MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/ Analysis of population subdivision
|
|
|
- Emil Hruška
- před 6 lety
- Počet zobrazení:
Transkript
1 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/ Analysis of population subdivision (mezipopulační variabilita)
2 Hierarchická populační struktura Druh populace subpopulace (demy) Předpoklady studia populačně-genetické struktury lokusy používané pro analýzu populační struktury jsou neutrální vůči selekci klasický populačně-genetický přístup = jednotlivé populace jsou předem známy (např. chceme zjistit úroveň genetických rozdílů mezi dvěma lokalitami = populacemi)
3 Genetická struktura populací drift, mutace Drift diferenciace subpopulací díky změnám frekvencí (až fixaci) alternativních alel Aa aa Aa AA Aa AA aa Aa Aa aa Aa AA Aa aa AA Aa drift Mutace mohou zvýšit diferenciaci (nebo ne homoplázií) AA AB AA aa aa AA aa AA AA aa aa ac AA aa Migrace (genový tok) Migrace (genový tok) - působí proti diferenciaci subpopulací
4 Vliv populační struktury na heterozygotnost Wahlundův princip Dvě izolované subpopulace s fixovanými alelami Subpopulace v HW, celkově v populaci však nedostatek heterozygotů AA AA AA AA AA AA AA AA AA B A R I É R A aa aa aa aa aa aa aa aa aa aa
5 Wahlundův princip (isolate breaking) Pokles homozygotnosti při sloučení subpopulací AA AA AA AA AA AA AA AA AA Aa Aa Aa Aa Aa Aa Aa Aa Aa aa aa aa aa aa aa aa aa aa aa
 172/172 Total p 2pq (=He) Přítok 50 0")
 (33.1) (48.9) (18.1) p 2 = 0.575 2 q 2 = 0.")
6 Wahlundův princip - příklad Jezero Bunnersjöarna (severní Švédsko) brown trout 2 alely na jednom znaku 170/ /172 (= Ho) 172/172 Total p 2pq (=He) Přítok 50 0 (0) Odtok 1 13 (0.26) Celé (0.13) jezero (expected) (33.1) (48.9) (18.1) p 2 = q 2 = Ryman et al. 1979
7 Factorial correspondence analysis - each locus as one variable, reduction of number of variables - Genetix orientační zjištění strukturovanosti populace - individuals vs. populations
8 F-statistiky Masatoshi Nei *1931 Sewall Wright Wright, Nei F IS, F ST, F IT Popisují heterozygotnost (odchylky od HW) na různých měřítkách
9 Odhad vlivu populační struktury na genetický make-up populace C e l k o v á p o p u l a c e I 1 3 I 1 7 I 1 6 I 2 1 I 2 2 I 3 1 I 3 2 I 1 8 I 2 3 I 1 4 I 1 5 I 1 9 I 2 9 I 2 4 I 3 3 I 1 1 I 2 5 I 2 6 I 3 6 I 3 5 I I 1 2 I 3 4 I 2 7 I 2 8 I I 3 7 S 1 S 2 S 3 T 3 úrovně (T, S, I) x subpopulací (x = 1 až k; zde k = 3) každá subpopulace má N x jedinců AA, AB, BB odlišný symbol př. I1-13 = 13. jedinec z první subpopulace
10 Koncept heterozygotnosti H I průměrná pozorovaná heterozygotnost jedince v subpopulaci H S - očekávaná heterozygotnost jedince v subpopulaci za předpokladu náhodného páření H T - očekávaná heterozygotnost jedince v celé populaci za předpokladu náhodného páření H I = k x= 1 H x k H x = pozorovaná heterozygotnost v subpopulaci x j i= 1 H S = p i 2 1 p i,x2 = frekvence i-té, x alely v subpopulaci x H H T = 2pq 0 0 p o = frekvence alely v celé populaci S = k x = 1 H k S průměrná oček. heterozygotnost v populaci (tj. průměr ze všech subpopulací) pouze pro dvě alely na jednom lokusu (Wright 1931) pro více alel je výpočet složitější (Nei 1987)
11 F statistiky F IS = H H S H S I Snížení heterozygotnosti jedince kvůli nenáhodnému páření v subpopulaci (~ HWE) F ST = H H T H T S Vliv rozdělení populace na subpopulace F IT = H H T H T I Celkový koeficient inbreedingu F IT - měří redukci heterozygotnosti jedince ve vztahu k celkové populaci (1-F IT )= (1-F ST )(1-F IS ) Weir & Cockerham (1984) f, θ, F Korekce na velikost vzorku a počet subpopulací
12 Výpočet F statistik - příklad Průměrná frekvence alely A v celé populaci Subpopulace 1 (N 1 =40) Subpopulace 2 (N 2 =20) Lokus AA AB BB p 1(j) AA AB BB p 2(j) p 0(j) Pozn. Loc I H.-W. rovnováha Loc II deficit heterozygotů Loc III přebytek heterozygotů Loc IV alternativně fixované alely Výpočet alelových frekvencí Pozorovaná heterozygotnost Očekávaná heterozygotnost Wrightova F-statistika Lokus H 1 (j) H 2 (j) H I (j) H S (j) H T (j) F IS (j) F ST (j) F IT (j) Loc I Loc II Loc III Loc IV Průměr Průměrné hodnoty F statistik mohou maskovat odlišnou evoluční historii na různých lokusech
13 F-statistiky F IS snížení heterozygotnosti v lokální subpopulaci vysoké hodnoty inbreeding F IT souhrnná hodnota, heterozygotnost v celé populaci F ST míra rozdělenosti = snížení toku mezi subpopulacemi (tj. existence bariéry Wahlundův princip) Vliv driftu fixuje odlišné alely v subpopulacích


 1000 x")


 35.")
14 Bootstrap test významnosti Fst 1. Skutečné populace 2. Sloučení jedinců x náhodně vytvořené populace Absolutní hodnoty závisí na heterozygotnosti!!! Nutno Reálné standardizovat: Fst F ST = F ST /F STmax (Hedrick 2005) 1000 x nasimulované Fst za nepřítomnosti bariéry Silně ovlivněno nulovými alelami (např. software FREENA) Fst = 0,072 Fst = 0,0013 0,80 % simulovaných hodnot větších než skutečné Fst p = 0,008 (tj. významný rozdíl) % simulovaných hodnot větších než skutečné Fst p = 0,354 (tj. nevýznamný rozdíl)
15 Výpočet F ST - příklad A/A A/B (=Ho) B/B Total p 2pq (=He) Přítok 50 0 (0) Odtok 1 13 (0.26) Celé jezero (0.13) (expected) (33.1) (48.9) (18.1) F ST = H H H T H T S = = V důsledku bariéry toku genů je heterozygotnost o 72.8% nižší než by byla v panmiktické populaci Ryman et al. 1979
16 Zdroje chyb při analýze Fst Global vs. pairwise indices Absolutní hodnoty závisí na heterozygotnosti!!! (např. nelze srovnávat alozymy s mikrosatelity) Nutno standardizovat: F ST = F ST /F STmax (Hedrick 2005) např. GenAlex Nutno korigovat na přítomnost nulových alel (většinou uměle zvyšují Fst zvýšení homozygotnosti) - FreeNA 1/1 1/3 3/5 2/5 1/2 1/1 1/3 3/3 2/2 1/2 1/5 2/2 1/1 2/2 1/1 1/1 4/5 2/4 2/2 1/1 1/1 4/4 2/4 2/2 1/5 2/2 1/1 2/2


17 Panda velká 192 vzorků trusu 136 genotypů 53 unikátních genotypů rozdělení řekou (cca 26 tisíc let) a cestami (recentně) i cesty jsou významná bariéra, i když menší (Zhu et al., 2011)
18 Populační cykly hrabošovitých hlodavců (Berthier et al. 2006) Výrazné cyklické změny v početnosti populací střídají se období izolovanosti s kontinuálním rozšířením Arvicola terrestris Nízká hustota extrémní vliv driftu S nárůstem populační hustoty se zvyšuje intenzita toku genů mezi subpopulacemi a narůstá genetická variabilita subpopulací Snižování Fst
19 G ST (Nei 1973) Analogie F ST Haploidní (haplodiploidní) organismy, sekvence mtdna Počítá s gene diversity místo heterozygotnosti Pracuje tedy jen s frekvencemi alel, Pracuje tedy jen s frekvencemi alel, ne s procentem heterozygotů
20 Myotis bechsteinii Kerth et al Letní kolonie samic (15-40 jedinců) Vzorky - jen Nukleární i mitochondriální mikrosatelity filopatrie disperze
21 R ST Obdoba F ST Pracuje však s velikostí alel (počet repeatů u mikrosatelitů) Předpoklad známého mutačního modelu jen při platnosti SMM (stepwise mutation model) Paměť mutací v minulosti R ST >F ST větší vliv mutací R ST =F ST větší vliv driftu Potvrzení významnosti rozdílu randomizačními testy (Hardy et al. 2003, program SPAGeDi 1.1)

 alel Program ARLEQUIN Data: sekvence")
22 AMOVA Excoffier et al Analýza Molekulární Variance Analýza variance alelických frekvencí (již dříve Cockerham & Weir 1987,1993) Započítává se ale rozdílnost (mutace) alel Program ARLEQUIN Data: sekvence mikrosatelity (jen při platnosti SMM stepwise mutation model)



23 Bombus pascuorum Widmer & Schmid-Hempel 1999 Mikrosatelity, AMOVA Nejvíc vysvětlují Alpy
24 Genetické distance počítané z mikrosatelitů Cavali-Sforza distance Nei distance matice vzdáleností použita jako podklad pro shlukovou analýzu dendrogram

25 Genetic distance - Neighbour-joining
26 AMOVA a F statistika popis výsledku nikoliv příčin možná alternativní vysvětlení
27 Tok genů metody 1. Přímé metody: Capture-Mark-Recapture + parentage analýzy
28 2. Nepřímé metody stanovení toku genů populačně-genetické modely Island model (Wright) Stejně velké subpopulace Symetrický tok genů Stejná pravděpodobnost výměny mezi jakýmikoliv subpopulacemi Stepping-stone model (Kimura) Výměna jen mezi sousedními subpopulacemi
29 Tok genů metody 1. Přímé metody: Capture-Mark-Recapture + parentage analýzy 2. Nepřímé metody na základě distribuce genetické variability N e m = počet migrantů / subpopulace / generace ( island model!) Jde o hrubý odhad ve škále: málo, středně, hodně! Privátní alely (Slatkin 1985) vhodné pro vysoce polymorfní znaky Alely vyskytující se jen v jedné subpopulaci p(1) frekvence privátních alel lnp(1) = -0,505 ln(n e m) F statistika F ST N = (jen pro Fst > ) e m
30 Předpoklady použití N e m: island model (= infinite number of populations, absence of selection, the same size of all populations etc.) migration-drift equilibrium (= no range expansions, habitat fragmentation or population bottlenecks) Co ale rozhodně NE! Dvě hodně vzdálené populace F ST vždy bude nenulové N e m v minulosti došlo k výměně jedinců I populace, které si nikdy nevyměnily migranty mohou mít nenulové N e m



31 BayesASS Panda velká - Bayesian estimates of gene flow - doporučení pro ochranářský management stavba koridorů
32 Modely toku genů (alel) Island model (Wright) Stejně velké subpopulace Symetrický tok genů Stejná pravděpodobnost výměny mezi jakýmikoliv subpopulacemi Stepping-stone model (Kimura) Výměna jen mezi sousedními subpopulacemi Isolation by distance Tok slábne se vzrůstající vzdáleností subpopulací
33 Isolation by distance rozumné geografické měřítko (závisí na schopnosti disperze) musí být ustanovena rovnováha mezi migrací a driftem IBD (isolation-by-distance) nebude u velmi recentně izolovaných populací u zcela izolovaných populací při značné migraci




34 Isolation by distance Crotaphytus collaris Hutchinson & Templeton 1999 desítky tisíc let nejsou bariéry rovnováha mezi driftem a migrací postglaciálně nejsou bariéry vliv migrace postglaciálně fragmentace vliv driftu postglaciálně vzrůstající fragmentace vliv driftu na velkých měřítkách rovnováha na malých měřítkách
35 Detekce IBD korelace mezi maticí genetických a geografických vzdáleností Mantelův test např. Genepop
36 Příklad: IBD u netopýrů rodu Pipistrellus
37 Genetika metapopulací Metapopulace: časté lokální extinkce a kolonizace Jednoduchá populační struktura vs. metapopulace populační genetika stabilní vs. nestabilní populačně-genetický pattern

38 Atlantic salmon Sainte Marguerite river (Quebec) (Garant et al. 2000) výrazná populační struktura mezi řekami v rámci jedné řeky různé mikrohabitaty obývané (1) demy nebo (2) metapopulacemi? dvouleté sledování, mikrosatelity, hierarchická AMOVA lokálně adaptované demy, ale i metapopulace (např. po povodních je mikrohabitat obsazen náhodně a ne jedinci z původního demu) long-term (stable) vs. short-term (metapopulations) struktura Variance component D.f. % of total variance F-statistic P Mezi přítoky 1 0,1 0,0014 0,167 Mezi lokalitami 6 0,9 0,0085 0,001 Mezi roky uvnitř lokality 7 2,5 0,0255 < 0,0001 Uvnitř vzorku ,6 0,0337 < 0,0001
39 Který typ metapopulační struktury? classical mainland-island source and sinks ad a) Cristatella mucedo - Bryozoa (Freeland et al. 2000) longdistance migration (birds), no IBD, 5 ms for clone identification black rats in Senegal transport by traffic no IBD (Konečný et al. 2012)
 různé rybníky mají různý reprodukční úspěch klasické subpopulace (Area 1) i")
 Klasická subpopulační struktura + IBD Mainland-island")
40 ad b) Mainland-island model Bufo calamita (Rowe et al. 2000) různé rybníky mají různý reprodukční úspěch klasické subpopulace (Area 1) i metapopulace (Area 2); křížek vyšší tok genů než by se předpokládalo (jsou v nich obsaženy vysoce úspěšné rybníky = mainland ) Klasická subpopulační struktura + IBD Mainland-island metapopulace F ST = F ST = N m
41 Populační struktura - shrnutí Connected populations (gene flow) Isolated populations (no gene flow) N e Genetic drift Genetic diversity Population differentiation
42 1. vnitropopulační variabilita deskriptivní statistiky 2. popis populačně-genetické struktury, bariéry toku genů 3. Population assignments Klasické problémy populační genetiky Populace dány, jedinci předem zařazeni do populací, zajímají nás vlastnosti populací (Fstatistiky) ad (1) nebo (2) Populace sice definovány, ale chceme k nim přiřadit jedince neznámého původu Kryptická populační struktura = předem není dáno Kryptická populační struktura = předem není dáno nic chci zjistit klastry (tj. přirozené populace) a rozřadit individua do klastrů (population assignments)
43 Př.: Unraveling migratory connectivity (a) strong vs. (b) weak connectivity
44 A. Direct methods morphological variation (geographical races) leg-bands or similar markers (ex. over one million Ficedula hypoleuca have been ringed in UK and Sweden only six recaptured on wintering grounds in Africa satellite telemetry expensive, not useful for small animals B. Biogeochemical approaches ratios of stable isotopes of naturally occurring elements (C, H, N, Sr) vary across the landscape determined by the relative frequency of C3 and C4 plants, climate, and bedrock (1) geographical structure of isotopic ratio distributions (2) knowledge about where animals incorporate isotopes (3) tissue samples from individuals at different parts of their annual cycle
45 C. Genetic approaches «very few birds have bands, but all have genotypes» genetic data on population structure problems: (1) week genetic differentiation among populations (widespread dispersal), (2) lack of differentiation in northern temperate zone recent postglaciation expansion Řešení: (1) použití více genetických znaků, (2) studium DNA parazitů (např. avian malaria) mají rychlejší evoluci, jsou více diferencovaní
46 Population assignment tests program GeneClass (Piry et al. 2004) calculates the probability that an individual s genotype might exist in a particular population identifikace recentních migrantů nebo zvířat neznámého původu (pytláctví) can combine data from multiple genetic marker types 5 microsatellite loci 5 microsatellite loci F st = 0.14 F st = % assigned correctly 90.2% assigned correctly
47 Individual-based assignments Kryptická populační struktura Neznámý počet skupin (klastrů) Úroveň jedince Vytvořit klastry a současně k nim přiřadit jedince K dispozici máme individuální genotypy (případně i souřadnice) Data: msat (jiné kodominantní lokusy, SINE), AFLP








48
49 Dendrogram založený na mikrosatelitových distancích (Cavali-Sforza) Může být biased pokud máme málo znaků
50 Bayesian clustering approach STRUCTURE - Pritchard et al Neznámý počet populací charakterizovaných různými frekvencemi alel počet populací a frekvence zjišťuji Současně přiřazuji individua do populací Lokusy, které nejsou ve vazbě, HW uvnitř subpopulací (např. mikrosatelity, SNPs) Model se snaží vyložit HW nebo vazebnou nerovnováhu zavedením populační struktury Místo přímého výpočtu odhad pomocí Markov chain Monte Carlo nutno opakovat několik běhů pro každý model
51 Turdus helleri Fragmenty deštného pralesa Lokality Chawia, Ngangao, Mbololo, Yale (Kenya) 7 mikrosatelitových lokusů Neighbour-joining * špatně zařazení jedinci Shlukovací analýza na základě mikrosatelitových distancí
52 Program STRUCTURE - Bayesiánský přístup recentní migranti hybrid? Q-values (pravděpodobnost přiřazení k danému clusteru)
53 Admixture model umožňuje zařazení jedince do více klastrů Barplot pro K = 7 Q-value Proporce genomu každého jedince náležející určitému clusteru
54 Stanovení počtu přirozených subpopulací ? 10? Ln Likel lihood K (number of clusters)
55 K=5
56 K = 7 K = 10 Proporce genomu každého jedince náležející určitému clusteru
57 Un nited Kingdom W Europe Un nited Kingdom W Europe Un nited Kingdom W Europe Un nited Kingdom W Europe N France W Europe SW Spain Portugal N Italy SW Italy Switzerland W Europe W Europe W Europe W Europe W Europe W Iran SE Iran Slovenia Slovakia Kosovo N Bulgaria N Bulgaria E Bulgaria S Bulgaria E Greece Cyprus S Russia S Turkey N Syria W Syria S Syria E Syria W Syria N Syria C China C China N China NE China Japan E E E E E E E E E Europe Europe Europe Europe Europe Europe Europe Europe Europe West Asia West Asia West Asia West Asia West Asia West Asia West Asia West Asia East Asia East Asia East Asia East Asia East Asia West Asia West Asia Alternativní vizualizace výsledků ze STRUCTURE K forced clustering Zobrazení hierarchické struktury mezi populacemi K = 2 K = 3 K = 4 K = 5 K = 6 K = 7 K = 8 K = 9 K = 10 = 11 K = 12 K = 13 K = 14 K = 15 K = 16
58 NewHYBRIDS STRUCTURE NEWHYBRIDS
59 4. Spatially explicit analyses = spatial genetics = landscape genetics vychází z Bayesian clustering approach (typu STRUCTURE) individual-based models do modelování genetické informace přidává i geografické koordináty např. programy BAPS, TESS, Geneland např. programy BAPS, TESS, Geneland (automaticky stanovují nejlepší počet populací K)
60 Spatial models tesselation, Voronoi polygons
61 The example of very fragmented populations: the best model in BAPs for Central and Southern Dinaromys populations (spatial clustering of groups of individuals): K=13 (i.e. evidence of very high structuration) Best Partition: Cluster 1: {C9, C13} Cluster 2: {S6} Cluster 3: {C8, C14} Cluster 4: {C4} Cluster 5: {C1, C2} Cluster 6: {S1, S2, S3, S4} Cluster 7: {C6} Cluster 8: {C3, C15} Cluster 9: {C5, C7} Cluster 10: {C10} Cluster 11: {C11, C12} Cluster 12: {S5} Cluster 13: {C16} Př. program BAPS







62 Př.: Geneland

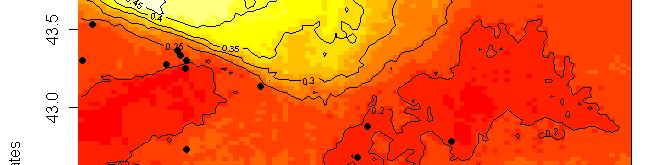
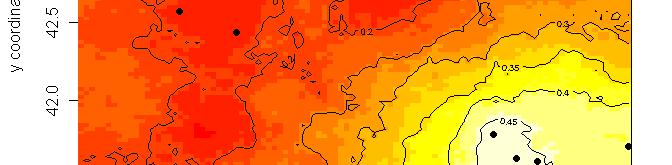
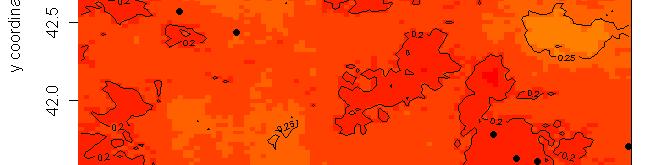
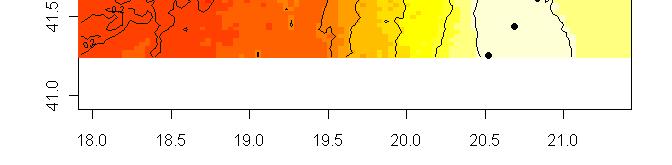
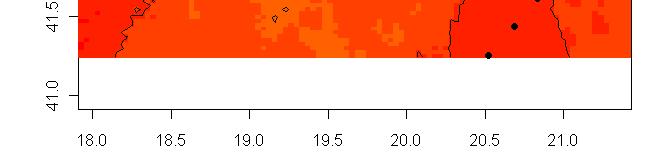

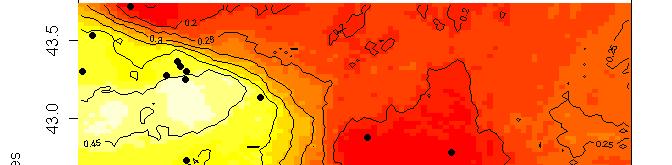
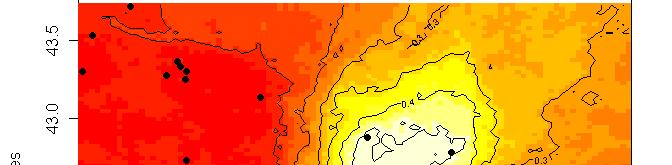
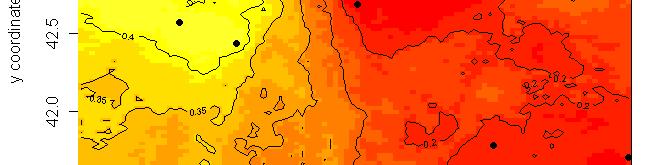
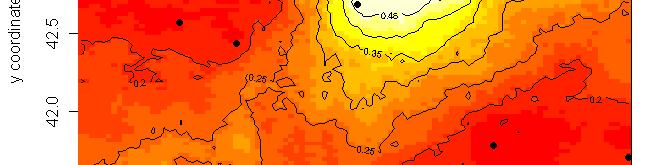
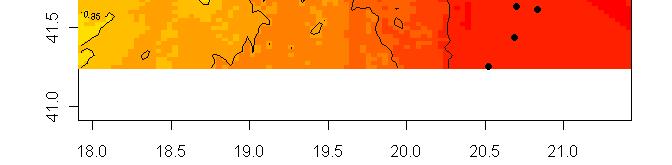
63 R platform Posterior probability maps
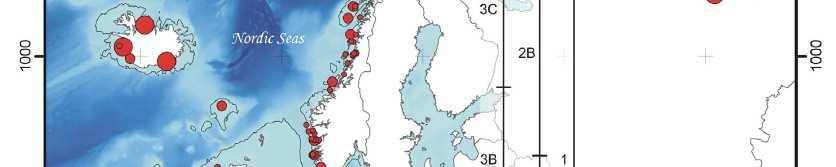
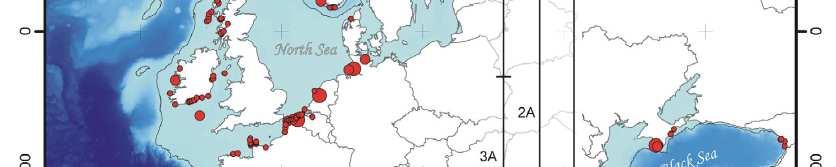
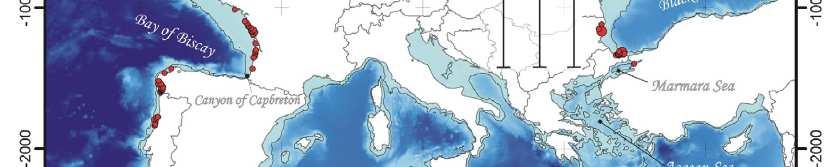


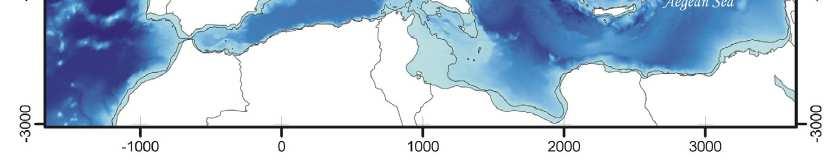

64 Spatial population genetics Fontaine et al Phocoena phocoena
Populačně-genetická data
 Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
Populačně-genetická data Základy analýzy diploidních kodominantních znaků (Mendelovská dědičnost) mikrosatelity T jaderné sekvence (např. SSCP) CCGATCAATGCGGCAA G CCGATCACTGCGGCAA SNPs Typ získaných dat
Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).
: RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae).") Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Populační studie Fisher M. & al. (2000): RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). American Journal of Botany 87(8): 1128
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.
: Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis.") Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Populační studie Kameyama Y. et al. (2001): Patterns and levels of gene flow in Rhododendron metternichii var. hondoense revealed by microsatellite analysis. Molecular Ecology 10:205 216 Proč to studovali?
Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).
: Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae).") Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Populační studie Tomimatsu H. &OharaM. (2003): Genetic diversity and local population structure of fragmented populations of Trillium camschatcense (Trilliaceae). Biological Conservation 109: 249 258.
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Genetika vzácných druhů zuzmun
 Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Genetika vzácných druhů Publikace Frankham et al. (2003) Introduction to conservation genetics Časopis Conservation genetics, založeno 2000 (máme online) Objekt studia Genetická diversita Rozložení genetické
Konzervační genetika INBREEDING. Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.
 Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Konzervační genetika INBREEDING Dana Šafářová Katedra buněčné biologie a genetiky Univerzita Palackého, Olomouc OPVK (CZ.1.07/2.2.00/28.0032) Hardy-Weinbergova rovnováha Hardy-Weinbergův zákon praví, že
Využití molekulárních markerů v systematice a populační biologii rostlin. 12. Shrnutí,
 Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Využití molekulárních markerů v systematice a populační biologii rostlin 12. Shrnutí, Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length Polymorphism) založené
Tok GI v buňce. Genetický polymorfizmus popis struktury populací. Organizace genetického materiálu. Definice polymorfismu
 Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Genetický olymorfizmus ois struktury oulací Tok GI v buňce Dr. Ing. Urban Tomáš ÚSTAV GEETIKY MZLU Brno urban@mendelu.cz htt://www.mendelu.cz/af/genetika/ Seminář doktorského grantu 53/03/H076 : Molekulárn
Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna
 Mendelova genetika v příkladech Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna Roman Longauer, Ústav zakládání a pěstění lesů, MENDELU
Mendelova genetika v příkladech Populační genetika a fylogeneze jedle bělokoré analyzována pomocí izoenzymových genových markerů a variability mtdna Roman Longauer, Ústav zakládání a pěstění lesů, MENDELU
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Populační genetika Obor zabývající se genetickou variabilitou v populacích a procesy, které ji
Genetická variabilita. rostlinných populací
 Genetická variabilita rostlinných populací Literatura Fox & Wolf eds. (2006) Evolutionary Genetics. Genetická variabilita Základ pro evoluci, možnosti selekce Kvalitativní vs. kvantitativní znaky Kvalitativní
Genetická variabilita rostlinných populací Literatura Fox & Wolf eds. (2006) Evolutionary Genetics. Genetická variabilita Základ pro evoluci, možnosti selekce Kvalitativní vs. kvantitativní znaky Kvalitativní
Genetika populací. KBI / GENE Mgr. Zbyněk Houdek
 Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
Genetika populací KBI / GENE Mgr. Zbyněk Houdek Genetika populací Populace je soubor genotypově různých, ale geneticky vzájemně příbuzných jedinců téhož druhu. Genový fond je společný fond gamet a zygot
Jak měříme genetickou vzdálenost a co nám říká F ST
 Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Jak měříme genetickou vzdálenost a co nám říká F ST 1) Genetická vzdálenost a její stanovení Pomocí genetické rozmanitosti, kterou se populace liší, můžeme určit do jaké míry jsou si příbuznější jaká je
Úvod do populační genetiky
 Úvod do populační genetiky 1 Termíny Gene Locus (loci) Allele alternative forms of a gene Genome single copy of all of the genes Diploid - 2 alely pro každý gen (výjimka Chr Y, cpdna a mtdna) 2 genomy
Úvod do populační genetiky 1 Termíny Gene Locus (loci) Allele alternative forms of a gene Genome single copy of all of the genes Diploid - 2 alely pro každý gen (výjimka Chr Y, cpdna a mtdna) 2 genomy
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele)
") Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Drift nejen v malých populacích (nebo při bottlenecku resp. efektu zakladatele) Nově vzniklé mutace: nová mutace většinou v 1 kopii u 1 jedince mutace modelovány Poissonovým procesem Jaká je pravděpodobnost,
Populační genetika Radka Reifová
 Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Populační genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013.
Využití molekulárních markerů v systematice a populační biologii rostlin. 2. Přehled aplikací a otázek
 Využití molekulárních markerů v systematice a populační biologii rostlin 2. Přehled aplikací a otázek Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length
Využití molekulárních markerů v systematice a populační biologii rostlin 2. Přehled aplikací a otázek Přehled molekulárních markerů 1. proteiny isozymy 2. DNA markery RFLP (Restriction Fragment Length
Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany
: Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany") Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
Populační studie Tribsch A., Schönswetter P. & Stuessy T. (2002): Saponaria pumila (Caryophyllaceae) and the Ice Age in the European Alps. American Journal of Botany 89(12): 2024 2033 Proč to studovali?
Vyhodnocování multilokusových dat II verze (T. Fér, F. Kolář)
") Vyhodnocování multilokusových dat II verze 2013-12-18 (T. Fér, F. Kolář) 1. PGDSpider převod mezi různými formáty dat 2. FSTAT - zjištění základních populačně genetických parametrů kodominantních dat 3.
Vyhodnocování multilokusových dat II verze 2013-12-18 (T. Fér, F. Kolář) 1. PGDSpider převod mezi různými formáty dat 2. FSTAT - zjištění základních populačně genetických parametrů kodominantních dat 3.
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Důsledky selekce v populaci - cvičení
 Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
TEORIE OSTROVNÍ BIOGEOGRAFIE (TOB)
") Organismy v krajině krajinné uspořádání a jeho vliv na organismy, populace a společenstva teorie ostrovní biogeografie metapopulační teorie zdroje a propady 34 TEORIE OSTROVNÍ BIOGEOGRAFIE (TOB) MacArthur
Organismy v krajině krajinné uspořádání a jeho vliv na organismy, populace a společenstva teorie ostrovní biogeografie metapopulační teorie zdroje a propady 34 TEORIE OSTROVNÍ BIOGEOGRAFIE (TOB) MacArthur
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Hardy-Weinbergův zákon - cvičení
 Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
AFLP. protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech
 AFLP AFLP protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech AFLP protokoly kit Invitrogen bez použití kitu AmpliTaq Gold (ABI) možnost
AFLP AFLP protokoly standardizace AFLP hodnocení primárních dat dnes používané metody hodnocení fylogenetický signál v AFLP datech AFLP protokoly kit Invitrogen bez použití kitu AmpliTaq Gold (ABI) možnost
Selekce v populaci a její důsledky
 Genetika a šlechtění lesních dřevin Selekce v populaci a její důsledky Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Selekce v populaci a její důsledky Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Základy genetiky populací
 Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Coalesce spojit se, splynout, sloučit se. Didaktická simulace Coalescence = splynutí linií
 Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Koalescence 1 2 Coalesce spojit se, splynout, sloučit se Didaktická simulace http://www.coalescent.dk/ Coalescence = splynutí linií 3 Koalescence Matematický model, který popisuje průběh genealogií. Postupujeme
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost divergence druhů
Teorie neutrální evoluce a molekulární hodiny
 Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Teorie neutrální evoluce a molekulární hodiny Teorie neutrální evoluce Konec 60. a začátek 70. let 20. stol. Ukazuje jak bude vypadat genetická variabilita v populaci a jaká bude rychlost evoluce v případě,
Genetická diverzita masného skotu v ČR
 Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Genetická diverzita masného skotu v ČR Mgr. Jan Říha Výzkumný ústav pro chov skotu, s.r.o. Ing. Irena Vrtková 26. listopadu 2009 Genetická diverzita skotu pojem diverzity Genom skotu 30 chromozomu, genetická
Příklady z populační genetiky lesních dřevin
 Obecná genetika Příklady z populační genetiky lesních dřevin Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem a Státním
Obecná genetika Příklady z populační genetiky lesních dřevin Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním fondem a Státním
Genetika populací. kvalitativních znaků
 Genetika populací kvalitativních znaků Úroveň studia genetických procesů Molekulární - struktura a funkce nukleových kyselin Buněčná buněčné struktury s významem pro genetiku, genetické procesy na buněčné
Genetika populací kvalitativních znaků Úroveň studia genetických procesů Molekulární - struktura a funkce nukleových kyselin Buněčná buněčné struktury s významem pro genetiku, genetické procesy na buněčné
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce jak lze studovat evoluci a jak funguje mutace, přírodní výběr, genový posun a genový tok 3)
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce jak lze studovat evoluci a jak funguje mutace, přírodní výběr, genový posun a genový tok 3)
Příbuznost a inbreeding
 Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Příbuznost a inbreeding Příbuznost Přímá (z předka na potomka). Souběžná (mezi libovolnými jedinci). Inbreeding Inbrední koeficient je pravděpodobnost, že dva geny přítomné v lokuse daného jedince jsou
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Evoluční (populační) genetika Radka Reifová
 genetika Radka Reifová") Evoluční (populační) genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Evoluční genetika Obor zabývající se genetickou variabilitou v populacích a procesy,
Evoluční (populační) genetika Radka Reifová Prezentace ke stažení: http://web.natur.cuni.cz/~radkas v záložce Courses Evoluční genetika Obor zabývající se genetickou variabilitou v populacích a procesy,
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK
 MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/15.0204 Molecular Ecology J. Bryja, M. Macholán MU, P. Munclinger - UK Co je molekulární ekologie? Uměle vytvořený obor vymezený technickým
S = c.a z. log(s) = log(c) + z.log(a) Rovnovážná teorie ostrovní biogeografie. The species-area relationship. The species-area relationship
 = log(c) + z.log(a) Rovnovážná teorie ostrovní biogeografie. The species-area relationship. The species-area relationship") Rovnovážná teorie ostrovní biogeografie equilibrium theory of island biogeography Robert MacArthur & Edward Osborne Wilson (1963, 1967) proč druhů přibývá s plochou ostrova? proč druhů ubývá s izolovaností
Rovnovážná teorie ostrovní biogeografie equilibrium theory of island biogeography Robert MacArthur & Edward Osborne Wilson (1963, 1967) proč druhů přibývá s plochou ostrova? proč druhů ubývá s izolovaností
Obecné principy KE v praxi
 Obecné principy KE v praxi BIOLOGICKÉ PRINCIPY OCHRANY HABITATU NA ÚROVNI KRAJINY Princip 1 Princip 2 Princip 3 Princip 4 Princip 5 Princip 6 Princip 7 Udržování velkých, nedotčených ploch (plošek) původní
Obecné principy KE v praxi BIOLOGICKÉ PRINCIPY OCHRANY HABITATU NA ÚROVNI KRAJINY Princip 1 Princip 2 Princip 3 Princip 4 Princip 5 Princip 6 Princip 7 Udržování velkých, nedotčených ploch (plošek) původní
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
PROSTOROVÁ EKOLOGIE, metapopulace, kvantitativní změny populace
 Partnerství pro rozvoj vzdělávání a komunikace v ochraně přírody reg. číslo: CZ.1.07/2.4.00/17.0073 PROSTOROVÁ EKOLOGIE, metapopulace, kvantitativní změny populace Filip Harabiš Katedra ekologie, FŽP,
Partnerství pro rozvoj vzdělávání a komunikace v ochraně přírody reg. číslo: CZ.1.07/2.4.00/17.0073 PROSTOROVÁ EKOLOGIE, metapopulace, kvantitativní změny populace Filip Harabiš Katedra ekologie, FŽP,
Migrace. 1) Jednosměrná migrace. 2) Obousměrná migrace. 3) Genový tok a historie populací. 4) Migrace a genetická odlišnost mezi populacemi
 Jednosměrná migrace. 2) Obousměrná migrace. 3) Genový tok a historie populací. 4) Migrace a genetická odlišnost mezi populacemi") P m p 0 1) Jednosměrná migrace 2) Obousměrná migrace 3) Genový tok a historie populací 4) Migrace a genetická odlišnost mezi populacemi 5) Odhady velikosti migrace 6) Přerušení izolace a Wahlundův princip
P m p 0 1) Jednosměrná migrace 2) Obousměrná migrace 3) Genový tok a historie populací 4) Migrace a genetická odlišnost mezi populacemi 5) Odhady velikosti migrace 6) Přerušení izolace a Wahlundův princip
Hodnocení multilokusových molekulárních dat II. Tomáš Fér & Filip Kolář Katedra Botaniky PřF UK, Praha
 Hodnocení multilokusových molekulárních dat II Tomáš Fér & Filip Kolář Katedra Botaniky PřF UK, Praha Multilokusová data dominantní nejsme schopni odlišit heterozygoty od dominantních homozygotů binární
Hodnocení multilokusových molekulárních dat II Tomáš Fér & Filip Kolář Katedra Botaniky PřF UK, Praha Multilokusová data dominantní nejsme schopni odlišit heterozygoty od dominantních homozygotů binární
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY
 GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
Koncept heritability
 Genetika kvantitativních znaků Koncept heritability doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Heritabilita míra Použití korelačních koeficientů může podporovat hypotézu, že daná kontinuální vlastnost
Genetika kvantitativních znaků Koncept heritability doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Heritabilita míra Použití korelačních koeficientů může podporovat hypotézu, že daná kontinuální vlastnost
UTILIZATION OF DNA MICROSATELLITES USED IN PARENTITY PANEL IN EVALUATION OF DIVERZITY AND DISTANCES BETWEEN THE BREEDS OF PIGS IN CZECH REPUBLIC
 UTILIZATION OF DNA MICROSATELLITES USED IN PARENTITY PANEL IN EVALUATION OF DIVERZITY AND DISTANCES BETWEEN THE BREEDS OF PIGS IN CZECH REPUBLIC VYUŽITÍ DNA MIKROSATELITŮ POUŽÍVANÝCH V PANELU NA URČOVÁNÍ
UTILIZATION OF DNA MICROSATELLITES USED IN PARENTITY PANEL IN EVALUATION OF DIVERZITY AND DISTANCES BETWEEN THE BREEDS OF PIGS IN CZECH REPUBLIC VYUŽITÍ DNA MIKROSATELITŮ POUŽÍVANÝCH V PANELU NA URČOVÁNÍ
Metody studia historie populací. Metody studia historie populací
 1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
1) Metody studia genetické rozmanitosti komplexní fenotypové znaky, molekulární znaky. 2) Mechanizmy evoluce mutace, přírodní výběr, genový posun a genový tok 3) Anageneze x kladogeneze - co je vlastně
VARIABILITY IN H-FABP, C-MYC, GH, LEP, LEPR GENES IN LARGE WHITE, LANDRACE AND DUROC BREEDS OF PIGS
 VARIABILITY IN H-FABP, C-MYC, GH, LEP, LEPR GENES IN LARGE WHITE, LANDRACE AND DUROC BREEDS OF PIGS VARIABILITA GENŮ H-FABP, C-MYC, GH, LEP, LEPR U PLEMEN PRASAT BÍLÉ UŠLECHTILÉ, LANDRASE A DUROK Mikolášová
VARIABILITY IN H-FABP, C-MYC, GH, LEP, LEPR GENES IN LARGE WHITE, LANDRACE AND DUROC BREEDS OF PIGS VARIABILITA GENŮ H-FABP, C-MYC, GH, LEP, LEPR U PLEMEN PRASAT BÍLÉ UŠLECHTILÉ, LANDRASE A DUROK Mikolášová
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/
 Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Základy populační genetiky Osnova 1. Genetická struktura populace 2. Způsob reprodukce v
Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Základy populační genetiky Osnova 1. Genetická struktura populace 2. Způsob reprodukce v
= oplození mezi biologicky příbuznými jedinci
 = oplození mezi biologicky příbuznými jedinci Jestliže každý z nás má 2 rodiče, pak má 4 prarodiče, 8 praprarodičů... obecně 2 n předků tj. po 10 generacích 2 10 = 1024, po 30 generacích = 1 073 741 824
= oplození mezi biologicky příbuznými jedinci Jestliže každý z nás má 2 rodiče, pak má 4 prarodiče, 8 praprarodičů... obecně 2 n předků tj. po 10 generacích 2 10 = 1024, po 30 generacích = 1 073 741 824
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Základní pojmy I. EVOLUCE
 Základní pojmy I. EVOLUCE Medvěd jeskynní Ursus spelaeus - 5 mil. let? - 10 tis. let - 200 tis. let? Medvěd hnědý Ursus arctos Medvěd lední Ursus maritimus Základní otázky EVOLUCE Jakto, že jsou tu různé
Základní pojmy I. EVOLUCE Medvěd jeskynní Ursus spelaeus - 5 mil. let? - 10 tis. let - 200 tis. let? Medvěd hnědý Ursus arctos Medvěd lední Ursus maritimus Základní otázky EVOLUCE Jakto, že jsou tu různé
Bioinformatika a výpočetní biologie. KFC/BIN VII. Fylogenetická analýza
 ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
ioinformatika a výpočetní biologie KF/IN VII. Fylogenetická analýza RNr. Karel erka, Ph.. Univerzita Palackého v Olomouci Fylogeneze Vznik a vývoj jednotlivých linií organismů Vývoj člověka phylogenetic
World of Plants Sources for Botanical Courses
 Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Speciace a extinkce Speciace Pojetí speciace dominuje proces, při němž vznikají nové druhy organismů z jednoho předka = kladogeneze, štěpná speciace jsou možné i další procesy hybridizace (rekuticulate
Populační genetika II. Radka Reifová
 Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Populační genetika II Radka Reifová Literatura An Introduction to Population Genetics. Rasmus Nielsen and Montgomery Slatkin. 2013. (v knihovně) Elements of Evolutionary Genetics (2010) Brian Charlesworth
Genetické mapování. v přírodních populacích i v laboratoři
 Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Genetické mapování v přírodních populacích i v laboratoři Funkční genetika Cílem je propojit konkrétní mutace/geny s fenotypem Vzniklý v laboratoři pomocí mutageneze či vyskytující se v přírodě. Forward
Mikrosatelity (STR, SSR, VNTR)
") Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Mikrosatelity (STR, SSR, VNTR) Repeats Více než polovina našeho genomu Interspersed (transposony) Tandem (mini- a mikrosatelity) Minisatellites (longer motifs 10 100 nucleotides) mikrosatelity Tandemová
Vytvořen. ení genetické databanky vybraných druhů savců ČR ití pro udržitelný rozvoj dopravy. Tomáš. Libosvár
 Vytvořen ení genetické databanky vybraných druhů savců ČR k využit ití pro udržitelný rozvoj dopravy Tomáš Libosvár TA02031259 Vytvořen ení genetické databanky vybraných druhů savců ČR k využit ití pro
Vytvořen ení genetické databanky vybraných druhů savců ČR k využit ití pro udržitelný rozvoj dopravy Tomáš Libosvár TA02031259 Vytvořen ení genetické databanky vybraných druhů savců ČR k využit ití pro
Populační genetika III. Radka Reifová
 Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Populační genetika III Radka Reifová Genealogie, speciace a fylogeneze Genové genealogie Rodokmeny jednotlivých kopií určitého genu v populaci. Popisují vztahy mezi kopiemi určitého genu v populaci napříč
Genetika kvantitativních znaků. - principy, vlastnosti a aplikace statistiky
 Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
GENETIKA Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální dědičnost
 Polygenní dědičnost Multifaktoriální dědičnost") GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích
 Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Mikroevoluce = vznik a osud genetické variability na druhové a nižší úrovni děje a mechanismy v populacích Evoluce = genetická změna populací v čase a prostoru Evoluce = změna frekvence alel v populacích
Genetika populací. Doposud genetika na úrovni buňky, organizmu
 Doposud genetika na úrovni buňky, organizmu - jedinec nás nezajímá - pouze jeho gamety a to jako jedny z mnoha = genofond = soubor všech gamet skupiny jedinců Populace mnoho různých definic - skupina organizmů
Doposud genetika na úrovni buňky, organizmu - jedinec nás nezajímá - pouze jeho gamety a to jako jedny z mnoha = genofond = soubor všech gamet skupiny jedinců Populace mnoho různých definic - skupina organizmů
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Projekční algoritmus. Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění. Jan Klíma
 Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Urychlení evolučních algoritmů pomocí regresních stromů a jejich zobecnění Jan Klíma Obsah Motivace & cíle práce Evoluční algoritmy Náhradní modelování Stromové regresní metody Implementace a výsledky
Inovace studia molekulární a buněčné biologie
 Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Investice do rozvoje vzdělávání Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. Investice do rozvoje vzdělávání
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské. doc. RNDr. Ivan Mazura, CSc.
 Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Genetický polymorfismus jako nástroj identifikace osob v kriminalistické a soudnělékařské praxi doc. RNDr. Ivan Mazura, CSc. Historie forenzní genetiky 1985-1986 Alec Jeffreys a satelitní DNA 1980 Ray
Heritabilita. Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability
 Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
Heritabilita Heritabilita = dědivost Podíl aditivního rozptylu na celkovém fenotypovém rozptylu Výpočet heritability h 2 = V A / V P Výpočet genetické determinance znaku h 2 = V G / V P Heritabilita závisí
KBI / GENE Mgr. Zbyněk Houdek
 Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
Dědičnost komplexních a kvantitativních znaků KBI / GENE Mgr. Zbyněk Houdek Komplexní znaky Komplexní fenotypy mohou být ovlivněny genetickými faktory a faktory prostředí. Mezi komplexní znaky patří např.
Vícerozměrné statistické metody
 Vícerozměrné statistické metody Podobnosti a vzdálenosti ve vícerozměrném prostoru, asociační matice II Jiří Jarkovský, Simona Littnerová Vícerozměrné statistické metody Práce s asociační maticí Vzdálenosti
Vícerozměrné statistické metody Podobnosti a vzdálenosti ve vícerozměrném prostoru, asociační matice II Jiří Jarkovský, Simona Littnerová Vícerozměrné statistické metody Práce s asociační maticí Vzdálenosti
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Mgr. et Mgr. Lenka Falková. Laboratoř agrogenomiky. Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita
 Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Mgr. et Mgr. Lenka Falková Laboratoř agrogenomiky Ústav morfologie, fyziologie a genetiky zvířat Mendelova univerzita 9. 9. 2015 Šlechtění Užitek hospodářská zvířata X zájmová zvířata Zemědělství X chovatelství
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,
: Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread,") Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Malcomber S.T. (2000): Phylogeny of Gaertnera Lam. (Rubiaceae) based on multiple DNA markers: evidence of a rapid radiation in a widespread, morphologically diverse genus. Evolution 56(1):42-57 Proč to
Evropské mapování znečištění ovzduší za rok 2005
 Evropské mapování znečištění ovzduší za rok 2005 Jan Horálek Tvorbě evropských map a rozvoji mapování je několikátým rokem věnovv nován úkol Spatial air quality data v rámci ETC/ACC - úkol 5.3.1.2. Implementačního
Evropské mapování znečištění ovzduší za rok 2005 Jan Horálek Tvorbě evropských map a rozvoji mapování je několikátým rokem věnovv nován úkol Spatial air quality data v rámci ETC/ACC - úkol 5.3.1.2. Implementačního
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/..00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG) Tento
Crossing-over. Synaptonemální komplex. Crossing-over a výměna genetického materiálu. Párování homologních chromosomů
 Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Vazba genů Crossing-over V průběhu profáze I meiózy Princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem Synaptonemální komplex Zlomy a nová spojení chromatinových řetězců
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat
 Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Genetické markery ve studiu genetické diverzity v populacích hospodářských zvířat Bakalářská
Mendelova zemědělská a lesnická univerzita v Brně Agronomická fakulta Ústav morfologie, fyziologie a genetiky zvířat Genetické markery ve studiu genetické diverzity v populacích hospodářských zvířat Bakalářská
Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz
 Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická
Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická
Real Estate Investment 2019
 Real Estate Investment 2019 Máme se bát obchodní války? Dopady Reality v nové kontextu vlny protekcionismu globální a lokální ekonomiky David DubenMarek 2019 Reality v investičním a ekonomickém kontextu
Real Estate Investment 2019 Máme se bát obchodní války? Dopady Reality v nové kontextu vlny protekcionismu globální a lokální ekonomiky David DubenMarek 2019 Reality v investičním a ekonomickém kontextu
Vícerozměrné statistické metody
 Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Vícerozměrné statistické metody Shluková analýza Jiří Jarkovský, Simona Littnerová FSTA: Pokročilé statistické metody Typy shlukových analýz Shluková analýza: cíle a postupy Shluková analýza se snaží o
Genetické rozdíly mezi populacemi aneb něco o migracích a genovém toku. Genetické rozdíly mezi populacemi
 Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Genetické rozdíly mezi populacemi Genetické rozdíly mezi populacemi 1) Genetická vzdálenost populací a její příčiny 3) Proč jsou subsaharské africké populace geneticky vzdálenější od populací ostatních?
Problémy malých populací
 Problémy malých populací Rozložení velikostí populací Velikost populace Pocet pripadu 1 2 3 4 5 6 7 3 52 12 151 21 25 3 349 399 448 497 547 596 646 695 745 794 843 893 942 992 141 191 114 119 1239 1288
Problémy malých populací Rozložení velikostí populací Velikost populace Pocet pripadu 1 2 3 4 5 6 7 3 52 12 151 21 25 3 349 399 448 497 547 596 646 695 745 794 843 893 942 992 141 191 114 119 1239 1288
Evoluční algoritmy. Podmínka zastavení počet iterací kvalita nejlepšího jedince v populaci změna kvality nejlepšího jedince mezi iteracemi
 Evoluční algoritmy Použítí evoluční principů, založených na metodách optimalizace funkcí a umělé inteligenci, pro hledání řešení nějaké úlohy. Populace množina jedinců, potenciálních řešení Fitness function
Evoluční algoritmy Použítí evoluční principů, založených na metodách optimalizace funkcí a umělé inteligenci, pro hledání řešení nějaké úlohy. Populace množina jedinců, potenciálních řešení Fitness function
Genetická variabilita v populacích
 - je nástrojem studia populační (evoluční) genetiky 1) Populace, genofond 2) Fenotypová a genotypová variabilita populací 3) Vznik a zdroje genetické variability 4) Odhad genetické struktury populací (výpočet
- je nástrojem studia populační (evoluční) genetiky 1) Populace, genofond 2) Fenotypová a genotypová variabilita populací 3) Vznik a zdroje genetické variability 4) Odhad genetické struktury populací (výpočet
CLP ANALYSIS OF MOLECULAR MARKERS DIGITAL IMAGE ANALYSIS OF ELECTROPHOEROGRAMS CZECH VERSION
 CLP ANALYSIS OF MOLECULAR MARKERS DIGITAL IMAGE ANALYSIS OF ELECTROPHOEROGRAMS CZECH VERSION DIGITÁLNÍ OBRAZOVÁ ANALÝZA ELEKTROFORETICKÝCH GELŮ *** Vyhodnocování získaných elektroforeogramů: Pro vyhodnocování
CLP ANALYSIS OF MOLECULAR MARKERS DIGITAL IMAGE ANALYSIS OF ELECTROPHOEROGRAMS CZECH VERSION DIGITÁLNÍ OBRAZOVÁ ANALÝZA ELEKTROFORETICKÝCH GELŮ *** Vyhodnocování získaných elektroforeogramů: Pro vyhodnocování
3) Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR
 Analýza mtdna mitochondriální Eva, kdy a kde žila. 8) Haploskupiny mtdna a chromozomu Y v ČR") Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Hledání našeho společného předkap 1) Jak to, že máme společného předka 2) Metodika výzkumu mtdna 3) Analýza mtdna mitochondriální Eva, kdy a kde žila 4) Problémy a názory proti 5) Analýza chromozomu Y
Zpracování studie týkající se průzkumu vlastností statistických proměnných a vztahů mezi nimi.
 SEMINÁRNÍ PRÁCE Zadání: Data: Statistické metody: Zpracování studie týkající se průzkumu vlastností statistických proměnných a vztahů mezi nimi. Minimálně 6 proměnných o 30 pozorováních (z toho 2 proměnné
SEMINÁRNÍ PRÁCE Zadání: Data: Statistické metody: Zpracování studie týkající se průzkumu vlastností statistických proměnných a vztahů mezi nimi. Minimálně 6 proměnných o 30 pozorováních (z toho 2 proměnné
Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc.
 Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická statistika 010 1.týden (0.09.-4.09. ) Data, typy dat, variabilita, frekvenční analýza
Pravděpodobnost a matematická statistika Doc. RNDr. Gejza Dohnal, CSc. dohnal@nipax.cz Pravděpodobnost a matematická statistika 010 1.týden (0.09.-4.09. ) Data, typy dat, variabilita, frekvenční analýza
Metody studia historie populací. Metody studia historie populací. 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.
 Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky.") 1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva
1) Metody studiagenetickérozmanitosti komplexní fenotypové znaky, molekulární znaky. 2)Mechanizmy evoluce mutace, p írodnívýb r, genový posun a genový tok 3) Anagenezex kladogeneze-co je vlastn druh 4)Dva