHC odd.kn Liberec
NEUROPLASTICITA PLASTICITA MOZKU A MOTORICKÉ UČENÍ Je schopnost adaptace, přestavby a reorganizace CNS v jeho molekulární formě a funkci. IMPLIKACE PRO NEUROREHABILITACI Bc. Hana Kafková RHC odd.kn Liberec
Neurorehabilitační péče po CMP
 Neurorehabilitační péče po CMP As. MUDr. Martina Hoskovcová Rehabilitační oddělení Neurologické kliniky 1. LF UK a VFN v Praze Ucelená rehabilitace výcvik nebo znovuzískání co možná nejvyššího stupně funkčních
Neurorehabilitační péče po CMP As. MUDr. Martina Hoskovcová Rehabilitační oddělení Neurologické kliniky 1. LF UK a VFN v Praze Ucelená rehabilitace výcvik nebo znovuzískání co možná nejvyššího stupně funkčních
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
(VIII.) Časová a prostorová sumace u kosterního svalu. Fyziologický ústav LF MU, 2016 Jana Svačinová
 Časová a prostorová sumace u kosterního svalu. Fyziologický ústav LF MU, 2016 Jana Svačinová") (VIII.) Časová a prostorová sumace u kosterního svalu Fyziologický ústav LF MU, 2016 Jana Svačinová Kontrakce příčně pruhovaného kosterního svalu Myografie metoda umožňující registraci kontrakce svalů
(VIII.) Časová a prostorová sumace u kosterního svalu Fyziologický ústav LF MU, 2016 Jana Svačinová Kontrakce příčně pruhovaného kosterního svalu Myografie metoda umožňující registraci kontrakce svalů
Metodika studia chování a paměti u zvířat
 Metodika studia chování a paměti u zvířat Vybrané metodické přístupy ke studiu různých typů chování, učení a paměťových fenoménů, především u laboratorních hlodavců Metodika studia chování a paměti u zvířat
Metodika studia chování a paměti u zvířat Vybrané metodické přístupy ke studiu různých typů chování, učení a paměťových fenoménů, především u laboratorních hlodavců Metodika studia chování a paměti u zvířat
Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách
 Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách Difuze Vyrovnávání koncentrací látek na základě náhodného pohybu Osmóza (difuze rozpouštědla) Dva roztoky o rúzné koncentraci oddělené
Membránový potenciál, zpracování a přenos signálu v excitabilních buňkách Difuze Vyrovnávání koncentrací látek na základě náhodného pohybu Osmóza (difuze rozpouštědla) Dva roztoky o rúzné koncentraci oddělené
9. Léčiva CNS - úvod (1)
") 9. Léčiva CNS - úvod (1) se se souhlasem souhlasem autora autora ál školy koly -techlogic techlogické Jeho Jeho žit bez bez souhlasu souhlasu autora autora je je ázá Nervová soustava: Centrální nervový
9. Léčiva CNS - úvod (1) se se souhlasem souhlasem autora autora ál školy koly -techlogic techlogické Jeho Jeho žit bez bez souhlasu souhlasu autora autora je je ázá Nervová soustava: Centrální nervový
Bp1252 Biochemie. #11 Biochemie svalů
 Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
Bp1252 Biochemie #11 Biochemie svalů Úvod Charakteristickou funkční vlastností svalu je schopnost kontrakce a relaxace Kontrakce následuje po excitaci vzrušivé buněčné membrány je přímou přeměnou chemické
Projekt FRVŠ 745/2011. Modernizace výukové laboratoře pro studium patofyziologie centrálního nervového systému. Doc. MUDr. František Vožeh, CSc.
 Projekt FRVŠ 745/2011 Modernizace výukové laboratoře pro studium patofyziologie centrálního nervového systému Řešitel: Doc. MUDr. František Vožeh, CSc. Teoretický základ a principy vyšetření úlekové reakce
Projekt FRVŠ 745/2011 Modernizace výukové laboratoře pro studium patofyziologie centrálního nervového systému Řešitel: Doc. MUDr. František Vožeh, CSc. Teoretický základ a principy vyšetření úlekové reakce
Etiologie epilepsie. Epilepsie nevychází z centra jizvy nebo postmalatické pseudocysty, ale spíše z jejího okraje, kde přežívají poškozené neurony.
 Epilepsie Epilepsie Chronické mozkové onemocnění charakterizované opakujícím se výskytem (nevyprovokovaných) epileptických záchvatů. Ojedinělý epileptický záchvat může být vyprovokován i u člověka bez
Epilepsie Epilepsie Chronické mozkové onemocnění charakterizované opakujícím se výskytem (nevyprovokovaných) epileptických záchvatů. Ojedinělý epileptický záchvat může být vyprovokován i u člověka bez
Zpracování informace v NS Senzorická fyziologie
 Zpracování informace v NS Senzorická fyziologie doc. MUDr. Markéta Bébarová, Ph.D. Fyziologický ústav, Lékařská fakulta, Masarykova univerzita Tato prezentace obsahuje pouze stručný výtah nejdůležitějších
Zpracování informace v NS Senzorická fyziologie doc. MUDr. Markéta Bébarová, Ph.D. Fyziologický ústav, Lékařská fakulta, Masarykova univerzita Tato prezentace obsahuje pouze stručný výtah nejdůležitějších
Vlastnosti neuronových sítí. Zdeněk Šteffek 2. ročník 2. LF UK v Praze
 Vlastnosti neuronových sítí Zdeněk Šteffek 2. ročník 2. LF UK v Praze 7. 3. 2011 Obsah Neuronální pooly Divergence Konvergence Prolongace signálu, kontinuální a rytmický signál Nestabilita a stabilita
Vlastnosti neuronových sítí Zdeněk Šteffek 2. ročník 2. LF UK v Praze 7. 3. 2011 Obsah Neuronální pooly Divergence Konvergence Prolongace signálu, kontinuální a rytmický signál Nestabilita a stabilita
Senzorická fyziologie
 Senzorická fyziologie Čití - proces přenosu informace o aktuálním stavu vnitřního prostředí a zevního okolí do formy signálů v CNS Vnímání (percepce) - subjektivní vědomá interpretace těchto signálů na
Senzorická fyziologie Čití - proces přenosu informace o aktuálním stavu vnitřního prostředí a zevního okolí do formy signálů v CNS Vnímání (percepce) - subjektivní vědomá interpretace těchto signálů na
Regulace metabolických drah na úrovni buňky
 Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
Regulace metabolických drah na úrovni buňky EB Obsah přednášky Obecné principy regulace metabolických drah na úrovni buňky regulace zajištěná kompartmentací metabolických dějů změna absolutní koncentrace
PATOFYZIOLOGIE UČENÍ A PAMĚTI. Ústav patologické fyziologie LF UK v Plzni
 PATOFYZIOLOGIE UČENÍ A PAMĚTI Ústav patologické fyziologie LF UK v Plzni 1 Učení = změna chování na základě předchozí zkušenosti = vstup do paměti = postupné vytváření paměťové stopy opakováním podnětů
PATOFYZIOLOGIE UČENÍ A PAMĚTI Ústav patologické fyziologie LF UK v Plzni 1 Učení = změna chování na základě předchozí zkušenosti = vstup do paměti = postupné vytváření paměťové stopy opakováním podnětů
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
Struktura a funkce biomakromolekul KBC/BPOL 10. Struktury signálních komplexů Ivo Frébort Typy hormonů Steroidní hormony deriváty cholesterolu, regulují metabolismus, osmotickou rovnováhu, sexuální funkce
10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální
 10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální iktální periodické Evokované potenciály sluchové (AEP) zrakové
10. PŘEDNÁŠKA 27. dubna 2017 Artefakty v EEG Abnormální EEG abnormality základní aktivity paroxysmální abnormality epileptiformní interiktální iktální periodické Evokované potenciály sluchové (AEP) zrakové
Neurobiologie chování a paměti
 Neurobiologie chování a paměti Semestrální přednáška na katedře Fyziologie živočichů PřF UK. 3 kredity, zakončená písemnou zkouškou Přednášející: Aleš Stuchlík, Hosté: Eduard Kelemen Kamil Vlček Štěpán
Neurobiologie chování a paměti Semestrální přednáška na katedře Fyziologie živočichů PřF UK. 3 kredity, zakončená písemnou zkouškou Přednášející: Aleš Stuchlík, Hosté: Eduard Kelemen Kamil Vlček Štěpán
Elektrofyziologické metody a studium chování a paměti
 Elektrofyziologické metody a studium chování a paměti (EEG) oscilace v mozku Théta rytmus Gamma rytmus Sharp waves ripples Pomalé oscilace Spánková vřetena Hans Berger První nahrávání EEG u lidí (1924)
Elektrofyziologické metody a studium chování a paměti (EEG) oscilace v mozku Théta rytmus Gamma rytmus Sharp waves ripples Pomalé oscilace Spánková vřetena Hans Berger První nahrávání EEG u lidí (1924)
Buněčné a molekulární mechanismy paměti. Vyšší kognitivní funkce
 Buněčné a molekulární mechanismy paměti Vyšší kognitivní funkce.. Mozkový kmen prodloužená mícha, most a střední mozek Funkce: koordinace a přenos informace mezi mozkovými centry Prodloužená mícha - centra
Buněčné a molekulární mechanismy paměti Vyšší kognitivní funkce.. Mozkový kmen prodloužená mícha, most a střední mozek Funkce: koordinace a přenos informace mezi mozkovými centry Prodloužená mícha - centra
NEUROBIOLOGIE CHOVÁNÍ A PAMĚTI
 NEUROBIOLOGIE CHOVÁNÍ A PAMĚTI 1/66 Semestrální přednáška na katedře Fyziologie živočichů PřFUK. 3 kredity, zakončená písemnou zkouškou (25 otázek, multiple choice) Přednášející: Aleš Stuchlík, Eduard
NEUROBIOLOGIE CHOVÁNÍ A PAMĚTI 1/66 Semestrální přednáška na katedře Fyziologie živočichů PřFUK. 3 kredity, zakončená písemnou zkouškou (25 otázek, multiple choice) Přednášející: Aleš Stuchlík, Eduard
kognitivního deficitu schizofrenie (repetitivní transkraniáln lní magnetická stimulace)
") Nefarmakologické přístupy ovlivnění kognitivního deficitu schizofrenie (repetitivní transkraniáln lní magnetická stimulace) Radovan PřikrylP Psychiatrická klinika LF MU a FN Brno 2009 Obsah sdělen lení
Nefarmakologické přístupy ovlivnění kognitivního deficitu schizofrenie (repetitivní transkraniáln lní magnetická stimulace) Radovan PřikrylP Psychiatrická klinika LF MU a FN Brno 2009 Obsah sdělen lení
Prostorová navigace, kognitivní kontrola, flexibilita chování a jejich vztah k neurodegeneraci
 Prostorová navigace, kognitivní kontrola, flexibilita chování a jejich vztah k neurodegeneraci Aleš Stuchlík Karel Valeš Štěpán n Kubík Fyziologický ústav AV ČR Oddělen lení neurofyziologie paměti Tématická
Prostorová navigace, kognitivní kontrola, flexibilita chování a jejich vztah k neurodegeneraci Aleš Stuchlík Karel Valeš Štěpán n Kubík Fyziologický ústav AV ČR Oddělen lení neurofyziologie paměti Tématická
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
Spánek, anestézie, paměťové stopy.
 Spánek, anestézie, paměťové stopy. Radomir Čumlivski Orthopaedic Hospital Vienna, Austria Dept. of Anaesthesia 9. Křivánkovy dny Pardubice 2015 Je praktické představovat si účinek celkových anestetik na
Spánek, anestézie, paměťové stopy. Radomir Čumlivski Orthopaedic Hospital Vienna, Austria Dept. of Anaesthesia 9. Křivánkovy dny Pardubice 2015 Je praktické představovat si účinek celkových anestetik na
Poruchy paměti, mírná kognitivní porucha, demence. Jakub Hort, Jan Laczó
 Poruchy paměti, mírná kognitivní porucha, demence Jakub Hort, Jan Laczó Kognitivní funkce paměť, řeč, pozornost, exekutivní, gnostické, praktické, visuo-konstruktivní a další MoCA test - senzitivní Senzitivita
Poruchy paměti, mírná kognitivní porucha, demence Jakub Hort, Jan Laczó Kognitivní funkce paměť, řeč, pozornost, exekutivní, gnostické, praktické, visuo-konstruktivní a další MoCA test - senzitivní Senzitivita
Toxikologie PřF UK, ZS 2016/ Toxikodynamika I.
 Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
Toxikodynamika toxikodynamika (řec. δίνευω = pohánět, točit) interakce xenobiotika s cílovým místem (buňkou, receptorem) biologická odpověď jak xenobiotikum působí na organismus toxický účinek nespecifický
VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ
 FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
FUNKCE PROTEINŮ 1 VÝZNAM FUNKCE PROTEINŮ V MEDICÍNĚ Příklad: protein: dystrofin onemocnění: Duchenneova svalová dystrofie 2 3 4 FUNKCE PROTEINŮ: 1. Vztah struktury a funkce proteinů 2. Rodiny proteinů
Neurobiologie chování a paměti
 Neurobiologie chování a paměti Semestrální přednáška na katedře Fyziologie živočichů PřF UK. 3 kredity, zakončená zkouškou Přednášející: Aleš Stuchlík + hosté Odd. neurofyziologie paměti FgÚ AV ČR Obsah
Neurobiologie chování a paměti Semestrální přednáška na katedře Fyziologie živočichů PřF UK. 3 kredity, zakončená zkouškou Přednášející: Aleš Stuchlík + hosté Odd. neurofyziologie paměti FgÚ AV ČR Obsah
PAMĚŤ. Definice: Schopnost vštípit, uchovat, vybavit, rozpoznat, event. zapomenout informace Nejednotný proces několik druhů dělení:
 Paměť Emoce Mnestické funkce PAMĚŤ Definice: Schopnost vštípit, uchovat, vybavit, rozpoznat, event. zapomenout informace Nejednotný proces několik druhů dělení: Dle smyslových analyzátorů (zraková, sluchová,
Paměť Emoce Mnestické funkce PAMĚŤ Definice: Schopnost vštípit, uchovat, vybavit, rozpoznat, event. zapomenout informace Nejednotný proces několik druhů dělení: Dle smyslových analyzátorů (zraková, sluchová,
Rozdělení svalových tkání: kosterní svalovina (příčně pruhované svaly) hladká svalovina srdeční svalovina (myokard)
 hladká svalovina srdeční svalovina (myokard)") Fyziologie svalstva Svalstvo patří ke vzrušivým tkáním schopnost kontrakce a relaxace veškerá aktivní tenze a aktivní pohyb (cirkulace krve, transport tráveniny, řeč, mimika, lidská práce) 40% tělesné
Fyziologie svalstva Svalstvo patří ke vzrušivým tkáním schopnost kontrakce a relaxace veškerá aktivní tenze a aktivní pohyb (cirkulace krve, transport tráveniny, řeč, mimika, lidská práce) 40% tělesné
Řízení svalového tonu Martina Hoskovcová
 Řízení svalového tonu Martina Hoskovcová Neurologická klinika a Centrum klinických neurověd Universita Karlova v Praze, 1. lékařská fakulta a Všeobecná fakultní nemocnice v Praze Svalový tonus Reflexně
Řízení svalového tonu Martina Hoskovcová Neurologická klinika a Centrum klinických neurověd Universita Karlova v Praze, 1. lékařská fakulta a Všeobecná fakultní nemocnice v Praze Svalový tonus Reflexně
Hořčík. Příjem, metabolismus, funkce, projevy nedostatku
 Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Hořčík Příjem, metabolismus, funkce, projevy nedostatku Příjem a pohyb v rostlině Příjem jako ion Mg 2+, pasivní, iont. kanály Mobilní ion v xylému i ve floému, možná retranslokace V místě funkce vázán
Paměť. u lidí, neuropsychologie paměti. 20. dubna 2010
 Paměť u lidí, neuropsychologie paměti Kamil Vlček 20. dubna 2010 Co je to paměť ěť? Změna chování na základě předchozí zkušenosti Schopnost uložit, uchovat a vybavit informaci Hypotetický sklad informací
Paměť u lidí, neuropsychologie paměti Kamil Vlček 20. dubna 2010 Co je to paměť ěť? Změna chování na základě předchozí zkušenosti Schopnost uložit, uchovat a vybavit informaci Hypotetický sklad informací
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU
 LYMFOCYTŮ PREZENTACE ANTIGENU") PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
PREZENTACE ANTIGENU A REGULACE NA ÚROVNI Th (A DALŠÍCH) LYMFOCYTŮ PREZENTACE ANTIGENU Podstata prezentace antigenu (MHC restrikce) byla objevena v roce 1974 V současnosti je zřejmé, že to je jeden z klíčových
LIMBICKÝ PŘEDNÍ MOZEK A AMYGDALÁRNÍ JÁDRA
 LIMBICKÝ PŘEDNÍ MOZEK A AMYGDALÁRNÍ JÁDRA Účast ve vytváření nejrudimentálnějších a nejzákladnějších lidských emocí zahrnujících strach,sexuální touhu, záchvat zuřivosti, náboženskou extázi nebo bazální
LIMBICKÝ PŘEDNÍ MOZEK A AMYGDALÁRNÍ JÁDRA Účast ve vytváření nejrudimentálnějších a nejzákladnějších lidských emocí zahrnujících strach,sexuální touhu, záchvat zuřivosti, náboženskou extázi nebo bazální
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY
 PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
PŘENOS SIGNÁLU DO BUŇKY, MEMBRÁNOVÉ RECEPTORY 1 VÝZNAM MEMBRÁNOVÝCH RECEPTORŮ V MEDICÍNĚ Příklad: Membránové receptory: adrenergní receptory (receptory pro adrenalin a noradrenalin) Funkce: zprostředkování
Kognitivní poruchy u RS. Eva Havrdová 1.LF UK a VFN
 Kognitivní poruchy u RS Eva Havrdová 1.LF UK a VFN Celkový přehled Výskyt kognitivních poruch: 43 70 % pacientů s RS ve všech stadiích choroby Lépe koreluje s progresí nemoci, atrofií CNS Důležitá determinanta
Kognitivní poruchy u RS Eva Havrdová 1.LF UK a VFN Celkový přehled Výskyt kognitivních poruch: 43 70 % pacientů s RS ve všech stadiích choroby Lépe koreluje s progresí nemoci, atrofií CNS Důležitá determinanta
Glykolýza Glukoneogeneze Regulace. Alice Skoumalová
 Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Glykolýza Glukoneogeneze Regulace Alice Skoumalová Metabolismus glukózy - přehled: 1. Glykolýza Glukóza: Univerzální palivo pro buňky Zdroje: potrava (hlavní cukr v dietě) zásoby glykogenu krev (homeostáza
Vybrané parametry neurodegenerace u animálních modelů i lidí. Psychiatrické centrum Praha
 Vybrané parametry neurodegenerace u animálních modelů i lidí Psychiatrické centrum Praha RNDr. Jan Říčný MUDr. Aleš Bartoš MUDr. Anna Kubešová Mgr. Michala Kolářová Mgr. Lenka Hromádková MUDr. Anna Kubešová
Vybrané parametry neurodegenerace u animálních modelů i lidí Psychiatrické centrum Praha RNDr. Jan Říčný MUDr. Aleš Bartoš MUDr. Anna Kubešová Mgr. Michala Kolářová Mgr. Lenka Hromádková MUDr. Anna Kubešová
VYŠETŘENÍ NERVOVÉHO SYSTÉMU. seminář z patologické fyziologie
 VYŠETŘENÍ NERVOVÉHO SYSTÉMU seminář z patologické fyziologie Osnova Morfologické vyšetřovací metody (zobrazovací diagnostika) 1 Počítačová (výpočetní) tomografie 2 Pozitronová emisní tomografie (PET) 3
VYŠETŘENÍ NERVOVÉHO SYSTÉMU seminář z patologické fyziologie Osnova Morfologické vyšetřovací metody (zobrazovací diagnostika) 1 Počítačová (výpočetní) tomografie 2 Pozitronová emisní tomografie (PET) 3
Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem. MUDr.
 Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem MUDr. Tomáš Veverka Neurologická klinika Lékařské fakulty Univerzity Palackého a Fakultní
Spasticita jako projev maladaptivní plasticity CNS po ischemické cévní mozkové příhodě a její ovlivnění botulotoxinem MUDr. Tomáš Veverka Neurologická klinika Lékařské fakulty Univerzity Palackého a Fakultní
Ambasadoři přírodovědných a technických oborů. Ing. Michal Řepka Březen - duben 2013
 Ambasadoři přírodovědných a technických oborů Ing. Michal Řepka Březen - duben 2013 Umělé neuronové sítě Proč právě Neuronové sítě? K čemu je to dobré? Používá se to někde v praxi? Úvod Umělé neuronové
Ambasadoři přírodovědných a technických oborů Ing. Michal Řepka Březen - duben 2013 Umělé neuronové sítě Proč právě Neuronové sítě? K čemu je to dobré? Používá se to někde v praxi? Úvod Umělé neuronové
Kosterní svalstvo tlustých a tenkých filament
 Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
Kosterní svalstvo Základní pojmy: Sarkoplazmatické retikulum zásobárna iontů vápníku - depolarizace membrány uvolnění vápníku v blízkosti kontraktilního aparátu vazba na proteiny zajišťující kontrakci
PŘENOS SIGNÁLU V BUŇCE. Nela Pavlíková
 PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
PŘENOS SIGNÁLU V BUŇCE Nela Pavlíková nela.pavlikova@lf3.cuni.cz Odpovědi na otázky Co za ligand aktivuje receptor spřažený s G-proteinem obsahující podjednotku α T? Opsin. Co prochází otevřenými CNGC
ÚLOHY AKTIVNÍHO A PASIVNÍHO VYHÝBÁNÍ SE MÍSTU: SLIBNÉ NÁSTROJE V KOGNITIVNÍCH NEUROVĚDÁCH
 ÚLOHY AKTIVNÍHO A PASIVNÍHO VYHÝBÁNÍ SE MÍSTU: SLIBNÉ NÁSTROJE V KOGNITIVNÍCH NEUROVĚDÁCH ACTIVE AND PASSIVE PLACE AVOIDANCE TASKS: PROMISING TOOLS IN COGNITIVE NEUROSCIENCE Aleš Stuchlík Fyziologický
ÚLOHY AKTIVNÍHO A PASIVNÍHO VYHÝBÁNÍ SE MÍSTU: SLIBNÉ NÁSTROJE V KOGNITIVNÍCH NEUROVĚDÁCH ACTIVE AND PASSIVE PLACE AVOIDANCE TASKS: PROMISING TOOLS IN COGNITIVE NEUROSCIENCE Aleš Stuchlík Fyziologický
Regulace enzymové aktivity
 Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Regulace enzymové aktivity MUDR. MARTIN VEJRAŽKA, PHD. Regulace enzymové aktivity Organismus NENÍ rovnovážná soustava Rovnováha = smrt Život: homeostáza, ustálený stav Katalýza v uzavřené soustavě bez
Neuronové sítě (11. přednáška)
") Neuronové sítě (11. přednáška) Machine Learning Naučit stroje se učit O co jde? Máme model výpočtu (t.j. výpočetní postup jednoznačně daný vstupy a nějakými parametry), chceme najít vhodné nastavení parametrů,
Neuronové sítě (11. přednáška) Machine Learning Naučit stroje se učit O co jde? Máme model výpočtu (t.j. výpočetní postup jednoznačně daný vstupy a nějakými parametry), chceme najít vhodné nastavení parametrů,
Fyziologie svalové činnosti. MUDr. Jiří Vrána
 Fyziologie svalové činnosti MUDr. Jiří Vrána Syllabus 2) Obecný úvod 4) Kosterní svaly a) funkční stavební jednotky b) akční pot., molek. podklad kontrakce, elektromech. spřažení c) sumace, tetanus, závislost
Fyziologie svalové činnosti MUDr. Jiří Vrána Syllabus 2) Obecný úvod 4) Kosterní svaly a) funkční stavební jednotky b) akční pot., molek. podklad kontrakce, elektromech. spřažení c) sumace, tetanus, závislost
*Mléko a mléčné výrobky obsahují řadu bioaktivních
 www.bileplus.cz Mléko a mléčné výrobky obsahují řadu bioaktivních látek (vápník, mastné kyseliny, syrovátka, větvené aminokyseliny) ovlivňující metabolismus tuků spalování tuků Mléčné výrobky a mléčné
www.bileplus.cz Mléko a mléčné výrobky obsahují řadu bioaktivních látek (vápník, mastné kyseliny, syrovátka, větvené aminokyseliny) ovlivňující metabolismus tuků spalování tuků Mléčné výrobky a mléčné
Obecná fyziologie smyslů. Co se děje na membránách.
 Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Korové evokované potenciály jako metoda pro farmakologické studie
 Korové evokované potenciály jako metoda pro farmakologické studie Cortical Evoked Potentials as a Tool for Pharmacological Studies Gregory Tsenov, Pavel Mareš Fyziologický ústav Akademie věd České republiky,
Korové evokované potenciály jako metoda pro farmakologické studie Cortical Evoked Potentials as a Tool for Pharmacological Studies Gregory Tsenov, Pavel Mareš Fyziologický ústav Akademie věd České republiky,
Obecná fyziologie smyslů. Co se děje na membránách.
 Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
Obecná fyziologie smyslů Co se děje na membránách. Svět smyslů úloha mozku. Paralelní dráhy specializované na určitou vlastnost (kvalitu). V rámci dráhy ještě specializace na konkrétní hodnotu. Transformace
PATOFYZIOLOGIE MOZEČKU. Ústav patologické fyziologie LF UK v Plzni
 PATOFYZIOLOGIE MOZEČKU Ústav patologické fyziologie LF UK v Plzni 1 STRUKTURA MOZEČKU Kůra - stratum moleculare (A) - stratum gangliosum (B) - stratum granulosum (C) Bílá hmota Mozečková jádra - nc. dentatus
PATOFYZIOLOGIE MOZEČKU Ústav patologické fyziologie LF UK v Plzni 1 STRUKTURA MOZEČKU Kůra - stratum moleculare (A) - stratum gangliosum (B) - stratum granulosum (C) Bílá hmota Mozečková jádra - nc. dentatus
Monitorování léků. RNDr. Bohuslava Trnková, ÚKBLD 1. LF UK. ls 1
 Monitorování léků RNDr. Bohuslava Trnková, ÚKBLD 1. LF UK ls 1 Mechanismus působení léčiv co látka dělá s organismem sledování účinku léčiva na: - orgánové úrovni -tkáňové úrovni - molekulární úrovni (receptory)
Monitorování léků RNDr. Bohuslava Trnková, ÚKBLD 1. LF UK ls 1 Mechanismus působení léčiv co látka dělá s organismem sledování účinku léčiva na: - orgánové úrovni -tkáňové úrovni - molekulární úrovni (receptory)
Alzheimerova choroba. senility nádoba? Helena Janíčková 8.3.2012, Krásný Ztráty
 Alzheimerova choroba senility nádoba? Helena Janíčková 8.3.2012, Krásný Ztráty Alzheimerova choroba senilita demence stařecká demence další typy demence... Peter Falk Charles Bronson Charlton Heston Marice
Alzheimerova choroba senility nádoba? Helena Janíčková 8.3.2012, Krásný Ztráty Alzheimerova choroba senilita demence stařecká demence další typy demence... Peter Falk Charles Bronson Charlton Heston Marice
OBSAH. 1. ÚVOD il 3. MOZEK JAKO ORGÁNOVÝ ZÁKLAD PSYCHIKY POZORNOST 43
 OBSAH 1. ÚVOD il 1.1 VYMEZENÍ OBECNÉ PSYCHOLOGIE 11 1.2 METODY POUŽÍVANÉ K HODNOCENÍ PSYCHICKÝCH PROCESŮ A FUNKCÍ 12 1.3 DÍLČÍ OBLASTI, NA NĚŽ JE ZAMĚŘENA OBECNÁ PSYCHOLOGIE 14 1.3.1 Psychologie poznávacích
OBSAH 1. ÚVOD il 1.1 VYMEZENÍ OBECNÉ PSYCHOLOGIE 11 1.2 METODY POUŽÍVANÉ K HODNOCENÍ PSYCHICKÝCH PROCESŮ A FUNKCÍ 12 1.3 DÍLČÍ OBLASTI, NA NĚŽ JE ZAMĚŘENA OBECNÁ PSYCHOLOGIE 14 1.3.1 Psychologie poznávacích
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Patofyziologie srdce. 1. Funkce kardiomyocytu. Kontraktilní systém
 Patofyziologie srdce Funkce kardiomyocytu Systolická funkce srdce Diastolická funkce srdce Etiopatogeneze systolické a diastolické dysfunkce levé komory a srdečního selhání 1. Funkce kardiomyocytu Kardiomyocyty
Patofyziologie srdce Funkce kardiomyocytu Systolická funkce srdce Diastolická funkce srdce Etiopatogeneze systolické a diastolické dysfunkce levé komory a srdečního selhání 1. Funkce kardiomyocytu Kardiomyocyty
ENZYMY. RNDr. Lucie Koláčná, Ph.D.
 ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
ENZYMY RNDr. Lucie Koláčná, Ph.D. Enzymy: katalyzátory živé buňky jednoduché nebo složené proteiny Apoenzym: proteinová část Kofaktor: nízkomolekulová neaminokyselinová struktura nezbytně nutná pro funkci
Digitální učební materiál
 Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Nervová soustava Společná pro celou sadu oblast
Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Nervová soustava Společná pro celou sadu oblast
1. ZÁKLADY NEUROBIOLOGY A NEUROCHEMIE Zdeněk Fišar 1.1 Neurony 1.2 Glie 1.3 Membrány 1.3.1 Struktura a funkce 1.3.2 Složení biomembrán 1.3.
 1. ZÁKLADY NEUROBIOLOGY A NEUROCHEMIE 1.1 Neurony 1.2 Glie 1.3 Membrány 1.3.1 Struktura a funkce 1.3.2 Složení biomembrán 1.3.3 Membránový transport 1.3.4 Receptory 1.3.4.1 Regulace 1.3.4.2 Adaptace 1.3.4.3
1. ZÁKLADY NEUROBIOLOGY A NEUROCHEMIE 1.1 Neurony 1.2 Glie 1.3 Membrány 1.3.1 Struktura a funkce 1.3.2 Složení biomembrán 1.3.3 Membránový transport 1.3.4 Receptory 1.3.4.1 Regulace 1.3.4.2 Adaptace 1.3.4.3
The cell biology of rabies virus: using stealth to reach the brain
 The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
The cell biology of rabies virus: using stealth to reach the brain Matthias J. Schnell, James P. McGettigan, Christoph Wirblich, Amy Papaneri Nikola Skoupá, Kristýna Kolaříková, Agáta Kubíčková Historie
Neurobiologie paměti Neurobiology of memory
 Neurobiologie paměti Neurobiology of memory PØEHLEDNÉ ÈLÁNKY Hynie S., Klenerová V. Laboratoø biochemické neurofarmakologie, Ústav lékaøské biochemie, 1. lékaøská fakulta, Univerzita Karlova v Praze, Praha
Neurobiologie paměti Neurobiology of memory PØEHLEDNÉ ÈLÁNKY Hynie S., Klenerová V. Laboratoø biochemické neurofarmakologie, Ústav lékaøské biochemie, 1. lékaøská fakulta, Univerzita Karlova v Praze, Praha
Specifická imunitní odpověd. Veřejné zdravotnictví
 Specifická imunitní odpověd Veřejné zdravotnictví MHC molekuly glykoproteiny exprimovány na všech jaderných buňkách (MHC I) nebo jenom na antigen prezentujících buňkách (MHC II) u lidí označovány jako
Specifická imunitní odpověd Veřejné zdravotnictví MHC molekuly glykoproteiny exprimovány na všech jaderných buňkách (MHC I) nebo jenom na antigen prezentujících buňkách (MHC II) u lidí označovány jako
10. Reflexy a reakční doba.
 1 10. Reflexy a reakční doba. Živé organismy musí registrovat změny ve svém okolí a adekvátně reagovat. Reflexní přizpůsobení zahrnuje receptory, které registrují změnu, sensorické neurony, které posílají
1 10. Reflexy a reakční doba. Živé organismy musí registrovat změny ve svém okolí a adekvátně reagovat. Reflexní přizpůsobení zahrnuje receptory, které registrují změnu, sensorické neurony, které posílají
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL
 STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
STRUKTURNÍ SKUPINY ADHEZIVNÍCH MOLEKUL - INTEGRINY LIGANDY) - SELEKTINY (SACHARIDOVÉ LIGANDY) - ADHEZIVNÍ MOLEKULY IMUNOGLOBULINOVÉ SKUPINY - MUCINY (LIGANDY SELEKTIN - (CD5, CD44, SKUPINA TNF-R AJ.) AKTIVACE
Protinádorová imunita. Jiří Jelínek
 Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Protinádorová imunita Jiří Jelínek Imunitní systém vs. nádor l imunitní systém je poslední přirozený nástroj organismu jak eliminovat vlastní buňky které se vymkly kontrole l do boje proti nádorovým buňkám
Use of dynamic environments in the study of spatial memory
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Obor: Biologie Anna Staňková Využití dynamických prostředí ve výzkumu prostorové paměti Use of dynamic environments in the study
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Obor: Biologie Anna Staňková Využití dynamických prostředí ve výzkumu prostorové paměti Use of dynamic environments in the study
Bunka a bunecné interakce v patogeneze tkánového poškození
 Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
Bunka a bunecné interakce v patogeneze tkánového poškození bunka - stejná genetická výbava - funkce (proliferace, produkce látek atd.) závisí na diferenciaci diferenciace tkán - specializovaná produkce
DUŠEVNÍ PORUCHY A KVALITA PÉČE
 DUŠEVNÍ PORUCHY A KVALITA PÉČE Sborník přednášek a abstrakt VIII. sjezdu Psychiatrické společnosti ČLS JEP s mezinárodní účastí Tribun EU 2010 Pořadatelé sborníku prof. MUDr. Jiří Raboch, DrSc. prim. MUDr.
DUŠEVNÍ PORUCHY A KVALITA PÉČE Sborník přednášek a abstrakt VIII. sjezdu Psychiatrické společnosti ČLS JEP s mezinárodní účastí Tribun EU 2010 Pořadatelé sborníku prof. MUDr. Jiří Raboch, DrSc. prim. MUDr.