cluster clusters cluster cluster hierarchické klastrování: => strom je jedním z grafických znázornění hierarchického klastrování:
|
|
|
- Marek Kovář
- před 5 lety
- Počet zobrazení:
Transkript
1 Fylogenetika

2 klastrovat na základě podobnosti lze cokoliv, vůbec nemusí jít o evoluční záležitosti (= lidstvo potřebuje popisovat a škatulkovat), je to obecný matematický základ pro analýzy dat, data mining, atd. => pro studium evoluce pak vznikl obor fylogenetiky cluster hierarchické klastrování: clusters Clustering: group the data based on the similarities. = příklad z nebiologické praxe rozdělení zákazníků na skupiny dle parametrů nakupování => cílená reklama => strom je jedním z grafických znázornění hierarchického klastrování: cluster cluster
3 idea fylogenetického stromu (tj. evolučního), stromu života: Darwin, 1859: (mmch. jediný obrázek v jeho knize o původu druhů) Heckel, 1879: Tree of Life
4 fylogenetický strom = záznam evoluce zkoumaných taxonů: fylogeneze (to štěpení) vs. fylogenetika (přístup, který to zkoumá) kořen = společný předek všech kořen = společný předek všech
5 Typy fylogenetických stromů: sdělení těchto stromů je identické speciace kladogram (bez časové informace) ultrametrický strom (končí ve stejném čase = molek. hodiny) strom (s délkami větví)
6

 toto jsou v zásadě grafické clustery.")
7 programy pro práci s fylogenetickými stromy (formáty newick, nexus): závorková konvence: FigTree TreeView = A,B, C,D,E = (((A:1,B:1):1,(C:1,D:1):1):1.2,E:3.2) toto jsou v zásadě grafické clustery... = (((A,B),(C,D)),E) přidaná informace délka větví, nebo třeba podpora uzlů (dozvíme se dále)

8 náhled skutečného stromu:
9 rozřešený strom založen na dichotomickém větvení: reálný strom: polytomie => v uzlu, kde je polytomie nemáme dostatek informace abychom byli schopni vyřešit vztahy mezi potomky uzlu; jen velmi vyjímečně může jít o biologický jev (vyvinulo se více druhů najednou z 1 předka = např. adaptivní radiací)
10 znaky např. morfologie, anatomie, fyziologie, atd. může to být cokoliv apomorfie (nově odvozený znak, nese informaci o příbuznosti), funkčně jde o homologii naopak: např. mnohobuněčnost by byla tzv. pleziomorfie, tedy zděděným znakem od předků, který v tomto kontextu nemá žádnou informaci pro rekonstrukci evolucem jelikož ho nesou všichni
11 znaky např. morfologie, anatomie, fyziologie, atd. může to být cokoliv netopýr netopýr přítomnost znaků bránice a srst nám pomůže odhalit to, že křídla nevznikla jednou, ale dvakrát (a tedy že nejsou homologní, ale je to tzv. homoplázie) křídla!!polarizace znaků co je ancestrální víme až poté co známe fylogenezi, založenou i na jiných znacích...
12 volba znaků je důležitá! vynecháme netopýr netopýr křídla v tomto případě námi zvolená sada poskytuje špatnou fylogenetickou informaci, protože je založena na neodhalené homoplázii => a toto riziko je v našich datech vždy!!polarizace znaků co je ancestrální víme až poté co známe fylogenezi, založenou i na jiných znacích...
13 volba znaků je důležitá! netopýr toto je tzv. matice znaků!!polarizace znaků co je ancestrální víme až poté co známe fylogenezi, založenou i na jiných znacích...
14 znaky molekulární = sekvence DNA alignment = seřazení do matice tvorba matice pro následné analýzy = srovnání sekvencí pod sebe... při alignmentu: program hledá nejlepší rozmístění, dostává body: za každou shodnou bázi plusové body, za každou mezeru mínus body, za otevření mezery speciální mínus body... => cílem je získat co nejvíce bodů!
15 metody výpočtu fylogenetických stromů: 1) distanční metody 2) maximální úspornost = parsimonie 3) maximální věrohodnost = likelihood 4) Bayesovské metody výpočtu

16 Distanční metody: UPGMA (Sokal & Michener 1958), Neighbor-Joining (Saitou and Nei, 1987) čistě hierarchické clusterování již částečné zanesení evoluční info o potenciálně odlišné rychlosti změn v různých větvích z DNA či morfologie: příklad

17 distanční metody výpočtu stromů: - výpočet vzdáleností každé sekvence od každé vznikne matice vzdáleností: nejmenší vzdálenost = spojení
18 distanční metody výpočtu stromů:
19 distanční metody výpočtu stromů: => matice se pak znovu přepočítá, králík+člověk se teď berou jako jeden taxon s průměrnou hodnotou
20 distanční metody výpočtu stromů: nezakořeněný strom: -nyní se vezmou druhy s nejmenší vzdáleností a spojí se, mají společnou vzdálenost a pokračuje se, dokud nejsou všechny druhy ve stromu...
21 zakořeněné a nezakořeněné fylogenetické stromy: pro zakořenění je nutné přidat druh, který nepatří do skupiny, tzv. outgroup, tj. zde např. druh, který není obratlovec...
22 zakořeněné a nezakořeněné fylogenetické stromy:
23 zakořeněné a nezakořeněné fylogenetické stromy: zakořeněné stromy: předek současnost předek současnost
24 co se stane, když zvolíme špatný outgroup?
25 co se stane, když zvolíme špatný outgroup?
26 všechno je totéž... rotace uzlů je volná a nemění smysl!
27 Metoda maximální parsimonie = maximální úspornosti - máme 3 zvířata: vlaštovka, netopýr a kočka - bez jakékoliv matice: existují 3 možné způsoby, jak si mohou být příbuzní: - maximální parsimonie = nejmenší počet změn znaků

28 - maximální parsimonie = nejmenší počet změn znaků: zdroj: Jan Zrzavý - evoluce
29 Metoda maximální parsimonie = maximální úspornosti vítězí scénář č. 1 5 změn 7 změn 8 změn
30 2) parsimonie jak distanční metoda, tak metoda maximální parsimonie zde dávají shodný strom evoluční čas

31 ACCTRAN/DELTRAN optimalizace pro situace, kdy je počet záměn stejný, ale jsou různé varianty ACCTRAN = preference změny co nejdříve DELTRAN = změna co nejpozději zpětná změna změna co nejdříve
!/2 7 (7)! = 2 027 025 stromů exponenciální nárůst počtu potenciálních stromů 12 taxonů: (24-3)!/2 10 (10)!")
32 problém: všechny stromy jdou udělat do max. 11 druhů... (2n 3)! 2 n-2 (n-2)! 3 taxony: (6-3)!/2(1)! = 6/2 = 3 stromy 6 taxonů: (12-3)!/2 4 (4)! = 945 stromů heuristický přístup když nelze prohledat všechno: 9 taxonů: (18-3)!/2 7 (7)! = stromů exponenciální nárůst počtu potenciálních stromů 12 taxonů: (24-3)!/2 10 (10)! = stromů => maximální počet, kdy je reálné prohledávat všechny kombinace je 11 taxonů
33 heuristický přístup když nelze prohledat všechno: není tedy možné prohlédnout všechny stromy, spočítat pro ně počty evolučních změn a vybrat ten nejlepší heuristické hledání stromů tj. - vytvoří se náhodný strom, - spočítají se evoluční změny, - náhodně se v něm přehodí dvě větve, - spočítají se evoluční změny - dál postupuje jen ten, který měl méně změn!!! atd. atd. - na konci řady je strom s nejméně změnami výběr náhodného stromu se opakuje několikrát, porovnání výsledných stromů - může být i několik nejlepších stromů - heuristický postup se uplatňuje u všech dalších metod, tj. nejen parsimonie, ale i max. likelihood a Bayesovském přístupu
34 metody výpočtu fylogenetických stromů: 1) distanční metody - na základě distancí, výsledkem 1 strom 2) maximální úspornost = parsimonie více možných stromů, bereme konsensus 3) maximální věrohodnost = likelihood výsledkem 1 strom 4) Bayesovské metody výpočtu výsledkem 1 strom vč. statistické podpory tyto dvě metody jsou statisticky nejodvozenější, používají obecný statistický aparát nevyvinutý přímo pro fylogenetiku - existují myšlenkové školy, které uznávají jen první dvě metody, které mají dle nich jasné evoluční zadání: buď používám znaky a chci dosáhnout nejmenšího počtu změn, nebo dělám vzdálenosti (např. Willi Hennig Society, časopis Cladistics; jde rozhodně o menšinový názor, vadí jim i toto: - metoda 3 a 4 jsou momentálně nejodvozenější a nejčastěji využívané
35 metoda Maximum Likelihood = maximální věrohodnosti = pravděpodobnost pozorování našich dat za předpokladu, že platí náš model, P(D M) model = strom + parametry (více na dalším slidu) příklad s házením mincí: hodím 10x mincí. Jaká je pravděpodobnost (likelihood, L), že mi padne 5 x hlava a 5 x orel? L=(n!/(k!(n-k)!)) p k (1-p) n-k = n = počet hodů k = počet hlav p = pravděpodobnost pádu hlavy vs. orla (0.5) pokud by byla mince nesouměrná a měnila by se pravděpodobnost (p) padnutí hlavy, pak se zmenšuje i likelihood padnutí 5 hlav + 5 orlů paralela k fylogenezi: máme model (strom + parametry evoluce) a zkoumáme jaká je pravděpodobnost, že na ně budou sedět naše data testujeme postupně stromy (analýza tedy vezme 1 strom, spočítá L, pak vezme další, a pokud ten je lepší tak pokračuje s ním, pokud ne tak s původním) počet kroků = počet generací => vyhledáme tímto nejlepší strom N L = L(1) x L(2)... x L(N) = L(j) j=1 výsledný likelihood je součtem všech znaků znak1 = třeba nukleotidová pozice 1
36 evoluční model pro ML (ale i Bayesovskou analýzu): v modelu (paralela cinknutosti mince) jsou matematicky zainkorporovány: 1) pravděpodobnosti jednotlivých záměn (A->T, A->C, A->G, T->C, T->G, C->G), maximálně může být až šest různých pravděpodobností; (= šest různých substitučních typů) 2) poměry jednotlivých bází 3) poměr nevariabilních míst vůči variabilním 4) pravděpodobnost záměny jednotlivých pozic (např. morfologie stejná pro všechny znaky; DNA různá, např. dle pozic v kodónu atd.) 5) korekce distance (vychází i z bodu 3). to vše je vlastně popis evoluce DNA
37 purin pyrimidin purin pyrimidin transice je v datech mnohem častější než transverze, protože molekula je stabilnější a protože nevede tak často ke změně aminokyseliny, tedy zůstává v datech zachována

38 p-distance vs. g-distance pokus o optimální korekci; saturace AATGCCGTATCGCGTTAATTTGAGCGCTTTCGAT AATGCCATATCGCGTTAAGTTGAGCCCTATCGAT AAAGCCATATCGCATTAAGTTGAGCGCTATCGAT tato mutace nebude detekována, protože je zpětná, tedy změnila se na původní stav => saturované sekvence již nesou hodně šumu, saturované pozice pak málo fylogenetické informace. Nejčastěji jsou satuorvány 3. pozice kodónů. Model se snaží korigovat tento jev tím, že předpoví míru saturace
 BT = pravděpodobnost stromu/uzlu při datech")
39 Bayesovská fylogenetika Bayesův teorém (Bayesova věta): Thomas Bayes (18. století) vymyslel statistickou metodu a tzv. Bayesův teorém pro nás uživatele je to mírně modifikovaná forma likelihoodu velmi zjednodušeně: Maximum Likelihood = pravděpodobnost dat ze stromu (modelu) BT = pravděpodobnost stromu/uzlu při datech = > využívá k výpočtu tzv. inverzní pravděpodobnost (také hledá nejlepší strom) P(A B) = P(B A)P(A) P(B) = pravděpodobnost jevu A za předpokladu, že platí B P(strom data) = P(uzel data) = P(data strom) P(strom) P(data) P(data uzel) P(uzel) P(data)
40 Bayesovská fylogenetika Bayesův teorém (Bayesova věta): odbočka = příklad pro vysvětlení Bayesovy věty: Mám 2 pytlíky s kuličkami. Pytlík 1 ( bílý ) obsahuje 3x více bílých, pytlík 2 ( černý ) - 3x více černých kuliček. Vyberu náhodně 1 pytlík. Vytáhnu 5 kuliček (vždy po vytažení každou vrátím). Výsledkem je 4 bílé a 1 černá kulička. Jaká je pravděpodobnost, že mnou vybraný pytlík byl bílý Pytlík (1)? data: 4x bílá, 1x černá otázka: s jakou pravděpodobností to byl bílý pytlík?: výběr pytlíku byl náhodný, tj. pravděpodobnost na počátku = 1/2 a priori = 1/2 P(BílýPytlík data) = P(data BílýPytlík) P(BílýPytlík) P(data) celkový součet všech možností, tj. Pstí pro data jak z bílého tak z černého pytlíku P(BílýPytlík data) = P(BílýPytlík data) = bílá kulička Pst pro data z bílého pytlíku P(data BílýPytlík) P(BílýPytlík) P(data BílýPytlík) P(BílýPytlík) + P(data ČernýPytlík) P(ČernýPytlík) P(data BílýPytlík) 1/2 P(data BílýPytlík) 1/2 + P(data ČernýPytlík) 1/2 P(data BílýPytlík) = 5 3/4 4 1/4 1 = 405/1024 = pst, že nastane pozorovaná sestava (4B+1Č) při tahu z tohoto pytlíku P(data ČernýPytlík) = 5 1/4 4 3/4 1 = 12/1024 = pst, že nastane pozorovaná sestava (4B+1Č) při tahu z tohoto pytlíku P(BílýPytlík data) = černá kulička 405/1024 1/2 405/1024 1/2 + 12/1024 1/2 = (to je odpověď)
41 Bayesovská fylogenetika Bayesův teorém (Bayesova věta): odbočka = příklad pro vysvětlení Bayesovy věty: Mám 2 pytlíky s kuličkami. Pytlík 1 ( bílý ) obsahuje 3x více bílých, pytlík 2 ( černý ) - 3x více černých kuliček. Vyberu náhodně 1 pytlík. Vytáhnu 5 kuliček (vždy po vytažení každou vrátím). Výsledkem je 4 bílé a 1 černá kulička. Jaká je pravděpodobnost, že mnou vybraný pytlík byl bílý Pytlík (1)? data: 4x bílá, 1x černá otázka: s jakou pravděpodobností to byl bílý pytlík?: výběr pytlíku byl náhodný, tj. pravděpodobnost na počátku = 1/2 a priori = 1/2 P(BílýPytlík data) = P(data BílýPytlík) P(BílýPytlík) P(data) celkový součet všech možností, tj. Pstí pro data jak z bílého tak z černého pytlíku P(BílýPytlík data) = Pst pro data z bílého pytlíku P(data BílýPytlík) P(BílýPytlík) P(data BílýPytlík) P(BílýPytlík) + P(data ČernýPytlík) P(ČernýPytlík) celé je to krásná paralela pro naše P(data BílýPytlík) hledání optimálního 1/2 stromu: teď si představte, že P(BílýPytlík data) = pytlíky jsou různé alternativní topologie uzlů (tedy např. různé kombinace druhů v uzlu). P(data BílýPytlík) 1/2 + P(data ČernýPytlík) 1/2 bílá kulička Máme tedy naše data (třeba černá sekvence kulička DNA, nebo sadu morfologických znaků) a ptáme P(data BílýPytlík) = 5 3/4 4 1/4 1 = 405/1024 = pst, že nastane pozorovaná sestava (4B+1Č) při tahu z tohoto pytlíku P(data ČernýPytlík) = 5 1/4 4 3/4 1 = 12/1024 = pst, že nastane pozorovaná sestava (4B+1Č) při tahu z tohoto pytlíku se: jaká je pravděpodobnost, že platí jeden z uzlů na základě našich dat? Analýza jede pro každý uzel ve stromu a 405/1024 pravděpodobnosti 1/2 = likelihoody se pak sčítají... (jen je to o P(BílýPytlík data) = = (odpověď) něco složitější i zde vstupují další parametry evolučního modelu, jako u ML metody) 405/1024 1/2 + 12/1024 1/2
42 heuristické hledání v Bayesovské analýze: hledání provádějí tzv. Markovovy řetězce (Markov-chain Monte Carlo) krok 1: máme 4 řetězce, které se vydaly hledat do krajiny nejlepší strom... hledání je opět heuristické, tedy zkusím strom, spočítám jeho L, zkusím další, posunu se pouze, je-li nový strom lepší... větší likelihood = adaptivní krajina, tedy vizualizace optimálních stromů čím výše, tím vhodněji strom odpovídá datům, algoritmus analýzy krajinu postupně prohledává (a samozřejmě neví, co je kde za kopce a údolí )
43 1 řetězec je studený tzn. konzervativní, posune se pouze nahoru tedy pokud je další strom lepší 3 řetězce jsou teplé tzn. mohou se vrátit i dolů + skáčou náhodně na jiná místa teplé řetězce volají studeného, pokud najdou lepší strom = vyšší vrcholek, než na které se usídlil studený větší likelihood větší likelihood
44 heuristické hledání v Bayesovské analýze: větší likelihood héééj
45 heuristické hledání v Bayesovské analýze: při dostatečném počtu generací (tj. hledacích kroků) najde studený řetězec nejvyšší vrchol v krajině, tj. strom s nejlepším Likelihoodem.!!problém uvíznutí v lokálním maximu je rizikem všech typů analýz, které využívají heuristický přístup (tedy nemají možnost projít všechny potenciální stromy).
46 likelihood dostatečný počet generací + několik nezávislých běhů je důležitý po čase je vrchol nalezen a čím je vyšší než ostatní, tím lépe bude náš výsledek podpořen => tedy Baysovská analýza probíhá vždy na dostatečném počtu generací, které časem konvergují ke shodnému nálezu ; počáteční fázi potom z výsledků vyhazujeme: 2 milióny generací
47 statistické podpory míra důvěryhodnosti topologie; bootstrapování a BPP statistická podpora existence uzlu

48 likelihood statistické podpory BPP = Bayesian Posterior Probability Bayesovská analýza zpracuje všechny stromy v plateau fázi = kolik % stromů obsahuje daný uzel 2 milióny generací
49 statistická podpora bootstrapování využívá se pro metody Maximální parsimonie a Max. Likelihood jaká je statistická podpora jednotlivých uzlů?
50 statistická podpora bootstrapování představte si situaci, kdy je některý uzel podpořen téměř všemi znaky, a jiný jen některými, které zrovna převáží...
51
52
53 1 2 bootstrap1.jpg 3
54 Bootstrap - pseudomatice:
55 Bootstrap výsledný strom: v kolika % z 1000 stromů z pseudomatic se uzel vyskytl? morfologie i DNA některé znaky se opakují a jiné se do pseudomatice nedostanou, během 1000 opakování by mělo padnout dostatečné množství kombinací pokud je topologie (=uzel) založena jen na málo znacích, bude podpora bootstrapu malá (pravděpodobnost, že se vylosují tyto znaky do pseudomatice je menší než když je topologie založena na mnoha znacích napříč datasetem).
56 potenciální problémy: přitahování dlouhých větví - long-branch attraction:
57 evoluční historie není vždy přímočará...
58 molekulární fylogeneze založená na sekvenci DNA různých genů: - máme-li více genů, pak můžeme buď všechny sekvence jednoduše spojit za sebe (=konkatenovat) nebo počítat strom pro každý gen zvlášť - počítat zvlášť je správnější poté se udělá konsensus z jednotlivých genů gene trees vs. species tree:


59 fylogenomika = fylogenetika na celých genomech! topologie se může měnit podél chromozomů Neurospora detekce introgrese a nerekombinujícího úseku v genomu Martin, S. H., & Van Belleghem, S. M. (2017). Exploring evolutionary relationships across the genome using topology weighting. Genetics, 206(1),
. Exploring evolutionary relationships across the genome using topology weighting.")
60 fylogenomika = fylogenetika na celých genomech! topologie se může měnit podél chromozomů Martin, S. H., & Van Belleghem, S. M. (2017). Exploring evolutionary relationships across the genome using topology weighting. Genetics, 206(1),

61 alternativní topologie podpořené různými částmi genomu: Johansson, Frank, et al. "Phylogeography and larval spine length of the dragonfly Leucorhinia dubia in Europe." PloS one12.9 (2017): e
")
: 1817-1828.")
62 hybridizace vede k introgresi a k smíšenému signálu z genomu: celá teorie za fylogenetikou předpokládá, že druhy vznikají bifurkací a pak se nekříží, pak už tedy záleží, co chceme ukázat (tedy - ukázat 1 strom by byl problém) Martin, Simon H., et al. "Genome-wide evidence for speciation with gene flow in Heliconius butterflies." Genome Research23.11 (2013): hybridizace => křížení způsobí, že se genomový signál namíchá, polovina genomu podporuje jinou topologii
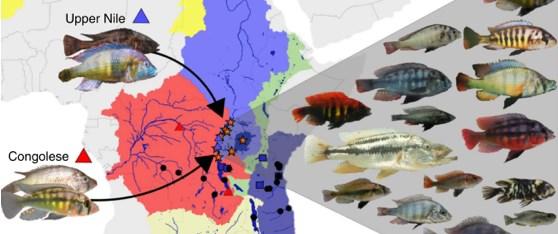
63 co hybridní druhy? nebo dokonce celé množství druhů, které vznikly díky hybridizaci?? - revoluční metody v sekvenaci DNA (next-generation sequencing) nám umožnily studovat celé genomy rutinně, tedy teď začínají vycházet najevo zákonitosti, které jsme neznali... celá druhová diverzita cichlid z jezera Viktoria vznikla pravděpodobně díky počáteční hybridizaci 2 vzdáleně příbuzných linií, z Konga a z Nilu genomy různých druhů jsou různé mozaiky Meier, Joana I., et al. "Ancient hybridization fuels rapid cichlid fish adaptive radiations." Nature Communications 8 (2017).
 nám umožnily studovat celé genomy rutinně, tedy teď začínají vycházet najevo zákonitosti, které jsme neznali.")
 nebudeme nikdy mít jeden klasický fylogenetický strom, ale fylogenetický přístup se stále bude používat např.")
64 co hybridní druhy? nebo dokonce celé množství druhů, které vznikly díky hybridizaci?? - revoluční metody v sekvenaci DNA (next-generation sequencing) nám umožnily studovat celé genomy rutinně, tedy teď začínají vycházet najevo zákonitosti, které jsme neznali... celá druhová diverzita cichlid z jezera Viktoria vznikla pravděpodobně díky počáteční hybridizaci 2 vzdáleně příbuzných linií, z Konga a z Nilu bude asi nutné akceptovat, že pro tyto ryby (a všechny ostatní, u kterých se ukáže něco podobného) nebudeme nikdy mít jeden klasický fylogenetický strom, ale fylogenetický přístup se stále bude používat např. při zkoumání jednotlivých genů a jejich funkcí (a důvodu proč byla vyselektována zrovna tato varianta...), atd. genomy různých druhů jsou různé mozaiky Meier, Joana I., et al. "Ancient hybridization fuels rapid cichlid fish adaptive radiations." Nature Communications 8 (2017).
 a jejich kříženců")
65 schéma fylogeneze citrusů (přírodních forem) a jejich kříženců (vyšlechtěných)
66 fylogenetické sítě = phylogenetic networks zobrazují vztahy ne jako strom (tedy rozdvojováním a společným uzlem vždy pro 2 taxony), ale jako síť, kde jeden uzel může mít více výstupů, a zároveň každý taxon může být zapojen ve více uzlech jsou tam mezistavy, tedy nepozorované, ale předpověděné uzly

67 haplotypová síť: - každá spojnice spojuje haplotypy lišící se jedinou mutací - velikost kruhu = počet jedinců íťové propojení nepozorovaný, ale předpovězený haplotyp fylogeografický vzor u sekavce C. strumicae z Balkánu
68 haplotypová síť lidská mtdna: Gandini, Francesca, et al. "Mapping human dispersals into the Horn of Africa from Arabian Ice Age refugia using mitogenomes." Scientific reports 6 (2016):
69 fylogenetické sítě = phylogenetic networks Gouzelou, Evi, et al. "Genetic diversity and structure in Leishmania infantum populations from southeastern Europe revealed by microsatellite analysis." Parasites & vectors 6.1 (2013): 342.









70 cichlidy z kráterového jezera Barombi Mbo v Kamerunu: Myaka myaka Sarotherodon caroli Sarotherodon linnellii pravděpodobně stále probíhající genový tok nebo velmi mladá speciace situace na počátku evoluce této skupiny nejasná, šlo pravděpodobně o rychlou adaptivní radiaci Sarotherodon lohbergeri Stomatepia mongo Sarotherodon steinbachi Konia eisentrauti Stomatepia pindu speciace se vznikem tří druhů nebo 2 speciace velmi rychle po sobě, nedetekovatelné daty Pungu maclareni Konia dikume RAD-seq (9280 SNPs) Stomatepia mariae SplitsTree4: NeighbourNet
71 potenciální zdroj chybného či zavádějícího signálu genová duplikace (o které nevíme) je tedy kritické nespojovat alfu a betu dohromady, což ale často nevíme! = je nutné dávat pozor když vyrábíme dataset a hledáme v něm homologie
72 některé geny mají velké množství kopií: hemoglobinový cluster tilápie: 7x hemoglobin beta a 12x hemoglobin alfa = těžké určit co je s čím homologní napříč druhy...

73 genová konverze jev probíhající většinou na genových duplikátech v rámci jedince! jde o jakýsi horizontální přenos, takže genetická informace např. mezi velmi starými kopiemi genů (např. vzniklých u předka všech obratlovců) se promíchá... chyba v rekombinaci
 skutečný evoluční čas ideální je fosílie co nejblíže kořenu stromu, jinak jsou odhady s velkými konfidenčními intervaly fosilní taxony jejchž stáří známe")
74 molekulární hodiny, fosílie a ultrametrické stromy běžný strom používá délku větví k znázornění rychlosti evoluce (např. mutací) skutečný evoluční čas ideální je fosílie co nejblíže kořenu stromu, jinak jsou odhady s velkými konfidenčními intervaly fosilní taxony jejchž stáří známe použijeme pro kalibraci, příp. přímo zařadíme do matice

75 díky molekulárním hodinám jsme pak schopni interpretovat například vznik nějakého znaku, atd. zde například určitý vzor v retrotransposonech fosílie
76 jiný typ molekulárních hodin: virová evoluce, koncové větve nekončí stejně chřipkový virus a rezistence na něj
Typy fylogenetických analýz
 Typy fylogenetických analýz Distanční metody: Neighbor-Joining Minimum Evolultion UPGMA,... Maximum Likelihood Bayesian Inference Maximum Parsimony Genetické distance, substituční modely pro výpočet fylogenetických
Typy fylogenetických analýz Distanční metody: Neighbor-Joining Minimum Evolultion UPGMA,... Maximum Likelihood Bayesian Inference Maximum Parsimony Genetické distance, substituční modely pro výpočet fylogenetických
Fylogeneze a diverzita obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Fylogeneze a diverzita obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Charles
Systém a evoluce obratlovců I.Úvod
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Systém a evoluce obratlovců I.Úvod literatura taxonomie a systematika znaky a klasifikace Carl Linné Willy Hennig Literatura 2007
Populační genetika III. Radka Reifová
 Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,
: Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,") Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Pravděpodobnost, náhoda, kostky
 Pravděpodobnost, náhoda, kostky Radek Pelánek IV122, jaro 2015 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností krátké
Pravděpodobnost, náhoda, kostky Radek Pelánek IV122, jaro 2015 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností krátké
Pravděpodobnost, náhoda, kostky
 Pravděpodobnost, náhoda, kostky Radek Pelánek IV122 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností připomenutí, souvislosti
Pravděpodobnost, náhoda, kostky Radek Pelánek IV122 Výhled pravděpodobnost náhodná čísla lineární regrese detekce shluků Dnes lehce nesourodá směs úloh souvisejících s pravděpodobností připomenutí, souvislosti
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
2. Maximální úspornost (Maximum Parsimony, MP)
") 2. Maximální úspornost (Maximum Parsimony, MP) Ze všech metod konstrukce fylogenetických stromů byly donedávna nejpoužívanější metody maximální úspornosti (parsimonie). Důvodem pro jejich mimořádnou oblibu
2. Maximální úspornost (Maximum Parsimony, MP) Ze všech metod konstrukce fylogenetických stromů byly donedávna nejpoužívanější metody maximální úspornosti (parsimonie). Důvodem pro jejich mimořádnou oblibu
Základy fylogenetiky a konstrukce fylogenetických stromů
 EKO/MEM Molekulární ekologie mikroorganismů Základy fylogenetiky a konstrukce fylogenetických stromů Iva Buriánková Katedra ekologie PřF UP Kde vyrostl první fylogenetický strom? Charles Darwin (1809 1882)
EKO/MEM Molekulární ekologie mikroorganismů Základy fylogenetiky a konstrukce fylogenetických stromů Iva Buriánková Katedra ekologie PřF UP Kde vyrostl první fylogenetický strom? Charles Darwin (1809 1882)
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany
: Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany") Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Využití DNA markerů ve studiu fylogeneze rostlin
 Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Využití DNA markerů ve studiu fylogeneze rostlin Ing. Petra VESELÁ Ústav lesnické botaniky, dendrologie a geobiocenologie LDF MENDELU Brno Tento projekt je spolufinancován
 Evoluční genetika KBI/GENE Mgr. Zbyněk Houdek Evoluční teorie Evoluční teorii vyslovil Ch. Darwin v díle O původu druhů (1859), kde ukazoval, že druhy se postupně měnily v dlouhých časových periodách.
Evoluční genetika KBI/GENE Mgr. Zbyněk Houdek Evoluční teorie Evoluční teorii vyslovil Ch. Darwin v díle O původu druhů (1859), kde ukazoval, že druhy se postupně měnily v dlouhých časových periodách.
matematika v biologii: fylogenetika David Černý
 matematika v biologii: fylogenetika David Černý David Černý Úvod Linné (1735), Systema Naturae: živá příroda vykazuje hierarchické uspořádání Darwin (1859), On the Origin of Species: příčinou je společný
matematika v biologii: fylogenetika David Černý David Černý Úvod Linné (1735), Systema Naturae: živá příroda vykazuje hierarchické uspořádání Darwin (1859), On the Origin of Species: příčinou je společný
Modely teorie grafů, min.kostra, max.tok, CPM, MPM, PERT
 PEF ČZU Modely teorie grafů, min.kostra, max.tok, CPM, MPM, PERT Okruhy SZB č. 5 Zdroje: Demel, J., Operační výzkum Jablonský J., Operační výzkum Šubrt, T., Langrová, P., Projektové řízení I. a různá internetová
PEF ČZU Modely teorie grafů, min.kostra, max.tok, CPM, MPM, PERT Okruhy SZB č. 5 Zdroje: Demel, J., Operační výzkum Jablonský J., Operační výzkum Šubrt, T., Langrová, P., Projektové řízení I. a různá internetová
Jak se matematika poučila v biologii
 Jak se matematika poučila v biologii René Kalus IT4Innovations, VŠB TUO Role matematiky v (nejen) přírodních vědách Matematika inspirující a sloužící jazyk pro komunikaci s přírodou V 4 3 r 3 Matematika
Jak se matematika poučila v biologii René Kalus IT4Innovations, VŠB TUO Role matematiky v (nejen) přírodních vědách Matematika inspirující a sloužící jazyk pro komunikaci s přírodou V 4 3 r 3 Matematika
Aplikace DNA markerů v mykologii a molekulárni taxonomii
 Mendelova genetika v příkladech Aplikace DNA markerů v mykologii a molekulárni taxonomii doc. RNDr. Michal Tomšovský, Ph.D., Ústav ochrany lesů a myslivosti, LDF MENDELU, Brno Tento projekt je spolufinancován
Mendelova genetika v příkladech Aplikace DNA markerů v mykologii a molekulárni taxonomii doc. RNDr. Michal Tomšovský, Ph.D., Ústav ochrany lesů a myslivosti, LDF MENDELU, Brno Tento projekt je spolufinancován
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů)
") Počítačováčást 1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů) Pavel Munclinger, Petr Synek 2. fylogenetická
Počítačováčást 1. seznámení s on-line databázemi, nástroji a softwarem (databáze, vyhledání sekvencí, základní manipulace se sekvencemi, navržení primerů) Pavel Munclinger, Petr Synek 2. fylogenetická
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Pravděpodobnost vs. Poměr šancí. Pravděpodobnostní algoritmy: Bayesova věta. Bayesova teorie rozhodování. Bayesova věta (teorém) Vzorec. ...
 Vzorec. ...") ravděpodobnostní algoritmy: Bayesova věta Fantasy is hardly an escape from reality. It is a way of understanding it. LLoyd Alexander ravděpodobnost vs. oměr šancí ravděpodobnost - poměr počtu jedinců surčitým
ravděpodobnostní algoritmy: Bayesova věta Fantasy is hardly an escape from reality. It is a way of understanding it. LLoyd Alexander ravděpodobnost vs. oměr šancí ravděpodobnost - poměr počtu jedinců surčitým
Bayesovské metody. Mnohorozměrná analýza dat
 Mnohorozměrná analýza dat Podmíněná pravděpodobnost Definice: Uvažujme náhodné jevy A a B takové, že P(B) > 0. Podmíněnou pravěpodobností jevu A za podmínky, že nastal jev B, nazýváme podíl P(A B) P(A
Mnohorozměrná analýza dat Podmíněná pravděpodobnost Definice: Uvažujme náhodné jevy A a B takové, že P(B) > 0. Podmíněnou pravěpodobností jevu A za podmínky, že nastal jev B, nazýváme podíl P(A B) P(A
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Vícerozměrné statistické metody
 Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Paleogenetika člověka
 Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Kdo je náš předek?
Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Budeme se snažit najít odpověď na možná nejstarší otázku člověka: Kdo jsme a odkud pocházíme? Kdo je náš předek?
VYUŽITÍ PRAVDĚPODOBNOSTNÍ METODY MONTE CARLO V SOUDNÍM INŽENÝRSTVÍ
 VYUŽITÍ PRAVDĚPODOBNOSTNÍ METODY MONTE CARLO V SOUDNÍM INŽENÝRSTVÍ Michal Kořenář 1 Abstrakt Rozvoj výpočetní techniky v poslední době umožnil také rozvoj výpočetních metod, které nejsou založeny na bázi
VYUŽITÍ PRAVDĚPODOBNOSTNÍ METODY MONTE CARLO V SOUDNÍM INŽENÝRSTVÍ Michal Kořenář 1 Abstrakt Rozvoj výpočetní techniky v poslední době umožnil také rozvoj výpočetních metod, které nejsou založeny na bázi
Vytěžování znalostí z dat
 Pavel Kordík, Jan Motl (ČVUT FIT) Vytěžování znalostí z dat BI-VZD, 2012, Přednáška 7 1/27 Vytěžování znalostí z dat Pavel Kordík, Jan Motl Department of Computer Systems Faculty of Information Technology
Pavel Kordík, Jan Motl (ČVUT FIT) Vytěžování znalostí z dat BI-VZD, 2012, Přednáška 7 1/27 Vytěžování znalostí z dat Pavel Kordík, Jan Motl Department of Computer Systems Faculty of Information Technology
Grafy. doc. Mgr. Jiří Dvorský, Ph.D. Katedra informatiky Fakulta elektrotechniky a informatiky VŠB TU Ostrava. Prezentace ke dni 13.
 Grafy doc. Mgr. Jiří Dvorský, Ph.D. Katedra informatiky Fakulta elektrotechniky a informatiky VŠB TU Ostrava Prezentace ke dni 13. března 2017 Jiří Dvorský (VŠB TUO) Grafy 104 / 309 Osnova přednášky Grafy
Grafy doc. Mgr. Jiří Dvorský, Ph.D. Katedra informatiky Fakulta elektrotechniky a informatiky VŠB TU Ostrava Prezentace ke dni 13. března 2017 Jiří Dvorský (VŠB TUO) Grafy 104 / 309 Osnova přednášky Grafy
Projekční algoritmus. Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění. Jan Klíma
 Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague
 1 / 23 Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague 2 / 23 biologové často potřebují najít často se opakující sekvence DNA tyto sekvence bývají relativně krátké,
1 / 23 Faculty of Nuclear Sciences and Physical Engineering Czech Technical University in Prague 2 / 23 biologové často potřebují najít často se opakující sekvence DNA tyto sekvence bývají relativně krátké,
Typologická koncepce druhu
 Speciace Co je to druh? Nebudu zde ani probírat různé definice pojmu druh. Žádná z nich až dosud neuspokojila všechny přírodovědce, ale každý přírodovědec zhruba ví, co míní tím, když mluví o druhu. (Charles
Speciace Co je to druh? Nebudu zde ani probírat různé definice pojmu druh. Žádná z nich až dosud neuspokojila všechny přírodovědce, ale každý přírodovědec zhruba ví, co míní tím, když mluví o druhu. (Charles
MOLEKULÁRNÍ TAXONOMIE 10
 MOLEKULÁRNÍ TAXONOMIE 10 Molekulární hodiny Skutečnost, že počet substitučních událostí vzrůstá s časem, vedla velmi brzy ke snahám využít sekvencí k datování stáří uzlů na fylogenetických stromech. Jako
MOLEKULÁRNÍ TAXONOMIE 10 Molekulární hodiny Skutečnost, že počet substitučních událostí vzrůstá s časem, vedla velmi brzy ke snahám využít sekvencí k datování stáří uzlů na fylogenetických stromech. Jako
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Genetické algoritmy. Informační a komunikační technologie ve zdravotnictví
 Genetické algoritmy Informační a komunikační technologie ve zdravotnictví Přehled přednášky Úvod Historie Základní pojmy Principy genetických algoritmů Možnosti použití Související metody AI Příklad problém
Genetické algoritmy Informační a komunikační technologie ve zdravotnictví Přehled přednášky Úvod Historie Základní pojmy Principy genetických algoritmů Možnosti použití Související metody AI Příklad problém
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
p 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y 6) Jak jsme osídlili svět podle
Nové směry v evoluční biologii. Jaroslav Flegr Katedra filosofie a dějin přírodních věd Přírodovědecká Fakulta UK Praha
 Nové směry v evoluční biologii Jaroslav Flegr Katedra filosofie a dějin přírodních věd Přírodovědecká Fakulta UK Praha 2014 Genetika věda o dědění znaků Mendelismus původně spíše antidarwinistický
Nové směry v evoluční biologii Jaroslav Flegr Katedra filosofie a dějin přírodních věd Přírodovědecká Fakulta UK Praha 2014 Genetika věda o dědění znaků Mendelismus původně spíše antidarwinistický
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Pravděpodobnost Podmíněná p. Úplná p. III. Pravděpodobnost. III. Pravděpodobnost Statistika A (ZS 2015)
") III Pravděpodobnost Pravděpodobnost Podmíněná p. Úplná p. Odkud se bere pravděpodobnost? 1. Pravděpodobnost, že z balíčku zamíchaných karet vytáhmene dvě esa je přibližně 0:012. Modely a teorie. 2. Pravděpodobnost,
III Pravděpodobnost Pravděpodobnost Podmíněná p. Úplná p. Odkud se bere pravděpodobnost? 1. Pravděpodobnost, že z balíčku zamíchaných karet vytáhmene dvě esa je přibližně 0:012. Modely a teorie. 2. Pravděpodobnost,
Rekonstrukce biogeografické historie: outline přednášky
 Rekonstrukce biogeografické historie: outline přednášky tradiční přístupy (do 80-ých let) a jejich slabiny Croizatova panbiogeografie a její slabiny Hennigovo progression rule a jeho slabiny disperzní
Rekonstrukce biogeografické historie: outline přednášky tradiční přístupy (do 80-ých let) a jejich slabiny Croizatova panbiogeografie a její slabiny Hennigovo progression rule a jeho slabiny disperzní
Státnice odborné č. 20
 Státnice odborné č. 20 Shlukování dat Shlukování dat. Metoda k-středů, hierarchické (aglomerativní) shlukování, Kohonenova mapa SOM Shlukování dat Shluková analýza je snaha o seskupení objektů do skupin
Státnice odborné č. 20 Shlukování dat Shlukování dat. Metoda k-středů, hierarchické (aglomerativní) shlukování, Kohonenova mapa SOM Shlukování dat Shluková analýza je snaha o seskupení objektů do skupin
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
Matematika III. 4. října Vysoká škola báňská - Technická univerzita Ostrava. Matematika III
 Vysoká škola báňská - Technická univerzita Ostrava 4. října 2018 Podmíněná pravděpodobnost Při počítání pravděpodobnosti můžeme k náhodnému pokusu přidat i nějakou dodatečnou podmínku. Podmíněná pravděpodobnost
Vysoká škola báňská - Technická univerzita Ostrava 4. října 2018 Podmíněná pravděpodobnost Při počítání pravděpodobnosti můžeme k náhodnému pokusu přidat i nějakou dodatečnou podmínku. Podmíněná pravděpodobnost
Jak měříme genetickou vzdálenost a co nám říká F ST
 Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Molekulární základy genetiky 1/76 GENY Označení GEN se používá ve dvou základních významech: 1. Jako synonymum pro vlohu
Dobývání znalostí. Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze
 Dobývání znalostí Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze Dobývání znalostí Pravděpodobnost a učení Doc. RNDr. Iveta Mrázová,
Dobývání znalostí Doc. RNDr. Iveta Mrázová, CSc. Katedra teoretické informatiky Matematicko-fyzikální fakulta Univerzity Karlovy v Praze Dobývání znalostí Pravděpodobnost a učení Doc. RNDr. Iveta Mrázová,
World of Plants Sources for Botanical Courses
 Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
VK CZ.1.07/2.2.00/
 Robotika Tvorba map v robotice - MRBT 3. března 2015 Ing. František Burian Komplexní inovace studijních programů a zvyšování kvality výuky na FEKT VUT v Brně OP VK CZ.1.07/2.2.00/28.0193 v pojetí mobilní
Robotika Tvorba map v robotice - MRBT 3. března 2015 Ing. František Burian Komplexní inovace studijních programů a zvyšování kvality výuky na FEKT VUT v Brně OP VK CZ.1.07/2.2.00/28.0193 v pojetí mobilní
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Prohledávání do šířky = algoritmus vlny
 Prohledávání do šířky = algoritmus vlny - souběžně zkoušet všechny možné varianty pokračování výpočtu, dokud nenajdeme řešení úlohy průchod stromem všech možných cest výpočtu do šířky, po vrstvách (v každé
Prohledávání do šířky = algoritmus vlny - souběžně zkoušet všechny možné varianty pokračování výpočtu, dokud nenajdeme řešení úlohy průchod stromem všech možných cest výpočtu do šířky, po vrstvách (v každé
Genetika pro začínající chovatele
 21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
4. Úvod do kladistiky. kladogram podobnost a příbuznost homologie (sym)plesiomorfie, (syn)apomorfie polarizace znaků kritérium parsimonie
plesiomorfie, (syn)apomorfie polarizace znaků kritérium parsimonie") 4. Úvod do kladistiky kladogram podobnost a příbuznost homologie (sym)plesiomorfie, (syn)apomorfie polarizace znaků kritérium parsimonie Willi Hennig (1913-1976) německý entomolog 1950: Grundzüge einer
4. Úvod do kladistiky kladogram podobnost a příbuznost homologie (sym)plesiomorfie, (syn)apomorfie polarizace znaků kritérium parsimonie Willi Hennig (1913-1976) německý entomolog 1950: Grundzüge einer
Binární vyhledávací stromy pokročilé partie
 Binární vyhledávací stromy pokročilé partie KMI/ALS lekce Jan Konečný 30.9.204 Literatura Cormen Thomas H., Introduction to Algorithms, 2nd edition MIT Press, 200. ISBN 0-262-5396-8 6, 3, A Knuth Donald
Binární vyhledávací stromy pokročilé partie KMI/ALS lekce Jan Konečný 30.9.204 Literatura Cormen Thomas H., Introduction to Algorithms, 2nd edition MIT Press, 200. ISBN 0-262-5396-8 6, 3, A Knuth Donald
Využití metod strojového učení v bioinformatice David Hoksza
 Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
Využití metod strojového učení v bioinformatice David Hoksza SIRET Research Group Katedra softwarového inženýrství, Matematicko-fyzikální fakulta Karlova Univerzita v Praze Bioinformatika Biologické inspirace
Hemoglobin a jemu podobní... Studijní materiál. Jan Komárek
 Hemoglobin a jemu podobní... Studijní materiál Jan Komárek Bioinformatika Bioinformatika je vědní disciplína, která se zabývá metodami pro shromážďování, analýzu a vizualizaci rozsáhlých souborů biologických
Hemoglobin a jemu podobní... Studijní materiál Jan Komárek Bioinformatika Bioinformatika je vědní disciplína, která se zabývá metodami pro shromážďování, analýzu a vizualizaci rozsáhlých souborů biologických
Úvodem Dříve les než stromy 3 Operace s maticemi
 Obsah 1 Úvodem 13 2 Dříve les než stromy 17 2.1 Nejednoznačnost terminologie 17 2.2 Volba metody analýzy dat 23 2.3 Přehled vybraných vícerozměrných metod 25 2.3.1 Metoda hlavních komponent 26 2.3.2 Faktorová
Obsah 1 Úvodem 13 2 Dříve les než stromy 17 2.1 Nejednoznačnost terminologie 17 2.2 Volba metody analýzy dat 23 2.3 Přehled vybraných vícerozměrných metod 25 2.3.1 Metoda hlavních komponent 26 2.3.2 Faktorová
Bioinformatika a výpočetní biologie. KFC/BIN VII. Fylogenetická analýza
 ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
5.1. Klasická pravděpodobnst
 5. Pravděpodobnost Uvažujme množinu Ω všech možných výsledků náhodného pokusu, například hodu mincí, hodu kostkou, výběru karty z balíčku a podobně. Tato množina se nazývá základní prostor a její prvky
5. Pravděpodobnost Uvažujme množinu Ω všech možných výsledků náhodného pokusu, například hodu mincí, hodu kostkou, výběru karty z balíčku a podobně. Tato množina se nazývá základní prostor a její prvky
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Úvod do optimalizace, metody hladké optimalizace
 Evropský sociální fond Investujeme do vaší budoucnosti Úvod do optimalizace, metody hladké optimalizace Matematika pro informatiky, FIT ČVUT Martin Holeňa, 13. týden LS 2010/2011 O čem to bude? Příklady
Evropský sociální fond Investujeme do vaší budoucnosti Úvod do optimalizace, metody hladké optimalizace Matematika pro informatiky, FIT ČVUT Martin Holeňa, 13. týden LS 2010/2011 O čem to bude? Příklady
Taxonomický systém a jeho význam v biologii
 Taxonomie Taxonomický systém a jeho význam v biologii -věda zabývající se tříděním organismů (druhů, rodů, ), jejich vzájemnou příbuzností a podobností. 3 úrovně: 1) charakteristika, pojmenování, vymezení
Taxonomie Taxonomický systém a jeho význam v biologii -věda zabývající se tříděním organismů (druhů, rodů, ), jejich vzájemnou příbuzností a podobností. 3 úrovně: 1) charakteristika, pojmenování, vymezení
Výuka genetiky na Přírodovědecké fakultě UK v Praze
 Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
ZÁKLADY STATISTICKÉHO ZPRACOVÁNÍ ÚDAJŮ 5. hodina , zapsala Veronika Vinklátová Revize zápisu Martin Holub,
 ZÁKLADY STATISTICKÉHO ZPRACOVÁNÍ ÚDAJŮ 5. hodina - 22. 3. 2018, zapsala Revize zápisu Martin Holub, 27. 3. 2018 I. Frekvenční tabulky opakování z minulé hodiny Frekvenční tabulka je nejzákladnější nástroj
ZÁKLADY STATISTICKÉHO ZPRACOVÁNÍ ÚDAJŮ 5. hodina - 22. 3. 2018, zapsala Revize zápisu Martin Holub, 27. 3. 2018 I. Frekvenční tabulky opakování z minulé hodiny Frekvenční tabulka je nejzákladnější nástroj
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Studijní materiály pro bioinformatickou část ViBuChu. úloha II. Jan Komárek, Gabriel Demo
 Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
Studijní materiály pro bioinformatickou část ViBuChu úloha II Jan Komárek, Gabriel Demo Adenin Struktura DNA Thymin 5 konec 3 konec DNA tvořena dvěmi řetězci orientovanými antiparalelně (liší se orientací
BOOTSTRAPPINGOVÉ METODY VE FYLOGENETICE
 VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
VYSOKÉ UČENÍ TECHNICKÉ V BRNĚ BRNO UNIVERSITY OF TECHNOLOGY FAKULTA ELEKTROTECHNIKY A KOMUNIKAČNÍCH TECHNOLOGIÍ ÚSTAV BIOMEDICÍNSKÉHO INŽENÝRSTVÍ FACULTY OF ELECTRICAL ENGINEERING AND COMMUNICATION DEPARTMENT
Genetika zvířat - MENDELU
 Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).
: Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).") Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
TEORIE PRAVDĚPODOBNOSTI. 2. cvičení
 TEORIE RAVDĚODONOSTI 2. cvičení Základní pojmy Klasická def. Statistická def. Geometrická def. odmíněná prav. ayesův teorém Test Základní pojmy Náhodný pokus - je každý konečný děj, jehož výsledek není
TEORIE RAVDĚODONOSTI 2. cvičení Základní pojmy Klasická def. Statistická def. Geometrická def. odmíněná prav. ayesův teorém Test Základní pojmy Náhodný pokus - je každý konečný děj, jehož výsledek není
Pravděpodobnost a statistika (BI-PST) Cvičení č. 1
 Cvičení č. 1") Pravděpodobnost a statistika (BI-PST) Cvičení č. 1 Katedra aplikované matematiky Fakulta informačních technologií České vysoké učení technické v Praze ZS 2014/2015 (FIT ČVUT) BI-PST, Cvičení č. 1 ZS 2014/2015
Pravděpodobnost a statistika (BI-PST) Cvičení č. 1 Katedra aplikované matematiky Fakulta informačních technologií České vysoké učení technické v Praze ZS 2014/2015 (FIT ČVUT) BI-PST, Cvičení č. 1 ZS 2014/2015
7. přednáška Systémová analýza a modelování. Přiřazovací problém
 Přiřazovací problém Přiřazovací problémy jsou podtřídou logistických úloh, kde lze obecně říci, že m dodavatelů zásobuje m spotřebitelů. Dalším specifikem je, že kapacity dodavatelů (ai) i požadavky spotřebitelů
Přiřazovací problém Přiřazovací problémy jsou podtřídou logistických úloh, kde lze obecně říci, že m dodavatelů zásobuje m spotřebitelů. Dalším specifikem je, že kapacity dodavatelů (ai) i požadavky spotřebitelů
Cvičení ze statistiky - 4. Filip Děchtěrenko
 Cvičení ze statistiky - 4 Filip Děchtěrenko Minule bylo.. Dokončili jsme deskriptivní statistiku Tyhle termíny by měly být známé: Korelace Regrese Garbage in, Garbage out Vícenásobná regrese Pravděpodobnost
Cvičení ze statistiky - 4 Filip Děchtěrenko Minule bylo.. Dokončili jsme deskriptivní statistiku Tyhle termíny by měly být známé: Korelace Regrese Garbage in, Garbage out Vícenásobná regrese Pravděpodobnost
Základní pojmy I. EVOLUCE
 Základní pojmy I. EVOLUCE Medvěd jeskynní Ursus spelaeus - 5 mil. let? - 10 tis. let - 200 tis. let? Medvěd hnědý Ursus arctos Medvěd lední Ursus maritimus Základní otázky EVOLUCE Jakto, že jsou tu různé
Základní pojmy I. EVOLUCE Medvěd jeskynní Ursus spelaeus - 5 mil. let? - 10 tis. let - 200 tis. let? Medvěd hnědý Ursus arctos Medvěd lední Ursus maritimus Základní otázky EVOLUCE Jakto, že jsou tu různé
Příklad 1. Řešení 1 ŘEŠENÉ PŘÍKLADY Z MV2 ČÁST 11
 Příklad 1 Vyhláška Ministerstva zdravotnictví předpokládala, že doba dojezdu k pacientovi od nahlášení požadavku nepřekročí 17 minut. Hodnoty deseti náhodně vybraných dob příjezdu sanitky k nemocnému byly:
Příklad 1 Vyhláška Ministerstva zdravotnictví předpokládala, že doba dojezdu k pacientovi od nahlášení požadavku nepřekročí 17 minut. Hodnoty deseti náhodně vybraných dob příjezdu sanitky k nemocnému byly:
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Využití DNA sekvencování v
 Využití DNA sekvencování v taxonomii prokaryot Mgr. Pavla Holochová, doc. RNDr. Ivo Sedláček, CSc. Česká sbírka mikroorganismů Ústav experimentální biologie Přírodovědecká fakulta Masarykova univerzita,
Využití DNA sekvencování v taxonomii prokaryot Mgr. Pavla Holochová, doc. RNDr. Ivo Sedláček, CSc. Česká sbírka mikroorganismů Ústav experimentální biologie Přírodovědecká fakulta Masarykova univerzita,
Genetické rozdíly mezi populacemi aneb něco o migracích a genovém toku. Genetické rozdíly mezi populacemi
 Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Vztah genotyp fenotyp
 Evoluce fenotypu II Vztah genotyp fenotyp plán? počítačový program? knihovna? genotypová astrologie (Jablonka a Lamb) Modely RNA - různé vážení: A-U, G-C, G-U interakcí, penalizace za neodpovídající si
Evoluce fenotypu II Vztah genotyp fenotyp plán? počítačový program? knihovna? genotypová astrologie (Jablonka a Lamb) Modely RNA - různé vážení: A-U, G-C, G-U interakcí, penalizace za neodpovídající si
Příbuznost a inbreeding
 Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Vícerozměrné statistické metody
 Vícerozměrné statistické metody Podobnosti a vzdálenosti ve vícerozměrném prostoru, asociační matice II Jiří Jarkovský, Simona Littnerová Vícerozměrné statistické metody Práce s asociační maticí Vzdálenosti
Vícerozměrné statistické metody Podobnosti a vzdálenosti ve vícerozměrném prostoru, asociační matice II Jiří Jarkovský, Simona Littnerová Vícerozměrné statistické metody Práce s asociační maticí Vzdálenosti
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Lékařská biofyzika, výpočetní technika I. Biostatistika Josef Tvrdík (doc. Ing. CSc.)
") Lékařská biofyzika, výpočetní technika I Biostatistika Josef Tvrdík (doc. Ing. CSc.) Přírodovědecká fakulta, katedra informatiky josef.tvrdik@osu.cz konzultace úterý 14.10 až 15.40 hod. http://www1.osu.cz/~tvrdik
Lékařská biofyzika, výpočetní technika I Biostatistika Josef Tvrdík (doc. Ing. CSc.) Přírodovědecká fakulta, katedra informatiky josef.tvrdik@osu.cz konzultace úterý 14.10 až 15.40 hod. http://www1.osu.cz/~tvrdik
Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak)
") MOLEKULÁRNÍ TAXONOMIE - 6 (2015) Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak) Nyní si ukážeme obecný princip, jakým se obohacují substituční modely o parametry tak,
MOLEKULÁRNÍ TAXONOMIE - 6 (2015) Uvolňování parametrů v substitučních modelech (opakování z minula, trochu jinak) Nyní si ukážeme obecný princip, jakým se obohacují substituční modely o parametry tak,
Simulační modely. Kdy použít simulaci?
 Simulační modely Simulace z lat. Simulare (napodobení). Princip simulace spočívá v sestavení modelu reálného systému a provádění opakovaných experimentů s tímto modelem. Simulaci je nutno považovat za
Simulační modely Simulace z lat. Simulare (napodobení). Princip simulace spočívá v sestavení modelu reálného systému a provádění opakovaných experimentů s tímto modelem. Simulaci je nutno považovat za
Odhady - Sdružené rozdělení pravděpodobnosti
 Odhady - Sdružené rozdělení pravděpodobnosti 4. listopadu 203 Kdybych chtěl znát maximum informací o náhodné veličině, musel bych znát všechny hodnoty, které mohou padnout, a jejich pravděpodobnosti. Tedy
Odhady - Sdružené rozdělení pravděpodobnosti 4. listopadu 203 Kdybych chtěl znát maximum informací o náhodné veličině, musel bych znát všechny hodnoty, které mohou padnout, a jejich pravděpodobnosti. Tedy
Ústav teorie informace a automatizace. J. Vomlel (ÚTIA AV ČR) Úvod do bayesovských sítí 30/10/ / 28
 Úvod do bayesovských sítí 30/10/ / 28") Úvod do bayesovských sítí Jiří Vomlel Ústav teorie informace a automatizace Akademie věd České republiky http://www.utia.cz/vomlel 30. října 2008 J. Vomlel (ÚTIA AV ČR) Úvod do bayesovských sítí 30/10/2008
Úvod do bayesovských sítí Jiří Vomlel Ústav teorie informace a automatizace Akademie věd České republiky http://www.utia.cz/vomlel 30. října 2008 J. Vomlel (ÚTIA AV ČR) Úvod do bayesovských sítí 30/10/2008
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Operátory ROLLUP a CUBE
 Operátory ROLLUP a CUBE Dotazovací jazyky, 2009 Marek Polák Martin Chytil Osnova přednášky o Analýza dat o Agregační funkce o GROUP BY a jeho problémy o Speciální hodnotový typ ALL o Operátor CUBE o Operátor
Operátory ROLLUP a CUBE Dotazovací jazyky, 2009 Marek Polák Martin Chytil Osnova přednášky o Analýza dat o Agregační funkce o GROUP BY a jeho problémy o Speciální hodnotový typ ALL o Operátor CUBE o Operátor
Řešení: PŘENESVĚŽ (N, A, B, C) = přenes N disků z A na B pomocí C
 = přenes N disků z A na B pomocí C") Hanojské věže - 3 kolíky A, B, C - na A je N disků různé velikosti, seřazené od největšího (dole) k nejmenšímu (nahoře) - kolíky B a C jsou prázdné - úkol: přenést všechny disky z A na B, mohou se odkládat
Hanojské věže - 3 kolíky A, B, C - na A je N disků různé velikosti, seřazené od největšího (dole) k nejmenšímu (nahoře) - kolíky B a C jsou prázdné - úkol: přenést všechny disky z A na B, mohou se odkládat
Moderní systémy pro získávání znalostí z informací a dat
 Moderní systémy pro získávání znalostí z informací a dat Jan Žižka IBA Institut biostatistiky a analýz PřF & LF, Masarykova universita Kamenice 126/3, 625 00 Brno Email: zizka@iba.muni.cz Bioinformatika:
Moderní systémy pro získávání znalostí z informací a dat Jan Žižka IBA Institut biostatistiky a analýz PřF & LF, Masarykova universita Kamenice 126/3, 625 00 Brno Email: zizka@iba.muni.cz Bioinformatika:
oddělení Inteligentní Datové Analýzy (IDA)
") Vytěžování dat Filip Železný Katedra počítačů oddělení Inteligentní Datové Analýzy (IDA) 22. září 2014 Filip Železný (ČVUT) Vytěžování dat 22. září 2014 1 / 25 Odhad rozdělení Úloha: Vstup: data D = {
Vytěžování dat Filip Železný Katedra počítačů oddělení Inteligentní Datové Analýzy (IDA) 22. září 2014 Filip Železný (ČVUT) Vytěžování dat 22. září 2014 1 / 25 Odhad rozdělení Úloha: Vstup: data D = {