Nishikigoi. Luděk Štěch ml. koi.cz
|
|
|
- Bohuslav Kolář
- před 5 lety
- Počet zobrazení:
Transkript
1 Nishikigoi Luděk Štěch ml. koi.cz
2 ÚVOD Člověk dokázal vyšlechtit mnoho typů ů zbarvení u několika druhů ryb jako jsou barevné formy akvarijních ryb např.u živorodek duhových (gupek) (Poecilia reticulata), mečovek mexických (Xiphophorus heleri), ) plat skvrnitých (Xiphophorus maculatus), živorodek velkoploutvých (Poecilia velifera), terčovcůčervených (Symphysodon y p y discus), skalár amazonských (Pterophyllum scalare), vrubozubců pavých (Astronotus ocellatus), parmiček čtyřpruhých (Puntius tetrazona mechové zbarvení) a dalších jako u karase stříbřitého (Carassius auratus), lína obecného (Tinca tinca), albinotické formy u sumce velkého (Silurus glanis) a u dalších známých i méně známých mnoha druhů ryb.
3 Barevné mutace byly yynalezeny u několika čeledí ryb, u některých znich byly dále šlechtěny a jsou stále chovány. Barevné mutace se vyskytují např. u čeledí: jeseterovití (Acipenseridae), lososovití (Salmonidae), kaprovití (Cyprinidae), sumcovití (Siluridae), keříčkovcovití (Clariidae), vrubozobcovití (Cichlidae). U většiny těchto čeledí existují pouze albinotická zbarvení, nebo částečná ztráta pigmentu vkůži, ale u kaprovitých bylo zatím vyšlechtěno nejvíce barevných mutací ze všech čeledí, počínaje barevnými (zlatými, modrými, alampickými) líny, zlatými perlíny, albinotickými (zlatými) amury, zlatými a modrými jeseny a konče zlatými karasy, karasy Shubunkin, Sarasa a závojnatkami. Zlaté mutace se u kaprovitých vyskytují sporadicky vpřírodě i u dalších druhů (plotice, slunka).
4 Nejvyšší množství íbarevných mutací u chovaných druhů kaprovitých ryb je zřejmě zdůvodu toho, že jejich chov je jednoduchý, jsou nenároční na životní prostředí a prostor a chovají se již velmi dlouho ať už pro potřeby potravní, potravinářské, nebo okrasné. Jedním z nejkrásnějších a zároveň nejdéle šlechtěných druhů ryb je kapr obecný (Cyprinus carpio). )
5 Historie a původ chovu Koi Kořeny chovu kapra Koi sahají do doby před 2500 lety, kdy se v Číně zabývali nejen chovem obyčejného kapra, ale také kapra barevného. Nám známý název Koi, který je názvem pro kapra v Japonsku, pochází zřejmě již z této doby. Barevnýkapr, japonsky,,nishikigoi
6 V roce 470 př.n.l. byl sepsán rukopis,,yang Yu Jing, g, který napsal Tao Zhu Gong, g,jež mimo jiné referuje o chovu kapra Koi. Z Číny, která se začne zaobírat více chovem závojnatek se kapr Koi přesouvá do Japonska, které se stává jeho novým domovem. Další už japonské písemné zmínky jsou z let 713 n.l.,, Hitachi Fudoki a v roce 720 n.l.,,nihon Shoki (Japonská kronika).
7 Koije v Japonsku ryba válečníků, č symbolem odvahy a statečnosti, ale také lásky, síly, mužnosti a štěstí. K velkému rozmachu chovu Koi dochází až v době Bunka a Bunsei ( ). V prefektuře Niigata dochází ke vzniku nových barevných mutací a Niigata je centrem chovu Koi. V éře Taisho ( ) vznikají typy Kohaku, Ki Utsuri a Sanshoku (Taisho Sanke) V době Showa pak variety Showa a Ginrin
8 Roční produkce Koi v Japonsku se odhaduje na 100 miliónů kusů Cena ryb je různá, mladší kategorie ryb jsou levnější, u starších kusů mohou dosahovat až astronomických výšin Dovoz probíhá nejsnadněji přes zprostředkovatelskou firmu, která vykupuje od chovatelů ryby a exportuje je Dovozy pak probíhají najednou jd i s dlší dalšími obchodníky
9 Dovoz probíhá v PE pytlích a kartonových krabicích Trvá až hodin od nabalení ryb Dováží se všechny velikosti
10 Historie chovu v Evropě a v ČR V 70. letech se dostává ákapr Koi do Evropy a to do Anglie Začátkem let 80. se dostává i do okolních zemí Do východního bloku se nejdříve dostávají v roce 1983 do Maďarska V roce 1985 se nishikigoi rozšiřují i do ČR. Před tím se u nás vyskytoval zlatý kapr vznikl mutací z našich chovů, měl žluté ploutve a žlutě lemované šupiny
11 V 90. letech hbli byli přivezeni iprvní kvalitnější kapři Koi od firmy Ogata do Rybníkářství Hluboká (1991) Poté se k barevným kaprům dostává více chovatelů, zprvu hlavně velké rybářské firmy a posléze i drobnochovatelé Větší rozmach chov Koi v ČR prožívá po roce 2000
12 Tvar těla kaprů Koi U divokého kapra je tvar těla nízký, protažený a na průřezu téměř kruhovitý Tvar těla u Koi se dělí na: Shinshu h mírně ě vyklenutý hřbet i břicho, průřez je téměř kruhovitý, jako u divokého kapra, klábonosá hlava v čelní části. Nevýhodný k dalšímu chovu Ishugrudel vysokohřbetost, podobný tvar těla jako u kapra konzumního, nevhodný k dalšímu chovu Třetím je tvar těla podobný divokému kapru, příliš nízký se špičatější hlavou, nevýhodný ýk chovu
13 Ideálem je tvar těla s mírně a ladně vyklenutým hřbetem za hlavou až do úrovně začátku hřbetní ploutve a posléze s pravidelně a elegantně se snižující linií k ocasní ploutvi nazýváme ho japonský tvar těla. Břicho může být mírně ě vyklenuté, průřez těla je však šk požadován co nejvíce kruhovitý. Poměr výšky těla k délce (bez ocasní ploutve) těla by měla být od 1:2,6 až do 1:3.
14 Zbarvení kůže Vzájemné vrstvení chromatoforů má za následek vznik výsledného, strukturálního zbarvení. Toto zbarvení vzniká jako výsledek kombinací a vrstvení výše uvedených čtyř typů pigmentových buněk ve škáře. Jednotlivé barvy vznikají takto: stříbřitá vmístech kde jsou melanofory uloženy pod iridocyty, šedá melanofory jsou na povrchu, iridocyty pod nimi, melanin je částečně roztažený
15 černá melanofory se široce roztaženým melaninem překrývají iridocyty modrá iridocyty jsou nad melanofory sroztaženým melaninem (trvale modré zbarvení vzniká pevným spojením stříbřitých a černých buněk vtzv. melaniridocyty), zelená vzniká kombinací třech pigmentových buněk, nahoře jsou xantofory, pod nimi iridocyty, dole melanofory, žlutá lesklá xantofory leží nad iridocyty,
16 žlutá nelesklá tvořena pouze xantofory s roztaženým barvivem, žlutohnědá melanofory s částečně roztaženým melaninem leží nad xantofory s roztaženým barvivem, červenohnědá melanofory s částečně roztaženým melaninem leží nad erytrofory s roztaženým barvivem, červená lesklá erytrofory jsou nad iridocyty, červená matná tvořena pouze erytrofory s roztaženým barvivem, oranžová vzniká promícháním xantoforů s erytrofory, bílá je způsobena pouze iridocyty (Dubský a kol., 2003
17 Základy dědičnosti barev ryb Barva je typickým kvalitativním znakem organismů. Takové znaky obecně kóduje jeden nebo několik málo definovaných genů velkého účinku. Tyto geny jsou nazývány monogeny (jeden pár alel llna jd jednom lokusu určuje č jd jeden znak) či oligogeny (2 a více párů alel, zpravidla ne více než 4, umístěných ě ýhna dvou a více lokusech určují č jeden znak).
18 Jejich fenotypový projev je obvykle alternativní (znak se buď projeví nebo neprojeví). U ryb se mezi kvalitativní znaky zahrnuje právě zbarvení těla nebo oka, barevné znaky na trupu, hlavě nebo ploutvích, ošupení, tvarploutvíapod apod. Vliv prostředí na expresi genů velkého účinku je obvykle nevýznamný nebo málo významný. (Flajšhans a kol., 2008)
19 Základy šlechtění barevných mutací ryb Barevné mutace lína Výskyt zlatého a modrého zbarvení u lína obecného byl blpoprvé zmíněn ě Kluppem (1985) a Geldhauserem (1988) vněmeckých chovech. Dědičnost zlatého a modrého zbarvení u lína, jejich genotypy a jejich fenotypový projev popsal Kvasnička a kol. (1996). Na základě analýz štěpných poměrů zbarvení potomstva vf 1, F 2, B 1 a B 2 generacích bylo zjištěno, že modré a zlaté zbarvení jsou mutace dvou různých autozomálních genů (b a g), které nejsou ve vazbě (Flajšhans a kol., 1997).
20 Obě zbarvení se mohou projevit pouze v homozygotní konstituci (bbg modré zbarvení: B gg zlaté zbarvení) a jsou recesivní vůči divokému zbarvení (B G ) G ). Modely křížení, rodičovské genotypy a fenotypy, a fenotypové štěpné poměry ě F 1 potomstva t jsou popsány vtabulce 10. Dosud ověřené štěpné poměry potomstva F 1, F 2, B 1 a B 2 generací (B = zpětné křížení) jsou popsány p v tabulce 11 (Flajšhans a kol., 1997).
21 Modré zbarvení lína Modré zbarvení je způsobeno mutací, negativně ovlivňující produkci červených ý a žlutých pigmentů. Křížením modrých jedinců mezisebou dostaneme 100% modrého potomstva (Flajšhans a kl kol., 1997).
22 Zlaté zbarvení lína Zlaté zbarvení íje způsobeno ů mutací, ovlivňující ň í negativně produkci melaninu a guaninu nebo koncentraci melanoforů a guanoforů. Sytost zbarvení od světle žluté po tmavě červenou je dána distribucí červených a žlutých pigmentů, ovlivnitelnou vnějšími vlivy, zatímco tmavé skvrnyna na hlavách, ploutvích a na hřbetní partii ryb jsou dány distribucí melaninu. Křížením zlatých jedinců mezi sebou dostaneme 100% zlatého potomstva. Není li u potomstva žádoucí výskyt černých znaků na zlatém podkladě, vybíráme rodičovské jedince bez těchto znaků (Flajšhans a kol., 1997).
23
24 Divoké (zelené) zbarvení lína Divoký (zelený) fenotyp odpovídá teoretickým genotypům BBGG (dominantní homozygot v obou vlohách); BBGg (homozygot ve vloze pro modré a heterozygot ve vloze pro zlaté zbarvení); BbGG (heterozygot ve vloze pro modré a homozygot ve vloze pro zlaté zbarvení), nebo BbGg (heterozygot v obou vlohách; Flajšhans a kol., 1997)
25 Alampie u lína Alampií se rozumí stav, kdy jedinec postrádá všechny typy kožní pigmentace pg na těle i ploutvích (Fox, 1957; Crozier, 1974). Oko jevšak pigmentováno. Tyto jedince získáme při křížení F 2 potomstva po křížení zlatých a modrých jedinců (tab11; Flajšhans a kol., 1997).
26 Tab. 10: Modely křížení, rodičovské genotypy a fenotypy, a fenotypové štěpné poměry F 1 potomstva (Flajšhans a kol., 1997). Křížení číslo Jikernačka RODIČE Mlíčák Fenotyp Genotyp Fenotyp Genotyp POTOMSTVO Fenotypový štěpný poměr 1 zlatá BBgg zlatý BBgg 100 % zlatých 2 modrá bbgg modrý bbgg 100 % modrých 3 zlatá ltá BBgg divoký BBGG 100 % divokých 4 zlatá BBgg divoký BBGg 50 % divokých : 50 % zlatých 5 zlatá BBgg divoký BbGG 100 % divokých 6 zlatá BBgg divoký BbGg 50 % divokých : 50 % zlatých 7 modrá bbgg divoký BBGG 100 % divokých 8 modrá bbgg divoký BBGg 100 % divokých 9 modrá bbgg divoký BbGG 50 % divokých : 50% modrých 10 modrá bbgg divoký BbGg 50 % divokých : 50% modrých
27 Barevné mutace u karase stříbřitého Njí Nejvíce základního studia genetiky u karase stříbřitého bylo uskutečněno japonskými a čínskými vědci. Chen (1928, 1934) ukázal, že modré a hnědé barvy charakteristické pro některé rázy, stejně jako průhlednost kožního krytí, jsou děděny malým počtem genů. ů Absolutní a mozaiková průhlednost je dána (podle Matsui 1934) kombinací dvou párů genů. Polodominantní gen T pro absolutní průhlednost je epistatický v homozygotním stavu, v postavení k recesivnímu n genu (z jiného páru) působí na tvorbu umístění barevných a nebarevných sekcí
28 . Oči č karase mohou být teleskopické kékvůli přítomnosti recesivního genu, zatímco dlouhé rozdvojené ploutve vznikají u karase působením interakce dvou nebo tří párů genů (Matsui, 1934a a c). Mnoho zvláštností u karase stříbřitého ještě nebylo objasněno z genetického hlediska, ačkoli některé variety karasů existují po mnoho staletí, zvláště červené formy, které byly vyšlechtěny v 10. století (Chen, 1956). Mnoho mutací ve všech rázech karase se nahromadily během posledních staletí. Z toho důvodu je jasné, že rozdíly mezi rázy musí být polygenního charakteru a je velmi obtížné je analyzovat (Kirpičnikov, 1971).
29
30
31 Dědičnost zbarvení xantorického rázu u pstruha duhového První pokus o křížení í dospělých jdi jedinců ů uvnitř žlutě (xantoricky) zbarveného chovu (speciálně dovezeného od Maliszewského), dopadl tak, že vprvní F 1 generaci mělo celé potomstvo zbarvení xantorického rázu (žlutého rázu). Od ryb zjastarniea Olesnice s xantorickým zbarvením po dosažení pohlavní dospělosti, se pokusy dělaly v sérii křížících experimentů uvnitř tří barevných rázů. Potomstvo získané ztěchto křížení bylo všechno sxantorickýmzbarvením. Dále byly provedeny pokusy vkřížení mezi xantorickým zbarvením a divokým zbarvením, uvnitř rázů i mezi nimi.v tabulce 7 jsou uvedeny výsledky pokusů (Backiel a kol.,2007).
32 Tab.7: Výsledky pokusů křížení xantorického (žlutého) a divokého zbarvení. Křížení divokého zbarvení a zbarvení Xantorického, zbarvení potomstva linie/zbarvení samice linie/zbarvení samec zbarvení potomstva Žluté/Xantorické Žluté/Xantorické Xantorické Jastarnia/Xantorické Jastarnia/Xantorické Xantorické Olesnica/Xantorické Olesnica/Xantorické Xantorické Jastarnia/Xantorické Žluté//Xantorické Xantorické Olesnica/Xantorické Žluté//Xantorické Xantorické Žluté/Xantorické Jastarnia/Divoké Divoké/Xantorické Žluté/Xantorické Olesnica/Divoké Divoké Jastarnia/Divoké Žluté//Xantorické Divoké Jastarnia/Divoké Olesnica/Xantorické Divoké Jastarnia/Divoké Olesnica/Divoké Divoké/Xantorické Jastarnia/Divoké Jastarnia/Divoké Divoké/Xantorické Olesnica/Divoké Jastarnia/Divoké Divoké
33 Podle výsledků zbarvení potomstva se předpokládá, že xantorické zbarvení je dědičná mutace genu, který ovládá divoké zbarvení u Pstruha duhového. Projev genu odpovědný za divoké zbarvení je označen symbolem A, zatímco gen odpovědný za xantorické zbarvení, které jepodlo podloženo výsledky v tabulce, je vůči divokému zbarvení (A) recesivní, jeoznačen symbolem a (Backiel a kol., 2007).
34 Barevné mutace u dalších chovaných druhů ryb Jelec jesen (Leuciscusi idus), ) šlechtěný v některých evropských zemích, je albinotickou formou běžného jelce jesena. Evidentně je albinotické zbarvení jelce jesena výsledkem recesivní mutace vybírané a dál šlechtěné člověkem. Stejné albinotické formy byly nalezeny i u sumečka skvrnitého (Ictalurus punctatus), který je nyní hlavním druhem chovaným v teplovodní akvakultuře v USA. Nd Nedostatek ttkčerného barviva je u tohoto t druhu způsoben ů přítomností recesivního genu (Nelson, 1958). Albinotické mutace byly nalezeny také i u dalších rybničních druhů ryb (Kirpičnikov, 1971).
35
36
37 Šlechtění barevných mutací kapra 261J Jednobarevné étypy kapra Koi. Základní barevné mutace původního divokého ( tj. šedožlutozeleného) zbarvení kapra, zahrnující bílý, modrý a zlatý fenotyp, byly již několikrát popsány (Probst, 1949; Moav a Wohlfarth, 1968; Katasonov 1973, 1974, 1976, 1978; Merla, 1982; Kirpičnikov, 1981; Linhart, 1987). Podle těchto autorů je modré zbarvení podmíněno genem BL a řízeno alelou bl v recesivní formě (Linhart, 1987). Zlaté zbarvení íje podmíněno ě genem G a řízeno alelou l g v recesivní formě (Kirpičnikov, 1981). Ztěchto údajů by bylo možno usuzovat na stejný mechanismus dědičnosti těchto barev, jak bylo dokázáno u lína obecného.
38 Přímo u kapra Koi byly popsány dva znaky : světlézbarvení, podmíněné genem L a řízené alelou L v dominantní formě (Katasonov, 1973, 1974, 1976); a světle žluté obrazce na hlavě podmíněné genem D a řízené alelou D v dominantní íformě ě (Katasonov, 1973, 1974).
39 Křížením í kteréhokoli k typu kapra Koi s divoce zbarvenými jedinci kterékoli chované populace nebo linie bylo vždy získáno 100% divoce zbarveného potomstva. Byla tedy potvrzena dominance divokého zbarvení. Jedinou výjimku tvoří pozorování Rothbarda a Wohlfartha (1995), při kterém při křížení typu Asagi s kaprem divokého fenotypu vznikla forma Shusui. Není ale jasné zda autoři neměli výrazem Shusui na mysli pouze fenotyp ošupení í( (u typu Asagije doitsu lysá áforma nazývána právě Shusui). Křížením typu Orange ogon x Orange ogon bylo získáno 100% šupinatých červenozlatých homozygotů, křížením typu Orange ogon x Doitsu Orange Ogon i u potomstva reciprokého křížení bylo získáno100% červenozlatých šupinatých heterozygotů.
40 Křížením í Shiro Ogon (bílý) x Platinum ogon (bílý metalický) tliký) bylo získáno po 50% zobou rodičovských fenotypů. Štěpný poměr 1:1 by odpovídal křížení heterozygota s recesivním homozygotem za předpokladu, že by projev čistě bílé a bílé metalické barvy byl řízen jedním genem. Křížením Karasugoi x Platinum ogon bylo získáno48% černě zbarveného potomstva a 52% potomstva typu Platinum ogon. V obou případech se však škpouze 08% 0,8% 12% 1,2% potomstva blížila standardní kvalitě zbytek byl nestandardní kvality. V tomto případě se může jednat o odchylku od štěpného poměru 1:1, svědčící o křížení černého heterozygota s bílým recesivním homozygotem (Flajšhans a kol., 1997).
41 Dvoubarevné, tříbarevné a vícebarevné typy kapra koi Výsledky křížení í dvou a tříbarevvných typů ů Koi s jednobarevnými typy, mezi sebou a s tříbarevnými typy potvrzují, že nelzehovořit o dědičnosti složitých barevných typů Koi v pravém slova smyslu. Není tedy možné předpovědět p zbarvení potomstva na základě barev rodičů. Sled provedených křížení byl sestaven do tab. 13 a pro úplnost doplněn o literární údaje Szweigmana a kol. (1992) a Rothbarda a Wohlfartha (1995). Tyto údajejsouoznačeny jsou označeny hvězdičkou.
42 Na základě vlastních pokusů autorů a na základě literárních údajů lze vyvodit následující závěry: 1. Genotyp divokého zbarvení je dominantní 2. Genotyp modrého zbarvení je recesivní a je řízen alelou bl v homozygotní konstituci 3. Genotyp zlatého zbarvení je recesivní a je řízen alelou g v recesivní konstituci 4. V sérii barev černá zlatá bílá existuje vztah: Černá ( dominantní; převaha melanoforů) Šedá ( pomalejší vývoj melaninové pigmentace) pg Žlutá řada (převaha xantoforů) Bílá ( recesivní; absence melanoforů a xantoforů)
43 5. St Sytost tbarev červená oranžová á ž á zlatá ltážlutá může být ovlivněna příjmem karotenoidů vpotravě. Vzhledem ktomu, že ve žlutě zbarvené části potomstva jednotlivých křížení zpravidla nebyl pozorován plynulý přechod sytosti barvy, ale docházelo kvyštěpení podobných počtů červených, ý, oranžových a žlutých jedinců, není možné předem vyloučit ani geneticky podmíněný vliv. 6. Zvýše uvedeného vyplývá, že při křížení typů Koi, obsahující ve svém zbarvení černé znaky (Taisho sanke, Showa, Bekko, Utsuri) s jedno nebo dvoubarevnými typy bez černých znaků, dojde v potomstvu kvyštěpení černých nebo šedých šdýhznaků. Stejně ě tak tkpři křížení í dvoubarevných typů s červenými, nebo oranžovými znaky (Kohaku, Hariwake) s jednobarevnými bílými Koi (Platinum ogon), dojde kvyštěpení červených, oranžových nebo žlutých znaků u potomstva.
44 Tab. 13: Výsledky křížení dvou- a tříbarevných typů Koi s jednobarevnými typy, mezi sebou a s tříbarevnými typy (Flajšhans a kol., 1997) Samci Samice Kohaku Tancho Taisho sanke Showa sanshoku Bekko shiro/ki Asagi Shusui Utsuri Shiro/ki Platinum ogon Hajiro Karasug oi divoké zbarvení Kohaku bílý, červený, Kohaku, vzácně Taisho sanke, Tancho* Kohaku, ogon, Taisho sanke,s Bekko Kohaku, ogon Taisho sanke, Kohaku Aigorom, Koromo*, Kohaku Showa Ogon, Hariwake, Shusui divoké Tancho Taisho sanke Kohaku, ogon, Taisho sanke,s Bekko Goshiki, Koromo divoké Showa sanshoku Kohaku, Ogon Ogon*, Hariwake ne showa Koromo Kohaku* Kin/Gin showa* divoké Bekko shiro/ki Taisho sanke*, Kohaku Khk Kohaku, Taisho sanke, Bekko* divoké Asagi Aigoromo, Koromo, Kohaku* Goshiki* Koromo shusui Shusui Kohaku, Ogon, Hariwake Utsuri Shiro/ki Showa* Kohaku Kohaku, Sh. Bekko divoké Platinum ogon Ogon, Hariwake, Shusu Showa* Kohaku, Ogon, Hariwake Ogon Kabuto, Ogon, Hariwake Karasugo i, Ogon Hajiro Karasugoi Karasugoi, ogon divoké zbarvení divoké divoké* divoké divoké Shusui* divoké Kabuto divoké
45 Kohaku Základní barevné typy
46 Taisho Sanke
47 Showa Sanshoku








48
49 Utsurimono








50
51







52 Bekko
53 Koromo
54 Asagi
55 Shusui






56 Goshiki
57 Kumonryu








58 Kawarimono
59








60
61
























62





















63
























64 Hikari Mujimono (Ogon)
65
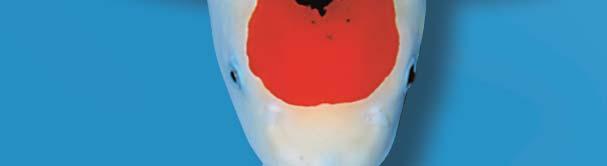
66
67






















68 Hikari Moyomono
























69
70






















71 Hikari Utsurimono
72 Kujaku























73 Tancho
74























75 Kinginrin
76
77 Závojnatí koi
78
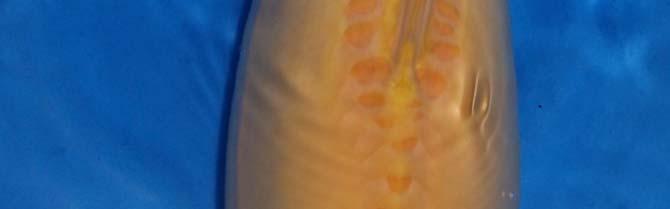
79 Pigmentaci sledujeme už u váčkového plůdku Pigmentace je sledována, ld jk jako melaninově ě pigmentovaný a melaninově nepigmentovaný (černě ě pigmentovaný a zlatý váčkový plůdek). U melaninově pigmentovaného pg váčkového plůdku je předpoklad budoucího zbarvení typu: Shiro utsuri, Hi utsuri a Showa (spíše (p tmavší typy). Viz. obr.:
80 Obr.: 1
81 Obr.: 2.
82 Obr.: 3.
83 Segregace zbarvení v F1 u typu při křížení v parentální generaci dvou ryb typu Showa P: X F 1 :
84 Při křížení dvou ryb v parentální generaci, stejného zbarvení typu Showa,, se vyštěpily py tyto fenotypové projevy u potomstva vf1 generaci: Shiro Muji, Kohaku, Shiro Utsuri, Hi Utsuri, Showa, Aka Muji, Taisho Sanke a nezařaditelné ř ryby do typu (odpadní d zbarvení), které nejsou uvedeny na obrázku.
85 Segregace barevných typů u plůdku z rybničních odchovů poukazuje na fakt, že při křížení ryb v rodičovské generaci se stejným zbarvením typu Showa se u potomstva objevuje několik jiných typů, než jsou rodičovské fenotypy. Showa se vyskytuje v potomstvu vmnožství mezi 17 19% (může se měnit ě v závislosti áil ina kvalitě ě rodičovské generace a dalších podmínkách). Ostatní vyštěpená zbarvení jako jsou Kohaku, Shiro Utsuri, Hi Utsuri nebo Taisho Sanke, jsou také typy plnohodnotné a štěpí se v potomstvu v kvalitním vzhledu.
86 Ryby v parentální generaci by měly být co nejkvalitněji j zbarvené. Přínosem kvality rodičů by mohlo být i zvýšení kvality zbarvení u potomstva. V potomstvu se však i přes to, že rodičovská linie je japonského původu od renomovaných chovatelů, vyskytovaly ryby v typech, které jsou nepoužitelné pro další odchov nebo prodej. Tyto ryby nepatří do standardu.
87 í jdůl ži ější dá bý řídě í Zatím nejdůležitějším se zdá být třídění vprůběhu odchovu ryb.
88 Vystavování a hodnocení ryb Nj Nejprestižnější výstavy jsou v Japonsku Výstavy organizují spolky a sdružení chovatelů Koi, nejznámější je v Japonsku Zen Nippon Airinkai i i (Z.N.A.) Tato organizace pořádá v Japonsku ročně kolem 100 výstav Nejznámější je Z.N.A. All Japan Nishikigoi Show, která je uznávaná jako světová výstava hobby chovatelů. Může zde vystavovat každý člen Z.N.A. Odroku Koi
89 Další je All Japan Combined Shinkokai Tokyo Koi Show Výstava profesionálních chovatelů a obchodníků ks Koi V Tki Tokiu Z této výstavy vychází Grand Champion je považována za nejlepší exponát na světě Dále áevýstava a mladých Koi, od roku ou1982 V Ojiya
90 Hodnocení ídříve: forma 30, barva 20, skvrny 20, celková kvalita ryby 10, ladnost a elegance 10, image (dojem) 10. Celkový počet bodů byl 100 Dnešní hodnocení: forma těla 50 %, barva 30%, rozložení skvrn 20%.
91 Negativní vliv Kapři Koi (plůdek, násada i generační č ryby) b)jsou mnohem citlivější knepříznivým faktorům prostředí než běžné obsádky kapra, a proto i vysazení ryb vdobrém výživném i zdravotním stavu a vdobré kondici do kvalitního komorového rybníka a kvalitní péče o něj není vždy zárukou, že se na jaře sloví podobné množství Koi a v podobném zdravotním stavu, jako je chovatel zvyklý při komorování kapra. I tak dosahují ztráty při komorování vprůměru 60 90% podle toho, o který barevný typ se jedná. K nejodolnějším typům Koi patří jednobarevní metaličtí Ogoni. Je vhodné kapra Koi v kategorii K 1 zimovat v objektech vnádržích soteplenou vodou (Flajšhans a kol., 1997).
92
93
94
Nishikigoi. Luděk Štěch ml. www.alcedor.cz www.kapr-koi.cz
 Nishikigoi Luděk Štěch ml. www.alcedor.cz www.kapr-koi.cz ÚVOD Člověk dokázal vyšlechtit mnoho typů zbarvení u několika druhů ryb jako jsou barevné formy akvarijních ryb např.u živorodek duhových (gupek)
Nishikigoi Luděk Štěch ml. www.alcedor.cz www.kapr-koi.cz ÚVOD Člověk dokázal vyšlechtit mnoho typů zbarvení u několika druhů ryb jako jsou barevné formy akvarijních ryb např.u živorodek duhových (gupek)
Mendelistická genetika
 Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Mendelistická genetika Základní pracovní metodou je křížení křížení = vzájemné oplozování organizmů s různými genotypy Základní pojmy Gen úsek DNA se specifickou funkcí. Strukturní gen úsek DNA nesoucí
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Mendelovská genetika - Základy přenosové genetiky Základy genetiky Gregor (Johann)
Základní genetické pojmy
 Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Základní genetické pojmy Genetika Věda o dědičnosti a proměnlivosti organismů Používá především pokusné metody (např. křížení). K vyhodnocování používá statistické metody. Variabilita v rámci druhu Francouzský
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
VYBRANÉ GENETICKÉ ÚLOHY II.
 VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které
VYRNÉ GENETICKÉ ÚLOHY II. (Nemendelistická dědičnost, kodominance, genové interakce, vazba genů) ÚLOHY 1. Krevní skupiny systému 0 -,,, 0 - jsou určeny řadou alel (mnohotná alelie, alelická série), které
Cvičení č. 8. KBI/GENE Mgr. Zbyněk Houdek
 Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
Cvičení č. 8 KBI/GENE Mgr. Zbyněk Houdek Genové interakce Vzájemný vztah mezi geny nebo formami existence genů alelami. Jeden znak je ovládán alelami působícími na více lokusech. Nebo je to uplatnění 2
12. Mendelistická genetika
 12. Mendelistická genetika Genetika se zabývá studiem dědičnosti a proměnlivosti organismů proměnlivost (variabilita) odraz vlivu prostředí na organismus potomků klasická dědičnost schopnost rodičů předat
12. Mendelistická genetika Genetika se zabývá studiem dědičnosti a proměnlivosti organismů proměnlivost (variabilita) odraz vlivu prostředí na organismus potomků klasická dědičnost schopnost rodičů předat
MENDELOVSKÁ DĚDIČNOST
 MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
MENDELOVSKÁ DĚDIČNOST Gen Část molekuly DNA nesoucí genetickou informaci pro syntézu specifického proteinu (strukturní gen) nebo pro syntézu RNA Různě dlouhá sekvence nukleotidů Jednotka funkce Genotyp
Deoxyribonukleová kyselina (DNA)
") Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Genetika Dědičností rozumíme schopnost rodičů předávat své vlastnosti potomkům a zachovat tak rozličnost druhů v přírodě. Dědičností a proměnlivostí jedinců se zabývá vědní obor genetika. Základní jednotkou
Nauka o dědičnosti a proměnlivosti
 Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Nauka o dědičnosti a proměnlivosti Genetika Dědičnost na úrovni nukleových kyselin molekulární buněk organismů populací Předávání vloh z buňky na buňku Předávání vlastností mezi jednotlivci Dědičnost znaků
Základní barvy holuba domácího
 Základní barvy holuba domácího Ing. Juraj Kafka Národní komise pro standardy holubů ČSCH 5 Popis původního zbarvení Columba livia, (L.) modré černopruhé 4 3 1 2 1. Krk část opeření s rozprostřenými pigmentovými
Základní barvy holuba domácího Ing. Juraj Kafka Národní komise pro standardy holubů ČSCH 5 Popis původního zbarvení Columba livia, (L.) modré černopruhé 4 3 1 2 1. Krk část opeření s rozprostřenými pigmentovými
Cvičeníč. 10 Dědičnost a pohlaví. Mgr. Zbyněk Houdek
 Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Cvičeníč. 10 Dědičnost a pohlaví Mgr. Zbyněk Houdek Dědičnost a pohlaví Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů, ale i další geny. V těchto
Genetika ryb popis druhů
 Genetika ryb popis druhů Vít Baránek Vytvořeno s podporou projektu FRVŠ 2030/2007 pro přípravu semináře Genetika ryb jsme si vybrali tři perspektivní druhy: lín obecný (Tinca tinca) candát obecný (Sander
Genetika ryb popis druhů Vít Baránek Vytvořeno s podporou projektu FRVŠ 2030/2007 pro přípravu semináře Genetika ryb jsme si vybrali tři perspektivní druhy: lín obecný (Tinca tinca) candát obecný (Sander
Genetika zvířat - MENDELU
 Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Genetika zvířat Gregor Mendel a jeho experimenty Gregor Johann Mendel (1822-1884) se narodil v Heinzendorfu, nynějších Hynčicích. Během období, v kterém Mendel vyvíjel svou teorii dědičnosti, byl knězem
Mendelistická genetika
 Mendelistická genetika Distribuce genetické informace Základní studijní a pracovní metodou v genetice je křížení (hybridizace), kterým rozumíme vzájemné oplozování jedinců s různými genotypy. Do konce
Mendelistická genetika Distribuce genetické informace Základní studijní a pracovní metodou v genetice je křížení (hybridizace), kterým rozumíme vzájemné oplozování jedinců s různými genotypy. Do konce
Geny p řevážně nepůsobí izolovan ě izolovan ale, v kontextu s okolním prostředím (vnitřním i vnějším) ě a v souladu souladu s ostatními g eny geny.
 ě a v souladu souladu s ostatními g eny geny.") Genové interakce Geny převážně nepůsobí izolovaně, ale v kontextu s okolním prostředím (vnitřním i vnějším) a v souladu s ostatními geny. Genové interakce -intraalelické -interalelické A a intraalelické
Genové interakce Geny převážně nepůsobí izolovaně, ale v kontextu s okolním prostředím (vnitřním i vnějším) a v souladu s ostatními geny. Genové interakce -intraalelické -interalelické A a intraalelické
Základy genetiky 2a. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Základy genetiky 2a Přípravný kurz Komb.forma studia oboru Všeobecná sestra Základní genetické pojmy: GEN - úsek DNA molekuly, který svojí primární strukturou určuje primární strukturu jiné makromolekuly
Akvarijní výstavy Akvarijní výstavy. Hodnocení ryb - soutěže. Akvaristika
 Cvičení #3 Akvarijní výstavy Akvarijní výstavy Akvaristika Jiří Patoka patoka@af.czu.cz 2017 Expoziční Soutěžní Prodejní (trhy) Součást veletrhů Pouze živočichové a rostliny Nebo též doplňky technika,
Cvičení #3 Akvarijní výstavy Akvarijní výstavy Akvaristika Jiří Patoka patoka@af.czu.cz 2017 Expoziční Soutěžní Prodejní (trhy) Součást veletrhů Pouze živočichové a rostliny Nebo též doplňky technika,
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
S v a z c h o v a t e l ů k o n í K i n s k ý c h
 ZBARVENÍ A DĚDIČNOST BARVY U KINSKÉHO KONĚ Prof. Ing. Václav Jakubec, DrSc., Česká zemědělská univerzita, Praha, Česká republika Dr. Monika Reissmann, Humboldt-Universität zu Berlin, Německo Ing. Josef
ZBARVENÍ A DĚDIČNOST BARVY U KINSKÉHO KONĚ Prof. Ing. Václav Jakubec, DrSc., Česká zemědělská univerzita, Praha, Česká republika Dr. Monika Reissmann, Humboldt-Universität zu Berlin, Německo Ing. Josef
Genetika pro začínající chovatele
 21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
21.4.2012 Praha - Smíchov Genetika pro začínající chovatele včetně několika odboček k obecným základům chovu Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních
Genetická determinace zbarvení vlasů u člověka. Genetická determinace zbarvení očí u člověka
 Genetická determinace zbarvení vlasů u člověka Genetická determinace zbarvení očí u člověka znaky polygenní, které však při studiu dědičnosti v rodinách vykazují zdánlivě jednoduchou dědičnost výzkumem
Genetická determinace zbarvení vlasů u člověka Genetická determinace zbarvení očí u člověka znaky polygenní, které však při studiu dědičnosti v rodinách vykazují zdánlivě jednoduchou dědičnost výzkumem
Genetika mnohobuněčných organismů
 Genetika mnohobuněčných organismů Metody studia dědičnosti mnohobuněčných organismů 1. Hybridizační metoda představuje systém křížení, který umožňuje v řadě generací vznikajících pohlavní cestou zjišťovat
Genetika mnohobuněčných organismů Metody studia dědičnosti mnohobuněčných organismů 1. Hybridizační metoda představuje systém křížení, který umožňuje v řadě generací vznikajících pohlavní cestou zjišťovat
Dědičnost zbarvení srsti u psů se zaměřením na plemeno Cane Corso
 Dědičnost zbarvení srsti u psů se zaměřením na plemeno Cane Corso V průběhu domestikace vlka, která začala před 40 000 lety, bylo postupně vyšlechtěno přibližně tisíc dnes známých plemen psů, přičemž většina
Dědičnost zbarvení srsti u psů se zaměřením na plemeno Cane Corso V průběhu domestikace vlka, která začala před 40 000 lety, bylo postupně vyšlechtěno přibližně tisíc dnes známých plemen psů, přičemž většina
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Základy genetiky - Alelové a Genové interakce Intra-alelické interakce = Interakce
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Základy genetiky - Alelové a Genové interakce Intra-alelické interakce = Interakce
Základy genetiky populací
 Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základy genetiky populací Jedním z významných odvětví genetiky je genetika populací, která se zabývá studiem dědičnosti a proměnlivosti u velkých skupin jedinců v celých populacích. Populace je v genetickém
Základní pravidla dědičnosti
 Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
Mendelova genetika v příkladech Základní pravidla dědičnosti Tento projekt je spolufinancován Evropským sociálním fondem a Státním rozpočtem ČR InoBio CZ.1.07/2.2.00/28.0018 Mendelovy zákony dědičnosti
9.12.2012 Brno - Lužánky Základy chovatelství a genetiky potkanů
 9.12.2012 Brno - Lužánky Základy chovatelství a genetiky potkanů Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních a nejedná se o má díla. Prezentace nejsou
9.12.2012 Brno - Lužánky Základy chovatelství a genetiky potkanů Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních a nejedná se o má díla. Prezentace nejsou
Obecná genetika a zákonitosti dědičnosti. KBI / GENE Mgr. Zbyněk Houdek
 Obecná genetika a zákonitosti dědičnosti KBI / GENE Mgr. Zbyněk Houdek Důležité pojmy obecné genetiky Homozygotní genotyp kdy je fenotypová vlastnost genotypově podmíněna uplatněním páru funkčně zcela
Obecná genetika a zákonitosti dědičnosti KBI / GENE Mgr. Zbyněk Houdek Důležité pojmy obecné genetiky Homozygotní genotyp kdy je fenotypová vlastnost genotypově podmíněna uplatněním páru funkčně zcela
Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
 Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Výukový materiál zpracován v rámci projektu EU peníze školám Název školy: Střední zdravotnická škola a Obchodní akademie, Rumburk, příspěvková organizace Registrační číslo projektu: CZ.1.07/1.5.00/34.0649
Barevné formy zebřiček a jejich genetika - část II. příklady
 Barevné formy zebřiček a jejich genetika - část II. příklady Tyto příklady se váží k předchozímu článku o obecných zákonitostech genetiky. K napsaní těchto detailů mne inspiroval jeden dotaz, který určuje
Barevné formy zebřiček a jejich genetika - část II. příklady Tyto příklady se váží k předchozímu článku o obecných zákonitostech genetiky. K napsaní těchto detailů mne inspiroval jeden dotaz, který určuje
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny
 Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
Obecná genetika Základní pojmy obecné genetiky, kvalitativní a kvantitativní znaky, vztahy mezi geny Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ing. Roman LONGAUER, CSc. Ústav zakládání a pěstění lesů LDF MENDELU
O B R A T L O V C I - SLADKOVODNÍ RYBY
 VODNÍ O B R A T L O V C I - SLADKOVODNÍ RYBY 1. Kapr obecný Délka: 110 cm, váha: 21 kg, místo nálezu: Španělsko délka života: 40 let pohlavní dospělost: 3-5. rok doba rozmnožování: květen-červen běžná
VODNÍ O B R A T L O V C I - SLADKOVODNÍ RYBY 1. Kapr obecný Délka: 110 cm, váha: 21 kg, místo nálezu: Španělsko délka života: 40 let pohlavní dospělost: 3-5. rok doba rozmnožování: květen-červen běžná
Štika obecná. Pstruh obecný. latinsky: Esox lucius slovensky: Šťuka obyčajná anglicky: Pike, v USA Northern Pike německy: Hecht hovorově: zubatá
 Štika obecná latinsky: Esox lucius slovensky: Šťuka obyčajná anglicky: Pike, v USA Northern Pike německy: Hecht hovorově: zubatá řád: Bezostní čeleď: Štikovití potrava: dravec délka života: 15 let pohlavní
Štika obecná latinsky: Esox lucius slovensky: Šťuka obyčajná anglicky: Pike, v USA Northern Pike německy: Hecht hovorově: zubatá řád: Bezostní čeleď: Štikovití potrava: dravec délka života: 15 let pohlavní
Reprodukce a genetika
 Reprodukce a genetika Jeden z hlavních kritérií při výběru druhů pro chov, je existence vhodné technologie pro řízenou reprodukci, nebo snadná dostupnost larev nebo juvenilních stádií v přírodě. I když
Reprodukce a genetika Jeden z hlavních kritérií při výběru druhů pro chov, je existence vhodné technologie pro řízenou reprodukci, nebo snadná dostupnost larev nebo juvenilních stádií v přírodě. I když
V F 2. generaci vznikají rozdílné fenotypy. Stejné zabarvení značí stejný fenotyp.
 Cvičení č. 6: Mendelovy zákony KI/GENE Mgr. Zyněk Houdek Mendelovy zákony Při pohlavním rozmnožování se může z každého rodiče přenést na jeho potomka vždy pouze jediná alela z páru. Vyslovil v roce 1865
Cvičení č. 6: Mendelovy zákony KI/GENE Mgr. Zyněk Houdek Mendelovy zákony Při pohlavním rozmnožování se může z každého rodiče přenést na jeho potomka vždy pouze jediná alela z páru. Vyslovil v roce 1865
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací. KBI/GENE: Mgr. Zbyněk Houdek
 Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
Cvičeníč. 9: Dědičnost kvantitativních znaků; Genetika populací KBI/GENE: Mgr. Zbyněk Houdek Kvantitativní znak Tyto znaky vykazují plynulou proměnlivost (variabilitu) svého fenotypového projevu. Jsou
KŮŽE, ŠUPINY A ZBARVENÍ RYB Rybí tělo je kryto kůží, která je tvořena pokožkou (epidermis) a škárou (corium), složenou ze tří vrstev, (stratum
 a škárou (corium), složenou ze tří vrstev, (stratum") KŮŽE, ŠUPINY A ZBARVENÍ RYB Rybí tělo je kryto kůží, která je tvořena pokožkou (epidermis) a škárou (corium), složenou ze tří vrstev, (stratum vasculare, stratum compactum a subcutis). Kůže je zpravidla
KŮŽE, ŠUPINY A ZBARVENÍ RYB Rybí tělo je kryto kůží, která je tvořena pokožkou (epidermis) a škárou (corium), složenou ze tří vrstev, (stratum vasculare, stratum compactum a subcutis). Kůže je zpravidla
pásmo cejnové dolní tok řek (velmi pomalý tok řeky) pásmo parmové střední tok řek pásmo lipanové podhorské potoky a řeky
 pásmo parmové střední tok řek pásmo lipanové podhorské potoky a řeky") Ryby sladkovodní Sladkovodní ryby žijí jak ve stojatých, tak i tekoucích vodách, které podle rychlosti proudu, teploty a obsahu kyslíku rozdělujeme do čtyř pásem, které jsou pojmenovány podle typického
Ryby sladkovodní Sladkovodní ryby žijí jak ve stojatých, tak i tekoucích vodách, které podle rychlosti proudu, teploty a obsahu kyslíku rozdělujeme do čtyř pásem, které jsou pojmenovány podle typického
Karas stříbřitý. (Carassius auratus) Libor Mikl, HYDR I.
 Libor Mikl, HYDR I.") Karas stříbřitý (Carassius auratus) Libor Mikl, HYDR I. Řád: Máloostní Čeleď: Kaprovití Charakteristika potrava: všežravec délka života: 10 let pohlavní dospělost: 3. rok doba rozmnožování: květen-červenec
Karas stříbřitý (Carassius auratus) Libor Mikl, HYDR I. Řád: Máloostní Čeleď: Kaprovití Charakteristika potrava: všežravec délka života: 10 let pohlavní dospělost: 3. rok doba rozmnožování: květen-červenec
Hardy-Weinbergův zákon - cvičení
 Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Hardy-Weinbergův zákon - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Degenerace genetického kódu
 AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
AJ: degeneracy x degeneration CJ: degenerace x degenerace Degenerace genetického kódu Genetický kód je degenerovaný, resp. redundantní, což znamená, že dva či více kodonů může kódovat jednu a tutéž aminokyselinu.
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162
 Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 ZŠ Určeno pro Sekce Předmět Téma / kapitola Prameny 8. třída (pro 3. 9. třídy)
Rozvoj vzdělávání žáků karvinských základních škol v oblasti cizích jazyků Registrační číslo projektu: CZ.1.07/1.1.07/02.0162 ZŠ Určeno pro Sekce Předmět Téma / kapitola Prameny 8. třída (pro 3. 9. třídy)
Genotypy absolutní frekvence relativní frekvence
 Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Genetika populací vychází z: Genetická data populace mohou být vyjádřena jako rekvence (četnosti) alel a genotypů. Každý gen má nejméně dvě alely (diploidní organizmy). Součet všech rekvencí alel v populaci
Metody plemenitby. plemenitba = záměrné a cílevědomé připařování + rozmnožování zvířat zlepšování tvarových + především užitkových vlastností
 Metody plemenitby plemenitba = záměrné a cílevědomé připařování + rozmnožování zvířat zlepšování tvarových + především užitkových vlastností Metody plemenitby využívající 1. podobnosti rodičů + jejich
Metody plemenitby plemenitba = záměrné a cílevědomé připařování + rozmnožování zvířat zlepšování tvarových + především užitkových vlastností Metody plemenitby využívající 1. podobnosti rodičů + jejich
Působení genů. Gen. Znak
 Genové interakce Působení genů Gen Znak Dědičnost Potomek získává predispozice k vlastnostem z rodičovské buňky nebo organismu. Vlastnosti přenášené do další generace nemusí být zcela totožné s vlastnostmi
Genové interakce Působení genů Gen Znak Dědičnost Potomek získává predispozice k vlastnostem z rodičovské buňky nebo organismu. Vlastnosti přenášené do další generace nemusí být zcela totožné s vlastnostmi
Nové systematické členění
 Cvičení 4 Nové systematické členění Řád: Cypriniformes (Máloostní) Čeleď: Cyprinidae (Kaprovití) Rod: Rutilus (Plotice) Druh: Rutilus rutilus (Plotice obecná) Druh: Rutilus pigus (Plotice lesklá) Rod:
Cvičení 4 Nové systematické členění Řád: Cypriniformes (Máloostní) Čeleď: Cyprinidae (Kaprovití) Rod: Rutilus (Plotice) Druh: Rutilus rutilus (Plotice obecná) Druh: Rutilus pigus (Plotice lesklá) Rod:
Rili Shrimp a Orange Sakura - dvoubarevná a oranžová novinka druhu Neocaridina heteropoda
 Rili Shrimp a Orange Sakura - dvoubarevná a oranžová novinka druhu Neocaridina heteropoda Neocaridina heteropoda je nejčastěji chovaným druhem sladkovodní krevety u nás. Předností tohoto druhu je především
Rili Shrimp a Orange Sakura - dvoubarevná a oranžová novinka druhu Neocaridina heteropoda Neocaridina heteropoda je nejčastěji chovaným druhem sladkovodní krevety u nás. Předností tohoto druhu je především
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje
 Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje Mgr. Siřínková Petra březen 2009 Mendelovy zákony JOHANN GREGOR MENDEL Narodil se 20. července 1822 v
Tento výukový materiál vznikl za přispění Evropské unie, státního rozpočtu ČR a Středočeského kraje Mgr. Siřínková Petra březen 2009 Mendelovy zákony JOHANN GREGOR MENDEL Narodil se 20. července 1822 v
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Genetika populací Studium dědičnosti a proměnlivosti skupin jedinců (populací)
Genetické určení pohlaví
 Přehled GMH Seminář z biologie Genetika 2 kvalitativní znaky Genetické určení pohlaví Téma se týká pohlavně se rozmnožujících organismů s odděleným pohlavím (gonochoristů), tedy dvoudomých rostlin, většiny
Přehled GMH Seminář z biologie Genetika 2 kvalitativní znaky Genetické určení pohlaví Téma se týká pohlavně se rozmnožujících organismů s odděleným pohlavím (gonochoristů), tedy dvoudomých rostlin, většiny
Národní program uchování a využívání genetických zdrojů zvířat
 METODIKA CHOVU GENETICKÝCH ZDROJŮ KRÁLÍKŮ První zmínky o chovu králíků na území Čech pocházejí ze 13. století, chovatelství jako takové se však začíná rozvíjet na počátku 19. století. V polovině 19. století
METODIKA CHOVU GENETICKÝCH ZDROJŮ KRÁLÍKŮ První zmínky o chovu králíků na území Čech pocházejí ze 13. století, chovatelství jako takové se však začíná rozvíjet na počátku 19. století. V polovině 19. století
Genetika kvantitativních znaků. - principy, vlastnosti a aplikace statistiky
 Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Genetika kvantitativních znaků Genetika kvantitativních znaků - principy, vlastnosti a aplikace statistiky doc. Ing. Tomáš Urban, Ph.D. urban@mendelu.cz Genetika kvantitativních vlastností Mendelistická
Bochov Dědičnost bílých znaků
 4.11.2011 Bochov Dědičnost bílých znaků Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních a nejedná se o má díla. Prezentace nejsou určeny ke komerčnímu
4.11.2011 Bochov Dědičnost bílých znaků Obrázky použité v prezentaci byly postahovány z různých zdrojů na internetu z důvodů ilustračních a nejedná se o má díla. Prezentace nejsou určeny ke komerčnímu
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST
 INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST I. ročník, letní semestr 13. týden 14. - 18.5.2007 Aleš Panczak, ÚBLG 1. LF a VFN Krátké opakování: Jednotková dědičnost podíl alel téhož genu (lokusu) při
INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST I. ročník, letní semestr 13. týden 14. - 18.5.2007 Aleš Panczak, ÚBLG 1. LF a VFN Krátké opakování: Jednotková dědičnost podíl alel téhož genu (lokusu) při
TEST. na přezkoušení uchazeče. pro získání kvalifikace pro vydání prvního rybářského lístku. 1. O jaký druh ryby se jedná.
 TEST na přezkoušení uchazeče pro získání kvalifikace pro vydání prvního rybářského lístku 1. O jaký druh ryby se jedná a] úhoř říční b] lipan podhorní c] hrouzek obecný 2. O jaký druh ryby se jedná a]
TEST na přezkoušení uchazeče pro získání kvalifikace pro vydání prvního rybářského lístku 1. O jaký druh ryby se jedná a] úhoř říční b] lipan podhorní c] hrouzek obecný 2. O jaký druh ryby se jedná a]
Genetika na úrovni mnohobuněčného organizmu
 Genetika na úrovni mnohobuněčného organizmu Přenos genetické informace při rozmnožování Nepohlavní rozmnožování: - nový jedinec vzniká ze somatické buňky nebo ze souboru somatických buněk jednoho rodičovského
Genetika na úrovni mnohobuněčného organizmu Přenos genetické informace při rozmnožování Nepohlavní rozmnožování: - nový jedinec vzniká ze somatické buňky nebo ze souboru somatických buněk jednoho rodičovského
- Zákl. metodou studia organismů je křížení (hybridizace)- rozmn. dvou vybraných jedinců, umožnuje vytváření nových odrůd rostlin a živočichů
- rozmn. dvou vybraných jedinců, umožnuje vytváření nových odrůd rostlin a živočichů") Otázka: Základní zákonitosti dědičnosti Předmět: Biologie Přidal(a): Kateřina P. - Zákl. zákonitosti dědičnosti zformuloval Johann Gregor Mendel - Na základě svých pokusů křížením hrachu- popsal a vysvětlil
Otázka: Základní zákonitosti dědičnosti Předmět: Biologie Přidal(a): Kateřina P. - Zákl. zákonitosti dědičnosti zformuloval Johann Gregor Mendel - Na základě svých pokusů křížením hrachu- popsal a vysvětlil
Schopnost organismů UCHOVÁVAT a PŘEDÁVAT soubor informací o fyziologických a morfologických (částečně i psychických) vlastnostech daného jedince
 vlastnostech daného jedince") Genetika Genetika - věda studující dědičnost a variabilitu organismů - jako samostatná věda vznikla na počátku 20. století - základy položil J.G. Mendel již v druhé polovině 19. století DĚDIČNOST Schopnost
Genetika Genetika - věda studující dědičnost a variabilitu organismů - jako samostatná věda vznikla na počátku 20. století - základy položil J.G. Mendel již v druhé polovině 19. století DĚDIČNOST Schopnost
Genetika kvantitativních znaků
 Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
Genetika kvantitativních znaků Kvantitavní znaky Plynulá variabilita Metrické znaky Hmotnost, výška Dojivost Srstnatost Počet vajíček Velikost vrhu Biochemické parametry (aktivita enzymů) Imunologie Prahové
BARVY BORDER COLLIÍ. Na konci tohoto dokumentu naleznete schéma hlavních barev podle lokusů.
 BARVY BORDER COLLIÍ Barva psí srsti je dána geneticky. Pro všechny border collie (snad až na vzácné výjimky) platí, že ve své genetické výbavě nesou alelu Si, která determinuje irské zbarvení (bílé znaky)
BARVY BORDER COLLIÍ Barva psí srsti je dána geneticky. Pro všechny border collie (snad až na vzácné výjimky) platí, že ve své genetické výbavě nesou alelu Si, která determinuje irské zbarvení (bílé znaky)
INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST
 INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST I. ročník, letní semestr 13. týden 12. - 16.5.2008 Aleš Panczak, ÚBLG 1. LF a VFN Krátké opakování: Jednotková dědičnost podíl alel téhož genu (lokusu) při
INTERAKCE NEALELNÍCH GENŮ POLYGENNÍ DĚDIČNOST I. ročník, letní semestr 13. týden 12. - 16.5.2008 Aleš Panczak, ÚBLG 1. LF a VFN Krátké opakování: Jednotková dědičnost podíl alel téhož genu (lokusu) při
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním
 1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
1. Téma : Genetika shrnutí Název DUMu : VY_32_INOVACE_29_SPSOA_BIO_1_CHAM 2. Vypracovala : Hana Chamulová 3. Vytvořeno v projektu EU peníze středním školám Genetika - shrnutí TL2 1. Doplň: heterozygot,
Molekulární genetika, mutace. Mendelismus
 Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
Molekulární genetika, mutace 1) Napište komplementární řetězec k uvedenému řetězci DNA: 5 CGTACGGTTCGATGCACTGTACTGC 3. 2) Napište sekvenci vlákna mrna vzniklé transkripcí molekuly DNA, pokud templátem
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY
 GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
GENETIKA POPULACÍ ŘEŠENÉ PŘÍKLADY 5. Speciální případy náhodného oplození PŘÍKLAD 5.1 Testováním krevních skupin systému AB0 v určité populaci 6 188 bělochů bylo zjištěno, že 2 500 osob s krevní skupinou
Důsledky selekce v populaci - cvičení
 Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován
Genetika a šlechtění lesních dřevin Důsledky selekce v populaci - cvičení Doc. Ing. RNDr. Eva Palátová, PhD. Ing. R. Longauer, CSc. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován

Genetika BIOLOGICKÉ VĚDY EVA ZÁVODNÁ
 BIOLOGICKÉ VĚDY EVA ZÁVODNÁ Genetika - věda studující dědičnost a variabilitu organismů - jako samostatná věda vznikla na počátku 20. století - základy položil J.G. Mendel již v druhé polovině 19. století
BIOLOGICKÉ VĚDY EVA ZÁVODNÁ Genetika - věda studující dědičnost a variabilitu organismů - jako samostatná věda vznikla na počátku 20. století - základy položil J.G. Mendel již v druhé polovině 19. století
Leucismus u ještěrky obecné Lacerta agilis L.
 Leucismus u ještěrky obecné Lacerta agilis L. Tomáš Mazuch*, Tomáš Huňáček * VETERINÁRNÍ A FARMACEUTICKÁ UNIVERZITA BRNO ÚSTAV PARAZITOLOGIE Fyziologie zbarvení plazů Kožní systém je tvořen 3 vrstvami
Leucismus u ještěrky obecné Lacerta agilis L. Tomáš Mazuch*, Tomáš Huňáček * VETERINÁRNÍ A FARMACEUTICKÁ UNIVERZITA BRNO ÚSTAV PARAZITOLOGIE Fyziologie zbarvení plazů Kožní systém je tvořen 3 vrstvami
Selekce v populaci a její důsledky
 Genetika a šlechtění lesních dřevin Selekce v populaci a její důsledky Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Genetika a šlechtění lesních dřevin Selekce v populaci a její důsledky Doc. Ing. RNDr. Eva Palátová, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je spolufinancován Evropským sociálním
Národní program uchování a využívání genetických zdrojů zvířat
 Národní program uchování a využívání genetických zdrojů zvířat METODIKA CHOVU GENETICKÝCH ZDROJŮ RYB K 1.1.2006 jsou genetickými zdroji ryb tyto druhy ve schválených formách a liniích: kapr obecný (Žďárský
Národní program uchování a využívání genetických zdrojů zvířat METODIKA CHOVU GENETICKÝCH ZDROJŮ RYB K 1.1.2006 jsou genetickými zdroji ryb tyto druhy ve schválených formách a liniích: kapr obecný (Žďárský
Ing. Mgr. Denisa Nechanská
 Ing. Mgr. Denisa Nechanská nechanska@af.czu.cz mimotělní oplození velké množství gamet (řádově tisíce) vývoj embryí ve vodě přirozená synchronizace (podmínka úspěchu) období, místo a způsob druhově rozdílné
Ing. Mgr. Denisa Nechanská nechanska@af.czu.cz mimotělní oplození velké množství gamet (řádově tisíce) vývoj embryí ve vodě přirozená synchronizace (podmínka úspěchu) období, místo a způsob druhově rozdílné
Seminář genotyp, fenotyp, krevní skupiny MONOHYBRIDISMUS
 Seminář genotyp, fenotyp, krevní skupiny MONOHYBRIDISMUS Úkol č.1: Sestavte kombinační čtverce pro následující hybridizace jedinců. Uveďte jejich genotypové a fenotypové štěpné poměry. Fenotypové štěpné
Seminář genotyp, fenotyp, krevní skupiny MONOHYBRIDISMUS Úkol č.1: Sestavte kombinační čtverce pro následující hybridizace jedinců. Uveďte jejich genotypové a fenotypové štěpné poměry. Fenotypové štěpné
á ý á á ú ú ř ý ý ů ě ů ř á á á á ě ě š ř ů á ě ě ě ů ř š ý š ě ů ž ář ř ř š ý ář á ě ř á ý ě ů á á á ě á ž ě ě ů ě ý ě ř ě šť Č ý á á ř á ě á ř ý ý á
 É Ř Á Ý Ý Ý ů Ř Ý Ě ů ě ář Ú ř ě ě ě ě ě á ý á á ú ú ř ý ý ů ě ů ř á á á á ě ě š ř ů á ě ě ě ů ř š ý š ě ů ž ář ř ř š ý ář á ě ř á ý ě ů á á á ě á ž ě ě ů ě ý ě ř ě šť Č ý á á ř á ě á ř ý ý á á ě ú ř ě
É Ř Á Ý Ý Ý ů Ř Ý Ě ů ě ář Ú ř ě ě ě ě ě á ý á á ú ú ř ý ý ů ě ů ř á á á á ě ě š ř ů á ě ě ě ů ř š ý š ě ů ž ář ř ř š ý ář á ě ř á ý ě ů á á á ě á ž ě ě ů ě ý ě ř ě šť Č ý á á ř á ě á ř ý ý á á ě ú ř ě
Základní pravidla dědičnosti - Mendelovy a Morganovy zákony
 Obecná genetika Základní pravidla dědičnosti - Mendelovy a Morganovy zákony Ing. Roman LONGAUER, CSc. Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je
Obecná genetika Základní pravidla dědičnosti - Mendelovy a Morganovy zákony Ing. Roman LONGAUER, CSc. Doc. RNDr. Ing. Eva PALÁTOVÁ, PhD. Ústav zakládání a pěstění lesů LDF MENDELU Brno Tento projekt je
QTL u koní. Kmen je skupina koní v rámci plemene, odlišných morfologických a užitkových vlastností (šlechtění na tažné a jezdecké využití).
.") QTL u koní Dnešní plemena koní se odvozují od divokých předků, od: Equus przewalskii (kůň Převalského-kertaka) Equus gmelini (kůň západní) Equus gracilis (kůň severský) Pojmy plemenitby Plemeno je skupina
QTL u koní Dnešní plemena koní se odvozují od divokých předků, od: Equus przewalskii (kůň Převalského-kertaka) Equus gmelini (kůň západní) Equus gracilis (kůň severský) Pojmy plemenitby Plemeno je skupina
Výukový materiál zpracován v rámci projektu EU peníze školám
 http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
http://vtm.zive.cz/aktuality/vzorek-dna-prozradi-priblizny-vek-pachatele Autorem materiálu a všech jeho částí, není-li uvedeno jinak, je Mgr. Eva Strnadová. Dostupné z Metodického portálu www.rvp.cz ;
Ceník akvarijních ryb
 Ceník akvarijních ryb - dostupné - dočasně nedostupné Název zboží cena/ks teplota tvrdost ph Akara hnědá 20,- 24-30 C 11-30 dgh 6,0-8,0 Black molly 15,- 24-30 C 11-30 dgh 6,0-8,0 Black molly - "Black/Gold"
Ceník akvarijních ryb - dostupné - dočasně nedostupné Název zboží cena/ks teplota tvrdost ph Akara hnědá 20,- 24-30 C 11-30 dgh 6,0-8,0 Black molly 15,- 24-30 C 11-30 dgh 6,0-8,0 Black molly - "Black/Gold"
GENETIKA Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální dědičnost
 Polygenní dědičnost Multifaktoriální dědičnost") GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální
GENETIKA vědecké studium dědičnosti a jejich variant studium kontinuity života ve vztahu ke konečné délce života individuálních organismů Monogenní dědičnost (Mendelovská) Polygenní dědičnost Multifaktoriální

Ceník akvarijních ryb
 Ceník akvarijních ryb Pro jistotu dostupnosti požadovaných ryb, nás prosím předem kontaktujte - dostupnost se může velice rychle měnit. - dostupné - dočasně nedostupné Název zboží cena/ks teplota tvrdost
Ceník akvarijních ryb Pro jistotu dostupnosti požadovaných ryb, nás prosím předem kontaktujte - dostupnost se může velice rychle měnit. - dostupné - dočasně nedostupné Název zboží cena/ks teplota tvrdost
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
ž ř ž ř ý é é č ů ý ý ň ý ý ň ň é č ř ř ř é č é ř é
 Ý ý ř ý ů ú ř ž ý ř ý é Ý é ý ý é ř č ú ý ř ý č é ž ý ň ň ž é ř é ř é ř č ř ý é č é ý ý é ř É Á Á Í Á É Ý Í Ů Š Á Ž Ě Ý É Á Ř Ý Á Á ž ř ž ř ý é é č ů ý ý ň ý ý ň ň é č ř ř ř é č é ř é ů ý é ř ů ř é čů
Ý ý ř ý ů ú ř ž ý ř ý é Ý é ý ý é ř č ú ý ř ý č é ž ý ň ň ž é ř é ř é ř č ř ý é č é ý ý é ř É Á Á Í Á É Ý Í Ů Š Á Ž Ě Ý É Á Ř Ý Á Á ž ř ž ř ý é é č ů ý ý ň ý ý ň ň é č ř ř ř é č é ř é ů ý é ř ů ř é čů
Záchyty virů u kaprovitých ryb v ČR
 Funded by the European Union s Seventh Framework Programme Záchyty virů u kaprovitých ryb v ČR Tomáš Veselý Výzkumný ústav veterinárního lékařství, Brno NRL pro virové choroby ryb Vlivy na vznik onemocnění
Funded by the European Union s Seventh Framework Programme Záchyty virů u kaprovitých ryb v ČR Tomáš Veselý Výzkumný ústav veterinárního lékařství, Brno NRL pro virové choroby ryb Vlivy na vznik onemocnění
ů ř Ž ý ý ř ď ř
 ř ů ř ů ř ř ý ů ř ů ů ř ť ý Ž ř ř ř ř Ž ř ú ý Ž ř ů ů ť Ř ý ř ř ř ů ý ý ř ý ň Ž ý ů ř Ž ý ý ř ď ř Á ů ó ř Í ř ý ř ý ř ř ř ř ř ř ř ř ř ý ř ť ř ř ř ý ť ř ď ú É ř ť ý ů ř ý ď ř ř Ž ý ý Í ý ó ů ý ý ř ř Í ř
ř ů ř ů ř ř ý ů ř ů ů ř ť ý Ž ř ř ř ř Ž ř ú ý Ž ř ů ů ť Ř ý ř ř ř ů ý ý ř ý ň Ž ý ů ř Ž ý ý ř ď ř Á ů ó ř Í ř ý ř ý ř ř ř ř ř ř ř ř ř ý ř ť ř ř ř ý ť ř ď ú É ř ť ý ů ř ý ď ř ř Ž ý ý Í ý ó ů ý ý ř ř Í ř
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy
 "Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
"Učení nás bude více bavit aneb moderní výuka oboru lesnictví prostřednictvím ICT ". Základy genetiky, základní pojmy 1/75 Genetika = věda o dědičnosti Studuje biologickou informaci. Organizmy uchovávají,
Základy produkce kapra a dalších druhů ryb
 Základy produkce kapra a dalších druhů ryb prof. Dr. Ing. Jan Mareš,Odd. rybářství a hydrobiologie Mendelova univerzita v Brně www.rybarstvi.eu, mares@mendelu.cz Produkce kapra Historie chovu kapra v českých
Základy produkce kapra a dalších druhů ryb prof. Dr. Ing. Jan Mareš,Odd. rybářství a hydrobiologie Mendelova univerzita v Brně www.rybarstvi.eu, mares@mendelu.cz Produkce kapra Historie chovu kapra v českých
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/
 Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky Populační genetika (KBB/PG)
Základní škola praktická Halenkov VY_32_INOVACE_03_03_03. Ryby
 Základní škola praktická Halenkov VY_32_INOVACE_03_03_03 Ryby Číslo projektu CZ.1.07/1.4.00/21.3185 Klíčová aktivita III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Zařazení učiva v rámci ŠVP Přírodopis
Základní škola praktická Halenkov VY_32_INOVACE_03_03_03 Ryby Číslo projektu CZ.1.07/1.4.00/21.3185 Klíčová aktivita III/2 Inovace a zkvalitnění výuky prostřednictvím ICT Zařazení učiva v rámci ŠVP Přírodopis
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Pohlavní typy Drosophila Protenor Člověk Lymantria/Abraxas (bekyně) Habrobracon/haplodiploidie
Vejcorodé ryby. Dělení ryb podle způsobu rozmnožování 19/02/2017. Úvod. Úvod. Rozmnožování akvarijních ryb. Akvaristika.
 9/0/07 Úvod Akvaristika 07 Rozmnožování akvarijních ryb Přednáška #6 Jiří Patoka patoka@af.czu.cz Úvod Dělení ryb podle způsobu rozmnožování Vejcorodé ryby 9/0/07 Vejcorodé ryby třecí vyrážka samců kaprovitých
9/0/07 Úvod Akvaristika 07 Rozmnožování akvarijních ryb Přednáška #6 Jiří Patoka patoka@af.czu.cz Úvod Dělení ryb podle způsobu rozmnožování Vejcorodé ryby 9/0/07 Vejcorodé ryby třecí vyrážka samců kaprovitých
ň ú Ú ů é é ň ů ž ů ů ů ů é é é é ú ň é ú ú ů é é ů ů Č é ň ú ú ů é é ů Ť ň é ů ů ú ň é ú ť ť é é é ů é é ů é é ť ň ú ú ů é é ů ů ú é ů é ů é ů ť ů ú
 Í ÁŇ Ý ÚŘ ú ů é é Č ó ž ů ú é ú ú ť é é é é ž Č ů é é ů ů ň ť ú Í ů é é ť é ň é ů ů ú ú Í é é é ú Ú ů Í Č Č ú ň ú Ú ů é é ň ů ž ů ů ů ů é é é é ú ň é ú ú ů é é ů ů Č é ň ú ú ů é é ů Ť ň é ů ů ú ň é ú ť
Í ÁŇ Ý ÚŘ ú ů é é Č ó ž ů ú é ú ú ť é é é é ž Č ů é é ů ů ň ť ú Í ů é é ť é ň é ů ů ú ú Í é é é ú Ú ů Í Č Č ú ň ú Ú ů é é ň ů ž ů ů ů ů é é é é ú ň é ú ú ů é é ů ů Č é ň ú ú ů é é ů Ť ň é ů ů ú ň é ú ť
PRAŽSKÝ KRYSAŘÍK - STANDARD
 PRAŽSKÝ KRYSAŘÍK - STANDARD CELKOVÝ VZHLED Malý hladkosrstý pes, téměř kvadratické, kompaktní tělesné stavby. Vzdor svému malému vzrůstu dobře pohyblivý a vytrvalý. Dobrý pohlavní výraz psa i feny. CHOVÁNÍ
PRAŽSKÝ KRYSAŘÍK - STANDARD CELKOVÝ VZHLED Malý hladkosrstý pes, téměř kvadratické, kompaktní tělesné stavby. Vzdor svému malému vzrůstu dobře pohyblivý a vytrvalý. Dobrý pohlavní výraz psa i feny. CHOVÁNÍ