Funkčnístruktury chromosomu
|
|
|
- Miluše Burešová
- před 8 lety
- Počet zobrazení:
Transkript
1 Cytogenetika03 Funkčnístruktury chromosomu Magda Zrzavá František Marec

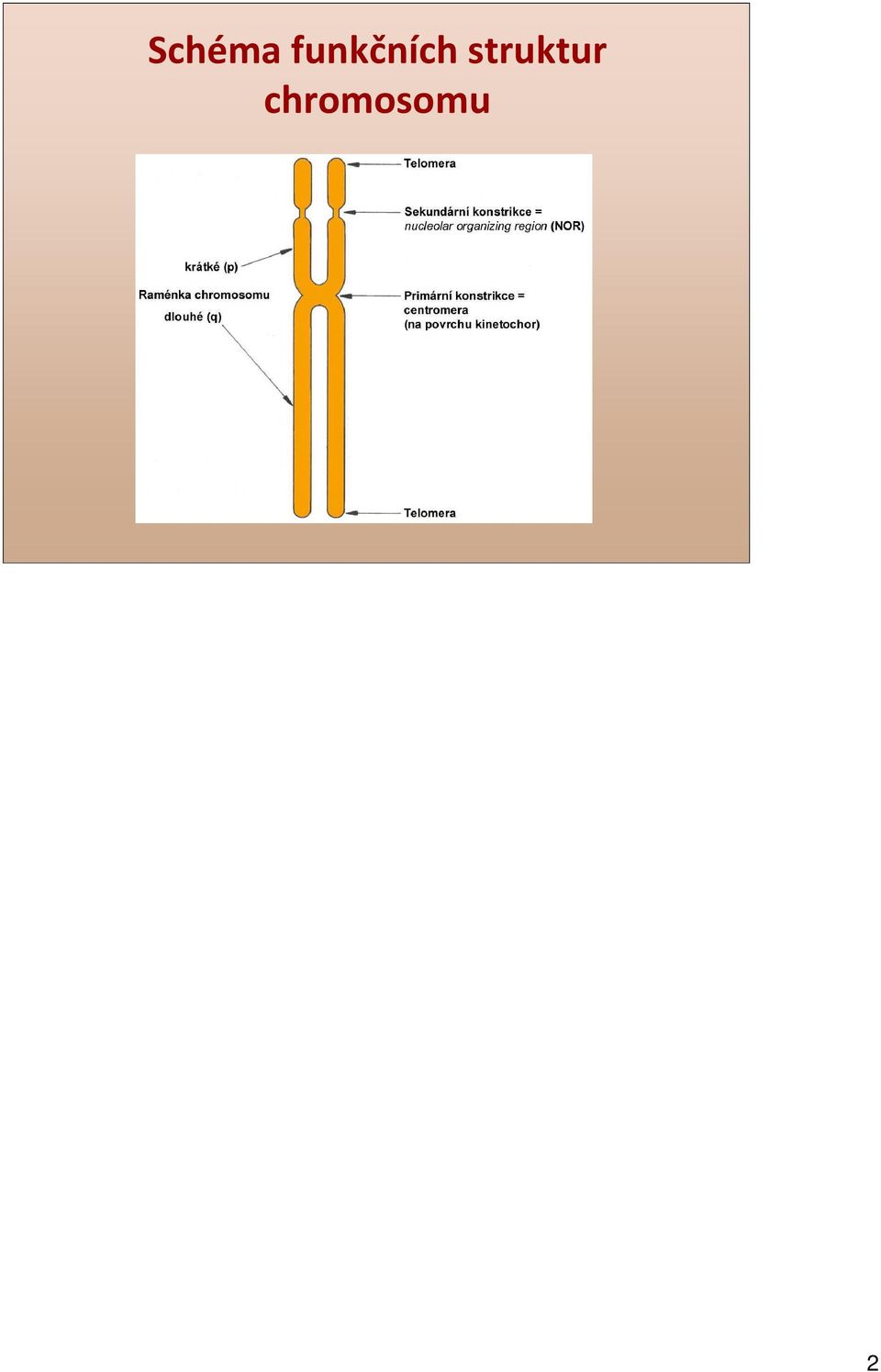
2 Schéma funkčních struktur chromosomu 2
3 Centromery

4 Centromera = primární konstrikce - rozděluje chromosom na raménka - místo nasedání kinetochoru - funkce: bezchybný rozchod chromosomů - ztráta centromery ztráta chromosomu - fúze chromosomů vznik dicentrického chromosomu zastaveníbuněčného dělení smrt buňky

5 Děleníchromosomůpodle typu centromery Monocetrické chromosomy centromerickáaktivita soustředěna do jednoho místa Podle velikosti dělíme na: - bodové(point) u Saccharomyces cerevisiae -regionální(regional) např. Schizosaccharomycespombe, člověk, kur, Drosophila Holocetrické chromosomy tzv. difuzní centromera-kinetochorrozptýlen po celém chromosomu -např. Caenorhabditiselegans, motýli, mšice Centromery Podle velikosti centromery rozdělujeme chromosomy na monocentrické a holocentrické (holokinetické). Monocentrické centromery jsou lokalizované v jediném místě na chromosomu a podle jejich délky je lze rozdělit na dva podtypy: - bodová (point) centromera, kdy je formování kinetochoru vázáno na určitou sekvenci DNA (např. Saccharomyces cerevisiae), zaujímá velice malou oblast (1 nukleosom), na kterou se upíná jeden mikrotubul. U S. cerevisiae je centromerická DNA tvořena 125 bp sekvencí složené ze tří elementů (CDEI, CDEII a CDEIII). - regionální (regional) centromera, kde je oblast centromery sice na jednom místě, ale zabírá větší oblast. Přestože se v těchto oblastech často nacházejí určité typy sekvencí DNA (u vyšších eukaryot je to často tandemové repetice, např. alfa satelitní DNA u primátů), zdá se, že fungování centromery je vymezeno spíše epigeneticky. Najdeme je např. u člověka, kura, drozofily a Schizosaccharomyces pombe. Lidská regionální centromera je komplexní. Dvě rodiny alfa satelitní DNA (červené a šedé šipky) se nacházejí pouze v centromeře. Alfa satelitní DNA typu I (červená šipka) je složena z chromosomálně specifických repetitivních jednotek vyššího řádu, obsahuje CENP-B boxy a nasedá na ní CENP-A. Alfa satelitní DNA typu II (šedé šipky) se skládá z různých alfoidních monomerů (různě barevné trojúhelníky). Kinetochor váže mikrotubulů. U většiny studovaných eukaryot je funkčníčást centromerické DNA (jejíž délka je vřádu desítek kilobází) obklopena pericentromerickým heterochromatinem, který může mít i několik megabází, přičemž sekvence v těchto oblastech mohou být stejné (Plohl et al. 2014). Holokinetické chromosomy mají kinetochor lokalizovaný po celé nebo téměř celé délce chromosomu. Vyskytují se např. u motýlů, mšic, Caenorhabditis elegans. Obrázky z práce Varnagelli et al
 centromera, kdy je formování kinetochoru vázáno na")
6 Monocentrické versus holokinetické chromosomy Výskyt: od primitivních organismů po vyšší rostliny a živočichy Metafáze -chromatidy tvar X, V Nízkáodolnost vůči ionizačnímu záření a klastogenům obecně Výskyt: některé skupiny organismů (prvok Nyctotherus, Nematoda; rostliny čeledíjuncaceae, Cyperaceaea Liliaceae; roztoči, Hemiptera, Trichoptera, Lepidoptera) Metafáze chromatidy paralelně Vysokáodolnost vůči ionizačnímu záření a klastogenům obecně 6
 Metafáze chromatidy paralelně Vysokáodolnost vůči ionizačnímu záření a")
7 Monocentrické versus holokinetické chromosomy Výhodou kinetochoru lokalizovaného podél (témeř) celého chromosomu je to, že dojde-li k zlomu chromosomu, oba vzniklé fragmenty se mohou přichytit k vláknům dělícího vřeténka a mohou se dostat do dceřiné buňky. Na druhou stranu centromera lokalizovaná do jednoho malého místa, má při chromosomálním zlomu za následek vznik acentrického chromosomu (nemá žádnou centromeru), a protože konce chromosomů bez telomer jsou lepivé a mají silnou tendenci se spojit, vznikne z druhého fragmentu dicentrický chromosom se dvěma chromatidami. Acentrický chromosom se nemůže přichytit na dělící vřetýnko a je při buněčném dělení ztracen. Dicentrický chromosom je pro buňku fatální, protože pokud nedojde k inaktivaci jedné centromery, obě centromery mohou táhnout chromosom k opačným pólům buňky (tzv. anafázní můstek), což vede k zastavení dělení a smrti buňky. 7

8 Centromerické proteiny Řada funkcí: - tvorba kinetochoru - vazba mikrotubulů - kontrolní bod mezi metafází a anafází - spojení sesterských centromer - motorové proteiny Centromerické proteiny S centromerami je asociovány proteiny, které zajišťujířadu funkcí (účastní se tvorby kinetochoru, vazby mikrotubulů, kontrolního bodu mezi metafází a anafází, spojení sesterských chromatid, atd.). Některé z nich se označují CENP, což neznamená nic jiného než CENtromerický Protein, nemají společnou funkci ani strukturu a byly izolovány pomocí protilátek izolovaných z jedinců s autoimunitními chorobami. Některé jsou markery aktivních centromer a jsou nezbytné pro jejich fungování (např. CENP-A, CENP-C,...), jiné jsou zbytné a nacházejí se i na neaktivních centromerách (např. CENP-B). Na obázku a na další snímku jsou savčí centromerické proteiny, z nichž některé mají homology i u bezobratlých a kvasinek. Obrázek podle Sumner
, jiné jsou zbytné a nacházejí se i na neaktivních centromerách (např. CENP-B).")
9 Savčí centromerické proteiny CENP-A centromerickávarianta histonu H3 (cenh3) -markeraktivních centromer(i neocentromer a umělých centromer) - lokalizován na vnitřní ploše kinetochoru CENP-B vazba na CENP-B boxy v cen. DNA -i na neaktivních cen., chybína Y -rozhodujeo vnikunovýchcentromera inaktivaci stávajících (povolává CENP- A/zprostředkuje H3K9Me3) CENP-C a CENP-E markery aktivních centromer, součásti kinetochoru - potřeba pro seřazení metafázních chromosomů do roviny hbub1, hbubr1, zw10 a hmad-kontrolníbod mezi metafázía anafází MCAK a dynein- kinetochorové motorové proteiny, na vnější straně kinetochoru INCENps a CLiPs- drží pohromadě sesterské chromatidy CENP-A je markerem aktivních centromer, je centromerickou variantou histonu H3 (též zvaný CenH3, Cse4 u S. cerevisiae, CID u D. melanogaster, cehcp-3 u C. elegans a HTR12 u Arabadopsis thaliana). Je přítomen nejen u klasických centromer, ale i u přirozených nebo umělých neocentromer. Jeho knock-out/knock-down způsobí neschopnost centromer vykonávat svou funkci. Je lokalizován na vnitřní ploše kinetochoru (inner plate) a zřejmě centromerickému chromatinu uděluje větší pevnost (rigiditu), což je potřeba, aby centromera odolala tahu mikrotubulů při rozchodu sesterských chromatid. CENP-B je protein savčích centromer, na jejichž DNA se váže v místech specifických sekvencí DNA (CENP-B boxů). Může se nacházet i na neaktivních centromerách, naopak v některých aktivních chybí (např. lidský a myší chromosom Y), není tedy nutný pro fungování centromer (myši bez CENP-B přežívají, mají jen menší váhu a méně spermií). CENP-B je tedy zřejmě potřebný pro ustavené nové centromery, ale není potřeba k jejímu udržení. Ukazuje se, že CENP- B má dvojí roli: 1) je schopen povolat k centromerické DNA CENP-A, což je klíčové pro vznik nových centromer (např. u umělých chromosomů), 2) navodit reprimující epigenetickou modifikaci H3K9Me3, která vede k formování heterochromatitu. Tyto antagonistické síly rozhodují o tom, jestli se satelitní sekvence stane centromerou nebo neaktivním heterochromatinem. CENP-C a CENP-E jsou markery aktivních centromer, jsou součástí kinetochoru a jsou potřeba pro seřazení metafázních chromosomu do roviny. Při jejich selhání buňka nemůže vstoupit do anafáze. Mají homology u S. cerevisiae a rostlin (Hordeum vulgare a Vicia faba). hbub1, hbubr1, zw10 a hmad se účastní kontrolního bodu mezi metafází a anafází. BUB a MAD mají homology u S. cerevisiae a drozofily. MCAK (mitotic centromere-associated kinesin) a dynein (stejně jako CENP-E) jsou kinetochorové motorové proteiny umístěné na vnější straně kinetochoru. INCENps a CLiPs (chromatid linking proteins) drží pohromadě sesterské chromatidy, CLiPs jsou specifické pro sesterské centromery. Info z Sumner 2001, Varnagelli et al

10 Sclerodermia a studium kinetochorů Sclerodermia - autoimunitní multisystémová choroba poškozeníkůže, škáry, kloubů, pojiva vnitřních orgánů, plic, ledvin Celkovásklerodermie (Sclerodermia diffusa) Lokalizovaná sklerodermie Pacient tvoří protilátky proti centromerám Protilátky proti centromerám izolované ze séra pacientů se sklerodermií byly využívány k výzkumu kinetochorových proteinů u živočichů i rostlin kinetochorové proteiny jsou velmi konzervativní Velký průlom ve výzkumu kinetochorů nastav v roce 1980, kdy se přišlo na to, že pacienti s CREST syndromem (forma sklerodermie, autoimunitní choroba) produkují protilátky proti centromerám. Protilátky byly použity pro výzkum kinetochorových proteinů u živočichů i rostlin, což svědčí o tom, že jsou tyto proteiny velmi konzervativní. 10
 produkují protilátky proti centromerám.")
11 Centromerická DNA - Často tvořena repetitivní DNA obtížné sekvenovat -Co víme: 1) Často satelitní DNA/mobilní elementy, ale není pravidlem 2) Rychle se vyvíjí(concerted evolution), koevoluce s cent. proteiny reprodukční bariera speciace 3) Neníznáma univerzálníeukaryotnícentromerickásekvence, ale některé motivy jsou společné pro skupinu (CENP-B box u savců). 4) Funkčnícentromera(desítky kbp)často obklopena pericentromerickým heterochromatinem(i > Mb) Centromerická DNA Sekvenování centromerické DNA a heterochromatinu obecně je problematická záležitost, protože obsahují repetitivní DNA, jejíž sekvence je obtížné bioinformaticky složit. Proto i u jinak osekvenovaných genomů centromery jsou oblasti, kteréčasto osekvenované nejsou. Nicméně u některých druhů rostlin, hub a živočichů sekvence centromerické DNA známe, a z nich vyplývá, že: 1) velmi často je centromera tvořena satelitní DNA nebo mobilními elementy, 2) centromerická DNA se rychle vyvíjí a liší se i mezi příbuznými druhy, 3) není známa žádná univerzální centromerická sekvence, ale některé motivy jsou sdíleny v rámci skupiny, např. tzv. CENP-B box, 17bp sekvence, na kterou se váže CENP-B a je klíčová pro formování kinetochoru. CENP-B box se vyskytuje v alfa-satdna člověka a různých alfoidních satelitů v centromerách savců a podobná sekvence se stejnou funkcí byla nalezena i u některých bezobratlých a rostlin. SatDNA se vyvíjí podle principu tzv. concerted evolution, kdy jsou mutace mezi monomery sjednocovány nerovnoměrným crossing-overem, genovou konverzí, replikací valivou kružnicí a transpozicí. Těmito mechanismy se nová mutace může rychle rozšířit do všech monomerů satdna chromosomu nebo chromosomů. Rychlá evoluce centromerické DNA vyžaduje koevoluci centromerických proteinů. Tyto změny mohou vést až k reprodukční izolaci a speciaci (Plohl et al. 2014). Lidské centromery V centromerách lidských chromosomů jsou dva typy alfa-satdna: monomerická a repetice vyššího řádu (HOR, higher-order repeat). HORs se skládají z 2-20 různých 171 bazových monomerů a jsou obvykle chromosomálně specifické. Vyskytují se v aktivní centromeře i v pericentromerickém heterochtomatinu a jsou obklopené monomerickou alfa-satdna (Plohl et al. 2014). Sondy k HORs se používají v lidské cytogenetice k identifikaci jednotlivých chromosomů.

12 Kinetochory se podílejí na: Kinetochor 1) přichycenímikrotubulůdělícího vřeténka a srovnání metafázních chromosomů do roviny 2) opravěchybného přichycenímikrotubulůna chromosom 3) metafáznímkontrolním bodu, kdy buňka nepokračuje do anafáze, dokud nejsou obě chromatidyvšech chromosomůpřichycenéna dělící vřeténko 4) napětívcentromerách, kteréje signálem pro opuštěnímetafázníhokontrolního bodu a vstup do anafáze 12
 napětívcentromerách, kteréje signálem pro opuštěnímetafázníhokontrolního bodu a vstup do")
13 Typy kinetochorů - U některých organismů není patrný, mikrotubuly vstupují přímo do chromatinu (kvasinky) - Typy kinetochorů: 1) ballandcup vyššírostliny a některý hmyz (Orthoptera) kinetochor = kulička sedícína prohlubni (centromeře) EM řezu metafázníhochromosomu s kinetochorem Ball and cup 2) třívrstvý(trilaminar) nižšírostliny, většina živočichů -vnějšía vnitřnívrstva husté, prostřednířídká. - vnější vrstva vazba mikrotubulů - vnitřní nasedá na centromeru Model trojvrstevného kinetochoru: chromatinovávlákna spojujívnitřnía vnější vrstvu Kinetochor je struktura, která zprostředkovává kontakt mezi centromerou a mikrotubuly dělícího vřeténka. Některé organizmy (např. kvasinky) kinetochor nemají (nebo není patrný) a mikrotubuly vstupují přímo do chromatinu. Kinetochory jsou dvou typů: 1) ball and cup, kdy se kinetochor jeví jako nepravidelná masa (kulička) sedící na prohlubni, kterou je centromera (pohárek). Tento typ je přítomen u vyšších rostlin a u některých skupin hmyzu (Orthoptera). Druhým typem je třívrstvý kinetochor, který je složen z husté vnější vrstvy, husté vnitřní vrstvy sedící na centromeře, a řídké prostřední vrstvy. Většina mikrotubulů se upíná k vnější vrstvě, ale některé pronikají až do vnitřní vrstvy. Třívrstvý kinetochor je přítomen v pozdní profázi a metafázi, i když některé jeho složky lze detekovat i v interfázi. Nachází se u nižších rostlin a většiny živočichů. 13

14 Typy kinetochorů EM řez monocentrickým metafázním chromosomem řasy Oedogonium EM řez holokinetickým metafázním chromosomem mikrojádra prvoka Nyctotherus ovalis

15 NOR

16 Oblast organizátoru jadérka nucleolusorganizingregion (NOR) Je tvořena rdna geny místo intenzivnítranskripce ribosomální RNA (=> kódují rrna) rrnatvoříspolu s proteiny ribosomy=>nor přímo zodpovídáza vznik jadérka (nucleolus) Počet NOR druhověspecifický (člověk 5 párůautosomůnese NOR) Místo, kde je lokalizována rdna (geny pro 18S, 28S a 5,8S rrna), je nazýváno organizátor jadérka (NOR, nucleolus organizer region), protože se kolem něj v interfázním jádře formuje jadérko. U člověka je rdna lokalizována na p- raménkách pěti párů akrocentrických chromosomů, které nesou sekundární konstrikci, což je místo, kde je právě lokalizovaná 45S rdna. To, proč se toto místo jeví jako konstrikce není jasné, ale protože rdna je transkribována až do začátku profáze, je možné, že se tato oblast jednoduše začne později kondenzovat. 16

17 Schéma rdna transkripčníjednotky vyšších eukaryot (tato struktura vysoce konzervovaná) rdna jsou klastry genů kódujících ribosomální RNA. Klastr obsahuje stovky kopií tzv. 45S rdna, což je název pro operon obsahující kromě genů pro 18S, 5,8S a 28S rrna i přepisující se oblasti ETS (external transcribed spacer) a ITS (internal transcribed spacer). Kopie 45S jsou odděleny nepřepisující se oblastí (nontranscribed spacer, NTR). Počty ribosomálních genů se u různých organizmů liší. Ty, které mají malý genom, mají méně než 100 rrna genů, naopak některé rostliny a obojživelníci, kteří mají velmi velké genomy, mohou mít až kopií rrna genů. Geny pro 5S rrna jsou lokalizované mimo klastr, často na jiném chromosomu než 45S, a jsou srovnatelně početné jako 45S. 17

18 Metody lokalizace NOR a jadérka -BarveníAgNO 3 vizualizujeněkteréproteiny v jadérku (interfáze) a v NORech, které byly v interfázi aktivní (mitóza, meióza) -FISH s rdna sondou lokalizuje i neaktivní NORy NORy na metafázních chromosomech Jadérko v interfázním jádře - YOYO fluorochrom barvícídna i RNA rdna-fish se sondou pro 18S rdna pozdní profáze (YOYO-1) Detekce rdna na chromosomech Klasickou metodou vizualizace NORů a jadérek je barvení dusičnanem stříbrným. Tato metoda detekuje jadérka v interfázi a na metafázních chromosomech NORy, které byly aktivní v předchozí interfázi. Z toho ovšem vyplývá, že takto nemusejí (a obvykle nejsou) detekovány všechny NORy (u člověka obvykle jen 7-8 z 10). Barvení AgNO 3 detekuje některé proteiny, které byly součástí jadérka a zůstaly asociované s NOR i během buněčného dělení. Jsou to především RNA polymeráza I a transkripční faktory. Další metodou vizualizace rdna je FISH. Výhodou je to, že lze detekovat i neaktivní rdna. Obrázky z Bombarová et al a Novotná et al
 detekovány všechny NORy (u člověka obvykle jen 7-8 z 10).")
19 Jadérko v interfázi Ultrastruktura: Fibrillarcentre lokalizace rdna a RNA polymerázy I syntéza rrna Densefibrillarcomponent asociace rrna s ribosomálními proteiny Granular component skládání preribosomů Jadérko přítomno jen v interfázi a v profázi (do rozpuštění jaderné membrány) Jadérko existuje pouze v interfázi a na začátku buněčného dělení (profáze) do rozpuštění jaderné membrány. Funkcí jadérka je transkripce 45S rdna, rozdělení vzniklé RNA na 18S, 5,8S a 28S RNA a jejich spojení s ribosomálními proteiny v pre-ribosomech. Jadérko se skládá z tří vrstev: vláknité centrum (fibrillar centre, FC), obklopené tenkou vrstvou o vysoké hustotě (dense fibrillar component, DFC) a vnější hmotou (granular component, GC). V FC jsou lokalizované rdna a RNA polymeráza I, v DFC se rrna váže na proteiny a v GC dochází k skládání pre-ribosomů. Obrázek ultrastruktury jadérka z 19
, obklopené tenkou vrstvou o vysoké hustotě (dense fibrillar component, DFC) a vnější hmotou (granular component, GC).")
20 Telomery

21 = fyzické konce eukaryotních chromosomů - řada funkcí: Telomery 1) ochrana chromosomů před fúzemi 2) řeší problém replikace konců 3) omezuje počet buněčných dělení role ve stárnutí a rakovině Telomery jsou DNA-proteinové struktury na koncích eukaryotních chromosomů. Majířadu funkcí: 1) Telomery chrání chromosomy před fúzemi, protože chromosomy bez nich mají tendenci se k sobě lepit a tvořit dicentrické chromosomy. Zároveň zajišťují, že konce chromosomů nejsou rozpoznávány reparačními mechanismy buňky jako dvouřetězcové zlomy, což by mohlo vést až ke smrti buňky. 2) Telomeryřeší problém replikace konců, tj. fakt, že díky neschopnosti DNA polymerázy začít syntézu DNA bez volné OH- skupiny při každé replikaci ubyde kousek DNA (viz dále). 3) Počet dělení, která může buněčná linie absolvovat, je omezená (Hayflickův limit), což slouží jako ochrana před rakovinným bujením. Negativní stránkou tohoto mechanismu je stárnutí, kdy v tkáních nemohou být nahrazeny odumřelé buňky. Telomery v tomto mechanismu díky postupnému ubývání při každé replikaci DNA slouží jako počítadlo buněčných dělení, která buňka již absolvovala. 21
22 Objev telomer Barbara McClintock - Přirozené konce chromosomů se chovají jinak než chromosomální zlomy -Telomery (telos konec a meros část) Chromosomy chráněné telomerami Napojeníchromosomůna dělící vřeténko Úspěšný rozchod chromosomů Chybějící telomery Vznik dicentrického chromosomu Rozchod chromosomů znemožněn Smrt buňky Barbara McClintock pozorovala u kukuřice, že se přirozené konce chromosomů chovají jinak než dvouřetězcové zlomy, a že tedy musejí mít nějaký ochranný mechanismus, který brání jejich fúzím. Tyto konce byly pojmenovány telomery z řeckého telos konec a meros část (název vymyslel Herman Muller, který objevil telomery u drozofily paralelně s McClintock). Až později byla objevena role telomer v zachování integrity genomu, jejich struktura a způsob fungování. B. McClintock - americká cytogenetička, 1983 dostala Nobelovu cenu za fyziologii a medicínu za objev mobilních elementů u kukuřice. Studovala centromery a telomer, rekombinaci, vytvořila první genetickou mapu chromosomů kukuřice. 22
23 Jak telomery vypadají Elisabeth Blackburn práce s modelovým organismem Tetrahymena thermophila Prvok Tetrahymenathermophila-2 jádra: Micronucleus - předáváno do další generace (ochrana DNA) - transkripčně neaktivní Macronucleus - genetický materiál modifikován nedědí se - transkripčně aktivní - masivní fragmentace DNA syntéza telomer Mochizuki 2010 Objev první telomerické sekvence(ttgggg) V roce 1978 se podařilo Elisabeth Blackburn identifikovat v telomerách u prvoka Tetrahymena thermophila (Ciliophora, Ciliata, Chromalveolata) sekvenci TTGGGG, která je v telomeře v mnoha tandemově uspořádaných kopiích. Výběr nálevníka byl geniální v tom, že tato skupina má dva typy jader. Mikronukleus obsahuje normální chromosomy, které ale zůstávají neaktivní a slouží jen k meióze a k přenosu genetické informace do další generace. Makronukleus vzniká z mikronuklea fragmentací chromosomů, delecemi, amplifikacemi a syntézou nových telomer. Výsledkem je polyploidní jádro plné transkripčně aktivních acentrických fragmentů DNA, z nichž každý je opatřen vlastními telomerami. Při vzniku makronuklea je tedy telomerická sekvence zastoupena ve vysoké koncentraci. O pár let později prokázali Blackburn a Jack Szostak, že telomerická sekvence z Tetrahymeny může iniciovat vznik nové telomery u kvasinky. 23
24 Podoba telomer je velice konzervativní Nalezeno několik variant sekvencí často stejné v rámci velkých skupin: TTTAGGG většina rostlin TTAGGG většina mnohobuněčných živočichů TTAGG většina členovců ALE existujívýjimky jinésekvence a mechanismy prodlužování telomer Po objevu telomerické sekvence u Tetrahymeny se podařilo identifikovat několik dalších telomerických sekvencí. Zajímavé je, že jsou často společné pro velké skupiny eukaryot (např. TTTAGGG u většiny rostlin, TTAGG u většiny členovců). Byly ale objeveny i druhy, které nemají žádnou ze známých telomerických sekvenci, takže mohou mít buď dosud nenalezenou variantu nebo vynalezly jiný mechanismus udržování telomer. Dva z těchto alternativních mechanismů se podařilo prozkoumat, oba u dvoukřídlého hmyzu 1) transpozice retroelementů u drozofily a 2) rekombinace mezi satelitní DNA (Chironomus, Anopheles) (viz dále). 24
25 Struktura telomer Struktura telomer DNA: Jednořetězcový převis bohatý na guanin (je prodlužován telomerázou) invaduje do dvouřetězcové oblasti a vytvoří T-smyčku (tzv. telomerickou smyčku). T-smyčka tak ukryje konec telomery před exonukleázami a reparačními mechanismy buňky, které by volný konec považovaly za zlomenou DNA. Proteiny: Na telomery se váže multiproteinový komplex shelterin. Ten obsahuje jak proteiny, které se váží na jednořetězcový převis (Pot1 (protection of telomeres) +TPP1 heterodimer), tak na dvouřetězcovou oblast: TRF1 a TRF2 (telomere repeat binding factors), jejich kofaktory Rap1 (repressor activator protein 1) a Tin2 (TRF1-interacting nuclear protein 2). TRF1 a jeho interagující proteiny zřejmě regulují prodlužování telomer tím, že ovlivňují přístup telomerázy. TRF2 a Pot1 také ovlivňují délku telomer a navíc brání spojování konců chromosomů. Obrázek z Martínez a Blasco 2010
26 Struktura telomer u drozofily - Telomery prodlužovány transpozicí -Non-LTR retrotransposony vmezeřujíse do telomery -Konec chráněn čepičkou proteinovým komplexem konec nenírozpoznán jako zlom DNA + chráněn před degradací Telomera u drozofily je tvořena non-ltr retrotransposony Het-A, TART a TAHRE, které se do ní vmezeřují. Úplný konec telomery nese čepičku proteinový komplex, díky kterému je konec chromosomu chráněn před degradací a není rozpoznán jako dvouřetězcový zlom DNA. Obrázek z Mason et al
27 Prodlužování telomer rekombinací Chromosomy dvoukřídlého hmyzu zakončeny dlouhými satelitními repeticemi- např.: pakomár Chironomus komár Anopheles Alternativnímechanismus prodlužovánítelomer(alt): REKOMBINACE Druhým alternativním mechanismem prodlužování telomer je rekombinace mezi různě dlouhými telomerickými sekvencemi. Podle tohoto modelu se telomery prodlužují invazí 3 konce řetězce DNA s krátkou telomerou do dvoušroubovice s dlouhou telomerou. Dlouhá telomera pak poslouží jako templát pro syntézu DNA (čili k prodloužení telomery invadujícího řetězce). Po návratu toto prodloužené vlákno poslouží jako templát pro prodloužení 5 konce druhého vlákna s krátkou telomerou. Obrázek z Mefford & Trask,
28 Telomeryřešíproblém replikace konců DNA James Watson Při replikaci DNA docházíke zkrácenímolekuly (problém replikace konců) Schéma replikace DNA DNA polymerázaneumízačít syntézu nového vlákna DNA, RNA polymeráza ano S každou replikacíse DNA o kus zkrátí Řešení: dlouhénekódujícíúseky DNA na koncích chromosomů neboli telomery. Ale i telomery se musejí nějak obnovovat! dvoušroubovice DNA replikace DNA začíná od RNA primerů syntéza nových vláken DNA podle původních vláken odbourání RNA primerů - nová vlákna nejsou kompletní V 70. letech poznamenal James Watson, že způsob, jakým se DNA replikuje, neumožňuje zreplikovat i její úplné konce, takže by se při každém dělení buňky měla kousek zkrátit. Nazval tento jev problémem replikace konců. Přibližně ve stejnou dobu pochopil Alexej Olovnikov, že zkracování konců chromosomů může být příčinou omezeného počtu dělení normálních buněk a našel tak souvislost mezi problémem replikace konců a Hayflickovým limitem. Zkracování konců chromosomů funguje jako hodiny, které odměřují počet dělení, která již buňka absolvovala a kolik jich ještě absolvovat může. Problém replikace konců je založen na faktu, že DNA polymeráza potřebuje k syntéze DNA volnou OH- skupinu na deoxyribóze, bez níž nedokáže začít. Replikace DNA začíná tedy syntézou krátkých RNA primerů, protože RNA polymeráza volný OH- konec nepotřebuje a může začít syntézu RNA kdekoli. Na vzniklé primery nasedne DNA polymeráza a začne je prodlužovat vznikající DNA. RNA primery jsou následně odstraněny, DNA polymeráza vyplní vzniklé mezery DNA (využívá OH-skupiny z DNA, kterou syntetizovala předtím pomocí RNA primerů) a řetězce jsou spojené ligázou. Jedinou výjimkou jsou úplné začátky chromosomů, kde po odstranění RNA primeru nemá DNA polymeráza kam nasednout a kde se tedy DNA nezreplikuje. Každou replikací DNA by se tedy molekula DNA měla o kus zkrátit. Problém replikace konců a jejich prodlužování je dobře znázorněn na tomto videu:
29 Telomeráza - Objevena u Tetrahymena thermophila - Enzym doplňující telomerickou DNA - Reverzní transkriptáza syntetizuje DNA podle RNA - RNA matrice je součástí enzymu 2009 udělena Nobelova cena za fyziologii a medicínu za objev telomerázy Elizabeth Blackburn Carol W. Greider Jack W. Szostak V roce 1985 Blackburn a Carol Greider objevily telomerázu, enzym, který prodlužuje telomery a kompenzuje tím jejich zkracování během replikace DNA. Telomeráza byla izolována z Tetrahymeny po páření v době formování makronuklea, kdy se masově syntetizují telomery. Telomerázu původně pojmenovaly telomerická terminální transferáza, později po její purifikaci zjistily, že enzym v sobě obsahuje RNA složku, komplementární k sekvenci telomery, která slouží jako templát pro syntézu telomerické DNA, zatímco proteinová složka (TERT) má enzymatickou aktivitu. Telomeráza je tedy reverzní transkriptáza, enzym, který syntetizuje DNA podle RNA matrice. Telomeráza složena z několika podjednotek: TERT reverzní transkriptáza TR (TERC) RNA templát
30 Telomery- stárnutí vs. nesmrtelnost August Weissmann - stárnutí je způsobeno omezeným množstvím buněčných dělení A. Weissmann Alexis Carrel -kultivoval kuřecífibroblasty34 let stárnutíje vlastnostítěla, ne buněk - nepodařilo se zopakovat Paul Moorhead a Leonard Hayflick -kultivace lidských fibroblastů počet buněčných dělení omezen na Hayflickův limit L. Hayflick A. Carrel V roce 1881 přišel německý biolog August Weismann s myšlenkou, že smrt je důsledkem omezenému množství buněčných dělení a tím pádem i neschopnosti tkání se donekonečna obnovovat. Tuto myšlenku napadl Alexis Carrel (nositel Nobelovy ceny za fyziologii a medicínu 1912), který ve své laboratoři pěstoval buněčnou kulturu z kuřecích fibroblastů kontinuálně 34 let, a tvrdil, že buňky se mohou dělit donekonečna a to, že to nedělají, je způsobeno chybnou kultivací. Stárnutí bylo podle něj znakem mnohobuněčného těla jako celku. Jeho práci se ale nikomu nepodařilo zopakovat a jeho koncept nesmrtelných buněk byl opuštěn po publikování práce Paula Moorheada a Leonarda Hayflicka, kteří prokázali, že počet dělení buněk je omezen na konkrétní počet. Kultivovali lidské fibroblasty izolované z kůže a zjistili, že se tyto buňky přestali dělit po děleních. Počet dělení, které může buňka absolvovat, se nazývá Hayflickův limit.
31 Hayflickův limit neplatí pro všechny Platí Neplatí ryby většina savců plazi obojživelníci ptáci bezobratlí
32 Hayflickůvlimit obrana proti rakovině Alexej Olovnikov Zkracováníkoncůchromosomůmůže být příčinou omezeného počtu dělenínormálních buněk našel tak souvislost mezi problémem replikace koncůa Hayflickovýmlimitem (1971). Hayflickůvlimit (1961) počet dělení, kterábuňka může absolvovat telomery= markerstáříbuňky (mnoho dělení = krátké telomery) Ale co telomeráza? Alexej Olovnikov je ruský biolog, který v roce 1971 postuloval teorii, že příčinou omezeného množství dělení buněk je zkracování telomer. Ty tedy slouží jako biologické hodiny odměřující kolik dělení ještě může buňka absolvovat. Přestože byl první, kdo tuto teorii publikoval, nebyl navržen na Nobelovu cenu, ale v roce 2009 dostal Demidovovu cenu, prestižní ocenění udělované členům Ruské akademie věd.
33 Telomeráza, kmenovébuňky a rakovina U člověka je telomeráza aktivní pouze během embryonálního vývoje a v dospělosti v kmenových buňkách Cca 85% lidských nádorůmáaktivovanou telomerázu. Model role telomer v stárnutí a rakovině založený na kmenových buňkách Kmenové buňky mají, narozdíl od většiny somatických buňek, aktivní telomerázu. Množství telomerázy je ale malé, takže ztrátu telomer jen zpomaluje, ale není schopná jí zcela kompenzovat. Kmenové buňky se v tkáních nacházejí v nikách. Jak buňky v tkáních odumírají, kmenové buňky proliferují, aby ztrátu kompenzovaly. To ale vede ke zkracování jejich telomer, takže když je dosaženo Hayflickova limitu, kriticky krátké telomery jsou rozpoznávány jako poškozená DNA a kmenové buňky přestanou odpovídat na svou mobilizaci. Tkáň přestane regenerovat, což nakonec vede k selhání orgánu. Pokud se kmenové buňky naučí doplňovat si telomery, může to vést ke vzniku rakoviny. Obrázek z Donate a Blasco 2011 V roce 1994 byla vyvinuta TRAP assay, metoda, která je schopna detekovat přítomnost telomerázy v buňce. S její pomocí se zjistilo, že 85% nádorů obsahuje aktivní telomerázu, která je od té doby předmětem výzkumu protinádorových terapií.
34 Telomery a stárnutí Délka telomeru člověka je kolem bp, ale záležína: -věku(novorozenci majítelomerydelšíi o několik tisíc bazínežstoletístařečci) Hayflickův limit Telomeráza je u člověka aktivní během embryonálního vývoje, čímž je kompenzováno zkracování telomer při masivní proliferaci buněk v rostoucím embryu. U dospělého člověka je telomeráza aktivní jen v některých buňkách. Proto nejdelší telomery má novorozenec a jejich délka se během života zkracuje. 34
35 Telomery a stárnutí Délka telomeru člověka je kolem bp, ale záležína: -věku(novorozenci majítelomerydelšíi o několik tisíc bazínežstoletístařečci) -tkáni Např: Epidermis 33 bp/rok Střevní sliznice 59 bp/rok Játra 120 bp/rok Endotel 147 bp/rok Srdce nedetekovatelné Mozek - nedetekovatelné
36 Telomery a stárnutí Délka telomeru člověka je kolem bp, ale záležína: -věku(novorozenci majítelomerydelšíi o několik tisíc bazínežstoletístařečci) -tkáni - genetické dispozici
37 Těžký život bez telomerázy Myši s deaktivovanou telomerázou měly: - kratší život - nestabilní genom - zvýšený výskyt nádorů - zkrácené telomery Mnoho informací o roli telomerázy pochází z myší, které měly deletovanou RNA komponentu z telomerázy. I když chybějící telomeráza nezpůsobovala všechny projevy stárnutí, myši některé projevy měly: zkrácené telomery, genetickou nestabilitu, sníženou reakci na stimulaci hematopoezy, kratší život a zvýšené množství maligních nádorů.
38 Těžký život bez telomerázy Lidská onemocnění způsobena nedostatkem funkční telomerázy: Kongenitální dyskeratóza - pigmentové skvrny na kůži, bílé skvrny v ústech, anémie - úmrtí většinou na selhání kostní dřeně - způsobena mutacemi v genech pro telomerázu nebo asociované proteiny Aplastická anémie -útlum kostnídřeně málokrevních buněk - může být způsobena mutacemi v genech pro telomerázu Idiopatická plicní fibróza - zjizvení plic snížená kapacita chronický kašel, zadýchávání - může být způsobena mutacemi v genech pro telomerázu Dyskeratosis congenita (DKC, Kongenitální dyskeratóza) je dědičné onemocnění, které se projevuje hyperpigmentací (tmavé skvrny na kůži), orální leukoplakií (bílé skvrny v ústech a nehtovou dystrofií (chybějící nehty). Úmrtí ve většině případů nastává selháním kostní dřeně, dále imunodeficiencí, dýchacími obtížemi a rakovinou. Autosomálně dominantní forma je způsobena mutací v genech kódujících TR nebo TERT, které redukují aktivitu telomerázy a tím i schopnost proliferace kmenových buněk. To vede i nízkému množství bílých a červených krvinek (aplastická anemie). Rodiny s DKC vykazují zhoršování projevů z generace na generaci kvůli zkracujícím se teloomerám, které jsou předávány potomkům. Aplastic anemia (AA) je většinou získaná choroba, ale vzácně s objevují rodiny, kde je děděná. Její příčinou je pak mutace v genech pro TR nebo TERT. Často je spojena s DKC, ale v některých případech chybějí typické projevy. Při AA má pacient malé množství krevních buněk a smrt nastává většinou v důsledku selhání kostní dřeně. Idiopathic pulmonary fibrosis (IPF) idiopathic = vyskytující se spontánně bez jasné příčiny IPF je onemocnění, jehož příčina je často neznámá. Projevuje se vazivovými jizvami na plicích, což zmenšuje jejich kapacitu. Symptomy jsou chronický kašel a zadýchání bez fyzické námahy. V některých rodinách je toto onemocnění dědičné a je způsobeno mutací v genech pro TERT a TR.
39 Další choroby spojené s redukcí telomer Lidskáonemocněníbez mutacív telomeráze, ale s krátkými telomerami, např: Wernerův syndrom -předčasnéstárnutípo 10. roce (šedivévlasy, slabákůže, diabetes, arteroskleróza, rakovina) - rychlé zkracování telomer a nestabilita genomu - nejčastější příčina smrti infarkt nebo rakovina -mutace vgenu WRN(DNA helikáza) interagujes telomerickýmproteinem Trf2 rozplétánítelomerpři replikaci? Wernerův syndrom WS je autosomálně recesivní onemocněním jehož jedinou známou příčinou je mutace v genu WRN, který kóduje DNA helikázu. Při tomto onemocnění bylo pozorováno rychlé zkracování telomer a nestabilita genomu. První příznaky nastávají obvykle po 10. roce, po pubertě následuje rychlé stárnutí. Projevy jsou: chybějící růstové sprouty, šedivění vlasů, slabá kůže, diabetes, katarakta, hypogonadismus, arteroskleróza a rakovina. Častou příčinou smrti je infarkt nebo rakovina. Není známo propojení mezi funkcí helikázy a telomerami, ale tato helikáza má silnou afinitu k telomerickému proteinu Trf2 (TTAGGG repeat binding factor 2) a její funkcí by mohlo být rozplétání telomer pro replikaci (Opresko et al. 2002)
40 Další choroby spojené s redukcí telomer Lidskáonemocněníbez mutacív telomeráze, ale s krátkými telomerami, např: Fanconiho anemie -vzácnágenetickáchoroba, průměrnádoba života 30 let -způsobenávyřazením některého z15 genůpro opravu DNA -často nízkápostava, abnormality kůže, rukou, hlavy,..., - rakovina (nejčastěji AML) -telomeryse zkracujíodlamováním spojováníchromosomů nestabilita genomu Fanconiho anemie FA je vzácná genetická choroba (častější u Židů Aškenazi a v Jižní Africe), způsobená vyřazením některého z 15 genů, které se účastní reparace DNA. Většina pacientů dostane rakovinu (většinou akutní myeloidní leukemii), průměrný věk úmrtí je 30 let. Většina pacientů má i další problémy, často nízká postava, abnormality kůže, rukou, hlavy,... U FA bylo prokázáno, že dochází k progresivnímu zkracování telomer vlivem zlomů v telomerických oblastech (fragmenty telomer pozorovány po FISH mimo chromosomy), důsledkem je 10x vyšší frekvence spojování chromosomů než u zdravých lidí, a tím i vysoká nestabilita genomu (Callén et al. 2002).
41 Telomery a stárnutí Délka telomeru člověka je kolem bp, ale záležína: -věku(novorozenci majítelomerydelšíi o několik tisíc bazínežstoletístařečci) -tkáni - genetické dispozici - životním stylu
42 Negetickéfaktory ovlivňujícídélku telomer Vysoký krevní tlak Kardiovaskulární choroby Obezita Zánět tlustého střeva Dlouhodobé vystavení cigaretovému kouři Dlouhodobý stres Na délku telomer u člověka mají vliv i negenetické faktory prostředí a biochemie. Na krátké telomery mají vliv ulcerózní kolititida (zánět tlustého střeva), obezita, vysoký krevní tlak, kardiovaskulární choroby a chronické vystavení cigaretovému kouři. Rovněž vysoká produkce stresových hormonů (glukokortikoidů) má negativní vliv na udržování telomer. E. Blackburn a její tým ukázali, že staří lidé s krátkými telomerami mají kratší život a méně zdravých let.
43 Směry výzkumu telomer Telomery a stárnutí potenciálníaplikace v kosmetice krémy/nápoje aktivující/dodávající telomerázu -genováterapie -aktivace telomerázyv dospělých somatických buňkách problematické, riziko rakoviny
44 Směry výzkumu telomer Telomery a stárnutí potenciálníaplikace v kosmetice krémy aktivující/dodávající telomerázu -genováterapie -aktivace telomerázyv dospělých somatických buňkách problematické, riziko rakoviny Klonování zvířat ze somatických buněk - délka telomer klonovaných zvířat je různá - záleží na použité buňce
45 Telomery a klonování savců Dolly prvnívytvořený savčíklon -narozenav r klon z buňky mléčné žlázy Dolly zemřela mladá(6,5 roku místo let) Její telomery byly kratší, než odpovídalo věku Dolly byla biologicky starší než její skutečný věk! Robert Lanzaa kolektiv (2000): 6 telat klonovanýchz dělících se nediferencovaných buněk (fibroblastů) - dle délky telomer dokonce biologicky mladší!!! Klonování savců se provádí tak, že ze somatické buňky dospělého jedince je odebráno jádro, které je vloženo do vajíčka zbaveného vlastního pronuklea. Z této diploidní buňky se začne vyvíjet embryo, které je implantováno náhradní matce, která je donosí a porodí. Mládě nese stejnou genetickou informaci jako jedinec, kterému byla odebrána somatická buňka. Klonování savců je problematické, protože má velmi malou účinnost a i narozené klony často trpí zdravotními problémy nebo umírají bez zjevné příčiny. Prvním klonovaným savcem byla ovce Dolly, která vnikla ze somatické buňky odebrané z vemene dospělé ovce. Dolly zemřela poměrně mladá (cca v polovině věku, kterého se ovce dožívají). Při analýze délky jejích telomer se ukázalo, že neodpovídají věku Dolly, ale věku Dolly plus věku ovce, ze které pocházela somatická buňka použitá ke klonování. Na druhou stranu to, že klon bude mít krátké telomery, není pravidlem. Jiná práce, popisující klonování krávy, ukázala, že narozené klony měly telomery ještě delší, než by měla přirozeně počatá telata (Lanza et al. 2000). 45
46 Směry výzkumu telomer Telomery a stárnutí potenciálníaplikace v kosmetice krémy aktivující/dodávající telomerázu -genováterapie -aktivace telomerázyv dospělých somatických buňkách problematické, riziko rakoviny Klonování zvířat ze somatických buněk - délka telomer klonovaných zvířat je různá - záleží na použité buňce Telomery a rakovina -inhibice telomerázyv nádorech často toxickéin vivo nebo neefektivní - nejslibnější očkování proti telomeráze Vývoj léků Protože telomeráza a udržování telomer hraje roli ve vývoji různých malignit, pracuje se na vývoji léků, které cílí na telomerázu nebo telomery. Navzdory velkému úsilí jen minimum léků se dostalo až do klinických zkoušek, protože většina vykazovala buď malou efektivitu nebo toxicitu in vivo. Roli hraje samozřejmě i to, že o regulaci telomerázy a jejích funkcích víme stále velice málo. Zatím nejslibnější je vakcinace pacientů s rakovinou proti htert epitopům (Vonderheide 2008/2009). 46
47 Prodlužování telomer rekombinací aneb proč nás vypnutí telomerázy neochrání před rakovinou Alternativnímechanismus prodlužovánítelomer(alt): REKOMBINACE -nalezen i u člověka Nepotřebují telomerázu 85% lidských nádorů má aktivní telomerázu zbytek používá alternativní prodlužování telomer V roce 1990 byl u kvasinek objeven způsob prodlužování telomer nezávislý na telomeráze, v roce 1994 bylo totéž objeveno u savců. Ukázalo se, že 10-15% nesmrtelných buněčných linií tento ALT (Alternative Lengthening of Telomeres) systém využívá. Protože 85% lidských nádorů má aktivní telomerázu, je tato slibným cílem pro nádorovou terapii, nicméně už teď je jasné, že na zbývajících 15% tato terapie fungovat nebude. 47
48 Co si odnést z této přednášky 1) Centromery místo nasedáníkinetochoru, funkce: bezchybný rozchod chromosomů 2) Monocentrické vs. holokinetické chromosomy 3) DNA centromer rychláevoluce, často repetitivnídna, důležité jsou epigenetické markery 4) Centromericképroteiny řada funkcí(vazba mikrotubulů, motorové proteiny, kontrolní bod,...) 5) NOR (organizátor jadérka) rdna, různépočty + různý počet genů 6) Jadérko transkripce rdna, jen v interfázi a profázi
49 Co si odnést z této přednášky 7) Telomery konce eukaryotních chromosomů, jsou nezbytné pro fungováníbuňky (replikace konců, nelepivékonce, Hayflickův limit). 8) Telomeryjsou složenéz DNA a proteinů. Sekvence telomerickédna je často společnápro velkéskupiny organismů. Ale existují výjimky! 9) Telomeráza, enzym prodlužující telomery, je u člověka aktivní pouze během embryonálního vývoje a v dospělosti v kmenových buňkách. Jejíaktivace v dospělých somatických buňkách může vést k rakovině. 10) Na délku telomermávliv i životnístyl. Takže sportujte, jezte zdravě a nestresujte se.
50 Don t worry, be happy!
Exprese genetické informace
 Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Exprese genetické informace Stavební kameny nukleových kyselin Nukleotidy = báze + cukr + fosfát BÁZE FOSFÁT Nukleosid = báze + cukr CUKR Báze Cyklické sloučeniny obsahující dusík puriny nebo pyrimidiny
Dědičnost pohlaví Genetické principy základních způsobů rozmnožování
 Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
Dědičnost pohlaví Vznik pohlaví (pohlavnost), tj. komplexu znaků, vlastností a funkcí, které vymezují exteriérové i funkční diference mezi příslušníky téhož druhu, je výsledkem velmi komplikované série
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života?
 6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
6. Kde v DNA nalézáme rozdíly, zodpovědné za obrovskou diverzitu života? Pamatujete na to, co se objevilo v pracích Charlese Darwina a Alfreda Wallace ohledně vývoje druhů? Aby mohl mechanismus přírodního
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH. Michaela Nesvadbová
 DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
DNA TECHNIKY IDENTIFIKACE ŽIVOČIŠNÝCH DRUHŮ V KRMIVU A POTRAVINÁCH Michaela Nesvadbová Význam identifikace živočišných druhů v krmivu a potravinách povinností každého výrobce je řádně a pravdivě označit
Globální pohled na průběh replikace dsdna
 Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Globální pohled na průběh replikace dsdna 3' 5 3 vedoucí řetězec 5 3 prodlužování vedoucího řetězce (polymerace ) DNA-ligáza směr pohybu enzymů DNA-polymeráza I DNA-polymeráza III primozom 5' 3, 5, hotový
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk. Aleš Hampl
 Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
Molekulární mechanismy diferenciace a programované buněčné smrti - vztah k patologickým procesům buněk Aleš Hampl Tkáně Orgány Živé buňky, které plní různé funkce (podpora struktury, přijímání živin, lokomoce,
ONKOGENETIKA. Spojuje: - lékařskou genetiku. - buněčnou biologii. - molekulární biologii. - cytogenetiku. - virologii
 ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
ONKOGENETIKA Spojuje: - lékařskou genetiku - buněčnou biologii - molekulární biologii - cytogenetiku - virologii Důležitost spolupráce různých specialistů při detekci hereditárních forem nádorů - (onkologů,internistů,chirurgů,kožních
Dědičnost a pohlaví. KBI/GENE Mgr. Zbyněk Houdek
 Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Dědičnost a pohlaví KBI/GENE Mgr. Zbyněk Houdek Dědičnost pohlavně vázaná Gonozomy se v evoluci vytvořily z autozomů, proto obsahují nejen geny řídící vznik pohlavních rozdílů i další jiné geny. V těchto
Buněčný cyklus. Replikace DNA a dělení buňky
 Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Buněčný cyklus Replikace DNA a dělení buňky 2 Regulace buněčného dělení buněčný cyklus: buněčné dělení buněčný růst kontrola kvality potomstva (dceřinných buněk) bránípřenosu nekompletně zreplikovaných
Molekulární biotechnologie č.8. Produkce heterologního proteinu v eukaryontních buňkách
 Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Molekulární biotechnologie č.8 Produkce heterologního proteinu v eukaryontních buňkách Eukaryontní buňky se využívají v případě, když Eukaryontní proteiny syntetizované v baktériích postrádají biologickou
Spermatogeneze saranče stěhovavé (Locusta migratoria)
") Spermatogeneze saranče stěhovavé (Locusta migratoria) Vývoj pohlavních buněk u živočichů zahrnuje několik dějů, které zajistí, že dojde k redukci a promíchání genetického materiálu a vzniklé buňky jsou
Spermatogeneze saranče stěhovavé (Locusta migratoria) Vývoj pohlavních buněk u živočichů zahrnuje několik dějů, které zajistí, že dojde k redukci a promíchání genetického materiálu a vzniklé buňky jsou
Genetický polymorfismus
 Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Genetický polymorfismus Za geneticky polymorfní je považován znak s nejméně dvěma geneticky podmíněnými variantami v jedné populaci, které se nachází v takových frekvencích, že i zřídkavá má frekvenci
Cytogenetika. chromosom jádro. telomera. centomera. telomera. buňka. histony. páry bazí. dvoušroubovice DNA
 Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
Cytogenetika telomera chromosom jádro centomera telomera buňka histony páry bazí dvoušroubovice DNA Typy chromosomů Karyotyp člověka 46 chromosomů 22 párů autosomů (1-22 od největšího po nejmenší) 1 pár
Populační genetika II
 Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Populační genetika II 4. Mechanismy měnící frekvence alel v populaci Genetický draft (genetické svezení se) Genetický draft = zvýšení frekvence alely díky genetické vazbě s výhodnou mutací. Selekční vymetení
Těsně před infarktem. Jak předpovědět infarkt pomocí informatických metod. Jan Kalina, Marie Tomečková
 Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Těsně před infarktem Jak předpovědět infarkt pomocí informatických metod Jan Kalina, Marie Tomečková Program, osnova sdělení 13,30 Úvod 13,35 Stručně o ateroskleróze 14,15 Měření genových expresí 14,00
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
O původu života na Zemi Václav Pačes
 O původu života na Zemi Václav Pačes Ústav molekulární genetiky Akademie věd ČR centrální dogma replikace transkripce DNA RNA protein reverzní transkripce translace informace funkce Exon 1 Intron (413
O původu života na Zemi Václav Pačes Ústav molekulární genetiky Akademie věd ČR centrální dogma replikace transkripce DNA RNA protein reverzní transkripce translace informace funkce Exon 1 Intron (413
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince. Co bylo dřív? Slepice nebo vejce?
 REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
REPRODUKCE A ONTOGENEZE Od spermie s vajíčkem až po zralého jedince Co bylo dřív? Slepice nebo vejce? Rozmnožování Rozmnožování (reprodukce) může být nepohlavní (vegetativní, asexuální) pohlavní (sexuální;
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. MBIO1/Molekulární biologie 1 Tento projekt je spolufinancován
NUKLEOVÉ KYSELINY. Základ života
 NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
NUKLEOVÉ KYSELINY Základ života HISTORIE 1. H. Braconnot (30. léta 19. století) - Strassburg vinné kvasinky izolace matiére animale. 2. J.F. Meischer - experimenty z hnisem štěpení trypsinem odstředěním
Genetika pohlaví genetická determinace pohlaví
 Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
Genetika pohlaví Genetická determinace pohlaví Způsoby rozmnožování U nižších organizmů může docházet i k ovlivnění pohlaví jedince podmínkami prostředí (např. teplotní závislost pohlavní determinace u
BUŇEČNÝ CYKLUS A JEHO KONTROLA
 BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
BUŇEČNÝ CYKLUS A JEHO KONTROLA MITOSA - fáze: Profáze - kondensace chromosomů - 30 nm chromatine fibres vázané na matrix Rozpad Metafáze - párové ( sesterské ) chromatidy - vázané centromerou, seřazené
Tematické okruhy k SZZ v bakalářském studijním oboru Zdravotní laborant bakalářského studijního programu B5345 Specializace ve zdravotnictví
 Tematické okruhy k SZZ v bakalářském studijním oboru Zdravotní laborant bakalářského studijního programu B5345 Specializace ve zdravotnictví Dle čl. 7 odst. 2 Směrnice děkana pro realizaci bakalářských
Tematické okruhy k SZZ v bakalářském studijním oboru Zdravotní laborant bakalářského studijního programu B5345 Specializace ve zdravotnictví Dle čl. 7 odst. 2 Směrnice děkana pro realizaci bakalářských
Struktura a funkce biomakromolekul
 Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Struktura a funkce biomakromolekul KBC/BPOL 7. Interakce DNA/RNA - protein Ivo Frébort Interakce DNA/RNA - proteiny v buňce Základní dogma molekulární biologie Replikace DNA v E. coli DNA polymerasa a
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/28.0032
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Charakteristika chromozomové výbavy 2n = 46,XY Karyotyp - Karyogram - Idiogram
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 Charakteristika chromozomové výbavy 2n = 46,XY Karyotyp - Karyogram - Idiogram
http://www.accessexcellence.org/ab/gg/chromosome.html
 3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
3. cvičení Buněčný cyklus Mitóza Modifikace mitózy 1 DNA, chromosom genetická informace organismu chromosom = strukturní podoba DNA během dělení (mitózy) řetězec DNA (chromonema) histony další enzymatické
Bakteriální transpozony
 Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Bakteriální transpozony Transpozon = sekvence DNA schopná transpozice, tj. přemístění z jednoho místa v genomu do jiného místa Transpozice = proces přemístění transpozonu Transponáza (transpozáza) = enzym
Vzdělávací materiál. vytvořený v projektu OP VK CZ.1.07/1.5.00/34.0211. Anotace. Biosyntéza nukleových kyselin. VY_32_INOVACE_Ch0219.
 Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Vzdělávací materiál vytvořený v projektu OP VK Název školy: Gymnázium, Zábřeh, náměstí Osvobození 20 Číslo projektu: Název projektu: Číslo a název klíčové aktivity: CZ.1.07/1.5.00/34.0211 Zlepšení podmínek
Genetická kontrola prenatáln. lního vývoje
 Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Genetická kontrola prenatáln lního vývoje Stádia prenatáln lního vývoje Preembryonální stádium do 6. dne po oplození zygota až blastocysta polární organizace cytoplasmatických struktur zygoty Embryonální
Radiobiologický účinek záření. Helena Uhrová
 Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Radiobiologický účinek záření Helena Uhrová Fáze účinku fyzikální fyzikálně chemická chemická biologická Fyzikální fáze Přenos energie na e Excitace molekul, ionizace Doba trvání 10-16 - 10-13 s Fyzikálně-chemická
Základy molekulární a buněčné biologie. Přípravný kurz Komb.forma studia oboru Všeobecná sestra
 Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Základy molekulární a buněčné biologie Přípravný kurz Komb.forma studia oboru Všeobecná sestra Genetický aparát buňky DNA = nositelka genetické informace - dvouvláknová RNA: jednovláknová mrna = messenger
Zaměření bakalářské práce (témata BP)
") Zaměření bakalářské práce (témata BP) Obor: Buněčná a molekulární diagnostika - zadává katedra - studenti si témata losují Obor: molekulární biologie a genetika - témata BP vychází z vybraného tématu DP
Zaměření bakalářské práce (témata BP) Obor: Buněčná a molekulární diagnostika - zadává katedra - studenti si témata losují Obor: molekulární biologie a genetika - témata BP vychází z vybraného tématu DP
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno
 Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Co nás učí nádory? Prof. RNDr. Jana Šmardová, CSc. Ústav patologie FN Brno Přírodovědecká a Lékařská fakulta MU Brno Brno, 17.5.2011 Izidor (Easy Door) Osnova přednášky 1. Proč nás rakovina tolik zajímá?
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Vztah struktury a funkce nukleových kyselin. Replikace, transkripce
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Vztah struktury a funkce nukleových kyselin. Replikace, transkripce Nukleová kyselina gen základní jednotka informace v živých systémech,
EPIGENETIKA reverzibilních změn funkce genů, Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
 EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
EPIGENETIKA Epigenetika se zabývá studiem reverzibilních změn funkce genů, aniž by při tom došlo ke změnám v sekvenci jaderné DNA. Epigenetické faktory ovlivňují fenotyp bez změny genotypu. Epigenetická
Věda v prostoru. Voda v pohybu. Buněční detektivové. Svědkové dávné minulosti Země
 6+ Věda v prostoru Jak vědci pracují v laboratoři? Proč je zelená víc než jen obyčejná barva? Jak můžeme použít prášek do pečiva ke sfouknutí svíčky? Získejte odpovědi na všechny otázky v tomto vzrušujícím
6+ Věda v prostoru Jak vědci pracují v laboratoři? Proč je zelená víc než jen obyčejná barva? Jak můžeme použít prášek do pečiva ke sfouknutí svíčky? Získejte odpovědi na všechny otázky v tomto vzrušujícím
UNIVERZITA KARLOVA V PRAZE 3. LÉKAŘSKÁ FAKULTA (tématické okruhy požadavků pro přijímací zkoušku)
") UNIVERZITA KARLOVA V PRAZE 3. LÉKAŘSKÁ FAKULTA (tématické okruhy požadavků pro přijímací zkoušku) B I O L O G I E 1. Definice a obory biologie. Obecné vlastnosti organismů. Základní klasifikace organismů.
UNIVERZITA KARLOVA V PRAZE 3. LÉKAŘSKÁ FAKULTA (tématické okruhy požadavků pro přijímací zkoušku) B I O L O G I E 1. Definice a obory biologie. Obecné vlastnosti organismů. Základní klasifikace organismů.
BAKTERIÁLNÍ GENETIKA. Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc.
 BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
BAKTERIÁLNÍ GENETIKA Lekce 12 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. -dědičnost u baktérií principiálně stejná jako u komplexnějších organismů -genom haploidní a značně menší Bakteriální genom
GENETIKA 1. Úvod do světa dědičnosti. Historie
 GENETIKA 1. Úvod do světa dědičnosti Historie Základní informace Genetika = věda zabývající se dědičností a proměnlivostí živých soustav sleduje variabilitu (=rozdílnost) a přenos druhových a dědičných
GENETIKA 1. Úvod do světa dědičnosti Historie Základní informace Genetika = věda zabývající se dědičností a proměnlivostí živých soustav sleduje variabilitu (=rozdílnost) a přenos druhových a dědičných
Digitální učební materiál
 Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Odborná biologie, část biologie Společná pro
Digitální učební materiál Projekt CZ.1.07/1.5.00/34.0415 Inovujeme, inovujeme Šablona III/2 Inovace a zkvalitnění výuky prostřednictvím ICT (DUM) Tematická Odborná biologie, část biologie Společná pro
Transpozony - mobilní genetické elementy
 Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
Transpozony - mobilní genetické elementy Tvoří pravidelnou součást genomu prokaryot i eukaryot (až 50% genomu) Navozují mutace genů (inzerční inaktivace, polární mutace, změny exprese genů) Jsou zodpovědné
44 somatických chromozomů pohlavní hormony (X,Y) 46 chromozomů
 46 chromozomů") Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Buněčný cyklus MUDr.Kateřina Kapounková Inovace studijního oboru Regenerace a výţiva ve sportu (CZ.107/2.2.00/15.0209) 1 DNA,geny genom = soubor všech genů a všechna DNA buňky; kompletní genetický materiál
Genetický screening predispozice k celiakii
 VETERINÁRN RNÍ A FARMACEUTICKÁ UNIVERZITA BRNO Farmaceutická fakulta Ústav humánn nní farmakologie a toxikologie Genetický screening predispozice k celiakii RNDr. Ladislava Bartošov ová,ph.d. 1, PharmDr.
VETERINÁRN RNÍ A FARMACEUTICKÁ UNIVERZITA BRNO Farmaceutická fakulta Ústav humánn nní farmakologie a toxikologie Genetický screening predispozice k celiakii RNDr. Ladislava Bartošov ová,ph.d. 1, PharmDr.
2. Z následujících tvrzení, týkajících se prokaryotické buňky, vyberte správné:
 Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
Výběrové otázky: 1. Součástí všech prokaryotických buněk je: a) DNA, plazmidy b) plazmidy, mitochondrie c) plazmidy, ribozomy d) mitochondrie, endoplazmatické retikulum 2. Z následujících tvrzení, týkajících
7. Regulace genové exprese, diferenciace buněk a epigenetika
 7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
7. Regulace genové exprese, diferenciace buněk a epigenetika Aby mohl mnohobuněčný organismus efektivně fungovat, je třeba, aby se jednotlivé buňky specializovaly na určité funkce. Nový jedinec přitom
Struktura chromatinu. Co je to chromatin?
 Struktura chromatinu Buněčné jádro a genová exprese Lenka Rossmeislová struktura-význam-modifikace Co je to chromatin? hmota, ze které jsou vytvořeny chromozomy DNA asociovaná s proteiny, které napomáhají
Struktura chromatinu Buněčné jádro a genová exprese Lenka Rossmeislová struktura-význam-modifikace Co je to chromatin? hmota, ze které jsou vytvořeny chromozomy DNA asociovaná s proteiny, které napomáhají
Apoptóza. Veronika Žižková. Ústav klinické a molekulární patologie a Laboratoř molekulární patologie
 Apoptóza Veronika Žižková Ústav klinické a molekulární patologie a Laboratoř molekulární patologie Apoptóza Úvod Apoptóza vs nekróza Role apoptózy v organismu Mechanismus apoptózy Metody detekce Úvod -
Apoptóza Veronika Žižková Ústav klinické a molekulární patologie a Laboratoř molekulární patologie Apoptóza Úvod Apoptóza vs nekróza Role apoptózy v organismu Mechanismus apoptózy Metody detekce Úvod -
Typy nukleových kyselin. deoxyribonukleová (DNA); ribonukleová (RNA).
; ribonukleová (RNA).") Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Typy nukleových kyselin Existují dva typy nukleových kyselin (NA, z anglických slov nucleic acid): deoxyribonukleová (DNA); ribonukleová (RNA). DNA je lokalizována v buněčném jádře, RNA v cytoplasmě a
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Inovace studia molekulární a buněčné biologie I n v e s t i c e d o r o z v o j e v z d ě l á v á n í reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Modul IB. Histochemie. CBO Odd. histologie a embryologie. MUDr. Martin Špaček
 Modul IB Histochemie CBO Odd. histologie a embryologie MUDr. Martin Špaček Histochemie Histologická metoda užívaná k průkazu různých látek přímo v tkáních a buňkách Histochemie Katalytická histochemie
Modul IB Histochemie CBO Odd. histologie a embryologie MUDr. Martin Špaček Histochemie Histologická metoda užívaná k průkazu různých látek přímo v tkáních a buňkách Histochemie Katalytická histochemie
Přípravný kurz z biologie MUDr. Jana Kolářová, CSc. témata 1 Mgr. Kateřina Caltová témata 3-5 doc. PharmDr. Emil Rudolf, Ph.D. 2 + 6-10 materiály k
 Přípravný kurz z biologie MUDr. Jana Kolářová, CSc. témata 1 Mgr. Kateřina Caltová témata 3-5 doc. PharmDr. Emil Rudolf, Ph.D. 2 + 6-10 materiály k přípravnému kurzu: stránka Ústavu lékařské biologie a
Přípravný kurz z biologie MUDr. Jana Kolářová, CSc. témata 1 Mgr. Kateřina Caltová témata 3-5 doc. PharmDr. Emil Rudolf, Ph.D. 2 + 6-10 materiály k přípravnému kurzu: stránka Ústavu lékařské biologie a
Vytvořilo Oddělení lékařské genetiky FN Brno
 GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
GONOSOMY GONOSOMY CHROMOSOMY X, Y Obr. 1 (Nussbaum, 2004) autosomy v chromosomovém páru homologní po celé délce chromosomů crossingover MEIÓZA Obr. 2 (Nussbaum, 2004) GONOSOMY CHROMOSOMY X, Y ODLIŠNOSTI
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA
 RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
RIGORÓZNÍ OTÁZKY - BIOLOGIE ČLOVĚKA 1. Genotyp a jeho variabilita, mutace a rekombinace Specifická imunitní odpověď Prevence a časná diagnostika vrozených vad 2. Genotyp a prostředí Regulace buněčného
Kontrola genové exprese
 Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
Základy biochemie KBC/BC Kontrola genové exprese Inovace studia biochemie prostřednictvím e-learningu CZ.04.1.03/3.2.15.3/0407 Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem
od eukaryotické se liší svou výrazně jednodušší stavbou a velikostí Dosahuje velikosti 1-10 µm. Prokaryotní buňku mají bakterie a sinice skládá se z :
 Otázka: Buňka Předmět: Biologie Přidal(a): konca88 MO BI 01 Buňka je základní stavební jednotka živých organismů. Je to nejmenší živý útvar schopný samostatné existence a rozmnožování. Každá buňka má svůj
Otázka: Buňka Předmět: Biologie Přidal(a): konca88 MO BI 01 Buňka je základní stavební jednotka živých organismů. Je to nejmenší živý útvar schopný samostatné existence a rozmnožování. Každá buňka má svůj
Říše Prvoci. (Protozoa) Autor: Katka www.nasprtej.cz Téma: Prvoci Ročník: 2. Opora, ochrana. Pohyb. o Pouze pokud nemají pelikulu.
 Autor: Katka www.nasprtej.cz Téma: Prvoci Ročník: 2. Opora, ochrana. Pohyb. o Pouze pokud nemají pelikulu.") Říše Prvoci (Protozoa) - Mikroorganismy - Jednobuněční - Jedná se o živočišnou buňku s dalšími (rozšiřujícími) strukturami Opora, ochrana - Pelikula - tuhá blanka na povrchu (nemají ji měňavky) - Schránka
Říše Prvoci (Protozoa) - Mikroorganismy - Jednobuněční - Jedná se o živočišnou buňku s dalšími (rozšiřujícími) strukturami Opora, ochrana - Pelikula - tuhá blanka na povrchu (nemají ji měňavky) - Schránka
Struktura a organizace genomů
 CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
CG020 Genomika Přednáška 8 Struktura a organizace genomů Markéta Pernisová Funkční genomika a proteomika rostlin, Mendelovo centrum genomiky a proteomiky rostlin, Středoevropský technologický institut
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1
 Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
Beličková 1, J Veselá 1, E Stará 1, Z Zemanová 2, A Jonášová 2, J Čermák 1 1 Ústav hematologie a krevní transfuze, Praha 2 Všeobecná fakultní nemocnice, Praha MDS Myelodysplastický syndrom (MDS) je heterogenní
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti. Translace, techniky práce s DNA
 Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Evropský sociální fond Praha & EU: Investujeme do vaší budoucnosti Translace, techniky práce s DNA Translace překlad z jazyka nukleotidů do jazyka aminokyselin dá se rozdělit na 5 kroků aktivace aminokyslin
Deriváty karboxylových kyselin, aminokyseliny, estery
 Deriváty karboxylových kyselin, aminokyseliny, estery Zpracovala: Ing. Štěpánka Janstová 29.1.2012 Určeno pro 9. ročník ZŠ V/II,EU-OPVK,42/CH9/Ja Přehled a využití derivátů organických kyselin, jejich
Deriváty karboxylových kyselin, aminokyseliny, estery Zpracovala: Ing. Štěpánka Janstová 29.1.2012 Určeno pro 9. ročník ZŠ V/II,EU-OPVK,42/CH9/Ja Přehled a využití derivátů organických kyselin, jejich
Genetika na úrovni mnohobuněčného organizmu
 Genetika na úrovni mnohobuněčného organizmu Přenos genetické informace při rozmnožování Nepohlavní rozmnožování: - nový jedinec vzniká ze somatické buňky nebo ze souboru somatických buněk jednoho rodičovského
Genetika na úrovni mnohobuněčného organizmu Přenos genetické informace při rozmnožování Nepohlavní rozmnožování: - nový jedinec vzniká ze somatické buňky nebo ze souboru somatických buněk jednoho rodičovského
Molekulární procesy po fertilizacinormální či abnormální po ART?
 Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
Molekulární procesy po fertilizacinormální či abnormální po ART? Aleš Hampl Již více jak MILION dětí bylo na světě počato pomocí ART ART jako zdroj zvýšeného rizika:? Kongenitální malformace (Ericson and
Základní škola a Mateřská škola G.A.Lindnera Rožďalovice. Za vše mohou geny
 Základní škola a Mateřská škola G.A.Lindnera Rožďalovice Za vše mohou geny Jméno a příjmení: Sandra Diblíčková Třída: 9.A Školní rok: 2009/2010 Garant / konzultant: Mgr. Kamila Sklenářová Datum 31.05.2010
Základní škola a Mateřská škola G.A.Lindnera Rožďalovice Za vše mohou geny Jméno a příjmení: Sandra Diblíčková Třída: 9.A Školní rok: 2009/2010 Garant / konzultant: Mgr. Kamila Sklenářová Datum 31.05.2010
Typy chromosomů. A telocentrický B akrocentrický C submetacentrický D metacentrický. Člověk nemá typ telocentrický!
 Karyologie Typy chromosomů A telocentrický B akrocentrický C submetacentrický D metacentrický Člověk nemá typ telocentrický! Chromosom chromosom telomera jádro centomera telomera buňka histony dvoušroubovice
Karyologie Typy chromosomů A telocentrický B akrocentrický C submetacentrický D metacentrický Člověk nemá typ telocentrický! Chromosom chromosom telomera jádro centomera telomera buňka histony dvoušroubovice
Svět RNA a bílkovin. RNA svět, 1. polovina. RNA svět. Doporučená literatura. Struktura RNA. Transkripce. Regulace transkripce.
 RNA svět, 1. polovina Struktura RNA Regulace transkripce Zrání pre-mrna Svět RNA a bílkovin Sestřih pre-mrna Transport a lokalizace RNA Stabilita RNA Doporučená literatura RNA svět Alberts B., et al.:
RNA svět, 1. polovina Struktura RNA Regulace transkripce Zrání pre-mrna Svět RNA a bílkovin Sestřih pre-mrna Transport a lokalizace RNA Stabilita RNA Doporučená literatura RNA svět Alberts B., et al.:
Oligobiogenní prvky bývají běžnou součástí organismů, ale v těle jich již podstatně méně (do 1%) než prvků makrobiogenních.
 než prvků makrobiogenních.") 1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
1 (3) CHEMICKÉ SLOŢENÍ ORGANISMŮ Prvky Stejné prvky a sloučeniny se opakují ve všech formách života, protože mají shodné principy stavby těla i metabolismu. Např. chemické děje při dýchání jsou stejné
Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN,
 Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN, CHRÓM, Calcium, Magnesium Organické Minerály ORGANICKÉ MINERÁLY Zásadní zvláštností všech přípravků linie «Organické minerály»
Česko ORGANICKÉ MINERÁLY BIOGENNÍ PRVKY VÁPNÍK, ŽELEZO, JÓD, ZINEK, SELÉN, CHRÓM, Calcium, Magnesium Organické Minerály ORGANICKÉ MINERÁLY Zásadní zvláštností všech přípravků linie «Organické minerály»
562 VESMÍR 79, říjen 2000 http://www.cts.cuni.cz/vesmir. 1. Nově replikovaná DNA lidské buňky HeLa značená modifikovaným
 1 1. Nově replikovaná DNA lidské buňky HeLa značená modifikovaným nukleotidem. Buněčné jádro v pozdní fázi S buněčného cyklu je narušeno působením detergentu. Z obrázku je patrná původní akumulace DNA
1 1. Nově replikovaná DNA lidské buňky HeLa značená modifikovaným nukleotidem. Buněčné jádro v pozdní fázi S buněčného cyklu je narušeno působením detergentu. Z obrázku je patrná původní akumulace DNA
VAKUOLA. membránou ohraničený váček membrána se nazývá tonoplast. běžná u rostlin, zvířata specializované funkce či její nepřítomnost
 VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
VAKUOLA membránou ohraničený váček membrána se nazývá tonoplast běžná u rostlin, zvířata specializované funkce či její nepřítomnost VAKUOLA Funkce: uložiště odpadů a uskladnění chemických látek (fenolické
EMBRYOLOGIE Učebnice pro studenty lékařství a oborů všeobecná sestra a porodní asistentka
 6pt;font-style:normal;color:grey;font-family:Verdana,Geneva,Kalimati,sans-serif;text-decoration:none;text-align:center;font-variant:n = = < p s t y l e = " p a d d i n g : 0 ; b o r d e r : 0 ; t e x t
6pt;font-style:normal;color:grey;font-family:Verdana,Geneva,Kalimati,sans-serif;text-decoration:none;text-align:center;font-variant:n = = < p s t y l e = " p a d d i n g : 0 ; b o r d e r : 0 ; t e x t
Tematický plán učiva BIOLOGIE
 Tematický plán učiva BIOLOGIE Třída: Prima Počet hodin za školní rok: 66 h 1. POZNÁVÁME PŘÍRODU 2. LES 2.1 Rostliny a houby našich lesů 2.2 Lesní patra 2.3 Živočichové v lesích 2.4 Vztahy živočichů a rostlin
Tematický plán učiva BIOLOGIE Třída: Prima Počet hodin za školní rok: 66 h 1. POZNÁVÁME PŘÍRODU 2. LES 2.1 Rostliny a houby našich lesů 2.2 Lesní patra 2.3 Živočichové v lesích 2.4 Vztahy živočichů a rostlin
Chromosomy a karyotyp člověka
 Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Chromosomy a karyotyp člověka Chromosom - 1 a více - u eukaryotických buněk uložen v jádře karyotyp - soubor všech chromosomů v jádře jedné buňky - tvořen z vláknem chromatinem = DNA + histony - malé bazické
Lze HCM vyléčit? Jak dlouho žije kočka s HCM? Je možné předejít hypertrofické kardiomyopatii?
 Nemoci srdce jsou, stejně jako u člověka, vrozené nebo získané v průběhu života. Ze získaných chorob srdce tvoří velkou část kardiomyopatie, což je onemocnění srdečního svalu spojené s jeho dysfunkcí,
Nemoci srdce jsou, stejně jako u člověka, vrozené nebo získané v průběhu života. Ze získaných chorob srdce tvoří velkou část kardiomyopatie, což je onemocnění srdečního svalu spojené s jeho dysfunkcí,
Biologie I. Buňka II. Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings
 Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Biologie I Buňka II Campbell, Reece: Biology 6 th edition Pearson Education, Inc, publishing as Benjamin Cummings BUŇKA II centrioly, ribosomy, jádro endomembránový systém semiautonomní organely peroxisomy
Telomery začátky a konce chromozomů. Jiří Fajkus
 Telomery začátky a konce chromozomů Jiří Fajkus Osnova: 1. Úvodní část (drozofila, kukuřice, Hayflickův limit) 2. Nástup molekulární biologie telomer 3. Příklady z praxe 4. Závěr H. J. Muller (1938) -
Telomery začátky a konce chromozomů Jiří Fajkus Osnova: 1. Úvodní část (drozofila, kukuřice, Hayflickův limit) 2. Nástup molekulární biologie telomer 3. Příklady z praxe 4. Závěr H. J. Muller (1938) -
Inovace studia molekulární a buněčné biologie
 Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
Inovace studia molekulární a buněčné biologie Tento projekt je spolufinancován Evropským sociálním fondem a státním rozpočtem České republiky. OBVSB/Obecná virologie Tento projekt je spolufinancován Evropským
REKOMBINACE Přestavby DNA
 REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
REKOMBINACE Přestavby DNA variace v kombinacích genů v genomu adaptace evoluce 1. Obecná rekombinace ( General recombination ) Genetická výměna mezi jakýmkoli párem homologních DNA sekvencí - často lokalizovaných
Gibbsovo samplování a jeho využití
 Gibbsovo samplování a jeho využití Regulace genů Hlavní pozornost výzkumů DNA je většinou věnována analýze genů Geny tvoří pouhá 3% lidské DNA Ukazuje se, že zbývající junk DNA má také velký význam Obsahuje
Gibbsovo samplování a jeho využití Regulace genů Hlavní pozornost výzkumů DNA je většinou věnována analýze genů Geny tvoří pouhá 3% lidské DNA Ukazuje se, že zbývající junk DNA má také velký význam Obsahuje
Crossing-over. over. synaptonemální komplex
 Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
Genetické mapy Crossing-over over v průběhu profáze I meiózy princip rekombinace genetického materiálu mezi maternálním a paternálním chromosomem synaptonemální komplex zlomy a nová spojení chromatinových
V roce 1981 byly v USA poprvé popsány příznaky nového onemocnění, které později dostalo jméno AIDS /Acquired Immune Deficiency Syndrome/ neboli
 Lenka Klimešová V roce 1981 byly v USA poprvé popsány příznaky nového onemocnění, které později dostalo jméno AIDS /Acquired Immune Deficiency Syndrome/ neboli Syndrom získaného imunodeficitu. V roce 1983
Lenka Klimešová V roce 1981 byly v USA poprvé popsány příznaky nového onemocnění, které později dostalo jméno AIDS /Acquired Immune Deficiency Syndrome/ neboli Syndrom získaného imunodeficitu. V roce 1983
ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA
 učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
učební texty Univerzity Karlovy v Praze ZÁKLADY BIOLOGIE a GENETIKY ČLOVĚKA Berta Otová Romana Mihalová KAROLINUM Základy biologie a genetiky člověka doc. RNDr. Berta Otová, CSc. MUDr. Romana Mihalová
Buněčné dělení ŘÍZENÍ BUNĚČNÉHO CYKLU
 BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
BUNĚČNÝ CYKLUS Buněčné dělení Cykliny a na cyklinech závislé proteinkinázy (Cyclin- Dependent Protein Kinases; Cdk-proteinkinázy) - proteiny, které jsou součástí řídícího systému buněčného cyklu 8 cyklinů
Terapeutické klonování, náhrada tkání a orgánů
 Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Transfekce, elektroporace, retrovirová infekce Vnesení genů Vrstva fibroblastů, LIF Terapeutické klonování, náhrada tkání a orgánů Selekce ES buněk, v nichž došlo k začlenění vneseného genu homologní rekombinací
Název: Hmoto, jsi živá? I
 Název: Hmoto, jsi živá? I Výukové materiály Téma: Obecné vlastnosti živé hmoty Úroveň: střední škola Tematický celek: Obecné zákonitosti přírodovědných disciplín a principy poznání ve vědě Předmět (obor):
Název: Hmoto, jsi živá? I Výukové materiály Téma: Obecné vlastnosti živé hmoty Úroveň: střední škola Tematický celek: Obecné zákonitosti přírodovědných disciplín a principy poznání ve vědě Předmět (obor):
rní tekutinu (ECF), tj. cca 1/3 celkového množstv
, tj. cca 1/3 celkového množstv") Představují tzv. extracelulárn rní tekutinu (ECF), tj. cca 1/3 celkového množstv ství vody v tělet (voda tvoří 65-75% váhy v těla; t z toho 2/3 vody jsou vázanv zané intracelulárn rně) Lymfa (míza) Tkáňový
Představují tzv. extracelulárn rní tekutinu (ECF), tj. cca 1/3 celkového množstv ství vody v tělet (voda tvoří 65-75% váhy v těla; t z toho 2/3 vody jsou vázanv zané intracelulárn rně) Lymfa (míza) Tkáňový
Sylabus témat ke zkoušce z lékařské biologie a genetiky. Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
, význam v medicíně") Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Sylabus témat ke zkoušce z lékařské biologie a genetiky Buněčná podstata reprodukce a dědičnosti Struktura a funkce prokaryot Struktura, reprodukce a rekombinace virů (DNA viry, RNA viry), význam v medicíně
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí. Reg. č.: CZ.1.07/2.2.00/
 Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 KBB/ZGEN Základy genetiky Dana Šafářová KBB/ZGEN Základy genetiky Rozsah: 2+1
Propojení výuky oborů Molekulární a buněčné biologie a Ochrany a tvorby životního prostředí Reg. č.: CZ.1.07/2.2.00/28.0032 KBB/ZGEN Základy genetiky Dana Šafářová KBB/ZGEN Základy genetiky Rozsah: 2+1
DUM č. 1 v sadě. 37. Bi-2 Cytologie, molekulární biologie a genetika
 projekt GML Brno Docens DUM č. 1 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Charakteristika buněčného cyklu eukaryot
projekt GML Brno Docens DUM č. 1 v sadě 37. Bi-2 Cytologie, molekulární biologie a genetika Autor: Martin Krejčí Datum: 02.06.2014 Ročník: 6AF, 6BF Anotace DUMu: Charakteristika buněčného cyklu eukaryot
METABOLISMUS TUKŮ VĚČNĚ DISKUTOVANÉ TÉMA
 METABOLISMUS TUKŮ VĚČNĚ DISKUTOVANÉ TÉMA Ing. Vladimír Jelínek V dnešním kongresovém příspěvku budeme hledat odpovědi na následující otázky: Co jsou to tuky Na co jsou organismu prospěšné a při stavbě
METABOLISMUS TUKŮ VĚČNĚ DISKUTOVANÉ TÉMA Ing. Vladimír Jelínek V dnešním kongresovém příspěvku budeme hledat odpovědi na následující otázky: Co jsou to tuky Na co jsou organismu prospěšné a při stavbě
Výuka genetiky na Přírodovědecké fakultě UK v Praze
 Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Výuka genetiky na Přírodovědecké fakultě UK v Praze Studium biologie na PřF UK v Praze Bakalářské studijní programy / obory Biologie Biologie ( duhový bakalář ) Ekologická a evoluční biologie ( zelený
Lekce 7 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. CYTOGENETIKA
 Lekce 7 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. CYTOGENETIKA Cytogenetika se zabývá studiem organizace genomu a struktury a funkce chromozómů u eukaryot. Genom je souhrn veškeré DNA buňky. Poskytuje
Lekce 7 kurzu GENETIKA Doc. RNDr. Jindřich Bříza, CSc. CYTOGENETIKA Cytogenetika se zabývá studiem organizace genomu a struktury a funkce chromozómů u eukaryot. Genom je souhrn veškeré DNA buňky. Poskytuje
Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354
 I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
I n v e s t i c e d o r o z v o j e v z d ě l á v á n í Inovace studia molekulární a buněčné biologie reg. č. CZ.1.07/2.2.00/07.0354 Tento projekt je spolufinancován Evropským sociálním fondem a státním
Genetika - maturitní otázka z biologie (2)
") Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):
Genetika - maturitní otázka z biologie (2) by jx.mail@centrum.cz - Ned?le, B?ezen 01, 2015 http://biologie-chemie.cz/genetika-maturitni-otazka-z-biologie-2/ Otázka: Genetika I P?edm?t: Biologie P?idal(a):
VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody
 VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody Čtvrté skupenství vody: Hexagonální voda: Na univerzitě ve Washingtonu bylo objeveno čtvrté skupenství vody, což může vysvětlit
VODA S ENERGIÍ Univerzita odhalila tajemství vody Objev hexagonální vody Čtvrté skupenství vody: Hexagonální voda: Na univerzitě ve Washingtonu bylo objeveno čtvrté skupenství vody, což může vysvětlit
Hematologie. Nauka o krvi Klinická hematologie Laboratorní hematologie. -Transfuzní lékařství - imunohematologie. Vladimír Divoký
 Hematologie Nauka o krvi Klinická hematologie Laboratorní hematologie -Transfuzní lékařství - imunohematologie Vladimír Divoký Fyzikální vlastnosti krve 3-4 X více viskózní než voda ph : 7.35 7.45 4-6
Hematologie Nauka o krvi Klinická hematologie Laboratorní hematologie -Transfuzní lékařství - imunohematologie Vladimír Divoký Fyzikální vlastnosti krve 3-4 X více viskózní než voda ph : 7.35 7.45 4-6
Funkce imunitního systému
 Téma: 22.11.2010 Imunita specifická nespecifická,, humoráln lní a buněč ěčná Mgr. Michaela Karafiátová IMUNITA je soubor vrozených a získaných mechanismů, které zajišťují obranyschopnost (rezistenci) jedince
Téma: 22.11.2010 Imunita specifická nespecifická,, humoráln lní a buněč ěčná Mgr. Michaela Karafiátová IMUNITA je soubor vrozených a získaných mechanismů, které zajišťují obranyschopnost (rezistenci) jedince
1 Biochemické animace na internetu
 1 Biochemické animace na internetu V dnešní době patří internet mezi nejužívanější zdroje informací. Velmi často lze pomocí internetu legálně stáhnout řadu již vytvořených výukových materiálů sloužících
1 Biochemické animace na internetu V dnešní době patří internet mezi nejužívanější zdroje informací. Velmi často lze pomocí internetu legálně stáhnout řadu již vytvořených výukových materiálů sloužících
Klonování DNA a fyzikální mapování genomu
 Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
Klonování DNA a fyzikální mapování genomu. Terminologie Klonování je proces tvorby klonů Klon je soubor identických buněk (příp. organismů) odvozených ze společného předka dělením (např. jedna bakteriální
CYTOLOGIE 3. týden. Jádro a jeho komponenty Buněčný cyklus, mitosa, meiosa. Ústav histologie a embryologie
 CYTOLOGIE 3. týden Jádro a jeho komponenty Buněčný cyklus, mitosa, meiosa Ústav histologie a embryologie MUDr. Radomíra Vagnerová, CSc. Předmět: Obecná histologie a obecná embryologie 02241 Přednášky 2.
CYTOLOGIE 3. týden Jádro a jeho komponenty Buněčný cyklus, mitosa, meiosa Ústav histologie a embryologie MUDr. Radomíra Vagnerová, CSc. Předmět: Obecná histologie a obecná embryologie 02241 Přednášky 2.