Pluripotentní, odvozené kmenové buňky. EpiSC
|
|
|
- Robert Němeček
- před 5 lety
- Počet zobrazení:
Transkript
 Embryonální zárodečné buňky")
 Kmenové buňky epiblastu (Epiblast stem cell EpiSC) Early-primitive")
1 Pluripotentní, odvozené kmenové buňky EPL ESC EpiSC EGC ECC Embryonální kmenové buňky (Embryonic stem cell ESC) Embryonální zárodečné buňky (Embryonic germ cell EGC) Embryonální nádorové buňky (Embryonal carcinoma cell ECC) Kmenové buňky epiblastu (Epiblast stem cell EpiSC) Early-primitive ectoderm-like - EPL
 - jsou pluripotentní - přirozeně neexistují, pouze in vitro - jsou nesmrtelné - mají schopnost si udržet stabilní genotyp (!")
2 Embryonální kmenové buňky (Embryonic stem cell) ESC - jsou odvozené z vnitřní buněčné masy blastocysty - fenotypem odpovídají přibližně buňkám vnitřní buněčné masy (mesc, ICM inner cell mass) nebo epiblastu (hesc, EpiSC) - jsou pluripotentní - přirozeně neexistují, pouze in vitro - jsou nesmrtelné - mají schopnost si udržet stabilní genotyp (!?) - po injikaci do imunitně tolerantního organismu tvoří teratomy (důkaz pluripotence) - po injikaci do blastocysty mají schopnost tvořit kompletní chiméry (známo jen u mesc, důkaz pluripotence)




3 Ontogeneze a diferenciační potenciál buněk vnitřní buněčné masy Gilbert 1997 / Bílek 2004



4 Izolace myších ES buněk 6-8 ddnů IVF 3,5 dní p.c. TE 4-5 dní ICM Imuno-chirurgie Vyseknutí a disociace ICM Fibroblastová výživná vrstva feeder - tvořená MEF Odstranění PEF Upraveno podle Kroupová 2004 Linie ES buněk na feederu MEF Homogenní populace ES buněk

 do blastocysty (příjemce) jsou směsí geneticky odlišných buněk na")
5 TERATOM Nádor, který obsahuje buňky více jak jednoho zárodečného listu, většinou všech tří. Tyto buňky mohou být fenotypu od časných stádií až po terminálně diferencované Teratomy jsou typické nádory původem ze zárodečných buněk (ovariální a testikulární teratomy. Teratomy jsou jak benigní, tak maligní (teratokarcinom) Kmenové buňky teratokarcinomu = embryonální nádorové buňky (Embryonal carcinoma (EC) cells) CHIMÉRA Organismus je směsí geneticky odlišných buněk / tkání / orgánů Chimeričtí jedinci vzniklí injikací ES buněk (dárce) do blastocysty (příjemce) jsou směsí geneticky odlišných buněk na ůrovni všech tkání, a tak také vytváří pohlavní buňky s genotypem jak dárce, tak příjemce!
6 Růst a kultivace ES buněk PROLIFERACE DIFERENCIACE - APOTÓSA Existence a charakter ES buněk je udržován kombinací účinků vnějších (extrinsic) a vnitřních (intrinsic) faktorů. Vnější faktory si ES buňky částečně syntetizují samy, ale ve větší míře musí být dodávány. Významným zdrojem těchto faktorů je výživná vrstva na které se ES buňky kultivují = FEEDER. Vnitřní faktory si ES buňky nesou jako pozůstatek svého embryonálního původu. FEEDER - výživná vrstva = zdroj růstových faktorů (cytokiny, ECM,..) a vhodný podklad - nejčastěji se používají myší embryonální fibroblasty (13d p.c., MEF (PEF)) - bez feederu z MEF = definované podmínky, ale horší ES buňky - tendence používat druhově identické MEF = sníženi rizika přenosu virů, - MEF lze nahradit i jinými typy fibroblastů případně jinými buňkami -MEF lze částečně nahradit komponenty ECM, specifickými cytokiny a přídavky, matrigelem,, obecně definovanějšími preparáty Důlëžitou komponentou kultivačního média pro ES jsou také nedefinované faktory séra, které lze nahradit dodáváním specifických růstových faktorů. Často se také používá tzv. Serum-replacement = lépe definovaná náhražka séra (patentované složení).
7 Obecný model inhibice diferenciace ES buněk - ES buňky spontáně diferencují, v kultuře je třeba této diferenciaci zabránit - diferenciace je často spojena s apoptózou - vhodnými kultivačními podmínkami, lze diferenciaci inhibovat - faktory inhibující diferenciaci ES buněk se částečně liší u různých druhů, ale existují výjimky i v rámci jednoho druhu! ES cell auto- a parakrinně působící factory mezoderm + entoderm A B feeder (výživná vrstva) ektoderm
8 Model inhibice diferenciace myších ES buněk ES cell IGF, FGF-4, Wnt mezoderm + entoderm LIF* Feeder* BMP-4 (BMP-2 /BMP-12 /FCS nebo SR*) ektoderm - existují i linie mes buněk nezávislé na LIF - zdá se, že z LIF závislé linie, lze vyselektovat LIF nezávislou (!?) -mesbuňky pěstované bez feederu ztrácí schopnost tvořit chiméry in vivo - některé linie nelze bez feederu pěstovat vůbec vlastnosti mes buněk jsou ovlivněny genotypem imbredního kmene myší z kterého byly izolovány! LIF leukemiainhibitory factor BMP-2,4,12 bone morphogenetic protein 2,4,12 FCS fetal calf serum FGF-4 fibroblast growth factor SR serum replacement *ABSOLUTNÍ OPTIMUM
9 Model inhibice diferenciace lidských ES buněk ES cell IGF,?, Wnt, FGF-2 mezoderm + entoderm? FGF-2 Activin/Nodal Noggin Wnt FCS? + MEF (feeder) nebo matrigel ektoderm FCS obsahuje jak difereciaci indukující, tak diferenciaci inhibující faktory. Kvalitu FCS také ovlivňuje titr protilátek a složek koplementu. FCS se liší mezi jednotlivými šaržemi = je třeba je testovat. Lépe je používat náhrady FCS se sníženou koncentrací negativně působících látek na kultivaci ES buněk, např. Serum- Replacement ( fy. Invitrogene-Gibco) OPTIMUM???
10 Vnitřní faktory charakterizující / regulující ES buňky (pluripotentní embryonální buňky) Oct-4 (Oct-3/4, Oct-3, Pou5f1 master of pluripotency) - transkripční faktor, homeoprotein - exprimuje se již u 2/4 (myš aktivace transkripce) buněčného embrya - ve stádiu blastuly je jeho exprese výrazně zvýšena - ve stádiu blastocysty je pouze v buňkách ICM - později jeho exprese vymizí, zachovává se pouze v PGC (a později v zárodečných buňkách), nízká exprese se předpokládá i v somatických kmenových buňkách?!? - nebyl nalezen u kuřat - reguluje expresi FGF-4 (Oct-4/Sox-2), PDGFα,. -v průběhu diferenciace za snížení jeho exprese odpovídá GCNF( germ cell nuclear factor, RA (retinoic acid), - V primitivním ektodermu je exprese Oct-4 podporována LRH-1 (liver receptor homologue 1) Nanog - transkripční faktor, homeoprotein - jeho exprese vede k udržení vysoké hladiny Oct-4 - objevuje se již ve vnitřních buňkách moruly Sox-2 - Transkripční faktor Sry-rodiny (sex-determining region Y protein) - Kooperuje s Oct-4 na vlastní expresi -V průběhu indukce neurální diferenciace se jeho exprese zvyšuje

11 Boiani & Scholer 2005
12 Potencionální mechanismus účinků LIF u mes buněk LIF/gp130/LIFR Jak2/STAT3 PI3K/Akt + Erk Klf4 Tbx3 Sox2 Nanog Oct4 Niwa 2009
 a) standard protocol b) 3i/LIF protocol c) 2i/LIF protocol LIF (indukce STAT3) SU5402 (inhibice FGF-R) PD0325901 /")
13 Alternativní kultivace mes protocol 2i/LIF, 3i/LIF (A. Smith laboratory) a) standard protocol b) 3i/LIF protocol c) 2i/LIF protocol LIF (indukce STAT3) SU5402 (inhibice FGF-R) PD / PD (inhibice Erk signalizace) CHIR99021 (inhibice GSK3, Wnt signalizace)
14 Wnt a mes Niwa 2011


, jak u mes tak hes.")
.")
15 Regulace transkripce faktory Oct-4, Nanog a Sox-2 Předpokládaný model vzájemné regulace exprese FoxD3, Nanog a Oct-4 u mes. FOxD3 není exprimován u hes. Vzájemná regulace Oct-4 a Nanog není dosud plně objasněna. Je však již jasné, že Oct-4 řídí transkripci Nanog přímou vazbou v jeho promotoru (společně se Sox-2), jak u mes tak hes. Pro self-renewal ES buněk je klíčové zachovat rovnováhu v hladinách Oct-4 a Nanog (viz. výše). Pan2006 Promotorové sekvence rozpoznávané Oct-4, Nanog a Sox-2 u hes. Model vzájemné regulace Oct-4, Nanog a Sox-2 a některé jimi řízené geny u hes. Proteiny Promotory genů Boyer2005
16 Ztráta pluripotence u ES buněk v důsledku deregulace rovnováhy mezi hladinou Oct-4 a Nanog Chamber, 2004 Johnson 2006
.")
17 ES BUŇKY a) myší ES (mes) x b) lidské ES (hes) (LIF / gp130* / STAT3 dependent) (LIF / gp130* / STAT3 independent) Toto rozdělení je pravděpodobně dáno možností některých živočišných druhů mít skrytou březost ve stádiu blastocysty (embryonální diapauza). Rao, 2004
 hesc/mepisc gp130 signalling dependent somatické SC?")
18 Signální dráha gp130 ->-> STAT3 ve vztahu k pluripotentním buňkám u člověka a myši mesc PGC (h/m) EGC (h/m) embryonální diapauza (myš / lasicovití) hesc/mepisc gp130 signalling dependent somatické SC?? pluripotent multipotent a méně mouse
19 Signální dráha TGFbeta / BMPs u ES buněk + mes - hes / mepisc - mes + hes / mepisc
20 Některé charakteristické znaky myších a lidských ES buněk
, TRA antigen odmítnutí nádoru (tumor-rejection antigen), NR nestudováno (not")
21 Glykolipidy, Keratan chondroitin sulfát proteoglycan, I Tetraspannin transmembránové proteiny, AP alkalická fosfatáza, EB embryoidní tělísko, ESC embryonální kmenové buňky, SSEA vývojově specifický embryonální antigen (stage-specific embryonic antigen), TRA antigen odmítnutí nádoru (tumor-rejection antigen), NR nestudováno (not reported)
 Brons, 2007 &")
22 Kmenové buňky epiblastu EpiSC (Epiblast stem cell) Brons, 2007 & Tesar, 2007

 -EpiSCčástečně")
 Brons, 2007 &")
23 EpiSC - neintegrují se do moruly a blastocysty => rozdíl s mes - netvoří kompletní chiméry - nebyla detekována aktivní alkalická fosfatáza (AP) (mes, hes, EC, EG, PGC, mají aktivní AP) -EpiSCčástečně vysvětlují rozdíly mezi mes a hes Kmenové buňky epiblastu EpiSC (Epiblast stem cell) Brons, 2007 & Tesar, 2007

 Brons, 2007")
24 Kmenové buňky epiblastu EpiSC (Epiblast stem cell) Brons, 2007 & Tesar, 2007
25 mes je možno reverzibilně převést na buňky připomínající buňky primitivního ektodermu tzv. EPL (early-primitive ectoderm-like) buňky. Tyto buňky již nemají podobně jako buňky primitivního ektodrmu schopnost tvořit buňky parietálního entodermu. Také některé jejich další schopnosti diferencovat, jsou oproti ES buňkám pozměněny (Pelton 2002). ES (- FGF-5*) +LIF -LIF + HepG2 buňkami kondiciované médium EPL (+ FGF-5*) Gadue 2005 *EPL buňky jsou podobně jako buňky primitivního ektodermu exprimují FGF-5 na rozdíl od ES buněk, které exprimují zejména FGF-4 (platí pro myš)


26 Epigenetika ES buněk Metylace DNA a kondenzace chromatinu u ES buněk Meshorer & Misteli 2006
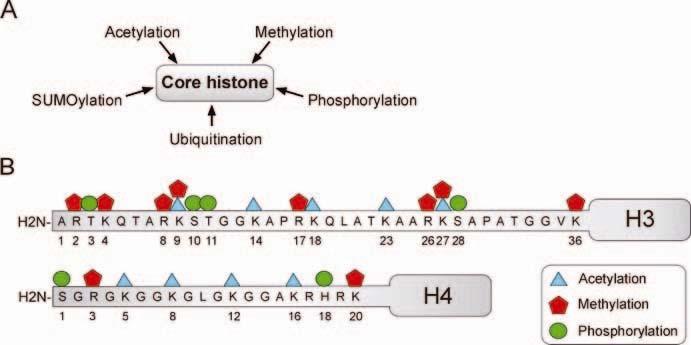
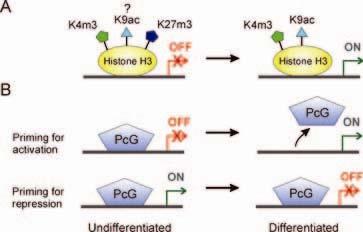
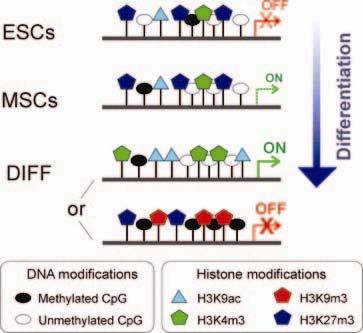
27 Modifikace histonů regulace transkripce

28 Profil postranslačních modifikací (lysinových zbytků) histonů H3 a H4 u ES (J1, V6.5), EC a MEF buněk Dai & Rasmussen 2007
 > modifikace histonů PT model promiskuitní transkripce (promiscuous transcription) > kombinace transkripčních faktorů a transdukce signálu Meshorer & Misteli")
29 Model změn trankripčního profilu v průběhu indukce diferenciace ES buněk HA model postupné (hierarchické) aktivace (hierarchical activation) > metylace DNA ETCM model povolené/umožněné transkripce (early transcription competence marks) > modifikace histonů PT model promiskuitní transkripce (promiscuous transcription) > kombinace transkripčních faktorů a transdukce signálu Meshorer & Misteli 2006
 Bibikova, 2006 cancer cells differ.")
30 Profil metylace DNA u různých typů buněk (40 genů, 49 CpG míst) Bibikova, 2006 cancer cells differ. cell SSC hes

31 Polycomb group (PcG) x Trithorax group (trxg) proteins PcG jsou odpovědné za inaktivaci transkripce cílových oblastí trxg jsou odpovědné za aktivaci transkripce cílových oblastí -obecně patří mezi skupinu proteinů modifikujících chromatin - regulují zejména transkripci homeotických genů význam v ontogenezi - jsou odpovědné za epigenetickou pamět genomu - rozpoznávají specifické a vzájemně málo homologní skvence DNA PRE PcG responsive element TRE trxg response element - jedná se o proteinové komplexy se základní jednotnou strukturou (PcG 4 základní skupiny komplexů), specifita těchto komplexů k daným PRE/TRE je dána dalšími asociujícími specifickými podjednotkami

32 Proliferace ES buněk S laskavým nedovolením z katalogu Santa Cruz Biotechnology, Inc.
33 - ES buňky relativně intenzivně proliferují (podobné nádorovým buňkám) - G1 fáze buněčného cyklu je krátká (u mes ~ 1.5 h), zdá se, že chybí G1-checkpoint* - velké procento buněk je v S fázi buněčného cyklu (mes > 60%, hes > 50%) - doubling time mes ~ 12h, hes ~ 24h - specifická charakteristika regulace buněčného cyklu (odolnost k p16, nízká hladina cyklinů D, vysoká hladina cyklinu E, není potřeba proteinů rodiny Rb) - inhibice proliferace (suboptimální podmíky) vede k diferenciaci a apoptóse, diferenciace a apoptósa ES buněk, však nemusí vést ke snížení proliferace jako takové *tzv. kontrolní bod R, o průchodu tímto bodem rozhodují zejména mechanismy rozpoznávající itegritu/neporušenost genomu, buňky s požkozenou DNA jsou za normálních okolností v tomto bodě zastaveny. Porucha tohoto kontrolního mechanismu je typická pro nádorové buňky.

34 Rozdílné proporce v jednotlivých fázích cyklu u časných embryonálních buněk


35 Jak vypadá správná ES buňka? Jsou všechny ES buňky v kultuře stejné? LIF -> STAT3 aktivita u mes buněk Microarray analýza exprese stemness genů Davey, 2006 Fortunel, 2003 pokračování pokračování
36 Model narůstající heterogenity v rostoucí kolonii ES buněk za dodržení známých optimálních kultivačních podmínek Myší ES 30<* 50<* 100<* růst kolonie ES buněk Lidské ES Buňky odpovídající buňkám ICM/epiblastu Časného neuroektodermu Buňky primitivního entodermu (morphologicky navíc tvoří také malá granula) ICM vývoj blastocysty epiblast *orientační počet buněk v kolonii blastocoel blastocoel trofoectoderm
37 Fortunel, 2003 Variabilita v analýze transkripčního profilu u ES buněk (ESC), neurálních progenitorů (NPC), progenitorů retiny (RPC) a hematopoetických kmenových buněk (HSC) u myši. Tři pracovní skupiny, jedna metoda. ESC x NPC + RPC/HSC ESC Fortunel NPC 798 Ramalho-Santos Ivanova 286 ESC + NPC
38 Existují geny kmenovosti, tzv. stemness geny? Vývojově specifické geny -potenciál buněk + Příslušné signální dráhy, specifický patern jejich aktivit - regulace diferenciace, proliferace, sebeobnovy
39 Chromosomální stabilita a ES buňky Draper, 2004; Hanson, dlouhodobá kultivace ES buněk vede k selekci odolnejších klonů a subpopulací - menší responzivnost na vnější signály, rychlejší proliferace, menší nároky na kultivaci, klíčové znaky často zachovány (Oct-4, Nanog, příslušné SSEA,...) - u myší snížena schopnost tvorby chimér a zéjména germline u těchto chimér - často spojeno s genetickou manipulací (knock-out, -in ES linie) - nestabilita chromosómů, polyploidie, trizomie, zlomy a přeskupování genů => adaptace na in vitro podmínky - hes, primární je trizomie chromozomu 12 nebo 17, vzácněji chromosomu 14 a 20 - často dochází i ke zdvojení dlouhého raménka chromosomu 17 a k translokaci této kopie na dlouhé raménko chromosomu 6 => posílení exprese genů na chromosomu 17 (Survivin proti apoptóse, STAT3 self-renewal mes a nadbytek u většiny nádorů, GRB2 a 7 (growth factor receptor bound protein, viz. signální transdukce)) - na chromosomu 12 je lokalizován nanog (Nanog) - časté i změny malých oblastí na chromosomech 1, 8, 18 a 20 -u mes se jedná o změny zejména chromosomů 8 a 11 (myší chromosom 11 je z části ekvivalentní k lidskému chromosomu 17)

40 Apoptická kontrola nestability karyotypu a ES buňky Checkpoint-apoptosis... of karyotipic instability. Mantel, u zdravých somatických buněk při poruchách mitotického aparátu během jejich dělení a tak vznikající chyby v mitóse vedou k jejich přechodu do senescentního stavu nebo k indukci apoptósy - ES buňky mají zvýšenou toleranci k poruchám mitózy, menší citlivost tzv. SAC (SAC splindle assembly checkpoint) => akumulace aneuploidních a polyploidních buněk v populaci - indukcí diferenciace lze tyto buňky částečně eliminovat (apoptósa) - tetraploidní buňky (blastomery) mohou tvořit jen trofoblast => G 1 MTA/tetraploidity checkpoint => využití při tvorbě embryí plně ES původu
41 Příklad nárůstu apoptósy s diferenciací a požkozeným mitotickým aparátem ES embryonální kmenové buňky EB ES diferencovanou formou embryoidních tělísek Noc/NOC nocodazol PAC - paclitaxel
42 VYUŽITÍ ES BUNĚK 1. Biologický a biomedicínský výzkum - Příprava geneticky modifikovaných organismů - Studium mechanismů časné embryogeneze / diferenciace - Studium mechanismů kancerogeneze - Studium embryotoxicity - Testování farmak 2. Lékařství - Buněčné a tkáňové terapie - Příprava biologicky aktivních preparátů - Nosiče biologicky aktivních látek (pathotaxe)
43 Příprava geneticky modifikovaných organismů -GMO Pro vytvoření linie GMO je potřeba, aby požadovaná genetická modifikace byly obsažena i v pohlavních buňkách. Tuto modifikaci je tedy potřeba provést na buňkách toti- nebo pluripotentních. Náhodným nebo cíleným(?) vložením požadované DNA do zygoty Náhodným nebo cíleným vložením požadované DNA do ES buněk - Díky prakticky neomezené možnosti kultivace ES buněk, lze mít prováděnou genetickou modifikace plně pod kontrolou, a také ji můžeme velice přesně naplánovat!!! - ES buňky jsou pluripotentní, po zpětné injikaci do blastocysty a vložení této blastocysty do dělohy pseudo-pregnantní myši, blastocysta pokračuje ve vývoji a vzniklý jedinec je chimérou buněk původni ICM a injikovaných ES na ůrovni všech tkání,, tedy i zárodez rodečné.

44 Příprava KO myší
45 ES buňky v buněčné terapii
46 Diferenciace ES buněk a) In vivo - teratomy: injikace suspenze ES buněk do vaskularizované tkáně imunitně tolerantního zvířete, popřípadě do zvířete s farmakologicky potlačenou imunitní odpovědí - chiméry: injikace ES buněk do blastocysty, navrácení takové blastocysty do pseudo-pregnantní myši = vznik chimerického jedince b) In vitro - metodiky korespondující s ontogenezí - metodiky získané empiricky (kopírující ontogenezi?) Musí buňka diferencující z ES buňky vždy kopírovat ontogenezi aby dosáhla určitého stavu?
 V monovrstvě + dobře definovné podmínky -malávýtěžnost - silně závislé na genotypu Rathjen 1998 / Bílek")
47 In vitro diferenciace ES buněk kultivace indukce selekce a) Embryoidní tělíska (Embryoid bodies -EB) +jednoduché, více buněčných typů + tolerující genotyp -špatně definované podmínky b) V monovrstvě + dobře definovné podmínky -malávýtěžnost - silně závislé na genotypu Rathjen 1998 / Bílek 2004

48 Loebel, 2003 Úloha specifických růstových faktorů v ontogenezi myši

49 Příklady diferenciace myších ES buněk kombinací typu jejich kultivace a specifických růstových faktorů s porovnáním úlohy těchto faktorů v myší ontogenezi pokračování Loebel, 2003
50 Guasch, 2005
51 Příklad účinků jednotlivých specifických růstových faktorů na indukci diferenciace u lidských ES buněk v kombinaci s tvorbou embryoidních tělísek Schuldiner, 2000
52 Příklad difernciace ES buněk do různých typů neurálních buněk NSC neural stem cell, DA/Ser/GABA dopaminergní/serotonergní/gabanergní neurony, MN - motoneurony Barberi 2003
53 Model regenerace poškozené hematopoesy v důsledku Rag2-/- mutace s použitím ES buněk, genetických manipulací a jaderného reprogramování Rideout, 2002

54 SOUČASNÉ PROBLÉMY S VYUŽITÍM ES BUNĚK V TERAPII
55 - hes se nedaří dlouhodobě kultivovat beze změn v genotypu / fenotypu - dlouhodobá kultivace za sub-optimálních podmínek vede k dosud neznámým, epigenetickým změnám snižujícím schopnost pluripotence (ireverzibilními i pro cytoplasmu zygoty, Amano 2006) - dosud není spolehlivě vyřešen potencionální vznik teratomů - biologie a diferenciační potenciál ES buněk nejsou dosud dobře prozkoumány - kultivace ES buněk je stále závislá na nedefinovaných faktorech - etika získávání nových lidských ES linií - finance, přes velice atraktivní potenciál, který v sobě ES buňky mají, není jisté, jestli současná společnost bude mít dost prostředků na jejich využití např. v buněčné terapii
56 Primordiální zárodečné buňky PGC (primordial germ cell) - PGC se u myši objevují již 6 dpc, pravděpodobně je jejich vznik indukovaný v průběhu gastrulace, a to vnějšími signály, zejména BMP (na rozdíl od žab, Drosophily a C. elegans) dpc migrují vně vlastní embryo, později (8.5 dpc <) migrují podél zadního střeva embrya do vytvořené zárodečné lišty. - PGC zanikají po usazení v zárodečné liště (10-13 dpc u myši), stávají se z nich zárodečné buňky. Prodělají ještě 2-3 mitózy a u samců vznikají prospermatogonie zastavené v G0/G1 fázi mitózy. U samic vstupují do meiotické profáze (obojí > 12.5 dpc). - Podobně jako ICM a ES buňky mají vysokou hladinu alkalické fosfatázy (ale TNAP ne GCAP/TNAP), Oct4, Nanog,.. - Významné jsou zdá se zejména Stella a Fragilis. - PGC nejsou pluripotentní! - PGC mají omezený počet dělení, u myši napočítáno kolem 1000 buněk, rozdíly v závislosti na imbrední lini Stella Také u buněk ES a epiblastu, později jen u PGC, podílí se na udržení jejich fenotypu, po jejich usazení v zárodečné liště jeho exprese vymizí. Fragilis Z rodiny IFN indukovaných genů, je silně exprimován při formování PGC, s jejich migrací jeho exprese klesá.

57 Cyklus zárodečných buněk u myši
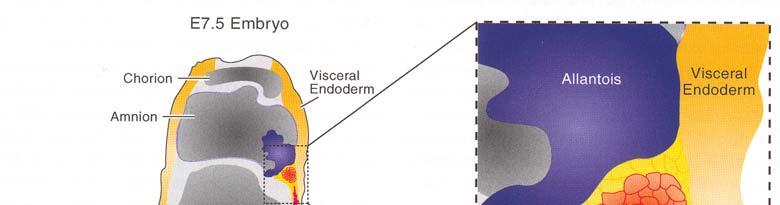
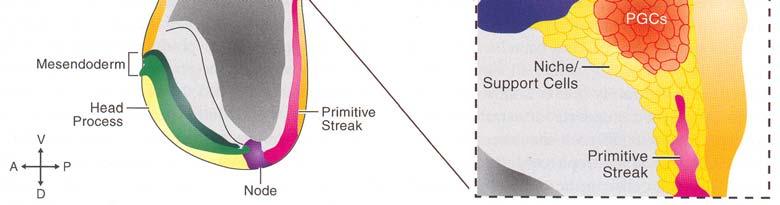
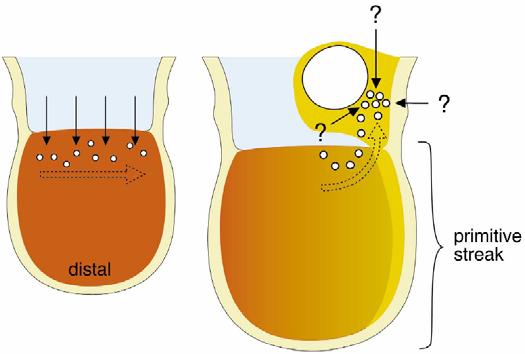

58 PGC primordiální zárodečné buňky ExpreseFragilismezi6 7 dpc Exprese Oct-4 v PGC usazených v zárodečné liště

59 Mechanismus vzniku PGC Hayashi, 2007

60 Blimp1 represor transkripce; Prmt5 - metyltransferáza Hayashi, 2007
61 Embryonální zárodečné buňky EGC (Embryonic germ cells) - EGC jsou odvozeny z primordiálních zárodečných buněk (PGC primordial germ cell). - Podobně jako ES buňky je lze expandovat in vitro, a jsou pluripotentní, jak dokazuje jejich schopnost diferencovat do buněk všech tří zárodečných listů jak in vitro (EB), tak in vivo (chiméry a teratomy). - Z epigenetického pohledu (DNA metylace) jsou však více podobné PGC než ES buňkám - Rozdíly v metylaci DNA se týkají zejména imprintovaných genů v závislosti na pohlaví, u EGC izolovaných z pozdějších embryonálních stádií se tento rozdíl zmenšuje. Tyto imprinting-free PGC, však již netvoří zdravé chimerické jedince. - U myší lze EGC izolovat z PGC mezi dpc, později to již nelze - PGC a následně EGC lze izolovat in vitro z ES buněk (lidských i myších) - m/hegc jsou závislé na LIF, během časné kultivace i na FGF2 a Stem cell faktoru (SCF) (+ feeder & FCS/SR). - Exprimují podobné markery jako ES buňky, lidské EGC jsou fenotypem více podobné mes než hes (morfologie + exprese SSEA-1!; u hes se SSEA-1 exprimuje až s jejich diferenciací)


62 Vznik EGC z PGC (Durcova-Hills 2008)

63
 R1, E14tg2a, TMA5 linie mes buněk TMA55G (male), TMA58G (female) EGC GS germ stem")
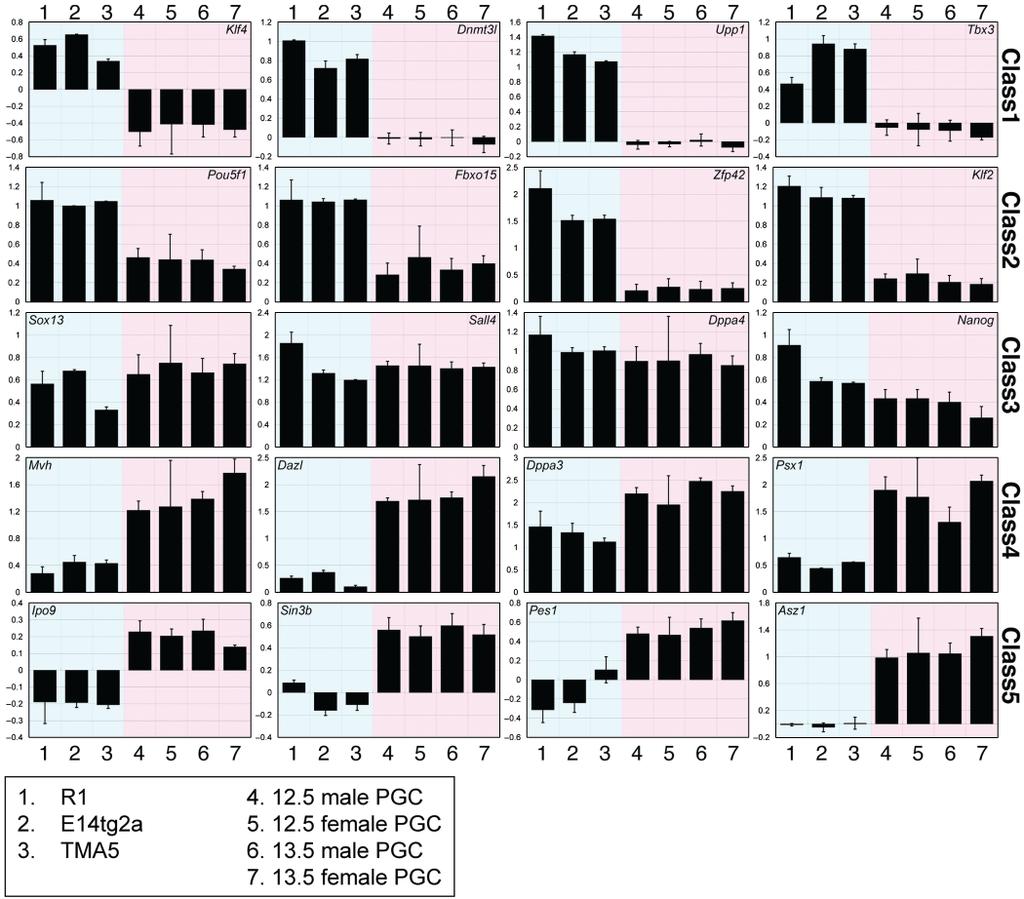
64 Porovnání transktiptomů mes, mpgc a některých tkání (Mise et al., 2008) R1, E14tg2a, TMA5 linie mes buněk TMA55G (male), TMA58G (female) EGC GS germ stem cell (derived from spermatogonia)

65 Analýza hlavních komponent expresních profilů blue ES specifický patern red PGC specifický patern purple - ES/PGC specifický patern green nevýznamné pro specifikaci ES/PGC
66
/ stem cell factor (SCF) v regulaci sel-renewal EG buněk Donovan, 2003")
67 Schema předpokládaného zapojení FGF, LIF a KL (c-kit ligand = Steel factor (SF)/ stem cell factor (SCF) v regulaci sel-renewal EG buněk Donovan, 2003
 - serine-threonine kinase mammalian target of rapamycin Donovan,")
68 Vztahy mezi pluripotentními buňkami a některé klíčové regulační komponenty těchto buněk Sl Steel locus, Ter Teratoma locus, pgct1 primordial germ cell tumor susceptibility locus, PTEN Phosphatase and tensin homologue deleted on chromosome 10, mtr (mtor) - serine-threonine kinase mammalian target of rapamycin Donovan, 2003

69 Není původ ES buněk v PGC??? Zwaka, 2005
70 Není původ ES buněk v PGC??? M- myš; H člověk; N/D netestováno; ES embryonální kmenové buňky; EGC časné primordiální zárodečné buňky (!); LGC pozdní primordiální zárodečné buňky; ICM vnitřní buněčná masa; PE primitivní ecdoderm/epiblast Zwaka, 2005
71 Kmenové buňky teratomu / teratokarcinomu - ECC Embryonální nádorové buňky (Embryonal carcinoma cells) - Izolované rozkultivováním a klonální selekcí buněk teratomu / teratokarcinomu - Lidské spontálně, myší indukované transplantací časných embryonálních buněk do dobře vyživované tkáně (varlata, ledviná kapsa, břišní dutina, ), musí být imunotolerance - Podobné vlastnosti jako ES buňky, ale méně závislé na specifických růstových faktorech (+) - Tvoří také chiméry, ale nedokonalé, většinou hynou v průběhu embryogeneze (-) - Většinou snížená schopnost pluripotence jak in vivo, tak in vitro (-) - Obecně nestabilní genotyp a časté aneuploidie (-) - Modelové studie genetické nestability a diferenciace, vzniku teratomů - Levnější alternativa k ES buňkám, lepší stabilita v experimentálních systémech jak ES (+)


72 Neurální lišta a kmenové buňky neurální lišty

73 Experiment potvrzujicí pluripotenci buněk neurální lišty
; Cdx2, Eomes, Errβ B) Kmenové buňky primitivního entodermu (hypoblastu) XEN buňky")

74 Kmenové buňky extraembryonálních tkání A) Kmenové buňky trofektodermu (trofoblastu) FGF-dependent (FGF4, FGFR2); Cdx2, Eomes, Errβ B) Kmenové buňky primitivního entodermu (hypoblastu) XEN buňky extraembryonálního entodermu zárodečný epiblast / embryoblast allantois primitivní ektoderm amnion ICM mezoderm žloutkového vaku primitivní entoderm viscerální entoderm blastomery parietální entoderm trofektoderm linie trofoblastu

75 Regulace kmenových buněk trofoblastu signály z epiblastu
Indukovaná pluripotence. Petr Vodička Liběchov 16/11/2016
 Indukovaná pluripotence Petr Vodička Liběchov 16/11/2016 Totipotentní Pluripotentní Lidské ES Myší ES LIF + FBS Feeder = vrstva podpůrných buněk Myší embryonální fibroblasty, SNL, STO bfgf + SR Feeder
Indukovaná pluripotence Petr Vodička Liběchov 16/11/2016 Totipotentní Pluripotentní Lidské ES Myší ES LIF + FBS Feeder = vrstva podpůrných buněk Myší embryonální fibroblasty, SNL, STO bfgf + SR Feeder
Úvod do studia biologie kmenových buněk. Jiří Pacherník tel:
 Úvod do studia biologie kmenových buněk Jiří Pacherník e-mail: jipa@sci.muni.cz tel: 532 146 223 Co jsou kmenové buňky? - buňky schopné vlastní obnovy (sebeobnova) - buňky schopné dávat vznik jiným typům
Úvod do studia biologie kmenových buněk Jiří Pacherník e-mail: jipa@sci.muni.cz tel: 532 146 223 Co jsou kmenové buňky? - buňky schopné vlastní obnovy (sebeobnova) - buňky schopné dávat vznik jiným typům
EMBRYONÁLNÍ KMENOVÉ BUŇKY
 Bi8120 Aplikovaná buněčná biologie EMBRYONÁLNÍ KMENOVÉ BUŇKY RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: vlastnosti kmenových buněk embryonální kmenové buňky linie
Bi8120 Aplikovaná buněčná biologie EMBRYONÁLNÍ KMENOVÉ BUŇKY RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: vlastnosti kmenových buněk embryonální kmenové buňky linie
Kmenové buňky, jejich vlastnosti a základní členění
 Kmenové buňky, jejich vlastnosti a základní členění O kmenových buňkách se v současné době mluví velmi často v nejrůznějších souvislostech. Je do nich vkládána naděje, že s jejich pomocí půjde vyléčit
Kmenové buňky, jejich vlastnosti a základní členění O kmenových buňkách se v současné době mluví velmi často v nejrůznějších souvislostech. Je do nich vkládána naděje, že s jejich pomocí půjde vyléčit
RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU
 Bi8120 Aplikovaná buněčná biologie i EMBRYONÁLNÍ KMENOVÉ BUŇKY RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU 1 Program přednášky: vlastnosti kmenových buněk embryonální kmenové buňky
Bi8120 Aplikovaná buněčná biologie i EMBRYONÁLNÍ KMENOVÉ BUŇKY RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU 1 Program přednášky: vlastnosti kmenových buněk embryonální kmenové buňky
Sekvenování genomu a bioinformatika Kmenové buňky vytvořené genetickou manipulací Materiálové a tkáňové inženýrství Editace genomu
 Sekvenování genomu a bioinformatika Kmenové buňky vytvořené genetickou manipulací Materiálové a tkáňové inženýrství Editace genomu Nobelova cena za fyziologii a medicínu 2012 Oct4 Sox2 c-myc Klf4 Kmenové
Sekvenování genomu a bioinformatika Kmenové buňky vytvořené genetickou manipulací Materiálové a tkáňové inženýrství Editace genomu Nobelova cena za fyziologii a medicínu 2012 Oct4 Sox2 c-myc Klf4 Kmenové
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
VÝVOJOVÁ BIOLOGIE. I. Úvod do vývojové biologie. II. Základní principy a mechanismy vývojové biologie. III. Kmenové buňky
 PŘEDNÁŠKOVÝ BLOK VÝVOJOVÁ BIOLOGIE I. Úvod do vývojové biologie II. Základní principy a mechanismy vývojové biologie III. Kmenové buňky IV. Růstové faktory a signální transdukce Kmenové buňky: definice
PŘEDNÁŠKOVÝ BLOK VÝVOJOVÁ BIOLOGIE I. Úvod do vývojové biologie II. Základní principy a mechanismy vývojové biologie III. Kmenové buňky IV. Růstové faktory a signální transdukce Kmenové buňky: definice
Embryonální období. Martin Špaček. Odd. histologie a embryologie
 Modul IB Embryonální období Martin Špaček Odd. histologie a embryologie Zdroje obrázků: Moore, Persaud: Zrození člověka Rarey, Romrell: Clinical human embryology Scheinost: Digitální zobrazování počátků
Modul IB Embryonální období Martin Špaček Odd. histologie a embryologie Zdroje obrázků: Moore, Persaud: Zrození člověka Rarey, Romrell: Clinical human embryology Scheinost: Digitální zobrazování počátků
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Nádorové kmenové buňky - CSCs (Cancer stem cells)
") Nádorové kmenové buňky - CSCs (Cancer stem cells) Původ CSCs? a) somatické kmenové buňky b) TA buňky (progenitory)* Podstatou je akumulace chyb v regulaci diferenciace, proliferace a apoptósy. Tyto chyby
Nádorové kmenové buňky - CSCs (Cancer stem cells) Původ CSCs? a) somatické kmenové buňky b) TA buňky (progenitory)* Podstatou je akumulace chyb v regulaci diferenciace, proliferace a apoptósy. Tyto chyby
(Vývojová biologie) Embryologie. Jiří Pacherník
 Embryologie. Jiří Pacherník") (Vývojová biologie) Embryologie Jiří Pacherník jipa@sci.muni.cz Podpořeno projektem FRVŠ 524/2011 buňka -> tkáně -> orgány -> organismus / jedinec Základní procesy na buněčné úrovni dělení buněk proliferace
(Vývojová biologie) Embryologie Jiří Pacherník jipa@sci.muni.cz Podpořeno projektem FRVŠ 524/2011 buňka -> tkáně -> orgány -> organismus / jedinec Základní procesy na buněčné úrovni dělení buněk proliferace
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Základní morfogenetické procesy
 Základní morfogenetické procesy 502 Základní morfogenetické procesy Mechanismy, které se uplatňují v ontogenesi, tedy při vývoji jedince od zygoty k mnohobuněčnému organismu Buněčná úroveň diferenciace
Základní morfogenetické procesy 502 Základní morfogenetické procesy Mechanismy, které se uplatňují v ontogenesi, tedy při vývoji jedince od zygoty k mnohobuněčnému organismu Buněčná úroveň diferenciace
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Přednáška v rámci cyklu přednášek pro střední školy
 Přednáška v rámci cyklu přednášek pro střední školy 1. 12. 2010 www.lf1.cuni.cz Není buňka jako buňka (některá je kmenová) Význam a výzkum kmenových buněk Eliška Krejčí Anatomický ústav 1.LF UK Není buňka
Přednáška v rámci cyklu přednášek pro střední školy 1. 12. 2010 www.lf1.cuni.cz Není buňka jako buňka (některá je kmenová) Význam a výzkum kmenových buněk Eliška Krejčí Anatomický ústav 1.LF UK Není buňka
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Neurální kmenové buňky NSCs (Neural stem cells)
") Neurální kmenové buňky NSCs (Neural stem cells) a) b) Gl glomerularní vrstva Gr granulární vrstva EPl vnější plexiformní vrstva Mi vrstva mitral buněk IPL vnitřní plexiformní vrstva cc - corpus callosum
Neurální kmenové buňky NSCs (Neural stem cells) a) b) Gl glomerularní vrstva Gr granulární vrstva EPl vnější plexiformní vrstva Mi vrstva mitral buněk IPL vnitřní plexiformní vrstva cc - corpus callosum
Lidské embryonální kmenové buňky Biologie, technologie a další
 Lidské embryonální kmenové buňky Biologie, technologie a další Aleš Hampl Ústav experimentální medicíny AV ČR, v.v.i. Lékařská fakulta, Masarykova Univerzita, Brno Centrum buněčné terapie a tkáňových náhrad,
Lidské embryonální kmenové buňky Biologie, technologie a další Aleš Hampl Ústav experimentální medicíny AV ČR, v.v.i. Lékařská fakulta, Masarykova Univerzita, Brno Centrum buněčné terapie a tkáňových náhrad,
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
BIOLOGICKÉ A MOLEKULÁRNÍ EFEKTY INHIBICE AKTIVITY CYKLIN DEPENDENTNÍCH KINÁZ U LIDSKÝCH EMBRYONÁLNÍCH KMENOVÝCH BUNĚK
 BAKALÁŘSKÁ PÍSEMNÁ PRÁCE MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA Ústav experimentální biologie Oddělení genetiky a molekulární biologie BIOLOGICKÉ A MOLEKULÁRNÍ EFEKTY INHIBICE AKTIVITY CYKLIN DEPENDENTNÍCH
BAKALÁŘSKÁ PÍSEMNÁ PRÁCE MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA Ústav experimentální biologie Oddělení genetiky a molekulární biologie BIOLOGICKÉ A MOLEKULÁRNÍ EFEKTY INHIBICE AKTIVITY CYKLIN DEPENDENTNÍCH
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčné dělení ŘÍZENÍ BUNĚČNÉHO CYKLU
 BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
Transgeneze u ptáků: očekávání vs. realita
 Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
Transgeneze u ptáků: očekávání vs. realita Proč ptáci? Kuře - základní model v anatomii, embryologii, vývojové biologii množství získaného proteinu nižší riziko cross reaktivity s tím spojená možnost produkce
VÝZNAM FYZIOLOGICKÉ OBNOVY BUNĚK V MEDICÍNĚ
 OBNOVA A REPARACE 1 VÝZNAM FYZIOLOGICKÉ OBNOVY BUNĚK V MEDICÍNĚ Příklad: Fyziologická obnova buněk: obnova erytrocytů Rychlost obnovy: 2 miliony nových erytrocytů/s (při průměrné době života erytrocytu
OBNOVA A REPARACE 1 VÝZNAM FYZIOLOGICKÉ OBNOVY BUNĚK V MEDICÍNĚ Příklad: Fyziologická obnova buněk: obnova erytrocytů Rychlost obnovy: 2 miliony nových erytrocytů/s (při průměrné době života erytrocytu
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Kmenové buňky a strategie buněčné terapie
 Kmenové buňky a strategie buněčné terapie 8. 3. 2013 Vyjmenujte co nejvíce rozdílů mezi embryonálními kmenovými buňkami a tzv. dospělými kmenovými buňkami Kmenové buňky: kriteria a definice Sebeobnova
Kmenové buňky a strategie buněčné terapie 8. 3. 2013 Vyjmenujte co nejvíce rozdílů mezi embryonálními kmenovými buňkami a tzv. dospělými kmenovými buňkami Kmenové buňky: kriteria a definice Sebeobnova
Molekulární procesy po fertilizacinormální či abnormální po ART?
 Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
AUG STOP AAAA S S. eukaryontní gen v genomové DNA. promotor exon 1 exon 2 exon 3 exon 4. kódující oblast. introny
 eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
eukaryontní gen v genomové DNA promotor exon 1 exon 2 exon 3 exon 4 kódující oblast introny primární transkript (hnrna, pre-mrna) postranskripční úpravy (vznik maturované mrna) syntéza čepičky AUG vyštěpení
Intracelulární detekce Foxp3
 Intracelulární detekce Foxp3 Ústav imunologie 2.LFUK a FN Motol Daniela Rožková, Jan Laštovička T regulační lymfocyty (Treg) Jsou definovány funkčně svou schopností potlačovat aktivaci a proliferaci CD4+
Intracelulární detekce Foxp3 Ústav imunologie 2.LFUK a FN Motol Daniela Rožková, Jan Laštovička T regulační lymfocyty (Treg) Jsou definovány funkčně svou schopností potlačovat aktivaci a proliferaci CD4+
VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ
 REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
REGULACE APOPTÓZY 1 VÝZNAM REGULACE APOPTÓZY V MEDICÍNĚ Příklad: Regulace apoptózy: protein p53 je klíčová molekula regulace buněčného cyklu a regulace apoptózy Onemocnění: více než polovina (70-75%) nádorů
Univerzita Karlova v Praze. Přírodovědecká fakulta
 Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Biologie Eliška Svobodová Charakterizace prasečích indukovaných pluripotentních kmenových buněk Chracterisation
Univerzita Karlova v Praze Přírodovědecká fakulta Studijní program: Biologie Studijní obor: Biologie Eliška Svobodová Charakterizace prasečích indukovaných pluripotentních kmenových buněk Chracterisation
Histogeneze příklady. 151 Kurs 5: Vývoj buněk a tkání
 Histogeneze příklady 151 Kurs 5: Vývoj buněk a tkání Kurs 5: Vývoj buněk a tkání 137 Kasuistika: Thalidomide 138 Základní morfogenetické procesy 139 Regenerace a reparace 140 Ženský reprodukční systém
Histogeneze příklady 151 Kurs 5: Vývoj buněk a tkání Kurs 5: Vývoj buněk a tkání 137 Kasuistika: Thalidomide 138 Základní morfogenetické procesy 139 Regenerace a reparace 140 Ženský reprodukční systém
Bi8120 Aplikovaná buněčná biologie DIFERENCIACE BUNĚK. RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU
 Bi8120 Aplikovaná buněčná biologie DIFERENCIACE BUNĚK RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: diferenciační potenciál buněk diferenciační dráhy růstové faktory
Bi8120 Aplikovaná buněčná biologie DIFERENCIACE BUNĚK RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: diferenciační potenciál buněk diferenciační dráhy růstové faktory
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Endocytóza o regulovaný transport látek v buňce
 . Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
. Endocytóza o regulovaný transport látek v buňce Exocytóza BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace
BUNĚČNÝ CYKLUS. OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky. Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí
 (1 BUNĚČNÝ CYKLUS BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace kontrolní body molekulární brzdy Jednobuněčné
(1 BUNĚČNÝ CYKLUS BUNĚČNÝ CYKLUS OMNIS CELLULA ET CELLULA - buňka vzniká jen z buňky Sled akcí, ve kterých buňka zdvojí svůj obsah a pak se rozdělí systém regulace kontrolní body molekulární brzdy Jednobuněčné
Výzkum kmenových buněk ve světle Úmluvy Martin Šolc 1/24
 Výzkum kmenových buněk ve světle Úmluvy 2. 11. 2017 Martin Šolc 1/24 Obsah Kmenové buňky Přípustnost embryodestruktivního výzkumu Úmluva Zákon č. 227/2006 Sb., o výzkumu embryonálních kmenových buňkách
Výzkum kmenových buněk ve světle Úmluvy 2. 11. 2017 Martin Šolc 1/24 Obsah Kmenové buňky Přípustnost embryodestruktivního výzkumu Úmluva Zákon č. 227/2006 Sb., o výzkumu embryonálních kmenových buňkách
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY
 BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
BUNĚČNÁ TRANSFORMACE A NÁDOROVÉ BUŇKY 1 VÝZNAM BUNĚČNÉ TRANSFORMACE V MEDICÍNĚ Příklad: Buněčná transformace: postupná kumulace genetických změn Nádorové onemocnění: kolorektální karcinom 2 3 BUNĚČNÁ TRANSFORMACE
8 cyklinů (A, B, C, D, E, F, G a H) - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů
 - v jednotlivých fázích buněčného cyklu jsou přítomny určité typy cyklinů") Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Buněč ěčné dělení BUNĚČ ĚČNÝ CYKLUS ŘÍZENÍ BUNĚČ ĚČNÉHO CYKLU cykliny a na cyklinech závislé proteinkinázy (Cyclin-Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího
Gametogenese a fertilizace. Vývoj 142
 Gametogenese a fertilizace Vývoj 142 Gamety pohlavní buňky Gametogenese diferenciace vysoce specializovaných pohlavních buněk schopných po fertilizaci vytvořit nového jedince Vajíčko (ovum) Spermie 1.
Gametogenese a fertilizace Vývoj 142 Gamety pohlavní buňky Gametogenese diferenciace vysoce specializovaných pohlavních buněk schopných po fertilizaci vytvořit nového jedince Vajíčko (ovum) Spermie 1.
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
Termíny z časného vývoje zárodku:
 Embryologie Termíny z časného vývoje zárodku: Ovulace 14.den menstruačního cyklu Oplodnění fertilizace vznik zygoty a dokončení 2. zracího dělení Rýhování mitotické dělení buněk (blastomer) a vznik moruly
Embryologie Termíny z časného vývoje zárodku: Ovulace 14.den menstruačního cyklu Oplodnění fertilizace vznik zygoty a dokončení 2. zracího dělení Rýhování mitotické dělení buněk (blastomer) a vznik moruly
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Embryologie III. Vývoj žloutkového váčku, amnion, chorion. Extraembryonální coelom. Ústav pro histologii a embryologii 1.LF Univerzity Karlovy
 Embryologie III Vývoj žloutkového váčku, amnion, chorion. Extraembryonální coelom. Ústav pro histologii a embryologii 1.LF Univerzity Karlovy Přednášející: Doc. MUDr. Tomáš Kučera, Ph.D. Předmět: Obecná
Embryologie III Vývoj žloutkového váčku, amnion, chorion. Extraembryonální coelom. Ústav pro histologii a embryologii 1.LF Univerzity Karlovy Přednášející: Doc. MUDr. Tomáš Kučera, Ph.D. Předmět: Obecná
IMUNOGENETIKA I. Imunologie. nauka o obraných schopnostech organismu. imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány
 IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
IMUNOGENETIKA I Imunologie nauka o obraných schopnostech organismu imunitní systém heterogenní populace buněk lymfatické tkáně lymfatické orgány lymfatická tkáň thymus Imunita reakce organismu proti cizorodým
Výskyt MHC molekul. RNDr. Ivana Fellnerová, Ph.D. ajor istocompatibility omplex. Funkce MHC glykoproteinů
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc = ajor istocompatibility omplex Skupina genů na 6. chromozomu (u člověka) Kódují membránové glykoproteiny, tzv. MHC molekuly, MHC molekuly
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Ontogeneze živočichů
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Ontogeneze živočichů postembryonální vývoj 1/73 Ontogeneze živočichů = individuální vývoj živočichů, pokud vznikají
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Ontogeneze živočichů postembryonální vývoj 1/73 Ontogeneze živočichů = individuální vývoj živočichů, pokud vznikají
MECHANIZMY EPIGENETICKÝCH PROCESŮ
 MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
MECHANIZMY EPIGENETICKÝCH PROCESŮ METYLACE DNA Metylace DNA Adice metylové skupiny (CH 3 ) na 5. uhlík cytosinu Obvykle probíhá pouze na cytosinech 5 vůči guanosinu (CpG) Cytosin NH 2 5-Metylcytosin NH
Gastrulace, neurulace, somitogenese 508
 Gastrulace, neurulace, somitogenese 508 Gastrulace Zásadní děj vývoje - 3. týden Tvorba intraembryonálního mesodermu: Proliferace epiblastu Kaudální morfogenetické centrum: o o Primitivní (Hensenův) uzel
Gastrulace, neurulace, somitogenese 508 Gastrulace Zásadní děj vývoje - 3. týden Tvorba intraembryonálního mesodermu: Proliferace epiblastu Kaudální morfogenetické centrum: o o Primitivní (Hensenův) uzel
Specifická imunitní odpověd. Veřejné zdravotnictví
 Specifická imunitní odpověd Veřejné zdravotnictví MHC molekuly glykoproteiny exprimovány na všech jaderných buňkách (MHC I) nebo jenom na antigen prezentujících buňkách (MHC II) u lidí označovány jako
Specifická imunitní odpověd Veřejné zdravotnictví MHC molekuly glykoproteiny exprimovány na všech jaderných buňkách (MHC I) nebo jenom na antigen prezentujících buňkách (MHC II) u lidí označovány jako
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Genetická kontrola prenatálního vývoje
 Genetická kontrola prenatálního vývoje Vývoj jedince Individuální vývoj začíná okamžikem oplození Předchází gametogeneze u obou pohlaví meióza, včetně procesu crosing over a rekombinace Vývoj jedince pak
Genetická kontrola prenatálního vývoje Vývoj jedince Individuální vývoj začíná okamžikem oplození Předchází gametogeneze u obou pohlaví meióza, včetně procesu crosing over a rekombinace Vývoj jedince pak
IV117: Úvod do systémové biologie
 IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
IV117: Úvod do systémové biologie David Šafránek 3.12.2008 Obsah Obsah Robustnost chemotaxe opakování model chemotaxe bakterií nerozliseny stavy aktivity represoru aktivita = ligandy a konc. represoru
Univerzita Karlova vpraze Přírodovědecká Fakulta
 Univerzita Karlova vpraze Přírodovědecká Fakulta Studijní program: Biologie Studijní obor: Buněčná a vývojová biologie Bc. Eliška Svobodová Příprava prasečích indukovaných pluripotentních kmenových buněk
Univerzita Karlova vpraze Přírodovědecká Fakulta Studijní program: Biologie Studijní obor: Buněčná a vývojová biologie Bc. Eliška Svobodová Příprava prasečích indukovaných pluripotentních kmenových buněk
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince. Co bylo dřív? Slepice nebo vejce?
 REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
ONKOGENETIKA. Spojuje: - lékařskou genetiku. - buněčnou biologii. - molekulární biologii. - cytogenetiku. - virologii
 ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
Variace Vývoj dítěte
 Variace 1 Vývoj dítěte 21.7.2014 16:25:04 Powered by EduBase BIOLOGIE ČLOVĚKA VÝVOJ DÍTĚTE OPLOZENÍ A VÝVOJ PLACENTY Oplození K oplození dochází ve vejcovodu. Pohyb spermií: 3-6 mm za minutu. Životnost
Variace 1 Vývoj dítěte 21.7.2014 16:25:04 Powered by EduBase BIOLOGIE ČLOVĚKA VÝVOJ DÍTĚTE OPLOZENÍ A VÝVOJ PLACENTY Oplození K oplození dochází ve vejcovodu. Pohyb spermií: 3-6 mm za minutu. Životnost
MUDr. Iva Slaninová, Ph.D. Biologický ústav LF
 MUDr. Iva Slaninová, Ph.D. Biologický ústav LF MU ipokorna@med. @med.muni.cz Hierarchie buněk v tkáních Kmenové buňky self-renewal - klon Progenitorové buňky s každým dělením více diferencované Progenitor
MUDr. Iva Slaninová, Ph.D. Biologický ústav LF MU ipokorna@med. @med.muni.cz Hierarchie buněk v tkáních Kmenové buňky self-renewal - klon Progenitorové buňky s každým dělením více diferencované Progenitor
Inovace profesní přípravy budoucích učitelů chemie
 Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Inovace profesní přípravy budoucích učitelů chemie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í CZ.1.07/2.2.00/15.0324 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
2.ročník - Zoologie. Rozmnožování Zárodečné listy (10)
") 2.ročník - Zoologie Rozmnožování Zárodečné listy (10) ROZMNOŽOVÁNÍ A VÝVIN ROZMNOŽOVÁNÍ: 1) Nepohlavní = zachována stejná genetická informace rodiče a potomka - založeno na schopnosti regenerace (obnovy
2.ročník - Zoologie Rozmnožování Zárodečné listy (10) ROZMNOŽOVÁNÍ A VÝVIN ROZMNOŽOVÁNÍ: 1) Nepohlavní = zachována stejná genetická informace rodiče a potomka - založeno na schopnosti regenerace (obnovy
Příprava vektoru IZOLACE PLASMIDU ALKALICKÁ LYZE, KOLONKOVÁ IZOLACE DNA GELOVÁ ELEKTROFORÉZA RESTRIKČNÍ ŠTĚPENÍ. E. coli. lyze buňky.
 Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
Příprava vektoru IZOLCE PLSMIDU LKLICKÁ LYZE, KOLONKOVÁ IZOLCE DN E. coli plasmidová DN proteiny proteiny + + vysrážená plasmidová lyze buňky + snížení ph chromosomální DN centrifugace DN chromosomální
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Buněčný cyklus a molekulární mechanismy onkogeneze
 Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Buněčný cyklus a molekulární mechanismy onkogeneze Imunofluorescence DAPI Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny s homologními oblastmi; jejich
Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR. Mgr. Silvie Dudová
 Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR Mgr. Silvie Dudová Centrum základního výzkumu pro monoklonální gamapatie a mnohočetný myelom, ILBIT LF MU Brno Laboratoř experimentální
Lekce z analýz genových expresních profilů u MM a návrh panelu genů pro ČR Mgr. Silvie Dudová Centrum základního výzkumu pro monoklonální gamapatie a mnohočetný myelom, ILBIT LF MU Brno Laboratoř experimentální
Hematologie. Nauka o krvi Klinická hematologie Laboratorní hematologie. -Transfuzní lékařství - imunohematologie. Vladimír Divoký
 Hematologie Nauka o krvi Klinická hematologie Laboratorní hematologie -Transfuzní lékařství - imunohematologie Vladimír Divoký Fyzikální vlastnosti krve 3-4 X více viskózní než voda ph : 7.35 7.45 4-6
Hematologie Nauka o krvi Klinická hematologie Laboratorní hematologie -Transfuzní lékařství - imunohematologie Vladimír Divoký Fyzikální vlastnosti krve 3-4 X více viskózní než voda ph : 7.35 7.45 4-6
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Výukové materiály: http://www.zoologie.upol.cz/osoby/fellnerova.htm Prezentace navazuje na základní znalosti Biochemie a cytologie. Bezprostředně
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Výukové materiály: http://www.zoologie.upol.cz/osoby/fellnerova.htm Prezentace navazuje na základní znalosti Biochemie a cytologie. Bezprostředně
8. Východoslovenský bioptický seminár SD IAP, Košice Květa Michalová
 8. Východoslovenský bioptický seminár SD IAP, Košice 2018 Květa Michalová Tumor levého varlete 65 let Bez udání velikosti Inhibin SALL4 Nanog OCT3/4 DMRT1 Diagnóza: Smíšený nádor z germinálních a
8. Východoslovenský bioptický seminár SD IAP, Košice 2018 Květa Michalová Tumor levého varlete 65 let Bez udání velikosti Inhibin SALL4 Nanog OCT3/4 DMRT1 Diagnóza: Smíšený nádor z germinálních a
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
INTRACELULÁRNÍ SIGNALIZACE II
 INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
INTRACELULÁRNÍ SIGNALIZACE II 1 VÝZNAM INTRACELULÁRNÍ SIGNALIZACE V MEDICÍNĚ Příklad: Intracelulární signalizace: aktivace Ras proteinu (aktivace receptorové kinázy aktivace Ras aktivace kinázové kaskády
FERTILIZACE A EMBRYOGENEZE
 Bi8120 Aplikovaná buněčná biologie FERTILIZACE A EMBRYOGENEZE RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: meióza a gametogeneze mechanismy fertilizace časná embryogeneze
Bi8120 Aplikovaná buněčná biologie FERTILIZACE A EMBRYOGENEZE RNDr. Jakub Neradil, Ph.D. Ústav experimentální biologie PřF MU Program přednášky: meióza a gametogeneze mechanismy fertilizace časná embryogeneze
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Apoptóza Onkogeny. Srbová Martina
 Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Apoptóza Onkogeny Srbová Martina Buněčný cyklus Regulace buněčného cyklu 1. Cyklin-dependentní kináza (Cdk) cyclin Regulace buněčného cyklu 2. Retinoblastomový protein (prb) E2F Regulace buněčného cyklu
Některé významné aspekty vývojové biologie v medicíně
 Některé významné aspekty vývojové biologie v medicíně - terapie infertility (in vitro oplození) - genetické poruchy vývoje člověka - malformace (Down, Waardenburg, Martin-Bell) - teratogeneze (alkohol,
Některé významné aspekty vývojové biologie v medicíně - terapie infertility (in vitro oplození) - genetické poruchy vývoje člověka - malformace (Down, Waardenburg, Martin-Bell) - teratogeneze (alkohol,
Aktuální výzkum kmenových buněk: ze zkumavky k terapeutickému využití
 Od fyziologie k medicíně Aktuální výzkum kmenových buněk: ze zkumavky k terapeutickému využití Tomáš Bárta, Dáša Doležalová, Aleš Hampl, Zuzana Holubcová, Josef Jaroš, Vladimír Vinarský Projekt Od fyziologie
Od fyziologie k medicíně Aktuální výzkum kmenových buněk: ze zkumavky k terapeutickému využití Tomáš Bárta, Dáša Doležalová, Aleš Hampl, Zuzana Holubcová, Josef Jaroš, Vladimír Vinarský Projekt Od fyziologie
Syllabus pro dosažení kvalifikace klinického embryologa
 Syllabus pro dosažení kvalifikace klinického embryologa Blok 1. Základní pojmy: buněčná biologie, molekulární biologie a genetika 1.1 Buňka Vnitřní organizace Řízení buněčného cyklu, kontrolní body Mitóza
Syllabus pro dosažení kvalifikace klinického embryologa Blok 1. Základní pojmy: buněčná biologie, molekulární biologie a genetika 1.1 Buňka Vnitřní organizace Řízení buněčného cyklu, kontrolní body Mitóza
VZTAH DÁRCE A PŘÍJEMCE
 TRANSPLANTAČNÍ IMUNITA Transplantace je přenos buněk, tkáně nebo orgánu z jedné části těla na jinou nebo z jednoho jedince na jiného. Transplantační reakce je dána genetickými rozdíly mezi dárcem a příjemcem.
TRANSPLANTAČNÍ IMUNITA Transplantace je přenos buněk, tkáně nebo orgánu z jedné části těla na jinou nebo z jednoho jedince na jiného. Transplantační reakce je dána genetickými rozdíly mezi dárcem a příjemcem.
Základy molekulární biologie KBC/MBIOZ
 Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
Základy molekulární biologie KBC/MBIOZ Mária Čudejková 2. Transkripce genu a její regulace Transkripce genetické informace z DNA na RNA Transkripce dvou genů zachycená na snímku z elektronového mikroskopu.
UNIVERZITA KARLOVA V PRAZE LÉKAŘSKÁ FAKULTA V PLZNI Ústav patologické fyziologie
 UNIVERZITA KARLOVA V PRAZE LÉKAŘSKÁ FAKULTA V PLZNI Ústav patologické fyziologie MOŽNOSTI A VÝZNAM DLOUHODOBÉ KULTIVACE EMBRYONÁLNÍCH BUNĚK RŮZNÝCH ŽIVOČIŠNÝCH DRUHŮ ABILITIES AND IMPORTANCE OF LONG-TERM
UNIVERZITA KARLOVA V PRAZE LÉKAŘSKÁ FAKULTA V PLZNI Ústav patologické fyziologie MOŽNOSTI A VÝZNAM DLOUHODOBÉ KULTIVACE EMBRYONÁLNÍCH BUNĚK RŮZNÝCH ŽIVOČIŠNÝCH DRUHŮ ABILITIES AND IMPORTANCE OF LONG-TERM
Od fyziologie k medicíně
 Milý čtenáři, publikace, kterou držíte v ruce je součástí olomouckého cyklu vzdělávacích materiálů vydávaných k projektu Od fyziologie k medicíně integrace vědy, výzkumu odborného vzdělávání a praxe. Projekt
Milý čtenáři, publikace, kterou držíte v ruce je součástí olomouckého cyklu vzdělávacích materiálů vydávaných k projektu Od fyziologie k medicíně integrace vědy, výzkumu odborného vzdělávání a praxe. Projekt
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer
 Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
Rekombinantní protilátky, bakteriofágy, aptamery a peptidové scaffoldy pro analytické a terapeutické účely Luděk Eyer Virologie a diagnostika Výzkumný ústav veterinárního lékařství, v.v.i., Brno Alternativní
RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU. Jana Novotná
 RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU Jana Novotná Co jsou to cytokiny? Skupina proteinů a peptidů (glykopeptidů( glykopeptidů), vylučovaných živočišnými buňkami a ovlivňujících buněčný růst (též růstové
RECEPTORY CYTOKINŮ A PŘENOS SIGNÁLU Jana Novotná Co jsou to cytokiny? Skupina proteinů a peptidů (glykopeptidů( glykopeptidů), vylučovaných živočišnými buňkami a ovlivňujících buněčný růst (též růstové
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1
 Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
MTI Cvičení č. 2 Pasážování buněk / Jana Horáková
 MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
MTI Cvičení č. 2 Pasážování buněk 15.11./16.11.2016 Jana Horáková Doporučená literatura M. Vejražka: Buněčné kultury http://bioprojekty.lf1.cuni.cz/3381/sylabyprednasek/textova-verze-prednasek/bunecnekultury-vejrazka.pdf
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Genová etiologie nemocí
 Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Genová etiologie nemocí 1. Obecná etiologie nemocí 1. Obecná etiologie nemocí 2. Mutace genů v germinativních a somatických buňkách 3. Molekulární fyziologie genu 4. Regulace aktivity genu (genové exprese)
Buňky s velkým potenciálem 1. Historie indukované pluripotence a metody přípravy ips buněk
 Kateřina Vodičková Kepková, Petr Vodička, Jan Motlík Buňky s velkým potenciálem 1. Historie indukované pluripotence a metody přípravy ips buněk Indukované pluripotentní buňky (ipsc, induced Pluripotent
Kateřina Vodičková Kepková, Petr Vodička, Jan Motlík Buňky s velkým potenciálem 1. Historie indukované pluripotence a metody přípravy ips buněk Indukované pluripotentní buňky (ipsc, induced Pluripotent
DUM č. 3 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
projekt GML Brno Docens DUM č. 3 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: chromatin - stavba, organizace a struktura
Obecná a srovnávací odontologie. Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód
 Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
Obecná a srovnávací odontologie Vývojové souvislosti 1: vznik a vývoj zubu jako produkt genetických regulačních kaskád, odontogenní regulační kód Vývojové souvislosti 1: vznik a vývoj zubu jako produkt
EXTRACELULÁRNÍ SIGNÁLNÍ MOLEKULY
 EXTRACELULÁRNÍ SIGNÁLNÍ MOLEKULY 1 VÝZNAM EXTRACELULÁRNÍCH SIGNÁLNÍCH MOLEKUL V MEDICÍNĚ Příklad: Extracelulární signální molekula: NO Funkce: regulace vazodilatace (nitroglycerin, viagra) 2 3 EXTRACELULÁRNÍ
EXTRACELULÁRNÍ SIGNÁLNÍ MOLEKULY 1 VÝZNAM EXTRACELULÁRNÍCH SIGNÁLNÍCH MOLEKUL V MEDICÍNĚ Příklad: Extracelulární signální molekula: NO Funkce: regulace vazodilatace (nitroglycerin, viagra) 2 3 EXTRACELULÁRNÍ
DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje.
 Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
Genomika DNA se ani nezajímá, ani neví. DNA prostě je. A my tancujeme podle její muziky. Richard Dawkins: Řeka z ráje. Obor genetiky, který se snaží stanovit úplnou genetickou informaci organismu a interpretovat
SKANÁ imunita. VROZENÁ imunita. kladní znalosti z biochemie, stavby membrán n a fyziologie krve. Prezentace navazuje na základnz
 RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Prezentace navazuje na základnz kladní znalosti z biochemie, stavby membrán n a fyziologie krve Rozšiřuje témata: Proteiny přehled pro fyziologii
RNDr. Ivana Fellnerová, Ph.D. Katedra zoologie, PřF UP Olomouc Prezentace navazuje na základnz kladní znalosti z biochemie, stavby membrán n a fyziologie krve Rozšiřuje témata: Proteiny přehled pro fyziologii
TERATOGENEZA ONTOGENEZA
 TERATOGENEZA ONTOGENEZA Vrozené vývojové vady (VVV) Jsou defekty orgánů, ke kterým došlo během prenatálního vývoje plodu a jsou přítomny při narození jedince. Postihují v různém rozsahu okolo 3-5 % novorozenců.
TERATOGENEZA ONTOGENEZA Vrozené vývojové vady (VVV) Jsou defekty orgánů, ke kterým došlo během prenatálního vývoje plodu a jsou přítomny při narození jedince. Postihují v různém rozsahu okolo 3-5 % novorozenců.
Buněčný cyklus, spojení se signálními cestami a molekulární mechanismy onkogeneze
 Buněčný cyklus, spojení se signálními cestami a molekulární mechanismy onkogeneze MUDr. Jiří Vachtenheim, CSc. Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny
Buněčný cyklus, spojení se signálními cestami a molekulární mechanismy onkogeneze MUDr. Jiří Vachtenheim, CSc. Přehled regulace buněčného cyklu Základní terminologie: Cycliny evolučně konzervované proteiny
Změny v parametrech imunity v průběhu specifické alergenové imunoterapie. Vlas T., Vachová M., Panzner P.,
 Změny v parametrech imunity v průběhu specifické alergenové imunoterapie Vlas T., Vachová M., Panzner P., Mechanizmus SIT Specifická imunoterapie alergenem (SAIT), má potenciál ovlivnit imunitní reaktivitu
Změny v parametrech imunity v průběhu specifické alergenové imunoterapie Vlas T., Vachová M., Panzner P., Mechanizmus SIT Specifická imunoterapie alergenem (SAIT), má potenciál ovlivnit imunitní reaktivitu
Václav Hořejší Ústav molekulární genetiky AV ČR. IMUNITNÍ SYSTÉM vs. NÁDORY
 Václav Hořejší Ústav molekulární genetiky AV ČR IMUNITNÍ SYSTÉM vs. NÁDORY PROTINÁDOROVÁ IMUNITA - HISTORIE 1891 W. Coley - otec imunoterapie 1957 F.M. Burnet hypotéza imunitního dozoru 1976 A.W. Bruce
Václav Hořejší Ústav molekulární genetiky AV ČR IMUNITNÍ SYSTÉM vs. NÁDORY PROTINÁDOROVÁ IMUNITA - HISTORIE 1891 W. Coley - otec imunoterapie 1957 F.M. Burnet hypotéza imunitního dozoru 1976 A.W. Bruce